Микроорганизм рода corynebacterium, обладающий l-изолейцин-продуцирующей способностью, и способ получения l-изолейцина с использованием этого микроорганизма - RU2698394C1
Код документа: RU2698394C1
Чертежи












































Описание
Область изобретения
Настоящее изобретение относится к микроорганизму рода Corynebacterium, обладающему L-изолейцин-продуцирующей способностью, и способу получения L-изолейцина с использованием этого микроорганизма.
Предшествующий уровень техники
К аминокислотам с разветвленной цепью относятся три аминокислоты: L-валин, L-лейцин и L-изолейцин, и они используются в промышленности в качестве пищевых добавок, фармацевтических агентов и так далее. В частности, L-изолейцин метаболизируется в мышцах для производства энергии и вовлечен в синтез гемоглобина, в то же время осуществляя функцию снятия усталости и стимуляции роста. Соответственно, L-изолейцин имеет разнообразные применения в форме жидкого препарата, пищевой добавки и так далее, а его использование в качестве добавок при спортивных тренировках возрастает.
L-изолейцин образуется биосинтезом из предшественников пирувата и 2-кетобутирата через три метаболических промежуточных соединения (Jinhwan Park et al., Appl. Microbial. Biotechnol. 85: 491-506, 2010).
Однако, в биосинтезе L-изолейцина оба фермента, треониндегидратаза (ген ilvA), которая катализирует образование 2-кетобутирата из L-треонина, и затем синтаза ацетогидроксикислот (ген ilvBN), подвержены ингибированию по типу обратной связи L-изолейцином.
Соответственно, известно, что регуляция ферментов, вовлеченных в биосинтез, путем освобождения от ингибирования по типу обратной связи L-изолейцином является важным элементом в получении штамма, продуцирующего L-изолейцин с высоким выходом (Jinhwan Park et al., Biotechnology Journal, 560-577, 2010). Кроме того, аминокислоты с разветвленной цепью синтезируются посредством тех же путей биосинтеза, что и пируват, и соответственно, массовая продукция L-изолейцина с высоким выходом требует отрегулированного предоставления L-треонина на стадии, предшествующей 2-кетобутирату (Патент Кореи No. 10-0823044). Техническая задача
Авторы настоящего изобретения исследовали новый путь биосинтеза, в котором не используется L-треонин в качестве предшественника, и ввели ген, обладающий активностью цитрамалатсинтазы, в путь биосинтеза L-изолейцина. В результате авторы настоящего изобретения подтвердили, что такое введение улучшает продуцирование L-изолейцина, тем самым создав настоящее изобретение.
Техническое решение
Задача настоящего изобретения заключается в том, чтобы предложить рекомбинантный микроорганизм, имеющий новый введенный путь биосинтеза и обладающий L-изолейцин-продуцирующей способностью.
Другая задача настоящего изобретения заключается в том, чтобы предложить способ получения L-изолейцина, включающий культивирование рекомбинантного микроорганизма, имеющего новый введенный путь биосинтеза, в среде и выделение L-изолейцина из микроорганизма или из среды.
Полезные эффекты изобретения
Микроорганизм рода Corynebacterium, обладающий L-изолейцин-продуцирующей способностью, по настоящему изобретению имеет введенную активность цитрамалатсинтазы и может продуцировать L-изолейцин с высоким выходом посредством нового пути биосинтеза, в котором L-треонин не используется в качестве предшественника.
Предпочтительное воплощение изобретения
В одном аспекте настоящего изобретения для достижения этих целей предложен микроорганизм рода Corynebacterium, обладающий L-изолейцин-продуцирующей способностью и содержащий белок, обладающий активностью цитрамалатсинтазы.
Термин "L-изолейцин" в настоящем изобретении относится к незаменимой аминокислоте и структурно к L-аминокислоте формулы НО2ССН(NH2)СН(СН3)СН2СН3, соответствующей аминокислоте с разветвленной цепью наряду с L-валином и L-лейцином.
Фраза "обладающий L-изолейцин-продуцирующей способностью" в настоящем изобретении указывает на то, что когда соответствующий микроорганизм культивируют в среде, он проявляет способность накапливать L-изолейцин в среде или в самом микроорганизме. Такая L-изолейцин-продуцирующая способность может наследоваться от штамма дикого типа или обеспечиваться или усиливаться путем модификации штамма.
Например, для обеспечения L-изолейцин-продуцирующей способности микроорганизма можно применять способы, используемые для модификации микроорганизма, раздельно или в комбинации, такие как получение ауксотрофного, устойчивого к аналогу мутанта или мутанта контроля метаболизма, или конструирование рекомбинантного штамма, у которого активность фермента, вовлеченного в биосинтез L-изолейцина, усилена.
Кроме того, в случае усиления активности фермента, вовлеченного в биосинтез L-изолейцина, ген-мишень, вовлеченный в биосинтез L-изолейцина, в частности гены ilvG, ilvM, ilvE, ilvD и ilvA, может быть усилен в отдельности или в комбинации из двух или более генов. Гены ilvG, ilvM, ilvE, ilvD и ilvA кодируют большие и малые субъединицы изофермента II синтазы ацетогидроксикислот, трансаминазы, дегидратазы дигидроксикислот и треониндезаминазы, соответственно.
Термин "цитрамалатсинтаза (ЕС2.3.1.182)" по настоящему изобретению означает каталитический фермент для трансформации ацетил-СоА и пирувата в цитрамалат и кофермент А. Этот фермент обнаружен в археях, таких как Methanocaldococcus jannaschii (Howell et al., J Bacteriol. 181: 331-333, 1999), Leptospira interogans (Xu et al., J Bacteriol. 186: 5400-5409, 2004) и Geobacter sulfurreducens, и вовлечен в треонин-независимый путь биосинтеза изолейцина (Risso et al., J Bacteriol. 190: 2266-2274, 2008). Этот фермент обладает высокой специфичностью в отношении пирувата как субстрата, и известно, что он обычно ингибируется изолейцином.
Термины "белок, обладающий активностью цитрамалатсинтазы" и "ген, кодирующий этот белок" по настоящему изобретению могут включать любые белки, обладающие активностью цитрамалатсинтазы, как описано выше, и любые гены, кодирующие эти белки, без ограничения.
В частности, белок, обладающий активностью цитрамалатсинтазы, может иметь происхождение из Methanocaldococcus, и более конкретно, Methanocaldococcus jannaschii. Цитрамалатсинтаза имеет аминокислотную последовательность SEQ ID NO: 1, и ген cimA, кодирующий эту последовательность, имеет нуклеотидную последовательность SEQ ID NO: 2. Аминокислотные последовательности белка, проявляющего активность, различаются в зависимости от вида или штамма микроорганизма, или множественные нуклеиновые кислоты, обладающие идентичной функцией, могут кодировать любые заданные белки вследствие вырожденности генетического кода, и соответственно, последовательности не ограничены SEQ ID NO, описанными выше.
Кроме того, любые белки, по существу обладающие активностью цитрамалатсинтазы, могут быть включены в объем настоящего изобретения без ограничения, при условии что белок по существу обладает активностью цитрамалатсинтазы, как аминокислотная последовательность, демонстрирующая 70% или больше гомологии с SEQ ID NO: 1, в частности 80% или больше гомологии с этой последовательностью, более конкретно 90% или больше гомологии с этой последовательностью, более конкретно 95% или больше гомологии с этой последовательностью и еще более конкретно 98% или больше гомологии с этой последовательностью.
Дополнительно, если аминокислотная последовательность, гомологичная этим последовательностям, по существу обладает биологической активностью идентичной или соответствующей белку с SEQ ID NO: 1, аминокислотная последовательность, часть которой делетирована, модифицирована, заменена или добавлена, также очевидно включена в объем настоящего изобретения.
Кроме того, ген, кодирующий цитрамалатсинтазу, может иметь нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 1. Здесь, вследствие вырожденности кодона или принимая во внимание кодон, предпочтитаемый организмом для экспрессии белка, полипептид может иметь разные преобразования в кодирующих областях в пределах объема настоящего изобретения, причем аминокислотная последовательность белка не изменена. Полинуклеотидная последовательность может, например, иметь полинуклеотидную последовательность SEQ ID NO: 2 и нуклеотидную последовательность, гомологичную ей на 80% или больше, в частности гомологичную ей на 90% или больше и более конкретно гомологичную ей на 99% или больше, но не ограничиваясь этим.
Кроме того, для выделения мутанта, обладающего повышенной активностью цитрамалатсинтазы, в настоящем изобретении предложена библиотека мутантов CimA(M), полученная с использованием подверженной ошибкам ПЦР на базе мутантов белка SEQ ID NO: 1.
В частности, библиотека мутантов CimA(M) по настоящему изобретению может включать следующее:
CimA(M)m1, имеющий аминокислотную последовательность SEQ ID NO: 3 (в частности, возможно кодируемую полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 4);
CimA(M)m2, имеющий аминокислотную последовательность SEQ ID NO: 5 (в частности, возможно кодируемую полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 6);
CimA(M)m3, имеющий аминокислотную последовательность SEQ ID NO: 7 (в частности, возможно кодируемую полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 8);
CimA(M)m4, имеющий аминокислотную последовательность SEQ ID NO: 9 (в частности, возможно кодируемую полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 10);
CimA(M)m5, имеющий аминокислотную последовательность SEQ ID NO: 11 (в частности, возможно кодируемую полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 12);
CimA(M)m6, имеющий аминокислотную последовательность SEQ ID NO: 13 (в частности, возможно кодируемую полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 14), и
CimA(M)m7, имеющий аминокислотную последовательность SEQ ID NO: 15 (в частности, возможно кодируемую полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 16).
Кроме того, любые белки, по существу обладающие активностью мутанта цитрамалатсинтазы, могут быть включены в объем настоящего изобретения без ограничения, при условии что этот белок по существу обладает активностью мутанта цитрамалатсинтазы, как аминокислотная последовательность, демонстрирующая 80% или больше гомологии с аминокислотными последовательностями SEQ ID NO, в частности 90% или больше, еще более конкретно 95% или больше и даже еще более конкретно 99% или больше.
Термин "гомология" в настоящем изобретении относится к степени сходства с данной аминокислотной или нуклеотидной последовательностью, и может быть представлен в процентах. В настоящем изобретении гомология с последовательностью, обладающей активностью идентичной или сходной с данной аминокислотной или нуклеотидной последовательностью, представлена как "% гомологии". Например, гомологию можно определить с использованием стандартного программного обеспечения, в частности BLAST 2.0, для расчета таких параметров как коэффициент (score), идентичность и сходство, или путем сравнения последовательностей путем Саузерн-гибридизации в определенных строгих условиях. Подходящие условия гибридизации можно определить из уровня техники (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 1989) и можно определить с использованием способов, хорошо известных специалисту в данной области техники.
В конкретном воплощении микроорганизм по настоящему изобретению может включать цитрамалатсинтазу с SEQ ID NO: 1, имеющую происхождение из Methanocaldococcus jannaschii, ее мутант и ген, кодирующий белок с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, 3, 5, 7, 9, 11, 13 и 15. Более конкретно, желаемый ген может экспрессироваться при трансформации рекомбинантных векторов, включающих каждый ген, в этот микроорганизм.
Термин "рекомбинантный вектор" в настоящем изобретении относится к ДНК-конструкции, содержащей нуклеотидную последовательность полинуклеотида, кодирующую желаемый белок, который функционально связан с соответствующей регуляторной последовательностью для экспрессии желаемого белка в подходящем хозяине. Регуляторная последовательность включает промотор, который может инициировать транскрипцию, произвольную операторную последовательность для регуляции такой транскрипции, последовательность, кодирующую подходящую мРНК сайта связывания с рибосомой, и последовательность, регулирующую терминацию транскрипции и трансляции. После того как рекомбинантным вектором трансформировали подходящую клетку-хозяин, он может реплицироваться или функционировать независимо от генома хозяина и может сам интегрировать в геном.
Рекомбинантный вектор, используемый в настоящем изобретении, специально не ограничен, при условии что он способен реплицироваться в клетке-хозяине, и можно использовать любые векторы, известные в данной области техники. Примеры традиционно используемых векторов могут включать природные или рекомбинантные плазмиду, космиду, вирус и бактериофаг. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A или им подобные. В качестве плазмидного вектора можно использовать тип pBR, тип pUC, тип pBluescriptII, тип pGEM, тип pTZ, тип pCL, тип рЕТ или им подобные. Векторы, применяемые в настоящем изобретении, специально не ограничены, и можно использовать любые известные векторы экспрессии. В частности, можно использовать вектор pECCG117, pDZ, pACYC177, pACYC184, pCL, pUC19, pBR322, pMW118 или pCClBAC или им подобные.
В одном примере воплощения полинуклеотид, кодирующий желаемый белок, может быть введен в хромосому клетки-хозяина с помощью вектора для вставки в хромосому. Введение полинуклеотида в хромосому может быть выполнено любым способом, известным в данной области техники, например, гомологичной рекомбинацией, но не ограничиваясь этим.
Термин "трансформация" в настоящем изобретении означает, что рекомбинантный вектор, содержащий полинуклеотид, кодирующий белок-мишень, введен в клетку-хозяина таким образом, что белок, кодируемый этим полинуклеотидом, может экспрессироваться в данной клетке-хозяине. Подвергнутый трансформации полинуклеотид может быть любым, при условии что полипептид может экспрессироваться в данной клетке-хозяине независимо от того, вставлен ли полинуклеотид в хромосому или расположен за пределами хромосомы.
Дополнительно, фраза "функционально связанный" относится к функциональной связи между промоторной последовательностью, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего белок-мишень по настоящему изобретению, и последовательностью гена.
Между тем, в конкретном воплощении настоящего изобретения микроорганизм рода Corynebacterium может дополнительно включать повышенные активности 3-изопропилмалатдегидрогеназы и 3-изопропилмалатдегидратазы.
Фраза "усиление активности белка" в настоящем изобретении относится к активности белка, усиленной по сравнению с таковой у белка дикого типа или предварительно модифицированного белка. В частности, усиление активности белка может включать увеличение активности, возникающее в результате улучшения активности эндогенного гена, кодирующего белок, амплификации эндогенного гена с помощью внутренних или внешних факторов, делеции регуляторного фактора, подавляющего экспрессию гена, увеличения числа копий гена, введения чужеродного гена, модификации последовательности регуляции экспрессии, в частности, замены или модификации промотора, или мутации в гене.
В частности, повышение активности 3-изопропилмалатдегидрогеназы или 3-изопропилмалатдегидратазы может быть осуществлено способом, выбранным из группы, состоящей из:
1) увеличения числа копий полинуклеотида, кодирующего указанный белок;
2) модификации последовательности регуляции экспрессии для увеличения экспрессии полинуклеотида;
3) модификации полинуклеотидной последовательности на хромосоме для повышения активности белка; делеции регуляторного фактора, подавляющего экспрессию гена; или
4) их комбинации; однако способ специально не ограничен этим.
Способ увеличения числа копий полинуклеотида может быть осуществлен в форме функциональной связи с вектором или путем вставки в хромосому в клетке-хозяине; однако, способ специально не ограничен этим. В частности, он может быть выполнен путем вставки вектора в клетку-хозяина, где вектор функционально связан с полинуклеотидом, кодирующим белок по настоящему изобретению, и может реплицироваться и функционировать независимо от клетки-хозяина. Альтернативно, он может быть выполнен способом увеличения числа копий полинуклеотида в хромосоме в клетке-хозяине путем вставки вектора в клетку-хозяина, где вектор функционально связан с полинуклеотидом и делает возможной вставку полинуклеотида в хромосому в клетке-хозяине.
Способ модификации последовательности регуляции экспрессии для увеличения экспрессии полинуклеотида может быть осуществлен путем индуцирования мутации в этой последовательности посредством делеции, вставки или неконсервативной или консервативной замены в последовательности нуклеиновой кислоты или их комбинации, так что активность последовательности регуляции экспрессии может быть усилена, или путем замены этой последовательности на последовательность нуклеиновой кислоты, обладающей усиленной активностью; однако способ специально не ограничен этим. Последовательность регуляции экспрессии включает промотор, операторную последовательность, последовательность, кодирующую сайт связывания с рибосомой, последовательность, регулирующую терминацию транскрипции и трансляции, или им подобные, но специально не ограничиваясь этим.
Сильный гетерологичный промотор вместо исходного промотора может быть присоединен выше по ходу транскрипции от единицы экспрессии полинуклеотида, и примеры сильного промотора могут включать промотор cj7 (Патент Кореи No. 10-0620092 и Международная публикация No. WO 2006/065095), промотор EF-Tu, промотор groEL, промотор асеА или асеВ, и более предпочтительно промотор cj7, как промотор, имеющий происхождение из Corynebacterium, функционально связан таким образом, что уровень экспрессии полинуклеотида, кодирующего этот белок, может быть повышен.
Дополнительно способ модификации полинуклеотидной последовательности в хромосоме может быть осуществлен путем индуцирования мутации в последовательности регуляции экспрессии посредством делеции, вставки, неконсервативной или консервативной замены в последовательности нуклеиновой кислоты, или их комбинации, так что активность полинуклеотидной последовательности может быть усилена, или путем замены последовательности на полинуклеотидную последовательность, модифицированную таким образом, что она имеет повышенную активность; однако способ специально не ограничен этим.
Термины "3-изопропилмалатдегидрогеназа" и "3-изопропилмалатдегидратаза" в настоящем изобретении относятся к ферментам, вовлеченным в пути биосинтеза L-лейцина, L-изолейцина и L-валина. Белок, обладающий активностью этих ферментов, может включать любые белки, обладающие активностью этих ферментов, и ген, кодирующий этот белок, может включать любые гены, кодирующие этот белок, без ограничения.
В частности, в настоящем изобретении 3-изопропилмалатдегидрогеназа (ЕС1.1.1.85) может иметь происхождение из микроорганизма рода Corynebacterium, более конкретно из Corynebacterium glutamicum. Между тем, 3-изопропилмалатдегидрогеназа в настоящем изобретении может иметь аминокислотную последовательность SEQ ID NO: 17. Кроме того, любой белок, по существу обладающий активностью 3-изопропилмалатдегидрогеназы, как аминокислотная последовательность по существу демонстрирующая 70% или более гомологии с SEQ ID NO: 17, в частности 80% или более, более конкретно 90% или более, еще более конкретно 95% или более и даже еще более конкретно 99% или более, может быть включен в объем настоящего изобретения без ограничения.
Кроме того, ген, кодирующий 3-изопропилмалатдегидрогеназу, может иметь нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 17. Аминокислотные последовательности белка, проявляющего активность, различаются в зависимости от вида или штамма микроорганизма, или множество нуклеиновых кислот, обладающих идентичной функцией, может кодировать заданные белки вследствие вырожденности генетического кода, и соответственно ген не ограничен раскрытыми SEQ ID NO. Например, ген может иметь нуклеотидную последовательность SEQ ID NO: 18 или нуклеотидную последовательность, гомологичную SEQ ID NO: 18 на 80%, в частности на 90% или более и более конкретно на 99% или более.
Дополнительно, 3-изопропилмалатдегидратаза (ЕС4.2.1.33) в настоящем изобретении может иметь происхождение из микроорганизма рода Corynebacterium, более конкретно из Corynebacterium glutamicum. Между тем, 3-изопропилмалатдегидратаза в настоящем изобретении состоит из двух малых/больших субъединиц, которые могут иметь аминокислотную последовательность SEQ ID NO: 19 и 21. Кроме того, любые белки, по существу обладающие активностью 3-изопропилмалатдегидратазы, могут быть включены в объем настоящего изобретения без ограничения, при условии что белок по существу обладает активностью 3-изопропилмалатдегидратазы, как аминокислотная последовательность, демонстрирующая 70% или более гомологии с SEQ ID NO: 19 и 21, в частности 80% или более, более конкретно 90% или более, еще более конкретно 95% или более и даже еще более конкретно 99% или более.
Дополнительно ген, кодирующий 3-изопропилмалатдегидратазу, может иметь нуклеотидную последовательность, кодирующую аминокислотные последовательности SEQ ID NO: 19 и 21. Аминокислотные последовательности белка, проявляющего эту активность, различаются в зависимости от вида или штамма микроорганизма, или множество нуклеиновых кислот, обладающих идентичной функцией, может кодировать любые заданные белки вследствие вырожденности генетического кода, и соответственно, ген не ограничен этими SEQ ID NO. Например, ген может иметь нуклеотидные последовательности SEQ ID NO: 20 и 22 или нуклеотидные последовательности, гомологичные этим последовательностям на 80%, в частности на 90% или более и более конкретно на 99% или более.
Между тем, в конкретном воплощении настоящего изобретения микроорганизм рода Corynebacterium может включать фермент, вовлеченный в путь биосинтеза L-изолейцина, дополнительно включающий повышенную активность.
В настоящем изобретении термин "фермент, вовлеченный в путь биосинтеза L-изолейцина" может включать аспартаткиназу (ген lysC), аспартат-β-полуальдегиддегидрогеназу (ген asd), гомосериндегидрогеназу (ген hom), гомосеринкиназу (ген thrB), треонинсинтазу (ген thrC), треониндегидратазу (ген ilvA), аминотрансферазу (ген ilvE) или им подобные, но не ограничиваясь ими.
Между тем, в конкретном воплощении настоящего изобретения микроорганизм рода Corynebacterium может дополнительно включать инактивированную пируватдегидрогеназу.
Термин "пируватдегидрогеназа (ЕС1.2.4.1)" в настоящем изобретении относится к ферменту, превращающему пируват в ацетил-СоА и СО2. Согласно настоящему изобретению можно получить штамм с делецией гена асеЕ, кодирующего этот фермент, для увеличения запаса ацетил-СоА, используемого в качестве предшественника гена cimA.
В частности, пируватдегидрогеназа (ЕС1.2.4.1) в настоящем изобретении может иметь происхождение из микроорганизма рода Corynebacterium, и более конкретно из Corynebacterium glutamicum. Между тем, пируватдегидрогеназа в настоящем изобретении может иметь аминокислотную последовательность SEQ ID NO: 25. Кроме того, любой белок, по существу обладающий активностью пируватдегидрогеназы, как аминокислотная последовательность, по существу демонстрирующая 70% или более гомологии с SEQ ID NO: 25, в частности 80% или более, более конкретно 90% или более, еще более конкретно 95% или более и даже еще более конкретно 99% или более, может быть включен в объем настоящего изобретения без ограничения.
Дополнительно ген, кодирующий пируватдегидрогеназу, может иметь нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO: 25. Аминокислотные последовательности белка, демонстрирующего эту активность, различаются в зависимости от вида или штамма микроорганизма, или множество нуклеиновых кислот, имеющих идентичную функцию, может кодировать любые заданные белки вследствие вырожденности генетического кода, и соответственно, ген не ограничен указанной SEQ ID NO. Например, ген может иметь нуклеотидную последовательность SEQ ID NO: 26, или нуклеотидные последовательности, гомологичные им на 80%, в частности на 90% или более, и более конкретно на 99% или более.
Термин "инактивация" в настоящем изобретении относится к случаям, когда ген, кодирующий релевантный полипептид, не экспрессируется, проявляет снижение активности вследствие неполной экспрессии или не продуцирует релевантный функциональный полипептид, даже несмотря на то, что ген экспрессируется.
Дополнительно, этот термин относится к случаю, когда ген, кодирующий релевантный полипептид, по существу не экспрессируется вследствие очень низкого уровня экспрессии, а также в случае, когда ген полностью инактивирован. Соответственно, инактивация гена может быть полной (нокаут) или частичной (например, ген может представлять собой гипоморф (hypomorph), экспрессирующий продукт на уровне ниже нормы или экспрессирующий продукт мутантного гена, демонстрирущего частичное снижение активности, связанное с геном).
Кроме того, "инактивация" релевантного полипептида по настоящему изобретению включает случаи элиминации активности полипептида, а также ее уменьшение по сравнению с немодифицированным штаммом. В частности, инактивация треониндегидратазы в настоящем изобретении может быть выполнена способом, выбранным из группы, состоящей из:
1) частичной или полной делеции полинуклеотида, кодирующего белок;
2) модификации последовательности регуляции экспрессии для уменьшения экспрессии полинуклеотида;
3) модификации полинуклеотидной последовательности в хромосоме для ослабления активности белка; или
4) их комбинации; однако способ специально не ограничен этим.
Способ частичной или полной делеции полинуклеотида, кодирующего белок, может быть осуществлен путем замены полинуклеотида, кодирующего эндогенный белок-мишень на хромосоме, на ген-маркер или полинуклеотид, в котором последовательность нуклеиновой кислоты была частично делетирована, с помощью вектора для вставки в хромосому бактерии. Как его используют здесь, термин "частично" может варьировать в зависимости от типов полинуклеотида, но может, в частности, относиться к фрагменту от 1 до 300 нуклеотидов, более конкретно от 1 до 100 нуклеотидов и еще более конкретно от 1 до 50 нуклеотидов.
Способ модификации последовательности регуляции экспрессии для уменьшения экспрессии полинуклеотида может быть осуществлен путем индуцирования мутации в последовательности посредством делеции, вставки или неконсервативной или консервативной замены в последовательности нуклеиновой кислоты, или их комбинации, так что активность последовательности регуляции экспрессии может быть снижена, или путем замены последовательности на последовательность нуклеиновой кислоты, обладающую пониженной активностью; однако способ специально не ограничен этим. Последовательность регуляции экспрессии включает промотор, операторную последовательность, последовательность, кодирующую сайт связывания с рибосомой, последовательность, регулирующую терминацию транскрипции и трансляции или им подобное; но не ограничиваясь этим.
Кроме того, способ модификации полинуклеотидной последовательности в хромосоме может быть осуществлен путем индуцирования мутации в последовательности посредством делеции, вставки или неконсервативной или консервативной замены полинуклеотидной последовательности, или их комбинации, или путем замены последовательности на полинуклеотидную последовательность, модифицированную для снижения активности; однако способ не ограничен этим.
В другом конкретном воплощении настоящего изобретения любой микроорганизм может быть включен в объем настоящего изобретения в качестве микроорганизма для получения L-изолейцина без ограничения, при условии что в этом микроорганизме может осуществляться экспрессия путем введения белка, обладающего активностью цитрамалатсинтазы. Примеры микроорганизма могут представлять собой микроорганизм рода Escherichia, Shigella, Citrobacter, Salmonella, Enterobacter, Yersinia, Klebsiella, Erwinia, Corynebacterium, Brevibacterium, Lactobacillus, Selenomanas, Vibrio, Pseudomonas, Streptomyces, Arcanobacterium, Alcaligenes или им подобного, и в частности рода Corynebacterium, более конкретно Corynebacterium glutamicum, но не ограничиваясь этим.
Еще в одном аспекте настоящего изобретения предложен способ получения L-изолейцина, включающий:
культивирование микроорганизма рода Corynebacterium, содержащего белок, обладающий активностью цитрамалатсинтазы, в среде; и
выделение L-изолейцина из микроорганизма или среды,
где микроорганизм рода Corynebacterium, имеющий введенный ген, кодирующий белок, обладающий активностью цитрамалатсинтазы, является таким как описано выше.
Кратко, микроорганизм рода Corynebacterium может иметь введенный ген, кодирующий цитрамалатсинтазу, имеющий происхождение из Methanocaldococcus, где цитрамалатсинтаза может, в частности, иметь аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, 3, 5, 7, 9, 11, 13 и 15.
Дополнительно, микроорганизм рода Corynebacterium по настоящему изобретению может дополнительно включать повышенные активности 3-изопропилмалатдегидрогеназы и 3-изопропилмалатдегидратазы. Кроме того, микроорганизм рода Corynebacterium может дополнительно включать инактивированную пируватдегидрогеназу.
В частности, микроорганизм рода Corynebacterium в примере настоящего изобретения может представлять собой Corynebacterium glutamicum, имеющий введенный ген, кодирующий цитрамалатсинтазу.
В данном способе культивирование микроорганизма рода Corynebacterium по настоящему изобретению может быть выполнено путем хорошо известного периодического культивирования, непрерывного культивирования и периодического культивирования с подпиткой, но специально не ограничиваясь этим.
В частности, для создания условий культивирования подходящий рН (например рН от 5 до 9, и в частности, рН от 6 до 8) может быть отрегулирован с использованием основного соединения (например гидроксида натрия, гидроксида калия или аммиака) или кислотного соединения (например фосфорной кислоты или серной кислоты), но специально не ограничиваясь этим. Дополнительно, образование пузырьков можно подавлять с помощью пеногасителя, такого как полигликолевый сложный эфир жирной кислоты.
Кислород или кислород-содержащую смесь газов можно вводить в культуру для поддержания аэробных условий, и температура культуры обычно составляет от 20°С до 45°С, в частности от 25°С до 40°С. Культивирование продолжают до получения максимального количества продуцированного L-изолейцина, и обычно достижение этого может занимать от 10 часов до 160 часов. L-изолейцин может секретироваться в среду или может оставаться в клетках.
Кроме того, в качестве источника углерода для используемой культуральной среды сахара и углеводы (например глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу), масла и жиры (например соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирные кислоты (например пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту), спирты (например глицерин и этанол), органические кислоты (например уксусную кислоту) или им подобное можно использовать индивидуально или в комбинации, но источник углерода не ограничен ими.
В качестве источника азота азот-содержащие органические соединения (например пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, кукурузный осадок, порошок из жмыха соевых бобов и мочевину), неорганические соединения (например сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония) или им подобные можно использовать по отдельности или в комбинации, но источник азота не ограничен ими.
В качестве источника фосфора дигидрофосфат калия, гидроортофосфат калия, соответствующие им натрийсодержащие соли или им подобное можно использовать по отдельности или в комбинации, но источник фосфора не ограничен ими.
В конкретном воплощении настоящего изобретения при культивировании микроорганизма рода Corynebacterium в способе получения L-изолейцина может быть дополнительно предоставлен ацетат.
В способе выделения L-изолейцина, продуцируемого путем культивирования микроорганизма в данном способе, для выделения целевой аминокислоты из культуральной жидкости можно использовать подходящий метод в зависимости от способа культивирования, известного в уровне техники, например, периодического культивирования, непрерывного культивирования, периодического культивирования с подпиткой и так далее. Например, можно использовать центрифугирование, фильтрование, анионообменную хроматографию, кристаллизацию, высокоэффективную хроматографию (ВЭЖХ) или им подобное, но способ выделения L-изолейцина не ограничен ими.
В еще одном аспекте настоящего изобретения предложено применение микроорганизма рода Corynebacterium, содержащего белок, обладающий активностью цитрамалатсинтазы, для получения L-изолейцина.
В этом применении микроорганизм рода Corynebacterium, имеющий введенный ген, кодирующий белок, обладающий активностью цитрамалатсинтазы, является таким как описано выше.
Как описано выше, микроорганизм рода Corynebacterium для получения L-изолейцина по настоящему изобретению может продуцировать L-изолейцин с высоким выходом посредством нового пути биосинтеза, в котором не используется L-треонин в качестве предшественника, ввиду наличия введенной активности цитрамалатсинтазы.
ПРИМЕРЫ
Пример 1: Конструирование рекомбинантного вектора, включающего ген cimA, имеющий происхождение из Methanocaldococcus
1-1. Получение фрагмента гена cimA, имеющего происхождение из Methanocaldococcus
Для получения фрагмента 1476 п. н., содержащего открытую рамку считывания (ORF) гена cimA (NC_000909.1), кодирующего цитрамалатсинтазу, геномную ДНК Methanocaldococcus jannaschii DSM 2661 выделяли с использованием системы Genomic-tip (Qiagen).
Цитрамалатсинтаза, имеющая происхождение из Methanocaldococcus jannaschii DSM 2661, имеет последовательность из 491 аминокислоты, представленную SEQ ID NO: 1, и ген cimA, кодирующий эту последовательность, имеет нуклеотидную последовательность SEQ ID NO: 2.
Полимеразную цепную реакцию (здесь и далее сокращенно обозначено "ПЦР") проводили с использованием выделенной геномной ДНК (гДНК) в качестве матрицы. ПЦР проводили с использованием пары праймеров с SEQ ID NO: 35 и 36 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 60 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем полосу, соответствующую желаемому размеру, элюировали для выделения и продукт обозначали как "фрагмент cimA(M)".

1-2. Конструирование рекомбинантного вектора p117-cj7-cimA(M)
ПЦР проводили с использованием p117-cj7-gfp, включающего промотор cj7, имеющий происхождение из известного микроорганизма рода Corynebacterium (патент Кореи No. 10-0620092) в качестве матрицы. Как его используют здесь, термин "р117" представляет собой pECCG117, который является Е. coli-Corynebacterium шаттл-вектором (Biotechnology Letters 13(10): 721-726, 1991). ПЦР проводили с использованием пары праймеров SEQ ID NO: 27 и 28 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 20 сек и элонгации при 72°С в течение 30 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем полосу 323 п. н. элюировали для выделения и продукт обозначали как "фрагмент cj7".

Сшивающую ПЦР проводили с использованием полученного фрагмента cj7 и фрагмента cimA(M), полученного в Примере 1-1, изложенном выше, в качестве матрицы. Сшивающую ПЦР проводили с использованием пары праймеров SEQ ID NO: 27 и 36 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 60 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем полосу 1799 п. н. элюировали для выделения и продукт обозначали как "фрагмент cj7-cimA(M)".
Полученный фрагмент cj7-cimA(M) обрабатывали ферментами рестрикции KpnI и XbaI и затем лигировали с линейным фрагментом р117, обработанным теми же ферментами рестрикции.
Клетки Е. coli DH5α трансформировали сконструированным вектором методом теплового шока, затем высевали на твердую среду LB, содержащую 25 мкг/мл канамицина, и культивировали в течение ночи при 37°С. Одну платиновую петлю с культивированной колонией инокулировали в 3 мл жидкой среды LB, содержащей 25 мкг/мл канамицина, и культивировали в течение ночи, и затем плазмидную ДНК выделяли с использованием набора для выделения плазмид "miniprep" (в каталоге No. 27104, Qiagen, здесь и далее так же).
Конструкцию рекомбинантного вектора подтверждали обработкой ферментами рестрикции KpnI и XbaI, и клон идентифицировали путем проведения ПЦР с использованием пары праймеров с SEQ ID NO: 29 и 30 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 90 сек. Выделенный рекомбинантный вектор обозначали как "p117-cj7-cimA(M)".

1-3. Получение фрагмента гена leuBCD, вовлеченного в путь биосинтеза 2-кетобутирата
Для получения рекомбинантного вектора для усиления гена в пути биосинтеза 2-кетобутирата фрагмент гена leuBCD, кодирующего 3-изопропилмалатдегидрогеназу и 3-изопропилмалатдегидратазу, имеющего происхождение из микроорганизма рода Corynebacterium, получали как описано ниже.
Для получения фрагмента 1359 п. н., содержащего ORF гена leuB, кодирующего 3-изопропилмалатдегидрогеназу (ЕС1.1.1.85), геномную ДНК Corynebacterium glutamicum АТСС13032 выделяли с использованием системы "Genomic-tip" (Qiagen).
3-Изопропилмалатдегидрогеназа, имеющая происхождение из Corynebacterium glutamicum АТСС13032, имеет последовательность из 340 аминокислот, представленных в SEQ ID NO: 17, и ген leuB, кодирующий эту последовательность, имеет нуклеотидную последовательность SEQ ID NO: 18.
ПЦР проводили с использованием выделенной гДНК в качестве матрицы. ПЦР проводили с использованием пары праймеров SEQ ID NO: 31 и 32 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 60 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем полосу, соответствующую желаемому размеру, элюировали для выделения и продукт обозначали как "фрагмент leuB".

Для получения фрагмента 2078 п. н., содержащего ORF гена leuCD, кодирующего 3-изопропилмалатдегидратазу (ЕС4.2.1.33), проводили ПЦР с использованием гДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы.
Ген LeuC кодирует большую единицу 3-изопропилмалатдегидратазы, и ген leuC, имеющий происхождение из Corynebacterium glutamicum АТСС13032, имеет нуклеотидную последовательность SEQ ID NO: 20, кодирующую полипептид, имеющий аминокислотную последовательность SEQ ID NO: 19. Кроме того, ген leuD кодирует малую субъединицу 3-изопропилмалатдегидратазы, и ген leuD, имеющий происхождение из Corynebacterium glutamicum АТСС13032, имеет нуклеотидную последовательность SEQ ID NO: 22, кодирующую полипептид, имеющий аминокислотную последовательность SEQ ID NO: 21.
ПЦР проводили с использованием пары праймеров SEQ ID NO: 33 и 34 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 80 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем полосу, соответствующую желаемому размеру, элюировали для выделения и продукт обозначали как "фрагмент leuCD".

Сшивающую ПЦР проводили с использованием фрагмента leuB и фрагмента leuCD, выделенных как описано выше, в качестве матрицы. Сшивающую ПЦР выполняли с использованием пары праймеров SEQ ID NO: 31 и 34 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 120 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем полосу 3437 п. н. элюировали для выделения и продукт обозначали как "фрагмент leuBCD".
1-4. Конструирование рекомбинантного вектора p117-cj7-cimA(M)-leuBCD
Для получения рекомбинантного вектора для усиления гена в пути биосинтеза 2-кетобутирата p117-cj7-cimA(M), который представляет собой рекомбинантный вектор, полученный в Примере 1-2 выше, обрабатывали ферментом рестрикции XbaI, фрагмент leuBCD, выделенный в Примере 1-3 выше, обрабатывали ферментом рестрикции XbaI, и затем оба фрагмента лигировали.
Клетки Е. coli DH5α трансформировали сконструированным вектором методом теплового шока, затем высевали на твердую среду LB, содержащую 25 мкг/мл канамицина, и культивировали в течение ночи при 37°С. Одну платиновую петлю с культивированной колонией инокулировали в 3 мл жидкой среды LB, содержащей 25 мкг/мл канамицина, и культивировали в течение ночи, и затем плазмидную ДНК выделяли с использованием набора для выделения плазмид "miniprep".
Конструкцию рекомбинантного вектора подтверждали путем обработки ферментом рестрикции XbaI, и клон идентифицировали путем осуществления ПЦР с использованием пары праймеров SEQ ID NO: 29 и 30 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 180 сек. Выделенный рекомбинантный вектор обозначали как "p117-cj7-cimA(M)-leuBCD".
1-5. Получение фрагмента мутанта cimA(M) с использованием подверженной ошибкам ПЦР
Чтобы обеспечить пул ДНК, имеющих случайную мутацию, введенную в ген cimA, проводили ПЦР с использованием фрагмента cimA(M), выделенного в Примере 1-1 выше, в качестве матрицы и набора для случайного мутагенеза методом ПЦР "diversify" (в каталоге No. 630703, Clonetech). ПЦР проводили в условиях 4 для реакции мутагенеза, изложенных в Таблице III в руководстве пользователя продукта, с использованием пары праймеров SEQ ID NO: 35 и 36 в следующих условиях: 25 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек и элонгации при 68°С в течение 30 сек.
В результате ПЦР был получен мутированный art cimA пул ДНК, имеющих модификацию путем введенных случайным образом нуклеотидных замен. Продукт ПЦР (обозначенный здесь и далее как "фрагмент cimA(M)m") подвергали электрофорезу в 1,0% агарозном геле и затем элюировали для выделения.

1- 6. Получение библиотеки мутантов p117-cj7-cimA(M)m
Рекомбинантный вектор p117-cj7-cimA(M)m, полученный в Примере 1-2 выше, обрабатывали ферментами рестрикции NdeI и XbaI, и лигировали вместе с фрагментом cimA(M)m, полученным в Примере 1-5 выше, и обрабатывали теми же ферментами рестрикции.
Клетки Е. coli DH5α трансформировали сконструированным вектором методом теплового шока, затем высевали на твердую среду LB, содержащую 25 мкг/мл канамицина, и культивировали в течение ночи 37°С. Культивированные колонии собирали и затем плазмидную ДНК выделяли с использованием набора для выделения плазмид "miniprep", тем самым получая "библиотеку мутантов p117-cj7-cimA(M)m".
Пример 2: Подтверждение L-изолейцин-продуцирующей способности при введении гена cimA
Для подтверждения того, может ли Corynebacterium glutamicum иметь L-изолейцин-продуцирующую способность при введении в него гена cimA, имеющего происхождение из Methanocaldococcus, сконструировали штамм, из которого был удален ген ilvA, кодирующий треониндегидратазу, и восстановление L-изолейцин-продуцирующей способности штамма изучали путем введения в него гена cimA.
Треониндегидратаза представляет собой фермент, превращающий L-треонин, являющийся предшественником в биосинтезе L-изолейцина, в 2-кетобутират. Треониндегидратаза, имеющая происхождение из Corynebacterium glutamicum АТСС13032, имеет аминокислотную последовательность SEQ ID NO: 23, и ген ilvA, кодирующий эту последовательность, имеет нуклеотидную последовательность SEQ ID NO: 24. Кроме того, любой белок, обладающий активностью треониндегидратазы, включен в объем настоящего изобретения без ограничения.
2-1. Конструирование вектора с делецией гена ilvA
Для получения фрагмента гена ilvA проводили ПЦР с использованием геномной ДНК, выделенной из Corynebacterium glutamicum АТСС13032 с использованием системы Genomic-tip (Qiagen), в качестве матрицы. ПЦР проводили с использованием каждой пары праймеров SEQ ID NO: 37 и 38 и SEQ ID NO: 39 и 40 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 45 сек.
Амплифицированные ПЦР-продукты подвергали электрофорезу в 1,0% агарозном геле и затем полосы 547 п. н. и 467 п. н. для каждой реакции ПЦР элюировали для выделения.
Сшивающую ПЦР проводили с использованием двух фрагментов гена ilvA, полученных как описано выше, в качестве матрицы. Сшивающую ПЦР проводили с использованием пары праймеров SEQ ID NO: 37 и 40 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 60 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем 1014 п. н. полосу элюировали для выделения и выделенный фрагмент с делецией гена ilvA обозначали как "DilvA".
Фрагмент DilvA, полученный как описано выше, обрабатывали ферментом рестрикции XbaI, вектор pDZ (патент Кореи No. 10-0924065) обрабатывали тем же ферментом рестрикции для получения линейного фрагмента pDZ и затем фрагменты DilvA и линейный pDZ лигировали для конструирования рекомбинантного вектора, имеющего делецию гена ilvA.
Клетки Е. coli DH5α трансформировали сконструированным рекомбинантным вектором методом теплового шока, затем высевали на твердую среду LB, содержащую 25 мкг/мл канамицина, и культивировали в течение ночи при 37°С. Одну платиновую петлю с культивированной колонией инокулировали в 3 мл жидкой среды LB, содержащей 25 мкг/мл канамицина, и культивировали в течение ночи, и затем выделяли плазмидную ДНК с использованием набора для выделения плазмид "miniprep".
Конструкцию рекомбинантного вектора подтверждали путем обработки ферментом рестрикции XbaI и клон идентифицировали путем проведения ПЦР с использованием пары праймеров SEQ ID NO: 41 и 42 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 90 сек. Выделенный рекомбинантный вектор обозначали как "pDZ-ilvA(Del)".

2-2. Получение штамма с делецией гена ilvA
Рекомбинантный вектор pDZ-ilvA(Del), сконструированный в Примере 2-1 выше, вводили в Corynebacterium glutamicum АТСС13032 в качестве родительского штамма путем электропорации, и высевали на твердую среду, содержащую 25 мкг/мл канамицина, для отбора отдельной колонии. Из этой колонии каждый штамм, имеющий вектор pDZ-ilvA(Del), введенный в хромосому, отбирали путем второго пересева и затем проводили ПЦР с использованием гДНК, выделенной из каждого из выбранных штаммов, в качестве матрицы. ПЦР проводили с использованием пары праймеров SEQ ID NO: 43 и 44 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 90 сек.
Как показали результаты ПЦР, был получен штамм с делецией гена ilvA, который обозначали как штамм "АТСС13032ΔilvA".

2-3. Получение фрагмента гена, вовлеченного в путь биосинтеза L-изолейцина
Для получения рекомбинантного вектора для усиления гена в пути биосинтеза L-изолейцина, фрагменты генов lysC, asd, hom, thrB, thrC и ilvA, имеющие происхождение из микроорганизма рода Corynebacterium, получали как описано ниже.
Во-первых, для получения фрагмента ДНК, содержащего ORF гена lysC-asd, кодирующего аспартаткиназу и аспартат-β-полуальдегид-дегидрогеназу, проводили ПЦР с использованием геномной ДНК, выделенной из Corynebacterium glutamicum АТСС13032, в качестве матрицы как описано в Примере 2-1. ПЦР проводили с использованием пары праймеров SEQ ID NO: 45 и 46 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 90 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем 2666 п. н. полосу элюировали для выделения и продукт обозначали как "фрагмент lysC-asd".

Для получения фрагмента ДНК, содержащего ORF гена hom-thrB, кодирующего гомосериндегидрогеназу и гомосеринкиназу, проводили ПЦР с использованием геномной ДНК, выделенной из Corynebacterium glutamicum АТСС13032, в качестве матрицы, как описано в Примере 2-1. ПЦР проводили с использованием пары праймеров SEQ ID NO: 47 и 48 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 90 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем 2633 п. н. полосу элюировали для выделения и продукт обозначали как "фрагмент hom-thrB".

Для получения фрагмента ДНК, содержащего ORF гена thrC, кодирующего треонин-синтазу, проводили ПЦР с использованием геномной ДНК, экстрагированной из Corynebacterium glutamicum АТСС13032, в качестве матрицы, как описано в Примере 2-1. ПЦР проводили с использованием пары праймеров с SEQ ID NO: 49 и 50 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 60 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем 1703 п. н. полосу элюировали для выделения и продукт обозначали как "фрагмент thrC".

Для получения фрагмента ДНК, содержащего ORP гена ilvA, кодирующего треониндегидратазу, проводили ПЦР с использованием геномной ДНК, выделенной из Corynebacterium glutamicum АТСС13032, в качестве матрицы, как описано в Примере 2-1. ПЦР проводили с использованием пары праймеров SEQ ID NO: 51 и 52 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 60 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем 1862 п. н. полосу элюировали для выделения и продукт обозначали как "фрагмент ilvA".

Сшивающую ПЦР проводили с использованием фрагмента thrC и фрагмента ilvA, полученных как описано выше, в качестве матрицы. Сшивающую ПЦР проводили и с использованием пары праймеров SEQ ID NO: 49 и 52 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 120 сек.
Амплифицированный ПЦР-продукт подвергали электрофорезу в 1,0% агарозном геле, затем 3565 п. н. полосу элюировали для выделения и продукт обозначали как "фрагмент thrC-ilvA".
2-4. Конструирование вектора для усиления гена в пути биосинтеза L-изолейцина
Для получения рекомбинантного вектора для усиления гена в пути биосинтеза L-изолейцина фрагмент lysC-asd, полученный в Примере 2-3 выше, обрабатывали ферментами рестрикции KpnI и SfoI и затем лигировали с линейным фрагментом р117, обработанным ферментами рестрикции KpnI и EcoRV.
Плазмидную ДНК выделяли таким же образом, как в Примере 1-2 выше. Для подтверждения конструкции рекомбинантного вектора клон идентифицировали путем проведения ПЦР с использованием пары праймеров SEQ ID NO: 29 и 30 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 100 сек. Полученный рекомбинантный вектор обозначали как "p117-lysC-asd".
Фрагмент hom-thrB, полученный в Примере 2-3 выше, обрабатывали ферментом рестрикции BamHI и затем лигировали с линейным фрагментом p117-lysC-asd, обработанным тем же ферментом рестрикции.
Плазмидную ДНК выделяли таким же образом, как в Примере 1-2 выше. Для подтверждения конструкции рекомбинантного вектора клон идентифицировали путем проведения ПЦР с использованием пары праймеров с SEQ ID NO: 29 и 30 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 180 сек. Выделенный рекомбинантный вектор обозначали как "p117-lysC-asd-hom-thrB".
Фрагмент thrC-ilvA, полученный в Примере 2-3 выше, обрабатывали ферментом рестрикции SpeI и затем лигировали с линейным фрагментом p117-lysC-asd-hom-thrB, обработанным тем же ферментом рестрикции.
Плазмидную ДНК выделяли таким же образом, как в Примере 1-2 выше. Для подтверждения конструкции рекомбинантного вектора клон идентифицировали путем проведения ПЦР с использованием пары праймеров с SEQ ID NO: 29 и 30 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 300 сек.
Полученный рекомбинантный вектор обозначали как "p117-IBGC (Isoleucine Biosynthesis Genes Cluster)", который представляет собой рекомбинантный вектор для усиления гена в пути биосинтеза L-изолейцина, включающий фрагменты генов lysC, asd, hom, thrB, thrC и ilvA, шести генов, вовлеченных в биосинтез L-изолейцина.
2-5. Сравнение продуктивности по L-изолейцину
Рекомбинантные векторы p117-cj7-cimA(M), p117-cj7-cimA(M)-leuBCD и p117-IBGC, сконструированные в Примерах 1-2, 1-4 и 2-4 выше, вводили в Corynebacterium glutamicum АТСС13032 путем электропорации и высевали на твердую среду, содержащую 25 мкг/мл канамицина для отбора отдельной колонии. Каждый из штаммов, отобранных из колонии, обозначали как Corynebacterium glutamicum "ATCC13032/p117-cj7-cimA(M)", "ATCC13032/p117-cj7-cimA(M)-leuBCD" и "ATCC13032/p117-IBGC".
Далее, p117-cj7-cimA(M) и p117-cj7-cimA(M)-leuBCD вводили в штамм Corynebacterium glutamicum ATCC13032ΔilvA, полученный в Примере 2-2 выше, путем электропорации и высевали на твердую среду, содержащую 25 мкг/мл канамицина, для отбора отдельной колонии. Каждый из штаммов, отобранных из колонии, обозначали как Corynebacterium glutamicum "ATCC13032ΔilvA/p117-cj7-cimA(M)" и "АТСС 13032ΔilvA/p117-cj7-cimA(M)-leuBCD".
Оценку титра продуктивности по L-изолейцину проводили с использованием Corynebacterium glutamicum АТСС 13032, ATCC13032/p117-IBGC, ATCC13032/p117-cj7-cimA(M), ATCC13032/p117-cj7-cimA(M)-leuBCD, ATCC13032ΔilvA, АТСС13032ΔilvA/p117-cj7-cimA(M) и ATCC13032ΔilvA/p117-cj7-cimA(M)-leuBCD.
Более подробно, каждый из штаммов инокулировали в колбу емкостью 250 мл, содержащую 25 мл глюкозосодержащей среды для титрования, имеющей состав, показанный в Таблице 1 ниже, и затем культивировали в инкубаторе при 32°С и 200 об/мин в течение 30 часов. Концентрации продуцированного L-изолейцина представлены в Таблице 2 ниже.


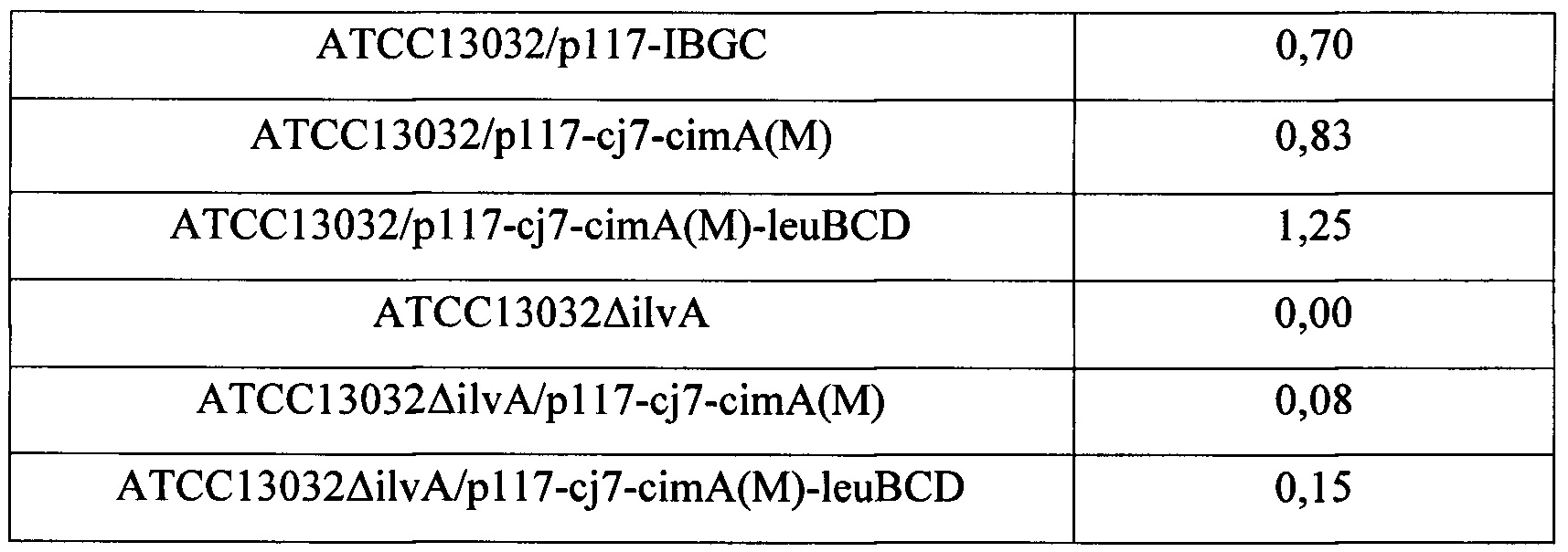
Как показано в Таблице 2 выше, штамм АТСС13032 дикого типа, имеющий ген ilvA, продуцировал 0,02 г/л L-изолейцина, a ATCC13032/p117-IBGC, трансформированный штамм, имеющий одновременно введенные гены пути биосинтеза L-изолейцина, продуцировал 0,70 г/л L-изолейцина, демонстрируя, что продуктивность по L-изолейцину увеличилась на 3400% по сравнению со штаммом дикого типа.
Кроме того, трансформированные штаммы ATCC13032/p117-cj7-cimA(M) и ATCC13032/p117-cj7-cimA(M)-leuBCD продуцировали 0,83 г/л и 1,25 г/л L-изолейцина, соответственно, демонстрируя, что продуктивность по L-изолейцину увеличилась на 4050% и 6150%, соответственно, по сравнению со штаммом дикого типа.
Результаты этого сравнения служат подтверждением того, что продуктивность по L-изолейцину ATCC13032/p117-cj7-cimA(M), штамма, трансформированного для введения гена cimA в существующий путь биосинтеза L-изолейцина, превосходит таковую для ATCC13032/p117-IBGC, штамма, трансформированного для усиления пути биосинтеза самого L-изолейцина. Более того, было подтверждено, что продуктивность по L-изолейцину ATCC13032/p117-cj7-cimA(M)-leuBCD, трансформированного штамма, дополнительно усиленного геном leuBCD, была дополнительно улучшена.
Между тем, штамм ATCC13032ΔilvA с делецией гена ilvA потерял продуктивность по L-изолейцину, в то время как штамм ATCC13032ΔilvA/p117-cj7-cimA(M), имеющий введенный ген cimA, продуцировал 0,08 г/л L-изолейцина, а штамм ATCC13032ΔilvA/p117-cj7-cimA(M)-leuBCD, имеющий введенный ген cimA и усиленный ген leuBCD, продуцировал 0,15 г/л L-изолейцина.
Вышеизложенные результаты показывают, что продуцирование L-изолейцина возможно с использованием нового пути биосинтеза L-изолейцина путем введения гена cimA, а не существующего пути биосинтеза L-изолейцина, использующего L-треонин в качестве предшественника.
Пример 3: Получение трансформированного рекомбинантного штамма и сравнение продуктивности по L-изолейцину
3-1. Получение рекомбинантного штамма с использованием микроорганизма рода Corynebacterium дикого типа
Для обеспечения ацетил-СоА, используемого в качестве предшественника гена cimA, был получен ауксотрофный по ацетату штамм дикого типа Corynebacterium glutamicum АТСС13032 с делецией гена асеЕ, кодирующего пируватдегидрогеназу, и он был обозначен как "АТСС13032ΔасеЕ" (Schreiner et al., J Bacteriol. 187: 6005-18, 2005).
Пируватдегидрогеназа, имеющая происхождение из Corynebacterium glutamicum АТСС13032, имеет аминокислотную последовательность SEQ ID NO: 25, и ген асеЕ, кодирующий ее, имеет нуклеотидную последовательность SEQ ID NO: 26.
Рекомбинантный вектор p117-cj7-cimA(M) и библиотеку мутантов p117-cj7-cimA(M)m, полученные в Примере 1 выше, вводили в штамм АТСС13032ΔасеЕ, полученный как изложено выше, путем электропорации. Полученные штаммы обозначали как "ATCC13032ΔaceE/p117-cj7-cimA(M)" и "библиотека мутантов ATCC13032ΔaceE/p117-cj7-cimA(M)m", соответственно.
АТСС13032ΔасеЕ, АТСС13032ΔасеЕ/р117-cj7-cimA(M) и библиотеку мутантов АТСС13032ΔасеЕ/p117-cj7-cimA(M)m высевали на твердую среду, содержащую 25 мкг/мл канамицина для отбора отдельной колонии, и выполняли оценку титра по продуктивности по L-изолейцину с использованием выбранных штаммов.
Более конкретно, штаммы инокулировали в колбы емкостью 250 мл, содержащие 25 мл среды для титрования, содержащей ацетат в форме ацетата аммония, имеющей состав, показанный в Таблице 3 ниже, и затем культивировали в инкубаторе при 32°С и 200 об/мин в течение 30 часов. В результате были отобраны семь типов колоний, имеющих повышенную концентрацию L-изолейцина, и их перечень и концентрации L-изолейцина представлены в Таблице 4 ниже.


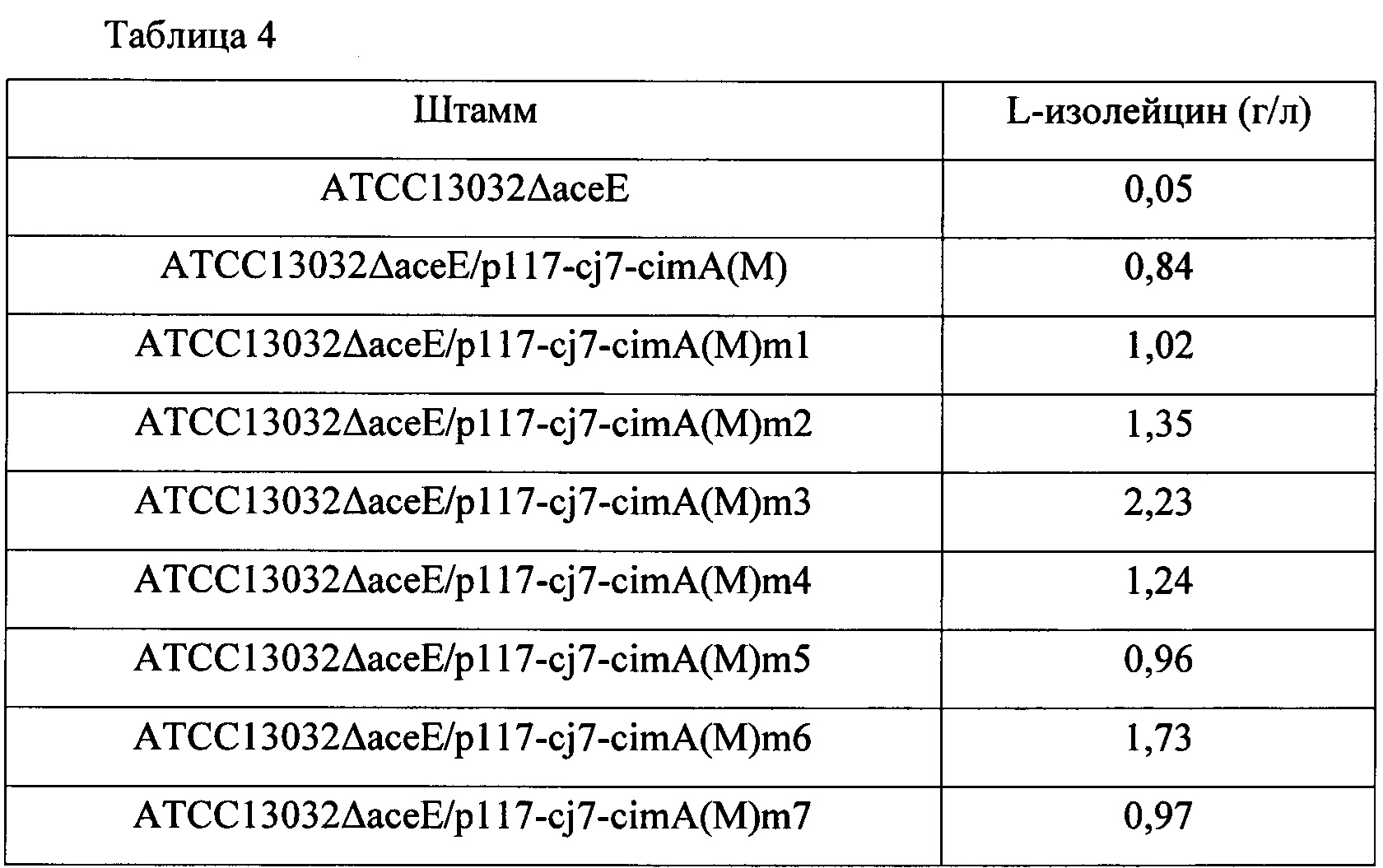
В результате, как показано в Таблице 4 выше, штамм АТСС13032ΔасеЕ как родительский штамм продуцировал 0,05 г/л L-изолейцина, а трансформированные штаммы продуцировали от 0,84 г/л до 2,23 г/л L-изолейцина, демонстрируя, что увеличение продуктивности по L-изолейцину составляло от 1580% до 4360%, соответственно, по сравнению с родительским штаммом. В частности, для трансформированного штамма ATCC13032ΔaceE/p117-cj7-cimA(M)m3 была зарегистрирована самая высокая продуктивность по L-изолейцину среди этих штаммов по сравнению с родительским штаммом АТСС13032ΔасеЕ.
Для выявления сайта мутации у мутантов по гену cimA(M), отобранных по увеличенной продуктивности по L-изолейцину, показанной в Таблице 4, и замещенных аминокислот при каждой мутации, было выполнено секвенирование.
В результате, как библиотека мутантов CimA(M)m,
CimA(M)m1 имеет аминокислотную последовательность SEQ ID NO: 3 и может кодироваться полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 4;
CimA(M)m2 имеет аминокислотную последовательность SEQ ID NO: 5 и может кодироваться полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 6;
CimA(M)m3 имеет аминокислотную последовательность SEQ ID NO: 7 и может кодироваться полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 8;
CimA(M)m4 имеет аминокислотную последовательность SEQ ID NO: 9 и может кодироваться полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 10;
CimA(M)m5 имеет аминокислотную последовательность SEQ ID NO: 11 и может кодироваться полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 12;
CimA(M)m6 имеет аминокислотную последовательность SEQ ID NO: 13 и может кодироваться полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 14, и
CimA(M)m7 имеет аминокислотную последовательность SEQ ID NO: 15 и может кодироваться полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 16.
3-2. Получение рекомбинантного штамма с использованием p117-cj7-cimA(M)m3, имеющего самую высокую активность выбранной библиотеки мутантов p117-cimA(M)m
Вектор p117-cj7-cimA(M)m3, идентифицированный как имеющий наиболее высокую продуктивность по L-изолейцину в Примере 3-1 выше, обрабатывали XbaI и лигировали с фрагментом ДНК, выделенным путем обработки фрагмента leuBCD, выделенного в Примере 1-3 выше, тем же ферментом рестрикции.
Клетки Е. coli DH5α трансформировали сконструированным вектором методом теплового шока, затем высевали на твердую среду LB, содержащую 25 мкг/мл канамицина, и культивировали в течение ночи при 37°С. Одну платиновую петлю с культивированной колонией инокулировали в 3 мл жидкой среды LB, содержащей 25 мкг/мл канамицина, и культивировали в течение ночи и затем выделяли плазмидную ДНК с использованием набора для выделения плазмид "miniprep".
Конструкцию рекомбинантного вектора подтверждали путем обработки ферментом рестрикции XbaI и клон идентифицировали путем проведения ПЦР с использованием пары праймеров SEQ ID NO: 29 и 30 в следующих условиях: 30 циклов, каждый из которых состоял из денатурации при 95°С в течение 30 сек, отжига при 56°С в течение 30 сек и элонгации при 72°С в течение 180 сек. Полученный рекомбинантный вектор обозначали как "p117-cj7-cimA(M)m3-leuBCD".
Каждый из 17-cj7-cimA(M) из Примера 1-2, p117-cj7-cimA(M)-leuBCD из Примера 1-4, p117-cj7-cimA(M)m3 из Примера 3-1 и p117-cj7-cimA(M)m3-leuBCD из Примера 3-2, изложенных выше, вводили в штамм АТСС13032ΔасеЕ, использованный в Примере 3-1, путем электропорации и высевали на твердую среду, содержащую 25 мкг/мл канамицина, для отбора отдельной колонии.
Выбранные штаммы инокулировали в колбы на 250 мл, содержащие 25 мл среды для титрования, содержащей глюкозу, имеющей состав, показанный в Таблице 3 выше, и затем культивировали в инкубаторе при 32°С и 200 об/мин в течение 30 часов для исследования продуктивности по L-изолейцину. Результаты показаны в Таблице 5 ниже.

Как показано в Таблице 5 выше, в то время как штамм АТСС13032ΔасеЕ как родительский штамм продуцировал 0,04 г/л L-изолейцина, трансформированный штамм ATCC13032ΔaceE/p117-cj7-cimA(M) продуцировал 0,98 г/л L-изолейцина, а ATCC13032ΔaceE/p117-cj7-cimA(M)-leuBCD продуцировал 2,09 г/л L-изолейцина, демонстрируя увеличение на 0,94 г/л и 2,05 г/л по сравнению с продуктивностью по L-изолейцину родительского штамма.
Далее, трансформированные штаммы ATCC13032ΔaceE/p117-cj7-cimA(M)m3 и ATCC13032ΔaceE/p117-cj7-cimA(M)m3-leuBCD, включающие мутантный ген cimA(M)m3, полученные в Примере 3-1, продуцировали 2,22 г/л и 3,76 г/л L-изолейцина, соответственно, что представляет собой увеличение продуцирования на 5450% и 9300% по сравнению с родительским штаммом. Соответственно, этот результат подтверждает, что мутант cimA(M)m3 является более эффективным, чем его штамм дикого типа, в увеличении продуктивности по L-изолейцину.
Из трансформированных штаммов, полученных в вышеизложенных Примерах, ATCC13032ΔaceE/p117-cj7-cimA(M)m3 обозначали как "Corynebacterium glutamicum СА10-1002", депонировали в Корейском Центре Культур Микроорганизмов (КССМ) согласно Будапештскому договору 27 февраля 2015 года с присвоением ему номера доступа No. КССМ11672Р.
3-3. Получение рекомбинантного штамма с использованием L-изолейцин-продуцирующего штамма КССМ11248Р
Каждый из рекомбинантных векторов p117-cj7-cimA(M), p117-cj7-cimA(M)-leuBCD, p117-cj7-cimA(M)m3 и p117-cj7-cimA(M)m3-leuBCD, используемых в Примерах 3-1 и 3-2, вводили в Corynebacterium glutamicum КССМ11248Р, L-изолейцин-продуцирующий штамм (патент Кореи No. 10-1335789) и высевали на твердую среду, содержащую 25 мкг/мл канамицина для отбора отдельной колонии.
Каждый из выбранных штаммов обозначали как KCCM11248P/p117-cj7-cimA(M), KCCM11248P/p117-cj7-cimA(M)-leuBCD, KCCM11248P/p117-cj7-cimA(M)m3 и KCCM11248P/p117-cj7-cimA(M)m3-leuBCD.
Продуктивность штаммов по L-изолейцину изучали с использованием среды для титрования L-изолейцина, имеющей состав, показанный в Таблице 6 ниже, и штаммы культивировали в колбах как описано ниже.


Более конкретно, родительский штамм КССМ11248Р и трансформированные штаммы KCCM11248P/p117-cj7-cimA(M), KCCM11248P/p117-cj7-cimA(M)-leuBCD, KCCM11248P/p117-cj7-cimA(M)m3 и KCCM11248P/p117-cj7-cimA(M)m3-leuBCD инокулировали в колбы на 250 мл, содержащие 25 мл среды для титрования, содержащей глюкозу, имеющей состав, показанный в Таблице 6, и затем культивировали в инкубаторе при 32°С и 200 об/мин в течение 60 часов для исследования продуктивности по L-изолейцину. Результаты показаны в Таблице 7 ниже.

Как показано в Таблице 7 выше, в то время как родительский штамм КССМ11248Р продуцировал 3,33 г/л L-изолейцина, трансформированные штаммы КССМ11248Р/р117-cj7-cimA(M) продуцировал 4,04 г/л L-изолейцина и KCCM11248P/p117-cj7-cimA(M)-leuBCD продуцировал 4,82 г/л L-изолейцина, демонстрируя увеличение на 0,71 г/л и 1,49 г/л по сравнению с продуктивностью по L-изолейцину родительского штамма.
Между тем, трансформированные штаммы KCCM11248P/p117-cj7-cimA(M)m3 и KCCM11248P/p117-cj7-cimA(M)m3-leuBCD продуцировали 4,71 г/л и 11,21 г/л L-изолейцина, соответственно, что свидетельствует об увеличении продуцирования на 41,4% и 236,6% по сравнению с родительским штаммом.
Как показано в результатах Примера 3-2 с использованием Corynebacterium glutamicum АТСС13032, этот результат дополнительно подтверждает, что мутант cimA(M)m3 является более эффективным, чем его штамм дикого типа в увеличении продуктивности по L-изолейцину.
Исходя из вышеизложенного, специалист в области техники, к которой принадлежит настоящее изобретение, может понять, что настоящее изобретение может быть воплощено в других конкретных формах без модификации технических идей или существенных характеристик настоящего изобретения. В этом отношении примеры воплощений, раскрытые здесь, предназначены исключительно для иллюстративных целей, и их не следует рассматривать как ограничивающие объем настоящего изобретения. Напротив, настоящее изобретение предназначено охватывать не только примеры воплощений, но также различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в идею и объем настоящего изобретения как определено в прилагаемой формуле изобретения.
Реферат
Группа изобретений относится к области биотехнологии. Предложен микроорганизм рода, обладающий L-изолейцин-продуцирующей способностью, где указанный микроорганизм содержит белок, обладающий активностью цитрамалатсинтазы. Предложен способ получения L-изолейцина с использованием микроорганизма рода, содержащего белок, обладающий активностью цитрамалатсинтазы. Группа изобретений позволяет увеличить продуктивность по L-изолейцину в модифицированном микроорганизме по сравнению с родительским немодифицированным микроорганизмом. 2 н. и 10 з.п. ф-лы, 7 табл., 3 пр.
Комментарии