Микроорганизм, продуцирующий о-ацетилгомосерин, и способ получения о-ацетилгомосерина с использованием этого микроорганизма - RU2614253C1
Код документа: RU2614253C1
Чертежи









































Описание
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к микроорганизму Escherichia sp., продуцирующему О-ацетилгомосерин и к способу получения О-ацетилгомосерина с высоким выходом с использованием этого микроорганизма.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
О-ацетилгомосерин действует как предшественник метионина, который является одной из незаменимых аминокислот в организме. Метионин широко используется в качестве компонента медицинских инфузионных растворов и сырья для медицинских продуктов, а также в качестве корма для животных и пищевой добавки.
Метионин может быть синтезирован биологически или химически. Недавно был описан двухстадийный способ, в котором предшественник L-метионина, полученный посредством ферментации, превращают в L-метионин посредством ферментативной реакции (международная публикация WO 2008/013432). В указанном выше двухстадийном способе О-сукцинил-гомосерин и О-ацетилгомосерин может быть использован в качестве предшественника метионина и для крупномасштабного высокорентабельного производства метионина важно получать О-ацетилгомосерин с высоким выходом
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая задача
Авторы настоящего изобретения, пытаясь улучшить получение О-ацетилгомосерина, обнаружили, что снижение экспрессии или активности белка цитратсинтазы может значительно увеличивать эффективность получения О-ацетилгомосерина, тем самым обеспечивая осуществление настоящего изобретения.
Техническое решение
Целью настоящего изобретения является создание О-ацетилгомосерин-продуцирующего микроорганизма с улучшенной способностью продуцировать О-ацетилгомосерин.
Другой целью настоящего изобретения является создание способа получения О-ацетилгомосерина с использованием этого микроорганизма.
Полезные эффекты
При использовании микроорганизма по настоящему изобретению, обладающего способностью продуцировать О-ацетилгомосерин, можно получать О-ацетилгомосерин с более высоким выходом и более экологически безопасным образом, чем химический синтез. Кроме того, О-ацетилгомосерин, полученный таким образом, можно использовать в качестве предшественника для синтеза метионина и уксусной кислоты при помощи О-ацетилгомосерин-сульфгидрилазы, тем самым обеспечивая возможность биоконверсии L-метионина, и L-метионин, преобразованный таким образом, можно широко использовать в производстве пищевых продуктов или пищевых добавок для людей, а также кормов для животных или пищевых добавок для животных.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 представлено изображение экспрессионной кассеты для конструирования микроорганизма с ослабленной активностью цитратсинтазы.
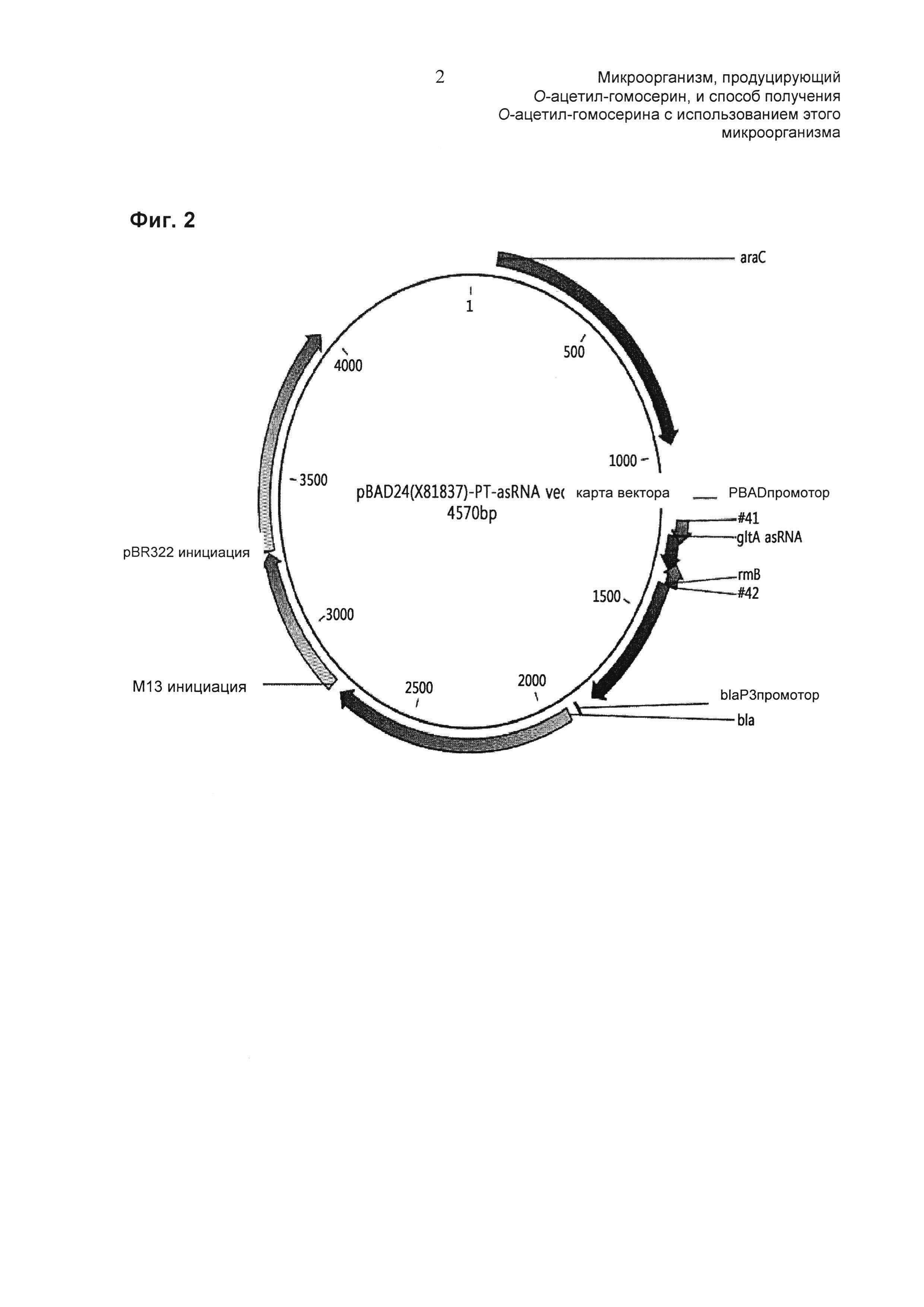
На Фиг. 2 представлена рестрикционная карта вектора pBAD24 с антисмысловой РНК (асРНК) цитратсинтазы.
ЛУЧШИЙ ВАРИАНТ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
В одном аспекте настоящего изобретения предложен микроорганизм Escherichia sp., продуцирующий О-ацетилгомосерин, с ослабленной или инактивированной активностью эндогенного белка цитратсинтазы.
При использовании в данном описании изобретения термин "О-ацетилгомосерин", являющийся конкретным промежуточным веществом в пути биосинтеза метионина микроорганизма, относится к ацетил-производному L-гомосерина. О-ацетилгомосерин может быть получен посредством ферментативной активности переноса ацетильной группы от ацетил-КоА на гомосерин с использованием гомосерина и ацетил-КоА в качестве субстратов.
При использовании в данном описании изобретения термин "микроорганизм, продуцирующий О-ацетилгомосерин" включает микроорганизм, который, являясь эукариотическим или прокариотическим микроорганизмом, продуцирующим О-ацетилгомосерин в живом организме, обеспечивает О-ацетилгомосерин-продуцирующую способность своему родительскому микроорганизму без О-ацетилгомосерин-продуцирующей способности, или микроорганизм, который эндогенно обладает О-ацетилгомосерин-продуцирующей способностью.
О-Ацетилгомосерин-продуцирующая способность может быть обеспечена или активизирована путем улучшения видов. Микроорганизмы, обладающие О-ацетилгомосерин-продуцирующей способностью, могут включать микроорганизмы, принадлежащие к Escherichia sp., Erwinia sp., Serratia sp., Providencia sp., Corynebacteria sp., Pseudomonas sp., Leptospira sp., Salmonella sp., Brevibacteria sp., Hypomononas sp., Chromobactehum sp.и Norcardia sp., или к грибам или дрожжам; конкретно, микроорганизмы, принадлежащие к Escherichia sp., Corynebacteria sp., Leptospira sp. и к дрожжам; и более конкретно, микроорганизмы, принадлежащие к Escherichia sp., в качестве конкретного примера к Escherichia coli. Микроорганизмами, обладающими О-ацетилгомосерин-продуцирующей способностью, могут быть микроорганизмы, продуцирующие L-лизин, L-треонин, L-изолейцин или L-метионин или их производные, но не ограниченные ими.
При использовании в данном описании изобретения термин "цитратсинтаза (Е.С.2.3.3.1)" относится к ферменту первой стадии цикла ТСА (цикла трикарбоновых кислот), который обеспечивает взаимодействие между оксалоацетатом и ацетил-КоА. В частности, цитратсинтаза опосредует реакцию конденсации между ацетатным остатком, имеющим два атома углерода, который находится в ацетил-КоА, и оксалоацетатом, имеющим четыре атома углерода, посредством чего образуется цитрат, имеющий шесть атомов углерода. В Escherichia coli цитратсинтаза обозначена как GltA, и "цитратсинтаза" и "GltA" в настоящем изобретении используются взаимозаменяемо.
ацетил-КоА + оксалоацетат + Н2О -> цитрат + КоА-SH
В частности, цитратсинтаза может представлять собой цитратсинтазу, происходящую из Escherichia sp. и, более конкретно, GltA, происходящую из Escherichia coli. Цитратсинтаза может быть белком, содержащим аминокислотную последовательность, представленную ка SEQ ID NO: 4, или последовательностями, имеющими 70%-ную или более гомологию с аминокислотной последовательностью SEQ ID NO: 4, конкретно 80% или более и более конкретно 90% или более. Кроме того, для последовательности, имеющей гомологию, если аминокислотная последовательность представляет собой последовательность, имеющую активность цитратсинтазы такую же или соответствующую представленной как SEQ ID NO: 4, очевидно, что аминокислотные последовательности с делецией, модификацией, замещением или добавлением в части последовательностей также должны быть включены в объем настоящего изобретения. Кроме того, учитывая вырожденность генетического кода, полинуклеотидные последовательности, кодирующие одинаковые аминокислотные последовательности и их варианты, также должны быть включены в объем настоящего изобретения.
При использовании в данном описании изобретения термин "эндогенная" активность относится к естественному состоянию белка в микроорганизме или состоянию активности соответствующего белка, имеющемуся в микроорганизме до модификации.
"Ослабление или инактивация активности белка по сравнению с его эндогенной активностью" относится к уменьшению или ликвидации активности белка по сравнению с активностью, имеющейся в его естественном состоянии. Ослабление представляет собой понятие, относящееся к случаю, когда активность белка уменьшена по сравнению с имеющейся исходно у микроорганизма из-за модификации гена, кодирующего белок, к случаю, когда уровень общей экспрессии белка ниже, чем у природного штамма микроорганизма, или к комбинации этих случаев, однако не ограничивается ими. Инактивация включает случай, когда ген, кодирующий белок, не экспрессируется совсем по сравнению с геном в природном штамме, и к случаю, когда ген экспрессируется, но не проявляет активности.
Ослабление или инактивация активности белка могут быть достигнуты различными способами, хорошо известными в данной области. Примеры способов могут включать способ замены гена, кодирующего белок, на хромосоме, геном, модифицированным таким образом, что ферментативная активность белка может быть уменьшена, включая случай ликвидации активности белка; способ введения модификации в последовательность, регулирующую экспрессию гена, кодирующего белок, на хромосоме; способ замены последовательности, регулирующей экспрессию гена, кодирующего белок, последовательностью, имеющей слабую активность или не имеющей активности; способ удаления части или всего гена, кодирующего белок, на хромосоме; способ введения антисмыслового олигонуклеотида (например антисмысловой РНК), который ингибирует трансляцию белка с мРНК посредством комплементарного связывания с транскриптом гена на хромосоме; способ, делающий невозможным прикрепление рибосомы в результате формирования вторичной структуры посредством искусственного добавления последовательности Шайна-Дальгарно (SD) и ее комплементарной последовательности на переднем конце последовательности SD гена, кодирующий белок; способ конструирования посредством обратной транскрипции (RTE), когда добавляют промотор таким образом, чтобы обеспечить обратную транскрипцию на 3'-конце открытой рамки считывания (ORF) соответствующей последовательности и т.д., а также включает комбинацию этих способов, но не ограничивается ими.
В частности, способ удаления части или всего гена, кодирующего белок, может быть осуществлен путем замены полинуклеотида, кодирующего эндогенный целевой белок, в хромосоме с помощью вектора для вставки хромосомы в микроорганизм полинуклеотидом или маркером, у которого удалена часть полинуклеотидной последовательности. Например, может быть использован способ делеции гена посредством гомологичной рекомбинации, без ограничения им. Кроме того, При использовании в данном описании изобретения термин "часть", хотя он может варьироваться в зависимости от видов полинуклеотида, может конкретно относиться к частям, составляющим от 1 нуклеотида до 300 нуклеотидов, более конкретно от 1 нуклеотида до 100 нуклеотидов и еще более конкретно от 1 нуклеотида до 50 нуклеотидов, но не ограничивается ими.
Кроме того, способ модификации последовательности, регулирующей экспрессию, может быть осуществлен путем индукции изменения последовательности, регулирующей экспрессию полинуклеотидной последовательности, путем делеции, вставки, консервативной замены, неконсервативной замены или их комбинации, чтобы дополнительно ослабить активность последовательности, регулирующей экспрессию; или путем замены полинуклеотидной последовательности полинуклеотидной последовательностью с более слабой активностью. Полинуклеотидная последовательность может включать последовательность промотора, последовательность оператора, последовательность, кодирующую домен связывания с рибосомами, и регулирующая последовательность окончания транскрипции и трансляции, но не ограничена ими.
Кроме того, способ модификации последовательности гена на хромосоме может быть осуществлен путем индукции изменения последовательности путем делеции, вставки, консервативной замены, неконсервативной замены или их комбинации, чтобы дополнительно ослабить активность последовательности, регулирующей экспрессию, или путем замены последовательности последовательностью гена, усовершенствованного так, чтобы иметь более слабую активность, или последовательностью гена, усовершенствованного так, чтобы не иметь активности, но не ограничивается ими.
В частности, для ослабления активности белка цитратсинтазы часть аминокислот(ы) в аминокислотной последовательности белка цитратсинтазы может быть заменена другими(ой) аминокислотами(ой). Более конкретно, может быть включена цитратсинтаза, имеющая аминокислотную последовательность, в которой 145-ая аминокислота или 167-ая аминокислота в аминокислотной последовательности белка цитратсинтазы, представленная тирозином (Y) или лизином (K), заменена на другую(ие) аминокислоту(ы). Еще более конкретно, цитратсинтаза может иметь последовательность гена, кодирующую модифицированный полипептид, в котором 145-ая аминокислота в аминокислотной последовательности белка цитратсинтазы, представленная тирозином (Y), заменена на аланин (A) и 167-ая аминокислота, представленная лизином (K), заменена на аланин (A). В частности, номер остатка аминокислоты был определен последовательно после обозначения аминокислоты, расположенной после метионина, который кодируется инициирующим кодоном, как 1-ой аминокислоты. Полипептид может, соответственно, иметь аминокислотную последовательность, представленную SEQ ID NO: 1 или 2. Кроме того, если активность цитратсинтазы более слабая, чем у дикого типа, цитратсинтаза может включать аминокислотные последовательности, имеющие 80%-ную или более гомологию с аминокислотной последовательностью SEQ ID NO: 1 или 2, например 90% или более, особенно 95% или более, и в особенности 97% или более. Что касается последовательности, имеющей гомологию, то, если аминокислотная последовательность имеет биологическую активность по существу такую же или соответствующую активности белка согласно SEQ ID NO: 1 или 2, очевидно, что аминокислотные последовательности с делецией, модификацией, заменой или добавлением в части последовательностей также должны быть включены в объем настоящего изобретения.
При использовании в данном описании изобретения термин "гомология" относится к проценту идентичности двух полинуклеотидов или полипептидных группировок. Гомология последовательностей от фрагмента к другому фрагменту может быть определена с помощью технологии, известной в данной области. Например, гомология может быть определена путем прямой систематизации информации о последовательностях двух различных полинуклеотидных молекул или двух различных полипептидов с использованием систематизации посредством компьютерной программы и легкого получения информации о последовательности. Компьютерная программа может включать BLAST (NCBI), CLC Main Workbench (CLC bio), MegAlignTM (DNASTAR Inc) и другие. Кроме того, гомологию полинуклеотидов можно определить путем гибридизации полинуклеотидов в условиях формирования стабильной двойной нити между гомологичными участками, расщепления с помощью нуклеазы, специфичной к одноцепочечным полипептидам, и определения расщепленных фрагментов.
При использовании в данном описании изобретения термин "гомология" относится к взаимоотношению между белками, имеющими "общее эволюционное происхождение", в том числе гомологичными белками, полученными из суперсемейства белков во всех их грамматических формах или с вариантами написания, и между белками, полученными из разных видов. Эти белки (и гены, кодирующие их) имеют гомологию последовательностей, отражающую высокие уровни сходства последовательности. Однако термин "гомология" для его общего использования и использования в настоящем изобретении будет относиться к сходству последовательностей, охарактеризованному прилагательным, таким как "очень высокое", вместо ссылки на общее эволюционное происхождение.
В типичном воплощении настоящего изобретения микроорганизм может быть таким, в котором активность цистатионин-гамма-синтазы (ЕС 2.5.1.48), гомосеринкиназы (ЕС 2.7.1.39) или активности их обоих ниже, чем их эндогенные активности, или они инактивированы.
При использовании в данном описании изобретения термин "цистатионин-гамма-синтаза" относится к ферменту, который может синтезировать цистатионин посредством химической реакции, описанной ниже, с использованием О-сукцинилгомосерина и L-цистеина в качестве субстратов. В настоящем изобретении цистатионин-гамма-синтаза из Е. coli обозначена как "MetB".
О-сукцинил-L-гомосерин + L-цистеин -> L-цистатионин + сукцинат
В частности, цистатионин-гамма-синтаза из Е. coli, без ограничения эти микроорганизмом, может представлять собой белок, содержащий аминокислотную последовательность, представленную SEQ ID NO: 9, или белки, имеющие гомологию 70% или более с аминокислотной последовательностью SEQ ID NO: 9, в частности 80% или более, и особенно 90% или более. Кроме того, в отношении последовательности, имеющей гомологию, если аминокислотная последовательность представляет собой последовательность, обладающую активностью гомосеринкиназы, такой же или соответствующей аминокислотной последовательности SEQ ID NO: 9, то очевидно, что аминокислотные последовательности с делецией, модификацией, заменой или добавлением в части последовательностей также должны быть включены в объем настоящего изобретения. Кроме того, учитывая вырожденность генетического кода, полинуклеотидные последовательности, кодирующие такую же аминокислотную последовательность, и их варианты также должны быть включены в объем настоящего изобретения.
Способ ослабления и инактивации активности цистатионин-гамма-синтазы может быть осуществлен способом, описанным выше.
При использовании в данном описании изобретения термин "гомосеринкиназа" относится к ферменту, вызывающему фосфорилирование гомосерина, который выполняет химическую реакцию, описанную ниже. В настоящем изобретении гомосеринкиназа из Е. coli обозначена как "ThrB".
ATP + L-гомосерин -> ADP + O-фосфо-L-гомосерин
В частности, гомосеринкиназа из Escherichia sp., без конкретного ограничения ею, может представлять собой белок, содержащий аминокислотную последовательность, представленную SEQ ID NO: 11, или последовательности, имеющие гомологию 70% или более с аминокислотной последовательностью SEQ ID NO: 11, в частности 80% или более, или особенно 90% или более. Кроме того, в отношении последовательности, обладающей гомологией, если аминокислотная последовательность является последовательностью, имеющей активность гомосеринкиназы такую же или соответствующую аминокислотной последовательностью SEQ ID NO: 11, то очевидно, что аминокислотные последовательности с делецией, модификацией, заменой или добавлением в части последовательностей должны быть включены в объем настоящего изобретения. Кроме того, учитывая вырожденность генетического кода, полинуклеотидные последовательности, кодирующие такую же аминокислотную последовательность, и их варианты также должны быть включены в объем настоящего изобретения.
Способ ослабления и инактивации активности гомосеринкиназы может быть осуществлен способом, описанным выше.
В конкретном аспекте настоящего изобретения микроорганизм может представлять собой такой микроорганизм, в котором активность гомосерин-О-ацетилтрансферазы дополнительно введена или усилена или эндогенная гомосерин-О-сукцинилтрансфераза дополнительно модифицирована с получением гомосерин-О-сукцинилтрансферазной активности.
При использовании в данном описании изобретения термин "гомосерин-О-ацетилтрансфераза (ЕС 2.3.1.31)" относится к ферменту, обладающему способностью переносить ацетильную группу из ацетил-КоА в гомосерин.
В частности, микроорганизм по настоящему изобретению может быть введен с активностью гомосерин-О-ацетилтрансферазы. Гомосерин-О-ацетилтрансфераза может происходить из различных видов микроорганизмов, например из микроорганизма, выбранного из Corynebacteria sp., Leptospira sp., Deinococcus sp., Deinococcus sp., Pseudomonas sp. и Mycobacterium sp. В частности, гомосерин-О-ацетилтрансфераза может включать аминокислотные последовательности, представленные как SEQ ID NO: 13 (Leptospira meyeri), SEQ ID NO: 14 (Corynebacterium glutamicum) или SEQ ID NO: 15 (Deinococcus radiodurans), но не ограничена ими. Кроме того, гомосерин-О-ацетилтрансфераза может представлять собой белок, содержащий указанные выше аминокислотные последовательности или имеющие гомологию 70% или более с вышеуказанными аминокислотными последовательностями, особенно 80% или более, или более конкретно 90% или более. Кроме того, учитывая вырожденность генетического кода, полинуклеотидные последовательности, кодирующие такую же аминокислотную последовательность, и их варианты также должны быть включены в объем настоящего изобретения.
Примеры гомосерин-О-ацетилтрансферазы для использования в настоящем изобретении раскрыты в публикации корейской патентной заявки 10-2011-0023703 и в публикации европейской патентной заявки ЕР 2290051, и все характеристики этих патентных документов могут быть включены в качестве ссылок в настоящее изобретение.
Кроме того, белок, в котором эндогенная гомосерин-О-сукцинилтрансфераза (ЕС 2.3.1.46) модифицирована с получением гомосерин-О-сукцинилтрансферазной активности, относится к полипептиду, у которого субстратная специфичность полипептида, имеющего гомосерин-О-сукцинилтрансферазную активность, изменена с сукцинил-КоА на ацетил-КоА. Кроме того, модифицированный белок, но без конкретного ограничения им, может представлять собой полипептид, обладающий гомосерин-О-ацетилтрансферазной активностью, в отличие от дикого типа, в результате замены части аминокислотной последовательности полипептида, имеющего гомосерин-О-сукцинилтрансферазную активность.
Примерами гомосерин-О-сукцинилтрансферазы может быть полипептид из Enterobacteria sp., Salmonella sp., Pseudomonas sp., Bacillus sp. или Escherichia sp., конкретно полипептид из Escherichia sp., имеющий гомосерин-О-сукцинилтрансферазную активность, например полипептид, имеющий гомосерин-О-сукцинилтрансферазную активность из Е. coli. Более конкретно, гомосерин-О-сукцинилтрансфераза из Е. coli может иметь аминокислотную последовательность, представленную SEQ ID NO: 16, но не ограничивается ею. Гомосерин-О-сукцинилтрансферазы из Е. coli обозначена как "MetA".
Модифицированная гомосерин-О-сукцинилтрансфераза может представлять собой вариант полипептида, в котором 111-ая аминокислота полипептида, представленного как SEQ ID NO: 16, или полипептидов, имеющих гомологию 95% или более с полинуклеотидной последовательностью SEQ ID NO: 16 заменена глутаминовой кислотой и, кроме того, 112-ая аминокислота заменена треонином или гистидином. В частности, вариант полипептида может представлять собой полипептид, имеющий любую аминокислотную последовательность из SEQ ID NO: 17-19. Кроме того, вариант полипептида может представлять собой белок, содержащий аминокислотную последовательность, имеющую гомологию 70% или более с вышеуказанной аминокислотной последовательностью, конкретно 80% или более, или особенно 90% или более. Кроме того, учитывая вырожденность генетического кода, полинуклеотидные последовательности, кодирующие такую же аминокислотную последовательность, и их варианты должны быть включены в объем настоящего изобретения. Информацию о модифицированной гомосерин-О-сукцинилтрансферазе можно получить из публикации корейской патентной заявки 10-2012-0070531 или из международной публикации WO 2012/087039, и все описания из этих патентных документов включены в настоящее изобретение в виде ссылок.
При использовании в данном описании изобретения термин "введение или усиление активности" относится к обеспечению активности конкретного белка у микроорганизма, который не имеет этого белка; или усиление внутриклеточной активности белка в микроорганизме, который имеет этот белок, и тому подобное, и относится к увеличению внутриклеточной активности белка по сравнению с эндогенной активностью белка.
При использовании в данном описании изобретения термин "введение или усиление активности белка" относится не только к получению большего эффекта, чем исходная функция, из-за увеличения активности самого белка, но также к увеличению активности белка за счет увеличения эндогенной активность гена, эндогенной амплификации гена при помощи внутренних и внешних факторов, увеличения числа копий, введения гена извне, увеличения ферментативной активности за счет замены, модификации или мутации и т.д., но не ограничен ими.
Среди указанного выше увеличение числа копий гена, хотя и не ограничено этим, может быть осуществлено в состоянии функциональной связи с вектором или путем включения в хромосому в клетке-хозяине. В частности, способ может быть осуществлен путем введения вектора, с которым полинуклеотид, кодирующий белок по настоящему изобретению, функционально связан и может реплицироваться и функционировать независимо от хозяина в клетке хозяина; или путем введения вектора, с которым полинуклеотид функционально связан, введения полинуклеотида в хромосому клетки-хозяина, в клетку-хозяин, тем самым увеличивая число копий гена внутри хромосомы клетки-хозяина.
Вектор представляет собой ДНК-конструкцию, содержащую полинуклеотидную последовательность полинуклеотида, кодирующего целевой белок, который функционально связан с подходящей регулирующей последовательностью, так что целевой белок может экспрессироваться в соответствующем хозяине, где регулирующая последовательность включает промотор, инициирующий транскрипцию, случайная последовательность оператора для регуляции транскрипции, последовательность, кодирующую подходящий домен связывания рибосом в мРНК, и последовательность регулирования транскрипции и трансляции. Вектор после трансформации в подходящую клетку-хозяина может быть реплицирован или функционировать независимо от генома хозяина, или он может быть интегрирован в сам геном хозяина.
Вектор, используемый в настоящем изобретении, может не быть конкретно ограниченным до тех пор, пока этот вектор реплицируется в клетке-хозяине, и можно использовать любой вектор, известный в данной области техники. Примеры вектора могут включать природные и рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, λMBL3, λMBL4, λIXII, λASHII, λAPII, λt10, λt11, Charon4A, Charon21A, и т.д.; и в качестве плазмидного вектора можно использовать векторы на основе pBR, на основе pUC, на основе pBluescriptII, на основе pGEM, на основе pTZ, на основе pCL, на основе рЕТ и т.д. В частности, можно использовать векторы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.д.
Кроме того, полинуклеотид, кодирующий эндогенный целевой белок, может быть замещен модифицированным полинуклеотидом с использованием вектора для встраивания в хромосому микроорганизма. Встраивание полинуклеотида в хромосому может быть осуществлено с использованием известного в данной области способа, например посредством гомологичной рекомбинации. Так как вектор по настоящему изобретению может быть встроен в хромосому посредством гомологичной рекомбинации, дополнительно может быть включен селективный маркер для подтверждения встраивания в хромосому. Селективный маркер используют для отбора трансформированных клеток, то есть для того, чтобы подтвердить, что целевой полинуклеотид был встроен, и могут быть использованы маркеры, обеспечивающие селектируемые фенотипы, такие как лекарственная резистентность, потребность в питательных веществах, резистентность к цитотоксическим агентам и экспрессия поверхностных белков, без ограничения ими. В условиях обработки селективными агентами только клетки, экспрессирующие селективные маркеры, могут выжить и экспрессировать другие фенотипические признаки и, таким образом, трансформированные клетки могут быть легко отобраны.
При использовании в данном описании изобретения термин "трансформация" относится к способу введения вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, посредством чего обеспечивается возможность экспрессии полинуклеотида, кодирующего белок в клетке-хозяине. Для трансформированного полинуклеотида не имеет значения, встроен ли он в хромосому клетки-хозяина и находится в ней или находится вне хромосомы, пока он может экспрессироваться в клетке-хозяине. Кроме того, полинуклеотид включает ДНК и РНК, которая кодирует целевой белок. Полинуклеотид может быть встроен в любой форме настолько, насколько он может быть введен в клетку-хозяина и экспрессироваться там. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, включающую все существенные элементы, необходимые для ее экспрессии. Экспрессионная кассета обычно может включать промотор, функционально связанный с полинуклеотидом, сигнал окончания транскрипции, домен связывания с рибосомой и сигнал окончания трансляции, и может находиться в форме экспрессионного вектора, способного к саморепликации. Кроме того, полинуклеотид может быть введен в клетку-хозяина как есть и функционально связан с последовательностью, необходимой для его экспрессии в клетке-хозяине. Кроме того, при использовании в данном описании изобретения термин "функционально связан" относится к функциональной связи между последовательностью промотора, который инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой белок, и последовательностью гена.
Кроме того, модификация последовательности, регулирующей экспрессию, для увеличения экспрессии полинуклеотида может быть осуществлена путем индукции вариации в полинуклеотидной последовательности посредством делеции, вставки, консервативной замены, неконсервативной замены или их комбинации, так чтобы дополнительно повысить активность последовательности, регулирующей экспрессию; или путем замены полинуклеотидной последовательности полинуклеотидной последовательностью с более сильной активностью, но конкретно не ограничена этими методами. Последовательность, регулирующая экспрессию, может включать промотор, последовательность оператора, последовательность, кодирующую домен связывания с рибосомой, и последовательность, регулирующую окончание транскрипции и трансляции и т.д. хотя конкретно и не ограничена ими. Кроме того, сильный экзогенный промотор, вместо исходного промотора, может быть соединен с верхним концом экспрессионной единицы полинуклеотида.
Кроме того, модификация полинуклеотидной последовательности на хромосоме, хотя и не ограничена конкретно ими, может быть осуществлена путем индукции вариации последовательности, регулирующей экспрессию полинуклеотидной последовательности посредством делеции, вставки, консервативной замены, неконсервативной замены или их комбинации, так чтобы усилить активность полинуклеотидной последовательности; или путем замены полинуклеотидной последовательности улучшенной полинуклеотидной последовательностью с более сильной активностью.
Как правило, введение и усиление активности белка может увеличивать активность или концентрацию соответствующего белка относительно активности или концентрацией белка дикого типа или в микроорганизме от по меньшей мере 1%, 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400% или 500% до максимального значения 1000% или 2000%.
Кроме того, микроорганизм может быть таким, у которого активность эндогенной гомосерин-О-сукцинилтрансферазы ослаблена или инактивирована по сравнению с ее эндогенной активностью, для того чтобы усилить путь биосинтеза О-ацетилгомосерина путем блокировании пути биосинтеза О-сукцинилгомосерина из гомосерина.
Ослабление и инактивация активности гомосерин-О-сукцинилтрансферазы может быть осуществлена в соответствии со способом, описанным выше.
В типичном воплощении настоящего изобретения О-ацетилгомосерин-продуцирующий микроорганизм может быть таким, в котором дополнительно введена или увеличена активность фермента, вовлеченного в путь биосинтеза от фосфоенолпирувата до гомосерина, для того чтобы дополнительно увеличить количество гомосерина, субстрата для биосинтеза О-ацетилгомосерина.
В частности, вышеуказанный микроорганизм может быть таким, в котором дополнительно введена или увеличена активность по меньшей мере одного белка, выбранного из группы, состоящей из фосфоенолпируваткарбоксилазы (ррс, ЕС 4.1.1.31), аспартатаминотрансферазы (aspC, ЕС 2.6.1.1) и аспартатполуальдегиддегидрогеназы (asd, ЕС 1.2.1.11).
Например, в микроорганизм может быть введен ген ppc, кодирующий фосфоенолпируваткарбоксилат, включающий аминокислотную последовательность, представленную SEQ ID NO: 20, ген aspC, кодирующий аспартатаминотрансферазу, включающий аминокислотную последовательность, представленную SEQ ID NO: 21, и ген asd, кодирующий аспартатполуальдегиддегидрогеназу, включающий аминокислоту, представленную SEQ ID NO: 22. Например, активности трех разных ферментов могут быть введены и усилены путем получения всех генов, кодирующих эти три разных фермента, присутствующих в хромосоме клетки-хозяина с числом копий по меньшей мере 2, без ограничения этим методом. Введение и увеличение активностей может быть осуществлено в соответствии с методом, описанным выше.
В типичном воплощении настоящего изобретения активность белка цитратсинтазы ослаблена или инактивирована различными способами, которые включают удаление гена цитратсинтазы в микроорганизме Е. coli, продуцирующем О-ацетилгомосерин; введение гена, кодирующего модифицированный белок цитратсинтазы, активность которого ослаблена по сравнению с активностью дикого типа, в положение гена цитратсинтазы; и введение экспрессионного вектора для антисмысловой РНК гена цитратсинтазы. В результате, О-ацетилгомосерин-продуцирующий микроорганизм, сконструированный таким образом, в котором активность белка цитратсинтазы ослаблена или инактивирована, продемонстрировал повышенную способность производить О-ацетилгомосерин по сравнению со способностью родительского микроорганизма (Примеры 1-4).
В другом аспекте настоящего изобретения предложен способ продуцирования О-ацетилгомосерина с использованием О-ацетилгомосерин-продуцирующего микроорганизма с улучшенной способностью продуцировать О-ацетилгомосерин. В частности, в настоящем изобретении предложен способ продуцирования О-ацетилгомосерина, включающий (а) культивирование микроорганизма; и (б) выделение О-ацетилгомосерина, полученного во время культивирования микроорганизма.
Способ культивирования Е. coli, обладающей способностью продуцировать О-ацетилгомосерин, по настоящему изобретению, может быть осуществлен в соответствии с подходящими условиями среды и культивирования, известными в данной области техники. Способ культивирования может быть легко адаптирован специалистом в данной области в зависимости от выбранного микроорганизма. В частности, поскольку микроорганизм по настоящему изобретению является микроорганизмом, у которого активность цитратсинтазы, представляющей собой фермент, опосредующий первую стадию цикла Кребса (ТСА), ослаблена или инактивирована, культуральная среда может включать глутамат, но конкретно не ограничена такой средой.
Примеры способов культивирования могут включать периодическую культуру, непрерывную культуру и культуру с подпиткой, но не ограничены ими. Эти различные методы раскрыты, например, в "Biochemical Engineering" James М. Lee, Prentice-Hall International Editions, pp 138-176.
Среда, используемая в культивировании, может подходящим образом удовлетворять требованиям конкретного микроорганизма. В частности, примеры культуральной среды для микроорганизмов раскрыты в "Manual of Methods for General Bacteriology", the American Society for Bacteriology, Washington, DC, 1981. Культуральная среда может быть такой, которая включает подходящий источник углерода, источник фосфора, неорганическое соединение, аминокислоту и/или витамины и т.д., и культивирование может быть осуществлено в аэробных условиях при установленной температуре, pH и т.д.
Примеры источника углерода могут включать углеводы, такие как глюкоза, лактоза, сахароза, молочная кислота, фруктоза, мальтоза, крахмал и целлюлоза; жиры, такие как соевое масло, подсолнечное масло, касторовое масло, аргановое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота. Эти источники углерода могут быть использованы по отдельности или в комбинации, но не ограничены ими.
Примеры источника азота могут включать органические источники азота, такие как пептон, дрожжевой экстракт, подливка, солодовый экстракт, жидкий кукурузный экстракт (CSL) и мука из бобов; и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота можно использовать по отдельности или в комбинации, но не ограничены ими. Культуральная среда может также включать, в качестве источника фосфора, дигидрофосфат калия, гидрофосфат калия и соответствующие натрий-содержащие соли. Культуральная среда может включать металлы, такие как сульфат магния и сульфат железа. Кроме того могут быть включены аминокислоты, витамины и подходящие предшественники. Эта культуральная среда или предшественники могут быть добавлены к культуре в виде периодической культуры или непрерывной культуры, без ограничения ими.
Кроме того, pH культуры может быть скорректирован путем добавления соединений, таких как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота во время культивирования подходящим способом. Кроме того, образование пузырьков может быть предотвращено во время культивирования с использованием пеногасителя, такого как полигликолевый эфир жирной кислоты. Кроме того, для поддержания аэробных условий в культуральной жидкости к культуре может быть добавлен газообразный кислород или газ (например воздух), содержащий газообразный кислород. Температура культуры может быть от 20°С до 45°С и, в частности, от 25°С до 40°С, но не ограничена ими. Культивирование может продолжаться до тех пор, пока производство О-ацетилгомосерина не достигнет намеченного уровня, и, в частности, в течение от 10 часов до 160 часов, без ограничения ими.
Способ получения О-ацетилгомосерина по настоящему изобретению может также включать выделение О-ацетилгомосерина из культивируемого микроорганизма или продукта его культивирования. Выделение целевого О-ацетилгомосерина может быть осуществлено при помощи способа культивирования микроорганизма по настоящему изобретению, например подходящего способа, известного в данной области, такого как периодическая культура, непрерывная культура и периодическая культура с подпиткой.
Выделение может включать стадию очистки.
Исходя из полученного таким образом О-ацетилгомосерина можно получать метионин посредством двухстадийного способа (корейский патент 10-0905381).
Двухстадийный способ включает процесс получения L-метионина и органической кислоты посредством ферментативной реакции с использованием фермента, обладающего О-ацетилгомосерин-сульфгидрилазной активностью, или микроорганизма, обладающего этим ферментом, при использовании О-ацетилгомосерина, который получен при помощи микроорганизма, продуцирующего предшественник L-метионина, и метилмеркаптана в качестве субстратов.
Более конкретно, в настоящем изобретении предложен способ получения L-метионина посредством ферментативной реакции О-ацетилгомосерин-сульфгидрилазы и т.д., с использованием О-ацетилгомосерина, который накапливается посредством предыдущего способа, в качестве субстрата.
На второй стадии, где О-ацетилгомосерин используется как предшественник L-метионина, можно использовать О-ацетилгомосерин-сульфгидрилазу, полученную из микроорганизма, в частности принадлежащего к Leptospira sp., Chromobacterium sp. и Hyphomonas sp. и более конкретно принадлежащего к Leptospira meyeri, Pseudomonas aurogenosa, Hyphomonas Neptunium и Chromobacterium Violaceum.
Реакция является такой же, как показано ниже.
CH3SH + О-ацетил-L-гомосерин <=> ацетат + метионин
Дополнительный способ получения метионина раскрыт в корейском патенте 10-0905381 и полное описание патента может быть включено в виде ссылки в настоящее изобретение.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано более подробно со ссылкой на следующие примеры. Однако, эти примеры приведены только в иллюстративных целях и изобретение не следует ограничивать этими примерами.
Базисный пример: Конструирование микроорганизма, продуцирующего О-ацетилгомосерин
<1-1> Удаление гена metB, происходящего из Е. coli дикого типа (международная публикация WO 2008/013432)
О-ацетилгомосерин-продуцирующий микроорганизм был сконструирован с использованием Е. coli, типичного микроорганизма Escherichia sp. С этой целью использовали Е. coli K12 W3110 дикого типа (АТСС27325), полученный из Американской коллекции клеточных культур (АТСС). Сначала, для того чтобы блокировать путь синтеза от О-сукцинил-L-гомосерина до цистатионина удаляли ген metB, кодирующий цистатионинсинтазу (SEQ ID NO: 10). В частности, ген metB, кодирующий цистатионинсинтазу, удаляли посредством метода FRT (флиппаза-распознаваемая мишень)-одностадийной PCR-делеции (PNAS (2000) vol 97: Р6640-6645).
В частности, делеционную кассету metB конструировали посредством PCR-реакции на основе вектора pKD3 (PNAS (2000) vol 97: Р6640-6645) в качестве матрицы, используя праймеры SEQ ID NO: 30 и 31 следующим образом: 30 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и удлинение при 72°С в течение 1 минуты. Полученный PCR-продукт подвергали электрофорезу на 1,0% агарозном геле и очищали полученную из него полосу ДНК из 1,2 т.н. Полученный фрагмент ДНК вставляли посредством электропорации в Е. coli (К12) W3110, который уже был трансформирован вектором pKD46 (PNAS (2000) vol 97: Р6640-6645). Для электропорации штамм W3110, трансформированный pKD46, культивировали в среде LB (литическая среда), содержащей 100 мкг/л ампициллина и 5 мМ арабинозы (L-арабиноза) при 30°С вплоть до OD600=0,6 и использовали после двукратного промывания стерильной дистиллированной водой и однократного промывания 10% глицерином. Электропорацию выполняли при 2500 В. Полученный штамм сеяли штрихом на чашку со средой LB, содержащей 25 мкг/л хлорамфеникола, культивировали при 37°С в течение ночи и отбирали штамм, демонстрирующий резистентность. Выбранный штамм подвергали PCR-реакции в таких же условиях на основе штамма в качестве матрицы с использованием тех же праймеров, и делецию гена metB подтверждали, наблюдая размер гена 1,2 т.н. на 1,0% агарозном геле. Полученный таким образом штамм культивировали в среде LB после еще одной трансформации с помощью вектора рСР20 (PNAS (2000) vol 97: Р6640-6645) и конечный штамм с удаленным геном metB, у которого размер гена был уменьшен до 150 п.о. на 1,0% агарозном геле, полученный посредством PCR при тех же условиях, был сконструирован и удаление хлорамфеникольного маркера было подтверждено. Таким образом сконструированный штамм был назван "W3-B".
<1-2> Удаление гена thrB (международная публикация WO 2008/013432)
Для увеличения синтеза О-сукцинилгомосерина из гомосерина был удален ген thrB gene, который представляет собой ген, кодирующий гомосеринкиназу. Для удаления гена thrB из штамма W3-B, сконструированного в Примере 1, использовали метод FRT-одностадийной PCR делеции, используемый для удаления гена metB.
Делеционную кассету thrB конструировали посредством PCR на основе вектора pKD4 (PNAS (2000) vol 97: Р6640-6645) в качестве матрицы, используя праймеры SEQ ID NO: 32 и 33 следующим образом: 30 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и удлинение при 72°С в течение 1 минуты.
Полученный PCR-продукт подвергали электрофорезу на 1,0%-ном агарозном геле и полученную из него 1,6 т.н. полосу ДНК очищали. Извлеченный фрагмент ДНК электропорировали в штамм W3-B, который уже был трансформирован вектором pKD46. Полученный штамм сеяли штрихом на чашку с LB, содержащую 50 мкг/л канамицина, культивировали при 37°С в течение ночи и отбирали штамм, обладающий резистентностью.
Отобранный штамм подвергали PCR-реакции в тех же условиях, непосредственно основанных на штамме, используемом в качестве матрицы, используя те же праймеры SEQ ID NO: 32 и 33, и подтверждали удаление гена thrB путем селекции штамма, имеющего размер гена 1,6 т.н., на 1,0% агарозном геле. Подтвержденный таким образом штамм культивировали в среде LB после еще одной трансформации вектором рСР20 и конечный штамм с удаленным геном thrB, у которого размер гена был уменьшен до 150 п.о. на 1,0% агарозном геле в результате осуществления PCR при тех же условиях, был сконструирован и удаление канамицинового маркера было подтверждено. Таким образом сконструированный штамм был назван "W3-BT.
<1-3> Вариант metA с гомосеринацетилтрансферазной активностью (международная публикация WO 2012/087039)
Для того чтобы усилить активность гомосеринацетилтрансферазы в штамме, полученном в Базисном примере <1-2>, предполагалось ввести мутантный ген типа metA (SEQ ID NO: 24 и 26), кодирующий гомосеринацетилтрансферазу.
Во-первых, для того чтобы сконструировать вариант гена metA с усиленной активностью, выполняли реакцию PCR на основе хромосомы штамма W3110 дикого типа в качестве матрицы, используя праймеры SEQ ID NO: 34 и 35, и амплифицировали ген metA, кодирующий гомосерин-О-сукцинилтрансферазу.
Праймеры, используемые в PCR-реакции, были получены на основе полинуклеотидной последовательности хромосомы Е. coli, NC_000913, зарегистрированной в NIH Gene Bank (банк генов Национального института здравоохранения США), и праймеры SEQ ID NO: 34 и 35 имели сайты рестрикции EcoRV и HindIII соответственно. Таким образом полученный PCR-продукт и плазмида pCL1920, включающая Pcj1, были соответственно обработаны с помощью EcoRV и HindIII, и PCR-продукт клонировали в плазмиду pCL1920. Е. coli DH5α трансформировали, используя клонированную плазмиду, и трансформированные Е. coli DH5α отбирали на чашках LB, содержащих 50 мкг/мл спектиномицина, и из них получали плазмиду. Полученная таким образом плазмида была названа "pCL_Pcj1_metA".
Затем 111ую аминокислоту, глицин (Gly), О-сукцинилтрансферазы заменяли глутаминовой кислотой (Glu) (G111E) на основе сконструированной выше плазмиды pCL_Pcj1_metA в качестве матрицы, используя набор для сайт-направленного мутагенеза (Stratagene, USA). Таким образом сконструированная плазмида, включающая вариант гена G111E metA, была названа "pCL_Pcj1_metA (EL)".
Кроме того, с целью замены 111ой аминокислоты глицина на глутаминовую кислоту в О-сукцинилтрансферазе и 112ой аминокислоты лейцина на гистидин, использовали праймеры SEQ ID NO: 38 и 39. Плазмида, включающая ген metA, в котором 111ая аминокислота глицин заменена на глутаминовую кислоту и 112ая аминокислота лейцин заменена на гистидин, была названа "pCL_Pcj1_metA (EH)".
Затем конструировали кассету замещения для замены metA (EH) в штамме посредством PCR, используя вектор pKD3 в качестве матрицы вместе с праймерами SEQ ID NO: 40 и 41 следующим образом: 30 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и удлинение при 72°С в течение 2 минут. Соответствующий PCR-продукт был получен с использованием pCL-Pcj1-metA (EH) в качестве матрицы для части metA (EH) кассеты замещения вместе с праймерами SEQ ID NO: 42 и 43, и праймерами SEQ ID NO: 42 и 45 для части metA дикого типа. Кассеты замещения metA (EH), включающие часть с хлорамфеникольным маркером, были сконструированы с использованием трех разных PCR-продуктов вместе с праймерами SEQ ID NO: 42 и 45 и были электропорированы в штамм W3-BT, который уже был трансформирован вектором pKD46, сконструированным в Базисном примере <1-2>. Таким образом подтвержденный штамм культивировали в среде LB после еще одной трансформации вектором рСР20, и штамм, у которого был удален хлорамфеникольный маркер и ген metA заменен metA (EH), был назван "W3-BTA".
<1-4> Конструирование штамма с 2 копиями генов ррс, aspC и asd (публикация европейской патентной заявки ЕР 2290051)
Для того чтобы увеличить О-ацетилгомосерин-продуцирующую способность штамма W3-BTA, сконструированного в Базисном примере <1-3>, биосинтез был усилен, как в ранее поданном патенте ЕР 2290051. Таким же образом, как в вышеуказанном патенте ЕР, был сконструирован штамм, имеющий 2 амплифицированные копии генов, то есть гена ррс, кодирующего фосфоенолпируваткарбоксилазу, с использованием SEQ ID NO: 46, 47, 48 и 49; гена aspC, кодирующего аспартатаминотрансферазу, с использованием праймеров SEQ ID NO: 50 и 51; и гена asd, кодирующего аспартатполуальдегиддегидрогеназу, с использованием праймеров SEQ ID NO: 52, 53, 54 и 55. В частности, вышеуказанный штамм с усиленным биосинтезом и при этом продуцирующий О-ацетилгомосерин был назван "W3-BTA2PCD" (также называется "WCJM").
<1-5> Эксперименты с культивированием в колбе
Количество О-ацетилгомосерина, продуцируемого штаммом, сконструированным в Базисных примерах <1-3> и <1-4>, тестировали при культивировании в колбе Эрленмейера.
В частности, штаммы W3110, W3-BTA и WCJM вносили в среду LB и культивировали при 33°С в течение ночи. Затем отдельную их колонию вносили в 3 мл среды LB, культивировали при 33°С в течение 5 часов, разбавляли в 200 раз в 250 мл колбе Эрленмейера, содержащей 25 мл О-ацетилгомосерин-производящей среды, культивировали при 33°С при скорости 200 оборотов в минуту в течение 30 часов и количество продуцируемого О-ацетилгомосерина исследовали с помощью анализа ВЭЖХ. Используемые составы среды приведены в Таблице 1 ниже и исследованное количество продуцируемого О-ацетилгомосерина показано в Таблице 2 ниже.


Результаты показали, что W3110 дикого типа не продуцирует О-ацетилгомосерин совсем, в то время как штамм W3-BTA продуцирует 0,9 г/л О-ацетилгомосерина и штамм WCJM, который был усилен в отношении биосинтеза, продуцирует 1,2 г/л of О-ацетилгомосерина.
Пример 1: Удаление цитратсинтазной активности
<1-1> Конструирование микроорганизма с удаленным геном цитратсинтазы в О-ацетилгомосерин-продуцирующем микроорганизме
Цитратсинтаза (GltA) представляет собой фермент первой стадии цикла ТСА и начинает с взаимодействия между оксалоацетатом и ацетил-КоА. Ингибирование роста посредством снижения цикла Кребса хорошо известно (Meded Rijksuniv Gent Fak Landbouwkd Toegep Biol Wet. 2001; 66(3a): 333-6). Тем не менее, для того чтобы увеличить количество ацетил-КоА, используемого в качестве субстрата для О-ацетилгомосерина, следовало получить штаммы W3-BTA и WCJM, где удалена цитратсинтазная активность.
В частности, ген цитратсинтазы в штаммах W3-BTA и WCJM был удален посредством PCR на основе вектора pKD4 в качестве матрицы с использованием праймеров SEQ ID NO: 56 и 57 следующим образом: 30 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд, и удлинение при 72°С в течение 2 минут. Полученный PCR-продукт подвергали электрофорезу на 1,0% агарозном геле, и было подтверждено что размер гена составляет 1,6 т.н., и его ДНК была очищена. Полученные фрагменты ДНК вводили посредством электропорации в штаммы W3-BTA и WCJM, которые уже были трансформированы вектором pKD46. Для электропорации штаммы W3-BTA и WCJM, трансформированные вектором pKD46, культивировали в среде LB, содержащей 100 мкг/л ампициллина и 5 мМ арабинозы при 30°С вплоть до OD600=0,6 (оптическая плотность при 600 нм), и дважды промывали дистиллированной водой и один раз 10%-ным глицерином для использования. Электропорацию выполняли при 2500 В. Полученные таким образом штаммы сеяли штрихом на LB чашки, содержащие 50 мкг/л канамицина, культивировали при 37°С и отбирали штаммы, демонстрирующие резистентность.
Выбранные штаммы были подвергнуты PCR с использованием праймеров SEQ ID NO: 58 и 59 в тех же условиях, подвергнуты электрофорезу на 1,0% агарозном геле и размер гена наблюдался равным 2,5 т.н., тем самым подтверждали, что делеционная кассета была встроена в часть гена цитратсинтазы на хромосоме. Подтвержденные таким образом штаммы трансформировали еще раз вектором рСР20, культивировали в среде LB и штаммы с делецией гена цитратсинтазы, размер которого был уменьшен до 1,1 т.н. на 1,0%-ном агарозном геле, конструировали посредством PCR, при этом было подтверждено, что канамициновые маркеры были удалены. Сконструированные таким образом штаммы были названы "W3-BТA-AD" и "WCJM-AD" соответственно.
<1-2> Оценка микроорганизма с удаленным геном цитратсинтазы в О-ацетилгомосерин-продуцирующем микроорганизме
Штаммы W3-BTA-AD и WCJM-AD могут расти в среде LB, но из-за делеции гена цитратсинтаза они не могут расти в среде, содержащей О-ацетилгомосерин. Для того чтобы определить количество продуцируемого О-ацетилгомосерина выполняли культивирование в колбе Эрленмейера в условиях добавления 3 г/л глутамата (Таблица 3 - состав с добавлением глутамата в среду) в существующий состав культуральной среды.
В частности, штаммы W3-BTA-AD и WCJM-AD вносили в среду LB и культивировали при 33°С в течение ночи. Затем отдельную их колонию вносили в 3 мл среды LB, культивировали при 33°С в течение 5 часов, разбавляли в 200 раз в 250 мл колбе Эрленмейера, содержащей 25 мл О-ацетилгомосерин-продуцирующей среды (с добавленным глутаматом), культивировали при 33°С при скорости 200 оборотов в минуту в течение 30 часов и количество продуцируемого О-ацетилгомосерина оценивали посредством ВЭЖХ-анализа. Определенное количество продуцируемого О-ацетилгомосерина показано в Таблице 4 ниже.



Результат определения продукции О-ацетилгомосерина посредством культивирования в колбе выявил, что штамм W3-BTA продуцирует 0,9 г/л О-ацетилгомосерина, W3-BTA-AD продуцирует 1,4 г/л О-ацетилгомосерина, что составляет 55,6%-ное увеличение, хотя он показывает уменьшение потребления глюкозы. Штамм WCJM продуцирует 1,3 г/л О-ацетилгомосерина, в то время как штамм WCJM-AD продуцирует 2,1 г/л О-ацетилгомосерина, таким образом подтверждая, что способность производить О-ацетилгомосерин улучшается на 61,5% благодаря делеции гена цитратсинтаза.
Пример 2: Ослабление активности белка цитратсинтазы
<2-1> Типы модификаций гена цитратсинтазы
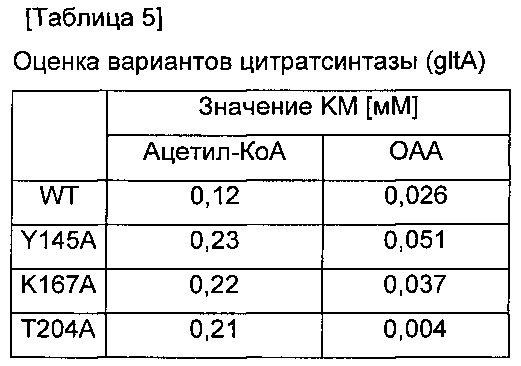
Штамм WCJM-AD, сконструированный в Примере <1-1>, показал низкую скорость культивирования и были выбраны три разных варианта, которые показывали ослабленную активность и уменьшенную способность связываться с ацетил-КоА в соответствии с различными модификациями цитратсинтазы, известными по многочисленным ссылкам (The Journal of Biological Chemistry, 2003, 278, 35435-35443). Информация о трех разных вариантах приведена в Таблице 5, где показаны модифицированные гены, в которых 145-ая аминокислота тирозин (Y) заменена на аланин (А), 167-ая аминокислота лизин (K) заменена на аланин (А) и 204-ая аминокислота треонин (Т), заменена на аланин (А).
<2-2> Конструирование микроорганизма с ослабленной активностью белка цитратсинтазы в О-ацетилгомосерин-продуцирующем микроорганизме
Авторы настоящего изобретения планировали увеличить производительность путем введения вариантов, в которых активность белка цитратсинтазы была ослаблена, как описано в Примере <2-1>, в О-ацетилгомосерин-продуцирующий микроорганизм.
Для того чтобы ввести три разных типа вариантов гена цитратсинтазы в штамм WCJM-AD была разработана кассета модифицирующей замены, как показано на Фиг. 1. Каждый вариант синтезировали путем замены праймера нуклеотидом, и каждая кассета была сконструирована в помощью 3 PCR-продуктов. Для части гена цитратсинтазы в качестве матрицы использовали штамм W3110 и осуществляли модификацию 145ой аминокислоты, PCR-реакции выполняли, используя праймеры SEQ ID NO: 60 и 63 и SEQ ID NO: 62 и 61 соответственно, и получали PCR-продукты с размером 514 п.о. и 1112 п.о.
Точно также, в результате модификации 167-ой аминокислоты получали PCR-продукты с размером 580 п.о. и 1046 п.о. с использованием праймеров SEQ ID NO: 60 и 65 и праймеров SEQ ID NO: 64 и 61, а в результате модификации 204-ой аминокислоты получали PCR-продукты с размером 688 п.о. и 936 п.о. с использованием праймеров SEQ ID NO: 60 и 67 и SEQ ID NO: 66 и 61. Для общей канамициновой части выполняли PCR-реакции на основе вектора pKD4 в качестве матрицы, используя праймеры SEQ ID NO: 68 и 69. В частности, для вставки в положение гена цитратсинтазы кассету конструировали таким образом, чтобы включить полинуклеотидную последовательность ниже гена цитратсинтазы в SEQ ID NO: 69, и PCR-продукт с размером 1571 п.о. получали посредством электрофореза. Сшивающую PCR-реакцию выполняли на основе ДНК фрагмента канамицина, который является общей частью для каждого из двух фрагментов ДНК, отобранных в соответствии с модификациями, соответственно, используя праймеры SEQ ID NO: 60 и 69, следующим образом: 30 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд, и удлинение при 72°С в течение 4 минут. Каждый из конечных PCR-продуктов был подтвержден на 1,0% агарозном геле и ДНК фрагменты с размером 3115 п.о. для трех разных типов кассет модификаций гена цитратсинтазы. Собранные фрагменты ДНК электропорировали в штамм WCJM-AD, который уже был трансформирован вектором pKD46. Для электропорации штамм W3110, трансформированный pKD46, культивировали в среде LB, содержащей 100 мкг/л ампициллина и 5 мМ арабинозы, при 30°С вплоть до OD600=0,6 и использовали после двукратного промывания стерильной дистиллированной водой и однократного промывания 10% глицерином. Электропорацию выполняли при 2500 В. Полученный штамм сеяли штрихом на чашку с LB, содержащей 25 мкг/л канамицина, культивировали при 37°С в течение ночи и отбирали штамм, демонстрирующий резистентность. Отобранный штамм подвергали PCR-реакции в тех же условиях на основе штамма в качестве матрицы, используя те же праймеры SEQ ID NO: 58 и 59, и подтверждали делецию гена metB, наблюдая размер гена 3,7 т.н. на 1,0% агарозном геле, тем самым подтверждая, что была встроена кассета модификации, в которой аминокислота гена цитратсинтазы была заменена. Подтвержденный таким образом штамм культивировали в среде LB после еще одной трансформации вектором рСР20 и были сконструированы три вариантных штамма в отношении активности цитратсинтазы, где размер гена был уменьшен до 2,5 т.н. на 1,0% агарозном геле в результате PCR, осуществленной при тех же условиях, и было подтверждено удаление канамицинового маркера. Полученные таким образом штаммы были названы "WCJM-A145", "WCJM-A167ʺ и "WCJM-A204" и информация о последовательности гена цитратсинтазы, введенного с модификациями, показана в SEQ ID NO: 1-3 (аминокислотные последовательности) и в SEQ ID NO: 5-7 (нуклеотидные последовательности) соответственно.
<2-3> Оценка микроорганизмов с ослабленной активностью цитратсинтазы в О-ацетилгомосерин-продуцирующих микроорганизмах
Культивирование в колбе Эрленмейера выполняли для того, чтобы определить количество О-ацетилгомосерина, продуцируемого тремя разными штаммами WCJM-A145, WCJM-A167 и WCJM-A204, где активность гена цитратсинтазы была ослаблена. Четыре типа штаммов, то есть штаммы WCJM-A145, WCJM-A167 и WCJM-A204, в том числе штамм WCJM, вносили в среду LB и культивировали при 33°С в течение ночи. Затем их одиночную колонию вносили в 3 мл среды LB, культивировали при 33°С в течение 5 часов, разбавляли в 200 раз в 250 мл колбе Эрленмейера, содержащей 25 мл О-ацетилгомосерин-продуцирующей среды, культивировали при 33°С со скоростью 200 оборотов в минуту в течение 30 часов, и количество продуцируемого О-ацетилгомосерина оценивали посредством ВЭЖХ-анализа. Результаты показаны в Таблице 6 ниже.
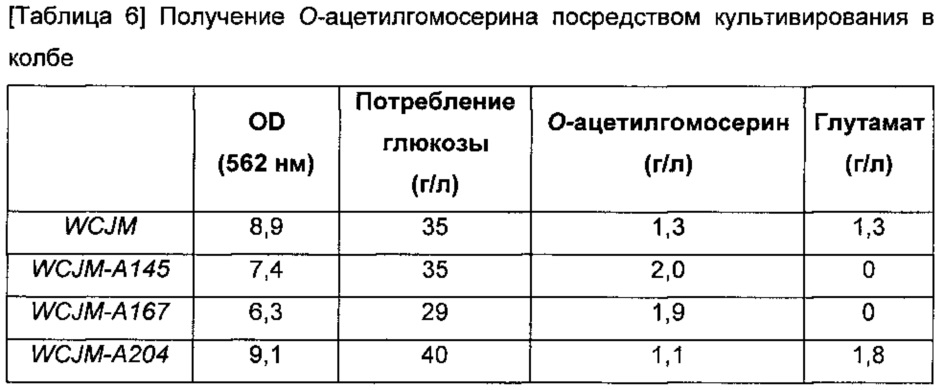
Результат продуцирования О-ацетилгомосерина в колбе выявил, что штамм WCJM продуцировал 1,3 г/л О-ацетилгомосерина, а два штамма WCJM-A145 и WCJM-A167 продуцировали 2,0 г/л и 1,9 г/л О-ацетилгомосерина соответственно, в то время как величина их потребления глюкозы уменьшалась наряду с уменьшением их поглощения (OD). Принимая во внимание специфическое уменьшение глутамата от 1,3 г/л до 0 г/л, было подтверждено, что этот результат обусловлен уменьшением потока в цикле Кребса, вызванного ослаблением активности цитратсинтазы. Однако штамм WCJM-A204 показал увеличение глутамата, в то же время демонстрируя уменьшение количества продуцированного О-ацетилгомосерина до 0,2 г/л, таким образом, подтверждая, что модификация усиливает активность.
Пример 3: Ослабление экспрессии белка цитратсинтазы
<3-1> Конструирование экспрессионного вектора для антисмысловой РНК (асРНК) гена цитратсинтазы
Авторы настоящего изобретения сделали попытку применить метод антисмысловой РНК (асРНК) для того, чтобы ослабить экспрессию белка цитратсинтазы. Метод антисмысловой РНК представляет собой способ уменьшения экспрессии белка путем нейтрализации связывания мРНК цитратсинтазы и рибосомы посредством сверхэкспрессии комплементарного участка связывания для мРНК цитратсинтазы гена-мишени. Преимуществом этого метода является то, что он может регулировать уровень ингибирования путем контролирования силы связывания с мРНК гена цитратсинтазы, и этот метод также полезен для конструирования рекомбинантного микроорганизма, так как этот метод может эффективно создавать и уменьшать экспрессию гена посредством антисмысловой РНК, контролирующей экспрессию гена, без необходимости в обычном процессе делеции гена.
Конструирование вектора выполняли в соответствии со ссылкой (Methods Mol Biol. 2012; 815:307-19. doi: 10.1007/978-1-61779-424-7_23.) и для сверхэкспрессии область антисмысловой РНК гена синтазы следовало ввести в плазмиду pBAD24, способную к индукции. Карта вектора pSAD24-асРНК цитратсинтазы показана на Фиг. 2. Область экспрессии, где экспрессировалась антисмысловая РНК гена цитратсинтазы, имеет размер 100 п.о., включая область промотора размером 52 п.о. и область размером 48 п.о. от инициирующего кодона цитратсинтазы, и структура спаренных концов (РТ)размером 38 п.о., которая снижает нестабильность антисмысловой РНК (асРНК), соединена с обеими фланкирующими областями. Область антисмысловой РНК гена цитратсинтазы была получена с использованием праймеров SEQ ID NO: 70 и 71 и сайты рестрикции NcoI и HindIII были включены для клонирования в вектор.
Полученный таким образом PCR-продукт имел размер 194 п.о., и этот PCR-продукт клонировали в плазмиду pBAD24 после обработки с помощью EcoRV и HindIII соответственно. Таким образом клонированную плазмиду использовали для трансформации Е. coli DH5α, и трансформированные Е. coli DH5α отбирали на LB чашках, содержащих 100 мкг/мл ампициллина, и из них получали плазмиду. Полученная таким образом плазмида была названа "pBAD24-gltA асРНК".
<3-2> Введение экспрессионного вектора антисмысловой РНК гена цитратсинтазы в О-ацетилгомосерин-продуцирующий микроорганизм и его оценка
Экспрессионный вектор антисмысловой РНК гена цитратсинтазы pBAD24-gltA-acPHK использовали для трансформации штамма WCJM, который представляет собой О-ацетилгомосерин-продуцирующий микроорганизм. Здесь трансформированный штамм был назван "WCJM/A-асРНК". В частности, была предпринята попытка контролировать величину экспрессии белка цитратсинтазы путем контролирования величины экспрессии антисмысловой РНК цитратсинтазы и здесь величину экспрессии антисмысловой РНК можно контролировать в соответствии с концентрацией арабинозы.
В результате было подтверждено, что количество продуцируемого О-ацетилгомосерина увеличивается при ослаблении активности цитратсинтазы, как в Примере 2.
Кроме того, выполняли культивирование в колбе Эрленмейера, чтобы установить, увеличивается ли количество продуцируемого О-ацетилгомосерина при уменьшении величины экспрессии цитратсинтазы.
В частности, штаммы WCJM и WCJM/A-асРНК вносили в среду LB и культивировали при 33°С в течение ночи. Затем отдельную их колонию вносили в 3 мл среды LB, культивировали при 33°С в течение 5 часов, разбавляли в 200 раз в 250 мл колбе Эрленмейера, содержащей 25 мл О-ацетилгомосерин-продуцирующей среды. В частности, чтобы контролировать величину экспрессии антисмысловой РНК цитратсинтазы, добавляли арабинозу в концентрациях 0 мМ, 2 мМ и 5 мМ и культивировали при 33°С со скоростью 200 оборотов в минуту в течение 15 часов и 30 часов. Количество продуцируемого О-ацетилгомосерина оценивали посредством ВЭЖХ-анализа, результаты показаны в приведенных ниже Таблицах 7 и 8.


В результате было подтверждено, что при культивировании в течение 15 часов штамм WCJM/A-асРНК показал уменьшение OD (оптической плотности) до примерно 1 в соответствии с концентрацией арабинозы, в то время как концентрация О-ацетилгомосерина была аналогичной. Однако при культивировании в течение 30 часов штамм WCJM, который представляет собой контрольный штамм, показал такую же OD и концентрацию О-ацетилгомосерина даже при увеличении концентрации арабинозы, в то время как штамм WCJM/A-асРНК, который представляет собой штамм с введенным экспрессионным вектором для антисмысловой РНК цитратсинтазы, показал значительную разницу при увеличении концентрации арабинозы. OD составляла 9,2 при концентрации арабинозы 0 мМ, в то время как OD составляла 7,1 при концентрации арабинозы 5 мМ, уменьшение 5,1, и количество О-ацетилгомосерина увеличилось на 30,8% несмотря на низкое потребление глюкозы. Эти результаты подтвердили, что не только ослабление активности цитратсинтазы, но также ослабление экспрессии белка демонстрирует такие же результаты.
Пример 4: Ослабление и инактивация активности цитратсинтазы в микроорганизме с высоким выходом продуцируемого О-ацетилгомосерина
<4-1> Конструирование микроорганизма с высоким выходом продуцируемого О-ацетилгомосерина с инактивированной активностью цитратсинтазы и его оценка
В международной публикации WO 2012/087039 подробно раскрыт способ конструирования О-ацетилгомосерин-продуцирующего микроорганизма из треонин-продуцирующего микроорганизма, происходящего от штамма W3110 дикого типа в результате мутации NTG. В частности, сконструированный штамм, продуцирующий с высоким выходом О-ацетилгомосерин, был депонирован в Корейском центре культур микроорганизмов (KCCM) под учетным номером KCCM 11146Р.
Штамм KCCM11146P может поглощать 40 г/л глюкозы во время культивирования в колбе, продуцирует примерно от 15 г/л до 16 г/л О-ацетилгомосерина и, таким образом, считается обладающим высокой производительностью в отношении О-ацетилгомосерина. Соответственно, чтобы определить продуцирует ли штамм более высокий выход О-ацетилгомосерина, когда цитратсинтазная активность удалена, то же самое было применено к штамму KCCM11146P. Способ конструирования был таким же как в Примере <1-1>, и с помощью этого способа штамм KCCM11146P с удаленной активностью цитратсинтазы был сконструирован и назван "KCCM11146P-AD".
Количество О-ацетилгомосерина, продуцируемого штаммом KCCM11146P, у которого была удалена активность цитратсинтазы, определяли путем культивирования в колбе Эрленмейера. Штамм KCCM11146P или KCCM11146P-AD вносили в среду LB и культивировали при 33°С в течение ночи. Затем одну колонию этих штаммов вносили в 3 мл среды LB, культивировали при 33°С в течение 5 часов, разбавляли в 200 раз в 250 мл колбе Эрленмейера, содержащей 25 мл О-ацетилгомосерин-продуцирующей среды (с добавленным глутаматом) и культивировали при 33°С со скоростью 200 оборотов в минуту в течение 30 часов. Количество продуцируемого О-ацетилгомосерина оценивали посредством ВЭЖХ-анализа и результаты представлены в Таблице 9 ниже.

Результат продуцирования О-ацетилгомосерина посредством культивирования в колбе выявил, что штамм KCCM11146P продуцирует 14,2 г/л О-ацетилгомосерина и штамм KCCM11146P-AD продуцирует 16,7 г/л О-ацетилгомосерина, с увеличением 17,6%, хотя он показывал уменьшение поглощения (OD).
<4-2> Конструирование микроорганизма с высоким выходом продуцируемого О-ацетилгомосерина с ослабленной активностью цитратсинтазы и его оценка
Для того чтобы оценить, продуцирует ли штамм KCCM11146P, который представляет собой штамм с высоким выходом произведенного О-ацетилгомосерина, более высокий выход О-ацетилгомосерина даже при ослабленной активности цитратсинтазы, к штамму KCCM11146P были применены модификация 145-ой аминокислоты (тирозин (Y) на аланин (А)) и модификация 167-ой аминокислоты (лизин (K) на аланин (А)), которые показывали самые высокие О-ацетилгомосерин-продуцирующие способности среди трех различных типов ослабления активности белка, описанных в Примере <2-1>.
Метод конструирования был таким же, как в Примере <2-2> и с помощью этого метода были сконструированы два штамма KCCM11146P, у которых активность цитратсинтазы была ослаблена, и они были названы "KCCM11146P-А145" и "KCCM11146P-А167" соответственно.
Количество продуцируемого О-ацетилгомосерина двумя штаммами KCCM11146P-А145 и KCCM11146P-А167, у которых активность цитратсинтазы была ослаблена, определяли путем культивирования в колбе Эрленмейера. Три штамма, то есть штаммы KCCM11146P-А145 и KCCM11146P-А167 и штамм KCCM11146P вносили в среду LB и культивировали при 33°С в течение ночи. Затем одну их колонию вносили в 3 мл среды LB, культивировали при 33°С в течение 5 часов, разбавляли в 200 раз в 250 мл колбе Эрленмейера, содержащей 25 мл О-ацетилгомосерин-продуцирующей среды и культивировали при 33°С со скоростью 200 оборотов в минуту в течение 30 часов. Количество продуцируемого О-ацетилгомосерина оценивали посредством ВЭЖХ-анализа, и результаты приведены в Таблице 10 ниже.

Результат продуцирования О-ацетилгомосерина посредством культивирования в колбе выявил, что штамм KCCM11146P продуцировал 15,0 г/л О-ацетилгомосерина и два штамма KCCM11746P-А145 и KCCM11146P-А167 показали аналогичные результаты, как в Примере <2-3>. Два штамма соответственно продуцировали 17,5 г/л и 17,3 г/л О-ацетилгомосерина, с увеличением примерно 16,7%, хотя оба они показали снижение оптической плотности (OD).
Штамм с высоким выходом продуцирования О-ацетилгомосерина также показал уменьшение глутамата от 1,6 г/л до 0 г/л, в соответствии с уменьшением потока в цикле Кребса, вызванного ослаблением активности цитратсинтазы.
Эти результаты демонстрируют, что активность цитратсинтазы позволяет производить О-ацетилгомосерин путем применения ослабляющей модификации. Кроме того, они также показывают, что при проведении реакции превращения на основе О-ацетилгомосерина, продуцированного в соответствии с международной публикацией WO 2008/013432 в качестве матрицы, и при использовании фермента превращения, который, кроме того, имеет активности цистатионин-гамма-синтазы, О-сукцинилгомосерин-сульфгидрилазы и О-ацетилгомосерин-сульфгидрилазы, можно одновременно синтезировать L-метионин и ацетат.
Авторы настоящего изобретения подтвердили, что штамм KCCM11146P, вариантный по 167-ой аминокислоте цитратсинтазы, обладает улучшенным продуцированием О-ацетилгомосерина, обозначили штамм KCCM11146P-А167 как "СА05-4007" и депонировали его в Корейском центре культур микроорганизмов (KCCM), международном органе по депонированию в соответствии с Будапештским договором, 22 ноября 2013 года (Учетный номер: KCCM11483P).
Из вышеизложенного специалисту в данной области, к которой относится настоящее изобретение, должно быть понятно, что настоящее изобретение может быть воплощено в других конкретных формах без изменения технических концепций или существенных характеристик настоящего изобретения. В связи с этим типичные воплощения, раскрытые здесь, предназначены только для иллюстративных целей и не должны быть истолкованы как ограничивающие объем настоящего изобретения. Напротив, предполагается, что настоящее изобретение охватывает не только типичные воплощения, но также и различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены, не выходя за рамки сущности и объема настоящего изобретения, как определено в прилагаемой формуле изобретения.
Реферат
Изобретение относится к области биохимии, генной инженерии и биотехнологии, в частности к трансформированному микроорганизму Escherichia sp., продуцирующему О-ацетилгомосерин. Настоящий микроорганизм характеризуется тем, что в нем ослаблена или инактивирована активность эндогенной цитратсинтазы. В результате указанной модификации этот микроорганизм способен продуцировать О-ацетилгомосерин с высоким выходом. Изобретение также относится к способу получения О-ацетилгомосерина. Настоящий способ предусматривает культивирование указанного микроорганизма и выделение О-ацетилгомосерина, продуцируемого при культивировании этого микроорганизма. Настоящее изобретение позволяет получать О-ацетилгомосерин с высоким выходом. 2 н. и 5 з.п. ф-лы, 2 ил., 10 табл., 4 пр.
Комментарии