Aav векторы с усовершенствованными rep-кодирующими последовательностями, используемыми в системах продукции на основе клеток насекомых - RU2457252C2
Код документа: RU2457252C2
Чертежи














Описание
Область техники, к которой относится изобретение
Настоящее изобретение относится к продукции аденоассоциированного вируса в клетках насекомых и к аденоассоциированному вирусу, который обладает улучшенными характеристиками экспрессии и стабильностью вирусных белков rep, что повышает продуктивность векторов на основе аденоассоциированных вирусов в клетках насекомых.
Предпосылки создания изобретения
Аденоассоциированный вирус (AAV) может рассматриваться как один из наиболее многообещающих вирусных векторов для использования в генной терапии человека. AAV обладает способностью эффективно инфицировать как делящиеся, так и неделящиеся клетки человека, при этом геном AAV интегрирует в единичный сайт хромосомы в геном клетки-хозяина, и что наиболее важно, наличие AAV в организме большого числа людей никогда не связывали с каким-либо заболеванием. Ввиду этих преимуществ, рекомбинантный аденоассоциированный вирус (rAAV) анализировали в рамках клинических испытаний для применения в генной терапии гемофилии B, злокачественной меланомы, муковисцидоза и других заболеваний.
Клетки-хозяева, которые поддерживают репликацию AAV in vitro, все получены из клеток различных типов, принадлежащих млекопитающим. Таким образом, rAAV, используемый в генной терапии, в основном получают на основе клеточных линий млекопитающих, таких как, например, клетки 293, COS клетки, HeLa клетки, KB клетки и другие клеточные линии млекопитающих (см., например, US 6156303, US 5387484, US 5741683, US 5691176, US 5688676, US 20020081721, WO 00/47757, WO 00/24916 и WO 96/17947). Как правило, векторы rAAV получают в этих культурах клеток млекопитающих путем введения ДНК-плазмид, содержащих терапевтический ген, фланкированный точкой начала репликации AAV (инвертированные концевые повторы или ITR), гены для репликации белков AAV Rep78, Rep68, Rep52 и Rep40, а также гены для образования вирионов или структурных белков VP1, VP2 и VP3. Кроме того, вводят плазмиду, содержащую ранние гены аденовируса (E2A, E40RF6, VARNA), для усиления экспрессии генов AAV и повышения выхода вектора (см., например, Grimm et al., 1998, Hum. Gene Ther. 9: 2745-2760). Однако в большинстве таких культур клеток млекопитающих число частиц AAV в расчете на клетку составляет порядка 104 частиц (см. обзор Clark, 2002, Kidney Int. 61 (Suppl. 1): 9-15). Для клинического испытания может потребоваться более 1015частиц rAAV. Для получения такого количества частиц rAAV требуется трансфицировать и культивировать примерно 1011 клеток 293 человека, что эквивалентно 5000 колб с массой клеток, соответствующей площади 175 см2, что, в свою очередь, означает проведение трансфекции 1011 клеток 293. Соответственно, как это уже было доказано, крупномасштабное получение rAAV на основе культуры клеток млекопитающих для получения материала для клинических испытаний является проблематичным, а коммерческий масштаб получения невозможен. Кроме того, имеется также риск того, что вектор для клинического использования, который образуется в культуре клеток млекопитающих, будет загрязнен нежелательным, возможно, патогенным материалом, присутствующим в клетках млекопитающего-хозяина.
Для преодоления такого рода трудностей, связанных с использованием систем клеток млекопитающих, в последнее время была разработана система продукции AAV с использованием клеток насекомых (Urabe et al., 2002, Hum. Gene Ther. 13: 1935-1943; US 20030148506 и US 20040197895). Для получения AAV в клетках насекомых необходимо сделать некоторые модификации для достижения корректной стехиометрии всех трех капсидных белков AAV (VP1, VP2 и VP3), которые основаны на сочетании чередующегося использования двух акцепторных сайтов сплайсинга и субоптимальном использовании ACG-инициирующего кодона для VP2, который не совсем точно репродуцируется клетками насекомых. Для сохранения корректной стехиометрии капсидных белков в клетках насекомых Urabe et al. (2002, supra) использовали конструкцию, которая транскрибируется в общий полицистронный мессенджер, способный экспрессировать все три белка VP, без необходимости сплайсинга и где большая часть инициирующего кодона против направления считывания информации замещена субоптимальным инициирующим кодоном ACG. В совместно рассматриваемой заявке (PCT/NL2005/050018) авторы настоящего изобретения также улучшили инфективность бакуловирусных векторов rAAV на основе продукции с последующей оптимизацией стехиометрии капсидных белков AAV в клетках насекомых.
Для экспрессии Rep белков AAV в системе клеток насекомых с экспрессией AAV, первоначально разработанной Urabe et al. (2002, supra), используют рекомбинантную бакуловирусную конструкцию, которая несет две независимые единицы экспрессии Rep (одну - для Rep78, а другую - для Rep52), где каждая находится под контролем независимого промотора клетки насекомого, промоторов ΔIЕ1 и PolH, соответственно. В этой системе для осуществления экспрессии Rep78 был выбран промотор ΔIE1, который значительно слабее промотора PolH, поскольку известно, что в клетках млекопитающих экспрессия Rep78 менее активна, по сравнению с Rep52, и приводит к высокому выходу вектора (Li et al., 1997, J Virol. 71: 5236-43; Grimm et al., 1998, supra).
Однако несколько позже Kohlbrenner et al. (2005, Mol. Ther. 12: 1217-25) сообщили, что бакуловирусная конструкция для экспрессии двух белков Rep, использованная Urabe et al., отличается присущей ей нестабильностью. При расщеплении палиндромной структуры двух генов Rep в исходном векторе, использованном Urabe, и при конструировании двух отдельных бакуловирусных векторов экспрессии Rep52 и Rep78, Kohlbrenner et al. (2005, supra) удалось повысить стабильность вектора при проведении пассажей. Однако, несмотря на устойчивую экспрессию Rep78 и Rep52 из двух независимых бакуловирусных конструкций на основе Rep в клетках насекомых в течение по меньшей мере 5 пассажей, выход вектора rAAV был в 5-10 раз ниже, по сравнению с исходной конструкцией бакуловируса на основе Rep, созданной Urabe et al. (2002, supra).
В этой связи существует потребность в преодолении вышеуказанных серьезных ограничений, свойственных крупномасштабной (коммерческой) продукции векторов AAV в клетках насекомых. Таким образом, задачей настоящего изобретения является создание средств и способов, при которых возможно получение стабильного и высокого крупномасштабного выхода векторов AAV в клетках насекомых.
Описание изобретения
Определения
В контексте настоящего описания термин «функционально связанный» относится к связи полинуклеотидных (или полипептидных) элементов в функциональную связь. Нуклеиновая кислота является «функционально связанной», если она находится в условиях функциональной связи с другой последовательностью нуклеиновой кислоты. Например, регуляторная последовательность транскрипции функционально связана с кодирующей последовательностью, если она влияет на транскрипцию указанной кодирующей последовательности. Термин «функционально связанный» означает, что связанные последовательности ДНК являются, как правило, непрерывными, и при необходимости соединения двух участков, кодирующих белок, являются также непрерывными и находятся в рамке считывания.
Фраза «контрольная последовательность экспрессии» относится к последовательности нуклеиновой кислоты, которая регулирует экспрессию нуклеотидной последовательности, с которой она функционально связана. Контрольная последовательность экспрессии является «функционально связанной» с нуклеотидной последовательностью, если такая контрольная последовательность экспрессии контролирует и регулирует транскрипцию и/или трансляцию нуклеотидной последовательности. Таким образом, контрольная последовательность экспрессии может включать промоторы, энхансеры, внутренние сайты входа в рибосому (IRES), терминаторы транскрипции, старт-кодон перед геном, кодирующим белок, сигнал сплайсинга для интронов и стоп-кодоны. Под термином «контрольная последовательность экспрессии» понимают последовательность, которая включает, как минимум, последовательность, присутствие которой необходимо для воздействия на экспрессию и которая может также включать дополнительные полезные компоненты. Так, например, лидерные последовательности и слитые последовательности представляют собой пример контрольных последовательностей экспрессии. Этот термин также может включать конструкцию последовательности нуклеиновой кислоты, в которой удалены нежелательные возможные кодоны инициации транскрипции как внутри, так и за пределами рамки считывания. Этот термин может также включать конструкцию последовательности нуклеиновой кислоты, в которой удалены нежелательные возможные сайты сплайсинга. Контрольная последовательность экспрессии также включает последовательности или последовательности полиаденилирования (pA), которые регулируют добавление полиА хвоста, то есть удлинение цепи адениновыми остатками на 3'-конце мРНК, последовательности, которые называют полиА последовательностями. Указанные контрольные последовательности могут быть сконструированы с тем, чтобы усиливать стабильность мРНК. Контрольные последовательности экспрессии, которые влияют на стабильность транскрипции и трансляции, например промоторы, а также последовательности, которые влияют на трансляцию, например последовательности Козака, известны для клеток насекомых. Природа контрольных последовательностей экспрессии может определять их способность модулировать нуклеотидную последовательность, с которой они функционально связаны, для снижения уровней экспрессии или повышения уровней экспрессии.
В контексте настоящего описания термин «промотор» или «регуляторная последовательность транскрипции» относится к фрагменту нуклеиновой кислоты, который контролирует транскрипцию одной или нескольких кодирующих последовательностей, и который расположен против направления считывания информации относительно направления транскрипции от сайта инициации транскрипции кодирующей последовательности, а также который структурно идентифицируется по наличию сайта связывания для ДНК-зависимой РНК-полимеразы, сайтов инициации транскрипции и других последовательностей ДНК, включающих, без ограничения, сайты связывания фактора транскрипции, сайты связывания репрессора и активатора белка, а также любые другие последовательности нуклеотидов, известные специалистам в данной области, которые непосредственно или опосредованно регулируют уровень транскрипции с данным промотором. «Конститутивный» промотор представляет собой такой промотор, который активен в большинстве тканей в обычных физиологических условиях и условиях развития. «Индуцибельный» промотор представляет собой промотор, который подвергается физиологической регуляции или регуляции в ходе развития, например, при воздействии химического индуктора. «Тканеспецифичный» промотор активен только в конкретных типах тканей или клеток.
Термины «по существу идентичные» или «по существу сходные» в контексте настоящего описания используют для обозначения того, что две пептидных или две нуклеотидных последовательности, при их оптимальном выравнивании, например, с помощью программ GAP или BESTFIT с использованием параметров по умолчанию, обладают по меньшей мере некоторым процентом идентичности по последовательности. В программе GAP используется алгоритм глобального выравнивания Needleman и Wunsch для выравнивания двух последовательностей по всей их длине, максимизации числа совпадений и минимизации числа пропусков. В основном, в программе GAP используются параметры по умолчанию, в соответствии с которыми штраф за пропуск составляет 50 (нуклеотидов)/8 (белков) и штраф за удлинение пропуска составляет 3 (нуклеотидов)/2 (белков). В случае нуклеотидов используют заданную балльную матрицу nwsgapdna, а для белков заданной по умолчанию балльной матрицей является Blosum62 (Henikoff & Henikoff, 1992, PNAS 89, 915-919). Очевидно, что в том случае, когда речь идет о том, что последовательности РНК по существу сходны или обладают определенной степенью идентичности последовательности с последовательностью ДНК, то в этом случае тимин (T) в последовательности ДНК рассматривается как урацил (U) в последовательности РНК. Выравнивание последовательностей и определение баллов процента идентичности последовательностей может быть проведено с использованием таких компьютерных программ, как программа GCG Wisconsin Package, версия 10.3, доступная от компании Accelrys Inc., 9685 Scranton Road, San Diego, CA 92121-3752 USA, или доступного из открытого источника пакета программ Emboss для Windows (современная версия 2.7.1-07). В альтернативном варианте, процент сходства или идентичности может быть определен путем поиска в соответствующих базах данных, таких как FASTA, BLAST и т.п.
Нуклеотидные последовательности, кодирующие парвовирусные белки Rep по настоящему изобретению, могут быть также определены по их способности гибридизоваться с нуклеотидной последовательностью SEQ ID NO:10, соответственно, в умеренных или, предпочтительно, в жестких условиях гибридизации. Жесткие условия гибридизации по настоящему изобретению представляют собой условия, которые позволяют гибридизоваться последовательностям нуклеиновой кислоты из по меньшей мере примерно 25, предпочтительно, примерно 50 нуклеотидов, 75 или 100 и, наиболее предпочтительно, примерно 200 или более нуклеотидов, при температуре около 65°C в растворе, содержащем примерно 1М соли, предпочтительно, в растворе 6 × SSC или в любом другом растворе, обладающем сравнимой ионной силой, и при промывке при температуре 65°C в растворе, содержащем примерно 0,1M соли или менее, предпочтительно, в растворе 0,2 × SSC или любом другом растворе, обладающем сравнимой ионной силой. Предпочтительно, гибридизацию проводят в течение ночи, то есть в течение по меньшей мере 10 часов, а промывку, предпочтительно, проводят в течение по меньшей мере одного часа с по меньшей мере двумя заменами промывочного раствора. Эти условия обычно позволяют достичь специфической гибридизации последовательностей с идентичностью последовательностей примерно 90% или больше.
Умеренные условия в соответствии с настоящим описанием определяются как условия, которые позволяют гибридизоваться последовательностям нуклеиновой кислоты по меньшей мере из 50 нуклеотидов, предпочтительно, примерно из 200 или более нуклеотидов, при температуре примерно 45°C в растворе, содержащем примерно 1М соли, предпочтительно, в растворе 6 × SSC или в любом другом растворе, обладающем сравнимой ионной силой, и при промывке при комнатной температуре в растворе, содержащем примерно 1М соли, предпочтительно, в растворе 6 × SSC или любом другом растворе, обладающем сравнимой ионной силой. Предпочтительно, гибридизацию проводят в течение ночи, то есть в течение по меньшей мере 10 часов, а промывку, предпочтительно, проводят в течение по меньшей мере одного часа с по меньшей мере двумя заменами промывочного раствора. Эти условия обычно позволяют достичь специфической гибридизации последовательностей, обладающих идентичностью последовательностей до 50%. Любой специалист в данной области может модифицировать описанные условия гибридизации для достижения специфической идентификации последовательностей, уровень идентичности которых изменяется от 50% до 90%.
Подробное описание изобретения
Настоящее изобретение относится к применению парвовирусов животных, в частности депендовирусов, таких как инфекционный AAV человека или обезьяны, и их компонентов (например, генома парвовируса животных) для применения в качестве векторов для введения и/или экспрессии нуклеиновых кислот в клетках млекопитающих. В частности, настоящее изобретение относится к повышению продуктивности таких парвовирусных векторов при образовании в клетках насекомых.
Вирусы семейства Parvoviridae представляют собой небольшие ДНК-содержащие вирусы животных. Семейство Parvoviridae может быть разделено на два подсемейства: Parvovirinae, представители которого инфицируют позвоночных животных, и Densovirinae, представители которого инфицируют насекомых. Представители подсемейства Parvovirinae в данном случае рассматриваются как парвовирусы и включают род Dependovirus. Как можно понять из названия этого рода, представители Dependovirus являются уникальными в том смысле, что для достижения продуктивной инфекции в клеточной культуре им обычно требуется одновременное инфицирование хелперным вирусом, таким как аденовирус или герпесвирус. Род Dependovirus включает AAV, который в норме инфицирует людей (например, серотипы 1, 2, 3А, 3B, 4, 5 и 6) или приматов (например, серотипы 1 и 4), а также родственные вирусы, которые инфицируют других теплокровных животных (например, коровий, собачий, лошадиный и овечий аденоассоциированные вирусы). Дополнительная информация по парвовирусам и другим представителям Parvoviridae описана в литературе (Kenneth I. Berns, "Parvoviridae: The Viruses and Their Replication", Chapter 69 in Fields Virology (3d Ed. 1996)). Для простоты настоящее изобретение далее описано только применительно к AAV. Однако следует понимать, что настоящее изобретение не ограничивается только AAV и может быть равным образом применимо к другим парвовирусам.
Геномная организация всех известных серотипов AAV очень сходна. Геном AAV представляет собой линейную одноцепочечную молекулу ДНК, которая содержит менее чем примерно 5000 нуклеотидов (нт) в длину. Инвертированные концевые повторы (ITR) фланкируют уникальные кодирующие нуклеотидные последовательности репликации неструктурных белков (Rep) и структурных белков (VP). Белки VP (VP1, -2 и -3) образуют капсид. Концевые 145 нуклеотидов являются самокомплементарными и организованы таким образом, что может быть сформирован энергетически стабильный внутримолекулярный дуплекс, образующий Т-образную шпилечную структуру. Такие шпилечные структуры функционируют как точки начала репликации ДНК вируса, являясь праймерами для клеточного ДНК-полимеразного комплекса. После инфекции клеток млекопитающих AAV дикого типа (wtAAV) гены Rep (например, Rep78 и Rep52) экспрессируются с помощью P5 промотора и P19 промотора, соответственно, и оба белка Rep выполняют определенную функцию в репликации генома вируса. Сплайсинг в открытой рамке считывания Rep (Rep ORF) приводит к экспрессии фактически четырех белков Rep (например, Rep78, Rep68, Rep52 и Rep40). Однако было показано, что несплайсированная мРНК, кодирующая белки Rep78 и Rep52, является достаточной для продукции вектора AAV в клетках млекопитающих. Аналогично, в клетках насекомых белки Rep78 и Rep52 являются достаточными для продукции вектора AAV.
Фраза «рекомбинантный парвовирусный или вектор AAV» (или «вектор rAAV») в контексте настоящего описания относится к вектору, содержащему одну или несколько интересующих полинуклеотидных последовательностей, интересующих генов или «трансгенов», которые фланкированы парвовирусными или инвертированными концевыми повторяющимися последовательностями (ITR) AAV. Такие векторы rAAV могут быть реплицированы и упакованы в инфекционные вирусные частицы, которые присутствуют в клетке-хозяине насекомого, экспрессирующей генные продукты rep и cap AAV (например, белки Rep и Cap). В том случае, если вектор rAAV встроен в более крупную конструкцию нуклеиновой кислоты (например, в хромосому или другой вектор, такой как плазмида или бакуловирус, используемый для клонирования или трансфекции), указанный вектор rAAV обычно рассматривается как «про-вектор», который может быть «незатронут» при репликации или инкапсидировании при наличии упаковочных функций и необходимых хелперных функций AAV.
В первом аспекте настоящее изобретение относится к нуклеотидной последовательности, содержащей открытую рамку считывания, которая содержит нуклеотидные последовательности, кодирующие белки Rep парвовирусов животных, где инициирующий кодон трансляции парвовирусного белка Rep78 представляет собой субоптимальный инициирующий кодон. Предпочтительно, субоптимальный инициирующий кодон представляет собой кодон инициации, который осуществляет частичный скиппинг экзонов. Частичный скиппинг экзонов в контексте настоящего описания означает, что по меньшей мере часть рибосом не начинает трансляцию при субоптимальном инициирующем кодоне белка Rep78, но при инициирующем кодоне, следующем по пути считывания информации, трансляция начинается, и в этой связи, предпочтительно, инициирующий кодон, следующий по пути считывания информации, представляет собой инициирующий кодон белка Rep52. Предпочтительно, субоптимальный инициирующий кодон влияет на частичный скиппинг экзона при экспрессии нуклеотидной последовательности в клетке насекомого. Предпочтительно, субоптимальный инициирующий кодон влияет на частичный скиппинг экзона в клетке насекомого, так что достигается продукция в клетке насекомого Rep78 и Rep52 в молярном соотношении в диапазоне от 1:10 до 10:1, от 1:5 до 5:1 или от 1:3 до 3:1, предпочтительно, примерно через 20-40 часов после инфекции, более предпочтительно, примерно через 30-40 часов после инфекции, используя систему экспрессии бакуловируса. Молярное соотношение Rep78 и Rep52 может быть определено вестерн-блоттингом, как описано в примере 1.1.3, предпочтительно, с использованием моноклональных антител, которые распознают общий эпитоп для Rep78 и Rep52, или с использованием антител, описанных в примере 1.1.3.
Термин «субоптимальный инициирующий кодон» в контексте настоящего описания относится не только к самому тринуклеотидному инициирующему кодону, но также к его соответствующему смысловому аналогу. Таким образом, субоптимальный инициирующий кодон может состоять из «оптимального» ATG кодона в субоптимальном контексте, то есть в контексте не-Козака. Однако более предпочтительными являются субоптимальные инициирующие кодоны, в случае которых тринуклеотидный инициирующий кодон сам является субоптимальным, то есть не является ATG. Термин «субоптимальный» в контексте настоящего описания означает, что такой кодон менее эффективен для инициации трансляции, тогда как в другом смысле он идентичен нормальному ATG кодону. Предпочтительно, эффективность субоптимального кодона составляет менее чем 90, 80, 60, 40 или 20% относительно эффективности нормального ATG кодона, тогда как в другом контексте они идентичны. Способы сравнения относительной эффективности инициации трансляции известны специалистам в данной области. Предпочтительные субоптимальные кодоны инициации могут быть выбраны из ACG, TTG, CTG и GTG. Более предпочтительным является ACG.
Под нуклеотидной последовательностью, которая кодирует белки Rep парвовирусов животных, в контексте настоящего изобретения следует понимать нуклеотидную последовательность, кодирующую неструктурные белки Rep, которые являются необходимыми и достаточными для продукции парвовирусного вектора в клетках насекомых, таких как белки Rep78 и Rep52. Нуклеотидную последовательность парвовирусов животных получают из депендовируса, более предпочтительно, из аденоассоциированного вируса человека или обезьяны (AAV) и, наиболее предпочтительно, из AAV, который обычно инфицирует людей (например, серотипы 1, 2, 3A, 3B, 4, 5 и 6) или приматов (например, серотипы 1 и 4). Пример нуклеотидной последовательности, кодирующей белки Rep парвовирусов животных, приведен в виде последовательности SEQ ID NO:10, которая представляет собой часть геномной последовательности AAV серотипа 2, кодирующей белки Rep. Кодирующая последовательность Rep78 включает нуклеотиды 11-1876, а кодирующая последовательность Rep52 включает нуклеотиды 683-1876. Следует понимать, что точный молекулярный вес белков Rep78 и Rep52, а также точное расположение инициирующих кодонов трансляции могут отличаться у разных парвовирусов. Однако специалистам в данной области известно, каким образом можно идентифицировать соответствующее положение в нуклеотидной последовательности из парвовирусов, отличных от AAV-2. Таким образом, нуклеотидная последовательность, кодирующая белки Rep парвовирусов животных, может быть определена как нуклеотидная последовательность:
a) которая кодирует полипептид, содержащий аминокислотную последовательность, которая идентична по меньшей мере на 50, 60, 70, 80, 88, 89, 90, 95, 97, 98 или 99% аминокислотной последовательности SEQ ID NO:11;
b) которая идентична по меньшей мере на 50, 60, 70, 80, 81, 82, 85, 90, 95, 97, 98 или 99% нуклеотидной последовательности на участке 11-1876 в SEQ ID NO:10;
c) комплементарная цепь которой гибридизуется с молекулой нуклеиновой кислоты, имеющей последовательность согласно пунктам (a) или (b);
d) нуклеотидные последовательности которой отличаются от последовательности молекулы нуклеиновой кислоты согласно пункту (c) благодаря вырожденности генетического кода.
Предпочтительно, нуклеотидная последовательность кодирует белки Rep парвовирусов животных, которые необходимы и достаточны для продукции парвовирусного вектора в клетках насекомых.
Другая предпочтительная нуклеотидная последовательность по настоящему изобретению содержит контрольную последовательность экспрессии, которая содержит девятинуклеотидную последовательность SEQ ID NO:7 или нуклеотидную последовательность, которая по существу гомологична SEQ ID NO:7, расположенную против направления считывания информации от инициирующего кодона в нуклеотидной последовательности, кодирующей парвовирусный белок Rep78. Последовательность, по существу идентичная нуклеотидной последовательности SEQ ID NO:7, которая будет способствовать повышению экспрессии парвовирусного белка Rep78, представляет собой, например, последовательность, которая по меньшей мере на 60%, 70%, 80% или 90% идентична девятинуклеотидной последовательности SEQ ID NO:7.
Потребность в удалении возможных ложных сайтов инициации трансляции в кодирующих последовательностях белка Rep, отличных от сайтов инициации трансляции Rep78 и Rep52, в других парвовирусах очевидна для специалистов в данной области, поскольку это будет устранение возможных сайтов сплайсинга, которые могут распознаваться в клетках насекомых. Различные модификации последовательностей парвовирусов дикого типа для соответствующей экспрессии в клетках насекомых получают при использовании известных методик генетического конструирования, таких как, например, описанные в работе Sambrook и Russell (2001) "Molecular Cloning: A Laboratory Manual (3rd edition), Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, New York. Специалистам в данной области хорошо известны другие модификации кодирующих участков белков Rep, с помощью которых можно повысить выход белка Rep. Такие модификации входят в объем настоящего изобретения.
В другом аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует белок Rep парвовируса, определенный выше. Предпочтительно, в указанной конструкции нуклеотидная последовательность, кодирующая белки Rep парвовируса, функционально связана с контрольными последовательностями экспрессии для экспрессии в клетке насекомого. Эти контрольные последовательности экспрессии включают по меньшей мере промотор, который активен к клетках насекомого. В практике осуществления настоящего изобретения могут быть использованы известные в данной области методики, которые применяются для экспрессии чужеродных генов в клетках-хозяевах насекомых. Стратегии молекулярного конструирования и экспрессии полипептидов в клетках насекомых описаны в литературе (см., например, Summers and Smith. 1986. A Manual of Methods for Baculovirus Vectors and Insect Culture Procedures, Texas Agricultural Experimental Station Bull. No. 7555, College Station, Tex.; Luckow. 1991. In Prokop et al., Cloning and Expression of Heterologous Genes in Insect Cells with Baculovirus Vectors' Recombinant DNA Technology and Applications, 97-152; King, L. A. and R. D. Possee, 1992, The baculovirus expression system, Chapman and Hall, United Kingdom; O'Reilly, D. R., L. K. Miller, V. A. Luckow, 1992, Baculovirus Expression Vectors: A Laboratory Manual, New York; W. H. Freeman and Richardson, С. D., 1995, Baculovirus Expression Protocols, Methods in Molecular Biology, volume 39; US 4,745,051; US2003148506 и WO 03/074714). Особенно подходящим промотором для транскрипции нуклеотидной последовательности по настоящему изобретению, которая кодирует белки Rep парвовируса, является, например, полиэдронный промотор. Однако в данной области известны также другие промоторы, которые активны в клетках насекомых, например промоторы p10, p35, IE-1 или ΔIE-1, а также другие промоторы, описанные в цитированных выше работах.
Предпочтительно, конструкция нуклеиновой кислоты для экспрессии белков Rep парвовируса в клетках насекомых представляет собой вектор, совместимый с клеткой насекомого. Под таким «вектором, совместимым с клеткой насекомого», или просто «вектором», следует понимать молекулу нуклеиновой кислоты, которая способна к трансформации или трансфекции насекомого или клетки насекомого. Примеры биологических векторов включают плазмиды, линейные молекулы нуклеиновой кислоты и рекомбинантные вирусы. При этом можно использовать любой вектор, если он совместим с клеткой насекомого. Такой вектор может быть интегрирован в геном клеток насекомого, но присутствие вектора в клетке насекомого необязательно должно быть постоянным, настоящее изобретение также включает временные эписомальные векторы. Указанные векторы могут быть введены любыми способами, например, путем химической обработки клеток, путем электропорации или инфекции. В предпочтительном варианте указанный вектор представляет собой бакуловирус, вирусный вектор или плазмиду. В более предпочтительном варианте указанный вектор представляет собой бакуловирус, например, указанная конструкция представляет собой бакуловирусный вектор. Бакуловирусные векторы и способы их использования описаны в цитированных выше работах по молекулярному конструированию клеток насекомых.
В другом аспекте настоящее изобретение относится к клетке насекомого, которая включает нуклеотидную последовательность не более одного типа, содержащую единственную открытую рамку считывания, которая кодирует белок Rep парвовируса. Предпочтительно, указанная единственная открытая рамка считывания кодирует один или несколько белков Rep парвовируса, более предпочтительно, указанная открытая рамка считывания кодирует все белки Rep парвовируса, наиболее предпочтительно, указанная открытая рамка считывания кодирует полноразмерный белок Rep 78, из которого в клетке насекомого предпочтительно экспрессируются по меньшей мере как белок Rep52, так и белок Rep78. Следует отметить, что клетка насекомого в контексте настоящего описания может включать более чем одну копию нуклеотидной последовательности одного типа, например, в составе мультикопийного эписомального вектора, и такие векторы представляют собой множественные копии по существу одной и той же молекулы нуклеиновой кислоты или по меньшей мере молекулы нуклеиновой кислоты, которая кодирует одну аминокислотную последовательность белка Rep, то есть молекулы нуклеиновой кислоты, которые незначительно отличаются друг от друга благодаря вырожденности генетического кода. Наличие только одного типа молекул нуклеиновой кислоты, кодирующих белки Rep парвовируса, препятствует рекомбинации гомологичных последовательностей, которая могла бы происходить в случае различных типов векторов, содержащих последовательности Rep, а это, в свою очередь, могло бы привести к образованию дефектных конструкций при экспрессии белка Rep и влиянию (в плане стабильности) на уровни продукции парвовирусов в клетках насекомых. Предпочтительно, нуклеотидная последовательность, которая находится в клетке насекомого и содержит единственную открытую рамку считывания, кодирующую один или несколько белков Rep парвовируса, представляет собой часть конструкции нуклеиновой кислоты, где указанная нуклеотидная последовательность функционально связана с контрольными последовательностями экспрессии для экспрессии в клетке насекомого. Другая предпочтительная клетка насекомого содержит, в качестве «первой» нуклеотидной последовательности, определенную выше нуклеотидную последовательность, которая кодирует белки Rep парвовируса, предпочтительно, кодирующую последовательность с субоптимальным инициирующим кодоном, определенную выше, или определенную выше конструкцию нуклеиновой кислоты, или клетка насекомого содержит, в качестве «первой» конструкции нуклеиновой кислоты, определенную выше конструкцию нуклеиновой кислоты, которая содержит такие нуклеотидные последовательности.
Любая клетка насекомого, в которой может реплицироваться рекомбинантный парвовирусный вектор (rAAV), и которая может поддерживаться в культуре, может быть использована по настоящему изобретению. Так, например, используемые клеточные линии могут быть получены из Spodoptera frugiperda, клеточных линий дрозофиллы, или клеточных линий комара, например клеточных линий, полученных из Aedes albopictus. Предпочтительные клетки насекомых или клеточные линии насекомых представляют собой клетки, полученные у вида насекомого, который является чувствительным к инфекции бакуловирусов, включая, например, Se301, SeIZD2109, SeUCR1, Sf9, Sf900+, Sf21, BTI-TN-5B1-4, MG-1, Tn368, HzAm1, Ha2302, Hz2E5, High Five (Invitrogen, CA, USA) и expresSF+® (US 6103526; Protein Sciences Corp., CT, USA).
Предпочтительная клетка насекомого по настоящему изобретению, кроме указанной выше «первой» нуклеотидной последовательности или конструкции нуклеиновой кислоты, содержит также:
a) вторую нуклеотидную последовательность, содержащую по меньшей мере одну нуклеотидную последовательность инвертированного концевого повтора (ITR) парвовируса; и
b) третью нуклеотидную последовательность, содержащую кодирующую последовательность белка Cap парвовируса, функционально связанную с контрольными последовательностями экспрессии для экспрессии в клетке насекомого.
В контексте настоящего описания под фразой «по меньшей мере одна нуклеотидная последовательность ITR парвовируса» следует понимать палиндромную последовательность, которая преимущественно содержит комплементарные, симметрично организованные последовательности, обозначаемые как участки "A", "B" и "C". ITR функционирует в качестве точки начала репликации, сайта, выполняющего «цис»-функцию в процессе репликации, то есть сайта распознавания трансактивных белков репликации, таких как, например, Rep78 (или Rep68), которые распознают палиндромные и специфические последовательности внутри палиндрома. Одним исключением в симметрии последовательности ITR является участок "D" в ITR. Он характеризуется уникальной структурой (не имеет комплемента внутри одной из последовательностей ITR). Точечный разрыв в одноцепочечной ДНК происходит в месте соединения участков A и D. Это тот участок, в котором происходит инициация синтеза ДНК. Участок D в норме находится на одной стороне палиндрома и определяет направленность нуклеиновой кислоты на стадии репликации. Парвовирус, реплицирующийся в клетке млекопитающего, как правило, содержит две последовательности ITR. Однако возможно сконструировать ITR таким образом, чтобы сайты связывания находились на обеих цепях участков A и D и располагались симметрично, то есть по одному на каждой стороне палиндрома. В случае двухцепочечной кольцевой ДНК-матрицы (например, в плазмиде), репликация нуклеиновой кислоты Rеp78 или Rеp68 происходит в двух направлениях и достаточно одной последовательности ITR для репликации кольцевого вектора парвовируса. Таким образом, в рамках настоящего изобретения можно использовать одну нуклеотидную последовательность ITR. Однако, предпочтительно, используют две или более упорядоченных ITR. Более предпочтительно, используют две последовательности ITR. Предпочтительной парвовирусной последовательностью ITR является ITR AAV. По соображениям безопасности может быть желательно сконструировать рекомбинантный парвовирусный вектор (rAAV), который не способен делиться после его введения в клетку. Такой механизм безопасности, направленный на ограничение нежелательного деления вектора в организме реципиента, может быть обеспечен за счет использования rAAV с химерной ITR, как описано в US2003148506.
Количество конструкций нуклеиновой кислоты, используемых в клетке насекомого для продукции рекомбинантного парвовирусного вектора (rAAV), в настоящем изобретении не ограничено. Например, можно использовать одну, две, три, четыре, пять или больше отдельных конструкций для продукции rAAV в клетках насекомых, в соответствии со способами по настоящему изобретению. Если используют пять конструкций, то одна конструкция кодирует VP1 AAV, другая конструкция кодирует VP2 AAV, еще одна конструкция кодирует VP3 AAV, дополнительная конструкция кодирует белок Rep, определенный выше, и последняя конструкция содержит по меньшей мере одну ITR AAV. Если используют меньше пяти конструкций, то указанные конструкции могут содержать различные сочетания по меньшей мере одной ITR AAV и VP1, VP2, VP3, а также кодирующие последовательности белка Rep. Предпочтительно, используют две конструкции или три конструкции, при этом, как описано выше, две конструкции являются более предпочтительными. Если используют две конструкции, то клетка насекомого предпочтительно содержит: (a) первую конструкцию нуклеиновой кислоты для экспрессии белков Rep, как описано выше, где указанная конструкция дополнительно содержит третью нуклеотидную последовательность, определенную выше в пункте (b) (которая содержит кодирующие последовательности белка Cap парвовируса, функционально связанные по меньшей мере с одной контрольной последовательностью экспрессии для экспрессии в клетке насекомого; см. также приведенное ниже описание); и (c) вторую конструкцию нуклеиновой кислоты, содержащую вторую нуклеотидную последовательность по пункту (a), выше, (которая содержит по меньшей мере одну нуклеотидную последовательность ITR AAV парвовируса). При использовании трех конструкций предпочтительно, чтобы две конструкции имели одинаковую конфигурацию, исключая, что используют отдельные конструкции для экспрессии капсидных белков и для экспрессии белков Rep. При этом, в каждой конструкции последовательности могут быть организованы в любом порядке относительно друг друга. Например, если одна конструкция содержит ITR и ORF, содержащую нуклеотидные последовательности, которые кодируют капсидные белки VP, то открытая рамка считывания (ORF) для VP может быть расположена на конструкции таким образом, что при репликации ДНК между последовательностями ITR указанная ORF для VP может реплицироваться или не реплицироваться. В качестве другого примера, следует указать, что кодирующие последовательности белков Rep и/или ORF, содержащие нуклеотидные последовательности, которые кодируют капсидные белки VP, могут находиться в конструкции в любом порядке. Следует понимать, что также вторая, третья и следующие конструкции нуклеиновой кислоты, предпочтительно, входят в состав векторов, совместимых с клетками насекомого, предпочтительно, в состав бакуловирусных векторов, описанных выше. Альтернативно, в клетке насекомого по настоящему изобретению одна или несколько последовательностей, выбранных из первой нуклеотидной последовательности, второй нуклеотидной последовательности, третьей нуклеотидной последовательности и четвертой нуклеотидной последовательности, а также необязательные другие нуклеотидные последовательности могут быть стабильно интегрированы в геном клетки насекомого. Любому специалисту в данной области известно, каким образом можно стабильно встроить нуклеотидную последовательность в геном насекомого и как идентифицировать клетку, содержащую такую нуклеотидную последовательность в своем геноме. Такое встраивание в геном может быть осуществлено с помощью, например, векторов, содержащих нуклеотидную последовательность с высокой гомологией к участкам генома насекомого. Использование специфических последовательностей, таких как транспозоны, представляет собой другой путь введения нуклеотидной последовательности в геном.
Согласно настоящему изобретению, третью нуклеотидную последовательность, содержащую кодирующую последовательность капсидного (Cap) белка парвовируса, следует понимать как последовательность, содержащую последовательности, кодирующие каждый из трех капсидных белков парвовируса, VP1, -2 и -3. Третья нуклеотидная последовательность, содержащая кодирующие последовательности капсидных белков, может присутствовать в разных формах. Так, например, могут использоваться отдельные кодирующие последовательности для каждого из капсидных белков VP1, -2 и -3, в соответствии с чем каждая кодирующая последовательность будет функционально связана с контрольными последовательностями экспрессии для экспрессии в клетке насекомого. Однако более предпочтительно, чтобы третья нуклеотидная последовательность содержала одну открытую рамку считывания, кодирующую все три капсидных белка VP1, VP2 и VP3 парвовирусов (AAV) животных, где инициирующий кодон трансляции капсидного белка VP1 представляет собой субоптимальный инициирующий кодон, который не является ATG, как, например, описано Urabe et al. (2002, supra). Субоптимальный инициирующий кодон капсидного белка VP1 может представлять собой кодон, определенный выше для белка Rep78. Более предпочтительные субоптимальные инициирующие кодоны капсидного белка VP1 могут быть выбраны из ACG, TTG, CTG и GTG, где CTG и GTG являются наиболее предпочтительными. Предпочтительная третья нуклеотидная последовательность для экспрессии капсидных белков дополнительно содержит контрольную последовательность экспрессии, содержащую девятинуклеотидную последовательность SEQ ID NO:7 или нуклеотидную последовательность, которая по существу гомологична SEQ ID NO:7, расположенную против направления считывания от инициирующего кодона нуклеотидной последовательности, кодирующей капсидный белок VP1. Последовательность, которая по существу идентична нуклеотидной последовательности SEQ ID NO:7 и которая позволяет повысить экспрессию VP1, представляет собой, например, последовательность, которая идентична по меньшей мере на 60%, 70%, 80% или 90% девятинуклеотидной последовательности SEQ ID NO:7. Предпочтительной третьей нуклеотидной последовательностью для экспрессии капсидных белков также является последовательность, которая, предпочтительно, включает по меньшей мере одну модификацию в нуклеотидной последовательности, кодирующей капсидный белок VP1, выбранную из С в нуклеотидном положении 12, А в нуклеотидном положении 21 и С в нуклеотидном положении 24 (где положение 1 представляет собой первый нуклеотид от кодона инициации трансляции; см. SEQ ID NO:1). Способы удаления возможных некорректных кодонов инициации трансляции VP1 и других серотипов хорошо известны специалистам в данной области, так же как и способы удаления возможных сайтов сплайсинга, которые могут распознаваться в клетках насекомых. Специалистам в данной области известны различные модификации кодирующих участков для VP, которые могут либо повышать выход VP и вириона, либо могут демонстрировать другие желательные эффекты, такие как измененный тропизм или сниженная антигенность вириона. Эти модификации также входят в объем настоящего изобретения. Предпочтительная нуклеотидная последовательность по настоящему изобретению, которая кодирует капсидные белки парвовирусов, функционально связана с контрольными последовательностями экспрессии для экспрессии в клетке насекомого и содержит по меньшей мере один промотор, который активен в клетках насекомого. Такие контрольные последовательности, а также различные методики и материалы (например, векторы), используемые для экспрессии капсидных белков парвовирусов в клетках-хозяевах насекомых, описаны выше применительно к белкам Rep.
В предпочтительном варианте осуществления настоящего изобретения вторая нуклеотидная последовательность присутствует в клетках насекомого по настоящему изобретению, например, последовательность, содержащая по меньшей мере одну парвовирусную (AAV) ITR, дополнительно содержит по меньшей мере одну нуклеотидную последовательность, кодирующую интересующий генный продукт, в соответствии с чем, предпочтительно, по меньшей мере одна нуклеотидная последовательность, кодирующая интересующий генный продукт, встраивается в геном рекомбинантного парвовирусного вектора (rAAV), продуцируемого в клетке насекомого. Предпочтительно, по меньшей мере одна нуклеотидная последовательность, кодирующая интересующий генный продукт, является последовательностью экспрессии в клетке млекопитающего. Предпочтительно, вторая нуклеотидная последовательность содержит две нуклеотидных последовательности ITR парвовируса (AAV), где по меньшей мере одна нуклеотидная последовательность, кодирующая интересующий генный продукт, расположена между двумя нуклеотидными последовательностями ITR парвовируса (AAV). Предпочтительно, указанная нуклеотидная последовательность, кодирующая генный продукт, представляющий интерес (для экспрессии в клетке млекопитающего), будет включена в рекомбинантный парвовирусный (rAAV) вектор, продуцируемый в клетке насекомого, если она расположена между двумя регулярными ITR или расположена на любом краю ITR, сконструированной с использованием двух участков D.
Таким образом, определенная выше вторая нуклеотидная последовательность может содержать нуклеотидную последовательность, кодирующую по меньшей мере один «генный продукт», представляющий интерес для экспрессии в клетке млекопитающего, расположенную таким образом, что она встраивается в рекомбинантный парвовирусный (rAAV) вектор, реплицирующийся в клетке насекомого. Любая нуклеотидная последовательность может быть встроена для последующей экспрессии в клетке млекопитающего, трансфицированной рекомбинантным парвовирусным (rAAV) вектором, получаемым по настоящему изобретению. Указанная нуклеотидная последовательность может, например, кодировать белок и может экспрессировать РНКи, то есть молекулу РНК, которая способна к РНК-интерференции, такую как, например, shRNA (кшРНК, короткая (малая) шпилечная РНК)) или siRNA (киРНК, короткая (малая) интерферирующая РНК)). Термин «киРНК» означает малую интерферирующую РНК, которая представляет собой двухцепочечную РНК короткой длины, которая не токсична для клеток млекопитающих (Elbashir et al., 2001, Nature 411: 494-98; Caplen et al., 2001, Proc. Natl. Acad. Sci. USA 98: 9742-47). В предпочтительном варианте вторая нуклеотидная последовательность может содержать две нуклеотидных последовательности, каждая из которых кодирует один генный продукт, представляющий интерес для экспрессии в клетке млекопитающего. Каждая из двух нуклеотидных последовательностей, кодирующих интересующий продукт, расположена таким образом, что она встраивается в рекомбинантный парвовирусный (rAAV) вектор, реплицирующийся в клетке насекомого.
Продукт, представляющий интерес для экспрессии в клетке млекопитающего, является терапевтическим генным продуктом. Терапевтический генный продукт может представлять собой полипептид или молекулу РНК (siRNA, киРНК), или другой генный продукт, который при экспрессии в клетке-мишени обеспечивает желательный терапевтический эффект, такой как, например, удаление нежелательной активности, например, удаление инфицированной клетки, или может восполнять генетический дефект, например, вызванный недостаточной ферментативной активностью. Примеры полипептидных генных продуктов с терапевтической активностью включают CFTR, фактор IX, липопротеинлипазу (LPL, предпочтительно, LPL S447X; см. WO 01/00220), аполипопротеин A1, уридиндифосфат-глюкуронозилтрансферазу (UGT), белок, взаимодействующий с регулятором ГТФазы из Retinitis Pigmentosa (RP-GRIP), и цитокины или интерлейкины, такие как, например, IL-10.
Альтернативно или дополнительно, используемая в качестве второго генного продукта вторая нуклеотидная последовательность в соответствии с описанием может содержать нуклеотидную последовательность, кодирующую полипептид, который служит в качестве маркерного белка для оценки клеточной трансформации и экспрессии. Подходящими маркерными белками для этой цели являются, например, флуоресцентный белок GFP и селектируемые маркерные гены тимидинкиназы HSV (для селекции на среде HAT), бактериальная гигромицин-В-фосфотрансфераза (для селекции на гигромицине В), аминогликозидфосфотрансфераза Tn5 (для селекции на G418) и дигидрофолатредуктаза (DHFR) (для селекции на метотрексате), CD20, ген фактора роста нервных клеток с низким сродством. Источники получения указанных маркерных генов и способы их использования описаны в руководстве Sambrook и Russel (2001) "Molecular Cloning: A Laboratory Manual (3rd edition), Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, New York. Дополнительно, вторая нуклеотидная последовательность, определенная выше, может содержать нуклеотидную последовательность, кодирующую полипептид, что может служить безопасным механизмом, позволяющим освобождать субъект от клеток, трансдуцированных рекомбинантным парвовирусным (rAAV) вектором по настоящему изобретению, если это представляется необходимым. Такая нуклеотидная последовательность, часто называемая суицидным геном, кодирует белок, который способен превращать пролекарство в токсическое вещество, которое может уничтожать трансгенные клетки, в которые экспрессируется белок. Соответствующие примеры таких суицидных генов включают, например, ген цитозиндезаминазы из E.coli или один из генов тимидинкиназы из вируса Herpes simplex, цитомегаловируса и вируса ветряной оспы, где ганцикловир может быть использован в качестве пролекарства для уничтожения трансгенных клеток у индивида (см., например, Clair et al., 1987, Antimicrob. Agents Chemother. 31; 844-849).
В другом варианте один из генных продуктов, представляющих интерес, может быть белком AAV. В частности, это может быть белок Rep, такой как Rep78 или Rep68, или их функциональный фрагмент. Нуклеотидная последовательность, кодирующая Rep78 и/или Rep68, в случае ее наличия в геноме рекомбинантного парвовирусного вектора (rAAV) по настоящему изобретению и при ее экспрессии в клетке млекопитающего, трансдуцированной этим вектором, позволяет достичь интеграции рекомбинантного парвовирусного вектора (rAAV) в геном трансдуцированной клетки млекопитающего. Экспрессия Rep78 и/или Rep68 в rAAV-трансдуцированной или инфицированной клетке млекопитающего может обеспечивать определенные преимущества для некоторых направлений использования рекомбинантного парвовирусного вектора (rAAV) за счет того, что позволяет достичь длительной или постоянной экспрессии любого другого интересующего генного продукта, который был введен в клетку с помощью вектора.
В составе рекомбинантных парвовирусных векторов (rAAV) по настоящему изобретению по меньшей мере одна из нуклеотидных последовательностей, кодирующих генный продукт, представляющий интерес для экспрессии в клетке млекопитающего, предпочтительно функционально связана, по меньшей мере, с одной контрольной последовательностью экспрессии, совместимой с клеткой млекопитающего, например с промотором. В данной области известно множество таких промоторов (см. Sambrook and Russel, 2001, supra). Могут быть использованы конститутивные промоторы, которые достаточно широко представлены и экспрессируются в клетках разных типов, такие как промотор CMV. Однако более предпочтительны промоторы, которые могут индуцироваться, например тканеспецифичные промоторы, промоторы, специфичные для определенного типа клеток или специфичные для определенного клеточного цикла. Например, промотор, специфичный для экспрессии в клетках печени, может быть выбран из α1-антитрипсинового промотора, промотора глобулина, связывающего тироидный гормон, промотора альбумина, промотора LPS (тироксин-связывающего глобулина), промотора гибрида HCR-ApoCII, промотора гибрида HCR-hAAT и промотора аполипопротеина Е. Другие примеры включают промотор E2F, селективный для опухолевых клеток и, в частности, селективный для экспрессии в опухолевой неврологической клетке (Parr et al., 1997, Nat. Med. 3:1145-9), или промотор IL-2, используемый в мононуклеарных клетках крови (Hagenbaugh et al., 1997, J Exp Med; 185:2101-10).
Известно, что AAV способен инфицировать различные клетки млекопитающих. См., например, Tratschin et al. (1985, Mol. Cell Biol. 5: 3251-3260) и Grimm et al. (1999, Hum. Gene Ther. 10:2445-2450). Однако трансдукция AAV в синовиальные фибробласты человека происходит значительно более эффективно, чем в аналогичные мышиные клетки (Jennings et al., Arthritis Res, 3:1 (2001)), и клеточная тропность AAV различается у разных серотипов. См., например, Davidson et al. (2000, Proc. Natl. Acad. Sci. USA, 97: 3428-3432), где обсуждаются различия, имеющиеся между AAV2, AAV4 и AAV5, по их тропности к клеткам ЦНС млекопитающих и с точки зрения эффективности трансдукции.
Последовательности AAV, которые могут быть использованы по настоящему изобретению для продукции рекомбинантных векторов AAV в клетках насекомого, могут быть получены из генома AAV любого серотипа. В основном серотипы AAV содержат геномные последовательности с выраженной степенью гомологии на уровне аминокислот и нуклеиновых кислот, обеспечивая идентичный набор генетических функций, продукцию вирионов, которые по существу физически и функционально эквивалентны и которые могут реплицироваться и подвергаться сборке за счет практически идентичных механизмов. Геномные последовательности различных серотипов AAV и обзор по геномному сходству указаны ниже (см., например, GenBank, номер доступа U89790; GenBank, номер доступа J01901; GenBank, номер доступа AF043303; GenBank, номер доступа AF085716; Chlorini et al. (1997, J. Vir. 71: 6823-33); Srivastava et al. (1983, J. Vir. 45: 555-64); Chlorini et al. (1999, J. Vir. 73: 1309-1319); Rutledge et al. (1998, J. Vir. 72: 309-319); и Wu et al. (2000, J. Vir. 74: 8635-47)). Серотипы AAV 1, 2, 3, 4 и 5 представляют собой предпочтительные источники нуклеотидных последовательностей AAV для их использования в рамках настоящего изобретения. Предпочтительно, последовательности ITR AAV, используемые в рамках настоящего изобретения, получают из AAV1, AAV2 и/или AAV4. Аналогично, последовательности, кодирующие белки Rep (Rep78 и Rеp52), предпочтительно получают из AAV1, AAV2 и/или AAV4. Однако последовательности, кодирующие капсидные белки VP1, VP2 и VP3, которые используются в рамках настоящего изобретения, могут быть взяты из числа любых 42 известных серотипов, более предпочтительно, из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 или AAV9, или из новых сконструированных AAV-подобных частиц, полученных, например, по методике перегруппировки капсида и с использованием капсидных библиотек AAV.
Последовательности Rep AAV и ITR характеризуются особенно выраженной консервативностью для большинства серотипов. Так, например, белки Rep78 у разных серотипов AAV идентичны более чем на 89%, и объединенная идентичность на уровне нуклеотидной последовательности генома для AAV2, AAV3A, AAV3B и AAV6 составляет примерно 82% (Bantel-Schaal et al., 1999, J. Virol., 73(2):939-947). Кроме того, как известно, последовательности Rep и ITR многих серотипов AAV подвергаются эффективной кросс-комплементации (то есть могут подвергаться функциональному замещению) с соответствующими последовательностями других серотипов в процессе образования частиц AAV в клетках млекопитающих. В документе US2003148506 сообщается, что Rep AAV и ITR последовательности также эффективно подвергаются кросс-комплементации с другими Rep AAV и ITR последовательностями в клетках насекомых.
Известно, что белки VP AAV определяют клеточную тропность вириона AAV. Последовательности, кодирующие белки VP, обладают значительно меньшей консервативностью, чем белки и гены Rep разных серотипов AAV. Способность Rep и ITR последовательностей к кросс-комплементации с соответствующими последовательностями других серотипов позволяет достичь продукции псевдотипных частиц rAAV, содержащих капсидные белки определенного серотипа (например, AAV3) и последовательности Rep и/или ITR другого серотипа AAV (например, AAV2). Такие псевдотипные частицы rAAV составляют часть настоящего изобретения.
Модифицированные последовательности «AAV» также могут использоваться в рамках настоящего изобретения, например, для продукции векторов rAAV в клетках насекомых. Такие модифицированные последовательности, которые содержат, например, последовательности, обладающие по меньшей мере примерно 70%, по меньшей мере примерно 75%, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 95% или большей идентичностью на уровне нуклеотидной и/или аминокислотной последовательностей (например, это может быть последовательность, обладающая примерно 75-99% идентичностью на уровне нуклеотидной последовательности) с AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 или AAV9 ITR, Rep или VP, могут использоваться вместо ITR, Rep или VP последовательностей AAV дикого типа.
AAV5, являясь по многим аспектам близким к другим серотипам AAV, отличается от других серотипов АAV человека и обезьяны в большей мере, чем другие известные серотипы человека и обезьяны. Ввиду этого, продукция rAAV5 может отличаться от продукции других серотипов в клетках насекомых. Если способы по настоящему изобретению используют для получения rAAV5, предпочтительно, чтобы одна или несколько конструкций включала в целом, в случае более чем одной конструкции, нуклеотидную последовательность, содержащую ITR AAV5, нуклеотидную последовательность, содержащую кодирующую последовательность Rep AAV5 (например, нуклеотидную последовательность для Rep 78 AAV5). Такие последовательности ITR и Rep могут быть модифицированы в нужном направлении с достижением эффективной продукции rAAV5 или псевдотипных векторов rAAV5 в клетках насекомых. Так, например, старт-кодон в последовательностях Rep может быть модифицирован, сайты сплайсинга VP могут быть модифицированы или устранены, и/или старт-кодон VP1 и окружающие нуклеотиды могут быть модифицированы для того, чтобы повысить продукцию векторов rAAV5 в клетке насекомого.
Таким образом, в другом аспекте настоящее изобретение относится к способу получения рекомбинантного парвовирусного вириона (rAAV) (включающего рекомбинантный парвовирусный вектор (rAAV), описанный выше) в клетке насекомого. Предпочтительно, этот способ включает стадии: (a) культивирования клетки насекомого согласно приведенному выше описанию в условиях, позволяющих достичь продукции рекомбинантного парвовирусного вектора (rAAV); и (b) выделения рекомбинантного парвовирусного вектора (rAAV). Следует понимать, что рекомбинантный парвовирусный вектор (rAAV), продуцируемый по настоящему способу, предпочтительно представляет собой инфекционный парвовирусный или AAV вирион, который содержит нуклеиновые кислоты рекомбинантного парвовирусного вектора (rAAV). Условия выращивания клеток насекомых в культуре и получение гетерологичных продуктов в культуре клеток насекомых известны в данной области и описаны, например, в цитированных выше работах, относящихся к молекулярно-генетическим методам конструирования клеток насекомых.
Предпочтительно, этот способ дополнительно включает стадию аффинной очистки (с включением вирионов) рекомбинантного парвовирусного вектора (rAAV) с использованием антитела против AAV, предпочтительно, с использованием иммобилизованного антитела. Такое антитело против AAV, предпочтительно, представляет собой моноклональное антитело. Особенно подходящим антителом является одноцепочечное верблюжье антитело или его фрагмент, получаемый, например, от верблюдов или лам (см, например, Muyldermans, 2001, Biotechnol. 74: 277-302). Антитело, используемое для аффинной очистки rAAV, предпочтительно, представляет собой антитело, которое специфически связывается с эпитопом на капсидном белке AAV, и при этом, предпочтительно, данный эпитоп представляет собой эпитоп, который присутствует на капсидном белке AAV более чем одного серотипа. Так, например, указанное антитело может быть образовано или отобрано на основании специфического связывания с капсидом AAV2, но, в то же самое время, оно может также специфически связываться с капсидами AAV1, AAV3 и AAV5.
В другом аспекте настоящее изобретение относится к вириону rAAV, получаемому в рамках описанных выше способов, с использованием конструкций нуклеиновой кислоты и определенных выше клеток. Предпочтительно, вирион rAAV содержит в своем геноме по меньшей мере одну нуклеотидную последовательность, кодирующую интересующий генный продукт, где по меньшей мере одна нуклеотидная последовательность не является нативной нуклеотидной последовательностью AAV и в стехиометрическом соотношении капсидных белков VP1, VP2 и VP3 из AAV количество VP1: (a) составляет по меньшей мере 100, 105, 110, 120, 150, 200 или 400% от количества VP2; или (b) составляет по меньшей мере 8, 10, 10,5, 11, 12, 15, 20 или 40% от количества VP3; или (c) составляет по меньшей мере количество, определенное в пунктах (a) и (b). Предпочтительно, количество VP1, VP2 и VP3 определяют с использованием антитела, распознающего эпитоп, которое является общим для VP1, VP2 и VP3. В данной области доступны различные виды иммунологического анализа, которые позволяют определять относительные количества VP1, VP2 и/или VP3 (см., например, Using Antibodies, E. Harlow and D. Lane, 1999, Cold Spring Harbor Laboratory Press, New York). Подходящее антитело, распознающее эпитоп, который является общим для каждого из трех капсидных белков, может представлять собой, например, мышиное антитело против Cap B1 (такое как антитело, коммерчески доступное от Progen, Germany). Предпочтительный вирион rAAV по настоящему изобретению представляет собой вирион, содержащий в своем геноме по меньшей мере одну нуклеотидную последовательность, кодирующую интересующий генный продукт, где по меньшей мере одна нуклеотидная последовательность не является нативной нуклеотидной последовательностью AAV, и вирион AAV включает капсидный белок VP1, содержащий лейцин или валин в положении 1 аминокислотной последовательности. Более предпочтительный вирион AAV по настоящему изобретению характеризуется определенным выше соотношением капсидных белков и включает капсидный белок VP1, содержащий лейцин или валин в положении 1 аминокислотной последовательности.
В контексте данного описания и в прилагаемой формуле изобретения глагол «включать» и его варианты используются в неограничительном смысле для обозначения того, что следующее за ним слово включается, но это не означает, что исключаются те термины, которые конкретно не были указаны. Дополнительно, ссылки на элемент, приведенный с неопределенным артиклем, не исключают возможности того, что присутствует более чем один такой элемент, если из контекста явно не следует, что в данном случае может быть один и только один из таких элементов. Неопределенный артикль в контексте данного описания обозначает вариант «по меньшей мере один».
Описание фигур
Фиг.1: A) Организация экспрессии Rep в геноме AAV дикого типа. Гены Rеp78 и Rep 52 экспрессируются с помощью промотора P5 и P19, соответственно. Экспрессия Rep68 и Rep40 (которые представляют собой сплайсинг-варианты Rep78 и Rep52, соответственно) не показана. Обе единицы экспрессии содержат сайт инициации ATG.
B) Конструкция по настоящему изобретению содержит открытую рамку считывания Rep (ORF REP), функционирующую под контролем одного промотора (например, полиэдронного промотора (PolH)). Этот промотор осуществляет экспрессию как Rep78, так и Rep52, поскольку инициирующий кодон ATG для Rep78 преобразован в альтернативный инициирующий кодон ACG и частично проскакивает по рибосоме.
C) Исходная конструкция, созданная Urabe et al. (2002, supra), функционирует при экспрессии Rep78 и Rеp52 независимо от двух других промоторов (ΔIE1 и polH, соответственно).
Фиг.2: Вестерн-блот анализ белков Rep, экспрессированных на основе рекомбинантного бакуловируса, который был подвергнут пяти пассажам в клетках насекомых. Исходная конструкция бакуловируса, полученная Urabe et al., 2002 (исходный REP/Bac-Bac), приводит к медленному снижению уровней экспрессии Rep78/52 в течение 5 пассажей. Единица экспрессии Rep78 и 52, сконструированная Urabe et al., 2002, и встроенная в бакуловирусный скелет PSC (исходный REP/PSC), также приводит к снижению уровня экспрессии Rep78/52 после пассажей в клетках насекомых. Однако бакуловирус с единицей экспрессии REP, содержащий инициирующий кодон ACG в скелете PSC (REP-ACG/PSC), характеризуется стабильной экспрессией Rep78/52 в течение по меньшей мере 5 пассажей. Был проведен вестерн-блот анализ по методике, описанной в примере 1.1.3.
Фиг.3: Результаты, приведенные в таблице 1, проиллюстрированы в виде графика.
Фиг.4: Сравнение уровней стабильности различных конструкций rAAV в клетках насекомых. Вектор rAAV в клетках SF+ получают по методике, описанной выше в примере 1. В случае всех вариантов продукции, ITR-содержащий бакуловирус и бакуловирус, содержащий капсидный ген, были идентичны, использовалось такое же число пассажей, как и в случае бакуловирусов, содержащих ген Rep. Были использованы бакуловирусы, содержащие 4 разных гена Rep: 1) REP-ACG/PSC; 2) SLR: исходная конструкция, созданная Urabe et al. (2002, supra); 3) Rep52 + Rep78(B2B): два отдельных Bac-Bac бакуловируса, где один из них содержит ген Rep78, а другой содержит ген Rep52; 4) Rep52 + Rep78(PSC): два отдельных бакуловирусных белка, где один бакуловирус содержит ген Rep78, а другой содержит ген Rep52.
Фиг.5: Стабильность конструкций на основе бакуловируса REP-ACG/PSC при проведении 8 пассажей. Для продукции rAAV в клетках SF+ используют методику, описанную в примере 1.
Фиг.6: Сравнение влияния проводимых пассажей на экспрессию белка rep в случае исходной конструкции, созданной Urabe et al. (2002, supra), и конструкции REP-ACG/PSC по настоящему изобретению. Пассажи для продукции бакуловируса и вестерн-блоттинг были выполнены по методике, описанной в примере 1. В рамках обычной методики пассажей rep-бакуловирусов образцы отбирают через 40 часов после добавления бакуловируса к клеткам SF и проводят вестерн-блоттинг.
Примеры
Пример 1: Rep-конструкции
1.1. Материалы и методы
1.1.1 Конструкция бакуловирусной плазмиды
Для экспрессии Rep78 и Rep52 на основе одной бицистронной матричной РНК, инициирующий кодон ATG в Rep78, расположенный на векторе экспрессии pFastBacDualSLR (Urabe et al., 2002, supra), преобразуют в ACG. Используемые против направления считывания информации праймеры представляют собой:
BamHI
5'-cgcggatcctgttaagACGGCGGGGTTTTACGAGATTGTGATTAAGGTC-3'
(SEQ ID NO:8)
ПРЯМАЯ ПРАЙМЕРНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
3'-Праймер, который использовали в реакции ПЦР, был фланкирован геном REP78 и содержит сайт XbaI (TCTAGA):
XbaI
5'-AGGCTCTAGATTCGAAAGCGGCCCG-3'
(SEQ ID NO:9)
ОБРАТНАЯ ПРАЙМЕРНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
Последовательность, расположенную между указанным выше праймерным набором, амплифицируют по методу ПЦР (объем реакционной смеси 50 мкл; 1 × Pfx амплифицирующий буфер; 0,3мМ dNTP (дНТФ); 1мM MgSO4, 150мM прямой праймер; 150мМ обратный праймер; 2× энхансерный раствор; матрица 50 нг (pFastBacDualSLR); 1 Ед. Platinum Pfx (Invitrogen, Carlsbad, CA, USA), используя следующий протокол: 1 цикл при 95°C, 5 минут; 35 циклов при 95°C, 15 секунд; 55°C, 30 секунд; 72°C, 2 минуты; 1 цикл при 72°C, 10 минут; 4°C в случае последующих циклов. Продукт ПЦР клонируют в PCR-blunt II-TOPO, используя набор для клонирования Zero Blunt TOPO PCR (Invitrogen). Rep78 подвергают субклонированию в pFastBacDual (Invitrogen) с использованием сайтов рестрикции SpeI и XbaI. В итоге клонируют мутированную кассету экспрессии Rep (с использованием ферментов рестрикции BstZ17I и AvrII) в конструкции экспрессии бакуловирусов (с открытыми концами после обработки рестриктазами EcoRV и XbaI) pPSCIO (Protein Sciences Corporation, Meriden, CT, USA). Анализ последовательности в полученной конструкции подтверждают по методике Baseclear, Leiden, the Netherlands.
1.1.2 Получение рекомбинантного бакуловируса
Рекомбинантные бакуловирусы из вируса нуклеарного полиэдроза Autographa californica (AcNPV) получают с использованием набора GeneXpress BaculoKIT (Protein Sciences Corporation). Трансфекцию проводят следующим образом: в круглодонных пробирках объемом 14 мл смешивают 200 мкл среды GRACE с 6 мкл селлфектина (Invitrogen), а в эппендорфовской пробирке 200 мкл среды GRACE смешивают с 50 мкл вирусной ДНК (для исследуемого белка) и 2 мкг трансферной плазмиды (REP). Содержимое эппендорфовской пробирки добавляют ко второй пробирке и осторожно перемешивают. После инкубации в течение 30 минут при комнатной температуре к смеси для трансфекции добавляют 1300 мкл GRACE. Клетки насекомых в колбе Т25 промывают средой GRACE и к клеточному слою добавляют по каплям трансфицирующую смесь. После инкубации в течение 6 часов при температуре 28°C осторожно добавляют сыворотку SF900II с добавкой 10% FBS и колбу Т25 помещают в сушильный шкаф с температурой 28°C на 5 дней, после чего собирают полученный рекомбинантный бакуловирус.
1.1.3 Вестерн-блот анализ
Клетки насекомых (SF+) инфицируют конструкцией бакуловирус-REP. Через 16, 40 и 64 часа после инфекции клеток отбирают образцы, проводят лизирование клеток добавлением 0,1V 10 × Tрис лизирующего буфера (1,5M NaCl, 0,5M TRIS, 0,01M MgCl2, 1% Tритон X-100, pH8,5, при стерилизации фильтрованием) и инкубируют при температуре 28°C в течение 30 минут на качалке (Innova 44, New Brunswick). Выделенные ДНК и РНК расщепляют путем инкубации с бензоназой при температуре 37°C в течение 30 минут. Клеточный лизат центрифугируют (1900×g; 15 минут; 4°C). К образцу супернатанта добавляют буфер для образца NuPAGE LDS (4×, Invitrogen) и наносят все на 4-12% бис-Трис гель (120V). Белки подвергают блоттингу на PVDF-мембране (BioRad) в течение 30 минут, 10V (Semidry blotting). Проводят иммунохимический анализ по методике вестерн-блоттинга при блокировании мембраны блокирующим буфером Superblock-PBS (PIERCE) и последующем инкубировании с анти-Rep мышиным антителом (303,9, Progen, Germany; разведение
1:50) и кроличьим противомышиным-HRP антителом (DAKO, разведение1:500). Rep-белки визуализируют путем хемилюминесцентного окрашивания с использованием люминесцентной лампы и субстрата для вестерн-блоттинга (Roche).
1.2 Результаты
Картину, полученную для созданной новой Rep-конструкции по настоящему изобретению (REP-ACG/PSC), сравнивают с картинами, характерными для исходных Rep-конструкций, в следующих вариантах: 1) бакуловирус с PSC скелетом и 2) бакуловирус с Bac-to-Bac скелетом (Urabе et al., 2002). Все три конструкции подвергают серийным пассажам до 5 пассажа. Эксперименты по продукции AAV1-LPL проводят с использованием Rep-конструкции после 2, 3, 4 и 5 пассажей в сочетании с AAV-LPL и AAV-Cap рекомбинантным бакуловирусом, отобранным после 2, 3, 4 и 5 пассажей, соответственно (использованные при этом рекомбинантные бакуловирусы AAV-LPL и AAV-Cap описаны ниже в примере 2). По методике количественной ПЦР определяют выход продукта AAV1-LPL, и полученные результаты показаны в таблице 1. Исходный бакуловирус, сконструированный Urabe et al., 2002 (исходный REP/Bac-to-Bac), приводит к быстрому снижению продукции AAV после 5 пассажей. Единица экспрессии Rep, сконструированная Urabe et al., 2002, и встроенная в PSC скелет бакуловируса (исходный REP/PSC), также приводит к снижению продукции AAV после пассажей в клетках насекомых. Однако бакуловирус с единицей экспрессии REP, содержащий в скелете PSC инициирующий кодон ACG (REP-ACG/PSC), характеризуется стабильной продукцией AAV в течение по меньшей мере 5 пассажей. В связи с этим, воспроизводимый выход продукта AAV-LPL в течение нескольких пассажей (например, от 2 до 5) получают лишь в случае использования бакуловирусов, содержащих конструкцию REP-ACG.
В таблице 2 показаны результаты количественного теста, проводимого по методике ПЦР (qPCR), который был разработан для оценки единицы Rep-экспрессии в рекомбинантных бакуловирусах и для фланкирования открытой рамки считывания (ORF) бакуловируса (генные копии на мл; гк/мл). Соотношение параметров в количественной ПЦР (Q-PCR) указывает на наличие делеций в Rеp-бакуловирусе. Коэффициент соотношения 1 теоретически означает, что все бакуловирусы в данной партии содержат рекомбинантную последовательность Rep78 или 52. Как видно для исходного бакуловируса, разработанного Urabe et al., 2002 (исходный REP/Bac-to-Bac), значительные количества рекомбинантного бакуловируса после 5 пассажа содержат делеции в последовательностях Rep. Единица экспрессии Rep78 и 52, разработанная Urabe et al., 2002, и встроенная в скелет PSC бакуловируса (исходный REP/PSC), демонстрирует очень быстрое и резкое снижение уровней рекомбинантного бакуловируса. Однако бакуловирус с единицей экспрессии REP, содержащей инициирующий кодон ACG в скелете PSC (REP-ACG/PSC) (клон C4 и A3), демонстрирует наличие стабильных рекомбинантных бакуловирусов после проведения по меньшей мере 5 пассажей.
Пример 2: Cap конструкции
2.1.1 Конструкция бакуловирусной плазмиды
Для экспрессии VP1,2,3 из одной полицистронной матричной РНК была разработана конструкция бакуловирус-AAV-Cap (Urabe et al., 2002, supra). В общих чертах, процедура состояла в том, что инициирующий кодон ATG в VP1 был мутирован до ACG. Потенциальный инициирующий кодон ATG в положении 11 был изменен на ACG. Акцепторный сайт сплайсинга по направлению считывания информации от инициирующего кодона VP1 был расщеплен (мутация в положениях 21 и 24). Мутированная кассета экспрессии для Cap была клонирована в конструкции экспрессии бакуловируса pFastBacDual (pFBDAAVlVPm11) с сайтами рестрикции BamHI/StuI. Указанная плазмида (pFBDAAVlVPm11) представляет собой исходный материал для встраивания альтернативных инициирующих кодонов для VP1. Для введения указанных мутаций Urabe et al. (2002, supra) использовали следующий прямой праймер:

Приведенные ниже прямые праймеры были использованы для создания конструкций экспрессии с использованием pFBDAAVlVPm11 (Urabe et al., 2002, supra) в качестве исходного материала:
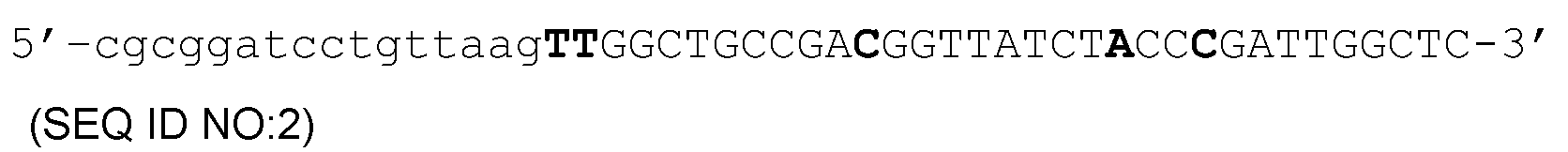



Обратный праймер, который был использован в реакциях ПЦР с указанными выше прямыми праймерами, направлен на положение, находящееся на расстоянии примерно 230 п.н. по направлению считывания информации от инициирующего кодона VP1, и содержит уникальный сайт для Stu I(AGGCCT).

Фрагменты амплифицируют с использованием указанных выше наборов прямых и обратных праймерных пар по методике ПЦР. После расщепления продуктов ПЦР с использованием BamHI и StuI ПЦР продукты субклонируют в сайтах BamHI/StuI pFBDAAVlvpm11 с получением различных тестируемых впоследствии конструкций бакуловирус-AAV-Cap. Идентичность полученных конструкций ДНК подтверждают анализом последовательности по Baseclear, Leiden, the Netherlands.
2.1.2 Получение рекомбинантного бакуловируса
Рекомбинантный бакуловирус, созданный на основе вируса нуклеарного полиэдроза Autographa californica (AcNPV), получают с использованием системы экспрессии бакуловируса Bac-to-Bac (Invitrogen). Полученный rBac-Cap амплифицируют путем инфекции 2×l06 клеток Sf9 на мл с показателем множественности заражения 0,1. Через три дня после инфекции клетки центрифугируют и оставляют супернатант, содержащий вирус.
2.1.3 Получение рекомбинантного AAV
Получают партии rAAV с использованием трех рекомбинантных бакуловирусов согласно методике, описанной Urabe et al., 2002. Однако в данном исследовании используют один бакуловирус, содержащий конструкцию экспрессии для LPLS447X-трансгена. Терапевтически активный агент, экспрессированный на основе трансгена, представляет собой природный вариант человеческой липопротеинлипазы, одноцепочечный полипептид из 448 аминокислот. LPLS447X содержит делецию из двух аминокислот на С-конце белка. Второй бакуловирус содержит конструкцию экспрессии для генов репликации AAV, Rep78 и Rep52. Tретий бакуловирус содержит последовательность капсида AAV1 с наличием ACG или TTG, CTG, GTG инициирующего кодона для VP1.
Партии rAAV млекопитающих получают с использованием системы плазмидной трансфекции согласно методике, описанной Grimm et al., 1998 (Novel tools for production and purification of recombinant adeno-associated virus vectors. Hum Gene Ther. 1998 Dec 10; 9(18):2745-60).
2.1.3 Вестерн-блот анализ
Клетки насекомых инфицируют конструкцией бакуловирус-Cap. Через три дня после инфекции клетки центрифугируют (3000 g; 15 минут). Супернатант фильтруют через фильтр Millex с размером пор 0,22 мкм. К образцу супернатанта добавляют буфер для образца NuPAGE LDS (Invitrogen) и наносят на 4-12% бис-Трис гель. Гель разгоняют в системе под напряжением 100В. Белки подвергают блоттингу на нитроцеллюлозной мембране (BioRad) в течение 1 часа, 100V, 350 мА. Иммунохимический анализ по методике вестерн-блоттинга проводят путем блокирования мембраны с использованием 1% марвела, высушенного обезжиренного молока, с последующей инкубацией с мышиным анти-Cap антителом (Bl от Progen, Germany; разбавление 1:50) и кроличьим противомышиным антителом-HRP (DAKO, разбавление 1:100). VP1, 2 и 3 визуализируют по наличию хемилюминесцентного окрашивания при использовании люминесцентной лампы и субстрата для вестерн-блоттинга (Roche).
2.1.4 Биохимические анализы
Активность человеческого LPLS447X тестируют, как было описано ранее, с использованием радиоактивного эмульсионного субстрата триолеоилглицерина (Nilsson-Ehle и Scholtz, 1976). Иммунореактивную массу человеческого LPLS447X оценивают методом сэндвич-ELISA с использованием куриного IgY и мышиного 5D2 анти-hLPL антител (Liu et al., 2000). Плазменные уровни триглицеридов определяют с использованием коммерческих наборов по протоколам производителя (Boehringer Mannheim, #450032).
2.2 Результаты
2.2.1 Конструкция рекомбинантного бакуловируса
С целью встраивания различных альтернативных инициирующих кодонов, для экспрессии VP1 в бакуловирусной плазмиде, разработанной Urabe et al. (2002 suprа), создают серию праймеров против направления считывания информации от сайта рестрикции BamHI и при наличии кодона TTG, ATT, GTG или CTG вместо инициирующего кодона ACG в VP1. ПЦР с использованием указанных праймеров в сочетании с праймером в направлении считывания информации от сайта StuI приводит к получению амплифицированных фрагментов, которые субклонируют в pFBDVPm11 с сайтом BamHI/StuI (Bac-Cap). Полученные при этом бакуловирусные плазмиды используют для получения рекомбинантных бакуловирусов с использованием бакуловирусной системы экспрессии Bac-to-Bac. Полученными рекомбинантными бакуловирусами инфицируют клетки насекомых с целью продукции капсидов AAV. Через три дня после инфекции методом вестерн-блоттинга определяют экспрессию вирусного белка в разных партиях бакуловируса. На основании результатов вестерн-блот анализа определяют, что бакуловирусная конструкция, содержащая инициирующий кодон TTG для VP1, экспрессирует данный белок на более высоком уровне по сравнению с ранее использованным инициирующим кодоном ACG. Было показано, что соотношение VP1 и VP2, в случае использования кодона TTG, составляет 1:1, что соответствует данным, полученным для AAV дикого типа (результаты не показаны).
2.2.2 Использование партий rAAV для инфекции культуры клеток
Для оценки инфективности капсидов AAV, полученных из рекомбинантных бакуловирусов с инициирующим кодоном TTG, создают rAAV. Получают также партию rAAV путем трансфекции плазмиды в клетки HEK293 млекопитающих. Методом количественной ПЦР определяют титр векторных геномов для обеих партий rAAV. Этот титр используют для инфекции клеток HEK293 в микротитрационном планшете с возрастающим показателем множественности заражения. Через два дня после инфекции в среде инфицированных клеток проводят тест (LPLS447X-масс-тест) для количественной оценки трансгенного продукта (LPLS447X). Результаты указанного теста показывают, что количество LPLS447X, продуцируемого бакуловирусным rAAV, аналогично количеству LPL, продуцируемого за счет плазмидного rAAV (данные не показаны).
2.2.3 Инъекция партий rAAV продукта мышам
Партии rAAV, полученные с использованием системы продукции на основе бакуловируса и с использованием традиционной системы продукции плазмиды в клетках млекопитающих, инъецируют внутримышечно мышам и затем определяют активность LPLS447X-белка и уровень триглицеридов в состоянии натощак in vivo. Через 3 дня, 7 дней и 2 недели после инъекции отбирают образцы крови и проводят анализ. В период между 3 и 7 днями после введения вируса образцы плазмы крови, взятые у мышей после инъекции rAAV, полученного в системе с использованием клеток млекопитающих, и у одной мыши, которой был инъецирован бакуловирус-rAAV, превращались из образцов молочного цвета в практически прозрачные. Образцы плазмы крови, полученные от одной мыши с инъецированной конструкцией бакуловирус-rAAV, все еще имели относительно молочный цвет, но уровень этого окрашивания явно снижался. Уровни триглицеридов (ТГ) снижались у всех обработанных мышей (данные не показаны). На 14 день уровни ТГ у мышей, которым вводили как AAV на основе клеток млекопитающего, так и бакуловирус-(TTG)-AAV, снижались на 96%. Образцы плазмы, взятые через 2 недели после введения вируса, выявили тот факт, что активность LPLS447X у мышей, которым вводили бакуловирус-AAV и AAV на основе клеток млекопитающего, были сходными (данные не показаны).
Пример 3: Стабильность конструкции rAAV с модифицированным инициирующим кодоном для Rep78 в клетках насекомых
3.1 Сравнение стабильности различных конструкций rAAV в клетках насекомых
Получение rAAV в клетках SF+ проводят по методике, описанной выше в примере 1. В случае всех вариантов получения, ITR-содержащий бакуловирус и бакуловирус, содержащий капсидный ген, были идентичны, число пассажей было таким же, как и в случае бакуловирусов, содержащих ген Rep. Были использованы бакуловирусы, содержащие 4 разных гена Rep: 1) REP-ACG/PSC; 2) SLR: исходная конструкция, описанная Urabe et al. (2002, supra); 3) Rep52 + Rep78(B2B): два отдельных бакуловируса Bac-to-Bac, где один из них содержит ген Rep78, а другой содержит ген Rep52; 4) Rep52 + Rep78(PSC): два отдельных бакуловирусных источника белка, где один содержит ген Rep78, а другой содержит ген Rep52.
Полученные результаты показаны на фиг.4. При пятом пассаже бакуловируса продукция rAAV повышается более чем в 10 раз, при использовании REP-ACG/PSC по настоящему изобретению, по сравнению с исходной конструкцией Rep и по сравнению с расщепленными конструкциями Rep.
3.2 Стабильность бакуловирусных конструкций при проведении 8 пассажей
Получают rAAV в клетках SF+ по методике, описанной в примере 1. В случае всех вариантов получения, ITR-содержащий бакуловирус и бакуловирус, содержащий капсидный ген, были идентичны, число пассажей было таким же, как в случае бакуловируса REP-ACG/PSC. Полученные результаты показаны на фиг.5. Бакуловирус REP-ACG/PSC сохраняет стабильность по меньшей мере до восьмого пассажа. Титры продукта rAAV в случае REP-ACG/PSC остаются стабильными по меньшей мере до 8 пассажа бакуловируса.
3.3 Влияние числа пассажей на экспрессию белка rep
Влияние количества пассажей на экспрессию белка Rep в случае исходной конструкции, созданной Urabe et al. (2002, supra), сравнивают с вариантом конструкции REP-ACG/PSC по настоящему изобретению. Пассажи бакуловируса и вестерн-блоттинг проводят по методике, описанной в примере 1. В ходе обычной процедуры пассажей Rep-бакуловирусов, образцы отбирают через 40 часов после добавления бакуловирусов к клеткам SF и проводят вестерн-блоттинг. На фиг.6 показано явное снижение экспрессии белка Rep при увеличении числа пассажей, по сравнению с более ранними вариантами пассажей применительно к исходной конструкции Urabe (SLR), тогда как экспрессия белка Rep в случае конструкции REP-ACG/PSC остается такой же при увеличении числа пассажей, как и при более ранних пассажах.
Реферат
Изобретение относится к области биотехнологии, генной инженерии и вирусологии. Изобретение относится к конструкциям нуклеиновой кислоты, используемым для получения рекомбинантных парвовирусных векторов в клетках насекомых, к клеткам насекомых, включающим такие конструкции, и к способам, в которых клетки используют для получения рекомбинантных парвовирусных вирионов. Клетки насекомых предпочтительно содержат первую нуклеотидную последовательность, кодирующую парвовирусные белки Rep, где инициирующий кодон трансляции парвовирусного белка Rep78 представляет собой субоптимальный инициирующий кодон, который влияет на частичный скиппинг экзона при экспрессии в клетках насекомых. Клетка насекомого дополнительно содержит вторую нуклеотидную последовательность, содержащую нуклеотидную последовательность, по меньшей мере с одним парвовирусным (AAV) инвертируемым концевым повтором (ITR), и третью нуклеотидную последовательность, содержащую последовательности, кодирующие капсидные белки парвовируса. 7 н. и 23 з.п. ф-лы, 6 ил., 2 табл., 3 пр.
Формула
a) вторую нуклеотидную последовательность, содержащую по меньшей мере одну нуклеотидную последовательность с инвертированным концевым повтором (ITR) парвовируса; и
b) третью нуклеотидную последовательность, содержащую последовательности, кодирующие капсидные белки парвовируса, функционально связанную с последовательностями, контролирующими экспрессию, для экспрессии в клетке насекомого.
a) первую конструкцию нуклеиновой кислоты, в которой нуклеотидная последовательность является частью конструкции нуклеиновой кислоты и функционально связана с последовательностями, контролирующими экспрессию, для экспрессии в клетках насекомых;
b) вторая конструкция нуклеиновой кислоты, содержащая по меньшей мере одну нуклеотидную последовательность инвертированного концевого повтора (ITR) парвовируса; и
c) третью конструкцию нуклеиновой кислоты, содержащую последовательности, кодирующие капсидные белки парвовируса, функционально связанные с последовательностями, контролирующими экспрессию, для экспрессии в клетке насекомого,
где первая, вторая и третья конструкция нуклеиновой кислоты предпочтительно является вектором, совместимым с клеткой насекомого, предпочтительно бакуловирусным вектором.
a. культивирования клетки насекомого по любому из пп.11-18 в условиях, при которых возможно получение рекомбинантного парвовирусного вириона; и
b. выделения рекомбинантного парвовирусного вириона.
a) культивирования клетки насекомого по любому из пп.11-18 в условиях, при которых возможно получение рекомбинантного парвовирусного вириона; и
b) выделения рекомбинантного парвовирусного вириона.
Комментарии