Выделение стволовых клеток/клеток-предшественников из амниотической мембраны пуповины - RU2416638C2
Код документа: RU2416638C2
Чертежи
































































Описание
Настоящее изобретение относится к способу извлечения стволовых/прародительских клеток из амниотической мембраны пуповины, причем способ включает отделение амниотической мембраны от других компонентов пуповины in vitro, культивирование ткани амниотической мембраны в условиях, обеспечивающих пролиферацию клеток, и извлечение стволовых/прародительских клеток из культур такни. В частности, изобретение относится к извлечению и культивированию стволовых клеток, имеющих эмбрионные свойства, таких как эпителиальные и/или мезенхимные стволовые/прародительские клетки, в условиях, позволяющих клеткам претерпевать митотическую экспансию. Кроме того, изобретение нацелено на способ дифференцировки извлеченных стволовых/прародительских клеток в эпителиальные и/или мезенхимные клетки и на терапевтические применения таких стволовых/прародительских клеток.
Уровень техники
Стволовые клетки - этот популяция клеток, обладающих способностью бесконечно самообновляться и дифференцироваться в разнообразные типы клеток или тканей. Эмбрионные стволовые клетки (приблизительно через приблизительно 3-5 дней после оплодотворения) бесконечно пролиферируют и могут спонтанно дифференцироваться во все типы тканей: поэтому их называют множественно потенциальными (плюрипотентными) стволовыми клетками (обзор по ним приведен, например, в работе Smith A.G. // Annu. Rev. Cell. Dev. Biol. 2001. Т.17. С.435-462). Стволовые клетки взрослых, однако, более тканеспецифичны и могут быть менее способны к репликации: поэтому их называют многопотенциальными (мультипотентными) стволовыми клетками (обзор по ним приведен, например, в работе Paul G. и др. // Drug Discov. Today. 2002. Т.7. С.295-302). В основе «пластичности» эмбрионных и взрослых стволовых клеток лежит их способность трансдифференцироваться в ткани иного, чем у них, происхождения и, возможно, через слои зачатка зародыша.
Способность стволовых клеток самообновляться необходима для их функционирования в качестве источника незрелых, недифференцированных клеток. Напротив, большинство соматических клеток имеют ограниченную способность к самообновлению вследствие укорачивания теломер (обзор см., например, в работе Dice J.F. // Physiol. Rev. 1993. Т.73. С.149-159). Поэтому способы терапии с использованием стволовых клеток могут быть полезны для лечения множества заболеваний человека и животных.
Стволовые клетки, а также стволовые/прародительские клетки можно получить из различных источников. Потенциал «множественности линий дифференцировки» эмбрионных и взрослых стволовых клеток был широко охарактеризован. Даже при том, что потенциал эмбрионных стволовых клеток необычайно широк, их применение заключает в себе множество этических проблем. Поэтому в качестве альтернативных источников были предложены неэмбрионные стволовые клетки, полученные из костного мозга, стромы, жировой ткани, дермы и крови пуповины. Эти клетки могут дифференцироваться in vitro, среди прочего, в хондроциты, адипоциты, остеобласты, миобласты, кардиомиоциты, астроциты и теноциты и претерпевают дифференцировку in vivo, что делает эти стволовые клетки (обычно называемые мезенхимными стволовыми клетками) многообещающими кандидатами для исправления дефектов мезодермы и устранения заболевания.
Однако при клиническом применении собирание таких мезенхимных стволовых клеток приводит к нескольким проблемам. Отбор таких клеток представляет собой для пациента психическую и физическую нагрузку, поскольку для получения таких клеток необходима хирургическая операция (например, отбор костного мозга - это инвазивная процедура, осуществляемая с помощью иглы для биопсии и требующая местного или даже общего наркоза). Кроме того, во многих случаях число извлеченных стволовых клеток довольно невелико. Более важно то, что из этих клеток не получаются или не дифференцируются клетки эпителия. Это побуждает к поиску других возможных источников стволовых клеток.
Было установлено, что кровь пуповины является богатым источником кроветворных стволовых/прародительских клеток. Однако существует полемика относительно существования мезенхимных стволовых/прародительских клеток. С одной стороны, такие клетки нельзя успешно извлечь или успешно культивировать из крови пуповиного канатика новорожденного (Mareschi К. и др. // Haematologica. 2001. Т.86. С.1099-1100). В то же самое время результаты, полученные Campagnoli С. и др. (II Blood. 2001. Т.98. С.2396-2402) и Erices А. и др. (II Br. J. Haematol. 2000. Т.109. С.235-242) позволяют предположить, что мезенхимные стволовые клетки имеются в некоторых органах плода и циркулируют в крови готовящихся к рождению плода вместе с кроветворными предшественниками. В соответствии с этим международная публикация WO 03/070922 раскрывает способы извлечения и культивирования мезенхимных стволовых/прародительских клеток из крови пуповины и способ дифференцировки таких клеток в различные мезенхимные ткани. Была описана эффективность извлечения около 60% (Bieback К. и др. // Stem Cells. 2004. Т.22. С.625-634). В этом же исследовании в качестве критических параметров для достижения такого выхода были определены и период времени от сбора крови пуповины до извлечения клеток, и используемый объем образца крови. Однако остается под вопросом, действительно ли эти стволовые/прародительские клетки происходят из ткани пуповины.
Недавно мезенхимные стволовые/прародительские клетки были успешно выделены из ткани пуповины, а именно из вартонова студня, матрикса пуповины (Mitchell K.E. и др. // Stem Cells. 2003. Т.21. С.50-60; патент США №5919702; заявка на патент США №2004/0136967). Было установлено, что эти клетки способны дифференцироваться, например, в клетки с фенотипом нейронов или, соответственно, в ткань хряща. Кроме того, мезенхимные стволовые/прародительские клетки были также извлечены из эндотелия и субэндотелиального слоя вены пуповины, одного из трех кровеносных сосудов (двух артерий и одной вены), находящихся внутри пуповины (Romanov Y.A. и др. // Stem Cells. 2003. Т.21. С.105-110; Covas D.T. и др. // Braz. J. Med. Biol. Res. 2003. Т.36. С.1179-1183).
Однако ни один из этих разработанных до сих пор подходов не привел к выделению или культивированию эпителиальных стволовых/прародительских клеток как источника для основанных на эпителиальных клетках лечебных приемов, таких как модификация поверхности кожи, восстановление печени, реконструкция ткани мочевого пузыря и других восстанавливаемых поверхностных тканей. Таким образом, все еще существует потребность в способах и надежных источниках, пригодных для извлечения и культивирования эпителиальных стволовых/прародительских клеток. Кроме того, для получения таких клеток в достаточных количествах для различных применений в восстановительной медицине и создании тканей все еще необходимы быстрые и эффективные способы извлечения эпителиальных и мезенхимных стволовых/прародительских клеток, применимые с этической точки зрения и не создающие биомедицинских трудностей для пациента.
Сущность изобретения
Изобретение предусматривает способ извлечения из амниотической мембраны пуповины стволовых/прародительских клеток, причем этот способ включает:
(а) отделение амниотической мембраны от других компонентов пуповины in vitro;
(б) культивирование полученной в этапе (а) ткани амниотической мембраны в условиях, обеспечивающих пролиферацию клеток; и
(в) извлечение стволовых/прародительских клеток.
В одном из вариантов осуществления изобретение предусматривает способ, дополнительно включающий:
(а") отделение клеток перед культивированием от ткани амниотической мембраны с помощью технических приемов, выбранных из группы, состоящей из ферментативного расщепления и прямого культивирования ткани эксплантата.
В одном из предпочтительных вариантов осуществления изобретение предусматривает способ извлечения стволовых/прародительских клеток, имеющих свойства, подобные свойствам эмбрионных стволовых клеток.
В другом предпочтительном варианте осуществления изобретение предусматривает способ извлечения эпителиальных и/или мезенхимных стволовых/прародительских клеток.
В ином варианте осуществления изобретение предусматривает способ, дополнительно включающий:
(г) культивирование стволовых/прародительских клеток в условиях, которые дают возможность клеткам претерпевать клональную экспансию.
Еще в одном варианте осуществления изобретение предусматривает способ, дополнительно включающий:
(д) культивирование стволовых/прародительских клеток в условиях, обеспечивающих дифференцировку указанных клеток в эпителиальные клетки и/или мезенхимные клетки; и
(е) извлечение дифференцированных клеток.
Еще в одном варианте осуществления изобретение предусматривает способ, дополнительно включающий:
(ж) консервацию извлеченных стволовых/прародительских клеток для последующего использования.
Еще в одном дополнительном варианте осуществления изобретение предусматривает способ культивирования стволовых/прародительских клеток согласно настоящему изобретению, включающий:
получение эксплантата ткани из амниотической мембраны пуповины;
культивирование эксплантата ткани в подходящей среде для культивирования и при подходящих условиях культивирования в течение подходящего периода времени.
В других вариантах осуществления изобретение нацелено на терапевтические применения стволовых/прародительских клеток или клеточных экстрактов из них. Один из этих вариантов осуществления изобретения предусматривает способ лечения имеющего расстройство индивидуума, включающий введение индивидууму эффективного количества стволовых/прародительских клеток, извлеченных заявляемым способом из описанных выше способов. Другой вариант осуществления изобретения предусматривает соответствующую фармацевтическую композицию.
Краткое описание чертежей
Лучшему пониманию изобретения способствует детальное описание, которое следует рассматривать вместе с неограничивающими примерами и чертежами которых:
Фиг.1 показывает проращивание эпителиальных клеток из амниотической мембраны пуповины методом прямого культивирования ткани эксплантата (увеличение 40 х) на 2-й день (фиг.1А) и на 5-й день (фиг.1Б, В) культивирования ткани. Пластиковые поверхности для культивирования клеток перед нанесением на их поверхность амниотической мембраны покрывали смесями коллаген-1/коллаген-4 (1:2; Becton Dickinson). Образцы амниотической мембраны погружали в 5 мл среды EpiLife или среды Medium 171 (обе от фирмы Cascade Biologies). Среду сменяли через каждые 2 или 3 дня, мониторинг прорастания клеток из эксплантата осуществляли с помощью световой микроскопии. Микрофотографии делали через различные интервалы времени, как указано выше. Наблюдавшаяся многоугольная (полиэдральная) морфология типична для эпителиальных клеток.
Фиг.2 описывает ферментативное расщепление сегментов пуповины, дающее такие же эпителиальные клетки (увеличение 40 х) на 2-й день (фиг.А и В) и на 5-й день (фиг.Б и Г). Амниотическую мембрану пуповины разделяли на маленькие кусочки размером 0,5 см × 0,5 см и переваривали в 0,1% растворе (вес к объему) коллагеназы 1-го типа (Roche Diagnostics) при 37°С в течение 8 ч. Образцы перемешивали во встряхивателе в течение 3 мин каждые 30 мин. Клетки собирали центрифугированием при 4000 об/мин в течение 30 мин. Осадки клеток ресуспендировали в среде EpiLife или среде Medium 171 (обе от фирмы Cascade Biologies), дополненной 50 мкг/мл инсулино-подобного фактора роста 1 (IGF-1), 50 мкг/мл полученного из тромбоцитов фактора роста ВВ (PDGF-ВВ), 5 мкг/мл трансформирующего фактора роста β1 (TGF-β1) и 5 мкг/мл инсулина (все от фирмы R&D Systems), подсчитывали и высевали на чашки для культур клеток диаметром 10 см, предварительно покрытые смесями коллагена-1 и коллагена-4 (1:2; Becton Dickinson) при плотности посева 1×106 клеток на чашку. Через 24 ч прикрепившиеся клетки промывали теплым солевым раствором в фосфатном буфере (PBS) и заменяли культуральную среду средой EpiLife или средой Medium 171 (обе от фирмы Cascade Biologies). Среду сменяли через каждые 2 или 3 дня, мониторинг прорастания клеток осуществляли с помощью световой микроскопии. Микрофотографии делали через различные интервалы времени, как указано выше. Клетки снова имели типичную для эпителиальных клеток многоугольную морфологию.
Фиг.3 показывает проросшие мезенхимные клетки, эксплантированные из амниотической мембраны пуповины. Прорастание клеток (увеличение 40х) наблюдалось уже через 48 ч после помещения их в чашки для роста культур ткани при использовании в качестве среды для культивирования среды DMEM, дополненной 10% фетальной телячьей сывороткой (FCS) (фиг.3А, В). Эксплантаты погружали в 5 мл среды DMEM (Invitrogen), дополненной 10% фетальной бычьей сывороткой FBS (Hyclone) (DMEM/10% FBS). Среду сменяли через каждые 2 или 3 дня. Мониторинг прорастания клеток осуществляли с помощью световой микроскопии. Микрофотографии делали через различные интервалы времени. Для клеток была характерна веретенообразная морфология, и in vitro они мигрировали и распространялись и легко, и быстро, что очень похоже на фибробласты (фиг.3Б, Г).
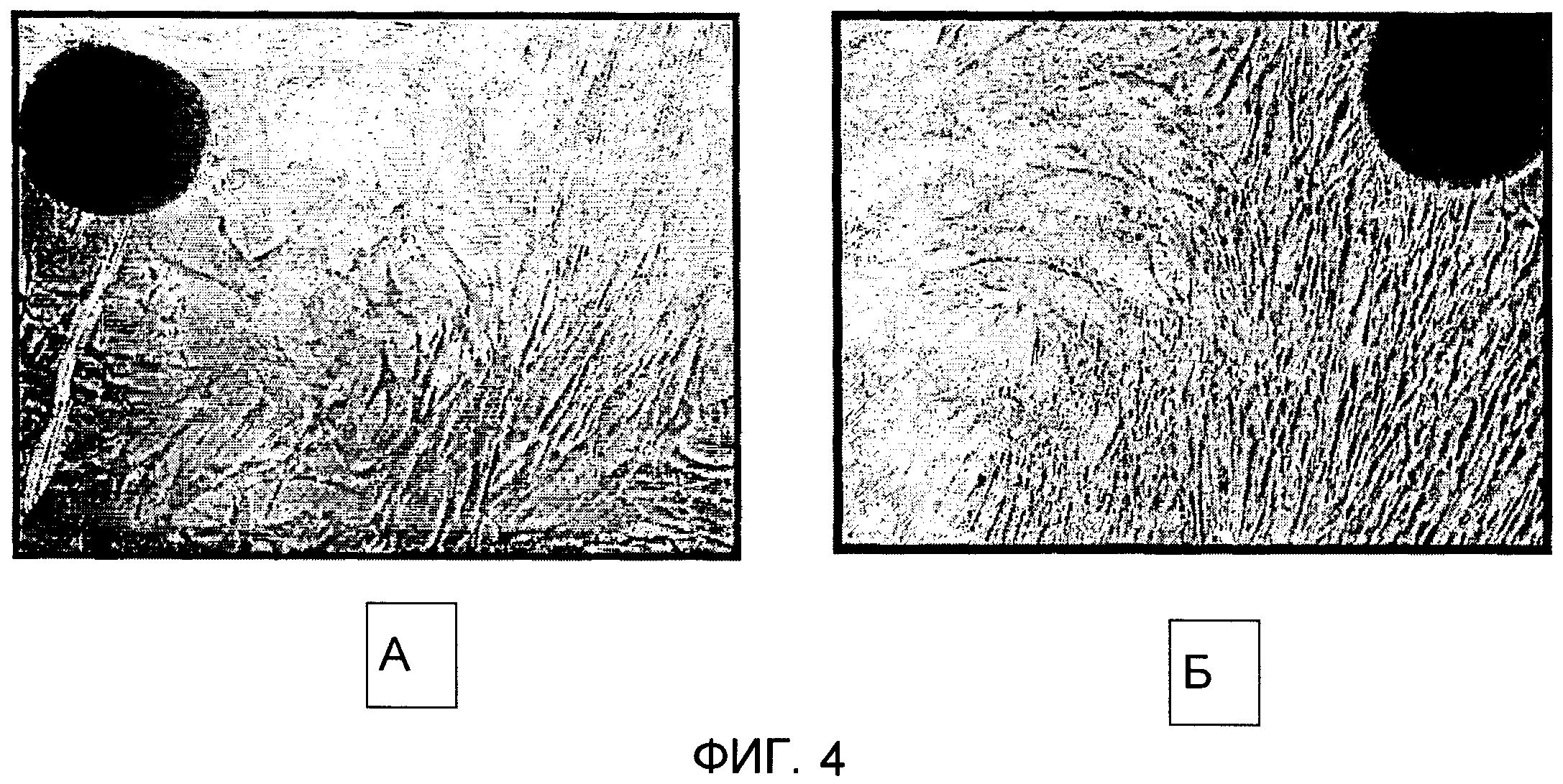
Фиг.4 (увеличение 40х) показывает мезенхимные клетки из клеток амниотической мембраны пуповины, извлеченные ферментативным расщеплением коллагеназой. Фиг.4А показывает мезенхимные клетки, извлеченные из амниотической мембраны пуповины на 2-й день. Пролиферация клеток наблюдалась на 5-й день (фиг.4Б). Амниотическую мембрану пуповины разделяли на маленькие кусочки размером 0,5 см ×0,5 см и переваривали в 0,1% растворе (вес к объему) коллагеназы 1-го типа (Roche Diagnostics) при 37°С в течение 6 ч. Образцы перемешивали во встряхивателе в течение 2 мин каждые 15 мин. Клетки собирали центрифугированием при 4000 об/мин в течение 30 мин. Осадки клеток ресуспендировали в среде DMEM/10% FBS, подсчитывали и высевали на чашки для культур ткани диаметром 10 см при плотности посева 1×106 клеток на чашку. Среду сменяли через каждые 2 или 3 дня. Мониторинг прорастания клеток осуществляли с помощью световой микроскопии. Микрофотографии делали через различные интервалы времени. Снова клетки обнаруживали веретенообразную морфологию, типичную для мезенхимных клеток в состоянии фибробластов.
Фиг.5 (увеличение 40х) описывает морфологию в культуральной среде без сыворотки (DMEM) и при культивировании с сывороткой (DMEM/10% FCS) мезенхимных клеток амниотической мембраны пуповины (umbilical cord mesenchymal cells, UCMC, фигуры 5Д, Е, Ж, З), извлеченных согласно способу в соответствии с настоящим изобретением, нормальных фибробластов кожи (клетки (NF109, фиг.5А, Б) и полученных из жировой ткани мезенхимных клеток (adipose-derived mesenchymal cells, ADMC, фиг.5В, Г). На фиг.5 видны изменения в морфологии клеток NF и ADMC, культивируемых в условиях голодания по сыворотке (только DMEM), что сказывается в появлении уплощенных клеток и менее плотной цитоплазме в сравнении с условиями в богатой сывороткой среде (DMEM/10%FCS), где клетки более круглые с плотной цитоплазмой (фиг.5А, Б, В, Г). Изменений в морфологии обеих групп UCMC, культивируемых в идентичных условиях в среде без сыворотки и в среде, богатой сывороткой (фиг.5Д, Е, Ж, З), обнаружено не было, что свидетельствует об ином поведении и иной морфологии этих последних мезенхимных клеток.
Фиг.6 (увеличение 40х) показывает извлеченные согласно настоящему изобретению UCMC, культивированные в DMEM/10%FCS без питающего слоя клеток 3Т3, на 3-й день и 7-й день. Видно, что клетки растут хорошо и формируют колонию (вертикальный рост) вместо радиального распространения. Это снова указывает на отличие в поведении этих мезенхимных клеток по сравнению с их более дифференцированными двойниками.
Фиг.7 (увеличение 40х) описывает формирование колоний эпителиальных клеток из пуповины (umbilical cord epithelial cells, UCEC), культивированных на питающем слое клеток 3T3, на 3-й и 7-й день. Их вид подобен виду полученных из нормальной кожи стволовых клеток эпителиальных кератиноцитов. У последних питающий слой 3Т3 сохраняет стволовой характер клеток.
Фиг.8 (увеличение 40х) показывает очевидное образование колоний на 3-й и 7-й день мезенхимными клетками из пуповины (UCMC), извлеченными согласно настоящему изобретению, культивируемыми на питающем слое 3T3. Питающий слой 3T3 обычно подавляет рост дифференцированных мезенхимных клеток как фибробластов кожи человека. Это вновь указывает на отличие в поведении этих мезенхимных клеток в сравнении с их более дифференцированными двойниками.
Фиг. с 9-1 по 9-27 показывают результаты анализа вестерн-блотированием, с помощью которого сравнивали экспрессию нескольких маркеров эмбрионных стволовых клеток в UCEC и UCMC, извлеченных согласно настоящему изобретению, с экспрессией этих маркеров в фибробластах кожи человека (NF), мезенхимных клетках костного мозга (BMSC) и полученных из жировой ткани мезенхимных клетках (ADMC). Фиг.9 показывает также высокую степень секреции активина А и фоллистатина, детектируемой методом ELISA в надосадочных жидкостях из культур мезенхимных и эпителиальных стволовых клеток пуповины в сравнении со стволовыми клетками из костного мозга и жировой ткани, фибробластами кожи человека и кератиноцитов эпидермиса.
Фиг.10 показывает результаты непрямого иммунофлуоресцентного анализа маркеров эпителиальных клеток, экспрессированных в эпителиальных стволовых клетках пуповины, таких как цитокератины (CK) в целом, CK17, CK6, CK10, CK19, CK18, CK16, CK15 (фиг.10-1); компоненты гемидесмосом - интегрин-α-6, интегрин-β-4, компоненты десмосом (фиг.10-2); компоненты базальной мембраны - ламинин 1, ламинин 5, коллаген IV, коллаген VII (фиг.10-3) и другие важные компоненты внеклеточного матрикса, такие как интегрин-бета и фибронектин (фиг.10-4).
Фиг.11 показывает результаты планшетного анализа цитокинов и факторов роста, секретируемых мезенхимными стволовыми клетками пуповины (UCMC), в сравнении с мезенхимными стволовыми клетками костного мозга человека.
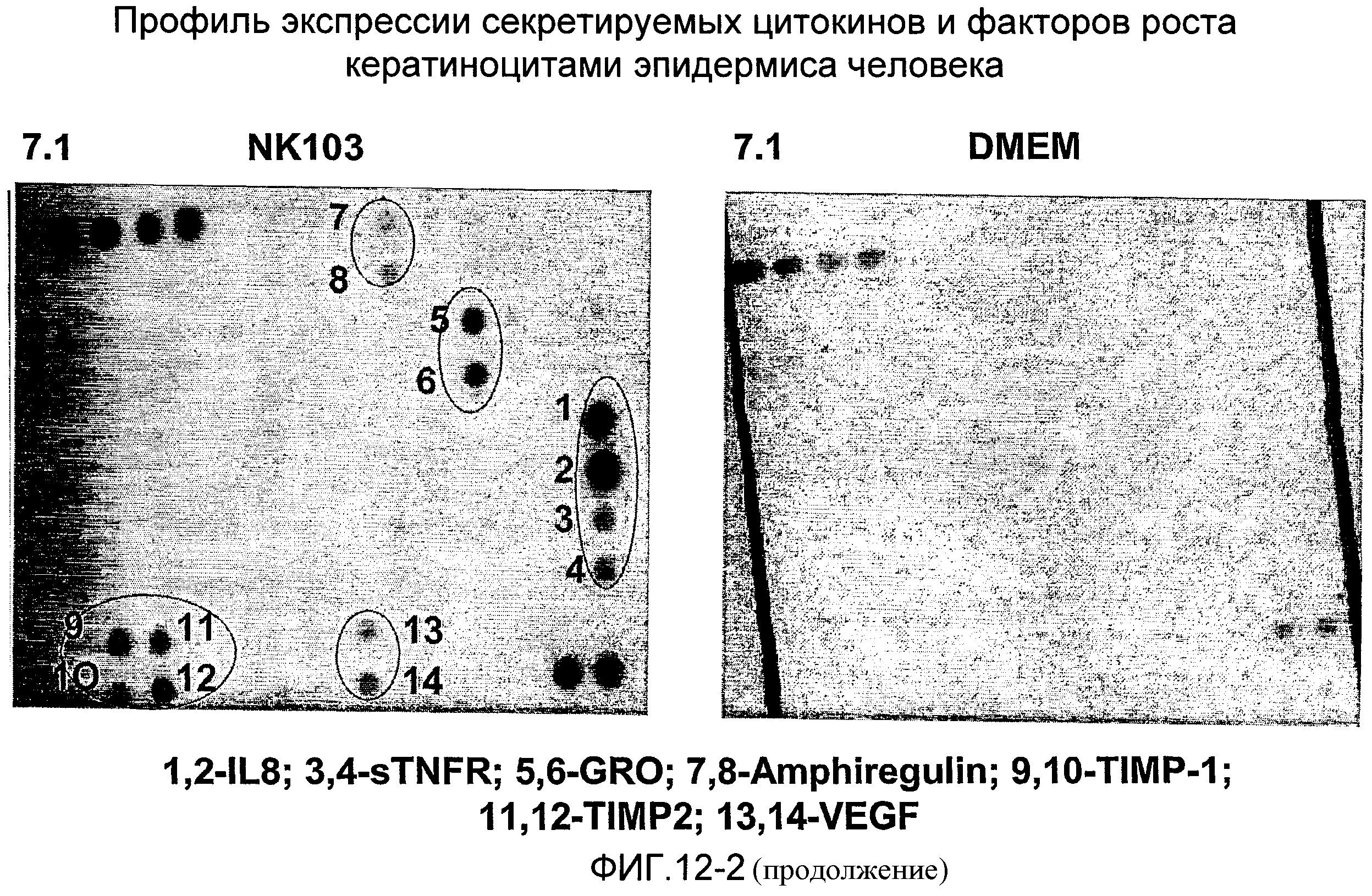
Фиг.12 показывает результаты планшетного анализа цитокинов и факторов роста, секретируемых эпителиальными стволовыми клетками пуповины (UCEC), в сравнении с кератиноцитами эпидермиса человека.
Фиг.13 показывает клетки UCMC, культивированные в среде DMEM, дополненной 10% сывороткой плода коровы (FCS) (фиг.13-1), в среде без сыворотки РТТ-1 (фиг.13-2), в среде без сыворотки РТТ-2 (фиг.13-3, фиг.13-4) и в среде без сыворотки РТТ-3 (фиг.13-5). На фиг.13 показан также рост в среде без сыворотки РТТ-3 полученных из жировой ткани клеток стромы (фиг.13-6) и полученных из костного мозга клеток стромы (фиг.13-7).
Фиг.14 показывает общую экспрессию генов в эпителиальных и мезенхимных стволовых клетках из пуповины, анализированную с помощью микрочипов ДНК. UCEC экспрессировали всего 28055 генов, a UCMC экспрессировали всего 34407. Имеются 27308 перекрывающихся генов, экспрессируемых в обоих типах клеток. Из экспрессируеых генов 747 были уникальными для UCEC и 7099 были уникальными для UCMC. На этом чертеже представлены отобранные гены, представляющие интерес. Оба типа стволовых клеток экспрессировали 140 генов, относящихся к эмбрионным стволовым клеткам и развитию эмбриона.
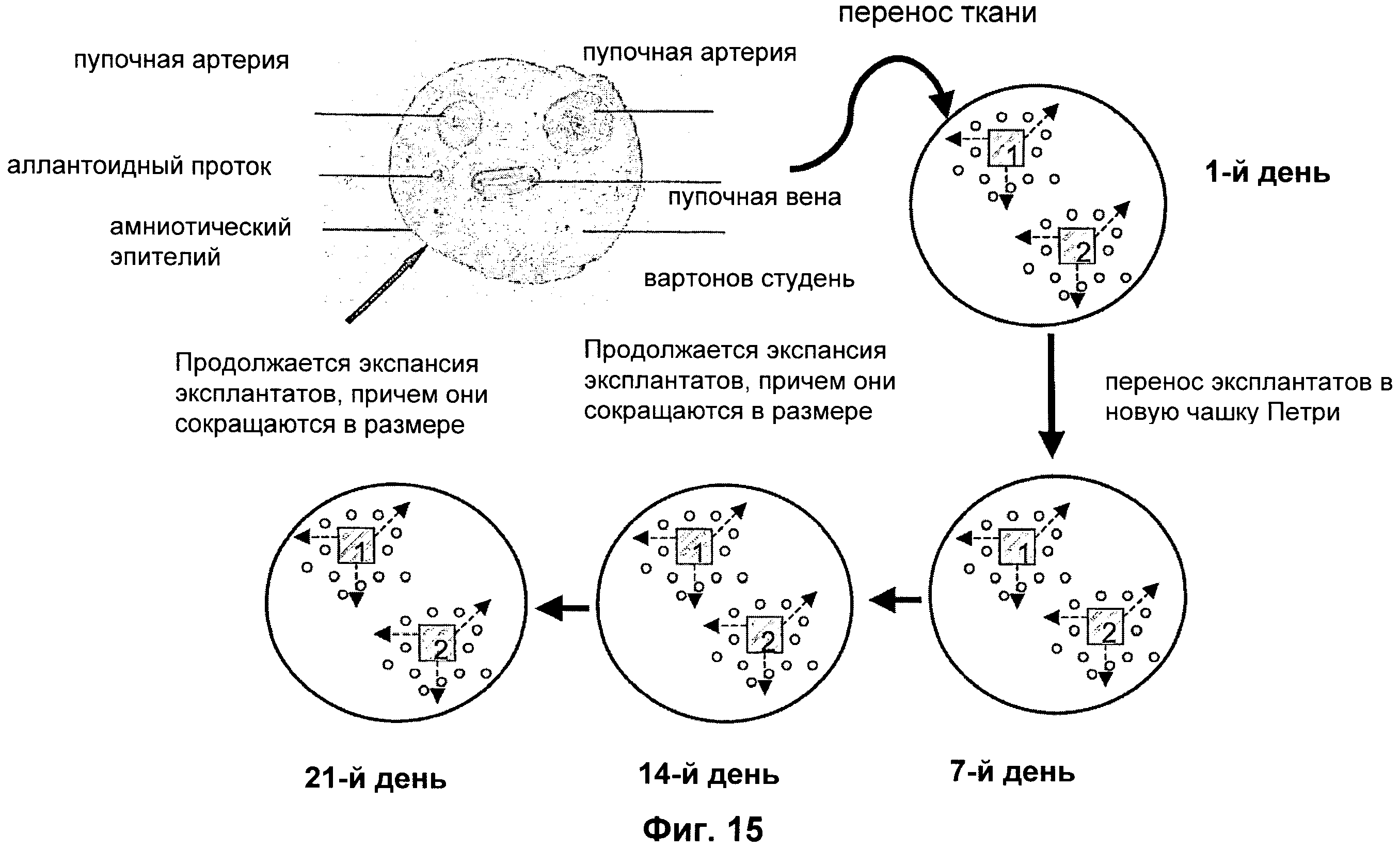
Фиг.15 дает схематическую иллюстрацию экспансии эпителиальных и мезенхимных стволовых клеток пуповины с использование повторных эксплантатов выстилающих пупочный канатик мембранных тканей.
Фиг.16 показывает поперечное сечение пуповины, демонстрирующее аминиотическую выстилающую пуповину мембрану (LM), содержащийся в ней вартонов студень (WJ), а также поддерживаемые внутри студня две пупочные артерии (UA) и одну пупочную вену (UV).
Подробное описание изобретения
Настоящее изобретение основано на неожиданном установлении того факта, что амниотическая мембрана пуповины представляет собой источник, из которого могут быть с успехом извлечены и подвергнуты проращиванию в условиях in vitro стволовые/прародительские клетки, такие как мезенхимные и эпителиальные стволовые/прародительские клетки. Даже более неожиданным является обнаружение того, что эти клетки проявляют характеристики, подобные характеристикам эмбрионных стволовых клеток. Амниотическая мембрана (которую называют также амниотической выстилающей мембраной), то есть лежащая глубоко внутри тонкая мембранная оболочка, заключающая в себе плаценту и развивающийся зародыш млекопитающих, была ранее использована как природный субстрат для реконструкции глазной поверхности и как биологический субстрат для проращивания стволовых клеток эпителия конечностей (см., например, Anderson D.F. и др. // Br. J. Ophthalmol. 2001. Т.85. С.567-575; Gruterich М. и др. // Surv. Ophthalmol. 2003. Т.48. С.631-646). Однако до сих пор не было описано никаких способов извлечения из амниотической мембраны стволовых/прародительских клеток (по крайней мере для людей) и не было описано использование амниотической мембраны, покрывающей пупочный канатик, в качестве источника стволовых клеток.
Изобретение предусматривает способ извлечения из амниотической мембраны пуповины стволовых/прародительских клеток, причем этот способ включает:
(а) отделение амниотической мембраны от других компонентов пуповины in vitro;
(б) культивирование полученной в этапе (а) ткани амниотической мембраны в условиях, обеспечивающих клеточную пролиферацию; и
(в) извлечение стволовых/прародительских клеток.
Термин «стволовая/прародительская клетка», как он использован здесь, относится к любой клетке, происходящей из пуповины и способной неограниченно самообновляться и дифференцироваться во многие типы клеток и тканей, таких как клетки эндотелия, клетки эпителия, фибробласты, миоциты или нейроны. Кроме того, клетки могут происходить из любого вида млекопитающих, такого как мышь, крыса, морская свинка, кролик, коза, собака, кошка, овца, обезьяна или человек, причем в одном из вариантов осуществления изобретения предпочтительными являются клетки человеческого происхождения.
Термин «свойства, подобные свойствам эмбрионных стволовых клеток», как он использован здесь, относится к способности клеток, происходящих из пуповины, почти как или в точности как эмбрионные стволовые клетки, спонтанно дифференцироваться во все типы тканей, что означает, что они являются плюрипотентными стволовыми клетками.
Термин «амниотическая мембрана», как он использован здесь, относится к лежащей глубоко внутри тонкой мембранной оболочке, заключающей в себе развивающийся зародыш млекопитающих. В течение беременности плод окружен и амортизирован жидкостью, которая называется амниотической жидкостью. Эта жидкость вместе с плодом и плацентой заключена в оболочку, называемую амниотической мембраной, которая покрывает также пуповину. Амниотическая жидкость важна по нескольким причинам. Она амортизирует и защищает плод, позволяя плоду свободно двигаться. Амниотическая жидкость дает также возможность пуповине плавать, защищая ее от передавливания, что отсекало бы снабжение плода кислородом и питательными веществами из крови, циркулирующей по кровеносным сосудам плаценты. Амниотическая оболочка содержит амниотическую жидкость, которая поддерживает гомеостаз окружения, защищая среду плода от внешнего мира. Кроме того, этот барьер защищает плод от организмов (таких как бактерии или вирусы), которые могут проникать во влагалище и быть причиной заражения.
Питательные среды и реагенты для культур ткани хорошо известны в данной области (см., например, Pollard J.W. and Walker J.M. // Basic Cell Culture Protocols, Second Edition. Humana Press, Totowa, NJ, 1997; Freshney R.I. // Culture of Animal Cells, Fourth Edition. Wiley-Liss, Hoboken, NJ, 2000). Примеры подходящих сред для инкубации/транспортировки образцов ткани пуповины включают (но не ограничиваются ими) модифицированную по Дульбекко среду Игла (Dulbecco's Modified Eagle Medium, DMEM), среду RPMI, сбаласированный солевой раствор Хэнкса (Hanks' Balanced Salt Solution, HBSS), забуференный фосфатом солевой физиологический раствор (phosphate buffered saline, PBS) и среду L-15, причем последняя среда предпочтительна в некоторых вариантах осуществления изобретения. Примеры подходящих сред для культивирования стволовых/прародительских клеток по изобретению включают (но не ограничиваются ими) среду DMEM, среды DMEM-F12 и RPMI, среду EpiLIfe и среду 171, причем последняя среда предпочтительна в некоторых вариантах осуществления изобретения. Среда может быть дополнена фетальной телячьей сывороткой (fetal calf serum, FCS) или фетальной бычьей сывороткой (fetal bovine serum, FBS), а также антибиотиками, факторами роста, аминокислотами, ингибиторами или подобными им веществами, которые полностью известны опытными специалистам в данной области.
В одном из вариантов осуществления изобретение предусматривает способ, дополнительно включающий:
(а") отделение этих стволовых/прародительских клеток от ткани амниотической мембраны перед культивированием с помощью техники ферментативного расщепления и/или прямого культивирования ткани эксплантата.
Термин «техника ферментативного расщепления», как он использован здесь, означает, что для освобождения клеток от основной массы ткани (здесь это амниотическая мембрана пуповины) добавляют ферменты. После этого собирают отделенные клетки. Термин «техника прямого культивирования ткани эксплантата», как он использован здесь, означает, что ткань вначале помещают в питательную среду без ферментов. Затем при щадящих условиях клетки сами по себе отделяются от основной массы ткани и их собирают для коллекции.
Методы отделения клеток конкретной ткани или конкретного органа путем обработки ферментами или прямого культивирования ткани эксплантата хорошо известны в данной области (см., например, Pollard J.VV. and Walker J.M. // Basic Cell Culture Protocols, Second Edition. Humana Press, Totowa, NJ, 1997; Freshney R.I. // Culture of Animal Cells, Fourth Edition. Wiley-Liss, Hoboken, NJ, 2000). Для осуществления способов согласно настоящему изобретению можно использовать любую катализируемую ферментом диссоциацию ткани. В предпочтительных вариантах осуществления изобретения для этой цели применяют коллагеназу. Фермент может быть использован либо в виде неочищенного препарата, либо в очищенной форме. Он может быть очищен из любого прокариотного или эукариотного организма (причем наиболее предпочтительным источником является Clostridium histolyticum) или получен рекомбинантным путем с помощью генной технологии. Можно применить любой тип коллагеназы, то есть тип 1, тип 2, тип 3, тип 4 или их любую комбинацию. В некоторых вариантах осуществления изобретения предпочтительно использование 1-го типа коллагеназы.
В одном из вариантов осуществления изобретение предусматривает способ извлечения стволовых/прародительских клеток, имеющих свойства, подобные свойствам эмбрионных стволовых клеток. Эти клетки могут в конце концов дифференцироваться по морфологии в эпителиальные или мезенхимные клетки (но без ограничения ими).
Соответственно, в другом варианте осуществления изобретение предусматривает способ извлечения эпителиальных и/или мезенхимных стволовых/прародительских клеток, причем в соответствии с раскрытым выше эти клетки могут иметь свойства, подобные свойствам эмбрионных стволовых клеток.
Эпителиальные стволовые/прародительские клетки включают любые клетки, обнаруживающие характерную для эпителиальных клеток морфологию (то есть многоугольную форму), которые могут дифференцироваться в любой тип эпителиальных клеток, такой как (но не ограничиваясь ими) эпителиальные клетки кожи, фолликулярные клетки волос, эпителиальные клетки роговицы, эпителиальные клетки конъюнктивы, эпителиальные клетки сетчатки, эпителиальные клетки печени, эпителиальные клетки почки, эпителиальные клетки поджелудочной железы, эпителиальные клетки пищевода, эпителиальные клетки тонкого кишечника, эпителиальные клетки толстой кишки, эпителиальные клетки легких и дыхательных путей, эпителиальные клетки мочевого пузыря или эпителиальные клетки матки.
Мезенхимные стволовые/прародительские клетки включают любые клетки, обнаруживающие морфологию, подобную морфологии мезенхимных клеток (то есть веретенообразную форму), которые могут дифференцироваться в любой тип мезенхимных клеток, такой как (но не ограничиваясь ими) фибробласты кожи, хондроциты, остеобласты, теноциты, фибробласты связок, кардиомиоциты, клетки гладкой мускулатуры, клетки мускулатуры скелета, адипоциты, происходящие из эндокринных желез клетки и все разнообразие и все производные нейроректодермальных клеток.
В другом варианте осуществления изобретение предусматривает способ, дополнительно включающий:
(г) культивирование стволовых/прародительских клеток в условиях, дающих возможность клеткам претерпевать клональную экспансию.
Термин «клональная экспансия» («clonal expansion») (иногда это называют также «митотическая клональная экспансия», «mitotic clonal expansion») относится к процессу, который происходит на ранней стадии программы дифференцировки клетки, путем которого стволовые/прародительские клетки направляются в сторону конкретного пути дифференцировки и затем претерпевают окончательную дифференцировку. В данной области хорошо известно, что для различных типов клеток условия, необходимые для индукции клональной экспансии прародительских клеток, могут существенно различаться. Без связи с конкретным методом индукция клональной экспансии обычно достигается путем культивирования стволовых/прародительских клеток в среде, которая была оптимизирована для пролиферации клеток. Такие среды предоставляются многими поставщиками. Неограничивающими примерами таких сред являются KGM®-Keratinocyte Medium (Cambrex), MEGM-Mammary Epithelial Cell Medium (Cambrex), EpiLife medium (Cascade Biologies) или Medium 171 (Cascade Biologies). В качестве альтернативы культуральная среда может быть дополнена реагентами, индуцирующими пролиферацию клеток, такими как факторы роста. Такие реагенты могут быть смешаны в одном растворе, таком как Human Keratinocyte Growth Supplement Kit (Cascade Biologies) (один из примеров) или могут быть внесены по отдельности. Такие реагенты включают (но не ограничиваются ими) факторы роста (такие как, например, фактор роста эпидермиса, инсулино-подобный фактор роста 1, происходящий из тромбоцитов фактор роста ВВ, трансформирующий фактор роста β1, инсулин), гормоны (такие, как экстракт бычьего гипофиза), гидрокортизон, трансферрин и подобные им в любой подходящей комбинации для индуцирования клональной экспансии данного типа клеток. Термин «клональная экспансия» включает также культивирование клеток in vivo, например, путем инъекции клеток млекопитающим, таким как люди, мыши, крысы, обезьяны, человекообразные обезьяны (чтобы ограничиться немногими примерами).
Еще в одном примере осуществления изобретение предусматривает способ, дополнительно включающий:
(д) культивирование стволовых/прародительских клеток в условиях, обеспечивающих дифференцировку указанных клеток в эпителиальные клетки и/или мезенхимные клетки; и
(е) извлечение дифференцированных клеток.
Еще в одном примере осуществления изобретение предусматривает способ, дополнительно содержащий:
(ж) консервацию извлеченных стволовых/прародительских клеток для дальнейшего использования.
Способы и протоколы для консервации и хранения эукариотических клеток и конкретно клеток млекопитающих хорошо известны в данной области (см., например, Pollard J.W. and Walker J.M. // Basic Cell Culture Protocols, Second Edition. Humana Press, Totowa, NJ, 1997; Freshney, R.I. // Culture of Animal Cells, Fourth Edition. Wiley-Liss, Hoboken, NJ, 2000). В связи с настоящим изобретением может быть использован любой метод, сохраняющий биологическую активность извлеченных эпителиальных или мезенхимных стволовых/прародительских клеток. В одном из предпочтительных вариантов осуществления изобретения стволовые/прародительские клетки поддерживают и хранят с использованием криоконсервации.
В соответствии с этим изобретение направлено также на прародительские/стволовые клетки, полученные из амниотической мембраны пуповины с помощью указанных выше способов. Кроме того, изобретение направлено также на клеточный банк, включающий или состоящий из одной или более прародительских/стволовых клеток, извлеченных так, как здесь описано. Это клеточный банк прародительских/стволовых клеток может быть аутологичным по отношению к индивидууму или сборным и может быть впоследствии применен путем дальнейшей дифференцировки, например, для регенеративной медицины, заживления и восстановления тканей.
В соответствии с указанным выше изобретение направлено также на фармацевтическую композицию, включающую стволовую/прародительскую клетку, извлеченную из амниотической мембраны пуповины указанным выше заявленным способом. Фармацевтическая композиция может быть любого вида и обычно включает стволовые/прародительские клетки или клеточный экстракт из них вместе с подходящим терапевтически приемлемым носителем/наполнителем. В некоторых вариантах осуществления изобретения фармацевтическая композиция приспособлена для системного или местного применения.
Приспособленная для местного применения фармацевтическая композиция может быть в жидкой или вязкой форме. Примеры этого включают мазь, крем, лосьон и подобные им. Примерами фармацевтических композиций, пригодных для системного применения, являются жидкие композиции, где стволовые/прародительские клетки или клеточный экстракт растворены, например, в буфере, приемлемом для инъекции или инфузии.
Соответственно, изобретение относится также к способу лечения индивидуума, имеющего расстройство. Это способ включает введение индивидууму эффективного количества либо стволовых/прародительских клеток, извлеченных так, как здесь объяснено, или полученного из таких клеток клеточного экстракта.
В принципе, любое (болезненное) состояние, подходящее для лечения стволовыми/прародительскими клетками, можно лечить клетками или клеточным экстрактом в соответствии с настоящим изобретением. В некоторых вариантах осуществления изобретения расстройство выбрано из группы, состоящей из опухолевого заболевания, ускоренного старения кожи, кожных расстройств, тканевых расстройств, эндокринных недостаточностей внутренних органов и нервных расстройств.
Подлежащее лечению тканевое расстройство может быть врожденным или приобретенным тканевым расстройством. Примеры эндокринной недостаточности внутренних органов, которую можно лечить клетками в соответствии с настоящим изобретением, включают (но не ограничиваются ими) связанный с инсулиновой недостаточностью сахарный диабет, дефицит тестостерона, анемию, гипогликемию, гипергликемию, панкреатическую недостаточность, недостаточность надпочечников и недостаточность щитовидной железы.
Примеры нервных расстройств, которые можно лечить, включают (но не ограничиваются ими) болезнь Альцгеймера, болезнь Паркинсона, болезнь Крейцфельда-Якоба, болезнь Лю Герига, болезнь Хантингтона и опухолевые болезненные неврологические состояния.
Примером заболевания кожи является рана или повреждение части кожи, например солнечный ожог кожи. Старение кожи здесь также рассматривается как заболевание кожи. Поэтому для заживления обожженной солнцем кожи может быть использовано местное или похожее введение стволовых/прародительских клеток согласно настоящему изобретению или клеточных экстрактов из них, например в качестве составной части лосьонов или кремов или любого другого подходящего носителя, и в добавление они могут также замедлить процесс старения кожи (противостарческая активность) путем пополнения и, следовательно, обогащения кожи недостающими факторами роста и родственными пептидными элементами, без которых старение кожи ускорилось бы. Стволовые/прародительские клетки могут также мигрировать к поврежденным участкам тела, таким как поверхность раны, чтобы сформировать требуемые клеточные элементы, необходимые для локальных процессов восстановления (см. // J. of Immunol. 2001. Т.166. С.7556-7562; или // Internat. I J. of Biochem. and Cell Biol. 2004. Т. 36. С.598-606).
Опухолевым заболеванием может быть рак, в особенности потому, что в недавних исследованиях было продемонстрировано, что стволовые клетки могут селективно нацеливаться на ткань новообразованной опухоли (// J. of the National Cancer Institute. 2004. Т. 96, №21. С.1593-1603), что обеспечивает направленную доставку к опухолевым очагам противоопухолевых средств, таких как интерферон. Раком может быть любой вид рака, в том числе те виды рака, при которых образуются твердые (солидные) опухоли, в диапазоне от рака кожи до рака внутренних органов. Примеры подлежащих лечению типов рака включают карциному сквамозных клеток, карциному протоков и долек молочной железы, карциному клеток печени, карциному носоглотки, рак легких, рак костей, рак поджелудочной железы, рак кожи, рак головы или шеи, кожную или внутриглазную злокачественную меланому, рак матки, рак яичников, рак прямой кишки, рак анальной области, рак желудка, рак толстой кишки, рак груди, рак яичек, карциному фаллопиевых труб, карциному эндометрия, карциному шейки матки, карциному влагалища, карциному вульвы, болезнь Ходжкина, лимфому не Ходжкина, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак околощитовидной железы, рак надпочечника, саркому мягких тканей, рак мочеиспускательного канала, рак пениса, рак простаты, хронические или острые лейкозы, детские твердые опухоли, лимфоцитарную лимфому, рак мочевого пузыря, рак почки или мочеточника, карциному клеток почки, карциному почечных лоханок, новообразования центральной нервной системы (ЦНС), первичную лимфому ЦНС, опухолевый ангиогенез, опухоли позвоночного столба, глиому ствола головного мозга, аденому гипофиза, саркому Капоши, эпидермоидный рак или любую комбинацию таких видов рака, в том числе их диссеминированные (метастазирующие) формы. В случае лечения опухолевого заболевания раскрытые здесь стволовые клетки из амниона пуповины и/или их клеточные экстракты могут быть введены системно или как прямое лечебное средство и/или как носитель. В последнем случае противоопухолевой терапии клетки включают противоопухолевое средство.
В другом фармацевтическом применении стволовые/прародительские клетки могут быть использованы для генной терапии. Для этой цели клетки могут быть трансформированы нуклеиновой кислотой, кодирующей белок, который должен продуцироваться в клетках. Нуклеиновую кислоту можно вводить в клетки согласно настоящему изобретению с помощью любого из различных методов, хорошо известных опытным специалистам. Например, это может быть сделано с применением вирусного вектора и/или содержащей липид трансфекционной композиции, такой как IBAfect (IBA GmbH, Gottingen, Germany), Fugene (Roche), GenePorter (Gene Therapy Systems), Lipofectamine (Invitrogen), Superfect (Qiagen), Metafecten (Biontex), или таких, какие описаны в публикации WO 01/015755). В связанном примере осуществления изобретения клетки согласно настоящему изобретению, после того, как они трансформированы нуклеиновой кислотой, кодирующей выбранный полипептид, могут быть использованы для рекомбинантной продукции этого полипептида.
Как указано выше, экстракты стволовых клеток богаты различными факторами роста и пептидами, имеющими отношение к нормальной клеточной физиологии. Таких факторов роста и/или пептидов может быть недостаточно в экспонированных частях тела, таких как кожа, которая является поверхностным слоем у всех человеческих существ, защищающим тело от внешних элементов для поддержания внутреннего гомеостаза. Поэтому в дальнейшем примере осуществления изобретения стволовые/прародительские клетки согласно настоящему изобретению или клеточные экстракты из них пригодны для воздействия на внутренний гомеостаз и/или поддержание внутреннего гомеостаза.
В дальнейшем варианте осуществления изобретения и в согласии с приведенным выше описанием стволовые/прародительские клетки согласно настоящему изобретению могут быть использованы для продукции любой биологической молекулы. Биологической молекулой может быть, например, любая молекула, которая в естественных условиях продуцируется клетками, или молекула, кодирующая которую нуклеиновая кислота была введена в клетки с помощью технологии рекомбинантной ДНК. Примеры молекул, которые могут продуцироваться клетками согласно настоящему изобретению, включают (но не ограничиваются ими) белок, такой как цитокин, фактор роста, такой как инсулино-подобный фактор роста (insulin-like growth factor, IGF), фактор роста эпидермиса (epidermal growth factor, EGF), трансформирующий фактор роста β (transforming growth factor β, TGF-β), активин А, морфогенетический белок костей (bone morphogenetic protein, BMP), полученный из тромбоцитов фактор роста (PDGF) или гормон (такой как инсулин или эритропоэтин), или транспортный белок (такой как трансферрин), пептид, такой как фактор роста или гормон (например, лютеинизирующий гормон (LSH), фолликулостимулирующий гормон (FSH)), низкомолекулярное органическое соединение, такое как стероидный гормон, олиго- или полисахарид, например гепарин или гепаран-сульфат (в отношении его см., например, WO 96/23003 или WO 96/02259), протеогликан гликопротеин, такой как коллаген или ламинин, или липид, чтобы ограничиться небольшим перечнем.
В дальнейшем аспекте и в согласии с последними подходами (см., например, Amit М. и др. Human feeder layers for human embryonic stem cells // Biol. Reprod. 2003. Т.68. С.2150-2156), описанные здесь стволовые/прародительские клетки могут быть применены в качестве питающего слоя для культивирования других эмбрионных стволовых клеток, в частности человеческих стволовых клеток. В одном из этих вариантов осуществления клетки согласно настоящему изобретению - это предпочтительно клетки человеческого происхождения, поскольку использование человеческих клеток в качестве питающих слоев сводит к минимуму риск загрязнения культуры клеток компонентами животного происхождения, такими как патогены или иммуногены животных. В этом отношении необходимо отметить, что клетки согласно настоящему изобретению можно культивировать в условиях отсутствия сыворотки. Этому соответствует применение клеток в качестве питающего слоя и культивация культур клеток в бессывороточной среде, такой как описанная, например, далее здесь или в работе Draper и др. (Culture and characterization of human embryonic stem cell lines // Stem Cells Dev. 2004. Т.13. С.325-336), или в международной публикации WO 98/30679.
В этой связи отмечается, что в трансплантационной хирургии и терапии на основе клеток решающим является большое количество прошедших малое число пассажей клеток с минимальным содержанием стареющих клеток (то есть с высокой долей клеток высокого качества), и эти клетки должны быть получены в течение как можно более короткого времени в ходе экспансии клеток. Например, мезенхимные стволовые клетки из костного мозга и крови пуповины имеются в небольшом количестве и поэтому требуют проращивания во многих пассажах в течение длительного периода времени, чтобы получить достаточное количество клеток для пересадки. Однако клетки, прошедшие большое число пассажей, имеют тенденцию к ухудшению качества, что может приводить к старению клеток или их трансформации в раковые клетки. Здесь было установлено, что большие количества клеток согласно настоящему изобретению можно получить малым числом пассажей, используя технику повторной эксплантации. Таким образом, настоящее изобретение относится также к способу культивирования стволовых/прародительских клеток согласно настоящему изобретению, причем этот способ включает:
получение эксплантата ткани из амниотической мембраны пуповины;
культивирование эксплантата ткани в подходящей культивационной среде и при подходящих условиях культивирования в течение подходящего периода времени.
По усмотрению, может быть проведено выдерживание эксплантата ткани в свежей культивационной среде и продолжение культивирования при подходящих условиях в течение подходящего периода времени (см. фиг.15).
Культивацию можно проводить в течение такого числа циклов (пассажей), как необходимо, и остановить по достижении необходимого количества клеток. Выдерживание эксплантата ткани в свежей культивационной среде может быть осуществлено путем удаления среды для культивации клеток из флакона, используемого для роста клеток, и добавления в этот флакон свежей среды. Вместо замены среды в используемом флаконе, выдерживание в свежей культивационной среде можно обеспечить переносом эксплантата ткани в новый флакон, заполненный культивационной средой. Эксплантат ткани, используемый для культивации/проращивания клеток, может быть получен любым пригодным методом, например техникой «прямого культивирования ткани эксплантата», как объяснено выше (в которой ткань вначале помещают в среду без ферментов, затем в щадящих условиях клетки сами по себе отделяются от основной массы ткани, и после этого клетки собирают для коллекции).
Культивирование эксплантатов ткани можно осуществить во многих средах, пригодных для культивирования клеток млекопитающих. Примеры включают обычные и имеющиеся в продаже среды, перечисленные выше в связи с культивированием или клональной экспансией клеток согласно настоящему изобретению, такие как (но без ограничения ими) KGM®-Keratinocyte Medium (Cambrex), MEGM-Mammary Epithelial Cell Medium (Cambrex), EpiLife medium (Cascade Biologies), Medium 171 (Cascade Biologies), DMEM, DMEM-F12 или среда RPMI. Культивирование обычно проводят в условиях (температура, атмосфера), которые обычно используют для культивирования клеток тех видов, из которых они получены (например, при 37°С в воздушной атмосфере с 5% CO2). В одном из примеров осуществления изобретения культивирование осуществляют в бессывороточной среде, в частности в среде без сыворотки крупного рогатого скота). Культивирование (в одном пассаже) проводят в течение любого необходимого времени, которое требуется клеткам для роста, как правило (но никоим образом не ограничиваясь этими сроками), в течение приблизительно от одного до нескольких дней (например, до приблизительно 7 или приблизительно 8 дней).
Иллюстративно описанные здесь изобретения могут быть подходящим образом осуществлены в отсутствие каких-либо элемента или элементов, ограничения или ограничений, не раскрытых здесь конкретно. Так, например, термины «включающий», «заключающий», «содержащий» и т.д. могут трактоваться широко и без ограничений. Кроме того, примененные здесь термины и выражения были использованы как термины описания, но не ограничения, и нет намерения использовать эти термины и выражения для исключения каких-либо эквивалентов показанных и описанных здесь признаков или их частей, и ясно, что в пределах заявляемого изобретения возможны различные модификации. Таким образом, следует понимать, что, хотя настоящее изобретение конкретно раскрывается в предпочтительных примерах осуществления с необязательными признаками, специалисты могут предложить модификации и вариации изобретения, и такие модификации и вариации рассматриваются как находящиеся в пределах объема настоящего изобретения.
Изобретение было описано здесь широко и обобщенно. Каждый из более узких разделов и более узких группировок, попадающих в обобщенное описание, также являются частью изобретения. Это включает обобщенное описание изобретения, с условием или негативным ограничением, удаляющим из общего любое частное, независимо от того, изложен ли здесь или нет конкретно исключаемый материал.
Другие примеры осуществления изобретения охватываются следующей далее формулой изобретения и неограничивающими примерами. Кроме того, если свойства или аспекты изобретения описываются в терминах групп Маркуша, опытные специалисты в данной области различат, что изобретение таким образом описано и в терминах любого из индивидуальных представителей или любой подгруппы представителей группы Маркуша.
Примеры
Пример 1
Отбор ткани пуповины
Ткань пуповины собирали сразу после рождения ребенка. Образец ткани отмывали дочиста и немедленно помещали в стерильный стеклянный флакон объемом 500 мл, содержащий среду для транспортирования культуры (среда L-15, дополненная 50 ед./мл пенициллина, 50 мкг/мл стрептомицина, 250 мкг/мл фунгизона, 50 мкг/мл гентамицина; все реагенты поставлены фирмой Invitrogen) перед транспортировкой в лабораторию. В лаборатории проводили экстракцию стволовых клеток в ламинарном боксе в стерильных условиях. Образец вначале переносили на стерильный поднос из нержавеющей стали. Всю оставшуюся в сосудах пуповины кровь удаляли несколькими промывками шприцем, используя теплый забуференный фосфатом солевой физиологический раствор (phosphate-buffered saline, PBS), дополненный 50 ед./мл гепарина (фирмы Sigma). Для последних промывок использовали обычный PBS без гепарина. Затем образец ткани пуповины разрезали на кусочки длиной 2 см и переносили в чашки для культур клеток диаметром 10 см, где осуществляли дальнейшую промывку и дезинфекцию 70% этанолом, после чего проводили несколько промывок PBS, содержащим смесь антибиотиков (50 ед./мл пеницилллина, 50 мкг/мл стрептомицина, 250 мкг/мл фунгизона, 50 мкг/мл гентамицина; все реагенты поставлены фирмой Invitrogen) до тех пор, пока раствор не становился прозрачным.
Пример 2.
Отделение и культивирование клеток
Сначала для отделения амниотической мембраны пуповины от вартонова студня (то есть от матрикса пуповины) и других внутренних компонентов производили иссечение ткани пуповины. Затем выделенную амниотическую мембрану разрезали на небольшие кусочки (0,5 см × 0,5 см) для извлечения клеток. Проращивание эксплантата производили, помещая кусочки амниотической мембраны пуповины на чашки для культивирования клеток при различных условиях культивирования клеток для получения либо эпителиальных, либо мезенхимных стволовых клеток.
Для извлечения/культивирования мезенхимных клеток эксплантаты погружали в 5 мл среды DMEM (Invitrogen), дополненной 10% сывороткой плода коровы (Hyclone) (DMEM/10% FBS) и инкубировали в термостате для культивирования клеток с CO2 при 37°С. Среду сменяли каждые 2-3 дня. Мониторинг прорастания клеток производили с помощью световой микроскопии. Отросшие клетки собирали трипсинизацией (0,125% трипсин/0,05% EDTA) для дальнейшего проращивания и крио-консервации в DMEM/10% FBS.
Для извлечения/культивирования эпителиальных клеток пластиковые поверхности для культивирования клеток перед тем как поместить на их поверхность образцы ткани, покрывали смесями коллагена 1 и коллагена 4 (1:2). Образцы ткани погружали в 5 мл среды EpiLife или среды Medium 171 (обе фирмы Cascade Biologies). Среду сменяли каждые 2-3 дня. Мониторинг прорастания клеток производили с помощью световой микроскопии. Отросшие клетки собирали трипсинизацией (0,125% трипсин/0,05% EDTA) в среде EpiLife или Medium 171.
Для метода ферментативной экстракции клеток амниотическую мембрану пуповины разрезали на небольшие кусочки (0,5 см × 0,5 см) и расщепляли в растворе 0,1% (вес к объему) коллагеназы 1-го типа (Roche Diagnostics) при 37°С в течение 6 ч. Образцы перемешивали в гомогенизаторе в течение 2 мин каждые 15 мин. Клетки собирали центрифугированием при 4000 об/мин в течение 30 мин. Для извлечения либо эпителиальных, либо мезенхимных стволовых клеток были применены 2 различных подхода.
Для извлечения эпителиальных стволовых клеток осадки клеток ресуспендировали в среде EpiLife или Medium 171 (обе фирмы Cascade Biologies), дополненной 50 мкг/мл инсулино-подобного фактора роста 1 (IGF-1), 50 мкг/мл полученного из тромбоцитов фактора роста ВВ (PDGF-BB), 5 мкг/мл трансформирующего фактора роста β1 (TGF-β1 и 5 мкг/мл инсулина (все получены от фирмы R&D Systems), клетки подсчитывали и высевали на чашки для культур ткани диаметром 10 см, предварительно покрытые смесями коллагена 1 и коллагена 4 (1:2; Becton Dickinson), при плотности 1×106 клеток на чашку. Через 24 ч прикрепившиеся клетки промывали теплым PBS и заменяли среду средой с добавками - EpiLife или Medium 171. Среду сменяли каждые 2-3 дня. Мониторинг роста клеток и образования клонов при экспансии производили с помощью световой микроскопии. При степени слияния клеток около 70% производили субкультивирование клеток с помощью трипсинизации (0,125% трипсин/0,05% EDTA) для дальнейшего проращивания и крио-консервации.
Для извлечения мезенхимных стволовых клеток осадки клеток ресуспендировали в DMEM/10% FBS, подсчитывали и высевали на чашки для культур ткани диаметром 10 см при плотности 1×106 клеток на чашку. Среду сменяли каждые 2-3 дня. Мониторинг роста клеток и экспансии производили с помощью световой микроскопии. При степени слияния клеток около 90% клетки субкультивировали, как указано выше.
Для культивирования эпителиальных и мезенхимных стволовых клеток на питающем слое выстилающую пуповину мембрану расщепляли коллагеназой, клетки подсчитывали и высевали на чашки для культур ткани диаметром 10 см, покрытые убитыми облучением или обработанными митомицином С фиборобластами 3T3 (питающий слой) в среде Грина. Культуральную среду сменяли каждые 2-3 дня. Мониторинг образования колоний проводили с помощью световой микроскопии, колонии фотографировали.
Пример 3
Идентификация стволовых/прародительских клеток
Эпителиальные клетки: На фиг.1 представлены изображения проросших эпителиальных клеток из амниотической мембраны пуповины, полученных способом с использованием эксплантата ткани (увеличение 40х). Фотографии сделаны на 2-й день (фиг.1А) и на 5-й день (фиг.1Б, В) культивирования ткани. Анализ морфологии клеток показывает подобные эпителиальным клетки многоугольной формы. Ферментативное расщепление сегментов пуповины дает похожие (фиг.2) эпителиальные клетки на 2-й день (фиг.А, В) и на 5-й день (фиг.Б, Г) (увеличение 40х). На фиг.7 показано образование колоний эпителиальных стволовых клеток из амниотической мембраны пуповины, культивированных на питающем слое по методу Грина (увеличение 40х). Между 3-м и 7-м днями наблюдается быстрая экспансия колонии клеток многоугольной формы, подобных эпителиальным.
Мезенхимные клетки: Прорастание мезенхимных клеток после эксплантации из амниотической мембраны пуповины наблюдалось уже через 48 ч после помещения в чашки для культивирования ткани при использовании в качестве культуральной среды DMEM, дополненной 10% сывороткой плода коровы (FCS) (фиг.3А, В) (увеличение 40х). Клетки характеризовались веретенообразной морфологией и мигрировали и прорастали in vitro легко и быстро, очень напоминая фибробласты (фиг.3Б, Г) (увеличение 40х). Подобные наблюдения были сделаны для группы клеток, извлеченных путем ферментативного расщепления коллагеназой (фиг.4). На фиг.4А показаны мезенхимные клетки, извлеченные из амниотической мембраны пуповины на 2-й день. Пролиферация клеток наблюдалась на 5-й день (фиг.4Б) (увеличение 40х). Фиг.6 и 8 показывают образование колоний мезенхимных стволовых клеток из амниотической мембраны пуповины, культивированных при условиях наличия непитающего слоя и питающего слоя в DMEM/10%FCS (увеличение 40х). Колонии подобных фибробластам клеток вытянутой формы быстро разрастались в период между 3-м и 7-м днями.
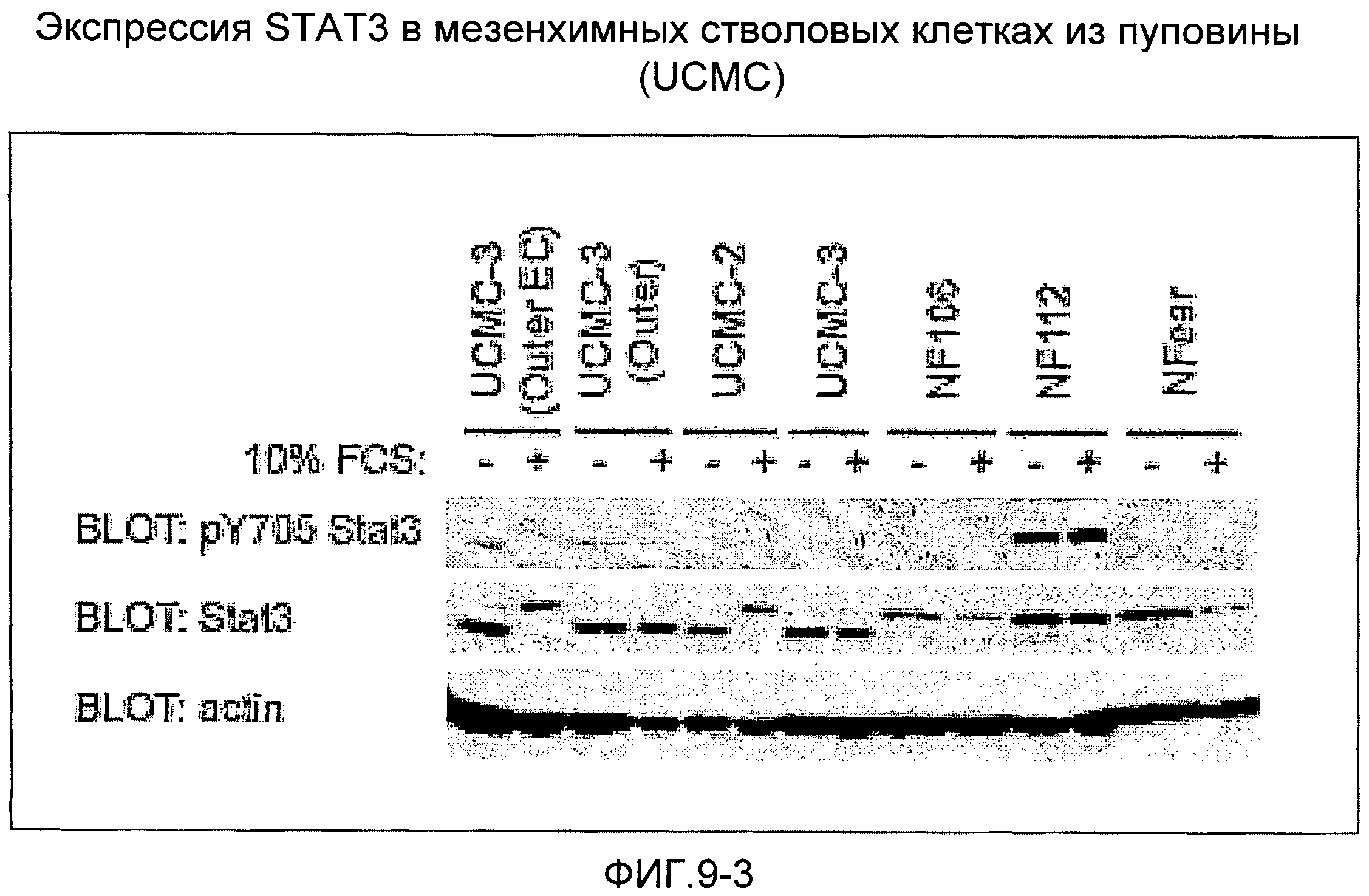
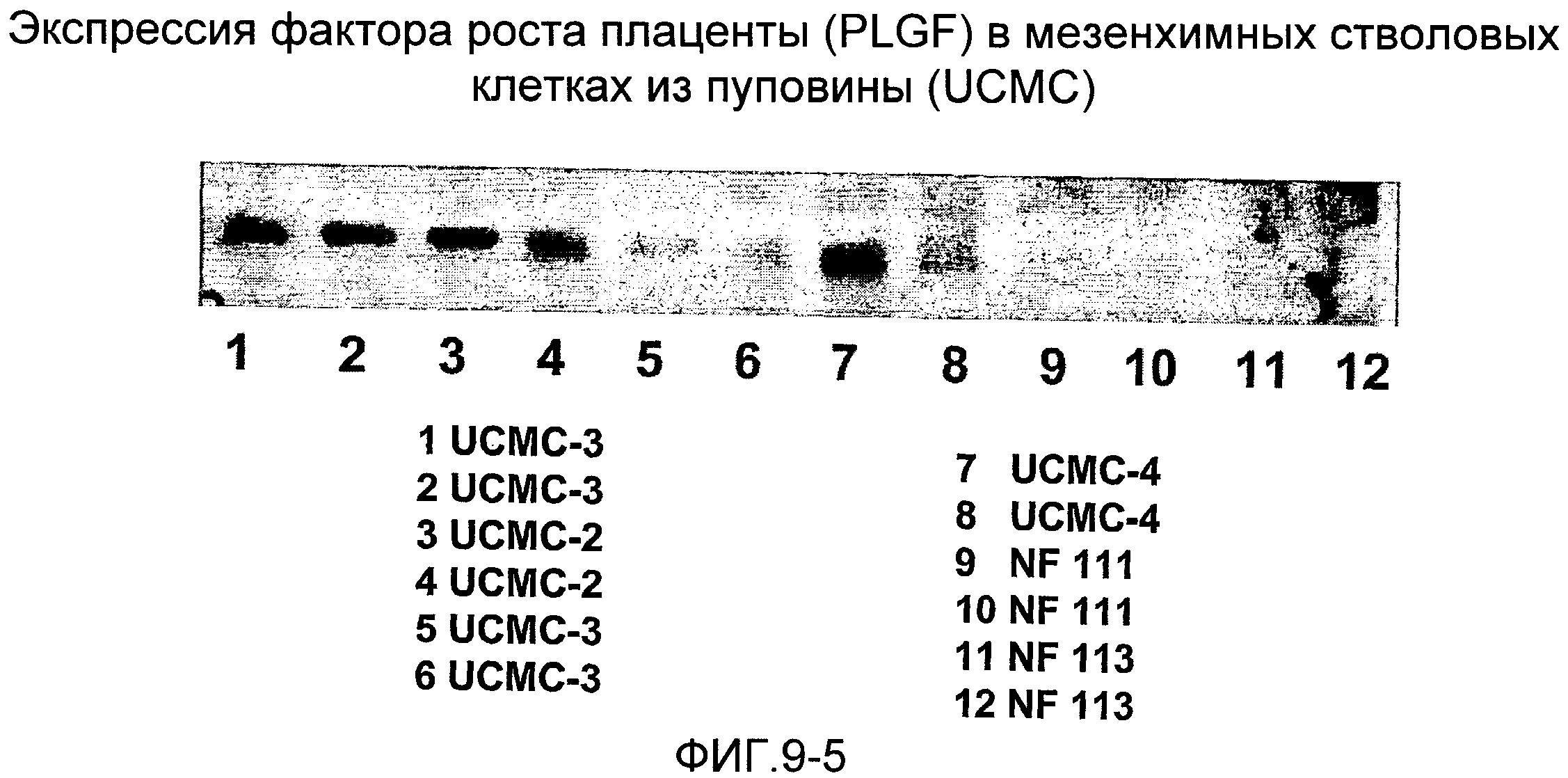
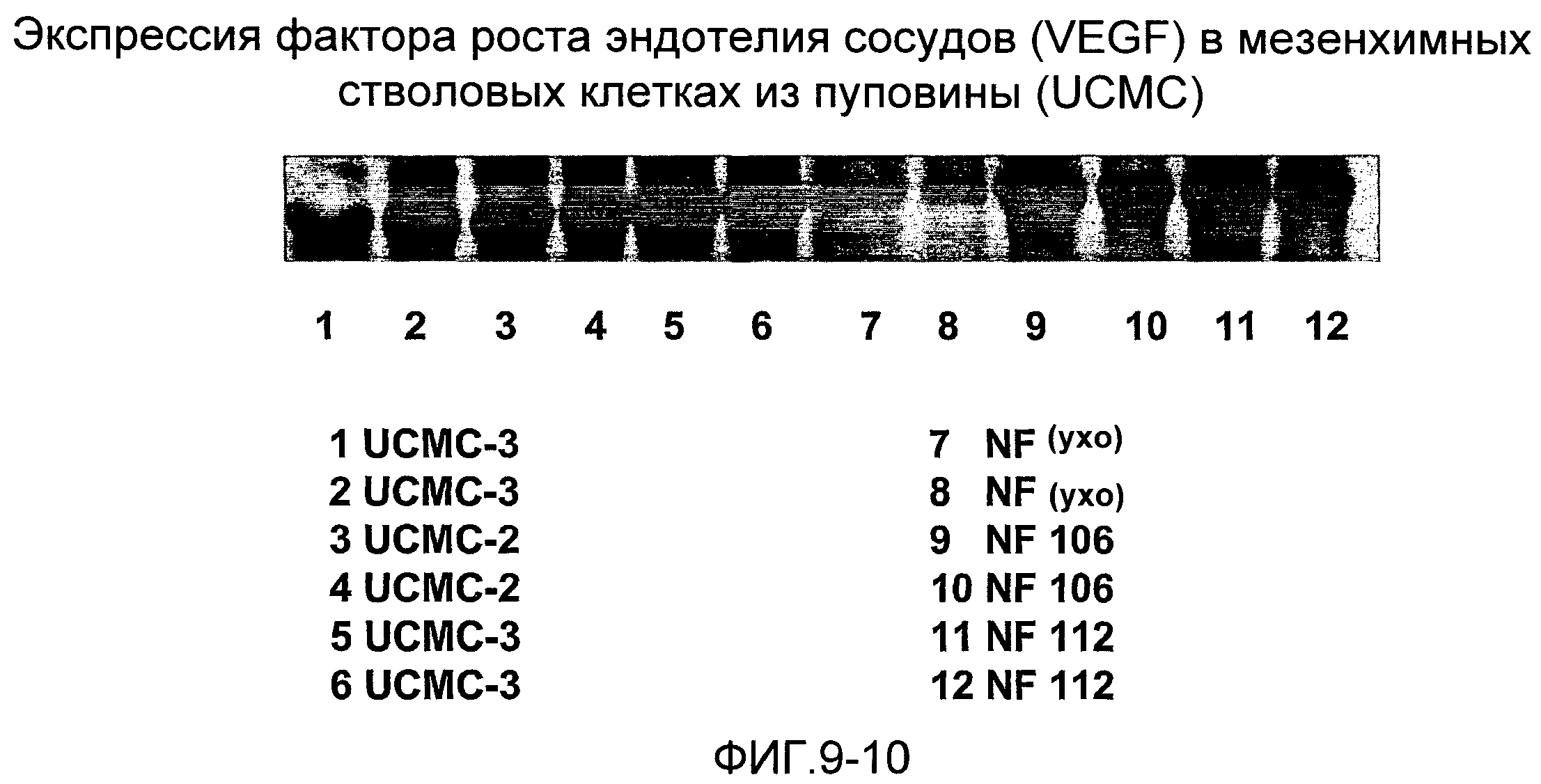
Анализ с помощью вестерн-блотирования (фиг.9) показывает, что мезенхимные стволовые клетки из амниотической мембраны пуповины (UCMC) и эпителиальные клетки из пуповины (UCEC), извлеченные согласно настоящему изобретению, экспрессируют ген POU5f1, который кодирует фактор транскрипции Octamer-4 (Oct-4) - специфический маркер эмбриональных стволовых клеток (см. Niwa H., Miyazaki J., Smith A.G. (2000). // Nat. Genet. 2000. Т.24. С.372-376), Таким образом, этот анализ указывает на подобные эмбрионным свойства этих стволовых клеток. Эти клетки обнаруживают также высокий уровень экспрессии других факторов роста, таких как фактор роста соединительной ткани (CTGF), фактор роста эндотелия сосудов (VEGF), плаценто-подобный фактор роста PLGF, STAT3, фактор стволовых клеток (SCF), происходящий из гепатомы фактор роста (HDGF), фактор роста 2 фибробластов (FGF-2), полученный из тромбоцитов фактор роста (PDGF), α-актин гладких мышц (α-SMA), фибронектин, декорин, синдекан-1,2,3,4. На фиг.9 показано сравнение экспрессии этих генов с экспрессией в фибробластах кожи человека, мезенхимных клетках костного мозга (BMSC) и мезенхимных клетках из жировой ткани (ADMC). Фиг.9 показывает также детектируемую методом ELISA в надосадочных жидкостях культур мезенхимных и эпителиальных стволовых клеток из пуповины высокую степень (в сравнении со стволовыми клетками из костного мозга и жировой ткани, фибробластами кожи человека и кератиноцитами кожи) секреции активина А и фоллистатина (хорошо известно, что оба этих белка способствуют репарации и регенерации тканей, усилению ангиогенеза и поддержанию культур эмбрионных стволовых клеток, так что экспрессия соответствующих генов служит признаком эмбрионных свойств и способности клеток к дифференцировке). Все эти результаты указывают на то, что клетки согласно настоящему изобретению являются многообещающими кандидатами для их терапевтического применения в таких связанных с клетками областях, как регенеративная медицина, медицина старения, восстановление тканей и конструирование тканей.
Мезенхимные клетки были дополнительно охарактеризованы путем анализа секретируемых цитокинов и факторов роста в сравнении с мезенхимными стволовыми клетками из костного мозга человека. Эпителиальные стволовые клетки из пуповины (UCEC) анализировали, сравнивая их с кератиноцитами эпидермиса человека. Этот анализ проводили следующим образом. Вкратце, UMMC, UCEC, фибробласты кожи, мезенхимные клетки костного мозга, кератиноциты эпидермиса культивировали в ростовой среде до 100% слияния (37°С, 5% CO2) и затем синхронизовали в голодной среде (DMEM без сыворотки) в течение 48 ч. На следующий день среду заменяли на новую среду, вновь DMEM без сыворотки, и после этого клетки культивировали еще в течение 48 ч. Среды после культивации собирали, концентрировали и анализировали на наличие цитокинов, используя набор Cytokine Array (RayBiotech, Inc, GA, USA).
Результаты этого анализа показывают, что UCMC секретируют интерлейкин-6 (IL-6); MCP1; фактор роста гепатоцитов (HGF); интерлейкин-8 (IL-8); sTNFRI; GRO; Т1МР1; TIMP2; TRAILR3; uPAR; ICAM1; IGFBP3; IGFBP6 (фиг.11), тогда как UCEC секретируют IGFBP-4; PARC; EGF; IGFBP-2; IL-6; ангиогенин; GCP-2; IL-1Rα; МСР-1; RANTES; SCF; TNFβ; HGF; IL-8; sTNFR; GRO; GRO-α; амфирегулин; IL-1R4/ST2; Т1МР1; TIMP2; uPAR; VEGF (фиг.12).
Поэтому это показывает, что оба типа клеток секретируют большие количества цитокинов и факторов роста, играющих важную роль в биологии развития, гомеостазе тканей, восстановлении и регенерации тканей и ангиогенезе. Это дополнительно демонстрирует многосторонность клеток согласно настоящему изобретению при использовании в соответствующих терапевтических применениях.
Кроме того, клетки согласно настоящему изобретению дополнительно испытывали на предмет показателей их безопасности, используя в качестве индикатора тест на образование мышиной тератомы. В этих опытах были взяты шесть мышей линии SCID. В бедренную мышцу каждой мыши SCID стерильной иглой 25G производили инъекцию суспензии, содержавшей более 2 миллионов клеток UCMC. Животных содержали до 6 месяцев и оценивали у них образование опухолей. У этих мышей не было обнаружено никакого образования опухолей (данные не приведены). Это указывает на то, что клетки согласно настоящему изобретению безопасны и не имеют никакой способности к образованию раковых опухолей, доброкачественных опухолей или чего-либо иного.
Пример 4
Культивирование стволовых/прародительских клеток в бессывороточной среде
Клетки UCMC культивировали в DMEM с 10% PCS и в средах без сыворотки - РТТ-1, РТТ-2 и РТТ-3. Эти три среды (РТТ-1, РТТ-2 и РТТ-3) были приготовлены одним из настоящих заявителей - д-ром Phan. Вкратце, эти три среды не содержали сыворотки плода коровы или сыворотки человека, но содержали различные цитокины и факторы роста, такие как IGF, EGF, TGF-β, активин А, факторы BMP, PDGF, трансферрин и инсулин. Компоненты факторов роста варьировали от среды к среде, чтобы получить различные характеристики роста. Культивирование проводили следующим образом. В базовые среды добавляли различные количества факторов роста и цитокинов. Клетки UCMC размораживали и выдерживали в этих средах в течение 10 дней. Мониторинг пролиферации клеток проводили с помощью световой микроскопии.
Фиг.13 показывает хороший рост клеток UCMC в 4 различных группах сред (фигуры с 13-1 по 13-5), причем морфология клеток UCMC различалась, в зависимости от соотношения или доли цитокинов или факторов роста, находящихся в соответствующей среде. Напротив, мезенхимные клетки, полученные из костного мозга или жировой ткани, не обнаруживали хорошего роста в этих бессывороточных средах (фигуры 13-6 и 13-7). Поэтому хороший рост клеток UCMC демонстрирует активность клеток согласно настоящему изобретению и их высокую жизнеспособность, что указывает на превосходство их ростовых характеристик над характеристиками полученных из обычных источников мезенхимных клеток (из костного мозга и жировой ткани). В этом отношении следует отметить, что в этих опытах была использована среда без сыворотки (крупного рогатого скота) и что большинство человеческих мезенхимных клеток не дает хорошего роста в системах с бессывороточной средой. Поэтому использование клеток согласно настоящему изобретению в сочетании с определенными технологиями бессывороточных сред является большим преимуществом для клеточной терапии, поскольку исключаются опасности, связанные с использованием для культивирования и проращивания клеток сыворотки коровьего плода (хотя использование сыворотки крупного рогатого скота практиковалось в течение долгого времени и, как правило, оптимизирует рост клеток, беспокойство по поводу ее использования возросло в связи с распространением зоонозов, таких как губчатая энцефалопатия крупного рогатого скота («коровье бешенство»)).
Пример 5
Оценка профиля экспрессии генов в эпителиальных и мезенхимных стволовых клетках пуповины
Профиль экспрессии генов в эпителиальных и мезенхимных стволовых клетках пуповины анализировали с помощью ДНК-микрочипов. Для этой цели клетки UCMC и UCEC культивировали в ростовой среде при 37°С, 5% CO2 до достижения 100% слияния. Клетки синхронизовали в базовой среде в течение еще 48 ч, а затем среду заменяли на свежую базовую среду еще на 48 ч. Выделяли общую РНК и направляли ее в Silicon Genetics Microarray Service. Анализ данных проводили с помощью программы GeneSpring 7.2. На фиг.14 суммированы данные по общей экспрессии генов. Клетки UCEC экспрессировали всего 28055 генов, а клетки UCMC экспрессировали всего 34407 генов. Имеется 27308 перекрывающихся генов, экспрессирумых в обоих типах клеток. Из экспрессируемых генов 747 генов были уникальными для UCEC, a 7099 генов были уникальными для UCMC. Отобранные представляющие интерес гены указаны на фиг.14.
Оба типа стволовых клеток экспрессировали 140 генов, относящихся к эмбрионным стволовым клеткам и развитию эмбриона, что дополнительно подтверждает, что клетки согласно настоящему изобретению имеют свойства, подобные свойствам эмбрионных стволовых клеток. Продукты этих генов следующие: Nanog; альфа-зародышевый белок; транскрипционный фактор 3 пред-В-клеточного лейкоза; ламинин-α5; подобный карциноэмбриональному антигену фактор 1; содержащий ангидролазный домен фактор 2; дельта-подобный фактор 3 (Drosophila); подобный Muscleblind фактор (Drosophila); комплексный локус GNAS; родственная карциноэмбрионному антигену молекула 3 клеточной адгезии; тиоэстераза 2 пальмитоилированного белка; специфический для беременности β-1-гликопротеин-2; подобный карциноэмбрионному антигену фактор 1; фактор развития эктодермы эмбриона; материнская эмбрионная киназа с лейциновой застежкой; хориальный соматомаммотропиновый гормон 2; рамка Forkhead box D3; гомолог фактора обрамления нервной веточки (Drosophila); представитель 1В семейства кинезинов; тяжелый полипептид 3 миозина скелетных мышц эмбриона; фактор нарушения различения верх/низ (эктродактилия) тип 3; представитель 3 семейства доменов TEA; α-1-ламинин; хориальный соматомаммотропиновый гормон 1; лактоген плаценты; рецептор 1 гормона высвобождения кортикотропина; тиротропный эмбрионный фактор; ядерный транслокатор 2 арил-углеводородного рецептора; родственный закручивающему мембрану белку фактор; нейрегулин 1'; α-1-коллаген тип XVI; нейрегулин 1; хориальный соматомаммотропиновый гормон 1 (лактоген плаценты), триплетный повтор CUG; связывающийся с РНК белок 1; подобный бустину хориальный соматомаммотропиновый гормон 1 (лактоген плаценты); ингибитор семейства MyoD (семейный ингибитор); индуцируемый ретинойной кислотой фактор 2; транскрипционный фактор 4 пред-В-клеточного лейкоза; α-2-ламинин (мерозин, фактор врожденной мышечной дистрофии); гомолог 1 SMAD - материнский фактор против DPP (Drosophila); транскрибируемая последовательность Homo sapiens с умеренным подобием белку pir:D28928 (Н.sapiens); D28928 - специфический для беременности у человека β-1-гликопротеин IB, абортивный фрагмент; подобный Bruno фактор 4; связывающийся с РНК белок (Drosophila); специфический для мозга зародыша белок; индуцируемый беременностью ингибитор роста; гомолог 5 SMAD - материнский фактор против DPP (Drosophila); хориальный соматомаммотропиновый гормон 2; активируемый аденилат-циклазой полипептид 1 гипофиза; родственная карциноэмбрионному антигену молекула клеточной адгезии; α-3-ламинин; белок о-фукозилтрансферазы 1; фактор Jagged 1 (синдром Alagille); изогнутый гомолог 1 фактора гаструляции (Drosophila); подобный ELAV фактор 3 (фактор летальности эмбриона, нарушение зрения, Drosophila) (человеческий антиген Нu antigen С); тиротропный эмбриональный фактор; фактор 3 семейства 43 переносчиков растворителя; Inversin; фактор 2 детского нефротуберкулеза; фактор инверсии направления поворота зародыша; транскрипционный вариант 2 (мРНК) инверсина Homo sapiens (INVS); транскрибированные последовательности Homo sapiens; последовательность Homeo box D8; ассоциированный с Fyn эмбриональный субстрат; подобный ELAV фактор 1 (фактор летальности эмбриона, нарушение зрения, Drosophila) (человеческий антиген Нu antigen R); фактор класса В2, содержащий основной домен спираль-петля-спираль; рецептор окситоцина; фактор роста 1 из тератокарциномы; родственная Fms тирозин-киназа 1 (фактор роста сосудов эндотелия/рецептор фактора сосудистой проницаемости); адреномедулин; триплетный повтор CUG коактиватора 6 ядерных рецепторов (связывающийся с РНК белок 1); родственная карциноэмбрионному антигену молекула клеточной адгезии 4; тип R рецептора белка тирозин-фосфатазы; ортолог минимального участка мышиного фактора Acrg летальности эмбриона; рецептор A3 ЕРН; Delta-подобный фактор 1 (Drosophila); назальный фактор LHRH эмбриона; подобный СР2 транскрипционный фактор 1; фактор нарушения различения верх/низ (эктродактилия) тип 3; фактор Jagged 2; фактор нарушения различения верх/низ (эктродактилия) тип 1; α-субъединица трифункционального белка оксиацил-кофермент А дегидрогеназа/3-кетоацил-кофермент А тиолаза/еноил-кофермент А гидратаза; фукозилтрансфераза (α-(1,3)-фукозилтрансфераза); родственная карциноэмбрионному антигену молекула клеточной адгезии 7; тип 2 фактора нуклеофосмин/нуклеоплазмин; α-рецептор, транспортер Fc-фрагмента IgG; фактор 3 Homo sapiens контакта мать-зародыш, подобный сортировщику 35 белков вакуолей (LOC146485), мРНК; мышиный гомолог фактора Т брахиурии; дизинтегрин А и домен 10 металлопротеиназы; рибосомный белок L29; конвертирующий эндотелин фермент 2; трофинин; α-4-ламинин; последовательность Homeo box В6; гипотетический белок FLJ13456; NACHT - богатый лейцином повтор и содержащий PYD фактор 5; фактор 1 транскрипции недифференцированных клеток эмбриона; паппализин 1 - ассоциированный с беременностью белок плазмы А; представитель 1 семейства 1А секретоглобинов (утероглобин); подобный паратироидному гормону гормон; родственная карциноэмбрионному антигену молекула 1 клеточной адгезии (желчный гликопротеин).
Оба типа стволовых клеток экспрессировали также тысячи генов, связанных с биологией развития, ростом и дифференцировкой клеток, клеточным гомеостазом, репарацией и регенерацией клеток и тканей. Примерами таких продуктов являются факторы роста и их рецепторы (G-CSF, факторы FGF, факторы IGF, KGF, NGF, факторы VEGF, PIGF, ангиопоэтин, CTGF, факторы PDGF, HGF, EGF, HDGF, TGF-β, активины и ингибины, фоллистатин, факторы BMP, SCF/c-Kit, LIF, факторы WNT, факторы SDF, онкостатин М, интерлейкины, хемокины и многие другие); белки ММР, белки внеклеточных матриксов TIMP (коллагены, ламинины, фибронектины, витронектины, тенасцины, интегрины, синдеканы, декорин, фибромодулин, протеогликаны, sparc/остеонектин, муцин, нетрин, глипикан, ассоциированный с хрящами белок, матрилин, гиалуронан, фибулин, ADAMTS, бигликан, дискоидин, компоненты десмосом, белки iCAM, кадерины, катенины и многие другие); цитокератины.
Существуют группы генов, присутствующих только в клетках UCMC. Эти гены связаны со следующим: с нормальными физиологическими процессами (инсулино-подобный фактор роста 1 (соматомедин С); инсулино-подобный фактор 4 из плаценты; релаксин 1; плазминоген; инсулино-подобный фактор 5; инсулино-подобный фактор роста 2 (соматомедин А), с гомеостазом (подобный радиальному spokehead фактор 1; фактор гемохроматоза; лиганд 5 хемокина (лейтмотив С-С); рецептор А интерлейкина-31; лиганд 12 хемокина (фактор 1 из стромальных клеток) (лейтмотив С-Х-С); представитель 2 группы С подсемейства 3 ядерных рецепторов; лиганд 23 хемокина (лейтмотив С-С); митохондриальный ферритин; коактиватор 1-α пролиферативно активируемого гамма-рецептора пероксисом; поверхностно-активное вещество - ассоциированный с легкими белок D; лиганд 11 хемокина (лейтмотив С-С); лиганд 3 хемокина (лейтмотив С-С); гомолог 2 эглонина (С.elegans); коактиватор 1-β пролиферативно активируемого гамма-рецептора пероксисом; лиганд 1 хемокина (лейтмотив С-С); α-2(+) полипептид транспортной Na+/K+АТФазы; лиганд 2 хемокина (лейтмотив С); гемопексин; рецептор 3 рианодина), с морфогенезом (α-спектрин - эритроцитный фактор 1 (фактор эллиптоцитоза 2); последовательность Homeo box D3; гомолог 1 фактора отсутствия глаз (Drosophila); представитель J семейства генов гомологов Ras; специфический для лейкоцитов транскрипт 1; рецептор эктодисплазина А2; глипикан 3; ген 7 спаренной последовательности; сериновая протеаза корин; гомолог 1 продукта гена Dishevelled (dsh, Drosophila); последовательность T-box 3 (локтевой синдром молочной железы); хондроитин-β-1,4-N-ацетилгалактозаминил-трансфераза; последовательность «box 10» SRY (область определения пола хромосомы Y); тяжелый полипептид 9 немышечного миозина; «бахромчатый» гомолог рецептора лютеинизирующего гормона/хориогонадотропина (Drosophila); секретируемый родственный бахромчатому рецептору белок 5; фактор 11 семейства сайтов интеграции MMTV (признак бескрылости); гомолог фактора 2 отсутствия глаз (Drosophila); фактор, родственный белку гена Muscleblind (Drosophila); последовательность T-box 5; подобный Mab-21 фактор 1 (С.elegans); специфический для торможения роста фактор 2; гомолог 1 фактора связанной с полом гребенчатости середины ног (Drosophila); последовательность T-box 6; связывающийся с филамином белок LIM-1; молекула адгезии клеток меланомы; гомолог 1 фактора закручивания (акроцефалосиндактилия 3, синдром Saethre-Chotzen (Drosophila); последовательность Homeo box A11; кератокан; кислый фактор роста 1 фибробластов; карбоксипептидаза М; эффекторный белок 4 CDC42 (связывающийся с Rho-ГТФазой); фактор транскрипции β-1 последовательности LIM homeobox; гомолог 1 продукта гена Engrailed; фактор роста 8 фибробластов (индуцируемый андрогенами); фактор роста 18 фибробластов; специфический для лейкоцитов транскрипт 1; эндотелин 3; подобный спаренному фактор транскрипции гомеодомена), с развитием эмбриона (специфический для беременности β-1-гликопротеин 3; подобный ELAV (embryonic lethal, abnormal vision, Drosophila) фактор 4 (человеческий антиген Нu antigen D); сопряженный с G-белком рецептор 10; рецептор эктодисплазина А2; фактор 4 подсемейства В кассеты связывания с АТФ (MDR/TAP); специфический для беременности β-1-гликопротеин 11; назальный фактор LHRH эмбрионов; релаксин 1; гомолог 4 продукта гена Notch (Drosophila); специфический для беременности β-1-гликопротеин 6; pih-2P; индуцируемый беременностью связанный с синдромом гипертензии белок Homo sapiens (PIH2); гликопротеин 1 яйцевода 120 кДа (муцин 9, овидуктин); связанный с прогестагеном белок эндометрия; легкий щелочной полипептид 4 предсердного миозина зародыша; пролактин; фактор транскрипции 1 пред-В-клеточного лейкоза; гомолог фактора окраинной бахромчатости (Drosophila); высвобождающий кортикотропин гормон; фактор 2 группы С подсемейства 3 ядерных рецепторов; нейрегулин 2; подобный продукту гена Muscleblind фактор (Drosophila); кДНК FLJ27401 fis Homo sapiens, клон WMC03071; последовательность, подобная homeobox 1, в экстраэмбрионном сперматогенезе; инсулино-подобный фактор 4 плаценты; экзон В2С гена человеческого процессированного, специфичного для ложной беременности гликопротеина PSG12, содержащий 3'-нетранслируемые участки двух альтернативных сайтов сплайсинга С1 и C2; связанная с Fms тирозин-киназа 1 (рецептор фактора роста эндотелия сосудов/фактора сосудистой проницаемости); специфический для беременности β-1-гликопротеин 3; родственная карциноэмбриональному антигену молекула 1 клеточной адгезии (желчный гликопротеин); микросомная стероидная сульфатаза (изофермент S арилсульфатазы С); последовательность Homeo box B6; протеин-O-фукозилтрансфераза 1; β-фактор 1 транскрипции последовательности LIM homeobox; β-полипептид фолликулостимулирующего гормона; ангиотензин (фактор 8 класса А (α-1-антипротеиназа, антитрипсин) ингибиторов сериновых или цистеиновых протеиназ); родственная карциноэмбриональному антигену молекула 6 клеточной адгезии (неспецифический перекрестно реагирующий антиген); α-связывающий белок протеин-киназы С; представитель 10 подсемейства коллектинов (лектин С-типа); α-1-ламинин; карбоксиэстераза 1 межклеточного пространства (сериновая эстераза 1 моноцитов/макрофагов); фибробластный фактор роста 5; прогастрицин (пепсиноген С); связанный со спермой антиген 11; пропротеин-конвертаза субтилизин/кексин, тип 2; связывающийся с гиалуронаном белок 2; секретируемый короткий основной домен 3F иммуноглобулина (домен Sema, семафорин); интерлейкин-2; подобные химотрипсину ферменты при заболевании Norrie (псевдоглиома); трахеобронхиальные/желудочные подтипы А и С муцина 5; карбоксипептидаза В2 (плазматическая карбоксипептидаза U); α-меприн А (РАВА пептид-гидролаза); предшественник 1 тахикинина (вещество K, вещество Р, нейрокинин 1, нейрокинин 2, нейромедин L, нейрокинин-α, нейропептид K, нейропептид-γ); фибробластный фактор роста 8 (индуцируемый андрогенами); фибробластный фактор роста 13; гемопексин; фактор 2 раннего начала рака молочной железы; фибробластный фактор роста 14; фактор 1 юношеского, связанного с Х-хромосомой ретиношизиса; подобный хитиназе 3 фактор 1 (гликопротеин-39 хрящей); дистонин; фактор 2 семейства 1D секретоглобулинов; ноггин; сердцевинный домен 2 WAP с 4 дисульфидными связями; подобный антигену CDS фактор (семейство богатых цистеином рецепторов-чистильщиков); ответственный за скрейпи белок 1; гомолог гремлина 1 (суперсемейство с цистеиновым узлом, Xenopus laevis); интерлейкин-16 (фактор хемоаттракции лимфоцитов); лиганд 26 хемокина (лейтмотив С-С); Nucleobindin 1; фибробластный фактор роста 18; связывающийся с инсулино-подобным фактором роста белок 1; поверхностно-активное вещество - связанный с легкими белок А1; подобный Delta гомолог 1 (Drosophila); регулируемый кокаином и амфетамином транскрипт; β-меприн А; интерлейкин-17F; фактор комплемента Н; богатый цистеином секреторный белок 2; дистонин; сердцевинный домен 1 WAP с 4 дисульфидными связями; пролактин; поверхностно-активное вещество - связанный с легкими белок В; фибробластный фактор роста 5; гомолог 2 Dickkopf (Xenopus laevis); ассоциированный со спермой антиген 11; лиганд хемокина 11 (лейтмотив С-С); α-меприн А (РАВА пептид-гидролаза); подобный хитиназе 3 фактор 2; индуцируемый C-fos фактор роста (фактор D роста эндотелия сосудов); лиганд 4 хемокина (лейтмотив С-С); рецептор вируса полиомы; гиалуроноглюкозаминидаза 1; лиганд 9 хемокина (лейтмотив С-Х-С); амелогенин (сцепленный с Х-хромосомой синдром amelogenesis imperfecta 1); релаксин 1; Sparc/osteonectin (подобный доменам cwcv и kazal протеогликан, тестикан); лиганд 26 хемокина (лейтмотив С-С); кислый фибробластный фактор роста; подобный ангиопоэтину фактор 2; дистонин; транскобаламин II; фактор макроцитной анемии; лиганд 1 хемокина (лейтмотив С-С); кислотолабильная субъединица инсулино-подобного фактора роста; фактор Н комплемента; специфический для беременности β-1-гликопротеин 6; гомолог Silver (мышь); протеогликан 4; фибробластный фактор роста 16; подобный цитокину белок С17; гранулизин; ангиопоэтин 2; хромогранин В (секретогранин 1); домен Sema 7A (домен иммуноглобулина и мембранный якорь GPI, семафорин); плейотропин (связывающийся с гепарином фактор роста 8, способствующий росту нейритов фактор 1); представитель 3 семейства Са-активируемых хлоридных каналов; представитель 1 семейства 1D секретоглобулинов; фибулин 1; рецептор 1 фосфолипазы А2, 180 кДа (и фактор 1 внеклеточного матрикса, подобный ADAMTS); периостин (остеобласт-специфический фактор); глипикан 5; богатый лейцином нейронный фактор 3 с повторами; трансглутаминаза 2 (С-полипептид, протеин глутамин-γ-глутамилтрансфераза); фактор 2, подобный металлопротеазе (типа репролизина) с лейтмотивом тромбоспондина 1 типа; ассоциированный с микрофибриллами белок 4; глипикан 3; α-3-коллаген, тип V; тканевый ингибитор металлопротеиназы 2; кератокан; олигомерный матриксный белок хряща; лумикан; связывающий гиалуронан с протеогликаном белок 3; статерин; фактор 3, подобный металлопротеазе (типа репролизина) с лейтмотивом тромбоспондина 1 типа; белок внеклеточного матрикса Spondin 1; α-3-коллаген, тип IV (антиген Goodpasture); представитель 7В семейства сайтов интеграции в синдроме MMTV типа Wingless; α-2 коллаген, тип VI; липокалин 7; связывающий гиалуронан с протеогликаном белок 4; фактор 5, подобный металлопротеиназе (типа репролизина) с лейтмотивом тромбоспондина 1 типа (аггреканаза-2); фибронектин 1; матрилин 1; белок матрикса хряща; гипотетический белок FLJ13710; хондроитин β-1,4-N-ацетилгалактозаминилтрансфераза; металлопротеиназа 16 матрикса (включенная в мембрану); фактор фон-Виллебрандта; α-2-коллаген, тип VI; сериновая трансмембранная протеаза 6; металлопротеиназа 23В матрикса; металлопротеиназа 14 матрикса (включенная в мембрану); нейронный фактор 3 с богатыми лизином повторами; подобный SPARC фактор 1 (mastQ, гевин); Sparc/osteonectin 3 (подобный доменам cwcv и kazal протеогликан, тестикан); дерматопонтин; α-1 коллаген, тип XIV (ундулин); связанный с Y-хромосомой амелогенин; нидоген (энактин); подобный ADAMTS фактор 2; связанный с гиалуронаном и протеогликаном белок 2; α-1-коллаген, тип XV; глипикан 6; металлопротеиназа 12 матрикса (эластаза макрофагов); фактор 15, подобный дизинетгрину, и металлопротеаза (типа репролизина) с лейтмотивом типа 1 тромбоспондина; сериновая трансмембранная протеаза 6; фактор 16, подобный дизинетгрину, и металлопротеаза (типа репролизина) с лейтмотивом типа 1 тромбоспондина; фактор 20, подобный дизинетгрину, и металлопротеаза (типа репролизина) с лейтмотивом типа 1 тромбоспондина; α-1-коллаген, тип XI; связывающийся с гиалуронаном и протеогликаном белок 1; хондроитин β-1,4-N-ацетилгалактозаминилтрансфераза; аспорин (LRR класс 1); α-1-коллаген, тип III (аутосомный детерминант, синдром Ehlers-Danlos, тип IV); секретируемый фосфопротеин 1 (остеопонтин, сиалопротеин I костей, фактор 1 ранней активации Т-лимфоцитов); белок Gla матрикса; фибулин 5; тканевый ингибитор мет аллопротеиназы 3 (псевдовоспалительная дистрофия Sorsby fundus); α-1-коллаген, тип XXV; олигомерный белок матрикса хряща; α-1-коллаген, тип VI; хондроадгерин; α-1-коллаген, тип XV; α-4-коллаген, тип IV; кислый фосфопротеин матрикса дентина; α-1-коллаген, тип IV; фактор 1, содержащий повтор тромбоспондина; металлопротеиназа 16 матрикса (включенная в мембрану); α-2-коллаген, тип I; β-текторин; специфическая к гликозилфосфатидил-инозитолу фосфолипаза D1; ген 1, регулируемый с усилением экспрессии при колоректальном раке), с цитоскелетом (β-филамин В, (связывающийся с актином белок 278); EF-сторонний белок 1, центрин; содержащий домен PERM фактор 3; межклеточный интегратор 3; γ-парвин; Rho-фактор 11 обмена гуаниновыми нуклеотидами (GEF); тирозинкиназа 2; подобный Kelch фактор 4 (Drosophila}; β-спектрин; эритроцитный взаимодействующий с Arg/Abl белок ArgBP2 (участвует в склероцитозе, клинический тип I); адвилин; содержащий спектриновый повтор белок 1 ядерной оболочки; δ-1-катенин (связанный с cadherin белок); белок 5, подобный мембранному белку эритроцитов, линия 4.1; α-2-катенин (связанный с cadherin белок); лиганд 3 хемокина (лейтмотив С-С); γ-саркогликан (ассоциированный с дистрофином гликопротеин 35 кДа); небулин; β-тимозин, идентифицированный в клетках нейробластомы; зависимая от 3-фосфоинозитида протеин-киназа 1; белок, взаимодействующий с белком синдрома Wiskott-Aldrich; белок 1, взаимодействующий с Huntingtin; продукт гена KIAA0316; мышечный тропомодулин 4; фактор 1, делегированный при раке печени; подобный Villin фактор; β-1-синтрофин (щелочной компонент 1 связанного с дистрофином белка А1, 59 кДа); cGMP-зависимая протеин-киназа, тип I; подобный кератину 8 белок Homo sapiens; цитокератин 8; мРНК цитоскелетного кератина 8 II-го типа (LOC345751); α-аддуцин 1; протеин-киназа С и субстрат 3 казеин-киназы в нейронах; группа крови Kell; взаимодействующий с филамином А белок 1; специфический фактор 2 запрета роста; открытая рамка считывания хромосомы 1; подобный Stathmin фактор 2; α-спектрин 1 эритроцитов (синдром эллиптоцитоза 2); ген FKSG44; представитель 1C семейства кинезинов; тензин; каптин (связывающийся с актином белок); нейрофибромин 2 (билатеральная акустическая неврома); имеющие гомологию с Pleckstrin домены Sec7 и клубковый домен 2 (цитогезин-2); родственный актину белок Т1 (синдром, подобный синдрому Wiskott-Aldrich); подобный Kelch фактор 4 (Drosophila); связывающийся с актином белок, гомолог 1 Fascin (Strongylocentrotus purpuratus); амфифизин (синдром Stiff-Man с аутоантигеном 128 кДа рака молочной железы); подобный белку 2 поликистоза почек фактор 1; нейронный анкирин 2; связывающаяся с CDC42 α-протеин-киназа (подобная DMPK); гипотетический белок FLJ36144; взаимодействующий с Arg/AbI белок ArgBP2; подобный формину фактор 3; β-1-катенин (связанный с cadherin белок 88 кДа); профилин 2; подобный синаптоподину 2 фактор; γ-2-синтрофин; фосфолипаза D2; фактор 2 поглощения и клеточной подвижности (гомолог ced-12, С.elegans); легкий полипептид 68 кДа нейрофиламентов; дистонин; подобный актину белок 7В; член 1C семейства кинезинов; домен 3 PDZ и LIM; β-2-аддуцин; обскурин (цитоскелетный кальмодулин и взаимодействующий с титином RhoGEF; β-полипептидный паралог тубулина; взаимодействующий с филамином А белок 1; талин 1; белок Homo sapiens, подобный [сегменту 1 из 2] белка Piccolo (Aczonin) (LOC375597); CDC42 эффекторный белок 4 (связывающийся с Rho ГТФазой); синдекан 1; α-филамин А (связывающийся с актином белок 280); профилин 2; подобная тензину содержащая домен С1 фосфатаза; гипотетический белок MGC33407; ГТФаза 1 семейства Rho; флавопротеин-оксидоредуктаза MICAL2; Са2+-зависимый активатор секреции; подобный рабфилину 3А фактор (без доменов С2); миозин XVA; цГМФ-зависимая протеин-киназа, тип I; взаимодействующий с регуляторной легкой цепью миозина белок; представитель 13В семейства кинезинов; мышечный гомолог онкогена RAS; не-эритроцитный β-спектрин-1; ТАО-киназа 2; β-филамин В (связывающийся с актином белок 278); нейрофибромин 2 (билатеральная акустическая фиброма); α-3-катенин (ассоциированный с cadherin белок); связывающийся с актином белок коронин 1А; не-эритроцитный β-спектрин-4; сцепленный с Y-хромосомой β-4-тимозин; тестикулярный тектин-2; представитель J семейства генов гомологов Ras; серин/треонин-киназа с доменами гомологии DbI и pleckstrin; β-дистробревин; γ-2-актин гладких брюшных мышц; подобный Tara белок; каспаза 8 (связанная с апоптозом цистеиновая протеаза); фактор 10, содержащий повтор Kelch и домен ВТВ (POZ); трансмембранный муцин-1; связанный с микротрубочками белок tau; тензин; представитель F семейства генов гомологов Ras (в филоподиях); α-4-актинин; мембранный белок эритроцитов, зона 4.1 (связанный с RH эллиптоцитоз 1); гомолог 2 Bicaudal D (Drosophila); анкирин 3, узел Ranvier (анкирин G); миозин VIIA (острый синдром 1В Usher, аутосомно-рецессивный); человеческий цитоскелетный белок (Homo sapiens) тип II, подобный кератину 8 (LOC285233); тестикулярный гомолог 3 фасцина (связывающийся с актином белок); структурный белок 2 бусинчатых волокон (факинин); десмин; миозин X; индуцируемый сигналом связанный с пролиферацией ген 1; скиндерин; подобный коактозину белок 1 (Dictyostelium); β-4-тубулин; содержащий домен FERM фактор 4А; α-1-актин скелетных мышц; талин 1; кальдесмон 1; α-1-синтрофин (кислый компонент связанного с дистрофином белка А1, 59 кДа); β-2-аддуцин; взаимодействующий с филамином А белок 1; белок 4В, подобный белку мембраны эритроцитов, зона 4.1; связывающийся с FYN белок (FYB-120/130); мостиковый интегратор), с внеклеточным пространством (подобный дизинтегрину белок 20 и металлопротеаза (типа репролизина) с лейтмотивом типа тромбоспондина 1-го типа; подобный SPARC белок 1 (mastQ, hevin); представитель 1 семейства ингибиторов сериновых (или цистеиновых) протеиназ, класс G (ингибитор С1) (наследственная ангиоэдема); подобный химотрипсину урокортин; β-полипептид фактора роста из тромбоцитов (гомолог онкогена вируса обезьяньей саркомы (v-sis)); белок-предшественник связывающегося с BMP эндотелиального регулятора; фактор комплемента Н; фактор 1, подобный соматомаммотропиновому гормону хориона; лиганд 18 (легочный и регулируемый с активацией) хемокина (лейтмотив С-С); фибронектин 1; специфический для беременности β-1-гликопротеин 3; подобный дизинтегрину белок 3 и металлопротеаза (типа репролизина) с лейтмотивом типа тромбоспондина 1-го типа; CocoaCrisp; подобный инсулину фактор 4 (плацента); представитель 11 семейства сайтов интеграции MMTV типа Wingless; олигомерный белок матрикса хряща; трансмембранная сериновая протеаза 6; индуцируемый C-fos фактор роста (фактор роста D эндотелия сосудов); член В семейства с подобием последовательностей 12 (при эпидидимите); регуляторная субъединица 9В протеин-фосфатазы 1 (спинофилин); транскобаламин II (макроцитная анемия); фактор коагуляции V (проакселерин, лабильный фактор); фосфолипаза А2, группа IID; α-индуцированный белок 6 фактора некроза опухоли; α-1-коллаген, тип XV; связывающий гиалуронан с протеогликаном белок 3; α-1-коллаген, тип XIV (ундулин); интерлейкин-19; ингибитор протеаз 15; β-полипептид 1 холинергического никотинового рецептора из мышц; подобный лизил-оксидазе белок 3; связывающийся с инсулино-подобным фактором роста белок 5; гормон роста 1; β-казеин; подобный NEL фактор 2 кур; комплемент фактора I; лиганд 23 хемокина (лейтмотив С-С); интерферон-α2; включенная в мембрану металлопротеаза 16 матрикса; металлопротеиназа 12 матрикса (эластаза макрофагов); глипикан 5; специфический для беременности β-1-гликопротеин 3; фибробластный фактор роста 6; гомолог гремлина 1, суперсемейство с цистеиновым узлом (Xenopus laevis); α-белок S; хондроитин β-1,4-N-ацетилгалактозаминил-трансфераза; специфическая к гликозилфосфатидил-инозитолу фосфолипаза D1; кислый фактор роста 1 из фибробластов; внеклеточный белок матрикса спондин 1; белок 1 морфогенеза костей; поверхностно-активное вещество, связанный с легкими белок В; кислый фосфопротеин матрикса дентина; липопротеин Lp(a); трансмембранный муцин 1; связывающая лектин с маннаном сериновая протеаза (активирующий компонент Ra-реактивного фактора); β-меприн А; представитель 1 семейства 1D секретоглобулинов; аспорин (класс 1 LRR); лиганд 25 хемокина (лейтмотив С-С); подобный цитокину белок С17; инсулино-подобный фактор 5; α-меприн А (гидролаза пептида РАВА); ответственный за скрейпи белок 1; фактор роста 18 из фибробластов; лиганд 9 хемокина (лейтмотив С-Х-С); β-ингибин В (β-полипептид активина АВ); индуцируемый андрогенами фактор роста 8 из фибробластов; гранулизин; регулируемый кокаином и амфетамином транскрипт; α-2 коллаген, тип I; лиганд 17 хемокина (лейтмотив С-С); лиганд 23 хемокина (лейтмотив С-С); Sparc/остеонектин (протеогликан 3 с доменами cwcv и подобным kazal, тестикан); β-3-А рецептор γ-аминомасляной кислоты (ГАМК); α-4-дефензин, кортикостатин; нейронный фактор 3 с богатыми лейцином повторами; глипикан 6; активируемая митогенами протеин-киназа 2; фактор коагуляции XI (предшественник тромбопластина плазмы); лиганд 5 хемокина (лейтмотив С-С); дистонин; родственный Frizzled белок; полипептид А1 фактора коагуляции XIII; инсулино-подобный фактор роста 1 (соматомедин С); гипотетический белок MGC45438; ассоциированный со спермой антиген 11; специфический фактор остеобластов периостин; α-2-макроглобулин; α-5-А рецептор γ-аминомасляной кислоты (ГАМК); представитель 3 семейства ингибиторов сериновых (или цистеиновых) протеиназ, класс А (α-1 антипротеиназа, антитрипсин); гомолог Silver мыши; родственный Frizzled белок; Chondroadherin; представитель С семейства рецепторов 3 5-окситриптамина (серотонина); α-2-коллаген, тип VI; подобный Toll рецептор 9; сцепленный с Y амелогенин; фактор роста В эндотелия сосудов; подобный Radial spokehead фактор 1; родственная Fms тирозин-киназа 1 (рецептор фактора роста сосудов эндотелия/рецептор фактора проницаемости сосудов); ингибитор 16 протеаз; интерлейкин-2; кластерин (ингибитор лизиса комплемента, SP-40,40, сульфированный гликопротеин 2, репрессируемый тестостероном фактор передачи 2 простаты, аполипопротеин J); β-полипептид фолликулостимулирующего гормона; фактор 16, подобный дизинетгрину и металлопротеазе (типа репролизина) с лейтмотивом тромбоспондина 1-го типа); лизоцим (амилоидоз почек); гомолог radical fringe (Drosophila); связывающийся с инсулино-подобным фактором роста белок 5; таксилин; аполипопротеин A-V; фактор роста С из тромбоцитов; подобный лиганду 3 хемокина (лейтмотив С-С) фактор 1; фактор роста 16 из фибробластов; α-2-коллаген, тип VI; представитель 1 класса С семейства ингибиторов сериновых (или цистеиновых) протеиназ (антитромбин); лиганд 11 хемокина (лейтмотив С-С); α-4-коллаген, тип IV; тирозин-киназа при агаммаглобулинемии Bruton; инсулино-подобный фактор роста 2 (соматомедин А); домен 1 ингибитора сериновых протеаз типа Kazal; α-полипептид А фибриногена; лиганд 1 хемокина (лейтмотив С-С); β-Е-ингибин; связывающийся с половым гормоном глобулин; α-1 коллаген, тип IV; лецитин-холестерин ацилтрансфераза; богатый цистеином секреторный белок 2; связывающий гиалуронан с протеогликаном белок 1; предшественник С Natriuretic пептида; представитель k6 семейства рибонуклеаз (РНКаз) А; фактор роста 14 из фибробластов; подобный ADAMTS фактор 2; α-3-коллаген, тип IV (антиген Goodpasture); ангиопоэтин 2; аполипопротеин L-3; лиганд 12 хемокина (лейтмотив С-Х-С) (фактор 1 клеток стромы); связывающийся с гиалуронаном белок 2; фактор коагуляции VII (ускоритель конверсии сывороточного протромбина); гликопротеин 1 яйцеводов, 120 кДа (муцин 9, овидуктин); белок матрикса хрящей Matrilin 1; трахеобронхиальные/желудочные подтипы А и С муцина 5; представитель 11b суперсемейства рецепторов фактора некроза опухолей (остеопротегерин); трансглутаминаза 2 (полипептид С, протеин-глутамин-γ-глутамилтрансфераза); кератокан; α-3-коллаген, тип V; домен 2 сердцевины WAP с 4 дисульфидными связями; лиганд 1 хемокина (лейтмотив С-Х3-С); представитель 1 класса D ингибиторов сериновых (или цистеиновых) протеиназ (кофактор гепарина); секреторный белок LOC348174; фактор коагуляции X; интерлейкин-16 (фактор хемоаттракции лимфоцитов); родственный панкреатической липазе белок 2; сериновая пептидаза 3 HtrA4; α-3 рецептор глицина; фактор, подобный CDS (семейство богатых цистеином рецепторов чистильщиков); гипотетический белок MGC39497; прокоагулянтный компонент фактора коагуляции VIII (гемофилия А); дерматопонтин; ноггин; содержащий секретируемый домен LY6/PLAUR фактор 1; подобный ADAMTS фактор 1; α-1-В гликопротеин; открытая рамка считывания 175 хромосомы 20; представитель 8В семейства сайтов интеграции ММТVтипа Wingless; фибулин 1; фибулин 6; катепсин S; нидоген (энактин); лиганд 26 хемокина (лейтмотив С-С); специфичная для клеток эндотелия молекула 1; подобный хитиназе 3 фактор 1 (гликопротеин-39 хрящей); β-1А рецептор γ-аминомасляной кислоты (ГАМК); представитель 2 семейства 1D секретоглобулинов; домен 7А Sema (домен иммуноглобулина и мембранный якорь, семафорин); фактор 15, подобный дизинтегрину и металлопротеазе с лейтмотивом тромбоспондина 1-го типа; субтилизин/кексин пропротеин-конвертазы 2-го типа; инсулино-подобный фактор роста 1 (соматомедин С); фактор 1 сцепленного с Х-хромосомой юношеского ретиношизиса; фактор 16, подобный дизинтегрину и металлопротеазе с лейтмотивом тромбоспондина 1-го типа; лиганд 2 хемокина (лейтмотив С); фактор роста 5 из фибробластов; связанный со спермой антиген 11; связанный с микрофибриллами белок 4; рецептор вируса полиомы; внеклеточная регулируемая сигналом киназа 8; сериновая трансмембранная протеаза 6; α-протеинкиназа С; подобный хитиназе 3 белок 2; интерлейкин-9; аполипопротеин L6; поверхностно-активное вещество - связанный с легкими белок А1; α-1-коллаген, тип VI; гипотетический белок FLJ 13710; карбоксипептидаза В2 плазмы (карбоксипептидаза U); подобный бактерицидному/повышающему проницаемость белку фактор 2; секретируемый фосфопротеин 1 (остеопорин, сиалопротеин 1 костей, фактор 1 ранней активации Т-лимфоцитов); сериновая пептидаза 3 HtrA; делегированный при раке печени фактор 1; специфическая для клеток эндотелия молекула 1; фактор Von Willebrand; фактор 5 (аггреканаза-2), подобный дизинтегрину и металлопротеазе (типа репролизина) с лейтмотивом тромбоспондина 1-го типа; короткий основной секретируемый домен 3А (домен Sema, домен иммуноглобулина, семафорин); лиганд 12 хемокина (лейтмотив С-Х-С) (фактор 1 из клеток стромы); статерин; тканевый ингибитор металлопротеиназы 3 (псевдовоспалительная дистрофия Sorsby fundus); фактор тромбоцитов 4 (лиганд 4 хемокина (лейтмотив С-Х-С)); поверхностно-активное вещество - связанный с легкими белок D; фактор комплемента Н; подобный Delta 1 гомолог (Drosophila); сердцевинный домен 1 WAP с 4 дисульфидными связями; кислотолабильная субъединица связывающегося с инсулино-подобным фактором роста белка; фактор раннего начала рака молочной железы 2; four-disulfide core domain 1; ген 1 пре-В лимфоцитов; кортикотропин высвобождающий гормон; гипотетический белок DKFZp434B044; индуцированный пролактином белок; RAS гуанил-высвобождающий белок 4; прогастрицин (пепсиноген С); короткий основной секретируемый домен 3F (домен Sema, домен иммуноглобулина, семафорин); ген 1, регулируемый с повышением активности при колоректальном раке; протеогликан 4; δ-полипептид никотинового холинергического рецептора; олигомерный белок матрикса хрящей; фактор крови группы АВО (трансфераза А, α-1-3-N-ацетилгалактозаминилтрансфераза; трансфераза В, α-1-3-галактозилтрансфераза); интерлейкин-12А (фактор 1 стимуляции естественных киллеров, фактор 1 созревания цитотоксических лимфоцитов, р35); фактор роста 7 из фибробластов (фактор роста кератиноцитов); родственник подобного IRRE фактора 3 (Drosophila); α-2-полипептид (нейрональный) никотинового холинергического рецептора; связанный с карциномой легких и эпителия носа палат; α-1-коллаген, тип XV; плейотропин (связывающийся с гепарином фактор роста 8, вызывающий рост нейритов фактор 1); ангиопоэтин-подобный фактор 2; заболевание Norrie (псевдоглиома); лиганд 3 хемокина (лейтмотив С-С); подобный хитиназе 3 фактор 1 (гликопротеин-39 хрящей); Inter-alpha (глобулин) ингибитор Н3; амелогенин (сцепленное с Х-хромосомой заболевание amelogenesis imperfecta 1); фактор роста эпидермиса (β-урогастрон); фактор роста 13 из фибробластов; представитель 7В семейства сайтов интеграции MMTV типа Wingless; γ-полипептид никотинового холинергического рецептора; специфический для беременности β-1-гликопротеин 6; включенная в мембрану металлопротеиназа 14; лиганд 26 хемокина (лейтмотив С-С); интерферон-α6; предшественник 1 тахикинина (вещество К, вещество Р, нейрокинин 1, нейрокинин 2, нейромедин L, нейрокинин-α, нейропептид К, нейропептид-γ); секретируемый относящийся к frizzled белок 5; связывающий гиалуронан с протеогликаном белок 4; компонент 4В комплемента; включенная в мембрану металопротеинаха 16; фактор роста 7 из фибробластов (фактор роста кератиноцитов); аполипопротеин C-II; представитель 3 семейства активируемых кальцием хлоридных каналов; тетранектин (связывающийся с плазминогеном белок); α-1-коллаген, тип III, (аутосомно-доминантный синдром Ehlers-Danlos, тип IV); белок KIAA0556; лиганд хемокина 4 (лейтмотив С-С); гемопексин; интер-альфа (глобулин) ингибитор Н1; релаксин 1; белок Gla матрикса; белок 2, подобный дизинтегрину и металлопротеазе (типа репролизина) с лейтмотивом тромбоспондина 1-го типа; рецептор 2 интерферонов α, β и ω; кислая фосфатаза из простаты; белок γ-8, связывающий гуаниновый нуклеотид (белок G); металлопротеиназа 23В матрикса; α-меприн А, (пептид-гидролаза РАВА); гиалуроноглюкозаминидаза 1; ангиотензиноген (фактор 8 класса А ингибиторов сериновых (или цистеиновых) протеиназ (α-1-антипротеиназа, антитрипсин)); белок промежуточного слоя хряща, нуклеотид-пирофосфогидролаза); пуринергический рецептор Р2Х, активируемый лигандом ионный канал 7; глипикан 3; β-тектроин; интерферон-α5; липокалин 7; вариант 1 фактора 4 тромбоцитов; Nucleobindin 1; α-1-коллаген, тип XI; желудочный ингибиторный полипептид; фактор 1, содержащий повтор тромбоспондина; фактор D семейства рецептора 3 окситриптамина (серотонина); α-1-коллаген, тип XXV; фактор 9 дифференцировки роста; гипотетический белок DKFZp434B044; эндотелин 3; лиганд 2 хемокина (лейтмотив С); прокинетицин 2; представитель 11b суперсемейства рецепторов фактора некроза опухоли (остеопротегерин); тканевый ингибитор металлопротеиназы; дистонин; хромогранин В (секретогранин 1); связывающий гиалуронан и протеогликан белок 2; нейрональный фактор с богатым лейцином повтором; лумикан; белок матрикса хряща матрилин 1; фосфолипаза А2, группа IIА (тромбоциты синовиальной жидкости); карбоксиэстераза 1 (сериновая эстераза 1 моноцитов/макрофагов); Sparc/остеонектин, протеогликан с доменами cwcv и подобным kazal (тестикан); гомолог 2 Dickkopf (Xenopus laevis); α-3 рецептор А γ-аминомасляной кислоты (ГАМК); специфический для беременности β-1-гликопротеин 11; связывающийся с инсулино-подобным фактором роста белок 1; дефензин β-106; интерлейкин-17F; субъединица активируемого лигандом ионного канала; рецептор 1 фосфолипазы А2, 180 кДа; фактор I (комплемент); гомолог 1 фактора LAG1 обеспечения длины (S. cerevisiae); пролактин; экспрессируемая в яичках последовательность 264; секретируемый домен 3D, домен Sema, домен иммуноглобулина, короткий основной домен (семафорин); секретируемый родственный frizzled белок 2; секретируемый родственный frizzled белок 4).
Имеется группа генов, присутствующих только в UCEC. Эти гены относятся к следующим процессам: гомеостаз (альбумин; чувствительный к кальцию рецептор; аквапорин 9; лактотрансферрин); морфогенез (последовательность Homeo box HB9; подобный эпителиальному V антиген 1); развитие эмбриона (релаксин 2; родственная карциноэмбрионному антигену молекула 8 клеточной адгезии; индоламин-пиррол 2,3-диоксигеназа; ЕРН рецептор A3; тиротропный фактор эмбриона; специфический к беременности β-1-гликопротеин 1; α-3-ламинин); внеклеточное пространство (поверхностно-активное вещество - ассоциированный с легкими белок А1; специфический к беременности β-1-гликопротеин 1; лактотрансферрин; TGF-α; альбумин; FGF-23; S100 связывающий кальций белок А9 (кальгранулин В)); внеклеточный матрикс (β-4-ламинин; α-3-ламинин; гликопротеин 4 Zona pellucida); активность структурных молекул (открытая рамка считывания 29 хромосомы 21; α-3-ламинин; связанный с микротрубочками белок 2; β-4-ламинин; кератин 6В; ладинин 1; кератин 6А; окклюдин; лорикрин; белок мембраны эритроцитов, зона 4.1 (связанный RH эллиптоцитоз 1); β-кристаллин А2; структурный белок глазного хрусталика; связанный с контактином подобный белку фактор 4; клодин 19; гипотетический белок LOC144501; кератин 6Е; кератин 6L; белок 2 внутренней мембраны хрусталика, 19 кДа); цитоскелет (ассоциированный с микротрубочками белок 2; фактор 5, подобный белку мембраны эритроцитов, зона 4.1; трихохиалин Homo sapiens (ТНН); кератин 6В; кератин 6А; подобный V антиген 1 эпителия; гомолог 1 Hook (Drosophila); лорикрин; белок мембраны эритроцитов, зона 4.1 (связанный RH эллиптоцитоз 1); тропомодулин 1; регулирующая сродство МАР/микротрубочки киназа 1; кератин 6Е; белок 2 семейства белков LIM, связывающихся с актином); молекулы клеточной адгезии (Cadherin 19, тип 2; миелоидно/лимфоидный или смешанного происхождения лейкоз; открытая рамка считывания хромосомы 21; фактор 2, относящийся к белкам рода IRRE; α-3-ламинин; сиалоадгезин; антиген CD84 (антиген лейкоцитов); растворимый связывающийся с галактозидами лектин 2 (галектин 2); V-подобный антиген 1 эпителия; антиген CD96; антиген промежуточного нефроза; родственная карциноэмбрионному антигену молекула 8 клеточной адгезии; IL-18; фактор 1 суперсемейства иммуноглобулинов; β-8-интегрин; орнитинарбамоил-трансфераза; β-6-интегрин; связанный с контактином белок-подобный фактор 4; α-1-коллаген, тип XVII; подобный Cadherin фактор 26; подобный муцину и cadherin фактор); белки клеточной дифференцировки (Z-полипептид 1 протеин-тирозинфосфатазы типа рецептора; α-3-ламинин; антиген CD84 (антиген лейкоцитов); EDRF2; связанный с эритроидной дифференцировкой фактор 2 Homo sapiens; фактор, подобный опухолевому белку р73; связанный с апоптозом/дифференцировкой NB4 белок; PNAS-133 Homo sapiens; подобный белку-7 при абсансе фактор 2; интерлейкин-24; кератин 6В; кератин 6А; представитель 9 семейства дегидрогеназ/редуктаз (SDR); β-5 белок соединения зазоров (коннексин 31.1); белок 4, относящийся к последовательности Iroquois homeobox; последовательность homeobox 2 передней брюшной стенки; лиганд 10 хемокина (лейтмотив С-Х-С); представитель 17 суперсемейства рецепторов фактора некроза опухоли; β-2-субъединица регулируемого потенциалом кальциевого канала; фактор 2 (parkin) юношеской, аутосомно-рецессивной болезни Паркинсона; калликреин 7 (химотрипсиновый фактор роговичного слоя); недостающий в глиальных клетках гомолог 2; α-АР-2; Z-полипептид 1 (протеин-тирозинфосфатаза рецепторного типа); тропонин Т1; Sciellin; глюкозаминил (N-ацетил) трансфераза 2, l-разветвляющий фермент; α-1-коллаген, тип XVII; супрессор 2 сигнальной функции цитокина; последовательность Distal-less homeo box 1; блокатор зиготы 1; интерлейкин-20; фактор дифференцировки роста 3; FGF-23; представитель 8А семейства сайтов интеграции MMTV типа Wingless); внеклеточные процессы (открытая рамка считывания 29 хромосомы 21; α-3-ламинин; β-4-ламинин; интерлейкин-24; специфический для беременности β-1-гликопротеин 1; лиганд 11 хемокина (лейтмотив С-Х-С); поверхностно-активное вещество - ассоциированный с легкими белок А1; препроноцицептин; рецептор 3В 5-окситриптамина (серотонина); родственная карциноэмбрионному антигену молекула 8 клеточной адгезии; лиганд 10 хемокина (лейтмотив С-Х-С); IL-18 (фактор индукции интерферона-γ); лактотрансферрин; альбумин; лиганд Fas (представитель 6 суперсемейства TNF); β-полипептид 4 семейства никотиновых холинергических рецепторов; антимикробный пептид Cathelicidin; трипсино-подобная протеаза Airway; S100 связывающий кальций белок А9 (кальгранулин В); TGF-α; калликреин 10; ингибитор 1 сериновых протеаз типа Куница; белок 3 индуцибельных сигнальных путей WNT1; релаксин 2; интерферон-κ; β-дефензин 103А; IL-20; гликопротеин 4 Zona pellucida; фактор 3 дифференцировки роста; FGF-23; представитель 8А семейства сайтов интеграции MMTV типа Wingless; фактор 5, родственный фактору Н комплемента); белки развития (ЕРН рецептор A3; родственная NIMA («never in mitosis gene а») киназа 2; белок с цинковыми пальцами 282; связывающаяся с TANK киназа 1; гомолог А фактора 11 мейотической рекомбинации MRE11; E2F фактор транскрипции 2; Z-полипептид 1 протеин-тирозинфосфатазы рецепторного типа; экспрессируемая в молочной железе мРНК из хромосомы Х клона 161455 Homo sapiens; α-3-ламинин; фактор 1, подобный гомологу онкогена v-myb вирусного птичьего миелобластоза; регулятор 11 сигнальной функции G-белка; связанный с микротрубочками белок 2; трансмембранный белок 16А; фактор 2 аденоматозного полипоза; последовательность Homeo box HB9; белок F центромеры, 350/400 кДа (митозин); антиген CD84 (антиген лейкоцитов); EDRF2; связанный с эритроидной дифференцировкой фактор 2 Homo sapiens; белок, подобный опухолевому белку р73; белок, родственный фактору апоптоза/дифференцировки NB4; PNAS-133 Homo sapiens; последовательность Forkhead box P2; связанный с желудочком экспрессируемый при дифференцировке белок YA61P (YA61) Homo sapiens; тенасцин N; открытая рамка считывания 49 хромосомы 6; белок с цинковыми пальцами 462; белок с цинковыми пальцами 71 (Cos26); последовательность box 7 области SRY (определяющая пол область хромосомы Y); фактор 4, подобный запускающему рецептору, экспрессируемому в миелоидных клетках; интерлейкин-24; специфический для беременности β-1-гликопротеин 1; хондроитинсульфат - протеогликан 5 (нейрогликан С); кератин 6В; кератин 6А; представитель 9 семейства дегидрогеназ/редуктаз (семейство SDR); антиген 1, подобный фактору V эпителия; белок β-5 заполнения просветов (коннексин 31.1); спаренный с G-белком рецептор 51; фактор 6 регуляции интерферона; нейтрофин 5 (нейтрофин 4/5); антиген CD96; белок 4 последовательности Iroquois homeobox; фактор 1, подобный рецептору интерлейкина-1; экспрессируемый в G-2 и S фазе фактор 1; представитель 3 группы Е подсемейства 2 ядерных рецепторов; последовательность Ventral anterior homeobox 2; белок с цинковыми пальцами 215; экспрессируемая последовательность сегмента ДНК 234 (уникального) на хромосоме 4; родственная карциноэмбрионному антигену молекула 8 клеточной адгезии; лиганд 10 хемокина (лейтмотив С-Х-С); IL-18; индоламин-пиррол 2,3-диоксигеназа; альбумин; чувствительный к кальцию рецептор (гипокальций-урическая гиперкальцемия 1, острый гиперпаратироидизм новорожденных); лиганд Fas (представитель 6 суперсемейства TNF); представитель 17 суперсемейства TNFR; β-2 субъединица зависящего от потенциала кальциевого канала; связанный с болезнью Паркинсона (юношеский аутосомно-рецессивный вариант) фактор 2 (parkin); калликреин 7 (химотриптический компонент слоя роговицы); утраченный в глиальных клетках гомолог 2; TGF-α; тиротропный фактор эмбрионов; АР-2а (активирующий связывающийся с энхансером белок 2α); калликреин 10; регулятор 7 сигнальной функции G-белка; Z-полипептид 1 протеин-тирозинфосфатазы типа рецептора; ингибитор 1 сериновых протеаз типа Куница; белок 3 индуцибельного пути WNT1 передачи сигналов; фактор 3 гетеротаксии 1 семейства Zic (гомолог нечетного спаривания, Drosophila); TTK протеин-киназа; скелетный медленный тропонин Т1; Sciellin; сцепленный с Х-хромосомой фактор, подобный индуцируемому TGFB фактору 2; калликреин 8 (нейропсин/овазин); глюкозаминил (N-ацетил) трансфераза 2, I-разветвляющий фермент; домен 30А анкиринового повтора; релаксин 2; α-1-коллаген, тип XVII; ген, дифференциально экспрессируемый в простате; регулятор 3 фосфатазы и актина; супрессор 2 сигнальной функции цитокина; представитель 3 группы А подсемейства 4 ядерных рецепторов; превращающий ангиотензин 1 фермент 1 (пептидил-дипептидаза А); гипотетический белок MGC17986; менее дистальная последовательность homeo box 1; гомолог 3 фактора обеспечения длины LAG1 (S. cerevisiae); фактор остановки зиготы 1; интерферон-κ; IL-20; ингибитор ICEBERG каспазы-1; фактор 3 дифференцировки роста; FGF-23; экспрессируемая в яичках последовательность 15; представитель 8А семейства сайтов интеграции MMTV типа Wingless; экспрессируемый в желудочке фактор 1, связанный с дефицитом карнитина; прокинетицин 1; фактор 3, подобный белку 3, связывающемуся с ответственным за CAMP элементом; представитель 15 семейства доменов мобилизации каспазы; белок FLJ23311).
Пример 6
Прямая дифференцировка эпителиальных стволовых клеток пуповины (UCEC) в кератиноциты эпидермиса кожи
Для дифференцировки в кератиноциты эпидермиса кожи эпителиальные стволовые клетки пуповины (клетки UCEC) культивировали в соответствии со стандартной методикой культивирования кератиноцитов. Техника извлечения клеток была такова, как описана выше. Затем UCEC культивировали в среде для роста кератиноцитов без сыворотки - KGM, KGM-2 (Cambrex), EpiLife (Cascade Biologies) или в среде Грина в присутствии облученных или обработанных митомицином С клеток мышиных эмбрионов 3T3 в качестве питающего слоя при 37°С, 5% CO2. Морфология клеток UCEC после дифференцировки таким способом напоминала морфологию кератиноцитов человеческого эпидермиса. Эпителиальные клетки имели подобную морфологию при световой микроскопии и могли быть легко превращены в фибробласты в обычных и имеющихся в продаже средах (см. фиг.2).
Иммунофлуоресцентный анализ показывает, что культивируемые клетки UCEC также экспрессируют молекулярные маркеры кератиноцитов эпидермиса, такие как кератины, десмосомы, гемидесмосомы и компоненты базальной мембраны (см. также фиг.10, которая показывает, что доказательством того, что клетки UCEC являются клетками эпителия, является в основном то, что они экспрессируют различные маркеры этих эпителиальных клеток). Поэтому эти результаты показывают, что эпителиальные стволовые/прародительские клетки из пуповины согласно настоящему изобретению могут быть дифференцированы в клетки кожи, такие как керотиноциты эпидермиса, которые могут быть использованы для залечивания ран и имеют высокие возможности для создания культивируемых эквивалентов кожи.
Пример 7
Проращивание эпителиальных и мезенхимных стволовых клеток пуповины с использованием повторных эксплантатов тканей выстилающей мембраны пуповины
Эпителиальные и мезенхимные стволовые клетки пуповины согласно настоящему изобретению проращивали, используя повторные эксплантаты ткани амниотической мембраны пуповины, следующим образом. Вкратце, в 1-й день процесса эксплантаты ткани помещали на чашки для культур ткани в ростовой среде (DMEM/10%FCS, EpiLife, KGM, KGM-2 или М171) при 37°С, 5% СО2; среду сменяли каждые 2-3 дня. Клетки начинали прорастать и продолжали мигрировать из эксплантатов в течение 7 дней. После этого эксплантаты ткани переносили на другие чашки, чтобы обеспечить дальнейшее прорастание клеток. Этот процесс продолжали до тех пор, пока эксплантат не уменьшался в размере до такой степени, что дальнейшая эксплантация становилась невозможной. В этой связи отмечено, что эксплантаты постепенно уменьшались в размере, пока они не становились слишком маленькими для дальнейшего проращивания тканевого эксплантата, поскольку в процессе прорастания клеток и их миграции из тканевых эксплантатов клетки продуцируют протеазы, переваривающие и разрушающие ткань. Фиг.16 схематически иллюстрирует быстрый и сильный процесс экспансии эпителиальных и мезенхимных стволовых клеток пуповины, достигаемый по этой прописи. Таким образом, это исследование демонстрирует, что из этого источника можно получить высокий выход клеток UCMC и UMEC, что дополнительно свидетельствует о высокой жизнеспособности и хороших характеристиках роста этих клеток в сравнении с полученными из других источников клетками, такими как стволовые клетки из костного мозга или жировой ткани. Кроме того, поскольку данная ткань плотная, успешная техника повторного проращивания эксплантата демонстрирует, что клетки согласно настоящему изобретению могут быть единообразно экстрагированы из всей ткани, а не только из некоторых порций. Это обеспечивает то, что максимальное количество клеток можно получить при малом числе пассажей вместо пропускания клеток через многие поколения, что приводит к ухудшению качества клеток.
Пример 8
Прямая дифференцировка мезенхимных клеток пуповины (UCMC) в фибробласты кожи
Для дифференцировки в фибробласты кожи мезенхимные стволовые клетки пуповины (UCMC) культивировали согласно стандартной прописи для культивирования фибробластов. Техника извлечения клеток была такая, как описана выше в примере 6. Затем клетки UCMC культивировали в среде DMEM или в имеющейся в продаже среде для роста фибробластов (FGM). Морфология дифференцированных таким способом клеток UCMC была похожа на морфологию фибробластов кожи человека. При световой микроскопии мезенхимные клетки обнаруживали такую же морфологию и могли легко быть превращены в фибробласты в обычных и имеющихся в продаже средах (см. фиг.3).
Реферат
Изобретение относится к области биотехнологии, клеточной инженерии и медицине. Способ извлечения стволовых/прародительских клеток из амниотической мембраны пуповины включает отделение амниотической мембраны от других компонентов пуповины in vitro. Далее проводят культивирование ткани амниотической мембраны в условиях, обеспечивающих пролиферацию клеток. В заключение осуществляют извлечение стволовых/прародительских клеток из культур ткани. Извлеченные стволовые клетки имеют свойства, подобные свойствам эмбрионных стволовых клеток, и могут быть использованы для различных терапевтических целей. 8 н. и 20 з.п. ф-лы, 68 ил.
Формула
(а) отделяют амниотическую мембрану от других компонентов пуповины in vitro;
(б) культивируют ткань амниотической мембраны, полученной в этапе (а), в условиях, дающих возможность клеточной пролиферации; и
(в) извлекают стволовые/прародительские клетки.
(а") отделяют клетки ткани амниотической мембраны перед культивированием способом, выбранным из группы, состоящей из ферментативного расщепления и прямого культивирования ткани эксплантата.
(г) культивируют стволовые/прародительские клетки в условиях, дающих возможность клеткам претерпевать клональную экспансию.
(д) культивируют стволовые/прародительские клетки в условиях, дающих возможность дифференцировки указанных клеток в эпителиальные клетки и/или мезенхимные клетки; и
(е) извлекают дифференцированные клетки.
(ж) консервируют извлеченные стволовые/прародительские клетки для дальнейшего использования.
получают эксплантат ткани из амниотической мембраны пуповины;
культивируют эксплантат ткани в подходящих среде для культивирования и условиях культивирования в течение подходящего периода времени.
Комментарии