Ретровирусный вектор на основе вируса мышиного лейкоза (mlv) (варианты) - RU2203321C2
Код документа: RU2203321C2
Чертежи



Описание
Изобретение относится к улучшенным ретровирусным векторам, применяемым в генотерапии. В частности, данное изобретение относится к безопасным и эффективным экспрессирующим векторам, в составе которых все ретровирусные гены, такие как гены gag, env и pol, полностью делетированы; в составе которых гетерологичный интрон, акцепторный сайт сплайсинга и/или некодирующая последовательность встроены выше сайта клонирования, предназначенного для чужеродного гена; в составе которого гетерологичный внутренний промотор или внутренний сайт связывания на рибосоме (здесь и далее обозначаемого как "IRES") встроены после сайта клонирования; и в составе которого полноразмерная U3-последовательность 5'-LTR (длинные концевые повторы) или мощный гетерологичный промотор контролирует экспрессию чужеродного гена.
ПРЕДПОСЫЛКИ
Ретровирусные векторы используются в применении к генотерапии чаще, чем любые другие типы
векторов: при этом они используются более чем в половине применяемых клинических методик в мире (Wiley - J. Gene Med. Website: http: //www. wiley. co. uk/genetherapy). Хотя преимущественно применяются
векторы, основанные на вирусе лейкоза мыши (здесь и далее обозначаемый как "MLV"), при этом существует достаточно много проблем в связи с применением ретровирусных векторов в клинической практике.
Наиболее серьезная проблема связана с вопросами безопасности: ретровирусный вектор является одним из вирусных векторов и, следовательно, может конвертироваться в клетках в способный реплицироваться
ретровирус (здесь и далее обозначаемый как "СРР"). Соответственно наибольшее беспокойство вызывает образование СРР в результате гомологичной рекомбинации.
Все доступные ретровирусные векторы включают значительные по длине вирусные кодирующие последовательности. Поскольку эти последовательности также присутствуют в составе генома "пакующих" клеток, в которых вирусы "упаковываются в оболочки" и из которых векторы выходят, то было подтверждено, что гомологичная рекомбинация может происходить между одинаковыми нуклеотидными последовательностями генома "упаковочной клетки" и вектора, в результате чего инициируется образование СРР, ранее описанное Миллером с соавт. (Miller et al., 1990, Human Gene Therapy, 1, 5).
Два типа основанных на вирусе MLV ретровирусных вектора (векторы серии LN и вектор MFG) наиболее часто находят использование в генотерапии (Miller & Roseman, 1989, Biotechniques, 1, 980-990; Dranoff et al., 1993, Proc. Natl. Acad. Sci. USA, 90, 3539-3543). Хотя экспрессия чужеродного гена в векторах серии LN находится под контролем внутреннего гете-рологичного промотора или LTR, то в вектор MFG уровень транскрипции контролируется LTR, а чужеродный ген экспрессируется в форме либо геномной РНК, либо сплайсированной субгеномной РНК. Векторы серии LN, которые обычно рассматриваются как ретровирусные векторы первого поколения, включают 420-нуклеотидную кодирующую последовательность гена gag. Хотя этот участок, как предполагается, играет важную роль в процессах упаковки вирусных частиц, было установлено, что сегмент gag может быть делетирован без какого-либо существенного проявления в процессе вирусной упаковки и в количестве вируса при соблюдении одинаковых условий (Kim et al., 1998, J. Virol., 71, 994-1004).
По сравнению с векторами серии LN для вектора MFG известно обеспечение более стабильной и происходящей на более высоком уровне экспрессии, а также выработка более высокого титравируса в большинстве клеточных линий, производных от клеток человека или мыши (Byun et al., 1996, Gene Therapy, 3, 780-788). Однако вектор MFG включает еще больше вирусных кодирующих последовательностей (420-нуклеотидная последовательность гена gag, 377-нуклеотидная последовательность гена ро1 и 99-нуклеотидная последовательность гена env): это указывает на вероятность еще более высокой частоты инициации образования СРР по сравнению с векторами серии LN.
Для преодоления этих ограничений, связанных со стандартными ретровирусными векторами, заявители данного изобретения сконструировали ретровирусный
вектор (заявка на корейский патент 97-48095), который характеризуется рядом таких особенностей:
транскрипты клонированных генов прошли эффективный сплайсинг с обеспечением высокого уровня
экспрессированности данного гена;
нуклеотидные последовательности генов gag и env были полностью делетированы без утраты вирусного титра;
IRES были использованы для обеспечения
одновременной экспрессии двух или большего числа генов в составе вектора;
мультиклональный сайт был встроен в состав такого вектора с целью облегчения клонирования чужеродного гена.
Поскольку последовательности генов gag и env были делетированы из состава ретровирусных векторов, безопасность этих векторов увеличилась по сравнению с таковой у других ретровирусных векторов. Однако этот вектор по-прежнему включает 377-нуклеотидную кодирующую последовательность гена ро1, которая содержит акцепторный сайт сплайсинга, равно как и ее нижерасположенную последовательность, включающую 284-нуклеотидную лидерную последовательность (транскрибируемую, но не транслируемую) гена env, поскольку делеция последовательности гена ро1 приводила бы к аномальному или неполному сплайсингу. Поскольку 377-нуклеотидная последовательность ро1 также присутствует в геноме "упаковочных" линий клеток, возможность гомологичной рекомбинации, приводящей к инициации образования СРР, сохраняется в случае с этим вектором.
Для создания новых ретровирусных векторов, характеризующихся повышенной эффективностью экспрессии генов, равно как и повышенной безопасностью, авторы сконструировали ретровирусные векторы, которые не включают каких-либо кодирующих ретровирусных последовательностей. Данное изобретение осуществлено путем создания ретровирусных векторов, в составе которых последовательность гена ро1, производная от генома вируса MLV, полностью делетирована, что исключает вероятность образования СРР, опосредуемой гомологичной рекомбинацией в "упаковочной" клеточной линии; в составе которых гетерологичный интрон, акцепторный сайт сплайсинга и/или некодирующая последовательность встроены выше сайта клонирования, предназначенного для чужеродного гена, что обеспечивает максимализацию эффективности генной экспрессии; в составе которых либо 5'-LTR, либо промотор MIEP главного немедленно раннего гена iel цитомегаловируса человека (здесь и далее обозначаемого как "HCMV") используются в качестве цисконтролирующего элемента, который регулирует экспрессию чужеродного гена; и в составе которых либо гетерологичный внутренний промотор, либо IRES встроены ниже расположения сайта клонирования, что обеспечивает одновременную экспрессию двух или большего числа чужеродных генов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Объектом данного изобретения является обеспечение безопасности ретровирусных векторов, в составе которых чужеродный ген (гены) экспрессирован на повышенных уровнях: соответственно они могут быть
эффективно использованы для целей генотерапии.
В соответствии с данным изобретением указанный выше объект может быть легко получен.
Данное изобретение представляет основанные на вирусе MLV ретровирусные векторы, при том что MLV-производные последовательности, кодирующие gag, env и pol, полностью делетированы.
В одном из аспектов изобретения MLV-производные ретровирусные векторы по данному изобретению могут включать гетерологичный интрон и/или гетерологичный акцепторный сайт сплайсинга, которые встроены выше расположения сайта клонирования, предназначенного для чужеродного гена.
В другом аспекте MLV-производные ретровирусные векторы по данному изобретению могут включать гетерологичную некодирующую последовательность, встроенную выше расположения сайта клонирования, предназначенного для чужеродного гена.
Еще в одном аспекте полноразмерная последовательность U3 (пары нуклеотидов [-419] -[-1] ) из состава 5'-LTR генома вируса MLV может быть замещена гетерологичным промотором в составе MLV-производных ретровирусных векторов по данному изобретению.
Еще в одном аспекте MLV-производные ретровирусные векторы по данному изобретению могут включать гетерологичный промотор, расположенный ниже сайта клонирования, предназначенного для чужеродного гена.
Данное изобретение также относится к штаммам Е. coli, трансформированным MLV-производными ретровирусными векторами по данному изобретению.
Другие характеристики данного изобретения будут ясны из дальнейшего описания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
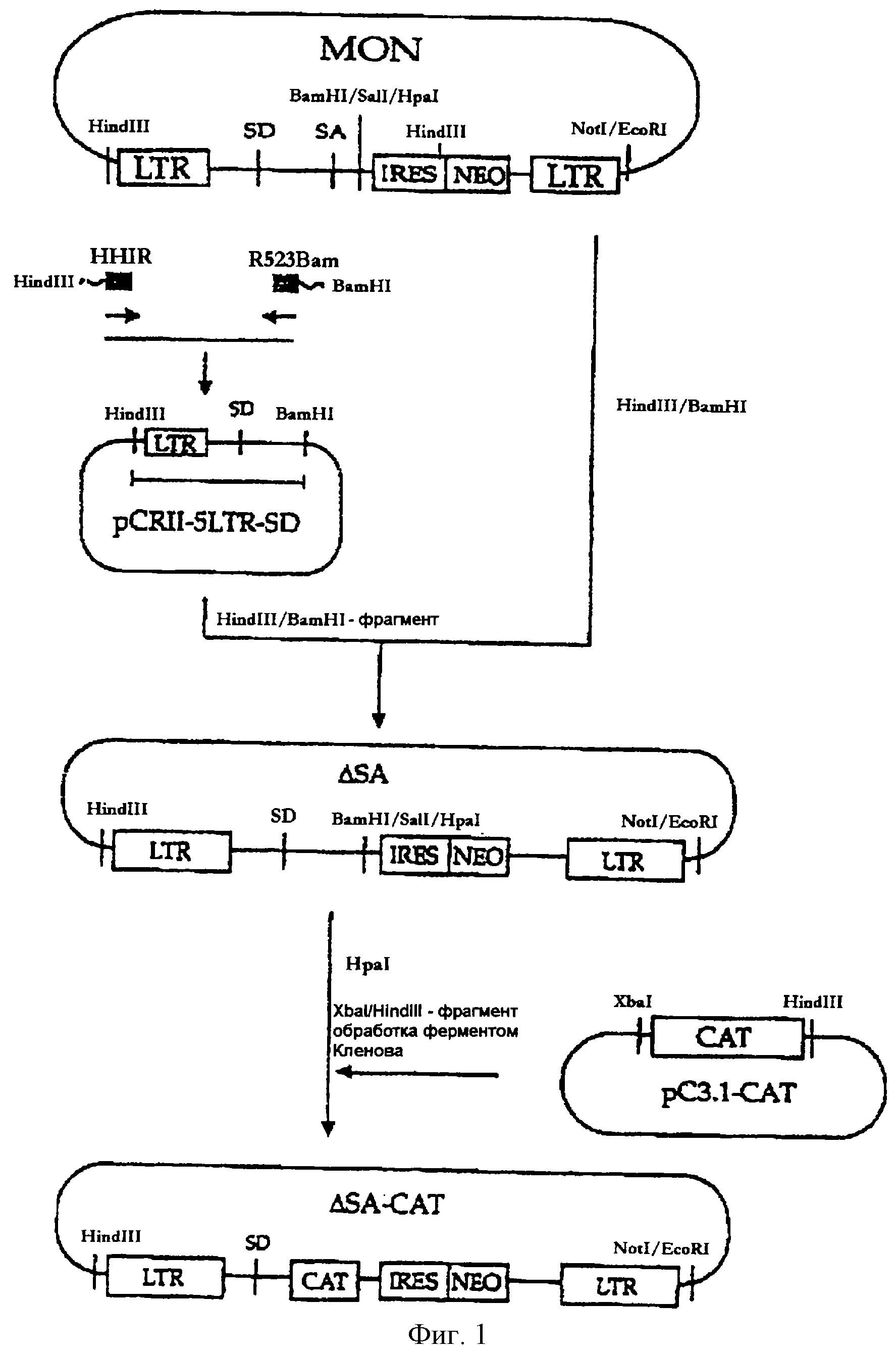
На фигуре 1 представлена методика, в соответствии с которой кодирующий сегмент гена ро1 делетируется
из вектора MON в ходе конструирования вектора ΔSА с последующим встраиванием гена CAT в состав вектора ΔSА, в результате чего образуется вектор ΔSА-САТ.
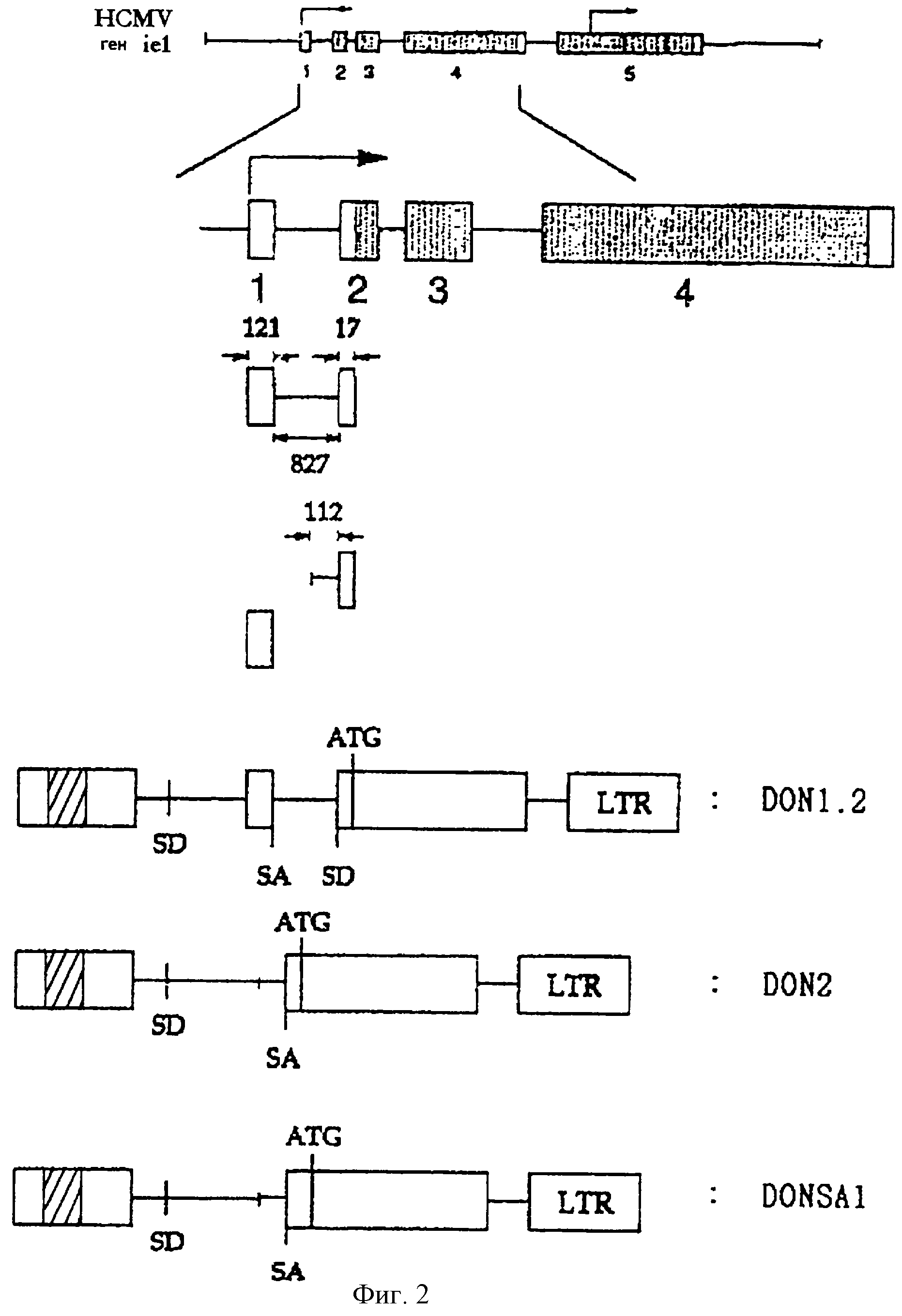
На фигуре 2 схематически изображены структуры ретровирусных векторов, включающих интрон и/или экзон гена iel (UL123) HCMV.
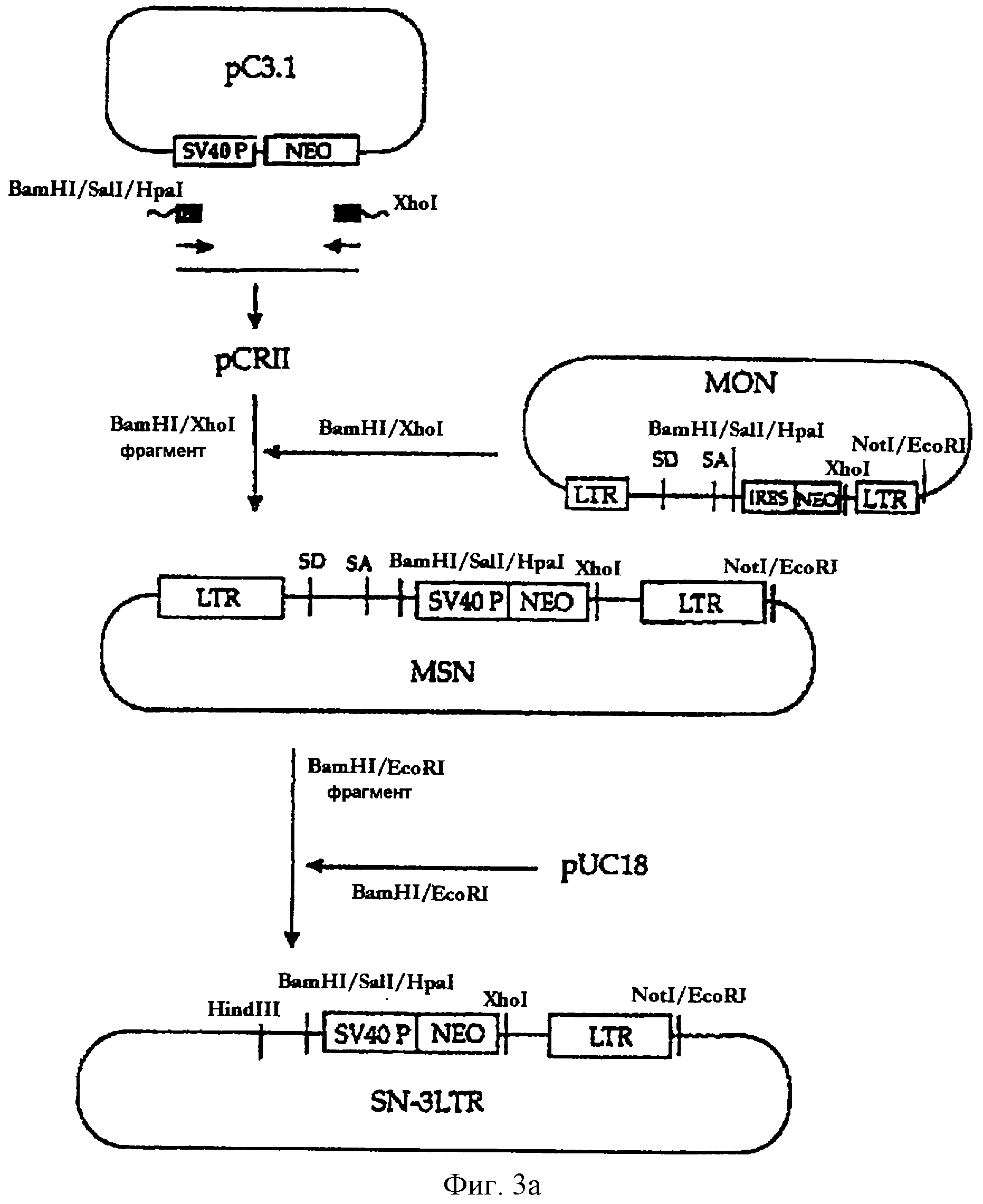
На фигуре 3а представлена методика, в соответствии с которой кассета "минимальный промотор вируса 3V4О/ген nео" замещается кассетой "IRE3-neo" с получением вектора MSN, а затем вектор 3N-3LTR конструируется путем встраивания фрагмента, включающего минимальный промотор SV40 и MLV-3'LTR в состав плазмиды pUC18.
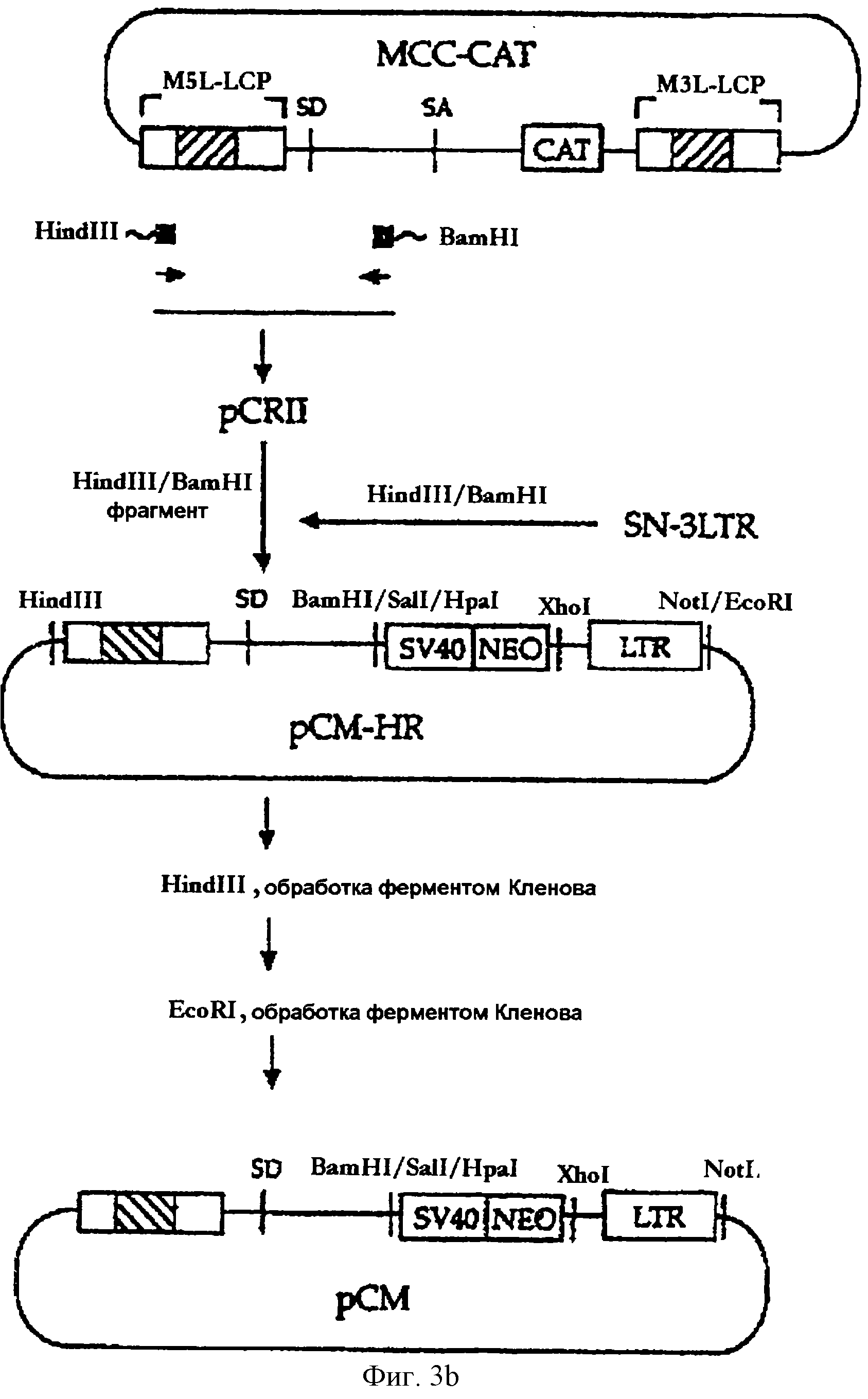
На фигуре 3b представлена методика, в соответствии с которой химерный сегмент LTR, включающий промотор главного немедленно раннего гена вируса HCMV, встраивается в состав вектора SN-3LTR.
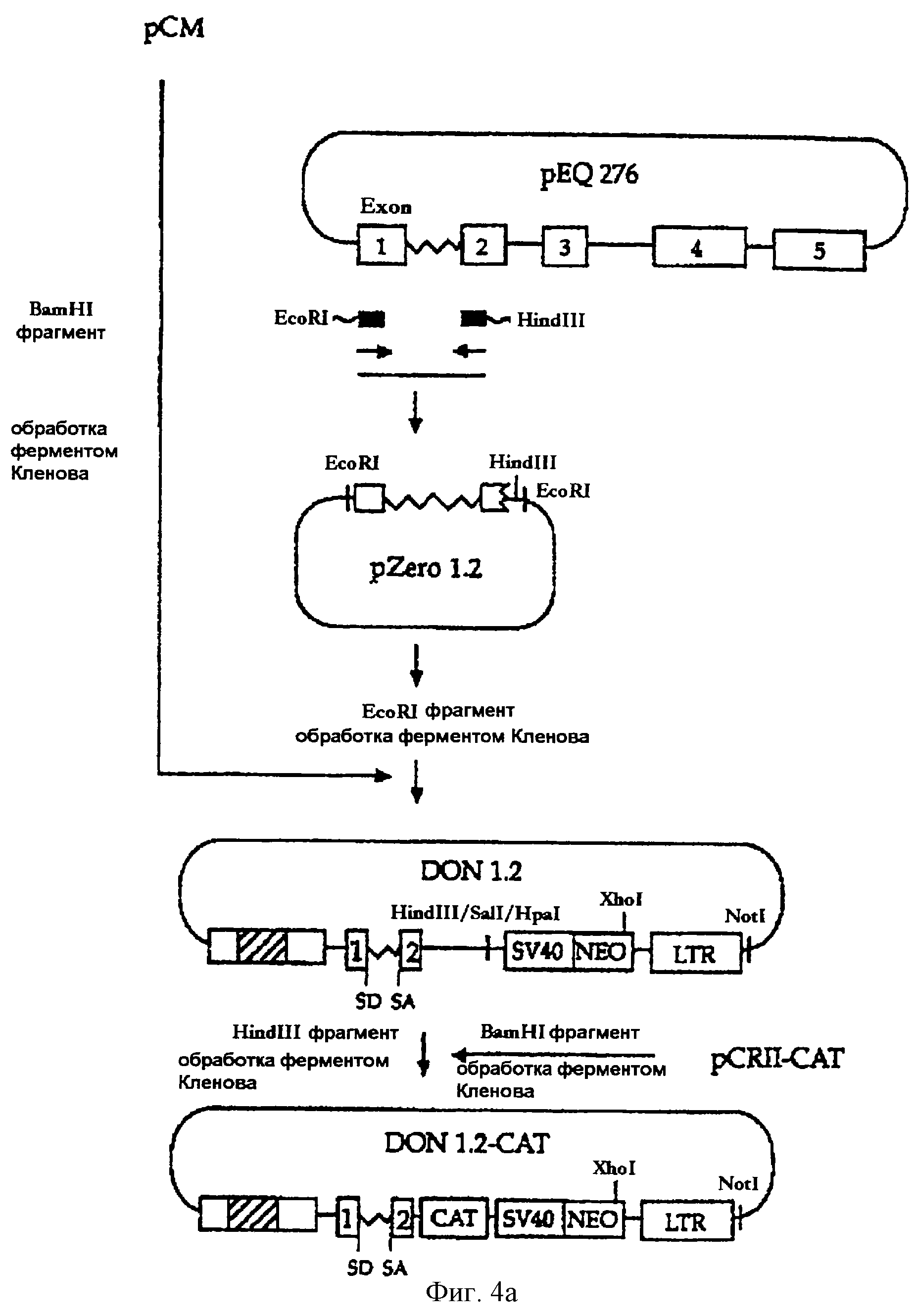
На фигуре 4а представлена методика, в соответствии с которой фрагмент ДНК, включающий 1-й экзон, интрон А и часть 2-го экзона гена iel (UL123) HCMV, вносится в состав вектора рСМ с конструированием вектора DON1.2, а затем бактериальный ген CAT встраивается в состав вектора DON1.2 с получением вектора DON1.2-CAT.
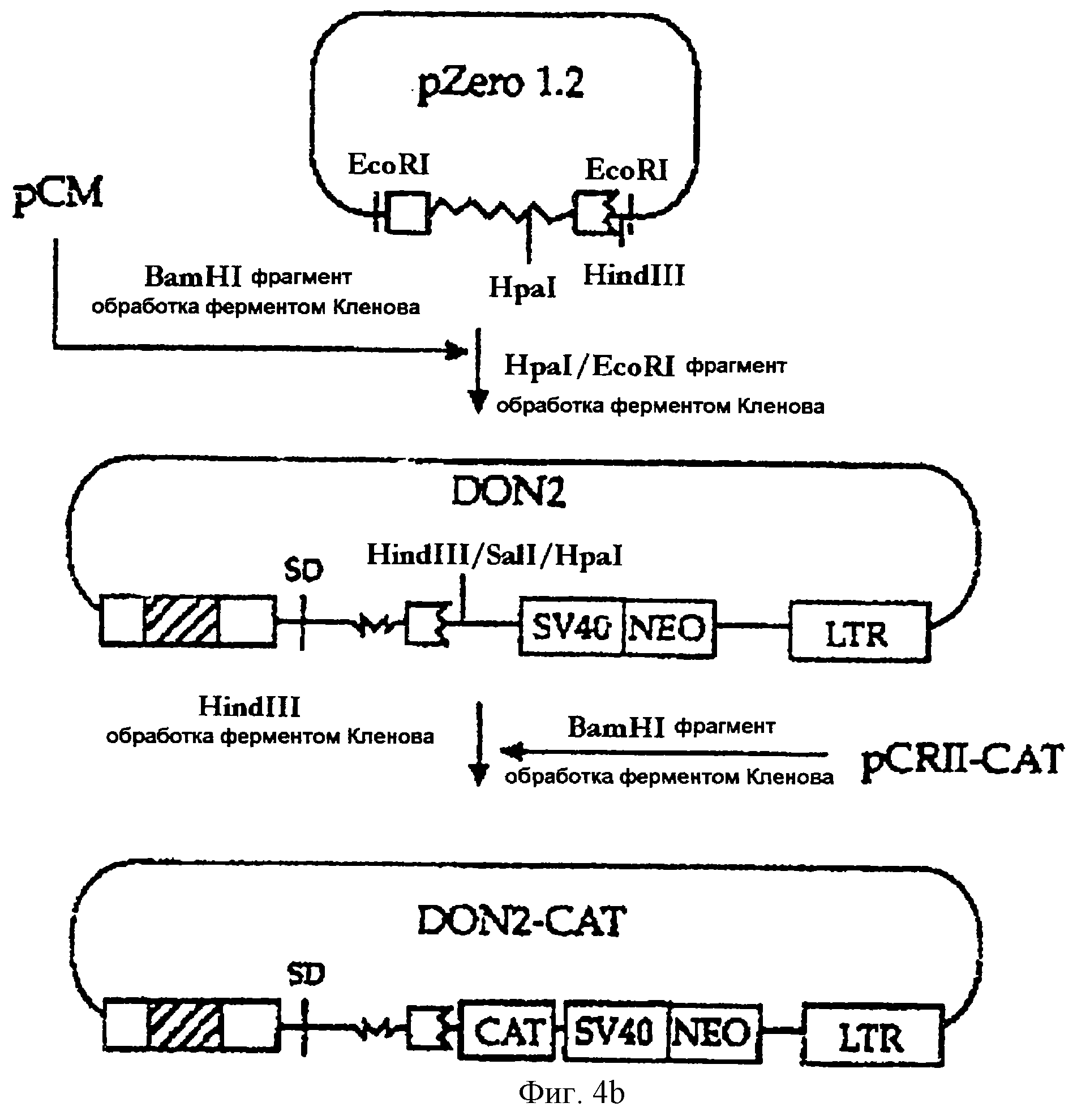
На фигуре 4b представлена методика, в соответствии с которой фрагмент ДНК, включающий часть интрона А и часть 2-го экзона гена iel (UL123) HCMV, вносится в состав вектора рСМ с конструированием вектора DON2, а затем бактериальный ген CAT встраивается в состав вектора DON2 с получением вектора DON2-CAT.
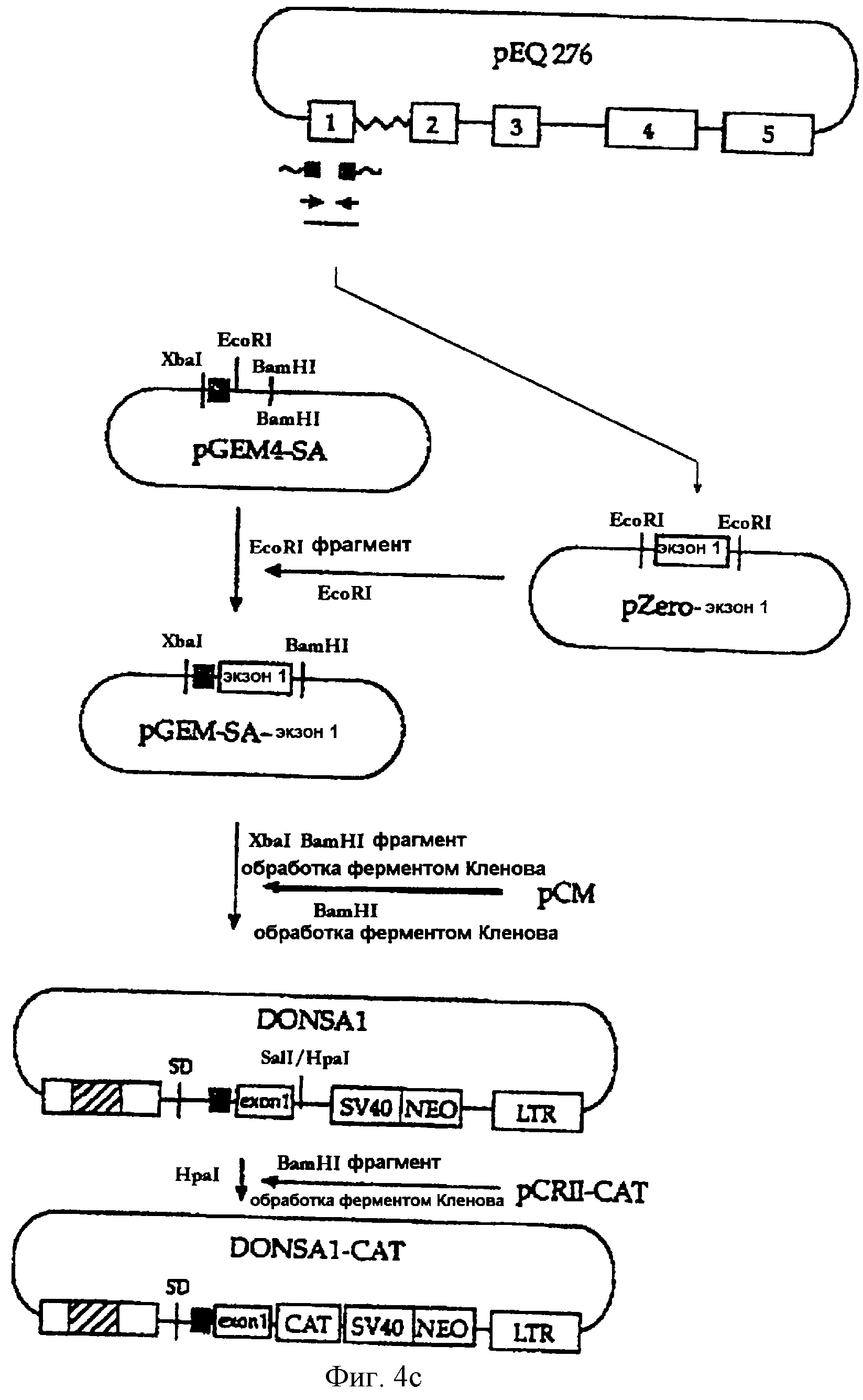
На фигуре 4с представлена методика, в соответствии с которой акцепторный сайт сплайсинга гена иммуноглобулина мыши и 1-й экзон гена iel (UL123) HCMV вносятся в состав вектора рСМ в ход конструирования вектора DONSA1, а затем бактериальный ген CAT встраивается в состав вектора DONSA1 с получением вектора DONSA1-CAT.
На фигуре 5 схематически представлены структуры ретровирусных вирусов, включающих интроны и экзоны генов человека.
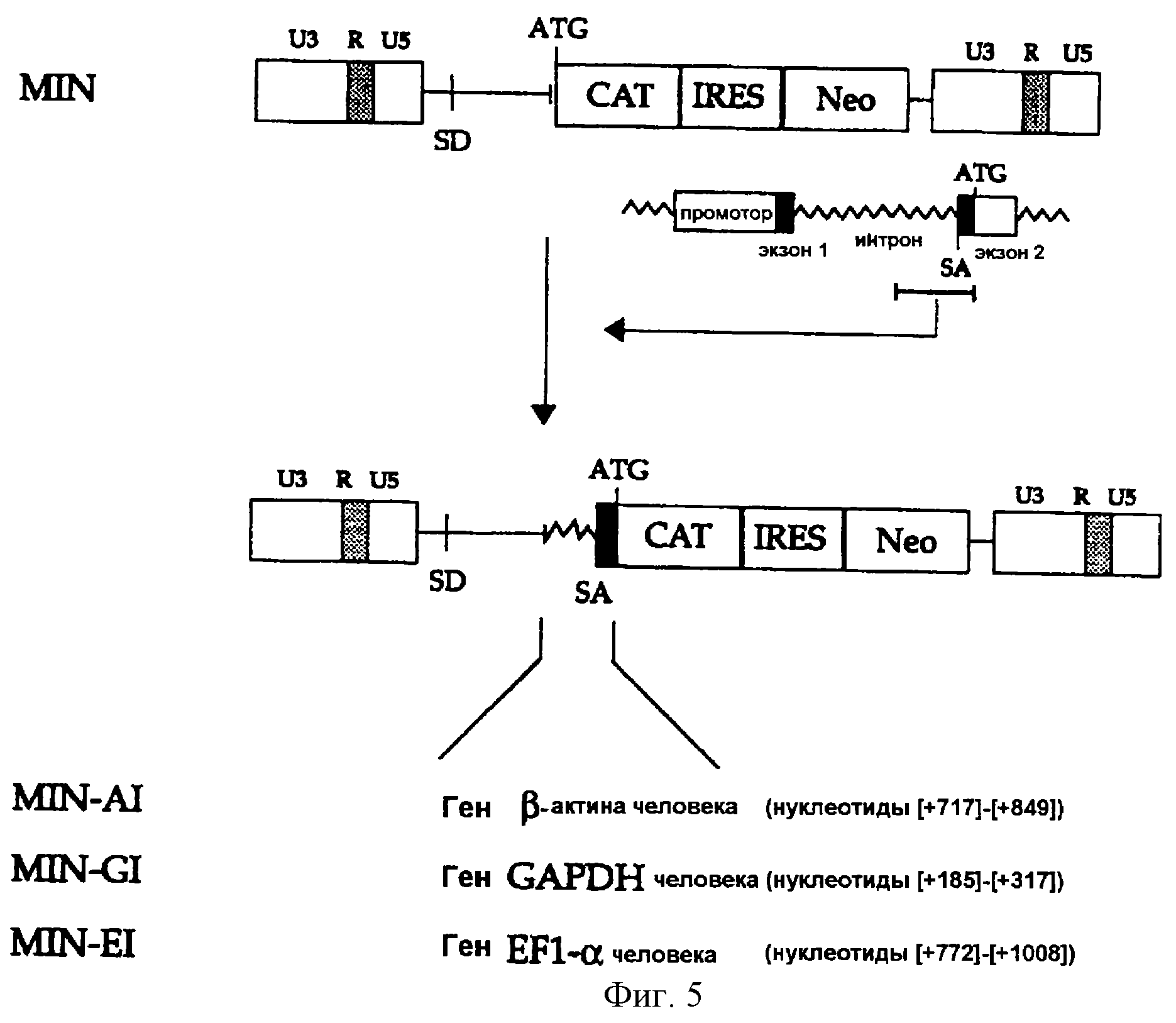
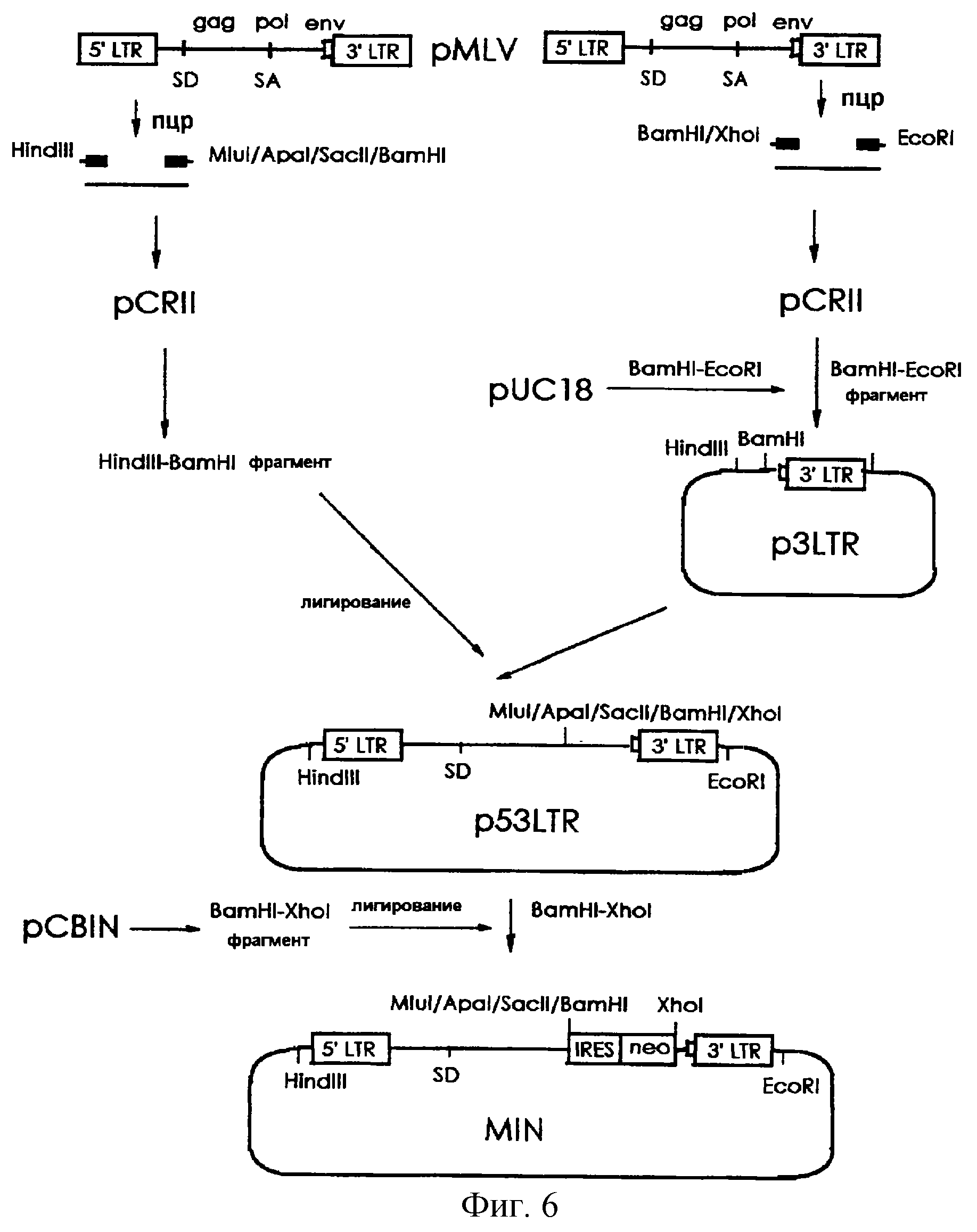
На фигуре 6 представлена методика, в соответствии с которой фрагмент ДНК, включающий последовательности MLV-5' и 3'-LTR, конструируется и встраивается в состав плазмиды pUC18 с получением вектора p53LTR, а затем вектор MIN конструируется путем встраивания кассеты IRES-neo, выделенной из плазмиды pCBIN, в состав p53LTR.
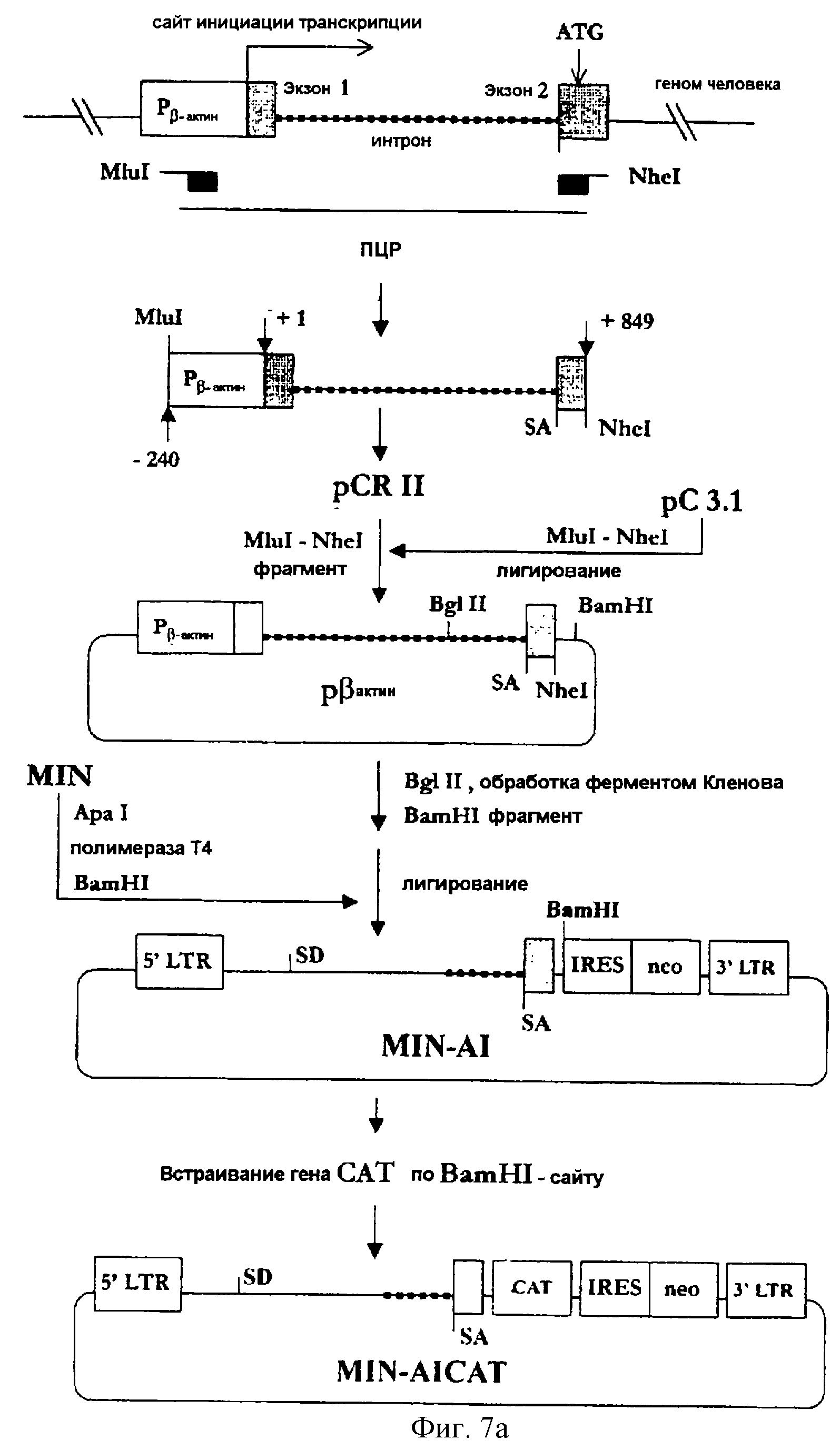
На фигуре 7а представлена методика конструирования MIN-AI, в соответствии с которой получают геномный ПЦР-продукт, включающий ген β-актина человека, а затем фрагмент ДНК, включающий часть 1-го интрона, акцепторный сайт сплайсинга и часть 2-го экзона гена β -актина человека встраивается в состав вектора MIN.
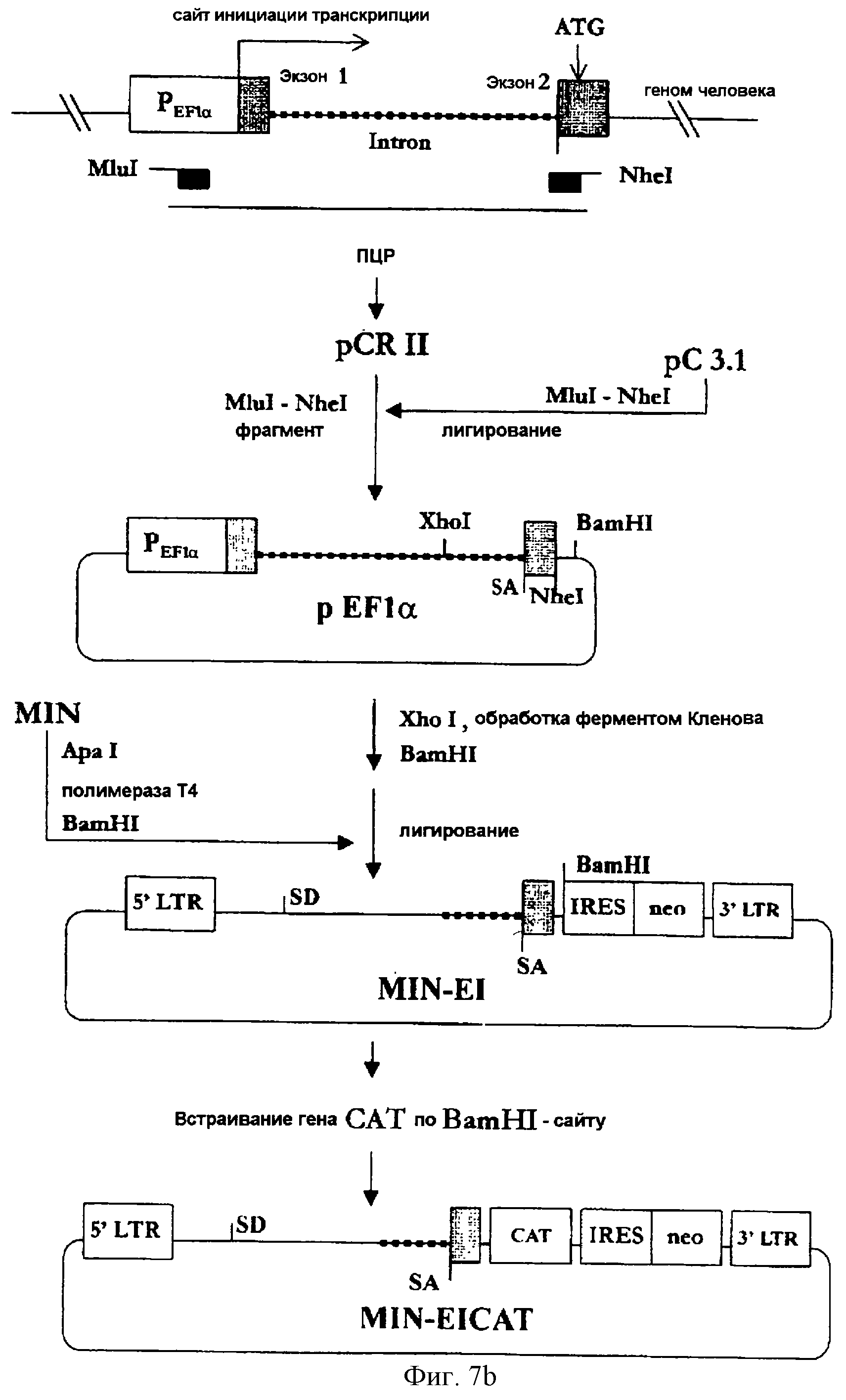
На фигуре 7b представлена методика конструирования MIN-EI, в соответствии с которой получают геномный ПЦР-продукт, включающий ген EF1α человека, а затем фрагмент ДНК, включающий часть 1-го интрона, акцепторный сайт сплайсинга и часть 2-го экзона гена EF1α человека встраивается в состав вектора MIN.
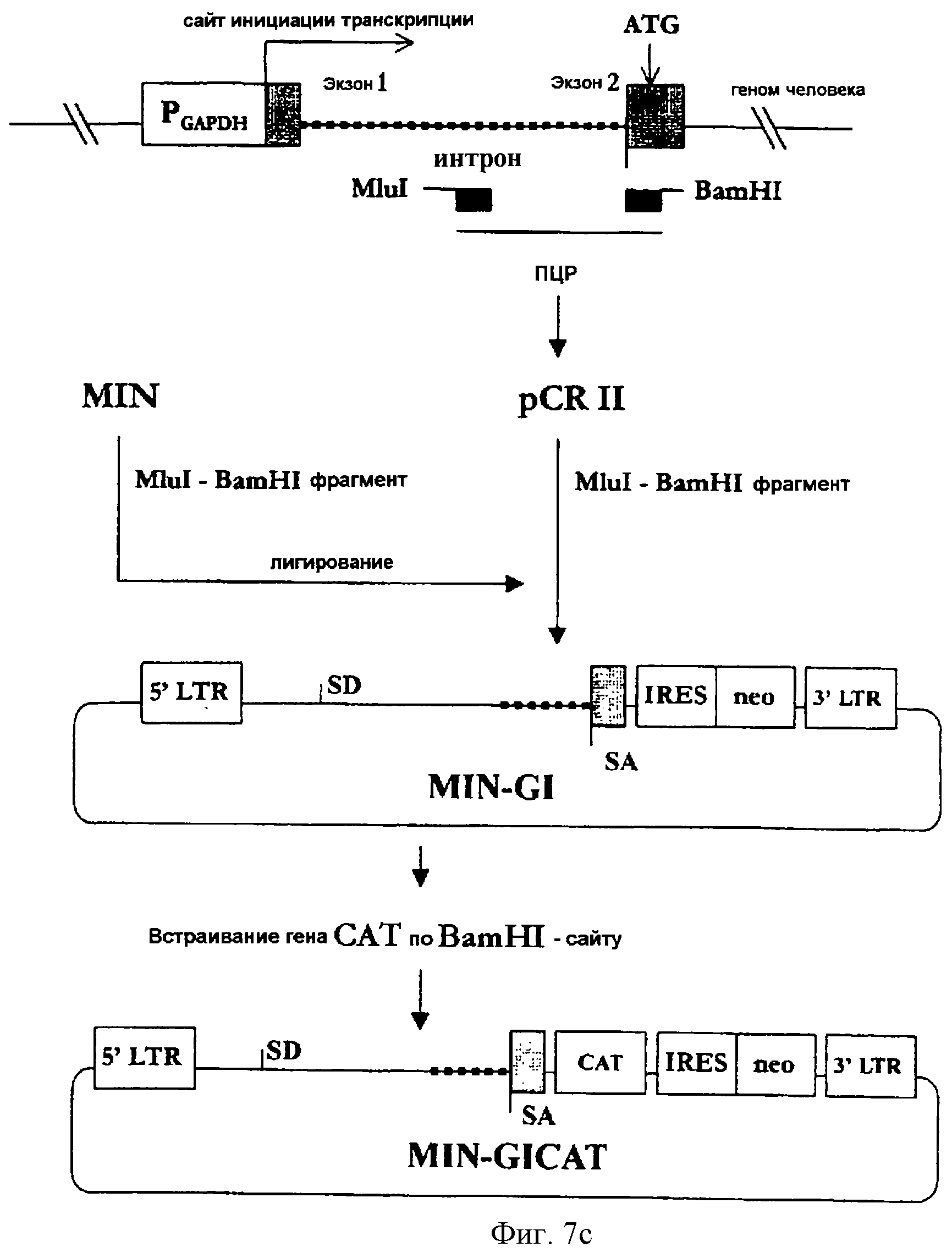
На фигуре 7c представлена методика конструирования MIN-GI, в соответствии с которой фрагмент ДНК, включающий часть 1-го интрона, акцепторный сайт сплайсинга и часть 2-го экзона гена GAPDH человека получают с применением геномной ПЦР и затем встраивают его в состав вектора MIN.
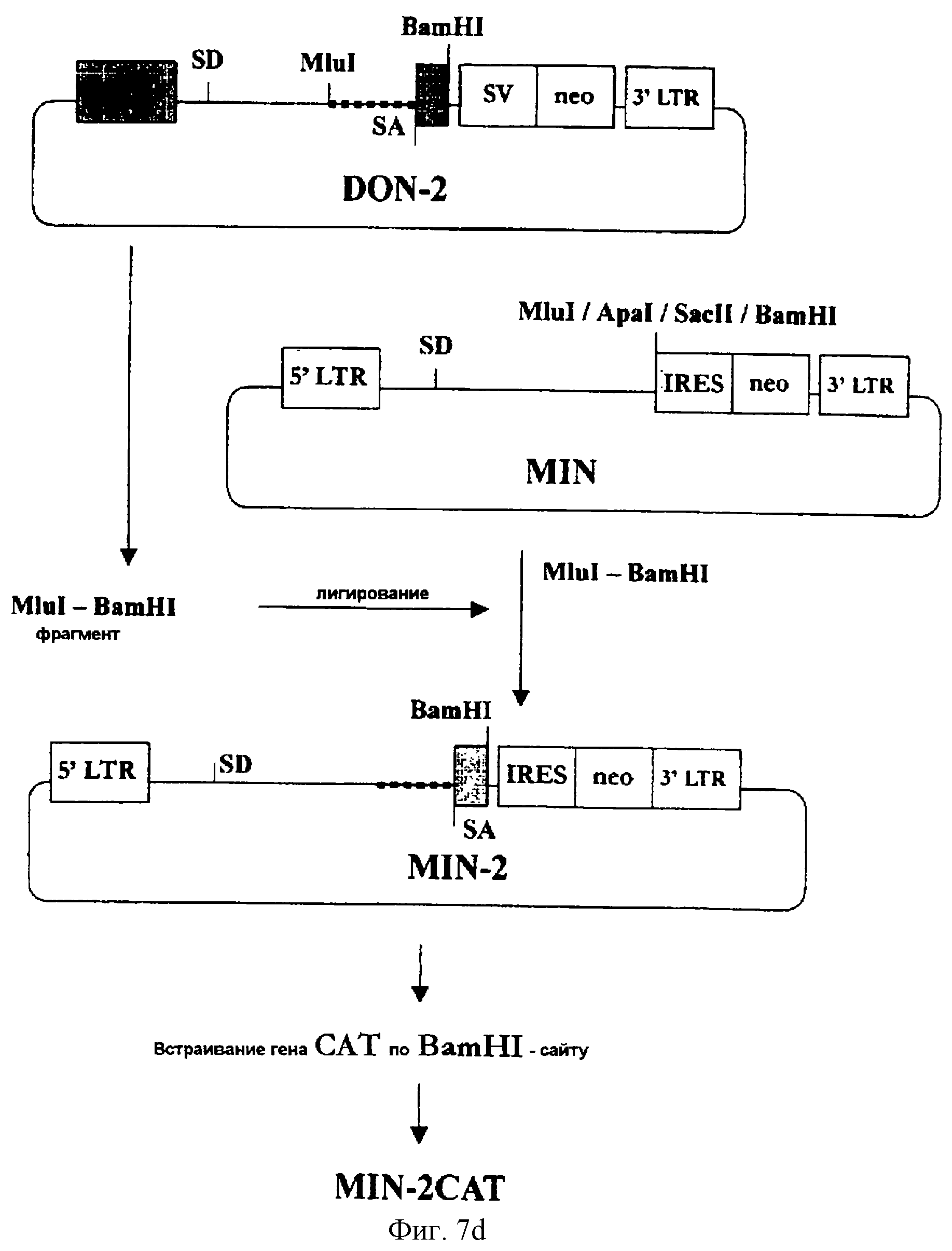
На фигуре 7d представлена методика конструирования MIN-2, в соответствии с которой фрагмент ДНК, включающий часть интрона А, акцепторный сайт сплайсинга и часть 2-го экзона гена iel (UL123) HCMV, встраивается в состав вектора MIN.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ
ОСУЩЕСТВЛЕНИЙ
Данное изобретение относится к безопасным и эффективным ретровирусным векторам, которые являются векторами на основе вируса MLV, конкретно являются производными от вирусов MON
или MIN. В векторах по данному изобретению ретровирусные гены (кодирующие нуклеотидные последовательности gag, env и pol) полностью делетированы; гетерологичный интрон, акцепторный сайт сплайсинга
и/или некодирующая последовательность встроены выше положения сайта клонирования, предназначенного для чужеродного гена; мощный гетерологичный промотор включает 5'-LTR; и гетерологичный промотор или
IRES помещены ниже положения, сайта клонирования.
В частности, поскольку кодирующая последовательность гена pol, включающая акцепторный сайт сплайсинга, полностью делетирована, вероятность гомологичной рекомбинации, которая является существенным недостатком стандартных ретровирусных векторов, при использовании векторов по данному изобретению может быть исключена.
Делетирование последовательносги гена pol приводит к снижению уровня генной экспрессии. Для восстановления сниженной эффективности экспрессии и вирусного титра данным изобретением
представляется ряд ретровирусных векторов:
Данное изобретение относится к ретровирусным векторам, в составе которых гетерологичный интрон и/или акцепторный сайт сплайсинга встроены выше
положения сайта клонирования, предназначенного для чужеродного гена с целью комплементировать делетированный акцепторный сайт сплайсинга, который перекрывается с кодирующей последовательностью гена
ро1. Все интроны и/или акцепторные сайты сплайсинга известных вирусных или клеточных генов могут быть использованы для данной цели, а предпочтительными являются интроны и/или акцепторные сайты
сплайсинга гена iel (UL123) HCMV, гена фактора элонгации 1α (здесь и далее обозначаемого как "EF1α"), гена глицеральдегид-3-фосфатдегидрогеназы (здесь и далее обозначаемого как "GAPDH"),
гена β-актина и т.п.
Данное изобретение также относится к ретровирусным векторам, в составе которых гетерологичная некодирующая последовательность встроена выше места положения сайта клонирования. Некодирующая последовательность используется для способствования эффективной трансляции и определяется как транскрибируемая последовательность ДНК, которая сама не транслируется в белок, включающая интрон и экзон с нетранслируемой последовательностью. Предпочтительно некодирующая последовательность выбирается из группы, которая включает некодирующие последовательности гена iel (UL123) HCMV, гена EF1α, гена GAPDH и гена β-актина. Встраивание гетерологичной некодирующей последовательности обеспечивает как эффективный сплайсинг транскрипта чужеродного гена, так и эффективную трансляцию субгеномной РНК.
Дополнительно данное изобретение относится к ретровирусным векторам, в составе которых гетерологичный внутренний промотор или IRES размещен ниже места положения сайта клонирования.
Наконец, данное изобретение относится к ретровирусным векторам, включающим либо полноразмерную последовательность U3 из состава 5'-LTR, либо мощный гетерологичный промотор.
Ниже дано подробное описание данного изобретения.
1. Делетирование кодирующей последовательности гена ро1: конструирование ΔSА
Для создания ретровирусных векторов, характеризующихся повышенной безопасностью, а именно ретровирусных векторов, не проявляющих активности по гомологичной рекомбинации, кодирующая последовательность
гена ро1, включая акцепторный сайт сплайсинга, была делетирована из состава вектора MON (заявка на корейский патент 97-48095), в результате чего был сконструирован мутантный делеционный вариант
- ΔSА. Дополнительно вектор ΔSА-САТ был сконструирован путем встраивания бактериального гена CAT (кодирует хлорамфениколацетилтрансферазу), используемого в качестве гена-репортера (см.
фиг.1). Далее клеточные линии, трансфицированные или трансдуцированные конструкцией ΔSА-САТ, были получены для последующего испытания в САТ-тесте. Данные САТ-теста показали, что вирусный титр
делеционного мутантного варианта был сравним с тем, который характерен для родительского вектора MON-CAT, однако уровень генной экспрессии был снижен (см. таблицу 1). Эти результаты позволили
предположить, что ген ро1, включающий акцепторный сайт сплайсинга, вовлечен в регуляцию экспрессии чужеродного гена в составе вектора.
Как ожидается, исходя из этих результатов, уровень экспрессии чужеродного гена должен повыситься тогда, когда гетерологичный акцепторный сайт сплайсинга и/или интрон будут встроены выше сайта клонирования, в котором встраивается чужеродный ген, за счет эффекта комплементирования сайта сплайсинга, делетированного вместе с последовательностью гена pol. Однако, если сплайсинг будет происходить слишком активно, соотношение субгеномной РНК и геномной РНК будет возрастать, это приведет к уменьшению вирусных титров, несмотря на высокие уровни генной экспрессии.
Для конструирования идеальных ретровирусных векторов соответственно векторы должны быть выстроены таким образом, чтобы уровни транскрипции, сплайсинга и трансляции могли бы быть сбалансированы в трансфицированных или трансдуцированных клеточных линиях. Другими словами, предпочтительными являются векторы, в составе которых чужеродный ген транскрибируется массово и количество геномной РНК сбалансировано с количеством субгеномной РНК так, чтобы геномная РНК вырабатывалась на уровне, достаточном для упаковки вирусных частиц, и чтобы субгеномная РНК могла быть транслирована в массовое количество белка, кодируемого чужеродным геном. В данном изобретении представляются такие ретровирусные векторы, в составе которых различные некодирующие последовательности, происходящие от вирусных или клеточных генов, встраиваются в состав делеционного мутантного вектора, а затем исследуются в связи с тем, обусловливает ли эта вставка какой-либо эффект в отношении титров вируса уровней генной экспрессии.
2. Встраивание гетерологичной
последовательности ДНК, усиливающей сплайсинг и эффективность трансляции: конструкции DON1.2, DON2 и DONSA1
С целью поддержания баланса уровня сплайсинга и интенсивности экспрессии гена,
равно как с целью увеличения общего выхода вирусной РНК, конструировали такие ретровирусные векторы, в которых сегмент U3 из последовательности 5'-LTR вируса MLV был замещен мощным промотором
- промотором главного немедленно раннего гена вируса HCMV, - и в которые была встроена гетерологичная некодирующая последовательность.
В предпочтительном варианте нетранслируемый экзон и/или интрон гена iel (UL123) вируса HCMV был использован в качестве гетерологичной некодирующей последовательности, которая была встроена выше сайта клонирования данного вектора (см. фиг. 2). Ранее было установлено, что экзонные и/или интронные последовательности гена iel (UL123) вируса HCMV повышают эффективность трансляции тогда, когда они встроены в состав экспрессирующего вектора.
В частности, вектор рСМ был сконструирован следующим образом: все кодирующие ретровирусные последовательности делетированы; полноразмерная последовательность U3 из 5'-LTR была замещена промотором главного немедленно раннего гена вируса HCMV при сохранении производной от MLV последовательности 3'-LTR; и кассета, состоящая из промотора SV40 и гена neo, была использована в качестве гетерологичного внутреннего промотора. Затем векторы DON1.2, DON2 и DONSA1 были сконструированы путем встраивания 1-го экзона, интрона А и/или 2-го экзона гена iel (UL123) вируса HCMV в состав рСМ (см. фиг.3a, фиг.3b фиг. 4а-4с). Затем векторы DON1.2-CAT, DON2-CAT и DONSA1-CAT были сконструированы путем встраивания репортерного гена CAT. Клеточные линии, трансфицированные или трансдуцированные этими векторами, анализировали с помощью САТ-теста. По результатам САТ-теста было подтверждено, что все три вектора характеризуются существенно более высокой эффективностью сплайсинга и экспрессии генов по сравнению с контрольным вектором L-CAT-SN (см. таблицу 2).
3. Встраивание клеточных последовательностей ДНК, которые могут способствовать сплайсингу и генной экспрессии:
конструкции MIN-AI, MIN-EI и MIN-GI
В другом предпочтительном варианте ретровирусные векторы были далее выстроены таким образом, чтобы балансировка уровня сплайсинга и интенсивность
генной экспрессии могли обусловливать более высокую эффективность как вирусных титров, так и экспрессии гена. В этих случаях экзонные и/или интронные последовательности генов человека были встроены
выше сайта клонирования, предназначенного для чужеродного гена, в составе ретровирусных векторов (см. фиг.5).
Для конструирования этих векторов вектор MIN был сформирован таким образом, что все ретровирусные кодирующие последовательности были делетированы; производные от MLV последовательности 5'- и 3'-LTR были сохранены; и кассета IRES-neo EMCV была использована в качестве гетерологичного внутреннего промотора. Конкретно вектор MIN был сформирован путем процедуры, включающей следующие этапы: амплификацию фрагмента ДНК, включающего сегмент 5'- и 3'-LTR MLV, встраивание этого фрагмента в плазмиду pUC18 и встраивание кассеты IRES-neo в состав рекомбинантного вектора (см. фиг.6). Фрагмент ДНК, включающий часть интрона и часть 2-го экзона гена β-актина, гена EF1α или гена GAPDH человека, был встроен выше положения сайта клонирования в составе вектора MIN, в результате чего были сконструированы векторы соответственно MIN-AI, MIN-EI или MIN-GI (см. фиг.7а-7с).
Для сравнения вектора, использующего клеточный ген в качестве акцепторного сайта сплайсинга (например, вектора MIN-AI и т.п.), с вектором, использующим вирусный ген (например, вектором DON1.2 и т.п.), был сконструирован еще один вектор на основе вируса MLV - вектор MIN-2. Конкретно последовательность ДНК, включающая часть интрона А и часть 2-го экзона гена iel (UL123) вируса HCMV, встраивается в состав вектора MIN (см. фиг.7d). Эта вставка идентична той, которая имеется в векторе DON2.
Затем векторы MIN-CAT, MIN-AICAT, MIN-EICAT, MIN-GICAT и MIN-2CAT были сконструированы путем встраивания репортерного гена CAT в состав соответствующего исходного вектора. Клеточные линии, трансфицированные или трансдуцированные этими векторами, были подвергнуты САТ-тесту. Результаты САТ-теста показали, что вирусные титры всех векторов по данному изобретению сходны с таковыми, характерными для контрольных векторов LXSN и MFG, в то время как уровни генной экспрессии в клеточных линиях, трансфицированных или трансдуцированных векторами MIN-2 и MIN-EI, существенно выше по сравнению с клеточными линиями, несущими контрольные векторы (см. табл. 3).
ПРИМЕРЫ
Практические и предпочтительные варианты данного изобретения являются иллюстративными, что видно из нижеследующих примеров.
Однако должно быть понятно, что специалисты в данной области техники, исходя из данной заявки, могут вносить модификации и улучшения, не отклоняющиеся от духа и масштаба данного изобретения.
ПРИМЕР 1
Делетирование гена ро1 из вектора MON
(1-1) Конструкция ΔSA
Для получения нижней последовательности 5'-LTR, лишенной кодирующей последовательности гена ро1, была
осуществлена ПЦР (полимеразная цепная реакция), при том что вектор MON (заявка на корейский патент 97-48095) использовался в качестве матрицы, а два одноцепочечных синтетических олигонуклеотида,
описанные в SEQ ID NO 1 (затравка HHIR) и в SEQ ID NO 2 (затравка R523Bam) в качестве затравок. Амплифицированные фрагменты ДНК соответствовали нуклеотидной последовательности от 5'-LTR MLV до
[+523]-го нуклеотида (см. фиг. 1). В этом случае обозначение [+1] соответствует начальному сайту генома вируса MLV, а последовательность на участке [+212]- [+523] является некодирующей
последовательностью, которая необходима в процессе упаковки геномной ДНК вируса.
ПЦР-продукт субклонировали в состав плазмиды pCRII (Invitrogen, CA, США) и затем HindIII/BamHI-фрагмент был выделен из полученного в результате вектора. Частичный HindIII/BamHI-фрагмент вектора MON был заменен на HindIII/BamHI-фрагмент ПЦР-продукта с получением вектора ΔSA. Этот вектор ΔSА используют в качестве элементарных базовых ретровирусных векторов, которые включают последовательности 5'- и 3'-LTR, некодирующую последовательность, включающую акцепторный сайт, кассету IRES-neo и полипуриновый тракт, при том что все кодирующие ретровирусные последовательности оказываются делетированными (см. фиг.1).
(1-2) Конструкция ΔSА-САТ
Для изучения влияния
описанных выше генетических манипуляций на экспрессию чужеродного гена и на упаковку ретровирусной геномной РНК в качестве репортерного гена был использован прокариотический ген CAT.
Ген CAT был получен с помощью метода ПЦР, в котором исходный вектор рСАТ3 (Promega, WI, США) был использован в качестве матрицы, а два синтетических олигонуклеотида, описанные в SEQ ID NO 3 (затравка CATATGN) и в SEQ ID NO 4 (затравка CATSTOP), были использованы в качестве затравок. ПЦР-продукт был встроен в плазмиду pCRII (Invitrogen, CA, США), а BamHI-фрагмент полученного в результате вектора, включающий ген CAT, был внесен в плазмиду pC3.1 (Invitrogen, CA, США) с получением рС3.1-САТ. Вектор ΔSA-CAT был сконструирован путем встраивания гена CAT (в виде обработанного ферментом Кленова XbaI/HindIII-фрагмента из состава рС3.1-САТ) по HpaI-сайту вектора ΔSA (см. фиг.1).
(1-3) Тест CAT
Вектор ΔSА-САТ и контрольный вектор (MON-CAT) вместе с
Env- и Gag-Pol-содержащими "упаковочными" векторами были трансфицированы в клетки 293Т (DuBridge et al. , 1987, Mol. Cell. Biol, 7, 379-387). После 48-часового культивирования трансфицированных клеток
цитоплазматические белки были экстрагированы с целью тестирования активности CAT, что является индикатором уровня экспрессии чужеродного гена. В этом случае бесклеточные вирусные супернатанты,
полученные путем отфильтровывания культуральной среды на фильтре 0,45 мкм, были использованы для трансдукции клеток NIH3T3 (АТСС CRL 1658). Уровень активности CAT был определен с применением белкового
экстракта, полученного через 48 часов после трансдукции.
Тест CAT был осуществлен следующим образом: сначала клетки были собраны и промыты 1 мл фосфатного буфера, а затем ресуспендированы в 0,25 М Трис-буфера (рН=7,5). Клетки лизировали путем шестикратного повторения процедуры замораживания (на сухом льду) и растаивания (на водяной бане при 37oС). После того, как полученный в результате клеточный экстракт был нагрет до 60oС на 7 минут с целью инактивации деацетилазы, его центрифугировали при 12000 об/мин в течение 10 минут с последующим отбором надосадочной фракции. Содержание белка в данном экстракте оценивали количественно по методу Бредфорда. Нормализованный экстракт был смешан с 1 мкл14С-хлорамфеникола (60 мКи/ммоль; 0,1 мКи/мл), 2 мкл ацетилкоэнзима А (40 мМ) и подходящий объем 0,25 М Трис (рН 7,5). Эту реакционную смесь инкубировали при 37oС в течение необходимого для реакции времени. После проведения этой реакции хлорамфеникол экстрагировали этилацетатом и концентрировали в условиях пониженного давления. Остаток ресуспендировали в 15 мкл этилацетата и загружали на пластину для тонкослойной хроматографии (ТСХ). После того, как ТСХ была осуществлена с использованием соответствующего растворителя (95% хлороформа, 5% метанола), ТСХ-пластину высушивали и затем подвергали рентгенографированию или анализу с помощью фосфорного анализатора, на основании чего может быть измерен уровень ацетилирования хлорамфеникола. Активность CAT в образце может быть подсчитана, исходя из радиоактивности ацетилированного хлорамфеникола по отношению к показателю общего хлорамфеникола.
В таблице 1 показано, что активность CAT в клетках линии 293Т, трансфицированных конструкцией Δ SА-САТ, лишенной гена ро1, была ниже, чем в контрольной линии, трансфицированной MON-CAT. Это подтверждает, что роl-кодирующая последовательность вовлечена в регуляцию экспрессии гена. Кроме того, активность CAT в трансдуцированной линии клеток проявила сходные параметры, что и в трансфицированной линии: это указывает на взаимосвязь эффективности упаковки вирусных частиц с эффективностью генной экспрессии.
ПРИМЕР 2
Встраивание промотора и экзона и/или интрона гена iel (UL123) вируса HCMV
С целью поддержания баланса уровня сплайсинга и интенсивности
трансляции, равно как и усиления общего выхода вирусной РНК, были сконструированы производные от MON ретровирусные векторы, включающие следующие три таких вектора:
вектор DON 1.2, в составе
которого полноразмерная последовательность U3 из 5'-LTR вируса MLV заменена промотором главного немедленно раннего гена MLV; и акцепторный сайт сплайсинга MLV заменен фрагментом ДНК, включающим 1-й
экзон, интрон А и часть 2-го экзона (которая заканчивается непосредственно перед старт-кодоном) гена iel (UL123) вируса HCMV;
вектор DON2, в составе которого полноразмерная последовательность
U3 из 5'-LTR вируса MLV заменена промотором главного немедленно раннего гена MLV; и акцепторный сайт сплайсинга MLV заменен фрагментом ДНК, включающим часть интрона А и часть 2-го экзона гена iel
(UL123) вируса HCMV;
вектор DONSA1, в составе которого полноразмерная последовательность U3 из 5'-LTR вируса MLV заменена промотором главного немедленно раннего гена MLV; и акцепторный сайт
сплайсинга MLV заменен фрагментом ДНК, включающим акцепторный сайт сплайсинга гена
иммуноглобулина мыши и 1-й экзон гена iel (UL123) вируса HCMV.
Подробное описание способа конструирования этих векторов приведено ниже (см. фиг.3а, фиг.3b и фиг.4а-4с).
(2-1) Конструирование SN-3LTR
В трех этих вариантах вектора кассета, включающая промотор SV40 и
ген nео, встраивается ниже сайта клонирования, предназначенного для чужеродного гена (см. фиг. 2). Для конструирования этих вариантных векторов вектор, включающий кассету "промотор SV40/ген nео", был
сформирован следующим образом (см. фиг.3а и 3b).
Для получения вектора, который экспрессирует ген nео, находящийся под контролем промотора SV40, кассету "промотор SV40/ген nео" формировали с помощью метода ПЦР. В этой ПЦР плазмиду рС3.1 (Invitrogen, CA, США) использовали в качестве матрицы, а два синтетических олигонуклеотида, описанные в SEQ ID NO 5 (затравка SV40-5) и в SEQ ID NO 6 (затравка Neo-3) были использованы в качестве затравок для ПЦР.
После того, как кассету "промотор SV40/ген nео" клонировали в состав pCRII (Invitrogen, СА, США), BamHI/XhoI-фрагмент полученного в результате вектора встраивали по BamHI/XhoI-сайту конструкции MON. Полученный в результате вектор MSN расщепляли рестриктазами BamHI и EcoRI, a BamHI/-EcoRI-фрагмент, включающий кассету "промотор SV4О/ген nео" и 3'-LTR, лигировали на BamHI/EcoRI-фрагмент плазмиды pUC18 с получением в результате вектора SN-3LTR (см. фиг.3а).
(2-2) Конструирование
рСМ
Метод ПЦР был применен с целью конструирования ретровирусного вектора, в составе которого последовательность 5'-LTR была заменена мощным гетерологичным промотором. В качестве матрицы для
ПЦР использовали ретровирусный вектор МСС-САТ (заявка на корейский патент 97-48095), включающий химерную LTR, в которой полноразмерная последовательность U3 (нуклеотиды [-419]-[-1]) из 5'-LTR MLV была
заменена полноразмерным промотором главного немедленно раннего гена вируса HCMV. Затравками для ПЦР были два синтетических олигонуклеотида, описанные в SEQ ID NO 1 (затравка HHIR) и в SEQ ID NO 2
(затравка R523Bam). ПЦР-продукты, включающие последовательность ДНК из химерной 5'-LTR до нуклеотида [+523], субклонировали в состав плазмиды pCRII (Invitrogen, CA, США). HindIII/BamHI-фрагмент
субклонированной последовательности был встроен по HindIII/BamHI-сайту в вектор SN-3LTR (сконструированный в примере 2-1) с получением вектора рСМ (см. фиг.3b). В векторе рСМ все кодирующие
ретровирусные последовательности делетированы, как и в векторе ΔSА, описанном в примере 1-1, в то время как 5'-LTR и кассета IRES-neo, имеющиеся в векторе ΔSA, заменены химерной LTR и
кассетой "промотор SV40/ген пео" соответственно. Вектор рСМ был использован в качестве исходного материала для конструирования векторов DON1.2, DON2 и DONSA1, описанных в примерах от 2-3 до 2-5.
(2-3) Конструирование DON1.2 и DON1.2-CAT
Для получения фрагмента ДНК, включающего 1-й экзон, интрон А и 2-й экзон (вплоть до старт-кодона) гена iel (UL123) вируса HCMV, была
осуществлена ПЦР. В качестве матрицы для ПЦР использовали вектор pEQ276 (Biegalke et al., 1991, Virology, 183, 381-385), включающий последовательность ДНК от промотора до 5-го экзона главного
немедленно раннего гена вируса HCMV. Две ПЦР-затравки были описаны в SEQ ID NO 7 (затравка RI5, гибридизующая с 1-м экзоном) и в SEQ ID NO 8 (затравка CMVexon2.3, гибридизующая со 2-м экзоном)
соответственно. Фрагмент ДНК длиной 1000 нуклеотидов был амплифицирован в методе ПЦР: он включал 1-й экзон, интрон А и частично 2-й экзон (который заканчивается непосредственно перед старт-кодоном)
гена iel (UL123) вируса HCMV.
ПЦР-продукт был встроен в вектор pZero-blunt (Invitrogen, CA, США) с получением вектора pZerol.2. Затем EcoRI-фрагмент последовательности pZerol.2 обрабатывали ферментом Кленова с получением "тупых концов". Вектор рСМ, сконструированный в примере 2-2, расщепляли рестриктазой BamHI и обрабатывали ферментом Кленова. Два фрагмента ДНК с "тупыми концами" лигировали друг на друга с получением вектора DON1.2. Кроме того, вектор DON1.2-CAT был сконструирован путем встраивания гена CAT в обработанный ферментом Кленова HindIII-фрагмент DON1.2 (см. фиг.4а).
(2-4) Конструирование DON2 и DON2-CAT
Был получен HpaI-EcoRI-фрагмент вектора pZerol.2, сконструированного в примере 2-3, который включает 112-нуклеотидный 3'-участок
интрона А и 5'-участок 2-го экзона (нуклеотиды [+837]-[+964]: непосредственно перед старт-кодоном). Затем этот фрагмент ДНК был обработан ферментом Кленова с получением "тупых концов". С другой
стороны, вектор рСМ, сконструированный в примере 2-2, был обработан рестриктазой BamHI и ферментом Кленова. Указанные выше два фрагмента ДНК, имеющие "тупые концы", лигировали друг на друга с
получением вектора DON2. Кроме того, вектор DON2-CAT был сконструирован путем встраивания гена CAT в обработанный ферментом Кленова HindIII-фрагмент DON2 (см. фиг.4b).
(2-5)
Конструирование DONSA1 и DONSA1-CAT
С целью получения акцепторного сайта сплайсинга гена иммуноглобулина мыши были синтезированы одноцепочечные олигонуклеотиды, которые описаны в SEQ ID NO 9
(олигонуклеотид SА Top) и в SEQ ID NO 10 (олигонуклеотид SА bottom) соответственно. В реакции отжига двух олигонуклеотидов были получены фрагменты акцепторного сайта сплайсинга, имеющие "липкие
концы". Вектор pGEM4-SA был сформирован путем встраивания данного фрагмента по BamHI-сайту вектора pGEM4 (Promega, WI, США) (см. фиг.4с).
Для амплификации последовательности 1-го экзона гена iel (UL123) вируса HCMV была проведена ПЦР, при том что вектор pEQ276, сконструированный в примере 2-3, был использован в качестве матрицы, а два синтетических олигонуклеотида, описанных в SEQ ID NO 7 (затравка RI5) и в SEQ ID NO 11 (затравка ехоп-13), были использованы соответственно в качестве затравок.
Амплифицированный фрагмент, включающий 1-й экзон, был субклонирован по EcoRI-сайту в состав вектора pZero-blunt (Invitrogen, CA, США) с получением вектора pZero-exonl. Затем EcoRI-фрагмент последовательности pZero-exonl был субклонирован по EcoRI-сайту плазмиды pGEM4-SA с получением вектора pGEM-SA-exonl.
XbaI/BamHI-фрагмент вектора GEM-SA-exon1 был обработан ферментом Кленова с получением "тупых концов" и затем лиги-рован на обработанный ферментом Кленова BamHI-фрагмент плазмиды рСМ (сконструированной в примере 2-2) с получением вектора DONSA1.
Кроме того, вектор DONSA1-CAT был сконструирован путем встраивания гена CAT по HpaI-сайту вектора DONSA1 (см. фиг.4с).
(2-6) Тест CAT
Векторы DON1.2-CAT, DON2-CAT, DONSA1-CAT и L-CAT-SN были использованы для трансфицирования
"упаковочной" линии клеток FlyA13 (Cosset et al., 1995, J. Virol, 69, 7430-7436). После того, как трансфицированные линии культивировали в течение 48 часов, бесклеточные вирусные надосадочные фракции
были приготовлены для последующей трансдукции клеток NIH3T3. Трансдуцированные линии клеток культивировали в течение 48 часов. Активность CAT в трансфицированных или трансдуцированных клеточных линиях
определяли в соответствии с методом, описанным в примере 1-3, тем самым оценивая относительные уровни интенсивности генной экспрессии.
Как показано в таблице 2, существенно более высокие уровни активности CAT были выявлены в линии FlyA13, трансфицированной векторами DON1.2-CAT или DON2-CAT по сравнению с контрольной линией (трансфицированной конструкцией L-CAT-SN). Кроме того, клетки NIH3T3, стабильно трансдуцированные векторами DON1.2-CAT или DON2-CAT, показали в 5-10 раз более высокую активность CAT по сравнению с контрольной линией. В том случае, когда клетки трансфицировали или трансдуцировали вектором DONSA1-CAT, ненамного более высокая активность CAT была отмечена по сравнению с контрольной линией. Полученные результаты подтвердили, что эффективность сплайсинга и эффективность генной экспрессии могут быть повышены тогда, когда гетерологичная некодирующая последовательность, вовлеченная в процессы сплайсинга, встраивается выше чужеродного гена в составе экспрессирующего вектора.
Штаммы кишечной палочки Е. coli, трансформированные векторами DON2 и DONSA1, были обозначены как TOP10-DON2 и TOP10-DONSA1 соответственно. Они были депонированы в Корейский Центр культур микроорганизмов 5 июня 1998 года (депозитарные КССМ-10128 и КССМ-10127 соответственно).
ПРИМЕР 3
Встраивание интрона и/или экзона гена
человека
В этом примере производные от MIN ретровирусные векторы MIN-AI, MIN-EI, MIN-GI и MIN-2 конструируются таким образом, чтобы специфически для эукариотических клеток обеспечить баланс
уровней транскрипции, сплайсинга и трансляции чужеродного гена. Ни одна из вирусных последовательностей не содержится в составе вектора MIN, равно как и вектора ΔSА, однако вектор MIN включает
кассету IRES-neo вместо кассеты "промотор SV40/ген nео" (см. фиг.5). Четыре названных выше вектора имеют следующие характеристики:
фрагмент ДНК, включающий интрон, акцепторный сайт сплайсинга
и часть 2-го экзона гена β-актина человека, был встроен выше чужеродного гена в составе вектора MIN-AI;
фрагмент ДНК, включающий интрон, акцепторный сайт сплайсинга и часть 2-го экзона
гена EFlα человека, был встроен выше чужеродного гена в составе вектора MIN-EI;
фрагмент ДНК, включающий интрон, акцепторный сайт сплайсинга и часть 2-го экзона гена GAPDH человека,
был встроен выше чужеродного гена в составе вектора MIN-GI.
Помимо этих конструкций был сконструирован вектор, в состав которого была встроена гетерологичная вирусная последовательность. Конкретно вектор MIN-2 был сформирован так, что фрагмент ДНК, включающий интрон, акцепторный сайт сплайсинга и часть 2-го экзона гена iel (UL123) вируса HCMV, был встроен выше сайта клонирования вектора MIN.
Вектор MIN и MIN-производные векторы MIN-AI, MIN-EI, MIN-GI и MIN-2 были сконструированы следующим образом.
(3-1) Конструирование MIN
Для получения сегмента 3'-LTR MLV была осуществлена ПЦР, в которой вектор pMLV (Shinnick et al., 1981, Nature, 293, 543-548) использовался в качестве матрицы, а два синтетических олигонуклеотида,
описанные в SEQ ID NO 12 (затравка 3LTR5) и в SEQ ID NO 13 (затравка 3LTR3) были использованы в качестве затравок. Амплифицированный ПЦР-продукт включал 3'-нетранслируемый сегмент, полипуриновый
тракт и последовательность 3'-LTR из генома вируса MLV. ПЦР-продукт субклонировали в состав вектора pCRII (Invitrogen, CA, США), расщепляли рестриктазами BamHI и EcoRI и полученный в результате
фрагмент встраивали по BamHI/EcoRI-сайту плазмиды pUC18 с получением вектора p3LTR (см. фиг.6).
С другой стороны, для некодирующей последовательности, включающей ретровирусный 5'-LTR и донорный сайт сплайсинга, была проведена ПЦР, в которой вектор pMLV использовали в качестве матрицы, а два синтетических олигонуклеотида, описанные в SEQ ID NO 1 (затравка HHIR) и в SEQ ID NO 14 (затравка 5LTR3), были использованы в качестве затравок. Амплифицированный ПЦР-продукт включал нуклеотидную последовательность от 5'LTR до 623-го нуклеотида (непосредственно перед кодирующей последовательностью гена gag) генома вируса MLV. После того, как ПЦР-продукт был субклонирован в состав вектора pCRII, HindIII/BamHI-фрагмент данного вектора был встроен по HindIII/BamHI-сайту вектора p3LTR с получением вектора p53LTR (см. фиг.6).
Наконец, кассета IRES-neo была выделена из вектора pCBIN (заявка на корейский патент 97-48095) путем расщепления рестриктазами BamHI/XhoI и затем встроена по BamHI/XhoI-сайту вектора p53LTR с получением вектора MIN (см. фиг.6).
Вектор MIN-CAT был также сконструирован путем встраивания гена CAT по BamHI-сайту вектора MIN таким образом, чтобы можно было анализировать эффективность экспрессии вектора MIN.
(3-2) Конструирование MIN-AI
Для получения нуклеотидных последовательностей человека,
которые бы можно было встроить в состав вектора MIN, геномную ДНК экстрагировали из клеток человека. Во-первых, моноядерные клетки периферической крови были отсепарированы из проб крови человека с
применением центрифугирования в градиенте Фиколл. После одно-двукратной промывки клетки лизировали с использованием ТЕЗ (10 мМ Трис-НСl - рН 7,0; 10 мМ EDTA, 0,7% SDS). К клеточному лизату добавляли
протеиназу-К (400 мкг/мл) и полученный лизат инкубировали при 50-55oС в течение 1-2 часов с последующей экстракцией в смеси фенола и хлороформа и преципитацией этанолом.
Выделенную геномную ДНК использовали в качестве матрицы в методе ПЦР так, что амплифицированный ПЦР-фрагмент включал промотор, 1-й экзон, интрон и часть 2-го экзона гена β-актина человека. Затравками для ПЦР были два синтетических олигонуклеотида, описанные в SEQ ID NO 15 (затравка beta-actin 5) и в SEQ ID NO 16 (затравка beta-actin 3). ПЦР-продукт был субклонирован в состав вектора pCRII, а затем MluI/NheI-фрагмент этого вектора был встроен по MluI/NheI-сайту в вектор рС3.1 (Invitrogen, СА, США) с получением вектора pβactin.
Обработанный ферментом Кленова BglI/BamHI-фрагмент pβactin (соответствующий нуклеотидам [+717] ≈[+849] гена β-актина человека) был встроен по обработанному полимеразой Т4 ApaI/BamHI-сайту вектора MIN (см. фиг.7а). Полученный в результате вектор был обозначен MIN-AI.
Кроме того, вектор MIN-AICAT был сконструирован путем встраивания гена CAT по BamHI-сайту вектора MIN-AI так, чтобы можно было анализировать эффективность экспрессии вектора MIN-AI.
(3-3) Конструирование MIN-EI
Для получения некодирующей последовательности гена EF1α человека была
осуществлена геномная ПЦР. Геномную ДНК, выделенную в примере 3-2, использовали в качестве матрицы в методе ПЦР, а два синтетических олигонуклеотида, описанные в SEQ ID NO 17 (затравка EF1α5) и
в SEQ ID NO 18 (затравка EF1α3), использовали в качестве затравок. Полученный ПЦР-продукт включал промотор, 1-й экзон, интрон и часть 2-го экзона гена EF1α человека. ПЦР-продукт был
субклонирован в состав вектора pCRII, а затем MluI/NheI-фрагмент этого вектора был встроен по MluI/NheI-сайту в вектор рС3.1 (Invitrogen, СА, США) с получением вектора pEF1α.
Обработанный ферментом Кленова XhoI/BamHI-фаргмент pEF1α (соответствующий нуклеотидам [+772]≈[+1008] гена EF1α человека) был встроен по обработанному полимеразой Т4
ApaI/BamHI-сайту вектора MIN (см. фиг.7b). Полученный в результате вектор был обозначен MIN-EI.
Кроме того, вектор MIN-EICAT был сконструирован путем встраивания кассеты CAT по BamHI-сайту вектора MIN-EI так, чтобы можно было анализировать эффективность экспрессии вектора MIN-EI. Вектор MIN-EICAT был внесен в клетки Е. coli штамма Тор10, а трансформант Е. coli был обозначен MIN-EICAT (Тор10) и депонирован в Корейский Центр культур микроорганизмов 2 июня 1999 года (депозитарным КССМ-10163).
(3-4) Конструирование MIN-GI
Для получения некодирующей
последовательности гена GAPDH человека была осуществлена геномная ПЦР. Геномную ДНК, выделенную в примере 3-2, использовали в качестве матрицы в методе ПЦР, а два синтетических олигонуклеотида,
описанные в SEQ ID NO 19 (затравка Gint5) и в SEQ ID NO 20 (затравка Gint3), использовали в качестве затравок. Полученный ПЦР-продукт включал интрон и часть 2-го экзона гена GAPDH человека, что
соответствовало нуклеотидам [+185] ≈[+317] гена GAPDH человека. ПЦР-продукт был субклонирован в состав вектора pCRII, а затем MluI/BamHI-фрагмент этого вектора был встроен по MluI/BamHI-сайту в
вектор MIN с получением вектора MIN-GI (см. фиг.7с).
Кроме того, вектор MIN-GICAT был сконструирован путем встраивания гена CAT по BamHI-сайту вектора MIN-GI так, чтобы можно было анализировать эффективность экспрессии вектора MIN-GI.
(3-5) Конструирование MIN-2
Был сформирован MluI/BamHI-фрагмент вектора DON2 (соответствует нуклеотидам [+837]≈
[+964] гена iel HCMV), который включал интрон, акцепторный сайт сплайсинга и часть 2-го экзона гена iel (UL123) вируса HCMV. Этот MluI/BamHI-фрагмент был встроен по Mlul/BamHI-сайту вектора MIN (см.
фиг. 7d). Полученный в результате вектор был обозначен MIN-2.
Кроме того, вектор MIN-2CAT был сконструирован путем встраивания гена CAT по BamHI-сайту вектора MIN-2 так, чтобы было можно анализировать эффективность экспрессии вектора MIN-2. Вектор MIN-2CAT был внесен в клетки Е. coli штамма Тор10. Трансформант Е. coli был обозначен MIN-2CAT (Тор10) и депонирован в Корейский Центр культур микроорганизмов 2 июня 1999 года (депозитарный КССМ-10164).
(3-6) Тест CAT
Для анализа эффективности экспрессии чужеродного гена и способности к нормальной
упаковке названных выше четырех векторов векторы были подвергнуты тесту CAT, при том что в качестве контрольных векторов были использованы хорошо известные рестровирусные векторы MFG и LXSN, в которые
был встроен ген-репортер CAT (Miller et al., 1989, Biotechniques, 7, 980-990; Ohashi et al. , 1992, Proc. Natl. Acad. Sci. USA, 89, 11332-11336). "Упаковочную" клеточную линию Phoenix (Kinsella
& Nolan, 19??, Human Gene Therapy, 7, 1405-1413) использовали для трансфицирования указанными векторами и затем культивировали на протяжении 48 часов. Затем бесклеточные вирусные надосадочные
фракции получали путем отфильтровывания культуральной среды через 0,45-мкм фильтр и использовали для трансдукции клеток NIH3T3 (АТСС CRL 1658). Уровень активности фермента CAT был определен с
использованием белкового экстракта через 2 дня после осуществления трансдукции. Активность CAT была определена в клеточной линии, трансфицированной или трансдуцированной каждым из векторов (см.
колонки "временно трансфицированные" и "временно трансдуцированные" в таблице 3), равно как в популяции резистентных к антибиотикам клеток (см. колонку "стабильно трансдуцированные" в таблице 3).
Кроме того, активность CAT была определена в популяции стабильных клеток, субкультивировавшихся в течение 4 недель (см. колонку "стабильно трансдуцированные (4 недели)" в таблице 3). Уровни генной
экспрессии и вирусные титры в трансфицированных линиях были определены, исходя из уровней активности CAT, которые были измерены в линиях, трансфицированных или трансдуци-рованных каждым из
векторов.
Как показано в таблице 3, все ретровирусные векторы проявляют более высокие уровни активности CAT в сравнении с вектором MIN, за исключением вектора LXSN. Однако уровни активности CAT варьировались в зависимости от встроенной гетерологичной последовательности. В частности, векторы MIN-EI и MIN-2 обеспечивали существенно более высокие уровни генной экспрессии. Было вполне ясно, что клетки, стабильно трансдуцированные векторами MIN-EI или MIN-2, проявляли значительно более высокие уровни генной экспрессии даже после дополнительного культивирования на протяжении еще 4 недель.
ПРОМЫШЛЕННОЕ ПРИМЕНЕНИЕ
Как было описано и подтверждено выше, данное изобретение представляет ретровирусные векторы, которые характеризуются многими
преимуществами с точки зрения генотерапии и т.п. Векторы по данному изобретению характеризуются следующими параметрами:
1. Поскольку все ретровирусные кодирующие последовательности
(последовательности gag, env и pol) делетированы, практически может быть исключена возможность того, что в результате гомологичной рекомбинации будут образовываться компетентные по репликации
ретровирусы.
2. Поскольку гетерологичные интрон, акцепторный сайт сплайсинга и/или некодирующая последовательность встроены выше сайта клонирования, предназначенного для чужеродного гена, то чужеродный ген в составе этих векторов может быть экспрессирован и стабильно, и эффективно.
3. Поскольку последовательность U3 из состава 5'-LTR, включающая гетерологичный промотор, который индуцирует интенсивную транскрипцию особенно в клетках человека, то производные от клеток человека "упаковочные" линии, трансфицированные указанными векторами, могут вырабатывать более высокие уровни РНК и таким образом обусловливать более высокие вирусные титры.
4. Последовательность IRES или гетерологичный промотор используются одновременно для обеспечения экспрессии двух или большего числа чужеродных генов в указанных векторах. В этом случае минимальный промотор может быть использован в качестве встроенного гетерологичного промотора с целью исключения помех внутреннему гетерологичному промотору и с целью клонирования чужеродного гена более крупного размера.
Для специалиста в данной области техники должно быть понятно, что концепции и конкретные варианты, описанные выше, могут быть легко применены в качестве исходного материала для модифицирования или разработки новых вариантов, призванных решать те же задачи данного изобретения. Также для специалиста в данной области техники должно быть понятно, что эквивалентные варианты не будут отклоняться от духа и масштаба данного изобретения в объеме представленной формулы изобретения.
Реферат
Данное изобретение относится к улучшенным ретровирусным векторам. Векторы конструируются на основе исходных векторов MON и MIN, основанных на MLV. Улучшенные векторы характеризуются делецией кодирующих областей MLV gag, pol и env, что позволяет избегать гомологичной рекомбинации. Данные вектора обеспечивают безопасную и эффективную экспрессию чужеродных генов. Векторы могут быть использованы для генотерапии. 5 с. и 8 з.п. ф-лы, 7 ил., 3 табл.
Комментарии