Способы и композиции для лечения болезни хантингтона - RU2693891C1
Код документа: RU2693891C1
Чертежи




























Описание
ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается приоритет на основании предварительной заявки на патент США № 61/902,704, поданной 11 ноября 2013 г., раскрытие которой полностью включено в настоящее описание посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0002] Настоящее раскрытие относится к области экспрессии генов и редактирования генома.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
[0003] Болезнь Хантингтона (БХ), также известная как хорея Хантингтона, представляет собой прогрессирующее заболевание с моторными, когнитивными и психиатрическими нарушениями. Средний возраст наступления для данного заболевания составляет возраст 35-44 лет, хотя приблизительно в 10% случаев, наступление происходит до 21 года, и средняя продолжительность жизни после диагностики заболевания составляет 15-18 лет. Частота заболевания составляет приблизительно от 3 до 7 случаев на 100000 людей западноевропейского происхождения.
[0004] Болезнь Хантингтона является примером нарушений, связанных с экспансией тринуклеотидных повторов, которые были впервые охарактеризованы в начале 1990-х годов (см., Di Prospero и Fischbeck (2005) Nature Reviews Genetics 6:756-765). Эти нарушения включают локализованную экспансию нестабильных повторов наборов из трех нуклеотидов и могут приводить к потере функции гена, в котором находится удлиненный повтор, приобретению токсической функции или к тому и другому. Тринуклеотидные повторы могут располагаться в любой части гена, включая некодирующие и кодирующие области гена. Повторы, расположенные внутри кодирующих областей, обычно включают либо повторяющийся триплет, кодирующий глутамин (CAG), или триплет, кодирующий аланин (CGA). Удлиненные области повторов внутри некодирующих последовательностей могут приводить к аберрантной экспрессии гена, в то время как удлиненные повторы внутри кодирующих областей (также известные нарушения реитерации кодона) могут вызвать неправильную укладку и агрегацию белка. Точная причина патофизиологии, ассоциированной с аберрантными белками, часто является неизвестной. Обычно, в генах дикого типа, которые являются объектом экспансии тринуклеотидных повторов, эти области содержат переменное число повторных последовательностей в нормальной популяции, но у пораженных болезнью популяций, число повторов может возрастать от удвоения до увеличения в логарифмическом порядке числа повторов. При БХ, повторы вставлены в N-концевую кодирующую область крупного цитозольного белка Хантингтина (Htt). Нормальные аллели Htt содержат 15-20 повторов CAG, в то время как аллели, содержащие 35 или более повторов, могут считаться аллелями, потенциально вызывающими БХ, и придают риск развития заболевания. Аллели, содержащие 36-39 повторов, считаются не полностью проявляющимися, и у тех индивидуумов, имеющих эти аллели, заболевание может развиваться или не развиваться (или могут развиваться симптомы позднее в жизни), в то время как аллели, содержащие 40 повторов или более, считаются полностью проявляющимися. Действительно, о таких асимптотических лицах, содержащих аллели БХ с этим числом мноогих повторов, не сообщалось. У индивидуумов с ювенильным наступлением БХ (в возрасте <21 года)) часто обнаруживают 60 или более повторов CAG. В дополнение к увеличению повторов CAG, также было показано, что БХ может включать +1 и +2 сдвиги рамки в последовательности повтора, таким образом, что область будет кодировать путь полисеринового полипептида (кодируемый повторами AGC в случае +1 сдвига рамки) в большей степени, чем полиглутаминовый путь (Davies and Rubinsztein (2006) Journal of Medical Genetics 43: 893-896).
[0005] При БХ, мутантный аллель Htt обычно наследуется от одного родителя как доминантный признак. Любой ребенок, рожденный от пациента с БХ, имеет 50% вероятность развития заболевания, если другой родитель не был поражен нарушением. В некоторых случаях, родитель может иметь промежуточный аллель БХ и быть бессимптомным, в то время как, вследствие экспансии повтора, у ребенка проявляется заболевание. В дополнение, аллель БХ может также проявлять явление, известное как антиципация, где возрастающую тяжесть заболевания или снижение возраста наступления наблюдают на протяжении нескольких поколений вследствие нестабильной природы области повтора в процессе сперматогенеза.
[0006] Кроме того, экспансия тринуклеотидных повторов в Htt приводит к потере нейронов в средних шипиковых нейронах с проекцией гамма-аминомасляной кислоты (ГАМК) в полосатом теле, с потерей нейронов, также происходящей в неокортексе. Средние шипиковые нейроны (MSN), которые содержат энкефалин, и, которые проецируются на внешний бледный шар (в так называемом “непрямом пути”) вовлечены в большей степени, чем нейроны, которые содержат вещество P и проецируются на внутренний бледный шар (в “прямом” пути), однако, оба типа MSN поражаются. MSN при БХ проявляют транскрипционную дисрегуляцию наряду с другими аномальными изменениями (аномальными агрегациями и включениями htt, биоэнергетическими дефектами, дефицитом нейротрофина, нарушениями аксонального транспорта и эксайтотоксичностью). Механизм транскриптомных эффектов может относиться к изменениям активностей растворимых ДНК-связывающих факторов транскрипции, аномалиями биохимии и организации хроматина и секвестрации агрегат-управляемого ядерного фактора транскрипции (см., Runne et al (2008) J Neurosci 28(39):9723-9731).
[0007] Другие области мозга, сильно поражаемые у людей с болезнью Хантингтона, включают черное вещество, кортикальные слои 3, 5, и 6, область CA1 гиппокампа, угловую извилину в теменной доле, клетки Пуркинье мозжечка, латеральные серобугорные ядра гипоталамуса, и центромедиалпарафасцикулярный комплекс таламуса (Walker (2007) Lancet 369:218-228).
[0008] Роль нормального белка Htt является плохо изученной, но он может быть вовлечен в нейрогенез, апоптотическую смерть клеток и миграцию везикул. В дополнение, имеются данные, указывающие на то, что Htt дикого типа стимулирует выработку нейротрофического фактора головного мозга (BDNF), фактор предварительного выживания для стриарных нейронов. Было показано, что прогрессирование БХ коррелирует со снижением экспрессии BDNF у мышиных моделей БХ (Zuccato et al (2005) Pharmacological Research 52(2): 133-139), и, что доставка либо BDNF, или нейротрофического фактора глиальной клеточной линии (GDNF) посредством аденоассоцированной вирусной (AAV) вектор-опосредованной доставки гена может защищать стриарные нейроны у мышиных моделей БХ (Kells et al, (2004) Molecular Therapy 9(5): 682-688).
[0009] Возможности терапии БХ в настоящее время очень ограничены. Несколько потенциальных методологий, разработанных для предотвращения токсичностей, ассоциированных с агрегацией белков, которая происходит посредством удлиненного полиглутаминового тракта, таких как сверхэкспрессия шаперонинов или индукция ответа на тепловой шок с использованием соединения гелданамицина, показали снижение этих токсичностей на моделях in vitro. Другие терапии выявляют роль апоптоза при клинических проявлениях заболевания. Например, было показано замедление симптомов заболевания посредством блокирования активности каспазы на животных моделях у потомства от спаривания мышей, где один родитель содержал аллель БХ, а другой родитель имел доминантный отрицательный аллель для каспазы 1. Дополнительно, расщепление мутантного Htt БХ каспазой может иметь значение для патогенности заболевания. Было обнаружено, что трансгенные мыши, несущие резистентный к каспазе-6 мутантный Htt, поддерживают нормальную нейрональную функцию, и у них не развивается стриарная нейродегенерация в сравнении с мышами, несущими не резистентный к каспазе мутантный аллель Htt (см. Graham et al (2006) Cell 125: 1179-1191). Было также показано, что молекулы, которые целенаправленно действуют на элементы апоптотического пути, имеют замедляющее воздействие на симптомологию. Например, было показано, что соединения zVAD-fmk и миноциклин, оба из которых ингибируют активность каспазы, замедляют проявление заболевания у мышей. Лекарственное средство ремацемид также применяли при небольших испытаниях по лечению БХ у людей, поскольку полагали, что соединение предотвращает связывание мутантного Htt с рецептором NDMA, чтобы предотвратить проявления токсических воздействий на нервные клетки. Однако никаких статистически значимых улучшений функции нейронов не наблюдали при этих испытаниях. Дополнительно, Группа по исследованиям болезни Хантингтона провела рандомизированное, двойное слепое исследование с использованием Кофермента Q. Несмотря на то, что наблюдали тенденцию в сторону более медленного прогрессирования заболевания среди пациентов, которых лечили с использованием кофермента Q10, не было никакого значительного изменения степени ухудшения общей функциональной способности. (Di Prospero и Fischbeck, там же). Публикации патентных заявок США 2011/0082093 и 20130253040 раскрывают нуклеазы, нацеленные на Htt.
[0010] Были описаны различные способы и композиции для направленного расщепления геномной ДНК. Такие направленные объекты расщепления могут применяться, например, чтобы индуцировать направленный мутагенез, индуцировать направленные делеции последовательностей клеточных ДНК, и способствовать направленной рекомбинации в заданном хромосомном локусе. См, например, Патенты США №№ 8,623,618; 8,034,598; 8,586,526; 6,534,261; 6,599,692; 6,503,717; 6,689,558; 7,067,317; 7,262,054; 7,888,121; 7,972,854; 7,914,796; 7,951,925; 8,110,379; 8,409,861; Публикации патентных заявок США 20030232410; 20050208489; 20050026157; 20060063231; 20080159996; 201000218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983 и 20130177960 и Патентную заявку США № 14/278,903, раскрытия которых полностью включены посредством ссылки для всех целей. Эти способы часто включают применение сконструированных систем расщепления для индуцирования двухцепочечного разрыва (DSB) или одноцепочечного разрыва в целевой последовательности ДНК, таким образом, чтобы репарация разрыва посредством процесса устранения ошибки, такая как негомологичное соединение концов (NHEJ), или репарация с использованием матрицы для репарации (гомологичная направленная репарация или HDR) могли бы приводить к выключению гена или вставке последовательности, представляющей интерес (целенаправленной интеграции). Данный метод также может применяться для введения сайт-специфичных изменений в геномную последовательность посредством использования донорного олигонуклеотида, включающего введение специфичных делеций геномных областей или специфичных точечных мутаций, или локализованных изменений (также известное как коррекция гена). Расщепление может происходить посредством применения специфичных нуклеаз, таких как сконструированные нуклеазы типа “цинковые пальцы” (ZFN), эффекторные нуклеазы подобные активатору транскрипции (TALEN), или с использованием системы CRISPR/Cas со сконструированной крРНК/тракр РНК (“единственным РНК-проводником”) для направления специфичного расщепления. Дополнительно, разрабатываются нуклеазы направленного действия на основе системы Аргонавт (например, из T. thermophilus, известные как “TtAgo”, см., Swarts et al (2014) Nature 507(7491): 258-261), которые также могут иметь потенциал для применений при редактировании генома и генной терапии.
[0011] Также были разработаны сконструированные гибридные белки для модуляции экспрессии адресного гена. Такие белки могут применяться, например, для усиления или подавления экспрессии желательного гена (см., например, Патенты США №№ 6,534,261; 6,607,882; 6,824,978; 6,933,113; 7, 013,219; 7,220,719; 8,268,618; 7,985,778; 8,586,526; Заявку на патент США 20120294838, раскрытия которых полностью включены посредством ссылки для всех целей).
[0012] Таким образом, остается потребность в композициях и способах, которые могут опираться на эти перспективные технологии, для лечения и предотвращения болезни Хантингтона.
КРАТКОЕ СОДЕРЖАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0013] В данном описании раскрыты способы и композиции для лечения болезни Хантингтона. Конкретно, в данном документе предоставлены способы и композиции для модификации (например, модуляции экспрессии) аллеля Htt БХ, таким образом, чтобы лечить болезнь Хантингтона. Также предоставлены способы и композиции для генерации животных моделей болезни Хантингтона.
[0014] Таким образом, в одном аспекте, предоставлены сконструированные (неприродного происхождения) ДНК-связывающие домены (например, белки “цинковые пальцы”, TAL эффекторные (TALE) белки или CRISPR/dCas-TF), которые модулируют экспрессию аллеля БХ (например, Htt). Сконструированные белки “цинковые пальцы” или белки TALE представляют собой белки “цинковые пальцы” или белки TALE неприродного происхождения, чьи ДНК-связывающие домены (например, спирали распознавания или RVD) были изменены (например, посредством селекции и/или целесообразного дизайна) для связывания с предварительно выбранным целевым сайтом. Любой из белков “цинковые пальцы”, описанных в настоящем документе, может включать 1, 2, 3, 4, 5, 6 или более цинковых пальцев, причем каждый цинковый палец имеет спираль распознавания, которая связывается с целевым субсайтом в выбранных последовательности (последовательностях) (например, гене(генах)). Аналогично, любой из белков TALE, описанных в данном документе, может включать любое число RVD TALE. В нескольких вариантах осуществления, по меньшей мере, один RVD имеет неспецифичное ДНК-связывание. В нескольких вариантах осуществления, по меньшей мере, одна спираль распознавания (или RVD) имеет неприродное происхождение. В некоторых вариантах осуществления, белки “цинковые пальцы” имеют спирали распознавания, показанные в Таблицах 1A и 1B. В других вариантах осуществления, белки “цинковые пальцы” связываются с целевыми последовательностями, показанными в Таблицах 2A и 2B. В нескольких вариантах осуществления, белки “цинковые пальцы” содержат спирали распознавания, показанные в Таблице 2C. В некоторых вариантах осуществления, белки “цинковые пальцы” составляют в фармацевтическую композицию, например, для введения субъекту.
[0015] Также предоставлены сконструированные (неприродного происхождения) системы CRISPR/Cas, которые модулируют экспрессию аллеля БХ (например, Htt). Может быть сконструирован конкретно домен нуклеазы Cas9 для потери активности расщепления ДНК (“dCAS”), и слит с функциональным доменом, способным к модуляции экспрессии гена (см., Perez-Pimera (2013) Nat Method 10(10):973-976) для создания dCas-TF. Когда dCas-TF поставляется вместе с Htt-специфичным РНК-проводником, система модулирует экспрессию гена Htt.
[0016] В одном аспекте, предоставлены репрессоры (ZFP-TF, CRISPR/dCas-TF или TALE-TF), которые связываются с последовательностями полностью или частично вне области повтора CAG в Htt. В одном другом аспекте, предоставлены репрессоры ZFP, Cas или TALE (ZFP-TF, CRISPR/dCas-TF или TALE-TF) которые связываются с последовательностями внутри области повтора CAG в Htt. В нескольких вариантах осуществления, эти ZFP-TF, CRISPR/dCas или TALE-TF предпочтительно связываются с удлиненными тринуклеотидными трактами относительно трактов с повторами, имеющим длину дикого типа, посредством чего достигается преимущественная репрессия удлиненного аллеля. В нескольких вариантах осуществления эти ZFP-TF, CRISPR/dCas-TF или TALE-TF включают домены белкового взаимодействия (или “домены димеризации”), которые обеспечивают мультимеризацию, при связывании с ДНК. В нескольких вариантах осуществления, эти ZFP-TF, CRISPR/dCas-TF или TALE TF достигают кооперативного связывания ДНК с последовательностью повтора, таким образом, что удлиненный аллель более эффективно связывается посредством большего числа ZFP, dCas или белков TALE, чем аллель дикого типа, что обеспечивает преимущественную репрессию мутантного аллеля. Эти кооперативно связывающие ZFP-TF, CRISPR/dCas-TF или TALE TF могут дополнительно содержать или могут не содержать домены белкового взаимодействия, которые обеспечивают мультимеризацию, когда связываются с ДНК. В нескольких вариантах осуществления, ZFP TF, CRISPR/dCas-TF или TALE TF образуют устойчивый комплекс из мультимеров данного размера, и, таким образом имеют способность к преимущественному взаимодействию с CAG трактом свыше определенного минимального размера, где такой минимальный размер больше, чем длина CAG тракта дикого типа.
[0017] В некоторых вариантах осуществления, ZFP, CRISPR/dCas-TF или белки TALE, описанные в данном документе, (например, двурукие, мультимеризующие и т.д.) преимущественно модифицируют экспрессию мутантного аллеля Htt. В нескольких вариантах осуществления, ZFP, CRISPR/dCas-TF или TALE специфически связывается с мутантными аллелями Htt, где удлиненный тракт кодирует поли-глутамин, в то время как в других вариантах осуществления, ZFP, CRISPR/dCas-TF или TALE специфически связывается с мутантным аллелем Htt, где удлиненный тракт кодирует поли-серин. Таким образом, в нескольких вариантах осуществления, ZFP-TF, CRISPR/dCas-TF или TALE-TF модулирует как формы дикого типа, так и мутантные формы аллеля HTT. В некоторых вариантах осуществления, ZFP, CRISPR/dCas-TF или TALE модулируют аллель Htt только дикого типа. В других вариантах осуществления, ZFP, CRISPR/dCas-TF или TALE модулируют только мутантную форму Htt.
[0018] В других вариантах осуществления, предоставлены репрессирующие ZFP-TF, CRISPR/dCas-TF или TALE-TF, которые предпочтительно связываются с известными SNP, ассоциированными с удлиненными аллелями Htt БХ. Следовательно, ZFP-TF, CRISPR/dCas-TF или TALE-TF являются специфичными для мутантных аллелей Htt, которые содержат SNP, обеспечивая специфичную репрессию мутантного аллеля Htt. В одном другом аспекте, предоставлены ZFP-TF, CRISPR/dCas-TF или TALE-TF, которые специфично активируют аллель Htt дикого типа посредством взаимодействия с SNP, ассоциированными с аллелями дикого типа. Таким образом, активируется аллель Htt только дикого типа.
[0019] В некоторых вариантах осуществления, белки “цинковые пальцы” (ZFP), dCas или белки TALE, описанные в данном документе, могут помещаться в функциональную связку с регуляторным доменом (или функциональным доменом) в качестве части гибридного белка. Функциональный домен может представлять собой, например, домен транскрипционной активации, домен транскрипционного репрессора и/или домен нуклеазы (расщепления). Посредством отбора либо домена активации или домена репрессии для слияния с ZFP, dCas или TALE, такие гибридные белки могут применяться, чтобы либо активировать или подавить экспрессию гена. В нескольких вариантах осуществления, предоставлен гибридный белок, содержащий ZFP, dCas или TALE, целенаправленный на мутантный Htt, описанный в данном документе, слитый с доменом транскрипционного репрессора, который может применяться для понижающей регуляции экспрессии мутантного Htt. В нескольких вариантах осуществления, предоставлен гибридный белок, содержащий ZFP, dCas или TALE, целенаправленный на аллель Htt дикого типа, слитый с доменом активации транскрипции, который может повышающе регулировать аллель Htt дикого типа. В некоторых вариантах осуществления, активность регуляторного домена регулируется экзогенной малой молекулой или лигандом таким образом, что взаимодействие с комплексом транскрипции клетки не будет происходить в отсутствии экзогенного лиганда. Такие внешние лиганды управляют степенью взаимодействия ZFP-TF, CRISPR/dCas-TF или TALE-TF с комплексом транскрипции. Регуляторные домен(ы) могут быть функционально связаны с любой частью (частями) одного или нескольких из ZFP, dCas или TALE, включая один или несколько из ZFP, dCas или TALE, внешних по отношению к одному или нескольким ZFP, dCas или TALE и любой их комбинации. Любой их гибридных белков, описанных в данном документе, может быть составлен в фармацевтическую композицию.
[0020] В нескольких вариантах осуществления, сконструированные ДНК-связывающие домены, описанные в данном документе, могут помещаться в функциональную связку с доменами нуклеазы (расщепления) в качестве части гибридного белка. В нескольких вариантах осуществления, нуклеаза содержит нуклеазу Ttago. В других вариантах осуществления, нуклеазные системы, такие как система CRISPR/Cas, могут использоваться вместе со специфичным единственным РНК-проводником для нацеливания нуклеазы на целевое расположение в ДНК. В некоторых вариантах осуществления, такие нуклеазы и нуклеазные гибриды могут использоваться для целенаправленного действия на мутантные аллели Htt в стволовых клетках, таких как индуцированные плюрипотентные стволовые клетки (iPSC), человеческие эмбриональные стволовые клетки (hESC), мезенхимальные стволовые клетки (MSC) или нейрональные стволовые клетки, где активность нуклеазного гибрида будет приводить к аллелю Htt, содержащего количество повторов CAG дикого типа. В некоторых вариантах осуществления, предоставлены фармацевтические композиции, содержащие модифицированные стволовые клетки.
[0021] В еще одном другом аспекте, предоставлен полинуклеотид, кодирующий любой из ДНК-связывающих белков, описанных в данном документе. В одном другом аспекте, предоставлены полинуклеотиды, кодирующие нуклеазу CRIPSR/Cas и единственный РНК-проводник. Такие полинуклеотиды могут вводиться субъекту, у которого желательным является лечение болезни Хантингтона.
[0022] В еще дополнительных аспектах, изобретение предоставляет способы и композиции для генерации специфичных модельных систем для исследования болезни Хантингтона. В некоторых вариантах осуществления, в данном документе предоставлены модели, в которых мутантные аллели Htt генерируют с использованием эмбриональных стволовых клеток для генерации линий клеток и животных, в которых тракты экспансии тринуклеотидных повторов с конкретной длиной (например, 50, 80, 109 и 180 повторов CAG) вводят в аллель Htt дикого типа с использованием управляемой нуклеазой “цинковые пальцы” (ZFN), TALE-нуклеазой (TALEN), Ttago или CRISPR/Cas нуклеазой целенаправленную интеграцию. В некоторых вариантах осуществления, модельные системы содержат клеточные линии in vitro, в то время как в других вариантах осуществления, модельные системы содержат трансгенных животных. В любой из животных моделей, описанных в данном документе, животное может представлять собой, например, грызуна (например, крысу, мышь), примата (например, примата, не являющегося человеком), или кролика. Таким образом, изобретение также включает клетку или линию клеток, в которой эндогенный ген, относящийся к БХ, модифицируют, например, в сравнении с последовательностью гена дикого типа в клетке. Клетки или линии клеток могут являться гетерозиготными или гомозиготными для модификации. Модификации могут включать вставки, делеции и/или их комбинации. В некоторых вариантах осуществления, ген (например, Htt) модифицируют посредством нуклеазы (например, ZFN, TALEN, системы CRISPR/Cas, системы Ttago и т.д.). В некоторых вариантах осуществления, модификация находится в сайте (сайтах) связывания и/или расщепления нуклеазы (нуклеаз) или рядом с ними, например, в интервале 1-300 (или любого значения между ними) пар оснований против хода транскрипции или по ходу транскрипции от сайта (сайтов) расщепления, более предпочтительно, в интервале 1-100 пар оснований (или любого значения между ними) с любой стороны от сайта (сайтов) связывания и/или расщепления, даже более предпочтительно, в интервале от 1 до 50 с любой стороны от сайта (сайтов) связывания и/или расщепления.
[0023] В других аспектах, изобретение включает доставку донорной нуклеиновой кислоты к целевой клетке. Донор может доставляться до, после, или наряду с нуклеиновой кислотой, кодирующей нуклеазу (нуклеазы). Донорная нуклеиновая кислота может содержать экзогенную последовательность (трансген) для интеграции в геном клетки, например, эндогенный локус. В нескольких вариантах осуществления, донор может содержать непроцессированный ген или его фрагмент, фланкированные областями, гомологичными с намеченным сайтом расщепления. В нескольких вариантах осуществления, у донора отсутствуют гомологичные области, и он интегрируется в целевой локус посредством независимого от гомологии механизма (т.е. NHEJ). Донор может содержать любую последовательность нуклеиновой кислоты, например, нуклеиновую кислоту, которая, при использовании в качестве субстрата для гомологичной направленной репарации индуцированного нуклеазой двухцепочечного разрыва, приводит к устанавливаемой донором делеции, генерируемой в эндогенном хромосомном локусе, или, альтернативно (или в дополнение к), созданию новых аллельных форм (например, точечным мутациям, которые устраняют сайт связывания фактора транскрипции) эндогенного локуса. В нескольких аспектах, донорная нуклеиновая кислота представляет собой олигонуклеотид, где интеграция приводит к событию коррекции гена, или целенаправленной делеции.
[0024] В нескольких вариантах осуществления, полинуклеотид, кодирующий ДНК-связывающий белок, представляет собой мРНК. В нескольких аспектах, мРНК может быть химически модифицирована (См., например, Kormann et al, (2011) Nature Biotechnology 29(2):154-157). В других аспектах, мРНК может содержать кэп ARCA (см., Патенты США 7,074,596 и 8,153,773). В дополнительных вариантах осуществления, мРНК может содержать смесь немодифицированных и модифицированных нуклеотидов (см. Патентную публикацию США 2012-0195936).
[0025] В еще одном другом аспекте, предоставлен вектор доставки гена, содержащий любой из полинуклеотидов, описанных в данном документе. В некоторых вариантах осуществления, вектор представляет собой аденовирусный вектор (например, вектор Ad5/F35), лентивирусный вектор (LV), включающий интеграционно компетентные или интеграционно дефективные лентивирусные векторы, или ассоциированный с аденовирусом вирусный вектор (AAV). В некоторых вариантах осуществления, вектор AAV представляет собой вектор AAV6. Таким образом, также в данном документе предоставлены аденовирусные (Ad) векторы, ассоциированные с LV или аденовирусом вирусные векторы (AAV), содержащие последовательность, кодирующую, по меньшей мере, одну нуклеазу (ZFN или TALEN) и/или донорную последовательность для целенаправленной интеграции в ген-мишень. В некоторых вариантах осуществления, Ad вектор представляет собой химерный Ad вектор, например, вектор Ad5/F35. В некоторых вариантах осуществления, лентивирусный вектор представляет собой интеграза-дефективный лентивирусный вектор (IDLV) или интеграционно компетентный лентивирусный вектор. В некоторых вариантах осуществления вектор является псевдотипированным с оболочкой VSV-G или с другими оболочками.
[0026] В нескольких вариантах осуществления, предоставлены модельные системы для болезни Хантингтона, где целевые аллели (например, мутантный Htt) являются меченными маркерами экспрессии. В некоторых вариантах осуществления, меченными являются мутантные аллели (например, мутантный Htt). В нескольких вариантах осуществления, меченным является аллель дикого типа (например, Htt дикого типа), и в дополнительных вариантах осуществления, аллели, как дикого типа, так и мутантные аллели являются меченными раздельными маркерами экспрессии. В некоторых вариантах осуществления, модельные системы содержат клеточные линии in vitro, в то время как, в других вариантах осуществления, модельные системы содержат трансгенных животных.
[0027] Дополнительно, также предоставлены фармацевтические композиции, содержащие нуклеиновые кислоты и/или белки (например, ZFP, Cas или TALE, или гибридные белки содержащие ZFP, Cas или TALE). Например, некоторые композиции включают нуклеиновую кислоту, содержащую последовательность, которая кодирует одну из ZFP, Cas или TALE, описанных в данном документе, функционально связанную с регуляторной последовательностью, комбинированную с фармацевтически приемлемым носителем или разбавителем, где регуляторная последовательность обеспечивает экспрессию нуклеиновой кислоты в клетке. В некоторых вариантах осуществления, кодируемые ZFP, CRISPR/Cas или TALE являются специфичными для аллели Htt БХ. В нескольких вариантах осуществления, фармацевтические композиции содержат ZFP, CRISPR/Cas или TALE, которые модулируют аллель Htt БХ, и ZFP, CRISPR/Cas или TALE, которые модулируют нейротрофный фактор. Композиции на основе белков включают одну из нескольких ZFP, CRISPR/Cas или TALE, раскрытых в данном описании, и фармацевтически приемлемый носитель или разбавитель.
[0028] В еще одном другом аспекте также предоставлена изолированная клетка, содержащая любые из белков, полинуклеотидов и/или композиций, описанных в данном документе.
[0029] В одном другом аспекте, в данном документе предоставлены способы лечения и/или предотвращения болезни Хантингтона с использованием способов и композиций, описанных в данном документе. В нескольких вариантах осуществления, способы включают композиции, где полинуклеотиды и/или белки могут доставляться с использованием вирусного вектора, невирусного вектора (например, плазмиды) и/или их комбинации. В нескольких вариантах осуществления, способы включают композиции, содержащие популяции стволовых клеток, содержащих ZFP или TALE, или измененных с использованием ZFN, TALEN, Ttago или нуклеазной системы CRISPR/Cas изобретения.
[0030] В нескольких вариантах осуществления, способы и композиции изобретения применяют для нормализации и/или модификации экспрессии некоторых генов в срединных шипиковых нейронах. Например, в нескольких вариантах осуществления, способ включает применение генетического репрессора и/или композиция представляет собой генетический репрессор, который изменяет экспрессию одного или нескольких биомаркеров, относящихся к БХ, включающих, но не ограниченных перечисленными, DARPP-32A, PDE10a, Drd1 и/или Drd2, для лечения или профилактики субъекта с болезнью Хантингтона. В нескольких вариантах осуществления, генетический репрессор является малой молекулой, нуклеиновой кислотой или белком, который ингибирует экспрессию белка хантингтина. В нескольких вариантах осуществления, генетический репрессор специфически связывается с геном хантингтина, в его геномной ДНК или в форме транскрипта, такой как мРНК. В нескольких вариантах осуществления, генетический репрессор является экзогенным и/или включает химическую модификацию и/или модификацию последовательности, которая не встречается в природе. В некоторых вариантах осуществления, генетический репрессор представляет собой ДНК-связывающий домен ZFP, TALE или Cas, который специфически связывается с кодирующей и/или некодирующей частью гена хантингтина, относительно связывания с другими генами в геноме. В некоторых других вариантах осуществления, генетический репрессор представляет собой антисмысловую нуклеиновую кислоту, которая включает последовательность, которая на 100%, или, по меньшей мере, на 90%, или, по меньшей мере, на 80%, или, по меньшей мере, на 70%, или, по меньшей мере, на 60% является идентичной с последовательностью, комплементарной с коодирующей и/или некодирующей частью гена хантингтина в его форме транскрипта, такой как мРНК. Часть гена хантингтина, с которой специфически связывается генетический репрессор, может иметь длину, равную, по меньшей мере, 8, по меньшей мере, 10, по меньшей мере, 12, по меньшей мере, 15, по меньшей мере, 18, по меньшей мере, 20, по меньшей мере, 22, по меньшей мере, 25, по меньшей мере, 30, по меньшей мере, 35, и/или, по меньшей мере, 40 нуклеотидам.
[0031] В нескольких вариантах осуществления, описанных в данном документе, применение генетического репрессора, такого как для лечения субъекта или для тестирования генетического репрессора, может приводить к увеличенной экспрессии одного или нескольких из DARPP-32A, PDE10a, Drd1 и/или Drd2 относительно контроля, не подвергаемого лечению. Необработанный контроль не получает генетический репрессор, например, не подвергаемый лечению субъект или необработанная контрольная клетка, такая как MSN. В нескольких аспектах, экспрессия DARPP-22 увеличивается в MSN (например, увеличивается на 10% или более, увеличивается на 20% или более, увеличивается на 30% или более, увеличивается на 40% или более, увеличивается на 50% или более, увеличивается на 60% или более, увеличивается на 70% или более, увеличивается на 80% или более, увеличивается на 90% или более, увеличивается на 100% или более, увеличивается на 125% или более, увеличивается на 150% или более, увеличивается на 175% или более, и/или увеличивается на 200% или более, или любое значение между ними) относительно необработанного контроля (например, не подвергаемого лечению субъекта или необработанной клетки). В других аспектах, уровень PDE10a увеличивается в MSN (например, увеличивается на 10% или более, увеличивается на 20% или более, увеличивается на 30% или более, увеличивается на 40% или более, увеличивается на 50% или более, увеличивается на 60% или более, увеличивается на 70% или более, увеличивается на 80% или более, увеличивается на 90% или более, увеличивается на 100% или более, увеличивается на 125% или более, увеличивается на 150% или более, увеличивается на 175% или более, и/или увеличивается на 200% или более, или любое значение между ними) относительно необработанного контроля (например, например, не подвергаемого лечению субъекта или необработанной клетки MSN). В еще дополнительных аспектах, уровень дофаминового рецептора Drd1 и/или Drd2 увеличивается в MSN (например, увеличивается на 10% или более, увеличивается на 20% или более, увеличивается на 30% или более, увеличивается на 40% или более, увеличивается на 50% или более, увеличивается на 60% или более, увеличивается на 70% или более, увеличивается на 80% или более, увеличивается на 90% или более, увеличивается на 100% или более, увеличивается на 125% или более, увеличивается на 150% или более, увеличивается на 175% или более, и/или увеличивается на 200% или более, или любое значение между ними) относительно необработанного контроля (например, например, не подвергаемого лечению субъекта или необработанной клетки MSN). В нескольких аспектах, уровень любой комбинации из DARPP-32A, PDE10a, Drd1 и Drd2 увеличивается, например, уровень комбинации из двух или более увеличивается, когда два могут представлять собой DARP32A и PDE10a, или DARP32A и Drd1 или любую такую комбинацию. В других аспектах, уровень любых трех из DARPP-32A, PDE10a, Drd1 и Drd2 увеличивается, в то время как в других аспектах, уровень всех из DARPP-32A, PDE10a, Drd1 и Drd2 увеличивается относительно необработанного контроля (например, например, не подвергаемого лечению субъекта или необработанной клетки MSN).
[0032] В нескольких вариантах осуществления, способы и композиции изобретения применяют для нормализации хватательного поведения у субъекта (например, животной модели) с БХ. Например, в нескольких аспектах, применение генетического репрессора, такое как для лечения субъекта или для тестирования генетического репрессора, может приводить к сниженному хватательному рефлексу у подвергнутого лечению субъекта или животной модели относительно необработанного контроля. В нескольких аспектах, модель представляет собой мышиную модель БХ R6/2. В нескольких аспектах, хватательный рефлекс снижается на 10% или более, снижается на 20% или более, снижается на 30% или более, снижается на 40% или более, снижается на 50% или более, снижается на 60% или более, снижается на 70% или более, снижается на 80% или более, снижается на 90% или более, или снижается на 100% или более, или любое значение между ними. В нескольких аспектах, снижение хватательного рефлекса может наблюдаться в интервале 30 секунд у тестируемого животного, которое находится в интервале 7 недель, в интервале 8 недель, в интервале 9 недель, в интервале 10 недель, в интервале 11 недель, или в интервале 12 недель от рождения животного и/или в интервале 2 недель, в интервале 3 недель, в интервале 4 недель, в интервале 5 недель, в интервале 6 недель или в интервале 7 недель от обработки генетическим репрессором.
[0033] Эти и другие аспекты будут очевидны для квалифицированного специалиста в области в свете раскрытия изобретения в целом.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
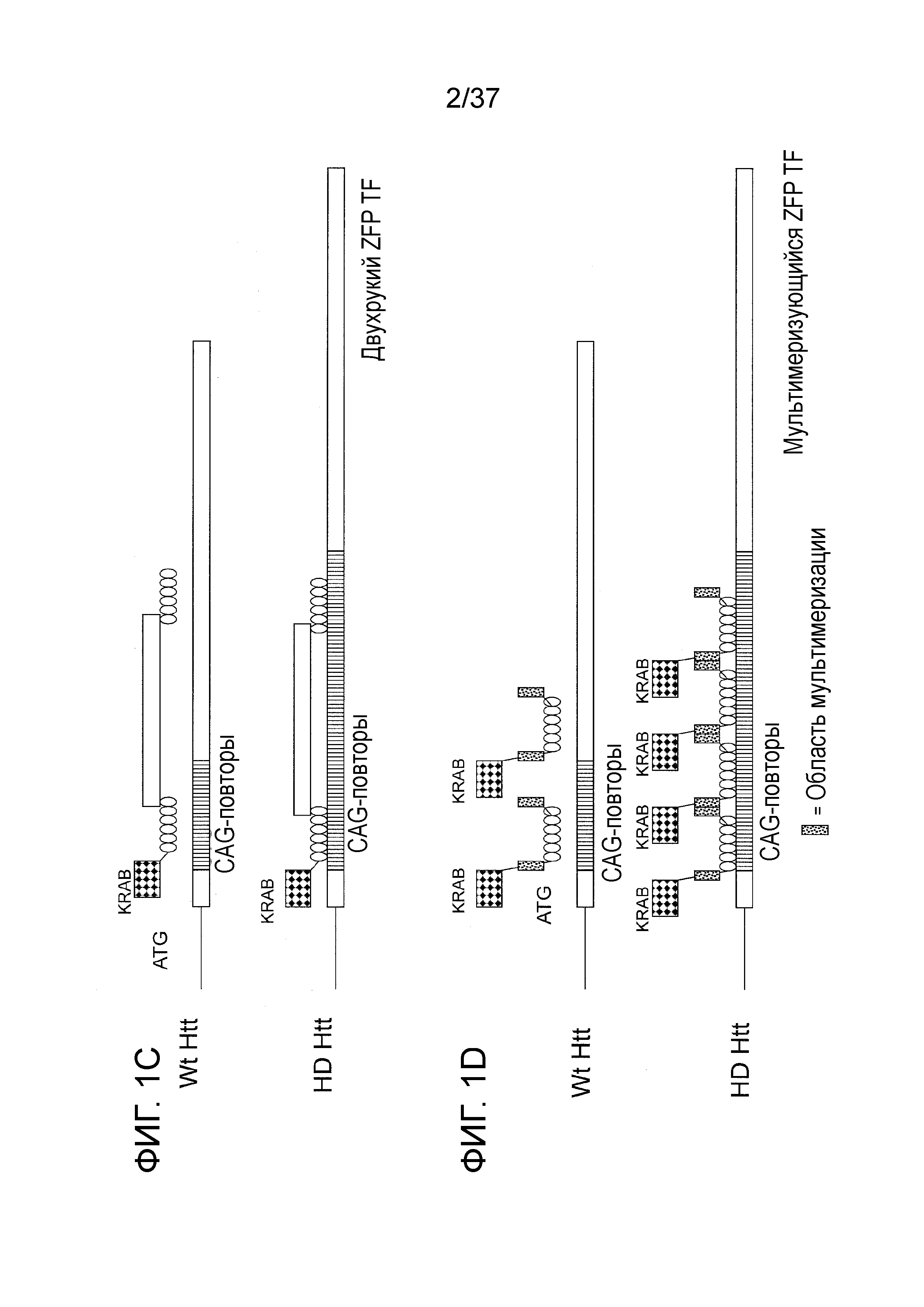
[0034] Фиг. 1A-1E представляют собой схематические изображения, отображающие аллель дикого типа и мутантный (болезнь Хантингтона, БХ) аллель хантингтина (Htt) и различные ZFP-TF, связывающиеся с этими аллелями. Фиг. 1A показывает конструкции ZFP, которые связываются вне CAG-повтора, и, следовательно, предполагается их связывание в равной степени с аллелем дикого типа и мутантным (БХ) аллелем. “KRAB” относится к домену репрессии KRAB из гена KOX1, и “ZFP” относится к ДНК-связывающему белку “цинковые пальцы”. “Стандартный ZFP TF” представляет собой гибридный белок ZFP с фактором транскрипции, в котором ДНК-связывающие домены цинковых пальцев связаны с доменом репрессии KRAB. Фиг. 1B показывает ZFP-TF, сконструированные для связывания внутри области CAG. Фиг. 1C отображает “двурукий ZFP TF,” который представляет собой ZFP-фактор транскрипции, в котором два кластера доменов цинковых пальцев разделены посредством жестких белковых последовательностей. Функциональный домен (репрессии) отображается снаружи по отношению к одному ZFP на этой Фиг., но будет очевидным, что функциональный домен может находиться между ZFP или снаружи по отношению к ZFP на любом конце белка. Фиг. 1D отображает “мультимеризующийся ZFP TF,” который представляет собой ZFP TF, который способен к мультимеризации через домен мультимеризации (изображенный в виде крапчатых коробочек). Фиг. 1E отображает конфигурацию ZFP-ZFP-KRAB, где два ДНК-связывающих домена цинковые пальцы связаны посредством гибкого линкера и также слиты с доменом KRAB. Для квалифицированного специалиста в области будет очевидным то, что во всех гибридных белках, функциональный домен может находиться по обоим концам ДНК-связывающего домена, и что ДНК-связывающий домен может содержать широкий числовой диапазон цинковых пальцев. Также на Фиг. 1 в виде коробочки с черными алмазами изображен функциональный домен (например, активации, репрессии, домен расщепления). Для квалифицированного специалиста в области будет очевидным то, что иллюстративные модели, представленные на Фиг., могут также применяться к TALE TF.
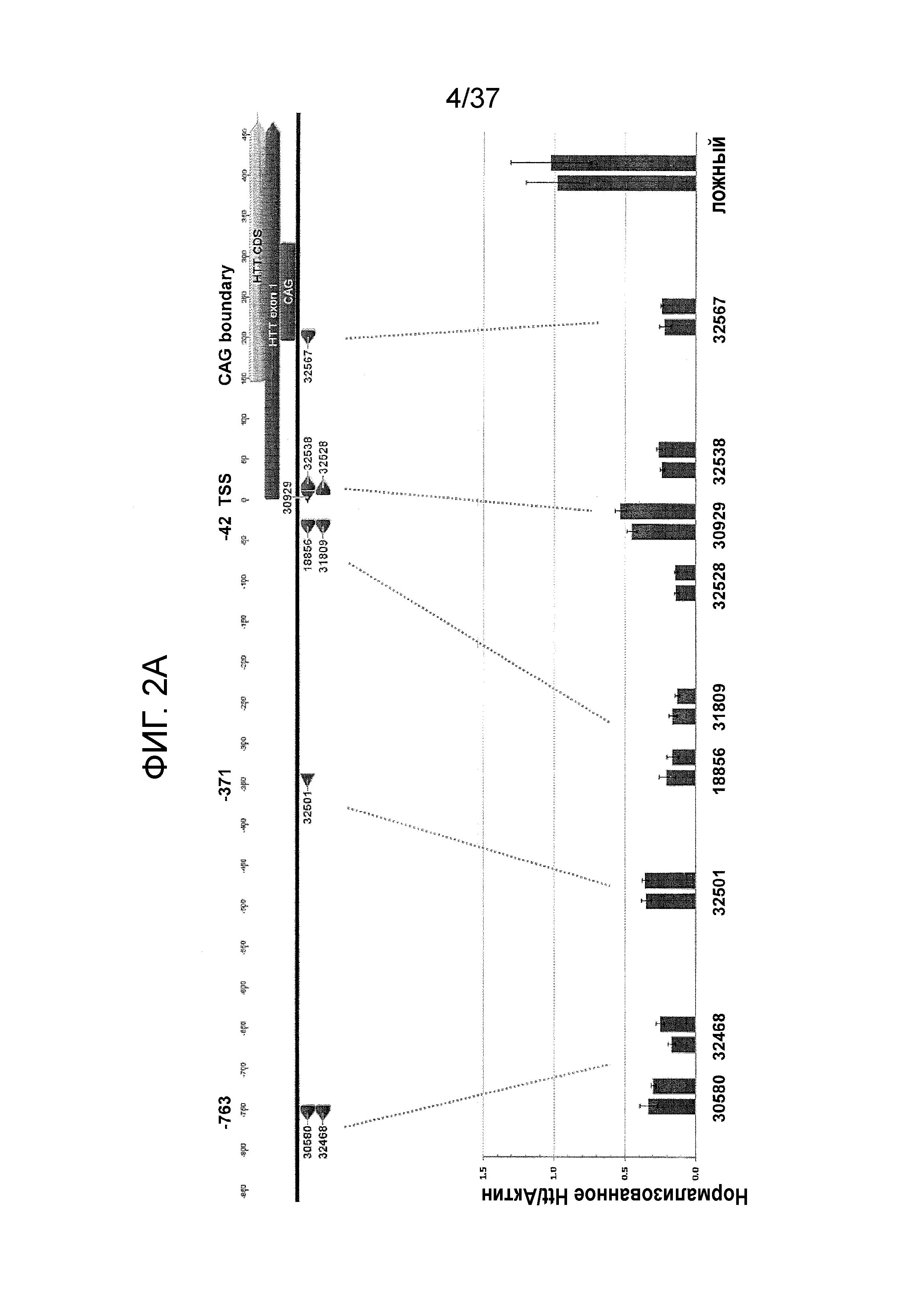
[0035] Фиг. 2A-2E отображают репрессию обоих аллелей Htt под действием ZFP, как описано в Фиг. 1A с использованием ZFP TF, которые не связываются с последовательностями CAG-повтора. Идентификационные номера ZFP, показанные в Таблицах 1A и 1B, указаны ниже столбиков. Фиг. 2A отображает репрессию человеческих аллелей Htt в клетках HEK293 с использованием ZFP, нацеленных на пять локусов в гене человека. Показана диаграмма человеческого гена Htt и показаны расположения сайтов связывания ZFP. Для каждой группы ZFP, каждый столбик представляет независимую трансфекцию. Фиг. 2B отображает Вестерн-блоттинг, показывающий уровни белка Htt в клетках HEK293, трансфицированных с использованием GFP-контроля или репрессора ZFP TF 18856 (содержащего домен репрессии KRAB из KOX1), где уровни NFκB p65 (“p65”) применяли для подтверждения равнозначной загрузки белка. Вестерн-блоттинг подтверждает репрессию экспрессию Htt посредством ZFP-TF. Фиг. 2C отображает аналогичный набор данных, как и Фиг. 2A для ZFP, специфичных к мышиному Htt в клетках Neuro2A. Как и на Фиг. 2A, показана диаграмма мышиного гена Htt и указаны расположения сайтов связывания ZFP. Фиг. 2D и 2E демонстрируют репрессию экспрессии мышиного гена Htt (РНК) в иммортализованных стриарных клетках, где для трансфекции применяли различные дозы ZFP-TF мРНК. Во всех случаях за исключением Фиг. 2B, уровни мРНК Htt измеряли посредством ПЦР в реальном масштабе времени и нормализовали к уровням мРНК Актина.
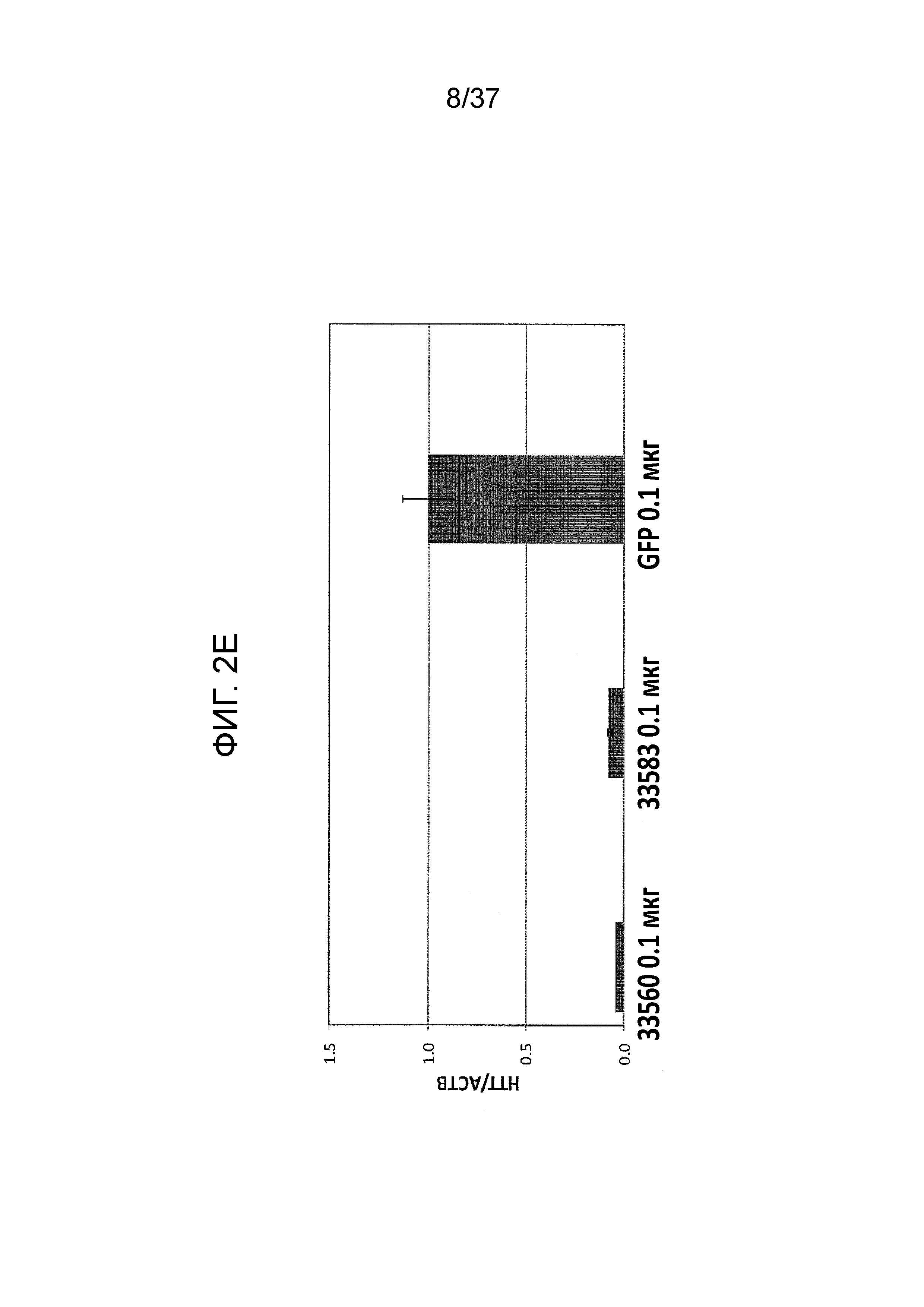
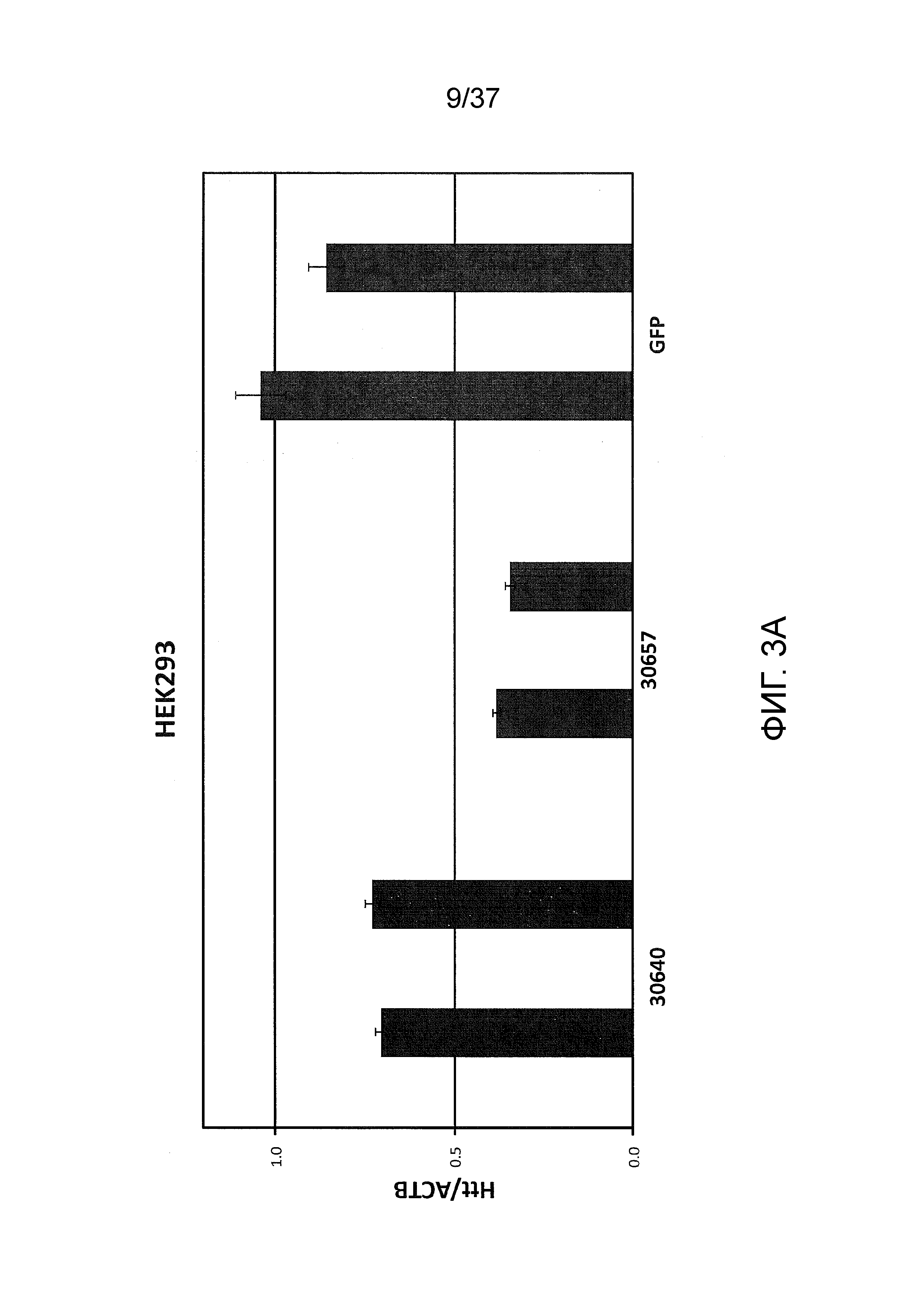
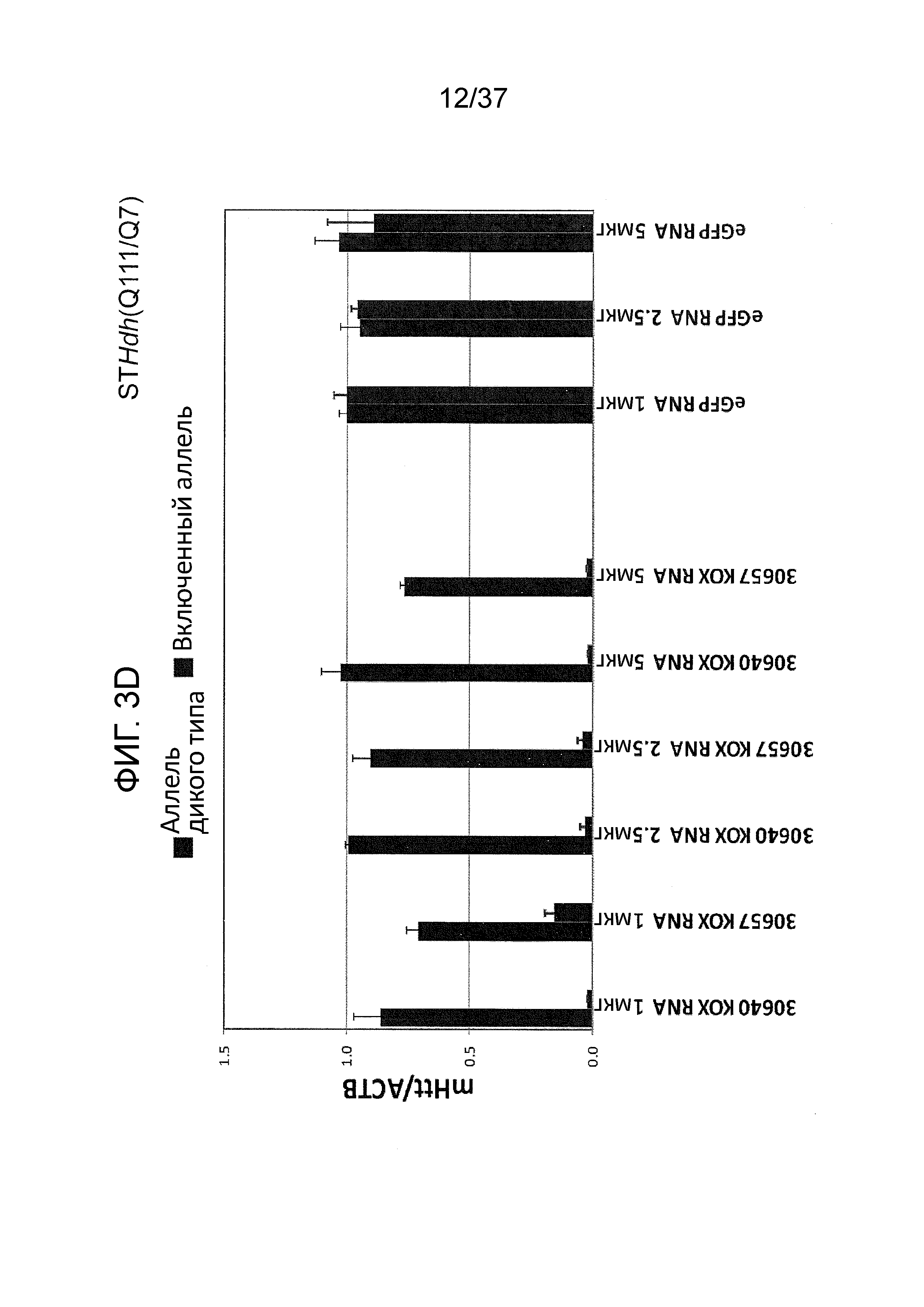
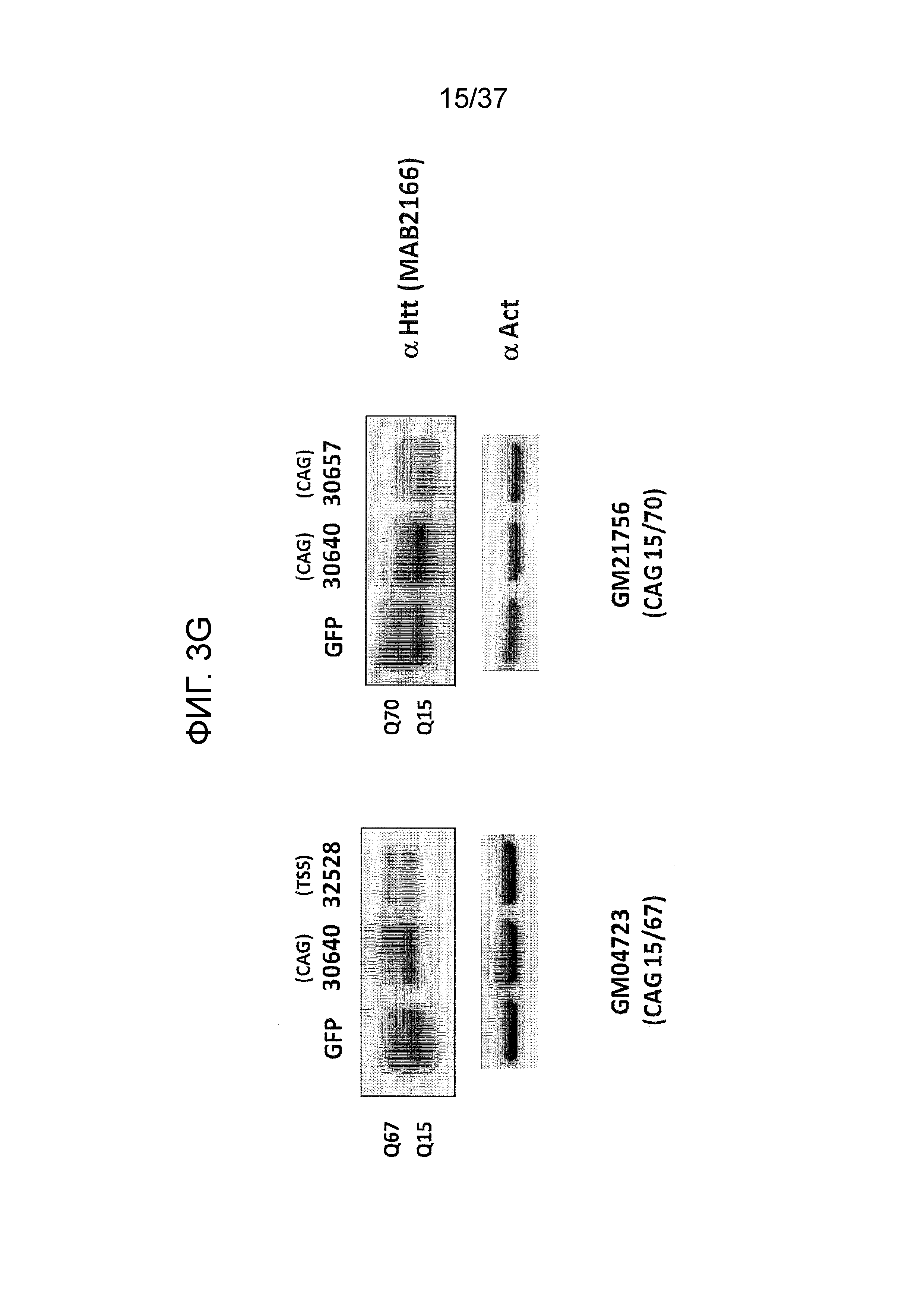
[0036] Фиг. 3A-3G отображают селективную репрессию мутантного Htt посредством использования ZFP, связывающихся внутри области повтора CAG, как проиллюстрировано на Фиг. 1B. Данная модель иллюстрирует, что более длинная область повтора CAG в мутантном аллеле обеспечивает увеличенное связывание CAG-нацеленных репрессорных молекул ZFP. Фиг. 3A отображает различные репрессорные активности по отношению к эндогенному гену Htt (с нормальной длиной CAG-повтора) под действием CAG-нацеленных ZFP в клетках HEK293. Фиг. 3B показывает репрессию люциферазных репортеров, управляемую фрагментами промотор/экзон1 Htt, содержащими CAG-повторы с изменяющейся длиной, в интервале от 10 до 47 CAG-повторов. CAG10 (левый-наибольший столбик для каждого из двух указанных условий) показывает результаты с 10 CAG-повторами; CAG17 (столбик, второй слева для каждого из двух указанных условий) показывает результаты с 17 CAG-повторами; CAG23 (столбик, второй справа для каждого из двух указанных условий) показывает результаты с 23 CAG-повторами; и CAG47 (правый-наибольший столбик для каждого из двух указанных условий) показывает результаты с 47 CAG-повторами. Схематическое указанное выше графическое изображение отображает расположение Htt-промотора, экзона 1, CAG-повторов и гена репортерной люциферазы, применяемых в данной системе. Данные демонстрируют, что возрастающее число CAG приводит к сниженной экспрессии от Htt-промотора под действием CAG-нацеленной ZFP. Кроме того, Фиг. 3C демонстрирует, что, в то время как относительно слабый CAG-нацеленный ZFP не подавляет люциферазный репортер, который содержит CAG-повтор с нормальной длиной, также как и сильный CAG-репрессор, он направляет аналогичную репрессию люциферазного репортера, который содержит удлиненный CAG-повтор, как сильный CAG-нацеленный ZFP при всех тестируемых дозах. “pRL-Htt-CAG23-интрон 1” (левый столбик из каждой пары) соответствует экспрессии из аллеля дикого типа, в то время как “pRL-HttCAG47-интрон 1” (правый столбик из каждой пары) коррелирует с экспрессией от мутантного удлиненного аллеля Htt (содержащего 47 CAG-повторов). Фиг. 3D представляет собой диаграмму, отображающую репрессию мутантного Htt (111 CAG) под действием CAG-нацеленных ZFP в иммортализованных мышиных стриарных клетках, полученных от мышей с включенным HdH(Q111/Q7). Экспрессия для дикого типа показана в левом столбике из каждой пары, и экспрессия включенного гена показана в правом столбике из каждой пары. ZFP-TF, содержащие установленную ZFP, слитую с доменом репрессии KRAB, тестировали с использованием трех различных концентраций мРНК ZFP при трансфекциях. Фиг. 3E отображает репрессию мутантного Htt под действием CAG-нацеленных ZFP в линии фибробластов, полученных от пациента с БХ, (CAG15/70). В этой линии фибробластов, аллель Htt дикого типа содержит 15 CAG-повторов (“099T(CAG15)”, средний столбик из каждого указанного условия), а мутантный удлиненный аллель Htt содержит 70 CAG-повторов (“099C(CAG70)”, правый столбик из каждого указанного условия). Фиг. 3F показывает селективную репрессию экспрессии мутантного Htt в 4 различных линиях клеток фибробластов, полученных от пациента с БХ. Числа сверху каждой группы указывают число CAG-повторов на аллеле Htt дикого типа (например, 15 или 18) и на мутантном аллеле (например, 70, 67, 45 и 44); где тестировали две различные дозы мРНК ZFP. Левый столбик из каждой пары показывает экспрессию дикого типа Htt, а правый столбик из каждой пары показывает экспрессию мутантного Htt. Фиг. 3G отображает экспрессию Htt в фибробластах, полученных от пациента с БХ, тестируемую посредством анализа методом Вестерн-блоттинга в присутствии ZFP-TF 30640, 32528 и 30657. Более медленно перемещающиеся полосы белков соответствуют белкам, продуцируемым удлиненными мутантными аллелями Htt. 32528 связывается с сайтом начала транскрипции из Htt (TSS) и, таким образом, ингибирует экспрессию от обоих аллелей, в то время как 30640 и 30657 связываются с CAG-повторами (CAG).
[0037] Фиг. 4A и 4B отображают репрессию мутантного Htt в линии фибробластов, полученных от пациента с БХ, посредством панели ZFP, нацеленных на CAG-повтор. Применяли интервал концентраций РНК от 0,1 нг до 3 мкг. На Фиг. 4A и 4B, левый столбик для каждого из указанных условий показывает общую экспрессию Htt, средний столбик показывает экспрессию Htt в фибробластах, в которых аллель Htt содержит 18 CAG-повторов (“099T(CAG18)”, а мутантный удлиненный аллель Htt содержит 45 CAG-повторов 099T (CAG45).
[0038] Фиг. 5 показывает воздействие CAG-нацеленных репрессоров ZFP на экспрессию Htt и других CAG-содержащих генов в фибробластах от пациента с БХ. Левый столбик под каждым указанным условием показывает результаты с 30640; средний столбик под каждым указанным условием показывает результаты с 30675; и правый столбик показывает ложные трансфекции.
[0039] Фиг. 6A и 6B отображают эксперимент, в котором исследуют полногеномную специфичность трех CAG-нацеленных ZFP. Фиг. 6A отображает анализ методом колПЦР репрессии Htt, выполняемый в шести биологических параллельных опытах (шесть раздельных трансфекций), БХ фибробластов (CAG18 (средние столбики)/CAG45, правые столбики) с использованием 30640, 30645, или 33074. Четыре наиболее сходных параллельных опыта посредством колПЦР затем отбирали для микроматричного анализа, и данные представлены на Фиг. 6B.
[0040] Фиг. 7 отображает репрессию Htt в нейрональных стволовых клетках CAG17/69 (NSC). Клетки трансфицировали мРНК ZFP при указанных дозах. Левые столбики под каждой из указанных доз показывают результаты в клетках CAG17, средние столбики показывают результаты в клетках дикого типа, правые столбики показывают результаты в клетках CAG69.
[0041] Фиг. 8 отображает экспрессию Htt в нейронах, дифференцированных из эмбриональных стволовых клеток БХ (ESC) (CAG 17/48), обработанных ZFP TF. Клетки трансфицировали с использованием РНК ZFP при указанных дозах.
[0042] Фиг. 9 отображает репрессию экспрессии мутантного трансгена Htt у мышей R6/2 после обработки ZFP TF 30640.
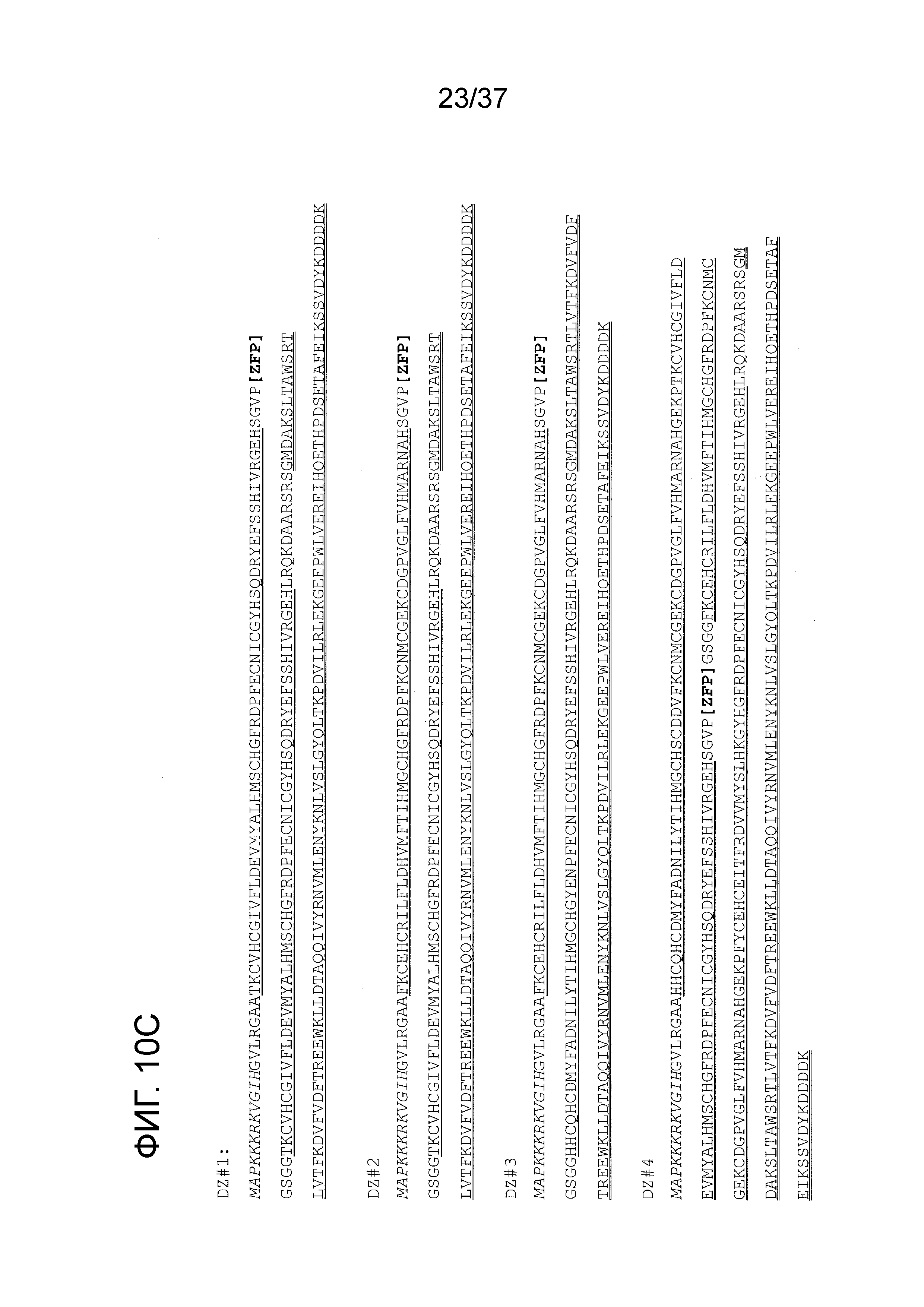
[0043] Фиг. 10A-10D отображают ZFP с доменами мультимеризации, которые специфически нацелены на удлиненные CAG-повторы, как проиллюстрировано на Фиг. 1D. Фиг. 10A показывает одиночную ZFP, которая имеет четыре компонента: (i) домен репрессора KOX (помеченный овалом “репрессор”); (ii) матрицу из 2-6 пальцев (два показаны, небольшие овалы с маркировкой “Z”), которая связывается с (CAG)N или пермутацией этой последовательности; и (iii) два домена димеризации (прямоугольники, помеченные “d1” и “d2”), которые взаимодействуют в антипараллельной конфигурации. Эти домены позволяют ZFP полимеризоваться внутри большой бороздки тракта CAG. Фиг. 10B показывает эскиз события связывания с мультимером из 3 ZFP. Будет очевидно, что может применяться любое число мультимеров, и, что функциональный домен может быть расположен в любом месте на одном или нескольких индивидуальных ZFP, и, что эти диаграммы также применимы к TALE-TF. Фиг. 10C показывает белковые последовательности четырех мономерных матриц ZFP, которые сконструированы, чтобы мультимеризоваться посредством взаимодействий между димеризующими цинковыми пальцами (DZ). Матрицы имеют названия DZ1 (SEQ ID NO:180), DZ2 (SEQ ID NO:181), DZ3 (SEQ ID NO:182) и DZ4 (SEQ ID NO:183). Домены димеризующих цинковых пальцев подчеркнуты, в то время как домен репрессии и последовательность ядерной локализации указаны жирным подчеркиванием и курсивом (соответственно). Фиг. 10D показывает белковые последовательности семи мономерных матриц ZFP, которые сконструированы, чтобы мультимеризоваться посредством взаимодействий между суперспиралями (CC). Матрицы имеют названия CC1 (SEQ ID NO:184), CC2 (SEQ ID NO:185), CC3 (SEQ ID NO:186), CC4 (SEQ ID NO:187), CC5 (SEQ ID NO:188), CC6 (SEQ ID NO:189) и CC7 (SEQ ID NO:190). Суперспиральные последовательности подчеркнуты, в то время как домен репрессии и последовательность ядерной локализации указаны жирным подчеркиванием и курсивом (соответственно). Положение области ZFP каждой матрицы, которое будет изменяться для конструкций, указано посредством “[ZFP]”. Положение (ДНК-связывающей) области ZFP каждой матрицы, которое будет изменяться для конструкций, указано посредством “[ZFP].”
[0044] Фиг. 11A и 11B отображают активность ZFP-TF с доменами димеризации. На Фиг. 11A, ZFP-TF с “суперспиральными” (CC) доменами тестировали с использованием люциферазных репортеров. pRL-Htt CAG17 (левый столбик из каждой пары) обозначает люциферазный репортер рениллы, управляемый фрагментом промотор/экзон1 человеческого Htt с 17 CAG; pGL3-Htt-CAG47 (правый столбик из каждой пары) обозначает люциферазный репортер светлячка, управляемый фрагментом промотор/экзон1 человеческого Htt с 47 CAG-повторами. См. текст в Примере 10 для описания различных доменов димеризации. На Фиг. 11B, ZFP с доменами димеризующих цинковых пальцев “DZ” тестировали с такими же люциферазными репортерами, и демонстрируется увеличенная репрессия с несколькими доменами димеризации ZFP-TF. Левый столбик в каждом дублете показывает экспрессию от аллеля Htt с 17 CAG-повторами, в то время как правый столбик показывает экспрессию от аллеля Htt с 47 CAG-повторами.
[0045] Фиг. 12A и 12B отображают репрессию Htt под действием белков ZFP-ZFP-KOX. Фиг. 12A отображает репрессию Htt под действием одиночных ZFP-TF 33088 и 33084, и репрессию под действием белков ZFP-ZFP-KOX 33088-33088 и 33088-33084 для дикого типа (левый столбик), CAG18 (средний столбик) и CAG45 (правый столбик) (Фиг. 12A) фибробластов при БХ; Фиг. 12B отображает репрессию Htt под действием ZFP-ZFP-KOX 33088-33088 и 33088-33084 для дикого типа (левый столбик), CAG 20 (средний столбик) и CAG41 (левый столбик) фибробластов при БХ.
[0046] Фиг. 13A-13E отображают активацию мышиного Htt. Фиг. 13A демонстрирует ZFP-TF-управляемую повышающую регуляцию мышиных генов Htt на уровне РНК в клетках Neuro2A с использованием ZFP, слитого с доменом активации p65. Двойные столбики указывают на дублирующие трансфекции. Фиг. 13B отображает Вестерн-блоттинг, демонстрирующий увеличенную выработку белка Htt, управляемую ZFP. Фиг. 13C отображает мышиный аллель Htt дикого типа и “включенный” аллель Htt, где мышиная последовательность (наибольшая часть экзона 1 и часть интрона 1, схематичная линия выше аллеля дикого типа) была заменена на соответствующую человеческую последовательность с использованием экспансии CAG (схематичная линия над включенным аллелем). Фиг. 13D отображает выравнивание между мышиной последовательностью (SEQ ID NO:191), которая была заменена на соответствующую человеческую последовательность (SEQ ID NO:192) таким образом, что включенный аллель имеет достаточную дивергенцию последовательности, чтобы обеспечить ZFP (показанному на A и B) конструирование для специфического связывания с мышиной последовательностью. Фиг. 13E отображает специфичную активацию мышиного аллеля Htt дикого типа в иммортализованных стриарных клетках полученных от мышей с включенным HdhQ111/Q7 мышей. Левый-столбик показывает результаты для клеток дикого типа, и правый столбик показывает результаты для клеток с включенным мутантном аллелем.
[0047] Фиг. 14A и 14B отображают результаты анализа Cel-I ошибок спаривания (Surveyor™, Transgenomeic) после обработки клеток K562 Htt-специфичными парами ZFN. Процент активности NHEJ (вставки-делеции) для активной ZFN показан в нижней части соответствующей полосы. “GFP” указывает клетки, которые были трансфицированы GFP-кодирующей плазмидой. Фиг. 14A отображает результаты для ZFN, которая расщепляет ранние экзоны Htt, в то время как Фиг. 14B отображает результаты для ZFN, которая расщепляет находящийся рядом стоп-кодон. Также наблюдали неактивные пары ZFN (полосы не сокращены процентным содержанием вставок-делеций).
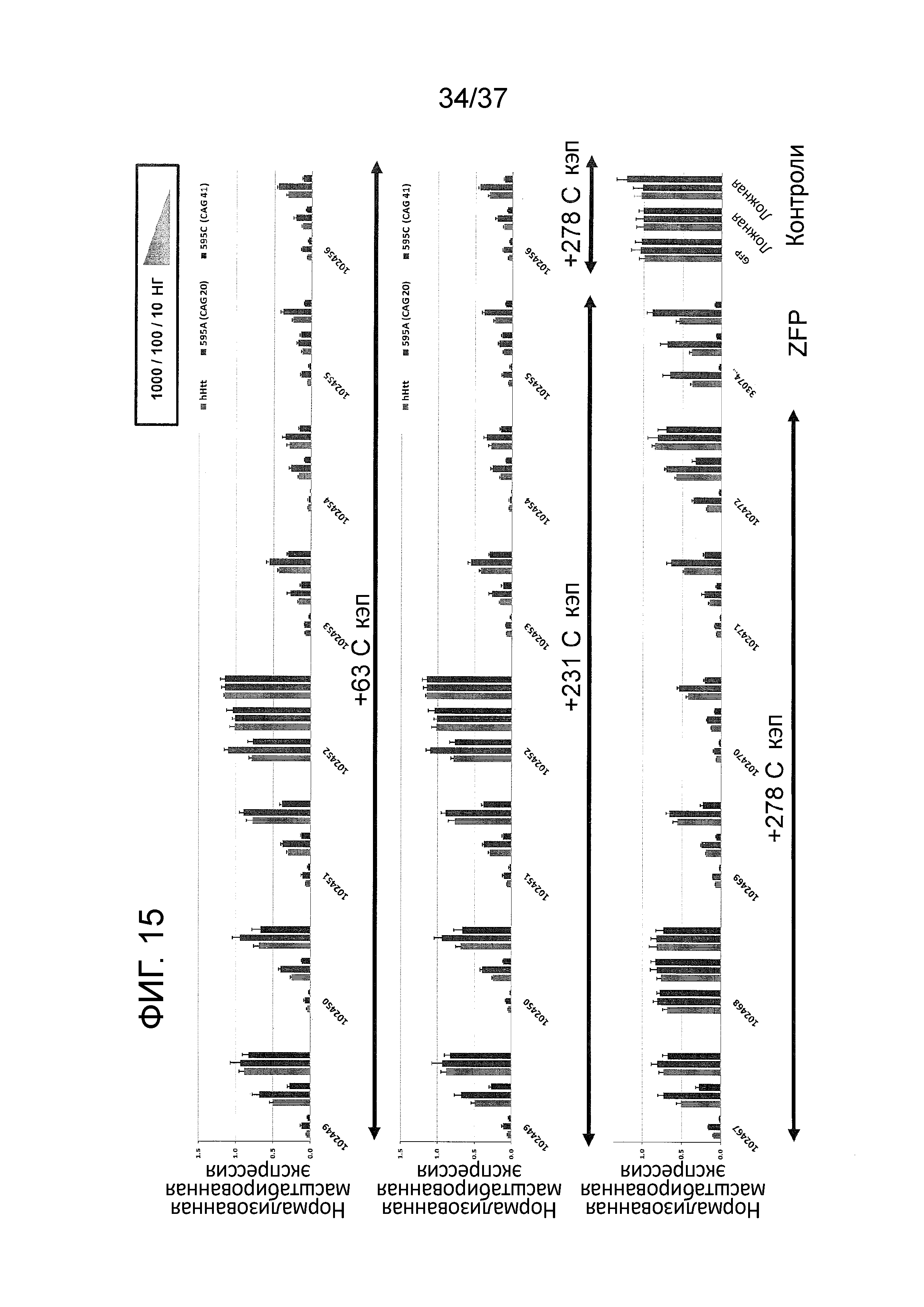
[0048] Фиг. 15 отображает графические изображения результатов репрессии Htt для нескольких кандидатных белков TALE-TF. TALE-TF тестировали на фибробластах от пациента с БХ (CAG 20/41). Результаты демонстрируют, что несколько из TALE TF были активными при подавлении общей экспрессии Htt, в то время как другие проявляли преимущественную репрессию мутантного Htt.
[0049] Фиг. 16 показывает две фотографии, отображающие хватательный рефлекс (Mangiarini et al (1996) Cell 87:493-605). Тест включает поднимание вверх тестируемых мышей за их хвост, где наблюдатель осторожно тянет их назад и вверх плавным движением, пока животное не окажется подвешенным над поверхностью на высоту примерно 12 дюймов. Животное затем оценивают баллами в течение 30 секунд. Мыши с БХ задействуют поведение ʹсжиманияʹ, как показано на фото справа, в то время как нормальные контрольные мыши проявляют более открытое поведение, показанное слева.
[0050] Фиг. 17 представляет собой графическое отображение хватательного рефлекса на мышиной модели БХ (R6/2). Мышей подвешивают в течение 30 секунд и оценивают баллами по типу и длине хватательного рефлекса в течение периода времени. Как можно наблюдать, мыши, обработанные ZFP, показывали снижение хватательного рефлекса для всех отметок времени на протяжении 12-недельного периода.
[0051] Фиг. 18 представляет собой графическое отображение изменения экспрессии четырех биомаркеров для срединных шипиковых нейронов, полученных из мозга мышей R6/2, обработанных ZFP. Все из четырех проанализированных биомаркеров, DARPP-32, PDE10a, DRD1 и DRD2, показали увеличение экспрессии относительно сигнала от мозга мышей R6/2, которые были обработаны вектором экспрессии GFP.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0052] В данном описании раскрыты композиции и способы для лечения болезни Хантингтона (БХ). В частности, предоставлены Htt-модулирующие факторы транскрипции, содержащие белки “цинковые пальцы” (ZFP-TF) или TALE (TALE-TF) и способы использования таких белков для применения при лечении или предотвращении болезни Хантингтона. Например, предоставлены ZFP-TF или TALE-TF, которые подавляют экспрессию мутантного аллеля Htt или активируют экспрессию аллеля Htt дикого типа. В дополнение, предоставлены нуклеазы “цинковые пальцы” (ZFN), нуклеазы TALE (TALEN), системы Ttago или нуклеазные системы CRISPR/Cas, которые модифицируют геномную структуру генов, ассоциированных с БХ. Например, предоставлены нуклеазные системы ZFN, TALEN или CRISPR/Cas, которые обладают способностью специфически изменять части мутантной формы Htt. Они включают композиции и способы с использованием сконструированных белков “цинковые пальцы” или сконструированных белков TALE, т.е. не встречающихся в природе белков, которые связываются с заданной целевой последовательностью нуклеиновой кислоты.
[0053] Таким образом, способы и композиции, описанные в данном документе, предоставляют способы для лечения и предотвращения болезни Хантингтона, и эти способы и композиции могут включать факторы транскрипции “цинковые пальцы” или факторы транскрипции TALE, которые обладают способностью к модулированию генов-мишеней, а также сконструированные нуклеазы “цинковые пальцы” и TALE, Ttago и нуклеазные системы CRISPR/Cas, способные к модификации или редактированию Htt.
Общая часть
[0054] В данном описании раскрыты практическая реализация способов, а также получение и применение композиций с использованием, если не указано иначе, общепринятых технологий в молекулярной биологии, биохимии, структуры и анализа хроматина, вычислительной химии, культивировании клеток, рекомбинантных ДНК и родственных областей, которые находятся в пределах квалификации в данной области. Эти технологии полностью объясняются в литературе. См, например, Sambrook et al. MOLECULAR CLONING: A LABORATORY MANUAL, Second edition, Cold Spring Harbor Laboratory Press, 1989 и Third edition, 2001; Ausubel et al., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, New York, 1987 и периодические обновления; серии METHODS IN ENZYMOLOGY, Academic Press, San Diego; Wolffe, CHROMATIN STRUCTURE AND FUNCTION, Third edition, Academic Press, San Diego, 1998; METHODS IN ENZYMOLOGY, Vol. 304, “Chromatin” (P.M. Wassarman и A. P. Wolffe, eds.), Academic Press, San Diego, 1999; и METHODS IN MOLECULAR BIOLOGY, Vol. 119, “Chromatin Protocols” (P.B. Becker, ed.) Humana Press, Totowa, 1999.
Определения
[0055] Термины "нуклеиновая кислота" "полинуклеотид" и "олигонуклеотид" применяют взаимозаменяемо и относят к дезоксирибонуклеотидному или рибонуклеотидному полимеру, в линейной или кольцевой конформации, и либо в одно- или двухцепочечной форме. Для целей настоящего раскрытия, эти термины не должны рассматриваться как ограничивающие по отношению к длине полимера. Термины могут охватывать известные аналоги природных нуклеотидов, а также нуклеотиды, которые модифицируют по основанию, сахарным и/или фосфатным фрагментам (например, фосфотиоатные остовы). Обычно, аналог конкретного нуклеотида имеет такую же специфичность по спариванию оснований; т.е. аналог A будет образовывать пару основания с T.
[0056] Термины "полипептид", "пептид" и "белок" применяют взаимозаменяемо для отнесения к полимеру аминокислотных остатков. Термин также применяют к аминокислотным полимерам, в которых одна или несколько аминокислот являются химическими аналогами или модифицированными производными соответствующих природных аминокислот.
[0057] "Связывание" относится к специфичному по отношению к последовательности, нековалентному взаимодействию между макромолекулами (например, между белком и нуклеиновой кислотой). Не все компоненты взаимодействия связывания обязательно являются специфичными по отношению к последовательности (например, контакты с фосфатными остатками в остове ДНК), поскольку взаимодействие в целом является специфичным по отношению к последовательности. Такие взаимодействия обычно характеризуются константой диссоциации (Kd), равной 10-6M-1 или ниже. “Аффинность” относится прочности связывания: увеличенная аффинность связывания коррелирует с более низкой Kd.
[0058] "Связывающий белок" представляет собой белок, который обладает способностью нековалентно связываться с еще одной другой молекулой. Связывающий белок может связываться, например, с молекулой ДНК (ДНК-связывающий белок), молекулой РНК (РНК-связывающий белок) и/или молекулой белка (белок-связывающий белок). В случае белок-связывающего белка, он может связываться сам с собой (с образованием гомодимеров, гомотримеров и т.д.) и/или он может связываться с одной или несколькими молекулами отличающегося белка или белков. Связывающий белок может иметь более одного типа активности связывания. Например, белки “цинковые пальцы” имеют ДНК-связывающую, РНК-связывающую и белок-связывающую активность.
[0059] "ДНК-связывающий белок типа цинковые пальцы" (или связывающий домен) представляет собой белок, или домен внутри более крупного белка, который связывается с ДНК специфичным по отношению к последовательности образом посредством одного или нескольких “цинковых пальцев”, которые представляют собой области аминокислотной последовательности внутри связывающего домена, структура которого стабилизируется посредством координации ионом цинка. Термин ДНК-связывающий белок типа “цинковые пальцы” часто сокращают как белок “цинковые пальцы” или ZFP.
[0060] “ДНК-связывающий домен TALE” или “TALE” представляет собой полипептид, содержащий одну или несколько повторяющихся доменов/единиц TALE. Повторяющиеся домены вовлечены в связывание TALE с его когнатной целевой последовательностью ДНК. Одиночная “повторяющаяся единица” (также именуемая как “повтор”) обычно имеет длину 33-35 аминокислот и проявляет, по меньшей мере, некоторую гомологию последовательности с другой повторяющейся последовательностью TALE внутри природного белка TALE. См, например, Патент США № 8,586,526.
[0061] Связывающие домены цинковые пальцы или ДНК-связывающие домены TALE могут быть "сконструированы" чтобы связываться с заданной нуклеотидной последовательностью, например посредством конструирования (изменения одной или нескольких аминокислот) области спирали распознавания природного белка “цинковые пальцы” или посредством конструирования RVD белка TALE. Следовательно, сконструированные белки “цинковые пальцы” или TALE представляют собой белки, которые являются неприродными. Неограничивающие примеры методов конструирования белков “цинковые пальцы” или TALE являются проектирование и селекция. Проектированный белок “цинковые пальцы” или TALE представляет собой белок, не встречающийся в природе, проект/состав которого в основном являются результатом обоснованных критериев. Обоснованные критерии для проектирования включают применение правил замещения и компьютеризованные алгоритмы для обработки информации в базе данных, хранящей информацию о существующих проектах ZFP и данных по связыванию. См, например, Патенты США 8,586,526; 6,140,081; 6,453,242; и 6,534,261; см., также WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[0062] "Прошедший селекцию" белок “цинковые пальцы” или TALE представляет собой белок, не обнаруживаемый в природе, получение которого является результатом прежде всего эмпирического процесса, такого как отображение фага, ловушка взаимодействия или селекция гибридов. См., например, 8,586,526; 5,789,538; Патенты США 5,925,523; 6,007,988; 6,013,453; 6,200,759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970 WO 01/88197, WO 02/099084.
[0063] “TtAgo” представляет собой прокариотный белок Аргонавт, который, как предполагают, вовлечен в сайленсинг генов. TtAgo получают из бактерий Thermus thermophilus. См, например, Swarts et al, там же, G. Sheng et al., (2013) Proc. Natl. Acad. Sci. U.S.A. 111, 652). “Система TtAgo” представляет собой все требуемые компоненты, включая, например, ДНК-проводники для расщепления ферментом TtAgo.
[0064] "Рекомбинация" относится к процессу обмена генетической информацией между двумя полинуклеотидами. Для целей данного раскрытия, "гомологичная рекомбинация (HR)" относится к специализированной форме такого обмена, которая имеет место, например, во время репарации двухцепочечных разрывов в клетках посредством гомологично направленных механизмов репарации. Данный процесс требует гомологии нуклеотидной последовательности, в нем используется молекула "донора" для создания матрицы репарации молекулы "мишени" (т.е. молекулы, которая перенесла двухцепочечный разрыв), и известен в различных вариантах как "некроссинговерная конверсия генов" или "конверсия генов короткого тракта", поскольку он приводит к переносу генетической информации от донора к мишени. Не желая быть связанными соответствием конкретной теории, авторы полагают, что, такой перенос может включать коррекцию ошибок спаривания гетеродуплексной ДНК, которые образуются между мишенью с разрывами и донором, и/или "зависимый от синтеза отжиг цепи", при котором донор используется для повторного синтеза генетической информации, которая станет частью мишени, и/или родственных процессов. Такая специализированная HR часто приводит к изменению последовательности молекулы-мишени таким образом, что часть последовательности или вся последовательность донорного полинуклеотида вводится в целевой полинуклеотид.
[0065] В способах раскрытия, одна или несколько целенаправленных нуклеаз, описанных в данном документе, создают двухцепочечный разрыв в целевой последовательности (например, клеточном хроматине) в заданном сайте, и “донорный” полинуклеотид, имеющий гомологию к нуклеотидной последовательности в области разрыва, может быть введен в клетку. Было показано, что присутствие двухцепочечного разрыва способствует интеграции донорной последовательности. Донорная последовательность может быть физически интегрирована или, альтернативно, донорный полинуклеотид используется в качестве матрицы для репарации разрыва посредством гомологичной рекомбинации, приводящей к введению всей нуклеотидной последовательности или ее части как в доноре в клеточный хроматин. Таким образом, первая последовательность в клеточном хроматине может изменяться и, в некоторых вариантах осуществления, может быть преобразована в последовательность, присутствующую в донорном полинуклеотиде. Таким образом, применение терминов “заменять” или “замена” следует понимать как представление замены одной нуклеотидной последовательности на другую, (т.е. замены последовательности в информационном смысле), и необязательно требует физической или химической замены одного полинуклеотида на другой.
[0066] В любом из способов, описанных в данном документе, дополнительные пары белков “цинковые пальцы” или белков TALE могут применяться для дополнительного двухцепочечного расщепления дополнительных целевых сайтов внутри клетки.
[0067] В некоторых вариантах осуществления способов целенаправленной рекомбинации и/или замены и/или изменения последовательности в области интереса в клеточном хроматине, хромосомная последовательность изменяется посредством гомологичной рекомбинации с экзогенной “донорной” нуклеотидной последовательностью. Такая гомологичная рекомбинация стимулируется присутствием двухцепочечного разрыва в клеточном хроматине, если присутствуют последовательности, гомологичные области разрыва.
[0068] В любом из способов, описанных в данном документе, первая нуклеотидная последовательность (“донорная последовательность”) может содержать последовательности, которые являются гомологичными, но не идентичными, с геномными последовательностями в области интереса, таким образом, стимулируя гомологичную рекомбинацию, чтобы вставить неидентичную последовательность в область интереса. Таким образом, в некоторых вариантах осуществления, части донорной последовательности, которые являются гомологичными с последовательностями в области интереса, проявляют приблизительно 80-99% (или имеющую любое числовое значение между ними) идентичность последовательности с геномной последовательностью, которую заменяют. В других вариантах осуществления, гомология между донорной и геномной последовательностью составляет выше, чем 99%, например, если донорная и геномная последовательности отличаются только на 1 нуклеотид на 100 смежных пар оснований. В некоторых случаях, негомологичная часть донорной последовательности может содержать последовательности, не присутствующие в области интереса, таким образом, что новые последовательности вводятся в область интереса. В этих случаях, негомологичная последовательность обычно фланкируется последовательностями из 50-1000 пар оснований (или имеющих любое числовое значение между ними) или любым числом пар оснований, свыше 1000, которые являются гомологичными или идентичными с последовательностями в области интереса. В других вариантах осуществления, донорная последовательность является негомологичной с первой последовательностью, и встраивается в геном посредством механизмов негомологичной рекомбинации.
[0069] Любые из способов, описанных в данном документе, могут применяться для частичной или полной инактивации одной или нескольких целевых последовательностей в клетке посредством целенаправленной интеграции донорной последовательности, которая нарушает экспрессию гена (генов), представляющих интерес. Также предоставлены линии клеток с частично или полностью инактивированными генами.
[0070] Кроме того, способы целенаправленной интеграции, описанные в данном документе, могут также применяться для интеграции одной или нескольких экзогенных последовательностей. Экзогенная последовательность нуклеиновой кислоты может включать, например, один или более генов или молекул кДНК, или любой тип кодирующей или некодирующией последовательности, а также один или несколько контролирующих элементов (например, промоторов). В дополнение, экзогенная последовательность нуклеиновой кислоты может продуцировать одну или более молекул РНК (например, малые шпилечные РНК (мшРНК), ингибирующие РНК (РНКинг), микроРНК (миРНК), и т.д.).
[0071] "Расщепление" относится к разрыванию ковалентного остова молекулы ДНК. Расщепление может быть инициировано различными методами, включающими, но не ограниченными приведенными, ферментативный или химический гидролиз фосфодиэфирной связи. Как одноцепочечное расщепление, так и двухцепочечное расщепление являются возможными, и двухцепочечное расщепление может происходить как результат двух различающихся событий одноцепочечного расщепления. Расщепление ДНК может приводить в результате к продуцированию либо “тупых” концов или ступенчатых концов. В некоторых вариантах осуществления, гибридные полипептиды применяют для целенаправленного расщепления двухцепочечной ДНК.
[0072] "Половинный домен расщепления" представляет собой полипептидную последовательность, которая, в сочетании со вторым полипептидом (либо идентичным или отличающимся) образует комплекс, имеющий активность расщепления (предпочтительно активность двухцепочечного расщепления). Термины “первый и второй половинные домены расщепления”; “+ и - половинные домены расщепления” и “правый и левый половинные домены расщепления” применяют взаимозаменяемо для обозначения пар половинных доменов расщепления, которые димеризуются.
[0073] “Сконструированный половинный домен расщепления” является половинным доменом расщепления, который был модифицирован таким образом, чтобы образовывать облигатные гетеродимеры с еще одним другим половинным доменом расщепления (например, еще одним сконструированным половинным доменом расщепления). См, также, Патентные публикации США №№ 2005/0064474, 20070218528, 2008/0131962 и 2011/0201055, полностью включенные в данное описание посредством ссылки.
[0074] Термин "последовательность" относится к нуклеотидной последовательности любой длины, которая может представлять собой ДНК или РНК; может быть линейной, круговой или разветвленной и может быть либо одноцепочечной или двухцепочечной. Термин "донорная последовательность" относится к нуклеотидной последовательности, которую вставляют в геном. Донорная последовательность может иметь любую длину, например, длину между 2 и 10000 нуклеотидами (или с любым целочисленным значением между ними или выше их), предпочтительно, длину между приблизительно 100 и 1000 нуклеотидами (или с любым целочисленным значением между ними), более предпочтительно, длину между приблизительно 200 и 500 нуклеотидами.
[0075] "Хроматин" является нуклеопротеиновой структурой, содержащей клеточный геном. Клеточный хроматин содержит нуклеиновую кислоту, прежде всего, ДНК, и белок, включая гистонные и негистонные хромосомные белки. Большая часть эукариотного клеточного хроматина существует в форме нуклеосом, где центральная часть нуклеосомы содержит приблизительно 150 пар оснований ДНК, ассоциированных с октамером, содержащим по два каждого из гистонов H2A, H2B, H3 и H4; и линкерную ДНК (с переменной длиной, зависящей от организма) проходит между центральными частями нуклеосом. Молекула гистона H1 обычно ассоциирована с линкерной ДНК. Для целей настоящего раскрытия, термин “хроматин” означает охват всех типов клеточного нуклеопротеина, как прокариотных, так и эукариотных. Клеточный хроматин включает как хромосомный, так и эписомальный хроматин.
[0076] "Хромосома" представляет собой хроматиновый комплекс, содержащий весь геном или часть генома клетки. Геном клетки часто характеризуется его кариотипом, который является коллекцией всех хромосом, которые составляют геном клетки. Геном клетки может содержать одну или более хромосом.
[0077] "Эписома" является реплицирующейся нуклеиновой кислотой, нуклеопротеиновым комплексом или другой структурой, содержащей нуклеиновую кислоту, которая не является частью хромосомного кариотипа клетки. Примеры эписом включают плазмиды и некоторые вирусные геномы.
[0078] "Сайт-мишень" или "последовательность-мишень" представляет собой последовательность нуклеиновой кислоты, которая определяет часть нуклеиновой кислоты, с которой будет связываться связующая молекула, если существуют достаточные условия для связывания.
[0079] "Экзогенная" молекула является молекулой, которая в норме не присутствует в клетке, но может быть введена внутрь клетки посредством одного или нескольких генетических, биохимических или других методов. “Нормальное присутствие в клетке” определяют по отношению к конкретной стадии развития и условий окружающей среды клетки. Таким образом, например, молекула, которая присутствует только во время эмбрионального развития мышцы, является экзогенной молекулой по отношению к взрослой мышечной клетке. Аналогично, молекула, индуцируемая тепловым шоком, является экзогенной молекулой по отношению к клетке, не подвергнутой воздействию теплового шока. Экзогенная молекула может включать, например, функционирующую версию неправильно функционирующей эндогенной молекулы или неправильно функционирующую версию нормально функционирующей эндогенной молекулы.
[0080] Экзогенная молекула может представлять собой, среди прочего, малую молекулу, такую как молекулу, генерируемую посредством процесса комбинаторной химии, или макромолекулу, такую как белок, нуклеиновую кислоту, углевод, липид, гликопротеин, липопротеин, полисахарид, любое модифицированное производное указанных выше молекул, или любой комплекс, содержащий одну или несколько из указанных выше молекул. Нуклеиновые кислоты включают ДНК и РНК, могут быть одно- или двухцепочечными; могут быть линейными, разветвленными или кольцевыми; и могут иметь любую длину. Нуклеиновые кислоты включают нуклеиновые кислоты, способные к образованию дуплексов, а также триплекс-образующие нуклеиновые кислоты. См, например, Патенты США №№ 5,176,996 и 5,422,251. Белки включают, но не ограничиваются перечисленным, ДНК-связывающие белки, факторы транскрипции, факторы реконструкции хроматина, метилированные ДНК-связывающие белки, полимерaзы, метилазы, деметилазы, ацетилазы, деацетилазы, киназы, фосфатазы, интегразы, рекомбиназы, лигазы, топоизомеразы, гиразы и геликазы.
[0081] Экзогенная молекула может иметь такой же тип молекулы, как эндогенная молекула, например, представлять собой экзогенный белок или нуклеиновую кислоту. Например, экзогенная нуклеиновая кислота может содержать геном инфицирующего вируса, плазмиду или эписому, вводимую внутрь клетки, или хромосому, которая обычно не присутствует в клетке. Методы введения экзогенных молекул в клетки известны специалистам в данной области и включают, но не ограничиваются перечисленным, липид-опосредованный перенос (т.е. липосомы, включающие нейтральные и катионные липиды), электропорацию, прямую инъекцию, слияние клеток, бомбардировку частицами, соосаждение с фосфатом кальция, перенос, опосредованный ДЭАЭ-декстраном, и перенос, опосредованный вирусным вектором. Экзогенная молекула также может иметь такой же самый тип молекулы, как и эндогенная молекула, но быть полученной из вида, отличающегося от вида, из которого получают клетку. Например, последовательность нуклеиновой кислоты человека может быть введена в линию клеток, исходно полученную от мыши или хомяка.
[0082] Напротив, "эндогенная" молекула представляет собой молекулу, которая обычно присутствует в конкретной клетке на конкретной стадии развития при конкретных условиях окружающей среды. Например, эндогенная нуклеиновая кислота может включать хромосому, геном митохондрии, хлоропласта или другой органеллы, или природную эписомальную нуклеиновую кислоту. Дополнительные эндогенные молекулы могут включать белки, например, факторы транскрипции и ферменты.
[0083] Термин “генетический репрессор” относится к любой молекуле, которая снижает в некоторой степени экспрессию одного или нескольких генов. Неограничивающие примеры генов, которые могут подвергаться репрессии, включают биомаркеры, относящиеся к БХ (например, DARPP-32A, PDE10a, Drd1 и/или Drd2). Такая генетическая репрессия может в результате приводить к лечению и/или предотвращению болезни Хантингтона у субъекта. Генетический репрессор может представлять собой любую молекулу, включая, но не ограничиваясь приведенными, малую молекулу, нуклеиновую кислоту и/или белок (например, белок, содержащий неприродный ДНК-связывающий домен, такой как белок “цинковые пальцы”, CRISRP/Cas или домен TALE), который ингибирует экспрессию гена (например, гена хантингтин). Генетические репрессорные белки могут содержать гибридные молекулы, например гибридные белковые факторы транскрипции (например, ZFP-TF, CRISPR/dCas-TF или TALE-TF), содержащие ДНК-связывающий домен и домен транскрипционной регуляции, или нуклеазы (например, ZFN, TALEN, CRISPR/Cas и/или нуклеазные системы Ttago), содержащие ДНК-связывающий домен и домен нуклеазы. В нескольких вариантах осуществления, генетический репрессор специфически связывается с геном хантингтина, в его геномной ДНК или в форме транскрипта, такой как мРНК.
[0084] "Гибридная" молекула представляет собой молекулу, в которой две или более молекул-субъединиц связаны, предпочтительно, ковалентно. Молекулы-субъединицы могут иметь одинаковый химический тип молекулы, или могут принадлежать к различным химическим типам молекул. Примеры гибридной молекулы первого типа включают, но не ограничиваются перечисленным, гибридные белки (например, гибрид между ДНК-связывающим доменом ZFP или TALE и одним или несколькими доменами активации) и гибридные нуклеиновые кислоты (например, нуклеиновую кислоту, кодирующую гибридный белок, описанный выше). Примеры второго типа гибридной молекулы включают, но не ограничиваются перечисленным, гибрид между триплекс-образующей нуклеиновой кислотой и полипептидом, и гибрид между белком, связывающимся с малой бороздкой и нуклеиновой кислотой.
[0085] Экспрессия гибридного белка в клетке может быть результатом доставки гибридного белка в клетку или доставки полинуклеотида, кодирующего гибридный белок, в клетку, где полинуклеотид транскрибируется, и транскрипт транслируется, чтобы генерировать гибридный белок. Транс-сплайсинг, расщепление полипептида и лигирование полипептида также могут быть вовлечены в экспрессию белка в клетке. Способы доставки полинуклеотидов и полипептидов в клетки представлены в других разделах в данном раскрытии.
[0086] “Домен мультимеризации”, (также именуемый как “домен димеризации” или “домен белкового взаимодействия”) представляет собой домен, включенный в амино-, карбокси- или амино- и карбокси- концевые области ZFP TF или TALE TF. Эти домены обеспечивают мультимеризацию множества единиц ZFP TF или TALE TF, таким образом, что более крупные тракты доменов тринуклеотидных повторов становятся преимущественно связанными мультимеризованными ZFP TF или TALE TF относительно более коротких трактов с длинами, соответствующими дикому типу. Примеры доменов мультимеризации включают лейциновые “молнии”. Домены мультимеризации могут также регулироваться малыми молекулами, где домен мультимеризации принимает нужную конформацию, чтобы обеспечить взаимодействие с еще одним доменом мультимеризации только в присутствии малой молекулы или внешнего лиганда. Таким способом, экзогенные лиганды могут применяться для регуляции активности этих доменов.
[0087] "Ген", для целей настоящего раскрытия, включает область ДНК, кодирующую генный продукт (см., ниже), а также все области ДНК, которые регулируют продуцирование генного продукта, независимо от того, являются ли такие регуляторные последовательности примыкающими к кодирующим и/или транскрибируемым последовательностям. Соответственно, ген включает, но необязательно ограничен перечисленными, промоторные последовательности, терминаторы, трансляционные регуляторные последовательности, такие как сайты связывания рибосом и участки внутренней посадки рибосом, энхансеры, сайленсеры, инсуляторы, граничные элементы, точки начала репликации, сайты присоединения матрикса и регуляторные области локусов.
[0088] "Экспрессия гена" относится к превращению информации, содержащейся в гене, в генный продукт. Генный продукт может представлять собой продукт прямой транскрипции гена (например, мРНК, тРНК, рРНК, антисмысловую РНК, рибозим, структурную РНК или любой другой тип РНК) или белок, продуцируемый посредством трансляции мРНК. Генные продукты также включают РНК, которые модифицируются, посредством процессов, таких как кэпирование, полиаденилирование, метилирование и редактирование, и белки, модифицируемые посредством, например, метилирования, ацетилирования, фосфорилирования, убиквитинирования, АДФ-рибозилирования, миристилирования и гликозилирования.
[0089] "Модуляция" экспрессии гена относится к изменению активности гена. Модуляция экспрессии может включать, но не ограничивается перечисленным, активацию гена и репрессию гена. Редактирование генома (например, расщепление, изменение, инактивация, случайная мутация) могут применяться для модуляции экспрессии. Инактивация гена относится к любому снижению экспрессии гена в сравнении с клеткой, которая не включает белки ZFP или TALE, описанные в данном документе. Таким образом, инактивация гена может быть частичной или полной.
[0090] "Область интереса" представляет собой любую область клеточного хроматина, такую как, например, ген или некодирующую последовательность внутри гена или примыкающую к гену, в которой желательно связать экзогенную молекулу. Связывание может служить для целей целенаправленного расщепления ДНК и/или целенаправленной рекомбинации. Область интереса может присутствовать в хромосоме, эписоме, геномы органелл (например, митохондриальном, хлоропластном), или, например, геноме инфицирующего вируса. Область интереса может находиться внутри кодирующей области гена, внутри транскрибируемых некодирующих областей, таких как, например, лидерные последовательности, трейлерные последовательности или интроны, или внутри нетранскрибируемых областей, находящихся либо против хода транскрипции или по ходу транскрипции от кодирующей области. Область интереса может быть настолько малой, как одиночная пара нуклеотидов или до 2000 пар нуклеотидов длиной, или любое целочисленное значение пар нуклеотидов.
[0091] "Эукариотные" клетки включают, но не ограничиваются перечисленным, клетки грибков (такие как дрожжи), клетки растений, клетки животных, клетки млекопитающих и человеческие клетки (например, T-клетки).
[0092] Термины "функциональная связка" и "функционально связанные" (или “функционально связан”) применяют взаимозаменяемо со ссылкой на соприкосновение двух или нескольких компонентов (таких как элементы последовательности), при котором компоненты располагаются таким образом, что оба компонента функционируют нормально и обеспечивают возможность того, что, по меньшей мере, один из компонентов может опосредовать функцию, которая проявляется в присутствии, по меньшей мере, одного из других компонентов. Для иллюстрации, транскрипционная регуляторная последовательность, такая как промотор, является функционально связанной с кодирующей последовательностью, если транскрипционная регуляторная последовательность управляет уровнем транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или нескольких транскрипционных регуляторных факторов. Транскрипционная регуляторная последовательность обычно является функционально связанной в цис-конфигурации с кодирующей последовательностью, но не обязательно непосредственно примыкает к ней. Например, энхансер представляет собой транскрипционную регуляторную последовательность, которая является функционально связанной с кодирующей последовательностью, даже, несмотря на то, что они не являются смежными.
[0093] По отношению к гибридным полипептидам, термин "функционально связанные" может относиться к тому факту, что каждый из компонентов выполняет одинаковую функцию в связке с другим компонентом, как если бы они не были связаны таким образом. Например, по отношению к гибридному полипептиду, в котором ДНК-связывающий домен ZFP или TALE слит с доменом активации, ДНК-связывающий домен ZFP или TALE и домен активации находятся в функциональной связке, если, в гибридном полипептиде, часть ДНК-связывающего домена ZFP или TALE способна связываться с ее сайтом-мишенью и/или ее сайтом связывания, в то время как домен активации способен регулировать с увеличением экспрессию гена. ZFP, слитые с доменами, способными к регуляции экспрессии гена обобщенно называют “ZFP-TF” или “цинковые пальцы факторы транскрипции”, в то время как TALE, слитые с доменами, способными к регуляции экспрессии гена обобщенно именуют “TALE-TF” или “TALE факторы транскрипции”. Когда гибридный полипептид, в котором ДНК-связывающий домен ZFP слит с доменом расщепления (“ZFN” или “нуклеаза цинковые пальцы”), ДНК-связывающий домен ZFP и домен расщепления находятся в функциональной связке, если, в гибридном полипептиде, часть ДНК-связывающего домена ZFP способна связываться с ее сайтом-мишенью и/или ее сайтом связывания, в то время как домен расщепления способен расщеплять ДНК вблизи от сайта-мишени. Когда гибридный полипептид, в котором ДНК-связывающий домен TALE слит с доменом расщепления (“TALEN” или “TALE нуклеаза”), ДНК-связывающий домен TALE и домен расщепления находятся в функциональной связке, если, в гибридном полипептиде, часть ДНК-связывающего домена TALE способна связываться с ее сайтом-мишенью и/или ее сайтом связывания, в то время как домен расщепления способен расщеплять ДНК вблизи от сайта-мишени.
[0094] "Функциональный фрагмент" белка, полипептида или нуклеиновая кислота представляют собой белок, полипептид или нуклеиновую кислоту, последовательность которых не является идентичной с непроцессированным белком, полипептидом или нуклеиновой кислотой, но все еще сохраняет те же самые функции, как непроцессированный белок, полипептид или нуклеиновая кислота. Функциональный фрагмент может обладать большим, меньшим, или одинаковым числом остатков, как и соответствующая нативная молекула, и/или может содержать одну или несколько аминокислотных или нуклеотидных замен. Методы определения функции нуклеиновой кислоты (например, кодирующей функции, способности к гибридизации с еще одной нуклеиновой кислотой) являются хорошо известными в данной области. Аналогично, методы определения функции белка являются хорошо известными. Например, ДНК-связывающая функция полипептида может быть определена, например, посредством связывания на фильтрах, изменения электрофоретической подвижности, или анализов иммунопреципитации. Расщепление ДНК может быть проанализировано посредством гель-электрофореза. См., Ausubel et al., выше. Способность белка взаимодействовать с еще одним другим белком может быть определена, например, посредством со-иммунопреципитации, двухгибридных анализов или комплементации, как генетической, так и биохимической. См, например, Fields et al. (1989) Nature 340:245-246; Патент США № 5,585,245 и PCT WO 98/44350.
[0095] "Вектор" обладает способностью к переносу генной последовательности к клеткам-мишеням. Обычно, "векторная конструкция", "вектор экспрессии" и "вектор переноса генов" обозначает любую конструкцию нуклеиновой кислоты, способную к направлению экспрессии гена, представляющего интерес, и которая может переносить генные последовательности к клеткам-мишеням. Таким образом, термин включает несущие среды для клонирования и экспрессии, а также интегрирующие векторы.
[0096] "Репортерный ген" или "репортерная последовательность" относится к любой последовательности, которая продуцирует белковый продукт, который легко измеряется, предпочтительно, хотя необязательно в рутинном аналитическом тесте. Подходящие репортерные гены включают, но не ограничиваются перечисленным, последовательности, кодирующие белки, которые опосредуют резистентность к антибиотикам (например, резистентность к ампициллину, резистентность к неомицину, резистентность к G418, резистентность к пуромицину), последовательности, кодирующие окрашенные или флуоресцентные или люминесцентные белки (например, зеленый флуоресцентный белок, улучшенный зеленый флуоресцентный белок, красный флуоресцентный белок, люциферазу), и белки, которые опосредуют увеличенные рост клеток и/или амплификацию генов (например, дигидрофолатредуктазу). Эпитопные метки включают, например, одну или несколько копий FLAG, His, myc, Tap, HA или любой обнаруживаемой аминокислотной последовательности. “Метки экспрессии” включают последовательности, которые кодируют репортеры, которые могут быть функционально связаны с желательной генной последовательностью, чтобы регистрировать экспрессию гена, представляющего интерес.
Биомаркеры болезни Хантингтона
[0097] Срединные шипиковые нейроны являются в высокой степени подверженными БХ. MSN, которые содержат энкефалин, и, которые проецируются на внешний бледный шар (в так называемом “непрямом пути”) являются вовлеченными в большей степени, чем нейроны, которые содержат вещество P и проецируются на внутренний бледный шар (в “прямом пути”), однако поражаются оба типа MSN. MSN при БХ проявляют аномальные агрегацию и включения htt, биоэнергетические дефекты, недостаток нейротрофина, транскрипционную дисрегуляцию, нарушения аксонального транспорта и эксайтотоксичность.
[0098] Например, DARPP-32, Дофамин- и -цАМФ-регулируемый фосфопротеин, ММ 32 кДа) является основной мишенью для дофамина и протеинкиназы A в полосатом теле, который действует как белок с двойной функцией в зависимости от состояния и положения фосфорилирования, в том, что он может действовать как ингибитор белка фосфатазы 1 (PP-1), когда DARP-32 фосфорилируется по Thr34, и как ингибитор PKA, когда фосфорилируется по Thr75 (Svenningsson et al, (2004) Annu Rev Pharmacol Toxicol 44:269-96). Он экспрессируется в 97% из MSN, в нескольких кортикальных слоях и в мозжечковых клетках Пуркинье. Однако на мышиных моделях БХ, экспрессия его мРНК понижающее регулируется в MSN в большей степени, чем в кортексе (см., Ehrlich (2012) Neurotherapeutics 9(2): 270-284).
[0099] Стриарно-специфическая циклонуклеотидфосфодиэстераза PDE10A действует в полосатом теле для ограничения сигнального пути циклических нуклеотидов посредством ферментативного гидролиза. Двойной субстрат PDE экспрессируется на высоких уровнях, как в прямых, так и непрямых MSN (см., Kleiman et al, 2011 J of Pharma and Exp Thera 336(1):64-78) и понижающее регулируется при БХ.
[0100] Сигнальный путь дофамина в полосатом теле регулируется отчасти дофаминовыми рецепторами, и непрямые MSN экспрессируют дофаминовые рецепторы D1 и D2 (Drd1 и Drd2, соответственно). D1, экспрессирующий клетки прямого пути, проецируется на черное вещество ретикулярной части, в то время как рецептор D2, экспрессирующий MSN непрямого пути, проецируется на средний бледный шар, и классическая модель функционирования базальных ганглиев предполагает, что способность полосатого тела выбирать дифференциальное действие дофамина при контроле движения обусловлена сегрегацией рецепторов D1 и D2 в две группы MSN (см., Gerfen et al (1990) Science 250:1429-1432). Таким образом, экспрессия обоих генов Drd1 и Drd2 является существенной функцией MSN и дисбаланс или дефект в экспрессии может вести к нарушению моторной функции, наблюдаемому при БХ.
[0101] Другие биомаркеры функции MSN при БХ, которые, по-видимому, повторяют биомаркеры, наблюдаемые в человеческом каудате при БХ, включают снижение PENK (проэнкефалина), RGS4 (регулятора сигнального пути 4 G-белка), фактора транскрипции NGFi-A, Кальций/кальмодулин-зависимой 3ʹ,5ʹ-циклонуклеотидфосфодиэстеразы 1B PDE1B, и CNR1 (каннабиноидного рецептора CB1) (Runne et al, там же). Дополнительно, экспрессию мутантного гена Htt также можно анализировать непосредственно.
[0102] Хватательный рефлекс на мышиных моделях является показателем заболевания. У относительно агрессивной мышиной модели R6/2, человеческий Htt промотор и фрагмент экзона 1, который содержит удлиненный CAG-повтор эктопически экспрессируется (Mangiarini et al. там же). Эти мыши проявляют БХ-подобные поведенческие изменения, а также потерю стриарных срединных шипиковых нейронов (MSNs), которая является отличительной чертой БХ. Возраст смерти обычно составляет между 10 и 13 неделями (несмотря на то, что смерть может происходить раньше или позже), и мыши проявляют прогрессирующий неврологический фенотип. Одним из первых симптомов является дискинезия конечностей, при удержании за хвост. Это прогрессирует в чередующееся сжимание вместе и высвобождение ступней, пока мыши не сожмут их ступни вместе сразу же после того, как их подняли, и не смогут более высвобождать это положение. Таким образом, хватательный рефлекс может быть оценен баллами, как мера степени фенотипического поведения при БХ.
[0103] Фенотипические улучшения при БХ могут быть измерены посредством сравнения обработанных и необработанных животных. Модели можно оценивать, измеряя хватательный рефлекс и изменения биомаркеров после умерщвления. Например, Rodriguez-Lebron et al. (2005, Mol Ther 12(4):618-633 измеряли эффективность действия кшРНК против htt с использованием анализа хватательного рефлекса и изменения экспрессии DARPP-32 и PENK.
ДНК-связывающие домены
[0104] В данном документе описаны композиции, содержащие ДНК-связывающий домен, который специфически связывается с последовательностью-мишенью в любом гене, содержащем тринуклеотидный повтор, включая, но не ограничиваясь приведенным, Htt. Любой ДНК-связывающий домен может применяться в композициях и способах, раскрытых в данном описании.
[0105] В некоторых вариантах осуществления, ДНК-связывающий домен содержит белок типа “цинковые пальцы”. Предпочтительно, белок “цинковые пальцы” является неприродным в том, что он сконструирован для связывания с выбранным сайтом-мишенью. См, например, Beerli et al. (2002) Nature Biotechnol. 20:135-141; Pabo et al. (2001) Ann. Rev. Biochem. 70:313-340; Isalan et al. (2001) Nature Biotechnol. 19:656-660; Segal et al. (2001) Curr. Opin. Biotechnol. 12:632-637; Choo et al. (2000) Curr. Opin. Struct. Biol. 10:411-416; Патенты США №№ 6,453,242; 6,534,261; 6,599,692; 6,503,717; 6,689,558; 7,030,215; 6,794,136; 7,067,317; 7,262,054; 7,070,934; 7,361,635; 7,253,273; и Патентные публикации США №№ 2005/0064474; 2007/0218528; 2005/0267061, все полностью включенные в данное описание посредством ссылки.
[0106] Сконструированный домен связывания цинковых пальцев может иметь новую специфичность связывания, в сравнении с природным белком “цинковые пальцы”. Способы конструирования включают, но не ограничиваются перечисленными, обоснованный проект и различные типы отбора. Обоснованный проект включает, например, использование баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и аминокислотные последовательности индивидуальных цинковых пальцев, в которых каждая триплетная или квадруплетная нуклеотидная последовательность является ассоциированной с одной или несколькими аминокислотными последовательностями цинковых пальцев, которые связывают конкретную триплетную или квадруплетную последовательность. См, например, совместные Патенты США 6,453,242 и 6,534,261, полностью включенные в данное описание посредством ссылки.
[0107] Иллюстративные методы отбора, включающие отображение фагов и двухгибридные системы, раскрыты в Патентах США 5,789,538; 5,925,523; 6,007,988; 6,013,453; 6,410,248; 6,140,466; 6,200,759; и 6,242,568; а также WO 98/37186; WO 98/53057; WO 00/27878; WO 01/88197 и GB 2,338,237. В дополнение, увеличение специфичности связывания для доменов, связывающих цинковые пальцы было описано, например, в совместной WO 02/077227.
[0108] В дополнение, как раскрыто в этих и других ссылках, домены цинковых пальцев и/или многопальцевые белки типа “цинковые пальцы” могут быть связаны вместе с использованием любых подходящей линкерных последовательностей, включающих например, линкеры из 5 или более аминокислот по длине. См, также, Патенты США №№ 6,479,626; 6,903,185; и 7,153,949 для иллюстративных линкерных последовательностей из 6 или более аминокислот по длине. Белки, описанные в данном документе, могут включать любую комбинацию подходящих линкеров между индивидуальными цинковыми пальцами белка. В дополнение, увеличение специфичности связывания для доменов, связывающих цинковые пальцы, были описаны, например, в совместной WO 02/077227.
[0109] Отбор сайтов-мишеней; ZFP и методы для дизайна и конструирования гибридных белков (и полинуклеотидов, кодирующих их) известны специалистам в данной области и подробно описаны в Патентах США №№ 6,140,081; 5,789,538; 6,453,242; 6,534,261; 5,925,523; 6,007,988; 6,013,453; 6,200,759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970 WO 01/88197; WO 02/099084; WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[0110] В дополнение, как раскрыто в этих и других ссылках, домены цинковых пальцев и/или многопальцевые белки типа “цинковые пальцы” могут быть связаны вместе с использованием любых подходящих линкерных последовательностей, включающих например, линкеры из 5 или более аминокислот по длине. См. также, Патенты США №№ 6,479,626; 6,903,185; и 7,153,949 для иллюстративных линкерных последовательностей с 6 или более аминокислотами по длине. Белки, описанные в данном документе, могут включать любую комбинацию подходящих линкеров между индивидуальными цинковыми пальцами белка.
[0111] В некоторых вариантах осуществления, ДНК-связывающий домен представляет собой сконструированный белок “цинковые пальцы”, который связывается (специфичным для последовательности образом) с сайтом-мишенью в гене Htt и модулирует экспрессию Htt. ZFP могут селективно связываться либо с мутантным аллелем Htt или последовательностью Htt дикого типа. Сайты-мишени Htt обычно включают, по меньшей мере, один цинковый палец, но могут включать множество цинковых пальцев (например, 2, 3, 4, 5, 6 или более пальцев). Обычно, ZFP включают, по меньшей мере, три пальца. Некоторые из ZFP включают четыре, пять или шесть пальцев, в то время как несколько ZFP включают 8, 9, 10, 11 или 12 пальцев. ZFP, которые включают три пальца, обычно распознают сайт-мишень, который включает 9 или 10 нуклеотидов; ZFP, которые включают четыре пальца, обычно распознают сайт-мишень, который включает от 12 до 14 нуклеотидов; в то время как ZFP, имеющий шесть пальцев может распознавать сайты-мишени, которые включают от 18 до 21 нуклеотидов. ZFP могут также представлять собой гибридные белки, которые включают один или несколько регуляторных доменов, которые могут быть доменами транскрипционной активации или доменами репрессии. В нескольких вариантах осуществления, гибридный белок содержит два ДНК-связывающих домена ZFP, связанных вместе. Такие белки “цинковые пальцы” могут, таким образом, содержать 8, 9, 10, 11, 12 или более пальцев. В нескольких вариантах осуществления, два ДНК-связывающих домена связаны через удлиняемый гибкий линкер таким образом, что один ДНК-связывающий домен содержит 4, 5, или 6 цинковых пальцев, а второй ДНК-связывающий домен содержит дополнительные 4, 5, или 5 цинковых пальцев. В нескольких вариантах осуществления, линкер представляет собой стандартный межпальцевый линкер, таким образом, что пальцевая матрица содержит один ДНК-связывающий домен, содержащий 8, 9, 10, 11 или 12 или более пальцев. В других вариантах осуществления, линкер представляет собой атипичный линкер, такой как гибкий линкер. ДНК-связывающие домены сливают по меньшей мере с одним регуляторным доменом и могут рассматриваться как архитектура ʹZFP-ZFP-TFʹ. Конкретные примеры этих вариантов осуществления могут именоваться “ZFP-ZFP-KOX”, которые содержат два ДНК-связывающие домены, связанные гибким линкером, и слитые с репрессором KOX, и “ZFP-KOX-ZFP-KOX”, где два гибридных белка ZFP-KOX слиты вместе через линкер.
[0112] Альтернативно, ДНК-связывающий домен может являться производным от нуклеазы. Например, известны последовательности распознавания хоуминг-эндонуклеаз и мегануклеаз, таких как I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. См. также Патент США № 5,420,032; Патент США № 6,833,252; Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble et al. (1996) J. Mol. Biol. 263:163-180; Argast et al. (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs. В дополнение, ДНК-связывающая специфичность хоуминг-эндонуклеаз и мегануклеаз может быть сконструирована для связывания с неприродными сайтами-мишенями. См, например, Chevalier et al. (2002) Molec. Cell 10:895-905; Epinat et al. (2003) Nucleic Acids Res. 31:2952-2962; Ashworth et al. (2006) Nature 441:656-659; Paques et al. (2007) Current Gene Therapy 7:49-66; Патентную публикацию США № 20070117128.
[0113] “Двурукие” белки типа “цинковые пальцы” представляют собой такие белки, в которых два кластера ДНК-связывающих доменов цинковые пальцы разделены промежуточными аминокислотами таким образом, что два домена цинковые пальцы связываются с двумя прерывистыми сайтами-мишенями. Примером двурукого типа связывающего белка “цинковые пальцы” является SIP1, где кластер из четырех цинковых пальцев расположен по амино-концу белка, а кластер из трех пальцев расположен по карбоксильному концу (см., Remacle et al., (1999) EMBO Journal 18 (18): 5073-5084). Каждый кластер цинковых пальцев в этих белках способен связываться с уникальной последовательностью-мишенью и пространство между двумя последовательностями-мишенями может содержать много нуклеотидов. Двурукие ZFP могут включать функциональный домен, например, слитый с одним или обоими ZFP. Таким образом, будет очевидно, что функциональный домен может быть присоединен к наружной области одного или обоих ZFP (см. Фиг. 1C) или может быть расположен между ZFP (присоединен к обоим ZFP) (см. Фиг. 4).
[0114] Конкретные примеры Htt-нацеленных ZFP раскрыты в Таблицах 1A и 1B. Первый столбец в данной таблице показывает внутреннее ссылочное название (номер) для ZFP и соответствует тому же названию в столбце 1 Таблиц 2A и 2B. “F” относится к пальцам, и число после “F” показывает, какой цинковый палец (например, “F1” относится к пальцу 1).
[0115] Последовательность и положение для сайтов-мишеней этих белков раскрыты в Таблицах 2A и 2B. Таблицы 2A и 2B показывают последовательности-мишени для указанных белков “цинковые пальцы”. Нуклеотиды в сайте-мишени, которые контактируют посредством спиралей распознавания указаны заглавными буквами; неконтактирующие нуклеотиды указаны строчными буквами.
[0116] В некоторых вариантах осуществления, ДНК-связывающий домен содержит природный или сконструированный (неприродный) ДНК-связывающий домен TAL эффектора (TALE). См. например, Патент США № 8,586,526, полностью включенный в данный документ посредством ссылки. Известно, что фитопатогенные бактерии рода Xanthomonas вызывают многие болезни у важных сельскохозяйственных культур. Патогенность Xanthomonas зависит от системы секреции консервативного типа III (T3S), которая вводит более 25 различных эффекторных белков в растительную клетку. Среди этих вводимых белков присутствуют эффекторы, подобные активатору транскрипции (TALE), которые имитируют растительные активаторы транскрипции и манипулируют транскриптом растения (см., Kay et al (2007) Science 318:648-651). Эти белки содержат ДНК-связывающий домен и домен транскрипционной активации. Одним из наиболее хорошо характеризованных TALE является AvrBs3 из Xanthomonas campestgris pv. Vesicatoria (см. Bonas et al (1989) Mol Gen Genet 218: 127-136 и WO2010079430). TALE содержат централизованный домен из тандемных повторов, причем каждый повтор содержит приблизительно 34 аминокислот, которые являются ключом к ДНК-связывающей специфичности этих белков. В дополнение, они содержат последовательность ядерной локализации и кислотный транскрипционный домен активации (см. обзор Schornack S, et al. (2006) J Plant Physiol 163(3): 256-272). В дополнение, у фитопатогенных бактерий Ralstonia solanacearum были обнаружены два гена, обозначенные brg11 и hpx17, которые являются гомологичными с семейством AvrBs3 Xanthomonas у R. solanacearum биовара 1 штамма GMI1000 и у биовара 4 штамма RS1000 (См., Heuer et al (2007) Appl и Envir Micro 73(13): 4379-4384). Эти гены являются на 98,9% идентичными по нуклеотидной последовательности друг с другом, но отличаются по делеции 1575 пар оснований в домене повтора из hpx17. Однако оба генных продукта имеет меньшую, чем 40%, идентичность последовательности с белками из Xanthomonas семейства AvrBs3.
[0117] Специфичность этих TALE зависит от последовательностей, обнаруженных в тандемных повторах. Повторяющаяся последовательность содержит приблизительно 102 пары оснований, и повторы обычно имеют 91-100% гомологию друг с другом (Bonas et al., там же). Полиморфизм повторов обычно находится в положениях 12 и 13, и, по-видимому, существует соответствие один-в-один между идентичностью гипервариабельных диостатков в положениях 12 и 13 и идентичностью смежных нуклеотидов в последовательности-мишени TALE (см., Moscou и Bogdanove, (2009) Science 326:1501 и Boch et al (2009) Science 326:1509-1512). Экспериментaльно был определен код для распознавания ДНК этих TALE, таким образом, что HD последовательность в положениях 12 и 13 приводит к связыванию с цитозином (C), NG связывается с T, NI с A, C, G или T, NN связывается с A или G, и IG связывается с T. Эти ДНК-связывающие повторы были собраны в белки с новыми комбинациями и числами повторов, чтобы создать искусственные факторы транскрипции, которые способны взаимодействовать с новыми последовательностями и активировать экспрессию неэндогенного репортерного гена в растительных клетках (Boch et al., там же). Сконструированные белки TAL были связаны с половинным доменом расщепления FokI с получением гибрида домена TAL эффектора и нуклеазы (TALEN), проявляющего активность в аналитическом тесте с дрожжевым репортером (мишенью на основе плазмиды). Christian et al ((2010)< Genetics epub 10.1534/genetics.110.120717). См. также Патент США № 8,586,526, полностью включенный в данный документ посредством ссылки.
[0118] Конкретные примеры сконструированных доменов димеризации для применения вместе с ZFP или белками TALE перечислены в Таблице 3. Приведены аминокислотные последовательности двух типов доменов, суперспирального домена (CC) и домена с димеризующими цинковыми пальцами (DZ).
Гибридные белки
[0119] Также предоставлены гибридные белки, содержащие ДНК-связывающие белки (например, ZFP или TALE), описанные в данном документе, и гетерологичный регуляторный (функциональный) домен (или его функциональный фрагмент). Общеизвестные домены включают, например, домены факторов транскрипции (активаторов, репрессоров, соактиваторов, сорепрессоров), сайленсеры, онкогены (например, члены семейств myc, jun, fos, myb, max, mad, rel, ets, bcl, myb, mos и т.д.); ферменты репарации ДНК и их ассоциированные факторы и модификаторы; ферменты пергруппировки ДНК и их ассоциированные факторы и модификаторы; белки, ассоциированные с хроматином и их модификаторы (например, киназы, ацетилазы и деацетилазы); и ферменты, модифицирующие ДНК, (например, метилтрансферазы, топоизомеразы, геликазы, лигазы, киназы, фосфатазы, полимерaзы, эндонуклеазы) и их ассоциированные факторы и модификаторы. Патенты США №№ 7,888,121 и 8,409,861, подробно касающиеся гибридов ДНК-связывающих доменов и доменов нуклеазного расщепления, полностью включены в данный документ посредством ссылки.
[0120] Подходящие домены для достижения активации включают домен активации HSV VP16 (см., например, Hagmann et al., J. Virol. 71, 5952-5962 (1997)) ядерные гормональные рецепторы (см., например, Torchia et al., Curr. Opin. Cell. Biol. 10:373-383 (1998)); субъединицу p65 ядерного фактора каппа B (Bitko & Barik, J. Virol. 72:5610-5618 (1998) и Doyle & Hunt, Neuroreport 8:2937-2942 (1997)); Liu et al., Cancer Gene Ther. 5:3-28 (1998)), или искусственные химерные функциональные домены, такие как VP64 (Beerli et al., (1998) Proc. Natl. Acad. Sci. USA 95:14623-33), и дегрон (Molinari et al., (1999) EMBO J. 18, 6439-6447). Дополнительные иллюстративные домены активации включают, Oct 1, Oct-2A, Sp1, AP-2 и CTF1 (Seipel et al., EMBO J. 11, 4961-4968 (1992) а также p300, CBP, PCAF, SRC1 PvALF, AtБХ2A и ERF-2. См., например, Robyr et al. (2000) Mol. Endocrinol. 14:329-347; Collingwood et al. (1999) J. Mol. Endocrinol. 23:255-275; Leo et al. (2000) Gene 245:1-11; Manteuffel-Cymborowska (1999) Acta Biochim. Pol. 46:77-89; McKenna et al. (1999) J. Steroid Biochem. Mol. Biol. 69:3-12; Malik et al. (2000) Trends Biochem. Sci. 25:277-283; и Lemon et al. (1999) Curr. Opin. Genet. Dev. 9:499-504. Дополнительные иллюстративные домены активации включают, но не ограничиваются перечисленными, OsGAI, HALF-1, C1, AP1, ARF-5,-6,-7 и -8, CPRF1, CPRF4, MYC-RP/GP, и TRAB1. См., например, Ogawa et al. (2000) Gene 245:21-29; Okanami et al. (1996) Genes Cells 1:87-99; Goff et al. (1991) Genes Dev. 5:298-309; Cho et al. (1999) Plant Mol. Biol. 40:419-429; Ulmason et al. (1999) Proc. Natl. Acad. Sci. USA 96:5844-5849; Sprenger-Haussels et al. (2000) Plant J. 22:1-8; Gong et al. (1999) Plant Mol. Biol. 41:33-44; и Hobo et al. (1999) Proc. Natl. Acad. Sci. USA 96:15,348-15,353.
[0121] Специалистам в данной области будет ясно, что при образовании гибридного белка (или нуклеиновой кислоты, кодирующей его) между ДНК-связывающим доменом и функциональным доменом, либо домен активации или молекула, которая взаимодействует с доменом активации, является подходящей в качестве функционального домена. По существу, любая молекула, способная к мобилизации активирующего комплекса и/или активирующей активности (такой как, например, ацетилирование гистонов) к гену-мишени, является применимой в качестве активирующего домена гибридного белка. Инсуляторные домены, домены локализации и белки реконструкции хроматина, такие как ISWI-содержащие домены и/или метил-связывающие доменные белки, подходящие для применения в качестве функциональных доменах в гибридных молекулах, описаны, например, в Патентах США №№ 6,919,204 и 7,053,264.
[0122] Иллюстративные домены репрессии включают, но не ограничиваются перечисленным, KRAB A/B, KOX, TGF-бета-индуцируемый ранним геном (TIEG), v-erbA, SID, MBD2, MBD3, члены семейства DNMT (например, DNMT1, DNMT3A, DNMT3B), Rb, и MeCP2. См. например, Bird et al. (1999) Cell 99:451-454; Tyler et al. (1999) Cell 99:443-446; Knoepfler et al. (1999) Cell 99:447-450; и Robertson et al. (2000) Nature Genet. 25:338-342. Дополнительные иллюстративные домены репрессии включают, но не ограничиваются перечисленными, ROM2 и AtБХ2A. См., например, Chem et al. (1996) Plant Cell 8:305-321; и Wu et al. (2000) Plant J. 22:19-27.
[0123] Гибридные молекулы конструируют методами клонирования и биохимического конъюгирования, которые являются хорошо известными специалистам в данной области. Гибридные молекулы содержат ДНК-связывающий домен и функциональный домен (например, домен активации транскрипции или домен репрессии). Гибридные молекулы также необязательно содержат сигналы ядерной локализации (такие как, например, сигналы из среды SV40 T-антигена) и эпитопные метки (такие как, например, FLAG и гемагглютинин). Гибридные белки (и нуклеиновые кислоты, кодирующие их) конструируют таким образом, что трансляционная рамка считыванияe сохраняется среди компонентов гибрида.
[0124] Гибриды между полипептидным компонентом функционального домена (или его функционального фрагмента) с одной сторны, и небелковым ДНК-связывающим доменом (например, антибиотиком, интеркалятором, белком, связывающимся с малой бороздкой, нуклеиновой кислотой) с другой, конструируют методами биохимического конъюгирования, известными специалистам в данной области. См., например, каталог Pierce Chemical Company (Rockford, IL). Способы и композиции для получения гибридов между белком, связывающимся с малой бороздкой, и полипептидом были описаны. Mapp et al. (2000) Proc. Natl. Acad. Sci. USA 97:3930-3935.
[0125] В некоторых вариантах осуществления, сайт-мишень, связанный посредством ДНК-связывающего домена, присутствует в доступной области клеточного хроматина. Доступные области могут быть определены, как описано, например, в совместной Международной Публикации WO 01/83732. Если сайт-мишень не присутствует в доступной области клеточного хроматина, одна или более доступных областей могут генерироваться, как описано в совместной WO 01/83793. В дополнительных вариантах осуществления, ДНК-связывающий домен гибридной молекулы способен к связыванию с клеточным хроматином независимо от того, находится ли сайт-мишень в доступной области или нет. Например, такие ДНК-связывающие домены являются способными к связыванию с линкерной ДНК и/или нуклеосомальной ДНК. Примеры этого типа "пионерского" ДНК-связывающего домена обнаруживаются в рецепторах некоторых стероидов и в гепатоцитарном ядерном факторе 3 (HNF3). Cordingley et al. (1987) Cell 48:261-270; Pina et al. (1990) Cell 60:719-731; и Cirillo et al. (1998) EMBO J. 17:244-254.
[0126] Гибридная молекула может быть составлена вместе с фармацевтически приемлемым носителем, как известно специалистам в данной области. См., например, Remingtonʹs Pharmaceutical Sciences, 17th ed., 1985; и совместную WO 00/42219.
[0127] Функциональный компонент/домен гибридной молекулы может быть отобран из любого разнообразия различных компонентов, способных к влиянию на транскрипцию гена, как только гибридная молекула связывается с целевой последовательностью через ее ДНК-связывающий домен. Следовательно, функциональный компонент может включать, но не ограничивается перечисленным, различные домены факторов транскрипции, таких как активаторы, репрессоры, соактиваторы, сорепрессоры и сайленсеры.
[0128] Дополнительные иллюстративные функциональные домены раскрыты, например, в совместных Патентах США №№ 6,534,261 и 6,933,113.
[0129] Функциональные домены, которые регулируются экзогенными малыми молекулами или лигандами, могут также быть отобраны. Например, может использоваться технология RheoSwitch®, где функциональный домен принимает свою активную конформацию только в присутствии внешнего лиганда RheoChem™ (см., например US 20090136465). Таким образом, ZFP или TALE могут быть функционально связаны с регулируемым функциональным доменом, где результирующая активность ZFP-TF или TALE-TF управляется внешним лигандом.
Нуклеазы
[0130] Также в данном документе описаны нуклеазы, такие как ZFN, TALEN, хоуминг-эндонуклеазы, CRISPR/Cas и/или Ttago РНК-направляемые системы, которые являются применимыми для генетической модификации in vivo, включающей вставки и/или делеции, сделанные после расщепления мишени нуклеазой (нуклеазами). В некоторых вариантах осуществления, гибридный белок содержит ДНК-связывающий домен и домен расщепления (нуклеазы). Сама по себе, генная модификация может достигаться с использованием нуклеаз, например сконструированных нуклеаз. Технология сконструированных нуклеаз основана на конструировании природных ДНК-связывающих белков. Например, было описано конструирование хоуминг-эндонуклеаз с привязанными ДНК-связывающими специфичностями (см, Chames et al. (2005) Nucleic Acids Res 33(20):e178; Arnould et al. (2006) J. Mol. Biol. 355:443-458). В дополнение, конструирование ZFP также было описано. См., например, Патенты США №№ 8,586,526; 6,534,261; 6,607,882; 6,824,978; 6,979,539; 6,933,113; 7,163,824; и 7,013,219.
[0131] В дополнение, ZFP и TALE были слиты с доменами нуклеаз для создания ZFN- и TALEN-функциональных молекул, которые способны распознавать их предназначенную мишень нуклеиновой кислоты посредством их сконструированных (ZFP или TALE) ДНК-связывающих доменов и вызывать разрезание ДНК рядом с ДНК-связывающим сайтом для ZFP или TALE посредством нуклеазной активности. См., например, Kim et al. (1996) Proc Natl Acad Sci USA 93(3):1156-1160. Нуклеазы (например, ZFN, TALEN, системы CRISPR/Cas и/или Ttago) применяли для модификации генома у различных организмов. Патенты США №№ 7,888,121; 7,972,854; 7,914,796; 7,951,925; 8,110,379; 8,409,861; 8,586,526; Патентные публикации США 20030232410; 20050208489; 20050026157; 20060063231; 20080159996; 201000218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983 и 20130177960 и Заявка США № 14/278,903.
[0132] Таким образом, способы и композиции, описанные в данном документе, имеют широкое применение и могут включать любую нуклеазу, представляющую интерес. Неограничивающие примеры нуклеаз включают системы CRISPR/Cas и/или Ttago, мегануклеазы, TALEN и нуклеазы “цинковые пальцы”. Нуклеаза может содержать гетерологичные ДНК-связывающий домен и домен расщепления (например, нуклеазы “цинковые пальцы”; TALEN; мегануклеазные ДНК-связывающие домены с гетерологичными доменами расщепления) или, альтернативно, ДНК-связывающий домен природной нуклеазы может быть изменен для связывания с выбранным сайтом-мишенью (например., мегануклеаза которая была сконструирована для связыванию с сайтой, отличным от когнатного сайта связывания).
[0133] В некоторых вариантах осуществления, нуклеаза является мегануклеазой (хоуминг-эндонуклеазой). Природные мегануклеазы распознают сайты расщепления, имеющие 15-40 пар оснований, и их обычно группируют в четыре семейства: семейство LAGLIDADG, семейство GIY-YIG, семейство His-Cyst коробочки и семейство HNH. Иллюстративные хоуминг-эндонуклеазы включают I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. Их последовательности распознавания являются известными. См., также Патент США № 5,420,032; Патент США № 6,833,252; Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388; Dujon et al. (1989) Gene 82:115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble et al. (1996) J. Mol. Biol. 263:163-180; Argast et al. (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs.
[0134] ДНК-связывающие домены из природных мегануклеаз, прежде всего из семейства LAGLIDADG, применяли для стимуляции сайт-специфичной геномной модификации у растений, дрожжей, Drosophila, клеток млекопитающих и мышей, но этот подход был ограничен модификацией любых гомологичных генов, которые сохраняют последовательность распознавания мегануклеазы (Monet et al. (1999), Biochem. Biophysics. Res. Common. 255: 88-93) или предварительно сконструированными геномами, в которую была введена последовательность распознавания (Route et al. (1994), Mol. Cell. Biol. 14:8096-106; Chilton et al. (2003) Plant Physiology 133:956-65; Puchta et al. (1996), Proc. Natl. Acad. Sci. USA 93: 5055-60; Rong et al. (2002), Genes Dev. 16: 1568-81; Gouble et al. (2006), J. Gene Med. 8(5):616-622). Соответственно, были сделаны попытки сконструировать мегануклеазы для проявления новой специфичности связывания в сайтах, применимых в медицинском отношении или биотехнологически (Porteus et al. (2005), Nat. Biotechnol. 23: 967-73; Sussman et al. (2004), J. Mol. Biol. 342: 31-41; Epinat et al. (2003), Nucleic Acids Res. 31: 2952-62; Chevalier et al. (2002) Molec. Cell 10:895-905; Epinat et al. (2003) Nucleic Acids Res. 31:2952-2962; Ashworth et al. (2006) Nature 441:656-659; Paques et al. (2007) Current Генн терапи 7:49-66; Патентные публикации США №№ 20070117128; 20060206949; 20060153826; 20060078552; и 20040002092). В дополнение, природные или сконструированные ДНК-связывающие домены из мегануклеаз также были функционально связаны с доменом расщепления из гетерологичной нуклеазы (например, FokI).
[0135] В других вариантах осуществления, нуклеаза является нуклеазой “цинковые пальцы” (ZFN). ZFN содержит белок “цинковые пальцы”, который был сконструирован для связывания с сайтом-мишенью в гене выбора, и домен расщепления или половинный домен расщепления.
[0136] Как подробно описано выше, могут быть сконструированы домены связывания с цинковыми пальцами для связывания с последовательностью выбора. См., например, Beerli et al. (2002) Nature Biotechnol. 20:135-141; Pabo et al. (2001) Ann. Rev. Biochem. 70:313-340; Isalan et al. (2001) Nature Biotechnol. 19:656-660; Segal et al. (2001) Curr. Opin. Biotechnol. 12:632-637; Choo et al. (2000) Curr. Opin. Struct. Biol. 10:411-416. Сконструированный домен связывания “цинковые пальцы” может иметь новую специфичность связывания, в сравнении с природным белком “цинковые пальцы”. Методы конструирования включают, но не ограничиваются перечисленным, обоснованный проект и различные типы отбора. Обоснованный проект включает, например, использование баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и индивидуальные аминокислотные последовательности цинковых пальцев, в которых каждая триплетная или квадруплетная нуклеотидная последовательность является ассоциированной с одной или несколькими аминокислотными последовательностями цинковых пальцев, которые связывают конкретную триплетную или квадруплетную последовательность. См., например, совместные Патенты США 6,453,242 и 6,534,261, полностью включенные в данное описание посредством ссылки.
[0137] Иллюстративные методы отбора, включающие фаговое отображения и двухгибридные системы, раскрыты в Патентах США 5,789,538; 5,925,523; 6,007,988; 6,013,453; 6,410,248; 6,140,466; 6,200,759; и 6,242,568; а также WO 98/37186; WO 98/53057; WO 00/27878; WO 01/88197 и GB 2,338,237. В дополнение, увеличение специфичности связывания для связывающих доменов с цинковыми пальцами было описано, например, в совместной WO 02/077227.
[0138] В дополнение, как раскрыто в этих и других ссылках, домены цинковых пальцев и/или многопальцeвые белки “цинковые пальцы” могут быть связаны вместе с использованием любых подходящих линкерных последовательностей, включающих например, линкеры из 5 или более аминокислот по длине (например, TGEKP (SEQ ID NO:135), TGGQRP (SEQ ID NO:136), TGQKP (SEQ ID NO:137), и/или TGSQKP (SEQ ID NO:138)). См., например, Патенты США №№ 6,479,626; 6,903,185; и 7,153,949 по иллюстративным линкерным последовательностям из 6 или более аминокислот по длине. Белки, описанные в данном документе, могут включать любую комбинацию из подходящих линкеров между индивидуальными цинковыми пальцами белка. См., также, Опубликованную предварительную патентную заявку США № 20110287512.
[0139] CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats)/Cas (CRISPR Associated) нуклеазная система представляет собой недавно сконструированную нуклеазную систему, основанную на бактериальной системе, которая может применяться для конструирования генома. Она основана на части адаптивного иммунного ответа многих бактерий и архебактерий. Когда вирус или плазмида захватывает бактерию, сегменты ДНК завоевателя превращаются в CRISPR РНК (крРНК) посредством ʹиммунногоʹ ответа. Эта крРНК затем ассоциируется, через область частичной комплементарности, с еще одним типом РНК, именуемым тракрРНК, чтобы провести нуклеазу Cas9 к области, гомологичной с крРНК в целевой ДНК, называемой “протоспэйсером.” Cas9 расщепляет ДНК с генерацией “тупых” концов по DSB на сайтам, установленным 20-нуклеотидной проводящей последовательностью, содержащейся внутри транскрипта крРНК. Для Cas9 требуются как крРНК, так и тракрРНК для сайт-специфичного распознавания и расщепления ДНК. Эта система была сконструирована в настоящее время таким образом, что крРНК и тракрРНК могут объединяться в одну молекулу (“единственного РНК-проводника”), и эквивалентная часть крРНК единственного РНК-проводника может быть сконструирована для проведения нуклеазы Cas9, чтобы целенаправленно воздействовать на любую желательную последовательность (см., Jinek et al (2012) Science 337, p. 816-821, Jinek et al, (2013), eLife 2:e00471, и David Segal, (2013) eLife 2:e00563). Таким образом, система CRISPR/Cas может быть сконструирована для создания DSB на желательной мишени в геноме, и на репарацию DSB можно влять посредством применения ингибиторов репарации, чтобы вызвать увеличение ошибочной репарации.
[0140] Как отмечено выше, расщепляющий (нуклеазный) домен нуклеаз может быть гетерологичным с ДНК-связывающим доменом, например ДНК-связывающий доменом с цинковыми пальцами и доменом расщепления из нуклеазного или мегануклеазного ДНК-связывающего домена и доменом расщепления из различных нуклеаз. Гетерологичные домены расщепления могут быть получены из любых эндонуклеаз или экзонуклеаз. Иллюстративные эндонуклеазы, из которых домен расщепления может быть получен, включают, но не ограничиваются перечисленным, эндонуклеазы рестрикции и хоуминг-эндонуклеазы. См., например, Каталог 2002-2003, New England Biolabs, Beverly, MA; и Belfort et al. (1997) Nucleic Acids Res. 25:3379-3388. Известны дополнительные ферменты, которые расщепляют ДНК (например, Нуклеаза S1; нуклеаза из золотистой фасоли; панкреатическая ДНКаза I; микрококковая нуклеаза; дрожжевая эндонуклеаза HO; см., также Linn et al. (eds.) Nycleases, Cold Spring Harbor Laboratory Press, 1993). Один или несколько из этих ферментов (или их функциональных фрагментов) могут применяться в качестве источника доменов расщепления и половинных доменов расщепления.
[0141] Аналогично, половинный домен расщепления может являться производным от любой нуклеазы или ее части, как приведено выше, что требует димеризации для расщепляющей активности. В общем случае, два гибридных белка требуются для расщепления, если гибридные белки содержат половинные домены расщепления. Альтернативно, может применяться единственный белок, содержащий два половинных домена расщепления. Два половинных домена расщепления могут быть получены из одной и той же эндонуклеазы (или ее функциональных фрагментов), или каждый половинный домен расщепления может быть получен из различных эндонуклеаз (или их функциональных фрагментов). В дополнение, сайты-мишени для двух гибридных белков предпочтительно располагаются, по отношению друг к другу, таким образом, что связывание двух гибридных белков с их соответствующими сайтами-мишенями помещает половинные домены расщепления в пространственную ориентацию друг к другу, которая обеспечивает половинным доменам расщепления образование функционального домена расщепления, например, посредством димеризации. Таким образом, в некоторых вариантах осуществления, рядом лежащие края сайтов-мишеней разделены 5-8 нуклеотидами или 15-18 нуклеотидами. Однако любое целое число нуклеотидов или пар нуклеотидов может быть расположено между двумя сайтами-мишенями (например, от 2 до 50 пар нуклеотидов или более). В целом, сайт расщепления лежит между сайтами-мишенями.
[0142] Эндонуклеазы рестрикции (ферменты рестрикции) присутствуют у многих видов и способны к специфичному по отношению к последовательности связыванию с ДНК (на сайте распознавания), и расщепляющей ДНК на сайте связывания или рядом с ним. Некоторые ферменты рестрикции (например, Типа IIS) расщепляют ДНК на сайтах, удаленных от сайта распознавания, и имеют разделяемые домены связывания и расщепления. Например, фермент Fok I Типа IIS катализирует двухцепочечное расщепление ДНК, на расстоянии 9 нуклеотидов от его сайта распознавания на одной цепи и на расстоянии 13 нуклеотидов от его сайта распознавания на другой цепт. См., например, Патенты США 5,356,802; 5,436,150 и 5,487,994; а также Li et al. (1992) Proc. Natl. Acad. Sci. USA 89:4275-4279; Li et al. (1993) Proc. Natl. Acad. Sci. USA 90:2764-2768; Kim et al. (1994a) Proc. Natl. Acad. Sci. USA 91:883-887; Kim et al. (1994b) J. Biol. Chem. 269:31,978-31,982. Таким образом, в одном варианте осуществления, гибридные белки содержат домен расщепления (или половинный домен расщепления) из по меньшей мере одного фермента рестрикции Типа IIS и одного или нескольких доменов связывания цинковых пальцев, которые могут быть сконструированными или несконструированными.
[0143] Иллюстративным ферментом рестрикции Типа IIS, чей домен расщепления является отделяемым от связывающего домена, является Fok I. Данный конкретный фермент является активным в виде димера. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10,570-10,575. Соответственно, для целей настоящего раскрытия, часть фермента Fok I, применяемая в раскрытых гибридных белках, считается половинным доменом расщепления. Таким образом, для целенаправленного двухцепочечного расщепления и/или целенаправленной замены клеточной последовательности с использованием гибридов цинковые пальцы- или TALE-Fok I, два гибридных белка, каждый из которых содержит половинный домен расщепления Fok I, могут применяться для восстановления каталитически активного домена расщепления. Альтернативно, также может применяться одиночная полипептидная молекула, содержащая домен связывания цинковых пальцев и два половинных домена расщепления Fok I. Параметры для целенаправленного расщепления и целенаправленного изменения последовательности с использованием гибридов цинковые пальцы- или TALE-Fok I предоставлены в других разделах в данном раскрытии.
[0144] Домен расщепления или половинный домен расщепления могут являться любой частью белка, которая сохраняет расщепляющую активность, или, которая сохраняет способность к мультимеризации (например, димеризации) с образованием функционального домена расщепления.
[0145] Иллюстративные ферменты рестрикции Типа IIS описаны в Международной Публикации WO 07/014275, полностью включенной в данный документ. Дополнительные ферменты рестрикции также содержат разделяемые домен связывания и домен расщепления, и они предусмотрены настоящим раскрытием. См., например, Roberts et al. (2003) Nucleic Acids Res. 31:418-420.
[0146] В некоторых вариантах осуществления, домен расщепления содержит один или более сконструированный половинный домен расщепления (также именуемые как мутанты домена димеризации), которые минимизируют или предотвращают гомодимеризацию, как описано, например, в Патентах США №№ 8,772,453; 8,623,618; 8,409,861; 8,034,598; 7,914,796; и 7,888,121, раскрытие всех из которых полностью включены в данный документ посредством ссылки. Все аминокислотные остатки в положениях 446, 447, 479, 483, 484, 486, 487, 490, 491, 496, 498, 499, 500, 531, 534, 537, и 538 из Fok I являются мишенями для влияния на димеризацию половинных доменов расщепления Fok I. Иллюстративные сконструированные половинные домены расщепления Fok I, которые образуют облигатные гетеродимеры, включают пару, в которой первый половинный домен расщепления включает мутации по аминокислотным остаткам в положениях 490 и 538 из Fok I, а второй половинный домен расщепления включает мутации по аминокислотным остаткам 486 и 499.
[0147] Таким образом, в одном варианте осуществления, a мутация в 490 заменяет Glu (E) на Lys (K); мутация в 538 заменяет Iso (I) на Lys (K); мутация в 486 заменяет Gln (Q) на Glu (E); и мутация в положении 499 заменяет Iso (I) на Lys (K). Конкретно, сконструированные половинные домены расщепления, описанные в данном документе, получали посредством проведения мутаций в положениях 490 (E→K) и 538 (I→K) в одном половинном домене расщепления с получением сконструированного половинного домена расщепления, обозначенного “E490K:I538K” и посредством проведения мутаций в положениях 486 (Q→E) и 499 (I→L) в другом половинном домене расщепления с получением сконструированного половинного домена расщепления, обозначенного “Q486E:I499L”. Сконструированные половинные домены расщепления, описанные в данном документе, представляют собой облигатные гетеродимерные мутанты, в которых аберрантное расщепление минимизируется или отменяется. См, например, Патенты США №№ 7,914,796 и 8,034,598, раскрытия которых полностью включены в данный документ посредством ссылки для всех целей. В некоторых вариантах осуществления, сконструированный половинный домен расщепления содержит мутации в положениях 486, 499 и 496 (нумерованные относительно Fok I дикого типа), например, мутации, которые заменяют остаток Gln (Q) дикого типа в положении 486 на остаток Glu (E), остаток Iso (I) дикого типа в положении 499 на остаток Leu (L) и остаток Asn (N) дикого типа в положении 496 на остаток Asp (D) или Glu (E) (также называемый доменами “ELD” и “ELE”, соответственно). В других вариантах осуществления, сконструированный половинный домен расщепления содержит мутации в положенииях 490, 538 и 537 (нумерованные относительно Fok I дикого типа), например, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 на остаток Lys (K), остаток Iso (I) дикого типа в положении 538 на остаток Lys (K), и остаток His (H) дикого типа в положении 537 на остаток Lys (K) или остаток Arg (R) (также называемый доменами “KKK” и “KKR”, соответственно). В других вариантах осуществления, сконструированный половинный домен расщепления содержит мутации в положенииях 490 и 537 нумерованные относительно Fok I дикого типа), например, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 на остаток Lys (K) и остаток His (H) дикого типа в положении 537 на остаток Lys (K) или остаток Arg (R) (также называемый доменами “KIK” и “KIR”, соответственно), Патент США № 8,772,453.
[0148] Сконструированные половинные домены расщепления, описанные в данном документе, могут быть получены с использованием любого подходящего способа, например, посредством сайт-направленного мутагенеза половинных доменов расщепления дикого типа (Fok I), как описано в Патентах США №№ 8,772,453; 8,623,618; 8,409,861; 8,034,598; 7,914,796; и 7,888,121.
[0149] Альтернативно, нуклеазы могут быть собраны in vivo на сайте-мишени нуклеиновой кислоты с использованием так называемой “сплит-ферментной” технологии (см., например, Патентную публикацию США № 20090068164). Компоненты таких сплит-ферментов могут экспрессироваться либо на раздельных конструкциях экспрессии, или могут быть связаны в одной открытой рамке считывания, где индивидуальные компоненты разделяются, например, посредством саморасщепляющего пептида 2A или последовательности IRES. Компоненты могут представлять собой индивидуальные домены связывания “цинковых пальцев” или домены связывания нуклеиновой кислоты мегануклеазы.
[0150] В нескольких вариантах осуществления, ДНК-связывающий домен представляет собой сконструированный домен из эффектора TAL, сходный с доменами, полученными из растительных патогенов Xanthomonas (см., Boch et al, (2009) Science 326: 1509-1512 и Moscou and Bogdanove, (2009) Science326: 1501) и Ralstonia (см., Heuer et al (2007) Applied and Environmental Microbiology 73(13): 4379-4384). Также, см. публикацию PCT WO2010/079430.
[0151] Нуклеазы (например, ZFN или TALEN, и т.д.) могут быть подвергнуты скринингу на активность перед применением, например, в хромосомной системе на основе дрожжей, как описано в 8,563,314. Экспрессия нуклеазы может находиться под управлением конститутивного промотора или индуцируемого промотора, например галактокиназного промотора, который активируется (дерепрессируется) в присутствии раффинозы и/или галактозы и репрессируется в присутствии глюкозы.
Доставка
[0152] Белки (например, ZFP, TALE, CRISPR/Cas, Ttago), полинуклеотиды, кодирующие их, и композиции, содержащие белки и/или полинуклеотиды, описанные в данном документе, могут быть доставлены к клетке-мишени посредством любого подходящего средства, включающего, например, инъекцию белков ZFP-TF, TALE-TF или применение мРНК, кодирующей ZFN или TALEN. Подходящие клетки включают, но не ограничены приведенными, эукариотные и прокариотные клетки и/или линии клеток. Неограничивающие примеры таких клеток или линий клеток, генерируемых из таких клеток, включают клетки COS, CHO (например, CHO-S, CHO-K1, CHO-DG44, CHO-DUXB11, CHO-DUKX, CHOK1SV), VERO, MDCK, WI38, V79, B14AF28-G3, BHK, HaK, NS0, SP2/0-Ag14, HeLa, HEK293 (например, HEK293-F, HEK293-H, HEK293-T), и perC6, а также клетки насекомых, таких как Spodoptera fugiperda (Sf), или клетки грибков, таких как Saccharomyces, Pichia и Schizosaccharomyces. В некоторых вариантах осуществления, линия клеток представляет собой линию клеток CHO-K1, MDCK или HEK293. Подходящие клетки также включают стволовые клетки, такие как, посредством примера, эмбриональные стволовые клетки, индуцированные плюрипотентные стволовые клетки, гемопоэтические стволовые клетки, нейрональные стволовые клетки и мезенхимальные стволовые клетки.
[0153] Способы доставки белков, включающих белки “цинковые пальцы”, описанных в данном документе, описаны, например, в Патентах США №№ 6,453,242; 6,503,717; 6,534,261; 6,599,692; 6,607,882; 6,689,558; 6,824,978; 6,933,113; 6,979,539; 7,013,219; и 7,163,824, раскрытие всех из которых полностью включено в данное описание посредством ссылки.
[0154] Белки типа “цинковые пальцы”, TALE или CRISPR/Cas, описанные в данном документе, могут также доставляться с использованием векторов, содержащих последовательности, кодирующие один или несколько белок (белки) “цинковые пальцы”, TALE или CRISPR/Cas. Могут применяться любые векторные системы включая, но не ограничиваясь приведенными, плазмидные векторы, ретровирусные векторы, лентивирусные векторы, аденовирусные векторы, поксвирусные векторы; герпесвирусные векторы и аденоассоциированные вирусные векторы, и т.д. См, также, Патенты США №№ 6,534,261; 6,607,882; 6,824,978; 6,933,113; 6,979,539; 7,013,219; и 7,163,824, полностью включенные в данное описание посредством ссылки. Кроме того, будет понятно, что любой из этиз векторов может содержать одну или несколько последовательностей, кодирующих белки “цинковые пальцы” или TALE. Таким образом, когда один или несколько белков ZFP, TALE или CRISPR/Cas вводят в клетку, последовательности, кодирующие белки ZFP, TALE или CRISPR/Cas, могут переноситься на том же самом векторе или на различных векторах. Когда применяют множество векторов, каждый вектор может содержать последовательность, кодирующую одну или несколько ZFP, TALE или систему CRISPR/Cass.
[0155] Общепринятые методы переноса генов на вирусной и невирусной основе могут применяться для введения нуклеиновых кислот, кодирующих сконструированные ZFP, TALE или систему CRISPR/Cass в клетках (например, клетках млекопитающих) и тканях-мишенях. Такие методы могут также применяться для введения нуклеиновых кислот, кодирующих ZFP, TALE или систему CRISPR/Cas, в клетки in vitro. В некоторых вариантах осуществления, нуклеиновые кислоты, кодирующие ZFP, TALE или систему CRISPR/Cas вводят для применений, связанных с генной терапией in vivo или ex vivo. Невирусные системы доставки векторов включают ДНК-плазмиды, голую нуклеиновую кислоту и нуклеиновую кислоту в комплексе со средой доставки, такой как липосома или полоксамер. Вирусные системы доставки векторов включают ДНК- и РНК-вирусы, которые имеют либо эписомальные или интегрированные геномы после доставки в клетку. Для обзора методик генной терапии, см., Anderson, Science 256:808-813 (1992); Nabel & Felgner, TIBTECH 11:211-217 (1993); Mitani & Caskey, TIBTECH 11:162-166 (1993); Dillon, TIBTECH 11:167-175 (1993); Miller, Nature 357:455-460 (1992); Van Brunt, Biotechnology 6(10):1149-1154 (1988); Vigne, Restorative Neurology and Neuroscience 8:35-36 (1995); Kremer & Perricaudet, British Medical Bulletin 51(1):31-44 (1995); Haddada et al., in Current Topics in Microbiology and Immunology Doerfler и Böhm (eds.) (1995); и Yu et al., Генн терапи 1:13-26 (1994).
[0156] Методы невирусной доставки нуклеиновых кислот включают электропорацию, липофекцию, микроинъекцию, биолистирование, виросомы, липосомы, иммунолипосомы, поликатионные конъюгаты или конъюгаты липид:нуклеиновая кислота, оголенную ДНК, оголенную РНК, искусственные вирионы, и усиленный агентом захват ДНК. Сонопорацию с использованием, например, системы Sonitron 2000 (Rich-Mar) также можно применять для доставки нуклеиновых кислот. В предпочтительном варианте осуществления, одну или более нуклеиновых кислот доставляют в виде мРНК. Также предпочтительным является применение кэппированных мРНК для увеличения трансляционной эффективности и/или стабильности мРНК. Особенно предпочтительными являются кэпы ARCA (аналог противообращенного кэпа) или их варианты. См., Патенты США US7074596 и US8153773, включенные в данный документ посредством ссылки.
[0157] Дополнительные иллюстративные системы доставки нуклеиновых кислот включают системы, предоставляемые Amaxa Biosystems (Cologne, Germany), Maxcyte, Inc. (Rockville, Maryland), BTX Molecular Delivery Systems (Holliston, MA) и Copernicus Therapeutics Inc., (см., например US6008336). Липофекция описана, например, в Патентах США №№ 5,049,386; 4,946,787; и 4,897,355) и реагенты для липофекции являются доступными для покупки (например, Трансфектам™ и Липофектин™ и Липофектамин™ RNAiMAX). Катионные и нейтральные липиды, которые являются подходящими для эффективной рецептор-распознающей липофекции полинуклеотидов включают описанные у Felgner, WO 91/17424, WO 91/16024. Доставка может проводиться в клетки (ex vivo введение) или ткани-мишени (in vivo введение).
[0158] Получение комплексов липид:нуклеиновая кислота, включающих липосомы направленного действия, такие как иммунолипидные комплексы, хорошо известно специалисту в данной области (см, например, Crystal, Science 270:404-410 (1995); Blaese et al., Cancer Gene Ther. 2:291-297 (1995); Behr et al., Bioconjugate Chem. 5:382-389 (1994); Remy et al., Bioconjugate Chem. 5:647-654 (1994); Gao et al., Gene Therapy 2:710-722 (1995); Ahmad et al., Cancer Res. 52:4817-4820 (1992); Патенты США №№ 4,186,183, 4,217,344, 4,235,871, 4,261,975, 4,485,054, 4,501,728, 4,774,085, 4,837,028, и 4,946,787).
[0159] Дополнительные методы доставки включают применение упаковывающих нуклеиновых кислот для доставки в несущих средах для доставки EnGeneIC (EDV). Эти EDV специфически доставляются к тканям-мишеням с использованием биспецифичных антител, где одно плечо антитела имеет специфичность к ткани-мишени, а другое плечо имеет специфичность к EDV. Антитело приносит EDV к поверхности клетки-мишени, и затем EDV вносится в клетку посредством эндоцитоза. При попадании в клетку, содержимое высвобождается (см., MacDiarmid et al (2009) Nature Biotechnology 27(7):643).
[0160] Применение систем, основанных на вирусных РНК или ДНК для доставки нуклеиновых кислот, кодирующих сконструированные ZFP, TALE или систему CRISPR/Cass имеет преимущество процессов с высоким выделением для целенаправленного действия вируса на специфичные клетки в организме и перемещения вирусной нагрузки в ядро. Вирусные векторы могут вводиться непосредственно пациентам (in vivo) или они могут применяться для обработки клеток in vitro, а модифицированные клетки вводят пациентам (ex vivo). Общепринятые системы на вирусной основе для доставки ZFP, TALE или систем CRISPR/Cas включают, но не ограничиваются перечисленным, ретровирусные, лентивирусные, аденовирусные, адено-ассоциированные вирусные векторы, вирусные векторы коровьей оспы и простого герпеса для переноса генов. Интеграция в геноме хозяина является возможной методами переноса генов с использованием ретровирусов, лентивирусов, и аденоассоциированных вирусов, часто приводя долговременной экспрессии введенного трансгена. Дополнительно, высокие эффективности трансдукции наблюдали во многих различных клеточных типах и тканях-мишенях.
[0161] Тропизм ретровируса может изменяться посредством введения чужеродных белков оболочки, расширяющих потенциальную целевую популяцию клеток-мишеней. Лентивирусные векторы представляют собой ретровирусные векторы, которые способны трансдуцировать или инфицировать неделящиеся клетки и обычно продуцируют высокие вирусные титры. Выбор ретровирусной системы переноса генов зависит от ткани-мишени. Ретровирусные векторы составлены из cis-действующих длинных терминальных повторов с плотностью упаковывания до 6-10 т.о. чужеродной последовательности. Минимальные cis-действующие LTR являются достаточными для репликации и упаковывания векторов, которые затем применяют для интеграции терапевтического гена в клетку-мишень для обеспечения перманентной экспрессии трансгена. Широко применяемые ретровирусные векторы включают векторы на основе вируса мышиной лейкемии (MuLV), вируса лейкоза гиббонов (GaLV), вируса иммунодефицита обезьян (SIV), вируса иммунодефицита человека (HIV), и их комбинации (см, например, Buchscher et al., J. Virol. 66:2731-2739 (1992); Johann et al., J. Virol. 66:1635-1640 (1992); Sommerfelt et al., Virol. 176:58-59 (1990); Wilson et al., J. Virol. 63:2374-2378 (1989); Miller et al., J. Virol. 65:2220-2224 (1991); PCT/US94/05700).
[0162] В областях применения, в которых временная экспрессия является предпочтительной, могут применяться системы на основе аденовирусов. Векторы на основе аденовирусов способны к очень высокой эффективности трансдукции у многих типов клеток и не требуют деления клеток. С такими векторами, были получены высокий титр и высокие уровни экспрессии. Этот вектор может быть получен в крупных количествах в относительно простой системе. Векторы аденоассоциированных вирусов (“AAV”) также применяют для трансдукции в клетки целевых нуклеиновых кислот, например, при in vitro получении нуклеиновых кислот и пептидов, и для in vivo и ex vivo методик генной терапии (см, например, West et al., Virology 160:38-47 (1987); Патент США № 4,797,368; WO 93/24641; Kotin, Human Gene Therapy 5:793-801 (1994); Muzyczka, J. Clin. Invest. 94:1351 (1994). Построение рекомбинантных векторов AAV описано в ряде публикаций, включающих Патент США № 5,173,414; Tratschin et al., Mol. Cell. Biol. 5:3251-3260 (1985); Tratschin, et al., Mol. Cell. Biol. 4:2072-2081 (1984); Hermonat & Muzyczka, PNAS 81:6466-6470 (1984); и Samulski et al., J. Virol. 63:03822-3828 (1989).
[0163] По меньшей мере, шесть подходов с применением вирусных векторов являются доступными в настоящее время для переноса генов при клинических испытаниях, в которых используются подходы, которые включают комплементацию дефектных векторов генами, вставленными в хелперные линии клеток для генерации трансдуцирующего средства.
[0164] pLASN и MFG-S являются примерами ретровирусных векторов, которые применялись при клинических испытаниях (Dunbar et al., Blood 85:3048-305 (1995); Kohn et al., Nat. Med. 1:1017-102 (1995); Malech et al., PNAS 94:22 12133-12138 (1997)). PA317/pLASN был первым терапевтическим вектором, применяемым в испытаниях генной терапии. (Blaese et al., Science 270:475-480 (1995)). Эффективности трансдукции, равные 50% или выше наблюдали векторов, упакованных в MFG-S. (Ellem et al., Immunol Immunother. 44(1):10-20 (1997); Dranoff et al., Hum. Gene Ther. 1:111-2 (1997).
[0165] Рекомбинантные аденоассоциированные вирусные векторы (rAAV) являются перспективными альтернативными системами доставки генов, основанными на дефектном и непатогенном парвовирусном адено-ассоциированном типа 2 вирусе. Все векторы получают из плазмиды, которая сохраняет только 145 пар оснований AAV инвертированных терминальных повторов фланкирующих кассету экспрессии трансгена. Эффективный перенос гена и стабильная доставка трансгена вследствие интеграции в геномы трансдуцированной клетки являются ключевыми признаками для этой векторной системы. (Wagner et al., Lancet 351:9117 1702-3 (1998), Kearns et al., Gene Ther. 9:748-55 (1996)). Другие серотипы AAV, включая AAV1, AAV3, AAV4, AAV5, AAV6, AAV8AAV 8.2, AAV9, и AAV rh10 и псевдотипированные AAV, таки как AAV2/8, AAV2/5 и AAV2/6 также могут применяться в соответствии с настоящим изобретением.
[0166] Рекомбинантные аденовирусные векторы (Ad) с дефективной репликацией могут быть получены с высоким титром и легко инфицировать ряд различных типов клеток. Большинство аденовирусных векторов сконструированы таким образом, что трансген заменяет гены Ad E1a, E1b, и/или E3; в последующем, вектор с дефективной репликацией распространяется в человеческих клетках 293, которые поставляют функцию гена, подвергнутую делеции, в trans. Векторы Ad могут трансдуцировать множество типов тканей in vivo, включающих неделящиеся, дифференцированные клетки, такие как клетки, обнаруженные в печени, почках и мышцах. Общепринятые Ad векторы имеют высокую несущую способность. Пример применения Ad вектора в клиническом испытании включал полинуклеотидную терапию для противоопухолевой иммунизации с использованием внутримышечной инъекции (Sterman et al., Hum. Gene Ther. 7:1083-9 (1998)). Дополнительные примеры применения аденовирусных векторов для переноса генов при клинических испытаниях включают Rosenecker et al., Infection 24:1 5-10 (1996); Sterman et al., Hum. Gene Ther. 9:7 1083-1089 (1998); Welsh et al., Hum. Gene Ther. 2:205-18 (1995); Alvarez et al., Hum. Gene Ther. 5:597-613 (1997); Topf et al., Gene Ther. 5:507-513 (1998); Sterman et al., Hum. Gene Ther. 7:1083-1089 (1998).
[0167] Упаковывающие клетки применяют для образования вирусных частиц, которые способны инфицировать клетку-хозяина. Такие клетки включают 293 клетки, которые упаковывают аденовирус, и ψ2 клетки или PA317 клетки, которые упаковывают ретровирус. Вирусные векторы, применяемые в генной терапии, обычно генерируют посредством линии клеток-производителей, которые упаковывают вектор нуклеиновой кислоты в вирусную частицу. Векторы обычно содержат минимальные вирусные последовательности, требуемые для упаковывания и последующей интеграции в хозяина (если применимо), причем другие вирусные последовательности заменяются на кассету экспрессии, кодирующую белок, подлежащий экспрессии. Отсутствие вирусных функций обеспечивается в ориентации trans посредством упаковывания линии клеток. Например, векторы AAV, применяемые в генной терапии, обычно обладают только последовательностями обращенного терминального повтора (ITR) из генома AAV, которые требуются для упаковывания и интеграции в геном хозяина. Вирусную ДНК упаковывают в линию клеток, которая содержит хелперную плазмиду, кодирующую другие гены AAV, а именно, rep и cap, но не содержит последовательности ITR. Линия клеток также инфицируется аденовирусом в качестве хелпера. Хелперный вирус инициирует репликацию вектора и экспрессию генов AAV из хелперной плазмиды. Хелперную плазмиду не упаковывают в значительных количествах из-за отсутствия последовательностей ITR. Загрязнение аденовирусом может быть снижено посредством, например, тепловой обработки, к которой аденовирус является более чувствительным, чем AAV.
[0168] Во многих областях применения генной терапии, является желательным, чтобы вектор генной терапии доставлялся с высокой степенью специфичности к конкретному типу ткани. Соответственно, вирусный вектор может быть модифицирован, чтобы иметь специфичность для данного клеточного типа, посредством экспрессии лиганд в качестве гибридного белка с белком вирусной оболочки на наружной поверхности вируса. Лиганд выбирают, чтобы он имел аффинность к рецептору, присутствие которого у клеточного типа, представляющего интерес, известно. Например, Han et al., Proc. Natl. Acad. Sci. USA 92:9747-9751 (1995), сообщают, что вирус Молони мышиной лейкемии может быть модифицирован, чтобы экспрессировать человеческий херегулин, слитый с gp70, и рекомбинантный вирус инфицирует клетки некоторых видов рака молочной железы человека, экспрессирующие рецептор человеческого эпидермального фактора роста. Этот принцип может быть распространен на другие клеточные пары вирус-мишень, в которых клетка-мишень экспрессирует рецептор и вирус экспрессирует гибридный белок, содержащий лиганд для рецептора клеточной поверхности. Например, может быть сконструирован нитевидный фаг для отображения фрагментов антитела (например, FAB или Fv), имеющих специфичную аффинность связывания для практически любого выбранного клеточного рецептора. Несмотря на то, что приведенное выше описание применяется прежде всего к вирусным векторам, такие же принципы могут применяться к невирусным векторам. Такие векторы могут быть сконструированы, чтобы содержать специфичные последовательности захвата, которые благоприятствуют захвату посредством специфичных клеток-мишеней.
[0169] Векторы генной терапии могут доставляться in vivo посредством введения индивидуальному пациенту, обычно посредством системного введения (например, внутривенной, внутрибрюшинной, внутримышечной, подкожной или внутричерепной инфузии) или местного применения, как описано ниже. Альтернативно, векторы могут быть доставлены в клетки ex vivo, такие как клетки, эксплантированные из индивидуального пациента (например, лимфоциты, аспираты костного мозга, тканевая биопсия) или универсальные донорные гемопоэтические стволовые клетки, с последующей реимплантацией клеток пациенту, обычно после селекции клеток, которые имеют включенный вектор.
[0170] Ex vivo клеточная трансфекция для диагностики, исследований или для генной терапии (например, посредством рееинфузии трансфицированных клетки в организм-хозяин) хорошо известна специалистам в данной области. В предпочтительном варианте осуществления, клетки выделяют из субъектного организма, трансфицированного нуклеиновой кислотой ZFP, TALE или системы CRISPR/Cas (ген, кДНК или мРНК), и реинфузируют обратно в субъектный организм (например, пациента). В предпочтительном варианте осуществления, одну или несколько нуклеиновых кислот доставляют в виде мРНК. Также предпочтительным является применение кэппированных мРНК для увеличения трансляционной эффективности и/или стабильности мРНК. Особенно предпочтительными являются кэпы ARCA (аналог противообращенного кэпа) или их варианты. См., Патенты США 7,074,596 и 8,153,773, полностью включенные в данное описание посредством ссылки. Различные клеточные типы, подходящие для ex vivo трансфекции, являются хорошо известными специалистам в данной области (см, например, Freshney et al., Culture of Animal Cells, A Manual of Basic Technique (3rd ed. 1994)) и ссылки, цитируемые в ней для обсуждения того, как изолировать и культивировать клетки от пациентов).
[0171] В одном варианте осуществления, стволовые клетки применяют в методиках ex vivo для клеточной трансфекции и генной терапии. Преимущество использования стволовых клеток состоит в том, что они могут быть дифференцированы в другие клеточные типы in vitro, или могут быть введены млекопитающему (такому как донору клеток), где они будут пересажены в костный мозг. Известны методы дифференцирования клеток CD34+ in vitro в клинически важные типы иммунных клеток с использованием цитокинов, таких как GM-CSF, IFN-γ и TNF-α (см., Inaba et al., J. Exp. Med. 176:1693-1702 (1992)).
[0172] Стволовые клетки изолируют для трансдукции и дифференциации с использованием известных методов. Например, стволовые клетки изолируют из клеток костного мозга посредством пэннинга клеток костного мозга с антителами, которые связывают нежелательные клетки, таки как CD4+ и CD8+ (T-клетки), CD45+ (panB клетки), GR-1 (гранулоциты), и Iad (дифференцированные антигенпрезентирующие клетки) (см., Inaba et al., J. Exp. Med. 176:1693-1702 (1992)).
[0173] Стволовые клетки, которые были модифицированы, также могут применяться в нескольких вариантах осуществления. Например, нейрональные стволовые клетки, которые были получены как резистентные к апоптозу, могут применяться в качестве терапевтических композиций, гле стволовые клетки также содержат ZFP TF изобретения. Резистентность к апоптозу может придаваться, например, посредством выключения BAX и/или BAK с использованием BAX- или BAK-специфичных TALEN или ZFN (см., Патентную публикацию США № 20100003756) в стволовых клетках, или тех, которые, которые были разрушены каспазой, например, повторно с использованием ZFN, специфичной к каспазе-6. Эти клетки могут быть трансфицированы с ZFP TF или TALE TF, которые, как известно, регулируют мутантный Htt или Htt дикого типа.
[0174] Векторы (например, ретровирусы, аденовирусы, липосомы и т.д.), содержащие терапевтические нуклеиновые кислоты ZFP, могут также непосредственно вводиться в организм для трансдукции клеток in vivo. Альтернативно, может вводиться оголенная ДНК. Введение проводят любым из путей, обычно применяемых для введения молекул в полный контакт с клетками крови или ткани, включая, но не ограничиваясь приведенными, инъекцию, инфузию, местное применение и электропорацию. Подходящие методы введения таких нуклеиновых кислот доступны и хорошо известны специалистам в данной области, и, несмотря на то, что может применяться более одного пути для введения конкретной композиции, конкретный путь часто может обеспечить более немедленную и более эффективную реакцию, чем другой путь.
[0175] Методы введения ДНК в гемопоэтические стволовые клетки раскрыты, например, в Патенте США № 5,928,638. Векторы, применимые для введения трансгенов в гемопоэтические стволовые клетки, например, CD34+ клетки, включают аденовирус Type 35.
[0176] Векторы, подходящие для введения трансгенов в иммунные клетки (например, T-клетки), включают неинтегрирующие лентивирусные векторы. См, например, Ory et al. (1996) Proc. Natl. Acad. Sci. USA 93:11382-11388; Dull et al. (1998) J. Virol. 72:8463-8471; Zuffery et al. (1998) J. Virol. 72:9873-9880; Follenzi et al. (2000) Nature Genetics 25:217-222.
[0177] Фармацевтически приемлемые носители определяются отчасти конкретной вводимой композицией, а также конкретным методом, используемым для введения композиции. Соответственно, существует большое разнообразие доступных подходящих готовых лекарственных форм фармацевтических композиций, как описано ниже (см, например, Remingtonʹs Pharmaceutical Sciences, 17th ed., 1989).
[0178] Как отмечено выше, раскрытые способы и композиции могут применяться к любым типам клеток, включающим, но не ограничиваясь приведенными, прокариотные клетки, грибковые клетки, клетки простейших, растительные клетки, клетки насекомых, клетки животных, клетки позвоночных, клетки млекопитающих и человеческие клетки. Подходящие линии клеток для экспрессии белка известны специалистам в данной области и включают, но не ограничиваются перечисленным COS, CHO (например, CHO-S, CHO-K1, CHO-DG44, CHO-DUXB11), VERO, MDCK, WI38, V79, B14AF28-G3, BHK, HaK, NS0, SP2/0-Ag14, HeLa, HEK293 (например, HEK293-F, HEK293-H, HEK293-T), perC6, клетки насекомых, таких как Spodoptera fugiperda (Sf), и грибковые клетки, такие как Saccharomyces, Pischia и Schizosaccharomyces. Потомство, варианты и производные этих линий клеток также могут применяться.
Применения
[0179] Раскрытые композиции и способы могут применяться для любых областей применения, в которых является желательным модулировать аллель Htt, включающих, но не ограниченных перечисленными, терапевтические и исследовательские области применения.
[0180] Заболевания и состояния, для которых подавляющие Htt ZFP TF или TALE TF могут применяться в качестве терапевтических средств, включают, но не ограничиваются перечисленными, болезнь Хантингтона. Дополнительно, способы и композиции, включающие ZFN или TALEN, специфичные для мутантных аллелей Htt, могут применяться в качестве терапевтического средств для лечения болезни Хантингтона.
[0181] ZFP-TF или TALE TF, которые подавляют аллель Htt БХ могут также применяться в сочетании с ZFP-TF или TALE-TF, которые активируют нейротрофические факторы, включая, но не ограничиваясь приведенными, GDNF и BDNF. Эти ZFP или TALE (или полинуклеотиды, кодирующие эти ZFP или TALE) могут вводиться одновременно (например, в одних и тех же фармацевтических композициях) или могут вводиться последовательно в любом порядке.
[0182] Способы и композиции для лечения болезни Хантингтона также включают композиции стволовых клеток, где мутантная копия аллеля HTT в стволовых клетках была модифицирована до аллеля Htt дикого типа с использованием Htt-специфичных ZFN или TALEN.
[0183] Способы и композиции изобретения являются также применимыми для разработки и реализации моделей in vitro и in vivo, например, животных моделей нарушений, связанных с тринуклеотидными повторами, которые обеспечивают исследование этих нарушений. Неограничивающие примеры подходящих моделей in vitro включают клетки или линии клеток из любого организма, включая фибробласты. Неограничивающие примеры подходящих животных для применения в качестве животных моделей включают, беспозвоночных (C. elegans, drosophila), грызунов (например, крыс или мышей), приматов (например, приматов, не являющихся людьми).
[0184] Композиции, описанные в данном документе, (например, белки и/или полинуклеотиды), по отдельности или в комбинации с другими подходящими компонентами (например, липосомами, наночастицами или другими компонентами, известными в данной области), могут быть изготовлены в виде аэрозольных готовых лекарственных форм (т.е. они могут быть “распылены”) для введения посредством ингаляции. Аэрозольные готовые лекарственные формы могут помещаться в находящиеся под давлением приемлемые пропелленты, такие как дихлордифторметан, пропан, азот и т.п. Готовые лекарственные формы, подходящие для парентерального введения, такого как, например, посредством внутривенного, внутримышечного, внутрикожного и подкожного путей, включают водные и неводные, изотонические стерильные инъекционные растворы, которые могут содержать антиоксиданты, буферы, бактериостатики и растворимые вещества, которые придают готовой лекарственной форме изотоничность с кровью предназначенного реципиента, и водные и неводные стерильные суспензии, которые могут включать суспендирующие средства, солюбилизаторы, загустители, стабилизаторы и консерванты. Композиции могут вводиться, например, посредством внутривенной инфузии, перорально, местно, внутрибрюшинно, интравезикально или интратекально. Готовые лекарственные формы соединений могут быть представлены в однодозовых или мультидозовых герметично закрытых контейнерах, таких как ампулы и виалы. Инъекционные растворы и суспензии могут быть получены из стерильных порошков, гранул и таблетка ранее описанного типа.
[0185] Доза, вводимая пациенту, должна быть достаточной, чтобы вызвать благоприятную терапевтическую реакцию у пациента с течением времени. Дозу определяют по эффективности действия и Kd конкретной используемой Htt-связывающей молекулы, клетки-мишени, и состоянию пациента, а также массе тела или площади поверхности тела пациента, подлежащего лечению. Размер дозы также определяют по наличию, природе и степени любых неблагоприятных побочных эффектов, которые сопровождают введение конкретного соединения или вектора у конкретного пациента.
ПРИМЕРЫ
Пример 1: Дизайн и конструкция Htt-нацеленных белков “цинковые пальцы”-факторов транскрипции (ZFP-TF) и ZFN
[0186] Белки “цинковые пальцы”, целенаправленно действующие на Htt, были сконструированы по существу, как описано в Патенте США № 6,534,261. Таблицы 1A и 1B показывают спирали распознавания ДНК-связывающего домена иллюстративного Htt-нацеленного ZFP, в то время как Таблицы 2A и 2B показывают целевые последовательности этих ZFP.
[0187] ZFP с одной смежной матрицей цинковых пальцев были спроектированы для сайтов-мишеней полностью в интервале области повтора CAG (Фиг. 1B). Такие ZFP могут связывать более длинные, мутантные тракты с более высокой аффинностью и/или более высокой чистой занятостью, достигая селективной репрессии мутантного аллеля. Были также спроектированы ZFN, которые целенаправленно действовали на сайты, которые лежат частично или полностью вне области CAG (Фиг. 1A), и, которые, следовательно, будут связываться с аллелем дикого типа и мутантным аллелем в равной степени, и регулировать экспрессию от обоих аллелей со сходной эффективностью. При проектировании белков “цинковые пальцы” для распознавания области, набор из одно- и двухпальцевых модулей может использоваться в комбинации “подбора по усмотрению”. Такие модули показаны ниже в Таблице 2C.
[0188] Мультимеризующиеся ZFP TF также конструируют, как описано выше, за исключением того, что вектор также содержит последовательности, кодирующие 1 или несколько доменов белкового взаимодействия (также именуемых доменами димеризации или белкового взаимодействия), которые обеспечивают мультимеризацию экспрессируемого белка вдоль тракта тринуклеотидных повторов, которые являются функционально связанными с последовательностями, кодирующими ZFP TF. См, Фиг. 1D и Фиг. 10. Таблица 3 показывает проекты домена димеризации, которые применяют с ZFP, нацеленными на область повтора CAG. Фиг. 10C показывает последовательности белка четырех мономерных матриксов ZFP, которые спроектированы, чтобы мультимеризоваться посредством взаимодействий между димеризующими цинковыми пальцами (DZ), DZ1-DZ4. Проекты основаны на работе, описанной в Mol. Syst. Biol. (2006) 2:2006.2011. Фиг. 10D показывает последовательности белков семи мономерных матриксов ZFP, которые спроектированы, чтобы мультимеризоваться посредством взаимодействий между суперспиралями (CC), CC1-CC7. Проект CC#1 основан на работе, описанной в (J. Am. Chem. Soc. (2001), 123:3151-3152), в то время как CC#2, CC#3 и CC#4 основаны на (J. Am. Chem. Soc. (2000), 122:5658-5659). Домены CC и DZ обеспечивают полимеризацию ZFP внутри большой бороздки тракта CAG (отображенной на Фиг. 10B). Посредством выбора матрикса пальцев и доменов димеризации с соответствующими свойствами связывания, эффективное связывание будет происходить только с удлиненным трактом CAG аллеля заболевания.
[0189] ZFP-TF были сконструированы как гибридные белки, содержащие последовательность ядерной локализации, сконструированный ДНК-связывающий домен цинковых пальцев (Таблицы 1A и 1B), нацеленный на аллель Htt, и домен репрессии KRAB из человеческого белка KOX1. См., Фиг. 1A, 1B и 1D. Спроектированные ДНК-связывающие домены содержат 3-6 пальцевых модулей, распознающих последовательности из 9-18 пар оснований (Таблицы 2A и 2B). Нуклеотиды в сайте-мишени, которые контактируют посредством спиралей распознавания, ZFP указаны буквами в верхнем регистре; неконтактирующие нуклеотиды указаны в нижнем регистре. Были также сконструированы молекулы ZFP-ZFP-TF, где два ДНК-связывающих домена ZFP были слиты с гибким линкером и слиты с доменом репрессии KRAB (Фиг. 1E). ДНК-связывающие домены выбирали из Таблиц 2A и 2B.
Пример 2: Репрессия обоих аллелей Htt в человеческих и мышиных клетках.
[0190] Для репрессии обоих аллелей Htt (аллель-неспецифичной), были спроектированы ZFP для связывания с промотором Htt и областью экзона 1, где сайт-мишень не находился полностью внутри CAG-повтора. См, Фиг. 1A. Для тестирования активности Htt, репрессирующие ZFP TF, ZFP TF трансфицировали в человеческие клетки, и экспрессию Htt регистрировали с использованием ПЦР в реальном масштабе времени.
[0191] Человеческие клетки HEK293 (Graham et al (1977) J Gen Virol 36:59-74) культивировали в DMEM, дополненной 10% FBS, и клетки 1e5трансфицировали с 1 мкг плазмидной ДНК, кодирующей указанные гибриды ZFP-KOX посредством Amaxa Нуклеофектор®, следуя инструкциям изготовителя.
[0192] Трансфицированные клетки инкубировали в течение 2 дней, и уровни эндогенного человеческого хантингтина (Htt) и бета-актина для контроля нормализации (ACTB) анализировали посредством ПЦР в реальном масштабе времени с использованием праймеров и зондов Hs00918176_m1 и 4352935E (Applied Biosystems), соответственно, в соответствии со стандартными протоколами. Уровни Экспресси Httровали как отношения Htt/ACTB, нормализованные к отношению для ложно-трансфицированных образцов (установленного как 1).
[0193] Как показано на Фиг. 2A, Htt-нацеленные ZFP подавляли экспрессию. Анализы методом вестерн-блоттинга проводили с использованием стандартных протоколов для подтверждения снижения уровня белка Htt (Фиг. 2B); белок p65 применяли в качестве контроля для нанесения.
[0194] Мышиные Htt-специфичные репрессоры ZFP TF временно трансфицировали в клетки Neuro2A (Klebe & Ruddle (1969) J. Cell Biol. 43: 69A) с использованием набора Lipofectamine® 2000 (Invitrogen), в соответствии с протоколами изготовителя. Уровни mHtt и ACTB мРНК измеряли через 48 часов после трансфекции с использованием набора ABI праймер/зонд Mm01213820 m1 и 4352933E от Taqman®, соответственно. Отношения mHtt/ACTB для образцов, трансфицированных ZFP нормализовали к отношению для контроля GFP (установленного, как 1).
[0195] Как показано на Фиг. 2C, ZFP подавлял экспрессию мышиного Htt. В дополнение, мышиные Htt-специфичные репрессоры ZFP-TF могут подавлять мышиный Htt в иммортализованных стриарных клетках, STБХh(Q111/Q7), полученных из мышей с активированным Htt (Trettel et al. (2000) Hum. Mol. Genet 9: 2799-2809). См, Фиг. 2D и 2E. мРНК для указанных ZFP генерировали с использованием набора mMessage mMachine (Ambion), и 0,1, 0,5 или 2 мкг этих мРНК трансфицировали с использованием нуклеофектора Amaxa, как описано выше. Клетки собирали через 48 часов после трансфекции для анализа экспрессии mHtt и ACTB, как описано выше. Наблюдали более значительную репрессию в стриарных клетках по сравнению с репрессией в клетках Neuro2A, и это может быть результатом усиленной эффективности трансфекции, достигаемой посредством трансфекции мРНК в стриарных клетках.
Пример 3: Селективная репрессия мутантного Htt в человеческих и мышиных клетках
[0196] Для достижения селективной репрессии мутантного аллеля Htt, были спроектированы ZFP для связывания внутри CAG-повтора. Фиг. 1B показывает один тип таких ZFP, со смежной матрицей цинковых пальцев, связанной с доменом репрессии (например, доменом KRAB из KOX1); эти ZFP могут быть спроектированы с соответствующей аффинностью, таким образом, что пороговая заполненность, требуемая для транскрипционной репрессии может быть установлена только на удлиненных CAG-повторах. Фиг. 1C, 1D и 1E показывают три других примера проекта ZFP, которые могут обеспечить специфичное связывание с удлиненными CAG-повторами.
[0197] ZFP, спроектированные, как проиллюстрировано на Фиг. 1B, вводили в клетки HEK293 и оценивали экспрессию Htt. ZFP-кодирующие конструкции трансфицировали в клетки HEK293 с использованием FugeneHD с использованием стандартных протоколов. Через семьдесят два часа после трансфекции полную РНК выделяли и уровни эндогенного человеческого хантингтина (Htt) относительно внутреннего контроля бета-актина (ACTB) анализировали посредством ПЦР в реальном масштабе времени с использованием праймеров и зондов Hs00918176_m1 и 4352935E (Applied Biosystems), соответственно. Отношения Htt/ACTB для образцов, трансфицированных ZFP, нормализовали к отношению для GFP-контроля (установленного как 1).
[0198] Как показано на Фиг. 3A, проектировали репрессоры ZFP (слитые с доменом репрессии KRAB) для связывания с CAG-повторами (Фиг. 1B), либо по верхней или по нижней цепи, в клетках HEK293, эффективно подавляли экспрессию Htt. Фиг. 3A отображает репрессоры транскрипции, гле экспрессию измеряли при двойных повторных трансфекциях (отдельные столбики на Фиг.), и проводили множественные анализы ПЦР в масштабе реального времени (планки погрешностей). Различные уровни репрессии от индивидуальных ZFP позволяют предположить, что они имеют различную аффинность к области повтора CAG. Поскольку аллели Htt в клетках HEK293 имеют 16 и 17 CAG, этот результат также предполагает, что “более слабые” ZFP, такие как 30640, не подавляют эффективно аллели Htt с CAG-повтором дикого типа (неудлиненным).
[0199] Для тестирования могут ли ZFP, такие как 30640 подавлять транскрипцию аллелей HTT с удлиненными CAG-повторами, конструировали люциферазные репортеры, управляемые фрагментом промотор/экзон 1 Htt, который содержит различные длины CAG-повтора. Сначала, фрагмент промотор/экзон 1 человеческого Htt амплифицировали из геномной ДНК HEK293 с использованием прямого праймера:
5ʹ GAAGATCTCACTTGGGGTCCTCAGGTCGTGCCGAC (SEQ ID NO:139)
и обратного праймера:
5ʹ GTATCCAAGCTTCAGCTTTTCCAGGGTCGCCTAGGCGGTCT (SEQ ID NO:140).
[0200] Прямой праймер вводит сайт BglII, обратный праймер изменяет первый ATG из Htt в TAG и создает сайт AvrII, и также включает сайт HindIII. Продукт ПЦР обрабатывали BglII и HindIII и лигировали к вектору pRL-TK (Proмега), который обрабатывали такими же ферментами, чтобы генерировать конструкцию pRL-Htt. Затем фрагмент экзона 1 человеческого Htt (кодирующая последовательность минус первый ATG) амплифицировали из геномной ДНК HEK293 или геномной ДНК от пациентов с БХ с удлиненными CAG-повторами с использованием прямого праймера:
5ʹ GCCTAGGCGACCCTGGAAAAGCTGATGAAGGCC (SEQ ID NO:141)
и обратный праймер: 5ʹ
5ʹ GTATCCAAGCTTGAGCTGCAGCGGGCCCAAACTCACG (SEQ ID NO:142).
[0201] Прямой праймер вводит сайт AvrII, обратный праймер вводит сайт HindIII. Продукт ПЦР обрабатывали AvrII и HindIII и лигировали к вектору pRL-Htt, который обрабатывали такими же ферментами. Клоны с 10, 17, 23 или 47 CAG-повторами (pRL-Htt-CAG(x)) идентифицировали посредством секвенирования.
[0202] Репортеры pRL-Htt-CAG(x) (300 нг) и репортер pGL3-промотора (100 нг, применяемый в качестве контроля нормализации, Progema) трансфицировали в клетки HEK293 со или 100 нг вектора экспрессии ZFP 30640 или без него. Активности люциферазы светляка (pGL репортер) и рениллы (pRL репортер) измеряли через 24 часа после трансфекции. Уровни люциферазы рениллы нормализовали до уровней люциферазы светляка из одного и того же трансфицированного образца, и дополнительно нормализовали до отношения ренилла/светляк “только репортерного” образца.
[0203] Как показано на Фиг. 3B, репрессия люциферазных репортеров под действием ZFP-TF 30640 увеличивается вместе с длиной CAG-повтора, позволяя предположить, что ZFP с ДНК-связывающими аффинностями, сходными с аффинностями для 30640, может подавлять активность Htt-промотора через удлиненный CAG-повтор, и уровень репрессии является зависимым от длины CAG-повторов.
[0204] Фиг. 3C показывает аналогичный эксперимент, как и на Фиг. 3B, за исключением того, что также тестировали “сильный” ZFP-TF 30657, а оба 30640 и 30657 тестировали при множестве доз, как указано. При каждом уровне дозы, 30640 давал больше репрессии репортера pRL-Htt-CAG47, чем репортера pRL-Htt-CAG23 (репрессия, зависимая от длины CAG-повтора), в то время как 30657 подавлял оба репортера до сходных уровней. На репортере pRL-Htt-CAG23, 30640 проявлял меньшую репрессию, чем 30657 при каждом уровне дозы, обобщая различие в их активностях на эндогенный аллель Htt с обычной длиной CAG-повтора (клетки HEK293, Фиг. 3A); но на репортер pRL-Htt-CAG47, 30640 и 30657 оказывали сходную репрессию при каждом уровне дозы, позволяя предположить, что “более слабые” ZFP, такие как 30640, могут эффективно подавлять промотор Htt через удлиненный CAG-повтор, наиболее вероятно потому, что только удлиненная CAG-мишень может обеспечить пороговую наполненность, требуемую для репрессии, устанавливаемой такими ZFP.
[0205] Фиг. 3D показывает, что ZFP-TF 30640 и 30657 (слитые с доменом репрессии KRAB из KOX1) может подавлять включенный аллель Htt (CAG111) в иммортализованных стриарных клетках, полученных из мышей с включенным HDh(Q111/Q7), демонстрируя, что ZFP, такаю как 30640, которая управляет зависимой от длины репрессией люциферазных репортеров в области CAG-повтора, может также подавлять экспрессию из эндогенного аллеля Htt, который имеет удлиненный CAG-повтор. мРНК для указанных ZFP генерировали с использованием набора mMessage mMachine kit (Ambion), и трансфицировали в клетки HDh(Q111/Q7) при указанных дохах с использованием нуклеофектора Amaxa. Для обнаружения экспрессии из мышиного аллеля Htt дикого типа, прямой праймер CAGGTCCGGCAGAGGAACC (SEQ ID NO:193) и обратный праймер TTCACACGGTCTTTCTTGGTGG (SEQ ID NO:194) применяли в ПЦР в реальном масштабе времени; для обнаружения экспрессии из включенного аллеля Htt, применяли прямой праймер GCCCGGCTGTGGCTGA (SEQ ID NO:195) и обратный праймер TTCACACGGTCTTTCTTGGTGG (SEQ ID NO:196).
[0206] Фиг. 3E показывает результат тестирования ZFP-TF 30640 и 30657 с полученной линией фибробластов от пациента с БХ (GM21756, Coriell), которая имеет 15 и 70 CAG в нормальном аллеле и мутантном аллеле Htt, соответственно. Сначала устанавливали аллель-специфичный аналитический тест с ПЦР в масштабе реального времени на основе SNP, чтобы обеспечить специфичное обнаружение от дикого типа или мутантного аллеля Htt. Фазирование SNP (rs363099 T/C) определяли по Carroll et al. (Mol Ther. (2011) 19:2178-85); “T” находится на нормальном аллеле, а “C” находится на мутантном аллеле. Для обнаружения экспрессии Htt из мутантного аллеля (099C), кДНК из фибробластов амплифицировали посредством ПЦР в реальном масштабе времени (SsoFast EvaGreen Supermix, Bio-Rad) с использованием прямого праймера 099C.F (5ʹAGTTTGGAGGGTTTCTC, SEQ ID NO:143) и обратного праймера 099.R5 (5ʹ TCGACTAAAGCAGGATTTCAGG, SEQ ID NO:144); температура отжига/удлинения была равна 55,6°C. Для обнаружения экспрессии Htt из аллеля дикого типа (099T), проводили ПЦР в масштабе реального времени кДНК фибробластов с использованием прямого праймера 099T.F (5ʹAGTTTGGAGGGTTTCTT, SEQ ID NO:145), обратного праймера 099.R5 и 3ʹ-фофорилированного блокатора олиго-099T.BL (5ʹAGGGTTTCTCCGCTCAGC-3ʹфос, SEQ ID NO:146); температура отжига/удлинения была равна 58,3°C. Уровни общего человеческого хантингтина (hHtt, как дикого типа, так и мутантного аллеля) и уровни контроля нормализации, бета-актина (ACTB), анализировали посредством ПЦР в реальном масштабе времени с использованием праймера/зонда Hs00918176_m1 и 4352935E (Applied Biosystems), соответственно. Для эксперимента, показанного на Фиг. 3E, мРНК для указанных ZFP генерировали с использованием набора mMessage mMachine (Ambion), 1 мкг мРНК трансфицировали с использованием нуклеофектора Amaxa, как указано выше. Клетки собирали через 48 часов после трансфекции; уровни мРНК от нормального (CAG15, 099T), мутантного (CAG70, 099C) аллеля Htt и общего Htt (hHtt) количественно оценивали, как описано выше, и нормализовали до уровней ACTB; отношения Htt/ACTB для каждого образца дополнительно нормализовали к отношению для ложно-трансфицированного образца. “Сильная” CAG-нацеленная ZFP 30657 подавляла оба аллеля, как ожидалось (на основании ее активности в клетках HEK293 Фиг. 3A). ZFP 30640, которая показывала зависимую от длины CAG-повтора репрессию репортеров, проявляла <10% репрессии аллеля дикого типа, в то время как подавляла мутантный аллель >90%. Уровни общего Htt в каждом образце соответствовали с такими уровнями для дикого типа (wt) и мутантного Htt в том же образце.
[0207] ZFP-30640 также тестировали на линии нормальных фибробластов, а также других линиях фибробластов при БХ, которые содержат CAG-повторы с различной длиной в гене Htt (см., Фиг. 3F). Экспрессию Htt от каждого аллеля обнаруживали, как описано выше. Никакой репрессии Htt не наблюдали в линии нормальных фибробластов (CAG18/18). Напротив, превосходное аллельное различие наблюдали у линий CAG 15/67 и CAG15/70 при как высокой, так и низкой дозе трансфицированной 30640 мРНК; аналогичные результаты были получены для двух линий БХ-фибробластов с промежуточной длиной CAG-повтора на мутантном аллеле (CAG 18/44 и CAG 18/45) - где удлиненный аллель подавлялся на ~80% при как высоких, так и низких дозах 30640, хотя аллель CAG18 оставался незатронутым. Взятые вместе, эти данные указывают на то, что аллель-специфичные репрессоры, такие как 30640, могут поддерживать высокую аллельную селективность по длине CAG в контексте более преобладающих генотипов заболевания, таких как CAG18/44 и CAG18/45.
[0208] Анализ методом вестерн-блоттинга применяли, чтобы показать, что ZFP, такие как 30640, селективно понижающе регулировали уровни мутантного белка Htt у двух полученных о пациента линий фибробластов, подтверждая аллель-специфичную регуляцию, которая была показана посредством анализов с колПЦР (см., Фиг. 3G). ZFP доставлялись посредством трансфекции мРНК (нуклеофекция Amaxa) при дозе, равной 300 нг, в 4 повтора клеток 1.5e5 и объединялись в пул перед размещением в 12-луночные планшеты. Через 48 часов, клетки промывали и собирали для получения белкового экстракта. Приблизительно 2,5 мкг экстракта загружали на гели с 5% Трис-ацетатом и обнаруживали посредством MAB2166 (Millipore). Дополнительно, те же самые образцы загружали на гели с 4-15% Tris-HCl (Bio-Rad) и переносили с использованием стандартных методов обнаружения посредством анти-B-актина (1:20000, Sigma) в качестве контролей для нанесения. На основании исследований с колПЦР, в которых измеряли мРНК Htt, 30640 представляет собой аллель-специфичный репрессор, целенаправленно действующий на CAG-повтор; 32528 представляет собой биаллельный репрессор, целенаправленно действующий на сайт начала транскрипции (TSS), и 30657 является CAG-нацеленным репрессором, который подавляет оба аллеля Htt при используемой дозе. Вестерн-блоттинг показал, что 30640 специфично снижал уровни мутантного Htt (верхняя полоса) в обеих линиях клеток, полученных от пациента с БХ, в то время как 32528 и 30657 подавляли оба аллеля сходным образом.
Пример 4: Дополнительные CAG-нацеленные конструкции ZFP, которые управляют аллель-специфичной репрессией Htt.
[0209] Фиг. 4A показывает результаты тестирования ZFP-TF 30640, 30643, 30645 и 33074 (все нацеленные на CAG-повтор и используют домен репрессии KRAB) на линии фибробластов CAG18/45 при БХ. Различные количества мРНК ZFP трансфицировали с использованием нуклеофектора Amaxa, как указано, экспрессию мутантного Htt (правый столбик), дикого типа Htt (средний столбик) и общего Htt (оба аллеля, левый столбик) измеряли, как описано выше, через 24 часа после трансфекции. ZFP 30640, 30645 и 33074 управляют аллель-специфичной репрессией на протяжении полного диапазона доз 3 мкг - 10 нг мРНК ZFP; в то время как 30643, по-видимому, значительно подавляет оба аллеля при дозах, которые составляют 30 нг или выше, и начинает проявлять селективность к аллелю при дозе 10 нг.
[0210] Фиг. 4B показывает результаты тестирования ZFP 30643, 30648, 30657 и 30658 (все нацеленные на CAG-повтор и используют домен репрессии KRAB) на линии фибробластов CAG15/70 при БХ. Различные количества мРНК ZFP трансфицировали с использованием нуклеофектора Amaxa, как указано, экспрессию мутантного Htt (правый столбик), дикого типа Htt (средний столбик) и общего Htt (оба аллеля, левый столбик) измеряли, как описано выше, через 24 часа после трансфекции. В сравнении с ZFP, который тестировали на предыдущей Фиг. (30640, 30645 и 33074), эти ZFP управляют специфичной репрессией мутантного Htt при более низких дозах. Эти результаты позволяют предположить, что в зависимости от уровней экспрессии, которые могут достигаться in vivo (например, в мозге пациентов с БХ), аллель-специфичная репрессия мутантного Htt может достигаться с использованием соответствующих конструкций ZFP.
Пример 5: Репрессия чередующихся CAG-содержащих генов
[0211] С использованием РНК, выделенной в Примере 3 (Фиг. 3E), анализировали репрессию других генов, содержащих CAG-повтор, и результаты отображены на Фиг. 5. Уровни экспрессии следующих генов исследовали с использованием ПЦР в масштабе реального времени и нормализовали к отношению для Актина: Атаксина 2 (“ATXN2”); Динамина (“DNM1”); F-коробочки только белка 11 (“FBXO11”), нитратредуктазы (“NAP”); точки распознавания комплекса субъединицы 4 (“ORC4”); фосфокиназы (“RNA”); белка OCT3/4 (“POU3”); домена THAP, содержащего ассоциированный с апоптозом белок 2 (“THAPII”); TATA-связывающего белка (“TBP”); и станнтокальцина 1 (“STC1”). В дополнение, отмечали расположение последовательности CAG-повтора относительно сайта начала транскрипции (TSS), и она указана на Фиг. 3F как “TSS@”, где “+” указывает положение оснований CAG-повторов, которые находятся по ходу транскрипции от TSS, и “-“ указывает положение оснований CAG-повторов, которые находятся против хода транскрипции от TSS. Также, число CAG-повторов (“#CAG”) указано для каждого гена.
[0212] Данные демонстрируют, что репрессия мутантного удлиненного аллеля Htt посредством 30640 является высокоспецифичной, и только подгруппа генов, содержащих CAG-повторы, чьи CAG-повторы находятся относительно близко к их соответствующим сайтам начала транскрипции, могут быть мишенями репрессии ZFP, таких как 30640.
Пример 6: Полногеномная специфичность аллель-специфичных ZFP репрессоров Htt
[0213] Фибробласты БХ (CAG18/45) трансфицировали для исследования полногеномной специфичности CAG-нацеленных ZFP посредством микроматричного анализа (Фиг. 6). ZFP доставляли посредством трансфекции мРНК (нуклеофекции Amaxa) при указанных дозах - ZFP 30640, 30645 и 33074 трансфицировали в шести параллельных опытах при дозе, равной 75 нг, 15 нг и 15 нг, соответственно; мРНК GFP-Kox (150 нг) трансфицировали в качестве контроля и применяли в качестве носителя, чтобы привести общее количество трансфицированной мРНК до 150 нг во всех образцах. Экспрессию от аллелей CAG18 (099T, средние столбики) и CAG45 (099C, правые столбики) измеряли посредством аллель-специфичных реагентов колПЦР через 24 часа после трансфекции, как описано выше, где каждый из образцов (1-6) представлял собой биологические повторы (раздельные трансфекции). Уровни Htt нормализовали до уровней для GAPDH. Мутантную аллель-специфичную репрессию Htt наблюдали для всех трех ZFP. Четыре наиболее сходных повтора затем были выбраны для микроматричного анализа (Affymetrix HGU133plus2.0) следующим образом: GFP повтор 1, 3, 4 и 6; 30640 повтор 2, 3, 5 и 6; 30645 повтор 2, 3, 5 и 6; и 33074 повтор 1, 3, 4 и 5 применяли для микроматричного анализа. Robust Multi-array Average (RMA) применяли для нормализации необработанных сигналов от каждого набора зондов; ZFP-трансфицированные образцы сравнивали с GFP-трансфицированными образцами с использованием T-теста; Запросы “изменения” были сделаны на генах (наборы зондов) с >2-кратным различием относительно контрольных образцов и имели в T-тесте P-значение<0,05. На основании этого критерия, 30640 подавлял только два гена, станниокальцина 1 (STC1) и удлиненного синаптотагмин-подобного белка 1 (ESYT1); каждый из 30645 и 33074 подавлял только один ген, STC1 и рецептора А интерлейкина 17 (ILR17RA), соответственно. Htt не обнаруживали как репрессированный (>2-кратная репрессия) ген, так как набор зондов на Htt на матрице обнаруживает мРНК как Htt дикого типа, так и мутантного Htt. Этот эксперимент демонстрирует, что мутантные Htt-специфичные ZFP, когда экспрессируются при уровнях, которые управляют эффективной аллель-специфичной репрессией Htt, могут функционировать с очень высокой полногеномной специфичностью.
Пример 7: Аллель-специфичная репрессия в нейрональных стволовых клетках (NSC) при БХ
[0214] iPSC/ESC БХ пассировали с аккутазой и культивировали на чашках, покрытых матригелем в среде E8 (Life Technologies). Нейрональные стволовые клетки были получены с использованием среды StemPro для нейрональной индукции (Life Technologies). Вкратце, iPSC/ESC высевали в покрытую гелтрексом 6-луночную чашку с 200000 клетками/лунке, и, при достижении слияния 10-20%, среду меняли на среду StemPro для нейрональной индукции. Среду меняли каждые 2 дня, и NSC собирали и размножали на 7-ой день. Среду StemPro NSC SFM (Life Technologies) применяли для культивирования NSC. NSC БХ (CAG17/69, полученные от Coriell GM23225 iPSC) трансфицировали с 1,5 или 0,5 мкг мРНК ZFP с использованием нуклеофекции. Через сорок восемь часов после трансфекции клетки собирали, и экспрессию количественно оценивали посредством РВ-ПЦР. Аллель-специфичное обнаружение экспрессии Htt проводили с использованием SNP (rs1143646)-основанного генотипирующего аналитического теста #4351376 (Applied Biosystems). При дозах ZFP, которые тестировали, 30640 давал аллель-специфичную репрессию мутантного Htt, 30643 давал ~50% репрессию Htt дикого типа и ~90% репрессию мутантного Htt, и 30648 подавлял оба аллеля (Фиг. 7); поведение этих ZFP совпадало с их поведением в фибробластах при БХ (Фиг. 4). Общие уровни Htt (средние столбики) для каждого образца соответствовали уровням мутантного Htt и Htt дикого типа.
Пример 8: Репрессия Htt в дифференцированных нейронах БХ
[0215] NSC БХ пассировали с аккутазой на чашках, покрытых гельтрексом. Дифференциацию нейронов индуцировали посредством смены среды на среду для нейрональной дифференциации, содержащую StemPRO NSC SFM среду без (bFGF и EGF). Среду меняли каждые 3-4 дня в течение периода до 21 дня. Нейроны получали из NSC (CAG17/48, полученной из БХ ESC) посредством культивирования в среде для нейрональной дифференциации. На 15-ый день после нейрональной индукции клетки трансфицировали с 1,0 или 0,5 мкг ZFP мРНК с использованием нуклеофекции. Через сорок восемь часов после трансфекции клетки собирали и экспрессию гена количественно оценивали посредством РВ-ПЦР. Поскольку линия клеток этого пациента не содержит SNP, который обеспечивает аллель-специфичное обнаружение дикого типа и мутантного Htt на основе колПЦР, могут быть измерены только общие уровни Htt. Так как авторы показали, что 30640 и 33074 не подавляет аллель CAG18 или CAG17 в фибробластах и NSC при БХ, уровни общего Htt, наблюдаемые в образцах, обработанных 30640 и 33074, находятся в соответствии с аллель-специфичной репрессией мутантного аллеля (CAG48). Более мощная репрессия под действием 30643 и 30648 при тестируемых дозах ZFP также соответствует поведению этих ZFP в фибробластах при БХ (Фиг. 8).
Пример 9: CAG-нацеленный репрессор подавляет мутантный трансген Htt у мышей R6/2
[0216] Мыши R6/2 (которые несут трансген экзона 1 мутантного человеческого Htt с ~120 CAG-повторами, см. Mangiarini et al, (1996) Cell 15:197) получали стереотаксические, билатеральные стриарные инъекции векторного генома 3e10 из рекомбинантного AAV2/6, кодирующего либо ZFP 30640-KOX или GFP под управлением CMV-промотора. Мышам вводили инъекцию в возрасте 5 недель и их умерщвляли для молекулярного анализа в возрасте 8 недель. Левое и правое полосатые тела иссекали из каждого полушария и мгновенно замораживали. Для оценки репрессии мутантного трансгена Htt, общую РНК экстрагировали из каждого полосатого тела с использованием TRIzol Plus (Life Technologies) с последующим синтезом кДНК с использованием High Capacity RT (Life Technologies). Последовательно, экспрессию трансгена R6/2 измеряли посредством колПЦР и нормализовали до геометрического среднего для трех референтных генов (Atp5b, Eif4a2, UbC), как описано прежде Benn et al. ((2008) Molecular Neurodegeneration: 3, 17). Авторы наблюдали статистически значимую репрессию (P<0,001) мутантного трансгена Htt в четырех ZFP-обработанных полосатых телах относительно четырех GFP-обработанных контрольных полосатых тел (Фиг. 9). Средняя репрессия R6/2 составляла 64,9% от GFP-обработанных контролей. Поскольку полный охват полосатого тела не был достигнут с использованием однократной стереотаксической инъекции, а AAV2/6 преимущественно трансдуцирует нейрональные клетки, кратность наблюдаемой репрессии (~35%) вероятно является заниженной по отношению к фактической репрессии в клетках, которые трансдуцировали вектором AAV.
Пример 10: Селективная репрессия мутантного Htt с использованием ZFP с доменами димеризации/мультимеризации
[0217] Чтобы сконструировать факторы транскрипции с цинковыми пальцами для лучшего различения между короткими CAG- и длинными GAG-повторами, авторы пытались, как уменьшить ДНК-связывающую аффинность индивидуальных факторов транскрипции с цинковыми пальцами, так и увеличить силу взаимодействия между различными копиями гибридного белка, связанными с примыкающими субсайтами внутри CAG-повтора. Чтобы уменьшить ДНК-связывающую аффинность индивидуальных факторов транскрипции с цинковыми пальцами, авторы генерировали домены цинковых пальцев с меньшими цинковыми пальцами и/или с аминокислотными последовательностями, для которых ожидали связывание ДНК с аффинностью меньше оптимальной. Чтобы увеличить силу взаимодействия между различными копиями гибридного белка, связанными с примыкающими субсайтами внутри CAG-повтора, авторы сливали различные домены димеризации с авторскими факторами транскрипции с цинковыми пальцами. Домены димеризации могут взаимодействовать "параллельным" образом и давать на выходе димеры гибридных белков типа "голова-к-голове" или "хвост-к-хвосту", которые содержат их. Одна потенциальная стратегия димеризации требует матрицы идентичных гибридов ZFP-фактор транскрипции, которые связываются в ориентации "голова-к-хвосту" и, таким образом, данная стратегия требует доменов димеризации, которые взаимодействуют "антипараллельным" образом. См. например, McClain et al. (2001) J. Am. Chem. Soc. 123:3151-3152) и пептиды с димеризующими цинковыми пальцами (Giesecke et al. (2006), Molecular Systems Biology 2:2006.2011).
[0218] Конструкции димеризации CC1 и CC2 были основаны на парах антипараллельных суперспиралей (McClain et al, там же, Ghosh et al. (2000) J Am Chem Soc 122:5658-5659). Конструкции димеризации CC3 и CC4 являлись укороченными версиями CC2, в которых отсутствуют либо 4 остатка или 7 остатков, соответственно. Конструкции димеризации DZ1, DZ2, DZ3, и DZ4 были основаны на паре димеризующих доменов цинковых пальцев (Giesecke et al., там же). В каждом случае, один члены пары был слит с N-концом ДНК-связывающим доменом с цинковыми пальцами, а другой член пары был слит с C-концом ДНК-связывающего домена с цинковыми пальцами. Короткие линкеры, обогащенные глицином и остатками серина, применяли для слияния домена димеризации с доменом связывания с цинковыми пальцами. В дополнительных вариантах осуществления изобретения используют линкеры с чередующимися длинами и/или аминокислотным составом. Линкеры с удаленными одним или несколькими остатками или с заменами одного или нескольких остатков глицина или серина на другие аминокислотные остатки будут снижать гибкость этих линкеров и могут приводить к улучшенному различению между длинными и короткими CAG-повторами.
[0219] Для достижения селективной репрессии мутантного аллеля Htt, были сконструированы ZFP, как проиллюстрировано на Фиг. 1D и Фиг. 10A и 4B. Фиг. 10C и 10D показывают последовательности доменов мультимеризации. Фиг. 11A и 11B отображают результаты таких экспериментов, запланированных, чтобы измерить способность ZFP-TF, содержащих домены CC и DZ, соответственно, подавлять их мишени. Для этих экспериментов, указанные конструкции ZFP (50 нг) сотрансфицировали с pRL-Htt-CAG17 (200 нг), pGL3-Htt-CAG47 (200 нг) и pVax-SEAP (секретируемой щелочной фосфатазой, 10 нг, применяемой как контроль нормализации) в клетки HEK293. Люциферазная активность и активности секретируемой щелочной фосфатазы измеряли через 24 часа после трансфекции. Отношения люциферазы Рениллы (CAG17)/SEAP и люциферазы светлячка, (CAG47)/SEAP, для каждого образца нормализовали до отношения в образцах, содержащих только репортер. Репортер pGL3-Htt-CAG47 был сконструирован таким же образом, что и репортер pRL-Htt-CAG47 (см. Пример 3), за исключением того, что конструкцию pGL-промотора (Promega) применяли вместо конструкции pRL-TK.
[0220] Как показано на Фиг. 11A, 3 ZFP, при тестировании в качестве одной или нескольких CC домен-содержащих конструкций, усиливали репрессию одного или обоих репортеров, при сравнении с конструкциями с таким же ZFP, но без CC доменов. Фиг. 11B показывает, что домены DZ1 и DZ3 усиливали репрессию 32220 на оба репортера.
[0221] Эти результаты позволяют предположить, что CC и DZ домены могут, в целом, увеличивать аффинность мультимеризованных ZFP, и, что конструкция ДНК-связывающего домена и домена димеризации могут давать оптимальное различение по длине CAG-повторов.
Пример 11: Селективная репрессия мутантного Htt конструкциями ZFP-ZFP-Kox
[0222] ZFP TF тестировали на фибробластах при БХ, которые имелт конструкцию ZFP-ZFP-KOX. В этих экспериментах, два ZFP ДНК-связывающих домена были связаны вместе гибким линкером (LRQKDAARGSAAMAERPFQ, SEQ ID NO:179) и слиты с доменом репрессии KOX. Линкер помещали между консервативными гистидинами и цистеинами. Белки тестировали, как описано выше, с использованием мРНК ZFP при указанных дозах. Эти результаты на линиях фибробластов CAG18/45 (Фиг. 12A) и CAG20/41 (Фиг. 12B) при БХ, демонстрировали, что связывание менее активных ZFP ДНК-связывающих доменов этим способом может приводить к композитным ZFP, которые управляют аллель-специфичной репрессией.
Пример 12: Активация Htt в мышиных клетках
[0223] ZFP, описанные в данном документе, также оценивали по их способности активировать экспрессию Htt. ZFP, нацеленные на область от +200 до +467 пар оснований (относительно сайта начала транскрипции) мышиного Htt сливали с доменом активации транскрипции субъединицы NFκB p65. Эта область нацеливания была выбрана, поскольку этот фрагмент заменяли на соответствующую последовательность (основная часть экзона 1 и некоторая часть последовательности интрона 1) из человеческого Htt в различных мышиных моделях с активацией гена БХ (Menalled et al. (2003) J. Comp. Neurol 4651:11-26; Wheeler et al. (2000) Hum Mol Genet 8:115-122), следовательно ZFP, нацеленные на эту область, могут селективно активировать аллель дикого типа у таких животных, но не активированный аллель.
[0224] ZFP трансфицировали в клетки Neuro2A (оба аллеля Htt являются аллелями дикого типа в этих клетках), уровни мышиного Htt и мРНК ACTB измеряли, как описано в Примере 2 (дублирующие трансфекции и множественные анализы).
[0225] Как показано на Фиг. 13, увеличение уровней мРНК Htt в сравнении с ложной трансфекцией обнаруживали с использованием обоих ZFP-TF. См. Фиг. 13A. Увеличенные уровни белка Htt подтверждали Вестерн-блоттингом. См. Фиг. 13B.
[0226] Генерация активированного аллеля Htt проиллюстрирована на Фиг. 13C; выравнивание последовательности (Фиг. 13D) показывает дивергенцию между мышиной последовательностью, которую заменяли, и соответствующей человеческой последовательностью. Фиг. 13E показывает, что когда такие активаторы ZFP трансфицировали в иммортализованные стриарные клетки, полученные от мышей с активированным HdhQ111/Q7, только Htt дикого типа селективно активировался.
Пример 13: Регуляция экспрессии Htt in vivo
[0227] Для тестирования эффективности действия Htt-специфичных ZFP TF in vivo, получали векторы AAV2, кодирующие ZFP. Эти конструкции на основе AAV2 затем доставляют в мозг мышей. Для ZFP TF специфичных к человеческому Htt, векторы AAV доставляют мышам R6/2 mice мышам BAC БХ (штаммы C57Bl/6 или FVB/N) для оценки репрессии человеческого трансгена, а также изменения в БХ-подобных фенотипах. Для ZFP, специфичных к мышиному Htt (активаторов или репрессоров), векторы AAV доставляют мышам дикого типа (C57Bl/6 или FVB/N) или мышам с активированным человеческим Htt (HdhQ111/Q7, HdhQ140/Q7 или HdhQ175/Q7) для оценки активации или репрессии экспрессии эндогенного мышиного Htt. Для ZFP, которые преимущественно нацелены на CAG-удлиненный аллель, векторы AAV доставляют мышам R6/2 или мышам с активированным человеческим Htt (HdhQ111/Q7, HdhQ140/Q7 или HdhQ175/Q7) для исследования селективной репрессии аллеля дикого типа в сравнении с удлиненным аллелем Htt. После умерщвления, ткани мозга анализируют на предмет экспрессии Htt посредством ПЦР Taqman в реальном масштабе времени, и демонстрируют, что гены Htt модулируются посредством ZFP-TF.
[0228] Линия трансгенных мышей R6/2 является хорошо охарактеризованной животной моделью БХ, в которой промотор человеческого Htt и фрагмент экзона 1, который содержит удлиненный CAG-повтор является эктопически экспрессируемым (Mangiarini et al. (1996) Cell 87, 493-506). Эти мыши проявляют БХ-подобные фенотипические поведенческие изменения, а также потерю стриарных срединных шипиковых нейронов (MSN), что является отличительной чертой БХ. В соответствии с дегенерацией MSN, сообщается о сниженной экспрессии маркеров MSN, DARPP-32 (Bibb et al. 2000, Proc. Natl. Acad. Sci. 97(12):6809-6814), фосфодиэстеразы 10a (PDE10a) (Hebb et al. 2004 Neuroscience, 123(4):967-81), дофаминового рецептора D1 (DRD1) и дофаминового рецептора D2 (DRD2) (Cha et al. 1998 Proc. Natl. Acad. Sci. 95(11): 6480-6485) у мышей R6/2.
[0229] Для тестирования эффективности действия in vivo генетического репрессора, ZFP-33074, который специфически подавляет мутантный аллель Htt в фибробластах от пациента с БХ и нейронах, AAV6-33074 (т.е. AAV6, содержащий генетический репрессор) билатерально доставляли в полосатое тело мышей R6/2 5-недельного возраста с использованием стереотаксической инъекции. Каждое полосатое тело получало инъекции AAV в два участка, координаты для передней и задней инфузии составляли A/P +1,4, M/L +/-1,7, D/V -3,5 и A/P +0,2, M/L +/-2,3, D/V -3,2, соответственно. Передний участок получал 5 мкл вектора AAV, а задний участок получал 4 мкл вектора. Титр вектора AAV6-33074 составлял 1x1013 векторного генома/мл. Контрольные животные получали такую же дозу AAV6-GFP (т.е. AAV6, содержащего зеленый флуоресцентный белок) в те же самые положения в полосатом теле.
[0230] Мышей R6/2, инъецированных AAV6-33074 или AAV6-GFP, тестировали еженедельно на хватательный рефлекс, который является хорошо установленным моторным нарушением, проявляемым этими животными (Mangiarini et al. 1996 Cell 87, 493-506). Вкратце, каждую мышь удаляли из ее клетки для содержания и помещали на крышку клетки. Наблюдатель затем осторожно тянул животное в заднем направлении и вверх плавным движением, пока животное не повисало над поверхностью на примерно 12 дюймов. Животное затем оценивали по балльной системе в течение 30 секунд. Если наблюдали сжимание только передних конечностей, животному присваивали 1 балл. Если наблюдали только сжимание задних конечностей, животному присваивали 2 балла. Если наблюдали сжимание как задних, так и передних конечностей, но не в одно и то же время, животному присваивали 3 балла. Полное сжимание, определяемое по одновременному сжиманию задних и передних конечностей вытягиваемых плотно к середине, получало оценку 4 балла (см. Фиг. 16). После 30-секундного подвешивания, животное возвращали в его клетку для содержания. Для каждой группы обработки, а также совпадающих по возрасту однопометных животных дикого типа, определяли долю животных, которые проявляли полное сжимание (4 балла) при каждом еженедельном наблюдении. В сравнении с животными, обработанными AAV6-GFP, AAV6-33074-обработанные мыши R6/2 проявляли сниженную частоту сжимания между 7 и 12 неделями возраста (когда животных умерщвляли для анализа экспрессии); снижение было статистически значимым (анализ распределения хи-квадрат, p<0,05) при возрасте 9 и 12 недель (Фиг. 17). Этот результат демонстрирует, что генетический репрессор мутантного Htt, ZFP-33074, улучшал хватательный рефлекс, хорошо охарактеризованное моторное нарушение у мышей R6/2.
[0231] Уровни экспрессии мутантного Htt и MSN-маркера DARPP-32 в полосатом теле мышей R6/2 измеряли посредством иммуногистохимии. Через 7 недель после инъекции AAV-33074, мыши R6/2 подвергали глубокой анестезии пентобарбиталом и перфузировали через восходящую часть дуги аорты изотоническим физраствором с последующей перфузией 250 мл ледяного 4% параформальдегида в 0,1M фосфатном буфере, pH 7.4. Мозг удаляли и постфиксировали в течение ночи в том же самом растворе и затем переносили в 25% сахарозу перед секционированием на микротоме со стадией замораживания при 40 мкм. Срезы промывали в PBS и блокировали в течение 4 часов при комнатной температуре в 0,3% Тритоне X-100 20% Нормальной Козьей сыворотки/PBS (блокирующий раствор). Кроличьи антитела против-DARPP-32 (Cell Signaling), и мышиные антитела против мутантного Htt (Millipore) инкубировали в течение 24 часов при 4°C в блокирующем растворе. Затем, срезы промывали в PBS, инкубировали в течение 2 часов в 0,3% Тритон X-100/PBS с ослиными Alexa555 против кроличьих (Life technologies) и козьими Alexa488 против мышиных (Life technologies). Окончательно, срезы промывали в PBS, инкубировали 10 минут с DAPI (Invitrogen) и устанавливали с покровными стеклами для анализа методом флуоресцентной микроскопии. Сниженное окрашивание мутантного Htt наблюдали в областях полосатого тела с инъецированным AAV-33074; важно, что, увеличенное окрашивание DARRP-32 наблюдали в той же самой области, позволяя предположить, что генетический репрессор мутантного Htt, ZFP-33074 регулирует с понижением дегенерацию MSN.
[0232] Экспрессию уровней маркеров мутантного Htt и MSN также измеряли в аналитических тестах с использованием ПЦР в масштабе реального времени (Taqman). Через 7 недель после инфузии вектора AAV (AAV-ZFP или AAV-GFP), полосатые тела мышей R6/2 выделяли и иссекали на переднюю, среднюю и заднюю секции и мгновенно замораживали индивидуально. Общую РНК выделяли из каждой стриарной секции с использованием набора для очистки РНК TRIzolPlus (Life Technologies), и 500 нг РНК применяли для генерации кДНК с использованием набора для высокопроизводительной обратной транскрипции кДНК, High Capacity cDNA Reverse Transcription kit (Life Technologies) в реакционной смеси с объемом 20-мкл. Одну сотую часть кДНК затем применяли в реакции ПЦР в масштабе реального времени для определения уровней генов, представляющих интерес (мышиного Htt дикого типа, трансгена мутантного Htt, DARPP-32, PDE10a, DRD1 и DRD2) и уровней генов нормализации (Atp5b, Eif4a2, UbC); гены нормализации были выбраны на основании Benn et al (2008) Molecular Neurodegeneration 3:17. Наборы праймер/зонд для мышиных DARPP-32, PDE10a, дофаминового рецептора D1, дофаминового рецептора D2, Atp5b, Eif4a2 и UbC были приобретены от IDT; праймер/зонд для мышиного Htt был приобретен от Life Technologies, и праймер/зонд для трансгена мутантного Htt был таким, как описано в Benn et al, там же. Последовательности наборов праймер/зонд перечислены в Таблице 3. Аналитические тесты с ПЦР в масштабе реального времени выполняли с использованием SsoFast Probes Supermix (Bio-Rad) в формате 384-луночного планшета на приборе CFX-384 для проведения ПЦР в масштабе реального времени (Bio-Rad) по инструкции изготовителя. Анализ экспрессии проводили с использованием программного обеспечения CFX manager (v3.0). Вкратце, уровень экспрессии каждого гена, представляющего интерес, и гена нормализации анализировали в реакциях с тремя повторами и количественно оценивали с использованием стандартных кривых, охватывающих 125-кратный диапазон концентраций (ряд пяти-кратных разведений). Уровень экспрессии каждого гена, представляющего интерес, нормализовали до средних уровней трех генов нормализации из того же образца (отношение ген интереса/ген нормализации); это отношение для каждого образца затем масштабировали до отношения, среднего для всех AAV-GFP-обработанных образцов (установленного как 1) и наносили на график. Статистическую достоверность оценивали с использованием T-теста с двусторонним парным критерием Стьюдента. Для DARPP-32, экспрессия составляла 178 процентов от экспрессии у мышей, получающих контрольный GFP, в то время как для PDE10a, Drd1 и Drd2 уровни экспрессии составляли 184%, 189%, и 162% от экспрессии при контрольном GFP, соответственно. AAV-ZFP-33074-обработанные полосатые тела показывали ~60% репрессии мутантного Htt относительно полосатых тел, обработанных AAV-GFP (P<0,001), в то время как экспрессия мышиного Htt дикого типа оставалась неизменной. ZFP-обработанные полосатые тела также показывали статистически значимое увеличение для всех маркеров срединных шипиковых нейронов (MSN), которые обследовали (DARPP-32, PDE10A, DRD1 и DRD2), предполагая, что генетический репрессор мутантного Htt, ZFP-33074, снижал дегенерацию стриарных MSN у мышей R6/2 (Фиг. 17).
Пример 14: Со-трансфекция нейротрофного фактора и аллеля Htt БХ-специфичного ZFP TF
[0233] Htt-специфичные ZFP TF, идентифицированные выше, coтрансфицировали с ZFP TF-специфичными к нейротрофическому мозговому фактору. Применяемые ZFP TF, специфичные к нейротрофическим факторам мозга, являются специфичными для обоих GDNF или BDNF.
Пример 15: Дизайн и конструкция Htt-нацеленных нуклеаз “цинковые пальцы” (ZFN)
[0234] ZFN, целенаправленно действующие на человеческий Htt и мышиный Htt были спроектированы для нацеливания на последовательности, фланкирующие CAG-повторы, последовательности рядом с первым кодирующим ATG, стоп-кодон, а также в ранних экзонах. ZFN были сконструированы и введены в плазмиды или аденовирусные векторы по существу так, как описано в Urnov et al. (2005) Nature 435(7042):646-651, Perez et al (2008) Nature Biotechnology 26(7): 808-816, и Патентной публикации США 2008/0131962.
Пример 16: Расщепляющая активность Htt-специфичных ZFN
[0235] Для тестирования расщепляющей активности, плазмиды, кодирующие пары человеческих Htt-специфичных ZFN описанных выше, трансфицировали в клетки K562. Клетки K562 были получены из Американской коллекции типовых культур и выращены, как рекомендовано, в среде F-12 (Invitrogen), дополненной 10% сертифицированной фетальной телячьей сывороткой (FCS, Cyclone). Клетки отсоединяли от пластиковой посуды с использованием протеазы TrypLE Select™ (Invitrogen). Для трансфекци, один миллион клеток K562 смешивали с 2 мкг плазмиды нуклеазы “цинковые пальцы” и 100 мкл Раствора Т Amaxa. Клетки трансфицировали в Нуклеофекторе II™ Amaxa с использованием программы U-23 и извлекали в 1,4 мл теплой среды F-12+10% FCS.
[0236] Геномную ДНК собирали, и часть локуса Htt, охватывающую намеченный сайт расщепления амплифицировали с использованием ПЦР. ПЦР с использованием полимерaзы Accuprime HiFi от InVitrogen проводили следующим образом: после первоначальной 3-минутной денатурации при 94°C, 30 циклов ПЦР выполняли с 30-секундной стадией денатурации при 94°C с последующей 30-секундной стадией отжига при 58°C с последующей 30-секундной стадией удлинения при 68°C. После завершения 30 циклов, реакционную смесь инкубировали при 68°C в течение 7 минут, затем при 10°C неопределенное время.
[0237] Геномную ДНК из клеток K562, обработанных Htt-специфичными ZFN, исследовали посредством нуклеазы Surveyor™ (Transgenome) как описано, например, в Патентных публикациях США №№ 20080015164; 20080131962 и 20080159996.
[0238] Плазмиды, кодирующие пары мышиных Htt-специфичных ZFN, тестировали аналогичным образом на клетках Neuro-2a.
[0239] Фиг. 14A и B показывают, что ZFN обладали способностью к целенаправленному действию на гены Htt с эффективностью генной модификации между 8-40%, анализируемые, как описано прежде, по количеству наблюдаемых инсерционно-делеционных мутаций.
Пример 17: Целенаправленная интеграция тринуклеотидных повторов с изменяющейся длиной
[0240] Htt-специфичные ZFN с наибольшей расщепляющей активностью для последовательностей, фланкирующих CAG-повтор, как описано выше, применяют в стратегии целенаправленной интеграции для введения переменных длин CAG-повтора в копию дикого типа Htt. Конструировали доноры, которые содержат 50, 80, 109 и 180 повторных единиц CAG. Эти доноры затем трансфицировали в клетки K562 с использованием плазмид, кодирующих Htt-специфичные ZFN, как описано выше. Верификации интеграции донора достигают посредством выделения геномной ДНК, амплификации при ПЦР (как описано выше) с последующим секвенированием области интереса.
[0241] ZFN, идентифицированные в клетках K562, которые приводят к целенаправленной интеграции донорных аллелей в аллель Htt, применяют для вставки донорных нуклеиновых кислот с переменной длиной в человеческие эмбриональные стволовые клетки (hESC). Успешную интеграцию донора верифицируют посредством выделения геномной ДНК, ПЦР и секвенирования, как описано выше.
Пример 18: Разрушение/выключение Htt дикого типа и/или мутантного Htt
[0242] ZFN, которые проводят расщепление в ранних экзонах, могут приводить к небольшим вставкам или делециям (вставкам-делециям), в результате негомологичного объединения концов (NHEJ), это может генерировать клеточные модели с разрушенными одним или обоими аллелями Htt.
[0243] Указанные пары ZFN получали, как описано выше, и тестировали на расщепляющую активность с использованием ошибочного спаривания нуклеотидов Cel I, как описано для Примера 8. Эти пары ZFN нацелены на ранние экзоны человеческого Htt, и, таким образом, могут применяться для выключения либо аллеля Htt дикого типа или мутантного аллеля Htt.
[0244] Как показано на Фиг. 14A, пары ZFP 29627/29628, 29631/29632 (экзон 12) и 29637/29638 (экзон 18) расщепляли ген Htt и могут, таким образом, использоваться для генерации выключенных линий клеток.
Пример 19: Экспрессионное мечение аллелей Htt дикого типа и аллелей БХ Htt.
[0245] ZFN с наибольшей расщепляющей активностью для первого или последнего кодирующего экзона применяют для мечения аллеля Htt дикого типа и мутантного аллеля Htt различными репортерными белками. Конструируют донорные ДНК для каждого репортера (A и B) на основании сайта расщепления лидерных пары (пар) ZFN для обеспечения целенаправленной интеграции репортерного гена, чтобы получить внутрирамочное слияние с Htt. Донорные ДНК со-трансфицируют с лидерными парой (парами) ZFN в клетки K562 для отбора конструкции донорной ДНК, которая дает наивысшую частоту интеграции.
[0246] Пары ZFN получали, как описано выше, и тестировали на расщепляющую активность с использованием ошибочного спаривания нуклеотидов Cel I, как описано для Примера 8. Для пар ZFN применяли целевой 3ʹ-конец последовательности, кодирующей Htt, и, таким образом, они могут применяться для целенаправленного действия на аллель Htt дикого типа или мутантный аллель Htt. Как показано на Фиг. 14B, пары ZFP 25917/25916, 25920/25921 и 25923/25922 были способны к расщеплению гена Htt, и могут, таким образом, использоваться для введения репортерной метки.
[0247] Отобранную конструкцию донорной ДНК для репортера A наряду с соответствующей ZFN доставляют в клетки, полученные от субъектов, несущих мутантный ген Htt (например, фибробласты, индуцированные плюрипотентные клетки). Клоны производят и скринируют на предмет целенаправленной интеграции репортера A. Гетерозиготные объекты являются желательными, и нацеленные аллели идентифицируют посредством ПЦР. Клоны, содержащие одиночный меченый репортером аллель Htt и немодифицированную целевую последовательность ZFN на другом аллеле, отбирают; донорную конструкцию для репортера B и соответствующую ZFN трансфицируют для мечения второго аллеля репортером B.
[0248] Полученный в результате клон мышиных эмбриональных стволовых клеток содержит аллель Htt дикого типа и мутантный аллель, меченные двумя различными маркерами, которые обеспечивают отслеживание экспрессии от каждого аллеля; эти клетки применяют для генерации мышиных моделей нарушений, связанных с тринуклеотидными повторами, с использованием стандартных протоколов.
Пример 20: Конструкция активных белков TALE-TF против Htt.
[0249] ДНК-связывающие домены TALE связывали с доменом репрессии KRAB из белка Kox1 (TALE TF) и применяли для тестирования репрессии гена Htt на фибробластах, полученных от пациента с БХ (CAG 20/41). Построение белков TALE проводили, как описано прежде (см. Патент США № 8,586,526 и Патентную публикацию США № 20130196373, оба из которых включены в данное описание посредством ссылки), и конструировали с тремя различными C-концевыми архитектурами: +63, +231 и +278, как описано в Патенте США № 8,586,526. Для построения плазмид экспрессии TALE TF, применяли плазмиды экспрессии TALEN, описанные прежде (см., Патент США № 8,586,526), за исключением того, что домен FokI, применяемый в TALEN заменяли на домен репрессии KRAB. Связки C-конца белка TALE и домена KRAB показаны ниже, где последовательность домена KRAB указана посредством подчеркивания. Текст с жирным шрифтом и выделенный курсивом указывает на тройной flag-маркер, Текст с жирным шрифтом указывает на ядерную локализацию последовательности, “[повторы]” указывает на положение матрицы единицы повтора TALE (полные повторы плюс C-концевой половинный повтор), и подчеркнутая волнистой линией часть показывает последовательность домена KRAB:
TALE-C63-Kox1:
MDYKDБХGDYKDБХIDYKDDDDKMAPKKKRKVGIHGVPMVDLRTLGYSQQQQEKIKPKVRSTVAQHHEALVGHGF
THAHIVALSQHPAALGTVAVKYQDMIAALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPPLQLDTGQLLKI
AKRGGVTAVEAVHAWRNALTGAPLN[повторы]GGRPALESIVAQLSRPDPALAALTNDHLVALACLGGRPALD
AVKKGLPHAPALIKRTNRRIPERTSHRVAGSGMDAKSLTAWSRTLVTFKDVFVDFTREEWKLLDTAQQIVYRNVM
LENYKNLVSLGYQLTKPDVILRLEKGEEPWLVEREIHQETHPDSETAFEIKSSV (SEQ ID NO:197)
TALE-C231-Kox1:
MDYKDБХGDYKDБХIDYKDDDDKMAPKKKRKVGIHGVPMVDLRTLGYSQQQQEKIKPKVRSTVAQHHEALVGHGF
THAHIVALSQHPAALGTVAVKYQDMIAALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPPLQLDTGQLLKI
AKRGGVTAVEAVHAWRNALTGAPLN[повторы]GGRPALESIVAQLSRPDPALAALTNDHLVALACLGGRPALD
AVKKGLPHAPALIKRTNRRIPERTSHRVADHAQVVRVLGFFQCHSHPAQAFDDAMTQFGMSRHGLLQLFRRVGVT
ELEARSGTLPPASQRWDRILQASGMKRAKPSPTSTQTPDQASLHAFADSLERDLDAPSPTHEGDQRRASSRKRSR
SDRAVTGPSAQQSFEVRAPEQRDALHLPLSWRVKRPRTSIGGGLPDPGSGMDAKSLTAWSRTLVTFKDVFVDFTR
EEWKLLDTAQQIVYRNVMLENYKNLVSLGYQLTKPDVILRLEKGEEPWLVEREIHQETHPDSETAFEIKSSV
(SEQ ID NO:198)
TALE-C278-Kox1
MDYKDБХGDYKDБХIDYKDDDDKMAPKKKRKVGIHGVPMVDLRTLGYSQQQQEKIKPKVRSTVAQHHEALVGHGF
THAHIVALSQHPAALGTVAVKYQDMIAALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPPLQLDTGQLLKI
AKRGGVTAVEAVHAWRNALTGAPLN[повторы]GGRPALESIVAQLSRPDPALAALTNDHLVALACLGGRPALD
AVKKGLPHAPALIKRTNRRIPERTSHRVADHAQVVRVLGFFQCHSHPAQAFDDAMTQFGMSRHGLLQLFRRVGVT
ELEARSGTLPPASQRWDRILQASGMKRAKPSPTSTQTPDQASLHAFADSLERDLDAPSPTHEGDQRRASSRKRSR
SDRAVTGPSAQQSFEVRAPEQRDALHLPLSWRVKRPRTSIGGGLPDPTPTAADLAASSTVMREQDEDPFAGAADD
FPAFNEEELAWLMELLPQGSGMDAKSLTAWSRTLVTFKDVFVDFTREEWKLLDTAQQIVYRNVMLENYKNLVSLG
YQLTKPDVILRLEKGEEPWLVEREIHQETHPDSETAFEIKSSV (SEQ ID NO:199)
[0250] Распознавание оснований достигалось с использованием соответствий канонических RVD-оснований ("код TALE”: NI для A, HD для C, NN для G (NK в половинном повторе), NG для T). В нескольких TALE TF, белок сконструирован для связывания со смысловой (5ʹ-3ʹ) цепью ДНК, в то время как в других, TALE TF сконструирован для связывания с антисмысловой (3ʹ-5ʹ) цепью. Этот набор из TALE TF был сконструирован для целенаправленного действия на CAG-повторы гена Htt. ДНК-связывающие белки TALE часто преимущественно взаимодействуют с ʹTʹ нуклеотидным основанием по 5ʹ-концу мишени, и, таким образом, поскольку мишенями являются области повтора CAG, можно предсказать, что белки, которые связываются с антисмысловой цепью ДНК, и, таким образом, последовательностью CTG-повтора с основанием ʹTʹ по 5ʹ-концу мишени, смогут иметь лучшую аффинность связывания и специфичность и, таким образом, репрессорную активность.
[0251] Мишени и числовые идентификаторы для тестированных TALE TF показаны ниже в Таблице 4. Числовые идентификаторы помечены “SBS#”, указана специфичность для Смысловой или Антисмысловой цепи (“S/A”), а также мишень, число единиц повтора или RVD и тип C-конца.
[0252] TALE TF в Таблице затем тестировали на предмет репрессии Htt в фибробластах, полученных от пациента с БХ (CAG 20/41), и результаты показаны на Фиг. 15. В этом эксперименте, клетки трансфицировали либо с 1000, 100 или 10 нг мРНК, кодирующих TALE-TF. Результаты для каждой проанализированной TALE TF показаны в группах по три, представляя три количества трансфицированной мРНК. В каждой группе также представлены три образца: левый столбик показывает полную экспрессию Htt, средний столбик показывает экспрессию от CAG20 аллеля Htt, и правый столбик показывает экспрессию от CAG41 аллеля Htt. Данные демонстрируют, что существуют несколько TALE TF, которые были способны подавлять оба аллеля Htt (см., например 102454), в то время как другие TALE TF были способно селективно ингибировать мутантный Htt с удлиненным повтором CAG (см., например 102451 и 102472).
[0253] Все патенты, патентные заявки и публикации, указанные в данном документе полностью включены в него посредством ссылки.
[0254] Несмотря на то, что раскрытие было представлено довольно подробно для иллюстрации и примеров с целью ясности понимания, для специалистов в данной области будет очевидным, что практически могут быть реализованы различные изменения и модификации без отступления от сущности или объема притязаний раскрытия. Соответственно, предшествующие описания и примеры не должны рассматриваться как ограничивающие.
Реферат
Изобретение относится к области биохимии, в частности к способу увеличения уровней по меньшей мере двух из PDE10a, DARPP-32, DRD1 и DRD2 в срединных шипиковых нейронах (MSN) субъекта-человека или субъекта-мыши. При этом способ включает введение в полосатое тело субъекту генетического репрессора, содержащего полинуклеотид, кодирующий ДНК-связывающий белок на основе «цинкового пальца» (ZFP), содержащий от 3 до 6 доменов «цинковых пальцев». Также раскрыт способ лечения болезни Хантингтона у субъекта-человека или субъекта-мыши, включающий увеличение уровней по меньшей мере двух из PDE10a, DARPP-32, DRD1 и DRD2 в срединных шипиковых нейронах (MSN) с помощью вышеуказанного способа. Изобретение позволяет эффективно осуществлять лечение болезни Хантингтона у субъекта-человека или субъекта-мыши. 2 н. и 7 з.п. ф-лы, 18 ил., 4 табл., 20 пр.
Комментарии