Способ получения белка с использованием трансляционной системы митохондрий - RU2201961C2
Код документа: RU2201961C2
Чертежи

Описание
Область изобретение
Данное изобретение относится к экспрессии белка рекомбинантными молекулами
нуклеиновой кислоты и, в частности, относится к продукции белков, включая вирусные белки, в тканях животных, культивируемых in vitro, путем инфицирования ткани-хозяина вирусом или трансфекции
ткани-хозяина рекомбинантной нуклеиновой кислотой в векторе экспрессии на основе вируса и с использованием трансляции в богатой митохондриями ткани.
Описание прототипа
Трансляция белков с трансфицированных нуклеиновых кислот в основном совершается с использованием универсальных трансляционных систем, присутствующих в прокариотных или эукариотных клетках (Sambrook
et
al., Molecular Cloning, A Laboratory Manual, 2nd Ed., Vol. 1-3, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989). Митохондрии, обнаруженные в эукариотных клетках, имеют
системы
транскрипции и трансляции для экспрессии эндогенной митохондриальной ДНК (мтДНК), в которых используется не универсальный генетический код. Митохондриальная система трансляции, однако, не
использовалась для трансляции чужеродных нуклеиновых кислот.
Митохондрии являются клеточными органеллами с многослойной мембраной, которые растут и делятся в координированном процессе, который требует участия генетической системы в ядре клетки и отдельной генетической системы, содержащейся в митохондриях (Alberts et al./ Molecular Biology of the Cell, 2nd Ed. , pp. 387-401, Garland Publishing Inc., New York, NY). Большинство митохондриальных белков кодируется ядерной ДНК, которая транскрибируется, транслируется в цитозоле и импортируется в митохондрии. По контрасту, некоторые митохондриальные белки транскрибируются с мтДНК и транслируются в самой органелле с использованием митохондриальной системы, которая включает две рибосомные РНК и 22 тРНК. Сравнение митохондриальных последовательностей генов с аминокислотными кодируемых белков показывает, что генетический код в митохондриях изменен по сравнению с универсальным кодом, используемым в ядре эукариотных клеток и у большинства прокариотов. Например, кодон UGA является стоп-кодоном для синтеза белка в универсальном коде, тогда как в митохондриях UGA кодирует триптофан, а кодоны AGA и AGG кодируют аргинин в универсальной системе, но являются стоп-кодонами в митохондриях млекопитающих.
Рекомбинантная ДНК может использоваться для продукции белков, которые транспортируются в митохондрии. В одной из систем экспрессии клетки почек обезьян (клетки COS-7) трансфицировали с помощью вектора экспрессии, содержащего кДНК, для введения митохондриального флавоэнзимного (MCAD) гена (Jensen et al., Biochim. et Biophys. Acta 1180: 65-72, 1992). РНК транскрипты и белок продуцировались с использованием систем транскрипции и трансляции трансфицированных клеток. Рекомбинантный белок MCAD обрабатывали и концентрировали в митохондриальной клеточной фракции, что указывало на то, что белок MCAD был транспортирован в митохондрий, где лидерный пептид был удален с белка, продуцируемого в цитозоле.
Репликация некоторых вирусов была связана с клеточными митохондриями или частицами с многослойной мембраной, обнаруженными в инфицированных клетках. В клетках почек обезьян, выращенных in vitro и инфицированных вирусом гепатита А (ВГА), вирусоподобные частицы были обнаружены в связанных с мембраной везикулярных тельцах-включениях, которые содержат антигены ВГА (Asher et al. , J. Virol. Meth. 15: 323-328, 1987). Фосфопротеин, необходимый для синтеза РНК вируса леса Семлики (ВЛС) (Semliki Forest virus [SFV]), был локализован в больших везикулоподобных структурах в инфицированных ВЛС клетках и в клетках COS, трансфицированных кДНК, кодирующей фосфопротеин (Peranen, J. Gen. Virol. 72: 195-199, 1991).
Нуклеозидные аналоги, которые подавляют репликацию вируса гепатита В (ВГВ), нарушают также функции митохондрий после хронического воздействия лекарств, что наводит на мысль о сходных механизмах репликации ДНК как для ВГВ, так и мтДНК. Аналоги 2',3'-дидезокси-3'-тиацитидина 5-фтор-2',3'-дидезокси-3'-тиацитидин и 1-(2'-дезокси-2'-фтор-β-арабинофуранозил)-5-иодурацил (т.е. фиалуридин) подавляют репликацию ВГВ (Doong et al., Proc. Natl. Acad. Sci. USA 88:8495-8499, 1991; Colacino et al., Antimicrobial Agents and Chemother. 38(9): 1997-2002, 1994). Из этих соединений, (+)-энантиомер 2', 3'-дидезокси-3'-тиацитидина, как было показано, значительно подавляет синтез мтДНК in vitro в изолированных митохондриях (Chang et al., J. Biol. Chem. 267(31): 22414-22420, 1992). ВГВ легко обнаруживаются в органах, которые содержат большие количества митохондрий, включая печень, поджелудочную железу и слюнные железы, но в трансфицированных ВГВ линиях клеток, которые содержат мало митохондрий, вирусные частицы ВГВ и его антигены обнаружить трудно. Кроме того, некоторые антигены ВГВ могут быть необходимы для репликации вируса потому, что линии клеток, которые не производят белки ВГВ, не продуцируют также и частицы Дана. Это может происходить из-за того, что митохондрий часто повреждаются во время обычного культивирования тканей или клеток, приводящего к ограниченному росту ВГВ в культивируемых клетках. Некоторые линии клеток (например, модифицированные гепатоциты взрослых, клетки гепатобластомы и фетальные гепатоциты), как было обнаружено, продуцируют антиген Нве в общепринятых системах культивирования тканей (Gripon et al., Virol. 192: 534-540, 1993; Ochiya et al., Proc. Natl. Acad. Sci. USA 86: 1875-1879, 1989). Такие линии клеток содержат достаточно митохондрий, чтобы была возможна продукция Нве при использовании общепринятых методов культивирования тканей.
Недавно были получены трансгенные мыши HBV и использованы для изучения сборки, транспорта, секреции и других функциональных свойств белков ВГВ (Guidotti et al., J. Virol. 69: 6158-6169, 1995; Araki et al., Proc. Natl. Acad. Sci. USA 86: 207-211, 1989). Антиген Нве, продуцируемый у таких трансгенных мышей, может быть результатом наличия плазмид, использованных для создания трансгеников или РНК, продуцируемой из этих плазмид, входящих в митохондрий. Возможность того, что эти плазмиды могут входить в митохондрии, основывается на том факте, что структура мембраны митохондрий похожа на структуру других мембран, что дает возможность прохождения нуклеиновых кислот в определенных условиях. Высокий уровень репликации ВГВ был обнаружен в тканях печени и почек у некоторых трансгенных мышей HBV, у которых имеются концевые избыточные конструкции ВГВ длиной больше чем геном (Guidotti et al. , J. Virol. 69: 6158-6169, 1995).
ВГВ, активно реплицирующийся у людей, в линиях клеток или у трансгенных животных, который продуцирует вирусные частицы, также всегда продуцирует Нве (Chisari F. V. , Hepatology 22: 1316-1325, 1996). Для репликации полностью функционального ВГВ могут быть необходимы как универсальные системы трансляции, так и системы трансляции митохондрий. В гепатоцитах, по-видимому, больше антигенов ВГВ продуцируется при использовании митохондриальной системы трансляции, чем с универсальной системой трансляции, так как наибольшая часть растворимых антигенов ВГВ обнаруживается во фракции митохондрий культивируемой ткани печени (Palk et al.. Abstract, Am. Assoc. For the Study of Liver Diseases, 1995). Однако, так как митохондрии часто повреждаются в обычных системах культивирования тканей, участие митохондриальной системы трансляции в сборке вируса и/или иммунные реакции in vivo было трудно определить. Этим повреждением митохондрий, связанным с обычными методами культивирования тканей, можно также объяснить, почему было трудно размножить ВГВ in vitro с использованием культур клеток.
Были описаны динамические органные системы культивирования, в которых жизнеспособность ткани печени может поддерживаться в течение примерно 24-48 ч регулируемых условиях (Smith P.F et al.. Life Sci. 36: 1367, 1985; S.S. Park, Inje Med. J. 14(3): 363-369, 1993). Использование тимусной органной культуры было описано в связи с методами идентификации потенциальных противовирусных средств (опубликованная заявка РСТ WO 9505453). В данном изобретении используется физиологическая система культивирования (продаваемая Leema Pharmed, Seoul, Korea) для культивирования тканей животных in vitro, где она эффективно инфицируется вирусом, включая человеческие ВГВ (HBV) или ВГС (HCV), для продукции вирусных антигенов с использованием эукариотной митохондриальной системы трансляции. Эта система может также использоваться для продукции других не митохондриальных белков, которые могут транслироваться в митохондриях, путем трансфекции культивируемых клеток вектором на основе человеческого вируса гепатита, содержащим рекомбинантную ДНК. Предпочтительный вектор содержит ДНК из ВГВ и/или комплементарные последовательности ВГС.
Краткое изложение изобретения
По этому
изобретению представлен
способ продукции вирусных антигенов в культивируемой ткани животного, включающий стадии: получения ткани органа от животного для того, чтобы она служила тканью-хозяином при
культивировании in vitro,
причем ткань-хозяин богата митохондриями; инфицирования ткани-хозяина in vitro вирусом; культивирования инфицированной ткани-хозяина in vitro с целью продукции вирусных
белков с использованием системы
трансляции митохондрий в ткани-хозяине и выделения вирусных белков из инфицированной и культивированной ткани-хозяина. При одном из воплощений этого способа
ткань-хозяин выделяется из ткани органа,
выбираемого из печени, почек, поджелудочной железы и слюнной железы. При другом воплощении животное выбирается из группы, состоящей из людей, крыс, мышей,
собак, цыплят и лягушек. В предпочтительном
воплощении вирус является человеческим вирусом, выбираемым из группы, состоящей из вируса гепатита А, вируса гепатита В, вируса гепатита С и вируса
энцефалита. При одном из воплощений вирусные
антигены продуцируются в митохондриях в ткани-хозяине. При предпочтительном осуществлении способ дополнительно включает введение выделенных вирусных
антигенов животному для индукции иммунного ответа.
При другом предпочтительном воплощении по этому способу продуцируются вирусные антигены, пригодные для использования в вакцине.
По другому аспекту этого изобретения представлен способ продукции белков в культивируемой ткани животного, включающий стадии: получения ткани органа от животного для того, чтобы она служила тканью-хозяином в культуре in vitro, причем ткань-хозяин богата митохондриями; трансфицирования ткани-хозяина in vitro вектором ДНК, содержащим вирусную ДНК и рекомбинантную ДНК; культивирования ткани-хозяина in vitro для продукции, белков, кодируемых трансфицированным вектором ДНК, с использованием системы трансляции митохондрий в ткани-хозяине и выделения белков, кодируемых трансфицированным вектором ДНК из культивированной и трасфицированной ткани-хозяина. При одном из воплощений этого способа ткань-хозяин выделяется из ткани органа, выбираемого из группы, состоящей из печени, почек, поджелудочной железы и слюнных желез. При еще одном воплощении животное выбирается из группы, состоящей из людей крыс, мышей, собак, цыплят и лягушек. При предпочтительном осуществлении вирусная ДНК является ДНК вируса гепатита В. Кроме того, способ может включать стадию трансфицирования ткани-хозяина хелперным (вспомогательным) вирусом. При одном из воплощений белки продуцируются в митохондриях в ткани-хозяине. По другому из воплощений этого способа продуцируются белки, пригодные для использования в вакцине. Предпочтительные осуществления включают белки, продуцируемые по этому способу, причем вирусная ДНК является ДНК вируса гепатита В, и причем вектор ДНК содержит рекомбинантную ДНК, встроенную в последовательность ДНК человеческого вируса, кодирующую неструктурный вирусный белок.
Краткое описание чертежей
Фиг. 1 представляет устройство для автоматизированного культивирования образцов тканей in vitro.
Фиг. 2 представляет в виде диаграммы вектор экспрессии на основе ВГВ (HBV).
Детальное описание изобретения
Данное изобретение представляет способы продукции
природных белков, которые не могут легко продуцироваться при использовании обычной технологии с
рекомбинантной ДНК, и белков из вирусов, при которых вирусная нуклеиновая кислота транслируется in
vitro в клетках, содержащих большое количество митохондрий, причем клетки сохраняются в
автоматической динамической системе культивирования.
Данное изобретение дает возможность перекрестно-видового инфицирования вирусами ткани, которая поддерживается in vitro для продукции белка из инфицирующего вируса. Это особенно важно для трансляции человеческих вирусов в клетках животных, но также применимо для перекрестно-видового инфицирования клеток любыми вирусами с использованием человеческих и нечеловеческих вирусов и тканей человека или нечеловеческих тканей в качестве ткани-хозяина. Например, слои печени крысы могут быть инфицированы человеческим ВГВ, и ткань печени может поддерживаться в автоматической динамической системе для культивирования, что дает возможность экспрессии вирусных антигенов in vitro.
Ткань органа выделяли от животного, такого как крыса, с использованием стандартных хирургических процедур. Обычно органом был тот орган, о котором известно, что он богат митохондриями, такой как печень, почки, поджелудочная железа или слюнная железа. Ткань разрезали на слои кусочками размером примерно 2 см2 толщиной примерно 260 мкм и инфицировали вирусом, таким как ВГВ путем инкубации слоев ткани с вирусом в культуральной среде. ВГВ получали из ткани, полученной при биопсии печени инфицированного больного человека. Опытным специалистам будет понятно, что ВГВ может быть заменен на другие вирусы, такие как вирус гепатита А, вирус гепатита С, вирус энцефалита и подобные вирусы животных. В качестве контроля культивировали в питательной среде срезы того же самого типа ткани животного, которые не контактировали с вирусом.
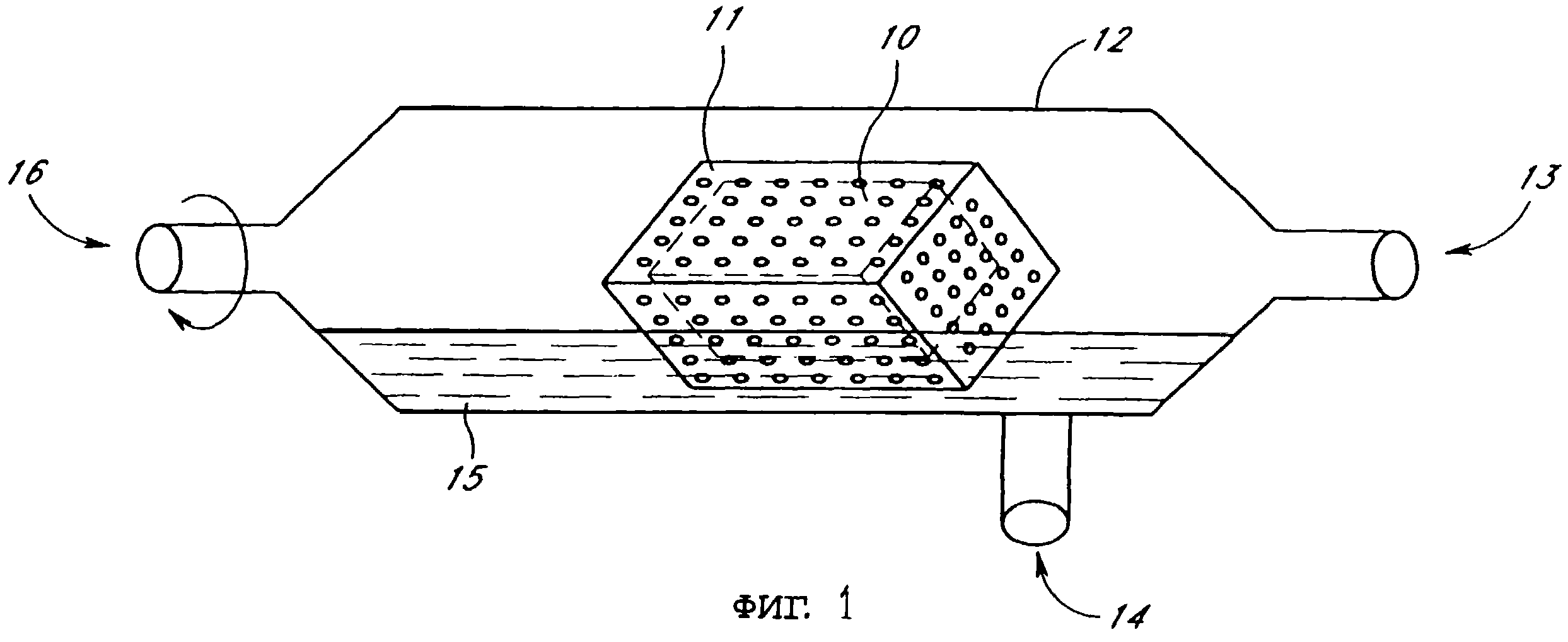
Инфицированные срезы органа культивировали в автоматической системе для культуры органов. Ссылаясь на фиг.1, в этой системе культивирования срезы ткани 10 культивируют в пористом контейнере 11, помещенном внутри трубки для культивирования 12, которая вращается (см. стрелку), чтобы дать возможность ткани периодически погружаться в среду для культуры ткани 15, когда трубка для культивирования 12 вращается. Газообмен в трубке для культивирования 12 происходит через регулярные интервалы, через которые газовую смесь вводят в трубку для культивирования через отверстия 13, 16, расположенные на концах трубки для культивирования 12. Удаление образцов для исследования или введение среды или других реагентов производится путем возможности доступа внутрь трубки для культивирования 12 через отверстие для образца 14, расположенного в стенке трубки для культивирования 12. В системе для культивирования поддерживалась при постоянной температуре, равной 37oС, путем помещения ее в термостат.
Срез ткани культивировали при 37oС в модифицированной среде для культивирования Waymouth's MB 752/1 с рН 7,0 при давлении газовой смеси из 5% CO2 и 95% O2, равном 1, 6-2 атм, хотя опытные специалисты поймут, что равным образом могут использоваться и другие среды и газовые смеси. Инкубация инфицированной вирусом ткани в основном происходит в течение 1-48 ч, предпочтительно в течение примерно 24 ч.
После завершения периода культивирования ткань собирали и использовали для исследования на белки или для получения белков с использованием хорошо известных специалистам стандартных методик. Например, стандартные иммунохимические методы использовали для контроля белков ВГВ в инфицированной ткани путем получения сечений ткани и окрашивания их антителами к НВsАg.
В основном вирусные белки определялись в клетках животных после менее 24 ч культивирования. Инфицированные ткани окрашивались антителами к НВsАg неравномерно, причем богатые митохондриями области в ткани окрашивались более интенсивно по сравнению с другими областями ткани. В контрольной ткани проявлялось только фоновое окрашивание.
Когда рассеченную инфицированную вирусом ткань животного исследовали с применением электронной микроскопии, определялись подобные митохондриям органеллы с многослойной мембраной, содержащие вирусные белки, что указывало на то, что эффективность вирусной инфекции связана с концентрацией митохондрий в ткани животного. Таким образом, было продемонстрировано перекрестно-видовое вирусное инфицирование человеческим вирусомткани животного с использованием ВГВ, так как интенсивное иммуноокрашивание ткани антителами к HBsAg показывает, что ВГВ может инфицировать орган животного, в котором достаточно митохондрий, чтобы обеспечить возможность репликации вируса, и реплицироваться в нем. Некоторые из иммуноокрашенных структур напоминали разрушенные гребешковые сечения митохондрий. Так как известно, что митохондрии имеют систему трансляции, отдельную от цитоплазматической системы трансляции, наличие HBsAg в органеллах, похожих на митохондрии, означает, что белок транслировался системой трансляции митохондрий. Такая трансляция должна продуцировать различные секреторные антигены из ВГВ по сравнению с трансляцией той же самой РНК с использованием универсальной кодоновой системы в клеточной цитоплазме.
Белки, выделенные из инфицированной ткани крысы, представляют тот же набор вирусных белков при применении стандартного метода электрофореза в полиакриламидном геле, который является более сложным, чем состав белков ВГВ, продуцируемых при стандартной технологии с рекомбинантной ДНК. Результаты иммуноокрашивания означают, что белки ВГВ, продуцируемые при данном способе, транслируются с использованием системы трансляции митохондрий, а не стандартной клеточной рибосомальной системы трансляции. Таким образом, белки, продуцируемые с использованием данного способа более похожи на вирусные белки, продуцируемые при нормальной инфекции, и поэтому обладают антигенными свойствами, которые существуют во время инфекции. Такие белки, продуцируемые при использовании данного способа, можно использовать для получения иммунного ответа у млекопитающих, и антигенные детерминанты могут иметь более близкое сходство с теми, которые продуцируются при инфекции, чем детерминанты на белках, продуцируемых с использованием стандартной технологии с рекомбинантной ДНК, которая основана на клеточной рибосомальной трансляции.
Изобретение также включает способ продукции белков с помощью клонированной ДНК, содержащейся в векторе на основе вируса, причем трансляция происходит in vitro в богатых митохондриями клетках животных, трансфицированных вектором, причем клетки содержатся в автоматической динамической системе культивирования. Эффективная система экспрессии на основе ВГВ используется для продукции белков, зависящей от трансляции в богатой митохондриями ткани. При этом воплощении изобретения вектор экспрессии на основе ВГВ, содержащий клонированную кодирующую последовательность ДНК, встроенную в структурный ген ВГВ, используется для прямой генной экспрессии клонированной ДНК в трансфицированной ткани органа животного, культивируемой in vitro с использованием предпочтительной автоматической системы для культивирования.
Двунитевая ДНК ВГВ (содержащая "минус" нить и "плюс" нить ДНК последовательностей) использована для построения кольцевого вектора ДНК, в который могут быть встроены другие кодирующие последовательности ДНК с использованием стандартных методов молекулярной биологии. Вектор на основе ВГВ также содержит последовательности из прокариотной плазмиды, которые дают возможность репликации вектора в прокариотах для амплификации ДНК. Вектор содержит ген лекарственной устойчивости для обеспечения маркера для селекции в трансфицированных клетках (например, устойчивости к гигромицину В). Вставочная кодирующая последовательность ДНК встраивается в структурный ген ВГВ, не требующий репликации в клетках животных. Вставочная кодирующая последовательность может быть последовательностью другого вирусного гена, кДНК, ДНК, амплифицированной с помощью полимеразной цепной реакции, или последовательностью синтетической ДНК, и вставка выполняется с использованием стандартных методов молекулярной биологии разрезания и лигирования на место вставочной ДНК в соответствующей рамке и ориентации для того, чтобы дать возможность экспрессии по последовательностям ВГВ. Так как было обнаружено, что репликация ВГВ происходит в ткани печени и почек у некоторых трансгенных мышей, имеющих удлиненные, длиной более чем геном, конструкции ВГВ (Guidotti et al., J. Virol. 69: 6158-6169, 1995), эти результаты свидетельствуют, что трансгенные конструкции могли быть трансфицированы скорее в митохондрии, чем в ядро. Таким образом, рекомбинантные конструкции, содержащие ВГВ длиной более чем геном, могут также применяться для трансфекции в ткань, поддерживаемую in vitro с использованием данной системы, и они считаются функционально эквивалентными конструкциям, обсуждаемым здесь для данного способа.
Изобретение может быть лучше понято с помощью последующих примеров, которые являются представлениями предпочтительных воплощений.
Пример 1
Инфицирование ВГВ ткани почек крысы in vitro
Белым
крысам смешанной линии делали анестезию в основном эфиром и хирургически вскрывали брюшную полость, используя известные специалистам
методики. Затем в аорту после разреза полой вены делали инъекцию 10
мл охлажденного (примерно 4oС) висконсинского (Wisconsin) раствора (Viaspan, DuPont) для обеспечения возможности
перфузии. Почки удаляли из обескровленной области и хранили в охлажденном
висконсинском растворе (примерно 4oС). Срезы ткани почки (например, кусочки 2 см2 толщиной примерно 260
мкм) готовили и хранили в охлажденной культуральной среде. Срезы
инкубировали с ВГВ, полученным из ткани печени, взятой путем биопсии у инфицированного больного человека. Посевной материал ВГВ
готовили, помещая биопсийный материал от больных, в крови которых
присутствовали поверхностные антигены вируса гепатита В, в модифицированную среду Веймаут (Waymouth's) MB 752/1 на 3 ч при 37oС; биопсийные образцы удаляли через 3 ч, а срезы ткани органа
крысы затем культивировали в среде. В основном соотношение количества биопсийной ткани и среды составляло 5020 г ткани на 10 мл
среды. В качестве контроля срезы ткани почки крысы культивировали в
среде, которая не контактировала с биопсийным материалом ткани печени человека.
Инфицированные срезы почки крысы культивировали в автоматической системе для культивирования ткани органов, которая показана на фиг.1, в которой вырезанный срез ткани органа 10 помещается внутри пористого контейнера 11, который помещается внутри трубки для культивирования 12, которая может вращаться и имеет по меньшей мере одно входное отверстие 13 для впуска газов, введения среды, факторов роста и тому подобного. Пористый контейнер 11 изготавливается из любого инертного вещества, включая, но не ограничивая этим, пластиковую сетку, найлоновую сетку или полупроницаемую мембрану, но предпочтительно является сеткой из нержавеющей стали в форме квадратной или прямоугольной коробочки, имеющей средний размер пор, равный примерно от 100 до примерно 500 мкм. Пробирка для культивирования 12 включает отверстие 14 для отбора образцов культуральной среды ткани 15, которое можно многократно запечатывать. Отверстие для отбора образцов 14 можно также использовать для ввода среды 15, вирусных частиц, факторов роста и других реагентов для культивирования или веществ для воздействия на образец ткани in vitro. Ткань органа 10 периодически погружается в среду для культивирования ткани 15, когда трубка для культивирования 12 вращается. Форма коробки пористого контейнера 11 лучше способствует переворачиванию образца, когда трубка для культивирования 12 вращается, чем когда контейнер остается в одном положении, а трубка для культивирования вращается вокруг него. Газообмен в пробирке для культивирования 12 происходит через интервалы, через которые газовая смесь вводится во входное отверстие 13, а газ вытесняется через выходное отверстие 16 трубки для культивирования. Трубка для культивирования 12 содержится при постоянной температуре, равной 37oС (например, в термостате, который не показан). Процесс культивирования ткани органа предпочтительно автоматизирован для содержания клеток в одних и тех же условиях в течение всего периода инкубации.
Срез ткани культивируют при 37oС в модифицированной культуральной среде Уеймаута МБ 752/1 с рН 7,0 при давлении в 1,6-2,0 атм газовой смеси из 5% CO2 и 95% O2. Культуральную среду готовили из порошкообразной среды Уеймаута MB 752/1 (Gibco), 10% фетальной бычьей сыворотки, 2,2% бикарбоната натрия, 25 мМ D-глюкозы, 1 мкг/мл кристаллического бычьего цинк-инсулина, смеси антибиотиков, содержащей 50 Ед/мл пенициллина и 50 мкг/мл стрептомицина (Gibco) и дистиллированной воды. Газообмен производили через интервалы в 2,5 мин, и ткань погружалась в культуральную среду 4,5 раза в мин с помощью вращения трубки для культивирования, показанной на фиг.1.
Инкубация инфицированной ВГВ ткани почек составляла в основном от примерно 1 до 48 ч, предпочтительно примерно 24 ч. Затем ткань обрабатывали, применяя стандартные методы иммунохимии, путем разрезания ткани и окрашивания с помощью антител к HBsAg (купленных у SIGMA, St. Louis, МО) для определения присутствия ВГВ в инфицированной ткани.
Обычно после менее 24 ч культивирования HBsAg обнаруживались в клетках почек. Инфицированная ткань почки окрашивалась антителами к HBsAg неоднородно, причем богатые митохондриями проксимальные канальцы демонстрировали большую интенсивность окрашивания по сравнению с относительно бедными митохондриями дистальными канальцами. Когда рассеченную инфицированную ВГВ ткань почки крысы исследовали с применением электронной микроскопии, в проксимальных канальцах определялась значительно более высокая концентрация похожих на митохондрии органелл с многослойной мембраной, содержащих HBsAg, чем в дистальных канальцах. Таким образом, эффективность инфицирования ВГВ связана с концентрацией митохондрии в ткани животного. Эти результаты также показывают, что в противоположность современным концепциям перекрестно-видовой вирусной инфекции ВГВ может инфицировать орган животного, который имеет достаточное количество митохондрий, чтобы дать возможность репликации и реплицироваться в нем. В дополнение к ткани почки крысы в автоматической системе для культивирования, описанной выше, успешно культивировали ткани печени собак, мышей, цыплят и лягушек. Опытным специалистам будет понятно, что подобные ткани животных также можно инфицировать ВГВ или другими человеческими или нечеловеческими вирусами (например, вирусами гепатита А и С или энцефалита), которые инфицируют богатые митохондриями ткани, чтобы дать возможность репликации вируса in vitro в этой системе. Опытным специалистам будет понятно, что такие ткани животных могут также включать ткань человека, инфицированную человеческим вирусом или вирусом животного.
Пример 2
Иифицирование ВГВ ткани печени крысы
локализуется в органеллах митохондрии
Ткань
печени хирургически забирали у белых крыс смешанной линии, как описано, для удаления почек в примере 1. Из ткани печени делали срезы и
инфицировали их ВГВ по существу так же, как описано в примере 1.
Инфицированную ткань печени крысы затем инкубировали в автоматической системе для культивирования в течение примерно 24 ч и затем
ткань исследовали на присутствие HBsAg и ее антигена ВГВ (НВеАg),
применяя иммуноферментный анализ, при котором выявляются эти антигены, с использованием известных специалистам методик (т. е. набора
для ИФА на ВГВ, поставляемого Abbott Laboratories). Инфицированную
ткань также исследовали на ДНК ВГВ с помощью ДНК-гибридизации с использованием стандартных методик саузерн-блоттинга (по существу
как описано у Guidotti et al., J. Virol. 69: 6158-6169, 1995).
Инфицированную ткань печени крыс сначала фракционировали на цитоплазматическую растворимую (цитозоль) фракцию и осадок, содержащий митохондрии, используя стандартный метод фракционирования клеток (по существу, как описано Jensen et al., Biochim. et Biophys. Acta 1180: 65-72, 1992). В кратком изложении, срезы инфицированной ткани гомогенизировали в буфере (0,25 М сахароза, 0,1 мМ ЭДТА и 1 мМ трис-HCl, рН 7,4) и центрифугировали при низкой скорости (700 х g) для удаления ядер и неразрушенных клеток (ядерная фракция). Супернатант центрифугировали при высокой скорости (12000 х g), чтобы отделить фракцию митохондрии (в осадке) и фракцию цитозоля (в супернатанте). Фракции ядер, митохондрии и цитозоля затем испытывали на наличие HBsAg и НВеАg, используя метод ИФА для определения этих двух антигенов.
Фракция митохондрии содержала по меньшей мере в 10 раз больше HBsAg, чем обнаружено в любой из ядерной или цитозольной фракций. Эти результаты показывают, что ВГВ реплицируется в ткани печени крысы главным образом в митохондриях или в митохондриеподобных органеллах, которые фракционируются вместе с только ограниченной репликацией ВГВ, происходящей в клеточных ядрах.
При применении стандартного разделения в геле и методик ДНК-гибридизации были обнаружены реплицирующиеся комплексы, состоящие из ДНК ВГВ размером менее или равным 2,1 т.п.о., во фракциях митохондрии. Не было обнаружено ДНК ВГВ во фракции цитозоля и незначительное количество (менее примерно 10%, обнаруженного во фракции митохондрии) было найдено во фракции ядер.
Пример 3
Сравнение HBsAg, выделенных из человеческой плазмы с HBsAg, продуцируемыми с
рекомбинантной ДНК
HBsAg в вакцине, произведенной из человеческой плазмы (гепавакс
[Hepavax], полученная из Синего Креста [Blue Cross] Корея) сравнили с HBsAg, полученными с помощью
технологии с рекомбинантной (полученными от JEIL-JEDANG, Сеул, Корея), используя электрофорез в
полиакриламидном геле с ДСН (ДСН-ЭПАГ, SDS-PAGE). Белки растворяли в буфере, содержащем 40 мМ Трис-HCl,
рН 6,8, 1% ДСН, 0,35% β-меркаптоэтанола, 5% глицерина и бромфеноловый синий, и кипятили в
течение 5 мин перед разделением в геле для ДСН-ЭПАГ с использованием стандартных методик (Laemmli U.K.,
Nature 227: 680-685, 1970). После электрофореза белки подвергали иммуноблоттингу с применением
хорошо известных методов и антител к HBsAg (полученных от SIGMA, St. Louis, МО).
HBsAg, продуцируемые с помощью технологии с рекомбинантной ДНК, давали только одну полосу для 23 кД, тогда как HBsAg, выделенные из человеческой плазмы, представляли широкий спектр поверхностных антигенов в широкой размазанной полосе в области от примерно 20 кД до примерно 30 кД. Эти результаты означают, что в митохондриях может продуцироваться много существующих в естественном состоянии антигенов ВГВ с использованием антигенных генов ядра и использования кодона, уникального для митохондрий по сравнению с единственным белком, продуцируемым по технологии с рекомбинантной ДНК. Так как вакцина, полученная из плазмы в основном более эффективна, чем вакцина, получаемая путем генной инженерии, эти результаты также означают, что многочисленные различные формы поверхностных антигенов ВГВ, продуцируемые во время инфекции, могут отдельно или вместе служить в качестве лучших иммуногенов, чем единственный антиген ВГВ, продуцируемый по технологии с рекомбинантной ДНК.
Пример 4
Продукция HBsAg в митохондриях с использованием митохондриальной системы трансляции
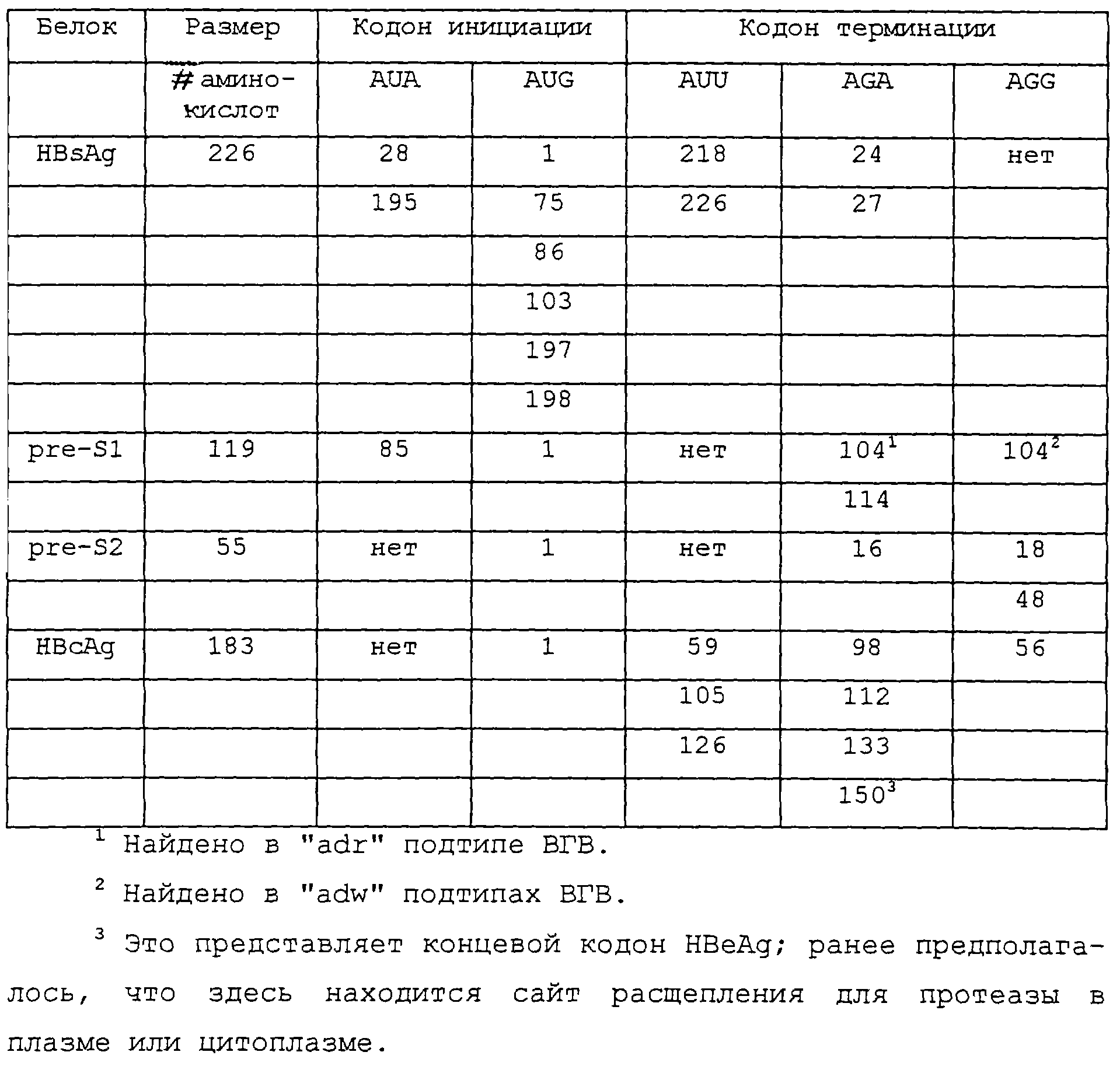
В системе использования кодонов митохондрий млекопитающих кодоны AGA и AGG служат в
качестве стоп-кодонов для прекращения трансляции. Ген для ядерного HBsAg содержит кодоны AGA и AGG, которые,
как предполагают, являлись сайтами расщепления для процессинга ядерного антигенного белка
в зрелый HBsAg. Однако при трансляции в митохондриях млекопитающих ген для ядерного HBsAg в естественных
условиях заканчивается у кодонов AGA и AGG. На основе использования генетического кодона
митохондрий существует несколько других предсказанных кодонов инициации и прекращения в гене HBsAg (суммированы
в таблице). Те же самые определения были сделаны для генов, кодирующих белки ВГВ,
называемые pre-S1 и prе-S2 и ядерный антиген (НВсАgд), и эти локусы кодонов инициации и терминации также показаны в
таблице (в отношении общего обсуждения белков ВГВ см. Lau and Wright, Lancet 342:
1335-1340, 1993).
Ткань печени крыс инфицировали ВГВ по существу, как описано в примере 2, и инфицированную ткань крыс культивировали in vitro в течение 12-48 ч. После инкубации инфицированную ткань крыс собирали и лизировали в буфере, содержащем 40 мМ Трис-HCl рН 6,8, 1% ДСН, 0,35% β -меркаптоэтанола, 5% глицерина и бромфеноловый синий. Лизат кипятили в течение 5 мин и разделяли в 10% полиакриламидном геле с ДСН путем электрофореза (ДСН-ЭФАГ), применяя стандартные методы (Laemmi U.K., Nature 227: 680-685, 1970). Для сравнения HBsAg, полученный с помощью технологии с рекомбинантной ДНК, включен в качестве контроля в соседнюю дорожку в гель для ДСН-ЭПАГ. После разделения белки подвергали иммуноблоттингу и определяли с помощью антител к HBsAg, используя хорошо известные методики.
HBsAg, продуцируемые в инфицированной ткани крысы, выращенной in vitro, содержат белки размером от примерно 20 кД до примерно 30 кД, подобные тем, которые определялись в плазме людей, хронически инфицированных ВГВ. Таким образом, транслирование генов ВГВ in vitro в богатой митохондриями ткани продуцирует ряд секреторных антигенов, которые имитируют антигены, которые естественно продуцируются у инфицированных людей. По контрасту с этим HBsAg, продуцируемые с помощью технологии с рекомбинантной ДНК, проявлялись в виде единственной полосы у примерно 23 кД. Многочисленные белки HBsAg, продуцируемые печенью крыс при инфицировании in vitro, выделены для использования в качестве вакцины против инфекции ВГВ.
Пример
5
Продукция белков в трансфицированной ткани животного с использованием вектора экспрессии на основе ВГВ
Эффективная
система экспрессии на основе ВГВ подобным же образом может
использоваться для продукции белков, зависимых от трансляции в богатых митохондриями тканях. То есть вектор экспрессии на основе ВГВ может
использоваться для прямой экспрессии генов клонированной ДНК
в трансфицированной ткани органа крысы, культивируемой in vitro, как описано в примерах 1 и 2.
ВГВ представляет вирусную ДНК, имеющую геном в 3200 оснований, включающий "минус" цепь и короткую "плюс" цепь, которые вместе составляют частично двунитевую кольцевую ДНК, которая кодирует структурные белки и белки, необходимые для репликации вируса (Lau and Wright, Lancet 342: 1335-1340, 1993).
Вектор на основе ВГВ содержит последовательности из прокариотной плазмиды pBR322, имеет репликацию, происходящую из ВГВ, усеченный ген полимеразы ВГВ и ген лекарственной устойчивости (например, ген гигромицин-B-фосфотрансферазы под контролем тимидинкиназных регуляторных последовательностей ВПГ (вируса простого герпеса), обеспечивающей устойчивость к гигромицину В).
Обращаясь к фиг.2, вектор на основе ВГВ, названный pHBVex, включает ДНК последовательности из прокариотного вектора pBR322 (обозначенный "pBR"), чтобы дать возможность репликации вектора в прокариотных клетках, включая Escherichiacoli, последовательности (обозначенный "AmpR"), которые несут устойчивость к ампициллину, когда экспрессируется в Е. coli; ген гигромицин-B-фосфотрансферазы (обозначенный "HYG") под контролем последовательностей тимидинкиназного промотора ВПГ (обозначенного "HSV TK pro") и последовательностей терминации (обозначенной "HSV TK"), которые делают эукариотные клетки экспрессирующими ген, резистентности к гигромицину В; вставочная ДНК последовательность (обозначенная "встДНК"), которая может быть геномной последовательностью или последовательностью кДНК, кодирующей белок, который нужно экспрессировать под контролем усеченного полимеразного гена ВГВ (обозначенного "HBVp"). Усечение полимеразного гена ВГВ и вставка чужеродной ДНК происходит в области между концевым белком для репликации и упаковки и началом гена pre-S1. Остаток этой плазмиды состоит из "минус" цепи ДНК ВГВ (обозначенной "HBV") и ее стандартной комплементарной последовательности ДНК, полученной путем стандартных генетических методик, включая обратную транскрипцию, ДНК-полимеризацию с синтетическим праймером и лигирование двунитевой ДНК, представляющей "минус" цепь ВГВ, в оставшиеся части вектора (Sambrook et al. Molecular Cloning, A Laboratory Manual (2nd Ed.), Vol. 1-3, Cold Spring Harbor, NY, 1989).
В векторе pHBVex часть кодирующей последовательности гена полимеразы ВГВ заменяется чужеродной последовательностью ДНК (или вирусного, или эукариотного гена, кДНК или ДНК, амплифицированную путем полимеразной цепной реакции) с применением стандартных методов молекулярной биологии (Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Ed., Vol. 1-3, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989) расщепления рестрикционными ферментами и легирования на место вставочной ДНК в соответствующей рамке и ориентации, чтобы дать возможность экспрессии с регуляторными последовательностями ВГВ. Стрелки внутри круга показывают ориентацию (направление транскрипции) последовательностей ДНК.
Другие последовательности ДНК в эквивалентном векторе pHBVex (не показан) могут включать последовательности, полученные из других прокариотных векторов, из вируса гепатита А, вируса гепатита С или других вирусов, включая вирус Эпштейна-Барра, вирусы простого герпеса (ВПГ) и вирусы энцефалита. Опытным специалистам будет понятно, что как эквиваленты для вектора, представленного в виде диаграммы на фиг.2, могут быть использованы другие векторы экспрессии на основе ВГВ. Например, вектор, подобный pHBVex, но содержащий конструкцию усеченного более чем одного генома ВГВ в векторе, может быть оптимальным для репликации или экспрессии гена, аналогично результатам, полученным у трансгенных мышей, имеющих усеченные конструкции ВГВ (Guidotti et al. , J. Virol. 69:6158-6169, 1995). Кроме того, опытным специалистам будет понятно, что трансфекция с использованием вектора pHBVex или эквивалентного вектора могло также включать сотрансфекцию или инфицирование хелперным вирусом для того, чтобы вызвать или усилить репликацию или экспрессию гена из векторной ДНК.
Ткань животного выделяют из богатых митохондриями органов и подготавливают для культивирования in vitro, по существу, как описано в примерах 1 и 2. Вектор pHBVex, содержащий вставочную ДНК, трансфицируют в богатую митохондриями ткань, применяя стандартную методологию трансфекции, включая осаждение фосфатом кальция, слияние клеток ткани с бактериальными протопластами, содержащими конструкцию pHBVex-встДНК, обработку ткани липосомами, содержащими последовательность pHBVex-встДНК, вызываемую ДЭАЭ-декстраном трансфекцию, электропорацию и микроинъекцию ДНК.
Срезы трансфицированной ткани культивируют in vitro в автоматической системе по существу так, как описано в примере 1, чтобы обеспечить возможность продукции белка, получаемой при экспрессии трансфицированной ДНК в богатой митохондриями ткани. Белок очищают, используя любой из множества стандартных методов, включая аффинную хроматографию. С использованием системы экспрессии на основе ВГВ могут продуцироваться другие вирусные антигены, которые имитируют антигены, продуцируемые во время естественного инфицирования вирусами, которые инфицируют богатую митохондриями ткань (например, другие гепатитные вирусы или вирусы энцефалита) для изготовления эффективных вакцин против этих возбудителей.
Пример 6
Продукция антигенов человеческого ВГС в трансфицированной ткани животного с использованием вектора экспрессии на основе ВГВ
Так как прямое культивирование ВГС
(HCV) в ткани животного в
динамической системе для культивирования ткани может все же быть неэффективным способом для получения достаточного количества антигенов ВГС (например, из-за того, что ВГС
реплицируется относительно
медленно), использование вектора на основе другого вектора является ценным вариантом для продукции антигенов ВГС in vitro. Вектор pHBVex использован для переноса генов,
кодирующих антигены
человеческого вируса гепатита С, в богатые митохондриями клетки для продукции природных антигенов с использованием системы трансляции митохондрий по существу, как описано в
примере 5. Так как вирус
гепатита С является РНК вирусом, РНК последовательность, кодирующая поверхностный антиген гепатита С (HCsAg) сначала подвергается обратному считыванию в кДНК с применением
хорошо известных методик
(Sambrook et al., Molecular Cloning, A Laboratory Manual (2nd Ed.), Vol. 1-3, Cold Spring Harbor, NY, 1989). КДНК HCsAg встраивается в усеченный полимеразный ген
ВГВ вектора pHBVex с
применением стандартных методик рестрикционного расщепления векторной ДНК и лигирования (с использованием соответствующих сайтов разрезания ферментом рестрикции или лигирования к
затупленным концам)
двунитевой кДНК, кодирующей HCsAg. Конструкцию pHBVex-HCsAg трансфицируют в выделенные срезы ткани печени крысы и культивируют in vitro в течение 24-48 ч, используя по существу
методы, описанные в
примерах 1, 2 и 5. После 24-48 ч культивирования ткань удаляют и белок HCsAg, произведенный в трансфицированной ткани, очищают, применяя стандартные методики очистки белков,
включая аффинную
хроматографию с использованием антител, которые связывают белок HCsAg.
Данное изобретение включает способ, применимый для получения белков, которые продуцируются в естественных условиях в богатых митохондриями клетках (например, белки, продуцируемые в печени или поджелудочной железе). Способ трансляции данного изобретения может использоваться для продукции природных немитохондриальных белков, которые транслируются в митохондриях. Это может быть особенно важно при продукции белков, которые обладают такими иммуногенными характеристиками, такими как процессинг или распознавание кодона, зависимые от трансляции в митохондриях. То есть данное изобретение применимо для продукции природных антигенов вирусов, которые реплицируются в митохондриях, или тех, которые реплицируются слишком медленно, когда культивируются с использованием обычных способов культивирования тканей, или тех, которые не могут продуцироваться с применением обычной технологии с рекомбинантной ДНК. Существует потребность в продукции белков из инфекционных возбудителей, особенно человеческих инфекционных возбудителей, в системе in vitro. Предпочтительной является перекрестновидовая, так как это ограничивает опасность загрязнения желаемого продукта нежелательным продуктом из того же самого вида. Например, способ инфицирования человеческим инфекционным возбудителем, который не зависит от человеческих клеток для роста инфекционного возбудителя, ограничивает опасность загрязнения другим человеческим инфекционным возбудителем (например, ВИЧ присутствующим в человеческой ткани). Подобным же образом, существует потребность в системе in vitro, которая эффективно имитирует инфекцию у человека, для продукции иммуногенов, которые похожи на иммуногены, продуцируемые при инфекции у человека, которые невозможно получить при использовании обычных методик, используемых для продукции белка с рекомбинантной ДНК.
Это изобретение представляет способ продукции белка с использованием рекомбинантного вектора на основе ВГВ, который применим для направления продукции других немитохондриальных белков в митохондрии трансфицированных клеток животных.
Изобретение также дает возможность выращивать вирус в системе in vitro, которая применима для открытия новых терапевтических средств для профилактики заболевания и улучшения современных методов лечения патологических состояний, вызываемых вирусной инфекцией у людей.
Реферат
Изобретение относится к медицине и касается способа продукции вирусных антигенов in vitro путем инфицирования ткани органа животного, богатого митохондриями, вирусом, включая вирус гепатита В (ВГВ), и культивирования инфицированной ткани человеческим вирусом. Раскрыт способ продукции белков in vitro путем трансфекции богатой митохондриями ткани животного рекомбинантным вектором на основе ВГВ и культивирования трансфицированной ткани в динамической системе для культивирования ткани. 4 с. и 19 з.п. ф-лы, 1 табл., 2 ил.
Комментарии