Способ получения l-лизина (варианты) и рекомбинантная днк, используемая для его осуществления (варианты) - RU2197528C2
Код документа: RU2197528C2
Чертежи
































Описание
Данное изобретение относится к способу получения L-лизина культивированием микроорганизма, полученного модификацией коринеформной бактерии, используемой для ферментативного получения аминокислоты или т.п., при помощи способа, основанного на генной инженерии.
Предпосылки изобретения
L-лизин, который используется в качестве добавки к корму, обычно получают ферментативным способом с
использованием L-лизин-продуцирующего мутантного штамма, принадлежащего к коринеформным бактериям. Различные известные в настоящее время L-лизин-продуцирующие бактерии являются бактериями,
создаваемыми при помощи искусственной мутации из штаммов дикого типа, относящихся к коринеформным бактериям.
Что касается коринеформных бактерий, описаны векторная плазмида, которая автономно реплицируется в бактериальных клетках и имеет маркерный ген устойчивости к лекарственному средству (см. United States Patent No. 4514502), и способ введения гена в бактериальные клетки (например, Japanese Patent Laid-open No. 2-207791). Также описана возможность размножения L-треонин- или L-изолейцин-продуцирующей бактерии с применением описанного выше способа (см. United States Patent No. 4452890 и 4442208). Что касается размножения L-лизин-продуцирующей бактерии, известен способ, в котором ген, участвующий в биосинтезе L-лизина, вводят в векторную плазмиду для амплификации этого гена в бактериальных клетках (например, Japanese Patent Laid-open No. 56-160997).
Известные гены для биосинтеза L-лизина включают, например, ген дигидродипиколинатредуктазы (Japanese Patent Laid-open No. 7-75578) и ген диаминопимелатдегидрогеназы (Ishino, S. et al. , Nucleic Acids Res., 15, 3917 (1987) (в этой работе клонирован ген, участвующий в биосинтезе L-лизина), а также ген фосфоенолпируваткарбоксилазы (Japanese Patent Laid-open No. 60-87788), ген дигидродипиколинатсинтазы (Japanese Patent Laid-open No. 6-55149) и ген диаминопимелатдекарбоксилазы (Japanese Patent Laid-open No. 60-62994, в котором амплификация гена влияла на продуктивность L-лизина).
Что касается ферментов, участвующих в биосинтезе L-лизина, известен случай для фермента, который подвергается ингибированию по типу обратной связи при использовании его в виде дикого типа. В этом случае продуктивность L-лизина улучшается введением гена фермента, имеющего такую мутацию, при которой ингибирование по типу обратной связи десенсибилизируется. К таким известным генам конкретно относится, например, ген аспартокиназы (International Publication Pamphlet of WO 94/25605).
Как описано выше, некоторые успешные результаты были получены путем амплификации генов для системы биосинтеза L-лизина или введением мутантных генов. Например, коринеформная бактерия, которая удерживает мутантный ген аспартокиназы с десенсибилизированным совместным ингибированием лизином и треонином, продуцирует значительное количество L-лизина (приблизительно 25 г/л). Однако эта бактерия страдает от снижения скорости роста по сравнению с бактерией, не удерживающей мутантного гена аспартокиназы. Также сообщалось, что продуктивность L-лизина улучшается дополнительно введением гена дигидродипиколинатсинтазы, кроме мутантного гена аспартокиназы (Applied and Environmental Microbiology. 57(6), 1746-1752 (1991)). Однако такая бактерия страдает от дополнительного снижения скорости роста.
Что касается гена дигидродипиколинатредуктазы, было показано, что активность дигидродипиколинатредуктазы увеличивается в коринеформной бактерии, в которую был введен этот ген, однако не сообщалось влияние на продуктивность L-лизина (Japanese Patent Laid-open No. 7-75578).
В настоящее время неизвестен ни один случай для коринеформных бактерий, когда кому-либо удалось значительное улучшение выхода L-лизина без ограничения роста путем комбинирования множества генов для биосинтеза L-лизина. Не сообщался ни один случай, в котором предусматривалось также улучшение роста путем усиления гена для биосинтеза L-лизина.
Описание изобретения
Целью данного изобретения является улучшение L-лизин-продуцирующей способности и
скорости роста коринеформной бактерии посредством использования генетических материалов последовательностей ДНК, кодирующих (каждая) аспартокиназу (далее называемую "АК", при условии, что ген,
кодирующий белок АК, далее называют "lysC, если необходимо), дигидродипиколинатредуктазу (далее называемую "DDPR", при условии, что ген, кодирующий белок DDPR, далее называют "dapB", если необходимо),
дигидродипиколинатсинтазу (далее называемую "DDPS", при условии, что ген, кодирующий белок DDPS, далее называют "dapA", если необходимо), диаминопимелатдекарбоксилазу (далее называемую "DDC", при
условии, что ген, кодирующий белок DDC, далее называют lysA, если необходимо), и диаминопимелатдегидрогеназу (далее называемую "DDH", при условии, что ген, кодирующий белок DDH, далее называют "ddh,
если необходимо), которые являются важными ферментами биосинтеза L-лизина в клетках коринеформных бактерий.
Когда целевое вещество получают ферментативно при помощи микроорганизма, скорость получения, а также выход целевого вещества относительно вводимого материала является крайне важным фактором. Целевое вещество можно получать очень дешево путем увеличения скорости продуцирования на единицу ферментативного оборудования. Поэтому в промышленном масштабе крайне важно, чтобы ферментативный выход и скорость продуцирования были совместимы друг с другом. Данное изобретение предлагает решение проблемы, описанной выше, для ферментативного получения L-лизина с использованием коринеформной бактерии.
Принцип данного изобретения основан на факте, что рост коринеформной бактерии может быть улучшен и скорость продуцирования L-лизина может быть улучшена посредством усиления при объединении dapB с мутантом lysC (далее называемым просто "мутантным lysC", если необходимо), кодирующим мутантную АК (далее называемую просто "АК мутантного типа", если необходимо), при котором совместное ингибирование лизином и треонином является десенсибилизированным по сравнению со случаем, когда lysC усиливается отдельно, и что скорость продуцирования L-лизина может быть дополнительно улучшена ступенчатым образом посредством усиления dapA, lysA и ddh.
А именно, данное изобретение основано на рекомбинантной ДНК, автономно реплицируемой в клетках коринеформных бактерий, содержащей последовательность ДНК, кодирующую аспартокиназу, для которой ингибирование по типу обратной связи по существу десенсибилизировано, и последовательность ДНК, кодирующую дигидродипиколинатредуктазу. Данное изобретение обеспечивает рекомбинантную ДНК, дополнительно содержащую последовательность ДНК, кодирующую дигидродипиколинатсинтазу, кроме каждой из описанных выше последовательностей ДНК. Данное изобретение обеспечивает рекомбинантную ДНК, дополнительно содержащую последовательность ДНК, кодирующую диаминопимелатдекарбоксилазу, кроме трех описанных выше последовательностей ДНК. Данное изобретение обеспечивает рекомбинантную ДНК, дополнительно содержащую последовательность ДНК, кодирующую диаминопимелатдегидрогеназу, кроме четырех описанных выше последовательностей ДНК.
В другом аспекте данное изобретение обеспечивает коринеформную бактерию, удерживающую аспартокиназу, в которой ингибирование по типу обратной связи L-лизином и L-треонином значительно десенсибилизировано, и содержащую усиленную ДНК, кодирующую дигидродипиколинатредуктазу. Данное изобретение обеспечивает коринеформную бактерию, дополнительно содержащую усиленную ДНК, кодирующую дигидропиколинатсинтазу в вышеупомянутой коринеформной бактерии. Данное изобретение обеспечивает коринеформную бактерию, дополнительно содержащую усиленную ДНК, кодирующую диаминопимелатдекарбоксилазу в вышеупомянутой коринеформной бактерии, кроме трех описанных выше ДНК. Данное изобретение обеспечивает коринеформную бактерию, дополнительно содержащую усиленную ДНК, кодирующую диаминопимелатдегидрогеназу в вышеупомянутой коринеформной бактерии, кроме четырех описанных выше ДНК.
Еще в одном варианте данное изобретение обеспечивает способ получения L-лизина, предусматривающий стадии культивирования любой из описанных выше коринеформных бактерий в подходящей среде, продуцирования и накопления L-лизина в культуре этой бактерии и сбор L-лизина из культуры.
Коринеформные бактерии, упоминаемые в данном изобретении, представляют собой группу микроорганизмов, описанных в Bergey's Manual of Determinative Bacteriology, 8th ed. ,p. 599 (1974), которые являются аэробными грамположительными палочками, не обладающими кислотоустойчивостью и спорообразующей способностью. Эти коринеформные бактерии включают бактерии, принадлежащие к роду Corinebacterium, бактерии, принадлежащие к роду Brevibacterium, классифицируемые до настоящего времени в род Brevibacterium, но объединенные в настоящее время в род Corinebacterium, и бактерии, принадлежащие к роду Brevibacterium, близкородственные бактериям, принадлежащим к роду Corinebacterium.
Данное изобретение будет объяснено подробно ниже.
<1> Получение генов
для биосинтеза L-лизина, используемых в данном изобретении
Гены для биосинтеза L-лизина, используемые в данном изобретении, получают соответственно получением хромосомной ДНК из бактерии в
качестве донора ДНК, конструированием библиотеки хромосомной ДНК с применением плазмидного вектора или т.п., отбора штамма, удерживающего желаемый ген, и извлечения из выбранного штамма рекомбинантной
ДНК, в которую был вставлен этот ген. Донор ДНК для гена для биосинтеза L-лизина, используемый в данном изобретении, не является специфически ограниченным, при условии, что желаемый ген для биосинтеза
L-лизина экспрессирует ферментный белок, который функционирует в клетках коринеформных бактерий. Однако донор ДНК предпочтительно является коринеформной бактерией.
Все гены lysC, dapA и dapB, происходящие из коринеформных бактерий, имеют известные последовательности. Поэтому они могут быть получены путем проведения амплификации в соответствии со способом полимеразной цепной реакции (PCR; см. White, Т. J. et al.. Trends Genet.. 5, 185 (1989)).
Каждый из генов для биосинтеза L-лизина, используемый в данном изобретении, можно получить в соответствии с определенными способами, приведенными в качестве примеров ниже.
(1) Получение мутантного lysC
Фрагмент ДНК, содержащий мутантный lysC, может быть получен из мутантного штамма, в котором
синергическое ингибирование по типу обратной связи активности АК L-лизином и L-треонином значительно десенсибилизировано (International Publication Pamphlet of WO 94/25605). Такой мутантный штамм
может быть получен, например, из группы клеток, происходящих из штамма дикого типа коринеформной бактерии, подвергнутых мутационной обработке путем приложения к ним обычной мутационной обработки,
такой как облучение ультрафиолетовым светом и обработка мутирующим агентом, таким как N-метил-N'-нитро-N-нитрозогуанидин. Активность АК может быть измерена при помощи способа, описанного Miyajima, R.
et al. в The Journal of Biochemistry (1968), 63(2), 139-148. Наиболее предпочтительным мутантным штаммом является продуцирующая L-лизин бактерия AJ3445 (FERM Р-1944), полученная мутационной обработкой
из штамма дикого типа Brevibacterium lactofermentum ATCC 13869 (имеющей измененное в настоящее время название Corinebacterium glutamicum).
Альтернативно, мутантный lysC можно получить также мутационной обработкой in vitro плазмидной ДНК, содержащей lysC дикого типа. В другом аспекте известна конкретная информация о мутации для десенсибилизации синергического ингибирования по типу обратной связи АК L-лизином и L-треонином ((International Publication Pamphlet of WO 94/25605). Поэтому мутантный lysC может быть также получен из lysC дикого типа на основании этой информации, например, по способу сайт-специфического мутагенеза.
Фрагмент, содержащий lysC, может быть выделен из коринеформной бактерии путем получения хромосомной ДНК в соответствии, например, со способом Saito and Miura (H. Saito and К. Miura, Biochim. Biophys. Acta,72,619 (1963) ), и амплификации lysC в соответствии со способом полимеразной цепной реакции (PCR; см. White, T.J. et al., Trends Genet., 5, 185 (1989)).
Примерами праймеров ДНК являются одноцепочечные ДНК 23-мера и 21-мера, имеющие нуклеотидные последовательности, показанные в SEQ ID 1 и SEQ ID 2 в Списке последовательностей, для амплификации, например, района приблизительно 1643 п. н., кодирующего lysC, на основе последовательности, известной для Corinebacterium glutamicum (см. Molecular Microbiology (1991), 5 (5) 1197-1204; Mol. Gen. Genet. (1990), 224, 317-324). ДНК может быть синтезирована в соответствии с обычным способом при помощи синтезатора ДНК модели 380В Applied Biosystems и при помощи фосфоамидитного способа (см. Tetrahedron Letters (1981), 22, 1859). ПЦР может быть выполнена с применением Термоциклера ДНК модели PJ2000, производимого Takara Shuzo, и с применением ДНК-полимеразы Taq согласно способу, предписанному поставщиком.
Предпочтительно, чтобы lysC, амплифицированный при помощи ПЦР, был лигирован с вектором ДНК, автономно реплицируемым в клетках Е.coli и/или коринеформных бактериях, для получения рекомбинантной ДНК, и эту рекомбинантную ДНК вводят в клетки E.coli заранее. Это облегчает последующие операции. Вектор, автономно реплицируемый в клетках Е.coli, предпочтительно является плазмидным вектором, который предпочтительно автономно реплицируется в клетках хозяина, в том числе, например, pUC19, pUC18, pBR322, pHSG299, pHSG399, pHSG398 и RSF 1010.
Когда фрагмент ДНК, способный позволять плазмиде автономно реплицироваться в коринеформной бактерии, вводят в эти векторы, их можно использовать в качестве так называемого челночного вектора, автономно реплицируемого как в Е.coli, так и в коринеформной бактерии.
Такими челночными векторами являются следующие векторы. Микроорганизмы, удерживающие каждый из векторов, и номера депозитов в международных средствах для депонирования указаны в скобках.
рНС4: Escherichia coli AJ12617 (FERM BP-3532)
pAJ655: Escherichia coii AJ11882
(FERM BP-136)
Corinebacterium glutamicum SR8201 (ATCC 39135)
pAJ 1844: Escherichia coli AJ 11883 (FERM BP-137)
Corinebacterium glutamicum SR8202 (ATCC 39136)
pAJ611: Escherichia coli AJ11884 (FERM BP-138)
pAJ3148: Corinebacterium glutamicum SR8203 (FERM 39137)
pAJ440: Bacillus subtilis AJ11901 (FERM BP-140)
Эти векторы могут быть
получены из депонированных микроорганизмов следующим образом. Клетки, собранные при логарифмической фазе роста, лизировали при помощи лизозима и ДСН с последующим отделением от лизата
центрифугированием при 30000 g с получением супернатанта, к которому добавляли полиэтиленгликоль и затем фракционировали и очищали центрифугированием в равновесном градиенте плотности хлорида
цезия/бромида этидия.
Е. coli может быть трансформирована введением плазмиды согласно, например, способу D.M. Morrison (Methods in Enzymology, 68, 326 (1979)) или способу, в котором реципиентные клетки обрабатывают хлоридом кальция для увеличения проницаемости для ДНК (Mandel, M. and Higa, A., J. Mol. Biol., 53, 159 ( 1970)).
LysC дикого типа получают при выделении lysC из содержащего АК штамма дикого типа, тогда как lysC выделяют из содержащего АК мутантного штамма в соответствии со способом, описанным выше.
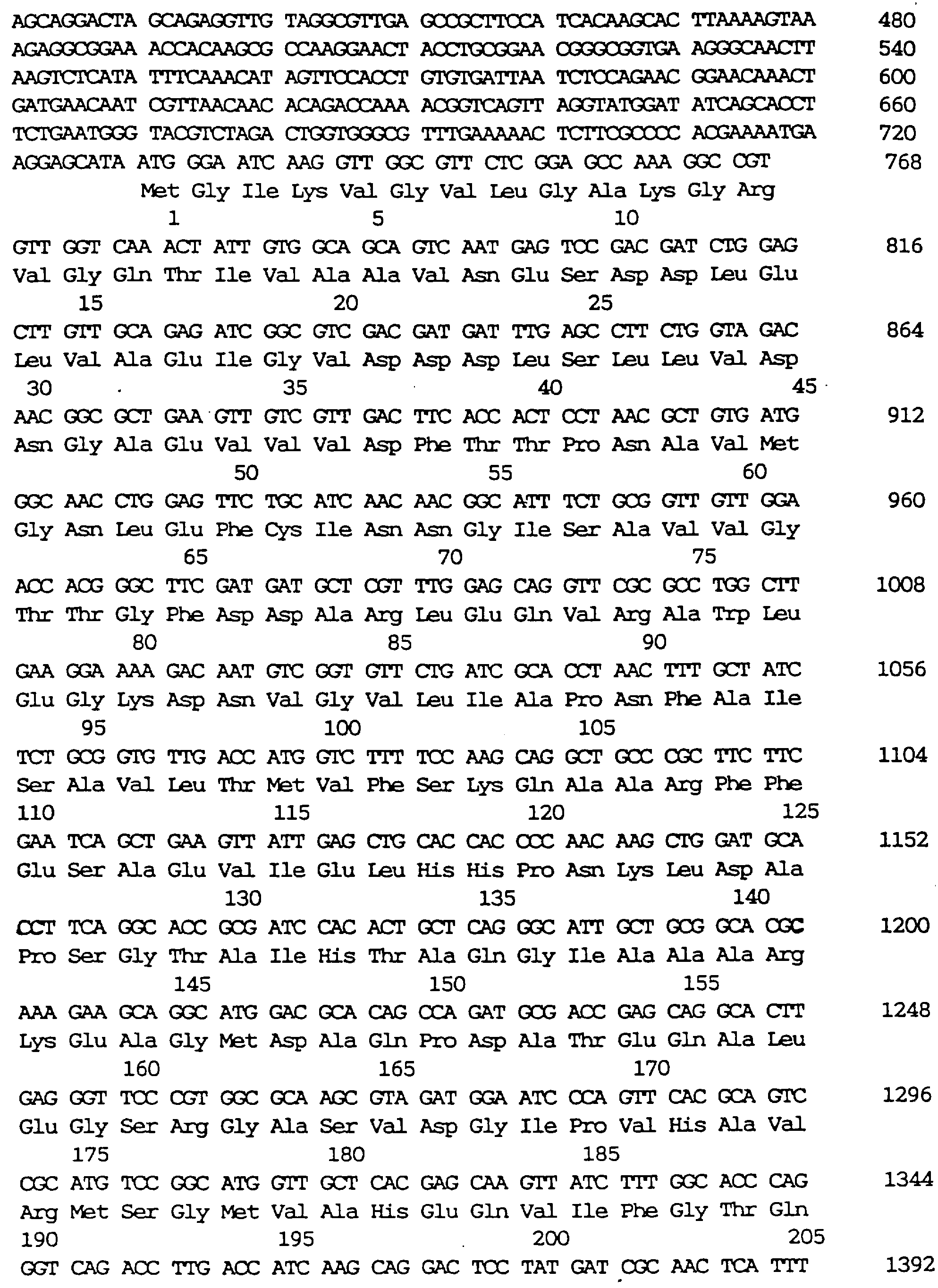
Пример нуклеотидной последовательности фрагмента ДНК, содержащего lysC, показан в SEQ ID 3 в Списке последовательностей. Аминокислотная последовательность α-субъединицы белка АК дикого типа расшифрована из нуклеотидной последовательности, показанной в SEQ ID 4 в Списке последовательностей вместе с последовательностью ДНК. Только аминокислотная последовательность показана в SEQ ID 5. Аминокислотная последовательность β-субъединицы белка АК дикого типа расшифрована из нуклеотидной последовательности ДНК, показанной в SEQ ID 6 в Списке последовательностей вместе с ДНК. Только аминокислотная последовательность показана в SEQ ID 7. В каждой из этих субъединиц GTG использован как инициирующий кодон и соответствующая аминокислота представлена метионином. Однако это представление относится к метионину, валину или формилметионину.
Мутантный lysC, используемый в данном изобретении, особо не ограничивается, при условии, что он кодирует АК, для которой синергическое ингибирование по типу обратной связи L-лизином и L-треонином является десенсибилизированным. Однако мутантный lysC представлен для примера геном, имеющим мутацию, в которой 279-ый остаток аланина при отсчете от N-конца заменен на аминокислотный остаток, отличающийся от аланина и не являющийся кислой аминокислотой, в α-субъединице, и 30-ый остаток аланина заменен на аминокислотный остаток, отличающийся от аланина и не являющийся кислой аминокислотой, в β-субъединице в аминокислотной последовательности АК дикого типа. Аминокислотная последовательность АК дикого типа включает в себя аминокислотную последовательность, показанную в SEQ ID 5 в Списке последовательностей, в качестве α-субъединицы и аминокислотную последовательность, показанную в SEQ ID 7 в Списке последовательностей, в качестве β-субъединицы.
Аминокислотными остатками, иными, чем аланин, и иными, чем кислая аминокислота, предпочтительно являются остатки треонина, аргинина, цистеина, фенилаланина, пролина, серина, тирозина и валина.
Кодон, соответствующий заменяемому аминокислотному остатку, особо не лимитирован относительно его типа, при условии, что он кодирует этот аминокислотный остаток. Предполагается, что аминокислотная последовательность имеющейся АК дикого типа может слегка отличаться в зависимости от различия в бактериальных видах и бактериальных штаммах. АК, которые имеют мутацию, основанную, например, на замене, делеции или инсерции одного или нескольких аминокислотных остатков в одном или нескольких положениях, не имеющих значения для активности фермента, описанных выше, могут также использоваться для данного изобретения. Другие АК, которые имеют мутацию, основанную, например, на замене, делеции или инсерции других одного или нескольких аминокислотных остатков, могут также использоваться при условии, что они не оказывают существенного влияния на активность АК и на десенсибилизацию синергического ингибирования по типу обратной связи L-лизином и L-треонином.
Штамм AJ12691, полученный введением плазмиды р399АК 9В с мутантным lysC в штамм AJ12036 (FERM ВР-734) в качестве штамма дикого типа Brevibacterium lactofermentum, был депонирован 10 апреля 1992 года под номером доступа FERM Р-12918 в National Institute of Bioscience and Human of Agency of Industrial Science and Technology of Ministry of International Trade and Industry (postal code: 305, 1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan), перенесен в международное депонирование на основе Budapest Treaty 10 февраля 1995 года и депонирован под номером доступа FERM ВР-4999.
(2) Получение dapB
Фрагмент ДНК, содержащий dapB, может быть получен из хромосомы коринеформной бактерии
при помощи ПЦР. Донор ДНК специфически не ограничивается, однако в качестве примера приводится штамм Brevibacterium lactofermentum ATCC 13869.
Последовательность ДНК, кодирующая DDPR, известна для Brevibacterium lactofermentum (Journal of Bacteriology, 175 (9), 2743-2749 (1993)), на основе этой последовательности ДНК могут быть получены праймеры ДНК для ПЦР. Примерами таких праймеров ДНК являются ДНК 23-меров, соответственно имеющие нуклеотидные последовательности, изображенные в SEQ ID 8 и SEQ ID 9 в Списке последовательностей. Синтез ДНК, ПЦР и приготовление плазмиды, содержащей полученный dapB, могут быть выполнены так же, как описано выше для lysC.
Нуклеотидная последовательность фрагмента ДНК, содержащего dapB, и аминокислотная последовательность, расшифрованная из этой нуклеотидной последовательности, представлены в SEQ ID 10. Только аминокислотная последовательность показана в SEQ ID 11. Кроме фрагментов ДНК, кодирующих эту аминокислотную последовательность, данное изобретение может равноценно использовать фрагменты ДНК, кодирующие аминокислотные последовательности, в основном такие же, какие показаны в SEQ ID 11, а именно аминокислотные последовательности, имеющие мутацию, основанную, например, на замене, делеции или инсерции одной или нескольких аминокислот, при условии, что при этом нет существенного влияния на активность DDPR.
Трансформированный штамм АJ13107, полученный введением плазмиды pCRDAPB, содержащей dapB, полученный в Примере, описанном ниже, в штамм Е.coli JM109, был депонирован международно с 26 мая 1995 года под депозитным номером FERM ВР-5114 в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology of Ministry of International Trade and Industry (postal code: 305, 1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan) на основе Budapest Treaty.
(3) Получение dapA
Фрагмент ДНК, содержащий dapA, может
быть получен из хромосомы коринеформной бактерии при помощи ПЦР. Донор ДНК особо не ограничивается, однако в качестве примера приведен штамм Brevibacterium lactofermentum ATCC 13869.
Последовательность ДНК, кодирующая DDPS, известна для Corinebacterium glutamicum (см. Nucleic Acids Research. 18 (21), 6421 (1990), EMBL accession No. X53993), на основе этой последовательности могут быть приготовлены праймеры ДНК для ПЦР. Специфическими примерами таких праймеров ДНК являются ДНК 23-меров, соответственно имеющие нуклеотидные последовательности, изображенные в SEQ ID 12 и SEQ ID 13 Списка последовательностей. Синтез ДНК, ПЦР и приготовление плазмиды, содержащей dapA, могут быть выполнены так же, как описано для lysC выше.
Нуклеотидная последовательность фрагмента ДНК, содержащего dapA, и аминокислотная последовательность, расшифрованная из этой нуклеотидной последовательности, приведены в виде примеров в SEQ ID 14. Только аминокислотная последовательность показана в SEQ ID 15. Кроме фрагментов ДНК, кодирующих эту аминокислотную последовательность, данное изобретение может равноценно использовать фрагменты ДНК, кодирующие аминокислотные последовательности, в основном такие же, что и аминокислотная последовательность, показанная в SEQ ID 15, а именно аминокислотные последовательности, имеющие мутацию, основанную, например, на замене, делении или инсерции одной или нескольких аминокислот, при условии, что при этом нет существенного влияния на активность DDPS.
Трансформированный штамм АJ13106, полученный введением плазмиды pCRDAPA, содержащей dapA, полученный в Примере, описанном ниже, в штамм Е. coli JM 109, был депонирован с 26 мая 1995 года под депозитным номером FERM BP-5113 в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology of Ministry of International Trade and Industry (postal code: 305, 1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan) на основе Budapest Treaty.
(4) Получение lysA
Фрагмент ДНК, содержащий lysA, может быть получен из хромосомы коринеформной бактерии при помощи ПЦР. Донор ДНК особо не
ограничивается, однако в качестве примера приведен штамм Brevibacterium lactofermentum ATCC 13869.
В коринеформных бактериях lysA образует оперон вместе с argS (геном аргинил-тРНК-синтазы) и lysA находится по ходу транскрипции от argS. Экспрессия lysA регулируется промотором, расположенным против хода транскрипции от argS (см. Journal of Bacteriology, Nov., 7356-7362 (1993)). Последовательности ДНК этих генов известны для Corinebacterium glutamicum (см. Molecular Microbiology, 4(11), 1819-1830 (1990); Molecular and General Genetics, 212, 112-119 (1988)), на основе этих последовательностей могут быть приготовлены праймеры ДНК для ПЦР. Такие праймеры ДНК приведены в качестве специфических примеров в виде 23-меров ДНК, соответственно имеющих нуклеотидные последовательности, показанные в SEQ ID 16 в Списке последовательностей (соответствующей номерам нуклеотидов 11-33 в нуклеотидной последовательности, описанной в Molecular Microbiology, 4(11), 1819-1830 (1990)), и SEQ ID 17 (соответствующей номерам нуклеотидов 1370-1392 в нуклеотидной последовательности, описанной в Molecular and General Genetics, 212, 112-119 (1988)). Синтез ДНК, ПЦР и приготовление плазмиды, содержащей полученный lysA, могут быть выполнены так же, как описано выше для lysC.
В Примере, описанном позже, фрагмент ДНК, содержащий промотор, argS и lysA, использовали для усиления lysA. Однако argS не является существенным для данного изобретения. Можно использовать фрагмент ДНК, в котором lysA лигирован непосредственно по ходу транскрипции от промотора.
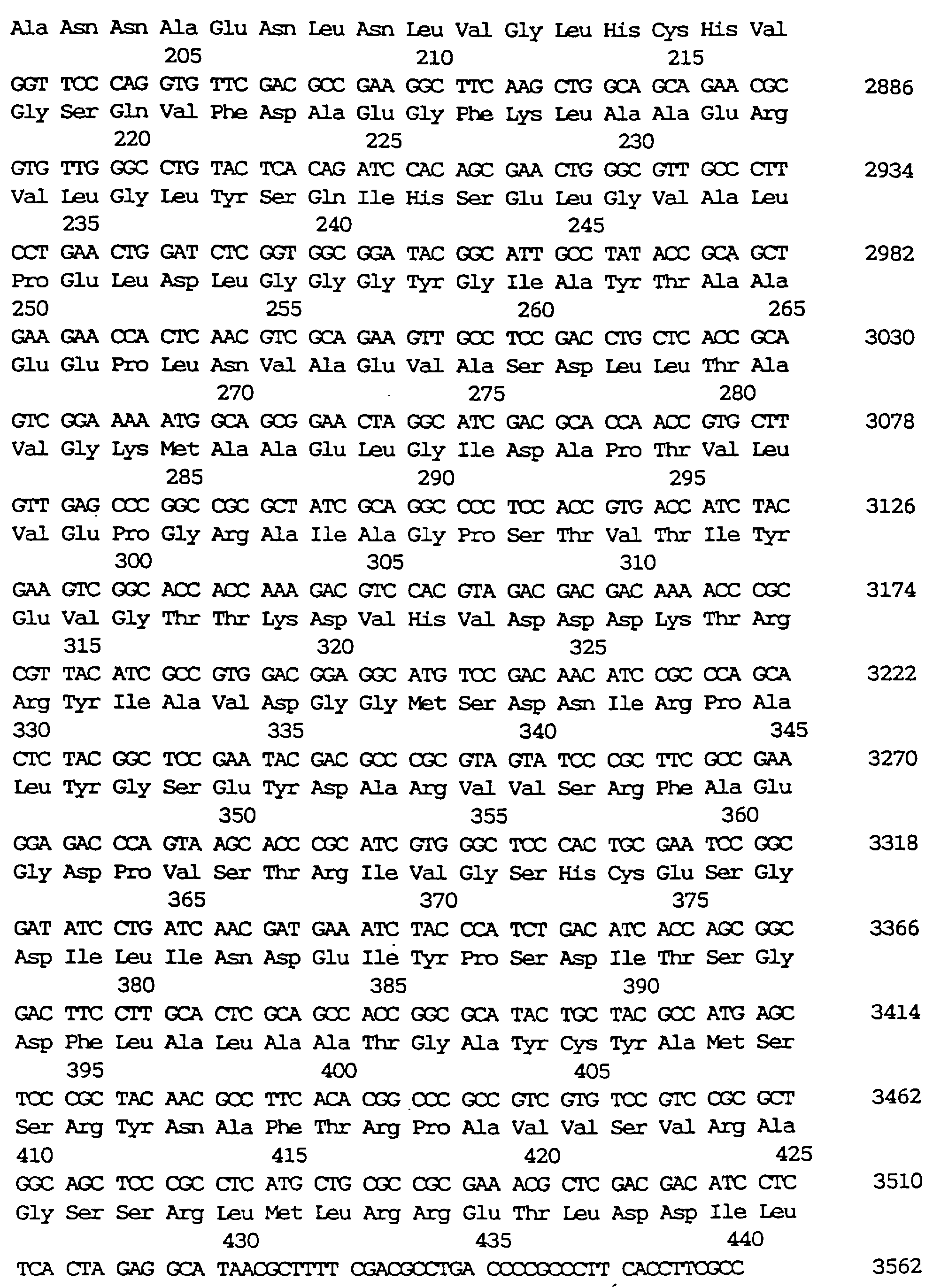
Нуклеотидная последовательность фрагмента ДНК, содержащего argS и lysA, и аминокислотная последовательность, расшифрованная из этой нуклеотидной последовательности, приведены в виде примеров в SEQ ID 18. Пример аминокислотной последовательности, кодируемой argS, показан в SEQ ID 19, и пример аминокислотной последовательности, кодируемой lysA, показан в SEQ ID 20. Кроме фрагментов ДНК, кодирующих эту аминокислотную последовательность, данное изобретение может равноценно использовать фрагменты ДНК, кодирующие аминокислотные последовательности, в основном такие же, что и аминокислотная последовательность, показанная в SEQ ID 20, а именно аминокислотные последовательности, имеющие мутацию, основанную, например, на замене, делении или инсерции одной или нескольких аминокислот, при условии, что при этом нет существенного влияния на активность DDC.
(5) Получение ddh
Фрагмент ДНК, содержащий ddh, может быть получен из хромосомы
коринеформной бактерии при помощи ПЦР. Донор ДНК особо не ограничивается, однако в качестве примера приведен штамм Brevibacterium lactofermentum ATCC 13869.
Ген DDH известен для Corinebacterium glutamicum (Ishino, S. et al., Nucleic Acids Res., 15, 3917 (1987)), на основе его последовательности могут быть приготовлены праймеры для ПЦР. Специфические примеры таких праймеров приведены в виде ДНК 20-меров, соответственно имеющих нуклеотидные последовательности, изображенные в SEQ ID 21 и SEQ ID 22 в Списке последовательностей. Синтез ДНК, ПЦР и приготовление плазмиды, содержащей полученный ddh, могут быть выполнены так же, как описано выше для lysC.
Нуклеотидная последовательность фрагмента ДНК, содержащего ddh, и аминокислотная последовательность, расшифрованная из этой нуклеотидной последовательности, приведены в виде примеров в SEQ ID 23. Только аминокислотная последовательность показана в SEQ ID 24. Кроме фрагментов ДНК, кодирующих эту аминокислотную последовательность, данное изобретение может равноценно использовать фрагменты ДНК, кодирующие аминокислотные последовательности, в основном такие же, что и аминокислотная последовательность, показанная в SEQ ID 24, а именно аминокислотные последовательности, имеющие мутацию, основанную, например, на замене, делении или инсерции одной или нескольких аминокислот, при условии, что при этом нет существенного влияния на активность DDH.
<2> Рекомбинантная ДНК и коринеформная бактерия данного изобретения
Коринеформная бактерия
данного изобретения удерживает аспартокиназу (мутантную АК), у которой ингибирование по типу обратной связи L-лизином и L-треонином по существу десенсибилизировано, причем ДНК (dapB), кодирующая
дигидродипиколинатредуктазу, усилена. В предпочтительном варианте коринеформная бактерия данного изобретения представляет собой коринеформную бактерию, в которой ДНК (dapA), кодирующая
дигидродипиколинтасинтазу, дополнительно усилена. В более предпочтительном варианте коринеформная бактерия данного изобретения представляет собой коринеформную бактерию, в которой ДНК (lysA),
кодирующая диаминопимелатдекарбоксилазу, дополнительно усилена. В более предпочтительном варианте коринеформная бактерия данного изобретения представляет собой коринеформную бактерию, в которой ДНК
(ddh), кодирующая диаминопимелатдегидрогеназу, дополнительно усилена.
Термин "усиленная" ДНК обозначает здесь тот факт, что внутриклеточная активность фермента, кодируемого этой ДНК, повышается, например, путем увеличения числа копий гена, использованием сильного промотора, использованием гена, кодирующего фермент, имеющий высокую удельную активность, или комбинированием этих средств.
Коринеформной бактерией, удерживающей мутантную АК, могут быть коринеформные бактерии, которые продуцируют мутантную аспартокиназу как результат мутации, или коринеформные бактерии, которые трансформированы введением мутантного гена lysC.
Примеры коринеформной бактерии, используемой для введения описанной выше ДНК, включают, например, следующие
продуцирующие лизин штаммы дикого типа:
Corinebacterium acetoacidophilum ANCC 13870;
Corinebacterium acetoglutamicum ATCC 15806;
Corinebacterium callunae ATCC 15991;
Corinebacterium glutamicum ATCC 13032;
(Brevibacterium divaricatum) ATCC 14020;
(Brevibacterium lactofermentum) ATCC 13869;
(Corinebacterium lilium) ATCC 15990;
(Brevibacterium flavum) ATCC 14067;
Corinebacterium melassecola ATCC 17965;
Brevibacterium saccharolyticum ATCC 14066;
Brevibacterium immariophilum ATCC 14068;
Brevibacterium roseum ATCC 13825;
Brevibacterium thiogenitalis ATCC 19240;
Microbacterium ammoniaphilum ATCC 15354;
Corinebacterium thermoaminogenes AJ12340 (FERM
BP-1539).
Другими бактериальными штаммами, кроме описанных здесь, используемыми в качестве хозяина, являются, например, мутантные штаммы, способные продуцировать L-лизин, произведенные из вышеуказанных штаммов. Такие искусственные мутантные штаммы включают: S-(2-аминоэтил)-цистеин (далее сокращаемый как "АЕС")-резистентные мутантные штаммы (Brevibacterium lactofermentum AJ11082 (NRRL B-1147), Japanese Patent Publication Nos. 56-1914, 56-1915, 57-14157, 57-14158, 57-30474, 58-10075, 59-4993, 62-35840, 62-24074, 62-36673, 5-11958, 7-112437 и 7-112438); мутантные штаммы, которые требуют аминокислоты, такой как L-гомосерин, для их роста (Japanese Patent Publication Nos. 48-28078 и 56-6499), мутантные штаммы, которые проявляют устойчивость к АЕС и требуют аминокислот, таких как L-лейцин, L-гомосерин, L-пролин, L-серин, L-аргинин, L-аланин и L-валин (United States Patent Nos. 3708395 и 3825472); L-лизин-продуцирующие мутантные штаммы, которые проявляют устойчивость к DL-α-ε-капролактаму, α-аминолауриллактаму, аналогу аспартата, сульфа-лекарстенным средствам, хиноиду и N-лауроиллейцину; L-лизин-продуцирующие мутантные штаммы, которые проявляют устойчивость к ингибиторам оксалацетатдекарбоксилазы или ферментов дыхательной системы (Japanese Patent Laid-open Nos. 50-53588, 50-31093, 52-102498, 53-9394, 53-86089, 55-9783, 55-9759, 56-32995 и 56-39778, и Japanese Patent Publication Nos. 53-43591 и 53-1833); L-лизин-продуцирующие мутантные штаммы, которые требуют инозита или уксусной кислоты (Japanese Patent Laid-open Nos. 55-9784 и 56-8692); L-лизин-продуцирующие мутантные штаммы, которые проявляют чувствительность к фторпировиноградной кислоте или температуре не менее 34oС (Japanese Patent Laid-open Nos. 55-9783 и 53-86090); и L-лизин-продуцирующие мутантные штаммы, принадлежащие к роду Brevibacterium или Corinebacterium, которые проявляют устойчивость к этиленгликолю и продуцируют L-лизин (United States Patent No. 4411997).
В характерном варианте для усиления генов для биосинтеза L-лизина в хозяине, описанном выше, гены вводят в хозяина с использованием плазмидного вектора, транспозона или фагового вектора или т.п. При введении ожидается получение усиления до некоторой степени даже при использовании вектора с низкой копийностью. Однако предпочтительно использовать тип вектора с высокой копийностью. Такой вектор включает, например, плазмидные векторы pAJ655, pAJ1844, pAJ611, pAJ3148 и pAJ440, описанные выше. Кроме того, транспозоны, произведенные из коринеформных бактерий, описаны в International Publication Pamphlets of WO02/02627 и WO93/18151, European Patent Publication No. 445385, Japanese Patent Laid-open No. 6-46867, Vertes, A.A. et al., Mol. Microbiol., 11, 739-746 (1994), Bonamy, С., et al., Mol. Microbiol. , 14, 571-581 (1994), Vertes, A. A. et al., Mol. Gen. Genet,, 245, 397-405 (1994), Jagar, W. et al. , FEMS Microbiology Letters, 126, 1-6 (1995), Japanese Patent Laid-open, No. 7-107976, Japanese Patent Laid-open No. 7-327680 и т. п.
В данном изобретении не является обязательным, чтобы мутантный lysC был обязательно усиленным. Можно использовать штаммы, которые имеют мутацию на lysC на хромосомной ДНК, или в которых мутантный lysC включен в хромосомную ДНК. Альтернативно, мутантный lysC может быть введен при помощи плазмидного вектора. С другой стороны, dapA, dapB, lysA и ddh предпочтительно усиливаются для эффективного продуцирования L-лизина.
Каждый из генов lysC, dapA, dapB, lysA и ddh может быть успешно введен в хозяина при помощи соответственно различных векторов. Альтернативно, два, три, четыре или пять из видов этих генов могут вводиться вместе при помощи одного вектора. При использовании различных векторов гены могут вводиться в любом порядке, однако предпочтительно использовать векторы, которые имеют стабильный механизм обмена и улавливания в хозяине и которые способны сосуществовать с каждым другим вектором.
Коринеформную бактерию, удерживающую мутантную АК и дополнительно содержащую усиленный dapB, получают, например, введением в коринеформную бактерию-хозяина рекомбинантной ДНК, содержащей мутантный lysC и dapB, автономно реплицируемые в клетках коринеформных бактерий.
Коринеформную бактерию, дополнительно содержащую dapA, кроме мутантного lysC и dapB, получают, например, введением в коринеформную бактерию-хозяина рекомбинантной ДНК, содержащей мутантный lysC, dapB и dapA, автономно реплицируемые в клетках коринеформных бактерий.
Коринеформную бактерию, дополнительно содержащую мутантный lysC, dapB и dapA, получают, например, введением в коринеформную бактерию-хозяина рекомбинантной ДНК, содержащей lysC, dapB, dapA и lysA, автономно реплицируемые в клетках коринеформных бактерий.
Коринеформную бактерию, дополнительно содержащую усиленный ddh, кроме мутантного lysC, dapB, dapA и lysA, получают, например, введением в коринеформную бактерию-хозяина рекомбинантной ДНК, содержащей lysC, dapB, dapA, lysA и ddh, автономно реплицируемые в клетках коринеформных бактерий.
Вышеупомянутые рекомбинантные ДНК могут быть получены, например, встраиванием каждого из генов, участвующих в биосинтезе L-лизина, в вектор, такой как плазмидный вектор, транспозон или фаговый вектор, как описано выше.
В случае использования в качестве вектора плазмиды рекомбинантную ДНК можно вводить в хозяина согласно электроимпульсному способу ( Sugimoto et al. , Japanese Patent Laidopen No. 2-207791). Амплификацию гена с использованием транспозона можно выполнять введением плазмиды, несущей транспозон, в клетку-хозяина и индуцированием транспозиции транспозона.
<3> Способ получения L-лизина
L-лизин может продуцироваться эффективно путем культивирования в подходящей среде коринеформной бактерии, содержащей
усиленные гены для биосинтеза L-лизина, как описано выше, продуцирования и накопления L-лизина в культуре этой бактерии и сбора L-лизина из культуры.
Примером используемой среды является обычная среда, содержащая источник углерода, источник азот, неорганические ионы и необязательно другие органические компоненты.
В качестве источника углерода можно использовать сахара, такие как глюкоза, фруктоза, сахароза, мелассы и гидролизат крахмала; и органические кислоты, такие как фумаровая кислота, лимонная кислота и янтарная кислота.
В качестве источника азота можно использовать неорганические соли аммония, такие как сульфат аммония, хлорид аммония и фосфат аммония; органический азот, такой как гидролизат сои; газообразный аммиак; и водный аммиак.
В качестве питательных веществ, предоставляемых в следовых количествах, желательно использовать в соответствующих количествах требующиеся вещества, такие как витамин B1 и L-гомосерин или дрожжевой экстракт или т.п. Кроме того, добавляют, если требуется, фосфат калия, сульфат магния, ион железа, ион марганца и т.д.
Культивирование предпочтительно проводят при аэробных условиях в течение приблизительно 30-90 часов. Температура культивирования предпочтительно регулируется при 25-37oС, и рН предпочтительно регулируется от 5 до 8 во время культивирования. Для доведения рН можно использовать неорганические или органические, кислые или щелочные вещества или газообразный аммиак или т.п. L-лизин может быть собран из культуры комбинированием обычного способа с ионообменной смолой и других известных способов.
Краткое описание чертежей.
Фиг. 1 иллюстрирует процесс конструирования плазмид p399AKYB и р399АК9В, содержащих мутантный lysC.
Фиг. 2 иллюстрирует процесс конструирования плазмиды pDPRB, содержащей dapB и Brevi.-ori. (точку начала репликации, ориджин).
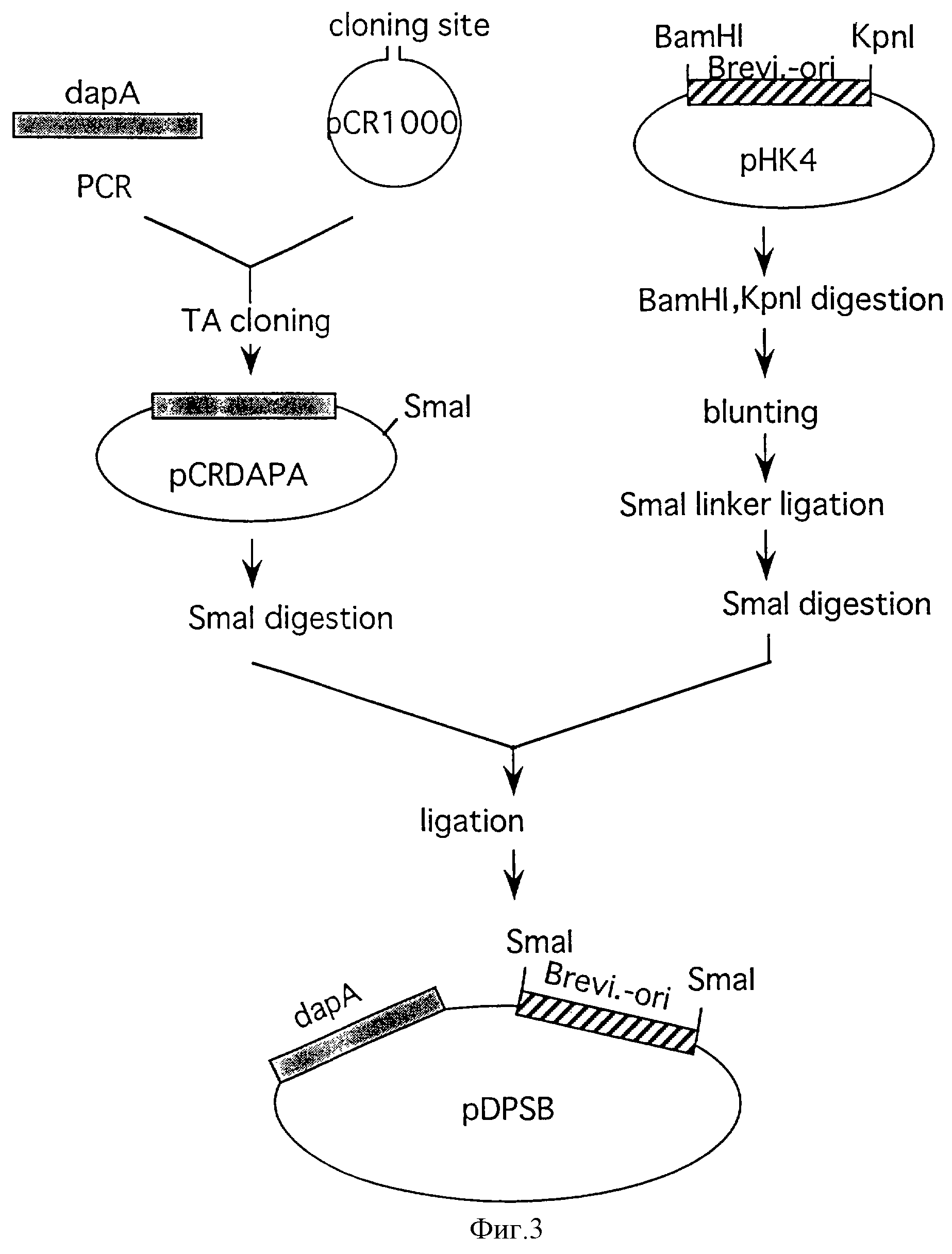
Фиг. 3 иллюстрирует процесс конструирования плазмиды pDPSB, содержащей dapA и Brevi.-ori.
Фиг.4 иллюстрирует процесс конструирования плазмиды p299LYSA, содержащей lysA.
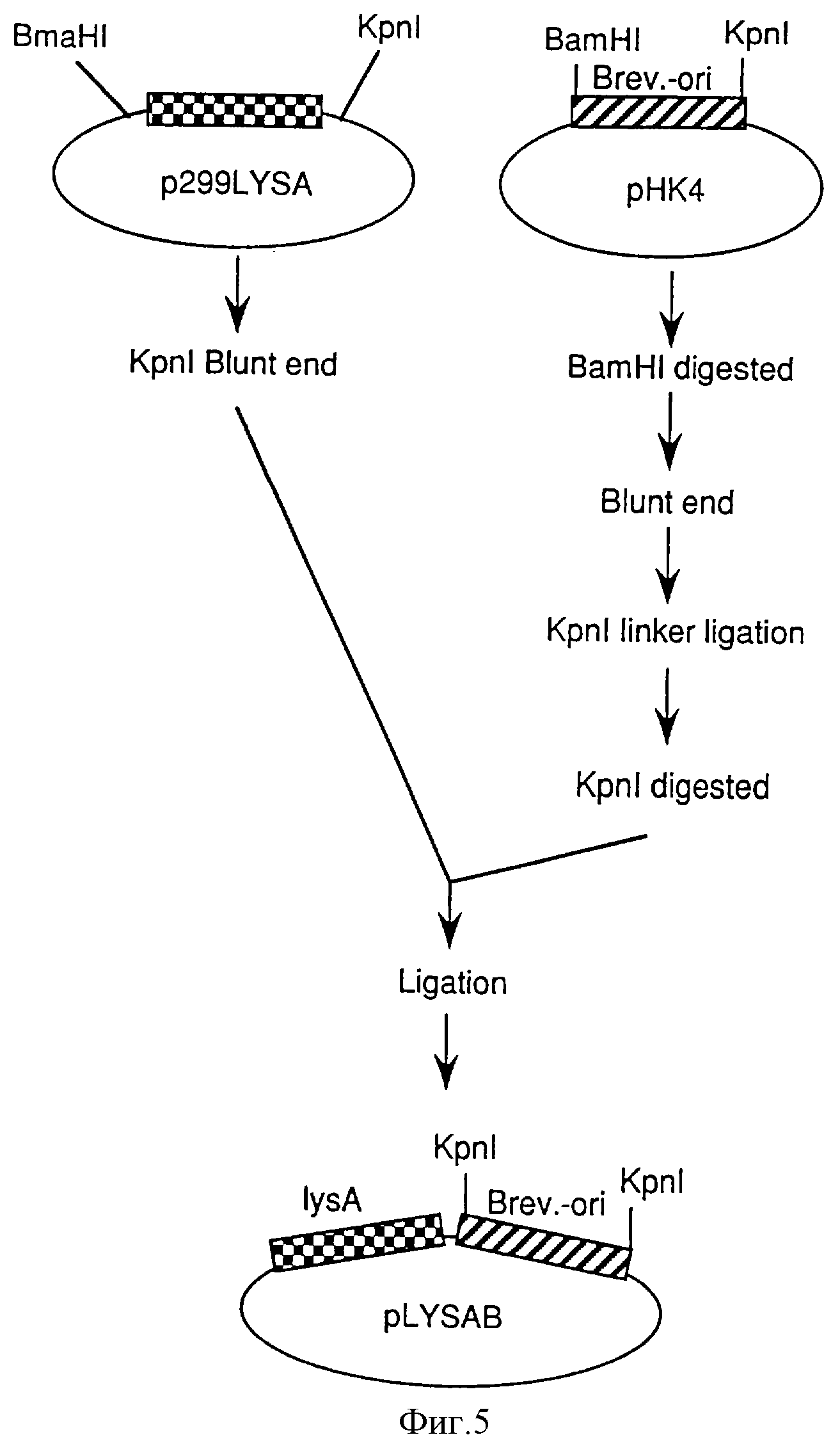
Фиг. 5 иллюстрирует процесс конструирования плазмиды pLYSAB, содержащей lysA и Brevi.-ori.
Фиг. 6 иллюстрирует процесс конструирования плазмиды рРК 4D, содержащей ddh и Brevi.-ori.
Фиг. 7 иллюстрирует процесс конструирования плазмиды pCRCAB, содержащей lysC, dapB и Brevi.-ori.
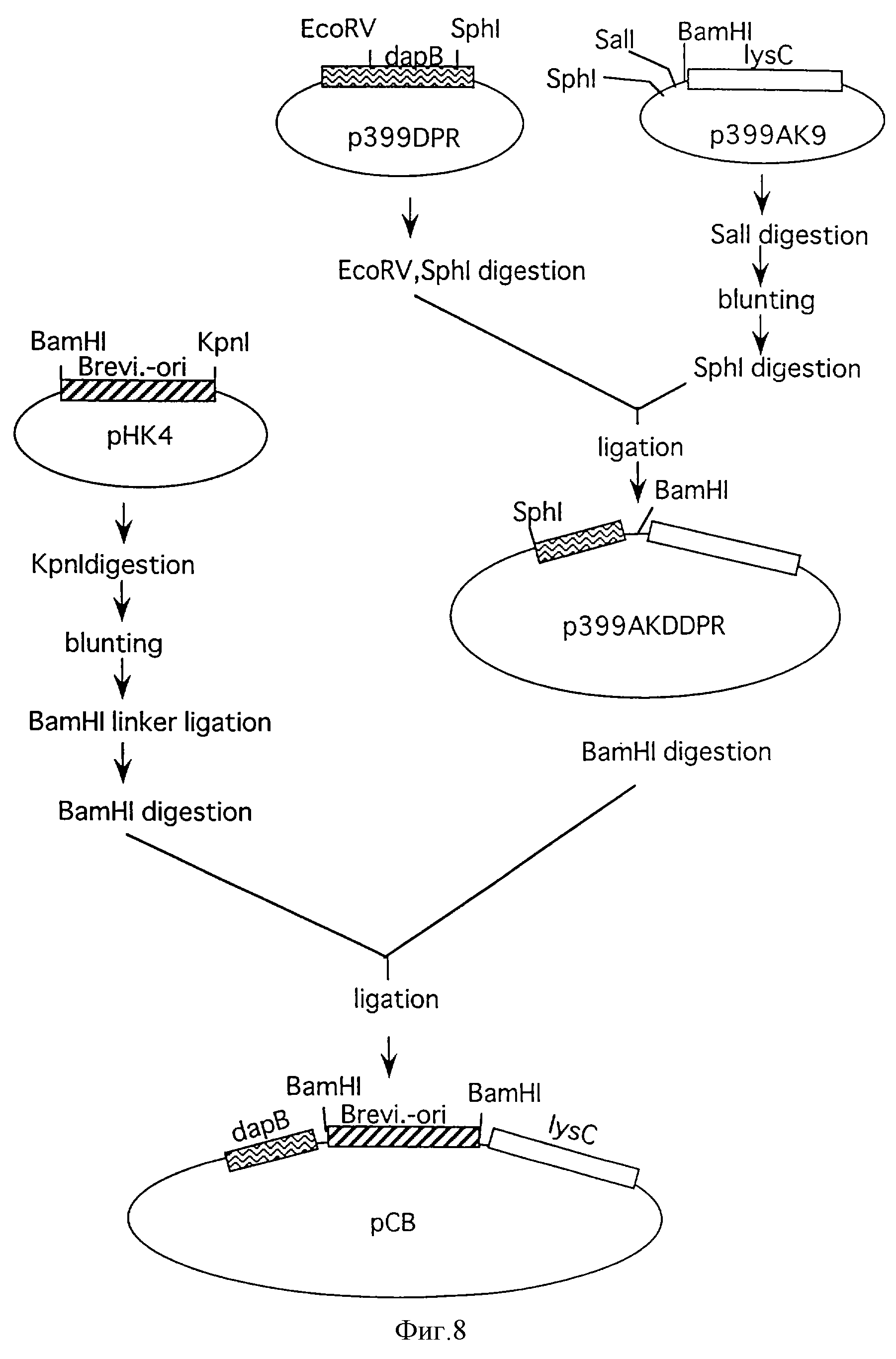
Фиг. 8 иллюстрирует процесс конструирования плазмиды рСВ, содержащей мутантный lysC, dapB и Brevi.-ori.
Фиг. 9 иллюстрирует процесс конструирования плазмиды рАВ, содержащей dapA, dapB и Brevi.-ori.
Фиг. 10 иллюстрирует процесс конструирования плазмиды p399DL, содержащей ddh и lysA.
Фиг. 11 иллюстрирует процесс конструирования плазмиды pDL, содержащей ddh, lysA и Brevi.-ori.
Фиг. 12 иллюстрирует процесс конструирования плазмиды рСАВ, содержащей мутантный lysC, dapA, dapB и Brevi.-ori.
Фиг. 13 иллюстрирует процесс конструирования плазмиды pCABL, содержащей мутантный lysC, dapA, dapB, lysA и Brevi.-ori.
Фиг. 14 иллюстрирует процесс конструирования плазмиды pCABDL, содержащей мутантный lysC, dapA, dapB, ddh, lysA и Brevi.-ori.
Описание предпочтительных вариантов
Данное изобретение будет более конкретно объяснено ниже со ссылкой на Примеры.
Пример 1: Получение гена lysC дикого типа и
мутантного гена lysC из Brevibacterium lactofermentum
<1> Получение lysC дикого типа и мутантного lysC и получение содержащих их плазмид
В качестве доноров хромосомной
ДНК использовали штамм Brevibacterium lactofermentum ATCC 13869 и L-лизин-продуцирующий мутантный штамм AJ34445 (FERM Р-1944), полученный из штамма ATCC 13869 мутационной обработкой. Штамм AJ3445
подвергали мутации таким образом, что lysC был изменен с появлением существенной десенсибилизации от совместного ингибирования лизином и треонином (Journal of Biochemistry, 68, 701-710 (1970)).
Фрагмент ДНК, содержащий lysC, амплифицировали из хромосомной ДНК в соответствии со способом ПЦР (полимеразная цепная реакция; см. White, T.J. et al. Trends Genet., 5, 185 (1989)). Что касается праймеров ДНК, используемых для амплификации, одноцепочечные ДНК 23-мера и 21-мера, имеющие нуклеотидные последовательности, показанные в SEQ ID 1 и SEQ ID 2, синтезировали для амплификации района приблизительно 1643 п. н. для lysC на основе последовательности, известной для Corinebacterium glutamicum (см. Molecular Microbiology (1991), 5(5), 1197-1204; и Mol. Gen. Genet. (1990), 224, 317-324). ДНК синтезировали согласно обычному способу с применением синтезатора ДНК модели 380В, производимого Applied Biosystems, и с применением фосфоамидитного способа (см. Tetrahegron Letters (1981), 22, 1859).
Ген амплифицировали при помощи ПЦР с использованием Термоциклера ДНК модели PJ2000, производимого Takara Shuzo, и с использованием ДНК-полимеразы Taq согласно способу, рекомендуемому поставщиком. Амплифицированный фрагмент гена 1643 т. п. н. подтверждали электрофорезом в агарозном геле. После этого фрагмент, вырезанный из геля, очищали согласно обычному способу и расщепляли его рестриктазами Nrul (производимой Takara Shuzo) и EcoRI (производимой Takara Shuzo).
pHSG399 (см. Takeshita, S. et al., Gene, (1987), 62, 63-74) использовали в качестве клонирующего вектора для фрагмента гена. pHSG399 расщепляли рестриктазами Smal (производимой Takara Shuzo) и EcoRI (производимой Takara Shuzo) и лигировали с амплифицированным фрагментом lysC. ДНК лигировали при помощи набора для лигирования ДНК (производимого Takara Shuzo) в соответствии с рекомендуемым способом. Так были получены плазмиды, в которых фрагменты lysC, амплифицированные из хромосом Brevibacterium lactofermentum, были лигированы с pHSG399 соответственно. Плазмида, содержащая lysC из АТСС 13869 (штамма дикого типа), была обозначена как p399AKY, а плазмида, содержащая lysC из AJ3463 (L-лизин-продуцирующей бактерии) была обозначена как р399АК9.
Фрагмент ДНК (далее называемый "Brevi.-ori."), обладающий способностью делать плазмиду автономно реплицируемой в бактериях, принадлежащих к роду Corinebacterium, вводили в p399AKY и р399АК9 соответственно для получения плазмид, несущих lysC, автономно реплицируемый в бактериях, принадлежащих к роду Corinebacterium. Brevi.-ori. получали из плазмидного вектора рНК4, содержащего Brevi. -ori. и автономно реплицируемого в клетках как Escherichia coli, так и в бактериях, принадлежащих к роду Corinebacterium. рНК4 конструировали расщеплением рНС4 при помощи Kpnl (производимой Takara Shuzo) и Bam HI (производимой Takara Shuzo), экстракцией фрагмента Brevi.-ori. и лигированием его с pHSG298, также расщепленной Kpnl и BamHI (см. Japanese Patent Laid-open No. 5-7491). рНК4 придает хозяину устойчивость к канамицину. Escherichia coli, удерживающую рНК4, обозначили как Escherichia coli AJ13136 и депонировали как Escherichia coli AJ13136 1 августа 1995 года под депозитным номером FERM ВР-5186 в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology of Ministry of International Trade and Industry (postal code: 305, 1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan).
pHK4 расщепляли рестриктазами Kpnl и BamHI и расщепленные края затупляли. Образование тупых концов выполняли с применением набора для затупления ДНК (производимого Takara Shuzo) в соответствии с предложенным способом. После образования тупых концов фосфорилированный линкер BamHI (производимый Takara Shuzo) лигировали для получения модификации так, чтобы фрагмент ДНК, соответствующий Brevi.-ori.-части, мог быть вырезан из pHK4 расщеплением только при помощи BamHI. Эту плазмиду расщепляли BamHI и образующийся фрагмент ДНК Brevi.-ori. лигировали с p399AKY и р399АК9, также уже расщепленными BamHI соответственно для получения плазмид, каждая из которых содержит ген lysC, автономно реплицируемый в бактериях, принадлежащих к роду Corinebacterium.
Плазмида, содержащая ген lysC дикого типа, происходящий из p399AKY, была
обозначена как p399AKYB, а плазмида, содержащая мутантный ген lysC, происходящий из р399АК9, была обозначена как р399АК9В. Процесс конструирования р399АК9В и p399AKYB показан в фиг. 1. Штамм AJ12691,
полученный введением плазмиды р399АК9В с мутантным lysC в штамм дикого типа
Brevibacterium lactofermentum (штамм AJ12036, FERM ВР-734), был депонирован 10 апреля 1992 года под депозитным
номером FERM Р-12918 в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology of Ministry of International Trade and Industry (postal code: 305, 1-3,
Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan), перенесен в международное депонирование на основе Budapest Treaty 10 февраля 1995 года под депозитным номером FERM ВР-4999.
<
2> Определение нуклеотидных последовательностей lysC дикого типа и мутантного lysC из Brevibacterium lactofermentum
Плазмиду p399AKY, содержащую lysC дикого типа, и плазмиду р399АК9,
содержащую мутантный lysC, получали из соответствующих трансформантов для определения нуклеотидных последовательностей lysC дикого типа и мутантного lysC. Определение нуклеотидных последовательностей
проводили по способу Sanger et al. (например, F. Sanger et al., Proc. Natl. Acad. Sci., 74, 5463 (1977)).
Нуклеотидная последовательность lysC дикого типа, кодируемого p399AKY, показана в SEQ ID 3 в Списке последовательностей. С другой стороны, нуклеотидная последовательность мутантного lysC, кодируемого р399АК9, имела только мутацию одного нуклеотида, т. е. 1051-ый G был заменен на А в SEQ ID 3 по сравнению с lysC дикого типа. Известно, что lysC Corinebacterium glutamicum имеет две субъединицы (α,β), кодируемые в идентичной рамке считывания на идентичной цепи ДНК (см. Kalinowski, J. et al., Molecular Microbiology (1991), 5(5), 1197-1204). На основе оценки гомологии предполагается, что секвенированный здесь ген также имеет две субъединицы (α, β), кодируемые в идентичной рамке считывания на идентичной цепи ДНК.
Аминокислотная последовательность α-субъединицы белка АК дикого типа, расшифрованная из нуклеотидной последовательности ДНК, показана в SEQ ID 4 вместе с последовательностью ДНК. Только аминокислотная последовательность показана в SEQ ID 5.
Аминокислотная последовательность β -субъединицы белка АК дикого типа, расшифрованная из нуклеотидной последовательности ДНК, показана в SEQ ID 6 вместе с последовательностью ДНК. Только аминокислотная последовательность показана в SEQ ID 7. В каждой из субъединиц GTG используют в качестве инициирующего кодона, и соответствующая аминокислота представлена метионином. Однако это представление относится к метионину, валину или формилметионину.
С другой стороны, мутация на последовательности мутантного lysC означает появление замены аминокислотного остатка, так что 279-ый остаток аланина α-субъединицы заменен на остаток треонина, а 30-ый остаток аланина β-субъединицы заменен на остаток треонина в аминокислотной последовательности белка АК дикого типа (SEQ ID 5, SEQ ID 7).
Пример 2: Получение dapB из Brevibacterium lactofermentum
<1> Получение dapB и конструирование плазмиды, содержащей dapB
В качестве донора хромосомной ДНК использовали
штамм дикого типа Brevibacterium lactofermentum ATCC 13869. Хромосомную ДНК получали из штамма ATCC 13869 согласно обычному способу. Фрагмент ДНК, содержащий dapB, амплифицировали из хромосомной ДНК
при помощи ПЦР. В качестве праймеров ДНК, используемых для амплификации, синтезировали ДНК 23-меров, имеющих нуклеотидные последовательности, изображенные в SEQ ID 8 и SEQ ID 9 в Списке
последовательностей соответственно для амплификации района приблизительно 2,0 т. п. н., кодирующего DDPR, на основе последовательности, известной для Brevibacterium lactofermentum (см. Journal of
Bacteriology, 157(9), 2743-2749 (1993)). Синтез ДНК, ПЦР проводили, как описано в Примере 1. pCR-Script (производимый Invitrogen) использовали в качестве клонирующего вектора для амплифицированного
фрагмента гена 2001 п. н., который лигировали с амплифицированным фрагментом dapB. Таким образом конструировали плазмиду, в которой фрагмент dapB 2001 п. н. , амплифицированный из хромосомы
Brevibacterium lactofermentum, лигировали с pCR-Script. Плазмида, полученная, как описано выше, которая имела dapB, происходящий из ATCC 13869, была обозначена как pCRDAPB. Трансформантный штамм
AJ13107, полученный введением pCRDAPB в штамм E.coli JM109, был международно депонирован с 26 мая 1995 года под депозитным номером FERM ВР-5114 в National Institute of Bioscience and Human Technology
of Agency of Industrial Science and Technology of Ministry of International Trade and Industry (postal code: 305, 1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan) на основе Budapest Treaty.
Фрагмент 1101 п. н., содержащий структурный ген DDPR, экстрагировали расщеплением pCRDAPB EcoRI и SphI. Этот фрагмент лигировали с pHSG399, расщепленной HincII и SphI для получения плазмиды. Полученная плазмида была обозначена как p399DPR.
Brevi. -ori. вводили в полученную p399DPR для конструирования плазмиды, несущей dapB, автономно реплицируемый в коринеформных бактериях. рНК4 расщепляли рестриктазой KpnI (производимой Takara Shuzo), и расщепленные края затупляли. Образование тупых концов выполняли с использованием набора для затупления ДНК (производимого Takara Shuzo) согласно предложенному способу. После образования тупых концов фосфорилированный линкер BamHI (производимой Takara Shuzo) лигировали для получения модификации так, чтобы фрагмент ДНК, соответствующий Brevi. -ori. -части, мог быть вырезан из рНК4 расщеплением только BamHI. Эту плазмиду расщепляли BamHI, и полученный фрагмент ДНК Brevi. -ori. лигировали с p399DPR, также расщепленной BamHI, для получения плазмиды, содержащей dapB, автономно реплицируемый в коринеформных бактериях. Полученная плазмида была обозначена как pDPRB. Процесс конструирования pDPRB показан в фиг.2.
<2> Определение нуклеотидной последовательности dapB из Brevibacterium lactofermentum
Плазмидную ДНК получали из штамма АJ13107, удерживающего p399DPR, и
ее нуклеотидную последовательность определяли, как описано в Примере 1. Определенная нуклеотидная последовательность и расшифрованная из нее аминокислотная последовательность показаны в SEQ ID 10.
Только аминокислотная последовательность показана в SEQ ID 11.
Пример 3: Получение dapA из Brevibacterium lactofermentum
<1> Получение dapA и конструирование
плазмиды, содержащей dapA
Штамм дикого типа Brevibacterium lactofermentum ATCC 13869 использовали в качестве донора хромосомной ДНК. Хромосомную ДНК получали из штамма ATCC 13869 в
соответствии с обычным способом. Фрагмент ДНК, содержащий dapA, амплифицировали из хромосомной ДНК при помощи ПЦР. В качестве праймеров ДНК, используемых для амплификации, синтезировали соответственно
ДНК 20-меров, имеющих нуклеотидные последовательности, показанные в SEQ ID 12 и SEQ ID 13 в Списке последовательностей, для амплификации района приблизительно 1,5 т. п. н., кодирующего DDPS, на основе
последовательности, известной для Corinebacterium. glutamicum (см. Nucleic Acids Research, 18(21), 6421 (1990); EMBL accession No. X53993). Синтез ДНК и ПЦР выполняли, как описано в Примере 1. pCR1000
(производимую Invitrogen, см. Bio/Technology, 9, 657-663 (1991)) использовали в качестве клонирующего вектора для амплифицированного фрагмента гена 1411 п. н., который лигировали с амплифицированным
фрагментом dapA. Лигирование ДНК выполняли с использованием набора для лигирования ДНК (производимого Takara Shuzo) согласно рекомендуемому способу. Так была сконструирована плазмида, в которой
фрагмент dapA 1411 п. н., амплифицированный из хромосомы Brevibacterium lactofermentum, был лигирован с pCR1000. Плазмида, полученная, как описано здесь, имеющая dapA, происходящий из АТСС 13869, была
обозначена как pCRDAPA.
Трансформированный штамм AJ3106, полученный введением pCRDAPA в штамм Е. coli, был международно депонирован 26 мая 1995 года под депозитным номером FERM BP-5113 в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology of Ministry of International Trade and Industry (postal code: 305, 1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan) на основе Budapest Treaty.
Brevi. -ori. вводили в полученную pCRDAPA для конструирования плазмиды, несущей dapA, автономно реплицируемый в коринеформных бактериях. рНК4 расщепляли рестриктазами КpnI и BamHI (производимой Takara Shuzo) и расщепленные края затупляли. Образование тупых концов выполняли с использованием набора для затупления ДНК (производимого Takara Shuzo) согласно предложенному способу. После образования тупых концов фосфорилированный линкер Smal (производимый Takara Shuzo) лигировали для получения модификации так, чтобы фрагмент ДНК, соответствующий Brevi.-ori.-части, мог быть вырезан из рНК4 расщеплением только Smal. Эту плазмиду расщепляли Smal и полученный фрагмент ДНК Brevi.-ori. лигировали с pCRDAPA, также расщепленной Smal, для получения плазмиды, содержащей dapA, автономно реплицируемый в коринеформных бактериях. Эта плазмида была названа pDPSB. Процесс конструирования pDPSB(Кmr) показан в фиг.3.
<2> Определение нуклеотидной последовательности dapA из Brevibacterium lactofermentum
Плазмидную ДНК получали из штамма АJ13106, несущего pCRDAPA, и ее
нуклеотидную последовательность определяли, как описано в Примере 1. Определенная нуклеотидная последовательность и расшифрованная из нее аминокислотная последовательность показаны в SEQ ID 14. Только
аминокислотная последовательность показана в SEQ ID 15.
Пример 4: Получение lysA из Brevibacterium lactofermentum
<1> Получение lysA и конструирование плазмиды,
содержащей lysA
Штамм дикого типа Brevibacterium lactofermentum ATCC 13869 использовали в качестве донора хромосомной ДНК. Хромосомную ДНК получали из штамма ATCC 13869 в соответствии с
обычным способом. Фрагмент ДНК, содержащий argS, lysA и промотор оперона, содержащего их, амплифицировали из хромосомной ДНК в соответствии с ПЦР. В качестве праймеров ДНК, используемых для
амплификации, применяли синтетические ДНК 23-меров, имеющие нуклеотидные последовательности, изображенные в SEQ ID 16 и SEQ ID 17 в Списке последовательностей соответственно для амплификации района
приблизительно 3,6 т. п. н. для аргинил-т-РНК-синтазы и DDC, на основе последовательности, известной для Corinebacterium glutamicum (см. Molecular Microbiology, 4(11), 1819-1830 (1990); Molecular and
General Genetics, 212, 112-119 (1988)). Синтез ДНК и ПЦР выполняли, как описано в Примере 1. pHSG399 использовали в качестве клонирующего вектора для амплификации фрагмента гена 3579 п. н. pHSG399
расщепляли рестриктазой Smal (производимой Takara Shuzo), которую лигировали с фрагментом ДНК, содержащим амплифицированный lysA. Плазмида, полученная, как описано выше, которая имела lysA,
происходящий из ATCC 13869, была обозначена как p399LYSA.
Фрагмент ДНК, содержащий lysA, экстрагировали расщеплением p399LYSA при помощи KpnI (производимой Takara Shuzo) и BamHI (производимой Takara Shuzo). Этот фрагмент ДНК лигировали с pHSG299, расщепленной KpnI и BamHI. Полученная плазмида была обозначена как p299LYSA. Процесс конструирования p299LYSA показан в фиг.4.
Brevi. -ori. вводили в полученную p299LYSA для конструирования плазмиды, несущей lysA, автономно реплицируемый в коринеформных бактериях. рНК4 расщепляли рестриктазами KpnI и BamHI и расщепленные края затупляли. Образование тупых концов выполняли с использованием набора для затупления ДНК (производимого Takara Shuzo) согласно предложенному способу. После образования тупых концов фосфорилированный линкер KpnI (производимый Takara Shuzo) лигировали для получения модификации так, чтобы фрагмент ДНК, соответствующий Brevi.-ori.-части, мог быть вырезан из рНК4 расщеплением только KpnI. Эту плазмиду расщепляли KpnI и полученный фрагмент ДНК Brevi.-ori. лигировали с p299LYSA, также расщепленной KpnI, для получения плазмиды, содержащей lysA, автономно реплицируемый в коринеформных бактериях. Полученная плазмида была названа pLYSAB. Процесс конструирования pLYSAB показан в фиг. 5.
<2> Определение нуклеотидной последовательности lysA
из Brevibacterium lactofermentum
Плазмидную ДНК p299LYSA получали и ее нуклеотидную последовательность определяли, как описано в Примере 1. Определенная нуклеотидная последовательность и
расшифрованная из нее аминокислотная последовательность показаны в SEQ ID 18. В отношении нуклеотидной последовательности аминокислотная последовательность, кодируемая argS, и аминокислотная
последовательность, кодируемая lysA, показаны в SEQ ID 19 и SEQ ID 20 соответственно.
Пример 5: Получение ddh из Brevibacterium lactofermentum
Ген ddh получали амплификацией
гена ddh из хромосомной ДНК Brevibacterium lactofermentum ATCC 13869 согласно способу ПЦР с применением двух олигонуклеотидных праймеров (SEQ ID 21 и SEQ ID 22), полученных на основе известной
нуклеотидной последовательности гена ddh Corinebacterium glutamicum (Ishino, S. et al., Nucleic Acids Res., 15, 3917 (1987) ). Полученный амплифицированный фрагмент ДНК расщепляли ЕсоТ221 и Aval и
расщепленные края затупляли. После этого этот фрагмент встраивали в сайт Smal pMW119 с получением плазмиды pDDH.
Затем pDDH расщепляли SalI и EcoRI с последующим образованием тупых концов. После этого полученный фрагмент лигировали с pUC18, расщепленной Smal. Полученная таким образом плазмида была обозначена как pUC18DDH.
Brevi. -ori. вводили в полученную pUC18DDH для конструирования плазмиды, несущей ddh, автономно реплицируемый в коринеформных бактериях. рНК4 расщепляли рестриктазами KpnI и BamHI и расщепленные края затупляли. Образование тупых концов выполняли с использованием набора для затупления ДНК (производимого Takara Shuzo) согласно предложенному способу. После образования тупых концов фосфорилированный линкер PstI (производимый Takara Shuzo) лигировали таким образом, чтобы он был вставлен в сайт PstI pHSG299. Плазмида, сконструированная таким образом, была обозначена как рРК4. Затем pUC18DDH расщепляли XbaI и KpnI и полученный фрагмент лигировали с рРК4, расщепленной KpnI и XbaI. Так была сконструирована плазмида, содержащая ddh, автономно реплицируемый в коринеформных бактериях. Эта плазмида была названа pPK4D. Процесс конструирования pPK4D показан в ф иг. 6.
Пример 6: Конструирование плазмиды, содержащей комбинацию мутантного lysC и dapA
Плазмиду, содержащую мутантный lysC, dapA и
начало репликации коринеформных бактерий, конструировали из плазмиды pCRDAPA, содержащей dapA, и плазмиды р399АК9В, содержащей мутантный lysC и Brevi.-ori. p399AK9B полностью разрушали SalI и затем
затупляли и лигировали с линкером EcoRI для конструирования плазмиды, в которой сайт SalI был модифицирован в сайт EcoRI. Полученная плазмида была обозначена как р399АК 9BSE. Мутантный lysC и
Brevi.-ori. вырезали в виде одного фрагмента частичной деградацией p399AK9BSE при помощи EcoRI. Этот фрагмент лигировали с pCRDAPA, расщепленной EcoRI. Полученная плазмида была обозначена как pCRCAB.
Эта плазмида автономно реплицировалась в Е.coli и коринеформных бактериях, и она придает устойчивость к канамицину хозяину, причем эта плазмида содержала комбинацию lysC и dapA. Процесс
конструирования pCRCAB показан в фиг.7.
Пример 7: Конструирование плазмиды, содержащей комбинацию мутантного lysC и dapB
Плазмиду, содержащую мутантный lysC и dapB,
конструировали из плазмиды р399АК9, имеющей мутантный lysC, и плазмиды p399DPR, имеющей dapB. Фрагмент 1101 п. н. , содержащий структурный ген DDPR, экстрагировали расщеплением p399DPR EcoRI и SphI.
Этот фрагмент лигировали с р399АК9, расщепленной SalI и затем затупленной и дополнительно расщепленной SphI, для конструирования плазмиды, содержащей комбинацию мутантного lysC и dapB. Эта плазмида
была обозначена как р399АКDDPR.
Затем Brevi. -ori. вводили в полученную p399AKDDPR. Плазмиду рНК4, содержащую Brevi.-ori., расщепляли рестриктазой КрnI (производимой Takara Shuzo) и расщепленные края затупляли. Образование тупых концов выполняли с применением набора для затупления ДНК (производимого Takara Shuzo) в соответствии с рекомендуемым способом. После образования тупых концов фосфорилированный линкер BamHI (производимый Takara Shuzo) лигировали для получения модификации так, чтобы фрагмент ДНК, соответствующий Brevi.-ori.-части, мог быть вырезан расщеплением только BamHI. Эту плазмиду расщепляли BamHI и полученный фрагмент ДНК с Brevi.-ori. лигировали с p399AKDDPR, расщепленной BamHI, для конструирования плазмиды, содержащей мутантный lysC и dapB, автономно реплицируемые в коринеформных бактериях. Сконструированная плазмида была обозначена как рСВ. Процесс конструирования рСВ показан в фиг.8.
Пример 8: Конструирование плазмиды,
содержащей комбинацию dapA и dapB
Плазмиду pCRDAPA, содержащую dapA, расщепляли KpnI и EcoRI для экстракции фрагмента ДНК, содержащего dapA, который лигировали с векторной плазмидой pHSG399,
расщепленной KpnI и EcoRI. Полученная плазмида была обозначена как p399DPS.
С другой стороны, плазмиду pCRDAPB, содержащую dapB, расщепляли SacII и EcoRI для экстракции фрагмента ДНК 2,0 т. п. н., содержащего район, кодирующий DDPR, который лигировали с p399DPS, расщепленной SacII и EcoRI, для конструирования плазмиды, содержащей комбинацию dapA и dapB. Полученную плазмиду обозначали как р399АВ.
Затем в р399АВ вводили Brevi.-ori. pHK4, содержащую Brevi.-ori., расщепляли рестриктазой BamHI (производимой Takara Shuzo), и расщепленные края затупляли. Образование тупых концов выполняли с использованием набора для затупления ДНК (производимого Takara Shuzo) согласно предложенному способу. После образования тупых концов фосфорилированный линкер KpnI (производимый Takara Shuzo) лигировали для получения модификации так, чтобы фрагмент ДНК, соответствующий Brevi. -ori. -части, мог быть вырезан из pHK4 расщеплением только KpnI. Эту плазмиду расщепляли KpnI и полученный фрагмент ДНК Brevi. -ori. лигировали с р399АВ, расщепленной KpnI, для конструирования плазмиды, содержащей dapA и dapB, автономно реплицируемые в коринеформных бактериях. Сконструированная плазмида была обозначена как рАВ. Процесс конструирования рАВ показан в фиг.9.
Пример 9: Конструирование плазмиды, содержащей комбинацию ddh и lysA
Плазмиду
pUC18DDH, содержащую ddh, расщепляли EcoRI и XbaI для экстракции фрагмента ДНК, содержащего ddh. Фрагмент ddh лигировали с плазмидой p399LYSA, содержащей lysA, расщепленной BamHI и XbaI, с затуплением
расщепленных краев после расщепления. Полученная плазмида была обозначена как p399DL. Процесс конструирования p399DL показан в фиг. 10.
Затем в p399DL вводили Brevi.-ori. рНК4 расщепляли XbaI и BamHI и расщепленные края затупляли. После образования тупых концов фосфорилированный линкер XbaI лигировали для получения модификации так, чтобы фрагмент ДНК, соответствующий Brevi. -ori. -части, мог быть вырезан из рНК4 расщеплением только XbaI. Эту плазмиду расщепляли XbaI и полученный фрагмент ДНК Brevi. -ori. лигировали с p399DL, расщепленной также XbaI, для конструирования плазмиды, содержащей ddh и lysA, автономно реплицируемые в коринеформных бактериях. Сконструированная плазмида была обозначена как pDL. Процесс конструирования pDL показан в фиг. 11.
Пример 10: Конструирование плазмиды, содержащей комбинацию мутантного lysC, dapA и dapB
p399DPS разрушали EcoRI и SphI с образованием тупых концов с последующей экстракцией фрагмента гена
dapA. Этот фрагмент лигировали с р399АК9, расщепленной SalI, и затупляли для конструирования плазмиды р399СА, в которой сосуществовали мутантный lysC и dapA.
Плазмиду pCRDAPB, содержащую dapB, расщепляли EcoRI и концы затупляли с последующим расщеплением SacI для экстракции фрагмента ДНК 2,0 т. п. н., содержащего dapB. Плазмиду р399СА, содержащую dapA и мутантный lysC, расщепляли SpeI и концы затупляли, а затем расщепляли SacI и лигировали с экстрагированным фрагментом dapB для получения плазмиды, содержащей мутантный lysC, dapA и dapB. Эта плазмида была обозначена как р399САВ.
Затем в р399САВ вводили Brevi.-ori. Плазмиду рНК4, содержащую Brevi.-ori. , расщепляли рестриктазой BamHI (производимой Takara Shuzo) и расщепленные края затупляли. Образование тупых концов выполняли с использованием набора для затупления ДНК (производимого Takara Shuzo) согласно предложенному способу. После образования тупых концов фосфорилированный линкер KpnI (производимый Takara Shuzo) лигировали для получения модификации так, чтобы фрагмент ДНК, соответствующий Brevi.-ori.-части, мог быть вырезан из рНК4 расщеплением только KpnI. Эту плазмиду расщепляли KpnI и полученный фрагмент ДНК Brevi.-ori. лигировали с р399САВ, расщепленной также KpnI, для конструирования плазмиды, содержащей комбинацию мутантного lysC, dapA и dapB, автономно реплицируемых в коринеформных бактериях. Сконструированная плазмида была обозначена как рСАВ. Процесс конструирования рСАВ показан в фиг.12.
Пример 11: Конструирование плазмиды, содержащей комбинацию
мутантного lysC, dapA, dapB и lysA
Плазмиду p299LYSA, содержащую lysA, расщепляли KpnI и BamHI и концы затупляли, а затем экстрагировали фрагмент гена lysA. Этот фрагмент лигировали с рСАВ,
расщепленной HpaI (производимой Takara Shuzo), и концы затупляли для конструирования плазмиды, содержащей комбинацию мутантного lysC, dapA, dapB и lysA, автономно реплицируемых в коринеформных
бактериях. Сконструированная плазмида была обозначена как pCABL. Процесс конструирования pCABL показан в фиг. 13. Следует заметить, что фрагмент гена lysA вводят в сайт HpaI во фрагменте ДНК,
содержащем ген dapB в pCABL, однако сайт HpaI локализован против хода транскрипции от промотора для гена dapB (номера нуклеотидов 611-616 в SEQ ID 10), и ген dapB не является отсоединенным.
Пример 12: Конструирование плазмиды, содержащей комбинацию мутантного lysC, dapA, dapB, ddh и lysA
pHSG299 расщепляли XbaI и KpnI, лигировали с p399DL, содержащей ddh и lysA,
расщепленной XbaI и KpnI. Сконструированная плазмида была обозначена как p299DL. p299DL расщепляли XbaI и KpnI и концы затупляли. После образования тупых концов экстрагировали фрагмент ДНК, содержащий
ddh и lysA. Этот фрагмент ДНК лигировали с плазмидой рСАВ, содержащей комбинацию мутантного lysC, dapA и dapB, расщепленной HpaI, и концы затупляли для конструирования плазмиды, содержащей комбинацию
мутантного lysC, dapA, dapB, lysA и ddh, автономно реплицируемых в коринеформных бактериях. Сконструированная плазмида была обозначена как pCABDL. Процесс конструирования pCABDL показан в фиг.14.
Пример 13: Введение плазмид, содержащих гены для биосинтеза L-лизина, в L-лизин-продуцирующую бактерию Brevibacterium lactofermentum.
Плазмиды, содержащие ген для биосинтеза L-лизина, сконструированные, как описано выше, а именно p399KK9B(Cmr), pDPSB(Kmr), pDPRB (Cmr), pLYSAB(Cmr), pPK4D(Cmr), pCRCAB(Kmr), pAB(Cmr), pCB(Cmr), pDL(Cmr), pCAB(Cmr), pCABL(Cmr) и pCABDL (Cmr), вводили в L-лизин-продуцирующую бактерию AJ11082 (NRRL B-11470) Brevibacterium lactofermentum соответственно. Штамм AJ11082 обладает устойчивостью к АЕС. Плазмиды вводили согласно электроимпульсному способу (Sugimoto et al. , Japanese Patent Laid-open No.2-207791). Трансформанты отбирали на основе маркеров устойчивости к лекарственному средству, которым обладают соответствующие плазмиды. Трансформанты отбирали на полной среде, содержащей 5 мкг/мл хлорамфеникола, при введении плазмиды, содержащей ген устойчивости к хлорамфениколу, или трансформанты отбирали на полной среде, содержащей 25 мкг/мл канамицина, при введении плазмиды, содержащей ген устойчивости к канамицину.
Пример 14: Получение L-лизина
Каждый из трансформантов, полученных в Примере 13, культивировали в среде для продуцирования L-лизина для оценки
продуктивности L-лизина трансформантом. Среда для продуцирования L-лизина имела следующий состав.
[Среда для продуцирования L-лизина]
Следующие компоненты, кроме карбоната
кальция, (на 1 л) растворяли с доведением рН 8,0 при помощи КОН. Среду стерилизовали при 115oС в течение 15 минут, после чего к ней добавляли карбонат кальция, стерилизованный отдельно в
горячем воздухе в сухом состоянии.
Глюкоза - 100 г
(NH4)SO4 - 55 г
КН2РO4 - 1 г
МgSO4•7Н2О - 1 г
Биотин - 500 мкг
Тиамин - 2000 мкг
FeSO4•7H2O - 0,01 г
MnSO4•7H2O - 0,01 г
Никотинамид - 5 мг
Гидролизат белка(Маmеnоu) - 30 мл
Карбонат кальция - 50 г
Каждый из различных типов трансформантов и родительский штамм инокулировали в среду, имеющую
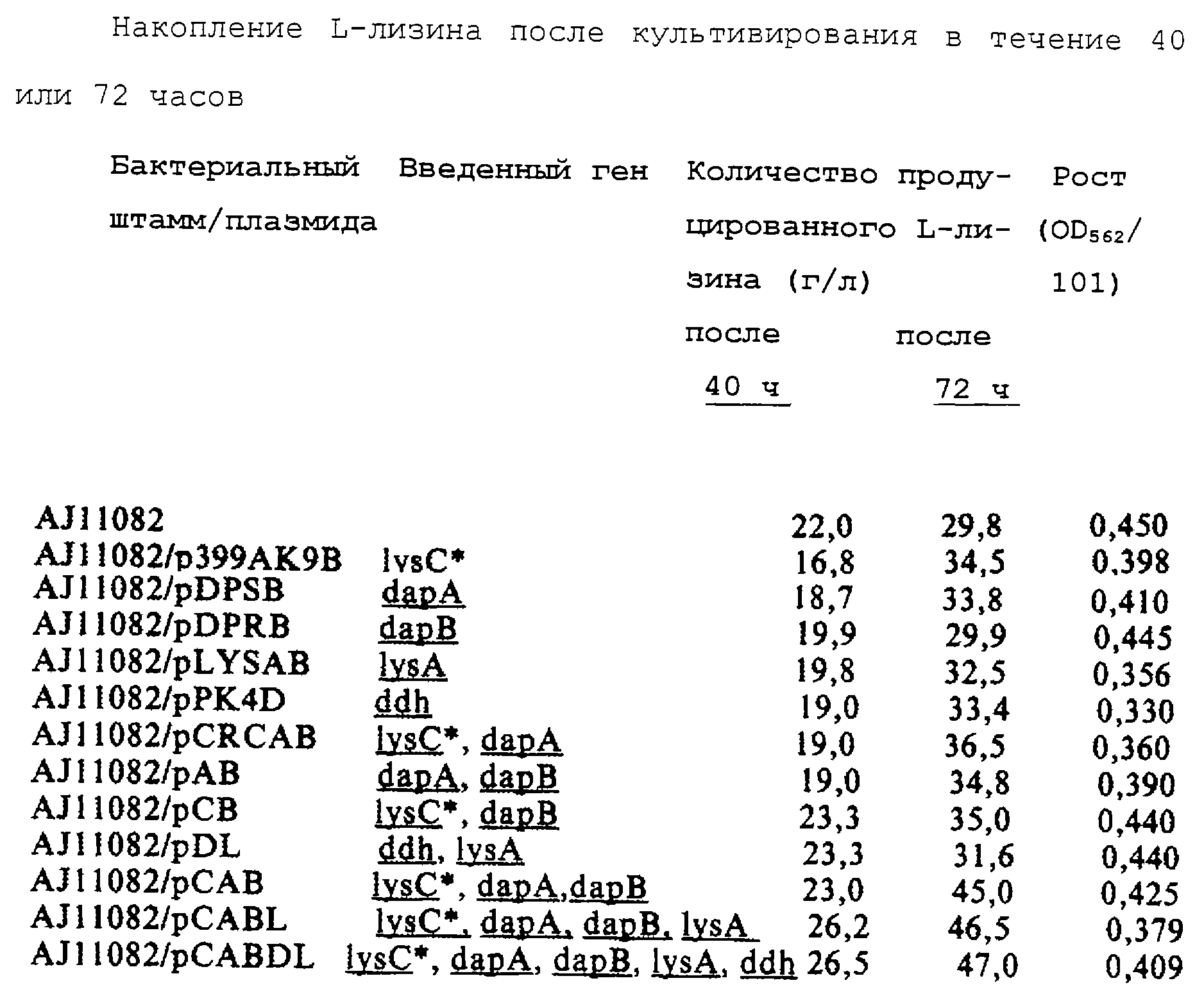
состав, описанный выше, для проведения культивирования при 31,5oС с возвратно-поступательным качанием. Количество продуцируемого L-лизина после 40 и 72 часов культивирования и рост после 72
часов (OD562) показаны в таблице. В этой таблице lysC* обозначает мутантный lysC. Рост определяли количественно путем измерения OD при 560 нм после 101-кратного разведения.
Как показано в таблице, при отдельном усилении мутантного lysC, dapA или dapB количество продуцируемого L-лизина было больше количества, продуцируемого родительским штаммом после 72 часов культивирования, или равноценно этому количеству, однако количество продуцируемого L-лизина было ниже количества, продуцируемого родительским штаммом после 40 часов культивирования. То есть скорость продуцирования L-лизина снижалась при культивировании в течение короткого периода. Подобным образом при усилении мутантного lysC и dapA или dapA и dapB в сочетании количество продуцируемого L-лизина было больше, чем количество, продуцируемое родительским штаммом, после 72 часов культивирования, однако количество продуцируемого L-лизина было меньше количества, продуцируемого родительским штаммом, после 40 часов культивирования. Таким образом, скорость продуцирования L-лизина снижается.
С другой стороны, при отдельном усилении lysC или ddh или при совместном усилении lysC и ddh количество продуцируемого L-лизина было больше, чем количество, продуцируемое родительским штаммом после 40 часов культивирования, однако количество продуцируемого L-лизина было в результате меньше, чем количество, продуцируемое родительским штаммом после длительного периода культивирования вследствие уменьшения в росте.
Напротив, в случае штамма, в котором dapB был усилен вместе с мутантным lysC, рост был улучшенным, скорость продуцирования L-лизина успешно восстанавливалась в коротком периоде культивирования, и накопленное количество L-лизина улучшалось в длительном периоде культивирования. В случае штамма, в котором одновременно усиливались три гена, мутантный lysC, dapA и dapB, продуктивность L-лизина дополнительно улучшалась. Как скорость продуцирования L-лизина, так и количество накапливаемого L-лизина улучшались ступенчатым образом путем успешного усиления lysA и ddh.
Промышленная применимость
Согласно данному изобретению способность
продуцирования L-лизина коринеформными бактериями может быть улучшена и может быть также улучшена скорость роста.
Скорость продуцирования L-лизина может быть улучшена и продуктивность может быть также улучшена в коринеформных продуцирующих L-лизин бактериях путем усиления dapB вместе с мутантным lysC. Скорость продуцирования L-лизина и продуктивность могут быть дополнительно улучшены путем успешного усиления dapA, lysA и ddh в дополнение к вышеуказанным генам.
Список последовательностей приведен в конце описания.
Реферат
Изобретение относится к биотехнологии и касается производства L-лизина. Способ получения L-лизина предусматривает выращивание коринеформных бактерий, трансформированных рекомбинантной ДНК (варианты), представленной в описании в виде перечня последовательностей. Рекомбинантная ДНК содержит а) последовательность ДНК, кодирующую аспартокиназу, у которой ингибирование по типу обратной связи L-лизином и L-треонином по существу десенсибилизировано, и б) последовательность ДНК, кодирующую дигидродипиколинатредуктазу. Рекомбинантная ДНК автономно реплицируется в клетках коринеформных бактерий. Выращивание трансформированных бактерий осуществляют в среде, подходящей для продуцирования и накопления L-лизина в культуре бактерий, и L-лизин извлекают из культуры. Использование изобретения позволяет улучшить L-лизинпродуцирующую способность и скорость роста продуцента. 8 с.п. ф-лы, 14 ил., 1 табл.

Комментарии