Гидролазы, кодирующие их нуклеиновые кислоты и способы их получения и применения - RU2525675C2
Код документа: RU2525675C2
Чертежи
















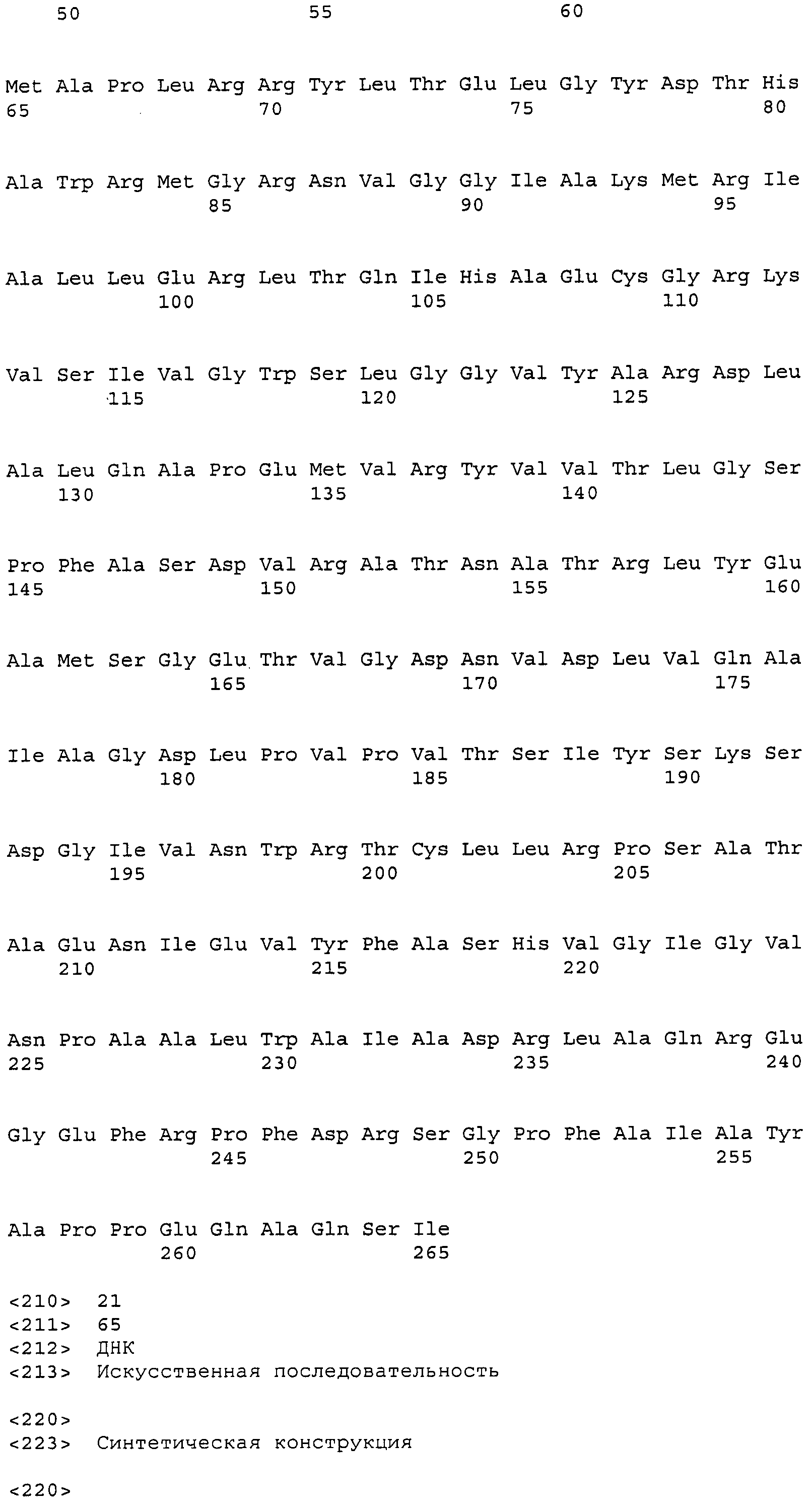


























Описание
ПЕРЕКРЕСТНАЯ ССЫЛКА НА СВЯЗАННУЮ ЗАЯВКУ
По настоящей заявке испрашивается приоритет по патентной заявке США № 12/202204, поданной 29 августа 2008 года, которая, таким образом, включена в полном объеме в настоящее описание в качестве ссылки.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка подана со списком последовательностей, представленным в виде файла с именем 011631-0045-228_SeqListing.txt размером 46086 байтов, который создан 25 августа 2009 года. Список последовательностей включен в настоящую заявку в качестве ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к полипептидам, которые обладают гидролазной активностью, включая липазную, сатуразную, пальмитазную и/или стеаратазную активность, кодирующим их полинуклеотидам и способам получения и применения этих полинуклеотидов и полипептидов. Также изобретение относится к пептидам и полипептидам, например ферментам, обладающим гидролазной активностью, например липазам, сатуразам, пальмитазам и/или стеаратазам, и способам обработки жиров и масел с использованием таких пептидов и полипептидов для получения продуктов гидролиза масел, таких как животные или растительные масла с низким содержанием насыщенных жирных кислот, например соевое масло или масло канолы, продуктов такой обработки масел и продуктов, содержащих такие обработанные масла.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
В основном гидролазы, например липазы, сатуразы, пальмитазы и/или стеаратазы, используют в промышленности, включая производство напитков и продуктов питания, в качестве ингибиторов черствения для хлебобулочных изделий, а также в производстве маргарина и других пастообразных смесей с натуральными ароматизаторами масла; в системах удаления отходов; и в фармацевтической промышленности, где их используют в качестве средств, способствующих пищеварению.
Переработанные масла и жиры представляют собой основные компоненты продуктов питания, пищевые добавки и пищевые технологические добавки, а также важное возобновляемое сырье для химической промышленности. Они доступны в больших количествах в результате переработки семян масличных культур, например, рисовой мучки, кукурузы, рапса, канолы, подсолнечника, оливы, пальмы или сои. Другие источники ценных масел и жиров включают рыбу, отходы ресторанов и топленые животные жиры. Эти жиры и масла представляют собой смеси триацилглицеридов или липидов, т.е. жирные кислоты (ЖК), этерифицированные на глицериновом остове. Каждое масло или жир содержит широкий спектр липидов с различной структурой, характеризующийся содержанием ЖК и их региохимического распределения на глицериновом остове. Эти свойства отдельных липидов определяют физические свойства чистого триацилглицерида. Таким образом, содержание триацилглицерида в жире или масле в большой степени определяет физические, химические и биологические свойства масла. Ценность липидов значительно возрастает в зависимости от их чистоты. Высокой чистоты можно добиться фракционной хроматографией или дистилляцией, отделив желаемый триацилглицерид от фоновой смеси источника жира или масла. Однако это имеет высокую стоимость, а выход часто ограничен тем, что в природе триацилглицерид встречается в низкой концентрации. Кроме того, очистка продукта часто осложняется присутствием множества структурно и физически или химически схожих триацилглицеридов в масле.
Альтернативой очистки триацилглицеридов или других липидов из природного источника является синтез липидов. Продукты таких процессов называют структурированными липидами, поскольку они содержат определенный набор жирных кислот, расположенных определенным образом на глицериновом остове. Также ценность липидов значительно возрастает за счет управления содержанием и распределением жирных кислот в липиде. Устранение из триглицеридов, жиров или масел нежелательных ЖК или замена ЖК с нежелательными свойствами на жирные кислоты с более хорошими или более желательными химическими, физическими или биологическими свойствами увеличивает ценность липидов. В частности, существует потребность в липазах, которые могут отщеплять посредством гидролиза, например, избирательно отщеплять посредством гидролиза, насыщенную жирную кислоту («сатураза»), или в липазах, которые, частности, могут отщеплять посредством гидролиза, например, избирательно отщеплять посредством гидролиза, пальмитиновую кислоту («пальмитаза») или стеариновую кислоту («стеаратаза») от глицеринового остова. Липазы, такие как сатуразы, например пальмитазы и/или стеаратазы, можно использовать для осуществления такого контроля, где удаляемые, добавляемые или замещаемые ЖК представляют собой насыщенные жирные кислоты, например пальмитиновую кислоту или стеариновую кислоту.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к полипептидам, которые обладают гидролазной активностью, включая липазную активность. В одном из аспектов изобретение относится к новым классам липаз, названным «сатуразы», «пальмитазы» и «стеаратазы». Также изобретение относится к полинуклеотидам, которые кодируют полипептиды, обладающие сатуразной, например пальмитазной и/или стеаратазной, активностью, и к способам получения и применения этих полинуклеотидов и полипептидов. В одном из аспектов изобретение относится к полипептидам, например ферментам, которые обладают гидролазной активностью, например липазной, сатуразной, пальмитазной и/или стеаратазной активностью и термостабильной и/или термостабильной ферментативной (каталитической) активностью. Ферментативные активности полипептидов и пептидов по изобретению включают (содержат или состоят из) сатуразную активность или липазную активность, включая гидролиз липидов, реакции ацидолиза (например, для замещения этерифицированной жирной кислоты свободной жирной кислотой), реакции трансэтерификации (например, обмен жирными кислотами между триацилглицеридами), синтез сложных эфиров, реакции переэтерификации и активность ацилгидролаз липидов (АГЛ). В другом аспекте полипептиды по изобретению используют для синтеза энантиомерно чистых хиральных продуктов.
Полипептиды по изобретению можно использовать в различных фармацевтических, сельскохозяйственных и промышленных ситуациях, включая производство косметических средств и нутрицевтиков. Дополнительно полипептиды по изобретению можно использовать в обработке пищевых продуктов, пивоварении, добавках для ванн, производстве спирта, синтезе пептидов, энантиоселективности, изготовлении шкур в кожевенной промышленности, обработке отходов и расщеплении животных отходов, восстановлении серебра в фотопромышленности, медицинской помощи, дегуммировании шелка, разложении биопленок, превращении биомассы в этанол, биозащите, противомикробных средствах и дезинфицирующих средствах, личной гигиене и косметических средствах, биотехнологических реактивах, в повышении выхода крахмала из зерна влажного помола и в качестве фармацевтических препаратов, таких как способствующие пищеварению средства и противовоспалительные средства.
В некоторых вариантах осуществления изобретение относится к композициям (например, липазы, сатуразы, пальмитазы и/или стеаратазы) и способам получения масел с низким содержанием насыщенных жирных кислот, например масел с низким содержанием насыщенных жирных кислот, содержащих масла с низким содержанием пальмитиновой, стеариновой, миристиновой, лауриновой или масляной жирной кислоты и/или каприловой кислоты (октановой кислоты). Любое растительное масло, например, масло канолы, соевое масло или животное масло или жир, например сало, можно обработать с использованием композиции или посредством способа по изобретению. Любые пищевые продукты, съестные продукты или печеные, жареные или термически обработанные продукты (например, соусы, маринады, приправы, масла для смазывания разбрызгиванием, маргарины, пекарные масла, майонез, кулинарные масла, масла для салата, заправки, которые можно черпать ложкой и наливать, и т.п., и продукты, полученные с их использованием) могут содержать растительное масло или животный жир, которые обработаны с использованием композиции или способа по изобретению. Модифицированные растительные масла со сниженным содержанием насыщенных жирных кислот можно использовать в любых продуктах питания, съестных продуктах или печеных или термических обработанных продуктах, например в соусах, маринадах, приправах, маслах для смазывания разбрызгиванием, маргаринах, пекарных маслах, майонезе, кулинарных маслах, маслах для салата, заправках, которые можно черпать ложкой и наливать и т.п. В одном из вариантов осуществления изобретение относится к маслам, таким как растительные масла, например масло канолы или соевое масло, и продукты питания или печеные или термически обработанные продукты, включая соусы, маринады, приправы, масла для смазывания разбрызгиванием, маргарины, майонез, пекарные масла, кулинарные масла, масла для жарки, масла для салата, заправки, которые можно черпать ложкой и наливать, и т.п., где масло или продукт питания, печеный или термически обработанный продукт модифицирован с использованием фермента по изобретению. В одном из аспектов эти растительные масла, например масло канолы, касторовое масло, кокосовое масло, кориандровое масло, кукурузное масло, хлопковое масло, масло лесного ореха, конопляное масло, льняное масло, масло пенника лугового, оливковое масло, пальмовое масло, косточковое пальмовое масло, арахисовое масло, рапсовое масло, рисовое масло, сафлоровое масло, масло сасанквы, соевое масло, масло подсолнечника, талловое масло, масло цубаки, различные обладающие измененным составом жирных кислот «натуральные» масла за счет генетически модифицированных организмов (ГМО) или традиционной «селекции», такие как масла с высоким содержанием олеиновой кислоты, с низким содержанием линоленовой кислоты или с низким содержанием насыщенных жирных кислот (масло канолы с высоким содержанием олеиновой кислоты, соевое масло с низким содержанием линоленовой кислоты или масла подсолнечника с высоким содержанием стеариновой кислоты), животные жиры (сало, лярд, жир коровьего масла и куриный жир), рыбий жир (жир тихоокеанского талеихта, жир печени трески, жир хоплостета, жир сардины, жир сельди и жир менхадена) или смеси любых указанных выше компонентов и продукты питания или печеные, жареные или термически обработанные продукты содержат масла со сниженным содержанием насыщенных жирных кислот, включая масла с низким содержанием пальмитиновой кислоты, миристиновой кислоты, лауриновой кислоты, стеариновой кислоты, каприловой кислоты (октановой кислоты) и т.д., обработанные посредством использования композиции или способа по изобретению.
В одном из аспектов изобретение относится к полипептидам, например, ферментам и каталитическим антителам, которые обладают гидролазной активностью, например липазной, сатуразной, пальмитазной и/или стеаратазной активностью, включая термостабильные и термостабильные ферментативные активности, и активностями со специфичностью к жирным кислотам или избирательностью к жирным кислотам, и ферментативными активностями с высокой или низкой устойчивостью к pH, и к полинуклеотидам, кодирующим эти полипептиды, включая векторы, клетки-хозяева, трансгенные растения и животные, не относящиеся к человеку, и к способам получения и применения этих полинуклеотидов и полипептидов.
В другом аспекте изобретение относится к выделенным, синтетическим или рекомбинантным нуклеиновым кислотам, которые содержат:
(a) нуклеиновую кислоту (полинуклеотид), которая кодирует по меньшей мере один полипептид, где нуклеиновая кислота содержит последовательность, которая обладает по меньшей мере приблизительно 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более или полной (100%) идентичностью последовательностей с:
(i) SEQ ID № 1, SEQ ID № 3, SEQ ID № 5, SEQ ID № 7, SEQ ID № 9, SEQ ID № 11, SEQ ID № 13, SEQ ID № 15, SEQ ID № 17, SEQ ID № 19, SEQ ID № 22 или SEQ ID № 23, или
(ii) нуклеиновой кислотой SEQ ID № 1, которая содержит одну или несколько нуклеотидных замен (или их эквиваленты), кодирующих одну, две, три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать, двенадцать, тринадцать, четырнадцать, пятнадцать, шестнадцать, семнадцать, восемнадцать, девятнадцать, двадцать, двадцать одну, двадцать две, двадцать три, двадцать четыре, двадцать пять или более или все аминокислотные замены (или их эквиваленты), как указано в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23,
где нуклеиновая кислота по п. (i) или (ii) кодирует по меньшей мере один полипептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, или кодирует полипептид или пептид, позволяющий создать специфическое антитело к гидролазе (например, к липазе, сатуразе, пальмитазе и/или стеаратазе) (полипептид или пептид, который выполняет функцию эпитопа или иммуногена),
(b) нуклеиновую кислоту (полинуклеотид) по п. (a), где идентичность последовательностей определяют: (a) посредством анализа с использованием алгоритма сравнения последовательностей или посредством визуального исследования, или (b) на участке длиной по меньшей мере приблизительно 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 100, 125, 150, 175, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300, 1350, 1400, 1450, 1500, 1550 или более остатков или по всей длине кДНК, транскрипта (мРНК) или гена,
(c) нуклеиновую кислоту (полипептид) по п. (a) или (b), где алгоритмом сравнения последовательностей является алгоритм BLAST версии 2.2.2, где установлены следующие настройки фильтрации blastall -p blastp -d «nr pataa» -F F, а все остальные опции имеют значения по умолчанию,
(d) нуклеиновую кислоту (полинуклеотид), кодирующую по меньшей мере один полипептид или пептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, где нуклеиновая кислота содержит последовательность, которая гибридизуется в строгих условиях с комплементарной последовательностью нуклеиновой кислоты по п. (a), (b) или (c), где строгие условия содержат стадию отмывания, которая включает отмывание в 0,2×SSC при температуре приблизительно 65°C в течение приблизительно 15 минут,
(e) нуклеиновую кислоту (полинуклеотид), кодирующую по меньшей мере один полипептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, где полипептид содержит последовательность SEQ ID № 2 или ее ферментативно активные фрагменты, которая содержит по меньшей мере одну, две, три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать, двенадцать, тринадцать, четырнадцать, пятнадцать, шестнадцать, семнадцать, восемнадцать, девятнадцать, двадцать, двадцать одну, двадцать две, двадцать три, двадцать четыре или более или все аминокислотные замены (или их эквиваленты), как указано в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23,
(f) нуклеиновую кислоту (полинуклеотид), кодирующую по меньшей мере один полипептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, где полипептид содержит последовательность SEQ ID № 2, SEQ ID № 4, SEQ ID № 6, SEQ ID № 8, SEQ ID № 10, SEQ ID № 12, SEQ ID № 14, SEQ ID № 16, SEQ ID № 18 или SEQ ID № 20 или ее ферментативно активные фрагменты,
(g) (a) нуклеиновую кислоту (полинуклеотид) по любому из п.п. с (a) до (f), кодирующую полипептид, содержащий по меньшей мере одну консервативную аминокислотную замену и сохраняющий свою гидролазную активность, например, липазную, сатуразную, пальмитазную и/или стеаратазную активность, или, (b) нуклеиновую кислоту по п. (g)(a), где по меньшей мере одна консервативная аминокислотная замена включает замену аминокислоты на другую аминокислоту с похожими характеристиками; или консервативная замена включает: замену алифатической аминокислоты на другую алифатическую аминокислоту; замену серина на треонин или наоборот; замену кислотного остатка на другой кислотный остаток; замену остатка, несущего аминогруппу, на другой остаток, несущий аминогруппу; замену основного остатка другим основным остатком; или замену ароматического остатка другим ароматическим остатком,
(h) нуклеиновую кислоту (полинуклеотид) по любому из п.п. с (a) до (g), кодирующую полипептид, который обладает гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, но не содержит сигнальную последовательность,
(i) нуклеиновую кислоту (полинуклеотид) по любому из п.п. с (a) до (h), кодирующую полипептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, который дополнительно содержит гетерологичную последовательность,
(j) нуклеиновую кислоту (полинуклеотид) по п. (i), где гетерологичная последовательность состоит из или содержит последовательность, кодирующую: (a) гетерологичную сигнальную последовательность, (b) последовательность по п. (a), где гетерологичную сигнальную последовательность получают из гетерологичного фермента, или, (c) метку, эпитоп, таргетинг-пептид, расщепляемую последовательность, поддающийся обнаружению фрагмент или участок, или
(k) последовательность нуклеиновой кислоты (полинуклеотид), полностью (абсолютно) комплементарную последовательности по любому п.п. с (a) до (j).
В одном из аспектов выделенная, синтетическая или рекомбинантная нуклеиновая кислота кодирует полипептид или пептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, которая является термостабильной. Полипептиды и пептиды, кодируемые нуклеиновыми кислотами по изобретению, или любой полипептид или пептид по изобретению, может сохранять ферментативную или связывающую активность (например, связывание с субстратом) в условиях, которые включают температурный диапазон приблизительно от -100°C приблизительно до -80°C, приблизительно от -80°C приблизительно до -40°C, приблизительно от -40°C приблизительно до -20°C, приблизительно от -20°C приблизительно до 0°C, приблизительно от 0°C приблизительно до 5°C, приблизительно от 5°C приблизительно до 15°C, приблизительно от 15°C приблизительно до 25°C, приблизительно от 25°C приблизительно до 37°C, приблизительно от 37°C приблизительно до 45°C, приблизительно от 45°C приблизительно до 55°C, приблизительно от 55°C приблизительно до 70°C, приблизительно от 70°C приблизительно до 75°C, приблизительно от 75°C приблизительно до 85°C, приблизительно от 85°C приблизительно до 90°C, приблизительно от 90°C приблизительно до 95°C, приблизительно от 95°C приблизительно до 100°C, приблизительно от 100°C приблизительно до 105°C, 5 приблизительно 105°C приблизительно до 110°C, приблизительно от 110°C приблизительно до 120°C, или 95°C, 96°C, 97°C, 98°C, 99°C, 100°C, 101 °C, 102°C, 103°C, 104°C, 105°C, 106°C, 107°C, 108°C, 109°C, 110°C, 111°C, 112°C, 113°C, 114°C, 115°C или более. Изобретение относится к термостабильным полипептидам, которые сохраняют гидролазную активность, например, липазную, сатуразную, пальмитазную и/или стеаратазную активность, при температуре в описанных выше диапазонах, приблизительно при pH 3,0, приблизительно при pH 3,5, приблизительно при pH 4,0, приблизительно при pH 4,5, приблизительно при pH 5,0, приблизительно при pH 5,5, приблизительно при pH 6,0, приблизительно при pH 6,5, приблизительно при pH 7,0, приблизительно при pH 7,5, приблизительно при pH 8,0, приблизительно при pH 8,5, приблизительно при pH 9,0, приблизительно при pH 9,5, приблизительно при pH 10,0, приблизительно при pH 10,5, приблизительно при pH 11,0, приблизительно при pH 11,5, приблизительно при pH 12,0 или более.
В одном из аспектов полипептиды по изобретению могут быть термостабильными и могут сохранять гидролазную активность, например липазную, сатуразную, пальмитазную и/или стеаратазную активность, после воздействия температуры в диапазоне приблизительно от -100°C приблизительно до -80°C, приблизительно от -80°C приблизительно до -40°C, приблизительно от -40°C приблизительно до -20°C, приблизительно от -20°C приблизительно до 0°C, приблизительно от 0°C приблизительно до 5°C, приблизительно от 5°C приблизительно до 15°C, приблизительно от 15°C приблизительно до 25°C, приблизительно от 25°C приблизительно до 37°C, приблизительно от 37°C приблизительно до 45°C, приблизительно от 45°C приблизительно до 55°C, приблизительно от 55°C приблизительно до 70°C, приблизительно от 70°C приблизительно до 75°C, приблизительно от 75°C приблизительно до 85°C, приблизительно от 85°C приблизительно до 90°C, приблизительно от 90°C приблизительно до 95°C, приблизительно от 95°C приблизительно до 100°C, приблизительно от 100°C приблизительно до 105°C, приблизительно от 105°C приблизительно до 110°C, приблизительно от 110°C приблизительно до 120°C, или 95°C, 96°C, 97°C, 98°C, 99°C, 100°C, 101°C, 102°C, 103°C, 104°C, 105°C, 106°C, 107°C, 108°C, 109°C, 110°C, 111°C, 112°C, 113°C, 114°C, 115°C или более.
В некоторых вариантах осуществления термостабильные полипептиды сохраняют гидролазную активность, например липазную, сатуразную, пальмитазную и/или стеаратазную активность, после воздействия температуры в описанных выше диапазонах, приблизительно при pH 3,0, приблизительно при pH 3,5, приблизительно при pH 4,0, приблизительно при pH 4,5, приблизительно при pH 5,0, приблизительно при pH 5,5, приблизительно при pH 6,0, приблизительно при pH 6,5, приблизительно при pH 7,0, приблизительно при pH 7,5, приблизительно при pH 8,0, приблизительно при pH 8,5, приблизительно при pH 9,0, приблизительно при pH 9,5, приблизительно при pH 10,0, приблизительно при pH 10,5, приблизительно при pH 11,0, приблизительно при pH 11,5, приблизительно при pH 12,0 или более.
В одном из вариантов осуществления выделенные, синтетические или рекомбинантные нуклеиновые кислоты содержат последовательность, которая гибридизуется в строгих условиях с нуклеиновой кислотой по изобретению, например, с образцовой нуклеиновой кислотой по изобретению, содержащей последовательность, как указано в SEQ ID № 1, SEQ ID № 3, SEQ ID № 5, SEQ ID № 7, SEQ ID № 9, SEQ ID № 11, SEQ ID № 13, SEQ ID № 15, SEQ ID № 17, SEQ ID № 19, SEQ ID № 22 или SEQ ID № 23, или последовательность, как указано в SEQ ID № 1, которая включает замены одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати или двенадцати или более или всех остатков (модификации последовательности SEQ ID № 1), изложенные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, или их фрагменты или подпоследовательности и последовательности, (полностью) комплементарные им. В одном из аспектов нуклеиновая кислота кодирует полипептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью. Длина нуклеиновой кислоты может составлять по меньшей мере приблизительно 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 100, 125, 150, 175, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700 или более остатков или соответствовать полной длине гена или транскрипта, содержащего SEQ ID № 1, и содержать последовательность, как указано в SEQ ID № 1, которая содержит замены одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати или двенадцати или более или всех остатков (модификации аминокислотной последовательности) SEQ ID № 1, которые изложены в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23; и последовательности, (полностью) комплементарные им. В одном из аспектов строгие условия включают стадию отмывания, содержащую отмывание в 0,2×SSC при температуре приблизительно 65°C в течение приблизительно 15 минут.
В одном из вариантов осуществления зонд из нуклеиновой кислоты, например, зонд для идентификации нуклеиновой кислоты, кодирующей полипептид, обладающей гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, включает зонд, содержащий или состоящий по меньшей мере приблизительно из 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000 или более последовательных оснований последовательности по изобретению, или ее фрагментов или подпоследовательностей, где зонд позволяет идентифицировать нуклеиновую кислоту посредством связывания или гибридизации. Зонд может содержать олигонуклеотид, содержащий по меньшей мере приблизительно от 10 до 50, приблизительно от 20 до 60, приблизительно от 30 до 70, приблизительно от 40 до 80 или приблизительно от 60 до 100 последовательных оснований последовательности, содержащей последовательность по изобретению, или ее фрагменты или подпоследовательности. Зонд может содержать олигонуклеотид, содержащий по меньшей мере приблизительно от 10 до 50, приблизительно от 20 до 60, приблизительно от 30 до 70, приблизительно от 40 до 80 или приблизительно от 60 до 100 последовательных оснований последовательности нуклеиновой кислоты по изобретению, или ее подпоследовательность.
В одном из вариантов осуществления пара последовательностей праймеров для амплификации нуклеиновой кислоты, кодирующей полипептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, содержит пару праймеров, содержащую или состоящую из пары праймеров, позволяющих амплифицировать нуклеиновую кислоту, содержащую последовательность по изобретению, или ее фрагменты или подпоследовательности. Один или оба члена пары последовательностей праймеров для амплификации могут содержать олигонуклеотид, содержащий по меньшей мере приблизительно от 10 до 50 последовательных оснований из последовательности.
В одном из вариантов осуществления способы амплификации нуклеиновой кислоты, кодирующей полипептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, содержат амплификацию матричной нуклеиновой кислоты с использованием пары последовательностей праймеров для амплификации, позволяющих амплифицировать последовательность нуклеиновой кислоты по изобретению, или ее фрагменты или подпоследовательности.
В одном из вариантов осуществления экспрессирующие кассеты содержат нуклеиновую кислоту по изобретению, или ее подпоследовательность. В одном из аспектов экспрессирующая кассета может содержать нуклеиновую кислоту, которая функционально связана с промоторами. Промоторы могут представлять собой промоторы вирусов, бактерий, млекопитающих или растений. В одном из аспектов промоторы растений могут представлять собой промоторы картофеля, риса, кукурузы, пшеницы, табака или ячменя. Промоторы могут представлять собой конститутивный промотор. Конститутивный промотор может содержать CaMV35S. В другом аспекте промоторы могут представлять собой индуцируемые промоторы. В одном из аспектов промоторы могут представлять собой тканеспецифичные промоторы или промоторы, регулируемые окружающей средой или регулируемые развитием. Таким образом, промоторы могут представлять собой специфические промоторы, например, семян, листьев, корней, стебля, или промоторы, индуцируемые сбрасыванием. В одном из аспектов экспрессирующая кассета может дополнительно содержать экспрессирующий вектор растения или вируса растения.
В одном из вариантов осуществления переносчики для клонирования содержат экспрессирующую кассету (например, вектор) по изобретению, или нуклеиновую кислоту по изобретению. Переносчик для клонирования может представлять собой вирусный вектор, плазмиду, фаг, фагемиду, космиду, фосмиду, бактериофаг или искусственную хромосому. Вирусный вектор может содержать аденовирусный вектор, ретровирусный вектор или аденоассоциированный вирусный вектор. Переносчик для клонирования может содержать бактериальную искусственную хромосому (BAC), плазмиду, вектор, полученный из бактериофага P1 (РАС), искусственную хромосому дрожжей (YAC) или искусственную хромосому млекопитающего (MAC).
В одном из вариантов осуществления трансформированные клетки содержат нуклеиновую кислоту по изобретению, или экспрессирующую кассету (например, вектор) по изобретению, или переносчик для клонирования по изобретению. В одном из аспектов трансформированная клетка может представлять собой бактериальную клетку, клетку млекопитающего, клетку гриба, клетку дрожжей, клетку насекомого или клетку растения. В одном из аспектов клетка растения может представлять собой клетку картофеля, пшеницы, риса, кукурузы, табака или ячменя. Трансформированная клетка может представлять собой любую клетку-хозяина, известную специалистам в данной области, включая прокариотические клетки, эукариотические клетки, такие как бактериальные клетки, клетки грибов, клетки дрожжей, клетки млекопитающих, клетки насекомых или клетки растений. Образцовые бактериальные клетки включают любые виды в пределах родов Escherichia, Bacillus, Streptomyces, Salmonella, Pseudomonas и Staphylococcus, включая, например, Escherichia coli, Lactococcus lactis, Bacillus subtilis, Bacillus cereus, Salmonella typhimurium, Pseudomonas fluorescens. Образцовые клетки грибов включают любые виды Aspergillus. Образцовые клетки дрожжей включают любые виды Pichia, Saccharomyces, Schizosaccharomyces или Schwanniomyces, включая Pichia pastoris, Saccharomyces cerevisiae или Schizosaccharomyces pombe. Образцовые клетки насекомых включают любые виды Spodoptera или Drosophila, включая Drosophila S2 и Spodoptera Sf9. Образцовые клетки животных включают линии клеток CHO, COS или меланомы Боуэса или любые линии клеток мыши или человека.
В одном из вариантов осуществления трансгенные растения содержат нуклеиновую кислоту по изобретению, или экспрессирующую кассету (например, вектор) по изобретению. Трансгенным растением может являться растение кукурузы, растение картофеля, растение помидора, растение пшеницы, растение масличной культуры, растение рапса, растение сои, растение риса, растение ячменя или растение табака.
В одном из вариантов осуществления трансгенные семена содержат нуклеиновую кислоту по изобретению, или экспрессирующую кассету (например, вектор) по изобретению. Трансгенные семена могут представлять собой рис, семена кукурузы, зерна пшеницы, семена масличных культур, семена рапса, семена сои, ядра кокосовых орехов, семена подсолнечника, семена кунжута, семена растений арахиса или табака.
В одном из вариантов осуществления выделенные, синтетические или рекомбинантные полипептиды, обладающие гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, или полипептиды, позволяющие получить специфический иммунный ответ к гидролазе, например, к липазе, сатуразе, пальмитазе и/или стеаратазе (например, эпитоп); и в альтернативных аспектах пептиды и полипептиды по изобретению содержат последовательность:
(a) которая обладает по меньшей мере приблизительно 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более или обладает 100% (полной) идентичностью последовательностей с:
(i) аминокислотной последовательностью SEQ ID № 2 или ее ферментативно активными фрагментами и содержит замены по меньшей мере одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати, двенадцати, тринадцати, четырнадцати, пятнадцати, шестнадцати, семнадцати, восемнадцати, девятнадцати, двадцати, двадцати одного, двадцати двух, двадцати трех, двадцати четырех или более или всех аминокислотных остатков (или их эквиваленты), как указано в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, или
(ii) аминокислотной последовательностью SEQ ID № 2, SEQ ID № 4, SEQ ID № 6, SEQ ID № 8, SEQ ID № 10, SEQ ID № 12, SEQ ID № 14, SEQ ID № 16, SEQ ID № 18, или SEQ ID № 20
где полипептид или пептид по п. (i) или (ii) обладает гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, или полипептид или пептид позволяет получить специфическое антитело к гидролазе (например, к липазе, сатуразе, пальмитазе и/или стеаратазе) (полипептид или пептид, который выполняет функцию эпитопа или иммуногена),
(b) полипептида или пептида по п. (a), где идентичность последовательностей определяют: (a) посредством анализа с использованием алгоритма сравнения последовательностей или посредством визуального исследования, или (В) на участке длиной по меньшей мере приблизительно 20, 25, 30, 35, 40, 45, 50, 55, 60, 75, 100, 150, 200, 250, 300 или более аминокислотных остатков или по всей длине полипептида или пептида или фермента и/или их ферментативно активных подпоследовательностей (фрагментов),
(c) полипептида или пептида по п. (b), где алгоритмом сравнения последовательностей является алгоритм BLAST версии 2.2.2, где установлены следующие настройки фильтрации blastall -p blastp -d «nr pataa» -F F, а все остальные опции имеют значения по умолчанию;
(d) аминокислотную последовательность, кодируемую нуклеиновой кислотой, предоставленной в настоящем документе, где полипептид обладает (i) гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, или (ii) обладает такой иммуногенной активностью, что она позволяет получить антитело, которое специфически связывается с полипептидом, содержащим последовательность по п. (a), и/или с его ферментативно активными подпоследовательностями (фрагментами);
(e) аминокислотную последовательность по любому п. с (a) до (d), включающую по меньшей мере одну замену консервативного аминокислотного остатка, но полипептид или пептид сохраняет гидролазную активность, например, липазную, сатуразную, пальмитазную и/или стеаратазную активность;
(f) аминокислотную последовательность по п. (e), где консервативная замена включает замену алифатической аминокислоты на другую алифатическую аминокислоту; замену серина на треонин или наоборот; замену кислотного остатка на другой кислотный остаток; замену остатка, несущего аминогруппу, на другой остаток, несущий аминогруппу; замену основного остатка другим основным остатком; или замену ароматического остатка другим ароматическим остатком или их сочетание,
(g) аминокислотную последовательность по п. (f), где алифатический остаток включает аланин, валин, лейцин, изолейцин или их синтетический эквивалент; кислый остаток включает аспарагиновую кислоту, глутаминовую кислоту или их синтетический эквивалент; остаток, содержащий амидную группу, включает аспарагин, глутамин или их синтетический эквивалент; основный остаток содержит лизин, аргинин, гистидин или их синтетический эквивалент; или ароматический остаток содержит фенилаланин, тирозин, триптофан или их синтетический эквивалент;
(h) полипептида по любому из п.п. с (a) до (f), обладающего гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, но не содержащего сигнальную последовательность,
(i) полипептида по любому из п.п. с (a) до (h), обладающего гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, который дополнительно содержит гетерологичную последовательность;
(j) полипептида по п. (i), где гетерологичная последовательность содержит или состоит из: (a) гетерологичной сигнальной последовательности, (b) последовательности по п. (a), где гетерологичную сигнальную последовательность получают из гетерологичного фермента, и/или (c) метки, эпитопа, таргетинг-пептида, расщепляемой последовательности, поддающегося обнаружению фрагмента или участка; или
(m) содержат аминокислотную последовательность, кодируемую любой последовательностью нуклеиновой кислоты по изобретению.
Образцовые полипептидные или пептидные последовательности по изобретению, включают SEQ ID № 2 и ее подпоследовательности и варианты, например, длиной по меньшей мере приблизительно 30, 35, 40, 45, 50, 75, 100, 150, 200, 250, 300, 350, 400, 450, 500 или более остатков или полноразмерный фермент, все содержат замены одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати или двенадцати или более или всех аминокислотных остатков (модификации аминокислотной последовательности SEQ ID № 2), которые изложены в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23. Образцовые полипептидные или пептидные последовательности по изобретению, содержат последовательность, кодируемую нуклеиновой кислотой по изобретению. Образцовые полипептидные или пептидные последовательности по изобретению, включают полипептиды или пептиды, с которыми специфически связывается антитело по изобретению. В одном из аспектов, полипептид по изобретению обладает по меньшей мере одним видом гидролазной активности, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью. В одном из аспектов активность представляет собой региоселективную и/или хемоселективную активность.
В одном из аспектов выделенный, синтетический или рекомбинантный полипептид может содержать полипептид по изобретению, который не содержит сигнальную (пептидную) последовательность, например, не содержит гомологичную сигнальную последовательность, и в одном из аспектов содержит гетерологичную сигнальную (пептидную) последовательность. В одном из аспектов выделенный, синтетический или рекомбинантный полипептид может содержать полипептид по изобретению, содержащий гетерологичную сигнальную последовательность, такую как гетерологичная сигнальная последовательность гидролазы или негидролазы (например, не липазы, не сатуразы или не пальмитазы). В одном из аспектов химерные белки содержат первый домен, содержащий сигнальную последовательность по изобретению, и по меньшей мере второй домен. Белок может представлять собой слитый белок. Второй домен может содержать фермент. Фермент может представлять собой гидролазу (например, липазу, сатуразу, пальмитазу и/или стеаратазу) по изобретению или другую гидролазу.
В одном из аспектов гидролазная (например, липазная, сатуразная, пальмитазная и/или стеаратазная) активность включает конкретную активность приблизительно при 37°C в диапазоне приблизительно от 100 приблизительно до 1000 единиц на миллиграмм белка. В другом аспекте гидролазная (например, липазная, сатуразная, пальмитазная и/или стеаратазная) активность включает конкретную активность приблизительно от 500 приблизительно до 750 единиц на миллиграмм белка. Альтернативно гидролазная активность включает конкретную активность при 37°C в диапазоне приблизительно от 500 приблизительно до 1200 единиц на миллиграмм белка. В одном из аспектов гидролазная активность включает конкретную активность при 37°C в диапазоне приблизительно от 750 приблизительно до 1000 единиц на миллиграмм белка. В другом аспекте термостабильность включает сохранение по меньшей мере половины конкретной активности гидролазы при 37°C после нагревания до повышенной температуры. Альтернативно термостабильность может включать сохранение конкретной активности при 37°C в диапазоне приблизительно от 500 приблизительно до 1200 единиц на миллиграмм белка после нагревания до повышенной температуры.
В одном из вариантов осуществления выделенные, синтетические или рекомбинантные полипептиды по изобретению содержат по меньшей мере один сайт гликозилирования. В одном из аспектов гликозилирование может представлять собой N-гликозилирование. В одном из аспектов полипептид можно гликозилировать после экспрессии в P. pastoris или S. pombe или в растениях, таких как масличные растения, например, соя, канола, рис, подсолнечник или генетически модифицированные (ГМО) варианты этих растений.
В одном из аспектов полипептид может сохранять гидролазную (например, липазную, сатуразную, пальмитазную и/или стеаратазную) активность в условиях, включающих приблизительно pH 6,5, pH 6, pH 5,5, pH 5, pH 4,5 или pH 4,0 или ниже. В другом аспекте полипептид может сохранять гидролазную (например, липазную, сатуразную, пальмитазную и/или стеаратазную) активность в условиях, которые включают приблизительно pH 7, pH 7,5, pH 8,0, pH 8,5, pH 9, pH 9,5, pH 10, pH 10,5, pH 11, pH 11,5, pH 12,0 или более.
В одном из вариантов осуществления препараты белка содержат полипептид по изобретению, где препарат белка содержит жидкость, твердое вещество или гель.
В одном из аспектов гетеродимеры по изобретению содержат полипептид и второй домен. В одном из аспектов второй домен может представлять собой полипептид и гетеродимер может представлять собой слитый белок. В одном из аспектов второй домен может представлять собой эпитоп или метку. В одном из аспектов гомодимеры по изобретению, содержат полипептид по изобретению.
В одном из вариантов осуществления иммобилизованные полипептиды по изобретению обладают гидролазной (например, липазной, сатуразной, пальмитазной и/или стеаратазной) активностью, где полипептид содержит полипептид по изобретению, полипептид, кодируемый нуклеиновой кислотой по изобретению, или полипептид, содержащий полипептид по изобретению, и второй домен. В одном из аспектов полипептид по изобретению, можно иммобилизовать на клетке, везикуле, липосоме, пленке, мембране, металле, смоле, полимере, керамике, стекле, микроэлектроде, графитовой частице, бусине, геле, планшете, кристалле, таблетке, драже, капсуле, порошке, агломерате, поверхности, пористой структуре, блоке или капиллярной трубке или на материалах, таких как зерна, кожура, кора, кожа, волосы, эмаль, кость, кожура и материалах, получаемых из них. Полинуклеотиды, полипептиды и ферменты по изобретению, можно ввести в состав в твердой форме, такой как порошок, лиофилизированный препарат, гранулы, таблетка, пластинка, кристалл, капсула, драже, пеллет, или в жидкой форме, такой как водный раствор, аэрозоль, гель, паста, суспензия, водно-масляная эмульсия, крем, капсула или суспензия везикул или мицелл.
В одном из вариантов осуществления пищевые добавки для животных содержат полипептид по изобретению, например, полипептид, кодируемый нуклеиновой кислотой по изобретению. В одном из аспектов полипептид в пищевой добавке может быть гликозилированным. В одном из вариантов осуществления съедобные матрицы для доставки фермента содержат полипептид по изобретению, например, полипептид, кодируемый нуклеиновой кислотой по изобретению. В одном из аспектов матрица для доставки содержит пеллет. В одном из аспектов полипептид может быть гликозилирован. В одном из аспектов гидролазная активность является термостабильной. В другом аспекте гидролазная активность является термостабильной.
В одном из вариантов осуществления способы выделения или идентификации полипептида, обладающего гидролазной (например, липазной, сатуразной, пальмитазной и/или стеаратазной) активностью, включают стадии: (a) получение антитела по изобретению; (b) получение образца, содержащего полипептиды; и (c) контакт образца со стадии (b) с антителом со стадии (a) в условиях, в которых антитело может специфически связываться с полипептидом, посредством чего осуществляют выделение или идентификацию полипептида, обладающего гидролазной (например, липазной, сатуразной, пальмитазной и/или стеаратазной) активностью.
В одном из вариантов осуществления способы получения антитела против гидролазы включают введение животному, не относящемуся к человеку, нуклеиновой кислоты по изобретению, или полипептида по изобретению, или их подпоследовательности в количестве, достаточном для создания гуморального иммунного ответа, посредством чего получают антитело против гидролазы. Изобретение относится к способам получения антитела к гидролазе, которые включают введение животному, не являющемуся человеком, нуклеиновой кислоты по изобретению, или полипептида по изобретению, или их подпоследовательности в количестве, достаточном для получения иммунного ответа.
В одном из вариантов осуществления способы получения рекомбинантного полипептида включают стадии: (a) получение нуклеиновой кислоты по изобретению, функционально связанной с промоторами; и (b) экспрессия нуклеиновой кислоты со стадии (a) в условиях, которые допускают экспрессию полипептида, посредством чего получают рекомбинантный полипептид. В одном из аспектов способ может дополнительно содержать трансформацию клетки-хозяина нуклеиновой кислотой со стадии (a) с последующей экспрессией нуклеиновой кислоты со стадии (a), посредством чего получают рекомбинантный полипептид в трансформированной клетке.
В одном из вариантов осуществления способы идентификации полипептида, обладающего гидролазной (например, липазной, сатуразной, пальмитазной и/или стеаратазной) активностью, включают следующие стадии: (a) получение полипептида по изобретению; или полипептида, кодируемого нуклеиновой кислотой по изобретению; (b) получение субстрата гидролазы; и (c) контакт полипептида или его фрагмента или варианта со стадии (a) с субстратом со стадии (b) и выявление снижения количества субстрата или увеличения количества продукта реакции, где снижение количества субстрата или увеличение количества продукта реакции выявляет полипептид, обладающий гидролазной (например, липазной, сатуразной, пальмитазной и/или стеаратазной) активностью.
В одном из вариантов осуществления способы идентификации субстрата гидролазы включают следующие стадии: (a) получение полипептида по изобретению; или полипептида, кодируемого нуклеиновой кислотой по изобретению; (b) получение тестируемого субстрата; и (c) контакт полипептида со стадии (a) с тестируемым субстратом со стадии (b) и выявление снижения количества субстрата или увеличения количества продукта реакции, где снижение количества субстрата или увеличение количества продукта реакции идентифицирует тестируемый субстрат как субстрат гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы).
В одном из вариантов осуществления способы определения специфического связывания тестируемого соединения с полипептидом включают следующие стадии: (a) экспрессия нуклеиновой кислоты или вектора, содержащего нуклеиновую кислоту в условиях, допускающих трансляцию нуклеиновой кислоты в полипептид, где нуклеиновая кислота содержит нуклеиновую кислоту по изобретению, или предоставление полипептида по изобретению; (b) получение тестируемого соединения; (c) контакт полипептида с тестируемым соединением; и (d) определение специфического связывания тестируемого соединения со стадии (b) с полипептидом.
В одном из вариантов осуществления способы идентификации модулятора гидролазной (например, липазной, сатуразной, пальмитазной и/или стеаратазной) активности включают следующие стадии: (a) получение полипептида по изобретению, или полипептида, кодируемого нуклеиновой кислотой по изобретению; (b) получение тестируемого соединения; (c) контакт полипептида со стадии (a) с тестируемым соединением со стадии (b) и измерение активности гидролазы, где изменение гидролазной активности измеряют в присутствие тестируемого соединения в сравнении с активностью в отсутствие тестируемого соединения, которое позволяет определить, что тестируемое соединение модулирует гидролазную активность. В одном из аспектов гидролазную (например, липазную, сатуразную, пальмитазную и/или стеаратазную) активность можно измерить путем получения субстрата гидролазы и выявления снижения количества субстрата или увеличения количества продукта реакции или выявления увеличения количества субстрата или снижения количества продукта реакции. Снижение количества субстрата или увеличение количества продукта реакции при использовании тестируемого соединения по сравнению с количеством субстрата или продукта реакции без использования тестируемого соединения идентифицирует тестируемое соединение как активатор гидролазной активности. Увеличение количества субстрата или снижение количества продукта реакции при использовании тестируемого соединения в сравнении с количеством субстрата или продукта реакции без использования тестируемого соединения идентифицирует тестируемое соединение как ингибитор гидролазной активности.
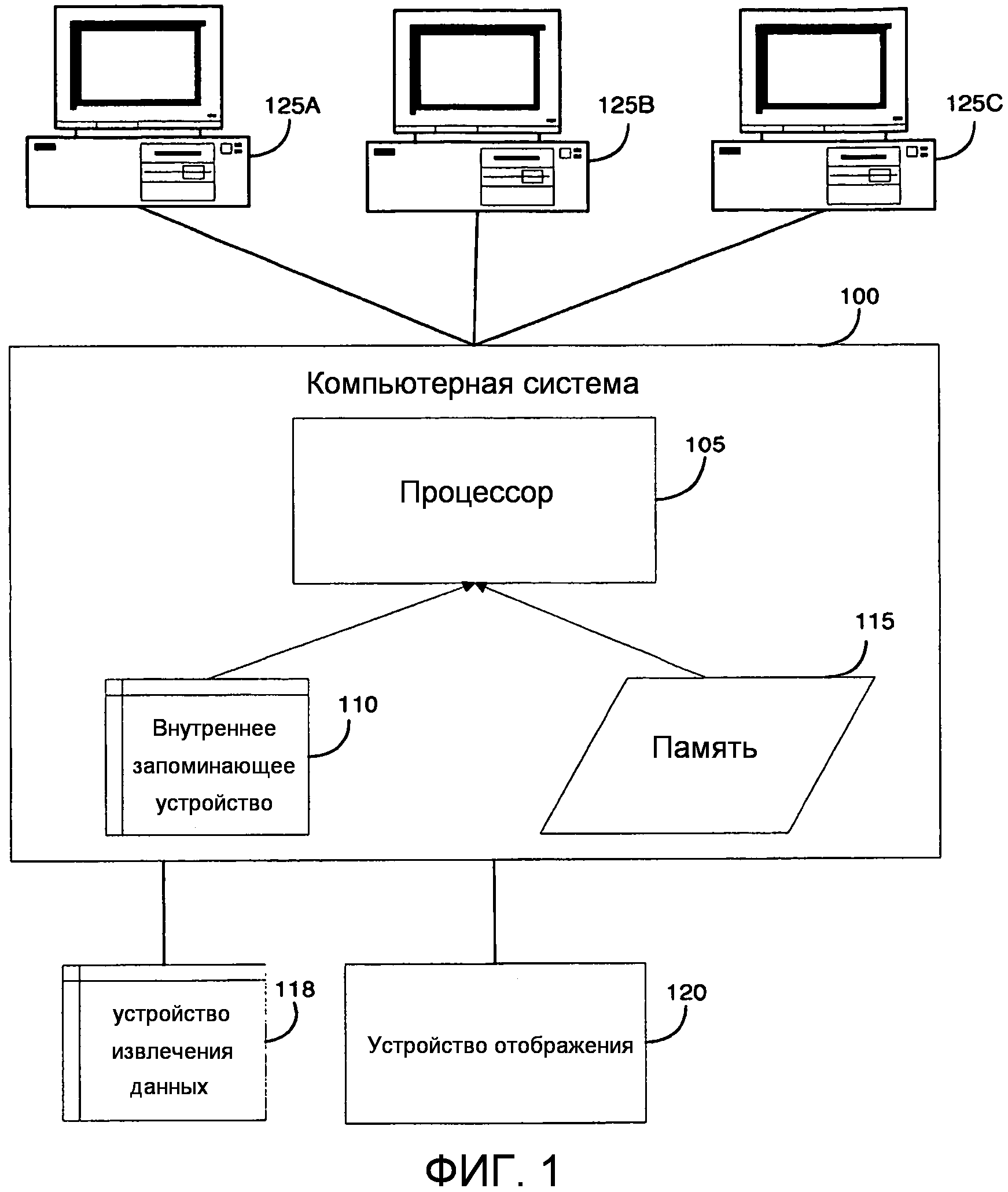
В одном из вариантов осуществления компьютерные системы содержат процессор и устройства хранения данных, где на указанном устройстве хранения данных хранят последовательность полипептида или последовательность нуклеиновой кислоты по изобретению, (например, полипептид, кодируемый нуклеиновой кислотой по изобретению). В одном из аспектов компьютерная система дополнительно может включать алгоритм сравнения последовательностей и устройство хранения данных, которое содержит хранящуюся на нем по меньшей мере одну эталонную последовательность. В другом аспекте алгоритм сравнения последовательностей включает компьютерную программу, которая отмечает полиморфизм. В одном из аспектов компьютерная система дополнительно может включать идентификатор, который идентифицирует один или несколько признаков в указанной последовательности. В одном из вариантов осуществления машиночитаемый носитель содержит хранящуюся на нем последовательность полипептида или последовательность нуклеиновой кислоты по изобретению.
В одном из вариантов осуществления способы идентификации признака в последовательности включают стадии: (a) считывание последовательности, используя компьютерную программу, которая идентифицирует один или несколько признаков в последовательности, где последовательность содержит последовательность полипептида или последовательность нуклеиновой кислоты по изобретению; и (b) идентификация одного или нескольких признаков в последовательности с использованием компьютерной программы.
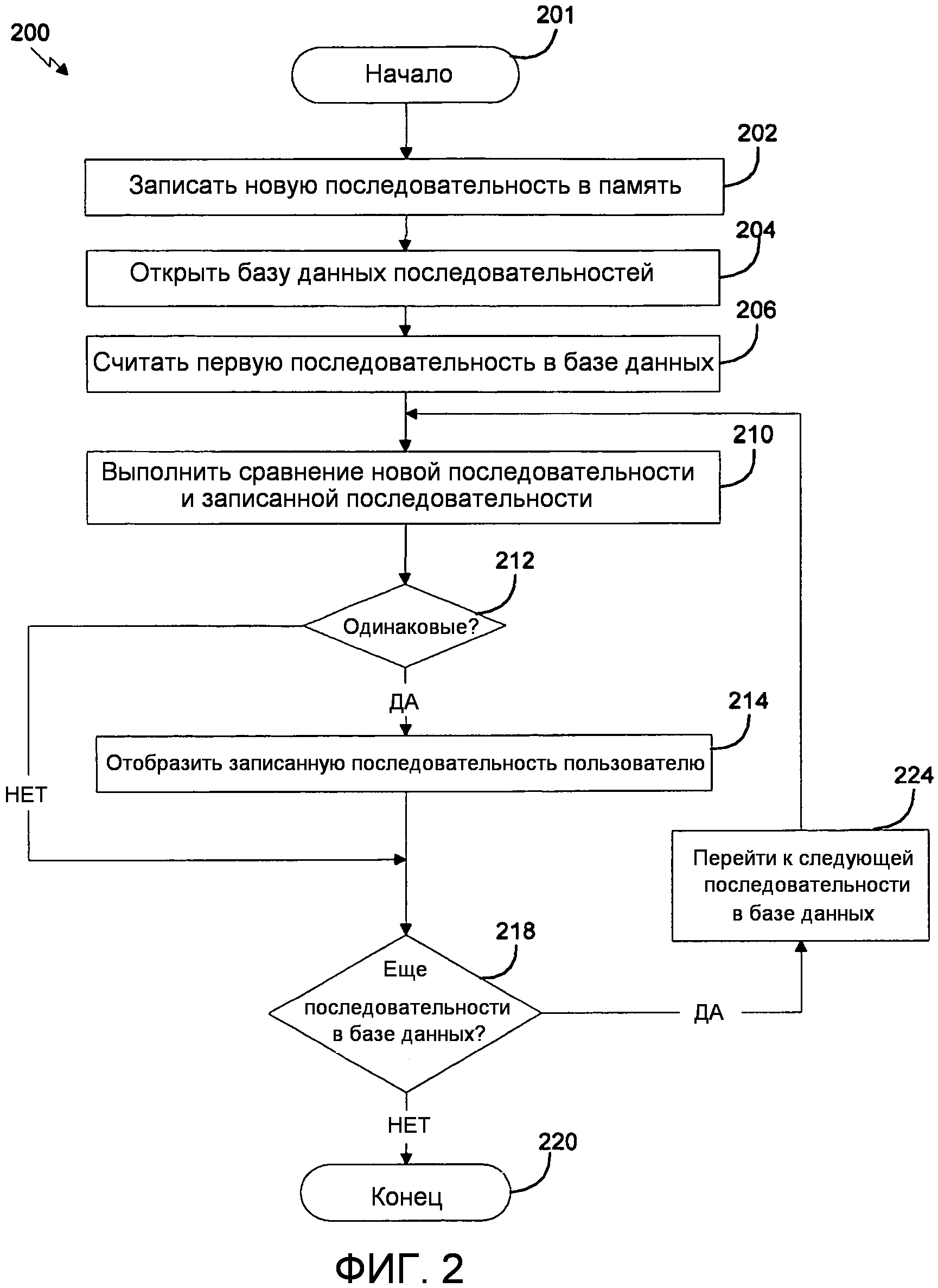
В другом варианте осуществления изобретение относится к способам сравнения первой последовательности со второй последовательностью, включающим стадии: (a) считывание первой последовательности и второй последовательности посредством компьютерной программы, которая сравнивает последовательности, где первая последовательность содержит последовательность полипептида или последовательность нуклеиновой кислоты по изобретению; и (b) определение различий между первой последовательностью и второй последовательностью с использованием компьютерной программы. Стадия определения различий между первой последовательностью и второй последовательностью дополнительно может содержать стадию идентификации полиморфизма. В одном из аспектов способ дополнительно может включать идентификатор, который идентифицирует один или несколько признаков в последовательности. В другом аспекте способ может включать считывание первой последовательности с использованием компьютерной программы и идентификацию одного или нескольких признаков в последовательности.
В одном из вариантов осуществления способы выделения или сбора нуклеиновой кислоты, кодирующей полипептид, обладающий гидролазной (например, липазной, сатуразной, пальмитазной и/или стеаратазной) активностью, из образца включают стадии: (a) получение пары последовательностей праймеров для амплификации нуклеиновой кислоты, кодирующей полипептид, обладающий гидролазной активностью, где пара праймеров позволяет амплифицировать нуклеиновую кислоту по изобретению; (b) выделение нуклеиновой кислоты из образца или обработка образца таким образом, что нуклеиновую кислоту в образце делают доступной для гибридизации с парой праймеров для амплификации; и, (c) объединение нуклеиновой кислоты со стадии (b) с парой праймеров для амплификации со стадии (a) и амплификация нуклеиновой кислоты из образца, посредством чего из образца выделяют или собирают нуклеиновую кислоту, кодирующую полипептид, обладающий гидролазной активностью. В одном из вариантов осуществления образец представляет собой образец окружающей среды, например, образец воды, образец жидкости, образец почвы, образец воздуха или биологический образец, например, клетку бактерии, клетку простейшего, клетку насекомого, клетку дрожжей, клетку растения, клетку гриба или клетку млекопитающего. Один или оба члена пары последовательностей праймеров для амплификации могут содержать олигонуклеотид, содержащий по меньшей мере приблизительно от 10 до 50 или более последовательных оснований из последовательности по изобретению.
В одном из вариантов осуществления способы увеличения термостабильности или термостабильности полипептида гидролазы включают гликозилирование полипептида гидролазы, где полипептид содержит по меньшей мере тридцать смежных аминокислот из полипептида по изобретению; или из полипептида, кодируемого последовательностью нуклеиновой кислоты по изобретению, посредством чего увеличивают термостабильность или термостабильность полипептида гидролазы. В одном из аспектов конкретная активность гидролазы может быть термостабильной или термостабильной при температуре в диапазоне приблизительно от более чем 37°C приблизительно до 95°C.
В одном из вариантов осуществления способы повышенной экспрессии полипептида рекомбинантной гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы) в клетке включают экспрессию вектора, содержащего нуклеиновую кислоту по изобретению, или последовательность нуклеиновой кислоты по изобретению, где идентичность последовательностей определяют посредством анализа с использованием алгоритма сравнения последовательностей или посредством визуального исследования, где повышенную экспрессию обеспечивают посредством использования промоторов с высокой активностью, дицистронного вектора или посредством амплификации гена вектора.
В одном из вариантов осуществления моющие составы, содержащие полипептид по изобретению, или полипептид, кодируемый нуклеиновой кислотой по изобретению, содержат гидролазную активность, например, липазную, сатуразную, пальмитазную и/или стеаратазную активность. В одном из аспектов гидролаза может представлять собой не поверхностно-активную гидролазу. В другом аспекте гидролаза может представлять собой поверхностно-активную гидролазу.
В одном из вариантов осуществления способы мытья объекта включают следующие стадии: (a) получение композиции, содержащей полипептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, где полипептид содержит: полипептид по изобретению, или полипептид, кодируемый нуклеиновой кислотой по изобретению; (b) получение объекта; и (c) контакт полипептида со стадии (a) с объектом со стадии (b) в условиях, в которых композиция может мыть объект.
В одном из вариантов осуществления способы получения трансгенного растения включают следующие стадии: (a) введение гетерологичной последовательности нуклеиновой кислоты в клетку растения, где гетерологичная последовательность нуклеиновой кислоты содержит последовательность нуклеиновой кислоты по изобретению, посредством чего получают трансформированную клетку растения; и (b) получение трансгенного растения из трансформированной клетки. В одном из аспектов стадия (a) дополнительно может содержать введение гетерологичной последовательности нуклеиновой кислоты посредством электропорации или микроинъекции протопластов клеток растения. В другом аспекте стадия (a) дополнительно может содержать введение гетерологичной последовательности нуклеиновой кислоты непосредственно в ткань растения посредством бомбардировки частицами ДНК. Альтернативно стадия (a) дополнительно может содержать введение гетерологичной последовательности нуклеиновой кислоты в клетку растения ДНК, используя хозяина Agrobacterium tumefaciens. В одном из аспектов клеткой растения может являться клетка картофеля, кукурузы, риса, пшеницы, табака или ячменя.
В одном из вариантов осуществления способы экспрессии гетерологичной последовательности нуклеиновой кислоты в клетке растения включают следующие стадии: (a) трансформация клетки растения гетерологичной последовательностью нуклеиновой кислоты, функционально связанной с промоторами, где гетерологичная последовательность нуклеиновой кислоты содержит нуклеиновую кислоту по изобретению; (b) выращивание растения в условиях, в которых происходит экспрессия гетерологичной последовательности нуклеиновой кислоты в клетках растения.
В одном из вариантов осуществления первый способ биокаталитического синтеза структурированного липида включает следующие стадии: (a) получение полипептида (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению; (b) получение композиции, содержащей триацилглицерид (ТАГ); (c) контакт полипептида со стадии (a) с композицией со стадии (b) в условиях, в которых полипептид осуществляет гидролитическое отщепление остатка карбоновой кислоты в положении Sn2 триацилглицерида (ТАГ), посредством чего получают 1,3-диацилглицерид (ДАГ); (d) предоставление R1-сложного эфира; (e) получение R1-специфической гидролазы, и (f) контакт 1,3-ДАГ со стадии (c) с R1-сложным эфиром со стадии (d) и R1-специфической гидролазой со стадии (e) в условиях, в которых R1-специфическая гидролаза катализирует этерификацию в положении Sn2, посредством чего получают структурированный липид. Гидролаза по изобретению, может представлять собой Sn2-специфическую липазу. Структурированный липид может содержать альтернативу масла какао (CBA), синтетическое масло какао, натуральное масло какао, 1,3-дипальмитоил-2-олеоилглицерин (POP), 1,3-дистеароил-2-олеоилглицерин (SOS), 1-пальмитоил-2-олеоил-3-стеароилглицерин (POS) или 1-олеоил-2,3-димиристоилглицерин (OMM).
В одном из вариантов осуществления второй способ биокаталитического синтеза структурированного липида включает следующие стадии: (a) получение гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению; (b) получение композиции, содержащей триацилглицерид (ТАГ); (c) контакт полипептида со стадии (a) с композицией со стадии (b) в условиях, в которых полипептид осуществляет гидролиз остатка карбоновой кислоты в положении Sn1 или Sn3 триацилглицерида (ТАГ), посредством чего получают 1,2-ДАГ или 2,3-ДАГ; и (d) содействие перегруппировке остатка карбоновой кислоты в 1,2-ДАГ или 2,3-ДАГ со стадии (c) в кинетически контролируемых условиях, посредством чего получают композицию содержащую 1,3-ДАГ.
Этот второй способ дополнительно может включать предоставление R1-сложного эфира и R1-специфической липазы и контакт 1,3-ДАГ со стадии (d) со R1-сложным эфиром и R1-специфической липазой в условиях, в которых R1-специфическая липаза катализирует этерификацию в положении Sn2, посредством чего получают структурированный липид. Гидролаза, например, липаза, сатураза, пальмитаза и/или стеаратаза по изобретению, может представлять собой Sn1- или Sn3-специфический фермент. Структурированный липид может содержать любое растительное масло, например, соевое масло, масло канолы, альтернативу масла какао (CBA), синтетическое масло какао, натуральное масло какао, 1,3-дипальмитоил-2-олеоилглицерин (POP), 1,3-дистеароил-2-олеоилглицерин (SOS), 1-пальмитоил-2-олеоил-3-стеароилглицерин (POS) или 1-олеоил-2,3-димиристоилглицерин (OMM).
R1-сложный эфир может содержать менее насыщенный фрагмент, чем гидролизованный остаток карбоновой кислоты, и в этом случае полученный таким образом структурированный липид представляет собой менее насыщенный жир или масло, чем исходный ТАГ. R1-сложный эфир может содержать одно или несколько веществ из омега-3 жирной кислоты, омега-6 жирной кислоты, мононенасыщенной жирной кислоты, полиненасыщенной жирной кислоты, фосфогруппы, сложного эфира фитостерина и оризанола. Более конкретно R1-сложный эфир может содержать фрагмент, выбранный из группы, состоящей из α-линоленовой кислоты, эйкозапентаеновой кислоты, докозагексаеновой кислоты, γ-линоленовой кислоты, дигомо-γ-линоленовой кислоты, арахидоновой кислоты, олеиновой кислоты, пальмолеиновой кислоты, холина, серин, β-ситостерина, куместрола, диэтилстильбэстрола и оризанола.
В одном из аспектов этого второго способа стадия (d) дополнительно включает использование ионообменных смол. Кинетически контролируемые условия могут включать неравновесные условия, которые ведут к получению конечного продукта, который обладает отношением 1,3-ДАГ к 2,3-ДАГ более 2:1. Композиция со стадии (b) может содержать флуорогенную жирную кислоту (ЖК). Композиция со стадии (b) может содержать умбеллифериловый эфир ЖК. Конечный продукт может быть энантиомерно чистым.
В одном из вариантов осуществления способ получения менее насыщенного жира или масла включает следующие стадии: (a) получение полипептида (гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению; (b) получение масла или жира, и (c) контакт полипептида со стадии (a) с маслом или жиром со стадии (b) в условиях, в которых гидролаза может модифицировать масло или жир, например, удалить по меньшей мере одну насыщенную жирную кислоту, например, пальмитиновую, стеариновую, лауриновую, каприловую кислоту (октановую кислоту) и т.п. Модификация может включать катализируемый гидролазой гидролиз жира или масла. Гидролиз может представлять собой полный или частичный гидролиз жира или масла. Гидролизованное масло может содержать глицериновый эфир полиненасыщенной жирной кислоты, который может заместить удаленную насыщенную жирную кислоту, или рыбий жир, животное или растительное масло. Растительное масло может содержать оливковое, пальмовое, соевое масло, масло канолы, подсолнечника или лауриновое масло или рисовое масло или их сочетание.
В одном из вариантов осуществления способ получения менее насыщенного жира или масла, которое может содержать незаменимые жирные кислоты, включает следующие стадии: (a) получение полипептида (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению; (b) получение композиции, содержащей триацилглицерид (ТАГ); (c) контакт полипептида со стадии (a) с композицией со стадии (b) в условиях, в которых полипептид гидролизует остаток карбоновой кислоты в положении Sn1 или Sn3 триацилглицерида (ТАГ), посредством чего получают 1,2-ДАГ или 2,3-ДАГ; и (d) содействие перегруппировке остатка карбоновой кислоты в 1,2-ДАГ или 2,3-ДАГ со стадии (c) при кинетически контролируемых условиях, посредством чего получают 1,3-ДАГ.
Способ дополнительно может включать предоставление R1-сложного эфира и R1-специфической липазы и контакт 1,3-ДАГ со стадии (d) с R1-сложным эфиром и R1-специфической липазой в условиях, в которых R1-специфическая липаза катализирует этерификацию в положении Sn2, посредством чего получают структурированный липид. R1-сложный эфир может содержать менее насыщенный фрагмент, чем гидролизованный остаток карбоновой кислоты, и в этом случае полученный таким образом структурированный липид представляет собой менее насыщенный жир или масло, чем исходный ТАГ. R1-сложный эфир может содержать омега-3 жирную кислоту (α-линоленовую, эйкозапентаеновую (ЭПК), докозагексаеновую (ДГК)), омега-6 жирную кислоту (γ-линоленовую, дигомо-γ-линоленовую (ДГЛК) или арахидоновую), мононенасыщенную жирную кислоту (олеиновую, пальмолеиновую и т.п.), фосфогруппы (холин и серин), сложные эфиры фитостерина (β-ситостерин, куместрол и диэтилстильбэстрол) и оризанол. Гидролаза, например, липаза, сатураза, пальмитаза и/или стеаратаза по изобретению, может являться Sn1- или Sn3-специфическим ферментом. Жир или масло со сниженным содержанием насыщенных жирных кислот можно получить посредством описанного выше гидролиза любого масла водорослей, растительного масла или животного жира или масла, например, масла Neochloris oleoabundans, масла Scenedesmus dimorphus, масла Euglena gracilis, масла Phaeodactylum tricornmutum, масла Pleurochrysis carterae, масла Prymnesium parvum, масла Tetraselmis chui, масла Tetraselmis suecica, масла Isochrysis galbana, масла Nannochloropsis salina, масла Botryococcus braunii, масла Dunaliella tertiolecta, масла видов Nannochloris, масла видов Spirulina, масла Chlorophycease (зеленые водоросли) и масла Bacilliarophy, масла канолы, касторового масла, кокосового масла, кориандрового масла, кукурузного масла, хлопкового масла, масла лесного ореха, конопляного масла, льняного масла, масла пенника лугового, оливкового масла, пальмового масла, косточкового пальмового масла, арахисового масла, рапсового масла, рисового масла, сафлорового масла, масла сасанквы, соевого масла, масла подсолнечника, таллового масла, масла цубаки, разнообразных «натуральных» масел, обладающих измененным составом жирных кислот за счет генетически модифицированных организмов (ГМО) или традиционной «селекции», такие как масла с высоким содержанием олеиновой кислоты, с низким содержанием линоленовой кислоты или с низким содержанием насыщенных жирных кислот (масло канолы с высоким содержанием олеиновой кислоты, соевое масло с низким содержанием линоленовой кислоты или масло подсолнечника с высоким содержанием стеариновой кислоты); животных жиров (сала, лярда, жира коровьего масла и куриного жира), рыбьего жира (жира тихоокеанского талеихта, жира печени трески, жира хоплостета, жира сардины, жира сельди и жира менхадена или смесей любых указанных выше. Полученные таким образом жир или масло со сниженным содержанием насыщенных жирных кислот можно использовать в продуктах питания или в печеных, жареных или термически обработанных продуктах, содержащих масла или жиры с низким содержанием некоторых жирных кислот, включая масла с низким содержанием пальмитиновой кислоты, олеиновой кислоты, лауриновой кислоты, стеариновой кислоты, каприловой кислоты (октановой кислоты) и т.д., которые обработаны с использованием композиции или способа по изобретению.
В одном из вариантов осуществления способ очистки лубриканта включает следующие стадии: (a) получение композиции, содержащей гидролазу (например, липазу, сатуразу, пальмитазу и/или стеаратазу) по изобретению; (b) получение лубриканта; и (c) обработка лубриканта гидролазой в условиях, в которых гидролаза (например, липаза, сатураза, пальмитаза и/или стеаратаза) по изобретению может осуществлять избирательный гидролиз масел в смазочном средстве, таким образом очищая его. Лубрикант может представлять собой гидравлическое масло.
В одном из вариантов осуществления способ обработки материи включает следующие стадии: (a) получение композиции, содержащей гидролазу (например, липазу, сатуразу, пальмитазу и/или стеаратазу) по изобретению где гидролаза может избирательно гидролизовать сложные эфиры карбоновых кислот; (b) получение материи; и (c) обработка материи гидролазой при условиях, в которых гидролаза может осуществлять избирательных гидролиз сложных эфиров карбоновых кислот, таким образом обрабатывая материю. Обработка материи может включать улучшение тактильных и драпировочных свойств конечной материи, окрашивания, получение замедления горения, получение водоотталкивающих свойств, получение оптического отбеливания или получение отделки смолой. Материя может содержать хлопок, вискозу, вискозное волокно, лиоцелл, лен, полотно, рами, любые их смеси или их смеси с полиэстерами, шерстью, полиамиды, акрилы или полиакрилы. В одном из вариантов осуществления материя, нить или пряжа содержит гидролазу по изобретению, которую можно адсорбировать, абсорбировать или иммобилизовать на поверхности материи, пряжи или волокна.
В одном из вариантов осуществления способ удаления или снижения количества пятен от пищевых продуктов или масла включает конакт гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению с пятном от продукта питания или масла в условиях, в которых гидролаза может осуществлять гидролиз масла или жира в пятне. Гидролаза (например, липаза, сатураза, пальмитаза и/или стеаратаза) по изобретению может обладать увеличенной устойчивостью к денатурации поверхностно-активными веществами и к тепловой деактивации. Гидролаза (например, липаза, сатураза, пальмитаза и/или стеаратаза) по изобретению может иметь детергент или раствор для стирки.
В одном из вариантов осуществления диетическая композиция содержит гидролазу (например, липазу, сатуразу, пальмитазу и/или стеаратазу) по изобретению. Диетическая композиция дополнительно может содержать питательную основу, содержащую жир. Гидролазу можно активировать солью желчной кислоты. Диетическая композиция дополнительно может содержать состав для грудных детей на основе коровьего молока. Гидролаза может осуществлять гидролиз жирных кислот с длинной цепью.
В одном из вариантов осуществления способ снижения содержания жира в молоке или диетических композициях на основе овощей включает следующие стадии: (a) получение композиции, содержащей гидролазу (например, липазу, сатуразу, пальмитазу и/или стеаратазу) по изобретению; (b) получение композиции, содержащей молоко или растительное масло, и (c) обработка композиции со стадии (b) гидролазой в условиях, в которых гидролаза может осуществлять гидролиз масла или жира в композиции. В одном из вариантов осуществления диетическая композиция для человека или животных с однокамерным желудком содержит питательную основу, где основа содержит жир и не содержит гидролазу или содержит небольшое количество гидролазы, и эффективное количество гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению, для увеличения всасывания жира и роста человека или животного с однокамерным желудком.
В одном из вариантов осуществления способ катализа реакции переэтерификации для получения новых триацилглицеридов включает следующие стадии: (a) получение композиции, содержащей полипептид (например, липазу, сатуразу, пальмитазу и/или стеаратазу) по изобретению, где полипептид может катализировать реакцию переэтерификации; (b) получение смеси триацилглицеридов и свободных жирных кислот; (c) обработка смеси со стадии (b) полипептидом в условиях, в которых полипептид может катализировать обмен свободной жирной кислоты и ацильных групп триацилглицеридов, посредством чего получают новые триацилглицериды, обогащенные присоединенными жирными кислотами. Полипептид может представлять собой Sn1,3-специфическую липазу.
В одном из вариантов осуществления способ переэтерификации для получения масла с низким содержанием транс-изомеров жирных кислот и низким содержанием жирных кислот со средней длиной цепи включает следующие стадии: (a) получение реакционной смеси для переэтерификации, которая содержит вещество источника стеариновой кислоты, выбранное из группы, состоящей из стеариновой кислоты, сложных моноэфиров стеариновой кислоты и одноатомных спиртов с низкой молекулярной массой и их смесей, (b) получение жидкого растительного масла; (c) получение полипептида (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению, где полипептид обладает 1,3-специфической липазной активностью; (d) переэтерификация вещества источника стеариновой кислоты и триацилглицерида растительного масла, (e) отделение компонентов переэтерифицированных свободных жирных кислот от глицеридных компонентов смеси переэтерификации, чтобы предоставить продукт переэтерификации масла маргарина и смесь жирных кислот, которая содержит жирные кислоты, сложные моноэфиры жирных кислот или их смеси, освобожденные от растительного масла, и (f) гидрогенизация смеси жирных кислот. В одном из вариантов осуществления способа переэтерификации реакцию переэтерификации продолжают до тех пор, пока в реакционной смеси присутствует значительное соотношение сложноэфирных групп в положениях 1, 3 глицеридного компонента и компонентов неглицеридных жирных кислот.
В одном из вариантов осуществления способ получения композиции, содержащей 1-пальмитоил-3-стеароил-2-монолеин (POSt) и 1,3-дистеароил-2-монолеин (StOSt), включает получение полипептида (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению, где полипептид позволяет осуществлять катализируемую 1,3-специфической липазой переэтерификацию 1,3-дипальмитоил-2-монолеина (POP) и стеариновой кислоты или тристеарина, и контакт указанного полипептида с композицией, содержащей указанный POP в присутствие источника стеарина, такого как стеариновая кислота или тристеарин для получения продукта, обогащенного 1-пальмитоил-3-стеароил-2-монолеином (POSt) или 1,3-дистеароил-2-монолеином (StOSt).
В одном из вариантов осуществления способ улучшения или предотвращения опосредованной липополисахаридами (ЛПС) токсичности включает введение пациенту фармацевтической композиции, содержащей гидролазу (например, липазу, сатуразу, пальмитазу и/или стеаратазу) по изобретению. В одном из вариантов осуществления способ детоксификации эндотоксина включает контакт эндотоксина с гидролазой (например, липазой, сатуразой, пальмитазой и/или стеаратазой) по изобретению. В одном из вариантов осуществления способ отщепления 2' или 3' цепи жирной кислоты деацилированием от липида A содержит контакт липида A с полипептидом по изобретению.
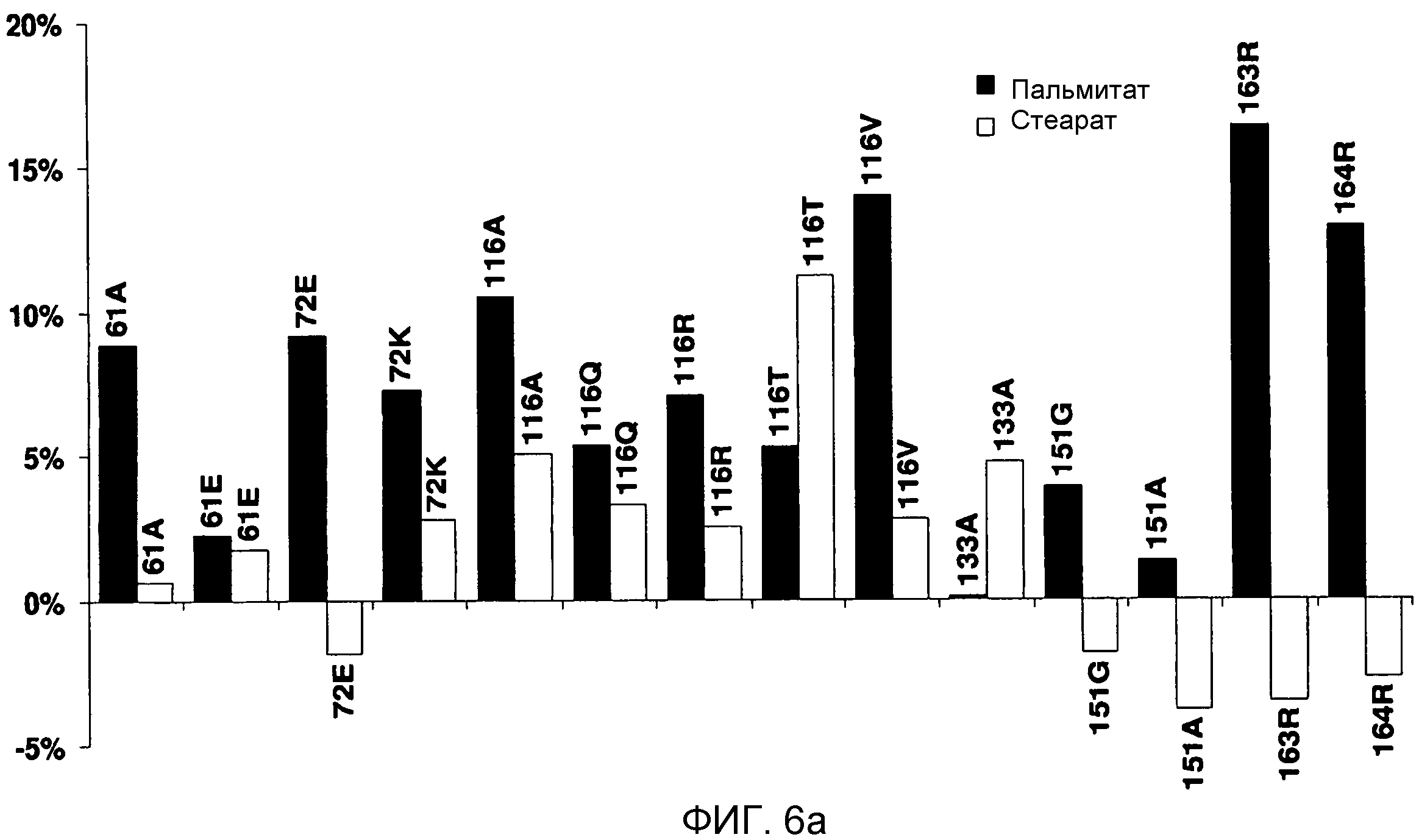
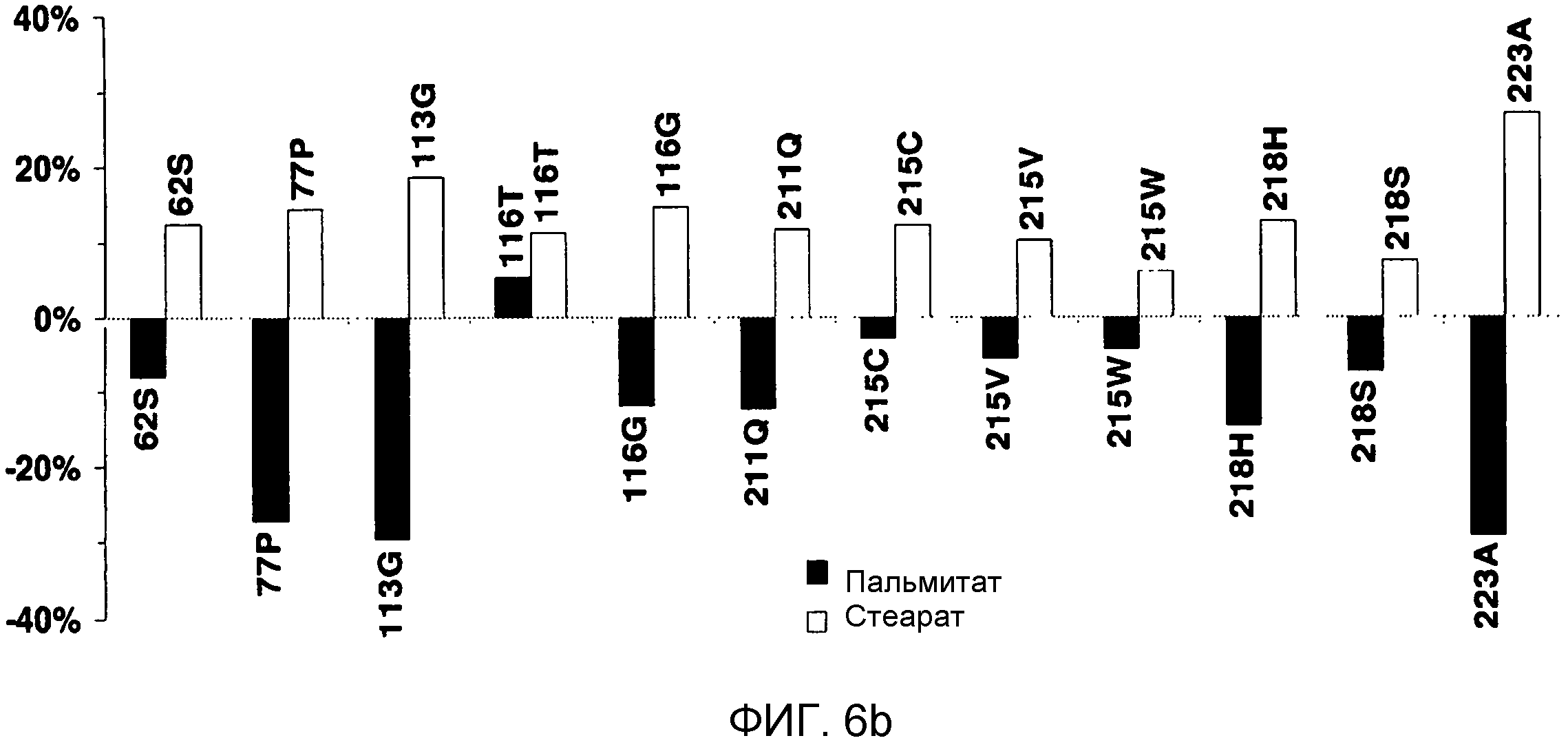
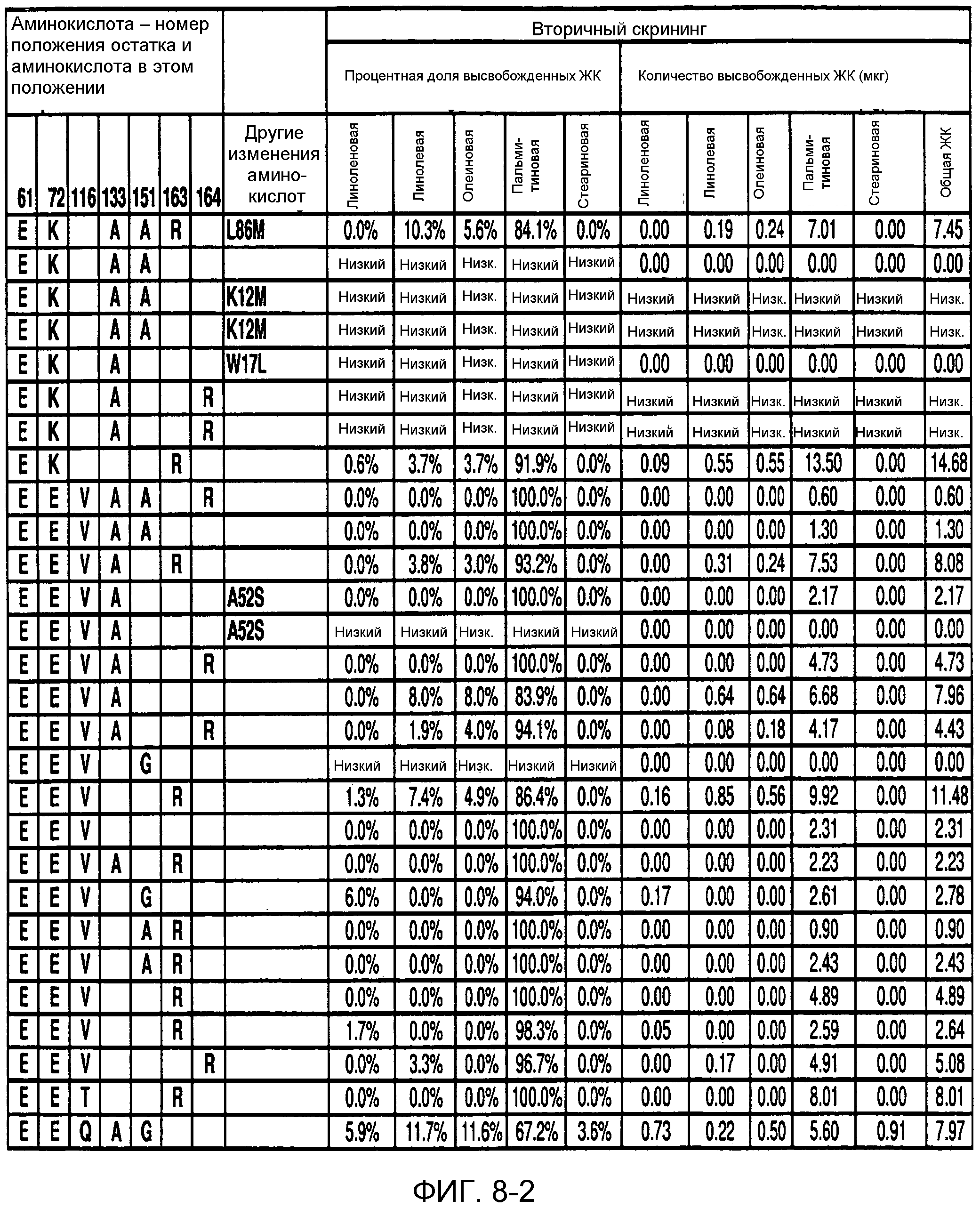
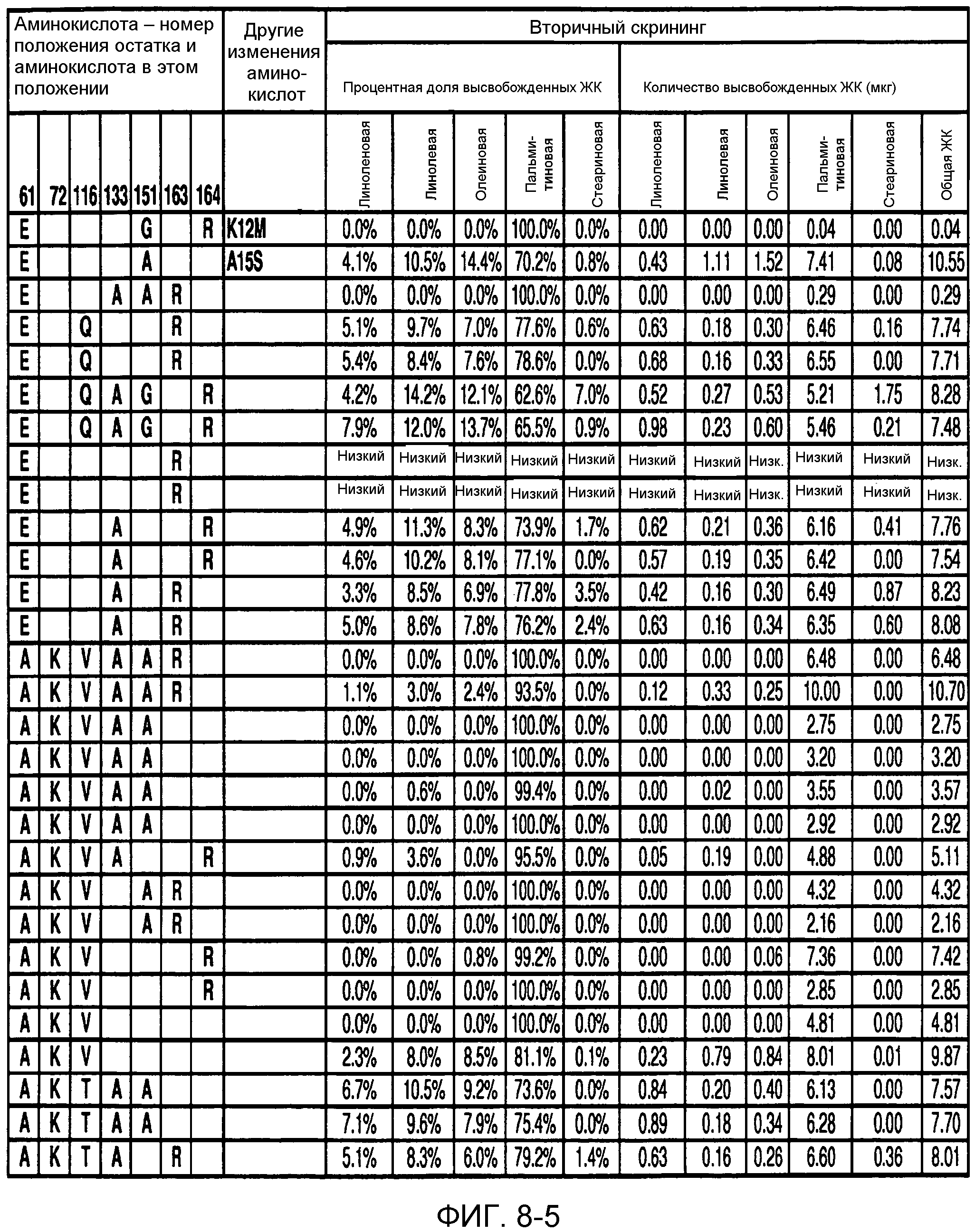
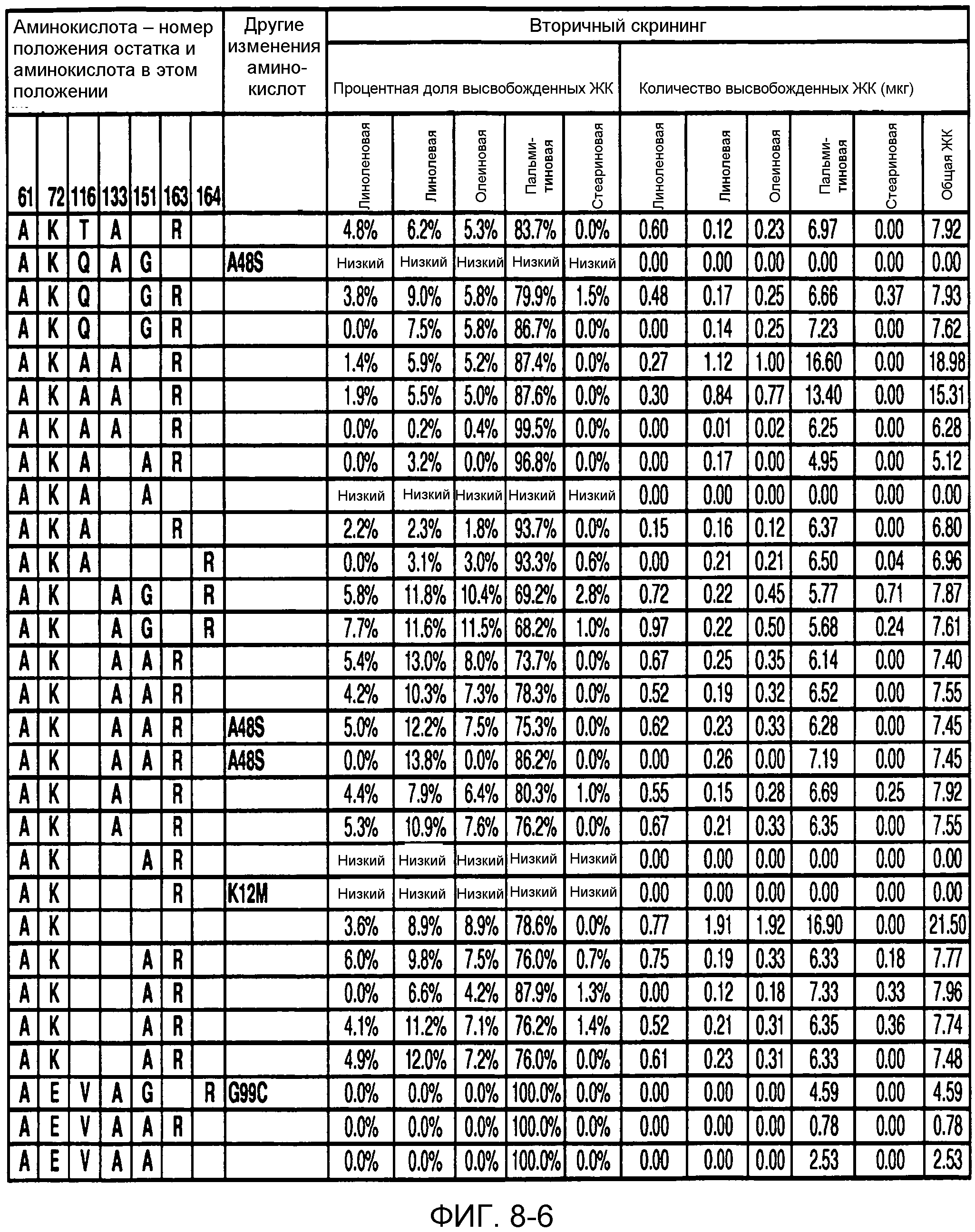
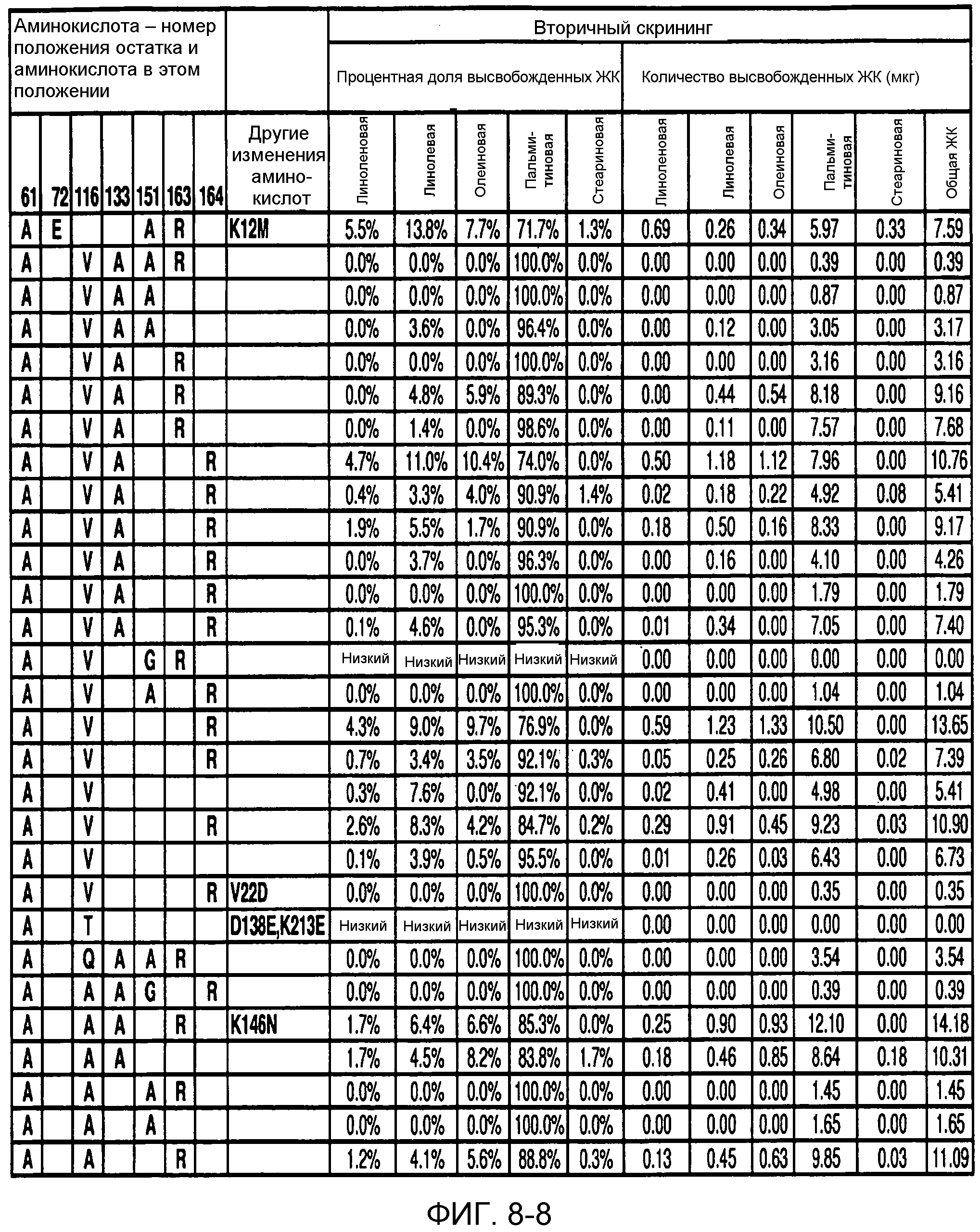
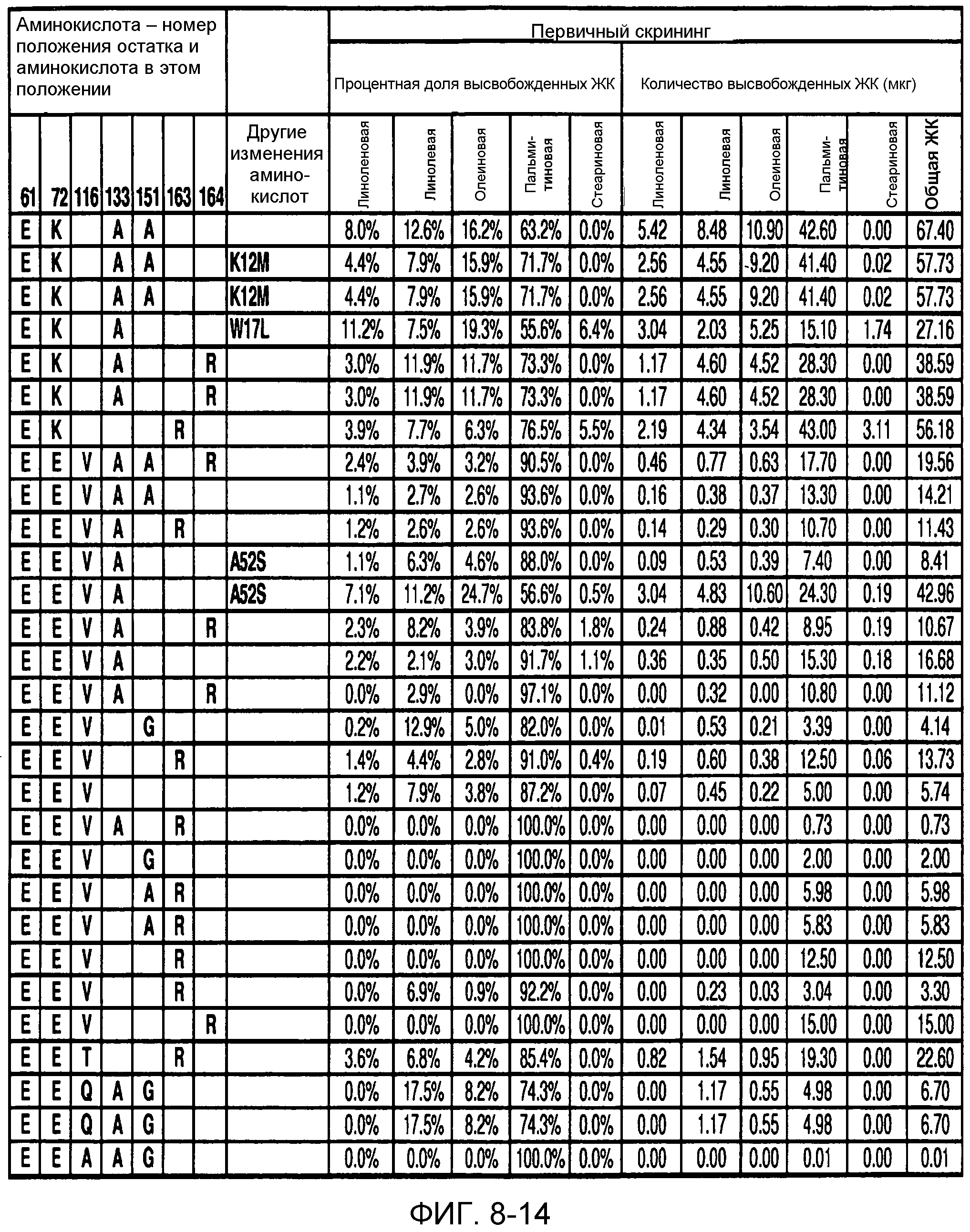
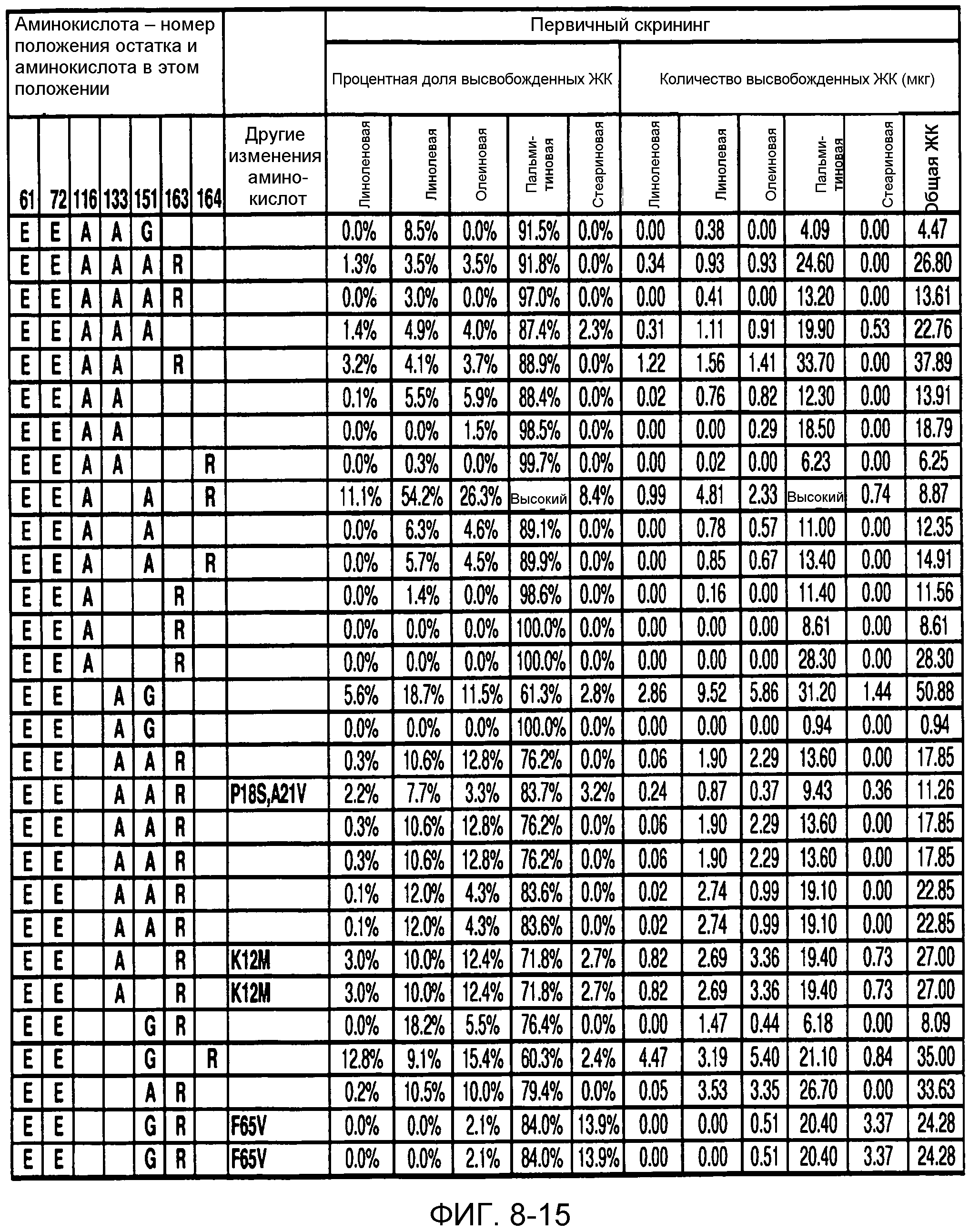
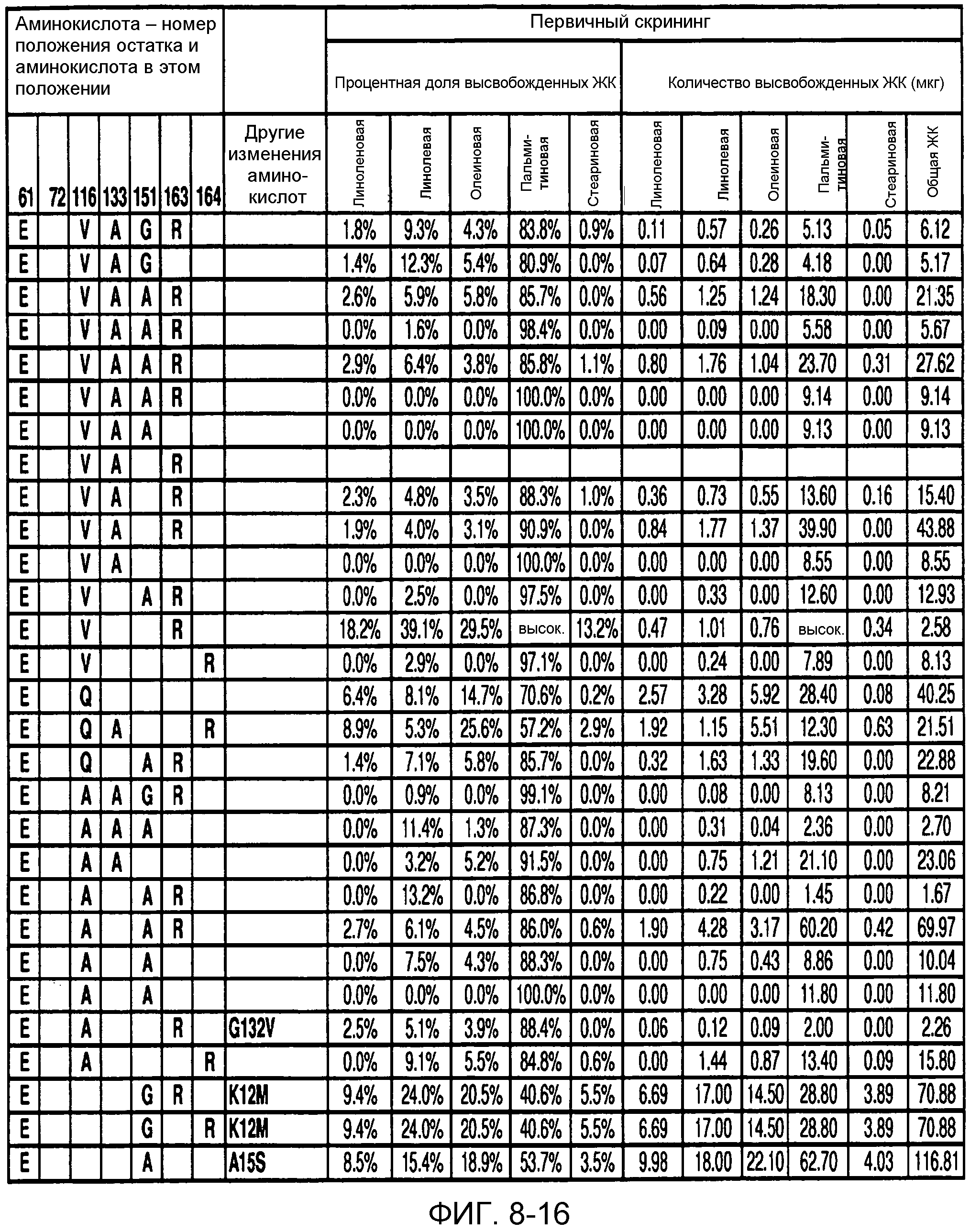
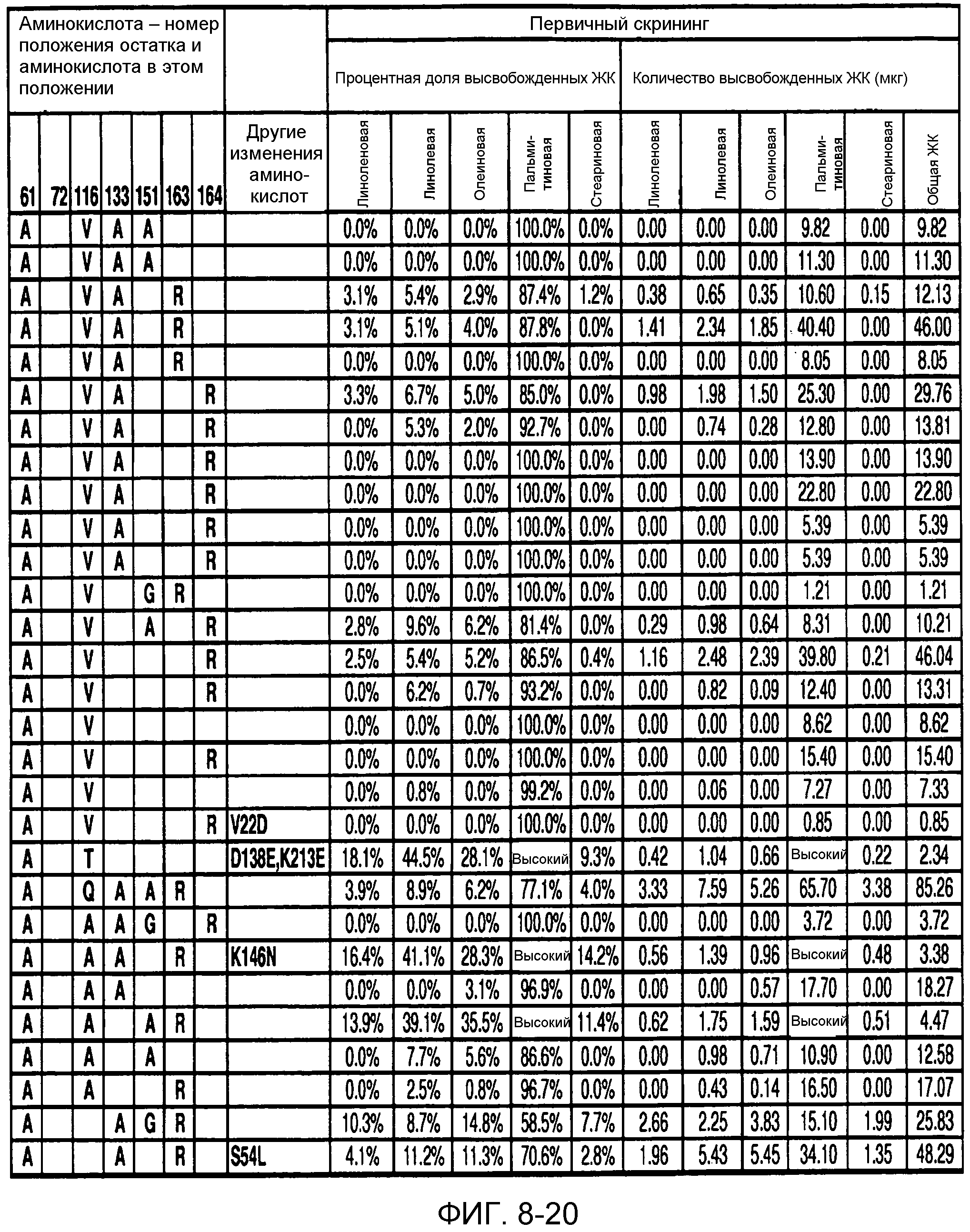
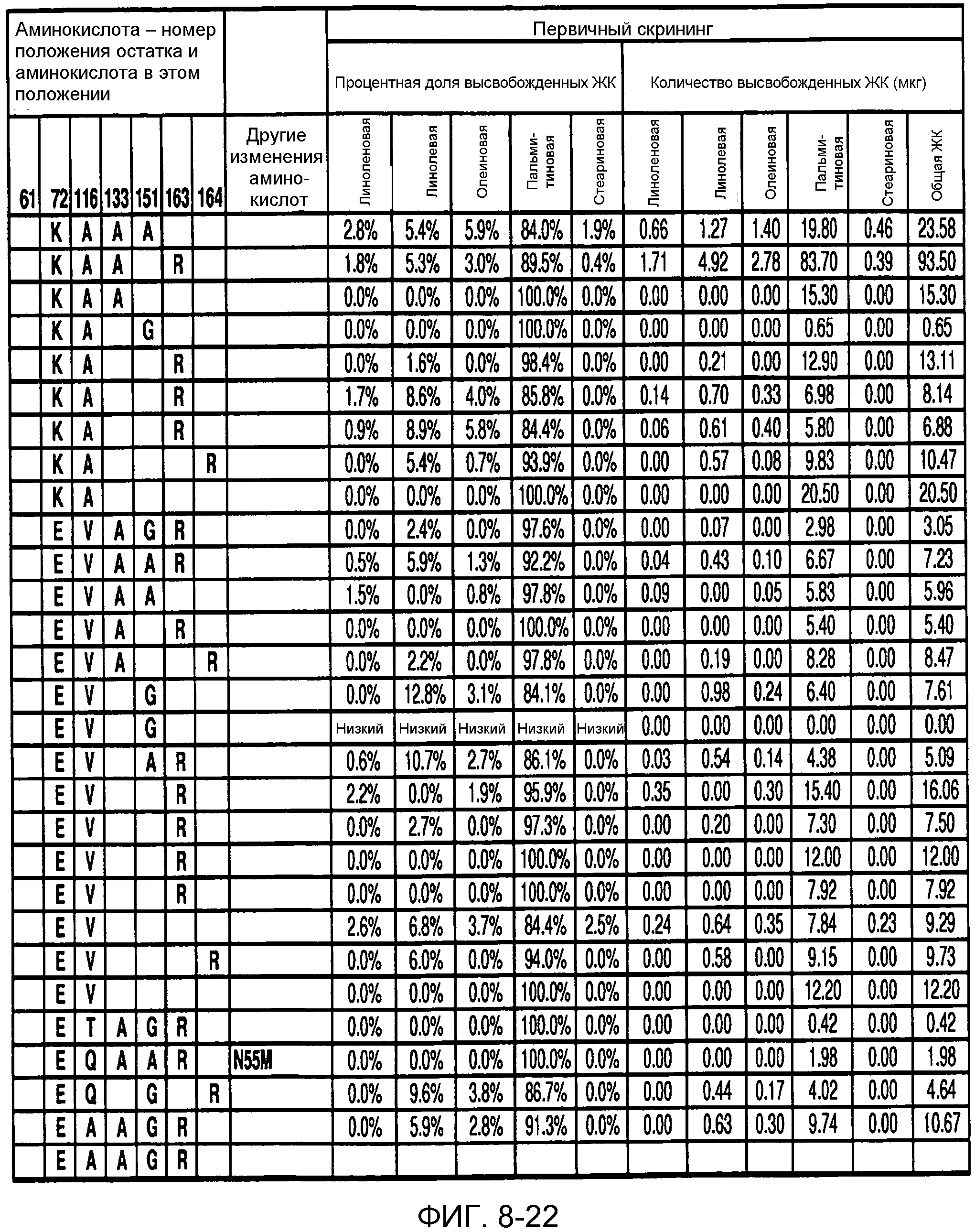
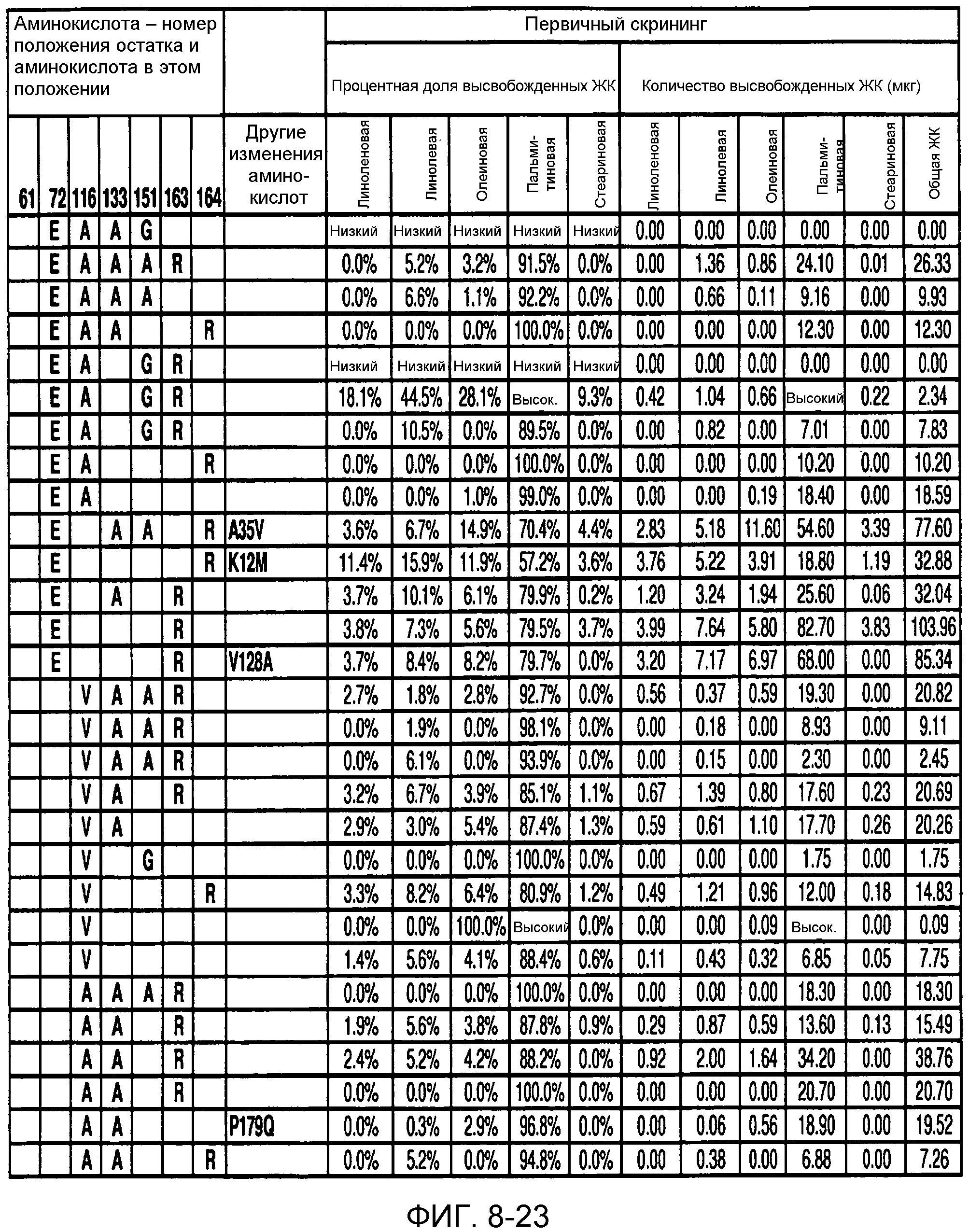
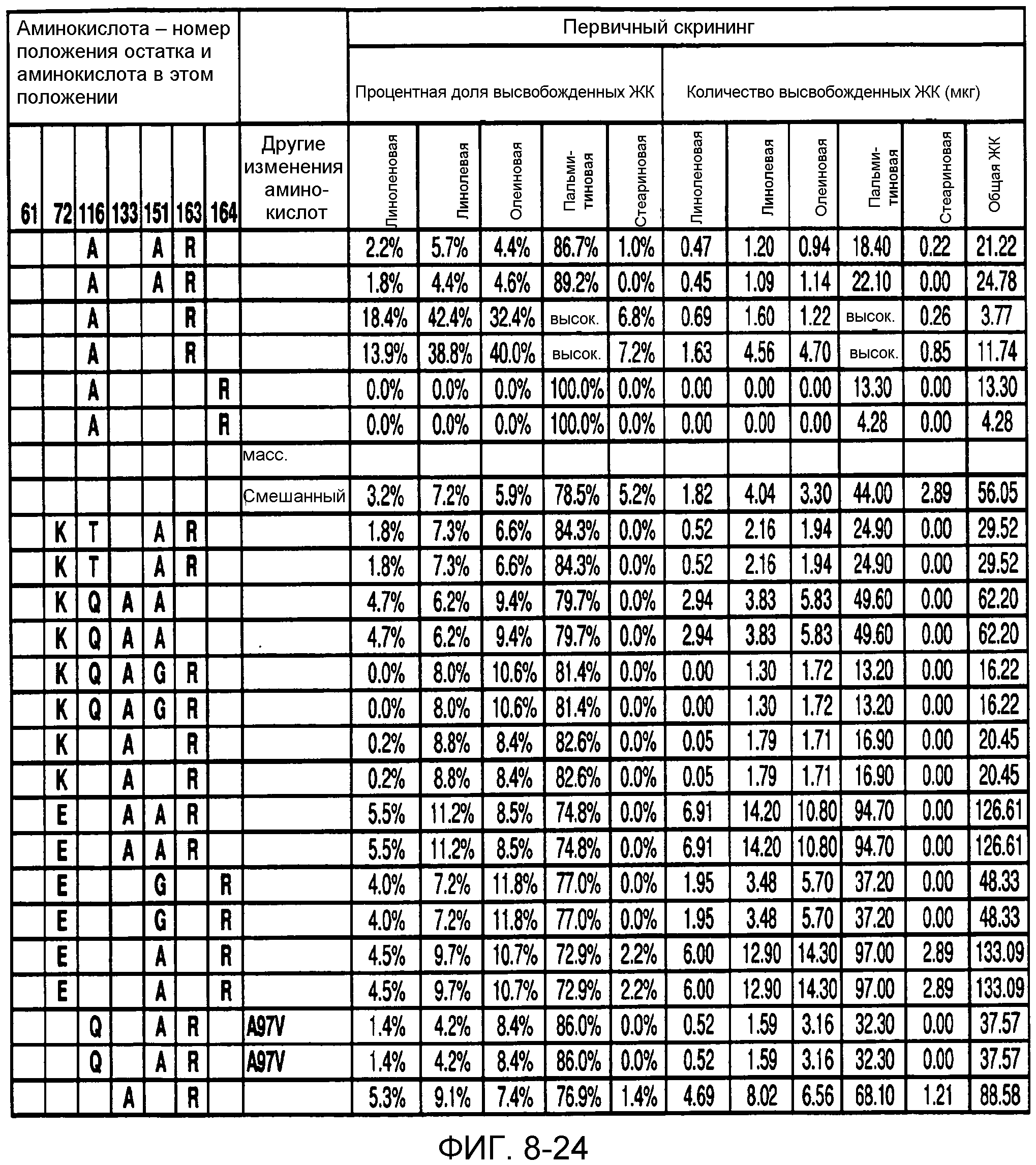
В одном из вариантов осуществления способы изменения субстратной специфичности или субстратного предпочтения исходного фермента липазы (гидролазы жирной кислоты), содержащего аминокислотную последовательность, соответствующую аминокислотной последовательности в SEQ ID № 2, включают стадию создания (введения) по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 или более мутаций аминокислотных остатков в SEQ ID № 2, как показано в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, посредством чего создают новый фермент гидролазу, обладающий модифицированной аминокислотной последовательностью и измененной субстратной специфичностью или субстратными предпочтениями по сравнению с исходной SEQ ID № 2 фермента липазы (гидролазы жирной кислоты). В одном из аспектов субстратная специфичность или субстратные предпочтения нового фермента липазы (гидролазы жирной кислоты) включает предпочтительное или повышенное гидролитическое отщепление пальмитиновой кислоты от масла, или субстратная специфичность или субстратные предпочтения нового фермента липазы (гидролазы жирной кислоты) включают предпочтительное или повышенное гидролитическое отщепление стеариновой кислоты от масла.
В одном из аспектов модифицированная аминокислотная последовательность (по сравнению с «исходной» SEQ ID № 2) содержит по меньшей мере одну модификацию аминокислоты A48C; D49R; D61A; D61E; R72E; R72K; V83M; R85Y; E95K; E116A; E116I; E116L; E116N; E116Q; E116R; E116T; E116V; S133A; A144I; E149H; A150I; I151G; I151A; P162G; P162K; V163R; D164R; R172H; R172L; или A225S, или их эквиваленты или их сочетание и/или по меньшей мере одну модификацию кодона (GCG)35(GCT); (GGC)45(GGA); (GCG)92(GCT), (GTG)102(GTT); (AGC)108(AGT); (CTG)117(CTT); (CTG)124(TTG); (CGG)126(AGG); (GTC)128(GTG); (AGT)133(TCT); (TTC)135(TTT); (GTG)183(GTT); (ACC)188(ACG) или их эквиваленты или их сочетание, а субстратная специфичность или субстратные предпочтения нового фермента липазы (гидролазы жирной кислоты) включают предпочтительное или повышенное гидролитическое отщепление пальмитиновой кислоты от масла. В одном из аспектов модифицированная аминокислотная последовательность (по сравнению с «исходной» SEQ ID № 2) содержит I20L; V62S; G77P; V83C; D88H; Y113G; E116T; E116G; H140K; K146S; I167S; L180E; E194M; A211Q; S212Y; G215C; G215V; G215W; A218H; A218S; V223A; A225M; A225Q или их сочетание, а субстратная специфичность или субстратные предпочтения нового фермента липазы (гидролазы жирной кислоты) включают предпочтительное или повышенное гидролитическое отщепление стеариновой кислоты от масла.
В одном из вариантов осуществления способы получения фермента, обладающего субстратной специфичностью или субстратными предпочтениями, включая предпочтительное или повышенное гидролитическое отщепление пальмитиновой кислоты от масла, включают стадии: (a) получение исходного фермента гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы), обладающего субстратной специфичностью или субстратными предпочтениями, включает предпочтительное гидролитическое отщепление пальмитиновой кислоты от масла, где исходный фермент гидролаза (например, липаза, сатураза, пальмитаза и/или стеаратаза) содержит последовательность по изобретению; и (b) создание по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 или более модификаций аминокислотных остатков в исходном ферменте гидролазе (например, липазе, сатуразе, пальмитазе и/или стеаратазе), где модификация аминокислотных остатков соответствуют мутациям аминокислотной последовательности в SEQ ID № 2, как показано в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, посредством чего создают фермент, обладающий субстратной специфичностью или субстратными предпочтениями, включая предпочтительное или повышенное гидролитическое отщепление пальмитиновой кислоты от масла.
В одном из вариантов осуществления способы получения фермента, обладающего субстратной специфичностью или субстратными предпочтениями, включая предпочтительное или повышенное гидролитическое отщепление стеариновой кислоты от масла, включают стадии: (a) получение исходного фермента гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы), обладающего субстратной специфичностью или субстратными предпочтениями, включая предпочтительное гидролитическое отщепление стеариновой кислоты от масла, где исходный фермент гидролаза (например, липаза, сатураза, пальмитаза и/или стеаратаза), содержит последовательность по изобретению; и (b) создание по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 или более модификаций аминокислотных остатков в исходном ферменте гидролазе (например, липазе, сатуразе, пальмитазе и/или стеаратазе), где модификации аминокислотных остатков соответствуют мутациям аминокислотной последовательности в SEQ ID № 2, как показано в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, посредством чего создают фермент, обладающий субстратной специфичностью или субстратными предпочтениями, включая предпочтительное или повышенное гидролитическое отщепление стеариновой кислоты от масла.
В одном из вариантов осуществления способы получения фермента гидролазы жирной кислоты (например, липазы, сатуразы, пальмитазы и/или стеаратазы), обладающего субстратной специфичностью или субстратными предпочтениями, включая предпочтительный гидролиз конкретной жирной кислоты, включают стадии (a) получения последовательности фермента гидролазы жирной кислоты (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению; (b) создания (введения) по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 или более мутаций остатков оснований в нуклеиновой кислоте, где мутации соответствуют тем изменениям последовательности, которые указаны в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23; и (c) тестирования активности вновь созданного фермента на субстратную специфичность или субстратные предпочтения, включая предпочтительное гидролитическое отщепление конкретной жирной кислоты, посредством чего получают новый фермент гидролазу жирной кислоты (например, липазу, сатуразу, пальмитазу и/или стеаратазу), обладающий субстратной специфичностью или субстратными предпочтениями, включая предпочтительный гидролиз конкретной жирной кислоты. В одном из аспектов фермент гидролаза жирной кислоты (например, липаза, сатураза, пальмитаза и/или стеаратаза) содержит последовательность, как указано в SEQ ID № 2. В одном из аспектов жирная кислота представляет собой линоленовую кислоту, линолевую кислоту, олеиновую кислоту, пальмитиновую кислоту или стеариновую кислоту.
В одном из вариантов осуществления способы получения фермента гидролазы жирной кислоты (например, липазы, сатуразы, пальмитазы и/или стеаратазы), обладающего субстратной специфичностью или субстратными предпочтениями, включая предпочтительный гидролиз конкретной жирной кислоты, включают стадии (a) получения последовательности нуклеиновой кислоты, кодирующей фермент гидролазу жирной кислоты (например, липазу, сатуразу, пальмитазу и/или стеаратазу) по изобретению; (b) создания (введения) по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 или более мутаций остатков оснований в нуклеиновую кислоту, где мутации соответствуют тем изменениям последовательности, которые указаны в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23; и, (c) экспрессии созданной нуклеиновой кислоты для получения нового фермента гидролазы жирной кислоты (например, липаза, сатуразы, пальмитазы и/или стеаратазы), посредством чего создают фермент гидролазу жирной кислоты (например, липазу, сатуразу, пальмитазу и/или стеаратазу), обладающий субстратной специфичностью или субстратными предпочтениями, включая предпочтительный гидролиз конкретной жирной кислоты.
В одном из аспектов последовательность, кодирующая фермент гидролазу жирной кислоты (например, липазу, сатуразу, пальмитазу и/или стеаратазу), содержит последовательность, как указано в SEQ ID № 1. В одном из аспектов жирной кислотой является линоленовая кислота, линолевая кислота, олеиновая кислота, пальмитиновая кислота или стеариновая кислота. В одном из аспектов новый фермент гидролаза жирной кислоты (например, липаза, сатураза, пальмитаза и/или стеаратаза) обладает субстратной специфичностью или субстратными предпочтениями к пальмитиновой кислоте по сравнению с субстратной специфичностью или субстратными предпочтениями исходного фермента гидролазы жирной кислоты (например, липазы, сатуразы, пальмитазы и/или стеаратазы) к стеариновой кислоте, или новый фермент гидролаза жирной кислоты (например, липаза, сатураза, пальмитаза и/или стеаратаза) обладает субстратной специфичностью или субстратными предпочтениями к стеариновой кислоте по сравнению с субстратной специфичностью или субстратными предпочтениями исходного фермента гидролазы жирной кислоты (например, липазы, сатуразы, пальмитазы и/или стеаратазы) к пальмитиновой кислоте.
В одном из вариантов осуществления липазы содержат аминокислотную последовательность, как указано в SEQ ID № 2, но также содержат по меньшей мере модификацию аминокислотного остатка A48C; D49R; D61A; D61E; R72E; R72K; V83M; R85Y; E95K; E116A; E116I; E116L; E116N; E116Q; E116R; E116T; E116V; S133A; A144I; E149H; A150I; I151G; I151A; P162G; P162K; V163R; D164R; R172H; R172L; или A225S, или их эквиваленты или их сочетание, и/или по меньшей мере одну модификацию кодона (GCG)35(GCT); (GGC)45(GGA); (GCG)92(GCT), (GTG)102(GTT); (AGC)108(AGT); (CTG)117(CTT); (CTG)124(TTG); (CGG)126(AGG); (GTC)128(GTG); (AGT)133(TCT); (TTC)135(TTT); (GTG)183(GTT); (ACC)188(ACG) или их эквиваленты или их сочетание.
В одном из аспектов субстратная специфичность или субстратные предпочтения новой липазы включают предпочтительное или повышенное гидролитическое отщепление жирной кислоты от масла по сравнению с «исходной» SEQ ID № 2. В одном из аспектов жирной кислотой является линоленовая кислота, линолевая кислота, олеиновая кислота, пальмитиновая кислота или стеариновая кислота.
Детали одного или нескольких вариантов осуществления изобретения, указаны на прилагаемых фигурах и в нижеследующем описании. Другие признаки, цели и преимущества изобретения, станут ясны из описания и фигур, а также из формулы изобретения.
Все публикации, патенты, патентные заявки, последовательности GenBank и депозиты ATCC, цитируемые в настоящем документе, таким образом, в явной форме включены ссылкой для всех целей.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Следующие фигуры только иллюстрируют варианты осуществления изобретения, но не ограничивают объем формулы изобретения.
Файл патента или заявки содержит по меньшей мере одну фигуру, выполненную в цвете. Копии публикации этого патента или патентной заявки с цветной фигурой(ами) предоставит Бюро по запросу и после уплаты необходимой пошлины.
На фиг. 1 представлена блочная диаграмма компьютерной системы.
На фиг. 2 представлена блок-схема, которая иллюстрирует один из аспектов процесса сравнения новой нуклеотидной или белковой последовательности с базой данных о последовательностях для того, чтобы определить уровни гомологии между новой последовательностью и последовательностями в базе данных.
На фиг. 3 представлена блок-схема, которая иллюстрирует один из аспектов процесса в компьютере для определения гомологии двух последовательностей.
На фиг. 4 представлена блок-схема, которая иллюстрирует один из аспектов процесса идентификации 300 для определения присутствия признака в последовательности.
На фиг. 5 представлен образцовый способ по изобретению, включающий использование липаз по изобретению, для обработки липида, например, липида из соевого масла, для избирательного гидролитического отщепления пальмитиновой кислоты для получения «соевого масла со сниженным содержанием пальмитиновой кислоты».
На фиг. 6a показано влияние образцовых мутаций GSSMSM пальмитазы на гидролиз пальмитата и стеарата относительно исходной SEQ ID № 2, которые подробно рассмотрены далее в примере 4. На 6b показано влияние образцовых мутаций GSSMSM стеаратазы на гидролиз пальмитата и стеарата относительно исходной SEQ ID № 2, которые подробно рассмотрены далее в примере 4.
На фиг. 7 показана SEQ ID № 2 с конкретными положениями влияющих на гидролиз пальмитата и стеарата мутаций, которые отмечены жирным начертанием увеличенного шрифта. Подчеркнутые мутации (например, 61A, E) представляют собой положения альтернативных аминокислотных остатков (альтернативные последовательности для альтернативных вариантов осуществления) для улучшения гидролиза пальмитата. Мутации, обозначенные курсивом, (например, 20L) представляют собой положения альтернативных аминокислотных остатков (альтернативные последовательности для альтернативных вариантов осуществления) для улучшения гидролиза стеарата. Положение 116 представляет собой положение мутации альтернативного аминокислотного остатка (альтернативная последовательность для альтернативного варианта осуществления) для улучшения гидролиза и пальмитата и стеарата.
На фиг. 8 показаны данные подтверждающего анализа соевого масла для отобранных клонов из библиотеки пальмитаз.
Одинаковые обозначения на разных чертежах обозначают одинаковые элементы.
ПОДРОБНОЕ ОПИСАНИЕ
Альтернативные варианты осуществления включают полипептиды, включая липазы, сатуразы, пальмитазы и/или стеаратазы, кодирующие их полинуклеотиды и способы получения и использования этих полинуклеотидов и полипептидов. Альтернативные варианты осуществления включают полипептиды, например, ферменты, обладающие гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, включая термостабильную и термостабильную гидролазную активность, и полинуклеотиды, кодирующие эти ферменты, а также получение и использование этих полинуклеотидов и полипептидов. Гидролазные активности полипептидов и пептидов по изобретению включают липазную активность (гидролиз липидов), реакции переэтерификации, синтез сложных эфиров, реакции переэтерификации, активность ацилгидролаз липидов (АГЛ) и связанную ферментативную активность. Для целей этой патентной заявки реакции переэтерификации могут включать реакции ацидолиза (включая реакцию жирной кислоты и триацилглицерида), алкоголиз (включая реакцию спирта и триацилглицерида), алкоголиз (включая реакцию глицерина и триацилглицерида) и реакции трансэтерификации (включая реакцию сложного эфира и триацилглицерида). Полипептиды по изобретению можно использовать в различных фармацевтических, сельскохозяйственных и промышленных ситуациях, включая производство косметических средств и нутрицевтиков. В другом аспекте полипептиды по изобретению используют для синтеза энантиомерно чистых хиральных продуктов.
В определенных вариантах осуществления ферменты по изобретению могут представлять собой высокоселективные катализаторы. Они могут обладать способностью к катализу реакций со стерео-, регио- и хемоселективностью, которая невозможна в стандартном химическом синтезе. В одном из вариантов осуществления ферменты по изобретению могут быть универсальными. В различных аспектах они могут функционировать в органических растворителях, действовать при предельных pH (например, при высоких pH и низких pH), предельных температурах (например, высоких температурах или низких температурах), предельных степенях засоленности (например, при высокой засоленности или низкой засоленности) и катализировать реакции с соединениями, которые структурно не связаны с их природными, физиологическими субстратами.
В одном из аспектов полипептиды по изобретению включают гидролазы, обладающие липазной, сатуразной, пальмитазной и/или стеаратазной активностью и их можно использовать, например, в биокаталитическом синтезе структурированных липидов (липидов, которые содержат определенный набор жирных кислот, распределенных определенным образом на глицериновом остове), включая любое растительное масло, например масло канолы, соевое масло, альтернативы соевого масла, альтернативы масла какао, 1,3-диацилглицериды (ДАГ), 2-моноацилглицериды (МАГ) и триацилглицериды (ТАГ), такие как 1,3-дипальмитоил-2-олеоилглицерин (POP), 1,3-дистеароил-2-олеоилглицерин (StOSt), 1-пальмитоил-2-олеоил-3-стеароилглицерин (POSt) или 1-олеоил-2,3-димиристоилглицерин (OMM), полиненасыщенные жирные кислоты (ПНЖК), полиненасыщенные жирные кислоты с длинной цепью, такие как арахидоновая кислота, докозагексаеновая кислота (ДГК) и эйкозапентаеновая кислота (ЭПК).
В определенном варианте осуществления ферменты и способы по изобретению можно использовать для удаления, добавления или замены любой жирной кислоты в композиции, например, для получения масла со сниженным содержанием насыщенных жирных кислот (например, масла «с низким содержанием насыщенных жирных кислот») или содержанием различных жирных кислот (например, превращение масла, содержащего «насыщенные» жирные кислоты в масло, содержащее альтернативные «ненасыщенные» жирные кислоты).
Примеры насыщенных жирных кислот, которые можно удалять, добавлять или «перегруппировывать» в липиде, например в масле, используя фермент или осуществляя на практике способ по изобретению, включают:
Уксусная кислота: CH3COOH
Масляная кислота: CH3(CH2)2COOH
Капроновая кислота: CH3(CH2)4СООН
Каприловая кислота: CH3(CH2)6COOH
Каприновая кислота: CH3(CH2)8СООН
Ундекановая кислота: CH3(CH2)9COOH
Лауриновая кислота: (додекановая кислота): CH3(CH2)10СООН
Миристиновая кислота: (тетрадекановая кислота):
CH3(CH2)12COOH
Пентадекановая кислота: CH3(CH2)13COOH
Пальмитиновая кислота: (гексадекановая кислота):
CH3(CH2)14COOH
Маргариновая кислота: CH3(CH2)15COOH
Стеариновая кислота (октадекановая кислота): CH3(CH2)16COOH
Арахиновая кислота (эйкозановая кислота): CH3(CH2)18COOH
Бегеновая кислота: CH3(CH2)20COOH
Примеры омега-3 ненасыщенных жирных кислот, которые можно удалять, добавлять или «перегруппировывать» в липиде, например в масле, используя фермент или осуществляя на практике способ по изобретению, включают:
α-линоленовая кислота (ALA):
CH3CH2CH=CHCH2CH=CHCH2CH=CH(CH2)7COOH
Стеаридоновая кислота (октадекатетраеновая кислота):
CH3CH2CH=CHCH2CH=CHCH2CH=CHCH2CH=CH(CH2)4COOH
Эйкозапентаеновая кислота (ЭПК):
CH3CH2CH=CHCH2CH=CHCH2CH=CHCH2CH=CHCH2CH=CH(CH2)3COOH
Докозагексаеновая кислота (ДГК):
CH3CH2CH=CHCH2CH=CHCH2CH=CHCH2CH=CHCH2CH=CHCH2CH=CH(CH2)2COOH
Примеры омега-6 ненасыщенных жирных кислот, которые можно удалять, добавлять или «перегруппировывать» в липиде, например в масле, используя фермент или осуществляя на практике способ по изобретению, включают:
Линоленовая кислота (9,12-октадекадиеновая кислота):
CH3(CH2)4CH=CHCH2CH=CH(CH2)7COOH
γ-линоленовая кислота (6,9,12-октадекатриеновая кислота):
СН3(CH2)4СН=CHCH2CH=CHCH2CH=СН(CH2)4СООН
Эйкозандиеновая кислота (11,14-эйкозадиеновая кислота):
CH3(CH2)4CH=CHCH2CH=CH(CH2)9COOH
Дигомо-γ-линоленовая кислота (8,11,14-эйкозатриеновая кислота):
CH3(CH2)4СН=CHCH2CH=CHCH2CH=СН(CH2)6СООН
Арахидоновая кислота (5,8,11,14-эйкозатетраеновая кислота):
CH3(CH2)4СН=CHCH2CH=CHCH2CH-CHCH2CH=СН(CH2)3СООН
Докозадиеновая кислота (13,16-докозадиеновая кислота):
CH3(CH2)4СН=CHCH2CH=СН(CH2)11COOH
Адреновая кислота (7,10,13,16-докозатетраеновая кислота):
CH3(CH2)4CH=CHCH2CH=CHCH2CH=CHCH2CH=CH(CH2)5COOH
Докозапентаеновая (4,7,10,13,16-докозапентаеновая кислота):
CH3(CH2)4СН=CHCH2CH=CHCH2CH=CHCH2CH=CHCH2CH=СН(CH2)2СООН
Примеры омега-9 жирных кислот, которые можно удалять, добавлять или «перегруппировывать» в липиде, например в масле, используя фермент или осуществляя на практике способ по изобретению, включают:
Олеиновая кислота (9-октадеценовая кислота):
CH3(CH2)7CH=CH(CH2)7СООН
Эйкозеноевая кислота (11-эйкозеноевая кислота):
CH3(CH2)7CH=CH(CH2)9COOH
(5Z,8Z,11Z)-эйкоза-5,8,11-триеновая кислота (5,8,11-эйкозатриеновая кислота):
CH3(CH2)7СН=CHCH2CH=CHCH2CH=СН(CH2)3СООН
Эруковая кислота (13-докозеновая кислота):
CH3(CH2)7CH=CH(CH2)11COOH
Нервоновая кислота (15-тетракозеновая кислота):
CH3(CH2)7CH=CH(CH2)13COOH.
Пальмитолеиновая кислота:
СН3(CH2)7СН=СН(CH2)5СООН
В одном из аспектов в настоящем документе предоставлены новые классы липаз, названные «сатуразами», например «пальмитазы» и «стеаратазы». Ранее в литературе термин «сатураза» использовали для обозначения фермента, который осуществляет насыщение конкретных связей в метаболическом пути, например, гидрогенизацию двойной связи (Moise, et. al., J Biol Chem, 2005, 280(30):27815-27825). Однако в настоящем документе предоставлены новые и прежде не описанные «сатуразы», где сатуразы, описанные в настоящем документе, гидролизуют сложные эфиры насыщенных жирных кислот, где гидролизованные сложные эфиры могут представлять собой сложные эфиры насыщенных жирных кислот и глицерина, умбиллиферола или других спиртов.
Также в настоящем документе предоставлены ранее не описанные «пальмитазы» и «стеаратазы», где пальмитазы и стеаратазы гидролитически отщепляют пальмитиновую кислоту и стеариновую кислоту, соответственно, например, от глицеринового остова. «Сатуразы», описанные в настоящем документе, также можно обозначить как «гидролазы насыщенных жирных кислот». Подобным образом, «пальмитазы», описанные в настоящем документе, также можно обозначить как «гидролазы пальмитата», а «стеаратазы», описанные в настоящем документе, также можно обозначить как «гидролазы стеарата».
В другом аспекте сатуразы, описанные в настоящем документе, избирательно гидролизуют по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% насыщенных жирных кислот. В другом аспекте пальмитазы, описанные в настоящем документе, избирательно гидролизуют жирные кислоты так, что по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гидролитически отщепленных жирных кислот представляют собой пальмитиновую кислоту. В другом аспекте стеаратазы, описанные в настоящем документе, избирательно гидролизуют жирные кислоты так, что по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гидролитически отщепленных жирных кислот представляют собой стеариновую кислоту.
В одном из аспектов, как показано на фиг. 5, способы использования фермента по изобретению позволяют обрабатывать липид, например, липид из соевого или другого растительного масла, для избирательного гидролитического отщепления насыщенной жирной кислоты, например, пальмитиновой или стеариновой кислоты, (например, от масла, содержащего эти насыщенные жирные кислоты) для получения «масла с низким (или сниженным) содержанием насыщенных жирных кислот», например, «масла со сниженным содержанием пальмитиновой кислоты», такого как «растительное масло со сниженным содержанием пальмитиновой кислоты», например, «соевое масло со сниженным содержанием пальмитиновой кислоты». Ферменты по изобретению также можно использовать для избирательного гидролитического отщепления любой жирной кислоты, конкретно насыщенных жирных кислот, от глицеринового остова для получения «масла с низким (или сниженным) содержанием насыщенных жирных кислот», включая избирательное гидролитическое отщепление насыщенной жирной кислоты, например пальмитиновой кислоты или стеариновой кислоты, от глицеринового остова в положении Sn1 или Sn2, в дополнение к гидролитическому отщеплению в положении Sn3 (например, гидролитическому отщеплению пальмитиновой кислоты в положении Sn3, изображенном на фиг. 5).
В одном из аспектов предоставлен образцовый синтез триглицеридов, масел или жиров с низким содержанием насыщенных жирных кислот. В этом образцовом синтезе можно использовать свободные жирные кислоты или сложные эфиры жирных кислот, в зависимости от используемого фермента. В одном из аспектов гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы по изобретению, используют для удаления или гидролитического отщепления насыщенных жирных кислот, таких как уксусная кислота, масляная кислота, капроновая кислота, каприловая кислота, каприновая кислота, ундекановая кислота, лауриновая кислота, миристиновая кислота, пентадекановая кислота, пальмитиновая кислота, маргариновая кислота, стеариновая кислота, арахиновая кислота или бегеновая кислота, от триглицерида, масла или жира. В одном из аспектов удаленные или гидролизованные жирные кислоты замещают жирными кислотами с усовершенствованной пользой для здоровья (например, со сниженной корреляцией с сердечно-сосудистыми заболеваниями) или с усовершенствованными химическими свойствами (такими как окислительная стабильность или реакционная способность) или усовершенствованными физическими свойствами (такими как температура плавления или тактильные ощущения в ротовой полости). В одном из аспектов присоединенные жирные кислоты представляют собой омега-3 ненасыщенные жирные кислоты, такие как α-линоленовая кислота, стеаридоновая кислота, эйкозапентаеновая кислота (ЭПК) или докозагексаеновая кислота (ДГК) или ПНЖК или жирные кислоты рыбьего жира. В одном из аспектов присоединенные жирные кислоты представляют собой омега-6 ненасыщенные жирные кислоты, такие как линолевая кислота, γ-линолевая кислота, эйкозадиеновая кислота, дигомо-γ-линолевая кислота, арахидоновая кислота, докозадиеновая кислота, адреновая кислота или докозапентаеновая кислота. В одном из аспектов присоединенные жирные кислоты представляют собой омега-9 ненасыщенные жирные кислоты, такие как олеинова кислота, эйкозаеновая кислота, (5Z,8Z,11Z)-эйкоза-5,8,11-триеновая кислота, эруковая кислота, невроновая кислота или пальмитолеиновая кислота. В одном из аспектов присоединенные жирные кислоты (например, омега-3, омега-6 или омега-9) присоединены посредством реакции жирных кислот с триглицеридами, маслами или жиром после удаления или гидролитического отщепления насыщенных жирных кислот посредством гидролаз, например, липаз, сатураз, пальмитаз и/или стеаратаз по изобретению. В одном из аспектов присоединенные жирные кислоты (например, омега-3, омега-6 или омега-9) присоединены посредством реакции сложных эфиров жирных кислот, включая глицериновые эфиры, или сложных этиловых или метиловых эфиров с триглицеридами, маслом или жиром после удаления или гидролитического отщепления насыщенных жирных кислот посредством гидролаз, например, липаз, сатураз, пальмитаз и/или стеаратаз по изобретению. В одном из аспектов реакцию присоединения жирных кислот (например, омега-3, омега-6 или омега-9) катализируют гидролазы или липазы, такие как неспецифическая липаза (включая нерегиоспецифическую и не обладающую специфичностью к жирной кислоте) или Sn1,3-специфическая липаза или Snl-специфическая липаза или Sn3-специфическая липаза или Sn2-специфическая липаза или липаза со специфичностью к жирной кислоте.
Способы и композиции (гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению можно использовать для получения нутрицевтиков (например, полиненасыщенных жирных кислот и масел), различных продуктов питания и пищевых добавок (например, эмульгаторов, заменителей жира, маргаринов и пастообразных смесей), косметических средств (например, эмульгаторов, кремов), средств доставки фармацевтических и лекарственных средств (например, липосом, таблеток, составов) и добавкам к животным кормам (например, полиненасыщенные жирные кислоты, такие как линолевая кислота).
В одном из аспектов липазы по изобретению могут воздействовать на флуорогенные сложные эфиры жирных кислот (ЖК), например, на умбеллифериловые эфиры ЖК. В одном из аспектов можно получить профили специфичностей к жирным кислотам липаз, полученных или модифицированных способами по изобретению, измеряя их относительные активности на ряде умбеллифериловых эфиров ЖК, таких как сложный пальмитиновый, стеариновый, олеиновый, лауриновый, ПНЖК или масляный эфир.
В одном из аспектов полипептид (например, антитело или фермент, например, липаза, сатураза, пальмитаза и/или стеаратаза) по изобретению для этих реакций иммобилизуют, например, как описано ниже. В альтернативных аспектах способы по изобретению, для осуществления которых не требуется органический растворитель, можно осуществлять при относительно высоких скоростях реакций. См., например, патенты США № 5552317; 5834259.
В определенных вариантах осуществления способы и композиции (липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению можно использовать для гидролиза (включая избирательный гидролиз) масел, таких как рыбий жир, животные и растительные масла, и липидов, таких как полиненасыщенные жирные кислоты. В одном из аспектов полипептиды по изобретению используют для получения масел с низким содержанием насыщенных жирных кислот, например, путем удаления (гидролитического отщепления) по меньшей мере одной жирной кислоты от масла; и гидролиз может представлять собой избирательный гидролиз, например удаление только конкретной жирной кислоты, такой как пальмитиновая, стеариновая или другие насыщенные жирные кислоты, или удаление жирной кислоты только из одного положения, например, из Sn1, Sn2 или Sn3. В одном из аспектов полипептиды по изобретению используют для обработки жирных кислот (таких как полиненасыщенные жирные кислоты), например, жирных кислот рыбьего жира, например, для использования в продуктах питания или в качестве пищевой или кормовой добавки, или кулинарного масла, масла для жарки или выпечки или пищевого масла. В другом варианте осуществления способы и композиции (липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению можно использовать для избирательного гидролиза насыщенных сложных эфиров относительно ненасыщенных сложных эфиров с получением кислот или спиртов. В другом варианте осуществления способы и композиции (липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению можно использовать для обработки латексов для множества различных целей, например, для обработки лактексов, используемых в композициях фиксаторов для волос для удаления неприятных запахов. В другом варианте осуществления способы и композиции (липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению можно использовать для лечения дефицита липазы у животного, например у млекопитающего, такого как человек. В другом варианте осуществления способы и композиции (липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению можно использовать для получения смазочных средств, таких как гидравлическое масло. В другом варианте осуществления способы и композиции (липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению можно использовать в получении и использовании детергентов. В другом варианте осуществления способы и композиции (липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению можно использовать в процессе химической отделки материи, волокон или пряжи. В одном из аспектов способы и композиции (липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению можно использовать для получения замедления горения материи, используя, например, галогенозамещенные карбоновые кислоты или их сложные эфиры, т.е. фторосодержащие, хлоросодержащие или бромосодержащие карбоновые кислоты или их сложный эфир. В одном из аспектов предоставлены способы создания липаз из природных библиотек.
В одном из вариантов осуществления «гидролазы» по изобретению включают полипептиды (например, антитела, ферменты) и пептиды (например, «активные центры»), обладающие любой гидролазной активностью, т.е. полипептиды по изобретению могут обладать любой гидролазной активностью, включая, например, липазную, сатуразную, пальмитазную и/или стеаратазную активность. В другом варианте осуществления «гидролазы» по изобретению включают все полипептиды, обладающие любой липазной, сатуразной, пальмитазной и/или стеаратазной активностью, включая липосинтетическую или липогидролитическую активность, т.е. полипептиды по изобретению могут обладать любой липазной, сатуразной, пальмитазной и/или стеаратазной активностью. В другом варианте осуществления липазы, сатуразы, пальмитазы и/или стеаратазы по изобретению включают ферменты, принимающие участие в биоконверсии липидов через катализ реакций гидролиза, алкоголиза, ацидолиза, этерификации и аминолиза. В одном из аспектов гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению могут гидролизовать эмульсии липидов. В одном из аспектов ферменты по изобретению могут действовать предпочтительно на связи Sn-1, Sn-2 и/или Sn-3 триацилглицеридов для высвобождения одной или нескольких жирных кислот из глицеринового остова. Например, гидролазная, липазная, сатуразная, пальмитазная и/или стеаратазная активность предоставленных в настоящем документе полипептидов включает синтез масла какао, полиненасыщенных жирных кислот (ПНЖК), 1,3-диацилглицеридов (ДАГ), 2-моноацилглицерид (МАГ) и триацилглицеридов (ТАГ). В другом варианте осуществления липазная, сатуразная, пальмитазная и/или стеаратазная активность предоставленных в настоящем документе полипептидов также содержит получение масел с низким содержанием насыщенных жирных кислот, например, соевого масла или масла канолы, посредством удаления жирной кислоты, например, пальмитиновой, олеиновой, лауриновой или стеариновой кислоты. В альтернативных аспектах ферменты по изобретению также могут гидролизовать и/или изомеризовать связи при высоких температурах, низких температурах, щелочных pH и кислых pH. В одном из аспектов гидролазы, например, липазы по изобретению представляют собой сатуразы, которые катализируют реакции гидролиза, алкоголиза, ацидолиза, этерификации и аминолиза, где карбоновая или жирная кислота в образованной или вступившей в реакцию молекуле представляет собой насыщенную жирную кислоту, такую как уксусная кислота, масляная кислота, лауриновая кислота, миристиновая кислота, пальмитиновая кислота, стеариновая кислота или арахиновая кислота. В одном из аспектов гидролаза, например липаза или сатураза по изобретению, представляет собой пальмитазу, которая катализирует реакции гидролиза, алкоголиза, ацидолиза, этерификации и аминолиза, где карбоновая или жирная кислота в образованной или вступившей в реакцию молекуле представляет собой пальмитиновую кислоту. В одном из аспектов гидролаза, например липаза или сатураза по изобретению, представляет собой стеаратазу, которая катализирует реакции гидролиза, алкоголиза, ацидолиза, этерификации и аминолиза, где карбоновая или жирная кислота в образованной или вступившей в реакцию молекуле представляет собой стеариновую кислоту.
В определенных вариантах осуществления в настоящем документе предоставлены ферменты, включая варианты ферментов гидролаз (например, «вариант липазы», «вариант сатуразы», «вариант пальмитазы» или «вариант стеаратазы») по изобретению; эти ферменты могут содержать аминокислотную последовательность, которая получена из аминокислотной последовательности «предшественника». Предшественник может содержать встречающуюся в природе гидролазу и/или рекомбинантную гидролазу. Аминокислотная последовательность варианта гидролаза «получена» из аминокислотной последовательности предшественника гидролазы посредством замены, делеции или вставки или инсерции одной или нескольких аминокислот в аминокислотной последовательности предшественника. Такую модификацию выполняют в «последовательности ДНК предшественника», которая кодирует аминокислотную последовательность предшественника липазы, а не манипулируют предшественником фермента гидролазы per se. Подходящие способы для таких манипуляций последовательностью ДНК предшественника включают способы, описанные в настоящем документе, а также способы, известные специалистам в данной области.
Создание и манипуляции с нуклеиновыми кислотами
В одном из аспектов в настоящем документе предоставлены нуклеиновые кислоты, включая экспрессирующие кассеты, такие как экспрессирующие векторы, кодирующие полипептиды (например, гидролазы, такие как липазы, сатуразы, пальмитазы и/или стеаратазы, и антитела). В другом аспекте в настоящем документе предоставлены нуклеиновые кислоты, обладающие последовательностью, как указано в SEQ ID № 1, и содержащие замены по меньшей мере одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати или двенадцати или более или всех остатков оснований, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23 (или их эквиваленты). В одном из вариантов осуществления в настоящем документе предоставлены нуклеиновые кислоты, кодирующие полипептиды, обладающие последовательностью, как указано в SEQ ID № 2, и содержащие изменения по меньшей мере одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати или двенадцати или более или всех аминокислотных остатков, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23 (или их эквиваленты).
SEQ ID № 1
ATGCTGAAACCGCCTCCCTACGGACGCCTGCTGCGCGAACTGGCCGATATC
CCGGCCATCGTGACGGCACCGTTCCGGGGCGCTGCGAAAATGGGCAAACTG
GCGGATGGCGAGCCGGTACTGGTGCTGCCCGGCTTCCTGGCCGACGACAAC
GCCACCTCGGTGCTGCGCAAGACCTTCGATGTCGCGGGCTTTGCCTGTTCG
GGCTGGGAACGCGGCTTCAACCTCGGCATTCGTGGCGACCTCGTGGACCGG
CTGGTCGACCGGCTGCGGGCGGTGTCGGAGGCGGCCGGTGGTCAGAAGGT
GATCGTGGTCGGCTGGAGCCTCGGCGGCCTCTATGCGCGCGAGCTGGGCCA
CAAGGCGCCCGAACTGATCCGGATGGTCGTCACGCTCGGCAGTCCGTTCGC
GGGCGACCTCCACGCCAACCATGCGTGGAAGATCTACGAGGCGATCAACAG
CCACACGGTCGACAACCTGCCGATCCCGGTCGATTTCCAGATTAAGCCGCC
GGTGCGCACCATCGCGGTGTGGTCGCCGCTCGACGGGGTGGTGGCGCCGG
AGACCTCGGAAGGCTCGCCCGAGCAGTCGGACGAGCGGCTAGAGCTGGCG
GTGACCCACATGGGCTTTGCCGCATCGAAGACCGGGGCCGAGGCTGTGGTC
CGGCTGGTCGCGGCGCGGCTCTAG
SEQ ID № 2 (кодируется посредством SEQ ID № 1):
1-буквенный код:
MLKPPPYGRLLRELADIPAIVTAPFRGAAKMGKLADGEPVLVLPGFLADDNATSVLR
KTFDVAGFACSGWERGFNLGIRGDLVDRLVDRLRAVSEAAGGQKVIVVGWSLGGL
YARELGHKAPELIRMVVTLGSPFAGDLHANHAWKIYEAINSHTVDNLPIPVDFQIKPP
VRTIAVWSPLDGVVAPETSEGSPEQSDERLELAVTHMGFAASKTGAEAVVRLVAAR
L-
3-буквенный код:
Met Leu Lys Pro Pro Pro Tyr Gly Arg Leu Leu Arg Glu Leu Ala Asp
Ile Pro Ala Ile Val Thr Ala Pro Phe Arg Gly Ala Ala Lys Met Gly
Lys Leu Ala Asp Gly Glu Pro Val Leu Val Leu Pro Gly Phe Leu Ala
Asp Asp Asn Ala Thr Ser Val Leu Arg Lys Thr Phe Asp Val Ala Gly
Phe Ala Cys Ser Gly Trp Glu Arg Gly Phe Asn Leu Gly Ile Arg Gly
Asp Leu Val Asp Arg Leu Val Asp Arg Leu Arg Ala Val Ser Glu Ala
Ala Gly Gly Gln Lys Val Ile Val Val Gly Trp Ser Leu Gly Gly Leu
Tyr Ala Arg Glu Leu Gly His Lys Ala Pro Glu Leu Ile Arg Met Val
Val Thr Leu Gly Ser Pro Phe Ala Gly Asp Leu His Ala Asn His Ala
Trp Lys Ile Tyr Glu Ala Ile Asn Ser His Thr Val Asp Asn Leu Pro
Ile Pro Val Asp Phe Gln Ile Lys Pro Pro Val Arg Thr Ile Ala Val
Trp Ser Pro Leu Asp Gly Val Val Ala Pro Glu Thr Ser Glu Gly Ser
Pro Glu Gln Ser Asp Glu Arg Leu Glu Leu Ala Val Thr His Met Gly
Phe Ala Ala Ser Lys Thr Gly Ala Glu Ala Val Val Arg Leu Val Ala
Ala Arg Leu
SEQ ID № 3:
ATGGCCGGCCACCAGGGCGCGCGGGGCCCCAAAGACGGTCCGCCGGCGATGGTG
ATCCCGGGCTTCCTCGCCCACGACAGGCACACGACACGATTGCGCCGGGAACTC
GCCGAGGCGGGGTTCAGGGTTCACCCCTGGCGGCAGGGCTGGAACATGGGAGCG
CGTGCCGACACGCTCGAGAAATTGAAGCGGGCAGTGGACCAGTGCGGTCATGAC
GAGCCGATCCTGCTGGTCGGCTGGAGTCTGGGCGGGCTCTACGCGAGGGAGGTC
GCGCGCGCCGAGCCGGATCAGGTGCGGGCGGTGGTCACTCTTGGTTCCCCGGTGT
CGGGCGACCGGCGCCGCTACACCAACGTGTGGAAGCTGTACGAATGGGTGGCGG
GTCACCCGGTGGACGACCCGCCGATCCCCGACAAGGAGGAAAAGCCGCCGGTGC
CGACCCTGGCTTTGTGGTCGGCGGATGACGGGATCGTCGGCGCCCCGTCGGCGCG
CGGGACTCAGTTATCTCACGACAAGGCGGTCGAGATGCGAACGAGCCACATGGG
CTTTGCCATGTCGGCGAAGAGCGCACGCTTTGTTGTCGCCGAGATCGTGAAGTTC
CTGAAGAAAACCGAAGGTTCCGAGTCGCACGATTGA
SEQ ID № 4 (кодируется посредством SEQ ID № 3):
MAGHQGARGPKDGPPAMVIPGFLAHDRHTTRLRRELAEAGFRVHPWRQGWNMGA
RADTLEKLKRAVDQCGHDEPILLVGWSLGGLYAREVARAEPDQVRAVVTLGSPVSG
DRRRYTNVWKLYEWVAGHPVDDPPIPDKEEKPPVPTLALWSADDGIVGAPSARGTQ
LSHDKAVEMRTSHMGFAMSAKSARFVVAEIVKFLKKTEGSESHD
SEQ ID № 5:
GTGAGCGAGAAAGGCGCACCCAAGGGAAGGCAGCGGCTGAAGGAGATCGGCGC
GCTTCTGTTCCACGCGCCTCGCAGCTTGGGCCATCTGGGCGCGCGCGGCCCCAAG
GACGGTCCTCCGGTGATGGTCATCCCGGGATTCCTCGCGCACGACTTGCATACGA
CGCAGTTGCGCCGGGCGCTCGCGAAGGCAGGCTTCCGAGTGCATCCGTGGCGGC
AGGGGATGAACCTTGGAGCGCGCGCCGATACGCTCGAAATTCTGAAGCGCGCGG
TGGATTCCTGCGGCTCGAGCGAGCCGATGCTGCTCGTCGGCTGGAGCCTGGGCGG
TCTCTATGCCCGGGAGATCGCGCGTGCGGAGCCGGACCGGGTGCGGGCGGTGGT
GACGATGGGATCGCCGGTGTGGGGCGACCGCAGGCGCTACACCAACGTGTGGAA
GCTGTACGAACGGATTGCCGGCCATCCGGTCGACAAGCCGCCGATCCCGGACAA
GAGCCAGAAGCCGCCGGTGCCGACTCTGGCTTTGTGGTCGCAGCATGATGGCATC
GTCGGCGCGCCCTCGGCGAGAGGGACGAAGAAGACCCGCGACAAGGCGGTCGC
CATCGACACGACTCACATGGGGTTTGCCATGTCGCCCAAGACGACGCGCGCGGC
AGTGCGTGAGATCGTGGGCTTTTTGAATGAAGTCGAAGGCGGTTCGTCACCCCGG
GCGTGA
SEQ ID № 6 (кодируется посредством SEQ ID № 5):
MSEKGAPKGRQRLKEIGALLFHAPRSLGHLGARGPKDGPPVMVIPGFLAHDLHTTQL
RRALAKAGFRVHPWRQGMNLGARADTLEILKRAVDSCGSSEPMLLVGWSLGGLYA
REIARAEPDRVRAVVTMGSPVWGDRRRYTNVWKLYERIAGHPVDKPPIPDKSQKPP
VPTLALWSQHDGIVGAPSARGTKKTRDKAVAIDTTHMGFAMSPKTTRAAVREIVGF
LNEVEGGSSPRA
SEQ ID № 7:
ATGAGGCTGCGCGAGGGGGGCGCGCTCGTATCGCGGGCCTATCGCGCCTTCGGG
CGCCTCGGCGAGCGCGGCCCGGCGGACGGGCCGCCGCTGATGGTGATCCCGGGC
TTCCTCGCCACCGATCGCACCACTTTGGGGCTGCAGCGGGCGCTGGCCAAGGGCG
GCTACAAGGTGACCGGATGGGGCATGGGCCTCAACAGCGGCGTCACCGAAGACA
TAGTCGACCGCATCGCCGCTCGGGTCGAAAGGTTTGGAGCCGGCCGCAAAGTGA
TCCTCGTCGGCTGGAGCCTCGGCGGACTCTACGCGCGCGTGGTCGCGCAGGAGC
GGCCGGATCTCGTCGACAAGGTGGTCACGCTCGGCTCGCCCTTTTCGGGCGACAG
GCGCCGCAACAACAATGTCTGGCGGCTCTACGAGTTCGTC
GCCGGCCATCCGGTCAACAGCCCGCCGATCGACAAGGACCCCGAGGTGAAGCCG
CCGGTGCCGACGCTCGCTATCTGGTCGCGGCGCGACGGCATCGTCTCTCCGGCGG
GCGCGCGCGGGCGGGAGGGAGAGCGCGACGCCGAGCTCGAGCTCGACTGCAGC
CACATGGGCTTTGCGGTCAGCGCCAGGGCTTATCCCAAGATCGTGGAGGCGGTG
CGGGCGTTTCCGGAAAACATCCGTTCGCGCTGA
SEQ ID № 8 (кодируется посредством SEQ ID № 7):
MRLREGGALVSRAYRAFGRLGERGPADGPPLMVIPGFLATDRTTLGLQRALAKGGY
KVTGWGMGLNSGVTEDIVDRIAARVERFGAGRKVILVGWSLGGLYARVVAQERPD
LVDKVVTLGSPFSGDRRRNNNVWRLYEFVAGHPVNSPPIDKDPEVKPPVPTLAIWSR
RDGIVSPAGARGREGERDAELELDCSHMGFAVSARAYPKIVEAVRAFPENIRSR
SEQ ID № 9:
ATGAAGCCGCCGCCCGGATGGATGAAGATCCGGGAGGCGGGCTCGCTCCTCGCG
CGCTTCTACCGCGCGTTCGGCAAGCTCGAGCCGCGCGGGCCGGCGGACGGGCCG
AAGCTGATGGTGATCCCGGGTTTCCTCGCGGGCGACAGGACGACGCTCGGGCTG
CAGCGAGCGCTGGCCGGCGGCGGCTACCGGGTCGCCGGCTGGGGGCTGGGGGTG
AACCGCGGCGTTTCGGAGGACGTGGTCGACCGGATCGGCCAGCAAGTCGCGCGG
TTCGGGGCGGGCGAGAAGGTGATCCTGGTCGGCTGGAGCCTTGGCGGGCTTTAT
GCGCGCGTGGTGGCGCAGGAGCGGCCCGACCTCGTCGAGAAGGTGGTGACCTTG
GGCTCGCCGTTTTCGGGCGACCGGCGGCGCAACAACAATGTGTGGCGGCTCTATG
AGTGGGTGGCTGGGCATCCGGTGAACGATCCGCCGATCGACAAGGACCCGGCGA
AGAAGCCCCCGGTGCCGACGCTCGCGATCTGGTCGCGGCGTGATGGGATCGTGG
CGGTCGAAGGCGCGCGGGGGCGGCCGGAGGAGCGGGATGCCGAGCTGGAGATC
GATTGCAGCCACATGGGGTTTGGGGTCAGCGGCAAGGCGTTTCCCCGAATCGTA
GAGGCGGTGAAGGGGTTCTAA
SEQ ID № 10 (кодируется посредством SEQ ID № 9):
MKPPPGWMKIREAGSLLARFYRAFGKLEPRGPADGPKLMVIPGFLAGDRTTLGLQR
ALAGGGYRVAGWGLGVNRGVSEDVVDRIGQQVARFGAGEKVILVGWSLGGLYAR
VVAQERPDLVEKVVTLGSPFSGDRRRNNNVWRLYEWVAGHPVNDPPIDKDPAKKPP
VPTLAIWSRRDGIVAVEGARGRPEERDAELEIDCSHMGFGVSGKAFPRIVEAVKGF
SEQ ID № 11:
GTGTTGGTGCTGCCGGCGTTCCTCGCCAACGACCTTCCCACTTCGCTTCTCCGCAG
GACGCTGAAGGCGAACGGGTTTCGCCCGTTCGGCTGGGCGAACGGTTTCAACTTA
GGTGCACGGCCGGACACGCTCCAGCGCCTGAGCGCACGGCTCGATGCGGTGGTT
CAGGAAGCGGGCAGGCCGGTTGCATTGATCGGCTGGAGCCTTGGCGGGCTTTAT
GCCCGAGAGCTGGCGAAACGCAGGTCGGCTGAGGTGTCGGCAGTGATCACGCTC
GGCACGCCCTTCTCGGTTGACCTCAGACGCAACAACGCCTGGAAGCTGTACGAG
CTCATCAACGATCATCCTGTCGATGCCCCTCCCTTGGATGTTCAGGTCGACGCGA
AGCCACCCGTCCGAACCTTCGCTTTGTGGTCGCGTCGCGACGGGATCGTAGCGCC
CGCGAGCGCGCACGGCATGGAGGGCGAGTTCGACCAGGCGATCGAGCTGCAGTG
CACGCACAACGAGATGGTCAGTGATCCGGAGGCCCTCTCCACGATCGTTACCTTG
CTGCGGGAAAATGTTGGCTCCTGA
SEQ ID № 12 (кодируется посредством SEQ ID № 11):
MLVLPAFLANDLPTSLLRRTLKANGFRPFGWANGFNLGARPDTLQRLSARLDAVVQ
EAGRPVALIGWSLGGLYARELAKRRSAEVSAVITLGTPFSVDLRRNNAWKLYELIND
HPVDAPPLDVQVDAKPPVRTFALWSRRDGIVAPASAHGMEGEFDQAIELQCTHNEM
VSDPEALSTIVTLLRENVGS
SEQ ID № 13:
GTGAATACAGCCGACCTATTGAAGCCACCACCCGCAAGCATGACAGTTCTCGAG
GCGAGAGCGCTGCTGGACATATGCAAGATGAGCGCCCCATTGGCGCGCTTGCTA
TTCAAAAAGAACTCGCCCTGGCGCAAACAACGGGTTCTCGTAATACCTGGCTTTG
GCGCTGATGATCGCTACACCTGGCCGTTGCGCAATTTCGTCCAGGCACAGGGCTA
TGCCACGACTGGCTGGGGCCTGGGCACCAACAAGGCAGGTCTCAATATGCCGCA
TCAACTATCCGACGTCCACCCCAGATGGAAGCTAAAACCCAAGACGCCGTACCG
TGGTGAGGCGGGCGTACCTTACGTGATTGACCGCTTGATCGAACGGTTTGACGAA
TTGGCATCGACGGATCCGCAACCCATCGCACTTATAGGTTGGAGTCTGGGTGGTT
TCATGGCCCGTGAAGTTGCCCGAGAGCGCCCAAACCAGGTGAGTCAGGTTATTA
CCCTCGGTTCTCCTGTCATCGGAGGCCCAAAATACACCCTCGCTGCATCGGCTTT
CATCCGGCGCAAATACGATTTGGACTGGGTGGAGCAAGTGATCGCGGAGCGGGA
AGATCGCCCCATTACTGTTCCTATTACAGCAATAGTCAGCCAGTCTGATGGCATC
GTCGGATATTCAGCGGCAATCGATCACCACAGTCCCGCTGTGCAGCATTTACATA
TGGATGTTGCCCATTTGGGCTTTCCTTACAACACGAGGGTTTGGTCAGAAATCGC
CAATGCGCTCAACTCTTTAGAGGTGGAGAAGGAGCGTGTTTAG
SEQ ID № 14 (кодируется посредством SEQ ID № 13):
MNTADLLKPPPASMTVLEARALLDICKMSAPLARLLFKKNSPWRKQRVLVIPGFGA
DDRYTWPLRNFVQAQGYATTGWGLGTNKAGLNMPHQLSDVHPRWKLKPKTPYRG
EAGVPYVIDRLIERFDELASTDPQPIALIGWSLGGFMAREVARERPNQVSQVITLGSPV
IGGPKYTLAASAFIRRKYDLDWVEQVIAEREDRPITVPITAIVSQSDGIVGYSAAIDHH
SPAVQHLHMDVAHLGFPYNTRVWSEIANALNSLEVEKERV
SEQ ID № 15:
ATGGAGCTCGCCAAGGTCACCGCCCTGATGAAGGCCACCGCCCTCGAGATCGCG
ATCCTCACCGGCCACCTCGTCCTCTACCCCTCCGGGATCGTGGCCGAGCGCCTCG
CGGCCGCCCCCTCTTCACCGTCCTCCCCGTCCGCGGGCCCGACGGGCCGACGTCC
GGTCGTCCTGCTGCACGGTTTCGTGGACAACCGCTCGGTCTTCGTCCTGCTGCGC
CGTGCCCTCACCCGGAGCGGCCGTGACTGCGTCGAGTCGCTCAACTACTCGCCGC
TCACCTGCGACCTGCGGGCCGCCGCCGAACTGCTGGGGCGCCGGGTGGACGAGA
TCCGCGCCCGGACCGGACACGCCGAGGTCGACATCGTCGGCCACAGCCTGGGCG
GGCTCATCGCCCGTTATTACGTACAGCGTCTCGGCGGTGACAGCCGGGTGCGCAC
CCTGGTCATGCTCGGCACCCCGCACTCCGGCACCACCGTGGCCCGGCTCGCCGAC
GCGCATCCGCTGGTGCGGCAGATGCGGCCGGGTTCGGAGGTGCTGCGGGAGCTC
GCCGCGCCCTCGCCCGGCTGCCGTACCCGGTTCGTGAGCTTCTGGAGCGACCTC
GACCAGGTGATGGTGCCGGTGGACACGGCCTGCCTGGACCACCCCGACCTGCTG
GTGCACAACGTCCGGGTCAGCGGGATCGGTCATCTCGCGCTGCCGGTCCATCCCA
CGGTGGCGGCCGGGGTCCGGGAGGCCCTCGACGCGAGCGGCGCGGGGGTCCCGG
GGGTGCGGGAGGAGGGGCCCGGCGCCGGCGCCGTGGCGTGA
SEQ ID № 16 (кодируется посредством SEQ ID № 15):
MELAKVTALMKATALEIAILTGHLVLYPSGIVAERLAAAPSSPSSPSAGPTGRRPVVL
LHGFVDNRSVFVLLRRALTRSGRDCVESLNYSPLTCDLRAAAELLGRRVDEIRARTG
HAEVDIVGHSLGGLIARYYVQRLGGDSRVRTLVMLGTPHSGTTVARLADAHPLVRQ
MRPGSEVLRELAAPSPGCRTRFVSFWSDLDQVMVPVDTACLDHPDLLVHNVRVSGI
GHLALPVHPTVAAGVREALDASGAGVPGVREEGPGAGAVA
SEQ ID № 17:
GTGGCCGCCGCGGACAGCGGGACGGCGGAAGGGCAAAGGCTTCGGCCGCCGAG
CCTGTTCCTGATGCTGGCCGAGGCGAGGGGCTTGCTCGAACTGAACTCGAGCCTG
TTGTTGTCGCCGCTGTTGTTGCGGGCGCCGAAGGGCGACGGACATCCGGTGCTGG
CGCTGCCGGGCTTTCTCGCCAGCGATCTGTCGATGGCGCCGATGCGGCGCTATCT
GAAAGAACTCGGCTACGATGCCCATGCGTGGAACATGGGCCGCAATCTCGGCGG
CGTCGCGTCCAAGCGCGAAGCCTTGCGCGACCTGTTGCGGCGCATTTACAGCCAG
ACGGGCCGCAAGGTCAGCCTGGTCGGCTGGAGTCTCGGCGGCGTCTATGCGCGC
GATCTCGCTTTGCAGGCGCCCGACATGGTGCGTTCCGTGATCACGCTCGGCAGTC
CGTTTGCCAGCGACATCAGGGCGACCAACGCCACGCGGCTCTACGAGGCGCTGT
CGGGAGAAAGGGTCGACGACAATCCGGAGTTAACAGCGGCGATCGCCGGCGACC
TGCCGGTGCCGGCGACCTCGATCTATTCCCGTACCGACGGTATCGTGAACTGGCA
CACCAGCCTGCTGCGTCCTTCCGCAACGGCTGAAAACATCGAGGTTTACTTCGCC
AGCCATATCGGGCTCGGCGTCAACCCGGCAGCGCTGTGGGCGGTGGCCGACCGC
CTGGCGCAGCCCGAGGGGGAATTTAAGCATTTTGACCGGTCGGGTCCCTTTGCCA
TTGCCTATGGCCCCCCTGAAAATGCACAATCCTGA
SEQ ID № 18 (кодируется посредством SEQ ID № 17):
MAAADSGTAEGQRLRPPSLFLMLAEARGLLELNSSLLLSPLLLRAPKGDGHPVLALP
GFLASDLSMAPMRRYLKELGYDAHAWNMGRNLGGVASKREALRDLLRRIYSQTGR
KVSLVGWSLGGVYARDLALQAPDMVRSVITLGSPFASDIRATNATRLYEALSGERV
DDNPELTAAIAGDLPVPATSIYSRTDGIVNWHTSLLRPSATAENIEVYFASHIGLGVNP
AALWAVADRLAQPEGEFKHFDRSGPFAIAYGPPENAQS
SEQ ID № 19:
ATGCCGGAGCGAAACGAAGCGCAGGCCCCGCCGCGTCTTCGTCCGCCGGGGCTC
GGGCTGTTCCTCGCCGAAGCGCGGGGCATTTTCGAGCTCAACGCGAGCCTGTTGC
TGTCGCCGCTTCTGTTGCGCGCGCCGCGCGGCGACGGCCATCCGGTGCTGGCGTT
GCCGGGCTTTCTTGCCAGTGATCTATCGATGGCGCCGTTGCGCCGCTACCTCACC
GAGCTCGGCTACGACACCCACGCCTGGCGCATGGGCCGCAATGTCGGCGGCATC
GCGAAGATGCGGATCGCGCTGCTCGAGCGGCTCACGCAGATCCATGCCGAGTGC
GGCCGCAAGGTCTCGATTGTCGGCTGGAGTCTCGGCGGCGTCTATGCGCGCGACC
TCGCGTTGCAGGCGCCCGAGATGGTGCGCTACGTCGTCACCCTCGGCAGCCCCTT
CGCCAGCGACGTCCGCGCCACCAATGCGACGCGGCTCTATGAGGCGATGTCGGG
CGAAACGGTCGGCGACAATGTCGACCTCGTGCAGGCGATTGCCGGCGACCTGCC
GGTTCCCGTGACCTCGATCTATTCGAAGAGCGACGGCATCGTGAACTGGCGGACC
TGCCTGCTGCGCCCGTCCGCGACCGCCGAGAATATCGAGGTCTATTTCGCGAGCC
ATGTCGGCATCGGCGTCAATCCGGCCGCGCTGTGGGCGATCGCGGACCGGCTGG
CCCAGCGGGAAGGCGAATTCCGCCCCTTCGACCGGTCCGGTCCTTTTGCCATTGC
CTACGCGCCCCCGGAACAGGCACAATCGATCTGA
SEQ ID № 20 (кодируется посредством SEQ ID № 19):
MPERNEAQAPPRLRPPGLGLFLAEARGIFELNASLLLSPLLLRAPRGDGHPVLALPGF
LASDLSMAPLRRYLTELGYDTHAWRMGRNVGGIAKMRIALLERLTQIHAECGRKVS
IVGWSLGGVYARDLALQAPEMVRYVVTLGSPFASDVRATNATRLYEAMSGETVGD
NVDLVQAIAGDLPVPVTSIYSKSDGIVNWRTCLLRPSATAENIEVYFASHVGIGVNPA
ALWAIADRLAQREGEFRPFDRSGPFAIAYAPPEQAQSI
В настоящем документе предоставлены способы обнаружения новых последовательностей гидролаз, используя нуклеиновые кислоты по изобретению. Также предоставлены способы модификации нуклеиновых кислот по изобретению, например, с помощью технологий GSSMSM и GeneReassemblySM. Нуклеиновые кислоты по изобретению можно получить, выделить и/или манипулировать ими посредством, например, клонирования и экспрессии библиотек кДНК, амплификации транскрипта или геномной ДНК посредством ПЦР, и т.п.
Исходный источник выбранных образцовых полипептидов и нуклеиновых кислот:
При практическом осуществлении способов по изобретению гомологичные гены можно модифицировать посредством манипуляции матричной нуклеиновой кислоты, как описано в настоящем документе. Заявленный объект изобретения можно осуществлять на практике в сочетании с любым способом или протоколом или устройством, известными в данной области, которые хорошо описаны в научной и патентной литературе.
Основные способы
В определенных вариантах осуществления в настоящем документе предоставлены нуклеиновые кислоты, включая РНК, интерферирующую РНК (например, миРНК, мкРНК), антисмысловую нуклеиновую кислоту, кДНК, геномную ДНК, векторы, вирусы или их гибриды, нуклеиновые кислоты, выделенные из различных источников, созданные способами генной инженерии, амплифицированные и/или экспрессированные/созданные рекомбинантными способами. Рекомбинантные полипептиды, созданные из этих нуклеиновых кислот, можно индивидуально выделять или клонировать и тестировать на желаемую активность (например, гидролазную, такую как, например, липазная, сатуразная, пальмитазная и/или стеаратазная активность). Можно использовать любую систему экспрессии рекомбинантных генов, включая системы экспрессии в клетках бактерий, млекопитающих, дрожжей, грибов, насекомых или растений.
Альтернативно, эти нуклеиновые кислоты можно синтезировать in vitro с помощью хорошо известных способов химического синтеза, например, как описано в Adams (1983) J. Am. Chem. Soc. 105:661; Belousov (1997) Nucleic Acids Res. 25:3440-3444; Frenkel (1995) Free Radic. Biol. Med. 19:373-380; Blommers (1994) Biochemistry 33:7886-7896; Narang (1979) Meth. Enzymol. 68:90; Brown (1979) Meth. Enzymol. 68:109; Beaucage (1981) Тетра. Lett. 22:1859; патенте США № 4458066.
Способы манипулирования нуклеиновыми кислотами, такие как, например, субклонирование, мечение зондами (например, мечение со случайными праймерами с использованием полимеразы Кленова, ник-трансляция, амплификация), секвенирование, гибридизация и т.п. Подробно описаны в научной и патентной литературе, см., например, Sambrook, ed., MOLECULAR CLONING: A LABORATORY MANUAL (2ND ED.), Vols. 1-3, Cold Spring Harbor Laboratory, (1989); CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, Ausubel, ed. John Wiley & Sons, Inc., New York (1997); LABORATORY TECHNIQUES IN BIOCHEMISTRY AND MOLECULAR BIOLOGY: HYBRIDIZATION WITH NUCLEIC ACID PROBES, Part I. Theory and Nucleic Acid Preparation, Tijssen, ed. Elsevier, N.Y. (1993).
Другими эффективными средствами получения и манипулирования нуклеиновыми кислотами, используемыми при практическом осуществлении способов по изобретению, являются клонирование из геномных образцов и, при желании, скрининг и повторное клонирование вставленных фрагментов, выделенных или амплифицированных, например, из геномных клонов или клонов кДНК. Источники нуклеиновых кислот, используемые в способах по изобретению, включают геномные библиотеки или библиотеки кДНК, содержащиеся, например, в искусственных хромосомах млекопитающих (MAC), см., например, патенты США №№ 5721118; 6025155; искусственных хромосомах человека, см., например, Rosenfeld (1997) Nat. Genet. 15:333-335; искусственных хромосомах дрожжей (YAC); искусственных хромосомах бактерий (BAC); искусственных хромосомах P1, см., например, Woon (1998) Genomics 50:306-316; векторах, полученных из P1 (PAC), см., например, Kern (1997) Biotechniques 23:120-124; космидах, рекомбинантных вирусах, фагах или плазмидах.
Фразы «нуклеиновая кислота» или «последовательность нуклеиновой кислоты» могут включать олигонуклеотид, нуклеотид, полинуклеотид или любой их фрагмент, ДНК или РНК (например, мРНК, рРНК, тРНК, интерферирующую РНК) геномного или синтетического происхождения, которые могут быть одноцепочечными или двухцепочечными и могут представлять смысловую или антисмысловую цепь, пептидную нуклеиновую кислоту (ПНК), или любое ДНК-подобное или РНК-подобное вещество, естественного или синтетического происхождения, включая, например, интерферирующую РНК (двухцепочечную «интерферирующую» РНК), рибонуклеопротеиды (например, iRNP). Термин охватывает нуклеиновые кислоты, т.е., олигонуклеотиды, содержащие известные аналоги природных нуклеотидов. Термин также охватывает структуры с синтетическими остовами, похожие на нуклеиновые кислоты, см., например, Mata (1997) Toxicol. Appl. Pharmacol. 144:189-197; Strauss-Soukup (1997) Biochemistry 36:8692-8698; Samstag (1996) Antisense Nucleic Acid Drug Dev 6:153-156.
Как применяют в настоящем документе, термин «промоторы» включает все последовательности, способные управлять транскрипцией кодирующей последовательности в клетке, например, клетке растения. Таким образом, промоторы, используемые в конструкциях по изобретению, включают транскрипционные управляющие элементы, действующие в цис-положении, и регуляторные последовательности, которые участвуют в регуляции или модуляции регулирования по времени и/или скорости транскрипции гена. Например, промоторы могут представлять собой транскрипционный управляющий элемент, действующий в цис-положении, включая энхансер, промоторы, терминатор транскрипции, участок начала репликации, последовательность интеграции в хромосому, 5' и 3'-нетранслируемые области или интронные последовательности, которые участвуют в регуляции транскрипции. Эти последовательности, действующие в цис-положении, как правило, взаимодействуют с белками или другими биологическими молекулами для осуществления (включения/выключения, регуляции, модуляции и т.д.) транскрипции. «Конститутивные» промоторы представляют собой промоторы, которые непрерывно управляют экспрессией в большинстве состояний окружающей среды и состояний развития или дифференциации клеток. «Индуцибельные» или «регулируемые» промоторы управляют экспрессией нуклеиновой кислоты по изобретению под влиянием условий окружающей среды или условий развития. Примеры условий окружающей среды, которые могут воздействовать на транскрипцию посредством индуцируемых промоторов, включают анаэробные условия, повышенную температуру, низкое содержание влаги или присутствие света.
«Тканеспецифичные» промоторы представляют собой транскрипционные управляющие элементы, которые активны только в части клеток или тканей или органов, например, в растениях или животных. Тканеспецифическую регуляцию можно осуществлять посредством определенных внутренних факторов, которые обеспечивают экспрессию генов, кодирующих белки, специфичные для данной ткани. Известно, что для того, чтобы конкретной ткани предоставить возможность развития, у млекопитающих и растений существуют такие факторы.
Термин «растение» включает целые растения, части растений (например, листья, стебли, цветки, корни и т.д.), протопласты растений, семена и клетки растений и их потомство. Класс растений, которые можно использовать в способе по изобретению, как правило, соответствует классу высших растений, восприимчивых к способам трансформации, включая покрытосемянные растения (односемядольные и двусемядольные растения), а также голосемянные растения. Он включает растения различной степени плоидности, включая полиплоидные, диплоидные, гаплоидные и гемизиготные состояния. Как применяют в настоящем документе, термин «трансгенное растение» включает растения или клетки растения, в которые введена гетерологичная последовательность нуклеиновой кислоты, например, нуклеиновые кислоты и различные рекомбинантные конструкции (например, экспрессирующие кассеты) по изобретению.
В одном из аспектов собирают нуклеиновую кислоту, кодирующую полипептид по изобретению, в соответствующей фазе и лидерную последовательность, которая может управлять секрецией транслированного полипептида или его фрагмента.
В одном из вариантов осуществления в настоящем документе предоставлены слитые белки и кодирующие их нуклеиновые кислоты. Полипептид по изобретению можно слить с гетерологичным пептидом или полипептидом, таким как N-концевые сигнальные пептиды, которые сообщают желаемые характеристики, такие как увеличенная стабильность или упрощенная очистка. Пептиды и полипептиды по изобретению также можно синтезировать и экспрессировать в виде слитых белков с присоединенным к ним одним или несколькими дополнительными доменами, например, для получения более иммуногенного пептида, для более легкого выделения пептида, синтезированного рекомбинантными способами, для идентификации и выделения антитела и В-клеток, экспрессирующих антитела, и т.п. Домены, содействующие определению и очистке, включают, например, пептиды, хелатирующие металлы, такие как полигистидиновые тракты и гистидин-триптофановые модули, которые предоставляют возможность очистки на иммобилизованных металлах, домены белка A, которые предоставляют возможность очистки на иммобилизованном иммуноглобулине, и домены, которые используют в системе экспрессии/аффинной очистки с FLAGS (Immunex Corp, Seattle WA). Включение последовательности расщепляемого линкера, таких как расщепляемые последовательности фактора Xa или энтерокиназы (Invitrogen, San Diego CA), между доменом очистки и мотив-содержащим пептидом или полипептидом, может содействовать очистке. Например, экспрессирующий вектор может содержать последовательность нуклеиновой кислоты, кодирующую эпитоп, которая соединена с шестью остатками гистидина, за которыми следует тиоредоксин и сайт расщепления энтерокиназы (см., например, Williams (1995) Biochemistry 34:1787-1797; Dobeli (1998) Protein Expr. Purif. 12:404-414). Остатки гистидина облегчают определение и очистку, тогда как сайт расщепления энтерокиназы обеспечивает средство очистки эпитопа от остальной части слитого белка. Способы, относящиеся к векторам, кодирующим слитые белки, и к применению слитых белков хорошо описаны в научной и патентной литературе, см., например, Kroll (1993) DNA Cell. Biol., 12:441-53.
Последовательности, управляющие транскрипцией и трансляцией
В другом варианте осуществления в настоящем документе предоставлены последовательности нуклеиновых кислот (например, ДНК, иРНК), функционально связанные последовательностью(ями), управляющей экспрессией (например, транскрипцией или трансляцией), например, промоторы или энхансеры, для управления или модуляции синтеза РНК/экспрессии. Последовательность, управляющая экспрессией, может представлять собой экспрессирующий вектор. Образцовые бактериальные промоторы включают lacI, lacZ, ТЗ, Т7, gpt, lambda PR, PL и trp. Образцовые эукариотические промоторы включают предранний промотор CMV, тимидинкиназу HSV, ранний и поздний SV40, LTR из ретровирусов и металлотионеин мыши.
Промоторы, подходящие для экспрессии полипептида в бактериях включают промоторы E. coli lac или trp, lacI промотор, промотор lacZ, промотор T3, промотор T7, промотор gpt, промотор lambda PR, промотор lambda PL, промоторы из оперонов, кодирующие гликолитические ферменты, такие как 3-фосфоглицераткиназа (PGK) и промотор кислой фосфатазы. Эукариотические промоторы включат предранний промотор CMV, промотор тимидинкиназы HSV, промоторы белков теплового шока, ранние и поздние промоторы SV40, LTR из ретровирусов и промотор металлотионеина-I мыши. Также можно использовать другие промоторы, о которых известно, что они управляют экспрессией генов в прокариотических или эукариотических клетках или их вирусах.
Тканеспецифичные промоторы растений
В одном из вариантов осуществления в настоящем документе предоставлены экспрессирующие кассеты, которые можно экспрессировать тканеспецифическим образом, например, которые позволяют экспрессировать гидролазу по изобретению тканеспецифическим образом. В другом варианте осуществления в настоящем документе предоставлены растения или семена, которые экспрессируют гидролазу по изобретению, тканеспецифическим образом. Тканеспецифичность может представлять собой специфическое действие в семенах, стеблях, листьях, корнях, фруктах и т.п.
В одном из аспектов можно использовать конститутивный промотор, такой как промотор CaMV 35S, для экспрессии в конкретных частях или семенах или во всем растении. Например, для повышенной экспрессии гидролазы по изобретению можно использовать фрагмент промотора растения, который будет управлять экспрессией нуклеиновой кислоты в некоторых или во всех тканях растения, например, регенерирующего растения. Такие «конститутивные» промоторы активны в большинстве условий окружающей среды и состояниях развития или дифференциации клеток. Примеры конститутивных промоторов включают участок инициации транскрипции вируса мозаики цветной капусты (CaMV) 35S, 1'- или 2'-промоторы, полученные из T-ДНК Agrobacterium tumefaciens, и другие известные специалистам участки инициации транскрипции из различных генов растений. Такие гены включают, например, ACT11 из Arabidopsis (Huang (1996) Plant Mol. Biol. 33:125-139); Cat3 из Arabidopsis (GenBankNo. U43147, Zhong (1996) Mol. Gen. Genet. 251:196-203); ген, кодирующий десатуразу белка-переносчика стеароил-ацила из Brassica napus (Genbank No. X74782, Solocombe (1994) Plant Physiol 104:1167-1176); GPc1 из маиса (GenBank No. XI5596; Martinez (1989) J. Mol. Biol 208:551-565); Gpc2 из маиса (GenBank No. U45855, Manjunath (1997) Plant Mol. Biol. 33:97-112); промоторы растений, описанные в в патентах США №№ 4962028; 5633440.
В одном из вариантов осуществления в настоящем документе предоставлены тканеспецифичные или конститутивные промоторы, полученные из вирусов, которые могут включать, например, субгеномные промоторы тобамовирусов (Kumagai (1995) Proc. Natl. Acad. Sci. USA 92:1679-1683; палочковидный вирус риса тунгро (RTBV), который реплицируется только в клетках флоэмы инфицированных растений риса, промоторы которого управляют выраженной специфичной для флоэмы экспрессией репортерного гена; промотор вируса мозаики жилок маниоки (CVMV), с наивысшей активностью в элементах жилкования, в мезофильных клетках листьев и в кончиках корней (Verdaguer (1996) Plant Mol. Biol. 31:1129-1139).
Альтернативно промотор растения может управлять экспрессией экспрессирующей гидролазу нуклеиновой кислоты в конкретной ткани, органе или типе клеток (т.е. тканеспецифичные промоторы) или может иным образом находиться под более точным управлением со стороны окружающей среды или состояния развития или находиться под управлением индуцируемого промотора. Примеры условий окружающей среды, которые могут воздействовать на транскрипцию, включают анаэробные условия, повышенную температуру, присутствие света или опрыскивание химическими веществами/гормонами. В одном из вариантов осуществления в настоящем документе предоставлены индуцируемые низким содержанием влаги промоторы маиса (Busk (1997), выше); индуцируемые холодом, низким содержанием влаги и высокой соленостью промоторы картофеля (Kirch (1997) Plant Mol. Biol. 33:897 909).
Тканеспецифичные промоторы могут содействовать транскрипции только в рамках определенного временного интервала стадии развития в пределах этой ткани. См., например, Blazquez (1998) Plant Cells 10:791-800, характеристику промотора гена LEAFY из Arabidopsis. Также см. Cardon (1997) Plant J 12:367-77, описание фактора транскрипции SPL3, который распознает последовательность консервативного мотива в участке промотора идентифицирующего гена AP1 меристемы цветков A. thaliana; и Mandel (1995) Plant Molecular Biology, том 29, стр. 995-1004, описание промотора меристемы eIF4. Можно использовать тканеспецифичные промоторы, которые активны на всем протяжении жизненного цикла конкретной ткани. В одном из аспектов нуклеиновые кислоты по изобретению функционально связаны с промотором, который активен, главным образом, только в клетках хлопкового волокна. В одном из аспектов нуклеиновые кислоты по изобретению функционально связаны с промотором, который активен, главным образом, в ходе стадий удлинения клеток хлопкового волокна, например, как описано автором Rinehart (1996) выше. Нуклеиновые кислоты могут быть функционально связаны с промотором гена Fb12A, чтобы экспрессировать предпочтительно в клетках хлопкового волокна (Ibid). Также см., John (1997) Proc. Natl. Acad. Sci. USA 89:5769-5773; John, et al., патенты США №№ 5608148 и 5602321, описание специфических промоторов хлопкового волокна и способов конструирования трансгенных растений хлопка. Для экспрессии нуклеиновых кислот по изобретению также можно использовать специфические промоторы корней. Примеры специфических промоторов корней включают промотор из гена алкогольдегидрогеназы (DeLisle (1990) Int. Rev. Cytol. 123:39-60). Другие промоторы, которые можно использовать для экспрессии нуклеиновых кислот по изобретению, включают, например, специфические промоторы семяпочки, зародыша, эндосперма, интегумента, семенной оболочки или какое-либо их сочетание; специфический промотор листьев (см., например, Busk (1997) Plant J. 11:1285 1295, описание специфического промотора листьев в маисе); промотор ORF13 из Agrobacterium rhizogenes (который проявляет высокую активность в корнях, см., например, Hansen (1997) выше); специфический промотор пыльцы маиса (см., например, Guerrero (1990) Mol. Gen. Genet. 224:161 168); промотор томата, активный в процессе созревания плодов, увядания и опадения листьев и, в меньшей степени, цветков, можно использовать (см., например, Blume (1997) Plant J. 12:731 746); специфический промотор пестика из гена SK2 картофеля (см., например, Ficker (1997) Plant Mol. Biol. 35:425 431); ген Blec4 гороха, который активен в эпидермальной ткани верхушек вегетативных побегов и цветоносных побегов трансгенной люцерны, что делает его эффективным инструментом для нацеленной экспрессии чужеродных генов в эпидермальном слое активно растущих побегов или волокон; специфический ген BEL1 семяпочки (см., например, Reiser (1995) Cell 83:735-742, GenBankNo. U39944); и/или, промотор в Klee, патент США № 5589583, описание промоторного участка растения, который способен обеспечить высокие уровни транскрипции в меристематической ткани и/или быстрое деление клеток.
Альтернативно промоторы растений, которые можно индуцировать под воздействием гормонов растений, таких как ауксины, используют для экспрессии нуклеиновых кислот по изобретению. В одном из вариантов осуществления в настоящем документе предоставлены промоторы, содержащие чувствительные к ауксину элементы фрагмента промотора E1 (AuxREs) в соевых бобах (Glycine max L.) (Liu (1997) Plant Physiol. 115:397-407); чувствительный к ауксину промотор GST6 из Arabidopsis (также чувствителен к салициловой кислоте и пероксиду водорода) (Chen (1996) Plant J. 10: 955-966); индуцируемый ауксином промотор parC из табака (Sakai (1996) 37:906-913); чувствительный к биотину элемент растений (Streit (1997) Mol. Plant Microbe Interact. 10:933-937); и промотор, чувствительный к гормону стресса абсцизиновой кислоте (Sheen (1996) Science 274:1900-1902).
Нуклеиновые кислоты по изобретению также могут быть функционально связаны с промоторами растений, которые можно индуцировать воздействием химических веществ, которые можно наносить на растение, такие как гербициды или антибиотики. Например, можно использовать промотор маиса In2-2, активируемый антидотами бензолсульфонамидых гербицидов (De Veylder (1997) Plant Cell Physiol. 38:568-577); применение различных антидотов гербицидов индуцирует четко выраженные профили генной экспрессии, включая экспрессию в корнях, гидратодах и апикальной меристеме побегов. Кодирующие последовательности могут находиться под управлением, например, индуцируемого тетрациклином промотора, например, как описано для трансгенных растений табака, содержащих ген аргининдекарбоксилазы Avena sativa L. (овес) (Masgrau (1997) Plant J. 11:465-473); или элемента, чувствительного к салициловой кислоте (Stange (1997) Plant J. 11:1315-1324). Используя индуцируемые химическими веществами (например, гормонами или пестицидами) промоторы, т.е. промоторы, чувствительные к химическим веществам, которые можно наносить на трансгенное растение на поле, на конкретной стадии развития растения можно индуцировать экспрессию полипептида по изобретению. В определенных вариантах осуществления в настоящем документе предоставлены трансгенные растения, содержащие индуцибельный ген, кодирующий полипептиды по изобретению, круг организмов-хозяев для которого ограничен целевыми видами растений, такими как кукуруза, рис, ячмень, пшеница, картофель и другие сельскохозяйственные культуры, которые можно индуцировать на любой стадии развития сельскохозяйственной культуры.
Тканеспецифичные промоторы растений могут управлять экспрессией функционально связанных последовательностей в тканях, отличающихся от целевой ткани. Таким образом, тканеспецифичный промотор представляет собой промотор, который управляет экспрессией предпочтительно в целевой ткани или типе клеток, а также может вести к некоторой экспрессии в других тканях.
Нуклеиновые кислоты по изобретению также могут быть функционально связаны с промоторами растений, которые можно индуцировать воздействием химических веществ. Эти вещества включают, например, гербициды, синтетические ауксины или антибиотики, которые можно наносить, например, распылять на трансгенные растения. Индуцируемая экспрессия нуклеиновых кислот для получения гидролаз по изобретению, позволяет растениеводу отбирать растения с оптимальным соотношением крахмал:сахар. Таким образом, можно контролировать развитие частей растения.
В одном из вариантов осуществления в настоящем документе предоставлены средства для облегчения уборки растений или частей растений. Например, в различных вариантах осуществления используют промотор маиса In2-2, активированный антидотами бензолсульфонамидных гербицидов, (De Veylder (1997) Plant Cell Physiol. 38:568-577); нанесение различных антидотов гербицидов индуцирует четко выраженные профили генной экспрессии, включая экспрессию в корнях, гидратодах и апикальной меристеме побегов. Кодирующие последовательности по изобретению также находятся под управлением индуцируемого тетрациклином промотора, например, как описано для трансгенных растений табака, содержащих ген аргининдекарбоксилазы Avena sativa L. (овес) (Masgrau (1997) Plant J. 11:465-473); или элемента, чувствительного к салициловой кислоте (Stange (1997) Plant J. 11:1315-1324).
При желании добиться правильной экспрессии полипептида в 3'-конец кодирующего участка следует включать участок полиаденилирования. Участок полиаденилирования можно получить из естественного гена, из множества других генов растений или из генов в T-ДНК из Agrobacterial.
Экспрессирующие векторы и переносчики для клонирования
В одном из вариантов осуществления в настоящем документе предоставлены экспрессирующие векторы, экспрессирующие кассеты и переносчики для клонирования, которые содержат нуклеиновые кислоты, например, последовательности, кодирующие гидролазы и антитела. Экспрессирующие векторы и переносчики для клонирования по изобретению могут включать вирусные частицы, бакуловирусы, фаги, плазмиды, фагемиды, космиды, фосмиды, искусственные хромосомы бактерий, вирусную ДНК (например, коровьей оспы, аденовируса, вируса оспы кур, вируса псевдобешенства и производных SV40), искусственные хромосомы на основе P1, плазмиды дрожжей, искусственные хромосомы дрожжей и любые другие векторы, обладающие специфичностью к конкретным хозяевам, представляющим интерес (такие как бакуловирусы, Aspergillus и дрожжи). Векторы по изобретению могут включать хромосомные, нехромосомные и синтетические последовательности ДНК. Специалистам в данной области известно большое число подходящих и коммерчески доступных векторов. Образцовые векторы включают: бактериальные: векторы pQE (Qiagen), плазмиды pBLUESCRIPT™, векторы pNH, (векторы lambda-ZAP (Stratagene); ptrc99a, pKK223-3, pDR540, pRIT2T (Pharmacia); эукариотические: pXT1, pSG5 (Stratagene), pSVK3, pBPV, pMSG, pSVLSV40 (Pharmacia). Однако любую другую плазмиду или другой вектор можно использовать при условии, что они поддаются репликации и сохраняют жизнеспособность в организме-хозяине. Можно использовать низкокопийные или высококопийные векторы.
В одном из вариантов осуществления «экспрессирующая кассета» по изобретению содержит нуклеотидную последовательность, которая позволяет осуществлять экспрессию структурного гена (т.е. последовательности, кодирующей белок, такой как гидролаза по изобретению) в организме-хозяине, совместимом с такими последовательностями. Экспрессирующие кассеты включают по меньшей мере промоторы, функционально связанные с последовательностью, кодирующей полипептид; и, необязательно, с другими последовательностями, например, сигналами терминации транскрипции. Также можно использовать дополнительные факторы, необходимые или полезные для осуществления экспрессии, например, энхансеры. «Функционально связанный», как применяют в настоящем документе, относится к присоединению промотора, расположенного в направлении считывания выше последовательности ДНК так, что промотор опосредует транскрипцию последовательности ДНК. Таким образом, экспрессирующие кассеты также включают плазмиды, экспрессирующие векторы, рекомбинантные вирусы, любые формы рекомбинантных векторов из «оголенной ДНК», и т.п. «Вектор» включает нуклеиновую кислоту, с помощью которой можно осуществлять инфекцию, трансфекцию, транзиторную или постоянную трансдукцию клетки. Следует признать, что вектор может представлять собой оголенную нуклеиновую кислоту или нуклеиновую кислоту в комплексе с белком или липидом. Вектор необязательно содержит вирусные или бактериальные нуклеиновые кислоты и/или белки и/или мембраны (например, клеточную мембрану, липидную оболочку вируса и т.д.). В качестве неограничивающих примеров векторы включают репликоны (например, репликоны РНК, бактериофаги), к которым можно присоединить фрагменты ДНК и реплицировать их. Таким образом, векторы включают в качестве неограничивающих примеров РНК, автономную самореплицирующуюся кольцевую или линейную ДНК или РНК (например, плазмиды, вирусы и т.п., см., например, патент США № 5217879), и включают как экспрессирующей, так и неэкспрессирующей плазмиды. Когда в качестве организма-хозяина для «экспрессирующего вектора» описывают рекомбинантный микроорганизм или клеточную культуру, это включает внехромосомную кольцевую и линейную ДНК и ДНК, встроенную в хромосому(ы) организма-хозяина. Когда вектор взаимодействует с клеткой-хозяином, может или происходить стабильная репликация вектора клетками в процессе митоза в качестве автономной структуры или встраивание в геном организма-хозяина.
Экспрессирующий вектор может содержать промоторы, участок связывания рибосомы для инициации трансляции и терминатор транскрипции. Вектор также может включать соответствующие последовательности для усиления экспрессии. Экспрессирующие векторы млекопитающих могут содержать участок начала репликации, любые необходимые участки связывания рибосом, участок полиаденилирования, донорные и акцепторные участки сплайсированных фрагментов, последовательности терминации транскрипции и 5'-фланкирующие нетранскрибируемые последовательности. В некоторых аспектах для предоставления необходимых нетранскрибируемых генетических элементов можно использовать последовательности ДНК, полученные из сплайсированного фрагмента SV40, и участки полиаденилирования.
В одном из аспектов экспрессирующие векторы содержат один или несколько генов селективных маркеров, чтобы предоставить возможность отбора клеток-хозяев, содержащих вектор. Такие селективные маркеры включают гены, кодирующие дигидрофолатредуктазу или гены, обеспечивающие устойчивость культуры эукариотических клеток к неомицину, гены, обеспечивающие устойчивость E. coli к тетрациклину или ампициллину, и ген TRP1 S. cerevisiae. Промоторные участки можно выбрать из любого желаемого гена, используя векторы, содержащие ген хлорамфеникол-трансферазы (ХАТ) или другие векторы, содержащие селективные маркеры.
Векторы для экспрессии полипептида или его фрагмента в эукариотических клетках также могут содержать энхансеры для повышения уровней экспрессии. Энхансеры представляют собой элементы ДНК, действующие в цис-положении, длиной обычно приблизительно от 10 приблизительно до 300 п.о., которые действуют на промотор для увеличения его транскрипции. Примеры включают энхансер SV40, расположенный от 100 до 270 п.о. на позднем крае точки начала репликации, энхансер раннего промотора цитомегаловируса, энхансер вируса полиомы на позднем крае точки начала репликации и энхансеры аденовируса.
Последовательность ДНК можно встроить в вектор с помощью различных процедур. В основном, последовательность ДНК лигируют в желаемом положение в векторе после расщепления вставки и вектора соответствующими рестрикционными эндонуклеазами. Альтернативно можно лигировать тупые концы, как во вставке, так и в векторе. В данной области известно множество способов клонирования, например, как описано в Ausubel and Sambrook. Предполагается, что такие и другие процедуры входят в возможности специалистов в данной области.
Вектор может быть представлен в форме плазмиды, вирусной частицы или фага. Другие векторы включают хромосомные, нехромосомные и синтетические последовательности ДНК, производные SV40; бактериальные плазмиды, ДНК фагов, бакуловирус, плазмиды дрожжей, векторы, полученные сочетанием ДНК плазмид и фагов, ДНК вирусов, таких как вирус коровьей оспы, аденовирус, вирус оспы кур и вирус псевдобешенства. Различные клонирующие и экспрессирующие векторы для использования в прокариотических и эукариотических организмах-хозяевах описаны, например, автором Sambrook.
Конкретные бактериальные векторы, которые можно использовать, включают коммерчески доступные плазмиды, содержащие генетические элементы хорошо известных клонирующих векторов pBR322 (ATCC 37017), pKK223-3 (Pharmacia Fine Chemicals, Uppsala, Sweden), GEM1™ (Promega Biotec, Madison, WI, USA) pQE70, pQE60, pQE-9 (Qiagen), pDIO, psiX174 Pbluescript II KS™, pNH8A, pNH16a, pNH18A, pNH46A (Stratagene), ptrc99a, pKK223-3, pKK233-3, DR540, pRIT5 (Pharmacia), pKK232-8 и pCM7. Конкретные эукариотические векторы включают pSV2CAT, pOG44, pXT1, pSG (Stratagene) pSVK3, pBPV, pMSG и pSVL (Pharmacia). Однако можно использовать любой другой вектор при условии, что он поддается репликации и остается жизнеспособным в клетке-хозяине.
Нуклеиновые кислоты по изобретению можно экспрессировать в экспрессирующих кассетах, векторах или вирусах и можно экспрессировать транзиторно или постоянно в клетках растений и семенах. В образцовой системе транзиторной экспрессии используют системы эписомальной экспрессии, например, вирусную РНК вируса мозаики цветной капусты (CaMV), созданную в ядре путем транскрипции эписомальной минихромосомы, содержащей суперскрученную ДНК, см., например, Covey (1990) Proc. Natl. Acad. Sci. USA 87:1633-1637. Альтернативно в геном растительной клетки-хозяина можно вводить кодирующие последовательности, т.е., целые последовательности или их подфрагменты по изобретению, которые становятся составным элементом хромосомной ДНК организма-хозяина. Таким образом, можно экспрессировать смысловые или антисмысловые транскрипты. Вектор, содержащий последовательности (например, промоторные или кодирующие участки) из нуклеиновых кислот по изобретению, может содержать маркерный ген, который обуславливает фенотип клетки растения или семени, поддающийся отбору. Например, маркер может кодировать устойчивость к биоциду, конкретно устойчивость к антибиотику, такую как устойчивость к канамицину, G418, блеомицину, гигромицину или устойчивость к гербициду, такую как устойчивость к хлорсульфурону или к Basta.
Экспрессирующие векторы, позволяющие экспрессировать нуклеиновые кислоты и белки в растениях, хорошо известны в данной области и могут включать, например, векторы из Agrobacterium spp., вируса X картофеля (см., например, Angell (1997) EMBO J. 16:3675-3684), вируса табачной мозаики (см., например, Casper (1996) Gene 173:69-73), вируса кустистой карликовости томатов (см., например, Hillman (1989) Virology 169:42-50), вируса гравировки табака (см., например, Dolja (1997) Virology 234:243-252), вируса золотистой мозаики бобов (см., например, Morinaga (1993) Microbiol Immunol. 37:471-476), вируса мозаики цветной капусты (см., например, Cecchini (1997) Mol. Plant Microbe Interact. 10:1094-1101), мобильного генетического элемента Ac/Ds маиса (см., например, Rubin (1997) Mol. Cell. Biol. 17:6294-6302; Kunze (1996) Curr. Top. Microbiol. Immunol. 204:161-194), и мобильного генетического элемента супрессора-мутатора маиса (Spm) (см., например, Schlappi (1996) Plant Mol. Biol. 32:717-725); и их производных.
В одном из аспектов экспрессирующий вектор может содержать две системы репликации, чтобы предоставить возможность его взаимодействия с двумя организмами, например, для экспрессии в клетках млекопитающего, дрожжей, грибов насекомого, и для клонирования и амплификации в прокариотическом организме-хозяиние. Кроме того, для интеграции экспрессирующих векторов, экспрессирующий вектор может содержать по меньшей мере одну последовательность, гомологичную геному клетки-хозяина. Он может содержать две гомологичные последовательности, которые фланкируют экспрессируемую конструкцию. Интегрирующий вектор может быть нацелен на конкретный локус в клетке-хозяине путем выбора соответствующей гомологичной последовательности для включения в вектор. Конструкции интегрирующих векторов хорошо известны в данной области.
Экспрессирующие векторы по изобретению также могут включать ген селективного маркера, чтобы предоставить возможность отбора бактериальных штаммов, которые прошли трансформацию, например, гены, которые делают бактерии устойчивыми к лекарственным средствам, таким как ампициллин, хлорамфеникол, эритромицин, канамицин, неомицин и тетрациклин. Селективные маркеры также могут включать гены, отвечающие за биосинтез, такие как гены путей биосинтеза гистидина, триптофана и лейцина.
Клетки-хозяева и трансформированные клетки
В одном из вариантов осуществления в настоящем документе предоставлены трансформированные клетки, содержащие последовательность нуклеиновой кислоты, например, последовательность, которая кодирует гидролазу или антитело, или вектор по изобретению. Клетка-хозяин может представлять собой любую клетку-хозяина, известную специалистам в данной области, включая прокариотические клетки, эукариотические клетки, такие как бактериальные клетки, клетки грибов, клетки дрожжей, клетки млекопитающих, клетки насекомых или клетки растений.
Ферменты по изобретению можно экспрессировать в любой клетке-хозяине, например, любой бактериальной клетке, любой клетке дрожжей, любых Saccharomyces или Schizosaccharomyces spp., любых Pichia spp., например, Pichia pastoris, Saccharomyces cerevisiae или Schizosaccharomyces pombe. Образцовые бактериальные клетки включают любые Streptomyces или Bacillus spp., например, E. coli, Lactococcus lactis, Bacillus subtilis, Bacillus cereus, Salmonella typhimurium или любые виды в пределах родов Bacillus, Streptomyces и Staphylococcus. Образцовые клетки насекомых включают Drosophila S2 и Spodoptera S/9. Образцовые клетки животных включают линии клеток CHO, COS или меланомы Боуэса или любые линии клеток мыши или человека. Специалисты в данной области способны выбрать подходящий организм-хозяин. Способы трансформации широкого спектра видов высших растений хорошо известны и описаны в технической и научной литературе. См., например, Weising (1988) Ann. Rev. Genet. 22:421-477, патент США № 5750870.
Вектор можно ввести в клетки-хозяева, используя любой из множества способов, включая трансформацию, трансфекцию, трансдукцию, вирусную инфекцию, генную пушку или Ti-опосредованный перенос генов. Конкретные способы включают трансфекцию с фосфатом кальция, опосредованную ДЭАЭ-декстраном трансфекцию, липофекцию или электропорацию (Davis, L., Dibner, M., Battey, I., Basic Methods in Molecular Biology, (1986)).
Когда это целесообразно, сконструированные клетки-хозяева можно культивировать в стандартных питательных средах, модифицированных соответствующим образом для активации промоторов, отбора трансформированных клеток или амплификации генов по изобретению. После трансформации подходящего штамма-хозяина и роста штамма-хозяина до подходящей плотности клеток, выбранный промотор можно индуцировать соответствующим средством (например, сдвигом температуры или химической индукцией) и клетки можно культивировать в течение дополнительного периода, чтобы позволить им синтезировать желаемый полипептид или его фрагмент.
В одном из аспектов нуклеиновые кислоты или векторы по изобретению вводят в клетки для скрининга, тем самым нуклеиновые кислоты попадают в клетки таким способом, который подходит для последующей экспрессии нуклеиновой кислоты. Способ введения в большой степени обусловлен целевым типом клеток. Образцовые способы включают преципитацию с CaPO4, слияние липосом, липофекцию (например, LIPOFECTIN™), электропорацию, вирусную инфекцию и т.д. Подходящие нуклеиновые кислоты можно стабильно интегрировать в геном клетки-хозяина (например, используя ретровирусное введение) или они могут находиться или временно или постоянно в цитоплазме (т.е. путем использования традиционных плазмид, используя стандартные регуляторные последовательности, маркеры селекции и т.д.). Альтернативные варианты осуществления содержат ретровирусные векторы, позволяющие осуществлять трансфекцию таких мишеней (например, клеток млекопитающих, человека) поскольку, например, для многих фармацевтически важных тестов необходимы мишени человека или модельные клеточные мишени млекопитающих.
Клетки можно собирать посредством центрифугирования, разрушать физическими или химическими средствами и полученный неочищенный экстракт сохранять для дальнейшей очистки. Клетки микробов, используемые для экспрессии белков, можно разрушать любым подходящим способом, включая цикл замерзание-оттаивание, разрушение ультразвуком, механическое разрушение или использование средств для лизиса клеток. Такие способы хорошо известны специалистам в данной области. Экспрессированный полипептид или его фрагмент можно собирать и очищать от культуры рекомбинантных клеток способами, которые включают преципитацию сульфатом аммония или этанолом, экстрагирование кислотой, анионо- или катионообменную хроматографию, хроматографию на фосфоцеллюлозе, хроматографию гидрофобного взаимодействия, аффинную хроматографию, хроматографию на гидроксиапатите и хроматографию на лектине. В завершении конфигурации полипептида по мере необходимости можно использовать стадии повторной укладки белка. При желании, можно использовать высокоэффективную жидкостную хроматографию (ВЭЖХ) на конечных стадиях очистки.
Для экспрессии рекомбинантного белка также можно использовать различные системы культивирования клеток млекопитающих. Примеры систем экспрессии клеток млекопитающих включают линии COS-7 фибробластов почки обезьяны и другие клеточные линии, позволяющие экспрессировать белки из совместимого вектора, такие как клеточные линии С127, 3T3, CHO, HeLa и BHK.
Конструкции в клетках-хозяевах можно использовать стандартным образом для получения продукта гена, кодируемого рекомбинантной последовательностью. В зависимости от организма-хозяина, используемого в процедуре получения рекомбинанта, полипептиды, полученные посредством клеток-хозяев, содержащих вектор, могут быть гликозилированными или негликозилированными. полипептиды по изобретению также могут содержать или не содержать начальный аминокислотный остаток метионина.
Для получения полипептида по изобретению также можно использовать бесклеточные системы трансляции. Бесклеточные системы трансляции могут использовать мРНК, транскрибированную с ДНК-конструкции, содержащей промоторы, функционально связанные с нуклеиновой кислотой, кодирующей полипептид или его фрагмент. В некоторых аспектах, ДНК-конструкцию можно перевести в линейную форму перед проведением реакции транскрипции in vitro. Затем транскрибированную мРНК инкубируют с подходящим бесклеточным экстрактом для трансляции, таким как экстракт ретикулоцитов кролика, для получения желаемого полипептида или его фрагмента.
Экспрессирующие векторы могут содержать один или несколько генов селективных маркеров для предоставления фенотипического признака для отбора трансформированных клеток-хозяев, такие как дигидрофолатредуктаза или устойчивость к неомицин для культуры эукариотических клеток или такие как устойчивость к тетрациклину или ампициллину у E. coli.
Амплификация нуклеиновых кислот
В другом варианте осуществления нуклеиновые кислоты по изобретению, кодирующие полипептиды или модифицированные нуклеиновые кислоты можно воспроизводить, например, посредством амплификации. В одном из вариантов осуществления в настоящем документе предоставлены пары праймеров для амплификации нуклеиновых кислот, кодирующих гидролазу, например, липазу, сатуразу, пальмитазу и/или стеаратазу, где пары праймеров позволяют амплифицировать последовательности нуклеиновых кислот по изобретению. Профессионал в данной области может сконструировать пары последовательностей праймеров для амплификации для полноразмерных последовательностей или любых их частей.
Также можно использовать реакции амплификации для определения количества нуклеиновой кислоты в образце (например, количество транскрипта в образце клеток), мечения нуклеиновой кислоты (например, для использования их в чипах или блоттинге), определения нуклеиновой кислоты или определения количества конкретной нуклеиновой кислоты в образце. В одном из аспектов по изобретению амплифицируют транскрипт, выделенный из клетки или библиотеки кДНК. Специалист в данной области может выбрать и сконструировать подходящие олигонуклеотидные праймеры для амплификации. Способы амплификации также хорошо известны в данной области и включают, например, полимеразную цепную реакцию, ПЦР (см., например, PCR PROTOCOLS, A GUIDE TO METHODS AND APPLICATIONS, ed. Innis, Academic Press, N.Y. (1990) и PCR STRATEGIES (1995), ed. Innis, Academic Press, Inc., N.Y.), лигазную цепную реакцию (ЛЦР) (см., например, Wu (1989) Genomics 4:560; Landegren (1988) Science 241:1077; Barringer (1990) Gene 89:117); амплификацию транскрипцией (см., например, Kwoh (1989) Proc. Natl. Acad. Sci. USA 86:1173); и самоподдерживающуюся репликацию последовательностей (см., например, Guatelli (1990) Proc. Natl. Acad. Sci. USA 87:1874); амплификацию репликазой Q-β (см., например, Smith (1997) J. Clin. Microbiol. 35:1477-1491), автоматизированный анализ амплификации репликазой Q-β (см., например, Burg (1996) Mol. Cell. Probes 10:257-271) и другие способы, опосредованные РНК-полимеразой (например, NASBA, Cangene, Mississauga, Ontario); также см. Berger (1987) Methods Enzymol. 152:307-316; Sambrook; Ausubel; патенты США №№ 4683195 и 4683202; Sooknanan (1995) Biotechnology 13:563-564.
В одном из вариантов осуществления в настоящем документе предоставлены пары праймеров для амплификации, содержащие последовательности по изобретению, например, где пара праймеров содержит первый член, содержащий последовательность, как указано, приблизительно из первых (5') 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39 или 40 или более остатков нуклеиновой кислоты по изобретению, и второй член, содержащий последовательность, как указано, приблизительно из первых (5') 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39 или 40 или более остатков комплементарной нити первого члена.
Определение степени идентичности последовательностей
В одном из вариантов осуществления в настоящем документе предоставлены нуклеиновые кислоты, содержащие по меньшей мере нуклеиновую кислоту, или последовательность, полностью (100%) идентичную нуклеиновой кислоте по изобретению, например, образцовой нуклеиновой кислоте по изобретению (например, содержащие последовательность, как указано в SEQ ID № 1, SEQ ID № 3, SEQ ID № 5, SEQ ID № 7, SEQ ID № 9, SEQ ID № 11, SEQ ID № 13, SEQ ID № 15, SEQ ID № 17, SEQ ID № 19, SEQ ID № 22 или SEQ ID № 23, или SEQ ID № 1, модифицированную для того, чтобы кодировать замены одного, двух, трех, четырех, пяти, шести, семи, восьми или более (нескольких) или всех оснований, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, или их эквиваленты); и полипептиды, обладающие по меньшей мере 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более или полной (100%) идентичностью последовательностей с полипептидом по изобретению, например, с образцовым полипептидом, содержащим последовательность, как указано в SEQ ID № 2, SEQ ID № 4, SEQ ID № 6, SEQ ID № 8, SEQ ID № 10, SEQ ID № 12, SEQ ID № 14, SEQ ID № 16, SEQ ID № 18, или SEQ ID № 20 или SEQ ID № 2, который содержит замены одной, двух, трех, четырех, пяти, шести, семи, восьми или более (нескольких) или всех аминокислот, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23 или их эквивалент. В альтернативных аспектах последовательности могут быть идентичны на участке длиной по меньшей мере приблизительно 5, 10, 20, 30, 40, 50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000 или более последовательных остатков или с полноразмерную нуклеиновую кислоту или полипептид. Степень идентичности (гомологии) последовательностей можно определить, используя любую компьютерную программу и связанные параметры, включая те, что описаны в настоящем документе, такую как BLAST 2.2.2. или FASTA версии 3.0t78, с параметрами по умолчанию. Как применяют в настоящем документе, термины «компьютер», «компьютерная программа» и «процессор» используют в их самом широком общем смысле и они включают все такие устройства, как подробно описано ниже.
Ниже в таблице описаны выбранные характеристики образцовых нуклеиновых кислот и полипептидов по изобретению, включая сравнение идентичности последовательностей для образцовых последовательностей с общедоступными базами данных для установления активности ферментов по изобретению посредством анализа гомологии (идентичности последовательностей). Все последовательности, описанные в таблице (все образцовые последовательности по изобретению), подвергали поиску с помощью BLAST (подробно описан ниже) в двух наборах баз данных. Первый набор баз данных доступен через NCBI (National Center for Biotechnology Information). Все результаты поиска в этих базах данных в колонках, названных «НИ описание», «НИ код доступа», «НИ ожидаемое значение» или «НИ организм». «НИ» относится к неизбыточной базе данных о нуклеотидах, поддерживаемой NCBI. Эта база данных представляет собой комбинацию GenBank, обновлений GenBank и обновлений EMBL. Записи в колонке «НИ описание» относятся к строке с определением в любой заданной записи NCBI, которая содержит описание последовательности, например, организм-источник, название гена/название белка или некоторое описание функции последовательности - таким образом устанавливают активность перечисленных образцовых ферментов по изобретению посредством анализа гомологии (идентичности последовательностей). Записи в колонке «НИ код доступа» относятся к уникальному идентификатору, присвоенному записи последовательности. Записи в колонке «НИ ожидаемое значение» относятся к ожидаемому значению, которое представляет вероятность того, что оценка выравнивания равна оценке выравнивания последовательности запроса (последовательности по изобретению) и последовательности из базы данных, которая будет вычислена при том же количестве сравнений между случайными последовательностями, которое выполнено в настоящем поиске посредством BLAST. Записи в колонке «НИ организм» относятся к организму-источнику последовательности, идентифицированной как ближайшее совпадение BLAST (гомология последовательностей). Второй набор баз данных в совокупности известен как база данных GENESEQ™, которая доступна через Thomson Derwent (Philadelphia, PA). Все результаты поиска в этой базе данных в колонках, названных «GENESEQ™ описание белка», «GENESEQ™ код доступа белка», «GENESEQ™ ожидаемое значение белка», «GENESEQ™ описание ДНК», «GENESEQ™ код доступа ДНК» или «GENESEQ™ ожидаемое значение ДНК». Сведения в этих колонках сравнимы с информацией, найденной в колонках НИ, описанных выше, за исключением того, что получили из поиска посредством BLAST в базе данных GENESEQ™ вместо баз данных NCBI. Колонки «Длина ДНК-запроса» и «Длина белкового запроса» относятся к числу нуклеотидов или числу аминокислот, соответственно, в последовательности по изобретению, которую искали или посредством которой выполняли запрос в базе данных NCBI или GENESEQ™. Колонки «Длина ДНК GENESEQ™ или НИ» и «Длина белка GENESEQ™ или НИ» относятся к числу нуклеотидов или числу аминокислот, соответственно, в последовательности наилучшего совпадения в поиске посредством BLAST. Результаты, предоставленные в этих колонках, из поиска, который вернул наименьше ожидаемое значение, в базах данных NCBI или базе данных GENESEQ. Колонки «GENESEQ™/НИ %ID белка» и «GENESEQ™/НИ %ID ДНК» относятся к проценту идентичности последовательностей между последовательностью по изобретению и последовательностью наилучшего совпадения BLAST. Результаты, представленные в этих колонках, из поиска, который вернул наименьшее ожидаемое значение, в базах данных NCBI или базе данных GENESEQ™.
Гомологичные последовательности также содержат последовательности РНК, в которых уридин замещает тимин в последовательностях нуклеиновых кислот. Гомологичные последовательности можно получить, используя любые процедуры, описанные в настоящем документе, или они могут быть результатом исправления ошибки секвенирования. Следует принимать во внимание, что последовательности нуклеиновых кислот, как указано в настоящем документе, можно представлять в традиционном однобуквенном формате (см., например, Stryer, Lubert. Biochemistry, 3rd Ed., W. H Freeman & Co., New York) или в любом другом формате, который позволяет записывать идентичность нуклеотидов в последовательности.
Для сравнения последовательностей можно использовать различные программы сравнения последовательностей, указанные в настоящем документе и известные специалисту в данной области. Идентичность (гомологию) последовательности белка и/или нуклеиновой кислоты можно оценить, используя любой из множества алгоритмов и программ сравнения последовательностей, известных в данной области. Такие алгоритмы и программы включают в качестве неограничивающих примеров TBLASTN, BLASTP, FASTA, TFASTA и CLUSTALW (Pearson and Lipman, Proc. Natl. Acad. Sci. USA 85(8):2444-2448, 1988; Altschul et al., J. Mol. Biol. 215(3):403-410, 1990; Thompson et al., Nucleic Acids Res. 22(2):4673-4680, 1994; Higgins et al., Methods Enzymol. 266:383-402, 1996; Altschul et al., J. Mol. Biol. 215(3):403-410, 1990; Altschul et al., Nature Genetics 3:266-272,1993).
Гомологию или идентичность можно измерить, используя программное обеспечение для анализа последовательностей (например, Sequence Analysis Software Package of the Genetics Компьютер Group, University of Wisconsin Biotechnology Center, 1710 University Avenue, Madison, WI 53705). Такое программное обеспечение находит похожие последовательности путем определения степеней гомологии различных делеций, замен и других модификаций. Термины «гомология» и «идентичность» в отношении двух или более последовательностей нуклеиновых кислот или полипептидов относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми или обладают установленным процентным содержанием совпадающих аминокислотных остатков или нуклеотидов при сравнении и выравнивании для достижения максимального соответствия на протяжении окна сравнения или определенного участка, которые измеряют, используя любое число алгоритмов сравнения последовательностей или выравнивание вручную и визуальное исследование. Для сравнения последовательностей одна последовательность может выполнять функцию эталонной последовательности (например, образцовой последовательности нуклеиновой кислоты или полипептида по изобретению), с которой сравнивают тестовые последовательности. При использовании алгоритма сравнения последовательностей, тестовую и эталонную последовательности вводят в компьютер, при необходимости, указывают координаты подпоследовательностей, и указывают параметры программы сравнения последовательностей. Можно использовать параметры программы по умолчанию или можно указать альтернативные параметры. Затем алгоритм сравнения последовательностей вычисляет процент идентичности последовательностей для тестовых последовательностей относительно эталонной последовательности на основе параметров программы.
«Окно сравнения», как применяют в настоящем документе, относится к сегменту любого числа смежных остатков. Например, в альтернативных аспектах по изобретению после оптимального выравнивания двух последовательностей смежные остатки, которые охватывают диапазон от 20 остатков до полноразмерной образцовой последовательности полипептида или нуклеиновой кислоты, сравнивают с эталонной последовательностью с тем же числом смежных положений. Если эталонная последовательность обладает необходимой идентичностью последовательностей с образцовой последовательностью полипептида или нуклеиновой кислоты, например, в альтернативных аспектах, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, или более или полной (100%) идентичностью последовательностей с образцовой последовательностью полипептида или нуклеиновой кислоты по изобретению, то последовательность входит в объем изобретения по изобретению. В альтернативных вариантах осуществления, подпоследовательности в диапазоне приблизительно от 20 до 600, приблизительно от 50 до 200 и приблизительно от 100 до 150 сравнивают с эталонной последовательностью с тем же числом смежных положений после оптимального выравнивания двух последовательностей. В данной области хорошо известны способы выравнивания последовательностей для сравнения. Оптимальное выравнивание последовательностей для сравнения можно проводить, например, посредством алгоритма локального сходства Smith & Waterman, Adv. Appl. Math. 2:482, 1981, посредством алгоритма гомологичного выравнивания Needleman & Wunsch, J. Mol. Biol. 48:443, 1970, посредством способа поиска сходства Person & Lipman, Proc. Nat'l. Acad. Sci. USA 85:2444, 1988, посредством компьютерной реализации этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI), или посредством выравнивания вручную и визуального исследования. Другие алгоритмы для определения гомологии или идентичности включают, например, в дополнение к программе BLAST (Basic Local Alignment Search Tool в National Center for Biological Information), ALIGN, AMAS (Analysis of Multiply Aligned Sequences), AMPS (Protein Multiple Sequence Alignment), ASSET (Aligned Segment Statistical Evaluation Tool), BANDS, BESTSCOR, BIOSCAN (Biological Sequences Comparative Analysis Node), BLIMPS (BLocks IMProved Searcher), FASTA, Intervals & Points, BMB, CLUSTAL V, CLUSTAL W, CONSENSUS, LCONSENSUS, WCONSENSUS, алгоритм Smith-Waterman, DARWIN, алгоритм Las Vegas, FNAT (Forced Nucleotide Alignment Tool), Framealign, Framesearch, DYNAMIC, FILTER, FSAP (Fristensky Sequence Analysis Package), GAP (Global ALIGNMENT PROGRAM), GENAL, GIBBS, GenQuest, ISSC (Sensitive Sequence Comparison), LALIGN (Local Sequence Alignment), LCP (Local Content Program), MACAW (Multiple Alignment Construction & Analysis Workbench), MAP (Multiple ALIGNMENT PROGRAM), MBLKP, MBLKN, PIMA (Pattern-Induced Multi-sequence Alignment), SAGA (Sequence Alignment by Genetic Algorithm) и WHAT-IF. Такие программы для выравнивания также можно использовать для скрининга геномных баз данных для идентификации последовательностей полинуклеотидов, содержащих последовательности со значительным сходством. Доступно множество геномных баз данных, например, значительная часть генома человека доступна в качестве части проекта секвенирования генома человека (Gibbs, 1995). Секвенировано несколько геномов, например, M. genitalium (Fraser et al., 1995), M. jannaschii (Bult et al., 1996), H. influenzae (Fleischmann et al., 1995), E. coli (Blattner et al., 1997) и дрожжей (S. cerevisiae) (Mewes et al., 1997), и D. melanogaster (Adams et al., 2000). Также значительный прогресс достигнут в секвенировании геномов модельных организмов, таких как мышь, С. elegans и Arabadopsis sp. Различные организации поддерживают базы данных, содержащие геномную информацию, аннотированную некоторыми функциональными сведениями, и доступ к ним можно получить через интернет.
Также используют алгоритмы BLAST, BLAST 2.0 и BLAST 2.2.2. Они описаны, например, в Altschul (1977) Nuc. Acids Res. 25:3389-3402; Altschul (1990) J. Mol. Biol. 215:403-410. Программное обеспечение для осуществления анализа BLAST общедоступно через National Center for Biotechnology Information. Этот алгоритм включает сначала идентификацию пар последовательностей с высокими оценками (HSP) посредством идентификации коротких слов длины W в последовательности запроса, которые или совпадают или соответствуют некоторому положительному значению пороговой оценки T при выравнивании со словом такой же длины в последовательности их базы данных. T обозначают как порог оценки соседнего слова (Altschul (1990), выше). Эти исходные совпадения соседних слов выполняют функцию затравки для инициации поиска содержащих их более длинных HSP. Совпадения слов расширяются в обоих направлениях вдоль каждой последовательности до тех пор, пока совокупную оценку выравнивания можно увеличивать. Совокупные оценки вычисляют, используя для нуклеотидных последовательностей параметры M (награда за пару совпадающих остатков; всегда >0). Для аминокислотных последовательностей используют матрицу замен для вычисления совокупной оценки. Расширение совпадений слов в каждом направлении останавливают, когда: совокупная оценка выравнивания снижается на величину X от максимального достигнутого значения; совокупная оценка доходит до нуля или ниже вследствие накопления одного или нескольких выравниваний остатков с отрицательными оценками; или достигают конца любой из последовательностей. Параметры W, T и X алгоритма BLAST определяют чувствительность и скорость выравнивания. По умолчанию в программе BLASTN (для нуклеотидных последовательностей) используют длину слова (W), равную 11, ожидание (E), равное 10, M=5, N=-4 и сравнение обеих нитей. Для аминокислотных последовательностей в программе BLASTP по умолчанию используют длину слова, равную 3, и ожидание (E), равное 10, и выравнивания (b) с матрицей замен BLOSUM62 (см. Henikoff & Henikoff (1989) Proc. Natl. Acad. Sci. USA 89:10915), равные 50, ожидание (E), равное 10, M=5, N=-4, и сравнение обеих нитей. Также алгоритм BLAST осуществляет статистический анализ сходства между двумя последовательностями (см., например, Karlin & Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873). Одной из мер сходства, предусмотренной алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), которая указывает на вероятность случайного совпадения между двумя нуклеотидными или аминокислотными последовательностями. Например, нуклеиновую кислоту считают похожей на эталонную последовательность, если наименьшая суммарная вероятность при сравнении тестовой нуклеиновой кислоты с эталонной нуклеиновой кислотой составляет менее чем приблизительно 0,2 или альтернативно менее чем приблизительно 0,01, или альтернативно менее чем приблизительно 0,001.
В одном из аспектов гомологию последовательностей белков и нуклеиновых кислот оценивают, используя Basic Local Alignment Search Tool («BLAST»). Например, можно использовать пять специальных программ BLAST для решения следующей задачи: (1) BLASTP и BLAST3 сравнивают аминокислотную последовательность запроса с последовательностью белка из базы данных; (2) BLASTN сравнивает нуклеотидную последовательность запроса с нуклеотидной последовательностью из базы данных; (3) BLASTX сравнивает продукты трансляции в шести возможных рамках считывания нуклеотидной последовательности запроса (обе нити) с последовательностью белка из базы данных; (4) TBLASTN сравнивает белковую последовательность запроса с нуклеотидной последовательностью из базы данных, транслированной во всех шести рамках считывания (обе нити); и (5) TBLASTX сравнивает продукты трансляции во всех шести рамках считывания нуклеотидной последовательности запроса с продуктами трансляции во всех шести рамках считывания нуклеотидной последовательности из базы данных.
В одном из аспектов программы BLAST идентифицируют гомологичные последовательности путем идентификации схожих сегментов, которые обозначают в настоящем документе как «пары сегментов с высокой оценкой», между аминокислотной или нуклеотидной последовательностью запроса и тестовой последовательностью, которую альтернативно получают из последовательности нуклеиновой кислоты или белка из базы данных. Пары сегментов с высокой оценкой альтернативно можно идентифицировать (т.е., выровнять) посредством матрицы замен, многие из которых известны в данной области. В одном из аспектов используют матрицу замен BLOSUM62 (Gonnet et al., Science 256:1443-1445, 1992; Henikoff and Henikoff, Proteins 17:49-61, 1993). В одном из аспектов также можно использовать матрицы РАМ или PAM250 (см., например, Schwartz and Dayhoff, eds., 1978, Matrices for Detecting Distance Relationships: Atlas of Protein Sequence and Структур*, Washington: National Biomedical Research Foundation).
В одном из аспектов для определения того, обладает ли нуклеиновая кислота необходимой идентичностью последовательностей, чтобы входить в объем изобретения по изобретению, используют программы NCBI BLAST 2.2.2, опции по умолчанию для blastp. В программе приблизительно BLAST 2.2.2 доступно приблизительно 38 настраиваемых опций. В этом образцовом аспекте по изобретению используют все значения по умолчанию, за исключением настройки фильтрации по умолчанию (т.е., все параметры установлены по умолчанию, за исключением фильтрации, которая имеет значение OFF); в этом месте используют настройку «-F F», которая отключает фильтрацию. Использование фильтрации по умолчанию части приводит к нарушениям Карлина-Альтшуля вследствие малой длины последовательности.
Значения по умолчанию, использованные в этом образцовом аспекте по изобретению, включают:
«Фильтр для низкой сложности: ON
Размер слова: 3
Матрица: Blosum62
Цены пропусков: Присутствие: 11
Удлинение: 1»
Другие настройки по умолчанию: фильтр для низкой сложности «OFF», размер слова для белка «3», матрица «BLOSUM62», штраф за присутствие пропуска «11» и штраф за удлинение пропуска «1». В одном из аспектов опция «-W» по умолчанию равна «0». Это обозначает, что без дополнительной настройки размер слова по умолчанию равен «3» для белков и «11» для нуклеотидов.
Компьютерные системы и компьютерные программные продукты
Для определения и идентификации идентичности последовательностей, структурной гомологии, мотивов и т.п. in silico последовательность по изобретению можно хранить, записывать и манипулировать ею в среде, которая предусматривает считывание и доступ посредством компьютера. В определенных вариантах осуществления в настоящем документе предоставлены компьютеры, компьютерные системы, машиночитаемый носитель, компьютерные программные продукты и т.п., которые включают (содержат) последовательности нуклеиновых кислот и полипептидов по изобретению, записанные или хранящиеся на них. Как применяют в настоящем документе, слова «записанный» и «хранящийся» относятся к процессу хранения информации в компьютерной среде. Специалист в данной области может быстро принимать любые известные способы для записи информации на машиночитаемый носитель для создания продуктов, содержащих одну или несколько последовательностей нуклеиновой кислоты и/или полипептида по изобретению.
Другой аспект по изобретению относится к машиночитаемому носителю, на котором записана по меньшей мере одна последовательность нуклеиновой кислоты и/или полипептида по изобретению. Машиночитаемый носитель включает магнитные носители, оптические носители, электронные носители и магнитооптические носители. Например, машиночитаемый носитель может представлять собой жесткий диск, гибкий диск, магнитную ленту, ПЗУ на компакт-диске, цифровой универсальный диск (DVD), оперативное запоминающее устройство (ОЗУ) или постоянное запоминающее устройство (ПЗУ), а также другие типы других носителей, известные специалистам в данной области.
Аспекты по изобретению включают системы, (например, системы, основанные на интернете), конкретно компьютерные системы, которые хранят и манипулируют последовательностями и информацией о последовательностях, которые описаны в настоящем документе. Один пример компьютерной системы 100 изображен в виде блочной диаграммы на фиг. 1. Как применяют в настоящем документе, «компьютерная система» относится к аппаратным компонентам, программным компонентам и компонентам накопления данных, которые используют для анализа нуклеотидной или полипептидной последовательности по изобретению. Компьютерная система 100 может содержать процессор для обработки, осуществления доступа и манипуляций данными о последовательностях. Процессор 105 может относиться к любому хорошо известному типу центрального процессора, такому как, например, Pentium III корпорации Intel, или аналогичный процессор компаний Sun, Motorola, Compaq, AMD или International Business Machines. Компьютерная система 100 представляет собой систему общего назначения, которая содержит процессор 105 и один или несколько внутренних компонентов накопления данных 110 для хранения данных и один или несколько устройств извлечения данных для извлечения данных, хранящихся на компонентах накопления данных. Специалист в данной области может с легкостью принимать во внимание, что подходит любая доступная в настоящий момент компьютерная система.
В одном из аспектов компьютерная система 100 включает процессор 105, соединенный с шиной, которая соединена с основной памятью 115 (альтернативно воплощенной в виде ОЗУ) и одним или несколькими внутренними устройствами хранения данных 110, такими как жесткий диск и/или другой машиночитаемый носитель, содержащий записанные на нем данные. Дополнительно компьютерная система 100 может содержать одно или несколько устройств извлечения данных 118 для считывания данных, хранящихся на внутренних устройствах хранения данных 110. Устройство извлечения данных 118 может представлять, например, привод гибких дисков, привод компактных дисков, привод магнитной ленты или модем, способный устанавливать соединение с удаленной системой хранения данных (например, через интернет) и т.д. В некоторых вариантах осуществления внутреннее устройство хранения данных 110 представляет собой съемный машиночитаемый носитель, такой как гибкий диск, компактный диск, магнитная лента и т.д., содержащий управляющую логику и/или хранящиеся на нем данные. Преимущественно компьютерная система 100 может содержать или быть запрограммирована посредством подходящего программного обеспечения на считывание управляющей логики и/или данных с компонента накопления данных после подключения к устройству извлечения данных. Компьютерная система 100 содержит устройство отображения 120, которое используют для отображения выходных данных пользователю компьютера. Также следует отметить, что компьютерную систему 100 можно соединить с другими компьютерными системами 125a-c в сеть или глобальную вычислительную сеть, чтобы предоставить централизованный доступ к компьютерной системе 100. Во время исполнения программное обеспечение для доступа и обработки нуклеотидных или аминокислотных последовательностей по изобретению может находиться в основной памяти 115. В некоторых аспектах компьютерная система 100 дополнительно может содержать алгоритм сравнения последовательностей для сравнения последовательностей нуклеиновых кислот по изобретению. Алгоритм и последовательность(и) можно хранить на машиночитаемом носителе. «Алгоритм сравнения последовательностей» относится к одной или нескольким программам, которые реализованы (локально или удаленно) в компьютерной системе 100, для сравнения нуклеотидной последовательности с другими нуклеотидными последовательностями и/или соединениями, хранящимися в средствах накопления данных. Например, алгоритм сравнения последовательностей может сравнивать нуклеотидные последовательности по изобретению, хранящиеся на машиночитаемом носителе, с эталонными последовательностями, хранящимися на машиночитаемом носителе, для идентификации гомологии или структурных мотивов.
Параметры, используемые с указанными выше алгоритмами, можно адаптировать в зависимости от длины последовательности и исследуемой степени гомологии. В некоторых аспектах параметры могут представлять собой параметры по умолчанию, используемые алгоритмами при отсутствии инструкций пользователя. На фиг. 2 представлена блок-схема, иллюстрирующая один аспект процесса 200 для сравнения новой нуклеотидной или белковой последовательности с базой данных последовательностей для того, чтобы определить уровни гомологии между новой последовательностью и последовательностями в базе данных. База данных последовательностей может представлять собой частную базу данных, которая хранится в компьютерной системе 100, или публичную базу данных, такую как GENBANK, которая доступна через интернет. Процесс 200 начинается в начальном состоянии 201 и затем переходит к состоянию 202, где новая последовательность, подлежащая сравнению, сохраняется в памяти в компьютерной системе 100. Как указано выше, память может представлять собой память любого типа, включая ОЗУ или внутреннее запоминающее устройство. Затем процесс 200 переходит к состоянию 204, в котором открывают базу данных последовательностей для анализа и сравнения. Затем процесс 200 переходит к состоянию 206, где первую последовательность, хранящуюся в базе данных, считывают в память на компьютере. Затем в состоянии 210 осуществляют сравнение, чтобы определить, являются ли первая последовательность и вторая последовательность одинаковыми. Важно отметить, что эта стадия не ограничена выполнением строгого сравнения между новой последовательностью и первой последовательностью в базе данных. Специалистам в данной области хорошо известны способы сравнения двух нуклеотидных или белковых последовательностей, даже если они не идентичны. Например, в одну последовательность можно вводить пропуски для того, чтобы повысить уровень гомологии между двумя протестированными последовательностями. Обычно пользователь компьютерной системы вводит параметры, которые определяют введение пропусков или других особенностей в последовательность в ходе сравнения. После выполнения сравнения двух последовательностей в состоянии 210, в состоянии принятия решения 212 определяют, являются ли две последовательности одинаковыми. Конечно, термин «одинаковый» не ограничен последовательностями, которые абсолютно идентичны. Последовательности, которые попадают в рамки параметров гомологии, введенных пользователем, будут помечены как «одинаковые» в процессе 200. Если установлено, что две последовательности являются одинаковыми, то процесс 200 переходит к состоянию 214, где название последовательности из базы данных отображают пользователю. Это состояние уведомляет пользователя о том, что последовательность, название которой отображено, соответствует введенным ограничивающим условиям гомологии. После отображения названия сохраненной последовательности пользователю, процесс 200 переходит к состоянию принятия решения 218, где определяют, есть ли в базе данных дополнительные последовательности. Если в базе данных больше нет последовательностей, то процесс 200 заканчивается в состоянии 220. Однако если в базе данных присутствуют дополнительные последовательности, то процесс 200 переходит к состоянию 224, где указатель перемещают к следующей последовательности в базе данных с тем, чтобы ее можно было сравнивать с новой последовательностью. Таким образом, новую последовательность выравнивают и сравнивают с каждой последовательностью в базе данных. Следует отметить, что если в состоянии принятия решения 212 установлено, что последовательности негомологичны, то процесс 200 переходит непосредственно к состоянию принятия решения 218 для того, чтобы определить, доступны ли для сравнения другие последовательности в базе данных. Таким образом, один аспект по изобретению представляет собой компьютерную систему, содержащую процессор, устройство хранения данных, на котором хранится последовательность нуклеиновой кислоты по изобретению, и блок сравнения последовательностей для осуществления сравнения. Блок сравнения последовательностей может показывать уровень гомологии между сравненными последовательностями или идентифицировать структурные мотивы, или он может идентифицировать структурные мотивы в последовательностях, которые сравнивают с этими кодами нуклеиновых кислот и кодами полипептидов. На фиг. 3 представлена блок-схема, которая иллюстрирует один из вариантов осуществления процесса 250 в компьютере для определения гомологии двух последовательностей. Процесс 250 начинается в исходном состоянии 252 и затем переходит в состояние 254, где первую последовательность, подлежащую сравнению, сохраняют в памяти. Затем в состоянии 256 вторую последовательность, подлежащую сравнению, записывают в памяти. Затем процесс 250 переходит в состояние 260, где считывают первый символ в первой последовательности, а затем в состояние 262, где считывают первый символ второй последовательности. Следует понимать, что если последовательность представляет собой нуклеотидную последовательность, то обычно символ будет представлять собой A, T, C, G или U. Если последовательность представляет собой последовательность белка, то она может представлять собой однобуквенный аминокислотный код с тем, чтобы можно было легко сравнить первую и вторую последовательности. Затем в состоянии принятия решения 264 определяют, являются ли два символа одинаковыми. Если они одинаковы, то процесс 250 переходит к состоянию 268, где считывают следующие символы в первой и второй последовательностях. Затем определяют, являются ли следующие символы одинаковыми. Если это так, то затем процесс 250 продолжает этот цикл до тех пор, пока два символа не окажутся не одинаковыми. Если установлено, что следующие два символа не одинаковы, то процесс 250 переходит в состояние принятия решения 274 для того, чтобы определить, есть ли в последовательности дополнительные символы для считывания. Если больше нет символов для считывания, то затем процесс 250 переходит в состояние 276, где пользователю отображают уровень гомологии между первой и второй последовательностями. Уровень гомологии определяют путем вычисления доли одинаковых символов между последовательностями из общего числа последовательностей в первой последовательности. Таким образом, если каждый символ в первой 100 нуклеотидной последовательности выровнен с каждым символом во второй последовательности, то уровень гомологии составит 100%.
Альтернативно, компьютерная программа может сравнивать эталонную последовательность с последовательностью по изобретению, чтобы установить, отличаются ли последовательности одним или несколькими положениями. Программа может записывать длину и идентичность вставленных, удаленных или замененных нуклеотидов или аминокислотных остатков относительно эталонной последовательности или последовательности по изобретению. Компьютерная программа может представлять собой программу, которая определяет, содержит ли эталонная последовательность однонуклеотидный полиморфизм (SNP) относительно последовательности по изобретению, или содержит ли последовательность по изобретению, SNP известной последовательности. Таким образом, в некоторых аспектах компьютерная программа представляет собой программу, которая идентифицирует SNP. Способ можно выполнять посредством компьютерных систем, описанных выше, и способа, изображенного на фиг. 3. Способ можно выполнять посредством считывания последовательности по изобретению и эталонных последовательностей путем применения компьютерной программы и идентификации различий с использованием компьютерной программы.
В других аспектах система на основе компьютера содержит идентификатор для идентификации признаков в нуклеиновой кислоте или полипептиде по изобретению. «Идентификатор» относится к одной или нескольким программам, которые идентифицируют конкретные признаки в последовательности нуклеиновой кислоты. Например, идентификатор может содержать программу, которая идентифицирует открытую рамку считывания (ORF) в последовательности нуклеиновой кислоты. На фиг. 4 представлена блок-схема, которая иллюстрирует один аспект процесса идентификации 300 для определения присутствия признака в последовательности. Процесс 300 начинается в исходном состоянии 302 и затем переходит в состояние 304, где первую последовательность, подлежащую проверке на признаки, сохраняют в памяти 115 в компьютерной системе 100. Затем процесс 300 переходит в состояние 306, где открывают базу данных признаков последовательностей. Такая база данных содержит список атрибутов каждого признака вместе с названием признака. Например, названием признака может быть «Инициирующий кодон» и атрибутом может быть «ATG». Другим примером будет название признака «TAATAA-бокс» и атрибутом признака будет «TAATAA». Пример такой базы данных создан генетической компьютерной группой университета Висконсина. Альтернативно признаками могут являться структурные мотивы полипептидов, такие как α-спирали, β-листы или функциональные мотивы полипептидов, такие как ферментативные активные центры, мотивы спираль-поворот-спираль или другие мотивы, известные специалистам в данной области. После открытия базы данных признаков в состоянии 306, процесс 300 переходит в состояние 308, где из базы данных считывают первый признак. Затем в состоянии 310 выполняют сравнение атрибута первого признака с первой последовательностью. Затем определяют в состоянии принятия решения 316, найден ли атрибут признака в первой последовательности. Если атрибут найден, то процесс 300 переходит в состояние 318, где название найденного признака отображают пользователю. Затем процесс 300 переходит к состоянию принятия решения 320, где определяют, есть ли в базе данных дополнительные признаки. Если в базе данных больше нет признаков, то затем процесс 300 прерывается в конечном состоянии 324. Однако если в базе данных еще есть признаки, то процесс 300 считывает следующий признак последовательности в состоянии 326 и возвращается к состоянию 310, где атрибут следующего признака сравнивают с первой последовательностью. Если атрибут признака не найден в первой последовательности в состоянии принятия решения 316, то процесс 300 переходит непосредственно к состоянию принятия решения 320 для того, чтобы определить, есть ли еще признаки в базе данных. Таким образом, в одном из аспектов предусмотрена компьютерная программа, которая идентифицирует открытые рамки считывания (ORF).
Последовательность полипептида или нуклеиновой кислоты по изобретению можно хранить или манипулировать ими в различных программах обработки данных в различных форматах. Например, последовательность можно хранить в виде текста в файле текстового процессора, таком как MICROSOFTWORD™ или WORDPERFECT™, или в виде файла ASCII в различных программах баз данных, хорошо знакомых специалистам в данной области, таких как DB2, SYBASE или ORACLE™. Кроме того, многие компьютерные программы и базы данных можно использовать в качестве алгоритмов сравнения последовательностей, идентификаторов или источников эталонных нуклеотидных последовательностей или полипептидных последовательностей, подлежащих сравнению с последовательностью нуклеиновой кислоты по изобретению. Программы и базы данных могут включать: MACPATTERN™ (EMBL), DISCOVERYBASE™ (Molecular Applications Group), GENEMINE™ (Molecular Applications Group), LOOK™ (Molecular Applications Group), MACLOOK™ (Molecular Applications Group), BLAST и BLAST2 (NCBI), BLASTN и BLASTX (Altschul et al, J. Mol. Biol. 215: 403, 1990), FASTA (Pearson and Lipman, Proc. Natl. Acad. Sci. USA, 85: 2444, 1988), FASTDB™ (Brutlag et al. Сотр. Арр. Biosci. 6:237-245, 1990), CATALYST™ (Molecular Simulations Inc.), CATALYST™/SHAPE™ (Molecular Simulations Inc.), CERIUS2.DBACCESS™ (Molecular Simulations Inc.), HYPOGEN™ (Molecular Simulations Inc.), Insight II, (Molecular Simulations Inc.), DISCOVER™ (Molecular Simulations Inc.), CHARMm™ (Molecular Simulations Inc.), FELIX™ (Molecular Simulations Inc.), DELPHI™ (Molecular Simulations Inc.), QUANTEMM™, (Molecular Simulations Inc.), HOMOLOGY™ (Molecular Simulations Inc.), MODELER™ (Molecular Simulations Inc.), ISIS™ (Molecular Simulations Inc.), Quanta/Protein Design (Molecular Simulations Inc.), WEBLAB™ (Molecular Simulations Inc.), WEBLAB™ Diversity Explorer (Molecular Simulations Inc.), GENE EXPLORER™ (Molecular Simulations Inc.), SEQFOLD™ (Molecular Simulations Inc.), базу данных MDL Available Chemicals Directory, базу данных MDL Drug Data Report, базу данных Comprehensive Medicinal Chemistry, базу данных Derwent's World Drug Index, базу данных BioByteMasterFile, базу данных Genbank, и базу данных Genseqn. Специалисту в данной области известны многие другие программы и базы данных, указанные в настоящем раскрытии.
Мотивы, которые можно обнаружить, используя указанные выше программы, включают последовательности, кодирующие лейциновые молнии, мотивы спираль-поворот-спираль, сайты гликозилирования, сайты убиквитинилирования, α-спирали и β-листы, сигнальные последовательности, кодирующие сигнальные пептиды, которые направляют секрецию кодируемых белков, последовательности, вовлеченные в регуляцию транскрипции, такие как гомеобоксы, кислые фрагменты секвенирования, активные центры ферментов, сайты связывания субстрата и сайты расщепления ферментов.
Гибридизация нуклеиновых кислот
В определенных вариантах осуществления в настоящем документе предоставлены выделенные, синтетические или рекомбинантные нуклеиновые кислоты, которые гибридизуются при строгих условиях с нуклеиновой кислотой, предоставленной в настоящем документе, например, с образцовой последовательностью, предоставленной в настоящем документе, например, последовательностью, как указано в SEQ ID № 1, SEQ ID № 3, SEQ ID № 5, SEQ ID № 7, SEQ ID № 9, SEQ ID № 11, SEQ ID № 13, SEQ ID № 15, SEQ ID № 17, SEQ ID № 19, SEQ ID № 22 или SEQ ID № 23 или в SEQ ID № 1, модифицированной для того, чтобы кодировать изменение одного, двух, трех, четырех, пяти, шести, семи, восьми или более (нескольких) или всех оснований, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, или их эквиваленты, и их подпоследовательности и комплементарные последовательности, или нуклеиновая кислота, которая кодирует полипептид по изобретению. Строгие условия могут представлять собой крайне условия, условия средней строгости, условия низкой строгости, включая условия высокой и пониженной строгости, описанные в настоящем документе.
«Гибридизация» относится к процессу, посредством которого нить нуклеиновой кислоты соединяется с комплементарной нитью через спаривание оснований. Реакции гибридизации могут быть чувствительными и избирательными с тем, чтобы можно было идентифицировать конкретную последовательность, представляющую интерес, даже в образцах, в которых она присутствует в низких концентрациях. Строгие условия могут определяться, например, концентрациями соли или формамида в растворах для предварительной гибридизации и гибридизации, или температурой гибридизации, и хорошо известны в данной области. Например, строгость можно увеличить посредством снижения концентрации соли, повышения концентрации формамида или повышения температуры гибридизации, изменения времени гибридизации, как описано в подробностях ниже. В альтернативных аспектах нуклеиновые кислоты по изобретению определяют по их способности к гибридизации при условиях различной строгости (например, высокой, средней, низкой), как указано в настоящем документе.
В альтернативных вариантах осуществления длина нуклеиновых кислот по изобретению, которые определены по их способности к гибридизации при строгих условиях, может составлять приблизительно от пяти остатков до полноразмерной нуклеиновой кислоты по изобретению; например, их длина может составлять по меньшей мере 5, 10, 15, 20, 25, 30, 35, 40, 50, 55, 60, 65, 70, 75, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000 или более остатков в длину. Также включены нуклеиновые кислоты, короче полноразмерных. Эти нуклеиновые кислоты можно использовать, например, в качестве зондов для гибридизации, зондов для мечения, олигонуклеотидных зондов для ПЦР, иРНК, антисмысловых или последовательностей, кодирующих пептиды, связываемые антителами (эпитопы), мотивы, активные центры и т.п.
В одном из аспектов нуклеиновые кислоты по изобретению определяют по их способности к гибридизации при условиях высокой строгости, включая приблизительно 50% формамид при температуре приблизительно от 37°C до 42°C. В одном из аспектов нуклеиновые кислоты по изобретению определяют по их способности к гибридизации при условиях низкой строгости, включая приблизительно от 35% до 25% формамид при температуре приблизительно от 30°C до 35°C.
Альтернативно, нуклеиновые кислоты по изобретению определяют по их способности к гибридизации при условиях высокой строгости при 42°C в 50% формамиде, 5×SSPE, 0,3% SDS и блокирующей нуклеиновой кислоте с повторяющейся последовательностью, такой как cot-1 или ДНК спермы лосося (например, 200 мкг/мл резанной и денатурированной ДНК спермы лосося). В одном из аспектов нуклеиновые кислоты по изобретению определяют по их способности к гибридизации при условиях пониженной строгости, включая 35% формамид при пониженной температуре 35°C.
После гибридизации фильтр можно промыть в 6×SSC, 0,5% SDS при 50°C. Эти условия рассматривают как «умеренные» условия при концентрации формамида более 25% и как «низкие» условия при концентрации формамида ниже 25%. Конкретными примером «умеренных» условий гибридизации является указанная выше гибридизация, когда ее проводят в 30% формамиде. Конкретным примером «низкой строгости» условий гибридизации является указанная выше гибридизация, когда ее проводят в 10% формамиде.
Температурный диапазон, соответствующий конкретному уровню строгости, можно дополнительно сузить, вычислив отношение пурина к пиримидину в нуклеиновой кислоте, представляющей интерес, и, таким образом, скорректировав температуру. Нуклеиновые кислоты по изобретению также определяют по их способности к гибридизации при высокой, средней и низкой строгости условий, как указано в Ausubel and Sambrook. Изменения указанных выше диапазонов и условий хорошо известны в данной области. Ниже дополнительно обсуждаются условия гибридизации.
Указанную выше процедуру можно модифицировать для идентификации нуклеиновых кислот, обладающих снижающимися уровнями гомологии с последовательностью зонда. Например, чтобы получить нуклеиновые кислоты со снижающейся гомологией с поддающимся обнаружению зондом, можно использовать менее строгие условия. Например, температуру гибридизации можно снижать с шагом в 5°C с 68°C до 42°C в гибридизационном буфере, содержащим Na+ в концентрации приблизительно 1M. После гибридизации фильтр можно промывать в 2×SSC, 0,5% SDS при температуре гибридизации. Эти условия считаются «умеренными» условиями при температуре 50°C и «низкими» условиями при температуре ниже 50°C. Конкретным примером «умеренных» условий гибридизации является указанная выше гибридизация, когда ее проводят при температуре 55°C. Конкретным примером условий гибридизации «низкой строгости» является указанная выше гибридизация, когда ее проводят при температуре 45°C.
Альтернативно гибридизацию можно осуществлять в буферах, таких как 6×SSC, содержащих формамид, при температуре 42°C. В этом случае концентрацию формамида в гибридизационном буфере можно снижать с шагом 5% с 50% до 0%, чтобы идентифицировать клоны, обладающие снижающимися уровнями гомологии с зондом. После гибридизации фильтр можно промывать в 6×SSC, 0,5% SDS при температуре 50°C. Эти условия считаются «умеренными» условиями при концентрации формамида выше 25% и «низкими» условиями при концентрации формамида ниже 25%. Конкретным примером «умеренных» условий гибридизации является указанная выше гибридизация, когда ее проводят при концентрации формамида 30%. Конкретным примером условий гибридизации «низкой строгости» является указанная выше гибридизация, когда ее проводят при концентрации формамида 10%.
Однако выбор вида гибридизации не критичен - строгость условий отмывания описывает условия, которые определяют, входит ли нуклеиновая кислота в объем изобретения по изобретению. Условия отмывания, которые используют для идентификации нуклеиновых кислот, входящих в объем изобретения, включают, например: концентрацию соли приблизительно 0,02 М при pH 7 и температуре по меньшей мере приблизительно 50°C или приблизительно от 55°C приблизительно до 60°C; или концентрацию соли приблизительно 0,15 M NaCl при 72°C в течение приблизительно 15 минут; или концентрацию соли приблизительно 0,2×SSC при температуре по меньшей мере приблизительно 50°C или приблизительно от 55°C приблизительно до 60°C в течение приблизительно от 15 приблизительно до 20 минут; или промывание гибридизационного комплекса два раза в растворе с концентрацией соли приблизительно 2×SSC, содержащей 0,1% SDS при комнатной температуре в течение 15 минут и затем промывание дважды в 0,1×SSC, содержащем 0,1% SDS, при температуре 68°C в течение 15 минут; или эквивалентные условия. См. описание буфера SSC и эквивалентных условий в Sambrook, Tijssen and Ausubel.
Эти способы можно использовать для выделения нуклеиновых кислот по изобретению.
Олигонуклеотидные зонды и способы их использования
В определенных вариантах осуществления в настоящем документе предоставлены зонды из нуклеиновых кислот для идентификации нуклеиновых кислот, кодирующих полипептид с гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью. В одном из аспектов зонд содержит по меньшей мере 10 последовательных оснований нуклеиновой кислоты по изобретению. Альтернативно зонд по изобретению может содержать по меньшей мере приблизительно 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 110, 120, 130, 150, 160, 170, 180, 190, 200 или более или приблизительно от 10 до 50, приблизительно от 20 до 60 приблизительно от 30 до 70 последовательных оснований из последовательности, как указано в нуклеиновой кислоте по изобретению. Зонды позволяют идентифицировать нуклеиновую кислоту посредством связывания и/или гибридизации. Зонды можно использовать в чипах по изобретению, см. обсуждение ниже, включая, например, капиллярные блоки. Зонды по изобретению также можно использовать для выделения других нуклеиновых кислот или полипептидов.
Зонды по изобретению можно использовать для определения содержания в биологическом образце, таком как образец почвы, организма, содержащего последовательность нуклеиновой кислоты по изобретению (например, нуклеиновой кислоты, кодирующей гидролазу) или организма, из которого получили нуклеиновую кислоту. В таких процедурах получают биологический образец, потенциально скрывающий организм, из которого выделена нуклеиновая кислота, и из образца получают нуклеиновые кислоты. Нуклеиновые кислоты приводят в контакт с зондом в условиях, которые позволяют зонду специфически гибридизоваться с любыми комплементарными последовательностями, представленными в образце. В случае необходимости, условия, которые позволяют зонду специфически гибридизоваться с комплементарными последовательностями, можно определить, приведя зонд в контакт с комплементарными последовательностями из образцов, которые точно содержат комплементарную последовательность, а также с контрольными последовательностями, которые не содержат комплементарную последовательность. Чтобы идентифицировать условия, которые позволяют зонду гибридизоваться специфически с комплементарными нуклеиновыми кислотами, можно менять условия гибридизации, такие как концентрация соли в гибридизационном буфере, концентрация формамида в гибридизационном буфере или температура гибридизации (см. обсуждение конкретных условий гибридизации).
Если образец содержит организм, из которого выделили нуклеиновую кислоту, то определяют специфическую гибридизацию зонда. Гибридизацию можно определить путем мечения зонда поддающимся обнаружению средством, таким как радиоактивный изотоп, флуоресцентный краситель или фермент, способный катализировать образование поддающегося обнаружению продукта. Специалистам в данной области известны многие способы использования меченых зондов для определения присутствия комплементарных нуклеиновых кислот в образце. Сюда входят саузерн-блоттинг, нозерн-блоттинг, процедуры гибридизации колоний и дот-блоттинг. Протоколы для всех этих процедур предоставлены в Ausubel and Sambrook.
Альтернативно в реакции амплификации можно использовать более, чем один зонд (по меньшей мере один из которых способен специфически гибридизоваться с любыми комплементарными последовательностями, которые присутствуют в образце нуклеиновой кислоты), чтобы определить, содержится ли в образце организм, содержащий последовательность нуклеиновой кислоты по изобретению (например, организм, из которого выделена нуклеиновая кислота). В одном из аспектов зонды содержат олигонуклеотиды. В одном из аспектов реакция амплификации может включать ПЦР. Протоколы ПЦР описаны в Ausubel and Sambrook (см. обсуждение реакций амплификации). В таких процедурах нуклеиновые кислоты в образце приводят в контакт с зондами, проводят реакцию амплификации и определяют любой полученный продукт амплификации. Продукт амплификации можно определить с помощью электрофореза продуктов реакции в геле и окрашивания геля интеркалятором, таким как бромистый этидий. Альтернативно один или несколько зондов можно метить радиоактивным изотопом, а присутствие радиоактивного продукта амплификации можно определить авторадиографией после электрофореза в геле.
Зонды, полученные из последовательностей рядом с 3'- или 5'-концами последовательности нуклеиновой кислоты по изобретению, также можно использовать в процедурах прогулки по хромосоме для идентификации клонов, содержащих дополнительные, например, геномные последовательности. Такие способы позволяют выделить из организма-хозяина гены, которые кодируют дополнительные белки, представляющие интерес.
В одном из аспектов последовательности нуклеиновых кислот по изобретению используют в качестве зондов для идентификации и выделения родственных нуклеиновых кислот. В некоторых аспектах идентифицированные таким образом родственные нуклеиновые кислоты могут представлять собой кДНК или геномную ДНК из организмов, отличных от тех, из которых изначально выделили нуклеиновую кислоту по изобретению. В таких процедурах образец нуклеиновой кислоты приводят в контакт с зондом в условиях, которые позволяют зонду специфически гибридизоваться с родственными последовательностями. Затем гибридизацию зонда с нуклеиновыми кислотами из родственного организма определяют, используя любой описанный выше способ.
В реакциях гибридизации нуклеиновых кислот условия, используемые для достижения конкретного уровня строгости, меняют в зависимости от природы гибридизуемых нуклеиновых кислот. Например, при выборе условий гибридизации можно учитывать длину, степень комплементарности, состав нуклеотидной последовательности (например, содержание GC или AT) и вид нуклеиновой кислоты (например, РНК или ДНК) гибридизующихся участков нуклеиновых кислот. Дополнительно учитывают, иммобилизована ли одна из нуклеиновых кислот, например, на фильтре. Гибридизацию можно проводить в условиях низкой строгости, умеренной строгости или высокой строгости. В качестве примера гибридизации нуклеиновых кислот, полимерную мембрану, содержащую иммобилизованные денатурированные нуклеиновые кислоты сначала предварительно гибридизуют в течение 30 минут при 45°C в растворе, состоящем из 0,9 M NaCl, 50 мМ NaH2PO4, pH 7,0, 5,0 мМ Na2EDTA, 0,5% SDS, 10× раствор Денхардта и 0,5 мг/мл полирибоадениловой кислоты. Затем в раствор добавляют приблизительно 2×107 имп./мин (точная активность 4-9×108 имп.×мкг/мин) олигонуклеотидного зонда, меченого P32 на конце. После 12-16 часов инкубации, мембрану промывают в течение 30 минут при комнатной температуре (КТ) в 1×SET (150 мМ NaCl, 20 мМ Tris гидрохлорид, pH 7,8, 1 мМ Na2EDTA), содержащем 0,5% SDS, после чего в течение 30 минут отмывают в свежем 1×SET при Tm-10°C для олигонуклеотидного зонда. Затем мембраной воздействуют на пленку для авторадиографии для определения сигналов гибридизации.
Меняя строгость условий гибридизации, используемых для идентификации нуклеиновых кислот, таких как кДНК или геномная ДНК, которые гибридизуются с поддающимся обнаружению зондом, можно идентифицировать и выделить нуклеиновые кислоты, обладающие различными уровнями гомологии с зондом. Строгость можно менять, проводя гибридизацию при различных температурах ниже температур плавления зондов. Температура плавления, Tm, представляет собой температуру (при определенной ионной силе и pH), при которой 50% целевой последовательности гибридизуется с абсолютно комплементарным зондом. Очень строгие условия выбирают равными или приблизительно на 5°C ниже Tm для конкретного зонда. Температуру плавления зонда можно вычислить, используя следующие образцовые формулы. Для зондов длиной от 14 до 70 нуклеотидов температуру плавления (Tm) вычисляют, используя формулу: Tm=81,5+16,6(log[Na+])+0,41 (фракция G+C)-(600/N), где N представляет собой длину зонда. Если гибридизацию осуществляют в растворе, содержащем формамид, то температуру плавления можно вычислить, используя уравнение: Tm=81,5+16,6(log[Na+])+0,41 (фракция G+C)-(0,63% формамид)-(600/N), где N представляет собой длину зонда. Предварительную гибридизацию можно проводить в 6×SSC, 5×растворе Денхардта, 0,5% SDS, 100 мкг денатурированной фрагментированной ДНК спермы лосося или 6×SSC, 5×растворе Денхардта, 0,5% SDS, 100 мкг денатурированной фрагментированной ДНК спермы лосося, 50% формамиде. Составы для SSC, раствора Денхардта и других растворов перечислены, например, в Sambrook.
В одном из аспектов, гибридизацию проводят посредством добавления поддающегося обнаружению зонда в перечисленные выше растворы для предварительной гибридизации. Когда зонд содержит двухцепочечную ДНК, перед добавлением в раствор для гибридизации ее денатурируют. Фильтр приводят в контакт с раствором для гибридизации в течение достаточного периода времени, чтобы позволить зонду гибридизоваться с кДНК или геномной ДНК, содержащей последовательности, комплементарные ему или гомологичные ему. Для зондов длиной более 200 нуклеотидов гибридизацию можно осуществлять при температуре на 15-25°C ниже Tm. Для более коротких зондов, таких как олигонуклеотидные зонды, гибридизацию можно проводить при температуре на 5-10°C ниже Tm. В одном из аспектов гибридизацию в 6×SSC проводят приблизительно при 68°C. В одном из аспектов гибридизацию в растворах, содержащих 50% формамид, проводят приблизительно при 42°C. Следует учитывать, что все упомянутые выше гибридизации проводят в условиях высокой строгости.
В одном из аспектов после гибридизации фильтр промывают для удаления любых неспецифически связанных поддающихся обнаружению зондов. Строгость, используемую для отмывания фильтров, также можно менять в зависимости от природы нуклеиновых кислот, подлежащих гибридизации, длины нуклеиновых кислот, подлежащих гибридизации, степени комплементарности, состава нуклеотидной последовательности (например, содержание GC и AT) и вида нуклеиновой кислоты (например, РНК или ДНК). Примеры условий отмывания постоянной возрастающей строгости: 2×SSC, 0,1% SDS при комнатной температуре в течение 15 минут (низкая строгость); 0,1×SSC, 0,5% SDS при комнатной температуре в течение от 30 минут до 1 часа (умеренная строгость); 0,1×SSC, 0,5% SDS в течение от 15 до 30 минут при температуре от температуры гибридизации до 68°C (высокая строгость); и 0,15M NaCl в течение 15 минут при 72°C (очень высокая строгость). Финальное отмывание при низкой строгости можно проводить в 0,1×SSC при комнатной температуре. Приведенные выше примеры лишь иллюстрируют один набор условий, которые можно использовать для отмывания фильтров. Профессионал в данной области знает, что существует множество прописей для отмывания различной строгости.
Нуклеиновые кислоты, гибридизованные с зондом, можно идентифицировать посредством радиоавтографии или других общепринятых способов. Указанную выше процедуру можно модифицировать для идентификации нуклеиновых кислот, обладающих снижающимися уровнями гомологии с последовательностью зонда. Например, для получения нуклеиновых кислот со снижающейся гомологией к поддающемуся обнаружению зонду, можно использовать менее строгие условия. Например, температуру гибридизации можно снижать с шагом 5°C с 68°C до 42°C в гибридизационном буфере, содержащим Na+ в концентрации приблизительно 1M. После гибридизации фильтр можно отмывать в 2×SSC, 0,5% SDS при температуре гибридизации. Эти условия считаются «умеренными» условиями при температуре выше 50°C и «низкими» условиями при температуре ниже 50°C. Примером «умеренных» условий гибридизации является указанная выше гибридизация, когда ее проводят при 55°C. Примером условий гибридизации «низкой строгости» является указанная выше гибридизация, когда ее проводят при 45°C.
Альтернативно гибридизацию можно проводить в буферах, таких как 6×SSC, содержащих формамид, при температуре 42°C. В этом случае, концентрацию формамида в гибридизационном буфере можно снижать с шагом 5% с 50% до 0% для идентификации клонов, обладающих снижающимися уровнями гомологии к зонду. После гибридизации фильтр можно отмывать в 6×SSC, 0,5% SDS при температуре 50°C. Эти условия считаются «умеренными» условиями при концентрации формамида более 25% и «низкими» условиями при концентрации формамида ниже 25%. Конкретным примером «умеренных» условий гибридизации является указанная выше гибридизация, когда ее проводят при концентрации формамида 30%. Конкретным примером условий гибридизации «низкой строгости» является указанная выше гибридизация, когда ее проводят при концентрации формамида 10%.
Эти зонды и способы по изобретению можно использовать для выделения или идентификации (например, используя чип) нуклеиновых кислот, обладающих последовательностью по меньшей мере приблизительно с 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или большей идентичностью последовательностей с последовательностью нуклеиновой кислоты по изобретению, содержащих по меньшей мере приблизительно 10, 15, 20, 25, 30, 35, 40, 50, 75, 100, 150, 200, 250, 300, 350, 400, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000 или более ее последовательных оснований, и последовательностей, комплементарных ей. Гомологию можно измерить, используя алгоритм выравнивания, как обсуждается в настоящем документе. Например, гомологичные полинуклеотиды могут содержать кодирующую последовательность, которая представляет собой встречающийся в природе аллельный вариант одной из кодирующих последовательностей, описанных в настоящем документе. Такие аллельные варианты могут содержать замену, делецию или вставку одного или нескольких нуклеотидов по сравнению с нуклеиновой кислотой по изобретению.
Дополнительно зонды и способы по изобретению можно использовать для выделения или идентификации (например, используя чип) нуклеиновых кислот, которые кодируют полипептиды, обладающие по меньшей мере приблизительно 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или большей идентичностью последовательностей (гомологией) с полипептидом по изобретению, содержащие по меньшей мере 5, 10, 15, 20, 25, 30, 35,40, 50, 75, 100 или 150 или более его последовательных аминокислот, что определяют, используя алгоритм выравнивания последовательностей, например, такой как алгоритм FASTA версии 3.0t78 с параметрами по умолчанию, или программу BLAST 2.2.2 с образцовыми настройками, как указано в настоящем документе.
Ингибирование экспрессии гидролаз
В определенных вариантах осуществления в настоящем документе предоставлены нуклеиновые кислоты, комплементарные (например, антисмысловые последовательности) последовательностям нуклеиновых кислот по изобретению, например, последовательностям, кодирующим гидролазы. Антисмысловые последовательности способны ингибировать транспорт, сплайсинг или транскрипцию генов, кодирующих гидролазы. Ингибирование можно осуществлять посредством нецеливания геномной ДНК или матричной РНК. Ингибирование можно осуществлять, используя ДНК, например, ингибирующий рибозим, или РНК, например, двухцепочечную иРНК, содержащую последовательность по изобретению. Транскрипцию или функцию целевой нуклеиновой кислоты можно ингибировать, например, посредством гибридизации и/или расщепления. В настоящем документе предоставлены наборы ингибиторов, содержащие олигонуклеотиды, способные связываться с геном и/или транскриптом гидролазы, в любом случае предотвращая или ингибируя образование или функцию гидролазы. Связывание может происходить посредством специфической гибридизации последовательностей. Другой эффективный класс ингибиторов включает олигонуклеотиды, которые вызывают инактивацию или расщепление транскрипта гидролазы. Олигонуклеотид, такой как рибозим, может обладать ферментативной активностью, которая является причиной такого расщепления. Олигонуклеотид можно химически модифицировать или конъюгировать с ферментом или композицией, способной расщеплять комплементарную нуклеиновую кислоту. Можно провести скрининг пула многих таких различных олигонуклеотидов для выявления олигонуклеотидов с желаемой активностью.
Антисмысловые олигонуклеотиды
В определенных вариантах осуществления в настоящем документе предоставлены антисмысловые олигонуклеотиды, способные связываться с транскриптом гидролазы, которые могут ингибировать гидролазную активность посредством нацеливания на мРНК или геномную ДНК. Стратегии конструирования антисмысловых олигонуклеотидов хорошо описаны в научной и патентной литературе, и специалист в данной области может сконструировать такие олигонуклеотиды гидролаз, используя новые реактивы по изобретению. Например, протоколы прогулки по гену/картирования РНК для скрининга эффективных антисмысловых олигонуклеотидов хорошо известны в данной области, см., например, Ho (2000) Methods Enzymol. 314:168-183, описание анализа картирования РНК, который основан на стандартных молекулярных способах для обеспечения легкого и надежного способа отбора активной антисмысловой последовательности. Также см. Smith (2000) Eur. J. Pharm. Sci. 11:191-198.
В одном из аспектов в качестве антисмысловых олигонуклеотидов используют полученные рекомбинантным способом или выделенные встречающиеся в природе нуклеиновые кислоты. Антисмысловые олигонуклеотиды могут иметь любую длину; например, в альтернативных аспектах антисмысловые олигонуклеотиды имеют длину приблизительно от 5 до 100, приблизительно от 10 до 80, приблизительно от 15 до 60, приблизительно от 18 до 40. Антисмысловые олигонуклеотиды могут представлять собой одноцепочечную или двухцепочечную РНК или ДНК. Оптимальную длину можно определить стандартным скринингом. Антисмысловые олигонуклеотиды могут присутствовать в любой концентрации. Оптимальную концентрацию можно определить стандартным скринингом. Известен широкий спектр синтетических, не встречающихся в природе аналогов нуклеотидов и нуклеиновых кислот, которые могут способствовать решению это возможной проблемы. Например, можно использовать пептидные нуклеиновые кислоты (ПНК), содержащие неионные остовы, такие как N-(2-аминоэтил)глициновые единицы. Также можно использовать антисмысловые олигонуклеотиды, содержащие фосфотиоатные связи, как описано в WO 97/03211; WO 96/39154; Mata (1997) Toxicol Appl Pharmacol 144:189-197; Antisense Therapeutics, ed. Agrawal (Humana Press, Totowa, N.J., 1996). В настоящем документе предоставлены антисмысловые олигонуклеотиды, содержащие синтетические аналоги остова ДНК, которые также могут включать фосфодитиоатные, метилфосфонатные, фосфорамидатные, алкилфосфотиоэфирные, сульфаматные, 3'-тиоацеталевые, метилен(метилимино), 3'-N-карбаматные и морфолинокарбаматные нуклеиновые кислоты, как описано выше.
Можно использовать способы комбинаторной химии для создания большого числа олигонуклеотидов, среди которых можно быстро проводить скрининг на конкретные олигонуклеотиды, которые обладают подходящей аффинностью и специфичностью связывания с любой мишенью, такой как смысловые и антисмысловые последовательности гидролаз по изобретению (см., например, Gold (1995) J. of Biol. Chem. 270:13581-13584).
Ингибирующие рибозимы
В определенных вариантах осуществления в настоящем документе предоставлены рибозимы, способные связывать транскрипт гидролазы, которые могут ингибировать гидролазную активность посредством нацеливания на мРНК. Стратегии конструирования рибозимов и отбора специфичной к гидролазе антисмысловой последовательности для нацеливания подробно описаны в научной и патентной литературе, и специалист в данной области может сконструировать такие рибозимы, используя новые реактивы по изобретению. Рибозимы действуют посредством связывания с целевой РНК через участок рибозима для связывания с целевой РНК, который расположен в непосредственной близости к ферментативному участку РНК, который расщепляет целевую РНК. Таким образом, рибозим распознает и связывает целевую РНК через спаривание комплементарных оснований, и после связывания с правильным участком выполняет функцию фермента для расщепления и инактивации целевой РНК. Таким образом, расщепление целевой РНК уничтожает ее способность управлять синтезом кодируемого белка, если расщепление происходит в кодирующей последовательности. После связывания и расщепления целевой РНК рибозимом, он, как правило, освобождается от этой РНК и, следовательно, может повторно связывать и расщеплять новые мишени.
В некоторых случаях ферментативная природа рибозима может быть предпочтительнее других способов, таких как антисмысловая технология (где молекула нуклеиновой кислоты просто связывает целевую нуклеиновую кислоту для блокирования ее транскрипции, трансляции или связывания с другой молекулой), поскольку эффективная концентрация рибозима, необходимая для осуществления терепевтического воздействия, может быть ниже концентрации антисмыслового олигонуклеотида. Это возможное преимущество отражает способность рибозима выполнять функцию фермента. Таким образом, одна молекула рибозима способна расщепить несколько молекул целевой РНК. Кроме того, как правило, рибозим представляет собой высокоспецифичный ингибитор, специфичность ингибирования которого зависит не только от механизма связывания на основе спаривания оснований, но также от механизма, посредством которого молекула ингибирует экспрессию РНК, с которой она связывается. То есть ингибирование обусловлено расщеплением целевой РНК и, следовательно, специфичность определяют как отношение скорости расщепления целевой РНК к скорости расщепления нецелевой РНК. Этот механизм расщепления зависит от факторов, дополняющих факторы, вовлеченные в спаривание оснований. Таким образом, специфичность действия рибозима может быть выше, чем у антисмыслового олигонуклеотида, связывающего тот же участок РНК.
РНК молекулу ферментативного рибозима можно сформировать в виде мотива молота, но также можно сформировать в виде мотива шпильки, вируса гепатита D, интрона I группы или РНК, похожей на РНКазу P (связанную с направляющей последовательностью РНК). Примеры таких мотивов молота описаны в публикации Rossi (1992) Aids Research and Human Retroviruses 8:183; мотивы шпильки в публикациях Hampel (1989) Biochemistry 28:4929, и Hampel (1990) Nuc. Acids Res. 18:299; мотив вируса гепатида D в публикации Perrotta (1992) Biochemistry 31:16; мотив РНКазы P в публикации Guerrier-Takada (1983) Cell 35:849; и интрон I группы в публикации Cech (патент США № 4987071). Перечисление этих конкретных мотивов не служит цели ограничения; специалистам в данной области понятно, что ферментативная молекула РНК по изобретению может иметь конкретный сайт связывания субстрата, комплементарный одному или нескольким участкам РНК целевого гена и содержит нуклеотидную последовательность внутри этого сайта связывания субстрата или вокруг него, которая наделяет молекулу расщепляющей РНК активностью.
РНК-интерференция (интерферирующая РНК)
В определенных вариантах осуществления в настоящем документе предоставлены ингибирующие молекулы РНК, так называемые молекулы «интерферирующей РНК», содержащие последовательность гидролазы по изобретению. Молекула интерферирующей РНК может содержать молекулу двухцепочечной РНК (дцРНК), например, миРНК и/или мкРНК. Интерферирующая РНК может ингибировать экспрессию гена или транскрипта гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы). В одном из аспектов молекула интерферирующей РНК, например, миРНК и/или мкРНК, имеет длину приблизительно в 10, 11,12,13, 14, 15, 16, 17, 18,19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 или более спаренных нуклеотидов. Несмотря на то, что изобретение не ограничено каким-либо конкретным механизмом действия, интерферирующая РНК может входить в клетку и вызывать разрушение одноцепочечной РНК (оцРНК) со схожей или идентичной последовательностью, включая эндогенную мРНК. Когда клетку подвергают воздействию двухцепочечной РНК (дцРНК), мРНК от гомологичного гена избирательно разрушается посредством процесса, который называют РНК-интерференция. Возможный основной механизм действия интерферирующей РНК состоит в разрушении двухцепочечной РНК (дцРНК), совпадающей с конкретной последовательностью гена, на короткие куски, которые называют малой интерферирующей РНК, которые запускают разрушение мРНК, которая совпадает с их последовательностью.
В одном из аспектов интерферирующую РНК по изобретению используют в лекарственных средствах для подавления экспрессии генов, см., например, Shuey (2002) Drug Discov. Today 7:1040-1046. В определенных вариантах осуществления в настоящем документе предоставлены способы избирательного разрушения РНК, используя интерферирующую РНК. Этот процесс можно осуществлять in vitro, ex vivo или in vivo. В одном из аспектов молекулы интерферирующей РНК по изобретению можно использовать для создания мутации с потерей функции в клетке, органе или животном. Способы получения и использования молекул интерферирующей РНК для избирательного разрушения РНК хорошо известны в данной области, см., например, патенты США № 6506559; 6511824; 6515109; 6489127.
Модификация нуклеиновых кислот
В определенных вариантах осуществления в настоящем документе предоставлены способы создания вариантов нуклеиновых кислот, например, кодирующих гидролазу или антитело по изобретению. Эти способы можно повторять или использовать в различных сочетаниях для создания гидролаз или антител, обладающих измененной или отличающейся активностью или измененной или отличающейся стабильностью относительно той гидролазы или антитела, которые кодирует матричная нуклеиновая кислота. Эти способы также можно повторять или использовать в различных сочетаниях, например, для создания изменений в экспрессии гена/транскрипта, трансляции транскрипта или стабильности транскрипта. В другом аспекте изменяют генетический состав клетки, например, посредством модификации гомологичного гена ex vivo, после чего его повторно вводят в клетку.
Термин «вариант» может включать полинуклеотиды или полипептиды по изобретению, модифицированные в одной или нескольких парах оснований, кодонах, интронах, экзонах или аминокислотных остатках (соответственно), при этом сохраняя биологическую активность гидролазы по изобретению. Варианты можно получить с помощью любого числа средств, включая такие способы как, например, допускающая ошибки ПЦР, шаффлинг, олигонуклеотид-специфический мутагенез, сборочная ПЦР, половой ПЦР мутагенез, мутагенез in vivo, кассетный мутагенез, рекурсивный множественный мутагенез, экспоненциальный множественный мутагенез, сайт-специфический мутагенез, GeneReassembly, GSSMSM и любое их сочетание. Способы получения вариантов гидролаз, обладающих активностью при pH или температуре, например, которые отличаются от таковых для гидролазы дикого типа, включены в настоящий документ.
Нуклеиновую кислоту по изобретению можно изменять любыми средствами. Например, случайные или стохастические способы или нестохастические способы или способы «направленной эволюции», см., например, патент США № 6361974. Способы внесения случайных мутаций в гены хорошо известны в данной области, см., например, патент США № 5830696. Например, можно использовать мутагены для внесения случайных мутаций в ген. Мутагены включают, например, ультрафиолетовый свет или γ-излучение или химический мутаген, например, митомицин, азотистую кислоту, фотоактивированные псоралены, отдельно или в сочетании, чтобы индуцировать дефекты ДНК, поддающиеся репарации посредством рекомбинации. Другие химические мутагены включают, например, бисульфит натрия, азотистую кислоту, гидроксиламин, гидразин или муравьиную кислоту. Другие мутагены представляют собой аналоги предшественников нуклеотидов, например, нитрозогуанидин, 5-бромурацил, 2-аминопурин или акридин. Эти средства можно добавлять в ПЦР вместо предшественника нуклеотида, посредством чего вносить мутации в последовательность. Также можно использовать интеркалирующие средства, такие как профлавин, акрифлавин, хинакрин и т.п.
Можно использовать любой способ из молекулярной биологии, например, случайный ПЦР-мутагенез, см., например, Rice (1992) Proc. Natl. Acad. Sci. USA 89:5467-5471; или множественный комбинаторный кассетный мутагенез, см., например, Crameri (1995) Biotechniques 18:194-196. Альтернативно нуклеиновые кислоты, например, гены, можно повторно собирать после случайного, или «стохастического», фрагментирования, см., например, патенты США №№ 6291242; 6287862; 6287861; 5955358; 5830721; 5824514; 5811238; 5605793. В альтернативных аспектах модификации, вставки или делеции вводят посредством допускающей ошибки ПЦР, шаффлинга, олигонуклеотид-специфического мутагенеза, сборочной ПЦР, полового ПЦР мутагенеза, мутагенеза in vivo, кассетного мутагенеза, рекурсивного множественного мутагенеза, экспоненциального множественного мутагенеза, сайт-специфического мутагенеза, Gene Site Saturation MutagenesisSM (GSSMSM), синтетической перегруппировки лигированием (SLR или GeneReassembly), рекомбинации, рекурсивной рекомбинации последовательностей, мутагенеза модифицированной фосфотиоатом ДНК, мутагенеза содержащей урацил матрицы, мутагенеза двойной спирали с разрывами, точечного мутагенеза при репарации ошибочно спаренных оснований, мутагенеза в штамме-хозяине с недостаточностью репарации, химического мутагенеза, радиогенного мутагенеза, мутагенеза делецией, мутагенеза рестрикцией-отбором, мутагенеза рестрикцией-очисткой, синтеза искусственных генов, множественного мутагенеза, создания мультимера химерной нуклеиновой кислоты и/или сочетания этих и других способов.
В следующих публикациях описаны различные процедуры и/или способы рекурсивной рекомбинации, которые можно встраивать в способы по изобретению: Stemmer (1999) «Molecular selection of viruses for targeting and other clinical properties» Tumor Targeting 4:1-4; Ness (1999) Nature Biotechnology 17:893-896; Chang (1999) «Evolution of a cytokine using DNA family shuffling» Nature Biotechnology 17:793-797; Minshull (1999) «Protein evolution by molecular selection» Current Opinion in Chemical Biology 3:284-290; Christians (1999) «Directed Evolution of thymidine kinase for AZT phosphorylation using DNA family shuffling» Nature Biotechnology 17:259-264; Crameri (1998) «DNA shuffling of a family of genes from diverse species accelerates directed evolution» Nature 391:288-291; Crameri (1997) «Molecular evolution of an arsenate detoxification pathway by DNA shuffling,» Nature Biotechnology 15:436-438; Zhang (1997) «Directed evolution of an effective fucosidase from a galactosidase by DNA shuffling and screening» Proc. Natl. Acad. Sci. USA 94:4504-4509; Patten et al. (1997) «Applications of DNA Shuffling to Pharmaceuticals and Vaccines» Current Opinion in Biotechnology 8:724-733; Crameri et al. (1996) «Construction and evolution of antibody-phage libraries by DNA shuffling» Nature Medicine 2:100-103; Gates et al. (1996) «Affinity selective isolation of ligands from peptide libraries through display on a lac repressor 'headpiece dimer'« Journal of Molecular Biology 255:373-386; Stemmer (1996) «Sexual PCR and Assembly PCR» In: The Encyclopedia of Molecular Biology. VCH Publishers, New York, pp.447-457; Crameri and Stemmer (1995) «Combinatorial multiple sassette nutagenesis creates all the permutations of mutant and wildtype cassettes» BioTechniques 18:194-195; Stemmer et al. (1995) «Single-step assembly of a gene and entire plasmid form large numbers of oligodeoxyribonucleotides» Gene, 164:49-53; Stemmer (1995) «The Evolution of Molecular Computation» Science 270: 1510; Stemmer (1995) «Searching Sequence Space» Bio/Technology 13:549-553; Stemmer (1994) «Rapid evolution of a protein in vitro by DNA shuffling» Nature 370:389-391; и Stemmer (1994) «DNA shuffling by random fragmentation and reassembly: In vitro recombination for molecular evolution.» Proc. Natl. Acad. Sci. USA 91:10747-10751.
Мутационные способы создания разнообразия включают, например, сайт-специфический мутагенез (Ling et al. (1997) «Approaches to DNA mutagenesis: an overview» Anal Biochem. 254(2): 157-178; Dale et al. (1996) «Oligonucleotide-directed random mutagenesis using phosphorothioate method» Methods Mol. Biol. 57:369-374; Smith (1985) «In vitro mutagenesis» Ann. Rev. Genet. 19:423-462; Botstein & Shortle (1985) «Strategies and applications of in vitro mutagenesis» Science 229:1193-1201; Carter (1986) «Site-directed mutagenesis» Biochem. J. 237:1-7; и Kunkel (1987) «The efficiency of oligonucleotide directed mutagenesis» in Nucleic Acids & Molecular Biology (Eckstein, F. and Lilley, D. M. J. eds., Springer Verlag, Berlin)); мутагенез с использованием матриц, содержащих урацил (Kunkel (1985) «Rapid and efficient site-specific mutagenesis without phenotypic selection» Proc. Natl. Acad. Sci. USA 82:488-492; Kunkel et al. (1987) «Rapid and efficient site-specific mutagenesis without phenotypic selection» Methods in Enzymol. 154, 367-382; и Bass et al. (1988) «Mutant Tip repressors with new DNA-binding specificities» Science 242:240-245); олигонуклеотид-специфический мутагенез, (Methods in Enzymol. 100: 468-500 (1983); Methods in Enzymol. 154: 329-350 (1987); Zoller & Smith (1982) «Oligonucleotide-directed mutagenesis using M13-derived vectors: an efficient and general procedure for the production of point mutations in any DNA fragment» Nucleic Acids Res. 10:6487-6500; Zoller & Smith (1983) «Oligonucleotide-directed mutagenesis of DNA fragments cloned into M13 vectors» Methods in Enzymol. 100:468-500; и Zoller & Smith (1987) Oligonucleotide-directed mutagenesis: a simple method using two oligonucleotide primers and a single-stranded DNA template » Methods in Enzymol. 154:329-350); мутагенез модифицированной фосфотиоатом ДНК (Taylor et al. (1985) «The use of phosphorothioate-modified DNA in restriction enzyme reactions to prepare nicked DNA» Nucl. Acids Res. 13: 8749-8764; Taylor et al. (1985) «The rapid generation of oligonucleotide-directed mutations at high frequency using phosphorothioate-modified DNA» Nucl. Acids Res. 13: 8765-8787 (1985); Nakamaye (1986) «Inhibition of restriction endonuclease NciI cleavage by phosphorothioate groups and its application to oligonucleotide-directed mutagenesis» Nucl. Acids Res. 14: 9679-9698; Sayers et al. (1988) «Y-T Exonucleases in phosphorothioate-based oligonucleotide-directed mutagenesis» Nucl. Acids Res. 16:791-802; и Sayers et al. (1988) «Strand specific cleavage of phosphorothioate-containing DNA by reaction with restriction endonucleases in the presence of ethidium bromide» Nucl. Acids Res. 16: 803-814); мутагенез с использованием двунитевой ДНК с пропусками (Kramer et al. (1984) «The gapped duplex DNA approach to oligonucleotide-directed mutation construction» Nucl. Acids Res. 12: 9441-9456; Kramer & Fritz (1987) Methods in Enzymol. «Oligonucleotide-directed construction of mutations via gapped duplex DNA» 154:350-367; Kramer et al. (1988) «Improved enzymatic in vitro reactions in the gapped duplex DNA approach to oligonucleotide-directed construction of mutations» Nucl. Acids Res. 16: 7207; and Fritz et al. (1988) «Oligonucleotide-directed construction of mutations: a gapped duplex DNA procedure without enzymatic reactions in vitro» Nucl. Acids Res. 16: 6987-6999).
Дополнительные протоколы, используемые в способах по изобретению, включают точечную репарацию ошибочно спаренных оснований (Kramer (1984) «Point Mismatch Repair» Cell 38:879-887), мутагенез с использованием штаммов-хозяев с недостаточностью репарации (Carter et al. (1985) «Improved oligonucleotide site-directed mutagenesis using M13 vectors» Nucl. Acids Res. 13: 4431-4443; и Carter (1987) «Improved oligonucleotide-directed mutagenesis using M13 vectors» Methods in Enzymol. 154: 382-403), мутагенез делециями (Eghtedarzadeh (1986) «Use of oligonucleotides to generate large deletions» Nucl. Acids Res. 14: 5115), рестрикцию-отбор и рестрикцию-отбор и рестрикцию-очистку (Wells et al. (1986) «Importance of hydrogen-bond formation in stabilizing the transition state of subtilisin» Phil. Trans. R. Soc. Lond. A 317: 415-423), мутагенез путем синтеза целого гена (Nambiar et al. (1984) «Total synthesis and cloning of a gene coding for the ribonuclease S protein» Science 223: 1299-1301; Sakamar and Khorana (1988) «Total synthesis and expression of a gene for the a-subunit of bovine rod outer segment guanine nucleotide-binding protein (transducin)» Nucl. Acids Res. 14: 6361-6372; Wells et al. (1985) «Cassette mutagenesis: an efficient method for generation of multiple mutations at defined sites» Gene 34:315-323; и Grundstrom et al. (1985) «Oligonucleotide-directed mutagenesis by microscale 'shot-gun' gene synthesis» Nucl. Acids Res. 13: 3305-3316), репарацию двухцепочечных разрывов (Mandecki (1986); Arnold (1993) «Protein engineering for unusual environments» Current Opinion in Biotechnology 4:450-455. «Oligonucleotide-directed double-strand break repair in plasmids of Escherichia coli: a method for site-specific mutagenesis» Proc. Natl. Acad. Sci. USA, 83:7177-7181). Дополнительные подробности относительно многих указанных выше способов можно найти в Methods in Enzymology Volume 154, в котором также описаны эффективные способы контроля для поиска и устранения проблем, связанных с различными способами получения мутаций.
Дополнительные протоколы, используемые в способах по изобретению, включают те, что рассмотрены в патентах США №№ 5605793 Stemmer (Feb. 25, 1997), «Methods for In vitro Recombination»; патенте США № 5811238 Stemmer et al. (Sep. 22, 1998) «Methods for Generating Polynucleotides having Desired Characteristics by Iterative Selection and Recombination»; патенте США № 5830721 Stemmer et al. (Nov. 3,1998), «DNA Mutagenesis by Random Fragmentation and Reassembly»; патенте США № 5834252 Stemmer, et al. (Nov. 10, 1998) «End-Complementary Polymerase Reaction»; патенте США № 5837458 Minshull, et al. (Nov. 17, 1998), «Methods and Compositions for Cellular and Metabolic Engineering»; WO 95/22625, Stemmer and Crameri, «Mutagenesis by Random Fragmentation and Reassembly»; WO 96/33207 Stemmer and Lipschutz «End Complementary Polymerase Chain Reaction»; WO 97/20078 Stemmer and Crameri «Methods for Generating Polynucleotides having Desired Characteristics by Iterative Selection and Recombination»; WO 97/35966 Minshull and Stemmer, «Methods and Compositions for Cellular and Metabolic Engineering»; WO 99/41402 Punnonen et al. «Targeting of Genetic Vaccine Vectors»; WO 99/41383 Punnonen et al. «Antigen Library Immunization»; WO 99/41369 Punnonen et al. «Genetic Vaccine Vector Engineering»; WO 99/41368 Punnonen et al. «Optimization of Immunomodulatory Properties of Genetic Vaccines»; EP 752008 Stemmer and Crameri, «DNA Mutagenesis by Random Fragmentation and Reassembly»; EP 0932670 Stemmer «Evolving Cellular DNA Uptake by Recursive Sequence Recombination»; WO 99/23107 Stemmer et al., «Modification of Virus Tropism and Host Range by Viral Genome Shuffling»; WO 99/21979 Apt et al., «Human Papillomavirus Vectors»; WO 98/31837 del Cardayre et al. «Evolution of Whole Cells and Organisms by Recursive Sequence Recombination»; WO 98/27230 Patten and Stemmer, «Methods and Compositions for Polypeptide Engineering»; WO 98/27230 Stemmer et al., «Methods for Optimization of Gene Therapy by Recursive Sequence Shuffling and Selection», WO 00/00632, «Methods for Generating Highly Diverse Libraries», WO 00/09679, «Methods for Obtaining in vitro Recombined Polynucleotide Sequence Banks and Resulting Sequences», WO 98/42832 Arnold et al., «Recombination of Polynucleotide Sequences Using Random or Defined Primers», WO 99/29902 Arnold et al., «Method for Creating Polynucleotide and Polypeptide Sequences», WO 98/41653 Vind, «An in vitro Method for Construction of a DNA Library», WO 98/41622 Borchert et al., «Method for Constructing a Library Using DNA Shuffling» и WO 98/42727 Pati and Zarling, «Sequence Alterations using Homologous Recombination».
Протоколы, которые можно использовать, (предоставлены подробности, касающиеся различных способов создания разнообразия) описаны, например, в патентной заявке США № (USSN) 09/407800, «SHUFFLING OF CODON ALTERED GENES» Patten et al., которая подана 28 сентября 1999 года; «EVOLUTION OF WHOLE CELLS AND ORGANISMS BY RECURSIVE SEQUENCE RECOMBINATION» del Cardayre et al., патенте США № 6379964; «OLIGONUCLEOTIDE MEDIATED NUCLEIC ACID RECOMBINATION» Crameri et al., патентах США №№ 6319714; 6368861; 6376246; 6423542; 6426224 и PCT/US00/01203; «USE OF CODON-VARIED OLIGONUCLEOTIDE SYNTHESIS FOR SYNTHETIC SHUFFLING» Welch et al., патенте США № 6436675; «METHODS FOR MAKING CHARACTER STRINGS, POLYNUCLEOTIDES & POLYPEPTIDES HAVING DESIRED CHARACTERISTICS» Selifonov et al., который подан 18 января 2000 года, (PCT/US00/01202) и, например, «METHODS FOR MAKING CHARACTER STRINGS, POLYNUCLEOTIDES & POLYPEPTIDES HAVING DESIRED CHARACTERISTICS» Selifonov et al., который подан 18 июля 2000 года (США № 09/618579); «METHODS OF POPULATING DATA STRECTURES FOR USE IN EVOLUTIONARY SIMULATIONS» Selifonov and Stemmer, который подан 18 января 2000 года (PCT/US00/01138); и «SINGLE-STRANDED NUCLEIC ACID TEMPLATE-MEDIATED RECOMBINATION AND NUCLEIC ACID FRAGMENT ISOLATION» Affholter, который подан 6 сенября 2000 года (США № 09/656549); и в патентах США №№ 6177263; 6153410.
Нестохастические способы, или способы «направленной эволюции», которые включают, например, Gene Site Saturation MutagenesisSM (GSSMSM), синтетическую перегруппировку лигированием (SLR или GeneReassembly) или их сочетание, используют для модификации нуклеиновых кислот по изобретению для создания гидролаз с новыми или измененными свойствами (например, активность при крайних кислых или щелочных условиях, высоких температурах и т.п.). Скрининг полипептидов, кодируемых модифицированными нуклеиновыми кислотами, на активность можно проводить перед тестированием на протеолитическую или другую активность. Можно использовать любой способ или протокол тестирования, например, использовать платформу капиллярного блока. См., например, патенты США №№ 6361974; 6280926; 5939250.
Насыщающий мутагенез или технология GSSMSM
В одном из аспектов по изобретению нестохастическую модификацию генов, «направленный эволюционный процесс», используют для создания гидролаз и антител с новыми или измененными свойствами. Разновидности этого способа названы «Gene Site Saturation Mutagenesis», «сайт-насыщающий мутагенез», «насыщающий мутагенез» или просто «GSSMSM». Их можно использовать в сочетании с другими процессами создания мутаций. В одном из аспектов в настоящем документе предоставлены способы получения ферментов и антител, используя технологию GSSMSM, например, как описано в настоящем документе, а также в патентах США №№ 6171820; 6579258; 6238884.
В одном из аспектов технология GSSMSM включает предоставление матричного полинуклеотида и нескольких олигонуклеотидов, где каждый олигонуклеотид содержит последовательность, гомологичную матричному полинуклеотиду, посредством чего осуществляют нацеливание на конкретную последовательность матричного полинуклеотида, и последовательность, которая является вариантом гомологичного гена; создание дочерних полинуклеотидов, содержащих нестохастические изменения последовательности посредством репликации матричного полинуклеотида с олигонуклеотидами, посредством чего создают полинуклеотиды, содержащие изменения последовательности гомологичного гена.
В одном из аспектов праймеры, содержащие кодоны с вырожденной последовательностью N,N,G/T, используют для введения точечных мутаций в полинуклеотид с тем, чтобы создать набор дочерних полипептидов, в которых представлен полный спектр замен одной аминокислоты в каждом положении аминокислоты, например, аминокислотного остатка в активном центре фермента или сайте связывания лиганда, на который нацелена модификация. Эти олигонуклеотиды могут содержать смежно первую гомологичную последовательность, вырожденную последовательность N,N,G/T, и необязательно вторую гомологичную последовательность. Дочерние продуты трансляции в 5'-3' направлении от использования таких олигонуклеотидов содержат все возможные аминокислотные замены в каждом аминокислотном положении вдоль полипептида, поскольку вырожденность последовательности N,N,G/T включает кодоны для всех 20 аминокислот. В одном из аспектов один такой вырожденный олигонуклеотид (содержащий, например, одно вырожденную кассету N,N,G/T) используют для введения в каждый исходный кодон в исходной полинуклеотидной матрице полного спектра замен кодонов. В другом аспекте используют по меньшей мере две вырожденные кассеты, или в одном и том же олигонуклеотиде или в разных, чтобы ввести по меньшей мере в исходных кодона в исходной полинуклеотидной матрице полный спектр замен кодонов. Например, один олигонуклеотид может содержать более одной последовательности N,N,G/T для введения аминокислотных мутаций более чем в один сайт. Эти несколько последовательностей N,N,G/T могут быть расположены непосредственно смежно или разделены одной или несколькими дополнительными нуклеотидными последовательностями. В другом аспекте олигонуклеотиды, пригодные для введения вставок и делеций, можно использовать или отдельно или в сочетании с кодонами, содержащими последовательность N,N,G/T, чтобы вводить в любом сочетании или комбинации аминокислотные вставки, делеции и/или замены.
В одном из аспектов выполняют одновременный мутагенез в двух или более смежных аминокислотных положениях с использованием олигонуклеотида, который содержит смежные триплеты N,N,G/T, т.е. вырожденную последовательность (N,N,G/T)n. В другом аспекте используют вырожденные кассеты, обладающие меньшей вырожденностью, чем последовательность N,N,G/T. Например, в некоторых случаях может быть желательным использовать (например, в олигонуклеотиде) вырожденную последовательность триплета, включающую только один N, где указанный N может находиться в первом, втором или третьем положении триплета. В остальных двух положениях триплета можно использовать любые другие основания, включая любые их сочетания и комбинации. Альтернативно, в некоторых случаях может быть желательным использовать (например, в олигонуклеотиде) вырожденную последовательность триплета N,N,N.
В одном из аспектов использование вырожденных триплетов (например, триплетов N,N,G/T) предоставляет возможность системного и легкого создания полного спектра возможных природных аминокислот (всего 20 аминокислот) в каждом и любом аминокислотном положении в полипептиде (в альтернативных аспектах способы также включают создание менее чем всех возможных замен в положении аминокислотного остатка или кодона). Например, для полипептида длиной 100 аминокислот можно создать 2000 различных видов (т.е. 20 возможных аминокислот в одном положении × 100 аминокислотных положений). Путем использования олигонуклеотида или набора олигонуклеотидов, содержащих вырожденный триплет N,N,G/T, 32 отдельные последовательности могут кодировать все 20 возможных природных аминокислот. Таким образом, в реакционном сосуде, в котором исходную полинуклеотид последовательность подвергают насыщающему мутагенезу с использованием по меньшей мере одного такого олигонуклеотида, создают 32 различных дочерних полинуклеотида, кодирующих 20 четко выраженных полипептидов. В отличие от этого, использование невырожденного олигонуклеотида в сайт-специфическом мутагенезе ведет только к одному дочернему полипептидному продукту в одном реакционном сосуде. Невырожденные олигонуклеотиды необязательно можно использовать в сочетании с описанными вырожденными праймерами; например, невырожденные олигонуклеотиды можно использовать для создания конкретных точечных мутаций в работающем полинуклеотиде. Это предоставляет средство для создания конкретных молчащих точечных мутаций, точечных мутаций, ведующих к соответствующим аминокислотным заменам, и точечных мутаций, которые являются причиной образования стоп-кодонов и соответствующей экспрессии полипептидных фрагментов.
В одном из аспектов каждый реакционный сосуд для насыщающего мутагенеза содержит полинуклеотиды, кодирующие по меньшей мере 20 таких дочерних полипептидных молекул (например, гидролаз, например, липаз, сатураз, пальмитаз и/или стеаратаз), что все 20 природных аминокислот представлены в одном конкретном аминокислотном положении, соответствующем положению кодона, мутировавшего в исходном полинуклеотиде (в других аспектах используют менее чем все 20 природных сочетаний). Можно провести клональную амплификацию 32-кратных вырожденных дочерних полипептидов, созданных в каждом реакционном сосуде для насыщающего мутагенеза, (например, клонировать в подходящий организм-хозяин, например, в организм-хозяин E. coli, используя, например, экспрессирующий вектор) и провести скрининг экспрессии. Когда посредством скрининга устанавливают, что отдельный дочерний полипептид проявляет благоприятное изменение свойства (по сравнению с исходным полипептидом, такое как повышенная избирательность гидролитического расщепления сложных эфиров пальмитиновой кислоты в сравнении со сложными эфирами олеиновой кислоты), его можно секвенировать для идентификации соответствующей благоприятной аминокислотной замены, содержащейся в нем.
В одном из аспектов, при введении мутаций в любое и каждое аминокислотное положение в исходном полипептиде с использованием насыщающего мутагенеза, как описано в настоящем документе, благоприятные аминокислотные замены можно идентифицировать более чем в одном аминокислотном положении. Можно создать одну или несколько новых дочерних молекул, которые содержат сочетание из всех или части этих благоприятных аминокислотных замен. Например, если идентифицируют 2 конкретных благоприятных аминокислотных замены в каждом из 3 аминокислотных положений в полипептиде, то комбинации включают 3 возможности в каждом положении (изменение исходной аминокислоты отсутствует и каждая из двух благоприятных замен) и 3 положения. Таким образом, возможны всего 3×3×3 или 27 возможностей, включая 7 ранее исследованных - 6 одиночных точечных мутаций (т.е. по 2 в каждом из трех положений) и отсутствие изменений в каком-либо положении.
В другом аспекте сайт-насыщающий мутагенез можно использовать вместе с другими стохастическими или нестохастическими средствами для изменения последовательности, например, с синтетической перегруппировкой лигированием (см. ниже), шаффлингом, химеризацией, рекомбинацией и другими процессами и средствами для создания мутаций. В настоящем документе предоставлен процесс(ы) для создания мутаций, включая насыщающий мутагенез, которые используют повторно.
Синтетическая перегруппировка лигированием (SLR)
В одном из аспектов в настоящем документе предоставлены системы нестохастической модификации генов, названные «синтетической перегруппировкой лигированием», или просто «SLR», также известные как технология «GeneReassembly», «направленный эволюционный процесс», для создания полипептидов, например, ферментов (таких как гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы) или антител по изобретению, с новыми или измененными свойствами. SLR представляет способ нестохастического лигирования олигонуклеотидных фрагментов вместе. Этот способ отличается от стохастического шаффлинга олигонуклеотидов тем, что строительные блоки нуклеиновой кислоты не перетасовывают, сцепляют или химеризуют случайным образом, а собирают нестохастически. См., например, патенты США №№ 6773900; 6740506; 6713282; 6635449; 6605449; 6537776.
В одном из аспектов SLR содержит следующие стадии: (a) предоставление матричного полинуклеотида, где матричный полинуклеотид содержит последовательность, кодирующую гомологичный ген; (b) предоставление нескольких полинуклеотидных строительных блоков, где полинуклеотидные строительные блоки сконструированы для перекрестной перегруппировки с матричным полинуклеотидом в предварительно определенной последовательности, и полинуклеотидный строительный блок содержит последовательность, которая представляет собой вариант гомологичного гена, и последовательность, гомологичную матричному полинуклеотиду, фланкирующую последовательность варианта; (c) объединение полинуклеотидного строительного блока с матричным полинуклеотидом так, что полинуклеотидный строительный блок перекрестно перегруппировывается с матричным полинуклеотидом для создания полинуклеотидов, содержащих изменения последовательности гомологичного гена.
SLR не зависит от наличия высоких уровней гомологии между полинуклеотидами, подлежащими перегруппировке. Таким образом, этот способ можно использовать для нестохастического создания библиотек (или наборов) дочерних молекул, содержащих более 10100 различных химер. SLR можно использовать для создания библиотек, содержащих более 101000 различных дочерних химер. В одном из аспектов в настоящем документе предоставлены нестохастические способы получения набора законченных химерных молекул нуклеиновых кислот, обладающих общим порядком сборки, который выбран посредством конструирования. Этот способ включает стадии создания посредством конструирования нескольких конкретных строительных блоков из нуклеиновых кислот, которые имеют пригодные взаимно совместимые лигируемые концы, и сборки этих строительных блоков из нуклеиновых кислот так, что достигается задуманный общий порядок сборки.
Взаимно совместимые лигируемые концы строительных блоков из нуклеиновых кислот, подлежащих сборке, считаются «пригодными» для этого типа упорядоченной сборки, если они позволяют соединять строительные блоки в предварительно определенном порядке. Таким образом, общий порядок сборки, в котором можно соединить строительные блоки из нуклеиновых кислот, точно определяется конструкцией лигируемых концов. Если используют более чем одну стадию сборки, то общий порядок сборки, в котором можно соединить строительные блоки из нуклеиновых кислот, также определяется последовательностью стадий сборки. В одном из аспектов ренатурированные строительные элементы обрабатывают ферментом, таким как лигаза (например, ДНК-лигаза T4), чтобы добиться ковалентного связывания строительных элементов.
В одном из аспектов конструкцию олигонуклеотидных строительных блоков получают посредством анализа набора родительских матричных последовательностей нуклеиновых кислот, которые выполняют функцию основы для получения дочернего набора законченных химерных полинуклеотидов. Таким образом, эти исходные олигонуклеотидные матрицы выполняют функцию источника информации о последовательности, которая помогает в конструировании строительных блоков из нуклеиновых кислот, которые подлежат введению мутаций, например, подлежат химеризации или шаффлингу. В одном из аспектов этого способа последовательности нескольких исходных матриц нуклеиновых кислот выравнивают для того, чтобы выбрать одну или несколько точек разграничения. Точки разграничения могут размещаться в области гомологии и состоять из одного или нескольких нуклеотидов. Альтернативно эти точки разграничения являются общими по меньшей мере для двух родительских матриц. Посредством этого точки разграничения можно использовать для определения границ олигонуклеотидных строительных блоков, которые должны быть созданы для того, чтобы перегруппировать исходные полинуклеотиды. Точки разграничения, идентифицированные и выбранные в родительских молекулах выполняют функцию возможных точек химеризации при сборке конечных химерных дочерних молекул. Точка разграничения может представлять собой область гомологии (состоящую по меньшей мере из одного гомологичного нуклеотидного основания), которая является общей по меньшей мере для двух исходных полинуклеотидных последовательностей. Альтернативно точка разграничения может представлять собой область гомологии, которая является общей по меньшей мере для половины исходных полинуклеотидных последовательностей, или она может представлять собой область гомологии, которая является общей по меньшей мере для двух третей исходных полинуклеотидных последовательностей. В альтернативных вариантах осуществления пригодная точка разграничения представляет собой область гомологии, которая является общей по меньшей мере для трех четвертей исходных полинуклеотидных последовательностей, или она может быть общей почти для всех исходных полинуклеотидных последовательностей. В одном из аспектов точка разграничения представляет собой область гомологии, которая является общей для всех исходных полинуклеотидных последовательностей.
В одном из аспектов, процесс перегруппировки лигированием выполняют в полной мере для того, чтобы создать исчерпывающую библиотеку дочерних химерных полинуклеотидов. Другими словами, все возможные упорядоченные сочетания строительных блоков из нуклеиновых кислот представлены в наборе законченных химерных молекул нуклеиновых кислот. В то же время в другом аспекте порядок сборки (т.е. порядок сборки всех строительных блоков в 5'-3' направлении в каждой законченной химерной нуклеиновой кислоте) в каждом сочетании является предумышленным (или нестохастически), как описано выше. В настоящем документе предоставлены нестохастические способы, которые снижают вероятность получения нежелательных побочных продуктов.
В другом аспекте способ перегруппировки лигированием выполняют систематично. Например, способ выполняют для того, чтобы создать систематично разделенную библиотеку дочерних молекул, с компартментами, скрининг которых можно проводить систематично, например, поочередно. В настоящем документе предоставлены способы, включающие избирательное и целесообразное использование конкретных строительных блоков из нуклеиновых кислот в сочетании с избирательным и целесообразным использованием последовательно идущих реакций сборки, и можно создать конструкцию, причем в каждом из нескольких реакционных сосудов получают конкретные наборы дочерних продуктов. Это позволяет проводить систематическое исследование и процедуру скрининга. Таким образом, эти способы позволяют систематично исследовать теоретически очень большое число дочерних молекул небольшими группами. По причине их способности выполнять химеризацию очень гибким, а также исчерпывающим и систематическим образом, конкретно при низком уровне гомологии с родительскими молекулами, эти способы предусматривают создание библиотеки (или набора), состоящей из большого числа дочерних молекул. В силу нестохастической природы текущих способов перегруппировки лигированием, созданные дочерние молекулы могут содержать библиотеку законченных химерных молекул нуклеиновой кислоты, обладающих общим порядком сборки, который выбран преднамеренно.
Насыщающий мутагенез и способы оптимизированной направленной эволюции также можно использовать для создания различных категорий дочерних молекул.
В одном из аспектов в настоящем документе способы предусматривают свободу выбора и контроль относительно выбора точек разграничения, размера и числа строительных блоков из нуклеиновых кислот и размера и конфигурации соединения. Требования к межмолекулярной гомологии можно значительно снизить. Фактически, точки разграничения можно выбрать даже в областях небольшой межмолекулярной гомологии или в областях, где гомология отсутствует. Например, в силу неоднозначности кодона, т.е. вырожденности кодонов, нуклеотидные замены можно вводить в строительные блоки из нуклеиновых кислот без изменения аминокислоты, исходно закодированной в соответствующей родительской матрице. Альтернативно кодон можно изменить так, что изменится кодирование исходной аминокислоты. В одном из аспектов замены можно вводить в строительный блок из нуклеиновой кислоты для того, чтобы увеличить содержание межмолекулярных гомологичных точек разграничения и, таким образом, увеличить число соединений, которое можно выполнить между строительными блоками, что в свою очередь позволяет создать большее число дочерних химерных молекул.
В другом аспекте синтетическая природа стадии, на которой создают строительные блоки, позволяет проектировать и вводить нуклеотиды (например, один или несколько нуклеотидов, которые могут представлять собой, например, кодоны или интроны или регуляторные последовательности), которые позднее можно необязательно удалить в процессе in vitro (например, посредством мутагенеза) или в процессе in vivo (например, используя способность организма-хозяина к сплайсингу генов). Важно, что во многих случаях введение этих нуклеотидов также может быть желательным по многим другим причинам в дополнение к потенциальной выгоде создания пригодной точки разграничения.
В одном из аспектов строительный блок из нуклеиновой кислоты используют для введения интрона. Таким образом, функциональные интроны вводят в синтетический ген, полученный согласно способам, описанным в настоящем документе. Искусственно введенный интрон(ы) может принимать участие в сплайсинге генов в клетках-хозяевах примерно таким же образом, как встречающиеся в природе интроны принимают участие в сплайсинге генов.
Система оптимизированной направленной эволюции
В определенных вариантах осуществления в настоящем документе предоставлены системы нестохастической модификации генов, названные «система оптимизированной направленной эволюции» для создания гидролазы и антител с новыми или измененными свойствами. Оптимизированная направленная эволюция направлена на использование повторяющихся циклов редуктивной пересортировки, рекомбинации и отбора, которые позволяют осуществлять направленную молекулярную эволюцию нуклеиновых кислот посредством рекомбинации. Оптимизированная направленная эволюция позволяет создавать большую популяцию эволюционировавших химерных последовательностей, где созданная популяция в значительной степени обогащена последовательностями, которые имеют предварительно определенное число событий перекреста.
Событие перекреста представляет собой точку в химерной последовательности, где в последовательности происходит переключение с одного исходного варианта на другой исходный вариант. Обычно такая точка расположена в месте соединения, где олигонуклеотиды от двух родителей лигируют вместе, чтобы получить одну последовательность. Этот способ позволяет вычислить правильные концентрации олигонуклеотидных последовательностей с тем, чтобы обогатить конечную химерную популяцию последовательностей выбранным числом событий перекреста. Это обеспечивает больший контроль в выборе химерных вариантов, обладающих предварительно определенным числом событий перекреста.
Кроме того, этот способ предоставляет подходящие средства для исследования пространства возможных вариантов белков устрашающих размеров по сравнению с другими системами. Раньше, создав в ходе реакции, например, 1013 химерных молекул, было чрезвычайно трудно тестировать такое большое число химерных вариантов на конкретную активность. Кроме того, значительная часть дочерней популяции будет иметь очень большое число событий перекреста, что приводило к белкам, которые с меньшей вероятностью обладали повышенными уровнями конкретной активности. Используя эти способы, популяцию химерных молекул можно обогатить теми вариантами, которые имеют конкретное число событий перекреста. Таким образом, несмотря на то, что в ходе реакции все же можно создать 1013 химерных молекул, каждая молекула, выбранная для дальнейшего анализа, с большей вероятностью имеет, например, только три события перекреста. Поскольку полученную дочернюю популяцию можно изменить в сторону предварительно определенного числа событий перекреста, границы функционального разнообразия между химерными молекулами сужены. Это позволяет лучше контролировать число переменных при вычислении того, какой олигонуклеотид из исходных полинуклеотидов может отвечать за изменение конкретного признака.
Один способ создания последовательности химерного дочернего полинуклеотида состоит в создании олигонуклеотидов, соответствующих фрагментам или частям каждой исходной последовательности. В альтернативных вариантах осуществления каждый олигонуклеотид включает уникальный участок перекрытия тем, чтобы смешивание олигонуклеотидов вместе приводило к новому варианту, который содержит каждый олигонуклеотидный фрагмент, расположенный в правильном порядке. Альтернативно протоколы для осуществления этих способов по изобретению можно найти в патентах США №№ 6773900; 6740506; 6713282; 6635449; 6605449; 6537776; 6361974.
Число олигонуклеотидов, созданных для каждого исходного варианта, дает связь с общим числом полученных перекрестов в химерной молекуле, созданной в конечном счете. Например, можно предоставить три варианта исходной нуклеотидной последовательности для проведения реакции лигирования для того, чтобы найти химерный вариант обладающий, например, более высокой активностью при высокой температуре. В качестве одного из примеров можно создать набор из 50 олигонуклеотидных последовательностей, соответствующих каждой части каждого исходного варианта. Таким образом, в ходе процесса перегруппировки лигированием в каждой из химерных последовательностей могут иметь место вплоть до 50 событий перекреста. Очень низка вероятность того, что каждый из созданных химерных полинуклеотидов будет содержать олигонуклеотиды из каждого исходного варианта в альтернативном порядке. Если каждый олигонуклеотидный фрагмент присутствует в реакции лигирования в равном молярном количестве, существует вероятность того, что в некоторых положениях олигонуклеотиды из одного и того же исходного полинуклеотида будут лигированы друг за другом и, таким образом, это не приведет к событию перекреста. Если в этом примере концентрацию каждого олигонуклеотида из каждой родительского полинуклеотида сохраняют постоянной в ходе стадии лигирования, существует вероятность, равная 1/3 (предполагая 3 родительских полинуклеотида), что олигонуклеотид из одного и того же исходного варианта будет лигирован в химерной последовательности и не приведет к образованию перекреста.
Таким образом, можно определить функцию плотности вероятности (ФПВ) для предсказания популяции событий перекреста, которые вероятно произойдут в ходе каждой стадии в реакции лигирования при заданном числе исходных вариантов, числе олигонуклеотидов, соответствующем каждому варианту, и концентрациях каждого варианта в ходе каждой стадии в реакции лигирования. Статистический и математический аспекты определения ФПВ описаны ниже. Используя эти способы, можно вычислить такую функцию плотности вероятности и, таким образом, обогатить химерную дочернюю популяцию предварительно определенным числом событий перекреста в результате конкретной реакции лигирования. Кроме того, предварительно можно определить целевое число событий перекреста, и затем запрограммировать систему на вычисление стартовых количеств каждого исходного олигонуклеотида в ходе каждой стадии в реакции лигирования, чтобы получить функцию плотности вероятности, которая сосредоточена на предварительно определенном числе событий перекреста. Эти способы направлены на использование повторяющихся циклов редуктивной пересортировки, рекомбинации и отбора, которые позволяют осуществлять направленную молекулярную эволюцию нуклеиновых кислот, кодирующих полипептиды, посредством рекомбинации. Эта систем позволяет создавать большую популяцию эволюционировавших химерных последовательностей, где созданная популяция в значительной степени обогащена последовательностями, которые имеют предварительно определенное число событий перекреста. Событие перекреста представляет собой точку в химерной последовательности, где в последовательности происходит переключение с одного исходного варианта на другой исходный вариант. Обычно такая точка расположена в месте соединения, где олигонуклеотиды от двух родителей лигируют вместе, чтобы получить одну последовательность. Способ позволяет вычислить правильные концентрации олигонуклеотидных последовательностей с тем, чтобы обогатить конечную химерную популяцию последовательностей выбранным числом событий перекреста. Это обеспечивает больший контроль в выборе химерных вариантов, обладающих предварительно определенным числом событий перекреста.
Определение событий перекреста
Аспекты по изобретению включают систему и программное обеспечение, которые в качестве входных данных получают желаемую функцию плотности вероятности (ФПВ) перекрестов, число родительских генов, подлежащих перегруппировке, и число фрагментов в перегруппировке. Выходными данными этой программы является «ФПВ фрагментов», которую можно использовать для определения состава для получения перегруппированных генов, и ФПВ предполагаемых перекрестов этих генов. Описанную в настоящем документе обработку альтернативно выполняют, используя MATLAB™ (The Mathworks, Natick, Massachusetts), язык программирования и среду разработки для промышленных вычислений.
Итеративные процессы
В определенных вариантах осуществления в настоящем документе предоставлены процессы, которые можно повторять итеративно. Например, идентифицируют нуклеиновую кислоту (или конкретную нуклеиновую кислоту), отвечающую за измененный фенотип гидролазы или антитела, повторно выделяют, снова модифицируют, повторно тестируют на активность. Этот процесс можно повторять итеративно до тех пор, пока не будет сконструирован желаемый фенотип. Например, в клетке можно сконструировать целый биохимический анаболический или катаболический путь, включающий протеолитическую активность.
Подобным образом, если установлено, что конкретный олигонуклеотид совсем не оказывает влияния на желаемый признак (например, новый фенотип гидролазы), его можно устранить в качестве переменной посредством синтеза более длинных исходных олигонуклеотидов, которые включают последовательность, подлежащую устранению. Поскольку встраивание последовательности внутри более длинной последовательности предотвращает любые события перекреста, в дочерних полинуклеотидах будут отсутствовать любые изменения, связанные с этой последовательностью. Этот итеративный способ определения тех олигонуклеотидов, которые в наибольшей степени связаны с желаемым признаком и которые не связаны, позволяет более эффективно исследовать все возможные варианты белков, которые могут обеспечивать конкретный признак или активность.
Шаффлинг in vivo
Шаффлинг in vivo молекул используют в способах по изобретению, которые предоставляют варианты предоставленных в настоящем документе полипептидов, например, антител, гидролаз и т.п. Шаффлинг in vivo можно осуществлять, используя природное свойство клеток осуществлять рекомбинацию мультимеров. Тогда как рекомбинация in vivo обеспечивает основной естественный путь к молекулярному разнообразию, генетическая рекомбинация остается относительно сложным процессом, который включает 1) распознавание гомологии; 2) расщепление нитей, внедрение нитей и метаболические стадии, ведущие к получению рекомбинантной хиазмы; и, наконец, 3) разрешение хиазмы на отдельные рекомбинированные молекулы. Формирование хиазмы требует распознавания гомологичных последовательностей.
В определенных вариантах осуществления в настоящем документе предоставлены способы получения гибридного полинуклеотида по меньшей мере из первого полинуклеотида и второго полинуклеотида. В других вариантах осуществления в настоящем документе предоставлены способы, используемые для получения гибридного полинуклеотида посредством введения по меньшей мере первого полинуклеотида и второго полинуклеотида, которые имеют по меньшей мере один общий участок частичной гомологии последовательностей, в подходящую клетку-хозяина. Участки частичной гомологии последовательностей содействуют процессами, которые ведут к реорганизации последовательностей с получением гибридного полинуклеотида. В одном из аспектов термин «гибридный полинуклеотид» охватывает любые нуклеотидные последовательности, полученные в результате способа по изобретению, и в одном из вариантов осуществления содержат последовательности по меньшей мере из двух исходных полинуклеотидных последовательностей. Такие гибридные полинуклеотиды можно получить в результате событий межмолекулярной рекомбинации, которые содействуют интеграции последовательностей между молекулами ДНК. Кроме того, такие гибридные полинуклеотиды можно получить в результате процесса внутримолекулярной редуктивной пересортировки, в котором используют повторяющиеся последовательности для изменения нуклеотидной последовательности в молекуле ДНК.
Получение вариантов последовательностей
В определенных вариантах осуществления в настоящем документе предоставлены способы получения вариантов последовательностей нуклеиновых кислот и последовательностей гидролаз и антител по изобретению, или выделения гидролаз с использованием нуклеиновых кислот и предоставленных в настоящем документе полипептидов. В определенных вариантах осуществления в настоящем документе предоставлены варианты гена гидролазы по изобретению, которые можно изменять любыми средствами, включая, например, случайные или стохастические способы, или нестохастические способы, или «направленную эволюцию», как описано выше.
В настоящем документе предоставлены способы создания варианта нуклеиновой кислоты, который кодирует полипептид, обладающий гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, включающие стадии: (a) предоставление матричной нуклеиновой кислоты, содержащей нуклеиновую кислоту по изобретению; и (b) модификация, удаление или добавление одного или нескольких нуклеотидов в матричной последовательности или их сочетание для создания варианта матричной нуклеиновой кислоты. В одном из аспектов способ дополнительно может содержать экспрессию варианта нуклеиновая кислота для создания варианта полипептида гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы. Модификации, вставки или делеции можно вводить способом, включающим допускающую ошибки ПЦР, шаффлинг, олигонуклеотид-специфический мутагенез, сборочную ПЦР, половой ПЦР мутагенез, мутагенез in vivo, кассетный мутагенез, рекурсивный множественный мутагенез, экспоненциальный множественный мутагенез, сайт-специфический мутагенез, Gene Site Saturation MutagenesisSM (GSSMSM), синтетическую перегруппировку лигированием (SLR или GeneReassembly) или их сочетание. В другом аспекте модификации, вставки или делеции вводят способом, включающим рекомбинацию, рекурсивную рекомбинацию последовательностей, мутагенез модифицированной фосфотиоатом ДНК, мутагенез содержащей урацил матрицы, мутагенез двойной спирали с разрывами, точечный мутагенез при репарации ошибочно спаренных оснований, мутагенез в штамме-хозяине с недостаточностью репарации, химический мутагенез, радиогенный мутагенез, мутагенез делецией, мутагенез рестрикцией-отбором, мутагенез рестрикцией-очисткой, синтез искусственных генов, множественный мутагенез, создание мультимера химерной нуклеиновой кислоты и их сочетание.
В одном из аспектов, способ можно повторять итеративно до тех пор, пока не будет получена гидролаза, например, липаза, сатураза, пальмитаза и/или стеаратаза, обладающая измененной или отличающейся активностью или измененной или отличающейся стабильностью относительно того полипептида, который кодирует матричная нуклеиновая кислота. В одном из аспектов вариант полипептида гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы, является термостабильным и частично сохраняет активность после воздействия повышенной температуры. В другом аспекте вариант полипептида гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы, обладает повышенным гликозилированием по сравнению с гидролазой, например, липазой, сатуразой, пальмитазой и/или стеаратазой, кодируемой матричной нуклеиновой кислотой. Альтернативно вариант полипептида гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы обладает гидролазной, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью при высокой температуре, где гидролаза, например, липаза, сатураза, пальмитаза и/или стеаратаза, кодируемая матричной нуклеиновой кислотой, не активна при высокой температуре. В одном из аспектов способ можно повторять итеративно до тех пор, пока не будет получена кодирующая последовательность гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы, обладающая измененными частотами использования кодонов относительно матричной нуклеиновой кислоты. В другом аспекте способ итеративно можно повторять до тех пор, пока не будет получен ген гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы, обладающий более высокими или более низкими уровнями экспрессии транскрипта или стабильностью относительно матричной нуклеиновой кислоты. В другом аспекте состав конечного гидролазного продукта, например, липазного, сатуразного, пальмитазного и/или стеаратазного продукта, дает возможность увеличения или модуляции характеристик гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы, в продукте.
Выделенные варианты могут встречаться в природе. Также варианты можно создать in vitro. Варианты можно создать с использованием способов генетической инженерии, таких как сайт-специфический мутагенез, случайный химический мутагенез, процедуры делеции экзонуклеазой III и стандартные способы клонирования. Альтернативно, такие варианты, фрагменты, аналоги или производные можно создать с использованием химического синтеза или процедур модификации. Другие способы получения вариантов также известны специалистам в данной области. Они включают процедуры, в которых последовательности нуклеиновых кислот, полученные из природных изолятов, модифицируют для создания нуклеиновых кислот, которые кодируют полипептиды, обладающие характеристиками, которые увеличивают их ценность для промышленных и лабораторных применений. В таких процедурах создают и характеризуют большое число вариантов последовательностей, обладающих отличиями в одном или нескольких нуклеотидах относительно последовательности, полученной из природного изолята. Эти отличия нуклеотидов могут вести к аминокислотным заменам относительно полипептидов, кодируемых нуклеиновыми кислотами из природных изолятов.
Например, можно создать варианты с использованием допускающей ошибки ПЦР. В допускающей ошибки ПЦР, ПЦР проводят в условиях, где ДНК полимераза обладает низкой точностью копирования, так что вдоль всей длины продукта ПЦР получают большую долю точечных мутаций. Допускающая ошибки ПЦР описана, например, в Leung, D.W., et al., Technique, 1:11-15, 1989) и Caldwell, R.С. & Joyce G.F., PCR Methods Applic, 2:28-33, 1992. В кратком изложении, в таких процедурах нуклеиновые кислоты, подлежащие мутагенезу, смешивают с праймерами для ПЦР, реакционным буфером, MgCl2, MnCl2, полимеразой Taq и dNTP в подходящих концентрациях для достижения большой доли точечных мутаций вдоль всей длины продукта ПЦР. Например, реакцию можно проводить с использованием 20 пмоль нуклеиновой кислоты, подлежащей мутагенезу, 30 пмоль каждого праймера для ПЦР, реакционного буфера, содержащего 50 мМ KCl, 10 мМ Tris HCl (pH 8,3) и 0,01% желатина, 7 мМ MgCl2, 0,5 мМ MnCl2, 5 единиц полимеразы Taq, 0,2 мМ dGTP, 0,2 мМ dATP, 1 мМ dCTP и 1 мМ dTTP. ПЦР можно выполнять в течение 30 циклов при температуре 94°C в течение 1 мин, 45°C в течение 1 мин и 72°C в течение 1 мин. Однако следует понимать, что эти параметры можно менять в случае необходимости. Мутировавшие нуклеиновые кислоты клонируют в подходящий вектор и оценивают активности полипептидов, кодируемых мутировавшими нуклеиновыми кислотами.
Также варианты можно создать с использованием олигонуклеотид-специфического мутагенеза для создания сайт-специфических мутаций в любой клонированной ДНК, представляющей интерес. Мутагенез олигонуклеотидов описан, например, в Reidhaar-Olson (1988) Science 241:53-57. В кратком изложении, в таких процедурах синтезируют и встраивают в клонированную ДНК, подлежащую мутагенезу, несколько двухцепочечных олигонуклеотидов, несущих одну или несколько мутаций для введения в клонированную ДНК. Собирают клоны, содержащие мутировавшую ДНК, и оценивают активности кодируемых ей полипептидов.
Другим способом создания вариантов является сборочная ПЦР. Сборочная ПЦР включает сборку продукта ПЦР из смеси маленьких фрагментов ДНК. В одном сосуде параллельно происходит большое число различных ПЦР, при этом продукты одной реакции являются праймерами для продуктов другой реакции. Сборочная ПЦР описана, например, в патенте США № 5965408.
Еще одним способом создания вариантов является половой ПЦР мутагенез. В половом ПЦР мутагенезе между молекулами ДНК с различными, но близкородственными последовательностями ДНК, происходит принудительная гомологичная рекомбинация in vitro в результате случайного фрагментирования молекулы ДНК на основе гомологии последовательностей, с последующей фиксацией и перекрестом посредством удлинения праймеров в реакции ПЦР. Половой ПЦР мутагенез описан, например, в Stemmer (1994) Proc. Natl. Acad. Sci. USA 91:10747-10751. В кратком изложении, в таких процедурах несколько нуклеиновых кислот, подлежащих рекомбинации, расщепляют ДНКазой для создания фрагментов со средней длиной 50-200 нуклеотиды. Фрагменты с желаемой средней длиной очищают и ресуспендируют в смеси для ПЦР. ПЦР проводят в условиях, которые содействуют рекомбинации между фрагментами нуклеиновых кислот. Например, ПЦР можно проводить, ресуспендировав очищенные фрагменты в концентрации 10-30 нг/л в растворе, содержащем по 0,2 мМ каждого dNTP, 2,2 мМ MgCl2, 50 мМ KCL, 10 мМ Tris HCl, pH 9,0 и 0,1% Triton X-100. Добавляют 2,5 единицы полимеразы Taq на 100:1 реакционной смеси и проводят ПЦР с использованием следующего режима: 94°C в течение 60 секунд, 94°C в течение 30 секунд, 50-55°C в течение 30 секунд , 72°C в течение 30 секунд (30-45 циклов) и 72°C в течение 5 минут. Однако следует понимать, что в случае необходимости эти параметры можно менять. В некоторых аспектах в ПЦР можно включить олигонуклеотиды. В других аспектах фрагмент Кленова ДНК полимеразы I можно использовать в первом наборе ПЦР, а полимеразу Taq можно использовать в последующем наборе ПЦР. Выделяют рекомбинантные последовательности и оценивают активности кодируемых ими полипептидов.
Также варианты можно создать посредством мутагенеза in vivo. В некоторых аспектах создают случайные мутации в последовательности, представляющей интерес, посредством клонирования последовательности, представляющей интерес, в бактериальном штамме, таком как штамм E. coli, который несет мутации в одном или нескольких путях репарации ДНК. Такие «мутаторные» штаммы обладают более высоким уровнем случайных мутаций по сравнению с родительским штаммом дикого типа. Клонирование ДНК в одном из этих штаммов в конечном счете ведет к созданию случайных мутаций в ДНК. Мутаторные штаммы, пригодные для использования в мутагенезе in vivo, описаны, например, в публикации PCT № WO 91/16427.
Также варианты можно создать, используя кассетный мутагенез. В кассетном мутагенезе небольшой участок двухцепочечной молекулы ДНК замещают синтетической олигонуклеотидной «кассетой» которая отличается от нативной последовательности. Часто олигонуклеотид содержит полностью и/или частично рандомизированную нативную последовательность.
Рекурсивный множественный мутагенез также можно использовать для создания вариантов. Рекурсивный множественный мутагенез представляет собой алгоритм для конструирования белков (белковый мутагенез), разработанный для получения разнообразных популяций фенотипически родственных мутантов, члены которых отличаются аминокислотными последовательностями. В этом способе используют механизм обратной связи для контроля последующих раундов комбинаторного кассетного мутагенеза. Рекурсивный множественный мутагенез описан, например, в Arkin (1992) Proc. Natl. Acad. Sci. USA 89:7811-7815.
В некоторых аспектах варианты создают с использованием экспоненциального множественного мутагенеза. Экспоненциальный множественный мутагенез представляет собой процесс для создания комбинаторных библиотек с высоким процентным содержанием уникальных и функциональных мутантов, где параллельно рандомизируют небольшие группы остатков для идентификации в каждом измененном положении аминокислот, которые ведут к функциональным белкам. Экспоненциальный множественный мутагенез описан, например, в Delegrave (1993) Biotechnology Res. 11:1548-1552. Случайный и сайт-специфический мутагенез описан, например, в Arnold (1993) Current Opinion in Biotechnology 4:450-455.
В некоторых аспектах варианты создают с использованием процедур шаффлинга, где части нескольких нуклеиновых кислот, которые кодируют определенные полипептиды, сливают вместе для создания химерных последовательностей нуклеиновых кислот, которые кодируют химерные полипептиды, как описано, например, в патентах США №№ 5965408; 5939250.
В настоящем документе предоставлены варианты полипептидов, содержащие последовательности, в которых один или несколько аминокислотных остатков (например, образцового полипептида, например, SEQ ID № 2, SEQ ID № 4, SEQ ID № 6, SEQ ID № 8, SEQ ID № 10, SEQ ID № 12, SEQ ID № 14, SEQ ID № 16, SEQ ID № 18 или SEQ ID № 20 или SEQ ID № 2, содержащего изменения одной, двух, трех, четырех, пяти, шести, семи, восьми или более (нескольких) или всех аминокислот, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, или их эквиваленты) заменены на консервативные или неконсервативные аминокислотные остатки (например, консервативные аминокислотные остатки) и такие замененные аминокислотные остатки могут кодироваться или могут не кодироваться генетическим кодом. Консервативные замены представляют собой замены, которые заменяют данную аминокислоту в полипептиде на другую аминокислоту с похожими характеристиками. Таким образом, полипептиды в настоящем документе включают полипептиды с консервативными заменами в последовательностях, например, в образцовых последовательностях по изобретению (например, в SEQ ID № 2, SEQ ID № 4, SEQ ID № 6, SEQ ID № 8, SEQ ID № 10, SEQ ID № 12, SEQ ID № 14, SEQ ID № 16, SEQ ID № 18 или SEQ ID № 20 или SEQ ID № 2, содержащей изменения одной, двух, трех, четырех, пяти, шести, семи, восьми или более (нескольких) или всех аминокислот, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, или их эквиваленты), включая в качестве неограничивающих примеров следующие замены: замена алифатической аминокислоты, такой как аланин, валин, лейцин и изолейцин, на другую алифатическую аминокислоту; замена серина на треонин или наоборот; замена кислого остатка, такого как аспарагиновая кислота и глутаминовая кислота, на другой кислый остаток; замена остатка, несущего аминогруппу, такую как аспарагин и глутамин, на другой остаток, несущий аминогруппу; замена основного остатка, такого как лизин и аргинин, на другой основной остаток; и замена ароматического остатка, такого как фенилаланин, тирозин или триптофан, на другой ароматический остаток. В других вариантах один или несколько аминокислотных остатков предоставленных в настоящем документе полипептидов содержат замещающую группу.
Другие варианты, входящие в объем изобретения, представляют собой те, в которых полипептид связан с другим соединением, таким как соединение для увеличения времени полужизни полипептида, например, с полиэтиленгликолем. Дополнительные варианты, входящие в объем изобретения, представляют собой те, в которых дополнительные аминокислоты слиты с полипептидом, такие как лидерная последовательность, секреторная последовательность, пропоследовательность белка или последовательность, которая облегчает очистку, обогащение или стабилизацию полипептида. В некоторых аспектах варианты, фрагменты, производные и аналоги предоставленных в настоящем документе полипептидов сохраняют ту же биологическую функцию или активность, что и образцовые полипептиды, например, протеолитическую активность, как описано в настоящем документе. В других аспектах вариант, фрагмент, производное или аналог включает такой пропротеин, что вариант, фрагмент, производное или аналог можно активировать отщеплением части пропротеина для получения активного полипептида.
Оптимизация кодонов для достижения высоких уровней экспрессии белка в клетках-хозяевах
В определенных вариантах осуществления в настоящем документе предоставлены способы для модификации нуклеиновых кислот, кодирующих гидролазы, чтобы модифицировать частоту использования кодонов. В одном из вариантов осуществления в настоящем документе предоставлены способы модификации кодонов в нуклеиновой кислоте, кодирующей гидролазу, для повышения или снижения ее экспрессии в клетке-хозяине, например, клетке бактерии, насекомого, млекопитающего, дрожжей или растения. Кроме того, в настоящем документе предоставлены нуклеиновые кислоты, кодирующие гидролазу, модифицированную для повышения ее экспрессии в клетке-хозяине, модифицированная таким образом гидролаза и способы получения модифицированных гидролаз. Способ включает идентификацию «непредпочтительного» или «менее предпочтительного» кодона в нуклеиновой кислоте, кодирующей гидролазу, и замену одного или нескольких этих непредпочтительных или менее предпочтительных кодонов на «предпочтительный кодон», кодирующий ту же аминокислоту, что и замененный кодон, и в нуклеиновой кислоте по меньшей мере один непредпочтительный или менее предпочтительный кодон замещен на предпочтительный кодон, кодирующий ту же аминокислоту. Предпочтительный кодон представляет собой кодон, более часто представленный в кодирующих последовательностях в генах в клетке-хозяине, а непредпочтительный или менее предпочтительный кодон представляет собой кодон, менее часто представленный в кодирующих последовательностях в генах в клетке-хозяине.
Клетки-хозяева для экспрессии нуклеиновых кислот, экспрессирующих кассеты и векторы по изобретению, включают клетки бактерий, дрожжей, грибов, растений, насекомых и млекопитающих. В определенных вариантах осуществления в настоящем документе предоставлены способы оптимизации частоты использования кодонов во всех этих клетках, нуклеиновые кислоты с измененными кодонами и полипептиды, полученные посредством нуклеиновых кислот с измененными кодонами. Образцовые клетки-хозяева включают грамм-отрицательные бактерии, такие как Escherichia coli и Pseudomonas fluorescens; грамм-положительные бактерии, такие как Lactobacillus gasseri, Lactococcus lactis, Lactococcus cremoris, Bacillus subtilis. Образцовые клетки-хозяева также включают эукариотические организмы, например, различные дрожжи, такие как Saccharomyces sp., включая Saccharomyces cerevisiae, Schizosaccharomyces pombe, Pichia pastoris и Kluyveromyces lactis, Hansenula polymorpha, Aspergillus niger и клетки и клеточные линии млекопитающих и клетки и клеточные линии насекомых. Другие образцовые клетки-хозяева включают бактериальные клетки, такие как E. coli, Streptomyces, Bacillus subtilis, Bacillus cereus, Salmonella typhimurium и различные виды родов Pseudomonas, Streptomyces и Staphylococcus, клетки грибов, таких как Aspergillus, дрожжей, таких как любые виды Pichia, Saccharomyces, Schizosaccharomyces, Schwanniomyces, включая Pichia pastoris, Saccharomyces cerevisiae или Schizosaccharomyces pombe, клетки насекомых, такие как Drosophila S2 и Spodoptera Sf9, клетки животных, такие как CHO, COS или меланома Боуэса, и аденовирусы. Выбор подходящего организма-хозяина находится в пределах возможностей специалистов в данной области. В определенных вариантах осуществления в настоящем документе предоставлены нуклеиновые кислоты и полипептиды, оптимизированные для экспрессии в этих организмах и видах.
Например, кодоны нуклеиновой кислоты, кодирующей гидролазу, выделенную из бактериальной клетки, модифицируют так, что нуклеиновая кислота оптимально экспрессируется в клетке бактерии, отличной от бактерии, из которых получена гидролаза, дрожжей, грибов, растения, насекомого или млекопитающего. Способы оптимизации кодонов хорошо известны в данной области, см., например, патент США № 5795737; Baca (2000) Int. J. Parasitol. 30:113-118; Hale (1998) Protein Expr. Purif. 12:185-188; Narum (2001) Infect. Immun. 69:7250-7253. Также см. Narum (2001) Infect. Immun. 69:7250-7253, описание оптимизации кодонов в мышиных системах; Outchkourov (2002) Protein Expr. Purif. 24:18-24, описание оптимизации кодонов в дрожжах; Feng (2000) Biochemistry 39:15399-15409, описание оптимизации кодонов в E. coli; Humphreys (2000) Protein Expr. Purif. 20:252-264, описание оптимизации частоты использования кодонов, которая влияет на секрецию в E. coli.
Трансгенные животные, не относящиеся к человеку
В определенных вариантах осуществления в настоящем документе предоставлены трансгенные животные, не относящиеся к человеку, содержащие нуклеиновую кислоту, полипептид (например, гидролазу или антитело по изобретению), экспрессирующую кассету, вектор, трансфицированную или трансформированную клетку по изобретению. Трансгенные животные, не относящиеся к человеку, могут являться, например, козами, кроликами, овцами, свиньями, коровами, крысами и мышами, содержащим нуклеиновые кислоты по изобретению. Этих животных можно использовать, например, в качестве in vivo для исследования гидролазной активности или в качестве моделей для скрининга средств, которые изменяют гидролазную активность in vivo. Кодирующие последовательности полипептидов, подлежащие экспрессии в трансгенных животных, не относящихся к человеку, можно сконструировать так, чтобы они были конститутивными, или находились под управлением тканеспецифических, зависящих от стадии развития или индуцибельных транскрипционных регуляторных факторов. Трансгенные животные, не относящиеся к человеку, можно сконструировать и создать с использованием любого известного в данной области способа; см., например, патенты США №№ 6211428; 6187992; 6156952; 6118044; 6111166; 6107541; 5959171; 5922854; 5892070; 5880327; 5891698; 5639940; 5573933; 5387742; 5087571, описание получения и использования трансформированных клеток и яйцеклеток и трансгенных мышей, крыс, кроликов, овец, свиней и коров. Также см., например, Pollock (1999) J. Immunol. Methods 231:147-157, описание получения рекомбинантных белков в молоке трансгенных молочных животных; Baguisi (1999) Nat. Biotechnol. 17:456-461, демонстрация получения трансгенных коз. В патент США № 6211428 описано получении и использование трансгенных млекопитающих, не относящихся к человеку, в мозге которых экспрессируется конструкция из нуклеиновой кислоты, содержащая последовательность ДНК. В патенте США № 5387742 описана инъекция клонированной рекомбинантной или синтетической последовательности ДНК в оплодотворенные яйцеклетки мышей, имплантация яйцеклеток после инъекции псевдобеременным самкам и выращивание трансгенных мышей, клетки которых экспрессируют белки, связанные с патогенезом болезни Альцгеймера. В патенте США № 6187992 описано получение и использование трансгенных мышей, геном которых содержит нарушение в гене, кодирующем предшественник белка амилоида (APP).
«Животных с нокаутом» также можно использовать для осуществления способов по изобретению. Например, в одном из аспектов трансгенные или модифицированные животные по изобретению включают «животное с нокаутом», например, «мышь с нокаутом», созданное не для экспрессии эндогенного гена, который замещают геном, экспрессирующим гидролазу или слитый белок, содержащий гидролазу по изобретению. Как указано выше, функциональный нокаут также можно создать, используя антисмысловые последовательности по изобретению, например, двухцепочечные молекулы интерферирующей РНК.
Трансгенные растения и семена
В определенных вариантах осуществления в настоящем документе предоставлены трансгенные растения и семена, содержащие нуклеиновую кислоту, полипептид (например, гидролазу или антитело по изобретению), экспрессирующую кассету или вектор или трансфицированную или трансформированную клетку по изобретению. трансгенное растение может представлять собой двудольное или однодольное растение. В одном из вариантов осуществления в настоящем документе предоставлены способы получения и использования этих трансгенных растений и семян. Трансгенное растение или клетку растения, экспрессирующие полипептид по изобретению, можно сконструировать в соответствии с любым известным в данной области способом. См., например, патент США № 6309872.
Нуклеиновые кислоты и экспрессирующие конструкции по изобретению можно ввести в клетку растения любыми способами. Например, нуклеиновые кислоты или экспрессирующие конструкции можно ввести в геном желаемого растения-хозяина или нуклеиновые кислоты или экспрессирующие конструкции могут представлять собой эписомы. Введение в геном желаемого растения можно осуществить таким образом, что синтез гидролазы организмом-хозяином будет управляться эндогенными транскрипционными или трансляционными управляющими элементами. В одном из аспектов в настоящем документе предоставлены «растения с нокаутом», где встраивание последовательности гена посредством, например, гомологичной рекомбинации нарушает экспрессию эндогенного гена. Средства для создания растений «с нокаутом» хорошо известны в данной области, см., например, Strepp (1998) Proc Natl. Acad. Sci. USA 95:4368-4373; Miao (1995) Plant J 7:359-365. См. обсуждение трансгенных растений ниже.
Нуклеиновые кислоты по изобретению можно использовать, чтобы придать желаемые признаки по существу любому растению, например, масличным растениям, включая рисовую мучку, рапс (канолу), подсолнечник, оливу, пальму или сою и т.п., или растениям, образующим глюкозу или крахмал, таким как кукуруза, картофель, пшеница, рис, ячмень и т.п. Нуклеиновые кислоты по изобретению можно использовать для манипуляции метаболическими путями растения для того, чтобы оптимизировать или изменить экспрессию гидролазы организмом-хозяином или субстрат или продукт гидролазы, например, масло, липид, такой как моно-, ди- или триацилглицерид и т.п. Это позволяет изменить соотношения липидов, превращение липидов и обмен в растении. Это может способствовать промышленной обработке растения. Альтернативно гидролазы по изобретению можно использовать в производстве трансгенного растения для получения соединения, которое растение обычно не образует. Это позволяет снизить стоимость производства или создать новый продукт.
В одном из аспектов первая стадия получения трансгенного растения включает создание экспрессирующей конструкции для экспрессии в клетке растения. Эти способы хорошо известны в данной области. Они могут включать выбор и клонирование промотора, кодирующей последовательности для облегчения эффективного связывания рибосом с мРНК и выбор подходящих последовательностей терминаторов генов. Одним из примеров конститутивного промотора является CaMV35S из вируса мозаики цветной капусты, который, как правило, дает высокий уровень экспрессии в растениях. Другие промоторы более специфичны и реагируют на стимулы из внутренней или внешней среды растения. Образцовый индуцируемый светом промотор представляет собой промотор из гена cab, который кодирует основной белок, связывающий хлорофилл a/b.
В одном из аспектов нуклеиновую кислоту модифицируют для достижения более высокого уровня экспрессии в клетке растения. Например, последовательность по изобретению, вероятно, содержит более высокую процентную долю пар нуклеотидов A-T по сравнению с тем, что наблюдают у растений, некоторые из которых предпочитают пары нуклеотидов G-C. Следовательно, нуклеотиды A-T в кодирующей последовательности можно заменить на нуклеотиды G-C без значительного изменения аминокислотной последовательности для усиления синтеза продукта гена в клетках растения.
Ген селективного маркера можно присоединить к генной конструкции для того, чтобы идентифицировать клетки или ткани растения, в которых успешно прошло встраивание трансгена. Это может быть необходимо, поскольку встраивание и экспрессия генов в клетки растения является редким событием, которое происходит лишь в нескольких процентах целевых тканей или клеток. Гены селективных маркеров кодируют белки, которые обеспечивают устойчивость к средствам, которые обычно токсичны для растений, таким как антибиотики или гербициды. Только клетки растения со встроенным геном селективного маркера выживут при выращивании в среде, содержащей соответствующий антибиотик или гербицид. Что касается других введенных генов, маркерным генам для правильного функционирования также требуется промоторные и терминирующие последовательности.
В одном из аспектов создание трансгенных растений или семян включает встраивание последовательности по изобретению и, необязательно, маркерных генов в целевую экспрессирующую конструкцию (например, в плазмиду, фаг), наряду с размещением промоторных и терминирующих последовательностей. Это может включать перенос модифицированного гена в растение подходящим способом. Например, конструкцию можно ввести непосредственно в геномную ДНК клетки растения, используя такие способы, как электропорация и микроинъекция в протопласты клеток растений, или конструкции можно ввести непосредственно в ткань растения, используя баллистические способы, такие как бомбардировка частицами ДНК. Например, см., например, Christou (1997) Plant Mol. Biol. 35:197-203; Pawlowski (1996) Mol. Biotechnol. 6:17-30; Klein (1987) Nature 327:70-73; Takumi (1997) Genes Genet. Syst. 72:63-69, обсуждение использования бомбардировки частицами для введения трансгенов в пшеницу; и Adam (1997) выше, использование бомбардировки частицами для введения YAC в клетки растений. Например, Rinehart (1997) выше, использование бомбардировки частицами для создания трансгенных растений хлопка. Устройство для ускорения частиц описано в патенте США № 5015580; и коммерчески доступен прибор для ускорения частиц BioRad (Biolistics) PDS-2000; также см., John, патент США № 5608148; и Ellis, патент США № 5681730, описание трансформации голосемянных, опосредованной частицами.
В одном из аспектов можно иммобилизовать протопласты и инъецировать в них нуклеиновые кислоты, например, экспрессирующую конструкцию. Несмотря на то, что в случае зерновых регенерация растений из протопластов не является простой задачей, регенерация растения возможна в случае бобовых, используя соматический эмбриогенез из каллюса, полученного из протопласта. Организованные ткани можно трансформировать с использованием оголенной ДНК и способа генной пушки, в котором ДНК наносят на вольфрамовые микроснаряды, частицы размером в 1/100 от размера клеток, которые переносят ДНК глубоко внутрь клеток и органелл. Затем индуцируют регенерацию трансформированной ткани, обычно посредством соматического эмбриогенеза. Этот способ эффективен в случае с некоторыми видами зерновых, включая маис и рис.
Нуклеиновые кислоты, например, экспрессирующие конструкции, также можно вводить в клетки растений, используя рекомбинантные вирусы. Клетки растений можно трансформировать, используя вирусные векторы, такие как, например, векторы, полученные из вируса табачной мозаики (Rouwendal (1997) Plant Mol. Biol. 33:989-999), см. Porta (1996) «Use of viral replicons for the expression of genes in plants», Mol. Biotechnol. 5:209-221.
Альтернативно нуклеиновые кислоты, например, экспрессирующую конструкцию, можно комбинировать с подходящими фланкирующими участками T-ДНК и ввести в стандартный вектор организма-хозяина Agrobacterium tumefaciens. Болезнетворные функции организма-хозяина Agrobacterium tumefaciens будут управлять встраиванием конструкции и прилежащего маркера в ДНК клетки растения, когда эту клетку инфицируют этими бактериями. Способы трансформации, опосредованные Agrobacterium tumefaciens, включающие обезвреживание и использование бинарных векторов, подробно описаны в научной литературе. См., например, Horsch (1984) Science 233:496-498; Fraley (1983) Proc. Natl. Acad. Sci. USA 80:4803 (1983); Gene Transfer to Plants, Potrykus, ed. (Springer-Verlag, Berlin 1995). ДНК в клетке A. tumefaciens содержится в бактериальной хромосоме, а также в другой структуре, известной как Ti-плазмида (опухолеиндуцирующая). Ti-плазмида содержит фрагмент секвенирования ДНК, названный T-ДНК (~20 т.п.н. в длину), который переносят в клетку растения в процессе инфицирования, и ряд генов vir (вирулентность), которые управляют процессом инфицирования. A. tumefaciens может инфицировать растение только через повреждения: повреждение корня или стебля растения является причиной определенных химических сигналов, в ответ на которые происходит активация генов vir в A. tumefaciens и управление рядом событий, необходимых для переноса T-ДНК из Ti-плазмиды в хромосому растения. Затем T-ДНК попадает в клетку растения через повреждение. Согласно одному из предположений, T-ДНК ждет до тех пор, пока не начнется репликация или транскрипция ДНК растения, а затем встраивается в незащищенную ДНК растения. Для того чтобы использовать A. tumefaciens в качестве трансгенного вектора, следует удалить опухолеиндуцирующий сегмент T-ДНК, но сохранить граничные участки T-ДНК и гены vir. Затем трансген встраивают между граничными участками T-ДНК, где он переносится в клетку растения и встраивается в хромосомы растения.
В определенных вариантах осуществления в настоящем документе предоставлены способы трансформации односемядольных растений с использованием нуклеиновых кислот по изобретению, включая важные зерновые культуры, см. Hiei (1997) Plant Mol. Biol. 35:205-218. Также см., например, Horsch, Science (1984) 233:496; Fraley (1983) Proc. Natl. Acad. Sci USA 80:4803; Thykjaer (1997) выше; Park (1996) Plant Mol. Biol. 32:1135-1148, обсуждение встраивания T-ДНК в геномную ДНК. Также см. D'Halluin, патент США № 5712135, описание процесса стабильного встраивания ДНК, содержащей ген, работающий в клетке зернового или другого односемядольного растения.
В одном из аспектов третья стадия может включать отбор и регенерацию целых растений, способных передавать встроенный целевой ген следующему поколению. Такие способы регенерации основаны на манипуляции определенными фитогормонами в среде для выращивания тканевой культуры, как правило, на основании биоцидного и/или гербицидного маркера, который введен вместе с желаемыми нуклеотидными последовательностями. Регенерация растения из культивируемых протопластов описана в Evans et al., Protoplasts Isolation and Culture, Handbook of Plant Cell Culture, стр. 124-176, MacMillilan Publishing Company, New York, 1983; и Binding, Regeneration of Plants, Plant Protoplasts, стр. 21-73, CRC Press, Boca Raton, 1985. Также можно добиться регенерации из каллюса, эксплантатов, органов растения или их частей. В целом такие способы регенерации описаны в Klee (1987) Ann. Rev. of Plant Phys. 38:467-486. Для получения целых растений из трансгенных тканей, таких как незрелые зародыши, их можно выращивать в контролируемых условиях окружающей среды в ряде сред, содержащих питательные вещества и гормоны, процесс известен как тканевая культура. После создания целых растений и получения семян начинают оценку потомства.
После стабильного встраивания экспрессирующей кассеты в трансгенные растения, ее можно ввести в другие растения посредством скрещивания. Можно использовать любое число стандартных способов селекции, в зависимости от видов, подлежащих скрещиванию. Поскольку трансгенная экспрессия нуклеиновых кислот по изобретению ведет к фенотипическим изменениям, растения, содержащие рекомбинантные нуклеиновые кислоты по изобретению, можно скрестить со вторым растением для получения конечного продукта. Таким образом, семена по изобретению можно получить из скрещивания двух трансгенных растений по изобретению или скрещивания растения по изобретению и другого растения. Желаемые эффекты (например, экспрессия предоставленных в настоящем документе полипептидов для получения растения с измененным, повышенным и/или сниженным содержанием липидов или масел) можно усилить, когда оба исходных растения экспрессируют полипептиды по изобретению. Желаемые эффекты можно передавать последующим поколениями растений стандартными средствами размножения.
Нуклеиновые кислоты и полипептиды по изобретению вводят или экспрессируют в любом растении или семенах. Трансгенные растения по изобретению могут быть двусемядольными или односемядольными. Примерами односемядольных трансгенных растений по изобретению являются травянистые растения, такие как мятлик луговой (голубая трава, стручок), кормовая трава, такая как овсяница, плевел, травы умеренной климатической зоны, такие как Agrostis, и зерновые, например, пшеница, овес, рожь, ячмень, рис, сорго и маис (кукуруза). Примерами двусемядольных трансгенных растений по изобретению являются табак, бобовые, такие как люпины, картофель, сахарная свекла, горох, бобы и соевые бобы, и крестоцветные растения (семейство Brassicaceae), такие как цветная капуста, рапсовое семя, и близкородственный модельный организм Arabidopsis thaliana. Таким образом, трансгенные растения и семена по изобретению включают широкий диапазон растений, в качестве неограничивающих примеров включая виды из родов Anacardium, Arachis, Asparagus, Atropa, Avena, Brassica, Citrus, Citrullus, Capsicum, Carthamus, Cocos, Coffea, Cucumis, Cucurbita, Daucus, Elaeis, Fragaria, Glycin, Gossypium, Helianthus, Heterocallis, Hordeum, Hyoscyamus, Lactuca, Linum, Lolium, Lupinus, Lycopersicon, Malus, Manihot, Majorana, Medicago, Nicotiana, Olea, Oryza, Panieum, Pannisetum, Persea, Phaseolus, Pistachio, Pisum, Pyrus, Prunus, Raphanus, Ricinus, Secale, Senecio, Sinapis, Solarium, Sorghum, Theobromus, Trigonella, Triticum, Vicia, Vitis, Vigna и Zea.
В альтернативных вариантах осуществления нуклеиновые кислоты по изобретению экспрессируют в растениях, которые содержат клетки волокон, включая, например, хлопок, капок настоящий пятитычиночный (капок, Сейба пятитычинковая), губовник линейный, креозотовый куст, Krascheninnikovia, бальзу, рами, кенаф, коноплю, розеллу, джут, сизаль, абаку и лен. В альтернативных вариантах осуществления трансгенные растения по изобретению могут являться членами рода Gossypium, включая членов любых видов Gossypium, таких как G. arboreum; G. herbaceum, G. Barbadense и G. hirsutum.
В настоящем документе в определенных вариантах осуществления трансгенные растения можно использовать для получения больших количеств полипептидов (например, антител, гидролаз) по изобретению. Например, см. Palmgren (1997) Trends Genet. 13:348; Chong (1997) Transgenic Res. 6:289-296 (получения белка β-казеина молока человека в трансгенных растениях картофеля с использованием ауксин-индуцибельного двунаправленного промотора маннопин-синтазы (mas1',2') с использованием способов трансформации листовой пластинки, опосредованных Agrobacterium tumefaciens).
Используя известные процедуры, профессионал может провести скрининг растений по изобретению посредством определения повышения или снижения трансгенной мРНК или белка в трансгенных растениях. Средства для обнаружения и количественного определения мРНК или белков хорошо известны в данной области.
В настоящем документе предоставлены жирные кислоты или производные жирных кислот из трансгенных растений по изобретению, например, трансгенных масличных растений. В одном из аспектов получают трансгенные масличные растения, содержащие по меньшей мере одну гидролазу по изобретению. В одном из аспектов трансгенное растение содержит ген гидролазы, функционально связанный с промотором, обеспечивающим экспрессию гена в клеточных, внеклеточных или тканевых компартментах, отличающихся от тех, в которых накапливаются липиды растения, или обеспечивающим экзогенную индукцию гидролазы. В одном из аспектов собирают семена и/или плоды, содержащие липиды растения, семена и/или плоды измельчают (в случае необходимости после обработки, индуцирующей ген гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы)) с тем, чтобы привести в контакт липиды и гидролазу по изобретению, содержащиеся в семенах и/или плодах. Смесь можно инкубировать, чтобы предоставить возможность протекания ферментативного гидролиза липидов измельченного материала посредством каталитического действия липазы по изобретению, содержащейся в измельченном материале. В одном из аспектов жирные кислоты, созданные посредством гидролиза, экстрагируют и/или подвергают превращению для того, чтобы получить желаемые производные жирных кислот.
В этом процессе ферментативного гидролиза по изобретению используют мягкие рабочие условия и его можно проводить в малом масштабе с использованием недорогих установок. В этом аспекте растение по изобретению индуцируют для получения гидролазы для трансформации липидов растения. Используя эту стратегию, предотвращают контакт фермента с накопленными липидами растения с тем, чтобы избежать любого риска преждевременного гидролиза («саморасщепление растения») до уборки. Блоки для измельчения и инкубации могут быть легкими и мелкомасштабными; многие из них известны в сельскохозяйственной промышленности и могут использоваться в местах уборки растений.
В одном из аспектов трансгенные растения по изобретению получены трансформацией природных масличных растений. Затем генетически трансформированные растения по изобретению воспроизводят половым размножением с тем, чтобы получить трансгенные семена по изобретению. Эти семена можно использовать для получения потомства трансгенных растений.
В одном из аспектов ген гидролазы функционально связан с индуцируемым промотором для предотвращения какого-либо преждевременного контакта гидролазы и липида растения. Этот промотор может управлять экспрессией гена в компартментах, которые отличаются от тех, в которых накапливаются липиды, или промотор может инициировать экспрессию гидролазы в желаемый момент времени посредством экзогенной индукции.
Полипептиды и пептиды
В определенных вариантах осуществления в настоящем документе предоставлены выделенные, синтетические или рекомбинантные полипептиды, обладающие идентичностью последовательностей (например, по меньшей мере 50% идентичностью последовательностей) с SEQ ID № 2, SEQ ID № 4, SEQ ID № 6, SEQ ID № 8, SEQ ID № 10, SEQ ID № 12, SEQ ID № 14, SEQ ID № 16, SEQ ID № 18, или SEQ ID № 20 или SEQ ID № 2, содержащей изменение одной, двух, трех, четырех, пяти, шести, семи, восьми или более (нескольких) или всех аминокислот, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, или их эквиваленты. В определенных вариантах осуществления в настоящем документе предоставлены нуклеиновые кислоты, кодирующие полипептиды, содержащие последовательность, как указано в SEQ ID № 2, SEQ ID № 4, SEQ ID № 6, SEQ ID № 8, SEQ ID № 10, SEQ ID № 12, SEQ ID № 14, SEQ ID № 16, SEQ ID № 18, или SEQ ID № 20 или SEQ ID № 2, содержащей изменение одной, двух, трех, четырех, пяти, шести, семи, восьми или более (нескольких) или всех аминокислот, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, или их эквиваленты.
Идентичность последовательностей может иметь место в полноразмерном полипептиде или идентичность может иметь место на участке длиной по меньшей мере приблизительно 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700 или более остатков. Полипептиды по изобретению также могут быть короче полноразмерных образцовых полипептидов. В одном из аспектов в настоящем документе предоставлены полипептиды, содержащие только подпоследовательность последовательности по изобретению, образцовые подпоследовательности могут состоять приблизительно из 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 100, 125, 150, 175, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700 или более остатков. В альтернативных аспектах размеры полипептидов (пептидов, фрагментов) могут находиться в диапазоне приблизительно от 5 остатков до полноразмерного полипептида, например, фермента по изобретению; образцовые размеры составляют приблизительно 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 100, 125, 150, 175, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700 или более остатков, например, смежных остатков образцовой гидролазы по изобретению. Пептиды по изобретению можно использовать, например, в качестве зондов для мечения, антигенов, толерогенов, мотивов, активных центров гидролаз.
Полипептиды по изобретению также включают антитела, способные связываться с гидролазой по изобретению.
Полипептиды по изобретению также включают аминокислотные последовательности, которые «по существу идентичны» последовательностям по изобретению, включая последовательности, которые отличаются от эталонной последовательности одной или несколькими консервативными или неконсервативными аминокислотными заменами, делециями или вставками, конкретно когда такая замена происходит в сайте, который не является активным центром молекулы, и при условии, что полипептид по существу сохраняет свои функциональные свойства. Консервативная аминокислотная замена, например, заменяет одну аминокислота на другую аминокислоту того же класса (например, замена гидрофобной аминокислоты, такой как изолейцин, валин, лейцин или метионин, на другую, или замена одной полярной аминокислоты на другую, такая как замена аргинина на лизин, глутаминовой кислоты на аспарагиновую кислоту или глутамина на аспарагин). Одну или несколько аминокислот можно удалить, например, из гидролазы, что ведет к модификации структуры полипептида, без изменения его биологической активности в значительной степени. Например, можно удалить амино- или C-концевые аминокислоты, которые не необходимы для гидролазной активности.
«Аминокислоту» или «аминокислотную последовательность» может содержать олигопептид, пептид, полипептид или последовательность белка, или фрагмент, часть или субъединица любой из перечисленных и встречающиеся в природе или синтетических молекул.
Термины «полипептид» и «белок» могут содержать аминокислоты, соединенные друг с другом посредством пептидных связей или модифицированных пептидных связей, т.е., пептидные изостеры, и могут содержать модифицированные аминокислоты, отличные от 20 аминокислот, кодируемых геном. Термин «полипептид» также содержит пептиды и полипептидные фрагменты, мотивы и т.п. Термин также содержит гликозилированные полипептиды. Пептиды и полипептиды по изобретению также включают все «миметические» и «пептидомиметические» формы, которые более подробно описаны ниже.
Полипептиды по изобретению включают гидролазы в активной или неактивной форме. Например, полипептиды по изобретению включают пропротеины перед «созреванием» или процессингом препро-последовательностей, например, с помощью фермента, осуществляющего процессинг пропротеина, такого как пропротеин конвертаза, для создания «активного» зрелого белка. Полипептиды по изобретению включают гидролазы, неактивные по другим причинам, например, перед «активацией» посредством события посттрансляционного процессинга, например, под действием эндо- или экзопептидазы или протеиназы, фосфорилирования, амидирования, гликозилирования или сульфатирования, димеризации и т.п. Способы идентификации «препро» последовательностей доменов и сигнальных последовательностей хорошо известны в данной области, см., например, Van de Ven (1993) Crit. Rev. Oncog. 4(2):115-136. Например, для идентификации препро-последовательности, белок выделяют из внеклеточного пространства и определяют N-концевую последовательность белка и сравнивают с формой до процессинга.
Полипептиды по изобретению включают все активные формы, включая активные подпоследовательности, например, каталитические домены или активные центры фермента по изобретению. В определенных вариантах осуществления в настоящем документе предоставлены каталитические домены или активные центры, как указано ниже. В других вариантах осуществления в настоящем документе предоставлены пептиды или полипептиды, содержащие или состоящие из домена активного центра, который предсказывают с помощью базы данных, такой как Pfam (которая представляет собой большую коллекцию множественных выравниваний последовательностей и скрытых марковских моделей, которая покрывает многие обычные семейства белков, The Pfam protein families data base, A. Bateman, E. Birney, L. Cerruti, R. Durbin, L. Etwiller, S.R. Eddy, S. Griffiths-Jones, K.L. Howe, M. Marshall, and E.L.L. Sonnhammer, Nucleic Acids Research, 30(l):276-280, 2002), или эквивалента.
В определенных вариантах осуществления в настоящем документе предоставлены полипептиды, содержащие или не содержащие сигнальную последовательность и/или препро-последовательность. В одном из вариантов осуществления в настоящем документе предоставлены полипептиды, содержащие гетерологичные сигнальные последовательности и/или препро-последовательности. Препро-последовательность (включает последовательность по изобретению, используемую в качестве гетерологичного препро-домена) можно расположить на N- или C-конце белка. В другом варианте осуществления в настоящем документе предоставлены выделенные, синтетические или рекомбинантные сигнальные последовательности, препро-последовательности и каталитические домены (например, «активные центры»), содержащие или состоящие из последовательностей по изобретению. Сигнальная последовательность, препро-домены и/или каталитический домен по изобретению могут являться частью слитого белка, например, в качестве гетерологичного домена в химерном белке. В определенных вариантах осуществления в настоящем документе предоставлены нуклеиновые кислоты, кодирующие эти каталитические домены (КД), препро-домены и сигнальные последовательности (СП, например, пептид, содержащий последовательность, содержащую/состояющую из N-концевых остатков полипептида по изобретению). В определенных вариантах осуществления в настоящем документе предоставлены сигнальные последовательности, содержащие пептид, содержащий/состоящий из последовательности, как указано в остатках с 1 до 12, с 1 до 13, с 1 до 14, с 1 до 15, с 1 до 16, с 1 до 17, с 1 до 18, с 1 до 19, с 1 до 20, с 1 до 21, с 1 до 22, с 1 до 23, с 1 до 24, с 1 до 25, с 1 до 26, с 1 до 27, с 1 до 28, с 1 до 28, с 1 до 30, с 1 до 31, с 1 до 32, с 1 до 33, с 1 до 34, с 1 до 35, с 1 до 36, с 1 до 37, с 1 до 38, с 1 до 39, с 1 до 40, с 1 до 41, с 1 до 42, с 1 до 43, с 1 до 44, с 1 до 45, с 1 до 46, с 1 до 47, с 1 до 48, с 1 до 49 или с 1 до 50 полипептида по изобретению.
Полипептиды и пептиды по изобретению можно выделить из природных источников, синтезировать или создать рекомбинантными способами. Пептиды и белки можно экспрессировать рекомбинантными способами in vitro или in vivo. Пептиды и полипептиды по изобретению можно получить и выделить, используя любой известный в данной области способ. Полипептид и пептиды по изобретению также можно синтезировать, полностью или частично, используя химические способы, которые хорошо известны в данной области. См., например, Caruthers (1980) Nucleic Acids Res. Symp. Ser. 215-223; Horn (1980) Nucleic Acids Res. Symp. Ser. 225-232; Banga, A.K., Therapeutic Peptides and Proteins, Formulation, Processing and Delivery Systems (1995) Technomic Publishing Co., Lancaster, PA. Например, пептидный синтез можно осуществлять, используя различные твердофазные способы (см., например, Roberge (1995) Science 269:202; Merrifield (1997) Methods Enzymol. 289:3-13), а автоматический синтез можно осуществлять, например, с использованием ABI431A Peptide Synthesizer (Perkin Elmer) в соответствии с инструкциями, предоставленными производителем.
Пептиды и полипептиды по изобретению также можно гликозилировать. Гликозилирование можно добавлять после трансляции или химически или посредством клеточных механизмов биосинтеза, где последние включают использование известных мотивов гликозилирования, которые могут присутствовать в последовательности в естественном виде или могут быть присоединены в виде пептида или присоединены в кодирующей последовательности нуклеиновой кислоты. Гликозилирование может представлять собой O-гликозилирование или N-гликозилирование.
«Рекомбинантные» полипептиды или белки относятся к полипептидам или белкам, полученным посредством технологии рекомбинантной ДНК; т.е., полученным из клеток, трансформированных экзогенной ДНК-конструкцией, кодирующей желаемый полипептид или белок. «Синтетические» нуклеиновые кислоты (включая олигонуклеотиды), полипептиды или белки по изобретению включают те, которые получены любым химическим синтезом, например, как описано ниже.
«Фрагменты», как применяют в настоящем документе, представляют собой части встречающихся в природе белков, которые могут существовать по меньшей мере в двух различных конформациях. Фрагменты могут обладать одинаковой или по существу одинаковой аминокислотной последовательностью, совпадающей со встречающимся в природе белком. «Ферментативно активные фрагменты», как применяют в настоящем документе, представляют собой части аминокислотной последовательности (кодирующей белок), которые сохраняют по меньшей мере один вид функциональной активности того белка, к которому они относятся. «По существу одинаковая» обозначает, что аминокислотная последовательность является одинаковой в значительной степени, но не полностью, но сохраняет по меньшей мере один вид функциональной активности последовательности, к которой они относятся. В целом, две аминокислотные последовательности являются «по существу одинаковыми» или «по существу гомологичными», если они идентичны по меньшей мере приблизительно на 85%. Также включены фрагменты, которые обладают трехмерными структурами, отличающимися от встречающегося в природе белка. Примером этого является «проформа» молекулы, например, пропротеин с низкой активностью, которые можно модифицировать расщеплением для получения зрелого фермента, обладающего значительно более высокой активностью.
Пептиды и полипептиды по изобретению, как определено выше, включают все «миметические» и «пептидомиметические» формы. Термины «миметический» и «пептидомиметический» относятся к синтетическому химическому соединению, которое обладает по существу одинаковыми структурными и/или функциональными характеристиками предоставленных в настоящем документе полипептидов. Миметик может или полностью состоять из синтетических, неприродных аналогов аминокислот, или представлять собой химерную молекулу, частично состоящую из природных пептидных аминокислот и частично из неприродных аналогов аминокислот. Миметик также может включать любое количество консервативных замен природных аминокислот, при условии, что такие замены также по существу не изменяют структуру и/или активность миметика. В случае предоставленных в настоящем документе полипептидов, которые представляют собой консервативные варианты, стандартные эксперименты позволят определить, входит ли миметик в объем изобретения, т.е., что его структура и/или функция по существу не изменены. Таким образом, в одном из аспектов композиция миметиков входит в объем изобретения, если она обладает гидролазной активностью.
Композиции полипептидных миметиков по изобретению могут содержать любое сочетание неприродных структурных компонентов. В альтернативном аспекте, композиции миметиков по изобретению, одну или все следующие три структурные группы: a) связующие группы остатков, отличающиеся от соединения природной амидной связью («пептидной связью»); b) неприродные остатки вместо встречающихся в природе аминокислотных остатков; или c) остатки, которые индуцируют вторичную структурную мимикрию, т.е., индуцируют или стабилизируют вторичную структуру, например, конформацию β-поворота, γ-поворота, β-листа, α-спирали и т.п. Например, полипептид по изобретению можно охарактеризовать как миметический, когда все или некоторые его остатки соединены химическими связями, отличающимися от природных пептидных связей. Отдельные остатки пептидомиметика можно соединять пептидными связями, другими химическими связями или соединительными средствами, такими как, например, глутаральдегид, сложные эфиры N-гидроксисукцинимида, бифункциональные малеимиды, N,N'-дициклогексилкарбодиимид (DCC) или N,N'-диизопропилкарбодиимид (DIC). Соединяющие группы, которые могут быть альтернативой обычному соединению амидной связью («пептидной связью»), включают, например, кетометилен (например, -C(=O)-CH2- для -C(=O)-NH-), аминометилен (CH2-NH), этилен, олефин (CH=CH), эфир (CH2-O), тиоэфир (CH2-S), тетразол (CN4-), тиазол, ретроамид, тиоамид или сложный эфир (см., например, Spatola (1983) in Chemistry and Biochemistry of Amino Acids, Peptides and Proteins, том 7, стр. 267-357, «Peptide Backbone Modifications», Marcell Dekker, NY).
Полипептид по изобретению также можно охарактеризовать как миметик за счет содержания всех или нескольких неприродных остатков вместо встречающихся в природе аминокислотных остатков. Неприродные остатки подробно описаны в научной и патентной литературе; ниже описаны некоторые образцовые неприродные композиции, которые можно использовать в качестве миметиков природных аминокислотных остатков, и указания. Миметики ароматических аминокислот можно создать замещением, например, D- или L-нафилаланином; D- или L-фенилглицином; D- или L-2 тиенилаланином; D- или L-1, -2, 3- или 4-пиренилаланином; D- или L-3 тиенилаланином; D- или L-(2-пиридинил)-аланином; D- или L-(3-пиридинил)-аланином; D- или L-(2-пиразинил)-аланином; D- или L-(4-изопропил)-фенилглицином; D-(трифторметил)-фенилглицином; D-(трифторметил)-фенилаланином; D-п-фторфенилаланином; D- или L-п-дифенилфенилаланином; D- или L-п-метоксидифенилфенилаланином; D- или L-2-индол(алкил)аланинами; и, D- или L-алкилаланинами, где алкил может представлять собой замещенный или незамещенный метил, этил, пропил, гексил, бутил, пентил, изопропил, изобутил, втор-бутил, изопентил, или некислые аминокислоты. Ароматические кольца неприродных аминокислот включают, например, ароматические кольца тиазола, тиофенила, пиразола, бензимидазола, нафталина, фурана, пиррола и пиридина.
Миметики кислых аминокислот можно создать замещением, например, некарбоксильными аминокислотами, сохранив отрицательный заряд; (фосфоно)аланином; сульфатированным треонином. Карбоксильные боковые группы (например, аспартил или глутамил) также можно избирательно модифицировать посредством реакции с карбодиимидами (R'-N-C-N-R'), такими как, например, 1-циклогексил-3(2-морфолинил-(4-этил) карбодиимид или 1-этил-3(4-азониа-4,4-диметолпентил)карбодиимид. Аспартил или глутамил также можно превратить в остатки аспарагина и глутамина посредством реакции с ионами аммония. Миметики основных аминокислот можно создать посредством замены, например, (в дополнение к лизину и аргинину) аминокислотами орнитин, цитруллин или (гуанидино)уксусной кислотой или (гуанидино)алкилуксусной кислотой, где алкил определен выше. Производным нитрила (например, содержащим фрагмент CN вместо COOH) можно заменить аспарагин или глутамин. Остатки аспарагина и глутамина можно дезаминировать до соответствующих остатков аспартила или глутамила. Миметики остатка аргинина можно создать посредством реакции аргинила, например, с одним или несколькими стандартными реактивами, включая, например, фенилглиоксаль, 2,3-бутандион, 1,2-циклогександион или нингидрин, альтернативно в щелочных условиях. Миметики остатка тирозина можно создать посредством реакции тирозила, например, с ароматическими соединениями диазония или тетранитрометаном. N-ацетилимидизол и тетранитрометан можно использовать для создания соединений O-ацетилтирозила и 3-нитропроизводных, соответственно. Миметики остатка цистеина можно создать посредством реакции остатков цистеина, например, с α-галоацетатами, такими как 2-хлоруксусная кислота или хлорацетамид и соответствующими аминами; для получения производных карбоксиметила или карбоксиамидометила. Миметики остатков цистеина также можно создать посредством реакции остатков цистеина, например, с бромтрифторацетоном, α-бром-β-(5-имидозоил)пропионовой кислотой; хлорацетилфосфатом, N-алкилмалеимидами, 3-нитро-2-пиридилдисульфидом; метил-2-пиридилдисульфидом; п-хлорртутьбензоатом; 2-хлорртуть-4-нитрофенолом или хлор-7-нитробензо-окса-1,3-диазолом. Миметики лизина можно создать (а также можно изменить N-концевые остатки) посредством реакции лизинила, например, с ангидридом янтарной или другой карбоновой кислоты. Миметики остатка лизина и других остатков, содержащих α-амин, также можно создать посредством реакций с имидоэфирами, такими как метилпиколинимидат, пиридоксальфосфат, пиридоксаль, хлорборогидрид, тринитробензолсульфоновая кислота, O-метилизомочевина, 2,4-пентандион, и посредством реакций с глиоксилатом, катализируемых трансамидазой. Миметики метионина можно создать посредством реакции, например, с метионинсульфоксидом. Миметики пролина включают, например, пипеколиновую кислоту, тиазолидин-карбоновую кислоту, 3- или 4-гидроксипролин, дегидропролин, 3- или 4-метилпролин, или 3,3,-диметилпролин. Миметики остатка гистидина можно создать посредством реакции гистидила, например, с диэтилпрокарбонатом или п-бромфенацилбромидом. Другие миметики включают, например, созданные гидроксилированием пролина и лизина; фосфорилированием гидроксильных групп остатков серина или треонина; метилированием α-аминогрупп лизина, аргинина и гистидина; ацетилированием N-концевого амина; метилированием основной цепи амидных остатков или заменой на N-метиламинокислоты; или амидированием C-концевых карбоксильных групп.
Остаток, например, аминокислота, в полипептиде по изобретению, также можно заменить аминокислотой (или пептидомиметическим остатком) противоположной хиральности. Таким образом, любую аминокислоту, встречающуюся в природе в L-конфигурации (которую также можно обозначать как R или S, в зависимости от структуры химического соединения) можно заменить на аминокислоту того же химического структурного типа или пептидомиметик, но с противоположной хиральностью, обозначаемую как D-аминокислота, которую также можно обозначать как R- или S-форма.
В определенных вариантах осуществления в настоящем документе предоставлены способы модификации предоставленных в настоящем документе полипептидов или посредством естественных процессов, таких как посттрансляционая обработка (например, фосфорилирование, ацилирование и т.п.), или посредством способов химической модификации, и получения модифицированных полипептидов. Модификации можно выполнять в любой части полипептида, включая пептидный остов, боковые цепи аминокислот и N- или C-конец. Следует понимать, что один и тот же тип модификации может быть представлен в одинаковой или различной степени в нескольких сайтах в данном полипептиде. Также данный полипептид может содержать много видов модификаций. Модификации включают ацетилирование, ацилирование, ADP-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение фрагмента гема, ковалентное присоединение нуклеотида или производного нуклеотида, ковалентное присоединение липида или производного липид, ковалентное присоединение фосфатидилинозита, замыкание кольца с образованием поперечных связей, образование дисульфидной связи, деметилирование, образование ковалентных поперечных связей, образование цистеина, образование пироглутамата, формилирование, γ-карбоксилирование, гликозилирование, образование гликозилфосфатидилинозитола, гидроксилирование, иодирование, метилирование, миристилирование, окисление, пэгилирование, протеолитическую обработку, фосфорилирование, пренилирование, рацемизацию, селеноилирование, сульфатирование и опосредованное т-РНК присоединение аминокислоты к белку, такое как аргинилирование. См., например, Creighton, Т.Е., Proteins - Structure and Molecular Properties 2nd Ed., W.H. Freeman and Company, New York (1993); Posttranslational Covalent Modification of Proteins, B.C. Johnson, Ed., Academic Press, New York, стр. 1-12 (1983).
Также можно использовать способы твердофазного химического синтеза пептидов, чтобы синтезировать полипептиды по изобретению или их фрагменты. Такой способ известен в данной области с начала 1960-х (Merrifield, R. В., J. Am. Chem. Soc, 85:2149-2154, 1963) (Также см. Stewart, J. M. and Young, J. D., Solid Phase Peptide Synthesis, 2nd Ed., Pierce Chemical Co., Rockford, 111., стр. 11-12)) и в последнее время используется в коммерчески доступных набора для конструирования и синтеза пептидов в лабораторных условиях (Cambridge Research Biochemicals). Такие коммерчески доступные лабораторные наборы, как правило, используют идеи H. M. Geysen et al, Proc. Natl. Acad. Sci., USA, 81:3998 (1984) и предоставляют возможность синтеза пептидов на кончиках нескольких «стержней» или «шпилек», которые соединены с одной пластиной. Когда используют такую систему, пластину со стержнями или шпильками переворачивают и вводят во вторую пластину с соответствующими лунками или емкостями, которые содержат растворы для присоединения или фиксации подходящей аминокислоты на кончиках шпилек или стержней. Повторяя эту стадию процесса, т.е. переворачивая и вводя кончики стержней и шпилек в подходящие растворы, из аминокислот собирают желаемые пептиды. Кроме того, доступно множество систем FMOC синтеза пептидов. Например, сборку полипептида или фрагмента можно осуществлять на твердофазной основе, используя автоматизированный пептидный синтезатор Applied Biosystems, Inc. Model 431 A™. Такое оборудование предоставляет быстрый доступ к пептидам по изобретению посредством или прямого синтеза или посредством синтеза ряда фрагментов, которые можно соединить, используя другие известные способы.
Ферменты
В определенных вариантах осуществления в настоящем документе предоставлены гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы, например, белки, обладающие по меньшей мере приблизительно 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более или полной (100%) идентичностью последовательностей с образцовым полипептидом по изобретению (например, с SEQ ID № 2, SEQ ID № 4, SEQ ID № 6, SEQ ID № 8, SEQ ID № 10, SEQ ID № 12, SEQ ID № 14, SEQ ID № 16, SEQ ID № 18, или SEQ ID № 20 или SEQ ID № 2, содержащей изменение одной, двух, трех, четырех, пяти, шести, семи, восьми или более (нескольких) или всех аминокислот, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23, или их эквиваленты, антитела, которые связываются с ними, и способы их получения и использования. Полипептиды по изобретению могут обладать любой гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью. В альтернативных аспектах активность фермента по изобретению включает гидролиз или синтез липидов или масел. Гидролазы по изобретению могут модифицировать масла посредством гидролиза, ацидолиза, алкоголиза, алкоголиза, этерификации, трансэтерификации и/или переэтерификации, включая реакции «усиленной миграции».
В альтернативных аспектах гидролазы по изобретению могут иметь модифицированные или новые активности по сравнению с образцовыми гидролазами или активностями, описанными в настоящем документе. В настоящем документе предоставлены гидролазы, содержащие или не содержащие сигнальные последовательности, и сами сигнальные последовательности. В настоящем документе предоставлены иммобилизованные гидролазы, антитела к гидролазам и их фрагменты. В настоящем документе предоставлены белки для ингибирования гидролазной активности, например, антитела, которые связываются с активным центром гидролазы. В настоящем документе предоставлены гомодимеры и гетерокомплексы, например, слитые белки, гетеродимеры и т.д., содержащие гидролазы по изобретению. В настоящем документе предоставлены гидролазы, обладающие активностью в широком диапазоне высоких и низких температур и pH (например, в кислых и щелочных водных средах).
В одном из аспектов одну или несколько гидролаз (например, липаз, сатураз, пальмитаз и/или стеаратаз) по изобретению используют для биокаталитического синтеза структурированных липидов, т.е., липидов, которые содержат определенный набор жирных кислот, распределенных определенным образом на глицериновом остове, включая альтернативы масла какао, полиненасыщенные жирные кислоты (ПНЖК), 1,3-диацилглицериды (ДАГ), 2-моноацилглицериды (МАГ) и триацилглицериды (ТАГ).
В настоящем документе предоставлены способы создания ферментов, обладающих измененным (повышенным или сниженным) Kcat/Km. В одном из аспектов сайт-специфический мутагенез используют для создания дополнительных ферментов гидролаз с альтернативными субстратными специфичностями. Это можно осуществить, например, путем повторной разработки участка связывания субстрата или активного центра фермента. В одном из аспектов гидролазы по изобретению более стабильны при высоких температурах, таких как от 80-85°C до 90-95°C, по сравнению с гидролазами из организмов, обитающих в стандартных или умеренных условиях.
Различные белки по изобретению обладают гидролазной активностью, например, липазной, сатуразной, пальмитазной и/или стеаратазной активностью, при различных условиях. В настоящем документе предоставлены способы получения гидролаз с различной каталитической эффективностью и стабильностью по отношению к температуре, окислителям и значениям pH. Эти способы можно использовать, например, способы сайт-специфического мутагенеза и/или случайного мутагенеза. В одном из аспектов направленную эволюцию можно использовать для получения гидролаз с альтернативными специфичностями и стабильностями.
Белки по изобретению используют в способах, которые позволяют идентифицировать модуляторы гидролаз, например, активаторы или ингибиторы. В кратком изложении, тестовые образцы (например, соединения, такие как члены пептидных или комбинаторных библиотек, бульоны, экстракты и т.п.) добавляют в анализы гидролаз для определения их способность к модуляции, например, ингибированию или активации, расщепления субстрата. Эти ингибиторы можно использовать в производстве и исследованиях для снижения или предотвращения нежелательной изомеризации. Модуляторы, выявленные при использовании способов по изобретению, можно использовать для изменения (например, для сужения или расширения) спектра активности гидролазы.
В одном из аспектов в настоящем документе предоставлены способы выявления гидролаз с использованием нуклеиновых кислот, полипептидов и антител по изобретению. В одном из аспектов проводят скрининг библиотек фага лямбда для выявления гидролаз на основе экспрессии. В настоящем документе предоставлены библиотеки фага лямбда для использования в скрининге, чтобы позволить определить токсичные клоны; усовершенствованный доступ к субстрату; сниженные потребности к конструированию организма-хозяина, избегая любой возможности отклонения в результате исключения массы из библиотеки; и более быстрый рост при низких плотностях клонов.
Скрининг библиотек фага лямбда можно проводить в жидкой фазе или в твердой фазе. В настоящем документе предоставлены способы скрининга в жидкой фазе. Это обеспечивает повышенную гибкость условий анализа; дополнительную субстратную гибкость; повышенную чувствительность для слабых клонов; и простоту автоматизации относительно твердофазного скрининга.
В других вариантах осуществления в настоящем документе предоставлены способы скрининга с использованием белков и нуклеиновых кислот по изобретению, в которых задействована роботизация. Это позволяет проводить многие тысячи биокаталитических реакций и скрининг-анализов за короткий период времени, например, за день, а также гарантирует высокий уровень точности и воспроизводимости (см. обсуждение чипов ниже). В результате приблизительно за неделю можно получить библиотеку производных соединений.
В определенных вариантах осуществления в настоящем документе предоставлены ферменты гидролазы, которые представляют собой не встречающиеся в природе гидролазы, обладающие отличающимися гидролазными активностями, стабильностями, субстратными специфичностями, профилями pH и/или эксплуатационными характеристиками по сравнению с не встречающейся в природе гидролазой. Эти гидролазы содержат аминокислотную последовательность, не найденную в природе. Их можно получить посредством замены нескольких аминокислотных остатков предшественника гидролаз на другие аминокислоты. Предшественник гидролазы может представлять собой встречающуюся в природе гидролазу или рекомбинантную гидролазу. В одном из аспектов варианты гидролаз охватывают замены любых встречающихся в природе L-аминокислот в обозначенных положениях аминокислотных остатков.
Сигнальные последовательности, препро- и каталитические домены гидролаз
В определенных вариантах осуществления в настоящем документе предоставлены сигнальные последовательности (например, сигнальные пептиды (СП)), препро-домены и каталитические домены (КД). СП, препро-домены и/или КД по изобретению могут представлять собой выделенные, синтетические или рекомбинантные пептиды или могут представлять собой часть слитого белка, например, в виде гетерологичного домена в химерном белке. В определенных вариантах осуществления в настоящем документе предоставлены нуклеиновые кислоты, кодирующие эти каталитические домены (КД), препро-домены и сигнальные последовательности (СП, например, пептид, содержащий последовательность, содержащую/состоящую из N-концевых остатков полипептида по изобретению). В определенных вариантах осуществления в настоящем документе предоставлены сигнальные последовательности, содержащие пептид, содержащий/состоящий из последовательности, как указано в остатках с 1 до 12, с 1 до 13, с 1 до 14, с 1 до 15, с 1 до 16, с 1 до 17, с 1 до 18, с 1 до 19, с 1 до 20, с 1 до 21, с 1 до 22, с 1 до 23, с 1 до 24, с 1 до 25, с 1 до 26, с 1 до 27, с 1 до 28, с 1 до 28, с 1 до 30, с 1 до 31, с 1 до 32, с 1 до 33, с 1 до 34, с 1 до 35, с 1 до 36, с 1 до 37, с 1 до 38, с 1 до 39, с 1 до 40, с 1 до 41, с 1 до 42, с 1 до 43, с 1 до 44 (или более длинный пептид) полипептида по изобретению. В одном из вариантов осуществления в настоящем документе предоставлены выделенные, синтетические или рекомбинантные сигнальные последовательности, содержащие/состоящие из сигнальной последовательности по изобретению, полученной из другого фермента по изобретению или другого вида фермента или полипептида.
Сигнальные последовательности (СП), КД и/или препро-последовательности гидролаз по изобретению могут представлять собой выделенные пептиды или последовательности, соединенные с другой гидролазой или полипептидом негидролазы, например, в виде слитого (химерного) белка. В определенных вариантах осуществления в настоящем документе предоставлены полипептиды, содержащие сигнальные последовательности гидролаз по изобретению. В одном из аспектов полипептиды, содержащие сигнальные последовательности СП, КД и/или препро гидролаз по изобретению, содержат последовательности, гетерологичные гидролазам по изобретению (например, слитый белок, содержащий СП, КД и/или препро по изобретению и последовательности из другой гидролазы или белка негидролазы). В настоящем документе предоставлены гидролазы по изобретению с гетерологичными СП, КД и/или препро-последовательностями, например, последовательностями с сигнальной последовательностью дрожжей. Гидролаза по изобретению может содержать гетерологичную СП и/или препро в векторе, например, в векторе серии pPIC (Invitrogen, Carlsbad, CA).
В одном из аспектов СП, КД и/или препро-последовательности по изобретению идентифицируют после идентификации новых полипептидов гидролаз. Пути, посредством которых происходит сортировка и транспортировка белков в их правильное местоположение в клетке часто обозначают как пути направления белков. Одним из наиболее важных элементов во всех этих системах направления является короткая аминокислотная последовательность на N-конце вновь синтезированного полипептида, которую называют сигнальной последовательностью. Эта сигнальная последовательность направляет белок в соответствующее ему местоположение в клетке и удаляется в процессе транспортировки или когда белок достигает своего пункта назначения. Большинство лизосомальных, мембранных или секретируемых белков содержит N-концевую сигнальную последовательность, которая обозначает, что их нужно перенести в просвет эндоплазматического ретикулума. Длина сигнальных последовательностей может меняться от 13 до 45 или более аминокислотных остатков. Различные способы распознавания сигнальных последовательностей известны специалистам в данной области. Например, в одном из аспектов сигнальные пептиды новых гидролаз идентифицируют способом, обозначаемым как SignalP. В SignalP использована комбинированная нейронная сет, которая распознает как сигнальные пептиды, так и их сайты расщепления. (Nielsen, et al., «Identification of prokariotic and eukaryotic signal peptides and prediction of their cleavage sites». Protein Engineering, том 10, № 1, стр. 1-6 (1997).
Следует понимать, что в некоторых аспектах гидролазы по изобретению могут не содержать СП и/или препро-последовательности и/или каталитические домены (КД). В одном из аспектов в настоящем документе предоставлены полипептиды (например, гидролазы), у которых частично или полностью отсутствует СП, КД и/или препро-домен. В другом аспекте в настоящем документе предоставлены нуклеиновые кислоты, кодирующие сигнальную последовательность (СП), КД и/или препро из одной гидролазы, функционально связанные с последовательностью нуклеиновой кислоты другой гидролазы, или необязательно желательными могут быть сигнальная последовательность (СП) и/или препро-домен из белка негидролазы.
В определенных вариантах осуществления в настоящем документе предоставлены выделенные, синтетические или рекомбинантные полипептиды, содержащие сигнальные последовательности (СП), препро-домен и/или каталитические домены (КД) по изобретению и гетерологичные последовательности. Гетерологичные последовательности представляют собой последовательности, которые в природе не связаны (например, с гидролазой) с СП, препро-доменом и/или КД. Последовательность, с которой СП, препро-домен и/или КД в природе не связаны, может быть расположена на N-конце, C-конце СП, препро-домена и/или КД и/или на обоих концах СП и/или КД. В определенных вариантах осуществления в настоящем документе предоставлены выделенные, синтетические или рекомбинантные полипептиды, содержащие (или состоящие из) полипептид, содержащий сигнальную последовательность (СП), препро-домен и/или каталитический домен (КД) по изобретению, с оговоркой, что они не связаны с какой-либо последовательностью, с которой они связаны в природе (например, с последовательностью гидролазы). В настоящем документе предоставлены выделенные или рекомбинантные нуклеиновые кислоты, кодирующие эти полипептиды. Таким образом, в одном из аспектов выделенная, синтетическая или рекомбинантная нуклеиновая кислота по изобретению содержит кодирующую последовательность для сигнальной последовательности (СП), препро-домена и/или каталитического домена (КД) по изобретению и гетерологичной последовательности (т.е., последовательности, которая в природе не связана с сигнальной последовательностью (СП), препро-доменом и/или каталитическим доменом (КД) по изобретению). Гетерологичная последовательность может находиться на 3'-конце, 5'-конце и/или на обоих концах СП, препро-домена и/или КД кодирующей последовательности.
В определенных вариантах осуществления в настоящем документе предоставлены слитые N-концом или C-концом подпоследовательности ферментов по изобретению (например, сигнальные последовательности, препро-последовательности), с другими полипептидами, активными белками или фрагментами белков. Фермент по изобретению (например, гидролазу, например, липазу, сатуразу, пальмитазу и/или стеаратазу) также можно получить посредством экспрессии фермента в виде неактивного слитого белка, который впоследствии активируют посредством акта протеолитического расщепления (с использованием эндогенной или экзогенной протеазной активности, например, трипсина), который ведет к отделению слитого белка-партнера и зрелого фермента, например, гидролазы по изобретению. В одном из аспектов слитый белок по изобретению экспрессируют из гибридной нуклеотидной конструкции, которая кодирует одну открытую рамку считывания, содержащую следующие элементы: нуклеотидная последовательность для слитого белка, последовательность линкера (определяется как нуклеотидная последовательность, которая кодирует гибкую аминокислотную последовательность, которая соединяет два менее гибких домена белка), сайт узнавания для расщепления протеазой и последовательность зрелого фермента (например, любого фермента по изобретению, например, гидролазы). В альтернативных аспектах слитый белок может содержать последовательность пектатлиазы, последовательность ксиланазы, последовательность фосфатидной кислой фосфатазы или другую последовательность, например, последовательность, которая, как было показано ранее, допускает сверхэкспрессию в системе-хозяине, предоставляющей интерес. Можно использовать любую систему-хозяина (см. обсуждение выше), например, E. coli или Pichia pastoris. Расположение нуклеотидных последовательностей в химерной нуклеотидной конструкции можно определить на основе уровней экспрессии белка, достигаемых при использовании каждой слитой конструкции. В одном из аспектов в направлении от 5'-конца нуклеотидной конструкции к 3'-концу конструкции нуклеотидные последовательности располагают в следующем порядке: сигнальная последовательность/слитый белок/последовательность линкера/сайт узнавания для расщепления протеазой/зрелый фермент (например, любой фермент по изобретению, например, гидролаза) или сигнальная последовательность/пропоследовательность/зрелый фермент/последовательность линкера/слитый белок. Экспрессия фермента (например, любого фермента по изобретению, например, гидролазы) в виде неактивного слитого белка может в целом усовершенствовать экспрессию последовательности фермента, может снизить любую возможную токсичность, связанную со сверхсинтезом активного фермента и/или может увеличить срок хранения фермента перед использованием, поскольку фермент неактивен до тех пор, пока слитый белок, например, пектатлиазу не отделят от фермента, например, гидролазы по изобретению.
В одном из вариантов осуществления в настоящем документе предоставлены конкретные составы для активации гидролазы по изобретению, экспрессированной в виде слитого белка. В одном из аспектов активацию активности гидролазы, исходно экспрессированной в виде неактивного слитого белка, выполняют с использованием протеолитической активности или потенциально протеолитической активности в сочетании с N-концевой или C-концевой пептидазой (пептидаза может представлять собой фермент по изобретению или другой фермент). Это событие активации можно выполнить различными путями и в различные моменты в процессе производства/хранения перед применением в рафинировании масла. Образцовые процессы по изобретению включают: расщепление посредством эндогенной активности, проявляемой производящим организмом-хозяином при секреции слитой конструкции в среды для брожения; расщепление посредством активности эндогенной протеазы, которая активируется или приходит в контакт с внутриклеточно экспрессированной слитой конструкцией при разрушении клеток-хозяев; пропускание неочищенной или очищенной слитой конструкции через колонку с активностью иммобилизованной протеазы для выполнения расщепления и активации фермента (например, гидролазы по изобретению, например, липазы, сатуразы, пальмитазы и/или стеаратазы) перед введением фермента в состав; обработка неочищенной или очищенной слитой конструкции с растворимым источником протеолитической активности; активация гидролазы (например, гидролазы по изобретению) при переработке нефти с использованием растворимого или нерастворимого источника протеолитической активности непосредственно перед использованием в процессе; и/или активация активности гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы по изобретению) посредством непрерывной циркуляции состава слитой конструкции через колонку с активностью иммобилизованной протеазы при сниженной температуре (например, при любой температуре между приблизительно 4°C и 20°C). Это событие активации можно выполнить перед доставкой в место использования или оно может происходить на месте при переработке нефти.
Гликозилирование
Пептиды и полипептиды по изобретению (например, гидролазы, антитела) также могут быть гликозилированы, например, в одном из аспектов могут содержать по меньшей мере один сайт гликозилирования, например, N-гликозилирования или O-гликозилирования. В одном из аспектов полипептид может быть гликозилирован после экспрессии в P. pastoris или S. pombe. Гликозилирование можно присоединять после трансляции или химически или посредством клеточных биосинтетических механизмов, где последние включают использование известных мотивов гликозилирования, которые могут быть нативными для последовательности или могут быть присоединены в виде пептида или добавлены в кодирующую последовательность нуклеиновой кислоты.
Гибридные гидролазы и библиотеки пептидов
В определенных вариантах осуществления в настоящем документе предоставлены гибридные гидролазы (например, синтетические белки) и слитые белки, включая библиотеки пептидов, содержащие последовательности по изобретению. Библиотеки пептидов по изобретению можно использовать для выделения пептидных модуляторов (например, активаторов или ингибиторов) мишеней. Библиотеки пептидов по изобретению можно использовать для идентификации правильных партнеров связывания для мишеней, таких как лиганды, например, цитокины, гормоны и т.п.
В одном из аспектов слитые белки по изобретению (например, пептидный фрагмент) конформационно стабильны (по отношению к линейным пептидам) для обеспечения более высокой аффинности связывания для мишеней. В другом аспекте в настоящем документе предоставлены слитые белки из гидролаз по изобретению и других пептидов, включая известные и случайные пептиды. Их можно слить таким образом, чтобы не нарушать структуру фермента или антитела (например, гидролазы) в значительной степени и метаболически или структурно конформационно стабилизировать пептид. Это позволяет создать библиотеку пептидов, в которой легко проводить мониторинг как их присутствия в клетках, так и их количества.
Варианты аминокислотных последовательностей по изобретению могут отличаться предварительно определенным характером изменения, признаком, который отделяет встречающуюся в природе форму, например, аллельную или межвидовую изменчивость последовательности гидролазы. В одном из аспектов варианты по изобретению проявляют качественно одинаковую биологическую активностью вместе со встречающимся в природе аналогом. Альтернативно можно отбирать варианты, которые обладают модифицированными характеристиками. В одном из аспектов, несмотря на то, что сайт или участок для введения изменения аминокислотной последовательности определен предварительно, мутацию per se не нужно определять предварительно. Например, для того, чтобы оптимизировать характеристики мутации в данном сайте, можно проводить случайный мутагенез в целевом кодоне или участке и скрининг экспрессированных вариантов гидролаз на предмет оптимального сочетания желаемой активности. Как обсуждается в настоящем документе, хорошо известны способы создания мутаций с заменами в предварительно определенных сайтах в ДНК, обладающей известной последовательностью, например, мутагенез праймером M13 и ПЦР мутагенез. Скрининг мутантов можно осуществлять, используя анализ протеолитической активности. В альтернативных аспектах замены аминокислот могут представлять собой отдельные остатки; вставки могут иметь длину порядка приблизительно от 1 до 20 аминокислот, хотя можно выполнять значительно более длинные вставки. Делеции могут находиться в диапазоне приблизительно от 1 приблизительно до 20, 30, 40, 50, 60, 70 остатков или более. Для получения конечного производного с оптимальными свойствами можно использовать замены, делеции, вставки или любое их сочетание. Как правило, эти изменения выполняют на нескольких аминокислотах, чтобы свести изменения молекулы к минимуму. Однако в некоторых случаях допустимы более значительные изменения.
В определенных вариантах осуществления в настоящем документе предоставлены гидролазы, где структура полипептидного остова, вторичная или третичная структура, например, структура α-спирали или β-листа модифицирована. В одном из аспектов модифицирован заряд или гидрофобность. В одном из аспектов модифицирован объем боковой цепи. Значительные изменения функции или иммунологической идентичности осуществляют посредством выбора замен, которые являются менее консервативными. Например, можно выполнять замены, которые в более значительной степени оказывают влияние на: структуру полипептидного остова в области изменения, например, на структуру α-спирали или β-листа; заряд или гидрофобный участок молекулы, который может находиться в активном центре; или на боковую цепь. В других вариантах осуществления в настоящем документе предоставлены белки, содержащие замены последовательности по изобретению, например, где (a) гидрофильные остатки, например, серил или треонил, заменяют на (или с использованием) гидрофобный остаток, например, лейцил, изолейцил, фенилаланил, валил или аланил; (b) цистеин или пролин заменяют на (или с использованием) любой другой остаток; (c) остаток, содержащий боковую цепь с положительным зарядом, например, лизил, аргинил или гистидил, заменяют на (или с использованием) остаток с отрицательным зарядом, например, глутамил или аспартил; или (d) остаток, обладающий объемной боковой цепью, например, фенилаланин, заменяют на (или с использованием) остаток, который не имеет боковой цепи, например, на глицин. Варианты могут проявлять качественно одинаковую биологическую активность (т.е. гидролазную активность), хотя по мере необходимости можно отбирать варианты для модификации характеристик гидролаз.
В одном из аспектов гидролазы по изобретению содержат эпитопы или метки для очистки, сигнальные последовательности или другие слитые последовательности и т.д. В одном из аспектов гидролазы по изобретению можно слить со случайным пептидом для создания слитого полипептида. Под «слитым» или «функционально связанным» в настоящем документе понимают то, что случайный пептид и гидролазу соединяют вместе таким образом, чтобы минимизировать нарушение стабильности структуры гидролазы, например, чтобы сохранить гидролазную активность. Слитый полипептид (или слитый полинуклеотид, кодирующий слитый полипептид) также может содержать дополнительные компоненты, включая несколько пептидов в нескольких петлях.
В одном из аспектов рандомизируют пептиды (например, подпоследовательности гидролаз) и кодирующие их нуклеиновые кислоты, или рандомизируют полностью или оказывают влияние на их рандомизацию, например, на частоту нуклеотидов/остатков в целом или в определенном положении. «Рандомизированный» обозначает, что каждая нуклеиновая кислота и пептид состоят по существу из случайных нуклеотидов и аминокислот, соответственно. В одном из аспектов нуклеиновые кислоты, дающие начало пептидам, можно синтезировать химическим путем и, таким образом, можно встроить любой нуклеотид в любое положение. Таким образом, когда для создания пептидов экспрессируют нуклеиновые кислоты, в любое положение можно встроить любой аминокислотный остаток. Можно разработать процесс синтеза для создания рандомизированных нуклеиновых кислот, чтобы предоставить возможность создания всех или почти всех возможных сочетаний по всей длине нуклеиновой кислоты, таким образом, создавая библиотеку рандомизированных нуклеиновых кислот. Библиотека может предоставить популяцию рандомизированных продуктов экспрессии с достаточным структурным разнообразием, чтобы затронуть вероятностно достаточный диапазон клеточных ответов, чтобы предоставить одну или несколько клеток, проявляющих желаемый ответ. В настоящем документе предоставлены библиотеки взаимодействия, достаточно большие для того, чтобы по меньшей мере один из их членов имел структуру, которая придает ему аффинность к определенной молекуле, белку или другому фактору.
Способы скрининга и устройства оперативного мониторинга
При осуществлении способов по изобретению можно использовать различные устройства и способы в сочетании с полипептидами и нуклеиновыми кислотами по изобретению, например, для скрининга гидролазной активности полипептидов, для скрининга соединений в качестве потенциальных активаторов или ингибиторов гидролазной активности (например, для потенциального скрининга лекарственных средств), антител, которые связываются с полипептидом по изобретению, нуклеиновых кислот, которые гибридизуются с нуклеиновой кислотой по изобретению, для скрининга клеток, экспрессирующих полипептид по изобретению, и т.п. См., например, патент США № 6337187.
Капиллярные блоки
Капиллярные блоки, такие как GIGAMATRIX™, Diversa Corporation, San Diego, CA, можно использовать в способах по изобретению. Нуклеиновые кислоты или полипептиды по изобретению можно иммобилизовать или нанести на чип, включая капиллярные блоки. Чипы можно использовать для скрининга или мониторинга способности библиотек композиций (например, низкомолекулярных соединений, антител, нуклеиновых кислот и т.д.) связываться с нуклеиновой кислотой или полипептидом по изобретению или модулировать их активность. Капиллярные блоки предоставляют другую систему для удержания и скрининга образцов. Например, устройство для скрининга образцов может включать несколько капилляров, сформированных в виде блока капилляров, расположенных рядом, где каждый капилляр содержит по меньшей мере одну стенку, определяющую просвет для удерживания образца. Дополнительно устройство может включать промежуточный материал, расположенный между соседними капиллярами в блоке, и один или несколько эталонных указателей, сформированных в промежуточном материале. Капилляр для скрининга образца, где капилляр приспособлен для крепления в блоке капилляров, может содержать первую стенку, определяющую просвет для удержания образца, и вторую стенку, созданную из фильтрующего материала, для фильтрации энергии возбуждения, которую подают в просвет для возбуждения образца.
Полипептид или нуклеиновая кислота, например, лиганд или субстрат, можно ввести в первый компонент по меньшей мере в часть капилляра капиллярного блока. Каждый капилляр капиллярного блока может содержать по меньшей мере одну стенку, определяющую просвет для удержания первого компонента. Позади первого компонента в капилляр можно ввести пузырек воздуха. В капилляр можно ввести второй компонент, где второй компонент отделяют от первого компонента посредством пузырька воздуха. В капилляр капиллярного блока можно ввести образец, представляющий интерес, в виде первой жидкости, меченой поддающейся обнаружению частицей, где каждый капилляр капиллярного блока содержит по меньшей мере одну стенку, определяющую просвет для удержания первой жидкости и поддающейся обнаружению частицы, и где по меньшей мере одну стенку покрывают связующим веществом для связывания поддающейся обнаружению частицы по меньшей мере с одной стенкой. Способ может дополнительно включать удаление первой жидкости из капиллярной трубки, где связанную поддающуюся обнаружению частицу удерживают внутри капилляра, и введение второй жидкости в капиллярную трубку.
Капиллярный блок может содержать несколько отдельных капилляров, содержащих по меньшей мере одну внешнюю стенку, определяющую просвет. Наружная стенка капилляра может представлять собой одну или несколько стенок, соединенных вместе. Подобным образом, стенка может определять просвет цилиндрической, прямоугольной, гексагональной или любой другой геометрической формы при условии, что стенки формируют просвет для удержания жидкости или образца. Капилляры капиллярного блока можно расположить вместе в непосредственной близости для создания плоской структуры. Капилляры можно соединить вместе, сплавив их (например, когда капилляры выполнены из стекла), склеив, связав или прижав бок о бок. Капиллярный блок можно создать из любого числа отдельных капилляров, например, в диапазоне от 100 до 4000000 капилляров. Капиллярный блок может формировать микротитровальный планшет, содержащий приблизительно 100000 или более отдельных капилляров, соединенных вместе.
Чипы или «биочипы»
Нуклеиновые кислоты или полипептиды по изобретению можно иммобилизовать или нанести на чип. Чипы можно использовать для скрининга или мониторинга библиотек композиций (например, низкомолекулярных соединений, антител, нуклеиновых кислот и т.д.) по их способности связываться или модулировать активность нуклеиновой кислоты или полипептида по изобретению. Например, в одном из аспектов по изобретению параметром мониторинга является экспрессия транскрипта гена гидролазы. Один или несколько или все транскрипты клетки можно измерить посредством гибридизации образца, содержащего транскрипты клетки или нуклеиновые кислоты, представляющие или комплементарные транскриптам клетки, посредством гибридизации с нуклеиновыми кислотами, иммобилизованными на чипе или «биочипе». Используя «набор» нуклеиновых кислот на микрочипе, можно одновременно определить количество некоторых или всех транскриптов клетки. Альтернативно чипы, содержащие геномную нуклеиновую кислоту, также можно использовать для определения генотипа вновь сконструированного штамма, полученного способами по изобретению. Полипептидные чипы также можно использовать для одновременного количественного определения нескольких белков. Настоящее изобретение можно осуществлять на практике, используя любой известных «чип», также обозначаемый как «микрочип» или «чип с нуклеиновыми кислотами» или «полипептидный чип» или «чип с антителами» или «биочип» или их вариант. В целом чипы представляют собой несколько «пятен» или «элементов мишени», каждый элемент мишени содержит определенное количество одной или нескольких биологических молекул, например, олигонуклеотидов, иммобилизованных на определенной области поверхности основы для специфического связывания с молекулами образца, например, с мРНК-транскриптами.
«Чипы» или «микрочипы» или «биочипы» по изобретению могут содержать несколько элементов мишени, каждый элемент мишени содержит определенное количество одного или нескольких полипептидов (включая антитела) или нуклеиновых кислот, иммобилизованных на определенной области поверхности основы.
В одном из аспектов гидролазы используют в виде иммобилизованных форм. Можно использовать любой способ иммобилизации, например, иммобилизацию на инертной основе, такой как диэтиламиноэтилцеллюлоза, пористое стекло, хитин или клетки. Клетки, которые экспрессируют гидролазы по изобретению, можно иммобилизовать посредством образования поперечных связей с поверхностью основы, например, используя глутаральдегид.
При осуществлении способов по изобретению полностью или частично можно включить любой известный чип и/или способ получения и использования чипов или их варианты, как описано, например, в патентах США №№ 6277628; 6277489; 6261776; 6258606; 6054270; 6048695; 6045996; 6022963; 6013440; 5965452; 5959098; 5856174; 5830645; 5770456; 5632957; 5556752; 5143854; 5807522; 5800992; 5744305; 5700637; 5556752; 5434049; также см., например, WO 99/51773; WO 99/09217; WO 97/46313; WO 96/17958; также см., например, Johnston (1998) Curr. Biol. 8:R171-R174; Schummer (1997) Biotechniques 23:1087-1092; Kern (1997) Biotechniques 23:120-124; Solinas-Toldo (1997) Genes, Chromosomes & Cancer 20:399-407; Bowtell (1999) Nature Genetics Supp. 21:25-32. Также см. Опубликованные патентные заявки США №№ 20010018642; 20010019827; 20010016322; 20010014449; 20010014448; 20010012537; 20010008765.
Антитела и способы скрининга на основе антител
В определенных вариантах осуществления в настоящем документе предоставлены выделенные, синтетические или рекомбинантные антитела которые специфически связываются с гидролазой по изобретению. Эти антитела можно использовать для выделения, идентификации или количественного определения гидролазы по изобретению или родственных полипептидов. Эти антитела можно использовать для выделения других предоставленных в настоящем документе полипептидов или других родственных гидролаз.
«Антитела» по изобретению могут содержать пептид(ы) или полипептид(ы), полученные из, смоделированные после или по существу кодируемые геном иммуноглобулина или генами иммуноглобулинов, или их фрагменты, способные специфически связывать антиген или эпитоп, см., например, Fundamental Immunology, Third Edition, W.E. Paul, ed., Raven Press, N.Y. (1993); Wilson (1994) J. Immunol. Methods 175:267-273; Yarmush (1992) J. Biochem. Biophys. Methods 25:85-97. Термин «антитело» включает антигенсвязывающие части, т.е. «антигенсвязывающие участки», (например, фрагменты, подпоследовательности, участки, определяющие комплементарность (CDR)), которые сохраняют способность связываться с антигеном, включая (i) Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab')2 фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, соединенные дисульфидным мостиком в шарнирной области; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела, (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH; и (vi) выделенный участок, определяющий комплементарность (CDR). Одноцепочечные антитела также включены по ссылке в термин «антитело». В настоящем документе предоставлены антитела, включая антигенсвязывающие участки и одноцепочечные антитела, которые специфически связываются с гидролазой по изобретению. При осуществлении способов по изобретению также можно использовать полипептиды, обладающие гидролазной активностью.
Антитела можно использовать в иммунопреципитации, окрашивании, иммуноаффинных колонках и т.п. При желании, последовательности нуклеиновой кислоты, кодирующие конкретные антигены, можно создать посредством иммунизации с последующим выделением полипептида или нуклеиновой кислоты, амплификации или клонирования и иммобилизации полипептида на чипе по изобретению. Альтернативно способы по изобретению можно использовать для модификации структуры антитела, полученного посредством клетки, подлежащей модификации, например, аффинность антитела можно повысить или понизить. Кроме того, способность создавать или модифицировать антитела может представлять собой фенотип, сконструированный в клетке посредством способов по изобретению.
Способы иммунизации, получения и выделения антител (поликлональных и моноклональных) известны специалистам в данной области и описаны в научной и патентной литературе, см., например, Coligan, CURRENT PROTOCOLS IN IMMUNOLOGY, Wiley/Greene, NY (1991); Stites (eds.) BASIC AND CLINICAL IMMUNOLOGY (7th ed.) Lange Medical Publications, Los Altos, С A («Stites»); Goding, MONOCLONAL ANTIBODIES: PRINCIPLES AND PRACTICE (2d ed.) Academic Press, New York, NY (1986); Kohler (1975) Nature 256:495; Harlow (1988) ANTIBODIES, A LABORATORY MANUAL, Cold Spring Harbor Publications, New York. Антитела также можно создавать in vitro, например, используя библиотеки фагового дисплея, экспрессирующие сайт связывания рекомбинантного антитела, в дополнение к традиционным способам с использованием животных in vivo. См., например, Hoogenboom (1997) Trends Biotechnol. 15:62-70; Katz (1997) Annu. Rev. Biophys. Biomol. Struct. 26:27-45.
Полипептиды или пептиды можно использовать для создания антитела, которое специфически связывается с предоставленными в настоящем документе полипептидами. Полученные антитела можно использовать в процедурах иммуноаффинной хроматографии для выделения или очистки полипептида или для определения присутствия полипептида в биологическом образце. В таких процедурах препарат белка, такой как экстракт или биологический образец, приводят в контакт с антителом, способным специфически связываться с одним из предоставленных в настоящем документе полипептидов.
В иммуноаффинных процедурах антитело прикрепляют к твердой основе, такой как бусы или другая матрица для колонки. Препарат белка приводят в контакт с антителом в условиях, в которых антитело специфически связывается с одним из предоставленных в настоящем документе полипептидов. После отмывания неспецифически связанных белков элюируют специфически связанные полипептиды.
Способность белков в биологическом образце связываться с антителом можно определить, используя любые из множества процедур, известных специалистам в данной области. Например, связывание можно определить посредством мечения антитела поддающейся обнаружению меткой, такой как флуоресцентное средство, ферментативная метка или радиоизотоп. Альтернативно связывания антитела с образцом можно определить, используя вторичное антитело, содержащее на себе такую поддающуюся обнаружению метку. Конкретные анализы включают анализ ELISA, сэндвич-анализ, радиоиммунологический анализ и Вестерн-блоттинг.
Поликлональные антитела к предоставленным в настоящем документе полипептидам можно получить посредством прямой инъекции полипептидов животному или посредством введения полипептидов животному, не относящемуся к человеку. Затем полученное таким образом антитело будет связывать сам полипептид. Таким образом, для создания антитела, которое может связываться с целым нативным полипептидом, можно использовать даже последовательность, кодирующую только фрагмент полипептида. Затем такие антитела можно использовать для выделения полипептида из клеток, экспрессирующих этот полипептид.
Для получения моноклональных антител можно использовать любой способ, который предоставляет антитела, полученные посредством непрерывных культур линий клеток. Примеры включают гибридомный способ, триомный способ, способ с B-клеточной гибридомой человека и способ с EBV-гибридомой (см., например, Cole (1985) in Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., стр. 77-96).
Описанные способы получения одноцепочечных антител (см., например, патент США № 4946778) можно приспособить для получения одноцепочечных антител к предоставленным в настоящем документе полипептидам. Альтернативно трансгенных мышей можно использовать для экспрессии гуманизированных антител к этим полипептидам или их фрагментам.
Созданные антитела к предоставленным в настоящем документе полипептидам (включая антиидиотипические антитела) можно использовать в скрининге схожих полипептидов из других организмов и образцов. В таких способах полипептиды из организма приводят в контакт с антителом и определяют те полипептиды, которые специфически связались с антителом. Любые описанные выше процедуры можно использовать для определения связывания с антителом.
Иммобилизованные гидролазы
В одном из аспектов гидролазы по изобретению, например, липазы, сатуразы, пальмитазы и/или стеаратазы, используют в виде иммобилизованных форм, например, для обработки липидов, в структурированном синтезе липидов, для расщепления белков и т.п. Иммобилизованные липазы по изобретению можно использовать, например, для гидролиза триацилглицеридов, диацилглицеридов или сложных эфиров или для этерификации или трансэтерификации жирных кислот, диацилглицеридов или триацилглицеридов, или в переэтерификации жиров. В одном из аспектов липаза обладает специфичностью для этерификации жирных кислот со спиртом, 1,3-специфичностью или специфичностью для гидролиза неполных глицеридов, сложных эфиров или триацилглицеридов. Иммобилизованные липазы по изобретению можно использовать в уплотненном слое для непрерывной трансэтерификации жиров в отсутствие растворителя. См., например, патент США № 4818695; 5569594.
Можно использовать любой способ иммобилизации или форму основы, например, чипы, бусины, основы капилляров и т.п., как описано выше. В одном из аспектов можно выполнять иммобилизацию гидролазы на инертной основе, такой как диэтиламиноэтилцеллюлоза, пористое стекло, хитин или клетки. Клетки, которые экспрессируют гидролазы по изобретению, можно иммобилизовать посредством образования поперечных связей с поверхностью основы, например, с использованием глутаральдегида. Можно получить иммобилизованные гидролазы по изобретению, содержащие гидролазу, связанную с сухой, пористой гидрофобной основе в форме частиц, с поверхностно-активным веществом, таким как сложный полиоксиэтиленсорбитановый эфир жирной кислоты или полиглицериновый эфир жирной кислоты. Основа может представлять собой алифатический олефиновый полимер, такой как полиэтилен или полипропилен, гомо- или сополимер стирола или их смесь или предварительно обработанную неорганическую основу. Эти основы можно выбрать из алифатических олефиновых полимеров, окисленные полимеры, смеси этих полимеров или предварительно обработанные неорганические основы для того, чтобы придать эти основам гидрофобные свойства. Эта предварительная обработка может включать силанирование органическим соединением кремния. Неорганическое вещество может представлять собой диоксид кремния, оксид алюминия, стекло или керамику. Основы можно выполнить из полистирола, сополимеров стирола, полиэтилена, полипропилена или из сополимеров, полученных из (мет)акрилатов. См., например, патент США № 5773266.
Ферменты гидролазы, их фрагменты и нуклеиновые кислоты, которые кодируют ферменты и фрагменты, можно прикрепить к твердой основе. Зачастую это повышает экономичность и эффективность использования гидролаз в промышленных процессах. Например, смесь или коктейль ферментов гидролаз (или их активных фрагментов), которые используют в конкретной химической реакции, можно прикрепить к твердой основе и окунать в производственную емкость. Может происходить ферментативная реакция. Затем для повторного использования твердую основу можно вынуть из емкости вместе с прикрепленными к ней ферментами. В одном из вариантов осуществления по изобретению, выделенную нуклеиновую кислоту по изобретению, прикрепляют к твердой основе. В другом варианте осуществления по изобретению твердую основу выбирают из группы, включающей гель, смолу, полимер, керамику, стекло, микроэлектрод и любое их сочетание.
Например, твердые основы, предоставленные в настоящем документе, включают гели. Некоторые примеры гелей включают Сефарозу™ (GE Healthcare, Piscataway, NJ), желатин, глутаральдегид, обработанный хитозаном глутаральдегид, альбумин-глутаральдегид, хитозан-ксантан, гель TOYOPEARL (полимерный гель), альгинат, альгинат-полилизин, каррагенан, агарозу, глиоксил агарозу, магнитную агарозу, декстран-агарозу, гидрогель из поли(карбамоилсульфоната), БСА-ПЭГ гидрогель, фосфорилированный поливиниловый спирт (ПВС), моноаминоэтил-N-аминоэтил (MANA), амино или любое их сочетание.
Другие твердые основы, предусмотренные в настоящем документе, включают смолы или полимеры. Некоторые примеры смол или полимеров включают целлюлозу, акриламид, нейлон, вискозу, полиэфир, анионообменную смолу, AMBERLITE™ XAD-7, AMBERLITE™ XAD-8, AMBERLITE™ IRA-94, AMBERLITE™ IRC-50 (Rohm and Haas, Philadelphia, PA), поливинил, полиакриловые соедиения, полиметакрилат или любое их сочетание.
Другой тип твердой основы, предусмотренный в настоящем документе, включает керамику. Некоторые примеры включают непористую керамику, пористую керамику, SiO2, Al2O3. Другим типом твердой основы, который можно применять в настоящем изобретении, является стекло. Некоторые примеры включают непористое стекло, пористое стекло, аминопропиловое стекло или любое их сочетание. Другим типом твердой основы, который можно использовать, является микроэлектрод. Примером является магнетит, покрытый полиэтиленимином. В качестве твердой основы можно использовать частицы графита.
Другой тип твердой основы, предусмотренный в настоящем документе, содержит продукты диатомовой земли и силикаты. Некоторые примеры включают диатомиты CELITE®, KENITE®, DIACTIV®, PRIMISIL®, DIAFIL® и синтетические силикаты кальция и магния MICRO-CEL®, CALFLO®, SILASORB™ и CELKATE® (World Minerals Inc., Santa Barbara, CA).
Другой пример твердой основы представляет собой или содержит клетку, такую как эритроцит.
Наборы
В определенных вариантах осуществления в настоящем документе предоставлены наборы, содержащие композиции, например, нуклеиновые кислоты, экспрессирующие кассеты, векторы, клетки, трансгенные семена или растения или части растений, полипептиды (например, гидролазы) и/или антитела по изобретению. Также наборы могут содержать инструкцию, в которой описаны способы и промышленные применения по изобретению, которые описаны в настоящем документе.
Промышленные и медицинские применения
Гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы), предоставленные в настоящем документе, имеют много промышленных применений и медицинских применений, и ниже описаны несколько образцовых применений и композиций. Лишь некоторые примеры применения включают процессы по изобретению, включающие превращение негидратируемого фосфолипида в гидратируемую форму, рафинирование масла, обработку продуктов питания, обработку масел (например, получение масла с низким содержанием насыщенных жирных кислот) из растений, рыбы, водорослей и т.п.
Обработка продуктов питания и кормов
В определенных вариантах осуществления в настоящем документе предоставлены процессы сыроварения с использованием гидролаз (например, липаз, сатураз, пальмитаз и/или стеаратаз) по изобретению. В других вариантах осуществления в настоящем документе предоставлены сыры, содержащие гидролазы. В одном из аспектов ферменты по изобретению (например, липазы, сатуразы, пальмитазы и/или стеаратазы или их сочетание) используют для обработки сыров для обогащения запаха, для увеличения выхода и/или для «стабилизации» сыров, например, посредством снижения склонности к «обезжириванию» или в одном из аспектов ферменты по изобретению, используют для получения сыра из молока для сыроделия. Эти процессы по изобретению могут включать любой способ или протокол, например, как описано, например, в патентах США №№ 6551635 и 6399121, WO 03/070013, WO 00/054601. Например, в одном из аспектов гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению используют для стабилизации жировой эмульсии в молоке или композициях, содержащих молоко, например, в сливках, и используют для стабилизации молочных композиций, например, для производства сливок или сливочных напитков. В одном из вариантов осуществления в настоящем документе предоставлены процессы для обогащения аромата сыра с использованием по меньшей мере одного фермента по изобретению, процесс включает инкубацию белка, жира и протеазы и липазы (например по изобретению) в водной среде в условиях, в которых происходит обогащение запаха сыра (например, снижение горечи), например, как описано в WO 99/66805. В одном из аспектов липазы по изобретению используют для обогащения запаха сыра (например, творога) путем смешивания с водой, протеазой и фосфолипазой при повышенной температуре, например, между приблизительно 75°C и 95°C, как описано, например, в патенте США № 4752483. В одном из аспектов липазы по изобретению используют для ускорения созревания сыра посредством добавления фермента по изобретению в сыр (например, в молоко для сыроделия) перед добавлением коагулянта в молоко или посредством добавления фермента (например, липазы) по изобретению в творог с солью перед прессованием, например, как описано, например, в патенте США № 4707364. В одном из аспектов липазу по изобретению используют для расщепления триацилглицерида в молочном жире для высвобождения свободных жирных кислот, что ведет к обогащению запаха. Фермент по изобретению также можно использовать в любом из этих процессов по изобретению, см., например, Brindisi (2001) J. of Food Sci. 66:1100-1107.
Структурированный синтез и обработка масел
В определенных вариантах осуществления в настоящем документе предоставлены способы структурированного синтеза масел, липидов и т.п. с использованием гидролаз (например, липаз, сатураз, пальмитаз и/или стеаратаз) по изобретению. Способы по изобретению, включают биокаталитический синтез структурированных липидов, т.е., липидов, которые содержат определенный набор жирных кислот, которые определенным образом распределены на остове, например, на глицериновом остов. Продукты, созданные с использованием гидролаз и с применением способов по изобретению, включают масла с низким содержанием насыщенных жирных кислот, например, масла из овощей (например, сои, канолы), животных, растений, рыбы, водорослей, где эти масла обработаны или переработаны с использованием полипептида по изобретению; и продукты питания, корма, добавки, фармацевтические препараты и т.п., содержащие масла с низким содержанием насыщенных жирных кислот, полученные путем применения способов и/или композиций (например, ферментов) по изобретению. Продукты, созданные с использованием гидролаз и с применением способов по изобретению, также содержат альтернативы масла какао, липиды, содержащие полиненасыщенные жирные кислоты (ПНЖК), липиды, содержащие незаменимые жирные кислоты, липиды, содержащие мононенасыщенные жирные кислоты, липиды, содержащие фосфохолин и фосфосерин, липиды, содержащие фитостеролы, 1,3-диацилглицериды (ДАГ), 2-моноацилглицериды (МАГ) и триацилглицериды (ТАГ).
Способы по изобретению позволяют синтезировать липиды или жирные кислоты с определенными региоселективностями и стереоселективностями. В настоящем документе предоставлены масла, липиды и т.п., и масла, которые можно использовать в продуктах питания и кормах и кулинарных продуктах (например, в кулинарных маслах, маслах для жарки, пекарных маслах, соусах, маринадах, приправах, маслах для смазывания разбрызгиванием, маргаринах, майонезе, заправках, которые можно черпать ложкой и наливать, альтернативах масла какао и т.п.), которые обработаны или переработаны с использованием полипептидов или пептидов (например, гидролаз, таких как липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению. В определенных вариантах осуществления в настоящем документе предоставлены фармацевтические препараты, нутрицевтики и косметические средства, содержащие полипептиды (например, гидролазы, такие как липазы, сатуразы, пальмитазы и/или стеаратазы; или пептиды или антитела) по изобретению.
В определенных вариантах осуществления в настоящем документе предоставлены способы обработки (модификации) масел, липидов и т.п. с использованием гидролаз по изобретению. Способы можно использовать для обработки масел из растений, животных, микроорганизмов. Способы по изобретению можно использовать в структурированном синтезе масел, схожих с теми, которые найдены в растениях, животных и микроорганизмах. Липиды и масла можно обработать для придания желаемой характеристики. Липиды и масла, которые можно обработать способами по изобретению (с использованием гидролаз по изобретению), включают альтернативы масла какао, липиды, содержащие полиненасыщенные жирные кислоты (ПНЖК), липиды, содержащие незаменимые жирные кислоты, липиды, содержащие мононенасыщенные жирные кислоты, липиды, содержащие фосфохолин и фосфосерин, липиды, содержащие фитостеролы, 1,3-диацилглицериды (ДАГ), 2-моноацилглицериды (МАГ) и триацилглицериды (ТАГ). В одном из аспектов обработанные и синтетические масла и жиры по изобретению (например, альтернативы масла какао и растительные масла) можно использовать в различных применениях, например, в производстве продуктов питания (например, кондитерских изделий, мучных кондитерских изделий) и в составе фармацевтических препаратов, нутрицевтиков и косметических средств. В настоящем документе предоставлены способы обработки жиров и масел из растений, например, из семян масличных культур, включая, например, канолу, клещевину, кокос, кориандр, кукурузу, семя хлопчатника, фундук, конопляное семя, льняное семя, пенник луговой, оливу, пальмовое масло, ядро кокосового ореха, арахис, семя рапса, рисовую мучку, сафлор, сасанкву, соевые бобы, подсолнечник, талловое масло, цубаки, различных «натуральных» масел, обладающих измененным составом жирных кислот посредством генетически модифицированных организмов (ГМО) или традиционной селекции, таких как масла с высоким содержанием олеиновой кислоты, с низким содержанием линоленовой кислоты или с низким содержанием насыщенных жирных кислот (канола с высоким содержанием олеиновой кислоты, соевые бобы с низким содержанием линоленовой кислоты или подсолнечник с высоким содержанием стеариновой кислоты) или смесей любых указанных выше с использованием гидролазы по изобретению.
В определенных вариантах осуществления в настоящем документе предоставлены способы обработки масел из животных, например, из рыбы (тихоокеанского талеихта, печени трески, хоплостета, сардин, сельди, менхадена и т.п.), млекопитающих (свинины, говядины и т.п.) и домашней птицы (кур и т.п.) с использованием гидролаз по изобретению. В определенных вариантах осуществления в настоящем документе предоставлены способы структурированного синтеза масел, схожих с обнаруженными в животных, например, в рыбе, домашней птице и млекопитающих, и в микроорганизмах с использованием гидролаз по изобретению. В одном из аспектов эти синтетические или переработанные масла используют в качестве кормовых добавок, продуктов питания, в качестве компонентов фармацевтических составов, нутрицевтиков или в косметических средствах. Например, в одном из аспектов гидролазы по изобретению используют для гидролитического отщепления жирных кислот от рыбьего жира с тем, чтобы можно было получить жирные кислоты и использовать в качестве кормовой добавки. В одном из аспектов гидролазы по изобретению можно использовать для обработки масла из отходов ресторанов и топленые животные жиры.
В других вариантах осуществления в настоящем документе предоставлены способы обработки жиров и масел, например, масел из водорослей, включая, например, масло Neochloris oleoabundans, масло Scenedesmus dimorphus, масло Euglena gracilis, масло Phaeodactylum tricornutum, масло Pleurochrysis carterae, масло Prymnesium parvum, масло Tetraselmis chui, масло Tetraselmis suecica, масло Isochrysis galbana, масло Nannochloropsis salina, масло Botryococcus braunii, масло Dunaliella tertiolecta, масло видов Nannochloris, масло Spirulina species, масло Chlorophycease (зеленые водоросли) и масло Bacilliarophy или смеси любых указанных жиров и масел.
В одном из аспектов гидролазы по изобретению представляют собой универсальные биокатализаторы в органическом синтезе, например, в структурированном синтезе масел, липидов и т.п. Ферменты по изобретению (включая гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы) могут взаимодействовать с широким диапазоном субстратов, включая вторичные и третичные спирты, например, из натуральных продуктов, такие как α-терпинеол, линалоол и т.п. В некоторых аспектах гидролазы по изобретению обладают энантиоспецифичностью (например, стереоспецифичностью) от хорошей до прекрасной.
В настоящем документе в определенных вариантах осуществления предоставлен процесс превращения масла (например, растительных масел, масел какао и т.п.), содержащий по меньшей мере один фермент (например, липазу, сатуразу, пальмитазу и/или стеаратазу) по изобретению. В одном из аспектов, процесс превращения масла включает контролируемый гидролиз и ацилирование, например, ацилирование глицерина, в результате чего можно получить широкий спектр продуктов высокой чистоты. В одном из аспектов гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению используют для получения диацилглицериновых масел и структурированных пищевых масел. В определенных вариантах осуществления в настоящем документе предоставлены процессы для этерификации пропиленгликоля с использованием фермента по изобретению, например, регио- и/или хемоселективной липазы для монозамещенной этерификации в положении Sn-1. В настоящем документе предоставлены процессы для структурированного синтеза масел с запланированными профилями насыщенных или ненасыщенных жирных кислот с использованием фермента по изобретению, например, регио- и/или хемоселективной липазы для удаления насыщенной жирной кислоты или для нацеленного присоединения жирной кислоты к глицериновому остову.
В одном из аспектов способы по изобретению дополнительно содержат процессы для избирательного удаления жирных кислот (например, нежелательных жирных кислот) из масел, например, для отделения насыщенных и/или ненасыщенных жирных кислот от масел, с использованием гидролазы (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению. Процесс по изобретению позволяет отделять насыщенные и/или ненасыщенные жирные кислоты от любого масла, например, от соевого масла. Фермент может быть хемоселективным и/или энантиоселективным. В одном из аспектов эти процессы позволяют создавать жиры и масла с высокой стабильностью, например, «полезные» масла для жарки. Этот образцовый процесс по изобретению можно использовать для создания масел с меньшим содержанием серы, например, с использованием процесса, включающего удаление серы из нерафинированного масла. Ферменты по изобретению также можно использовать в процессах переэтерификации для этих и других целей.
В одном из аспектов фермент по изобретению используют для создания масла, не содержащего транс-изомеры жирных кислот. В одном из аспектов масло, не содержащее транс-изомеры жирных кислот, получают из частично гидрогенезированного масла для получения масла, содержащего только цис-изомеры жирных кислот. Фермент может быть хемоселективным и/или энантиоселективным.
В других вариантах осуществления в настоящем документе предоставлены процессы для модификации масел какао с использованием фермента по изобретению. Приблизительно 80% масел какао содержат триацилглицериды POP, SOS и POS (P обозначает пальмитиновую жирную кислоту, О обозначает олеиновую жирную кислоту, S обозначает стеариновую жирную кислоту). Насыщенная-ненасыщенная-насыщенная структура жирной кислоты из масел какао придает ей характерные профили плавления, например, в шоколадах. В одном из аспектов процессы структурированного и прямого синтеза по изобретению применяют к маслам какао для уменьшения неоднородности масла какао или для получения синтетических масел какао («альтернативы масла какао»). В одном из аспектов хемоселективную и/или энантиоселективную (например, региоселективную) гидролазу (например, липазу или эстеразу) по изобретению используют для получения альтернативы масла какао, например, заменителя масла какао, заместителя масла какао и/или эквивалента масла какао. В настоящем документе предоставлены альтернативы масла какао, включая заменители масла какао, заместители масла какао и эквиваленты масла какао и их производственные промежуточные соединения, содержащие фермент по изобретению. Процесс по изобретению (с использованием фермента по изобретению) для получения альтернатив масла какао может содержать смешивание растительного масла, например, пальмового масла, с маслом ши или эквивалентом, маслом ореха бассии или эквивалентом и стеринами Sal или эквивалентом и обработку смешанных масел предоставленными в настоящем документе полипептидами. В одном из аспектов процесс по изобретению включает использование переэтерификации. Процесс по изобретению позволяет создавать композиционные или кристаллические формы, которые имитируют «натуральное» масло какао.
В определенных вариантах осуществления в настоящем документе предоставлены процессы (с использованием фермента по изобретению) для получения диацилглицерина (ДАГ), например, 1,3-диацилглицерина, с использованием растительного масла, например, дешевого масла. Фермент может быть хемоселективным и/или энантиоселективным. Процесс по изобретению позволяет получить ДАГ-содержащую композицию, обладающую хорошей стабильностью, длинным сроком хранения и эффективностью при высокой температуре.
Ферменты (гидролазы, например, липазы, сатуразы пальмитазы и /или стеаратазы) по изобретению и способы по изобретению также можно использовать в ферментативной обработке съедобных масел, как описано, например, в патенте США № 6025171. В этом образцовом способе ферменты по изобретению иммобилизуют посредством получения эмульсии, содержащей непрерывную гидрофобную фазу, такую как триацилглицеридное масло, и диспергированную водную фазу, содержащую алифатический фермент, такой как липаза по изобретению, и вещество-носитель, который частично растворен и частично не растворен в водной фазе, и удаления воды из водной фазы до тех пор, пока фаза не превратится в твердый фермент, нанесенный на частицы носителя. Нерастворенная часть вещества-носителя может представлять собой вещество, которое нерастворимо в воде и масле, или водорастворимое вещество в нерастворимой форме, поскольку водная фаза уже насыщена водорастворимым веществом. Можно сформировать водную фазу с использованием неочищенной липазной сбраживаемой жидкостью, содержащей остатки брожения и биомассу, которые могут выполнять функции веществ-носителей. Иммобилизованную липазу можно использовать для перегруппировки сложных эфиров и нейтрализации кислотности в маслах. После реакции иммобилизованный фермент можно регенерировать для последующей реакции путем добавления воды, чтобы добиться частичного растворения носителя, и испарения воды из полученного фермента и водной фазы, содержащей носитель, которая диспергирована в гидрофобной фазе, чтобы снова нанести фермент на частицы носителя.
Ферменты (например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению и способы по изобретению также можно использовать для получения трансэтерифицированных масел, как описано, например, в патенте США № 5288619. В настоящем документе предоставлены способы ферментативной трансэтерификации для получения маргаринового масла, содержащего жирные кислоты с низким содержанием транс-изомеров и с низким содержанием жирных кислот со средней длиной цепи. Способы включают стадии предоставления реакционной смеси для трансэтерификации, которая содержит вещество источника стеариновой кислоты и съедобное жидкое растительное масло, трансэтерификации вещества источника стеариновой кислоты и растительного масла с использованием липазы со специфичностью по положениям 1,3 и затем, в итоге, гидрогенизация смеси жирных кислот, чтобы предоставить повторно используемое вещество источника стеариновой кислоты для повторной реакции с растительным маслом. В настоящем документе предоставлен противоточный способ получения трансэтерифицированного масла. Способ включает стадии предоставления зоны реакции трансэтерификации, содержащей липазу со специфичностью по положениям 1,3, введения растительного масла в зону трансэтерификации, введения вещества источника стеариновой кислоты, пропускания противотока текучей среды сверхкритического газа или субкритического сжиженного газа, осуществления реакции трансэтерификации потока триацилглицерида с потоком стеариновой кислоты или сложного моноэфира стеариновой кислоты в зоне реакции, отведения потока трансэтерифицированного триацилглицеридного маргаринового масла, отведения противоточной текучей фазы, гидрогенизации трансэтерифицированной стеариновой кислоты или сложного моноэфира стеариновой кислоты чтобы предоставить гидрогенизировнное повторно используемое вещество источника стеариновой кислоты, и введения гидрогенизированного повторно используемого вещества источника стеариновой кислоты в зону реакции.
В одном из аспектов, чтобы предоставить ферменту по изобретению возможность действовать, обе фазы, масляную фазу и водную фазу, которая содержит фермент, следует тщательно перемешивать. Простого их перемешивания может оказаться не достаточно. Получению хорошей дисперсии фермента в масле способствует его растворение в небольшом количестве воды, например, 0,5-5% по массе (по отношению к маслу), и эмульсификация в масле в этой форме для формирования капелек диаметром менее 10 мкм (средневзвешенный). Капельки могут быть меньше 1 мкм. Турбулентное перемешивание можно выполнять при радиальных скоростях более 100 см/с. Также можно осуществлять циркуляцию масла в реакторе с использованием внешнего ротационного насоса. Тонкую дисперсию водной фазы, содержащей фермент, также можно получить под действием ультразвука. Можно использовать устройство для получения дисперсий.
В одном из аспектов ферментативная реакция по изобретению протекает на поверхности раздела между масляной фазой и водной фазой. Целью всех этих мероприятий по смешиванию является создание наибольшей возможной поверхности водной фазы, содержащей фермент. Добавление поверхностно-активных веществ увеличивает микродисперсию водной фазы. Следовательно, в некоторых случаях в раствор фермента добавляют поверхностно-активные вещества со значениями ГЛБ выше 9, такие как додецилсульфат натрия, как описано, например, в EP-A 0513709. Аналогичный эффективный способ для улучшения образования эмульсии заключается в добавлении лизолецитина. Добавляемые количества могут находиться в диапазоне от 0,001% до 1% по отношению к маслу. Температура в ходе ферментативной обработки не играет критической роли. Можно использовать температуры между 20°C и 80°C, но последние можно применять лишь в течение короткого времени. В этом аспекте используют липазу по изобретению, которая обладает хорошей температурной устойчивостью и/или устойчивостью к низкому pH. Оптимальным является применение при температуре между 30°C и 50°C. Период обработки зависит от температуры и может занимать меньше времени при повышении температуры. Как правило, достаточно периода времени длительностью от 0,1 до 10 часов или от 1 до 5 часов. В реакторе протекает реакция, которую можно разделить на этапы. Следовательно, возможна непрерывная работа наряду с периодической работой. Реакцию можно осуществлять на различных температурных этапах. Например, инкубация может протекать в течение 3 часов при 40°C, затем в течение 1 часа при 60°C. Если реакция протекает этапами, это также открывает возможность подбирать различные значения pH на отдельных этапах. Например, на первом этапе pH раствора скорректировать до 7, например, и на втором этапе до 2,5 путем добавления лимонной кислоты или других подходящих кислот. Однако по меньшей мере на одном этапе pH раствора фермента должен быть ниже 4 или ниже 3. Если pH впоследствии корректировали ниже этого уровня, то могли обнаружить ухудшение эффекта. Следовательно, можно добавить лимонную кислоту в раствор фермента перед смешиванием последнего с маслом.
Ферменты (гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы) по изобретению и способы по изобретению также можно использовать для получения масел, как описано, например, в патентной заявке США № 11/567318, включенной в настоящий документ в качестве ссылки в полном объеме. В настоящем документе предоставлены непрерывные процессы для ферментативной обработки липидов. Способ относится к процессу и устройству для непрерывной ферментативной переэтерификации композиций, содержащих липиды, с использованием нескольких реакторов с неподвижным слоем, где ток композиции, содержащей липид, через устройство может оставаться по существу постоянным даже при снижении ферментативной активности неподвижного слоя с течением времени, и даже когда неподвижный слой извлекают, например, для ремонта, замены или восполнения.
В настоящем документе в одном из вариантов осуществления предоставлен способ гидролиза масла или жира посредством реакции масла или жира с ферментом пальмитазой. В одном из вариантов осуществления гидролиз проводят в присутствие эмульгатора, обладающим ГЛБ выше 12. В одном из вариантов осуществления фермент пальмитазу кодирует последовательность нуклеиновой кислоты, обладающая по меньшей мере 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 99%, 99,5% или 100% идентичностью последовательностей с SEQ ID № 1 и содержащая i) нуклеотидную замену (или ее эквиваленты), кодирующую аминокислотный остаток в положении 95 (или в его эквиваленте), как указано в таблице 9, ii) нуклеотидные замены (или их эквиваленты), кодирующие аминокислотные остатки в положениях 85 и 172 (или в их эквивалентах), как указано в таблице 15, iii) нуклеотидную замену (или ее эквивалент), кодирующую аминокислотный остаток в положении 83 (или в его эквиваленте), как указано в таблице 16, и iv) следующие молчащие мутации 35GCT, 102GTT, 108AGT, 117CTT, 126AGG, 133TCT и 188ACG. В одном из вариантов осуществления последовательность нуклеиновой кислоты представляет собой последовательность SEQ ID № 1 и содержит i) нуклеотидную замену (или ее эквивалент), кодирующую аминокислотный остаток в положении 95 (или в его эквиваленте), как указано в таблице 9, ii) нуклеотидные замены (или их эквиваленты), кодирующие аминокислотные остатки в положениях 85 и 172 (или в их эквивалентах), как указано в таблице 15, iii) нуклеотидную замену (или ее эквивалент), кодирующую аминокислотный остаток в положении 83 (или в его эквиваленте), как указано в таблице 16, и iv) следующие молчащие мутации 35GCT, 102GTT, 108AGT, 117CTT, 126AGG, 133TCT и 188ACG. В одном из вариантов осуществления фермент пальмитаза представляет собой хит тепловой устойчивости 29, как описано в таблице 9, и содержит i) нуклеотидную замену (или ее эквивалент), кодирующую аминокислотный остаток в положении 95 (или в его эквиваленте), как указано в таблице 9, ii) нуклеотидные замены (или их эквиваленты), кодирующие аминокислотные остатки в положениях 85 и 172 (или в их эквивалентах), как указано в таблице 15, iii) нуклеотидную замену (или ее эквивалент), кодирующую аминокислотный остаток в положении 83 (или в его эквиваленте), как указано в таблице 16, и iv) следующие молчащие мутации 35GCT, 102GTT, 108AGT, 117CTT, 126AGG, 133TCT и 188ACG. В одном из вариантов осуществления фермент пальмитаза, используемый в способах, предоставленных в настоящем документе, представляет собой фермент 29 SM со следующими молчащими мутациями 35GCT, 102GTT, 108AGT, 117CTT, 126AGG, 133TCT и 188ACG, как описано в примере 12.
В определенных вариантах осуществления ГЛБ эмульгатор превышает 12, 14, 16 или 18. В определенных вариантах осуществления эмульгатор выбран из олеата натрия, олеата калия, линолеата натрия, линолеата калия, линолената натрия, линолената калия, лауреата натрия, лауреата калия, стеарата натрия, стеарат калия, пальмитата натрия, пальмитата калия, натриевой соли жирной кислоты пальмового масла, калиевой соли жирной кислоты пальмового масла или их сочетания. В определенных вариантах осуществления реакционная смесь содержит приблизительно от 1 до 20% воды от общей массы реагентов. В одном из вариантов осуществления реакционная смесь содержит приблизительно 1, 3, 5, 7, 10, 15, 17 или 20% воды от общей массы реагентов.
В определенных вариантах осуществления масло или жир смешивают с эмульгатором перед добавлением фермента пальмитазы. В определенных вариантах осуществления смесь масла/жира и эмульгатора гомогенизируют до и/или после добавления фермента пальмитазы, чтобы обеспечить однородную эмульсию.
В определенных вариантах осуществления реакцию проводят при температуре приблизительно от 20 до 70°C. В определенных вариантах осуществления реакцию проводят при температуре приблизительно 20, 30, 40, 50, 60 или 70°C. В определенных вариантах осуществления фермент пальмитаза, предоставленный в настоящем документе, снижает содержание пальмитата в масле/жире приблизительно до 5% или менее. В определенных вариантах осуществления фермент пальмитаза, предоставленный в настоящем документе, снижает содержание пальмитата в масле/жире приблизительно до 5, 4, 3, 2, 1% или менее. В определенных вариантах осуществления желаемое снижение содержания пальмитата наступает приблизительно за или приблизительно менее чем за 48 ч, 24 ч, 20 ч, 16 ч, 12 ч, 10 ч, 5 ч или 3 ч. В определенных вариантах осуществления способ дополнительно содержит предварительную обработку масла/жира для удаления смолы и водной фазы и для снижения содержания свободных жирных кислот. Можно использовать любые способы предварительной обработки, которые профессионал в данной области сочтет подходящими. В определенных вариантах осуществления способ дополнительно содержит добавление основания (добавление каустика) для получения мыла.
В определенных вариантах осуществления масло, используемое в реакции, представляет собой рафинированное масло или нерафинированное масло. В одном из вариантов осуществления реакция дополнительно включает присоединение фосфолипида. В реакциях можно использовать любой фосфолипид, который профессионал в данной области сочтет подходящим. В одном из вариантов осуществления фосфолипидом является лецитин. В одном из вариантов осуществления масло, используемое в реакциях, предусмотренных в настоящем документе, представляет собой рафинированное масло, а реакция содержит присоединение фосфолипида.
Нутрицевтики
В одном из аспектов композиции и способы по изобретению можно использовать для получения нутрицевтиков посредством обработки или синтеза липидов и масел с использованием ферментов по изобретению, например, гидролазы, например, липазы, сатуразы, пальмитазы и/или стеаратазы по изобретению. В одном из аспектов обработанные или синтезированные липиды или масла содержат полиненасыщенные жирные кислоты (ПНЖК), диацилглицериды, например, 1,3-диацилглицериды (ДАГ), моноацилглицериды, например, 2-моноацилглицериды (МАГ), и триацилглицериды (ТАГ). В одном из аспектов нутрицевтики получают посредством обработки диацилглицеридов, например, 1,3-диацилглицеридов (ДАГ), моноацилглицеридов, например, 2-моноацилглицеридов (МАГ), и/или триацилглицеридов (ТАГ) из растительных источников (например, из семян масличных культур) или из животных источников (например, рыбий жир). В определенных вариантах осуществления в настоящем документе предоставлены нутрицевтики (например, диетические композиции), содержащие полипептиды (например, ферменты, пептиды, антитела) по изобретению.
В одном из аспектов композиции и способы по изобретению можно использовать для обогащения диетических композиций, в особенности продуктов на основе коровьего молока, например, составов для новорожденных на основе коровьего молока, с использованием гидролаз, активируемых солями желчных кислот. Композиции, полученные конкретными способами, и композиции по изобретению можно использовать для кормления новорожденных и недоношенных новорожденных, включая введение гидролазы, активируемой солями желчных кислот по изобретению, для усиления усвоения жиров и, следовательно, скорости роста. В определенных вариантах осуществления в настоящем документе предоставлены композиции и способы для лечения субъектов с нарушенным образованием ферментов поджелудочной железы посредством введения гидролазы, активируемой солями желчных кислот, в сочетании с употреблением жиров; также см. обсуждение ниже.
В определенных вариантах осуществления в настоящем документе предоставлены диетические композиции, содержащие гидролазу, например, гидролазу, активируемую солями желчных кислот по изобретению. В определенных вариантах осуществления в настоящем документе предоставлены диетические композиции, содержащие питательную основу, содержащую жир и эффективное количество гидролазы, активируемой солями желчных кислот по изобретению. В одном из вариантов осуществления в настоящем документе предоставлены составы для новорожденных на основе коровьего молока, содержащие гидролазу, например, гидролазу, активируемую солями желчных кислот по изобретению. В одном из аспектов гидролаза по изобретению проявляет активность по расщеплению длинноцепочечных жирных кислот, например, с C12 до C22, которые составляют очень большую процентную долю в большинстве видов молока, например, 99% в грудном молоке человека. См., например, патент США № 5000975.
В определенных вариантах осуществления в настоящем документе предоставлены диетические композиции, содержащие растительное масло, жир и гидролазу по изобретению. В других вариантах осуществления в настоящем документе предоставлены способы обработки продуктов на основе молока и/или композиции, содержащие растительное масло, для получения диетических композиций. В одном из аспектов обработанные композиции содержат масло, содержащее лауриновую кислоту, масло, содержащее олеиновую кислоту, масло, содержащее пальмитиновую кислоту, и/или масло, содержащее линоленовую кислоту. В одном из аспектов рисовое масло, олеиновое масло подсолнечника и/или масло канолы можно использовать в качестве масел, содержащих олеиновую кислоту. В одном из аспектов для использования в нутрицевтиках и диетических композициях жиры и масла из растений, например, из семян масличных культур, включая, например, канолу, клещевину, кокос, кориандр, кукурузу, семя хлопчатника, фундук, конопляное семя, льняное семя, пенник луговой, оливу, пальму, ядро кокосового ореха, арахис, семя рапса, рисовую мучку, сафлор, сасанкву, соевые бобы, подсолнечник, талловое масло, цубаки, различные «натуральные» масла, обладающие измененным составом жирных кислот за счет генетически модифицированных организмов (ГМО) или традиционной «селекции», такие как масла с высоким содержанием олеиновой кислоты, с низким содержанием линоленовой кислоты, или с низким содержанием насыщенных жирных кислот (канола с высоким содержанием олеиновой кислоты, соевые бобы с низким содержанием линоленовой кислоты подсолнечник или с высоким содержанием стеариновой кислоты), смеси любых указанных выше обрабатывают или получают с использованием гидролазы по изобретению. См., например, патент США № 4944944.
В одном из аспектов ферменты по изобретению предоставлены в форме, которая сохраняет стабильность при хранении в составе и/или в желудке, но активна, когда состав достигает той части желудочно-кишечного тракта, в которой состав в норме усваивается. В данной области хорошо известны составы (например, микрокапсулы) для высвобождения в кишечнике, например, биоразрушаемые полимеры, такие как полилактид и полигликолид, как описано, например, в патентах США №№ 4767628; 4897268; 4925673; 5902617.
Кондитерские изделия, масло какао и продукты питания
В одном из аспектов композиции и способы по изобретению можно использовать для получения и обработки твердых масел, таких как масло какао. В другом аспекте в настоящем документе предоставлены кондитерские изделия, масло какао и продукты питания, содержащие полипептиды (например, ферменты, пептиды, антитела) по изобретению.
Композиции и способы по изобретению можно использовать для получения альтернатив масла какао посредством способов «структурированного» синтеза с использованием ферментов, например, гидролаз, например, липаз, сатураз, пальмитаз и/или стеаратаз по изобретению. Например, в одном из аспектов, способы по изобретению позволяют обрабатывать или синтезировать триацилглицериды, диацилглицериды и/или моноацилглицериды для применения в качестве, например, альтернатив масла какао. В одном из аспектов способы по изобретению позволяют получать твердое масло с определенной «пластической областью» для поддержания достаточной твердости при комнатной температуре или ниже. В одном из аспектов предусмотрена очень узкая «пластическая область» обработанного или синтезированного липида, например, в одном из аспектов, он быстро плавится приблизительно при температуре тела. Натуральное масло какао начинает размягчаться при температуре приблизительно от 30°C до 32°C и полностью плавится при температуре приблизительно 36°C. Натуральное масло какао может содержать три 1,3-динасыщенных 2-олеоилглицерина, которые представляют собой 1,3-дипальмитоил-2-олеоилглицерин (POP), 1-пальмитоил-2-олеоил-3-стеароилглицерин (POSt) и 1,3-дистеароил-2-олеоилглицерин (StOSt), в количестве 70% масс. или более. Эти три глицерина обладают похожими друг на друга характеристиками плавления и отвечают за свойства плавления масла какао, обладая очень узкой пластической областью. В определенных вариантах осуществления в настоящем документе предоставлены синтетические масла какао или обработанные масла какао (синтезированные или обработанные с использованием гидролазы по изобретению, все возможные композиции обозначают как альтернативные масла какао) с различными процентными долями 1,3-дипальмитоил-2-олеоилглицерина (POP), 1-пальмитоил-2-олеоилглицерина (POSt) и 1,3-дистеароил-2-олеоилглицерина (StOSt), в зависимости от желаемых свойств синтетического масла какао, и синтетические масла какао с содержанием трех 1,3-динасыщенных 2-олеоилглицеринов более или менее 70% масс. Синтетические масла какао по изобретению могут частично или полностью заменить натуральные или необработанные масла какао и могут сохранить или улучшить важные свойства твердого масла.
В определенных вариантах осуществления в настоящем документе предоставлены синтетические масла какао или обработанные масла какао (синтезированные или обработанные с использованием гидролазы по изобретению) с желаемыми свойствами для использования в кондитерских изделиях, хлебобулочных изделиях и фармацевтических препаратах. В других вариантах осуществления в настоящем документе предоставлены кондитерские изделия, хлебобулочные изделия и фармацевтические препараты и т.п., содержащие гидролазу по изобретению. В одном из аспектов способы по изобретению позволяют получать или обрабатывать липид (жир) из кондитерского изделия (например, из шоколада) или подлежащий использованию в кондитерском изделии. В одном из аспектов липид получают или обрабатывают так, что шоколад меньше пачкает пальцы, чем шоколад, полученный из натурального масла какао, при этом сохраняя определенные характеристики плавления во рту. В одном из аспектов липид получают или обрабатывают так, что кондитерское изделие (например, шоколад) можно получать при сравнительно высокой температуре окружающей среды или получать с использованием охлаждающей воды с относительно высокой температурой. В одном из аспектов липиды получают или обрабатывают так, что кондитерское изделие (например, шоколад) можно хранить в относительно теплых условиях, например, тропических или субтропических условиях или в зданиях с центральным отоплением. В одном из аспектов липиды получают или обрабатывают так, что кондитерское изделие (например, шоколад) будет иметь содержание липида (жира) твердой композиции и качества. Ферменты по изобретению можно использовать для предоставления композиции, заменяющей масло какао, которая может в значительной степени улучшить его тепловую стабильность и заменить его в широком спектре применений.
Получение маргаринов и разрыхлителей
В определенных вариантах осуществления в настоящем документе предоставлены синтетические или обработанные жиры, например, маргарин и разрыхлитель, синтезированные или обработанные с использованием гидролазы по изобретению. В других вариантах осуществления в настоящем документе предоставлены синтетические или обработанные жиры, например, маргарин и разрыхлитель, содержащие полипептиды (например, ферменты, пептиды, антитела) по изобретению.
В одном из вариантов осуществления в настоящем документе предоставлены обработанные жиры, включая растительное масло такого типа как масло из канолы, клещевины, кокоса, кориандра, кукурузы, семени хлопчатника, фундука, конопляного семени, льняного семени, пенника лугового, оливы, пальмового масла, ядра кокосового ореха, арахиса, семени рапса, рисовой мучки, сафлора, сасанквы, кунжута, соевых бобов, подсолнечника, талловое масло, цубаки, различные «натуральные» масла, обладающие измененным составом жирных кислот за счет генетически модифицированных организмов (ГМО) или традиционной «селекции», такие как масла с высоким содержанием олеиновой кислоты, с низким содержанием линоленовой кислоты или с низким содержанием насыщенных жирных кислот (канола с высоким содержанием олеиновой кислоты, соевые бобы с низким содержанием линоленовой кислоты или подсолнечник с высоким содержанием стеариновой кислоты), масла, синтезированные или обработанные с использованием гидролазы по изобретению. Синтетические или обработанные жиры, например, маргарин и разрыхлитель, обладают запланированной желаемой «пластичностью». Многие продукты пластичных жиров, такие как маргарин и разрыхлитель, получают из твердых продуктов и жидких масел в качестве сырья. Например, жидкие масла, такие как масло из канолы, клещевины, кокоса, кориандра, кукурузы, семени хлопчатника, фундука, конопляного семени, льняного семени, пенника лугового, оливы, пальмового масла, ядра кокосового ореха, арахиса, семени рапса, рисовой мучки, сафлора, сасанквы, кунжута, соевых бобов, подсолнечника, талловое масло, цубаки, различные «натуральные» масла, обладающие измененным составом жирных кислот за счет генетически модифицированных организмов (ГМО) или традиционной «селекции», такие как масла с высоким содержанием олеиновой кислоты, с низким содержанием линоленовой кислоты или с низким содержанием насыщенных жирных кислот (канола с высоким содержанием олеиновой кислоты, соевые бобы с низким содержанием линоленовой кислоты или подсолнечник с высоким содержанием стеариновой кислоты), смешивают с их отвержденными маслами (твердые продукты) и смесь доводят до подходящей консистенции (пластичности). Продукты пластичных жиров, такие как полученные таким образом маргарин и разрыхлитель, склонны вызывать образование относительно грубой кристаллической структуры, поскольку жиры и масла, используемые в качестве сырья, состоят из жирных кислот, обладающих углеродной цепью почти такой же длины. Другими словами, они обладают крайне единообразным составом жирных кислот. По этой причине можно поддерживать подходящую степень пластичности этих продуктов только в узком температурном диапазоне с тем, чтобы жидкие масла, содержащиеся в них имели склонность выделяться на поверхности. В настоящем документе предоставлены способы получения или обработки жиров, разработанные так, что они обладают разнообразным (и определенным) составом жирных кислот. Полученное масло, например, маргарин или разрыхлитель, может иметь более широкий диапазон пластичности.
В одном из аспектов способы и композиции по изобретению используют для получения или обработки растительных масел таких типов, как масло из канолы, клещевины, кокоса, кориандра, кукурузы, семени хлопчатника, фундука, конопляного семени, льняного семени, пенника лугового, оливы, пальмы, ядра кокосового ореха, арахиса, семени рапса, рисовой мучки, сафлора, сасанквы, кунжута, соевых бобов, подсолнечника, талловое масло, цубаки, различные «натуральные» масла, обладающие измененным составом жирных кислот за счет генетически модифицированных организмов (ГМО) или традиционной «селекции», такие как масла с высоким содержанием олеиновой кислоты, с низким содержанием линоленовой кислоты, или с низким содержанием насыщенных жирных кислот (канола с высоким содержанием олеиновой кислоты, соевые бобы с низким содержанием линоленовой кислоты или подсолнечник с высоким содержанием стеариновой кислоты) с использованием гидролаз по изобретению, включая переэтерификацию и ферментативную трансэтерификацию, см., например, патент США № 5288619 и патентную заявку США № 11/567318. Способы и композиции по изобретению можно использовать вместо случайной переэтерификации как описано, например, в патенте США № 3949105. В одном из аспектов способы и композиции по изобретению используют в ферментативной трансэтерификации для получения масла, например, маргаринового масла, обладающего как низким содержанием транс-изомеров жирных кислот, так и низким содержанием жирных кислот со средней длиной цепи.
В одном из аспектов симметричная структура масла, например, масла пальмового или лауринового типа модифицируют, например, с получением случайной структуры. Таким образом, способы по изобретению можно использовать для модификации свойств продуктов пластичных жиров. В одном из аспектов модификацию масел способами по изобретению можно предназначить для предотвращения медленного постепенного затвердения масла с течением времени, конкретно при хранении продуктов.
В одном из аспектов способы и композиции по изобретению в реакционной смеси для трансэтерификации содержат вещество источника стеариновой кислоты и съедобное жидкое растительное масло, трансэтерификацию вещества источника стеариновой кислоты и растительного масла с использованием липазы со специфичностью по положениям 1,3 по изобретению и затем гидрогенизацию смеси жирных кислот, чтобы предоставить повторно используемое вещество источника стеариновой кислоты для повторной реакции с растительным маслом. См., например, патент США № 5288619.
В одном из аспектов реакцию переэтерификации проводят с использованием липазы по изобретению. В одном из аспектов липаза по изобретению обладает избирательностью по положениям 1 и 3 триацилглицерида для замедления или сдерживания увеличения количества тринасыщенных триацилглицеридов в масле. В этой реакции по изобретению можно преодолеть недостаток стандартной случайной переэтерификации и затруднение переэтерификации с использованием неспецифической липазы, поскольку переэтерификацию проводят с помощью фермента по изобретению, который обладает специфичностью по положениям 1 и 3 триацилглицеридов. В одном из аспектов выделение жидких масел, содержащихся в продуктах, на поверхности замедляют или предотвращают при повышении температуры в реакции для ингибирования роста температуры плавления, вызванного увеличением количества тринасыщенных триацилглицеридов. Это позволяет решить проблему затвердения продуктов в течение длительного хранения.
Фармацевтические композиции и лечение недостаточностей гидролаз
В определенных вариантах осуществления в настоящем документе предоставлены способы и композиции (ферменты по изобретению, например, эстеразы, ацилазы, липазы, фосфолипазы или протеазы по изобретению), которое можно использовать для лечения недостаточности гидролазы у животного, например, у млекопитающего, такого как человек. Например, в одном из аспектов способы и композиции по изобретению используют для лечения пациентов, страдающих от недостаточности липазы поджелудочной железы. В одном из аспектов липазу вводят перорально. Фермент по изобретению можно доставлять вместо или вместе с препаратом фермента поджелудочной железы свиньи.
В определенных вариантах осуществления в настоящем документе предоставлены фармацевтические композиции, содержащие полипептиды (например, ферменты, пептиды, антитела) по изобретению. Эти фармацевтические композиции могут находиться в форме таблеток, драже, гелей, капсул, гидрогелей, спреев, порошков, аэрозолей, имплантатов, липосом, кремов, мазей, жидкостей, микросфер, частиц ядра в виде микрочастиц, эмульсии, суспензии, наноструктур и т.п. Фармацевтические композиции, содержащие полипептиды (например, ферменты, пептиды, антитела) по изобретению, можно вводить в любой форме, например, перорально, внутрикожно, интраперитонеально, внутривенно, местно и т.п. В одном из аспектов фармацевтические композиции по изобретению имеют состав для местной, сублингвальной, пероральной, внутривенной, подкожной, внутримышечной, чрескожной, внутриартериальной, внутрисуставной или внутрикожной доставки.
В одном из аспектов композиции по изобретению, используемые для такого лечения, обладают активностью в кислых условиях. В одном из аспектов композиции по изобретению вводят перорально в составах (например, таблетках, драже, гелях, капсулах, гидрогелях, спреях, порошках, аэрозолях), которые проходят через участки желудка с кислым pH и высвобождают фермент только в относительно щелочной среде тощей кишки. В одном из аспектов гидролаза по изобретению входит в состав, содержащий носитель, такой как лактоза, сахароза, сорбит, маннит, крахмал, производные целлюлозы или желатин или любой другой такой эксципиент. Также можно добавить лубрикант, такой как стеарат магния, стеарат кальция или полиэтиленгликолевый воск. В качестве покрытия можно добавить концентрированный сахарный раствор, который может содержать добавки, такие как тальк, диоксид титана, желатин или аравийскую камедь. Чтобы инкапсулировать гидролазу в виде жидкого или в виде твердого препарата, можно использовать мягкие или твердые капсулы. См., например, патент США № 5691181; 5858755.
Детергенты
В определенных вариантах осуществления в настоящем документе предоставлены способы и композиции (ферменты, например, липазы, сатуразы, пальмитазы и/или стеаратазы по изобретению), которые можно использовать в получении и использовании детергентов. Гидролазу по изобретению можно добавить, например, смешать с любым известным моющим составом, твердым или жидким, изменяя или не изменяя состава моющей композиции. Для примера, гидролазу по изобретению можно добавить в любое мыло, например, алифатические сульфаты, такие как алкил- или алкенилсульфаты неразветвленной или разветвленной цепью, сульфаты амидов, сульфаты алкиловых или алкениловых простых эфиров, содержащих алкиловую или алкениловую группу с неразветвленной или разветвленной цепью, к которой присоединяют один или несколько этиленоксидов, пропиленоксидов и бутиленоксидов, алифатические сульфонаты такие как алкилсульфонаты, сульфонаты амидов, диалкилсульфосукцинаты, сульфонаты α-олефинов, олефинов винилиденового типа и внутренних олефинов, ароматические сульфонаты, такие как алкилбензолсульфонаты с неразветвленной или разветвленной цепью, карбонаты или амиды алкиловых или алкениловых простых эфиров, содержащие алкиловую или алкениловую группу с неразветвленной или разветвленной цепью, к которой присоединяют один или несколько этиленоксидов, пропиленоксидов и бутиленоксидов, или амиды, соли или сложные эфиры α-сульфожирных кислот, поверхностно-активные вещества аминокислотного типа, фосфатные поверхностно-активные вещества, такие как кислые алкил- или алкенилфосфаты и алкил- или алкенилфосфаты, амфотерные поверхностно-активные вещества типа сульфоновой кислоты, амфотерные поверхностно-активные вещества типа бетаина, алкиловые или алкениловые простые эфиры или спирты, содержащие алкиловую или алкениловую группу с неразветвленной или разветвленной цепью, которой присоединяют один или несколько этиленоксидов, пропиленоксидов и бутиленоксидов, полиоксиэтиленалкилфениловые эфиры, содержащие алкильную группу с неразветвленной или разветвленной цепью, к которой присоединяют один или несколько этиленоксидов, пропиленоксидов и бутиленоксидов, алканоламиды высших жирных кислот или их продукты присоединения алкиленоксида, сложные эфиры сахарозы и жирной кислоты, сложные глицериновые моноэфиры жирных кислот, алкил- или алкениламиноксиды, катионные поверхностно-активные вещества типа солей тетраалкиламмония или их сочетание. См., например, патент США № 5827718.
В некоторых вариантах осуществления в настоящем документе предоставлены моющие композиции, содержащие один или несколько полипептидов (гидролаз) по изобретению. Можно использовать поверхностно-активные и/или не поверхностно-активные формы. В одном из аспектов общее количество гидролазы, поверхностно-активной и/или не поверхностно-активной, может составлять приблизительно от 0,0001% приблизительно до 1,0%, или приблизительно от 0,0002% приблизительно до 0,5% по массе моющей композиции. В одном из аспектов в моющей композиции поверхностно-активная гидролаза составляет приблизительно от 5% приблизительно до 67% и не поверхностно активная гидролаза составляет приблизительно от 33% приблизительно до 95% от общей гидролазной активности в ферментативной смеси. В одном из аспектов оптимальный pH всей ферментативной смеси составляет приблизительно от 5 приблизительно до 10,5.
В одном из аспектов моющие композиции по изобретению содержат щелочные гидролазы по изобретению, которые функционируют при щелочных значениях pH, поскольку в обычных условиях мытья pH моющего раствора может находиться в диапазоне щелочных pH. См., например, патент США № 5454971
Предоставленные в настоящем документе полипептиды (ферменты по изобретению) можно использовать в любых моющих композициях, которые хорошо известны в данной области, см., например, патенты США №№ 5069810; 6322595; 6313081. Например, в одном из аспектов предоставлены моющие композиции для стирки. Они могут содержать от 0,8 м.д. до 80 м.д. липазы по изобретению.
Любой способ получения и использования моющих композиций можно использовать с ферментами по изобретению, см., например, патенты США №№ 6413928; 6399561; 6365561; 6380147. Моющие композиции могут представлять собой одно- и двухкомпонентую водную композицию, неводную жидкую композицию, цельнолитую, гранулярную форму, композицию в форме частиц, прессованную таблетку, гелеобразную форма, порошок, гель, гидрогель, липосомы, аэрозоль, пасту и/или суспензию. Гидролазы по изобретению также можно использовать в качестве моющей добавки в твердой или жидкой форме. Также добавки предназначены для усиления или повышения эффективности стандартных моющих композиций и их можно добавлять на любом этапе процесса очистки.
В определенных вариантах осуществления в настоящем документе предоставлены способы, позволяющие удалять сильные пищевые загрязнения, пленки от остатков пищи и другие второстепенные пищевые композиции с использованием этих моющих композиций. Гидролазы по изобретению могут облегчать удаление пятен посредством каталитического гидролиза липидов, жиров или масел. Гидролазы по изобретению можно использовать в детергентах для мытья посуды и в детергентах для стирки текстиля.
Фактическое содержание активного фермента зависит от способа производства моющей композиции и не является критичным при условии, что моющая композиция обладает желаемой ферментативной активностью. В одном из аспектов количество гидролаз, присутствующих в конечной композиции, находится в диапазоне приблизительно от 0,001 мг до 0,5 мг на грамм моющей композиции. Выбор конкретного фермента для использования в процессе и продуктах, предоставленных в настоящем документе, зависит от условий конечного пользования, включая внешний вид продукта, используемый pH, используемой температуры и типов загрязнителей, подлежащих разрушению или изменению. Можно выбрать фермент для обеспечения оптимальной активности и стабильности для заданного набора подходящих условий применения. В настоящем документе в одном из аспектов предоставлены гидролазы, активные в диапазоне pH приблизительно от 4 приблизительно до 12 и в температурном диапазоне приблизительно от 20°C приблизительно до 95°C. Детергенты по изобретению могут содержать катионные, полуполярные неионные или цвиттерионные поверхностно-активные вещества; или их смеси.
В одном из вариантов осуществления ферменты по изобретению могут входить в состав порошкообразных и жидких детергентов, обладающих pH между 4,0 и 12,0 при концентрации приблизительно от 0,01 приблизительно до 5% (альтернативно от 0,1% до 0,5%) по массе. Эти моющие композиции также могут содержать другие ферменты, такие как протеазы, целлюлазы, липазы или эндогликозидазы, эндо-β-1,4-глюканазы, β-глюканазы, эндо-β-1,3(4)-глюканазы, кутиназы, пероксидазы, лакказы, амилазы, глюкоамилазы, пектиназы, редуктазы, оксидазы, фенолоксидазы, лигниназы, пуллуланазы, арабинаназы, гемицеллюлазы, маннаназы, ксилоглюканазы, ксиланазы, пектинацетилэстеразы, рамногалактуронанацетилэстеразы, полигалактуроназы, рамногалактуроназы, галактаназы, пектинлиазы, пектинметилэстеразы, целлобиогидролазы и/или трансглутаминазы. Эти моющие композиции также могут включать моющие компоненты и стабилизаторы.
Добавление гидролаз по изобретению в стандартные чистящие композиции не создает каких-либо конкретных ограничений использования. Другими словами, любые температура и pH, подходящие для детергента, также подходят для композиций по изобретению при условии, что фермент активен или устойчив к pH и/или температуре предполагаемого использования. Кроме того, гидролазы по изобретению можно использовать в чистящих композициях без детергентов, опять же в отдельности или в сочетании с моющими компонентами и стабилизаторами.
В определенных вариантах осуществления в настоящем документе предоставлены чистящие композиции, включая моющие композиции для чистки твердых поверхностей, моющие композиции для очистки тканей, композиции для мытья посуды, чистящие композиции для ухода за полостью рта, чистящие композиции для зубов и чистящие растворы для контактных линз.
В определенных вариантах осуществления в настоящем документе предоставлены способы мытья объекта, которые включают контакт объекта с полипептидом по изобретению, в условиях, достаточных для мытья. Гидролазу по изобретению можно включить в качестве моющей добавки. Моющие композиции по изобретению можно, например, ввести в состав в виде моющей композиции для ручной или машинной стирки, содержащие полипептид по изобретению. Добавка для стирки, подходящая для предварительной обработки испачканной ткани, может содержать полипептид по изобретению. Смягчающая композиция для ткани может содержать гидролазу по изобретению. Альтернативно гидролазу по изобретению можно ввести в состав в качестве моющей композиции для использования в основных хозяйственных операциях по очистке твердых поверхностей. В альтернативных аспектах моющие добавки и моющие композиции по изобретению могут содержать один или несколько других ферментов, таких как протеаза, липаза, кутиназа, другая протеаза, карбогидраза, целлюлаза, пектиназа, маннаназа, арабиназа, галактаназа, ксиланаза, оксидаза, например, лактаза, и/или пероксидаза (также см. выше). Выбирают фермент(ы) по изобретению со свойствами, совместимыми с выбранным детергентом (т.е. оптимальный pH, совместимость с другими ферментативными и неферментативными компонентами и т.д.), и фермент(ы) присутствует в эффективном количестве. В одном из аспектов ферменты по изобретению используют для устранения веществ с неприятным запахом из тканей. Различные моющие композиции и способы их получения, которые можно использовать, описаны, например, в патентах США №№ 6333301; 6329333; 6326341; 6297038; 6309871; 6204232; 6197070; 5856164.
Когда гидролазы по изобретению вводят в составы композиций, пригодных для использования в способе машинной стирки, они могут содержать как поверхностно-активное вещество, так и соединение моющего компонента. Они могут дополнительно содержать один или несколько компонентов детергента, например, органические полимерные соединения, отбеливатели, дополнительные ферменты, ингибиторы пенообразования, дисперсанты, дисперсанты кальциевого мыла, средства, суспендирующие загрязнители, средства против повторного отложения и ингибиторы коррозии. Композиции для стирки по изобретению также могут содержать смягчители в качестве дополнительных компонентов детергента. Композиции, содержащие гидролазы по изобретению могут обеспечить очистку ткани, удаление пятен, сохранение белизны, смягчение, появление цвета, ингибирование переноса краски и санитарную обработку, когда их вводят в составы в качестве моющих композиций для стирки.
Плотность моющих композиций для стирки по изобретению может находиться в диапазоне приблизительно от 200 до 1500 г/л или приблизительно от 400 до 1200 г/л или приблизительно от 500 до 950 г/л или от 600 до 800 г/л композиции; ее можно измерить приблизительно при 20°C.
«Компактная» форма моющих композиций для стирки по изобретению лучше всего отражена в плотности и, в пересчете на композицию, в количестве неорганической наполняющей соли. Неорганические наполняющие соли являются стандартными компонентами моющих композиций в форме порошка. В стандартных моющих композициях наполняющие соли присутствуют в значительных количествах, как правило, от 17% до 35% по массе от общей композиции. В одном из аспектов компактных композиций, наполняющая соль присутствует в количествах, не превышающих 15% от общей композиции или не превышающих 10% или не превышающих 5% по массе композиции. Неорганические наполняющие соли можно выбрать из сульфатов и хлоридов щелочных и щелочно-земельных металлов, например сульфат натрия.
Жидкие моющие композиции по изобретению также могут находиться в «концентрированной форме». В одном из аспектов жидкие моющие композиции могут содержать меньшее количество воды по сравнению со стандартными жидкими детергентами. В альтернативных аспектах содержание воды в концентрированном жидком детергенте составляет менее 40% или менее 30% или менее 20% по массе моющей композиции. Соединения детергентов по изобретению могут включать составы, как описано в WO 97/01629.
Гидролазы по изобретению можно использовать при составлении различных чистящих композиций. В качестве поверхностно-активных вещества подходит множество известных соединений, включая неионные, анионные, катионные или цвиттерионные детергенты, например, как раскрыто в патентах США №№ 4404128; 4261868; 5204015. Кроме того, ферменты по изобретению можно использовать, например, в применениях кускового или жидкого мыла, составах для ухода за блюдами, чистящих растворах или продуктах для контактных линз, гидролизе пептидов, обработке отходов, в текстильных применениях, в качестве ферментов, расщепляющих слитые белки, при получении белков и т.п. Гидролазы по изобретению могут обеспечить повышенные эксплуатационные характеристики в моющей композиции по сравнению с другой моющей протеазой, то есть, ферментативная группа может усилить очистку от определенных пятен, чувствительных к ферментам, таких как трава или кровь, как определяют посредством обычной оценки после стандартного цикла стирки. Гидролазы по изобретению можно ввести в состав известных порошковых и жидких детергентов, обладающих pH между 6,5 и 12,0 при концентрации приблизительно от 0,01 приблизительно до 5% (например, приблизительно от 0,1% до 0,5%) по массе. Эти моющие чистящие композиции также могут содержать другие ферменты, такие как другие известные эстеразы, фосфолипазы, протеазы, амилазы, целлюлазы, липазы или эндогликозидазы, а также моющие компоненты и стабилизаторы.
Обработка и производство продуктов питания
Гидролазы по изобретению можно использовать для отделения компонентов веществ клеток растений. Например, гидролазы по изобретению можно использовать в разделении богатого белком вещества (например, клеток растений) на компоненты, например, сахарозы из сахарной свеклы или крахмала или сахаров из картофеля, фракции мякоти или кожуры. В одном из аспектов гидролазы по изобретению можно использовать разделения сельскохозяйственных с высоким содержанием белка или масла на полезные фракции белка, масла и кожуры. Процесс разделения можно осуществлять посредством способов, известных в данной области.
Гидролазы по изобретению можно использовать в получении фруктовых или овощных соков, сиропов, экстрактов и т.п. для увеличения выхода. Гидролазы по изобретению можно использовать в ферментативной обработке (например, гидролизе белков) различных веществ, полученных из стенок клеток растений, или отходов, например, от производства вина или сока, или сельскохозяйственных остатков, таких как кожура овощей, кожура бобов, свекловичная пульпа, оливковая пульпа, картофельная пульпа и т.п. Гидролазы по изобретению можно использовать для модификации консистенции и внешнего вида обработанных фруктов или овощей. Гидролазы по изобретению можно использовать для обработки вещества растений для облегчения переработки вещества растений, включая продукты питания, облегчения очистки или экстракции компонентов растений. Гидролазы по изобретению можно использовать для повышения пищевой ценности, снижения способности к связыванию воды, улучшения разлагаемости в растениях сточных вод и/или улучшения превращения вещества растений в силос и т.п.
Животные корма и пищевые или кормовые добавки
В определенных вариантах осуществления в настоящем документе предоставлены способы обработки животных кормов и продуктов питания и пищевых или кормовых добавок с использованием гидролаз по изобретению, животные включают млекопитающих (например, человека), птиц, рыб и т.п. В других вариантах осуществления в настоящем документе предоставлены животные корма, продукты питания, кормовые и пищевые добавки и добавки, содержащие гидролазы по изобретению.
В определенных вариантах осуществления в настоящем документе предоставлены гидролазы для использования в модификации животного корма или пищи, например, для обработки пищи или корма или in vitro (посредством модификации компонентов корма или пищи) или in vivo. В другом аспекте гидролазу по изобретению можно доставлять посредством экспрессии ферментов непосредственно в трансгенных кормовых культурах (в виде, например, трансгенных растений, семян и т.п.), таких как кукуруза, соевые бобы, рапсовое семя, люпин и т.п. В одном из аспектов в настоящем документе предоставлены трансгенные растения, части растений и клетки растений, содержащие последовательность нуклеиновой кислоты, кодирующую полипептид по изобретению. В одном из аспектов нуклеиновую кислоту экспрессируют так, что гидролазу по изобретению получают в поддающихся извлечению количествах. Гидролазу можно получать из любого растения или части растения. Альтернативно растение или часть растения, содержащие рекомбинантный полипептид, можно использовать по существу для улучшения качества пищи или корма, например, повышения пищевой ценности, вкусовой привлекательности и реологических свойств или для разрушения антипитательного фактора.
Переэтерификация
В одном из аспектов способы и композиции, предоставленные в настоящем документе, можно использовать для модификации свойств смесей триацилглицеридов и, в одном из аспектов, их консистенции. В одном из аспектов фермент по изобретению можно использовать в присутствие катализатора, такого как металлический натрий или метоксид натрия для ускорения перегруппировки остатка карбоновой кислоты между молекулами глицерида так, что продукты состоят из смесей глицеридов, в которых остатки жирных кислот случайно распределены по молекулам глицеридов.
В одном из аспектов ферменты по изобретению можно использовать для получения продуктов переэтерификации при условиях реакции, в которых минимизирован гидролиз жира с тем, чтобы преобладающей реакцией стала переэтерификация, катализируемая липазой. Эти условия могут включать, например, ограничение количества воды в системе.
В одном из аспектов ферменты по изобретению можно использовать для катализа реакций переэтерификации с использованием смесей триацилглицеридов и свободных жирных кислот, как описано, например, в ЕР 0093602 B2. В этих случаях ацильные группы триацилглицеридов можно заменить свободной жирной кислотой для получения новых триацилглицеридов, обогащенных присоединенной жирной кислотой. В одном из аспектов 1,3-специфические липазы по изобретению можно использовать, чтобы ограничивать реакцию положениями 1 и 3 в глицеридах, что позволяет получить смесь триацилглицеридов, которые нельзя получить посредством химической переэтерификации или реакции с неспецифической липазой. В одном из аспектов неспецифические липазы используют для достижения результатов, похожих на химическую переэтерификацию.
Возможность получения смесей новых триацилглицеридов с использованием липаз со специфичностью по определенным положениям по изобретению можно использовать в производстве масел и жиров, поскольку некоторые из этих смесей обладают полезными свойствами. Одним из примеров является катализируемая 1,3-специфической липазой переэтерификация 1,3-дипальмитоил-2-монолеина (POP), который является основным триацилглицеридом средней фракции пальмового масла, со стеариновой кислотой или тристеарином для получения продуктов, обогащенных полезным 1-пальмитоил-3-стеароил-2-монолеином (POSt) и 1,3-дистеароил-2-монолеином (StOSt). POSt и StOSt являются важными компонентами масла какао. Таким образом, в одном аспекте по изобретению предусмотрена реакция переэтерификации для получения эквивалентов масла какао из дешевого исходного вещества.
В одном из аспектов в настоящем документе предоставлены способы получения заменителя твердого жира с использованием 1,3-специфических липаз по изобретению. В одном из аспектов заменитель твердого жира содержит среднюю фракцию пальмового масла и StOSt, POSt или StOSt/POSt с чистотой по меньшей мере 85%.
Далее настоящее изобретение описано со ссылкой на следующие примеры; однако следует понимать, что настоящее изобретение не ограничено такими примерами.
ПРИМЕРЫ
Пример 1: Образцовые анализы липазы-сатуразы
В следующем примере описаны образцовые анализы для скрининга гидролазной, например, липазной, сатуразной, пальмитазной и/или стеаратазной активности. В одном из аспектов эти образцовые анализы можно использовать в качестве повседневного скрининга для определения того, входит ли полипептид в объем изобретения. Такие анализы включают использование соединений индикаторов pH для определения отщепления жирных кислот от триацилглицеридов, спектрофотометрические способы, ВЭЖХ, ГХ, MS, TLC и другие. Jaeger (1994) FEMS Microbiol. Rev. 15:29-63; Ader (1997) Methods Enzymol. 286:351-386; Vorderwulbecke (1992) Enzyme Microb. Technol. 14:631-639; Renard (1987) Lipids 22:539-541.
Скрининг липазной/эстеразной активности
Колонии собирали стерильными зубочистками и использовали для однократной инокуляции каждой лунки 96-луночных микротитровальных планшетов. Лунки содержали по 250 мкл среды LB со 100 мкг/мл ампициллина, 80 мкг/мл метициллина и 10% об./об. глицерина (LB амп/мет, глицерин). Клетки выращивали в течение ночи при 37°C без встряхивания. Таким образом, каждая лунка содержала исходную культуру клеток E. coli, каждая из которых содержала pBLUESCRIPT™ с уникальной вставкой ДНК.
96-луночные планшеты использовали для многократной инокуляции одного планшета («конденсированный планшет»), содержащего в каждой лунке по 200 мкл LB амп/мет, глицерин. Эту стадию осуществляли с использованием High Density Replicating Tool (HDRT) из BIOMEK™ (Beckman Coulter, Inc., Fullerton, CA) с 1% отбеливателем, водой, изопропанолом, циклом стерилизации воздушной сушкой между каждой инокуляцией. Таким образом, каждая лунка конденсированного планшета содержала от 10 до 12 различных клонов pBLUESCRIPT™ из каждого планшета исходной библиотеки. Конденсированный планшет выращивали в течение 16 часов при 37°C и затем использовали для инокуляции двух белых 96-луночных микротитровальных дочерних планшетов (Polyfiltronics, Inc., Rockland MA), которые в каждой лунке содержали по 250 мкл LB амп/мет (без глицерина). Исходный конденсированный планшет помещали на хранение при -80°C. Два конденсированных дочерних планшета инкубировали при 37°C в течение 18 часов.
«600 мкМ исходный раствор субстрата» эстеразы с короткими цепями получали следующим образом: в подходящем объеме ДМСО растворяли по 25 мг каждого из следующих соединений для получения 25,2 мМ раствора. Использовали соединения 4-метилумбеллиферилпропионоат, 4-метилумбеллиферилбутират и 4-метилумбеллиферилгептаноат. 250 мкл каждого раствора в ДМСО добавляли приблизительно в 9 мл 50 мМ буфера HEPES, pH 7,5, который содержал 0,6% Triton X-100 и 0,6 мг/мл додецил мальтозида (Anatrace, Maumee, OH). Объем доводили до 10,5 мл буфером HEPES, чтобы получить слегка замутненную суспензию.
«600 мкМ исходный раствор субстрата» с длинными цепями получали следующим образом: в ДМСО растворяли по 25 мг каждого из следующих соединений до концентрации 25,2 мМ, как указано выше. Использовали соединения 4-метилумбеллиферилэлаидат, 4-метилумбеллиферилпальмитат, 4-метилумбеллиферилолеат и 4-метилумбеллиферилстеарат. Все их необходимо было непродолжительно нагревать в ванне при температуре 70°C, чтобы добиться растворения. 250 мкл каждого раствора в ДМСО добавляли в буфер HEPES и разбавляли до 10,5 мл, как указано выше. Все семь производных умбеллиферила получали из Sigma Chemical Co. (St. Louis, MO).
В каждую лунку белого конденсированного планшета добавляли 50 мкл «600 мкМ исходного раствора субстрата» эстеразы с длинными цепями или эстеразы с короткими цепями с использованием BIOMEK™ для получения конечной концентрации субстрата приблизительно 100 мкМ. Значения флуоресценции регистрировали (возбуждение=326 нм, испускание=450 нм) на флуориметре для считывания планшетов непосредственно после добавления субстрата. Планшет инкубировали при 70°C в течение 60 минут в случае субстратов с длинными цепями и в течение 30 минут при КТ в случае субстратов с короткими цепями. Снова регистрировали значения флуоресценции. Для определения присутствия активного клона сравнивали исходные и конечные значения флуоресценции.
Для выделения отдельного клона, обладающего активностью, планшеты Source GenBank оттаивали и использовали отдельные лунки для однократной инокуляции нового планшета, содержащего LB амп/мет. Как указано выше, для роста клеток планшет инкубировали при 37°C, добавляли 50 мкл 600 мкМ исходного раствора субстрата с использованием BIOMEK™ и определяли флуоресценцию. После идентификации активной лунки из исходного планшета, клетки из этой активной лунки переносили полосами на агар с LB/амп/мет и растили в течение ночи при 37°С для получения отдельных колоний. Стерильной зубочисткой собрали восемь отдельных колоний и использовали для однократной инокуляции лунок 96-луночного микротитровального планшета. Лунки содержали по 250 мкл LB амп/мет. Клетки растили в течение ночи при 37°C без встряхивания. Аликвоту 200 мкл удаляли из каждой лунки и анализировали с использованием подходящих субстратов с длинной или короткой цепью, как указано выше. Идентифицировали наиболее активный клон и оставшиеся 50 мкл культуры использовали для переноса полосами на чашку с агаром с LB/амп/мет. Восемь отдельных колоний собирали, выращивали и анализировали, как указано выше. Наиболее активный клон использовали для инокуляции 3 мл культур LB/амп/мет, которые растили в течение ночи. Плазмидную ДНК выделяли из культур и использовали для секвенирования.
Пример 2: Образцовые протоколы для определения профилей высвобожденных жирных кислот в результате ферментативного гидролиза растительного масла посредством LCMS
В следующем примере описаны образцовые способы (протоколы) проведения ферментативного гидролиза растительного масла, такого как соевое масло (использовали в этом примере), (включая получение фермента) с использованием, например, ферментов по изобретению. В этом примере также описаны образцовые способы (протоколы) качественного и количественного определения жирных кислот, высвобожденных из масла. Описан способ с использованием липазы SEQ ID № 2, но можно использовать другие ферменты, включая ферменты по изобретению, например, образцовые ферменты, обладающие последовательностями, как указано в SEQ ID № 2, и содержащие модификации одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати или двенадцати или более или всех аминокислотных остатков, описанные в таблице 3, таблице 4, таблице 9, таблице 10, таблице 11, таблице 16 или таблице 23.
Экспрессия белка в глубоком 96-луночном планшете:
1. Растить липазные клоны E. coli в течение ночи при 30°C в 1 мл среды ТВ, содержащей карбенициллин (100 мкг/мл), в глубоких 96-луночных планшетах. Регистрировать положение и идентичность клонов.
2. Инокулировать свежие глубокие 96-луночные планшеты, содержащие среду ТВ (1 мл; карбенициллин 100 мкг/мл), жидкими культурами (10 мкл/лунка).
3. Инкубировать культуру в течение ночи при 30°C при встряхивании со скоростью 200 об/мин.
4. Индуцировать экспрессию белка посредством переноса 500 мкл каждой ночной культуры в свежий 96-луночный планшет, содержащий среду ТВ (500 мкл/лунка; карбенициллин 100 мкг/мл) и безводный тетрациклин (200 нг/мл).
5. Инкубировать при 30°C в течение 2 часов при встряхивании со скоростью 200 об/мин.
6. Собрать клетки центрифугированием каждого планшета в течение 10 минут на скорости 3000×g. Удалить супернатант. Клеточные осадки можно использовать сразу в анализе масел или хранить при -20°C для дальнейшего использования.
Реакция ферментативного гидролиза масла:
1. В каждый клеточный осадок добавить 100 мкл B-PER™ (Pierce Chemical, Rockford, IL). Если осадки хранили при -20°C, то перед добавлением B-PER™ оставить оттаивать в течение 10 мин при комнатной температуре.
2. Добавить 400 мкл соевого масла в каждую лунку глубокого 96-луночного планшета.
3. В каждую лунку добавить несколько бусин (стекло 710-1180 мкм). Запечатать планшеты, используя CAPMATS™ (Whatman, Florham Park, NJ).
4. Лизировать клетки и получить эмульсию масло/фермент/буфер с использованием Mixer Mill (Retsch Inc., Newtown, PA). Поместить пару запечатанных планшетов в Mixer Mill и встряхивать в течение 30 секунд с частотой 30 циклов/секунда.
5. Заменить изоляцию CAPMATS™ на газопроницаемую изоляцию.
6. Инкубировать планшеты в течение 2 часов при 37°C при встряхивании на скорости 200 об/мин.
Экстракция жирных кислот:
1. В каждую лунку глубокого 96-луночного планшета добавить по 1 мл растворителя для экстракции (CHCl3:MeOH:4N HCl (2:1:0,075)).
2. Пипетировать смесь несколько раз до тех пор, пока она не приобретет гомогенный вид.
3. Закрыть планшеты изоляцией из алюминиевой фольги.
4. Центрифугировать в течение 5 минут при 3000×g. Нарушить изоляцию надрезом, используя лезвие бритвы.
5. Пройти концом пипетки через верхнюю фазу и перенести 5 мкл нижней фазы в новый глубокий 96-луночный планшет, содержащий MeOH 995 мкл/лунка (т.е. разведение нижней фазы 1/200). Избегать контаминации верхней фазой. Хранить отделенные экстракционные смеси при 4°C.
6. Перенести по 150 мкл разведений всех образцов 1/200 в полистироловый 96-луночный планшет.
7. Для предотвращения испарения запечатать планшеты термосваркой. Убедиться в отсутствии контакта MeOH с изоляцией, поскольку это будет препятствовать правильной адгезии.
8. Анализировать образцы посредством LC/MS.
Анализ LC/MS:
1. Образцы, представленные в формате 96-луночных планшетов, впрыскивали через автодозатор HTCPAL™ (LEAP Technologies, Carrboro, NC) в изократную смесь Н2О/MeCN (10/90, об./об.) и 0,1% муравьиную кислоту, подаваемые насосами LC-10ADVP™ (Shimadzu, Kyoto, Japan) со скоростью 1,2 мл/мин.
2. Разделение осуществляли с помощью 150×2,00 мм колонки SYNERGI MAX-RP™ (Phenomenex, Sutter Creek, CA) и определения. Количественное определение выполняли с помощью тройного квадрупольного масс-спектрометра API 4000™ (Applied Biosystems, Foster, CA) с использованием электрораспылительной ионизации (ESI) и мониторинга нескольких ионов с массами 277, 279, 281, 255, 283 в режиме отрицательно заряженных ионов.
3. Контроль оборудования и генерацию данных выполняли с использованием программного обеспечения ANALYST™ 1.3 (Applied Biosystems, Foster, CA).
4. LC/MS калибровали для каждой жирной кислоты в диапазоне от 0,5 до 50 мкг с использованием эталонных образцов (Sigma). Этот диапазон лучше всего соответствует калибровочной кривой квадратической регрессии, которую используют для вычисления количества каждой жирной кислоты, высвобожденной в образцах ферментов.
Пример 3: Образцовые протоколы для высокопроизводительного скрининга эволюционных библиотек липаз на повышенную избирательность гидролиза сложных эфиров пальмитиновой или стеариновой кислоты по сравнению со сложными эфирами олеиновой кислоты
В следующем примере описаны образцовые способы (протоколы) высокопроизводительного скрининга «эволюционных библиотек» липаз на повышенную избирательность к гидролизу сложных эфиров пальмитиновой или стеариновой кислоты по сравнению со сложными эфирами олеиновой кислоты. В этом образцовом способе (протокол/высокопроизводительный скрининг) описан скрининг эволюционных библиотек липаз, полученных из SEQ ID № 2, который можно применять и к другим ферментам, включая ферменты по изобретению, например, образцовые ферменты, содержащие последовательности, как указано в SEQ ID № 2, и обладающие модификациями одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати или двенадцати или более или всех аминокислотных остатков, описанными в таблице 3 или таблице 4; и этот образцовый способ (протокол) можно применять к другим типам библиотек.
Эти образцовые высокопроизводительные скрининги проводят с использованием двух флуорогенных субстратов: метилумбеллифериловые сложные эфиры пальмитиновой или стеариновой кислоты в сравнении с метилумбеллифериловым сложным эфиром олеиновой кислоты.
Порядок проведения высокопроизводительного скрининга:
1. Клоны из библиотек помещают в микротитровальные планшеты и анализируют в первичном высокопроизводительном скрининге.
2. Клоны, у которых установлена повышенная избирательность, обозначают как первичные хиты.
3. Первичные хиты повторно помещают в микротитровальные планшеты и анализируют во вторичном высокопроизводительном скрининге.
4. Клоны, у которых подтверждена повышенная избирательность, обозначают как вторичные хиты.
5. Вторичные хиты секвенируют для идентификации присутствующих мутаций последовательности и проводят анализ с маслом (см. отдельный протокол).
Протокол высокопроизводительного анализа
1. Нанести штрих-коды на черные 384-луночные планшеты для анализа; нанести штрих-коды на 384-луночные планшеты для роста и внести среду LB по 30 мкл/лунка (карбенициллин 100 мкг/мл).
2. Выборочно перенести клоны в планшеты для роста и растить в течение ночи при 30°C во влажном инкубаторе.
3. Индуцировать экспрессию липазы добавлением среды LB по 30 мкл/лунка (карбенициллин 100 мкг/мл), содержащей 4 мкг/мл безводного тетрациклина, и инкубировать 2 часа при 30°C.
4. Лизировать клетки добавлением 20 мкл/лунка B-PER™ (Pierce Chemical, Rockford, IL); хранить при комнатной температуре до тех пор, пока не поместят на робота.
5. Запустить анализ липазной активности на роботе (см. ниже).
6. Клоны, у которых установлена повышенная избирательность к MeUMB сложным эфирам пальмитиновой или стеариновой кислоты по сравнению с MeUMB сложным эфиром олеиновой кислоты, обозначают как хиты.
7. Выборочно перенести клоны хитов в глубокие 96-луночные планшеты, содержащие среду LB (1 мл/лунка; карбенициллин 100 мкг/мл) и растить в течение ночи при 30°C.
8. Во вторичном скрининге первичные хиты повторно помещают в 384-луночные планшеты и повторяют стадии 1-8; обозначают клоны хитов как вторичные хиты.
9. После стадии 8 вторичные хиты передают на секвенирование.
Пример протокола автоматизированного высокопроизводительного скрининга
1. Абрикос: смешать и перенести аликвоту (10 мкл) лизированных клеток из «планшета для роста» (см. стадии 1-4 выше) в каждый из двух отдельных планшетов для анализа (1 и 2).
2. MULTIDROP™ (Thermo Electron Corporation, Milford, MA): Добавить 70 мкл субстрата 1 (УМБ-16:0) в планшет для анализа 1; добавить 70 мкл субстрата 2 (УМБ-18:1) в планшет для анализа 2.
3. Инкубировать планшеты для анализа в течение 20 минут при 37°C.
4. Прочитать на флуориметре: возбуждение 360 нм и испускание 465 нм.
Клоны вторичных хитов, у которых определили наличие уникальных последовательностей, помещают и растят в 96-луночных планшетах и проводят анализ на соевом масле (см. ниже).
Структуры флуорогенных субстратов, используемые в высокопроизводительном скрининге

Пример 4: Образцовая эволюция для усовершенствованного гидролиза сложных эфиров пальмитиновой или стеариновой кислоты с использованием технологии GSSMSM
В следующем примере описаны и обобщены результаты образцовой «эволюции фермента» и протоколы скрининга, которые позволяют идентифицировать образцовые ферменты по изобретению, например, ферменты, содержащие последовательность, как указано в SEQ ID № 2, но также содержащие модификации остатков, как указано в таблице 3 или таблице 4; или ферменты, кодируемые нуклеиновой кислотой, содержащей последовательность, как указано в SEQ ID № 1, но также содержащие модификации остатков, как указано в таблице 3 или таблице 4. В одном из аспектов в образцовом скрининг-анализе для идентификации этих образцовых ферментов по изобретению используют соевое масло в качестве субстрата, а жирные кислоты, высвобожденные (гидролитически отщепленные) из соевого масла, характеризуют, например, как линоленовую кислоту, линолевую кислоту, олеиновую кислоту, пальмитиновую кислоту или стеариновую кислоту.
Соевое масло имеет следующее распределение жирных кислот: линоленовая = 8%; линолевая = 53%; олеиновая = 23%; пальмитиновая = 12%; стеариновая = 4%. Таким образом, если процент пальмитиновой кислоты, высвобожденной (гидролитически отщепленной) из соевого масла образцовым ферментом по изобретению, превышает 12%, то этот фермент обладает склонностью к гидролитическому отщеплению (высвобождению) пальмитиновой кислоты.
Скрининг пальмитаз: получение «библиотеки пальмитаз»
Библиотеку вариантов пальмитазы SEQ ID № 2 получали посредством технологии GSSMSM (патент № 6171820). Точечные мутации вводили с использованием вырожденных олигонуклеотидов в одно положение аминокислоты за один раз с тем, чтобы каждый исходный кодон заменить на каждую из 20 кодируемых природных аминокислот. Мутировавшие варианты трансформировали в организм-хозяин Escherichia coli TOP 10 (Invitrogen, USA) для экспрессии и скрининга. Библиотеку конструировали в экспрессирующем векторе pASK-5, который является модификацией вектора pASK-IBA (IBA GmbH, Germany). Чтобы получить pASK-5, исходный линкер клонирования заменяли новыми сайтами клонирования, а именно последовательность с XbaI до HindIII вектора pASK-IBA заменяли на следующую последовательность:

Экспрессию вариантов GSSMSM индуцировали ангидротетрациклином после достижения оптимальных плотностей клеток-хозяев.
Проводили скрининг ферментов, содержащих аминокислотные последовательности, созданные посредством технологии GSSMSM, используя протокол высокопроизводительного скрининга, например, протокол, описанный в примере 3, в котором определяли, какая жирная кислота предпочтительно подвергается гидролитическому отщеплению от жира - соевого масла в этом анализе. Цель эволюционного проекта состояла в усовершенствовании избирательности исходной последовательности, SEQ ID № 2, к пальмитату на масле. Анализ содержит приведение фермента с новой/модифицированной последовательностью в контакт с соевым маслом, которое содержит различные жирные кислоты, включая линоленовую кислоту, линолевую кислоту, олеиновую кислоту, пальмитиновую кислоту и стеариновую кислоту (см. распределение в %, приведенное выше) и измерение количества каждой жирной кислоты, гидролитически отщепленной посредством каждого модифицированного фермента. Идентифицировали «библиотеку» последовательностей, которые позволяли ферменту гидролитически отщеплять предпочтительно пальмитиновую кислоту (или стеариновую кислоту, см. ниже) от соевого масла (так называемая «библиотека пальмитаз»):
• Первичный и вторичный скрининг проводили с использованием высокопроизводительного скрининга, например, способа, описанного в примере 3;
• Секвенирование вторичных хитов позволило идентифицировать мутации аминокислот, которые ведут к усовершенствованной избирательности гидролитического отщепления пальмитата по сравнению с олеатом в высокопроизводительном скрининге в сравнении, например, с исходной последовательностью, SEQ ID № 2;
• Один клон для каждого варианта кодона, кодирующего мутацию аминокислоты, выбирали и переносили в 96-луночные планшеты для проведения анализа с маслом;
• В анализах с маслом получали избирательности мутировавших ферментов к пальмитату или стеарату или другим жирным кислотам (таблица 3);
• Для наиболее эффективного хита выход пальмитата составлял 59% от высвобожденных жирных кислот (ЖК) в сравнении с 43% для SEQ ID № 2 в том же анализе; это соответствует увеличению коэффициента избирательности от 3,6 до 4,9;
• Некоторые клоны показали увеличение избирательности к стеарату.
Ниже в таблице 1 суммированы мутации GSSMSM (см. выше), отобранные для включения в «библиотеку пальмитаз», которые подлежат объединению посредством технологии GeneReassemblySM (см. пример 5). В одном образцовом анализе идентифицировали четырнадцать (14) мутаций отдельных аминокислот в качестве приносящих наибольшее увеличение гидролитического отщепления пальмитата в анализах с маслом (также ниже см. таблицы 1, 3 и 4). Остатки помечали в соответствии с тем порядком, в котором они встречаются в родительской SEQ ID № 2 (см. фиг. 7), среди остатков, которые дают значительное увеличение гидролитического отщепления пальмитата или стеарата в анализах с маслом. Приведены «исходная AA» в SEQ ID № 2 и полезная мутация («новые аминокислоты»), т.е. образцовые последовательности по изобретению. В одном из аспектов, одиночные мутации в аргинин (R) в положениях остатков 163 и 164 можно включать попеременно так, что эта образцовая библиотека будет содержать клоны с последовательностями 163V-164D (SEQ ID № 2), 163R-164D и 163V-164R, но не с последовательностью 163R-164R.
На фиг. 6a показаны эффекты GSSMSM мутаций образцовой пальмитазы на гидролиз пальмитата и стеарата относительно исходной SEQ ID № 2. Для каждой из четырнадцати (14) одиночных мутаций аминокислот, отобранных для включения в библиотеку пальмитаз GeneReassembly, графически отображено изменение процентной доли высвобожденного пальмитата и стеарата относительно исходной SEQ ID № 2. Многие из этих мутаций дают значительные повышения гидролиза пальмитата, что сопровождается повышением гидролиза стеарата в небольшой или значительной степени. Однако некоторые мутации вызывают легкое снижение гидролиза стеарата. Звездочки обозначают мутации, идентифицированные как передающие повышенную избирательность сатуразного типа.
Скрининг стеаратаз: получение «библиотеки стеаратаз»
Библиотеку вариантов стеаратазы SEQ ID № 2 получали посредством технологии GSSMSM (патент № 6171820). Точечные мутации вводили с использованием вырожденных олигонуклеотидов, в одно положение аминокислоты за один раз, с тем, чтобы можно было заменить каждый исходный кодон на каждую из 20 кодируемых природных аминокислот. Мутировавшие варианты трансформировали в организм-хозяин Escherichia coli TOP 10 (Invitrogen, USA) для экспрессии и скрининга. Библиотеку конструировали в экспрессирующем векторе pASK-5 (как описано выше). Экспрессию вариантов GSSMSM индуцировали ангидротетрациклином после достижения оптимальных плотностей клеток-хозяев.
Проводили скрининг ферментов, содержащих аминокислотные последовательности, созданные посредством технологии GSSMSM, с использованием протокола высокопроизводительного скрининга, например, протокола, описанного в примере 3, который позволяет определить, какая жирная кислота предпочтительно гидролитически отщеплялась от жира, соевого масла в этом анализе. Анализ содержал приведение фермента с новой/модифицированной последовательностью в контакт с соевым маслом, которое содержит различные жирные кислоты, включая линоленовую кислоту, линолевую кислоту, олеиновую кислоту, пальмитиновую кислоту и стеариновую кислоту (см. % распределение, приведенное выше) и измерение количества каждой жирной кислоты, гидролитически отщепленной посредством каждого модифицированного фермента. Идентифицировали «библиотеку» последовательностей, которые позволяют ферменту предпочтительно гидролитически отщеплять стеариновую кислоту (или пальмитиновую кислоту, см. выше), от соевого масла (так называемая «библиотека стеаратаз»):
• Первичный и вторичный скрининг проводили с использованием высокопроизводительного скрининга, например, способа, который описан в примере 3;
• Секвенирование идентифицированных аминокислотных мутаций вторичных хитов, которые ведут к усовершенствованной избирательности гидролиза стеарата по сравнению с олеатом в высокопроизводительном скрининге в сравнении, например, с исходной последовательностью, SEQ ID № 2.
• Один клон для каждого варианта кодона, кодирующего мутацию аминокислоты, выбирали и переносили в 96-луночные планшеты для анализа с маслом.
• Анализы с маслом секвенированных вторичных хитов давали избирательности мутировавших ферментов к пальмитату или стеарату или к другим жирным кислотам (таблица 3).
• Наиболее эффективный хит давал стеарат в количестве 22% от высвобожденных ЖК в сравнении с 9% для SEQ ID № 2 в том же анализе; это соответствует увеличению коэффициента избирательности с 2,3 до 5,5;
• Несколько клонов также показали увеличение избирательности к пальмитату.
Ниже в таблице 2 суммированы мутации GSSMSM (см. выше), выбранные для включения в «библиотеку стеаратаз», которые подлежат объединению посредством технологии GeneReassemblySM. В одном образцовом анализе двадцать две (22) мутации отдельных аминокислот идентифицировали в качестве дающих наибольшее увеличение гидролитического отщепления стеарата в анализах с маслом (также ниже см. таблицы 2, 3 и 4). Остатки помечали в соответствии с порядком, в котором они встречаются в «исходной» SEQ ID № 2, среди остатков, которые дают значительное увеличение гидролиза пальмитата или стеарата в анализах с маслом. Приведены «Исходная аминокислота» в SEQ ID № 2 и полезные мутации («Новые аминокислоты»), т.е. образцовые последовательности по изобретению. В одном из аспектов одиночная мутация в аланин (a) в положении остатка 223 включена в качестве фиксированной мутации так, чтобы каждый клон в этой образцовой библиотеке содержал эту мутацию.
На фиг. 6b (также см. выше) показаны эффекты двенадцати (12) из двадцати двух (22) ведущих GSSMSM мутаций стеаратазы на гидролиз пальмитата и стеарата относительно исходной SEQ ID № 2. Для каждой из двенадцати (12) отдельных мутаций аминокислот, приведенных на фиг. 6b и выбранных для включения в библиотеку стеаратаз GeneReassembly, графически отображено изменение процентной доли высвобожденного пальмитата и стеарата относительно исходной SEQ ID № 2. Большинство этих мутаций давали значительные увеличения гидролиза стеарата, но легкое или значительное снижение гидролиза пальмитата. Звездочки обозначают мутации, идентифицированные как передающие повышенную избирательность сатуразного типа, т.е. увеличение избирательности гидролитического расщепления пальмитата и стеарата в сравнении с гидролитическим отщеплением ненасыщенных жирных кислот в масле, например, олеата, линолеата и линолената.
• Сводка
• Скрининг «библиотеки GSSMSM» (см. подробное описание технологии GSSMSM выше) на основе родительской SEQ ID № 2 дал клоны с одиночными мутациями аминокислот со значительным увеличением избирательности к пальмитату и стеарату и избирательности к сложным эфирам насыщенных жирных кислот, т.е. избирательности гидролиза пальмитата и стеарата (например, избирательного гидролитического отщепления пальмитата и/или стеарата от соевого масла);
• Выявлены клоны со значительным улучшением избирательности к стеарату (избирательного гидролитического отщепления стеариновой кислоты относительно других жирных кислот);
• Выявлены мутанты GSSMSM с увеличенной избирательностью к пальмитату (избирательное гидролитическое отщепление пальмитиновой кислоты относительно других жирных кислот) относительно фермента SEQ ID № 2.
Ниже в таблице 3 и таблице 4 описаны (дополнительно резюмированы) последовательности образцовых ферментов гидролаз по изобретению, например, образцовые ферменты, содержащие последовательность, как указано в SEQ ID № 2, и содержащие изменения по меньшей мере одного (одного, нескольких или всех) аминокислотного остатка, описанные в таблицах. В таблице 3 и таблице 4 также резюмированы данные об активности выбранных образцовых ферментов; данные включают сопоставление конкретных образцовых ферментов с их положительной гидролазной активностью, включая катализ гидролитического отщепления (высвобождения) пальмитиновой или стеариновой жирной кислоты от соевого масла, как установлено посредством протокола высокопроизводительного скрининга, как описано выше.
В таблице 3 и таблице 4 термин «Исходная аминокислота» обозначает намеченный аминокислотный остаток (указанный под «Аминокислотным остатком») в «родительском» ферменте SEQ ID № 2 («намеченный» для изменения); а термин «Новые аминокислоты» обозначает вновь спроектированный аминокислотный остаток (который заменяет соответствующий «намеченный» остаток в «старой последовательности») в образцовом (новом) ферменте по изобретению. Перечисление остатков «Новых аминокислот» в колонке «стеарат» против колонки «пальмитат» указывает на то, какой из двух высокопроизводительных скринингов жирных кислот (т.е., высвобождение пальмитиновой кислоты в одном скрининге и высвобождение стеариновой кислоты в другом скрининге, см. пример 3) использовали для определения (идентификации) конкретного фермента с указанным изменением остатка (последовательность нового фермента, остатки «Новая Аминокислота»).
Например, в первой строке в таблице 3 в 7 аминокислотном остатке тирозин (или «Y») из «родительского» фермента SEQ ID № 2 замещают аминокислотным остатком аргинина (или «R») и этот новый фермент (Y7R) обладает активностью, которая отличается от активности родительского фермента (см. таблицу 3); например, в «Данных о масле» резюмировано субстратное предпочтение (жирной кислоты) нового фермента (например, фермента Y7R) путем перечисления высвобожденных (гидролизованных) жирных кислот, полученных при воздействии (контакте) фермента с соевым маслом (анализы описаны выше); стоит отметить, что соевое масло субстрата содержит несколько доступных для гидролитического отщепления составляющих групп жирных кислот, включая линоленовую кислоту, линолевую кислоту, олеиновую кислоту, пальмитиновую кислоту, стеариновую кислоту.
Например, в первой строке для фермента Y7R 8,3% высвобожденных жирных кислот (из реагировавшего соевого масла) составила линоленовая кислота, 22,1% высвобожденных жирных кислот составила линолевая кислота; 19,7% высвобожденных жирных кислот составила олеиновая кислота; 41,5% высвобожденных жирных кислот составила пальмитиновая кислота; 8,4% высвобожденных жирных кислот составила стеариновая кислота (сумма этих четырех чисел составляет 100%).
В колонке P+S приведены суммы двух частных значений P и S, чтобы показать, какую часть от всех высвобожденных жирных кислот составляли пальмитиновая кислота и стеариновая кислота (41,5%+8,4%=49,9% гидролитически отщепленных жирных кислот составляли пальмитиновая кислота и стеариновая кислота, или «P+S»).



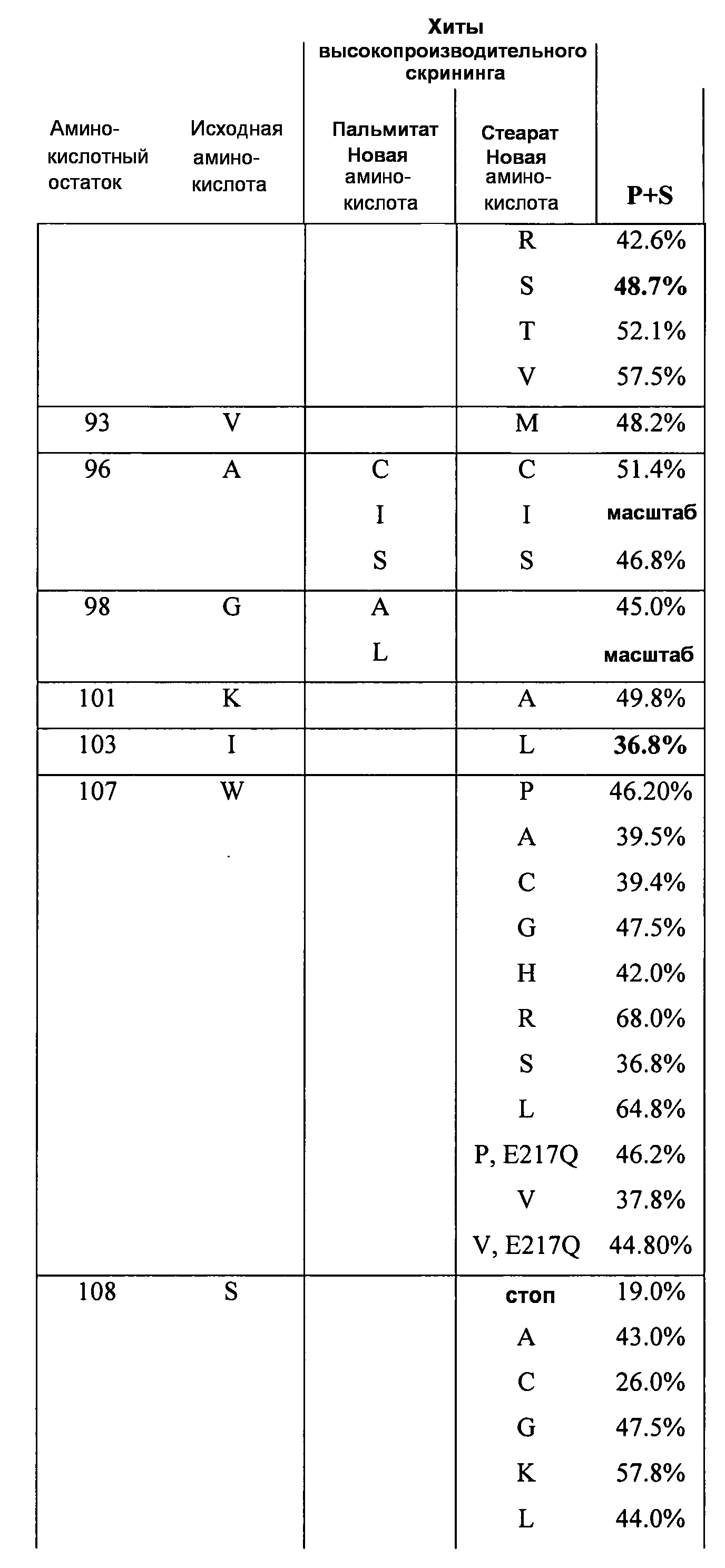
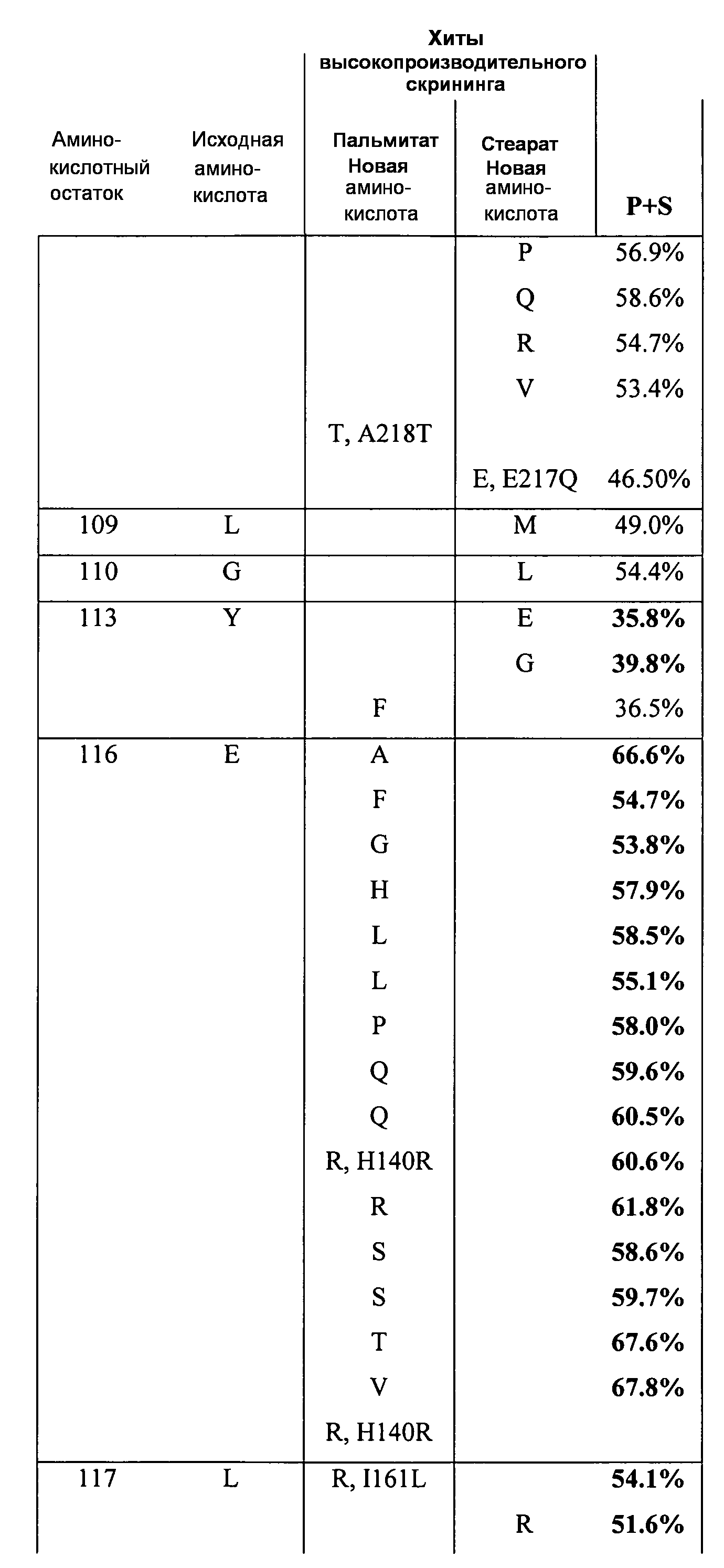

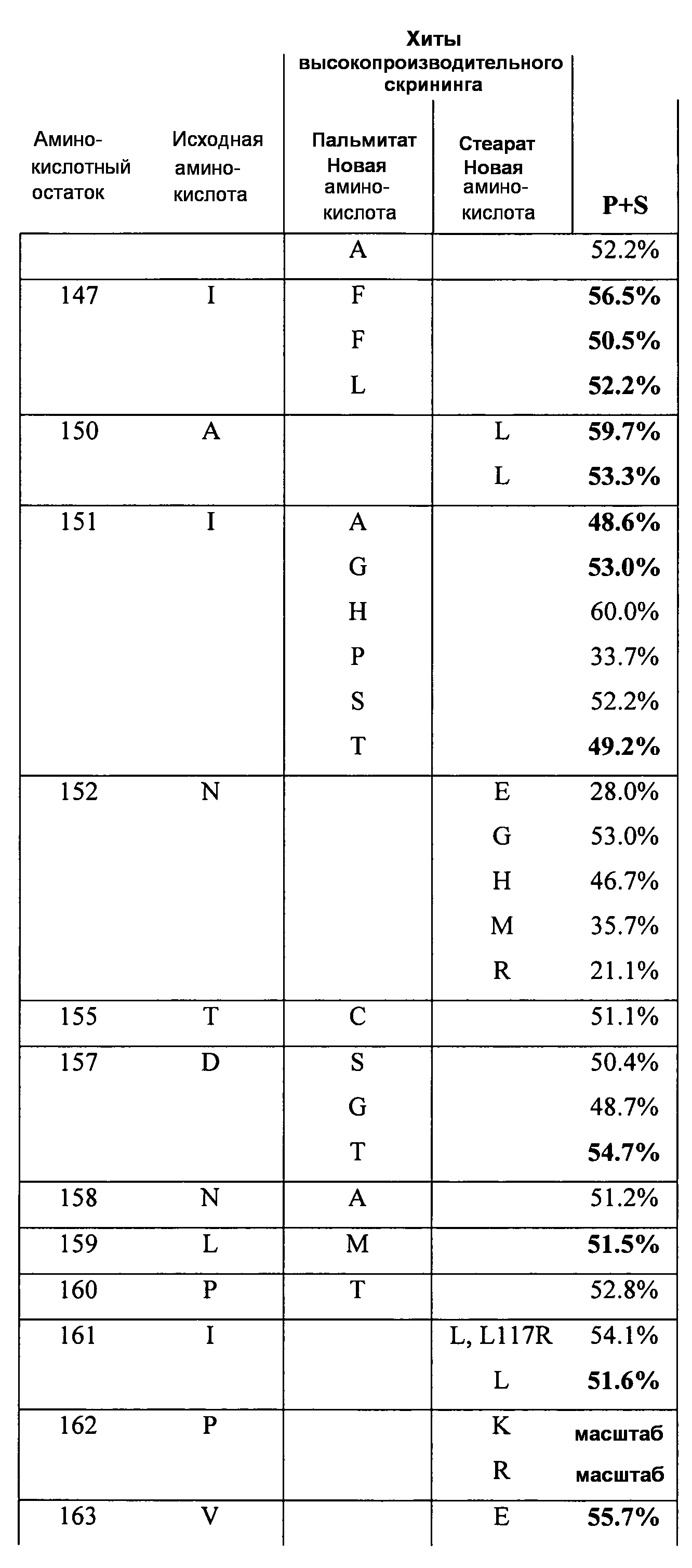

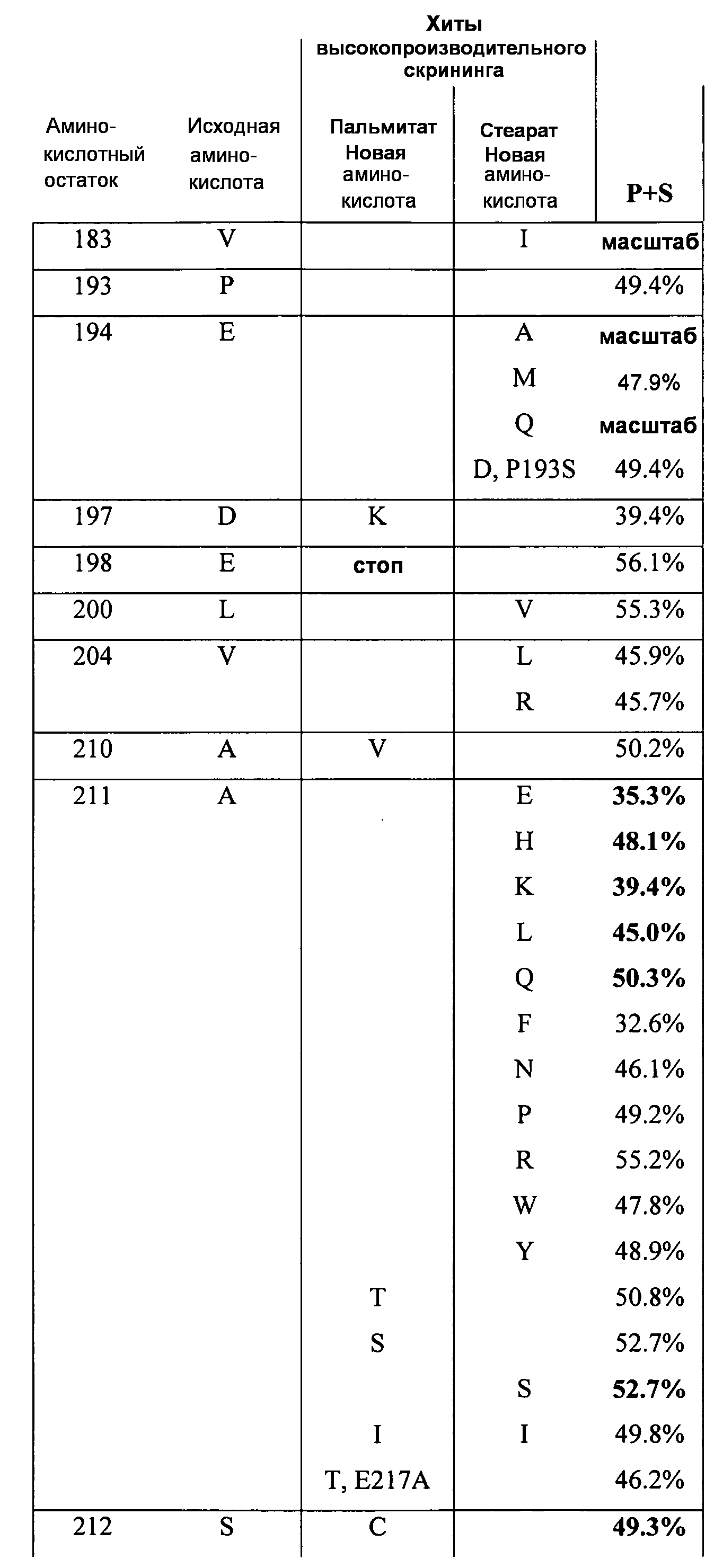




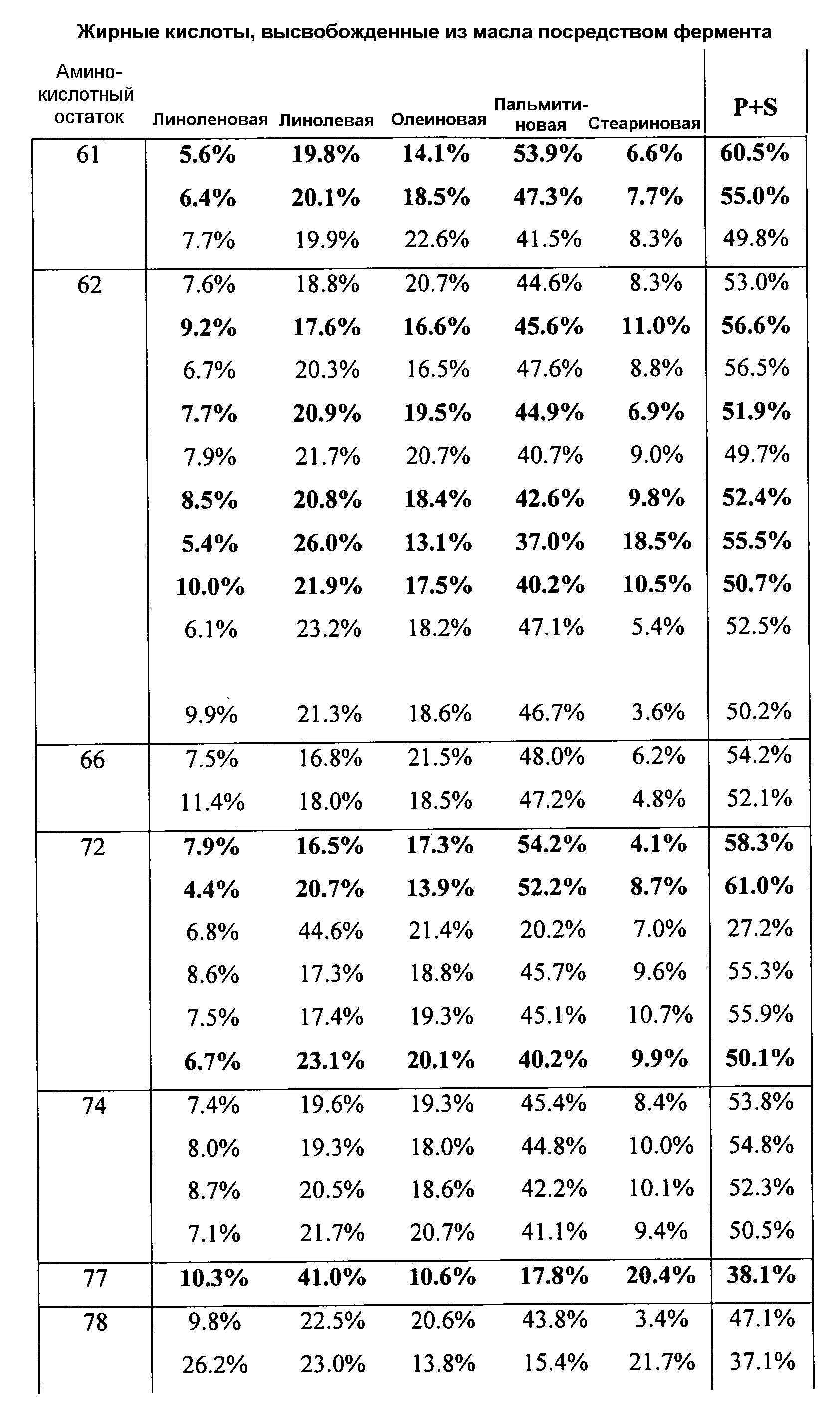
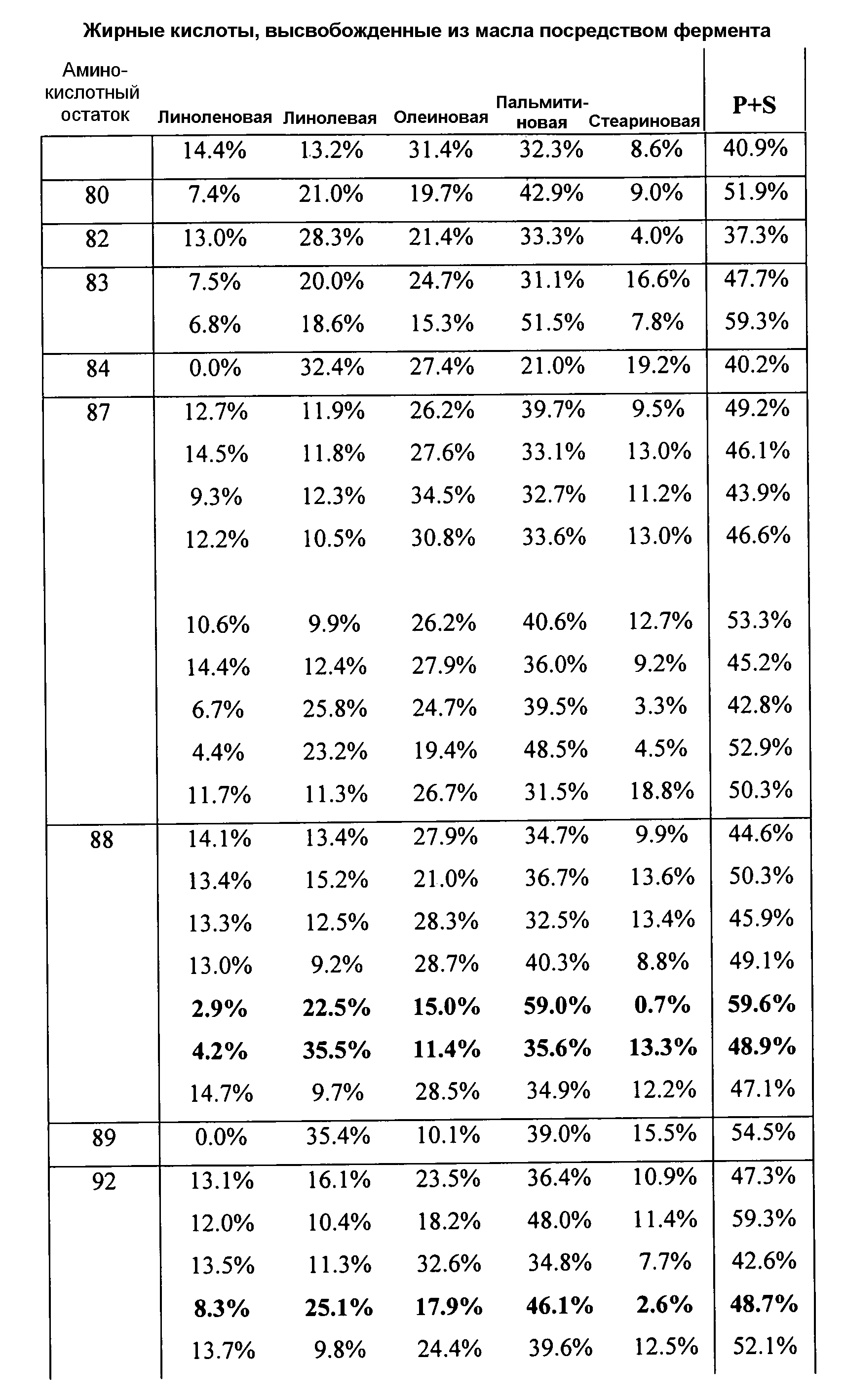
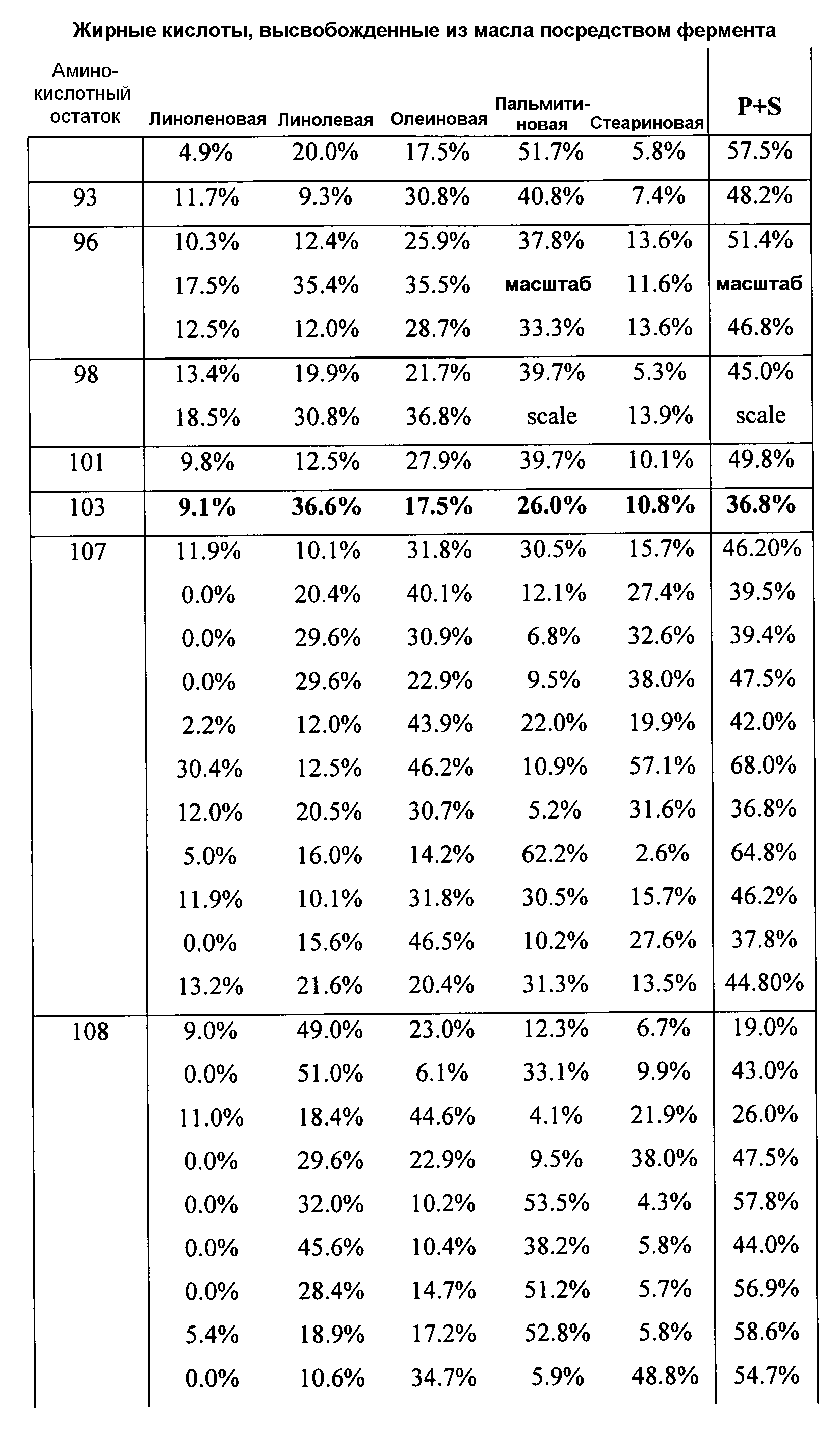

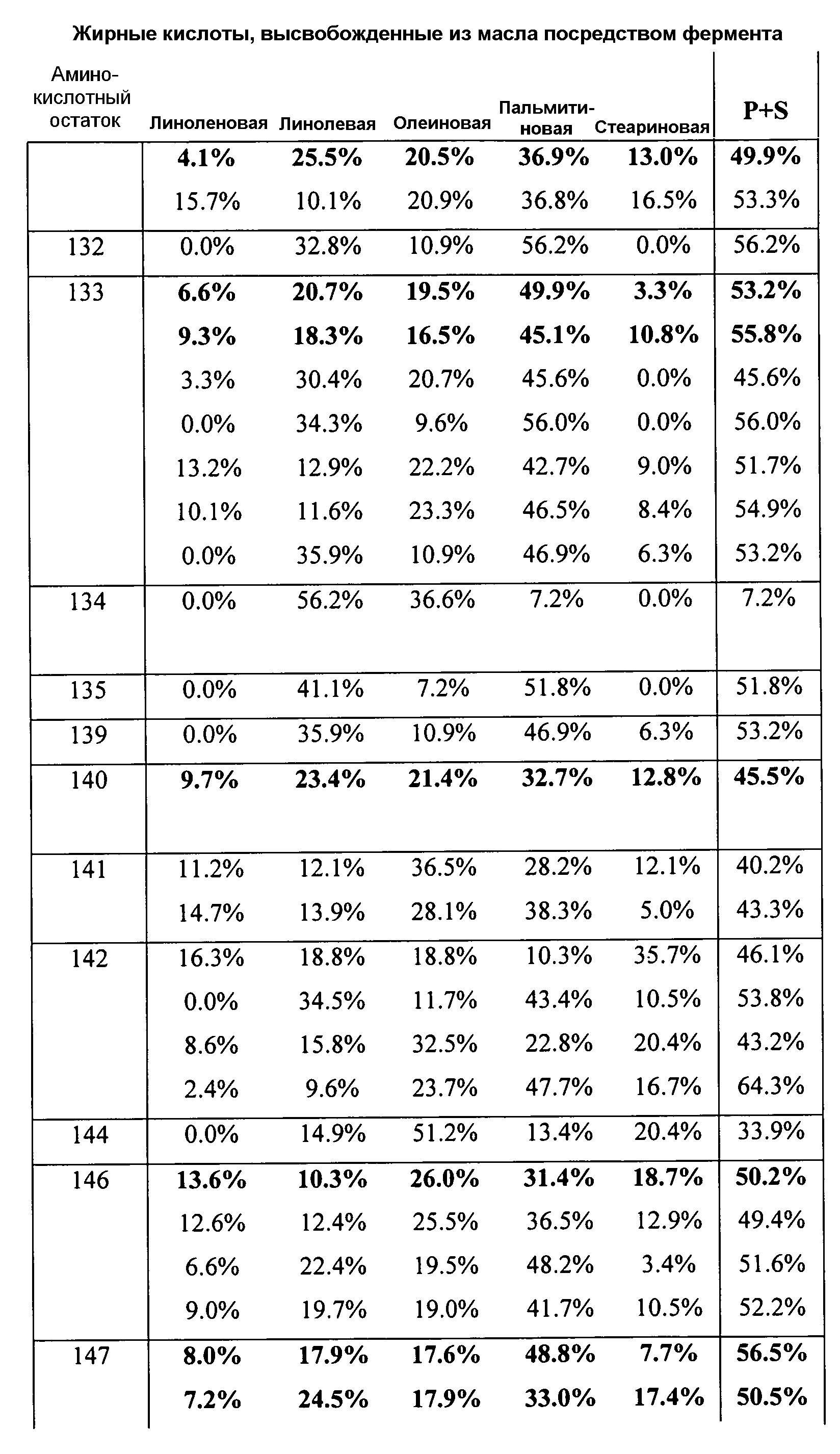

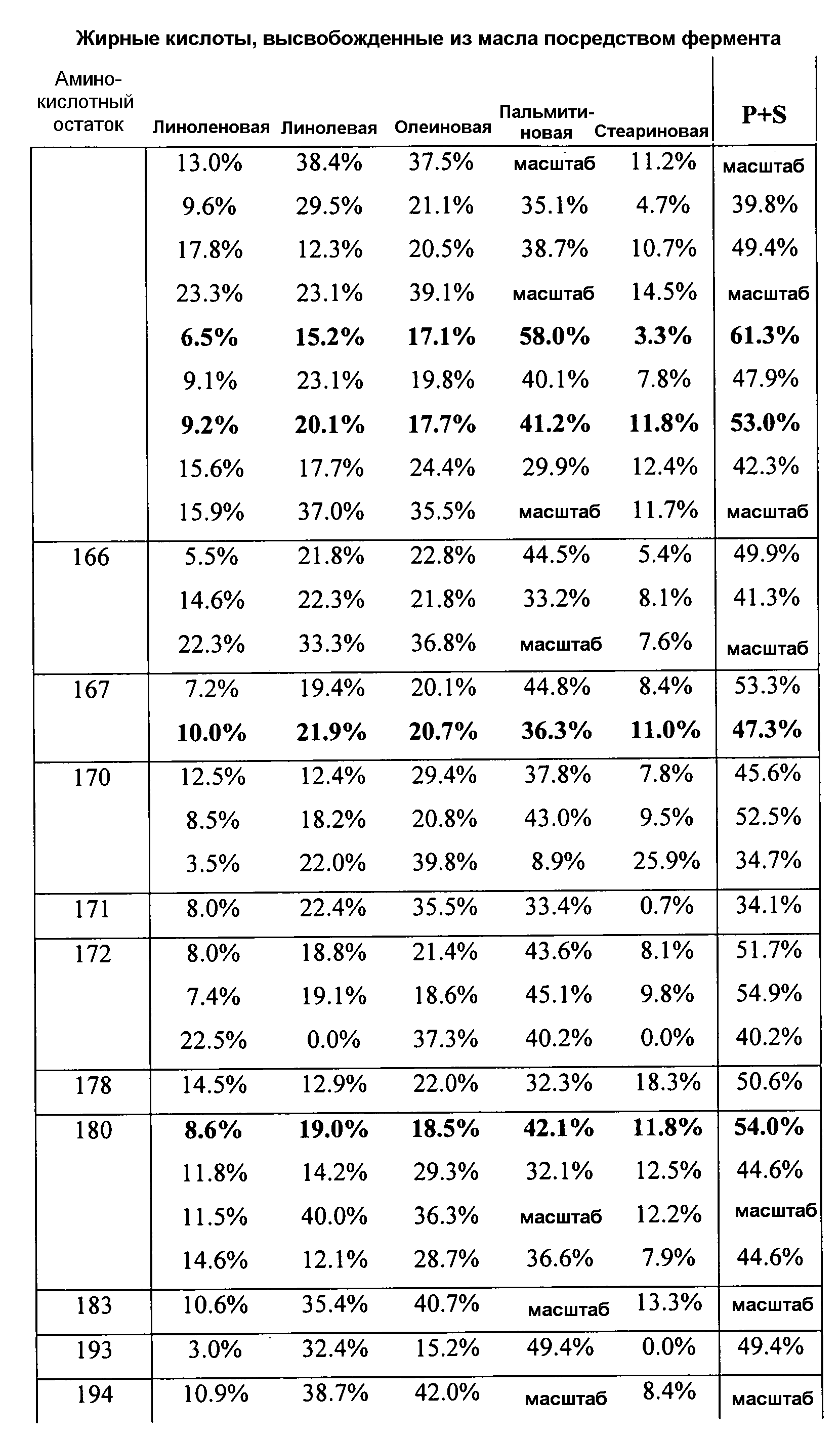


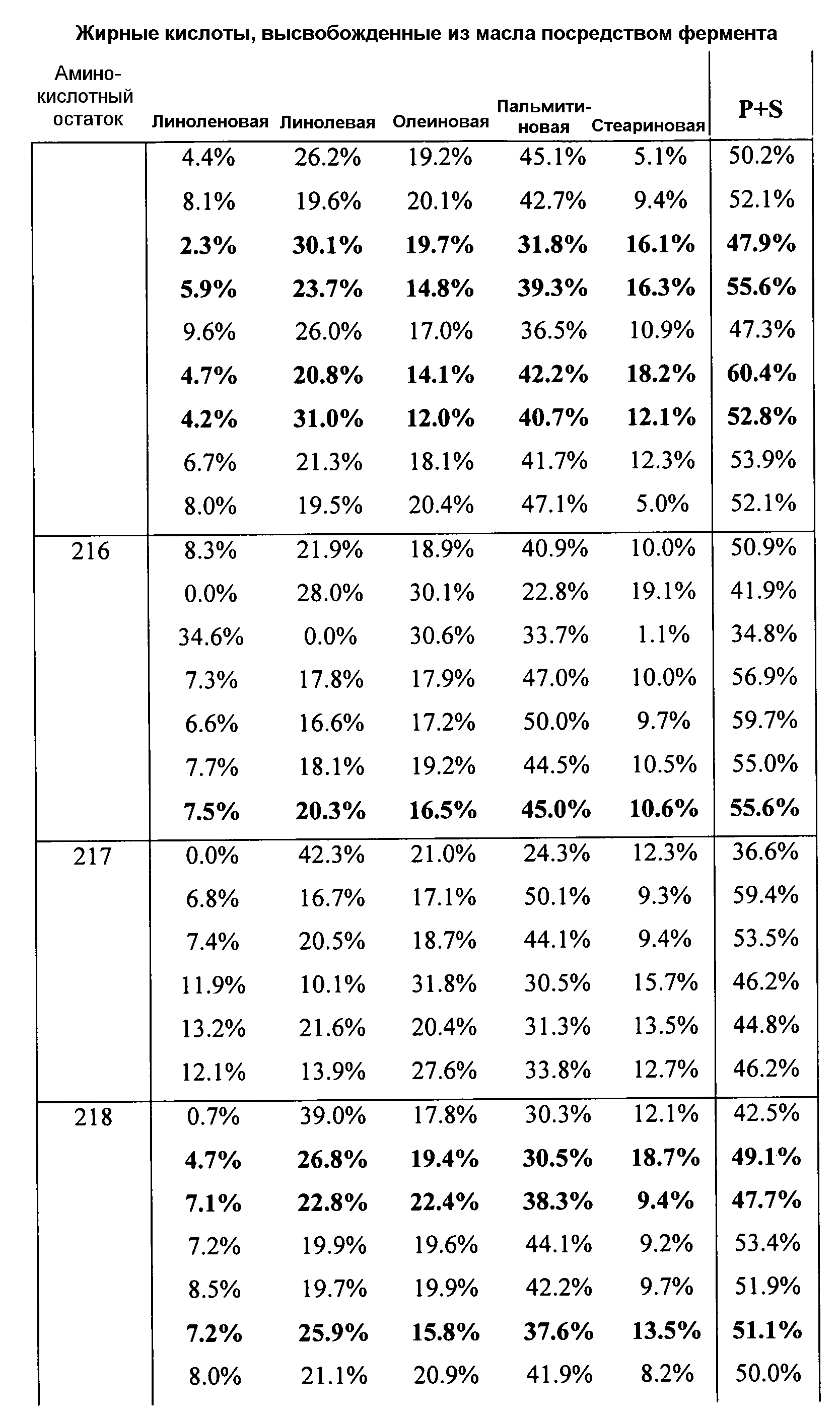
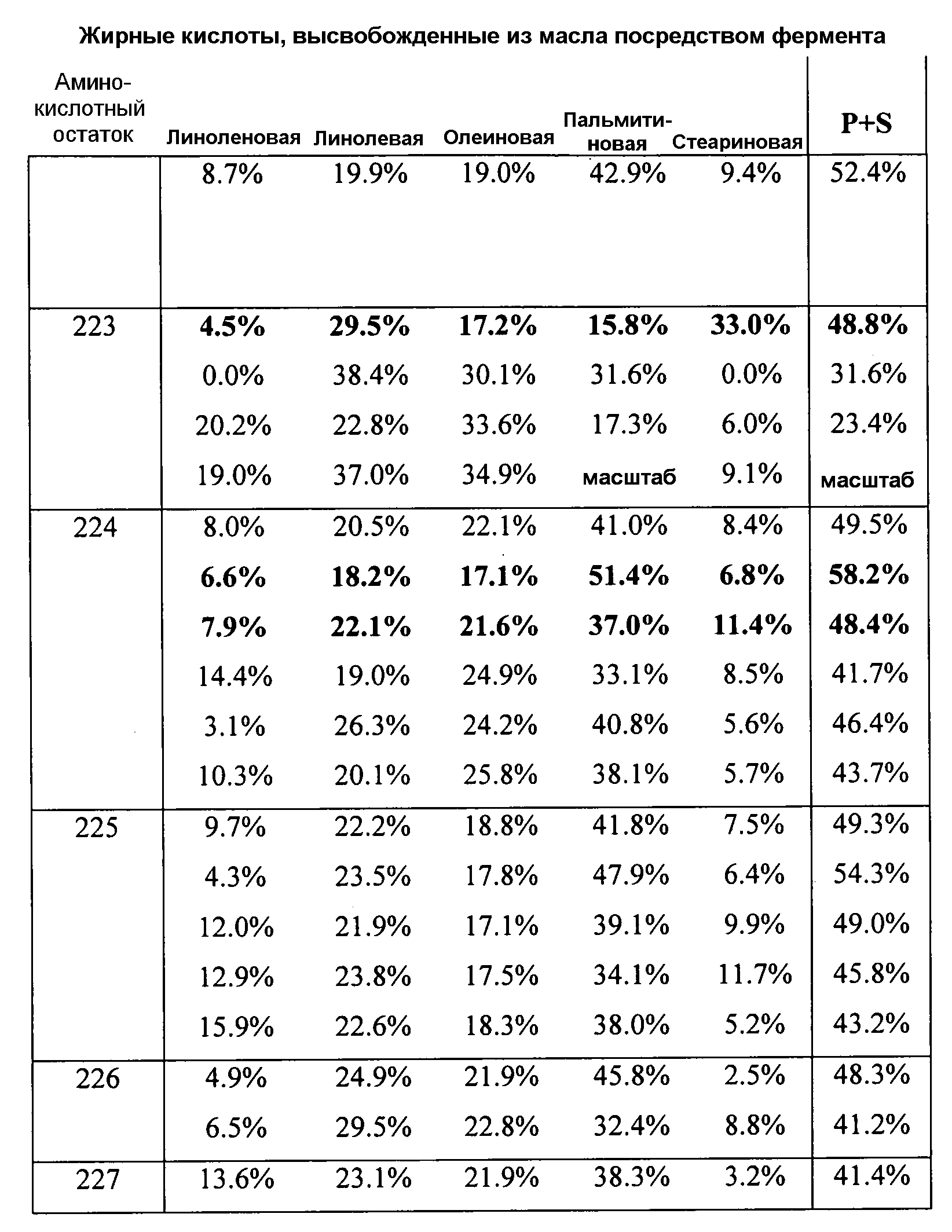
Таблица 4 представляет собой краткое изложение или дополнительную компиляцию данных, приведенных в таблице 3 (выше). Например, термин «положение» обозначает положение аминокислотного остатка в SEQ ID № 2; термин «Исходная аминокислота», как и в таблице 3, обозначает неизмененный «исходный» остаток, тогда как термин «Новая аминокислота», как и в таблице 3, обозначает измененный (новый) аминокислотный остаток в этом положении. Термины «WT_P» и «WT_S» обозначают субстратные (высвобождение жирных кислот) предпочтения «исходного» фермента, например, SEQ ID № 2, для конкретного субстрата (жирной кислоты), отражая количество высвобожденной (гидролитически отщепленной) жирной кислоты из соевого масла (как и в таблице 3), где «P» обозначает пальмитиновую кислоту, а «S» обозначает стеариновую кислоту.
В колонках «пальмитат» и «стеарат» показано количество пальмитиновой кислоты и стеариновой кислоты, высвобожденной (путем ферментативного гидролиза) из соевого масла, которое содержит линоленовую кислоту, линолевую кислоту, олеиновую кислоту, пальмитиновую кислоту, стеариновую кислоту, как указано выше. В «P+S» приведены объединенные количества гидролитически отщепленных жирных кислот пальмитиновая кислота и стеариновая кислота, или «P+S». Термины «delta_P» и «delta_S» обозначают изменение в предпочтении образцового фермента по изобретению (например, D61A из первой строки) относительно гидролитического отщепления пальмитиновой кислоты и стеариновой кислоты, соответственно, по сравнению с соответствующей активностью SEQ ID № 2. Термин «delta P+S» обозначает общее или суммарное изменение предпочтения образцового фермента по изобретению (например, D61A из первой строки) относительно гидролитического отщепления пальмитиновой кислоты и стеариновой кислоты по сравнению с соответствующей активностью SEQ ID № 2. В разделе «пальмитатные мутации» вкратце описаны образцовые ферменты по изобретению, обладающие предпочтительной активностью (гидролитическое отщепление жирных кислот) в отношении высвобождения пальмитиновой кислоты в сравнении с другими жирными кислотами. В разделе «стеаратные мутации» вкратце описан образцовый фермент по изобретению, обладающий предпочтительной активностью в отношении высвобождения стеариновой кислоты в сравнении с другими жирными кислотами (из соевого масла, анализ описан выше).
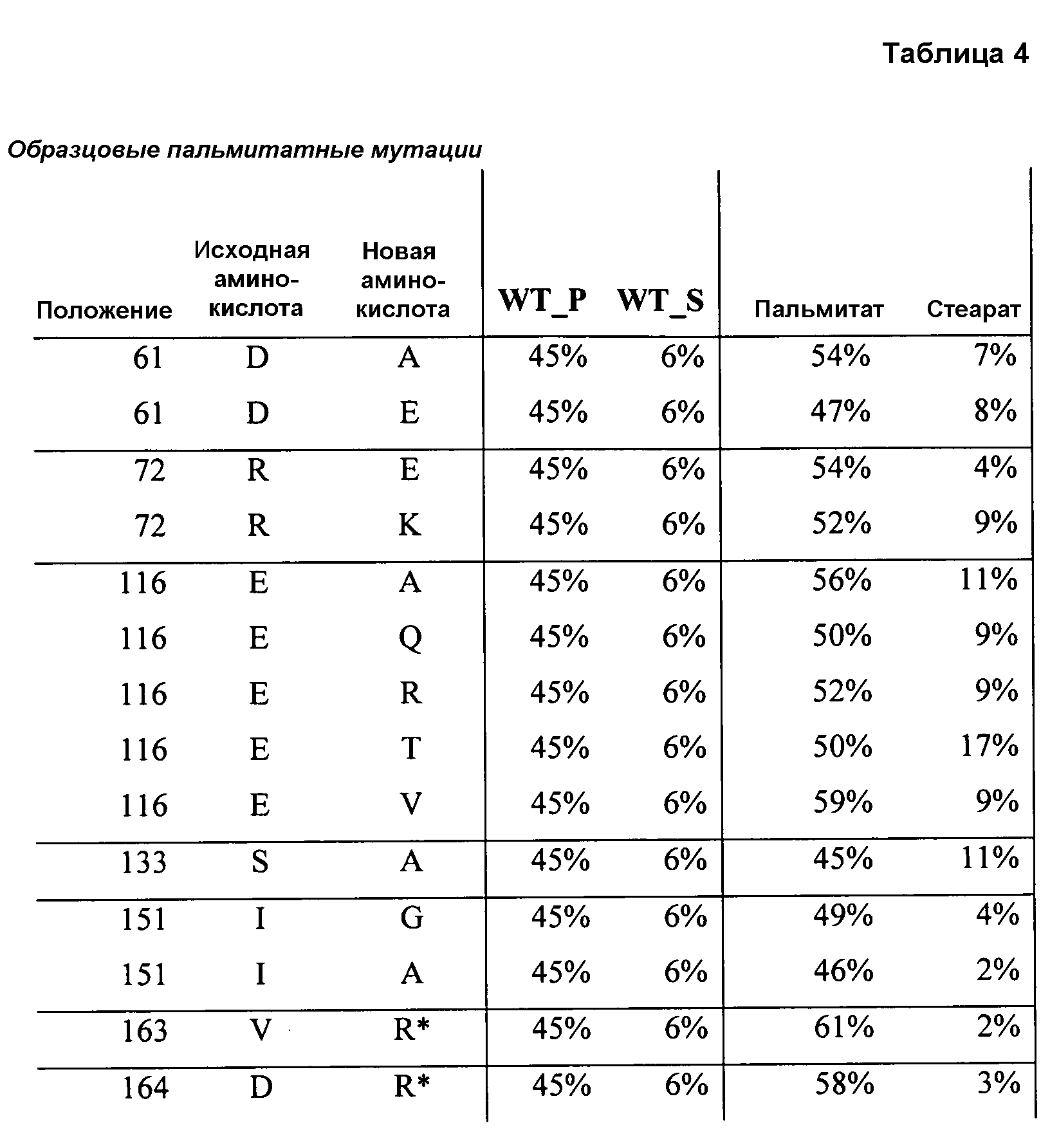
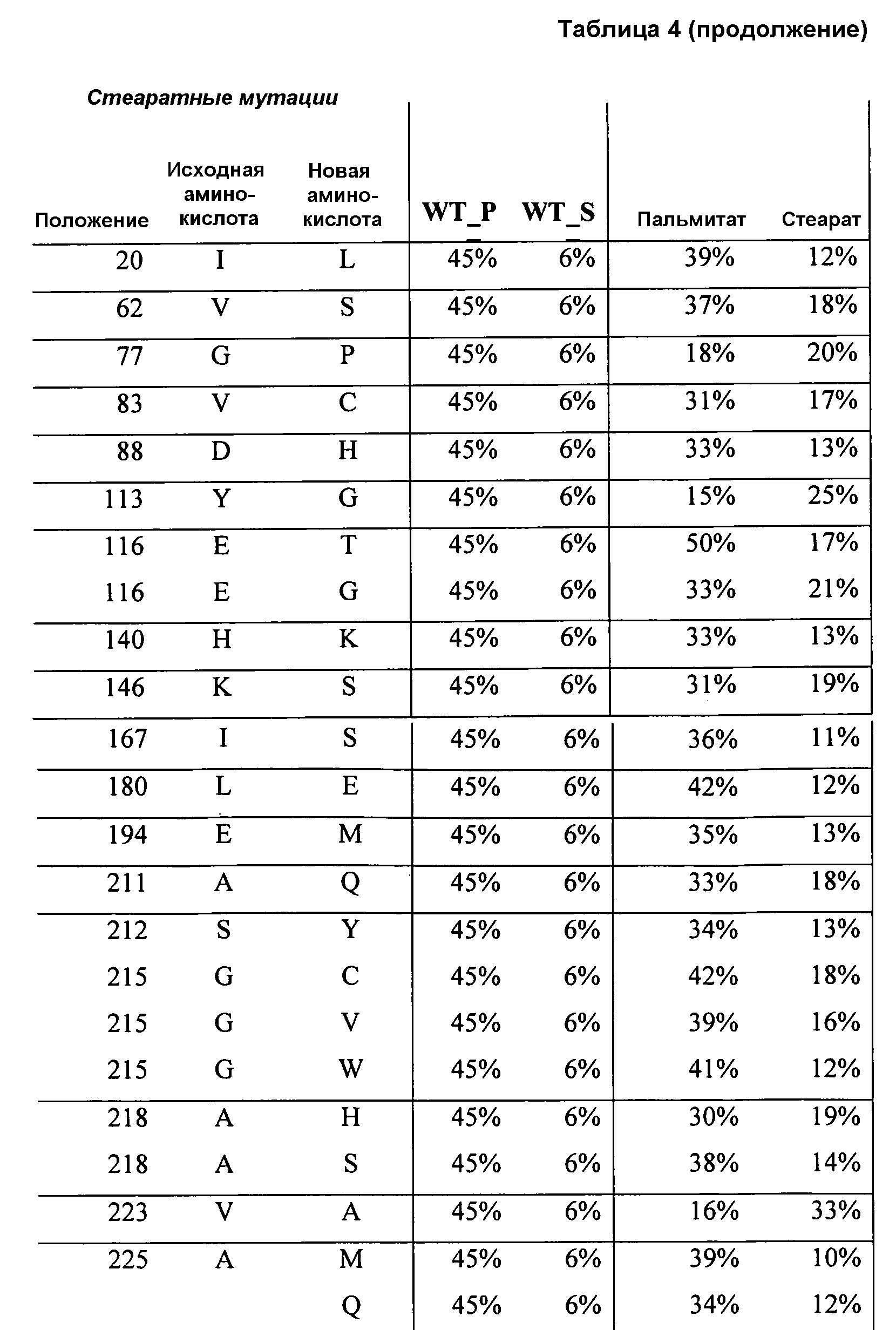
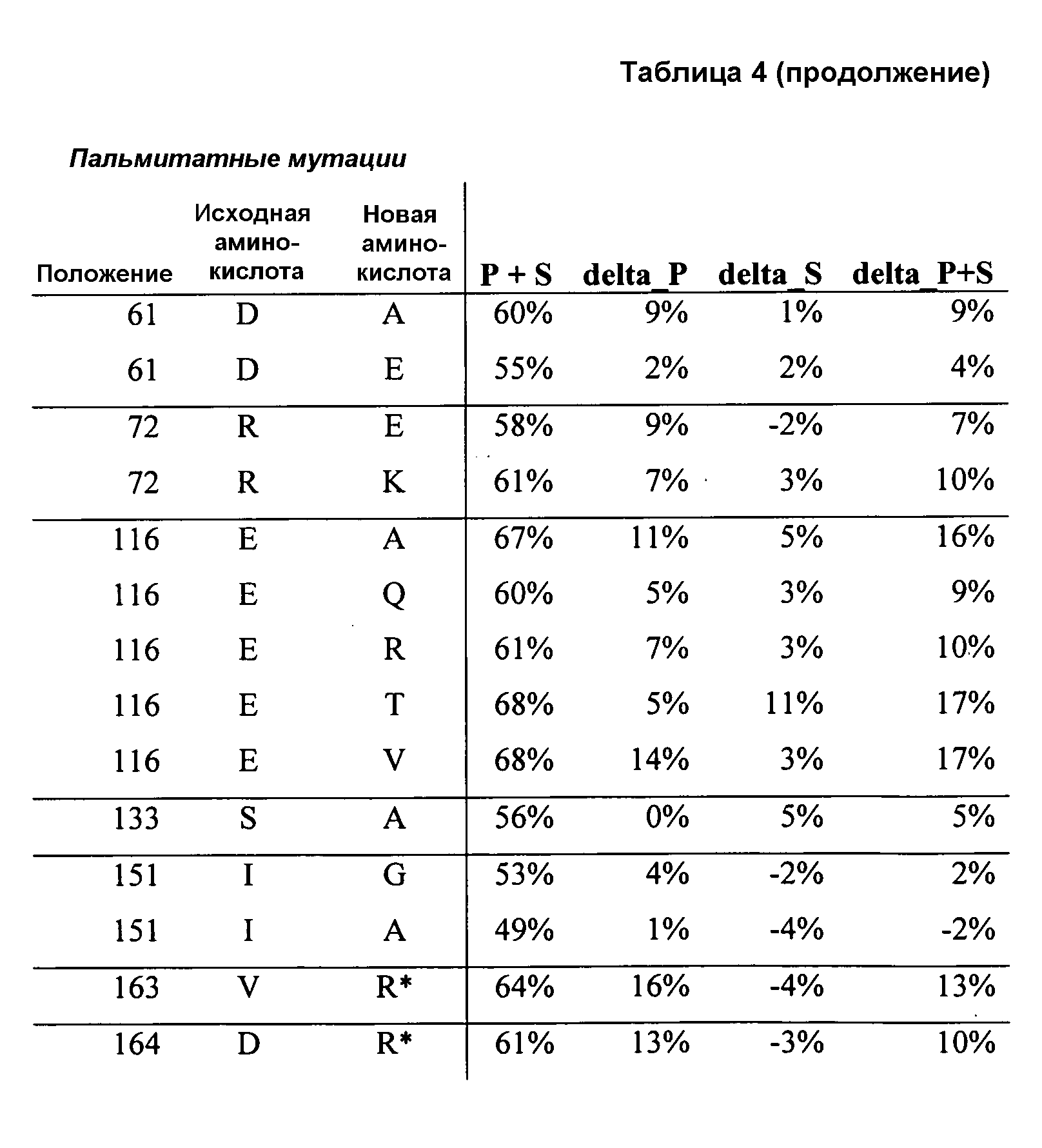

Пример 5: образцовая эволюция для усовершенствования гидролиза пальмитата с использованием технологии GeneReassemblySM
Четырнадцать (14) отдельных мутаций аминокислот, идентифицированных в скрининге GSSM, которые покрывают семь (7) положений аминокислот, комбинировали посредством технологии GeneReassemblySM (патент США № 6605449). Полноразмерные последовательности нуклеиновых кислот, созданные в фазе GeneReassembly, клонировали в экспрессирующий вектор pASK-5 (см. описание выше) для экспрессии в организме-хозяине Escherichia coli HMS175 (Novagen, USA). Экспрессию вариантов GeneReassembly индуцировали ангидротетрациклином после достижения оптимальных плотностей клеток-хозяев.
14 мутаций, которые давали наибольшие увеличения гидролиза пальмитата, идентифицированные в таблице 2, отбирали для включения в библиотеку пальмитаз GeneReassembly, созданную способами, описанными выше. Скрининг исходных клонов с умбеллиферилпальмитатом на активность дал приблизительно 145 клонов с уникальными последовательностями, которые анализировали на активность на соевом масле, как описано выше.
На фиг. 8 показаны данные первичного и вторичного скрининга для анализов на соевом масле для отобранных клонов из библиотеки пальмитаз. Клоны, которые давали пальмитат в количестве более 70% гидролитически отщепленных ЖК в первичном анализе (в условиях стандартного исходного уровня для способа анализа), отбирали для повторного анализа на соевом масле. Для каждого анализа на соевом масле, экстрагированные ЖК разбавляли в 50 и 100 раз для анализа посредством LCMS или ГХ. Если обнаруживали дополнительные, нецелевые мутации, это также обозначено. Представлены установленные степени гидролиза ЖК и количества каждой ЖК. На чертеже «высокое» и «низкое» обозначают величины, которые выходили за пределы калибровочной кривой. Строки отсортированы по процентной доле высвобожденного пальмитата во вторичном анализе, а затем по общему высвобожденному пальмитату. Множество клонов, показавших значительное увеличение избирательности к пальмитату (вплоть до 100%), сравнивали с родительской SEQ ID № 2 (61,2%).
Отобранные на основе вторичного анализа, описанного выше, лучше 25 хитов пальмитаз субклонировали в системы Pseudomonas (Dow Global Technologies Inc., публикация заявки на патент США № 20050130160, и Dow Global Technologies Inc., публикация заявки на патент США № 20050186666). Последовательность нуклеиновой кислоты, кодирующую фермент или полипептид, встраивали или в вектор pMYC (Dow Global Technologies Inc., публикация заявки на патент США № 20050130160) или в вектор pDOW1169 (Dow Global Technologies Inc., публикация заявки на патент США № 20080058262) и затем вводили в организм-хозяин Pseudomonas fluorescens посредством электропорации. трансформированные клетки отбирали или посредством роста на минимальной среде в случае с конструкциями pDOW1169 или на насыщенной среде с тетрациклином в случае с конструкциями pMYC. Экспрессию фермента или полипептида индуцировали, используя IPTG, после достижения оптимальных плотностей клеток-хозяев.
В таблице 5 приведены данные из анализов на соевом масле, проведенных дважды, для лучших 25 хитов, экспрессированных в системах Pseudomonas. 4 хита, встроенные в вектор pDOW1169, приведены в жирном подчеркнутом начертании, все другие хиты встраивали в вектор pMYC. К 5 г нерафинированного масла добавляли фермент, в результате чего конечное содержание воды составляло 20%. Затем смесь гомогенизировали с использованием 7 мм зонда и инкубировали в течение 40 часов при 25°C, помешивая с помощью мешалки. Брали аликвоты и проводили анализ ЖК посредством превращения ЖК в СМЭЖК и количественного определения СМЭЖК посредством ГХ, как описано в примере 8. 25 ферментов загружали в 5 г соевого масла, исходя из равного количества единиц активности в отношении УМБ-пальмитата. В этих реакциях содержание пальмитата в масле значительно снизилось с 11% в необработанном масле до 5% или менее в обработанных ферментами маслах, что указывает на увеличенное предпочтение относительно гидролиза пальмитата по сравнению с родительским ферментом SEQ ID № 2.
Ниже в таблице 6 приведены данные о термостабильности лучших 25 хитов пальмитаз, отобранных на основе вторичного анализа, описанного выше. Эти данные получены с использованием хитов, экспрессированных в организме-хозяине E. coli HMS174. Клоны помещали в 96-луночные планшеты и инкубировали в течение 10 минут при комнатной температуре (КТ), 45, 50 или 55°C, а затем проводили анализ с метилУМБ-пальмитатом при КТ. Процентную долю остаточной активности определяли посредством деления активности после инкубации при каждой температуре на активность после инкубации при КТ. Также для каждой пальмитазы приведены присутствующие мутации и примеры избирательности и активности в отношении пальмитата в соевом масле. SEQ ID № 2 сохраняла приблизительно 15% активности после инкубации в течение 10 мин при 50°C, но не обладала активностью после инкубации при 55°C.
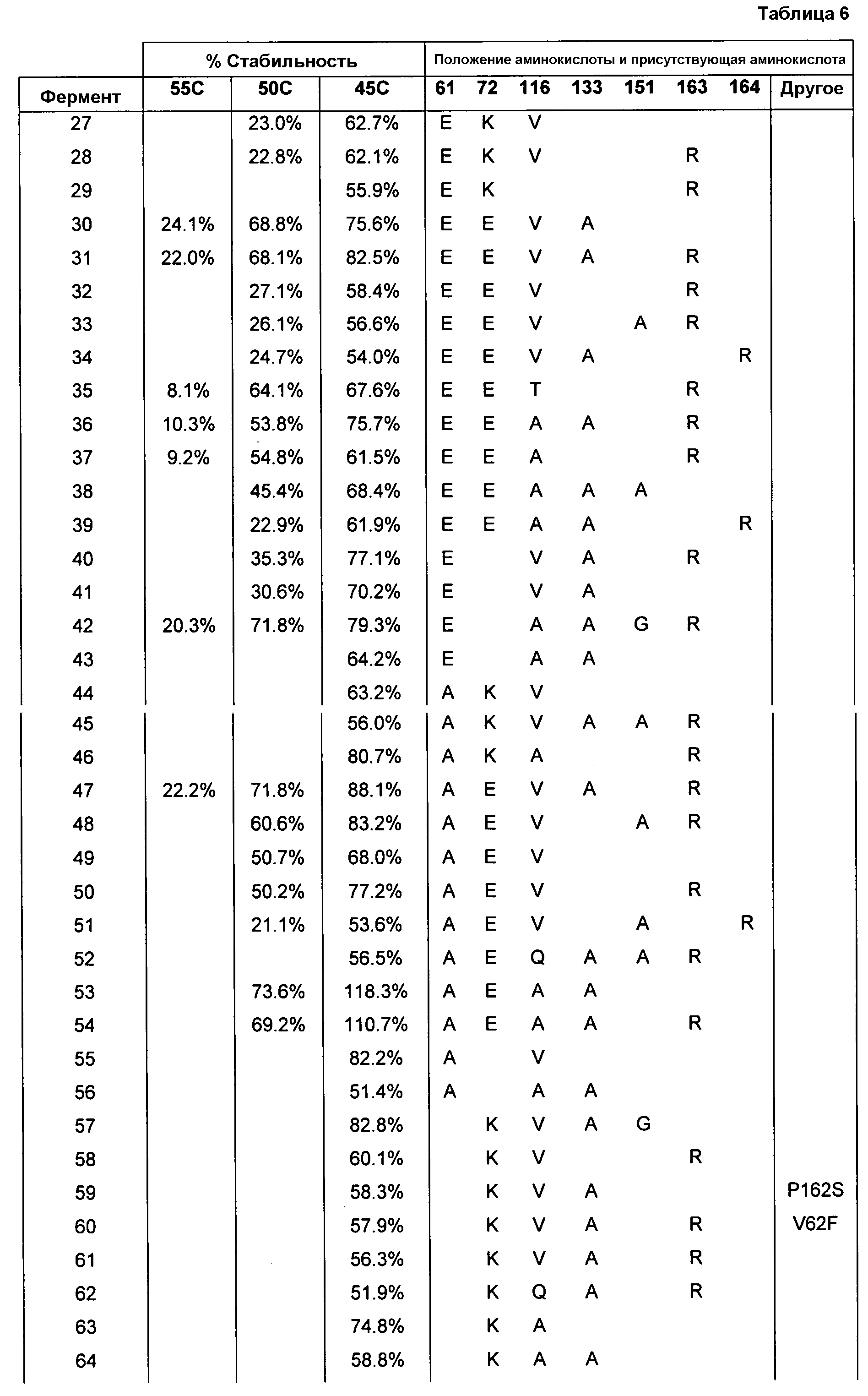
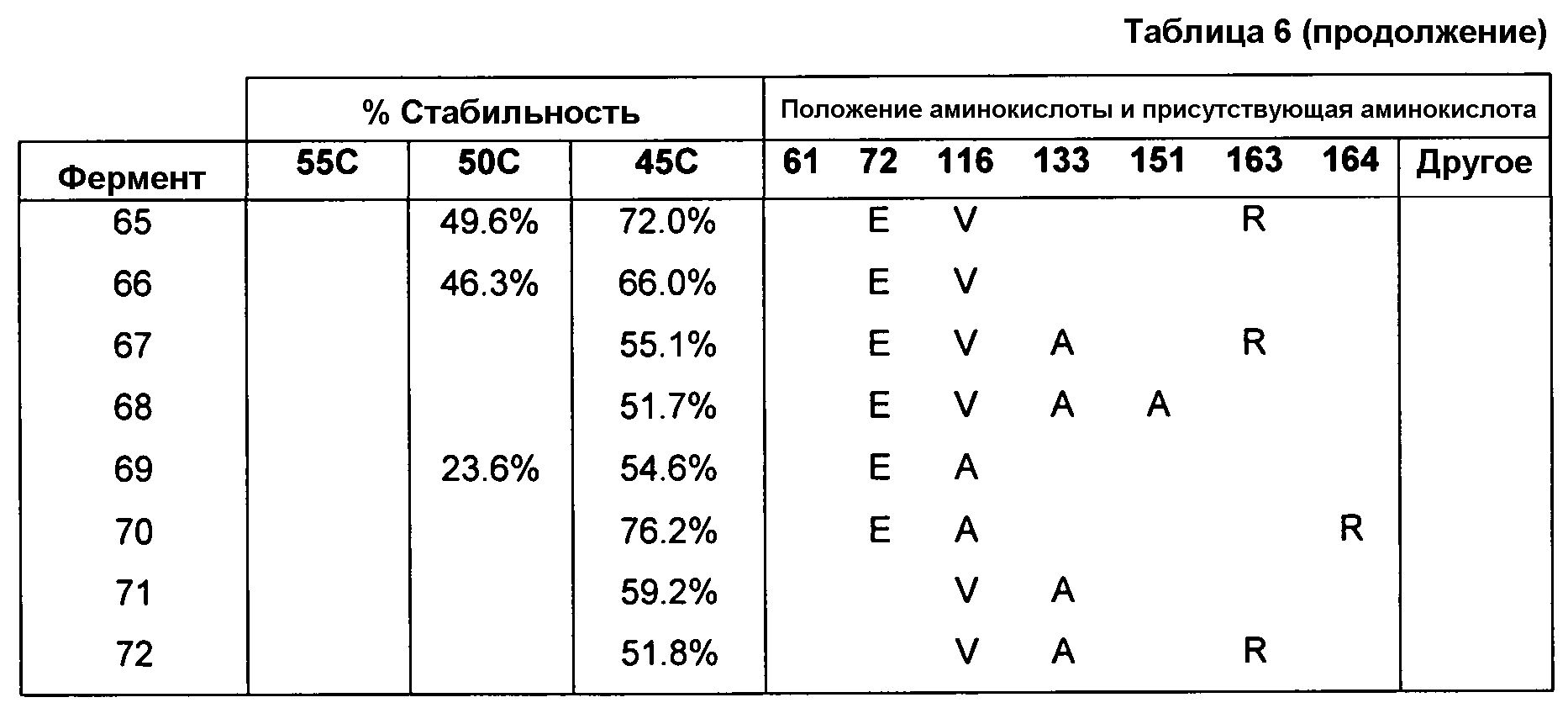
Пример 6: Лабораторный протокол для оценки подходящих ферментов пальмитаз, стеаратаз или сатураз
Образцовые ферменты и предоставленные в настоящем документе полипептид экспрессировали в системе Pseudomonas (Dow Global Technologies Inc., публикация заявки на патент США № 20050130160). Нуклеиновую кислоту, кодирующую фермент или полипептид, встраивали в вектор pMYC (Dow Global Technologies Inc., публикация заявки на патент США № 20050130160) и затем вводили в ауксотрофный организм-хозяин Pseudomonas fluorescens посредством электропорации. Трансформированные клетки отбирали выращиванием на минимальной среде. Экспрессию фермента или полипептида индуцировали, используя IPTG, после достижения оптимальных плотностей клеток-хозяев.
Следующую процедуру следует использовать для оценки способности фермента или другого полипептида по изобретению, гидролизовать образец масла. В 1 кг нерафинированного масла добавляли фермент пальмитазу, в результате чего конечное содержание воды составляло 20%. Затем смесь гомогенизировали, используя навесной смеситель, и инкубировали при комнатной температуре при постоянном перемешивании с использованием лопаточного смесителя. Брали аликвоты (0,5 мл) в моменты времени 0 ч, 21 ч, 43 ч, 65 ч и 72 ч и обрабатывали для превращения в СМЭЖК и ГХ анализа, как описано в примере 8.
В приведенной выше процедуре использовали SEQ ID № 2, а образец масла представлял собой нерафинированное соевое масло. Через 72 ч с использованием образцов необработанного масла и обработанного ферментом масла получали результаты, приведенные в таблице 7.
Результаты показывают значительное снижение количества пальмитиновой кислоты (C16:0), и такое снижение рассматривали как желаемое.
Пример 7: Оценка липаз, сатураз или пальмитаз, содержащих последовательности, гомологичные образцовому полипептиду SEQ ID № 2
Несколько гомологичных последовательностей липаз субклонировали в вектор pMAL-c2x (New England Biolabs, USA) способом xi-клонирования (Genlantis, USA). Конструкции, содержащие SEQ ID № 2, SEQ ID № 6, SEQ ID № 14 или SEQ ID № 16, трансформировали в организм-хозяин Escherichia coli ArcticExpress RP (Stratagene, USA) для экспрессии. Липазы под управлением промотора, который индуцировали с использованием IPTG, экспрессировали после достижения оптимальных плотностей клеток-хозяев. Рекомбинантные ферменты тестировали на соевом масле на избирательность к ЖК (таблица 8). Липазы, содержащие SEQ ID № 2, SEQ ID № 6, SEQ ID № 14, или SEQ ID № 16, экспрессировали и от метки слияния MBP с использованием стандартных условий. В среду LB, содержащую гентамицин 20 мкг/мл, инокулировали одну колонию и встряхивали со скоростью 200 об/мин в течение ночи при 30°C. Эту ночную культуру инокулировали в свежую среду LB, содержащую гентамицин 20 мкг/мл, до получения значения OD600, равного 0,05. Эту культуру встряхивали со скоростью 200 об/мин и при температуре 30°C до получения значения OD600, равного 0,5. Культуры переносили в 12°C, встряхивали со скоростью 200 об/мин и позволяли прийти в равновесие при пониженной температуре прежде, чем индуцировать экспрессию липазы путем добавления 0,5 мМ IPTG, за которым следовал дополнительный рост в течение 24 часов. Клетки собирали центрифугированием, суспендировали в буфере Tris pH 8, содержащем NaCl, CaCl2, ДНКазу I и лизоцим и затем лизировали, используя обработку ультразвуком. Клеточные лизаты осветляли посредством центрифугирования. Ферменты отщепляли от MBP посредством инкубации слитого белка липаза-MBP с фактором Xa в течение 6 часов при комнатной температуре, за которыми следовали дополнительные 18 часов при 12°C. Все осветленные лизаты с интактными активными рекомбинантными ферментами проявляли выраженные и схожие предпочтения относительно гидролиза пальмитата по отношению к другим ЖК при анализе на соевом масле (таблица 8).
Пример 8: Способ превращения свободных жирных кислот или триглицеридов в сложные метиловые эфиры жирных кислот (СМЭЖК) и количественное определение СМЭЖК газовой хроматографией.
Определить количество жирных кислот, высвобожденных из липидов, триглицеридов, жиров или масел под действием липаз, например, сатураз, пальмитаз и/или стеаратаз, можно напрямую посредством LCMS с использованием способа, описанного в примере 2. Альтернативно эти гидролитически отщепленные жирные кислоты можно превратить в сложные метиловые эфиры жирных кислот (СМЭЖК) с использованием катализируемого кислотой метанолиза, а затем определить количеством посредством газовой хроматографии (ГХ). В этом примере:
• Масло после реакции с липазами, например, сатуразами, пальмитазами и/или стеаратазами, обрабатывали посредством добавления 1 мл растворителя для экстракции (CHCl3:MeOH:4N HCl (2:1:0,075)) на 0,5 мл реакционного объема.
• Аликвоту экстрагированного масла 45 мкл переносили в 4 мл сосуд с винтовой крышкой. В каждый сосуд добавляли маленькую мешалку, а затем 2 мл гексана и 400 мкл 20% (об./об.) MeOH в HCl.
• Затем сосуды закупоривали и нагревали при перемешивании в течение 15 минут. Затем прекращали нагревание сосудов и оставляли их остывать перед добавлением 800 мкл H2O.
• Затем смесь перемешивали на вортексе и образец (500 мкл) верхнего гексанового слоя, содержащего СМЭЖК, переносили в сосуд автодозатора для ГХ. В каждый 500 мкл образец из 0,5 мг/мл C15:0 СМЭЖК добавляли в качестве внутреннего стандарта.
Затем СМЭЖК, синтезированные с использованием этого способа, анализировали посредством газовой хроматографии с использованием следующих эксплуатационных параметров:
• Оборудование Hewlett Packard 6890 Series GC с автодозатором
• Использовали колонку Supelco SP-2380 Fused Silica Capillary Column 30 м × 0,25 мм и толщина пленки 0,2 мкм
• Температуру инжектора и детектора устанавливали равной 260°C; поток несущего газа гелия устанавливали равным 0,6 мл/мин; исходную температуру печи устанавливали равной 150°C.
• Образцы (1 мл) впрыскивали с разделением потока 10:1. Использованный способ ГХ содержит:
- Скорость изменения температуры 1: 4°C/мин в течение 10 мин = 190°C
- Скорость изменения температуры 2: 15°C/мин в течение 4 мин = 250°C
- Удержание: 250°C в течение 2 мин
ЖК триглицеридов также можно анализировать посредством превращения СМЭЖК, даже в присутствие гидролитически отщепленных жирных кислот. Используя способ, приведенный выше, в сочетании со способом, приведенным ниже, это можно использовать для определения избирательности липазы, например, сатуразы, пальмитазы и/или стеаратазы, к жирным кислотам и действия фермента на масло. В способе анализа ЖК, связанных с глицерином (или с другими спиртами), используют катализируемый основанием метанолиз:
• Масло после реакции с липазами, например, сатуразами, пальмитазами и/или стеаратазами, обрабатывали посредством добавления 1 мл растворителя для экстракции (CHCl3:MeOH:4N HCl (2:1:0,075)) на 0,5 мл реакционного объема.
• Аликвоту экстрагированного масла 45 мкл переносили в пробирку для микроцентрифуги. Добавляли 500 мкл гептана, за которыми следовало 50 мкл 2N метанольной KOH.
• Смесь энергично перемешивали на вортексе в течение 30 секунд, а затем центрифугировали.
• Аликвоту (50 мкл) верхнего гептанового слоя, содержащего СМЭЖК, переносили в сосуд автодозатора и объединяли ее с 450 мкл гексана, содержащего внутренний стандарт C15:0.
• Анализ СМЭЖК посредством ГХ изложен выше.
Пример 9: Образцовая эволюция для усовершенствования тепловой устойчивости пальмитазы с использованием технологии GSSMSM
Образцовую пальмитазу по изобретению, которая содержит мутации D61E, R72K и V163R в SEQ ID № 2 (фермент 17, таблица 5, выше), выбрали в качестве лидирующего по избирательности мутанта из предыдущих раундов эволюции (примеры 4 и 5, выше). Проводили дополнительную эволюцию для усовершенствования тепловой устойчивости ведущего по избирательности фермента (SEQ ID № 2 с мутациями D61E, R72K и V163R) с использованием технологии GSSM (см., например, патент США № 6171820).
Вкратце, эволюцию GSSMосуществляют посредством введения точечных мутаций с использованием вырожденных олигонуклеотидов в одном положении аминокислоты за один раз с тем, чтобы можно было заменить каждый исходный кодон на каждую из 20 кодируемых природных аминокислот. Библиотеку конструировали в векторе pDOW-Kan, анализ проводили в агарозном геле, обрабатывали DPNI и затем трансформировали в компетентные клетки E. coli XL1-Blue. Колонии растили, собирали и секвенировали. Колонии объединяли, а ДНК получали с использованием набора Qiagen mini-prep (по каталогу № 27106, Qiagen, Valencia, CA).
Затем компетентные клетки Pseudomonas fluorescens трансформировали, используя ДНК. Организм-хозяин Pseudomonas fluorescens получали из Dow Global Technologies Inc. (публикация заявки на патент США № 20050130160, публикация заявки на патент США № 20050186666 и публикация заявки на патент США № 20060110747). Вектор pDOW-Kan конструировали посредством присоединения маркера устойчивости к канамицину к pDOW1169 (Dow Global Technologies Inc., публикация заявки на патент США № 20080058262). Клетки растили в минимальной среде M9 (Dow Global Technologies Inc., публикация заявки на патент США № 20050186666) с добавлением урацила и канамицина. Все примеры, которые соответствуют этому описанию, использовали вектор pDOW-kan, Pseudomonas fluorescens и среду M9, все относятся к тому же вектору pDOW-kan, Pseudomonas fluorescens и среде M9, описанным выше.
Скрининг библиотеки GSSM проводили в 384-луночном формате. Первичный и вторичный высокопроизводительный скрининг проводили с использованием УМБ-пальмитата (как описано выше в примере 3). Для того идентифицировать мутации, которые сохраняют активность фермента при повышенной температуре, проводили анализ посредством инкубации флуорогенного субстрата с полным клеточным лизатом при 54°C в течение 30 минут, которой предшествовала инкубация полного клеточного лизата с буфером в течение 30 минут при 54°C, чтобы гарантировать, что фермент достиг 54°C перед загрузкой субстрата. Измеряли флуоресценцию каждого мутанта (см. колонку 2 в таблице 9), и всех мутантов с показанием флуоресценции, превышающим контроль более чем на 2 стандартных отклонения, определяли в качестве «хитов» - то есть в качестве мутантов с достаточно увеличенной тепловой устойчивостью. В скрининге с УМБ-пальмитатом идентифицировали 117 хитов с уникальными аминокислотными заменами (ниже см. таблицу 9). Аминокислотные замены в 117 хитах с одиночными мутациями происходили в 50 остатках; 22% аминокислотных замен представленного белка. Дополнительно оценивали эффективность 117 хитов на нерафинированном масле в стандартном анализе с 5 г (см. пример 5) при 25°C и 45°C. Результаты приведены ниже в таблице 10.

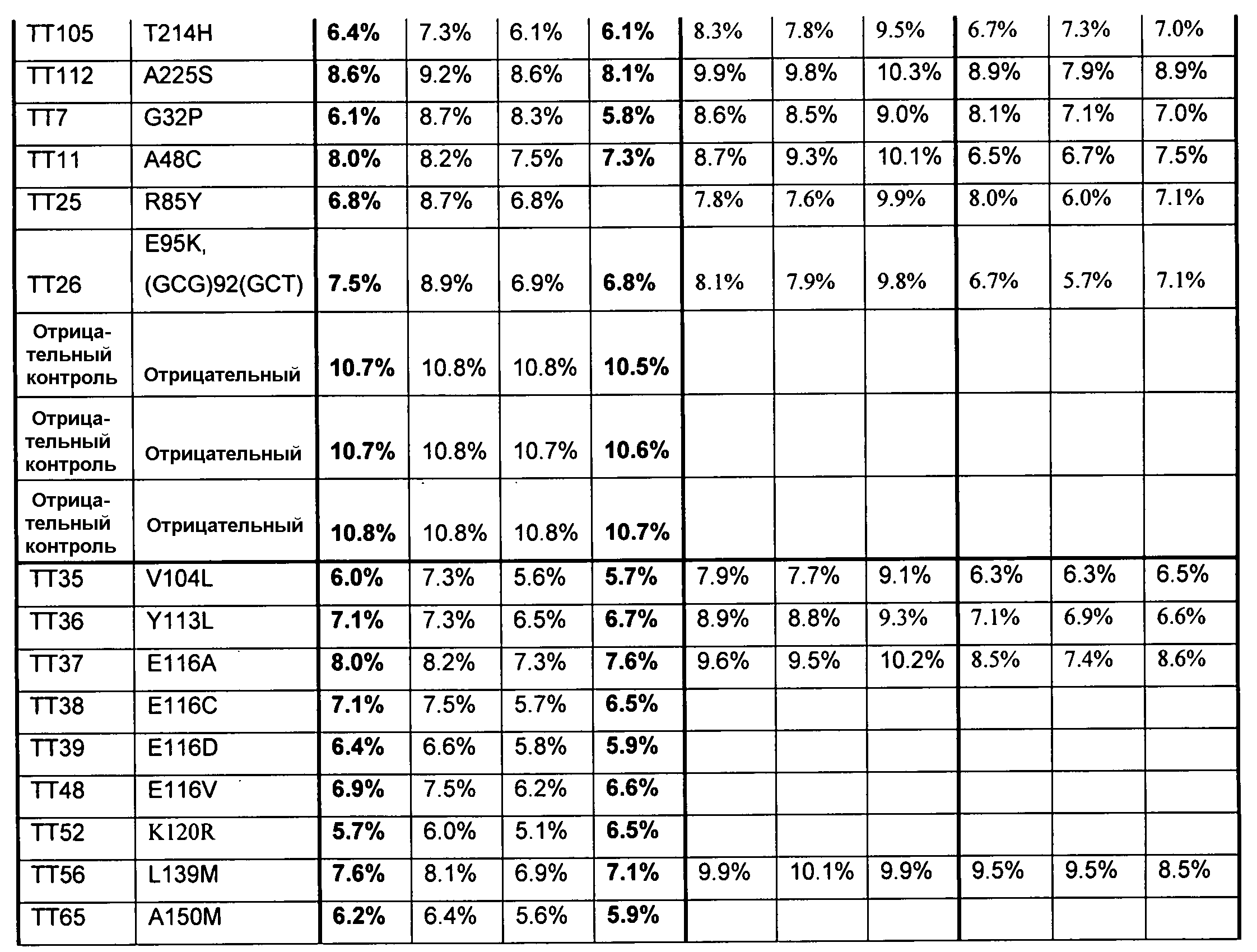

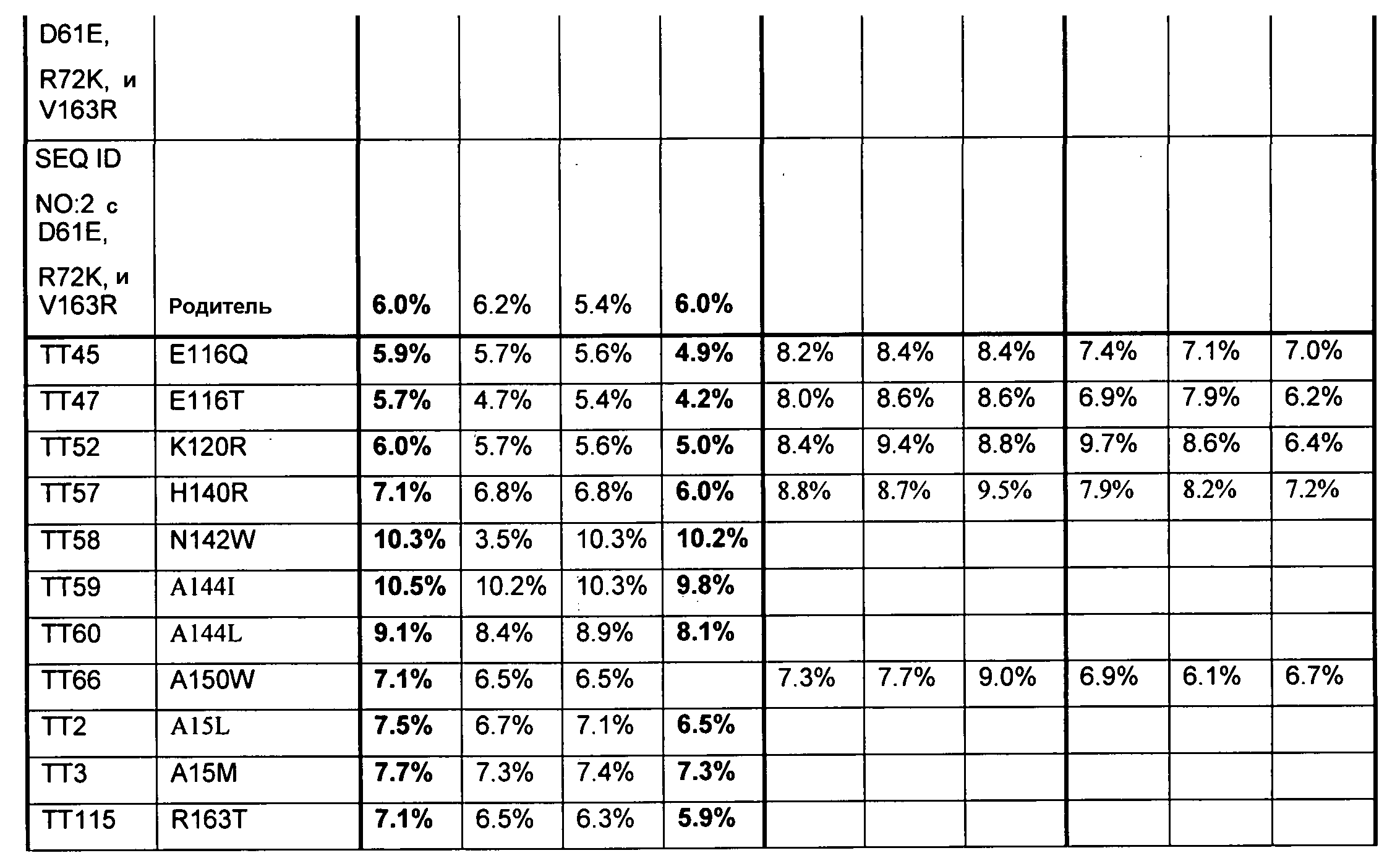
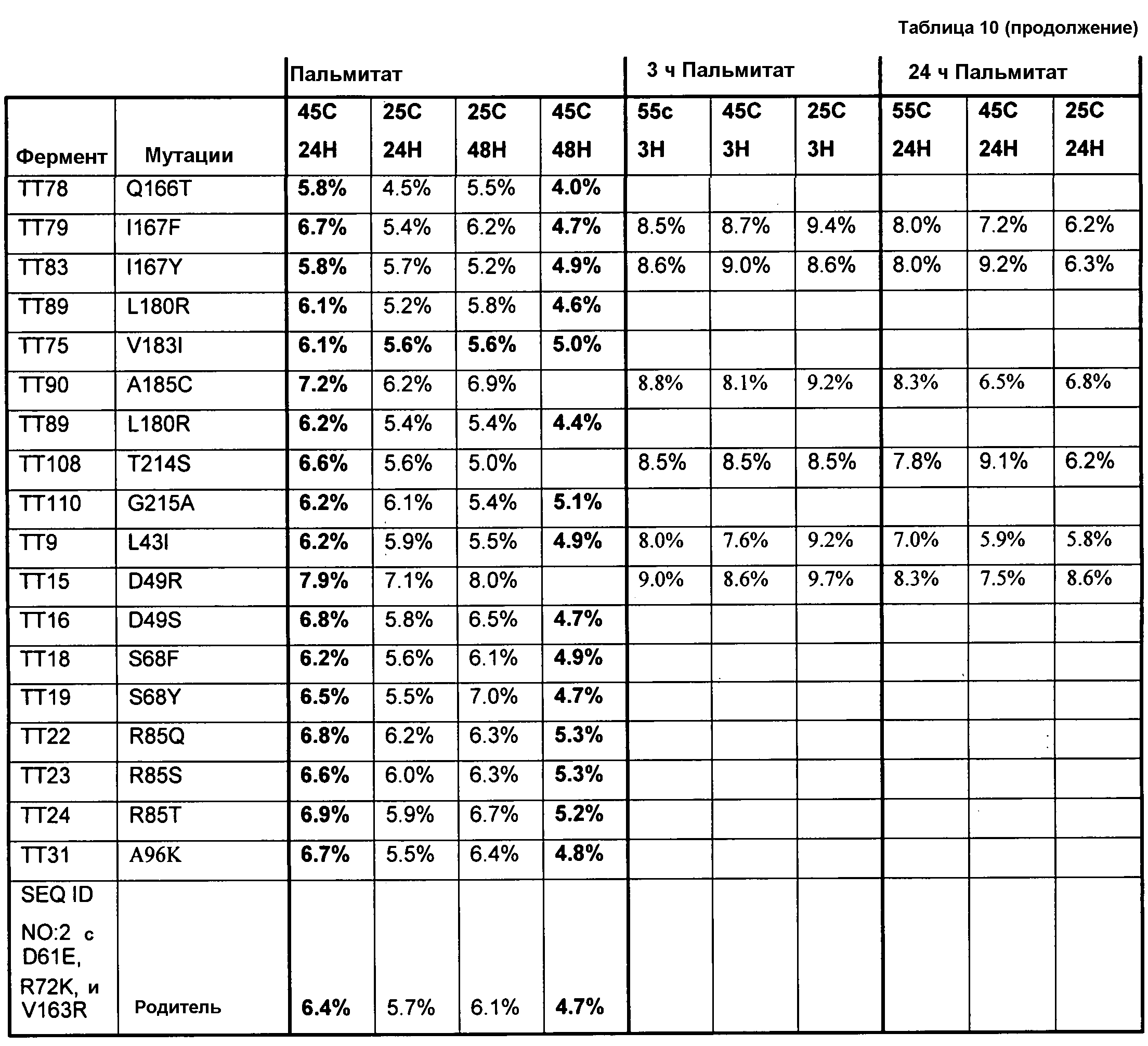


Замену кодона в данном остатке, которая не ведет к аминокислотной замене, обозначают как молчащую мутацию. Молчащие мутации получали в виде хитов вследствие увеличенной экспрессии по отношению к родительскому ферменту и наблюдали в 37 сайтах в скрининге GSSM (таблица 11, ниже).
Пример 10: Образцовая Эволюция для усовершенствования тепловой устойчивости пальмитазы с использованием технологии TMCASM
Наиболее эффективные мутации, идентифицированные в процессе эволюции GSSM (пример 9, выше) оценивали в первичном и вторичном анализе с маслом для включения в эволюцию TMCA. Анализы 117 уникальных хитов с 5 г нерафинированного масла проводили, как описано в примере 5, при температуре 25°C и 45°C, но при содержании воды 10%. Профиль масла оценивали после реакции в течение 24 и 48 часов. Первичные хиты подвергали вторичному анализу с маслом при 25°C, 45°C и 55°C. Для оценки профилей масла аликвоты брали в моменты времени 3, 24 и 48 часов. Наиболее эффективные ферменты перечислены ниже в таблице 12, где оставшийся в масле пальмитат указан в виде процента от всех связанных жирных кислот. Для оценки влияния промежуточного щелочного рафинирования, в анализах при 45°C проводили щелочное рафинирование посредством добавления 11% NaOH (оценивали для 7% FFA) и при непрерывном перемешивании нагревали до 60°C. Свежий фермент добавляли в рафинированное масло при содержании воды 20%, гомогенизировали и продолжали реакцию при перемешивании в течение дополнительных 24 часов при 45°C. Конечное содержание пальмитата в рафинированных щелочью образцах приведено в последней колонке, «ЩР 24 ч».
Среди наиболее эффективных ферментов, приведенных в таблице 12, для комбинирования с использованием технологии TMCA отобрали 15 мутанов (показаны жирным курсивным начертанием в таблице 12). Библиотеку TMCA конструировали, как описано в публикации PCT № WO 2009/018449 и в соответствии с нижеследующим дополнительным описанием. Эта библиотека содержала 9216 уникальных вариантов.

Технология Tailored Multi-Site Combinatorial Assembly (TMCA), технология TMCA (см. публикацию PCT № WO 09/018449), включает способ получения нескольких дочерних полинуклеотидов, содержащих различные сочетания разнообразных мутаций в нескольких сайтах. Способы можно выполнять отчасти посредством комбинирования по меньшей мере одной или нескольких следующих стадий:
Получение информации о последовательности («первого» или «матричного») полинуклеотида. Например, первая или матричная последовательность может представлять собой последовательность дикого типа (например, SEQ ID № 2 с мутациями D61E, R72K и V163R) или матировавшую последовательность. Информация о последовательности может относиться к полноразмерному полинуклеотиду (например, к гену или открытой рамке считывания) или к фрагментарным участкам, представляющим интерес, таким как последовательность, кодирующая сайт для связывания, специфичности связывания, катализа или субстратной специфичности.
Идентификация трех или более мутаций, представляющих интерес, в первой или матричной последовательности полинуклеотида. Например, в первой или матричной последовательности мутации могут присутствовать в 3, 4, 5, 6, 8, 10, 12, 20 или большем количестве положений. Положения можно предварительно определить посредством абсолютного положения или посредством контекста окружающих остатков или гомологии. Например, для TMCA полипептидов пальмитаз в качестве мутаций, представляющих интерес, включали лучшие термостабильные аминокислотные замены, которые приводят к усовершенствованной эффективности фермента. Могут быть известны последовательности, фланкирующие положения мутаций с любой стороны. Каждое положение мутации может содержать две или более мутации, например, для различных аминокислот. Такие мутации можно идентифицировать путем использования технологии Gene Site Saturation MutagenesisSM (GSSMSM), как описано в настоящем документе и, например, в патентах США №№ 6171820; 6562594 и 6764835.
Предоставление праймеров (например, синтетических олигонуклеотидов), содержащих мутации, представляющие интерес. В одном из вариантов осуществления предоставляют праймер для каждой мутации, представляющей интерес. Таким образом, первый или матричный полинуклеотид, содержащий 3 мутации, представляющие интерес, может использовать 3 праймера в этом положении. Также праймер можно предоставить в виде пула праймеров, содержащих вырожденное положение, с тем, чтобы мутация, представляющая интерес, представляла собой диапазон любых нуклеотидов или встречающихся в природе аминокислот или подмножество этого диапазона. Например, можно предоставить пул праймеров, которые способствуют образованию мутаций алифатических аминокислотных остатков. Праймеры можно получить в виде прямых или обратных праймеров или праймеры можно получить в виде по меньшей мере одного прямого праймера и по меньшей мере одного обратного праймера. Когда мутации расположены близко друг к другу, может оказаться целесообразным использовать праймеры, которые содержат мутации более чем для одного положения или различные сочетания мутаций в нескольких положениях.
Предоставление полинуклеотида, содержащего матричную последовательность. Первый или матричный полинуклеотид может быть кольцевым или может быть суперскрученным, таким как плазмида или вектор для клонирования, секвенирования или экспрессии. Полинуклеотид может быть одноцепочечным («оцДНК») или может быть двухцепочечным («дцДНК»). Например, в способе TCMA суперскрученную («сс») дцДНК матрицу подвергают стадии нагревания при 95°C в течение 1 мин (см. Levy, Nucleic Acid Res., 28(12):e57(i-vii) (2000)).
Присоединение праймеров к матричному полинуклеотиду в реакционной смеси. Праймеры и матричный полинуклеотид объединяют в условиях, которые позволяют праймерам гибридизоваться с матричным полинуклеотидом. В одном из вариантов осуществления протокола TMCA праймеры присоединяют к полинуклеотиду в одной реакционной смеси, но их можно присоединять и в нескольких реакциях.
Осуществление реакции(й) полимеразного удлинения. Продукты удлинения (например, в качестве «потомства» или «модифицированного удлиненного полинуклеотида») можно амплифицировать стандартными средствами. Можно анализировать длину, последовательность, желаемые свойства нуклеиновой кислоты продуктов или экспрессировать их в виде полипептидов. Другие способы анализа включают гибридизацию in situ, скрининг последовательностей или скрининг экспрессии. Анализ может включать один или несколько раундов скрининга и отбора по желаемому свойству. Также продукты можно трансформировать в клетку или другую или систему экспрессии, такую как бесклеточная система. Бесклеточная система может содержать ферменты, связанные с репликацией, репарацией, рекомбинацией, транскрипцией или трансляцией ДНК. Образцовые организмы-хозяева включают клетки бактерий, дрожжей, растений и животных и клеточные линии и включают E. coli, Pseudomonas fluorescens, Pichia pastoris и Aspergillus niger. Например, в качестве организмов-хозяев штаммы можно использовать E. coli XL1-Blue или Stbl2.
Способ по изобретению можно использовать с одинаковыми или различными праймерами при различных условиях реакции, чтобы содействовать образованию продуктов, содержащих различные сочетания или число мутаций.
При осуществлении образцового способа, описанного выше, этот протокол также предусматривает один или несколько полинуклеотидов, полученных посредством этого способа эволюции TMCA, которые затем можно подвергнуть скринингу или отбору по желаемому свойству. Один или несколько дочерних полинуклеотидов можно экспрессировать в виде полипептидов и необязательно подвергнуть скринингу или отбору по желаемому свойству. Таким образом, этот вариант осуществления протокола эволюции TMCA предусматривает полинуклеотиды и кодируемые полипептиды, а также библиотеки таких полинуклеотидов, кодирующих такие полипептиды. Этот вариант осуществления протокола эволюции TMCA дополнительно предусматривает скрининг библиотек посредством скрининга или отбора библиотеки для получения одного или нескольких полинуклеотидов, кодирующих один или несколько полипептидов, обладающих желаемой активностью.
Другой вариант осуществления протокола эволюции TMCA, описанный в публикации PCT № WO 2009/018449, включает способ получения нескольких модифицированных полинуклеотидов. Такие способы, как правило, включают (a) добавление по меньшей мере трех праймеров к двухцепочечному матричному полинуклеотиду в одной реакционной смеси, где по меньшей мере три праймера не перекрываются и где каждый из по меньшей мере трех праймеров содержит по меньшей мере одну мутацию, отличающуюся от других праймеров, где по меньшей мере один праймер представляет собой прямой праймер, который может гибридизоваться с минус-цепью матрицы, и по меньшей мере один праймер представляет собой обратный праймер, который может гибридизоваться с плюс-цепью матрицы, и (b) проведение реакции полимеразного удлинения в реакционной смеси, чтобы по меньшей мере из трех праймеров получить несколько удлиненных модифицированных полинуклеотидов.
Другой вариант осуществления протокола эволюции TMCA, описанный в публикации PCT № WO 2009/018449, включает способ, где клетку трансформируют несколькими удлиненными продуктами, которые не обрабатывали лигазой. В другом варианте осуществления изобретения несколько удлиненных модифицированных полинуклеотидов получают из клетки. В другом варианте осуществления несколько полученных удлиненных модифицированных полинуклеотидов анализируют, например, посредством экспрессии по меньшей мере одного из нескольких удлиненных модифицированных полинуклеотидов и анализа полипептида, экспрессированного с них. В другом варианте осуществления отбирают несколько удлиненных модифицированных полинуклеотидов, содержащих мутации, представляющие интерес. В другом варианте осуществления протокола эволюции TMCA получают информацию о последовательности, касающуюся матричного полинуклеотида, и по всей длине матричного полинуклеотида можно идентифицировать три или более мутации, представляющие интерес. В другом варианте осуществления продукты, полученные посредством полимеразного удлинения, можно анализировать перед трансформацией клетки несколькими удлиненными модифицированными продуктами.
В одном из вариантов осуществления протокола эволюции TMCA продукты, полученные посредством полимеразного удлинения обрабатывают ферментом, например, рестрикционным ферментом, таким как рестрикционный фермент DpnI, посредством чего разрушают последовательность матричного полинуклеотида. Обработанными продуктами можно трансформировать клетку, например, клетку E. coli. В одном из вариантов осуществления протокола эволюции TMCA можно использовать по меньшей мере два или по меньшей мере три или по меньшей мере четыре или по меньшей мере пять или по меньшей мере шесть или по меньшей мере семь или по меньшей мере восемь или по меньшей мере девять или по меньшей мере десять или по меньшей мере одиннадцать или по меньшей мере двенадцать или более праймеров. В одном из вариантов осуществления каждый праймер содержит одну точечную мутацию. В другом варианте осуществления два прямых или два обратных праймера содержат различные изменения в одном и том же положении в матричном полинуклеотиде. В другом варианте осуществления по меньшей мере один праймер содержит по меньшей мере два изменения в различных положениях в матричном полинуклеотиде. В еще одном другом варианте осуществления по меньшей мере один праймер содержит по меньшей мере два изменения в различных положениях и по меньшей мере два прямых или два обратных праймера содержат различные изменения в одном и том же положении в матричном полинуклеотиде. В одном из вариантов осуществления протокола эволюции TMCA прямые праймеры сгруппированы в прямую группу, а обратные праймеры сгруппированы в обратную группу, и праймеры в прямой группе и праймеры в обратной группе, независимо друг от друга, нормализованы относительно равной концентрации в соответствующей группе, независимо от положений в матричном полинуклеотиде, и где после нормализации в реакцию добавляют равное количество прямого и обратного праймеров. В этом способе нормализации, сочетание некоторых положений может быть смещенным. Смещение может происходить в результате, например, относительно низкой концентрации праймера в одном положении, содержащем один праймер, по сравнению с положением, содержащим несколько праймеров. «Смещение по положению» относится к полученным полинуклеотидам, которые проявляют выраженное предпочтение относительно встраивания праймеров в одном положении по отношению к другим положениям в пределах их группы прямых или обратных праймеров. Это ведет к сочетанию модифицированных полинуклеотидов, которые могут содержать высокую процентную долю мутаций в одном положении праймера, но низкую процентную долю мутаций в другом положении в их группе прямых или обратных праймеров. Такое смещение является неблагоприятным, когда задача TMCA состоит в создании дочерних полинуклеотидов, содержащих все возможные сочетания изменений в матрице. Смещение можно корректировать, например, посредством нормализации праймеров в виде пула в каждом положении для соблюдения равенства.
В одном из вариантов осуществления протокола эволюции TMCA нормализацию праймеров осуществляют посредством организации праймеров несколькими группами в зависимости от их местоположения на матричном полинуклеотиде, где праймеры, покрывающие один и тот же выбранный участок на матрице, находятся в одной группе; нормализации сгруппированных праймеров внутри каждой группы для достижения равной концентрации; объединения прямых праймеров внутри одной группы в прямую группу и нормализации концентрация между каждой группой прямых праймеров для соблюдения равенства; объединения обратных праймеров внутри одной группы в обратную группу и нормализация концентрации между каждой группой обратных праймеров для соблюдения равенства; и добавления равного количества объединенных прямых и обратных праймеров в реакцию. Для сочетаний положений отклонения не наблюдали. В одном из вариантов осуществления протокола эволюции TMCA предусмотрен набор вырожденных праймеров, каждый содержит вырожденное положение, где мутация, представляющая интерес, представляет собой спектр различных нуклеотидов в вырожденном положении. В другом варианте осуществления предусмотрен набор вырожденных праймеров, содержащих по меньшей мере один вырожденный кодон, соответствующий по меньшей мере одному кодону матричного полинуклеотида, и по меньшей мере одну смежную последовательность, которая гомологична последовательности, смежной с кодоном последовательности матричного полинуклеотида. В другом варианте осуществления вырожденный кодон представляет собой N,N,N и кодирует любые из 20 встречающихся в природе аминокислот. В другом варианте осуществления вырожденный кодон кодирует менее чем 20 встречающихся в природе аминокислот.
Другой вариант осуществления протокола эволюции TMCA, описанный в публикации PCT № WO 2009/018449, включает способ получения нескольких модифицированных полинуклеотидов, содержащих мутации, представляющие интерес. Такие способы, как правило, включают (a) добавление по меньшей мере двух праймеров к двухцепочечному матричному полинуклеотиду в одну реакционную смесь, где по меньшей мере два праймера не перекрываются и где каждый из по меньшей мере двух праймеров содержит по меньшей мере одну мутацию, отличную от другого праймера(ов), где по меньшей мере один праймер представляет собой прямой праймер, который может гибридизоваться с минус-цепью матрицы, и по меньшей мере один праймер представляет собой обратный праймер, который может гибридизоваться с плюс-цепью матрицы, (b) проведение реакции полимеразного удлинения в реакционной смеси, чтобы получить несколько удлиненных модифицированных полинуклеотидов из по меньшей мере двух праймеров, (c) обработку нескольких удлиненных модифицированных полинуклеотидов ферментом, посредством чего разрушают матричный полинуклеотид, (d) трансформацию клетки обработанными удлиненными модифицированными полинуклеотидами, которые не обрабатывали лигазой, (e) получение нескольких удлиненных модифицированных полинуклеотидов из клетки и (f) отбор нескольких удлиненных модифицированных полинуклеотидов, содержащих мутации, представляющие интерес.
В этом примере 15 мутаций, отобранные для включения в термостабильную библиотеку TMCA, представлены ниже в таблице 13. Использовали шесть ДНК-матриц: SEQ ID № 2 с мутациями D61E, R72K и V163R (родительская последовательность), родительская последовательность с мутацией E95K, родительская последовательность с мутацией P162G, родительская последовательность с мутацией P162K, родительская последовательность с мутациями E95K и P162G и родительская последовательность с мутациями E95K и P162K. Плазмиды родительской последовательности с мутацией E95K, родительской последовательности с мутацией P162G, родительской последовательности с мутацией P162K were пассировали посредством компетентных клеток E. coli XL1-Blue, чтобы метилировать ДНК. Проводили шесть отдельных эволюций TMCA, по одной на каждую из шести матриц в сочетании с олигонуклеотидами, перечисленными ниже в таблице 14.

Членов библиотеки амплифицировали в векторе pDOW-kan, анализировали в агарозном геле и обрабатывали с помощью DpnI. Затем образцы трансформировали в компетентные клетки E. coli XL1-Blue. Колонии собирали, растили и секвенировали. Результаты секвенирования показали, что мутация E116L и мутации аминокислот 144, 149 и 150 представлены в недостаточной мере по сравнению с теоретическим максимумом. Поэтому создавали другую библиотеку TMCA с использованием нормализованной смеси всех шести матриц. Реакции TMCA повторяли с использованием всех олигонуклеотидов, перечисленных в таблице 14, за исключением олигонуклеотидов для мутаций E116I и E116N. Удваивали количество праймеров, использованных для мутации E116L и для участка 4 (аминокислоты 144, 149 и 150). Образцы амплифицировали в векторе pDOW-kan, анализировали в агарозном геле и обрабатывали с помощью DpnI. Образец ДНК трансформировали в компетентные клетки E. coli XL1-Blue. Колонии собирали, растили и секвенировали. Усовершенствовали введение мутаций аминокислот 144, 149 и 150, E116L все еще была представлена в недостаточной мере по сравнению с теоретическим максимумом. ДНК из обеих библиотек трансформировали в компетентные клетки E. coli XL1-Blue. Колонии объединяли и получали ДНК с использованием набора Qiagen mini-prep. Затем ДНК использовали для трансформации компетентных клеток Pseudomonas fluorescens. Клетки растили в среде LB с добавлением урацила (750 мкг/мл) и канамицина (50 мкг/мл). Для библиотеки получали достаточное число колоний с 10-кратным избытком. Чтобы учесть представленную в недостаточной мере E116L, библиотеку наполняли образцами в 10-кратном избытке.
Полученную термостабильную библиотеку TMCA растили в минимальной среде M9 с добавлением урацила (750 мкг/мл) и канамицина (50 нг/мл) и проводили скрининг с использованием высокопроизводительного скрининга, который описан выше в примере 3. Библиотеку инкубировали при температуре 60°C в течение 30 минут до и после добавления УМБ-пальмитата. Затем 384-луночные планшеты считывали при длине волны возбуждения 365 нм и длине волны испускания 460 нм. Каждый планшет быстро охлаждали посредством центрифугирования, спектрофотометр предварительно нагревали, чтобы минимизировать артефакты в показаниях. Отображали значения флуоресценции по каждому 384-луночному планшету. Хиты идентифицировали по превышению положительного контроля более чем на 2 стандартных отклонения (одиночная мутация E116L). Анализ хитов первичного высокопроизводительного скрининга проводили посредством вторичного высокопроизводительного скрининга при двух температурах 60°C и 63°C. Вторичные хиты определяли по превышению усредненного положительного контроля более чем на 4 стандартных отклонения в планшете при 63°C и по сохранению >50% активности при температуре от 60°C до 63°C.
Идентифицировали 318 хитов вторичного высокопроизводительного скрининга с уникальными последовательностями. Эти хиты содержали от 2 до 9 мутаций. Используя осветленные лизаты 318 хитов, проводили стандартные анализы с 5 г нерафинированного масла (как описано в примере 5). Из 318 хитов вторичного высокопроизводительного скрининга в качестве «масляных хитов» с тепловой устойчивостью выбирали те хиты, которые в анализе с маслом снижали содержание пальмитата более эффективно при повышенных температурах (45°C или 60°C), чем при 25°C. Посредством дополнительных анализов с маслом идентифицировали и охарактеризовали 57 первичных масляных хитов с тепловой устойчивостью (ниже см. таблицы 15 и 16). Проводили дополнительные анализы с 5 г масла и идентифицировали 9 масляных хитов, которые обладали предпочтительными профилями температурной эффективности в дополнение к изменению уровней общей активности (ниже см. таблицу 17).

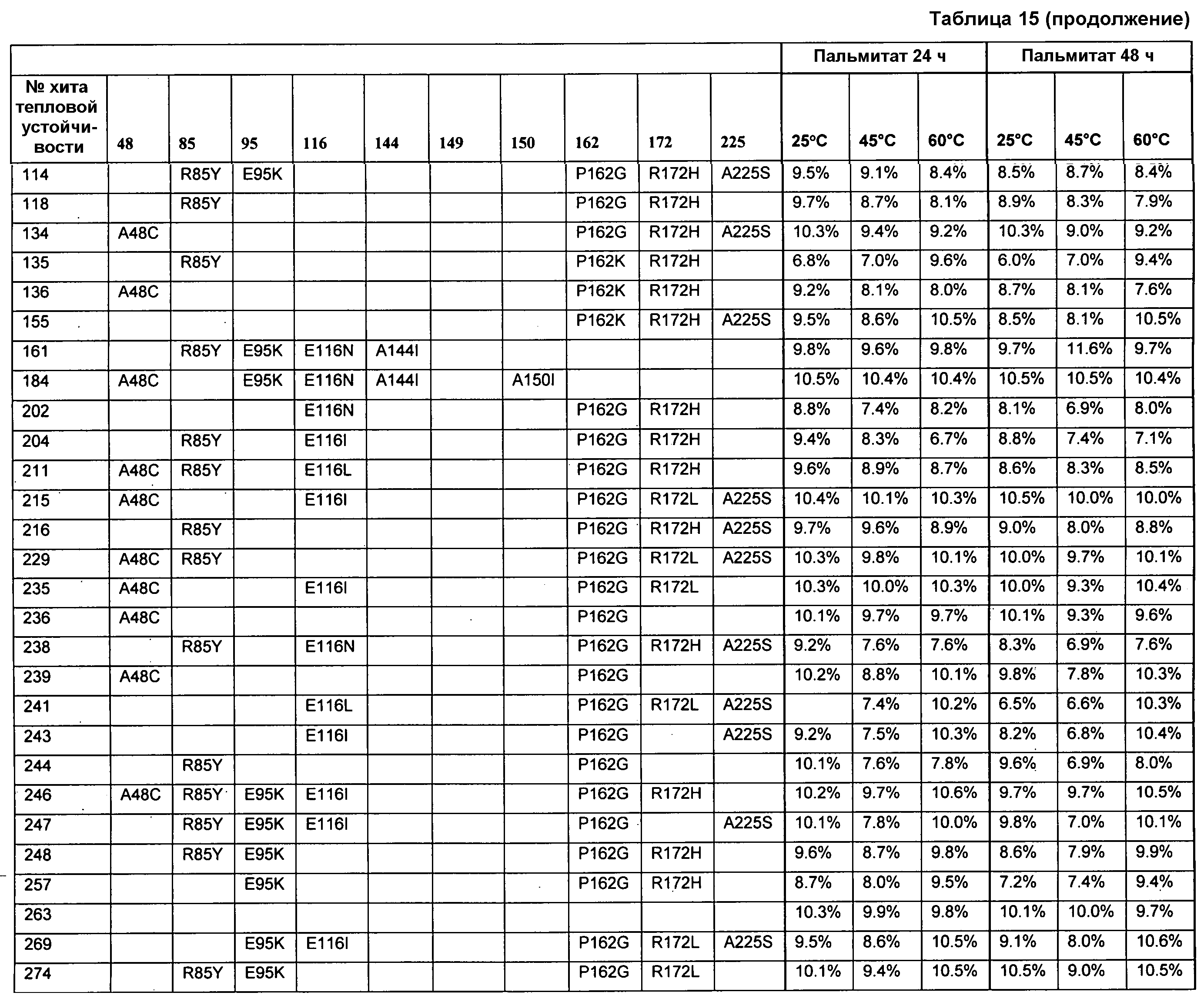

• Хиты тепловой устойчивости 29, 40, 41 и 74 также содержат дополнительные мутации (см. дополнительные мутации, перечисленные ниже в таблице 16).
Пример 11: Образцовая эволюция для усиления экспрессии пальмитазы с использованием технологии TMCASM
Оценивали экспрессию (см. ниже таблицу 18) 37 индивидуальных молчащих мутаций, идентифицированных в процессе скрининга GSSM (см. выше таблицу 11, пример 9). В колбах по 250 мл экспрессировали 50 мл культуры в среде M9 с добавлением урацила и канамицина. Хиты растили при 30°C в течение от 16 до 20 ч до и после индукции с использованием 0,3 мМ IPTG. Клетки осаждали центрифугированием и лизировали с использованием B-PER™ (по каталогу № 78248, Pierce Protein Research Products, Rockford, IL). Часть полного лизата центрифуриговали для осветления. Проводили анализ концентрации общего белка как в полном клеточном лизате, так и в осветленных лизатах с использованием Bio-Rad Protein Analysis (Bio-Rad, Hercules, CA, № по каталогу 500-0006) на основе способа Брэдфорда, и анализ активности на флуорогенном субстрате УМБ-пальмитат, и посредством SDS-PAGE. Значения нормализовали относительно значения родительского белка, принятого за 100%. Идентифицировали 16 хитов, которые демонстрировали повышенную активность относительно родительского белка).
Лучшие 11 индивидуальных молчащих мутаций (таблица 19, ниже) объединяли с использованием технологии TMCA, как описано в публикации PCT № WO 2009/018449 и дополнительно описано ниже).
Для эволюции TMCA использовали четыре ДНК-матрицы: (1) пальмитазу «Родитель A» (SEQ ID № 2 с мутациями D61E, R72K и V163R), (2) пальмитазу «Родитель B» (Родитель A с дополнительными мутациями (GGC)45(GGA) и (CTG)117(CTT)), (3) пальмитазу «Родитель C» (Родитель A с дополнительной мутацией (GGC)45(GGA)) и (4) «Родитель D» (Родитель A с дополнительной мутацией (CTG)117(CTT)). Комментарий: в мутациях указан исходный кодон, за которым следует номер положения модифицированной аминокислоты, за которым следует новый кодон. Например, в (GGC)45(GGA) указано, что кодон для аминокислоты в положении 45 из SEQ ID № 2 заменили с (GGC) на (GGA).
Первый раунд реакций TMCA выполняли с использованием каждой из четырех ДНК-матриц и праймеров для участков 1 и 4 (см. ниже таблицу 20; участок 1: аминокислота 35; участок 4: аминокислоты 183 и 188), посредством чего создавали четыре подбиблиотеки. Продукты подбиблиотек очищали с использованием Qiagen PCR Cleanup Kit. Затем каждую подбиблиотеку, содержащую смесь очищенных продуктов и, следовательно, содержащую несколько матриц, использовали в одной второй реакции TMCA на одну подбиблиотеку с праймерами для участков 2 и 3 (см. ниже таблицу 21; участок 2: аминокислоты 102 и 108; участок 3: аминокислоты 124, 126, 128 и 133).
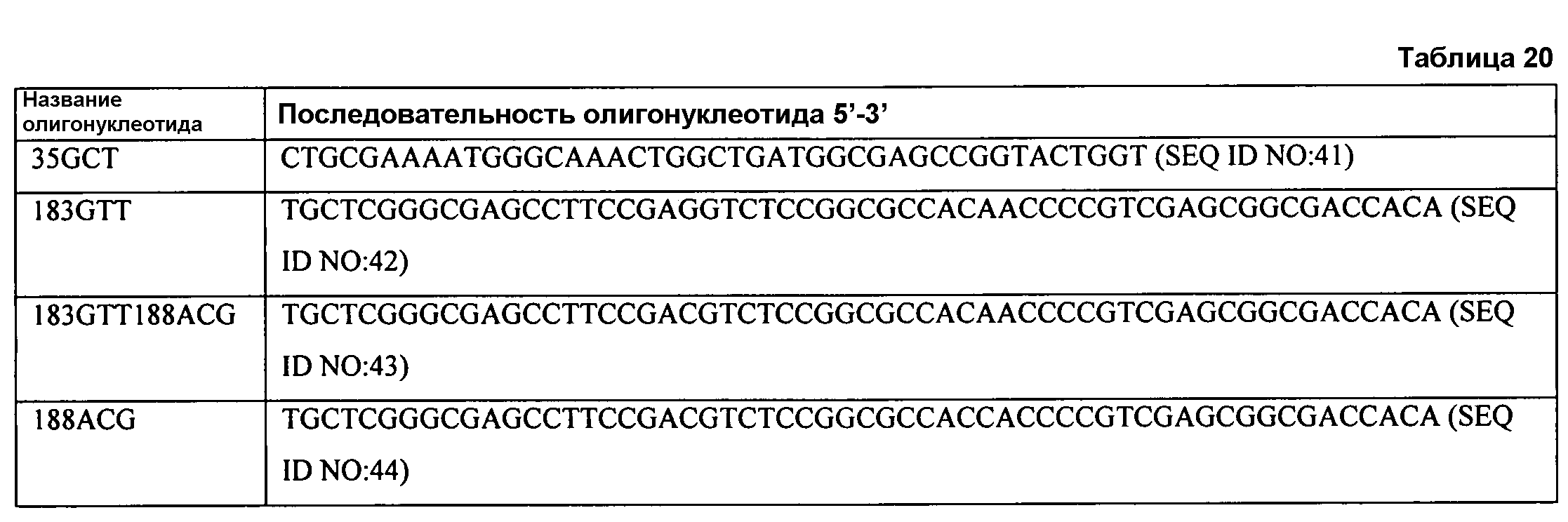
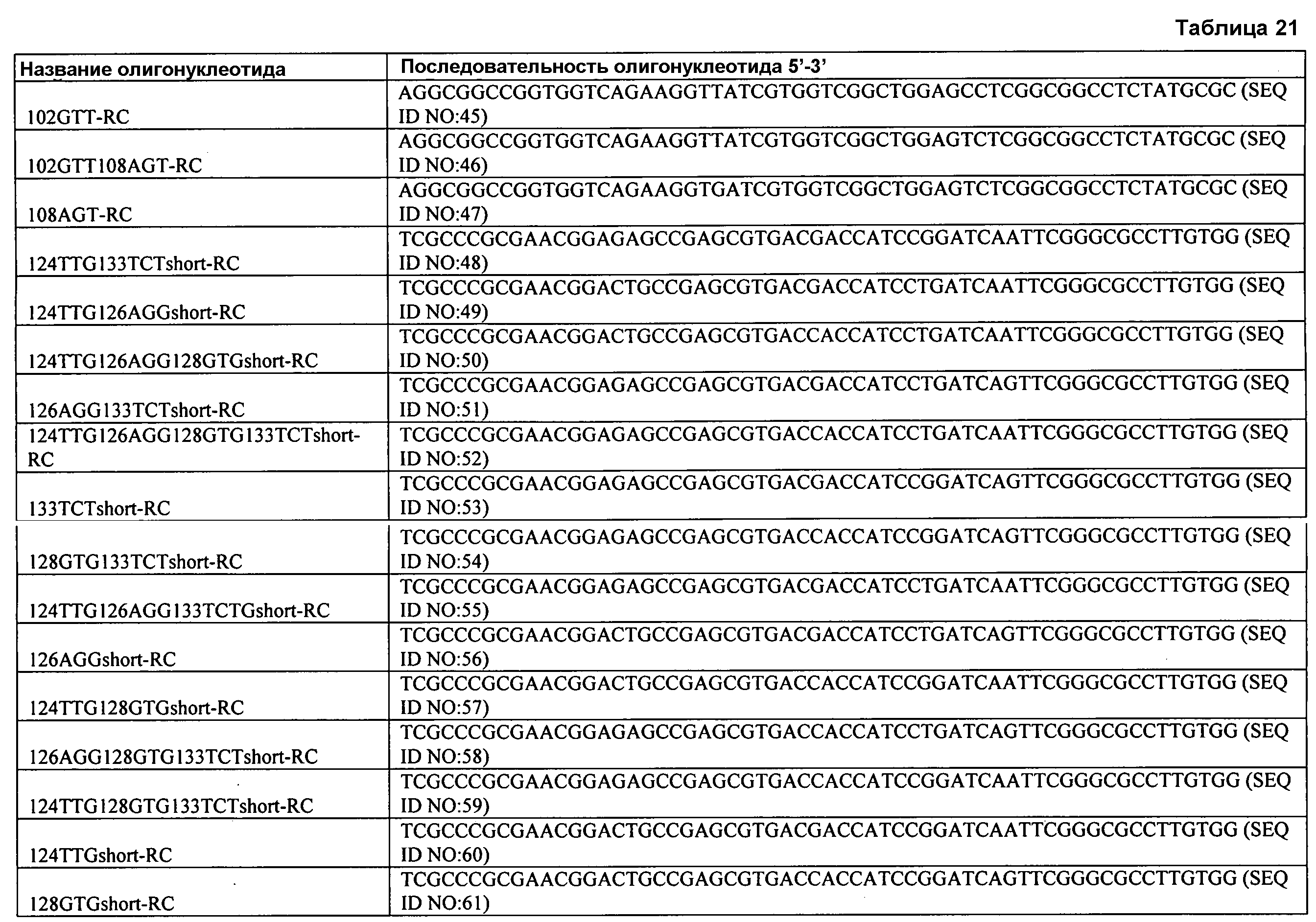
Образцы амплифицировали в векторе pDOW-kan, анализировали в агарозном геле, обрабатывали с использованием DPNI и затем трансформировали в компетентные клетки E. coli XL1-Blue. Колонии растили, собирали и секвенировали. Колонии объединяли и получали ДНК для каждой подбиблиотеки с использованием набора Qiagen mini-prep (по каталогу № 27106, Qiagen, Valencia, CA). Затем компетентные клетки Pseudomonas fluorescens трансформировали этой ДНК. Клетки растили в среде LB с добавлением урацила (750 мкг/мл) и канамицина (50 мкг/мл). Для библиотеки получали достаточное число колоний с 7-кратным избытком (по меньшей мере 14000 колоний).
Библиотеку упорядочили, растили в минимальной среде M9 с добавлением урацила (750 мкг/мл) и канамицина (50 мкг/мл) и проводили анализ с использованием 400 мкМ 4-метилумбеллиферилпальмитата в 80 мМ HEPES с pH 7,5. Образцы инкубировали в течение 30 минут при температуре 54°C до и после добавления субстрата. Флуоресценцию считывали при длине волны возбуждения 360 нм и длине волны испускания 465 нм. 46 образцов с молчащими мутациями дали уникальные последовательности (см. ниже таблицу 22). Показания флуоресценции для каждого из 46 образцов с молчащими мутациями также перечислены в таблице 22, в первой колонке «УМБ активность». В дополнение к молчащим мутациям, образцы с молчащими мутациями 2, 7, 9, 10, 13, 16, 23, 25, 40 и 44 содержали изменения аминокислот (а.к.) (см. ниже таблицу 23).



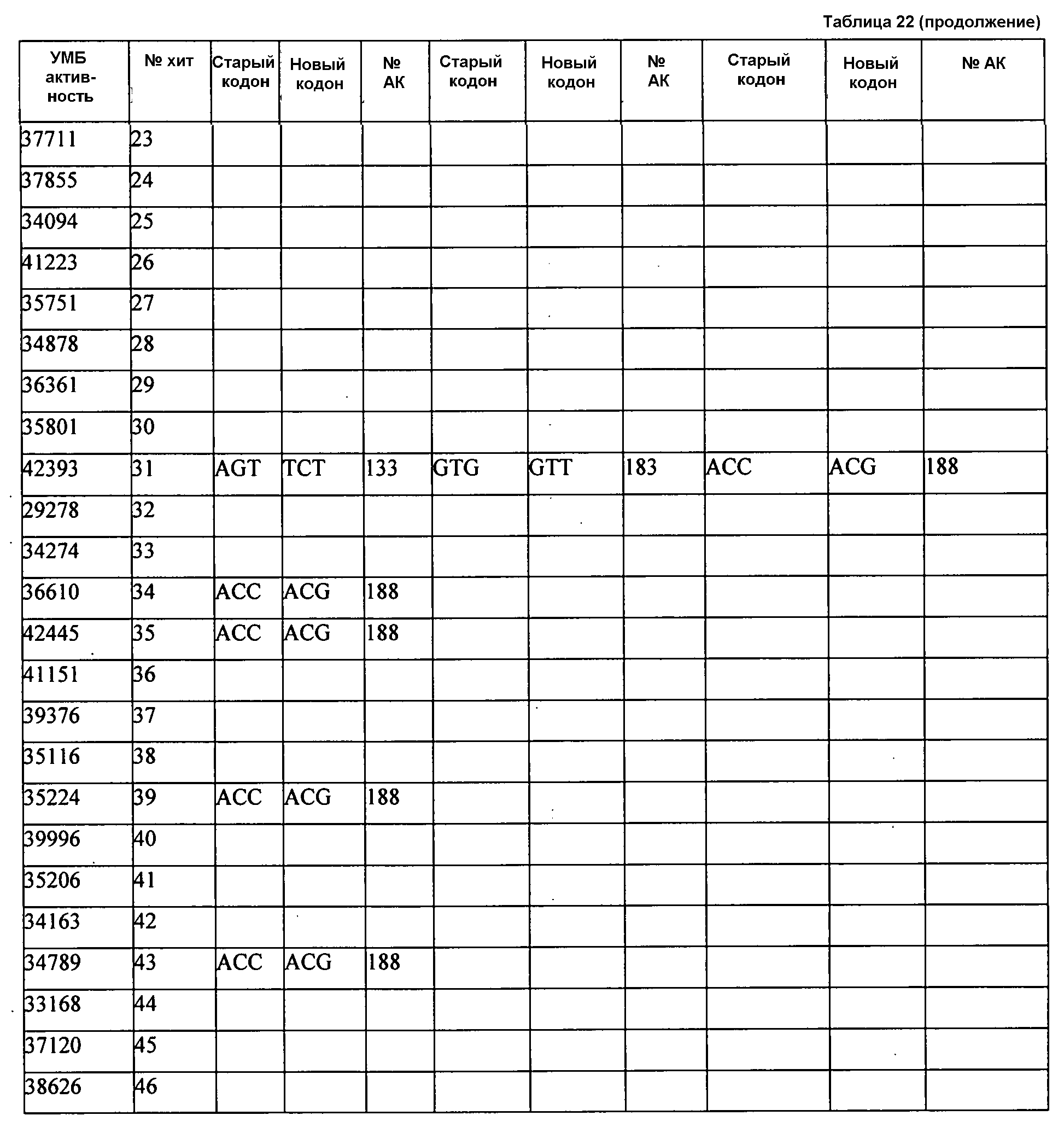
46 образцов с уникальными молчащими мутациями растили во встряхиваемых колбах объемом 250 мл. Проводили количественное определение осветленных лизатов с использованием Bio-Rad Protein Analysis (Bio-Rad, Hercules, CA, по каталогу № 500-0006) на основе способа Брэдфорда и затем нормализовали относительно экспрессии общего белка. Экспрессию анализировали посредством SDS-PAGE.
Родительский белок (SEQ ID № 2 с мутациями D61E, R72K и V163R), отрицательный контроль и лучше 4 хита с молчащими мутациями экспрессировали в масштабе 1 л. Каждый образец лизировали посредством микрофлюидайзера или в присутствие детергента BPER. Определение концентрации белка проводили для неочищенных и осветленных лизатов с использованием Bio-Rad Protein Analysis (Bio-Rad, Hercules, CA, по каталогу № 500-0006) на основе способа Брэдфорда. Затем на каждую дорожку SDS-PAGE геля загружали равное количество общего белка. Схожие уровни пальмитазы наблюдали при всех протестированных условиях растворения и лизиса. Хиты с молчащими мутациями 26, 34, 35 и 37 (таблицы 22 и 23, выше) показывали более высокую процентную долю экспрессии пальмитазы по сравнению с родительским белком (SEQ ID № 2 с мутациями D61E, R72K и V163R). Хит с молчащей мутацией 35 обладал самой высокой процентной долей экспрессии пальмитазы. Хит с молчащей мутацией 35 содержал следующие семь молчащих мутаций по сравнению с родительским белком: (GCG)35(GCT), (GTG)102(GTT), (AGC)108(AGT), (CTG)117(CTT), (CGG)126(AGG), (AGT)133(TCT) и (ACC)188(ACG).
Пример 12: Образцовая эволюция для повышения экспрессии термостабильных пальмитаз
Для оценки экспрессии 9 подходящих лидирующих термостабильных пальмитаз (пример 10, таблицы 15, 16 и 17), лучше 7 молчащих мутаций (пример 11) вводили в каждый термостабильную лидирующую пальмитазу, как описано ниже. В результате каждая термостабильная лидирующая пальмитаза (хиты тепловой устойчивости 29, 40, 41, 74, 81, 202, 204, 238 и 244) содержали следующие молчащие мутации (35GCT, 102GTT, 108AGT, 117CTT, 126AGG, 133TCT, 188ACG), введенные в каждую из них. Каждую термостабильную лидирующую пальмитазу с введенными молчащими мутациями переименовали путем добавления «SM» после номера термостабильного хита. Например, термостабильный хит 29 с введенными молчащими мутациями переименовали в «29 SM». Подобным образом, хит 40 переименовали в «40 SM», хит 41 переименовали в «41 SM», хит 74 переименовали в «74 SM», хит 81 переименовали в «81 SM», хит 202 переименовали в «202 SM», хит 204 переименовали в «204 SM», хит 238 переименовали в «238 SM», а хит 244 переименовали в «244 SM».
Для того чтобы ввести лучшие 7 молчащих мутаций в 9 лидирующих термостабильных пальмитаз, использовали технологию TMCA, как описано в публикации PCT № WO 2009/018449 и дополнительно описано ниже.
Для первого раунда эволюции TMCA лучшие 9 лидирующих термостабильных пальмитаз и лидирующий экспрессионный хит (хит с молчащей мутацией № 35) пассировали посредством компетентных клеток E. coli XL1-Blue для того, чтобы метилировать ДНК. Хит с молчащей мутацией № 35 использовали в качестве ДНК-матрицы. Олигонуклеотиды, перечисленные ниже в таблице 24, использовали для введения сочетаний мутаций в ДНК-матрицу.

Образцы амплифицировали в векторе pDOW-kan, анализировали в агарозном геле и обрабатывали с использованием DpnI. Образцы трансформировали в компетентные клетки E. coli XL1-Blue. Колонии собирали, растили и секвенировали. Посредством секвенирования идентифицировали несколько сочетаний мутаций. Однако некоторые сочетания были представлены в недостаточной мере. Следовательно, для следующего раунда эволюции TMCA для применения в качестве матриц отбирали некоторые новые конструкции из первого раунда эволюции TMCA. Использованные матрицы перечислены ниже в таблице 25.
Сочетания матриц и олигонуклеотидов, использованные во втором раунде эволюции TMCA, перечислены ниже в таблице 26. Для каждой строки в таблице 26A проводили отдельную реакцию TMCA и создавали отдельную библиотеку для каждой строки. Например, один цикл состоял из матрицы F8 и праймеров 2For и 7Rev. Второй цикл состоял из матрицы F8 и праймеров 2For и 6 Rev. Следует отметить, что использовали несколько тех же праймеров, что и в первом раунде. Два дополнительных праймера предназначены для второго раунда ПЦР. Эти олигонуклеотиды имеют названия 116LF и 162F. Олигонуклеотид 116LF содержит обратную и комплементарную олигонуклеотиду 4-Alt2-RevJC последовательность. Олигонуклеотид 162F содержит обратную и комплементарную олигонуклеотиду 5-RevJC последовательность.
Образцы амплифицировали в векторе pDOW-kan, анализировали в агарозном геле и обрабатывали с использованием DPNI. Образцы трансформировали в компетентные клетки E. coli XL1-Blue. Колонии собирали, растили и секвенировали. Посредством секвенирования установили, что 7 из 9 желаемых конструкций представлены в достаточно высоком количестве. Однако поскольку другие две конструкции представлены в недостаточной мере, то требовался дополнительный раунд эволюции TMCA для получения остальных лидирующих термостабильных пальмитаз в сочетании с семью молчащими мутациями. Для третьего раунда эволюции TMCA в качестве матриц использовали две новые конструкции. Эти матрицы перечислены ниже в таблице 27. Сочетания матриц и олигонуклеотидов, использованных в третьем раунде, перечислены ниже в таблице 28. Для каждой строки в таблице 28A проводили отдельные реакции TMCA и создавали для каждой строки отдельную библиотеку. Например, один цикл состоял из матрицы F3 и праймеров 162F и 7Rev. Второй цикл состоял из матрицы B5 и праймеров 162F и 6Rev.
Образцы амлифицировали в векторе pDOW-kan, анализировали в агарозном геле и обрабатывали с использованием DpnI. Образцы трансформировали в компетентные клетки E. coli XL1-Blue.
Колонии собирали, растили и секвенировали. Добились представления остальных двух конструкций в достаточном количестве. ДНК для всех желаемых конструкций E. coli XL1-Blue получали и трансформировали в компетентные клетки Pseudomonas fluorescens. Клетки растили в среде LB с добавлением урацила (750 нг/мл) и канамицина (50 мкг/мл). Образцы подтверждали секвенированием.
Для скрининга библиотеку растили в минимальной среде M9 с добавлением урацила (750 нг/мл) и канамицина (50 нг/мл). Стандартные анализы с маслом проводили посредством загрузки осветленных лизатов при содержании воды 10% с использованием фосфатного буфера в анализы с нерафинированным маслом при уровнях активности в отношении УМБ-пальмитата, перечисленных в левой колонке таблицы 29. Анализы с маслом загружали, гомогенизировали и впоследствии непрерывно смешивали, как для стандартных реакций, при температуре 25°C, 45°C и 60°C. Аликвоты 11% NaOH по 50 мкл добавляли через 3 и 20 ч для каждой впоследствии гомогенизированной реакции с маслом. Аликвоты брали через 20, 44 и 68 часов и подвергали стандартной экстракции хлороформом:метанолом и общему метанолизу с последующим анализом СМЭЖК посредством ГХ (как описано выше в примере 8). Для каждой аликвоты в таблице 29 приведено содержание пальмитата, оставшегося в масле. Термостабильный хит с молчащей мутацией 29 SM кардинально снижал содержание пальмитата на всем протяжении времени анализа при 25°C (таблица 29).
Пример 13: Калиевая соль олеиновой кислоты в качестве эмульгатора позволяет ферменту создавать масло с содержанием пальмитата 1%
Образцы масла инкубировали с 10% олеатом калия для оценки влияния этого эмульгатора на реакции масла с пальмитазой. В анализы с предварительно обработанным маслом (A и B) и нерафинированным маслом (c) загружали осветленные лизаты хита 29 SM или отрицательный контроль при содержании воды 5%. Предварительную обработку осуществляли посредством ферментативной обработки, нагревания, центрифугирования, а затем фильтрования масла для удаления смолы и водной фазы и для снижения содержания свободных жирных кислот, как описано в примере 14, для создания масла, не содержащего смолу, с содержанием пальмитата менее 5%. Масло гомогенизировали до и после добавления фермента, чтобы обеспечить гомогенность эмульсий. В 5 г масла отвешивали 0,5 г калиевой соли олеиновой кислоты, полученной в коммерческом источнике, перемешивали при 60°C для растворения, а затем гомогенизировали. Щелочную добавку для нейтрализации 1% FFA добавляли в реакции, помеченные как «щелочь». В анализы с маслом загружали фермент, повторно гомогенизировали и впоследствии непрерывно смешивали как для стандартных реакций. Через 3, 24, 48, 72 и 120 ч брали аликвоты. Аликвоты подвергали экстракции хлороформом:метанолом и общему метанолизу с последующим анализом СМЭЖК посредством ГХ (как описано выше в примере 8). Для каждой аликвоты пальмитат, оставшийся в масле, приведен ниже в таблице 30. В предварительно обработанных маслах уровень пальмитата успешно снизили до 1%.
Пример 14: использование промежуточного центрифугирования и эмульсификации олеатом позволяет получить масло с содержанием пальмитата 1% в 2 кг масштабе
Комбинированное влияние полипептида, промежуточной стадии фильтрования и добавления олеата калия для получения масла с содержанием пальмитата 1% оценивали в лабораторном масштабе. Проводили реакцию 2 кг нерафинированного масла с 29 SM при содержании воды 5%. Реакционную смесь с маслом гомогенизировали в течение 1 минуты с помощью IKA на 6-й скорости, достигая температуры 25°C. Навесной смеситель оборудовали лопаткой смесителя для краски и перемешивали реакционную смесь с маслом со скоростью 390 об/мин. После протекания реакции в течение 1 ч скорость смешивания увеличивали до 530 об/мин. Следили за температурой реакционной смеси, которая колебалась от 21°C до 29°C, при периодическом нагревании с помощью нагревательного блока. Образцы брали, чтобы следить за изменениями в профиле масла. После 28 ч реакционную смесь с маслом нагревали до 66°C и центрифугировали в гиротестере. Фракцию масла охлаждали до комнатной температуры в течение ночи при перемешивании мешалкой, затем охлаждали в ледяной ванне менее чем до 10°C. Добавляли диатомовую землю и отделяли ТАГ и ДАГ от свободных жирных кислот посредством фильтрования через охлажденную бюхнеровскую воронку с ватманской бумагой.
1 кг этого масла со сниженным содержанием пальмитата использовали для инициации реакций, используя олеат калия в качестве эмульгатора. В масло добавляли 100 г олеата калия. Реакционные смеси нагревали до 60°C при перемешивании, чтобы обеспечить растворимость. Масло охлаждали до 23°C, затем добавляли 50 мл фермента для достижения содержания воды 5%. Это масло гомогенизировали до и после добавления фермента 29 SM, чтобы обеспечить равномерные эмульсии. Аликвоты брали через 6, 21, 24, 28 ч, после фильтрования, после добавления олеата, после начала нового анализа и через 3, 20, и 22 ч во втором анализе. Аликвоты подвергали стандартной экстракции хлороформом:метанолом и общему метанолизу с последующим анализом СМЭЖК посредством ГХ (как описано выше в примере 8). Жирные кислоты, оставшиеся связанными с маслом, перечислены для каждой аликвоты ниже в таблице 31. Хит 29 SM способен снижать содержание пальмитата в масле до 1%.
Пример 15: Двойное центрифугирование и фильтрование дают масло с низким содержанием пальмитата
Ниже продемонстрирована способность отделять олеат калия после реакций с маслом. Проводили реакцию 2 кг нерафинированного масла и 29 SM при содержании воды 5%. Реакционную смесь с маслом гомогенизировали в течение 1 минуты с помощью IKA на 6-й скорости, достигая температуры 25°C. Навесной смеситель оборудовали лопаткой смесителя для краски и перемешивали реакционную смесь с маслом со скоростью 390 об/мин. После протекания реакции в течение 1 скорость смешивания увеличивали до 530 об/мин. Следили за температурой реакционной смеси, которая колебалась от 21°C до 29°C, при периодическом нагревании с помощью нагревательного блока. Образцы брали, чтобы следить за изменениями в профиле масла. После 28 ч реакционную смесь с маслом нагревали до 80°C и центрифугировали в гиротестере. Фракцию масла охлаждали до комнатной температуры в течение ночи при перемешивании мешалкой, затем охлаждали в ледяной ванне менее чем до 10°C. Добавляли диатомовую землю и отделяли ТАГ и ДАГ от свободных жирных кислот посредством фильтрования через охлажденную бюхнеровскую воронку с ватманской бумагой. 1 кг этого обработанного масла использовали для инициации реакций, используя олеат калия в качестве эмульгатора. В масло добавляли 50 г олеата калия для достижения 5%. Это перемешивали и гомогенизировали. 50 мл фермента загружали, чтобы достичь содержания воды 5%, и гомогенизировали в течение 1 минуты с помощью IKA на 6-й скорости, достигая температуры 26,6°C. Эту реакционную смесь с маслом перемешивали с использованием навесного смесителя, оборудованного лопаткой смесителя для краски, на скорости 400 об/мин. Использовали периодическое нагревание нагревательным блоком. Аликвоты брали через 3, 20, 24 и через 8, 28 и 48 часов после начала второго анализа. Аликвоты подвергали стандартной экстракции хлороформом:метанолом и общему метанолизу с последующим анализом СМЭЖК посредством ГХ (как описано выше в примере 8). Жирные кислоты, оставшиеся связанными в масле, перечислены для каждой аликвоты ниже в таблице 32. 29 SM снижал содержание пальмитата до 1,5%. Это масло нагревали до 85°C и центрифугировали для удаления остатков белков. Фракцию масла охлаждали до 6°C при перемешивании мешалкой, добавляли диатомовую землю и вещество разделяли на бюхнеровской воронке. Фильтрование продолжали при температуре 25°C.
Пример 16: Оптимизация кодонов
Версии с оптимизированными кодонами для экспрессии в Pseudomonas fluorescens лидирующего термостабильного хита с молчащей мутацией 29 SM (пример 12) конструировали двумя различными способами. В первой версии с оптимизированными кодонами заменяли все кодоны в хите 29 SM на кодоны, предпочтительные для Pseudomonas fluorescens, включая введенные лучшие 7 молчащих мутаций (пример 12). Эту версию назвали 29 SM-Pf (SEQ ID № 22). Предпочтительную частоту использования кодонов для Pseudomonas fluorescens определяли по обзору Базы данных о частоте использования кодонов, доступной в режиме он-лайн на веб-сайте Kazusa DNA Research Institute (2-6-7 Kazusa-kamatari, Kisarazu, Chiba 292-0818 JAPAN, получена в 2009 из интернета
Во второй версии с оптимизированными кодонами также заменяли все кодоны в хите 29 SM на кодоны, предпочтительные для Pseudomonas fluorescens, за исключением того, что сохранили 7 лидирующих молчащих мутаций (пример 11). Эту версию назвали 29 SM-Pf+SM (SEQ ID № 23). 29 SM-Pf+SM (SEQ ID № 23) является более предпочтительным кандидатом по сравнению с 29 SM-Pf (SEQ ID № 22).
SEQ ID № 22:
ATGCTCAAGCCCCCACCTTACGGCCGTCTGCTCCGCGAACTGGCTGATATCCC
GGCGATCGTGACTGCTCCGTTCCGCGGCGCAGCCAAAATGGGCAAACTGGCA
GATGGCGAGCCGGTACTGGTGCTGCCCGGCTTCCTGGCGGACGACAACGCGA
CCAGCGTGCTGCGGAAGACCTTCGAGGTCGCCGGCTTTGCGTGCAGCGGCTG
GGAAAAGGGCTTCAACCTCGGCATTCGTGGCGACCTCATGGACTACCTGGTCG
ACCGCCTGCGCGCCGTGAGCGAGGCCGCGGGGGGGCAGAAGGTAATCGTGG
TCGGCTGGTCCCTCGGCGGCCTCTACGCCCGGGAGTTGGGCCACAAGGCCCC
CGAACTGATCCGTATGGTCGTCACGCTCGGCTCCCCGTTCGCCGGCGACCTCC
ACGCGAACCATGCCTGGAAGATCTACGAGGCCATCAACTCCCACACGGTCGAC
AACCTGCCGATCCCGCGCGATTTCCAGATTAAGCCGCCGGTGCATACCATCGC
CGTGTGGAGCCCGCTCGACGGGGTGGTGGCCCCGGAGACGAGCGAAGGCAG
CCCCGAGCAGAGCGACGAGCGCTTGGAGCTGGCCGTGACCCACATGGGCTTT
GCGGCTAGCAAGACCGGGGCGGAGGCAGTGGTCCGCCTGGTCGCCGCCCGC
CTCTGA
SEQ ID № 23:
ATGCTCAAGCCCCCACCTTACGGCCGTCTGCTCCGCGAACTGGCTGATATCCC
GGCGATCGTGACTGCTCCGTTCCGCGGCGCAGCCAAAATGGGCAAACTGGCT
GATGGCGAGCCGGTACTGGTGCTGCCCGGCTTCCTGGCGGACGACAACGCGA
CCAGCGTGCTGCGGAAGACCTTCGAGGTCGCCGGCTTTGCGTGCAGCGGCTG
GGAAAAGGGCTTCAACCTCGGCATTCGTGGCGACCTCATGGACTACCTGGTCG
ACCGCCTGCGCGCCGTGAGCGAGGCCGCGGGGGGGCAGAAGGTTATCGTGG
TCGGCTGGAGTCTCGGCGGCCTCTACGCCCGGGAGCTTGGCCACAAGGCCCC
CGAACTGATCAGGATGGTCGTCACGCTCGGCTCTCCGTTCGCCGGCGACCTCC
ACGCGAACCATGCCTGGAAGATCTACGAGGCCATCAACTCCCACACGGTCGAC
AACCTGCCGATCCCGCGCGATTTCCAGATTAAGCCGCCGGTGCATACCATCGC
CGTGTGGAGCCCGCTCGACGGGGTGGTGGCCCCGGAGACGAGCGAAGGCAG
CCCCGAGCAGAGCGACGAGCGCTTGGAGCTGGCCGTGACCCACATGGGCTTT
GCGGCTAGCAAGACCGGGGCGGAGGCAGTGGTCCGCCTGGTCGCCGCCCGC
CTCTGA
Последовательности обеих версий с оптимизированными кодонами отсылали в DNA 2.0 Incorporated (Menlo Park, CA) для синтеза ДНК. Гены синтезировали в векторе pJ201 с участками рестрикции SpeI и XhoI с обеих сторон гена. После доставки синтезированных генов плазмиды резали рестрикционными ферментами. ДНК вектора pDOW-Kan также резали теми же рестрикционными ферментами и затем обрабатывали щелочной фосфатазой из кишечника теленка (New England Biolabs, № продукта M0290L). Все образцы очищали в геле и экстрагировали с использованием QIAquick gel extraction kit (Qiagen, № продукта 28706). Затем вектор и вставки лигировали с использованием Roche Rapid Ligation kit (Roche, № продукта 11635379001). Затем продукты лигировани трансформировали в компетентные клетки E. coli XL1-Blue. Колонии собирали, растили и получали ДНК с использованием Qiagen mini-prep kit. Подтверждали последовательности образцов ДНК, а затем использовали ДНК для трансформации Pseudomonas fluorescens.
Затем 29 SM-Pf (SEQ ID № 22), 29 SM-Pf+SM (SEQ ID № 23), 29 SM, родительскую последовательность (SEQ ID № 2 с мутациями D61E, R72K и V163R) и организм-хозяин/векторный контроль растили и индуцировали в 250 мл встряхиваемых колбах. Концентрацию в осветленных лизатах определяли с использованием Bio-Rad Protein Analysis (Bio-Rad, Hercules, CA, по каталогу № 500-0006) на основе способа Брэдфорда, а затем нормализовали по экспрессии общего белка. Экспрессию анализировали посредством SDS-PAGE, причем на каждую дорожку геля SDS-PAGE загружали равное количество общего белка. 29 SM-Pf и 29 SM-Pf+SM обладали большей процентной долей экспрессированного белка пальмитазы по сравнению с 29 SM и родительским белком. 29 SM-Pf+SM обладал слегка повышенной процентной долей экспрессии пальмитазы, чем 29 SM-Pf. Следовательно, в присутствие молчащих мутаций усовершенствование экпрессии выражено несколько больше.
Описаны многие варианты осуществления изобретения. Тем не менее, следует понимать, что можно выполнить различные модификации, не отступая от духа и объема изобретения. Таким образом, другие варианты осуществления входят в объем следующей формулы изобретения.
Реферат
Изобретение относится к области биотехнологии и представляет собой полипептиды, обладающие пальмитазной активностью. Изобретение также раскрывает способы гидролиза масел и жиров с низким содержанием насыщенных жирных кислот или с низким содержанием транс-изомеров с использованием указанных пальмитаз, способы биокаталитического синтеза липидов с использованием указанных пальмитаз, а также использование указанных пальмитаз для катализа реакции переэтерификации и способы получения триацилглицеридов (ТАГ), диацилглицеридов (ДАГ) и продуктов питания с использованием указанных реакций. Настоящее изобретение позволяет расширить ассортимент пальмитаз и использовать их для катализа реакций гидролиза. 12 н. и 44 з.п. ф-лы, 32 ил., 32 табл., 16 пр.
Формула
(а) получение композиции, содержащей масло или жир, где масло или жир можно гидролизовать посредством полипептида, обладающего гидролазной активностью, указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих гидролазной активностью, и указанный полипептид или
i) имеет аминокислотную последовательность, как указано в SEQ ID №2, но также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R, или
ii) имеет аминокислотную последовательность, как указано в SEQ ID №2, но также имеет одну модификацию аминокислотного остатка I20L; V62S; G77P; V83C; D88H; Y113G; Е116Т; E116G; Н140K; K146S; I167S; L180E; Е194М; A211Q; S212Y; G215C; G215V; G215W; А218Н; A218S; V223A; А225М; или A225Q;
(b) добавление полипептида со стадии (а) в композицию, содержащую указанное масло или жир, в достаточном количестве и в достаточных условиях для того, чтобы вызвать гидролиз масла или жира, посредством чего гидролизуют масло или жир.
(а) получение полипептида, обладающего пальмитазной активностью, причем указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих пальмитазной активностью, и указанный полипептид имеет аминокислотную последовательность, как указано в SEQ ID №2, но также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R;
(b) получение композиции, содержащей триацилглицерид (ТАГ);
(c) приведение полипептида со стадии (а) в контакт с композицией со стадии (b) в условиях, в которых полипептид гидролитически отщепляет остаток пальмитиновой кислоты в положении Sn2 триацилглицерида (ТАГ), посредством чего получают 1,3-диацилглицерид (ДАГ);
(d) получение R1-сложного эфира;
(e) получение R1-специфической гидролазы, и
(f) приведение 1,3-ДАГ со стадии (с) в контакт с R1-сложным эфиром со стадии (d) и R1-специфической гидролазой со стадии (е) в условиях, в которых R1-специфическая гидролаза катализирует этерификацию положения Sn2, посредством чего получают структурированный липид.
(а) получение полипептида, обладающего пальмитазной активностью, указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих пальмитазной активностью, и указанный полипептид имеет аминокислотную последовательность, как указано в SEQ ID №2, но также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R;
(b) получение композиции, содержащей триацилглицерид (ТАГ);
(c) приведение полипептида со стадии (а) в контакт с композицией со стадии (b) в условиях, в которых полипептид гидролитически отщепляет остаток пальмитиновой кислоты в положении Sn1 или Sn3 триацилглицерида (ТАГ), посредством чего получают 1,2-ДАГ или 2,3-ДАГ; и
(d) промотирование перегруппировке остатка карбоновой кислоты в 1,2-ДАГ или 2,3-ДАГ со стадии (с) в кинетически контролируемых условиях, посредством чего получают 1,3-ДАГ.
(а) получение композиции, содержащей полипептид, обладающий 1,3-специфической липазной активностью, указанный полипептид выбран из группы, состоящей из выделенных, синтетических или рекомбинантных полипептидов, обладающих пальмитазной активностью, и указанный полипептид имеет аминокислотную последовательность, как указано в SEQ ID №2, но также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R;
(b) получение смеси триацилглицеридов и свободных жирных кислот;
(c) обработка композиции со стадии (b) полипептидом в условиях, в которых полипептид может катализировать обмен свободных жирных кислот и групп пальмитиновой кислоты триацилглицеридов, посредством чего получают новые триацилглицериды, обогащенные указанными жирными кислотами.
(a) получение реакционной смеси для переэтерификации, которая содержит вещество источника стеариновой кислоты, выбранное из группы, состоящей из стеариновой кислоты, сложных стеариновых моноэфиров низкомолекулярных одноатомных спиртов и их смесей;
(b) получение продукта питания, корма или масла, содержащего триацилглицерид;
(c) получение полипептида, обладающего гидролазной активностью, указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих гидролазной активностью, и указанный полипептид или
i) имеет аминокислотную последовательность, как указано в SEQ ID №2, но также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R, или
ii) имеет аминокислотную последовательность, как указано в SEQ ID №2, но также имеет одну модификацию аминокислотного остатка I20L; V62S; G77P; V83C; D88H; Y113G; Е116Т; E116G; Н140K; K146S; I167S; L180E; Е194М; A211Q; S212Y; G215C; G215V; G215W; А218Н; A218S; V223A; А225М; или A225Q;
(d) переэтерификация вещества источника стеариновой кислоты и триацилглицерида из продукта питания, корма или масла; и
(е) отделение компонентов свободных жирных кислот от переэтерифицированных глицеридных компонентов смеси для переэтерификации, чтобы предоставить переэтерифицированный продукт масла и смесь жирных кислот, содержащую жирные кислоты, сложные моноэфиры жирных кислот или их смеси, высвобожденные из продукта питания, корма или масла.
(a) получение композиции масел, содержащей некоторое количество ТАГ;
(b) получение полипептида, обладающего гидролазной активностью, указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих гидролазной активностью, и указанный полипептид или
i) имеет аминокислотную последовательность, как указано в SEQ ID №2, но также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R, или
ii) имеет аминокислотную последовательность, как указано в SEQ ID №2, но также имеет одну модификацию аминокислотного остатка I20L; V62S; G77P; V83C; D88H; Y113G; Е116Т; E116G; H140K; K146S; I167S; L180E; Е194М; A211Q; S212Y; G215C; G215V; G215W; А218Н; A218S; V223A; А225М; или A225Q;
(с) приведение указанной композиции масел со стадии (а) в контакт с указанным полипептидом со стадии (b) в условиях, достаточных для гидролитического отщепления полипептидом остатка карбоновой кислоты на ТАГ для получения ДАГ.
(а) получение композиции, содержащей масло или жир, где масло или жир можно гидролизовать посредством полипептида, обладающего пальмитазной активностью, причем указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих пальмитазной активностью, и указанный полипептид имеет аминокислотную последовательность, как указано в SEQ ID №2, и мутации D61E, R72K и V163R, и также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R;
(b) добавление полипептида со стадии (а) в композицию, содержащую указанное масло или жир, в достаточном количестве и в достаточных условиях для того, чтобы вызвать гидролиз масла или жира, посредством чего гидролизуют масло или жир.
(а) получение полипептида, обладающего пальмитазной активностью, причем указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих пальмитазной активностью, и указанный полипептид имеет аминокислотную последовательность, как указано в SEQ ID №2, и мутации D61E, R72K и V163R, и также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R;
(b) получение композиции, содержащей триацилглицерид (ТАГ);
(c) приведение полипептида со стадии (а) в контакт с композицией со стадии (b) в условиях, в которых полипептид гидролитически отщепляет пальмитазный остаток карбоновой кислоты в положении Sn2 триацилглицерида (ТАГ), посредством чего получают 1,3-диацилглицерид (ДАГ);
(d) получение R1-сложного эфира;
(e) получение R1-специфической гидролазы, и
(f) приведение 1,3-ДАГ со стадии (с) в контакт с R1-сложным эфиром со стадии (d) и R1-специфической гидролазой со стадии (е) в условиях, в которых R1-специфическая гидролаза катализирует этерификацию положения Sn2, посредством чего получают структурированный липид.
(а) получение полипептида, обладающего пальмитазной активностью, причем указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих пальмитазной активностью, и указанный полипептид имеет аминокислотную последовательность, как указано в SEQ ID №2, и мутации D61E, R72K и V163R, и также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R;
(b) получение композиции, содержащей триацилглицерид (ТАГ);
(c) приведение полипептида со стадии (а) в контакт с композицией со стадии (b) в условиях, в которых полипептид гидролитически отщепляет пальмитазный остаток карбоновой кислоты в положении Sn1 или Sn3 триацилглицерида (ТАГ), посредством чего получают 1,2-ДАГ или 2,3-ДАГ; и
(d) промотирование перегруппировке остатка карбоновой кислоты в 1,2-ДАГ или 2,3-ДАГ со стадии (с) в кинетически контролируемых условиях, посредством чего получают 1,3-ДАГ.
(а) получение композиции, содержащей полипептид, обладающий 1,3-специфической липазной активностью, указанный полипептид выбран из группы, состоящей из выделенных, синтетических или рекомбинантных полипептидов, обладающих пальмитазной активностью, и указанный полипептид имеет аминокислотную последовательность, как указано в SEQ ID №2, и мутации D61E, R72K и V163R, и также имеет модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R;
(b) получение смеси триацилглицеридов и свободных жирных кислот;
(c) обработка композиции со стадии (b) полипептидом в условиях, в которых полипептид может катализировать обмен свободных жирных кислот и ацильных групп триацилглицеридов, посредством чего получают новые триацилглицериды, обогащенные указанными жирными кислотами.
(a) получение реакционной смеси для переэтерификации, которая содержит вещество источника стеариновой кислоты, выбранное из группы, состоящей из стеариновой кислоты, сложных стеариновых моноэфиров низкомолекулярных одноатомных спиртов и их смесей;
(b) получение продукта питания, корма или масла, содержащего триацилглицерид;
(c) получение полипептида, обладающего пальмитазной активностью, указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих пальмитазной активностью, и указанный полипептид имеет аминокислотную последовательность, как указано в SEQ ID №2, и мутации D61E, R72K и V163R, и также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R;
(d) переэтерификация вещества источника стеариновой кислоты и триацилглицерида продукта питания, корма или масла; и
(e) отделение компонентов свободных жирных кислот от переэтерифицированных глицеридных компонентов смеси для переэтерификации, чтобы предоставить переэтерифицированный продукт масла и смесь жирных кислот, содержащую жирные кислоты, сложные моноэфиры жирных кислот или их смеси, высвобожденные из продукта питания, корма или масла.
(a) получение композиции масел, содержащей некоторое количество ТАГ;
(b) получение полипептида, обладающего пальмитазной активностью, причем указанный полипептид выбран из группы, состоящей из выделенных, синтетических и рекомбинантных полипептидов, обладающих пальмитазной активностью, и указанный полипептид имеет аминокислотную последовательность, как указано в SEQ ID №2, и мутации D61E, R72K и V163R, и также имеет одну модификацию аминокислотного остатка D61A; D61E; R72E; R72K; Е116А; E116Q; E116R; Е116Т; E116V; S133A; I151G; I151A; V163R; или D164R;
(с) приведение указанной композиции масел со стадии (а) в контакт с указанным полипептидом со стадии (b) в условиях, достаточных для гидролитического отщепления полипептидом пальмитазного остатка карбоновой кислоты на ТАГ для получения ДАГ.
Комментарии