Способ получения l-аргинина и l-цитруллина с использованием бактерии семейства enterobacteriaceae, в которой ослаблена экспрессия гена pepa - RU2471870C2
Код документа: RU2471870C2
Чертежи






Описание
Область техники
Настоящее изобретение относится к микробиологической промышленности, в частности, к способу получения L-аргинина и L-цитруллина с использованием бактерии семейства Enterobacteriaceae, модифицированной таким образом, что экспрессия гена рерА, кодирующего аминопептидазу А, в указанной бактерии ослаблена.
Описание предшествующего уровня техники
Традиционно L-аминокислоты в промышленном масштабе могут быть получены методом ферментации с использованием штаммов микроорганизмов, полученных из природных источников, или их мутантов, специально модифицированных для того, чтобы увеличить продукцию L-аминокислот.
Описано множество методов увеличения продукции L-аминокислот, например, путем трансформации микроорганизма рекомбинантной ДНК (см., например, патент США 4278765). Другие методы основаны на повышении активности ферментов, вовлеченных в биосинтез аминокислот, и/или уменьшении чувствительности целевого фермента к обратному ингибированию продуцируемой L-аминокислотой (см., например, патенты США 4346170; 5661012 и 6040160).
Другим методом увеличения продукции L-аминокислот является ослабление экспрессии одного или нескольких генов, вовлеченных в деградацию целевой L-аминокислоты; генов, экспрессия которых ведет к отвлечению предшественников целевой аминокислоты от пути биосинтеза L-аминокислоты; генов, вовлеченных в перераспределение потоков углерода, азота и фосфора; генов, кодирующих токсины и т.д.
Ген рерА (синоним carP) Escherichia coli кодирует мультифункциональный фермент аминопептидазу А (РерА), которая сочетает каталитические и регуляторные свойства. Показано, что кроме участия в гидролизе N-концевых аминокислотных остатков белка, аминопептидаза А требуется для сайтспецифической рекомбинации ДНК и мономеризации плазмидных мультимеров (Stirling et al. (1989) EMBO J., 8, 1623-1627; Charlier et al (1995) J. Mol. Biol., 250, 392-406). Еще одна функция аминопептидазы А - регуляция транскрипции. Аминопептидаза А является ДНК-связывающим белком и вовлечена в репрессию транскрипции оперона carAB, контролирующего синтез карбамоилфосфатсинтазы (Roovers et al, (1988) J. Mol. Biol., 204, 857-865). Карбамоилфосфат является промежуточным соединением путей биосинтеза L-аргинина и пиримидина. Взаимодействие между карбамоилфосфатом и L-орнитином приводит к образованию L-цитруллина, превращающегося через L-аргининсукцинат в L-аргинин.
В настоящее время нет сообщений, описывающих использование ослабления экспрессии гена рерА для продукции L-аминокислот, таких как L-аргинин и L-цитруллин.
Описание изобретения
Цели настоящего изобретения включают повышение продуктивности штаммов-продуцентов L-аргинина или L-цитруллина и предоставление способа получения L-аминокислоты, такой как L-аргинин и L-цитруллин, с использованием этих штаммов.
Вышеупомянутые цели были достигнуты путем установления того факта, что инактивация гена рерА, кодирующего аминопептидазу А, приводит к дерепрессии синтеза карбамоилфосфата и может приводить к повышению продукции L-аминокислоты, такой как L-аргинин. Кроме того, поскольку L-цитруллин является промежуточным соединением пути биосинтеза L-аргинина, продукция этой L-аминокислоты также может быть увеличена.
Настоящее изобретение представляет бактерию, принадлежащую к семейству Enterobacteriaceae, обладающую повышенной способностью к продукции аминокислоты, такой как L-аргинин и цитруллин.
Целью настоящего изобретения является предоставление бактерии, принадлежащей к семейству Enterobacteriaceae, обладающей способностью к продукции L-аргинина и L-цитруллина, отличающейся тем, что указанная бактерия модифицирована таким образом, что экспрессия гена рерА, кодирующего аминопептидазу А, в указанной бактерии ослаблена.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что указанный ген рерА инактивирован.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что указанная бактерия принадлежит к роду Escherichia.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что указанная бактерия принадлежит к роду Pantoea.
Также целью настоящего изобретения является предоставление способа получения L-аминокислоты, выбранной из группы, состоящей из L-аргинина и L-цитруллина, включающего выращивание описанной выше бактерии в питательной среде для продукции и секреции указанной L-аминокислоты в культуральную жидкость и выделение указанной L-аминокислоты из культуральной жидкости. Более детально настоящее изобретение описано ниже.
1. Бактерия согласно настоящему изобретению
Бактерия согласно настоящему изобретению - это бактерия-продуцент L-аминокислоты, принадлежащая к семейству Enterobacteriaceae, отличающаяся тем что указанная бактерия модифицирована таким образом, что экспрессия гена рерА, кодирующего аминопептидазу А, в модифицированной бактерии ослаблена.
Фраза "бактерия-продуцент L-аминокислоты" означает бактерию, обладающую способностью к продукции и секреции L-аминокислоты в питательную среду, когда бактерия выращивается в указанной питательной среде.
Термин «бактерия-продуцент L-аминокислоты» также может означать бактерию, которая способна к продукции и накоплению L-аминокислоты в ферментационной среде в больших количествах по сравнению с природным или родительским штаммом Е. coli, таким как штамм Е. coli К-12, и также может означать, что бактерия способна накапливать в среде целевую L-аминокислоту в количестве не менее чем 0.5 г/л или не менее чем 1.0 г/л в другом примере. Термин «L-аминокислота» может включать L-аргинин и L-цитруллин.
Семейство Enterobacteriaceae включает в себя бактерии, принадлежащие к родам Escherichia, Enterobacter, Erwinia, Klebsiella, Pantoea, Photorhabdus, Providencia, Salmonella, Serratia, Shigella, Morganella, Yersinia и т.д. Более конкретно, могут быть использованы бактерии, классифицируемые как принадлежащие к семейству Enterobacteriaceae в соответствии с таксономией, используемой в базе данных NCBI (National Center for Biotechnology Information) (http://www.ncbi.nlm.nih.gov/htbinpost/Taxonomy/wgetorg?mode=Tree&id=1236&lvl=3&keep=1&srchmode=1&unlock). Предпочтительна бактерия, принадлежащая к роду Escherichia или Pantoea.
Термин "бактерия, принадлежащая к роду Escherichia", означает, что бактерия относится к роду Escherichia в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера микроорганизма, принадлежащего к роду Escherichia, использованного в настоящем изобретении, может быть упомянута бактерия Escherichia coli (Е.coli).
Круг бактерий, принадлежащих к роду Escherichia, которые могут быть использованы в настоящем изобретении, не ограничен каким-либо образом, однако, например, бактерии, описанные в книге Neidhardt, F.C. et al. (Escherichia coli and Salmonella typhimurium, American Society for Microbiology, Washington D.C., 1208, Таблица 1), могут быть включены в число бактерий согласно настоящему изобретению.
Термин «бактерия, принадлежащая к роду Pantoea» означает, что бактерия относится к роду Pantoea в соответствии с классификацией, известной специалисту в области микробиологии. Недавно несколько видов Enterobacter agglomerans были классифицированы как Pantoea agglomerans, Pantoea ananatis, Pantoea stewartii или подобные им, на основе анализа нуклеотидной последовательности 16S рРНК и т.д. (Int. J. Syst. Bacteriol., 43, 162-173 (1993)).
Термин «бактерия модифицирована таким образом, что экспрессия гена рерА, кодирующего аминопептидазу А, ослаблена» означает, что указанная бактерия модифицирована таким образом, что в результате модификации такая бактерия содержит пониженное количество аминопептидазы А по сравнению с немодифицированной бактерией, или это также может означать, что модифицированная бактерия не способна синтезировать аминопептидазу А.
Фраза «инактивация гена рерА» означает, что модифицированный ген кодирует полностью неактивный белок. Возможно также, что естественная экспрессия модифицированного участка ДНК невозможна из-за делеции части гена, сдвига рамки считывания, введения миссенс/нонсенс мутации(-ий) или модификации примыкающих к гену областей, которые включают последовательности, контролирующие экспрессию гена, такие как промоторы, энхансеры, аттенуаторы и т.д.
Наличие или отсутствие гена рерА на хромосоме может быть определено хорошо известными методами, включая ПЦР, блоттинг по Саузерну и т.п. Кроме того, уровень экспрессии гена можно оценить определением количества транскрибируемой с гена РНК с использованием различных известных методов, включая блоттинг по Нозерну, количественную ОТ-ПЦР и т.п. Количество и молекулярную массу белков, кодируемых генами, можно определить известными методами, включая электрофорез в SDS-ПААГ с последующим иммуноблоттингом (Вестерн-блоттинг) и т.п.
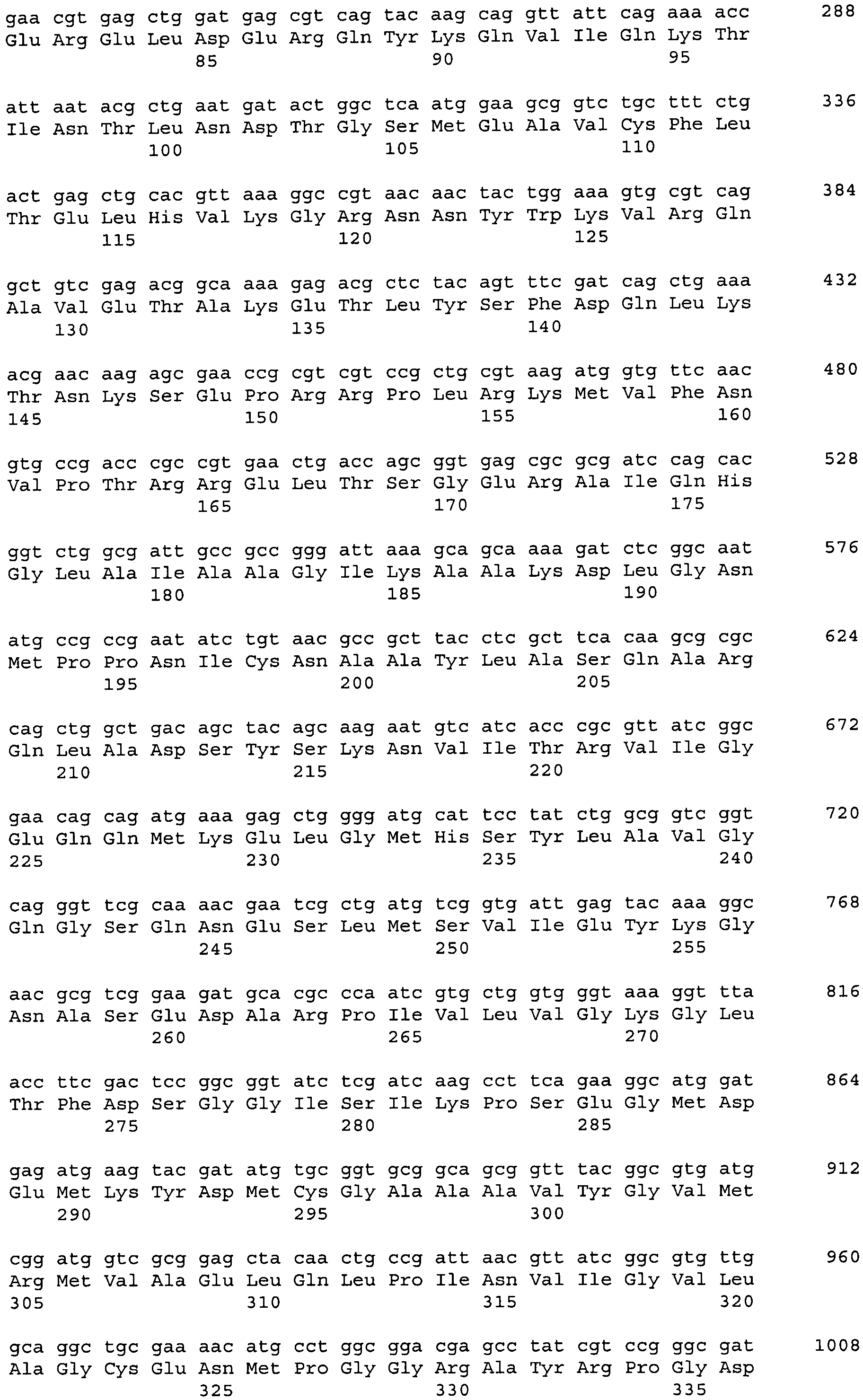
Последовательность гена рерА (синонимы: carP, b4260, ECK4253), кодирующего аминопептидазу A Escherichia coli, известна (нуклеотиды, комплементарные нуклеотидам с 4482463 по 4483974 в последовательности с инвентарным номером NC_000913.2 в базе данных GenBank). Ген рерА Е.coli расположен на хромосоме между генами holC и lptF. Нуклеотидная последовательность гена рерА и аминокислотная последовательность белка РерА, кодируемого геном рерА, приведены в Перечне последовательностей под номерами 1 (SEQ ID NO:1) и 2 (SEQ ID NO:2) соответственно.
Поскольку у представителей различных родов и штаммов семейства Enterobacteriaceae возможны некоторые вариации в нуклеотидных последовательностях, понятие инактивируемого гена не ограничивается геном, последовательность которого приведена в Перечне последовательностей под номером SEQ ID NO: 1, но также может включать и гены, гомологичные SEQ ID NO: 1, кодирующие вариант белка. Термин "вариант белка" может означать белок с изменениями в последовательности, будь то делеции, вставки, добавления или замены аминокислот, в котором сохраняется активность белка. Число изменений в варианте белка зависит от положения или типа аминокислотного остатка в третичной структуре белка. Оно может быть от 1 до 30, или в другом примере от 1 до 15, или в другом примере от 1 до 5. Данные изменения в вариантах могут иметь место в областях, не критичных для функции белка. Данные изменения возможны потому, что некоторые аминокислоты имеют высокую гомологию друг другу, поэтому такие изменения не влияют на третичную структуру или активность. Следовательно, вариант белка, кодируемого геном, может быть представлен белками с гомологией не менее 80%, или в другом примере не менее 90%, или в другом примере не менее 95%, по отношению к полной аминокислотной последовательности, приведенной в Перечне последовательностей под номером SEQ ID NO: 2 при условии, что до инактивации белок сохраняет активность аминопептидазы А.
Гомология между двумя аминокислотными последовательностями может быть определена с использованием известных методов, например, компьютерной программы BLAST 2.0, которая считает три параметра: число аминокислот, идентичность и сходство.
Кроме того, ген рерА может быть вариантом, который гибридизуется в жестких условиях с нуклеотидной последовательностью, приведенной в Перечне последовательностей под номером SEQ ID NO: 1, или с зондом, который может быть синтезирован на основе указанной нуклеотидной последовательности при условии, что до инактивации он кодирует функциональный белок. «Жесткие условия» включают такие условия, при которых специфические гибриды, например, гибриды с гомологией не менее 60%, в другом примере не менее 70%, в другом примере не менее 80%, в другом примере не менее 90%, в другом примере не менее 95%, образуются, а неспецифические гибриды, например, гибриды с меньшей гомологией, чем указано выше, - не образуются. Практическим примером жестких условий является однократная отмывка, в другом примере двух- или трехкратная, при концентрации солей 1×SSC, 0.1% SDS, предпочтительно 0.1×SSC, 0.1% SDS, при 60°С. Длина зонда может быть выбрана в зависимости от условий гибридизации, обычная длина около 100-1000 п.н.
Экспрессия гена может быть ослаблена введением в гены на хромосоме таких мутаций, что внутриклеточная активность кодируемого геном белка уменьшена по сравнению с таковой в немодифицированном штамме. Такой мутацией гена может быть замена одного или более оснований для аминокислотной замены в кодируемом геном белке («миссенс»-мутация), введение стоп-кодона («нонсенс »-мутация), делеция одного или более оснований для сдвига рамки считывания, вставка гена устойчивости к антибиотику, или делеция гена или его части (J. Biol. Chem., 1997, 272 (13); 8611-8617, J. Antimicrobial Chemotherapy, 2000, 46: 793-96). Экспрессия гена chaC или оперона chaBC также может быть ослаблена модификацией экспрессии регуляторных последовательостей, таких как промотор, последовательность Shine-Dalgarno (SD) и т.д. (заявка РСТ WO 95/34672; Carrier, T.A. and Keasling, J.D., Biotechnol Prog 15, 58-64 (1999)).
Например, следующие методы могут применяться для введения мутаций путем генной рекомбинации. Конструируется мутантный ген, кодирующий мутантный белок со сниженной активностью, и бактерия для ее модификации трансформируется фрагментом ДНК, содержащим мутантный ген. Затем нативный ген на хромосоме замещается гомологичной рекомбинацией мутантным геном, отбирается полученный штамм. Такое замещение гена с использованием гомологичной рекомбинации может быть проведено методом с использованием линейной ДНК, известный как "Red-зависимая интеграция" или "интеграция посредством Red-системы" (Datsenko, K.A., Wanner, B.L., Proc. Natl. Acad. Sci. USA. 97, 12, 6640-6645 (2000), заявка РСТ WO 2005/010175) или методом с использованием плазмиды, репликация которой чувствительна к температуре (патент США 6303383 или патентная заявка Японии JP 05-007491 A). Далее, введение сайтспецифической мутации путем замещения гена с использованием вышеупомянутой гомологичной рекомбинации может также быть осуществлено с использованием плазмиды с пониженной способностью к репликации в клетке хозяина.
Экспрессия гена также может быть ослаблена вставкой транспозона или IS-фактора в кодирующую область гена (патент США 5175107), или традиционными методами, такими как мутагенез с использованием УФ-излучения или обработка нитрозогуанидином (N-метил-N'-нитро-N-нитрозогуанидин), сайтспецифический мутагенез, разрушение гена с использованием гомологичной рекомбинации или/и мутагенеза за счет вставки-делеции (Yu, D. et al., Proc. Natl. Acad. Sci. USA, 2000, 97:12: 5978-83 and Datsenko, K.A. and Wanner, B.L., Proc. Natl. Acad. Sci. USA, 2000, 97:12: 6640-45), также называемого "Red-зависимая интеграция".
Методы приготовления плазмидной ДНК, рестрикции и лигирования ДНК, трансформации, выбора нуклеотидов в качестве праймера и т.п. могут быть обычными методами, известными специалисту в этой области. Эти методы описаны, например, в Sambrook, J., Fritsch, E.F., and Maniatis, Т., "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989).
Бактерия-продуцент L-аминокислоты
В качестве бактерии может быть использована бактерия, способная к продукции L-аминокислоты, предпочтительно L-аргинина и L-цитруллина.
Бактерия может быть получена путем инактивации гена рерА, кодирующего аминопептидазу А, в бактерии, уже обладающей способностью к продукции L-аминокислоты. С другой стороны, бактерия может быть получена путем придания бактерии, в которой ген рерА, кодирующий аминопептидазу А, инактивирован, способности к продукции L-аминокислоты.
Бактерия-продуцент L-аргинина
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-аргинина согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм Е. coli 237 (ВКПМ В-7925) (патентная заявка США 2002/058315 А1) и его производные, содержащие мутантную N-ацетилглутаматсинтазу (патентная заявка РФ 2001112869), штамм Е.coli 382 (ВКПМ В-7926) (Европейская патентная заявка ЕР 1170358 А1), штамм-продуцент аргинина, в который введен ген argA, кодирующий N-ацетилглутаматсинтетазу (Европейская патентная заявка ЕР 1170361 А1), и подобные им.
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-аргинина, согласно настоящему изобретению также включают в себя штаммы, в которых усилена экспрессия одного или нескольких генов, кодирующих ферменты биосинтеза L-аргинина. Примеры ферментов биосинтеза L-аргинина включают N-ацетилглутамилфосфатредуктазу (argC), орнитинацетилтрансферазу {argJ), N-ацетилглутаматкиназу (argB), ацетилорнитинтрансаминазу (argD), орнитинкарбамоилтрансферазу (argF), синтетазу аргининсукциниловой кислоты (argG), лиазу аргининсукциниловой кислоты (argH) и карбамоилфосфатсинтетазу (car AB).
Бактерия-продуцент L-цитруллина
Примеры родительских штаммов, используемых для получения бактерии-продуцента цитруллина согласно настоящему изобретению, включают в себя, но не ограничиваются ими, штаммы, принадлежащие к роду Escherichia, такие как штаммы Е. coli 333 (VKPM В-8084) и 374 (VKPM В-8086), содержащие мутантную устойчивую к ингибированию по типу обратной связи карбамоилфосфатсинтетазу (RU 2264459 С2), штаммы Е. coli, в которых увеличена активность α-кетоглутаратсинтазы, и дополнительно модифицированы активности ферредоксин-НАДФ+-редуктазы, пируватсинтазы или α-кетоглутаратдегидрогеназы (ЕР 2133417 А1), и штамм Р. ananantis NA1sucAsdhA, в котором снижены активности сукцинатдегидрогеназы и α-кетоглутаратдегидрогеназы (патентная заявка США No 2009286290) и подобные им.
Поскольку L-цитруллин является промежуточным соединением пути биосинтеза L-аргинина, примеры родительских штаммов, которые могут быть использованы для конструирования бактерии-продуцента L-цитруллина, включают штаммы, в которых усилена экспрессия одного или нескольких генов, кодирующих ферменты L-аргинина. Примеры таких генов включают, но не ограничиваются ими, гены, кодирующие N-ацетилглутаматсинтазу (argA), N-ацетилглутаматкиназу (argB), N-ацетилглутамилфосфатредуктазу (argC), ацетилорнитинтрансаминазу (argD), ацетилорнитиндеацетилазу (argE), орнитинкарбамоилтрансферазу (argF/I) и карбамоилфосфатсинтетазу (car AB) или их комбинации.
Примеры родительских штаммов, которые могут быть использованы для конструирования бактерии-продуцента L-цитруллина настоящего изобретения, могут также включать штаммы - продуценты L-аргинина, в которых активность аргининсукцинатсинтазы (ArgG), катализирующей конверсию L-цитруллина в L-аргинин, снижена по сравнению с родительским штаммом. Активность аргининсукцинатсинтазы может быть снижена путем инактивации соответствующего гена argG с использованием описанных выше традиционных методов.
2. Способ согласно настоящему изобретению.
Способом согласно настоящему изобретению является способ получения L-аминокислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-аминокислоты, такой как L-аргинин и L-цитруллин, в питательной среде, и выделения L-аминокислоты из культуральной жидкости.
Выращивание, выделение и очистка L-аминокислоты из культуральной среды могут быть осуществлены способом, подобным традиционным способам ферментации, в которых аминокислота продуцируется с использованием бактерии.
Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизмов. В качестве источника углерода могут использоваться различные углеводы, такие как глюкоза и сахароза, а также различные органические кислоты. В зависимости от формы ассимиляции выбранного микроорганизма может быть использован спирт, включая этанол и глицерин. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и т.п.
Выращивание осуществляется предпочтительно в аэробных условиях, таких как перемешивание культуральной жидкости на качалке, взбалтывание с аэрацией, при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 30 до 38°С. pH среды поддерживают в пределах от 5 до 9, предпочтительно от 6.5 до 7.2. pH среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Обычно выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем L-аминокислота может быть выделена и очищена методами ионообменной хроматографии, концентрирования и/или кристаллизации.
Краткое описание рисунков
На Фигуре 1 показано относительное положение праймеров Р1 и Р2 на плазмиде pMW118-attL-Cm-attR.
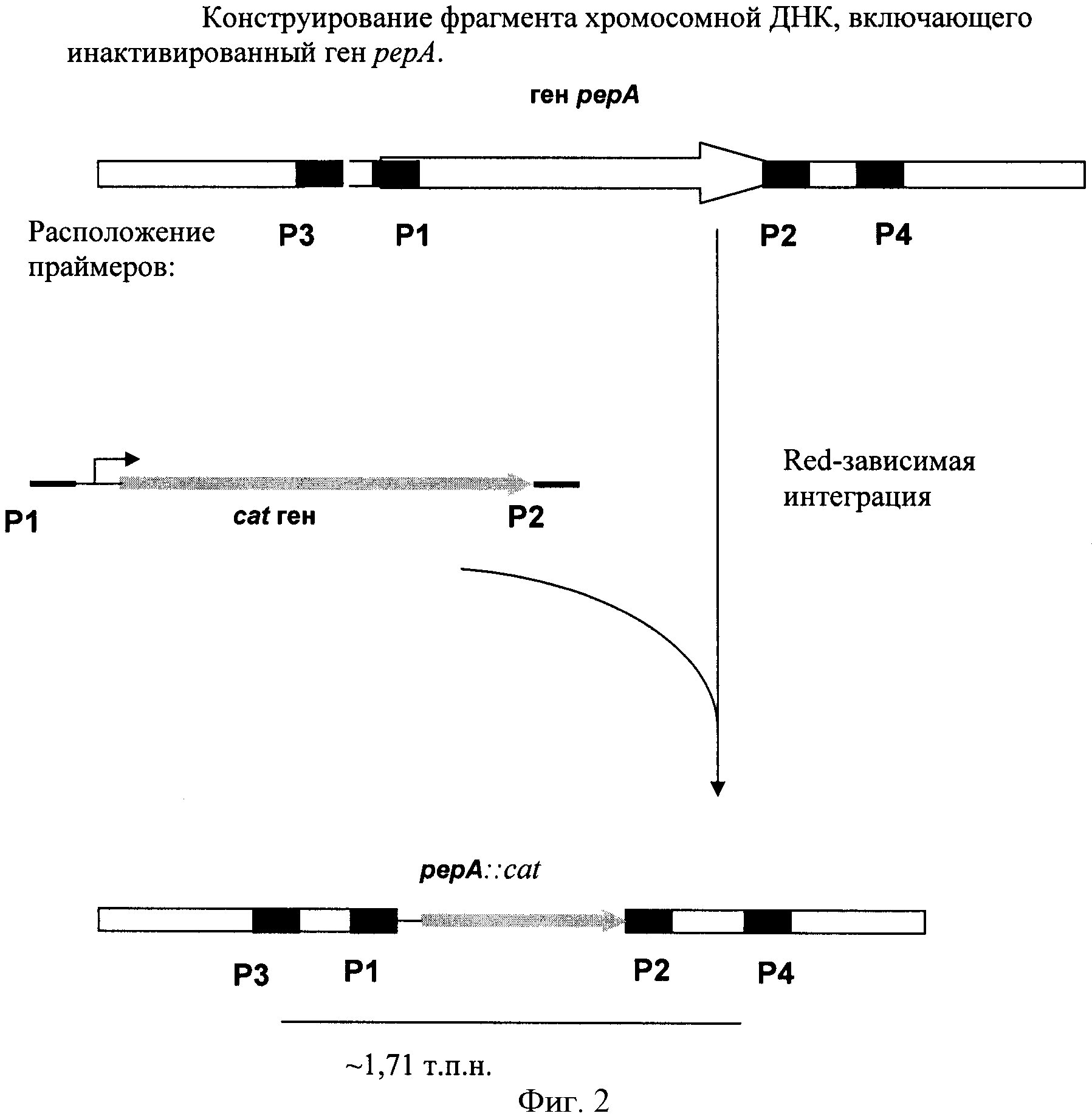
На Фигуре 2 показано конструирование фрагмента хромосомной ДНК, включающего инактивированный ген рерА.
Примеры
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение Примеры.
Пример 1. Конструирование штамма с инактивированным геном рерА.
1. Деления гена рерА.
Штамм, содержащий делецию гена рерА, был сконструирован с использованием методики, разработанной Datsenko, K.А. и Wanner, B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97(12), 6640-6645), известной как "Red-зависимая интеграция". В соответствии с этой процедурой были сконструированы праймеры для ПЦР P1 (SEQ ID NO: 3) и Р2 (SEQ ID NO: 4), гомологичные областям, прилегающим к гену рерА и к гену устойчивости к антибиотику на плазмиде, использованной в качестве матрицы для ПЦР. В качестве матрицы для ПЦР использовали плазмиду pMW118-attL-Cm-attR (WO 05/010175). Использовали следующий температурный профиль для ПЦР: денатурация при 94°С в течение 30 сек; последующие 25 циклов: денатурация - 30 сек при 94°С, отжиг - 30 сек при 55°С, элонгация - 1 мин 30 сек при 72°С; и заключительная полимеризация: 2 мин при 72°С.
Полученный ПЦР-продукт длиной 1.7 т.п.н. (Фиг.1) очищали в агарозном геле и использовали для электропорации в штамм Е. coli MG1655 (АТСС 700926), содержащий плазмиду pKD46 с термочувствительным репликоном. Плазмида pKD46 (Datsenko, K.A. and Wanner, B.L., Proc. Natl. Acad. Sci. USA, 2000, 97(12):6640-6645) содержит ДНК-фрагмент фага λ длиной 2154 п.н. (позиции с 31088 по 33241 нуклеотидной последовательности с инвентарным номером J02459 в базе данных GenBank), а также содержит гены λ Red-гомологичной системы рекомбинации (гены γ, β, exo) под контролем промотора ParaB, индуцируемого арабинозой. Плазмида pKD46 необходима для интеграции продукта ПЦР в хромосому штамма MG1655.
Электрокомпетентные клетки были получены следующим образом: ночную культуру штамма Е. coli MG1655 выращивали при 30°С в среде LB с добавкой ампициллина (100 мг/л), разводили в 100 раз, добавив 5 мл среды SOB (Sambrook et al, "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press, 1989), содержащей ампициллин и L-арабинозу (1 мМ). Полученную культуру растили с перемешиванием при 30°С до достижения OD600≈0.6, после чего делали клетки электрокомпетентными путем концентрирования в 100 раз и трехкратного отмывания ледяной деионизированной H2O. Электропорацию проводили с использованием 70 мкл клеток и ≈100 нг ПЦР-продукта. После электропорации клетки инкубировали в 1 мл среды SOC (Sambrook et al, "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press, 1989) при 37°C в течение 2.5 часов, после чего высевали на чашки с L-агаром, содержащим 30 мкг/мл хлорамфеникола и выращивали при 37°С для отбора CmR-рекомбинантов. Затем для удаления плазмиды pKD46 проводили 2 пассажа на L-агаре с Cm при 42°С и полученные колонии проверяли на чувствительность к ампициллину.
2. Подтверждение делеции гена рерА с помощью ПЦР.
Мутанты с делегированным геном рерА, содержащие ген устойчивости Cm, были проверены с помощью ПЦР с использованием локусспецифичных праймеров Р3 (SEQ ID NO:5) и Р4 (SEQ ID NO:6). Свежевыращенные колонии суспендировали в 20 мкл воды, и 1 мкл полученной суспензии использовали в ПЦР. Условия для ПЦР были следующие: денатурация при 94°С в течение 30 сек; профиль для 30 циклов: денатурация - 30 сек при 94°С, отжиг - 30 сек при 55°С, элонгация - 2 мин при 72°С; заключительная элонгация: 2 мин при 72°С. Длина продукта ПЦР, полученного в результате реакции с использованием в качестве матрицы клеток родительского штамма рерА+MG1655, составляет 1.65 т.п.н. Длина продукта ПЦР, полученного в результате реакции с использованием в качестве матрицы клеток мутантного штамма MG1655 ΔpepA::cat, составляет 1.71 т.п.н. (Фиг.2).
3. Удаление гена устойчивости к Cm (гена cat) из хромосомы штамма Е.coli MG1655 ΔpepA::cat
Ген устойчивости к Cm удаляли из хромосомы штамма Е. coli MG1655 pepA::cat с использованием int-xis системы. Для этого штамм Е. coli MG1655 pepA::cat трансформировали плазмидой pMWts-Int/Xis (международная заявка РСТ 05/010175). Колонии трансформантов отбирали на среде LB, содержащей 100 мкг/мл ампициллина. Чашки инкубировали в течение ночи при 30°С. Клоны трансформанотов излечивали от гена cat и плазмиды pMWts-Int/Xis путем получения отдельных клонов при 37°С (при этой температуре репрессор Cits частично инактивируется, и дерепрессируется транскрипция генов int/xis) с последующим отбором вариантов. Удаление гена cat из хромосомы указанного штамма подтверждали методом ПЦР с использованием локусспецифических праймеров Р3 (SEQ ID NO:5) и Р4 (SEQ ID NO:6). Условия для ПЦР были следующие: денатурация при 94°С в течение 30 сек; профиль для 30 циклов: денатурация - 30 сек при 94°С, отжиг - 30 сек при 55°С, элонгация - 2 мин при 72°С; заключительная элонгация: 2 мин при 72°С. Длина продукта ПЦР, полученного в результате реакции с использованием в качестве матрицы клеток штамма MG1655ΔpepA::cat, составляет 1.71 т.п.н. Длина продукта ПЦР, полученного в результате реакции с использованием в качестве матрицы клеток без гена cat, составляет 160 п.н. Таким образом был получен штамм с инактивированным геном рерА и удаленным геном cat.
Пример 2. Продукция L-аргинина штаммом Е. coli 382ΔрерА.
Для оценки влияния инактивации гена рерА на продукцию L-аргинина ДНК-фрагменты хромосомы описанного выше штамма Е.coli MG1655 ΔрерА были перенесены в штамм-продуцент L-аргинина Е.coli 382 с помощью P1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY) для получения штамма Е.coli 382 ΔрерА.
Штаммы Е. coli 382 и 382-ΔargT-hisP выращивали с перемешиванием при 37°С в течение 18 часов в 3 мл питательного бульона, по 0.3 мл полученных культур вносили в 3 мл ферментационной среды в пробирки размером 20×200 мм, и культуры выращивали при 32°С в течение 48 часов на роторной качалке.
После выращивания количество накопленного в среде L-аргинина определяли с помощью бумажной хроматографии, при этом использовали следующий состав подвижной фазы: бутанол: уксусная кислота: вода=4:1:1 (v/v). Раствор нингидрина (2%) в ацетоне использовали для визуализации. Пятно, содержащее L-аргинин, вырезали; L-аргинин элюировали 0.5% водным раствором CdCl2, после чего количество L-аргинина определяли спектрофотометрическим методом при длине волны 540 нм.
Состав ферментационной среды (г/л):
Глюкозу и сульфат магния стерилизовали раздельно. СаСО3 стерилизовали сухим жаром при 180°С в течение 2 часов. pH доводили до 7.0.
Результаты независимых пробирочных ферментации представлены в Таблице 1. Как следует из Таблицы 1, штамм с инактивированным геном рерА накапливал большее количество L-аргинина, чем родительский штамм-продуцент L-аргинина Е. coli 382.
Пример 3. Продукция L-цитруллина штаммом Е. coli 382ΔargGΔpepA
Для оценки влияния инактивации гена рерА на продукцию цитруллина ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔрерА могут быть перенесены в штамм-продуцент L-цитруллина Е. coli 382ΔargG с помощью Р1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY) с целью получения штамма 382AargGApepA. Штамм 382ΔargG может быть получен в результате делеции гена argG на хромосоме штамма 382 (ВКПМ В-7926) с использованием метода, предложенного Datsenko, K.A. and Wanner, B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97(12), 6640-6645), называемого "Red-зависимая интеграция". В соответствии с этой процедурой могут быть сконструированы ПЦР-праймеры, гомологичные как области, прилегающей к гену рерА, так и гену, отвечающему за устойчивость к антибиотику, на плазмиде, использующейся в качестве матрицы для ПЦР. В качестве матрицы в ПЦР может быть использована плазмида pMW118-attL-Cm-attR (WO 05/010175).
Оба штамма Е.coli, 382ΔargG и 382ΔargGΔpepA, могут быть выращены с перемешиванием при 37°С в течение 18 часов в 3 мл питательной среды, и 0.3 мл полученных культур могут быть перенесены в 2 мл ферментационной среды в пробирках 20×200-мм и могут быть культивированы при 32°С в течение 48 часов на роторной качалке.
После выращивания количество полученного цитруллина может быть определено с помощью бумажной хроматографии с использованием подвижной фазы следующего состава: бутанол-уксусная кислота-вода=4:1:1. Последующее окрашивание может быть выполнено нингидрином (2% раствор в ацетоне). Содержащее цитруллин пятно может быть вырезано, цитруллин может быть элюирован с использованием 0.5% водного раствора CdCl2, и количество цитруллина может быть определено спектрофотометрически при длине волны 540 нм.
Возможный состав ферментационной среды (г/л):
Глюкозу и сульфат магния стерилизуют отдельно. СаСО3 стерилизуют сухим жаром при 180°С в течение 2 часов. Значение pH доводят до 7.0.
Хотя указанное изобретение описано в деталях со ссылкой на наилучший способ осуществления изобретения, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
Каждому из упомянутых выше документов соответствует ссылка, и все цитируемые документы являются частью описания настоящего изобретения.
Реферат
Изобретение относится к области биотехнологии. Описана бактерия-продуцент L-аргинина и L-цитруллина, принадлежащая к роду Escherichia, модифицированная таким образом, что экспрессия гена pepА, кодирующего аминопептидазу А, ослаблена. Предложен способ получения L-аминокислоты, выбранной из L-аргинина и L-цитруллина, включающий выращивание указанной модифицированной бактерии в питательной среде для продукции и секреции указанной L-аминокислоты в культуральную жидкость и выделение указанной L-аминокислоты из культуральной жидкости. Изобретение позволяет получить L-аргинин и L-цитруллин в увеличенном объеме. 2 н. и 1 з.п. ф-лы, 2 ил., 1 табл., 3 пр.
Комментарии