Продукция гликопротеинов с модифицированным фукозилированием - RU2479629C2
Код документа: RU2479629C2
Чертежи


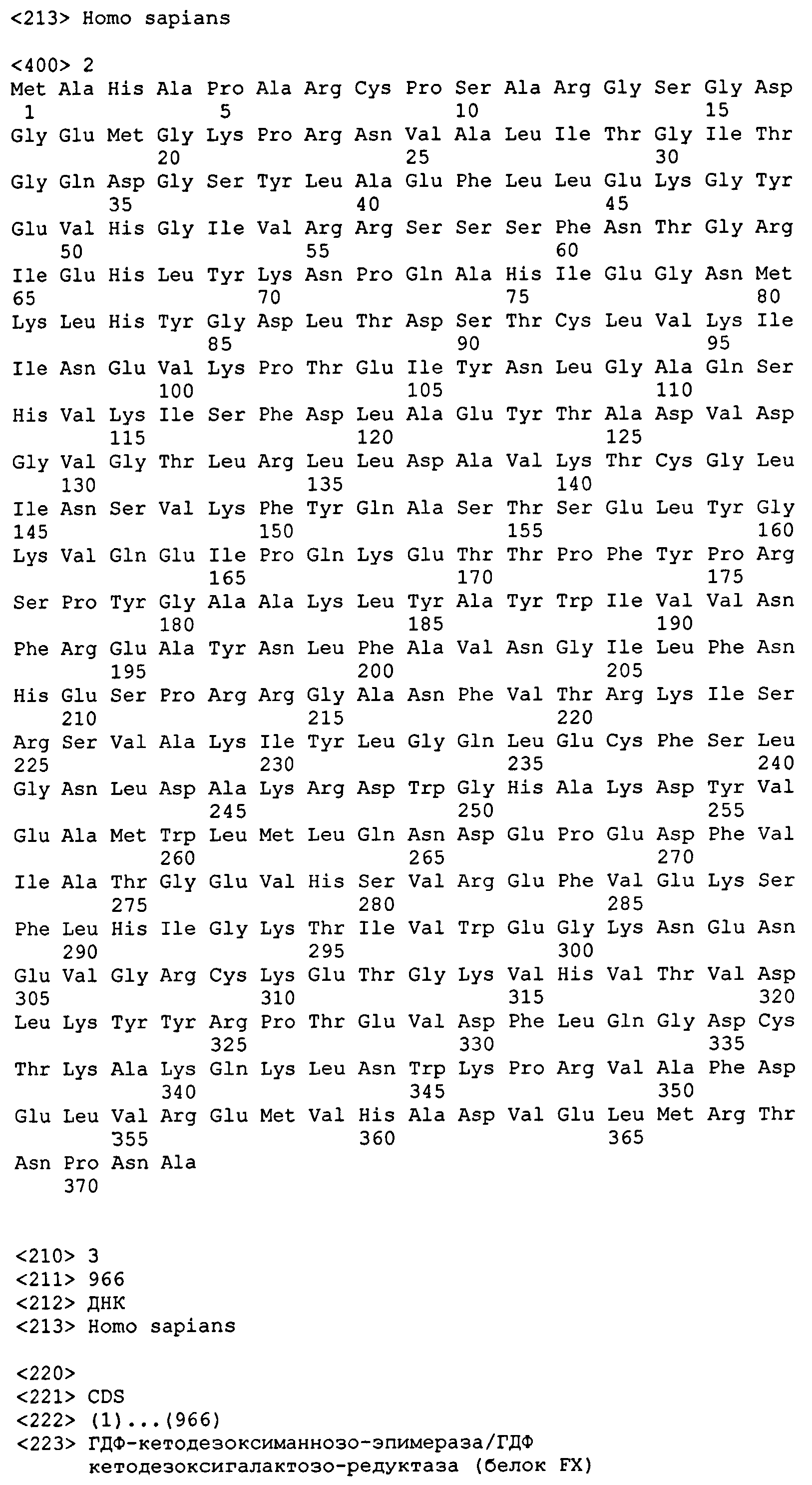






















Описание
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
(1) Область техники
Настоящее изобретение относится к области гликобиологии, в частности к способам создания методами генетической инженерии клеток-хозяев, в которых отсутствует эндогенный каскад реакций фукозилирования N-гликанов гликопротеинов, с приданием способности продуцировать гликопротеины с фукозилированными N-гликанами.
(2) Описание известного уровня техники
Предназначенные для применения для людей терапевтические белки, которые являются гликозилированными, должны иметь сложные специфические для людей характеры N-гликозилирования. Обычно могло бы быть выгодным продуцирование терапевтических белков, используя бактериальные или эукариотические микроорганизмы, из-за (а) возможности быстрого продуцирования высоких концентраций белка; (b) возможности использования стерильных, хорошо контролируемых условий продукции (например, условий ГМФ); (c) возможности использования простых, химически определенных сред для роста; (d) легкости генетической манипуляции; (e) отсутствия заражения патогенами людей или животных; (f) возможности экспрессии широкого выбора белков, в том числе белков, плохо экспрессируемых в культуре клеток вследствие токсичности и т.п.; и (g) легкости извлечения белка (например, через секрецию в среду для культивирования). Однако прокариоты и низшие эукариоты обычно не продуцируют белки, имеющие сложные характеры N-гликозилирования. Поэтому для продуцирования терапевтических белков, как правило, используют клетки животных, если желательно, чтобы белок имел сложный характер N-гликозилирования, напоминающий характер N-гликозилирования у людей. Но существует ряд значительных недостатков в использовании клеток животных для продуцирования терапевтических белков.
Только некоторые терапевтические белки подходят для экспрессии в клетках животных (например, белки, не обладающие каким-либо цитотоксическим эффектом или другим эффектом, неблагоприятным для роста). Системы культивирования клеток животных являются обычно очень медленными, часто требуя свыше одной недели роста при тщательно контролируемых условиях для продуцирования какого-либо полезного количества представляющего интерес белка. Выходы белка, тем не менее, трудно сопоставляются с выходами белка при способах ферментации в микроорганизмах. Кроме того, для систем культивирования клеток обычно требуются сложные и дорогостоящие питательные вещества и кофакторы, такие как бычья фетальная сыворотка. Более того, рост может ограничиваться запрограммированной гибелью клеток (апоптозом).
Более того, клетки животных (в частности, клетки млекопитающих) в высокой степени чувствительны к вирусной инфекции или заражению. В некоторых случаях вирус или другой инфекционный агент может подвергать риску рост культуры, в то время как в других случаях агент может быть патогенном человека, превращающим продукт в виде терапевтического белка в продукт, негодный для его намеченного применения. Кроме того, для способов культивирования многих клеток требуется использование сложных, термочувствительных, происходящих из животных компонентов сред для роста, которые могут нести патогены, такие как прионы губчатой энцефалопатии крупного рогатого скота. Такие патогены трудно обнаружить и/или трудно удалить или полностью уничтожить без подвергания риску среды для роста. В любом случае использование клеток животных для продуцирования терапевтических белков делает необходимыми дорогие контроли качества для гарантии безопасности продукта.
Недавно было установлено, что низшие эукариоты, в частности дрожжи, можно генетически модифицировать, так чтобы они экспрессировали белки, имеющие сложные характеры N-гликозилирования, которые напоминают характер N-гликозилирования у людей или являются гуманизированными. Такие генетически модифицированные низшие эукариоты можно получить устранением выбранных эндогенных ферментов гликозилирования, которые вовлечены в продукцию N-гликанов с высоким содержанием маннозы, и введением различных комбинаций экзогенных ферментов, вовлеченных в создание сложных N-гликанов. Способы создания методами генетической инженерии дрожжей, которые продуцируют сложные N-гликаны, описаны в патенте США № 7029872 и опубликованных заявках на патенты США № 2004/0018590, 2005/0170452, 2006/0286637, 2004/0230042, 2005/0208617, 2004/0171826, 2005/0208617 и 2006/0160179. Например, можно выбрать или создать клетку-хозяина, истощенную по 1,6-маннозилтрансферазным активностям, которые в противном случае могли бы добавлять остатки маннозы на N-гликан на гликопротеине, и затем создать клетку-хозяина, включающую каждый из ферментов, вовлеченных в продукцию сложных N-гликанов, напоминающих N-гликаны человека.
Клетки животных и человека имеют каскад реакций с участием фукозилтрансферазы, который добавляет остаток фукозы к остатку GlcNAc (N-ацетилглюкозамина) на восстанавливающем конце N-гликанов на белке. Каскад реакций фукозилирования у людей состоит из ГДФ-маннозо-дегидратазы и ГДФ-кетодезоксиманнозо-эпимеразы/ГДФ-кетодезоксигалактозо-редуктазы (белка FX), обе из которых находятся в цитоплазме, которые, действуя координированным образом с общей целью, превращают ГДФ-маннозу в ГДФ-фукозу; переносчика ГДФ-фукозы, находящегося в мембране аппарата Гольджи, который переносит ГДФ-фукозу в аппарат Гольджи, и фукозилтрансферазы (Fut8), которая переносит остаток фукозы посредством образования 1,6-связи в 6 положение остатка GlcNAc на восстанавливающем конце N-гликана. В противоположность высшим эукариотам у многих низших эукариот, например дрожжей, отсутствуют ферменты, вовлеченные в каскад реакций фукозилирования, они продуцируют гликопротеины, которые не содержат фукозу (смотри, например, Bretthauer/Catellino, Biotechnol. Appl. Biochem. 30: 193-200 (1999); Rabina et al., Anal. Biochem. 286: 173-178 (2000)). Однако было установлено, что отсутствие фукозы на гликопротеинах выгодно в некоторых случаях. Например, при продукции моноклональных антител, молекул иммуноглобулинов, и родственных молекул было установлено, что удаление сахара фукозы с N-гликана иммуноглобулинов увеличивает или изменяет их связывание с выбранными рецепторами Ig, что вызывает изменения свойств, таких как антителозависимая клеточноопосредованная цитотоксичность или ADCC (смотри, например, опубликованные заявки на патенты США № 2005/0276805 и US2003/0157108).
Однако, хотя удаление фукозы с N-гликанов иммуноглобулинов, по-видимому, увеличивает активность ADCC, фукозилированные N-гликаны оказываются важными для других гликопротеинов. Например, делеция гена фукозилтрансферазы у мышей вызывает серьезное замедление роста, раннюю смерть во время постнатального развития и напоминающие эмфизему изменения в легком. Эти Fut8-/--нулевые мыши были избавлены от эмфиземаподобного фенотипа путем введения экзогенного TGF-бета 1. Кроме того, нарушенная активация, опосредуемая рецептором, была устранена введением снова гена Fut8, что демонстрирует, что фукозилирование остова является очень важным для правильного функционирования рецепторов факторов роста, таких как TGF-бета 1 и EGF (Wang et al., Meth. Enzymol. 417: 11-22 (2006)). В ткани легкого, происходящей из мышей Fut8-/-, утрата фукозилирования остова нарушает функционирование родственного рецепторам липопротеинов низкой плотности (LDL) белка-1 (LRP-1), что приводит к уменьшению эндоцитоза связывающего инсулиноподобный фактор роста (IGF) белка-3 (IGFBP-3) (Lee et al., J. Biochem. (Tokyo) 139: 391-8 (2006)). Для клеток эмбриональных фибробластов мышей Fut8-/-опосредуемая интегрином α3β1 миграция клеток отменяется и снижается активация клеток, устанавливая, что остовая фукоза является существенной для функционирования белков (Zhao et al., J. Biol. Chem., 281: 38343-38350 (2006)). Кроме того, могут быть ситуации, при которых желательно продуцирование композиций антител, в которых по крайней мере часть антител являются фукозилированными для увеличения активности ADCC. Поэтому в особых случаях будет полезным обеспечение являющихся низшими эукариотами организмов и клеток, способных продуцировать фукозилированные гликопептиды. Соответственно, разработка способов и материалов для получения клеток-хозяев, являющихся низшими эукариотами, таких как грибы и дрожжи, и, в частности, таких дрожжей, как Pichia pastoris, K. lactis и другие, могла бы способствовать разработке генетически усиленных штаммов дрожжей для рекомбинантной продукции фукозилированных гликопротеинов.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Соответственно, настоящим изобретением обеспечиваются способы и материалы для создания систем экспрессии в низших эукариотах, которые можно использовать для продукции рекомбинантных, фукозилированных гликопротеинов. В частности, обеспечиваются векторы, содержащие гены, кодирующие один или несколько ферментов, вовлеченных в каскад реакций гликозилирования у млекопитающих, и клетки-хозяева, являющиеся низшими эукариотами, которые были трансформированы векторами для получения клеток-хозяев, которые способны продуцировать фукозилированные гликопротеины. Векторы, клетки-хозяева и способы особенно хорошо адаптированы к применению в экспрессионных системах, основанных на клетках-хозяевах, являющихся дрожжами и грибами, таких как Pichia pastoris.
В одном варианте осуществления настоящим изобретением обеспечиваются способы и материалы для трансформации клеток-хозяев, являющихся низшими эукариотами, одним или несколькими векторами, кодирующими ферментативные активности для превращения ГДФ-маннозы в ГДФ-фукозу и для присоединения фукозы к N-гликану, продуцируемому клеткой-хозяином. В дальнейших вариантах осуществления настоящее изобретение включает гибридные векторы, кодирующие слитый белок, включающий каталитический домен фермента каскада реакций фукозилирования, слитый с неприродной лидерной последовательностью, которая кодирует последовательность для направленной доставки, которая направляет слитый пептид в соответствующее положение в эндоплазматическом ретикулуме, аппарате Гольджи на ранней стадии или аппарате Гольджи на поздней стадии. Например, каталитический домен фукозилтрансферазы сливают с лидерным пептидом, который направляет каталитический домен в положение внутри эндоплазматического ретикулума, аппарата Гольджи на ранней стадии или аппарата Гольджи на поздней стадии. В дальнейших вариантах осуществления клетку-хозяина, являющегося низшим эукариотом, трансформируют вектором, кодирующим ГДФ-фукозо-трансферазу, которая переносит ГДФ-фукозу из цитоплазмы внутрь аппарата Гольджи.
Настоящим изобретением обеспечивается рекомбинантная клетка-хозяин, являющийся низшим эукариотом, включающая каскад реакций фукозилирования. В конкретных аспектах клеткой-хозяином являются дрожжи или нитчатый гриб, например дрожжи Pichia sp., такие как Pichia pastoris.
В дальнейших аспектах клетка-хозяин, кроме того, не демонстрирует активность α1,6-маннозилтрансферазы по отношению к N-гликану на гликопротеине и включает каталитический домен α1,2-маннозидазы, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности α1,2-маннозидазы в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина, посредством чего при прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу Man5GlcNAc2.
В дальнейших аспектах вышеуказанная клетка-хозяин, кроме того, включает каталитический домен GlcNAc-трансферазы I, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности GlcNAc-трансферазы I в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина, посредством чего при прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу GlcNAcMan5GlcNAc2.
В дальнейших аспектах вышеуказанная клетка-хозяин, кроме того, включает каталитический домен маннозидазы II, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности маннозидазы II в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина, посредством чего при прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу GlcNAcMan3GlcNAc2.
В дальнейших аспектах вышеуказанная клетка-хозяин, кроме того, включает каталитический домен GlcNAc-трансферазы II, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности GlcNAc-трансферазы II в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина, посредством чего при прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу GlcNAc2Man3GlcNAc2.
В дальнейших аспектах вышеуказанная клетка-хозяин, кроме того, включает каталитический домен галактозотрансферазы II, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности галактозотрансферазы II в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина, посредством чего при прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу Gal2GlcNAc2Man3GlcNAc2.
В дальнейших аспектах вышеуказанная клетка-хозяин, кроме того, включает каталитический домен сиалилтрансферазы, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности сиалилтрансферазы в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина, посредством чего при прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу NANA2Gal2GlcNAc2Man3GlcNAc2.
Трансформируя вышеуказанные клетки-хозяева нуклеиновой кислотой, кодирующей конкретный гликопротеин, можно продуцировать композиции гликопротеинов, которые включают множество гликоформ, при этом каждая гликоформа включает по крайней мере один присоединенный к ней N-гликан, причем композиция гликопротеинов, тем самым, включает множество N-гликанов, в котором преобладающая гликоформа включает желаемый фукозилированый N-гликан. В зависимости от конкретного желаемого гликопротеина можно использовать способы настоящего изобретения для получения композиций гликопротеинов, в которых преобладающая N-гликоформа присутствует в количестве, превышающем на 5-80 мольных процента количество следующей самой преобладающей N-гликоформы; в дальнейших вариантах осуществления преобладающая N-гликоформа может присутствовать в количестве, превышающем на 10-40 мольных процента, 20-50 мольных процента, 30-60 мольных процента, 40-70 мольных процента, 50-80 мольных процента количество следующей самой преобладающей N-гликоформы. В других вариантах осуществления преобладающая N-гликоформа является желаемой фукозилированной N-гликоформой и присутствует в количестве, составляющем более 25 мольных процентов, более 35 мольных процентов, более 50 мольных процентов, более 60 мольных процентов или более 70 мольных процентов от общего количества N-гликанов.
Таким образом, обеспечиваются клетки-хозяева для продуцирования композиций гликопротеинов, включающих множество гликоформ, при этом каждая гликоформа включает по крайней мере один присоединенный к ней N-гликан, причем композиция гликопротеинов, тем самым, включает множество N-гликанов, в котором преобладающий N-гликан выбирают из группы, состоящей из Man5GlcNAc2, GlcNAcMan5GlcNAc2, Man3GlcNAc2, GlcNAcMan3GlcNAc2, GlcNAc2Man3GlcNAc2, GalGlcNAc2Man3GlcNAc2, Gal2GlcNAc2Man3GlcNAc2, NANAGal2GlcNAc2Man3GlcNAc2, и NANA2Gal2GlcNAc2Man3GlcNAc2.
В дальнейших аспектах более чем 25 мольных процентов множества фукозилированных N-гликанов состоит, по существу, из фукозилированной гликоформы, которую выбирают из группы, состоящей из Man5GlcNAc2, GlcNAcMan5GlcNAc2, Man3GlcNAc2, GlcNAcMan3GlcNAc2, GlcNAc2Man3GlcNAc2, GalGlcNAc2Man3GlcNAc2, Gal2GlcNAc2Man3GlcNAc2, NANAGal2GlcNAc2Man3GlcNAc2 и NANA2Gal2GlcNAc2Man3GlcNAc2.
Во все еще дальнейших аспектах более чем 25 мольных процентов, более чем 35 мольных процентов, более чем 50 мольных процентов, более чем 60 мольных процентов, более чем 75 мольных процентов или более чем 90 мольных процентов множества N-гликанов состоит, по существу, из фукозилированной гликоформы, которую выбирают из группы, состоящей из Man5GlcNAc2, GlcNAcMan5GlcNAc2, Man3GlcNAc2, GlcNAcMan3GlcNAc2, GlcNAc2Man3GlcNAc2, GalGlcNAc2Man3GlcNAc2, Gal2GlcNAc2Man3GlcNAc2, NANAGal2GlcNAc2Man3GlcNAc2 и NANA2Gal2GlcNAc2Man3GlcNAc2.
В вышеуказанной композиции гликопротеинов фукоза находится в α1,3-связи с GlcNAc на восстанавливающем конце N-гликана, α1,6-связи с GlcNAc на восстанавливающем конце N-гликана, α1,2-связи с Gal на невосстанавливающем конце N-гликана, α1,3-связи с GlcNAc на невосстанавливающем конце N-гликана или α1,4-связи с GlcNAc на невосстанавливающем конце N-гликана.
Следовательно, в конкретных аспектах вышеуказанных композиций гликопротеинов фукоза гликоформы находится в α1,3-связи или α1,6-связи с образованием гликоформы, выбираемой из группы, состоящей из Man5GlcNAc2(Fuc), GlcNAcMan5GlcNAc2(Fuc), Man3GlcNAc2(Fuc), GlcNAcMan3GlcNAc2(Fuc), GlcNAc2Man3GlcNAc2(Fuc), GalGlcNAc2Man3GlcNAc2(Fuc), Gal2GlcNAc2Man3GlcNAc2(Fuc), NANAGal2GlcNAc2Man3GlcNAc2(Fuc) и NANA2Gal2GlcNAc2Man3GlcNAc2(Fuc); в α1,3-связи или α1,4-связи с образованием гликоформы, выбираемой из группы, состоящей GlcNAc(Fuc)Man5GlcNAc2, GlcNAc(Fuc)Man3GlcNAc2, GlcNAc2(Fuc1-2)Man3GlcNAc2, GalGlcNAc2(Fuc1-2)Man3GlcNAc2, Gal2GlcNAc2(Fuc1-2)Man3GlcNAc2, NANAGal2GlcNAc2(Fuc1-2)Man3GlcNAc2и NANA2Gal2GlcNAc2(Fuc1-2)Man3GlcNAc2; или в α1,2-связи с образованием гликоформы, выбираемой из группы, состоящей Gal(Fuc)GlcNAc2Man3GlcNAc2, Gal2(Fuc1-2)GlcNAc2Man3GlcNAc2, NANAGal2(Fuc1-2)GlcNAc2Man3GlcNAc2и NANA2Gal2(Fuc1-2)GlcNAc2Man3GlcNAc2.
В других аспектах композиции гликопротеинов настоящего изобретения включают композиции, в которых вышеуказанная N-гликоформа присутствует на уровне, превышающем на приблизительно 5-80 мольных процентов, 10-40 мольных процентов, 20-50 мольных процентов, 30-60 мольных процентов, 40-70 мольных процентов или 50-80 мольных процентов уровень следующей самой преобладающей N-гликоформы.
Определения
Используемые здесь термины «N-гликан» и «гликоформа» используются взаимозаменяемо и относятся к N-связанному олигосахариду, например олигосахариду, который присоединен с помощью связи аспарагин-N-ацетилглюкозамин к остатку аспарагина полипептида. N-связанные гликопротеины содержат остаток N-ацетилглюкозамина, связанный с амидным азотом остатка аспарагина в белке. Преобладающими сахарами, обнаруженными на гликопротеинах, являются глюкоза (Glc), галактоза (Gal), манноза (Man), фукоза (Fuc), N-ацетилгалактозамин (GalNAc), N-ацетилглюкозамин (GlcNAc) и сиаловая кислота (например, N-ацетилнейраминовая кислота (NANA)). Процессирование сахарных групп происходит котрансляционно в просвете эндоплазматического ретикулума и продолжается в аппарате Гольджи для N-связанных гликопротеинов.
N-гликаны имеют общий пентасахаридный остов Man3GlcNAc2. N-гликаны отличаются по числу ветвей (антенн), включающих периферические сахара (например, GlcNAc, галактозу, фукозу и сиаловую кислоты), которые добавляются к остовой структуре Man3GlcNAc2, которая также упоминается как «триманнозный остов», «пентасахаридный остов» или «пауциманнозный остов». N-гликаны классифицируют в соответствии с их разветвленными составными частями (например, с высоким содержанием маннозы, сложные или гибридные). Тип N-гликана «с высоким содержанием маннозы» имеет пять или более остатков маннозы. N-гликан «сложного» типа обычно имеет по крайней мере один GlcNAc, присоединенный к маннозному плечу «триманнозного» остова с помощью 1,3-связи, и по крайней мере один GlcNAc, присоединенный к маннозному плечу «триманнозного» остова с помощью 1,6-связи. Сложные N-гликаны могут также иметь остатки галактозы и N-ацетилгалактозамина, которые необязательно модифицированы сиаловой кислотой или производными (например, «NANA» или «NeuAc», где «Neu» относится к нейраминовой кислоте, а «Ac» относится к ацетилу). Сложные N-гликаны могут также иметь внутрицепочечные замены, включающие «биссекторный» GlcNAc и остовую фукозу («Fuc»). В качестве примера, когда N-гликан включает биссекторный GlcNAc в триманнозном остове, структуру можно представить как Man3GlcNAc2(GlcNAc) или Man3GlcNAc3. Когда N-гликан включает остовую фукозу, присоединенную к триманнозному остову, структуру можно представить как Man3GlcNAc2(Fuc). Сложные N-гликаны могут также иметь множественные антенны на «триманнозном остове», часто упоминаемые как «множественные антенные гликаны». «Гибридный» N-гликан имеет по крайней мере один GlcNAc, присоединенный к крайней маннозе маннозного плеча триманнозного остова с помощью 1,3-связи, и ноль или более манноз, присоединенных к маннозному плечу триманнозного остова с помощью 1,6-связи. Различные N-гликаны также упоминаются как «гликоформы».
Используемые здесь сокращения являются обычно используемыми в данной области техники сокращениями, смотри, например, сокращения сахаров выше. Другие обычные сокращения включают «PNGase», или «гликаназу», или «глюкозидазу, все из которых относятся к пептид-N-гликозидазе F (EC 3.2.2.18).
Используемый здесь термин «контролирующая экспрессию последовательность» относится к полинуклеотидным последовательностям, которые необходимы для оказания влияния на экспрессию кодирующих последовательностей, с которыми они функционально связаны. Контролирующие экспрессию последовательности являются последовательностями, которые контролируют транскрипцию, посттранскрипционные события и трансляцию последовательностей нуклеиновых кислот. Контролирующие экспрессию последовательности включают последовательность для соответствующей инициации, терминации транскрипции, промотор и последовательности - энхансеры, действенные сигналы процессирования РНК, такие как сигналы сплайсинга и полиаденилирования, последовательности, которые стабилизируют цитоплазматическую мРНК, последовательности, которые усиливают эффективность трансляции (например, сайты связывания рибосом), последовательности, которые усиливают стабильность белка, и, при желании, последовательности, которые усиливают секрецию белка. Природа таких контролирующих последовательностей различается в зависимости от организма хозяина, у прокариот такие контролирующие последовательности, как правило, включают промотор, сайт связывания рибосом и последовательность для терминации транскрипции. Подразумевается, что термин «контролирующие последовательности» включает, как минимум, все компоненты, присутствие которых необходимо для экспрессии, и также может включать дополнительные компоненты, присутствие которых является выгодным, например лидерные последовательности и последовательности партнеров по слиянию.
Подразумевается, что используемый здесь термин «рекомбинантная клетка-хозяин» («клетка-хозяин для экспрессии», «система хозяина для экспрессии», «экспрессионная система» или просто «клетка-хозяин») относится к клетке, в которую введен рекомбинантный вектор. Следует понимать, что такие термины, как подразумевается, относятся не только к конкретной рассматриваемой клетке, но также к потомству такой клетки. Поскольку определенные модификации могут происходить в последующих поколениях вследствие или мутации, или влияний окружающей среды, такое потомство может, в действительности, не быть идентичным родительской клетке, но все еще включаться в объем используемого здесь термина «клетка-хозяин». Рекомбинантная клетка-хозяин может быть выделенной клеткой или линией клеток, выращиваемой в культуре, или может быть клеткой, постоянно находящейся в живой ткани или организме.
Термин «эукариотическая» относится к содержащей ядро клетке или организму и включает клетки насекомых, клетки растений, клетки млекопитающих, клетки животных и клетки низших эукариот.
Термин «клетки низших эукариот» включает дрожжи, грибы, воротничковые жгутиконосцы, микроспоридии, альвеоляты (например, динофлагелляты), страменопилы (например, бурые водоросли, простейшие), родофиты (например, красные водоросли), растения (например, зеленые водоросли, клетки растений, мох) и другие одноклеточные организмы. Дрожжи и грибы включают, но без ограничения, Pichia pastoris, Pichia finlandica, Pichia trehalophila, Pichia koclamae, Pichia membranaefaciens, Pichia minuta (Ogataea minuta, Pichia lindneri), Pichia opuntiae, Pichia termotolerans, Pichia salictaria, Pichia guercuum, Pichia pijperi, Pichia stiptis, Pichia methanolica, Pichia sp., Saccharomyces cerevisiae, Saccharomyces sp., Hansenula polymorpha, Kluyveromyces sp., Kluyveromyces lactis, Candida albicans, Aspergillus nidulans, Aspergillus niger, Aspergillus oryzae, Trichoderma reesei, Chrysosporium lucknowense, Fusarium sp., Fusarium gramineum, Fusarium venenatum, Physcomitrella patens и Neurospora crassa, Pichia sp., любой Saccharomyces sp., Hansenula polymorpha, любой Kluyveromyces sp., Candida albicans, любой Aspergillus sp., Trichoderma reesei, Chrysosporium lucknowense, любой Fusarium sp., и Neurospora crassa.
Используемый здесь термин «пептид» относится к короткому полипептиду, например полипептиду, длина которого обычно меньше приблизительно 50 аминокислот и более обычно меньше приблизительно 30 аминокислот. Используемый здесь термин охватывает аналоги и миметики, которые воспроизводят структурную и, следовательно, биологическую функцию.
Предполагается, что используемый здесь термин «в преобладающей степени» или такие варианты, как «преобладающий» или «который является преобладающим», означают разновидность гликана, которая присутствует в наибольшем молярном проценте (%) общего количества N-гликанов после того, как гликопротеин был обработан N-гликозидазой и высвободившиеся гликаны проанализированы с помощью масс-спектрометрии, например MALDI-TOF MS. Другими словами, выражением «в преобладающей степени» определяется, что индивидуальный объект, такой как конкретная гликоформа, присутствует в большем молярном проценте, чем какой-либо другой индивидуальный объект. Например, если композиция состоит из разновидности А, присутствующей в 40 молярных процентах, разновидности В, присутствующей в 35 молярных процентах, и разновидности С, присутствующей в 25 молярных процентах, композиция включает в преобладающей степени разновидность А.
Кроме случаев, определенных особо, все технические и научные термины, используемые здесь, имеют значение, одинаковое со значением, в котором они обычно понимаются специалистом со средним уровнем компетентности в области техники, к которой относится это изобретение. Ниже описываются приводимые в качестве примеров способы и материалы, хотя при осуществлении на практике настоящего изобретения могут также использоваться способы и материалы, схожие с описанными здесь способами или материалами или эквивалентные им, и они будут очевидны квалифицированным в данной области техники специалистам. Все публикации и другие ссылки, упоминаемые здесь, полностью включены посредством ссылки. В конфликтной ситуации будут руководствоваться описанием настоящего изобретения, в том числе определениями. Материалы, способы и примеры являются только иллюстрацией и не означают ограничение.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 иллюстрируется каскад реакций фукозилирования, представленный во многих клетках высших эукариот.
На фиг. 2 демонстрируются стадии глико-инженерии, требуемые для получения рекомбинантных дрожжей, способных продуцировать фукозилированные гликопротеины. Эндогенная ГДФ-манноза, присутствующая в цитоплазме дрожжей, превращается в ГДФ-фукозу под действием ГДФ-маннозо-дегидратазы (GMD) и бифункционального фермента FX. Впоследствии продукт перемещается в аппарат Гольджи с помощью переносчика ГДФ-фукозы (GFTr), и фукоза переносится на акцепторный гликан с помощью α-1,6-фукозилтрансферазы (FUT8). Ферменты показаны голубым текстом, а промежуточные метаболические продукты - черным текстом. ГДФ-kdMan (ГДФ-4-кето-6-дезоксиманноза) и ГДФ-kdGal (ГДФ-4-кето-6-дезоксигалактоза) являются промежуточными продуктами при превращении ГДФ-маннозы в ГДФ-фукозу.
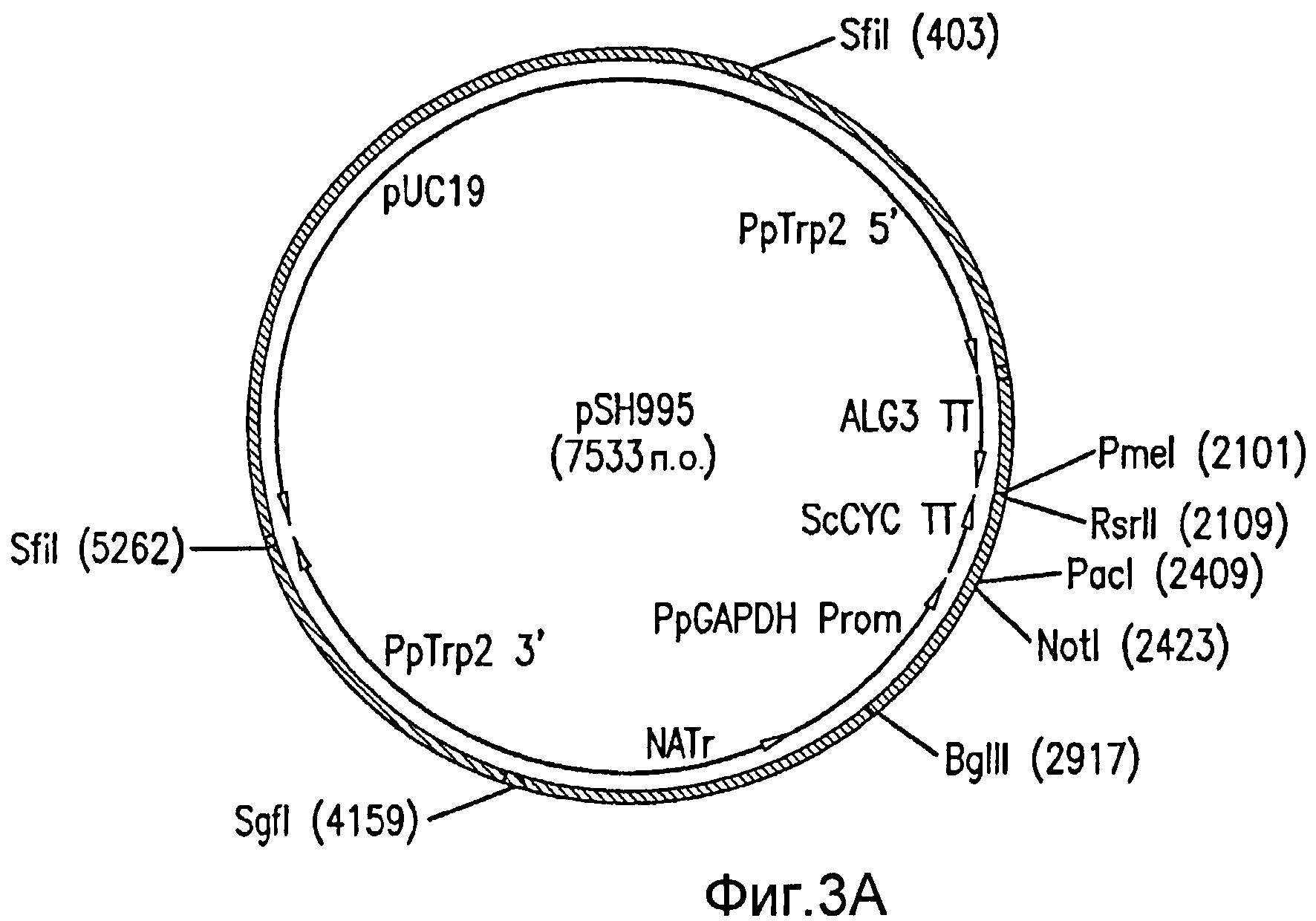
На фиг. 3А демонстрируются векторы, используемые при создании штаммов дрожжей, продуцирующих фукозилированные гликопротеины. Изображен экспрессионный вектор pSH995, в который введены гены биосинтеза и переноса фукозы. Введение генов, требуемых для биосинтеза и переноса фукозы, в pSH995 приводило к получению вектора pSH1022.
На фиг. 3В демонстрируется вектор pSH1022. В (В) показаны фланкирующие районы локусов TRP2, используемые для интеграции генов в геном Pichia; преобладающий селектируемый маркер NATr; экспрессирующая GAPDH-CYC кассета и основа плазмиды pUC19.
На фиг. 4А демонстрируется изображение, полученное при анализе MALDI-TOF N-гликанов, высвободившихся из EPO крысы, демонстрирующее, что штамм YSH661 Pichia pastoris (штамм RDP974, трансформированный вектором pSH1022, содержащим гены для каскада реакций фукозилирования) продуцировал rEPO, включающий N-гликаны Gal2GlcNAc2Man3GlcNAc2(Fuc) и Gal2GlcNAc2Man3GlcNAc2. N-гликаны Gal2GlcNAc2Man3GlcNAc2(Fuc) находятся внутри прямоугольника.
На фиг. 4В демонстрируется изображение, полученное при анализе MALDI-TOF N-гликанов, высвободившихся из EPO крысы, демонстрирующее, что контрольный штамм YSH660 (штамм RDP974, трансформированный контрольным вектором pSH995) продуцировал только rEPO с нефукозилированными N-гликанами или N-гликанами без фукозы Gal2GlcNAc2Man3GlcNAc2.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящим изобретением обеспечиваются способы и материалы для создания методами генетической инженерии клеток-хозяев, способных продуцировать белки гликопротеины, которые имеют фукозилированные N-гликаны. Хотя способы и материалы продемонстрированы с помощью примеров в дрожжах Pichia pastoris, которые не обладают эндогенным каскадом реакций фукозилирования, способы и материалы можно также использовать для создания методами генетической инженерии других низших эукариот, таких как грибы, прокариот и таких высших эукариот, которые не имеют эндогенного каскада реакций фукозилирования, например клеток насекомых. В других вариантах осуществления способы и материалы можно использовать для создания методами генетической инженерии клеток высших эукариот, которые не имеют эндогенного каскада реакций фукозилирования, но в которых желательно увеличить степень фукозилирования, присутствующую в гликопротеинах, продуцируемых такими клетками-хозяевами.
Как правило, способ настоящего изобретения включает получение клетки-хозяина, способной продуцировать фукозилированные гликопротеины, путем введения в клетку-хозяина нуклеиновых кислот, кодирующих такие ферменты или ферментативные активности, которые вовлечены в каскад реакций фукозилирования, которые при введении в клетку-хозяина будут предоставлять клетке способность продуцировать фукозилированные гликопротеины. Эти нуклеиновые кислоты включают, например, нуклеиновые кислоты, кодирующие ГДФ-маннозо-4,6-дегидратазную активность, ГДФ-кетодезоксиманнозо-эпимеразную активность/ГДФ-кетодезоксигалактозо-редуктазную активность, белок-переносчик ГДФ-фукозы и фукозилтрансферазную активность. Общий вид каскада реакций фукозилирования у высших эукариот показан на фиг. 1.
ГДФ-маннозо-4,6-дегидратаза (GMD) (EC 4.2.1.47), превращающая ГДФ-маннозу в ГДФ-4-кето-6-дезоксиманнозу в присутствии НАД, была идентифицирована в ряде видов. GMD человека (hGMD) кодируется нуклеотидной последовательностью, показанной в SEQ ID NO: 1, и имеет аминокислотную последовательность, показанную в SEQ ID NO: 2. Гомологичные гены с ГДФ-маннозо-дегидратазной активностью включают GMD свиньи (Broschat et al., Eur. J. Biochem., 153(2): 397-401 (1985)), GMD Caenorhabditis elegans и GMD Drosophila melanogaster (смотри, например, Rhomberg et al., FEBS J., 273: 2244-56 (2006)), Arabidopsis thaliana (смотри, например, Nakayama et al., Glycobiology, 13: 673-80 (2003)) и E.coli (Somoza et al., Structure, 8: 123-35 (2000)).
ГДФ-кетодезоксиманнозо-эпимераза/ГДФ-кетодезоксигалактозо-редуктаза (ГДФ-L-фукозосинтаза, EC 1.1.1.271) является бифункциональным ферментом, который был идентифицирован как у эукариот, так и у прокариот. ГДФ-кетодезоксиманнозо-эпимераза/ГДФ-кетодезоксигалактозо-редуктаза человека называется белком FX (также известным как hFX или GER). Нуклеотидная последовательность, кодирующая hFX, показана в SEQ ID NO: 3. Белок hFX имеет аминокислотную последовательность, показанную в SEQ ID NO: 4.
Переносчик ГДФ-фукозы был идентифицирован в нескольких видах. Установлено, что переносчик ГДФ-фукозы человека (hGFTr) имеет отношение к врожденным нарушениям гликозилирования-II (CDG-II) (Lubke et al., Nat. Genet. 28: 73-6 (2001)). Также известное как недостаток адгезии лейкоцитов II (LAD II), нарушение, как обнаружено, происходит в результате патологического отклонения в фукозилировании лигандов селектинов (Roos and Law, Blood Cells Mol. Dis. 27: 1000-4 (2001)). Нуклеотидная последовательность, кодирующая hGFTr, показана в SEQ ID NO: 5, а аминокислотная последовательность hGFTr показана в SEQ ID NO: 6. Были идентифицированы гомологичные гены с активностью переносчика ГДФ-фукозы в других видах, таких как Drosophila melanogaster (Ishikawa et al., Proc. Natl. Acad. Sci. U.S.A. 102: 18532-7 (2005)), печени крыс (Puglielli and Hirschberg, J. Biol. Chem. 274: 35596-60 (1999)), и предполагаемый гомолог СНО (Chen et al., Glycobiology, 15: 259-69 (2005)).
Был идентифицирован ряд фукозилтрансфераз (смотри Breton et al., Glycobiol. 8: 87-94 (1997); Becker, Lowe, Glycobiol. 13: 41R-53R (2003); Ma et al., Glycobiol. 16: 158R-184R (2006)), например, α1,2-фукозилтрасфераза (ЕС 2.4.1.69, кодируемая FUT1 и FUT2), α1,3-фукозилтрасфераза (гликопротеин-3-α-L-фукозилтрасфераза, ЕС 2.4.1.214, кодируемая FUT3-FUT7 и FUT9), α1,4-фукозилтрасфераза (ЕС 2.4.1.65, кодируемая FUT3) и α1,6-фукозилтрасфераза (гликопротеин-6-α-L-фукозилтрасфераза, ЕС 2.4.1.68, кодируемая FUT8). Как правило, α1,2-фукозилтрасфераза переносит фукозу на концевой остаток галактозы в N-гликане через образование α1,2-связи. Как правило, α1,3-фукозилтрасфераза и α1,4-фукозилтрасфераза переносят фукозу на остаток GlcNAc на восстанавливающем конце N-гликана.
Как правило, α1,6-фукозилтрасферазы переносят фукозу через образование α1,6-связи на остаток GlcNAc на восстанавливающем конце N-гликанов (связанный с аспарагином GlcNAc). Как правило, для α1,6-фукозилтрасферазы требуется концевой остаток GlcNAc на восстанавливающем конце по крайней мере одной ветви триманнозного остова для того, чтобы она могла добавить фукозу к GlcNAc на восстанавливающем конце. Однако была идентифицирована α1,6-фукозилтрасфераза, для которой требуется концевой галактозидный остаток на невосстанавливающем конце для того, чтобы она могла добавить фукозу к GlcNAc на восстанавливающем конце (Wilson et al., Biochim. Biophys. Res. Comm. 72: 909-916 (1976)) и Lin et al. (Glycobiol. 4: 895-901 (1994)). Было установлено, что в клетках яичника китайского хомячка с недостатком GlcNAc-трансферазы I α1,6-фукозилтрасфераза будет фукозилировать N-гликаны Man4GlcNAc2 и Man5GlcNAc2. Аналогичным образом, α1,3-фукозилтрасфераза переносит фукозу на остаток GlcNAc на восстанавливающем конце N-гликанов, но через образование α1,3-связи, обычно со специфичностью в отношении N-гликанов с одним незамещенным невосстанавливающим концевым остатком GlcNAc. N-гликановые продукты этого фермента присутствуют в растениях, у насекомых и некоторых других беспозвоночных (например, Schistosoma, Haemonchus, Lymnaea). Однако в патенте США № 7094530 описывается 1,3-фукозилтрасфераза, выделенная из линии моноцитарных клеток ТНР-1.
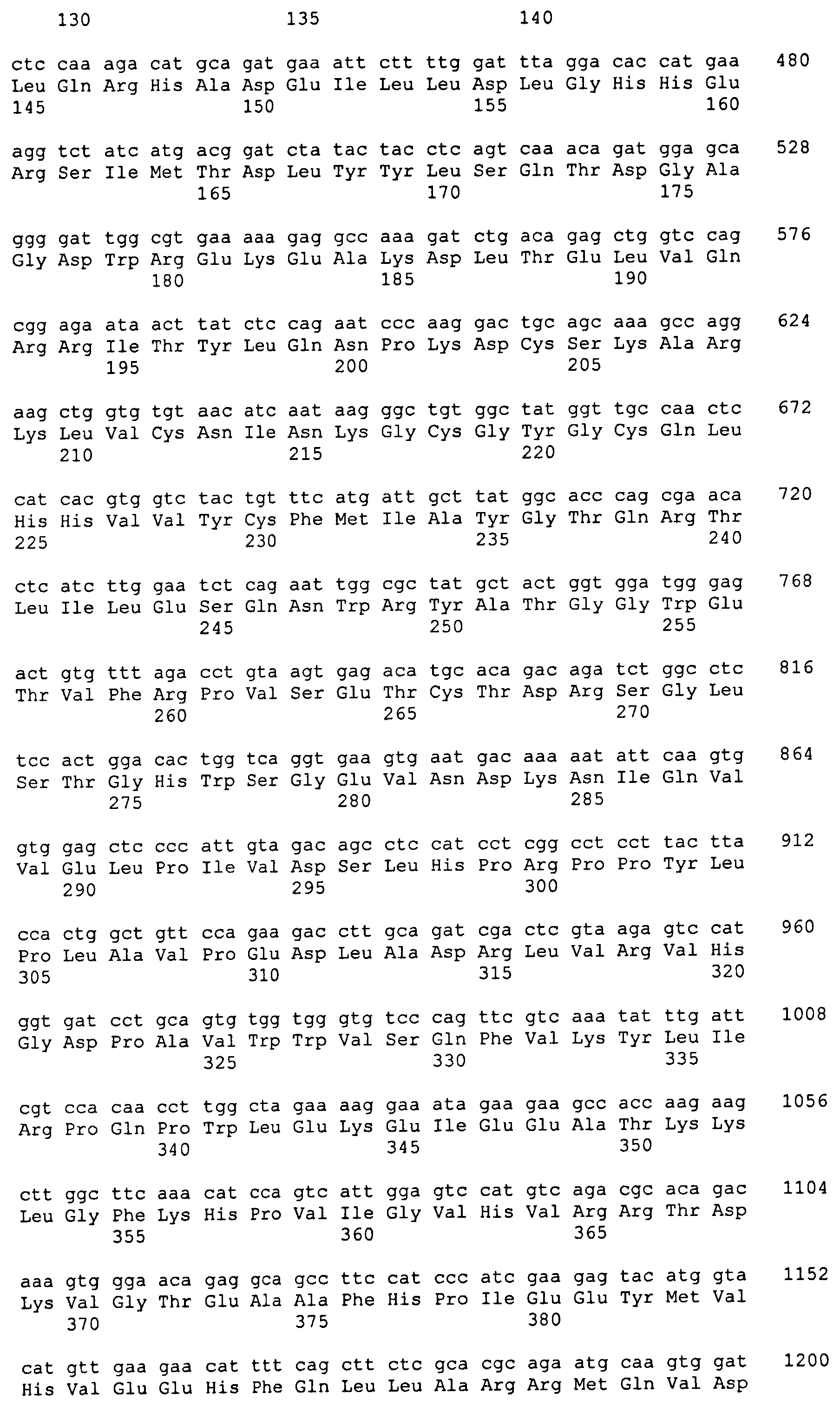
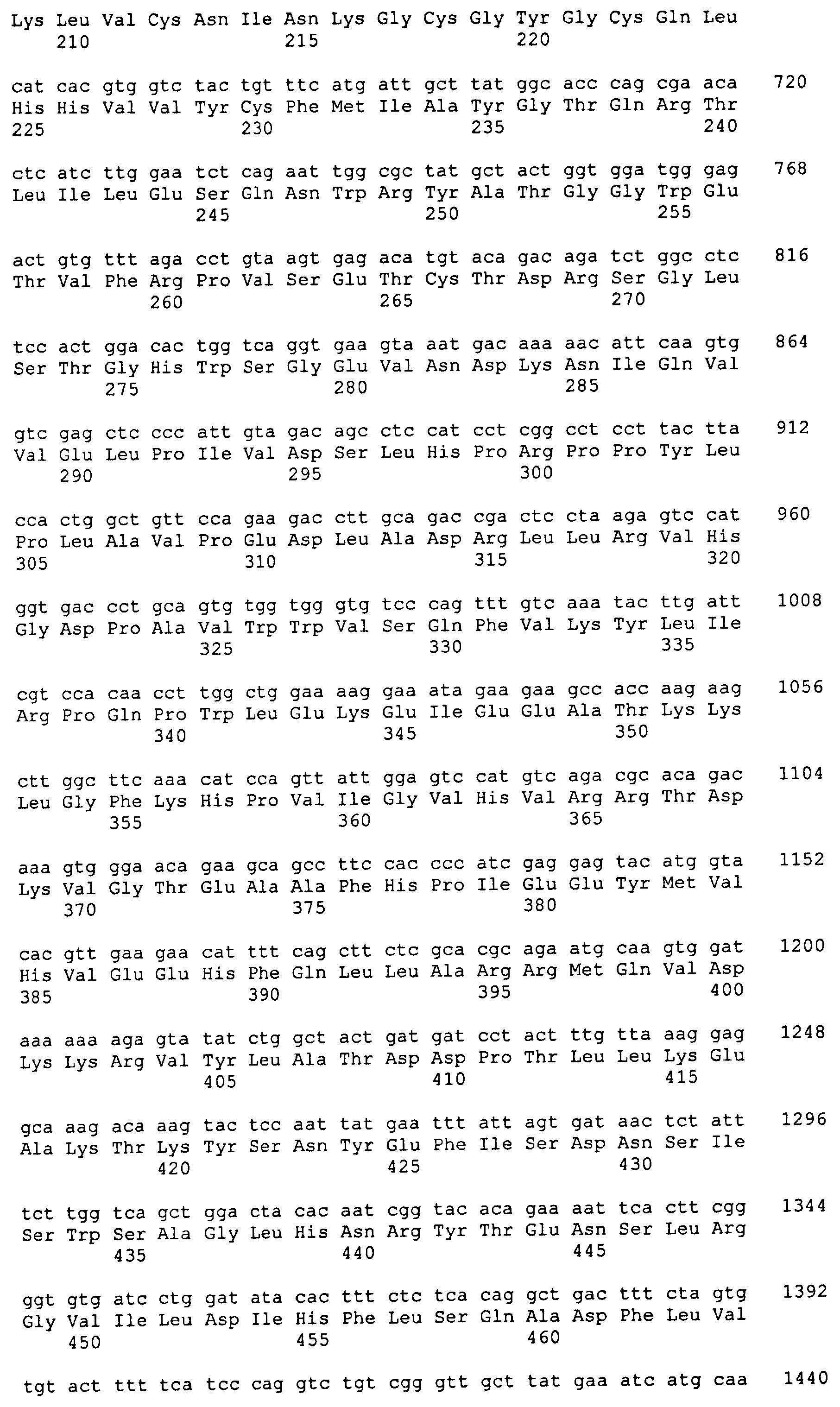
1,6-Фукозилтрасфераза человека (hFUT8) была идентифицирована Yamaguchi et al. (Cytogenet. Cell. Genet. 84: 58-6 (1999)). Нуклеотидная последовательность, кодирующая FUT8 человека, показана в SEQ ID NO: 7. Аминокислотная последовательность hFUT8 показана в SEQ ID NO: 8. Были идентифицированы гомологичные гены с активностью FUT8 в других видах, такие как FUT8 крысы (rFUT8), имеющая аминокислотную последовательность, показанную в SEQ ID NO: 10, и кодируемая нуклеотидной последовательностью, показанной в SEQ ID NO: 9, FUT8 мыши (mFUT8), имеющая аминокислотную последовательность, показанную в SEQ ID NO: 12, и кодируемая нуклеотидной последовательностью, показанной в SEQ ID NO: 11, и FUT8 свиньи (pFUT8), имеющая аминокислотную последовательность, показанную в SEQ ID NO: 14, и кодируемая нуклеотидной последовательностью, показанной в SEQ ID NO: 13. FUT8 была также идентифицирована в клетках СНО (Yamane-Ohnuki et al., Biotechnol. Bioeng. 87: 614-622 (2004)), клетках COS почки обезьяны (Clarke and Watkins, Glycobiol. 9: 191-202 (1999)) и клетках цыпленка (Coullin et al., Cytogenet. Genome Res. 7: 234-238 (2002)). Клонирование и характеристика фукозилтрансфераз из C. elegans и D. melanogaster описываются Paschinger и др. (Glycobiol. 15: 463-474 (2005)). Были идентифицированы предполагаемые 1,6-фукозилтрасферазы Ciona intestinalis, Drosophila pseudoobscura, Xenopus laevis и Danio rerio (входящие номера в GenBank AJ515151, AJ830720, AJ514872 и AJ781407, соответственно).
Вышеупомянутые ферменты или активности каскада реакций фукозилирования кодируются нуклеиновыми кислотами. Нуклеиновые кислоты могут быть ДНК или РНК, но обычно нуклеиновые кислоты являются ДНК, поскольку предпочтительно, чтобы нуклеиновые кислоты, кодирующие ферменты или активности каскада реакций фукозилирования, были стабильно интегрированы в геном клеток-хозяев. Каждая из нуклеиновых кислот, кодирующих ферменты или активности каскада реакций фукозилирования, функционально связана с регуляторными последовательностями, которые делают возможной экспрессию ферментов или активностей каскада реакций фукозилирования. Такие регуляторные последовательности включают промотор и необязательно энхансер, находящийся 5' от нуклеиновой кислоты, кодирующей фермент или активность каскада реакций фукозилирования, и сайт терминации транскрипции, находящийся 3' от нуклеиновой кислоты, кодирующей фермент или активность каскада реакций фукозилирования. Нуклеиновая кислота также, как правило, дополнительно включает 5' нетранслируемый район, имеющий сайт связывания рибосом, и 3' нетранслируемый район, имеющий сайт полиаденилирования. Нуклеиновая кислота часто является компонентом вектора, такого как плазмида, который способен реплицироваться в клетках, в которых экспрессируется фермент или активность каскада реакций фукозилирования. Вектор может также содержать маркер, делающий возможным отбор клеток, трансформированных вектором. Однако некоторые типы клеток, в частности дрожжи, можно успешно трансформировать нуклеиновой кислотой, в которой отсутствуют последовательности векторов.
Как правило, клетки-хозяева, трансформированные нуклеиновыми кислотами, кодирующими один или несколько ферментов или активностей каскада реакций фукозилирования, дополнительно включают одну или несколько нуклеиновых кислот, кодирующих желаемые гликопротеины. Как и для ферментов каскада реакций фукозилирования, нуклеиновые кислоты, кодирующие гликопротеины, функционально связаны с регуляторными последовательностями, которые делают возможной экспрессию гликопротеинов. Нуклеиновые кислоты, кодирующие гликопротеины, можно амплифицировать из линий клеток, которые, как известно, экспрессируют гликопротеин, используя праймеры к консервативным районам гликопротеина (смотри, например, Marks et al., J. Mol. Biol.: 581-596 (1991)). Нуклеиновые кислоты можно также синтезировать de novo на основе последовательностей в научной литературе. Нуклеиновые кислоты можно также синтезировать путем удлинения перекрывающихся олигонуклеотидов, охватывающих желаемую последовательность (смотри, например, Caldas et al., Protein Engineering, 13: 353-360 (2000)).
Тип структуры фукозилированного N-гликана, продуцируемого клеткой-хозяином, будет зависеть от каскада реакций фукозилирования в клетке-хозяине и конкретной фукозилтрансферазы. Например, α1,2-фукозилтрансфераза, как правило, добавляет фукозу к концевой галактозе в N-гликане. Как таковой, каскад реакций, в котором используется α1,2-фукозилтрансфераза, предпочтительно было бы внедрять в клетку-хозяина, которая способна продуцировать N-гликаны, имеющие гликоформу Gal2GlcNAc2Man3GlcNAc2. Продуцируемые N-гликаны будут иметь фукозу в α1,2-связи с концевыми остатками галактоз. Как α1,3-фукозилтрансфераза, так и α1,4-фукозилтрансфераза добавляют фукозу к одному или нескольким остаткам GlcNAc на невосстанавливающем конце или вблизи него через образование α1,3- или α1,4-связи соответственно и для некоторых α1,3-фукозилтрансфраз через образование α1,3-связи с остовым GlcNAc, связанным с остатком аспарагина гликопротеина. Как таковой, каскад реакций, в котором используется α1,3/4-фукозилтрансфераза, предпочтительно было бы внедрять в клетку-хозяина, которая способна продуцировать N-гликаны, имеющие по крайней мере гликоформу GlcNAcMan5GlcNAc2. Наконец, α1,6-фукозилтрансфераза, как правило, переносит фукозу через образование α1,6-связи на остовый остаток GlcNAc, связанный с остатком аспарагина гликопротеина. Как правило, каскад реакций, в котором используется α1,6-фукозилтрансфераза, предпочтительно было бы внедрять в клетку-хозяина, которая способна продуцировать N-гликаны, имеющие по крайней мере гликоформу GlcNAcMan5GlcNAc2, Man5GlcNAc2или Man4GlcNAc2.
Гликопротеины, которые можно продуцировать в соответствии с раскрытыми здесь способами, включают любой желаемый белок для терапевтических или диагностических целей, независимо от происхождения последовательности нуклеиновой кислоты для продуцирования гликопротеина. Например, моноклональные антитела, в которых N-гликан не является фукозилированным, обладают увеличенной активностью ADCC, однако увеличенная активность ADCC нежелательна для моноклональных антител, предназначенных для связывания лигандов для рецептора в качестве лечения нарушения, а не индицирования активности ADCC. Моноклональные антитела, продуцируемые в раскрытых здесь клетках-хозяевах, включающих каскад реакций фукозилирования, будут иметь фукозилированные N-гликаны и, как предполагается, уменьшенную активность ADCC. В качестве другого примера, агенты для иммунной адгезии (смотри, патенты США № 5428130, 5116964, 5514582 и 5455165, Capon et al. Nature 337: 525 (1989); Chamow and Ashkenazi, Trends Biotechnol. 14: 52-60 (1996); Ashkenazi and Chamow, Curr. Opin. Immunol. 9: 195-200 (1997)), которые включают экстраклеточную часть связанного с мембраной рецептора, слитую с частью Fc антитела, продуцируемую в раскрытых здесь клетках-хозяевах, включающих каскад реакций фукозилирования, будут иметь фукозилированные N-гликаны и, как предполагается, уменьшенную активность ADCC. Примеры гликопротеинов, которые можно продуцировать в соответствии со способами настоящего изобретения с наличием у них фукозилированных N-гликанов, включают, но без ограничения, эритропоэтин (ЕРО), цитокины, такие как интерферон-α, интерферон-β, интерферон-γ, интерферон-ω и гранулоцитарный CSF, факторы коагуляции, такие как фактор VIII, фактор IX и белок С человека, моноклональные антитела, α-цепь растворимого рецептора IgE, IgG, IgM, IgG, урокиназу, химазу и ингибитор трипсина, IGF-связывающий белок, фактор роста эпидермиса, фактор выброса гормона роста, слитый с аннексином V белок, ангиостатин, фактор-2 роста эндотелия сосудов, ингибиторный в отношении клеток-предшественников миелоидного ряда фактор-1, остеопротегерин, тканевой активатор плазминогена, G-CSF, GM-CSF, и TNF-рецептор.
В конкретных вариантах осуществления одна или несколько нуклеиновых кислот кодируют слитые белки, включающие каталитический домен белка каскада реакций фукозилирования, слитый с пептидом для направленной доставки, который направляет слитый белок в конкретную область внутри клетки. Как правило, пептид для направленной доставки будет направлять слитый белок в положение внутри секреторного пути. Термин «секреторный путь», таким образом, относится к органеллам и компонентам внутри клетки, в которых гликопротеины модифицируются при подготовке к секреции. Секреторный путь включает эндоплазматический ретикулум, аппарат Гольджи, транс-Гольджи-сеть и секреторные везикулы. Например, подходящие пептиды для клеточной доставки могут направлять каталитический домен в эндоплазматический ретикулум, аппарат Гольджи, транс-Голиджи-сеть или секреторные везикулы. Пептиды для направленной доставки, которые можно использовать в настоящем изобретении, включают пептиды, описанные в патенте США № 7029872. В одном варианте осуществления каталитический домен фукозилтрансферазы слит с пептидом для направленной доставки, который направляет слитый белок в аппарат Гольджи. Конкретный пептид для направленной доставки, слитый с каталитическим доменом фукозилтрансферазы, будет зависеть от клетки-хозяина, конкретной фукозилтрансферазы и продуцируемого гликопротеина. Примеры пептидов для направленной доставки, которые можно использовать для направленной доставки фукозилтрансферазы, описаны, например, в патенте США № 7029872 и опубликованных заявках на патенты США № 2004/0018590, 2004/0230042, 2005/0208617, 2004/0171826, 2006/0286637 и 2007/0037248.
Нуклеиновые кислоты, кодирующие ферменты или активности, вовлеченные в каскад реакций фукозилирования, лигируют в векторы, которые можно использовать для трансфекции клеток-хозяев. Как правило, векторы будут включать регуляторные элементы, которые выделены из того же вида клетки, что намеченная клетка-хозяин, или которые выделены из других видов, но о которых известно, что они будут функциональными при встраивании в намеченную клетку-хозяина. Как правило, эти регуляторные элементы включают 5' регуляторные последовательности, такие как промоторы, а также 3' регуляторные последовательности, такие как последовательности - терминаторы транскрипции. Векторы будут, как правило, включать по крайней мере один элемент - селектируемый маркер, который делает возможным отбор клеток-хозяев, которые были успешно трансформированы вектором. Векторы трансфецируют в намеченные клетки-хозяева и результирующие клетки скринируют на наличие селектируемого маркера, чтобы идентифицировать такие клетки-хозяева, которые были успешно трансфецированы вектором и которые будут, следовательно, нести вектор, кодирующий слитый белок.
Для экспрессии гликопротеинов предпочтительными часто являются низшие эукариоты, такие как дрожжи, поскольку их можно экономно культивировать, они дают высокие выходы белка и при соответствующей модификации способны продуцировать гликопротеины с конкретными преобладающими структурами N-гликанов. Дрожжи, в частности, характеризуются установленной генетикой, делающей возможными быстрые трансформации, методики локализации тестируемого белка и легкие методы выключения генов. Различные дрожжи, такие как K. lactis, Pichia pastoris, Pichia methanolica и Hansenula polymorpha, обычно используются для культур клеток и продукции белков, поскольку они способны расти до высоких плотностей клеток и секретировать большие количества рекомбинантного белка в промышленной масштабе. Подобным образом, нитчатые грибы, такие как Aspergillus niger, Fusarium sp., Neurospora crassa и другие, можно использовать для продукции гликопротеинов в промышленном масштабе.
Низшие эукариоты, в частности дрожжи, можно генетически модифицировать, так чтобы они экспрессировали гликопротеины, в которых характер гликозилирования является сложным или напоминает характер гликозилирования у людей или является гуманизированным. Такие генетически модифицированные низшие эукариоты можно получить устранением выбранных эндогенных ферментов гликозилирования, которые вовлечены в продукцию N-гликанов с высоким содержанием маннозы, и введением различных комбинаций экзогенных ферментов, вовлеченных в создание сложных N-гликанов. Способы создания методами генетической инженерии дрожжей, которые продуцируют сложные N-гликаны, описаны в патенте США № 7029872 и опубликованных заявках на патенты США № 2004/0018590, 2005/0170452, 2006/0286637, 2004/0230042, 2005/0208617, 2004/0171826, 2005/0208617 и 2006/0160179. Например, можно выбрать или создать клетку-хозяина, истощенную по 1,6-маннозилтрансферазным активностям, которые в противном случае могли бы добавлять остатки маннозы на N-гликан на гликопротеине. Например, в дрожжах ген ОСН1 кодирует 1,6-маннозилтрансферазную активность. Затем создают дополнительными методами клетку-хозяина, включающую один или несколько ферментов, вовлеченных в продукцию сложных N-гликанов, напоминающих N-гликаны человека.
В одном варианте осуществления клетка-хозяин, кроме того, включает каталитический домен α1,2-маннозидазы, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности α1,2-маннозидазы в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина. При прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу Man5GlcNAc2, например гликоформу Man5GlcNAc2(Fuc). В патенте США № 7029872 и опубликованных заявках на патенты США № 2004/0018590 и 2005/0170452 описываются клетки-хозяева, являющиеся низшими эукариотами, способные продуцировать гликопротеин, включающий гликоформу Man5GlcNAc2.
В дальнейшем варианте осуществления непосредственно предшествующая клетка-хозяин, кроме того, включает каталитический домен GlcNAc-трансферазы I (GnTI), слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности GlcNAc-трансферазы I в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина. При прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу GlcNAcMan5GlcNAc2, например гликоформу GlcNAcMan5GlcNAc2(Fuc). В патенте США № 7029872 и опубликованных заявках на патенты США № 2004/0018590 и 2005/0170452 описываются клетки-хозяева, являющиеся низшими эукариотами, способные продуцировать гликопротеин, включающий гликоформу GlcNAcMan5GlcNAc2. Продуцируемый в вышеуказанных клетках гликопротеин можно обработать in vitro гексаминидазой с получением рекомбинантного гликопротеина, включающего фукозилированную гликоформу Man5GlcNAc2(Fuc).
Во все еще дальнейшем варианте осуществления непосредственно предшествующая клетка-хозяин, кроме того, включает каталитический домен маннозидазы II, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности маннозидазы II в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина. При прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу GlcNAcMan3GlcNAc2, например гликоформу GlcNAcMan3GlcNAc2(Fuc). В опубликованной заявке на патент США № 2004/0230042 описываются клетки-хозяева, являющиеся низшими эукариотами, которые экспрессируют ферменты маннозидазы II и способны продуцировать гликопротеины, имеющие в преобладающей степени гликоформу GlcNAcMan3GlcNAc2. Продуцируемый в вышеуказанных клетках гликопротеин можно обработать in vitro гексаминидазой с получением рекомбинантного гликопротеина, включающего гликоформу Man3GlcNAc2(Fuc).
Во все еще дальнейшем варианте осуществления непосредственно предшествующая клетка-хозяин, кроме того, включает каталитический домен GlcNAc-трансферазы II (GnTII), слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности GlcNAc-трансферазы II в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина. При прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу GlcNAc2Man3GlcNAc2, например гликоформу GlcNAc2Man3GlcNAc2(Fuc). В патенте США № 7029872 и опубликованных заявках на патенты США № 2004/0018590 и 2005/0170452 описываются клетки-хозяева, являющиеся низшими эукариотами, способные продуцировать гликопротеин, включающий гликоформу GlcNAc2Man3GlcNAc2. Продуцируемый в вышеуказанных клетках гликопротеин можно обработать in vitro гексаминидазой с получением рекомбинантного гликопротеина, включающего гликоформу Man3GlcNAc2(Fuc).
Во все еще дальнейшем варианте осуществления непосредственно предшествующая клетка-хозяин, кроме того, включает каталитический домен галактозотрансферазы II, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности галактозотрансферазы II в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина. При прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу Gal2GlcNAc2Man3GlcNAc2, например Gal2GlcNAc2Man3GlcNAc2(Fuc). В опубликованной заявке на патент США № 2006/0040353 описываются клетки-хозяева, являющиеся низшими эукариотами, способные продуцировать гликопротеин, включающий гликоформу Gal2GlcNAc2Man3GlcNAc2. Продуцируемый в вышеуказанных клетках гликопротеин можно обработать in vitro галактозидазой с получением рекомбинантного гликопротеина, включающего фукозилированную гликоформу GlcNAc2Man3GlcNAc2(Fuc), например гликоформу GlcNAc2Man3GlcNAc2(Fuc).
Во все еще дальнейшем варианте осуществления непосредственно предшествующая клетка-хозяин, кроме того, включает каталитический домен сиалилтрансферазы, слитый с осуществляющим направленную клеточную доставку сигнальным пептидом, который обычно не связан с каталитическим доменом и выбирается для направленной доставки активности сиалилтрансферазы в эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина. При прохождении рекомбинантного гликопротеина через эндоплазматический ретикулум или аппарат Гольджи клетки-хозяина продуцируется рекомбинантный гликопротеин, включающий фукозилированную гликоформу NANA2Gal2GlcNAc2Man3GlcNAc2, например гликоформу NANA2Gal2GlcNAc2Man3GlcNAc2(Fuc). Для клеток-хозяев, являющихся низшими эукариотами, таких как дрожжи и нитчатые грибы, предпочтительно, чтобы клетка-хозяин, кроме того, включала средство для обеспечения ЦМФ-сиаловой кислоты для переноса на N-гликан. В опубликованной заявке на патент США № 2005/0260729 описывается способ создания методами генетической инженерии низших эукариот, имеющих каскад реакций для синтеза ЦМФ-сиаловой кислоты, а в опубликованной заявке на патент США № 2005/0286637 описывается способ создания методами генетической инженерии низших эукариот, продуцирующих сиалированные гликопротеины. Продуцируемый в вышеуказанных клетках гликопротеин можно обработать in vitro нейраминидазой с получением рекомбинантного гликопротеина, включающего фукозилированную гликоформу Gal2GlcNAc2Man3GlcNAc2, например гликоформу Gal2GlcNAc2Man3GlcNAc2(Fuc).
Любая из предшествующих клеток-хозяев может, кроме того, включать одну или несколько GlcNAc-трансфераз, выбираемых из группы, состоящей из GnTIII, GnTIV, GnTV, GnTVI и GnTIX, для продукции гликопротеинов, имеющих разветвленные и/или с множеством антенн структуры N-гликанов, таких как те, которые описаны в опубликованных заявках на патенты 2004/074458 и 2007/0037248. Различные предшествующие клетки-хозяева, кроме того, включают один или несколько переносчиков сахаров, таких как переносчики УДФ-GlcNAc (например, переносчики УДФ-GlcNAc Kluyveromyces lactis и Mus musculus), переносчики УДФ-галактозы (например, переносчик УДФ-галактозы Drosophila melanogaster), переносчик ЦМФ-сиаловой кислоты (например, переносчик сиаловой кислоты человека). Поскольку в клетках-хозяевах, являющихся низшими эукариотами, таких как дрожжи и нитчатые грибы, отсутствуют вышеуказанные переносчики, предпочтительно, чтобы были созданы методами генной инженерии клетки-хозяева, являющиеся низшими эукариотами, такие как дрожжи и нитчатые грибы, которые включают вышеуказанные переносчики.
В дальнейших вариантах осуществления вышеуказанных клеток-хозяев методами генетической инженерии создают клетки-хозяева, в которых устранены гликопротеины, имеющие устойчивые к α-маннозидазе N-гликаны, путем делеции или разрушения гена β-маннозилтрансферазы (ВМТ2) (смотри опубликованную заявку на патент США № 2006/0211085), и гликопротеины, имеющие остатки фосфоманноз, путем делеции или разрушения одного или обоих генов фосфоманнозилтрансфераз PNO1 и MNN4B (смотри, например, опубликованные заявки на патенты США № 2006/0160179 и 2004/0014170). Во все еще дальнейших вариантах осуществления вышеуказанных клеток-хозяев клетки-хозяева, кроме того, генетически модифицируют для исключения О-гликозилирования гликопротеина путем делеции или разрушения одного или нескольких генов Dol-P-белок(Ser/Thr)-маннозилтрансфераз (РМТ) (смотри патент США № 5714377).
Было установлено, что оптимизация кодонов генов или транскрипционных единиц, кодирующих конкретные полипептиды, приводит к увеличенной экспрессии кодируемых полипептидов, т.е. увеличенной трансляции мРНК, кодирующей полипептид. Следовательно, в случае раскрытых здесь клеток-хозяев увеличенная экспрессия кодируемых ферментов будет порождать больше кодируемых ферментов, что может приводить к увеличенной продукции N-гликанов, которые являются фукозилированными. В контексте оптимизации кодонов термин «экспрессия» и его варианты относятся к трансляции мРНК, кодирующей полипептид, а не к транскрипции полинуклеотида, кодирующего полипептид. Используемый здесь термин «ген» относится и к геномной ДНК или РНК, кодирующей полипептид, и к кДНК, кодирующей полипептид.
Оптимизация кодонов является способом, с помощью которого пытаются улучшить гетерологичную экспрессию гена, когда ген перемещают в чужеродную генетическую среду, которая проявляет использование состоящих из нуклеотидных звеньев кодонов, отличное от такого использования в природной генетической среде гена, или улучшить эктопическую экспрессию гена в его природной генетической среде, когда ген природно включает один или несколько состоящих из нуклеотидных звеньев кодонов, которые обычно не используются в природных относительно генетической среды генах, которые кодируют в высокой степени экспрессируемые гены. Другими словами оптимизация кодонов включает замену тех состоящих из нуклеотидных звеньев кодонов гена, которые используются с относительно низкой частотой в конкретной генетической среде или организме, состоящими из нуклеотидных звеньев кодонами, которые используются в генах, которые экспрессируются с большей частотой в генетической среде или организме. Таким образом, экспрессия (трансляция) продукта гена (полипептида) увеличивается. Допущением является то, что состоящие из нуклеотидных звеньев кодоны, которые обнаруживаются с большей частотой в высокой степени экспрессируемых генах, более эффективно транслируются, чем состоящие из нуклеотидных звеньев кодоны, которые обнаруживаются с низкой частотой.
Как правило, способы оптимизации состоящих из нуклеотидных звеньев кодонов для конкретного гена зависят от установления частоты использования в генах, являющихся в высокой степени экспрессируемыми в организме, состоящих из нуклеотидных звеньев кодонов для каждой из аминокислот и затем замены тех состоящих из нуклеотидных звеньев кодонов в представляющем интерес гене, которые используются с низкой частотой в высокой степени экспрессируемых генах, состоящими из нуклеотидных звеньев кодонами, которые, как установлено, используются в высокой степени экспрессируемых генах (смотри, например, Lathe, Synthetic Oligonucleotide Probes Deduced from Amino Acid Sequence Data: Theoretical and Practical Considerations, J. Molec. Biol.: 183: 1-12 (1985); Nakamura et al., Nuc. Acid Res. 28: 292 (2000); Fuglsang, Protein Expression & Purification 31: 247-249 (2003)). Существуют многочисленные компьютерные программы, которые будут в автоматическом режиме анализировать состоящие из нуклеотидных звеньев кодоны нуклеиновой кислоты организма, кодирующей ген, и предлагать состоящие из нуклеотидных звеньев кодоны для замены состоящих из нуклеотидных звеньев кодонов, которые встречаются с низкой частотой в организме, состоящими из нуклеотидных звеньев кодонами, которые обнаруживаются в генах, являющихся в высокой степени экспрессируемыми в организме.
Подразумевается, что следующие примеры содействуют дальнейшему пониманию настоящего изобретения.
ПРИМЕР 1
В этом примере демонстрируется конструирование штамма Pichia pastoris, способного продуцировать гликопротеины, которые включают фукозу в структуре N-гликана гликопротеина.
Для работы с рекомбинантными ДНК используют штаммы TOP10 или XL10-Gold Escherichia coli. N-гликозидазу F, ферменты для рестрикции и модификации получают от New England BioLabs (Beverly, MA) и используют согласно инструкциям производители. α1,6-Фукозидазу получают от Sigma-Aldrich (St. Louis, MO) и используют, как рекомендовано производителем. Олигонуклеотиды получают от Integrated DNA Technologies (Coralville, IA). Металлохелатирующую смолу «HisBind» получают от Novagen (Madison, WI). 96-луночные планшеты для выделения плазмидной ДНК из лизата - от Promega (Madison, WI). Связывающие белок 96-луночные планшеты - от Millipore (Bedford, MA). Соли и буферные агенты - от Sigma-Aldrich (St. Louis, MO).
Амплификация генов каскада реакций фукозилирования
На фиг. 1 продемонстрирован общий вид каскада реакций фукозилирования. Открытую рамку считывания (ORF) hGMD амплифицируют с кДНК печени человека (BD Biosciences, Palo Alto, CA) с использованием полимеразы Advantage 2, следуя процедуре, рекомендованной производителем. Вкратце, праймеры SH415 и SH413 (5'-GGCGG CCGCC ACCAT GGCAC ACGCA CCGGC ACGCT GC-3' (SEQ ID NO: 15) и 5'-TTAAT TAATC AGGCA TTGGG GTTTG TCCTC ATG-3' (SEQ ID NO: 16), соответственно) используют для амплификации продукта размером 1139 п.о. с кДНК печени человека, используя следующие условия: 97°С в течение 3 минут, 35 циклов, каждый из которых представляет 97°С в течение 30 секунд, 50°С в течение 30 секунд и 72°С в течение 2 минут, и 72°С в течение 10 минут. Впоследствии продукт клонируют в pCR2.1 (Invitrogen, Carlsbad, CA), секвенируют, а результирующей конструкции присваивают обозначение pSH985.
Используя очерченные выше условия, праймеры SH414 и SH411 (5'-GGCGG CCGCC ACCAT GGGTG AACCC CAGGG ATCCA TG-3' (SEQ ID NO: 17) и 5'-TTAAT TAATC ACTTC CGGGC CTGCT CGTAG TTG-3' (SEQ ID NO: 18), соответственно) используют для амплификации фрагмента размером 986 п.о. с кДНК почки человека (BD Biosciences, Palo Alto, CA), который соответствует ORF гена FX человека. Впоследствии этот фрагмент клонируют в pCR2.1, секвенируют и обозначают pSH988.
ORF GFTr человека амплифицируют с кДНК селезенки человека (BD Biosciences, Palo Alto, CA), используя очерченные выше условия и праймеры RCD679 и RCD680 (5'-GCGGC CGCCA CCATG AATAG GGCCC CTCTG AAGCG G-3' (SEQ ID NO: 19) и 5'-TTAAT TAATC ACACC CCCAT GGCGC TCTTC TC-3' (SEQ ID NO: 20), соответственно). Результирующий фрагмент размером 1113 п.о. клонируют в pCR2.1, секвенируют и обозначают pGLY2133.
Усеченную форму ORF FUT8 мыши, которая кодирует аминокислоты 32-575 и в которой отсутствуют нуклеотиды, кодирующие эндогенный трансмембранный домен, амплифицируют с кДНК головного мозга мыши (BD Biosciences, Palo Alto, CA), используя очерченные выше условия и праймеры SH420 и SH421 (5'-GCGGC GCGCC GATAA TGACC ACCCT GATCA CTCCA G-3' (SEQ ID NO: 21) и 5'-CCTTA ATTAA CTATT TTTCA GCTTC AGGAT ATGTG GG-3' (SEQ ID NO: 22), соответственно). Результирующий фрагмент размером 1654 п.о. клонируют в pCR2.1, секвенируют и обозначают pSH987.
Создание генов фукозилирования в кассетах для экспрессии в дрожжах
Открытые рамки считывания для GMD, FX и GFTr получают расщеплением вышеуказанных векторов ферментами рестрикции NotI и PacI с получением ДНК-фрагментов с NotI-совместимым 5'-концом и PacI-совместимым 3'-концом. Фрагмент FUT8 получают расщеплением ферментами рестрикции AscI и PacI с получением ДНК с AscI-совместимым 5'-концом и PacI-совместимым 3'-концом.
Для создания экспрессирующей GMD кассеты GMD клонируют в вектор для экспрессии в дрожжах pSH995, который содержит промотор GAPDH P. pastoris и последовательность-терминатор транскрипции CYC S. cerevisiae и предназначен для интеграции в геном Pichia 3' от ORF Trp2, используя маркер устойчивости к нур-сеотрицину. Этот вектор проиллюстрирован на фиг. 3А. Вектор pSH995 расщепляют NotI и PacI для выделения фрагмента размером 1,1 т.п.о., содержащего ORF GMD, который затем субклонируют в pSH995, предварительно расщепленный теми же ферментами. Результирующий вектор, содержащий GMD под контролем промотора GAPDH, обозначают pSH997.A.
Для создания экспрессирующей FX кассеты вектор pSH988 расщепляют NotI и PacI для выделения фрагмента размером 1 т.п.о., содержащего ORF FX, который обрабатывают Т4 ДНК-полимеразой для удаления одноцепочечных концевых избыточностей (J. Sambrook, D. W. Russell, Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, ed. 3rd, 2001)). Впоследствии этот фрагмент субклонируют в вектор pGLY359 (Hamilton et al., Science 313, 1441 (2006)), предварительно расщепленный NotI и AscI и обработанный Т4 ДНК-полимеразой. Результирующий вектор pSH994 содержит кассету для экспрессии FX, состоящую из ORF FX, функционально связанной на 5'-конце с промотором PMA1 P. pastoris (PpPMA1prom), а на 3'-конце с последовательностью-терминатором транскрипции PMA P. pastoris (PpPMA1tt). Экспрессионная кассета фланкирована рестрикционными сайтами для SwaI.
Экспрессирующую GFTr кассету создают расщеплением pGLY2133 NotI и PacI для выделения фрагмента размером 1,1 т.п.о., содержащего ORF GFTr, который обрабатывают Т4 ДНК-полимеразой. Впоследствии этот фрагмент субклонируют в вектор pGLY363 (Hamilton, выше), предварительно расщепленный NotI и PacI и обработанный Т4 ДНК-полимеразой. Результирующий вектор pGLY2143 содержит кассету для экспрессии GFTr, состоящую из ORF GFTr, функционально связанной на 5'-конце с PpPMA1prom, а на 3'-конце с PpPMA1tt. Экспрессионная кассета фланкирована рестрикционными сайтами для RsrII.
Для создания каталитического домена FUT8, слитого с сигналом, определяющим местонахождение в дрожжах, первые 36 аминокислот района Mnn2 для направленной доставки в S. cerevisiae анализируют с помощью программного обеспечения GeneOptimizer и подвергают оптимизации кодонов для экспрессии в P. pastoris (GeneArt, Regensburg, Германия). Результирующую синтетическую ДНК для аминокислот 1-36 ScMnn2 создают с совместимыми с рестрикционными ферментами NotI и AscI 5'- и 3'-концами, соответственно, клонируют в «челночный» вектор с получением плазмидного вектора pSH831. Впоследствии вектор pSH987 расщепляют AscI и PacI для выделения фрагмента размером 1,6 т.п.о, кодирующего ORF каталитического домена FUT8, который затем субклонируют в рамке с ДНК, кодирующей пептид ScMnn2 для направленной доставки, в вектор pSH831, предварительно расщепленный теми же ферментами. Результирующий вектор обозначают pSH989. Для создания экспрессирующей FUT8-ScMnn2 кассеты pSH989 расщепляют NotI и PacI для высвобождения фрагмента размером 1,8 т.п.о., который субклонируют в вектор pGLY361 (Hamilton et al., Science 313, 1441 (2006)), расщепленный теми же ферментами. Результирующий вектор pSH991 содержит кассету для экспрессии слитого белка FUT8-Mnn2, состоящую из ORF слияния FUT8-Mnn2, функционально связанной на 5'-конце с промотором TEF P. pastoris (PpTEFprom), а на 3'-конце с последовательностью-терминатором транскрипции TEF P. pastoris (PpTEFtt). Экспрессионная кассета фланкирована рестрикционными сайтами для SgfI.
Получение вектора для создания фукозилирования
Вектор pSH984 расщепляют SwaI для высвобождения фрагмента размером 2,5 т.п.о., содержащего экспрессирующую FX кассету, который субклонируют в вектор pSH997 (который содержит экспрессирующую GMD кассету), расщепленный PmeI. Результирующий вектор, в котором экспрессирующие PMA-FX и GAPDH-GMD кассеты располагаются в одном и том же направлении, обозначают pSH1009. Фрагмент размером 2,7 т.п.о., содержащий экспрессирующую PMA-GFTr кассету, выделяют из pGLY2143, используя фермент рестрикции RsrlI, и субклонируют в pSH1009, расщепленный этим же ферментом. Результирующий вектор, в котором экспрессирующие PMA-GFTr и GAPDH кассеты располагаются в одном и том же направлении, обозначают pSH1019. Наконец, кассету для экспрессии TEF-FUT8 размером 1,8 т.п.о. выделяют из pSH991, используя SgfI, и субклонируют в pSH1019, расщепленный этим же ферментом. Результирующий вектор, в котором экспрессирующие TEF-FUT8 и GAPDH кассеты располагаются в одном и том же направлении, обозначают pSH1022. Этот вектор проиллюстрирован на Фиг. 2В.
Создание вектора для экспрессии ЕРО крысы
Усеченную форму гена эритропоэтина Rattus norvegicus (rEPO), кодирующую аминокислоты 27-192, амплифицируют с кДНК почки крысы (BD Biosciences, Palo Alto, CA), используя полимеразу Advantage 2, как рекомендовано производителем. Вкратце, праймеры rEPO-прямой и rEPO-обратный (5'-GGGAA TTCGC TCCCC CACGC CTCAT TTGCG AC-3' (SEQ ID NO: 23) и 5'-CCTCT AGATC ACCTG TCCCC TCTCC TGCAG GC-3' (SEQ ID NO: 24), соответственно) используют для амплификации продукта размером 516 п.о. с кДНК почки крысы, используя следующие циклические условия: 1 цикл при 94°С в течение 1 минуты; 5 циклов, каждый из которых представляет 94°С в течение 30 секунд, 72°С в течение 1 минуты; 5 циклов, каждый из которых представляет 94°С в течение 30 секунд, 70°С в течение 1 минуты; 25 циклов, каждый из которых представляет 94°С в течение 20 секунд, 68°С в течение 1 минуты. Впоследствии продукт клонируют в pCR2.1 (Invitrogen, Carlsbad, CA), секвенируют, а результирующей конструкции присваивают обозначение pSH603. Для создания вектора для экспрессии в дрожжах pSH603 расщепляют EcoRI и XbaI для высвобождения фрагмента размером 506 п.о., который субклонируют в pPICZαA (Invitrogen, Carlsbad, CA), который был предварительно расщеплен теми же ферментами. Результирующий экспрессионный вектор обозначают pSH692. rEPO в pSH692 находится под контролем метанол-индуцибельного промотора АОХ.
Создание штаммов дрожжей и продукция ЕРО крысы.
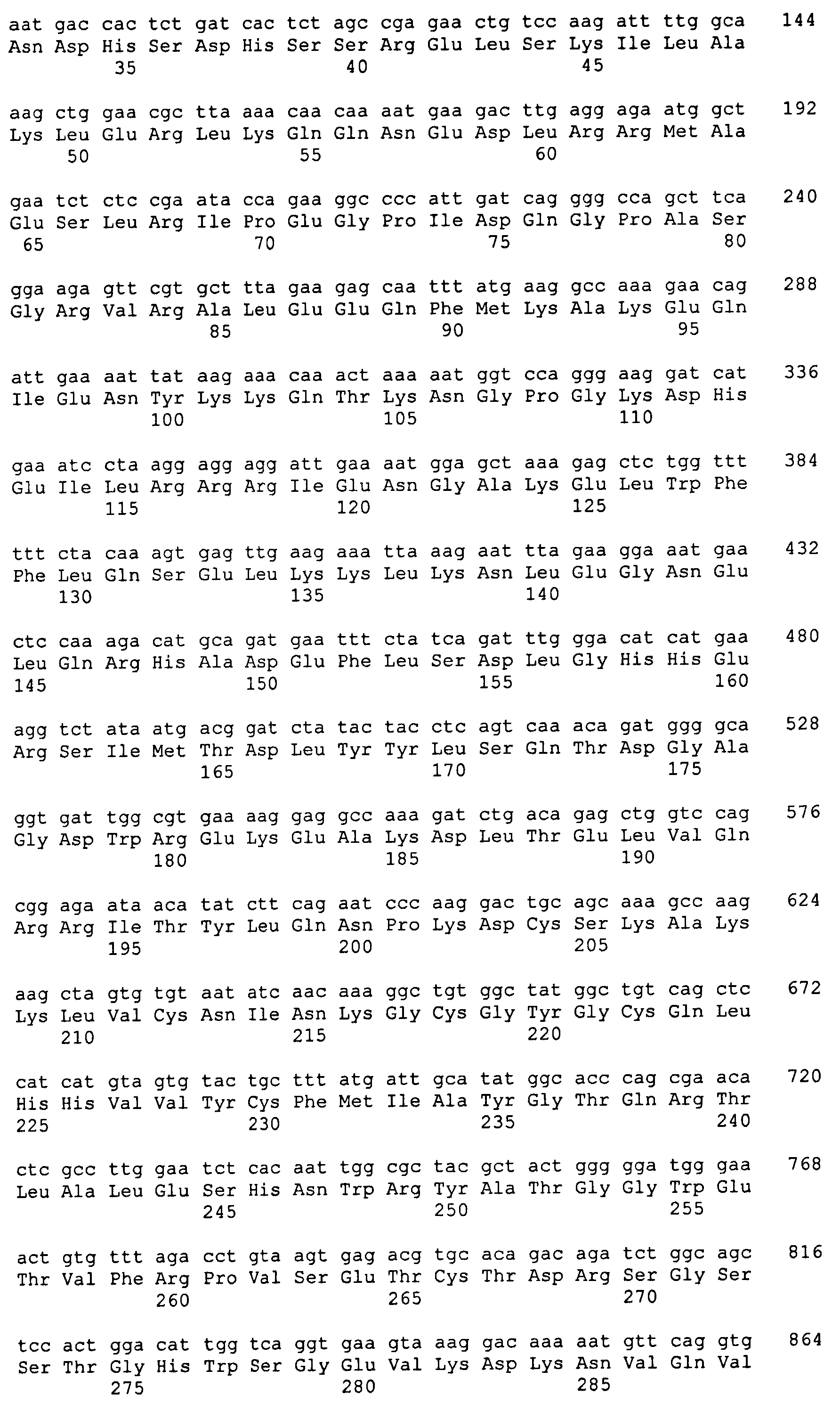
Созданную с помощью глико-инженерии линию клеток P. pastoris, YGLY1062, которая способна продуцировать рекомбинантные гликопротеины, имеющие в преобладающей степени N-гликаны Gal2GlcNAc2Man3GlcNAc2, (сходную со штаммами, описанными в опубликованной заявке на патент США № 2006/0040353, которые продуцируют гликопротеины, имеющие N-гликаны Gal2GlcNAc2Man3GlcNAc2) трансформируют вектором pSH692 с получением штамма RDP974, который продуцирует рекомбинантный ЕРО крысы (rEPO) с N-гликанами Gal2GlcNAc2Man3GlcNAc2. Штамм RDP974 схож со штаммом RDP762, описанным Hamilton и др. в Science 313, 1441-1443 (2006), который продуцирует ЕРО крысы, имеющий N-гликаны Gal2GlcNAc2Man3GlcNAc2.
Штамм RDP974 имеет делеции в генах OCH1, PNO1, MNN4B и BMT2 и включает ДНК, кодирующую полноразмерный переносчик УДФ-GlcNAc Kluyveromyces lactis, переносчик УДФ-GlcNAc M. musculus, УДФ-галактозо-4-эпимеразу S. cerevisiae и переносчик УДФ-галактозы D. melanogaster; и ДНК, кодирующую каталитический домен α1,2-маннозидазы I M. musculus, слитую с ДНК, кодирующей аминокислоты 1-36 лидерной последовательности MNN2 S. cerevisiae; ДНК, кодирующую каталитический домен β1,2-GlcNAc-трансферазы I (GnTI) H. sapiens, слитую с ДНК, кодирующей аминокислоты 1-36 лидерной последовательности MNN2 S. cerevisiae; ДНК, кодирующую каталитический домен маннозидазы II Drosophila melanogaster, слитую с ДНК, кодирующей аминокислоты 1-36 лидерной последовательности MNN2 S. cerevisiae; ДНК, кодирующую каталитический домен β1,2-GlcNAc-трансферазы II (GnTII) Rattus norvegicus, слитую с ДНК, кодирующей аминокислоты 1-97 лидерной последовательности MNN2 S. cerevisiae; и ДНК, кодирующую каталитический домен β1,4-галактозилтрансферазы (GalTI) H. sapiens, слитую с ДНК, кодирующей аминокислоты 1-58 лидерной последовательности KRE2 (MNTI) S. cerevisiae. В опубликованной заявке на патент США № 2006/0040353 описываются способы получения линий клеток Pichia pastoris, которые продуцируют галактозилированные гликопротеины в низших дрожжах (смотри также патент США № 7029872, опубликованные заявки на патенты США № 2004/0018590, 2004/0230042, 2005/0208617, 2004/0171826, 2006/0286637 и 2007/0037248 и Hamilton et al., Science 313, 1441-1443 (2006)).
Штамм RDP974 затем используют в качестве штамма-хозяина для введения ДНК для каскада реакций фукозилирования в векторе pSH1022. Вкратце, 10 мкг контрольной плазмиды pSH995 или плазмиды для каскада реакций фукозилирования pSH1022 расщепляют ферментом рестрикции SfiI для линеаризации вектора и трансформируют с помощью электропорации в штамм-хозяина RDP974. Трансформированные клетки помещают на чашки с YPD, содержащей 100 нг/мл нур-сеотрицина, и инкубируют при 26°С в течение пяти дней. Впоследствии несколько клонов отбирают и анализируют на перенос фукозы на N-гликаны rEPO. Штамм, трансформированный контрольным вектором, обозначают YSH660, в то время как штамм, трансформированный pSH1022 и демонстрирующий перенос фукозы, обозначают YSH661.
Как правило, экспрессию белков выполняют путем выращивания трансформированных штаммов при 26°С в 50 мл забуференной, комбинированной с глицерином среды (BMGY), состоящей из 1% дрожжевого экстракта, 2% пептона, 100 мМ калийфосфатного буфера, рН 6,0, 1,34% дрожжевого азотистого основания, 4×10-5% биотина и 1% глицерина, в качестве среды для роста. Индукцию экспрессии белка выполняют в 5 мл забуференной, комбинированной с метанолом среде (BMMY), состоящей из 1,5% метанола вместо глицерина в BMGY.
Рекомбинантный rEPO экспрессируют, как описано выше, и подвергают очистке на колонке с Ni-хелатным соединением, как описано Choi и др. (Proc. Natl. Acad. Sci. U.S.A. 100, 5022 (2003) и Hamilton и др. (Science 301, 1244 (2003)). Результирующий белок анализируют с помощью электрофореза в SDS-ПААГ (Laemmli, Nature 227, 680 (1970)) и окрашивают для визуализации кумасси синим. Фукозу удаляют in vitro расщеплением с помощью обработки α-1,6-фукозидазой (Sigma-Aldrich, St. Louis, MO), как рекомендовано производителем.
Для анализа гликанов гликаны высвобождают из rEPO обработкой N-гликозидазой F (Choi et al. (2003), Hamilton et al. (2003)). Высвободившиеся гликаны анализируют с помощью масс-спектрометрии MALDI/Time-of-flight (TOF) для подтверждения структуры гликанов (Choi et al. (2003)). Для определения относительного количества присутствующих фукозилированных гликанов высвободившиеся с помощью N-гликозидазы F гликаны подвергают мечению 2-аминобензидином (2-АВ) и анализируют с помощью HPLC (Choi et al. (2003)). Процент фукозилированных и нефукозилированных гликанов рассчитывают путем сравнения площади пика каждой разновидности до и после обработки фукозидазой.
Анализ N-гликанов, продуцируемых в штамме YSH661, полученном по существу, как описано выше, показал, что этот штамм продуцирует рекомбинантный rEPO, включающий N-гликаны Gal2GlcNAc2Man3GlcNAc2(Fuc). На фиг. 4А, на которой демонстрируются результаты анализа MALDI-TOF N-гликанов на rEPO, продуцируемом в штамме YSH661, демонстрируется, что этот штамм продуцировал N-гликаны, включающие N-гликаны Gal2GlcNAc2Man3GlcNAc2(Fuc) и Gal2GlcNAc2Man3GlcNAc2. N-гликаны Gal2GlcNAc2Man3GlcNAc2(Fuc) находятся внутри прямоугольника. На фиг. 4В, на которой демонстрируются результаты анализа MALDI-TOF N-гликанов на rEPO, продуцируемом в контрольном штамме YSH660 (без каскада реакций фукозилирования), демонстрируется, что этот штамм продуцировал только нефукозилированные N-гликаны, включающие только Gal2GlcNAc2Man3GlcNAc2.
ПРИМЕР 2
Штамм Pichia pastoris, способный продуцировать гликопротеины, имеющие N-гликаны NANA2Gal2GlcNAc2Man3GlcNAc2(Fuc), можно создать введением вектора pSH1022 в штамм Pichia pastoris, способный продуцировать гликопротеины, имеющие N-гликаны NANA2Gal2GlcNAc2Man3GlcNAc2. Например, вектор pSH1022, содержащий гены, кодирующие компоненты каскада реакций фукозилирования, можно трансформировать в штамм YSH597, который продуцирует ЕРО крысы, имеющий N-гликаны NANA2Gal2GlcNAc2Man3GlcNAc2, и описан в предварительной заявке на патент США № 60/801688 и Hamilton и др. в Science 313, 1441-1443 (2006). ЕРО крысы, продуцируемый в этом штамме при индукции, будет включать N-гликаны NANA2Gal2GlcNAc2Man3GlcNAc2(Fuc).
Следующим описанием обеспечивается предсказанный способ введения генов, кодирующих ферменты каскада реакций сиалирования, в штамм YSH661 примера 1.
Открытые рамки считывания для УДФ-N-ацетилглюкозамин-2-эпимеразы/N-ацетилманнозаминкиназы (GNE) Homo sapiens, N-ацетилнейраминат-9-фосфатсинтазы (SPS) H. sapiens, синтазы ЦМФ-сиаловой кислоты (CSS) H. sapiens, переносчика ЦМФ-сиаловой кислоты (CST) Mus musculus и аминокислоты 40-403 α-2,6-сиалилтрансферазы (ST) M. musculus анализируют с помощью программного обеспечения GeneOptimizer и подвергают оптимизации кодонов для экспрессии в P. pastoris (GeneArt, Regensburg, Германия). Результирующую синтетическую ДНК для GNE, SPS, CSS и CST создают с совместимыми с рестрикционными ферментами BsaI и HpaI 5'- и 3'-концами соответственно, клонируют в «челночный» вектор и обозначают pGLY368, 367, 366 и 369 соответственно. Синтетическую ДНК для ST создают с совместимыми с рестрикционными ферментами AscI и PacI 5'- и 3'-концами, соответственно, клонируют в «челночный» вектор и обозначают pSH660. Для создания экспрессирующих SPS, CSS и CST кассет векторы pGLY367, 366 и 369 расщепляют BsaI и HpaI для выделения фрагментов размерами 1,1, 1,3 и 1,0 т.п.о, которые обрабатывают Т4 ДНК-полимеразой для удаления одноцепочечных концевых избыточностей. Впоследствии эти фрагменты субклонируют в векторы pGLY359, 17 и 363, предварительно расщепленные NotI и AscI для первого из указанных векторов и NotI и PacI для двух последних из указанных векторов и обработанные Т4 ДНК-полимеразой. Результирующий вектор pSH819 содержит SPS в кассете PpPMA1prom-PpPMA1tt, фланкированной рестрикционными сайтами для PacI; pSH824 содержит CSS в кассете PpGAPDH-ScCYCtt, фланкированной рестрикционными сайтами для BglII и BamHI на 5'- и 3'-конце, соответственно; и pGLY372 содержит CST в кассете PpPMA1prom-PpPMA1tt, фланкированной рестрикционными сайтами для RsrII. Для создания каталитического домена ST, слитого с сигналом, определяющим местонахождение в дрожжах, район Mnt1 для направленной доставки в S. cerevisiae амплифицируют с геномной ДНК, используя Taq ДНК-полимеразу (Promega, Madison, WI) и праймеры ScMnt1-прямой и ScMnt1-обратный (5'-GGGCGGCCGCCACCATGGCCCTCTTTCTCAGTAAGAGACT GTTGAG-3' (SEQ ID NO: 25) и 5'-CCGGCGCGCCCGATGACTTGTTGTTCAGGGGATATAGATCCTG-3' (SEQ ID NO: 26), соответственно). Используемыми условиями являются 94°С в течение 3 минут, 1 цикл; 94°С в течение 30 секунд, 55°С в течение 20 секунд, 68°С в течение 1 минуты, 30 циклов; 68°С в течение 5 минут, 1 цикл. Результирующий фрагмент размером 174 п.о., содержащий рестрикционные сайты для NotI и AscI на 5'- и 3'-конце, соответственно, субклонируют в рамке 5' от оптимизированного в отношении использования кодонов ST, создавая вектор pSH861. Впоследствии этот вектор расщепляют NotI и PacI для выделения фрагмента размером 1,3 т.п.о., содержащего ST-слияние, обрабатывают Т4 ДНК-полимеразой и субклонируют в pGLY361, приготовленный расщеплением NotI и PacI и обработанный Т4 ДНК-полимеразой. Результирующий вектор, содержащий ST-слияние в кассете PpTEFprom-PpTEFtt, фланкированной рестрикционными сайтами для SgfI, обозначают pSH893.
Вектор для экспрессии в дрожжах pSH823, содержащий промотор GAPDH P. pastoris и терминатор транскрипции CYC S. cerevisiae, предназначен для интеграции в геном Pichia 3' от ORF Trp2. Фрагмент размером 2,6 т.п.о., кодирующий экспрессирующую PMA-CST кассету, вырезают из pGLY372, используя фермент рестрикции RsrII, и субклонируют в pSH823, расщепленный тем же ферментом. Результирующий вектор, в котором экспрессирующие PMA-CST и GAPDH кассеты располагаются в одном и том же направлении, обозначают pSH826. Впоследствии этот вектор расщепляют рестрикционными ферментами NotI и PacI и одноцепочечные концевые избыточности удаляют с использованием Т4 ДНК-полимеразы. В эту линеаризованную конструкцию субклонируют фрагмент размером 2,2 т.п.о. GNE, выделенный из pGLY368 путем расщепления BsaI и HpaI и обработанный Т4 ДНК-полимеразой для удаления одноцепочечных концевых избыточностей. Этот вектор обозначают pSH828. Впоследствии этот вектор расщепляют PacI, в который субклонируют PacI-фрагмент размером 2,7 т.п.о. pSH819, кодирующий экспрессирующую PMA-SPS кассету. Получаемый вектор, в котором экспрессирующая PMA-SPS кассета располагается в противоположном от экспрессирующей GAPDH кассеты направлении, обозначают pSH830. На этой стадии маркер URA5 замещают HIS1 путем вырезания фрагмента URA5 размером 2,4 т.п.о из pSH830, используя XhoI, и замены его фрагментом HIS1 размером 1,8 т.п.о. из pSH842, расщепленного тем же ферментом. Результирующий вектор, в котором ORF HIS1 располагается в одинаковом с экспрессирующей GAPDH-GNE кассетой направлении, обозначают pSH870. Впоследствии этот вектор расщепляют BamHI и субклонируют фрагмент размером 2,1 т.п.о. из pSH824, выделенный расщеплением BamHI и BglII, содержащий экспрессирующую GAPDH-CSS кассету. Образуемый вектор, в котором вновь введенная экспрессионная кассета располагается в противоположном от экспрессирующей GAPDH-GNE кассеты направлении, обозначают pSH872. Затем экспрессионную кассету размером 2,2 т.п.о., содержащую TEF-ST, вырезают с помощью SgfI из pSH893 и субклонируют в pSH872, расщепленный тем же ферментом. Образуемый вектор, в котором экспрессирующая TEF-ST кассета располагается в противоположном от экспрессирующей GAPDH-GNE кассеты направлении, обозначают pSH926.
Вектор pSH926 трансформируют в штамм YSH661, который затем способен продуцировать ЕРО крысы, имеющий N-гликаны NANA2Gal2GlcNAc2Man3GlcNAc2.
ПРИМЕР 3
Штамм Pichia pastoris, способный продуцировать ЕРО человека, имеющий N-гликаны NANA2Gal2GlcNAc2Man3GlcNAc2(Fuc), можно создать введением вектора pSH1022 в штамм Pichia pastoris, способный продуцировать ЕРО человека, имеющие N-гликаны NANA2Gal2GlcNAc2Man3GlcNAc2. Например, вектор pSH1022, содержащий гены, кодирующие компоненты каскада реакций фукозилирования, можно трансформировать в штамм, который способен продуцировать гликопротеины, имеющие N-гликаны NANA2Gal2GlcNAc2Man3GlcNAc2, такой как штамм YSH597, описанный Hamilton и др. в Science 313, 1441-1443 (2006), или YSH661 примера 2, включающий гены, кодирующие ферменты каскада реакций сиалирования, но с заменой ДНК, кодирующей ЕРО крысы, ДНК, кодирующей ЕРО человека. Этот штамм будет в таком случае продуцировать ЕРО человека, имеющий N-гликаны NANA2Gal2GlcNAc2Man3GlcNAc2(Fuc).
Хотя настоящее изобретение описывается здесь относительно иллюстрируемых вариантов осуществления, должно быть понятно, что настоящее изобретение не ограничивается ими. Специалисты, обладающие средним уровнем компетентности в данной области техники и обращающиеся к представленным здесь учениям, идентифицируют дополнительные модификации и варианты осуществления в пределах объема настоящего изобретения. Поэтому настоящее изобретение ограничивается только присоединенной здесь формулой изобретения.
Реферат
Изобретение относится к области биотехнологии, в частности к рекомбинантной клетке-хозяину Pichia pastoris и гибридному вектору, который обеспечивает получение данной клетки-хозяина. Рекомбинантная клетка-хозяин Pichia pastoris включает каскад реакций фукозилирования, где ферменты составляют указанный каскад реакций. Указанная клетка-хозяин Pichia pastoris включает нуклеиновые кислоты, кодирующие ГДФ-маннозо-дегидратазу (GMD), ГДФ-кетодезоксиманнозо-эпимеразы/ГДФ-кетодезоксигалактозо-редуктазу (FX), переносчик ГДФ-фукозы (GFTr) и слитый белок, который содержит каталитический домен α1,6-фукозилтрасферазы ЕС 2.4.1.68, слитый с аминокислотами 1-36 Mnn2 S.cerevisiae. Гибридный вектор включает регуляторные элементы ДНК, которые являются функциональными в клетке-хозяине и которые функционально связаны с кодирующей последовательностью ДНК, кодирующей слитый белок, включающий аминокислоты 1-36 Mnn2 S.cerevisiae, слитый с каталитическим доменом α1,6-фукозилтрасферазы ЕС 2.4.1.68. Указанным вектором трансформируют клетку-хозяина Pichia pastoris. Предложенное изобретение позволяет создать клетку-хозяина Pichia pastoris, которая будет продуцировать гликопротеины с фукозилированными N-гликанами. 2 н. и 6 з.п. ф-лы, 6 ил., 3 пр.
Формула
где указанный вектор, будучи совместно трансформированным в клетку Pichia pastoris с нуклеиновыми кислотами, кодирующими ГДФ-маннозо-дегидратазу (GMD), ГДФ-кетодезоксиманнозо-эпимеразы/ГДФ-кетодезоксигалактозо-редуктазу (FX) и переносчик ГДФ-фукозы (GFTr), обеспечивает получение клетки-хозяина Pichia pastoris no n.1.
Комментарии