Аналог инсулина, обладающий активностью снижения уровня глюкозы в крови - RU2109749C1
Код документа: RU2109749C1
Чертежи

































Описание
Изобретение касается аналогов инсулина, модифицированного в положении 29 аминокислоты B-цепи естественного человеческого инсулина и при желании в других положениях. Эти аналоги инсулина в меньшей степени склонны к димеризации или самопроизвольной ассоциации с более высокомолекулярными формами, в результате чего они более быстро повышают свою активность, сохраняя при этом биологическую активность естественного человеческого инсулина.
Изобретение охватывает аналоги инсулина формулы (I), приведенной на фиг. 1, в которой A21 представляет собой аланин, аспарагин, аспарагиновую кислоту, глутамин, глутаминовую кислоту, глицин, треонин или серин; B1 представляет собой фенилаланин или аспарагиновую кислоту или отсутствует; B2 представляет собой валин или может отсутствовать при отсутствии B1; B3 представляет собой аспарагин или аспарагиновую кислоту; B9 представляет собой серин или аспарагиновую кислоту; B10 представляет собой гистидин или аспарагиновую кислоту; B28 представляет собой любую аминокислоту; B29 представляет собой L-пролин, D-пролин, D-оксипролин, L-оксипролин, L-(N-метиллизин), D-линиз, L-(N-метиларгинин) или D-аргинин; B30 представляет собой аланин, треонин или отсутствует; Z представляет собой -OH, -NH2, -COH3 или -CH2CH3; X представляет собой Arg, Arg-Arg, Lys, Lys-Lys, Arg-Lys, Lys-Arg или отсутствует; Y может присутствовать лишь тогда, когда присутствует X, и если он присутствует, то представляет собой Glu или аминокислотную последовательность, которая содержит всю или часть последовательности -Glu-Ala-Glu-Asp-Leu-Gln-Val-Gly-Gln-Val-Glu-Leu-Gly-Gly-Gly-Pro-Gly-Ala-Gly-Ser-Leu-Gln-Pro-Leu-Ala-Leu-Glu-Gly-Ser-Leu-Gln-Lys-Ary, который начинается у N-терминального Glu этой последовательности.
Описан и заявлен также способ лечения гипергликемии путем ввода в организм пациента, нуждающегося в таком лечении, эффективного количества аналога инсулина формулы (I). Кроме того, описаны и заявлены фармацевтические композиции, содержащие эффективное количество аналога формулы (I) в комбинации с одним или несколькими фармацевтически пригодными эксципиентами.
На фиг. 2 изображены ограничительная точка и функциональная карта плазмиды pKC283; на фиг. 3 - ограничительная точка и функциональная карта плазмиды pKC283PX; на фиг. 4 - ограничительная точка и функциональная карта плазмиды pKC283-L; на фиг. 5 - ограничительная точка и функциональная карта плазмиды pKC283-LB; на фиг. 6 - ограничительная точка и функциональная карта плазмиды pKC283-PRS; на фиг. 7 - ограничительная точка и функциональная карта плазмиды pL32; на фиг. 8 - ограничительная точка и функциональная карта плазмиды pNM789; на фиг. 9 схематически представлено построение плазмиды 120; на фиг. 10 изображены ограничительная точка и функциональная карта плазмиды pL47; на фиг. 11 - ограничительная точка и функциональная карта плазмида pPR12; на фиг. 12 - ограничительная точка и функциональная карта плазмиды pPR12AR1; на фиг. 13 - ограничительная точка и функциональная карта плазмиды pL110; на фиг. 14 и 15 схематически представлено построение плазмиды pL110C; на фиг. 16 изображены ограничительная точка и функциональная карта плазмиды pCZR126S; на фиг. 17 показана нуклеотидная последовательность гена синтезированного человеческого проинсулина; на фиг. 18 изображены ограничительная точка и функциональная карта плазмиды pPB145; на фиг. 19 - ограничительная точка и функциональная карта плазмиды pRB164A; на фиг. 20 - ограничительная точка и функциональная карта плазмиды pRB172; на фиг.21 - ограничительная точка и функциональная карта плазмиды pRB173; на фиг. 22 - ограничительная точка и функциональная карта плазмиды pRB175.
Формула (I) воспроизводит с помощью сокращенных названий аминокислотную последовательность аналогов инсулина, отвечающих данному изобретению. Сокращенные названия аминокислот могут быть расшифрованы, как указано ниже.
Аббревиатура - Аминокислота
Aba - Аминомасляная кислота
Ala - Аланин
Arg - Аргинин
Asn
- Аспарагин
Asp - Аспарагиновая кислота
Cys - Цистеиновая кислота
Cys - Цистеины
Gln - Глутамин
Glu - Глутаминовая кислота
Gly - Глицин
His - Гистидин
Il - Изолейцин
Leu - Лейцин
Lys - Лизин
Met - Метионин
Nle - Норлейцин
Nva - Норвалин
Orn - Орнитин
Phe
- Фенилаланин
Pro - Пролин
Ser - Серин
Thr - Треонин
Trp - Триптофан
Tyr - Тирозин
Val - Валин
B28 может представлять собой любую
аминокислоту природного или неприродного происхождения. Эта аминокислота является предпочтительно аспарагиновой кислотой, валином, лейцином, изолейцином, норлейцином, пролином, аргинином, гистидином,
цитруллином, орнитином, лизином, фенилаланином, аланином или глицином. Из указанных выше аминокислот особенно предпочтительной подгруппой является аспарагиновая кислота, валин, лейцин, изолейцин,
норлейцин, пролин, аргинин, гистадин, орнитин или лизин. Лизин является самой предпочтительной аминокислотой для B28. B29 представляет собой предпочтительно L-пролин, D-пролин, D-оксипролин или
L-оксипролин. Особенно предпочтительным аналогом инсулина согласно изобретению является такой, в котором B28 представляет собой лизин и B29 представляет собой пролин, то есть инверсию аминокислотной
последовательности естественного человеческого инсулина в положениях 28 и 29 B-цепи.
Согласно следующей особенности изобретения, как указывалось выше, когда B28 представляет собой L-пролин, то положение B29 представляет собой предпочтительно L-(N-метиллизин), L-лизин, L-(N-метиларгинин) или L-аргинин. Предпочтительным аналогом согласно данной особенности является такой, в котором B28 представляет собой L-пролин и B29 представляет собой L-(N-метиллизин) или D-лизин.
Ниже рассматриваются также другие модификации аналогов инсулина согласно изобретению, то есть модификации аналогов инсулина в положениях иных, чем положения B28 и B29. В частности, может быть желаемой замена группы Asn в положении 21 цепи A (то есть карбоксильной концевой группы) группами Ala, Asp, Gln, Glu,Gly, Thr или Ser, и при такой замене предпочтительной замещающей группой является Ala. Аналогично этому может быть желательной замена группы в положении 3 цепи B аспарагиновой кислотой (Asp). Такие возможные замены приводят к увеличению стабильности аналогов при экстремальных значениях pH, поскольку Asn особенно чувствителен к реакциям деамидирования и перегруппировки как при низком, так и при высоком значении pH. Для специалистов в данной области должно быть ясно также, что глутаминовые группы аналогов инсулина могут быть одинаковым образом чувствительны к реакциям деамидирования и перегруппировки. Таким образом, замещение их глутаминовой кислотой также охватывается объемом настоящего изобретения. Возможные другие модификации аналогов инсулина согласно изобретению включают (в любой комбинации) замену гистидиновой группы в положении B10 аспарагиновой аминокислотой, замену фенилаланиновой группы в положении A1 аспарагиновой кислотой, замену треониновой группы в положении B30 аланином, замену сериновой группы в положении B9 аспарагиновой кислотой, делецию аминокислот в положении B1 (des-B1) либо как таковых, либо в комбинации с делецией в положении B2 (des-B2) и делецию треонина из положения B-30 (des-B30).
Аналоги инсулина согласно изобретению могут быть также модифицированы в концевых положениях B30 путем ввода любых из следующих аминокислот или дипептидов: Arg, Arg-Arg, Lys, Lys-Lys, Arg-Lys или Lys-Arg. В случае их присутствия они обозначаются в формуле (I) как X. Предпочтительным из них является Arg-Arg.
Кроме того, в случае наличия этих добавленных звеньев в положении B30 аналог может быть дополнительно модифицирован путем добавления к образуемой B30-удлиненной концевой группе глутаминовой кислоты (Glu) или аминокислотной последовательности, включающей всю или часть последовательности -Glu-Ala-Glu-Asp-Leu-Gln-Val-Gly-Gln-Val-Glu-Leu-Gly-Gly-Gly-Pro-Gly-Ala- Gly-Ser-Leu-Gln-Pro-Leu-Ala-Leu-Glu-Gly-Ser-Leu-Gln-Lys-Arg-, которая начинается в ее N-терминальной - Glu. Эта аминокислота или последовательность в случае ее присутствия обозначена в формуле (I) группой Y. Когда эта последовательность представляет лишь часть указанной выше, то она может представлять собой любую их тех частей, которые начинаются у N-конца данной последовательности, то есть у группы глутаминовой кислоты (Glu).
X или комбинация X и Y, которые определены выше, представляет собой полностью или часть соединяющего пептида, обнаруживаемого в человеческом проинсулине, молекуле, которая является биологическим источником образования естественного человеческого инсулина.
Предпочтительными последовательностями для Y являются
-Glu-Ala-Glu-Asp-Leu-Gln-Val-Gly-Gln-Val-Glu- Leu-Gly-Gly-Gly-Pro-Gly-Ala-Gly-Ser-Leu-Gln-Pro-Leu-Ala- Leu-Glu-Gly-Ser-Leu-Gln-Lys-Arg-;
-Glu-Ala-Glu-Asp-Leu-Gln-Val-Gly-Gln-Val-Glu- Leu-Gly-Gly-Gly-Pro-Gly-Ala-Gly-Ser-Leu-Gln-Pro-Leu-Ala- Leu-Glu-Gly-Ser-Leu-Gln-Les-;
Glu-Ala-Glu-Asp-Leu-Gln-Val-Gly-Gln-Val-Glu- Leu-Gly-Gly-Pro-Gly-Ala-Gly-Ser-Leu-Glu-Pro-Leu-Ala- Leu-Glu-Gly-Ser-Leu-Gln-;
-Glu-Ala-Glu-Asp-Leu-Gln-Val-Gly-Gln-Val-Glu- Leu-Gly-Gly-Gly-Pro-Gly-Ala-Gly-Ser-Leu-Gln-Pro-Leu-Ala- Leu-Glu-;
-Glu-Ala-Glu-Asp-Leu-Gln-Val-Gly-Gln-Val-Glu- Leu-Gly-Gly-Gly-Pro-Gly-Ala-Gly-Ser-Leu-Gln-Pro-Leu-Ala-;
-Glu-Ala-Glu-Asp-Leu-Gln-Val-Gly-Gln-Val-Glu-.
Кроме того, независимо от того, присутствует или отсутствует X отдельно или присутствуют или отсутствуют X и Y совместно, концевая группа Z может быть любой из числа следующих: -OH -NH2 -OCH3 или -OCH2CH3. Предпочтительным значением группы Z является -OH.
Как уже упоминалось выше, изобретение охватывает фармацевтически пригодные соли аналогов инсулина. Такими предпочтительными солями являются соли цинка, натрия, калия, магния и кальция.
Аналоги инсулина, отвечающие данному изобретению, могут быть получены одним из известных приемов синтеза пептидов, включая классические способы (в растворе), способы в твердофазных системах, способы полусинтеза и известные с недавнего времени способы рекомбинантной ДНК.
В способах с использованием твердофазных систем аминокислотная последовательность составляется из исходной нерастворимой C-терминальной аминокислоты со смоляным носителем. Эти твердофазные способы описываются T.Stewart и др. Твердофазный синтез пептидов. Freeman and Co., Сан-Франциско, 1969 г.
Обычно при осуществлении твердофазного способа аминокислота, соответствующая C-терминальной аминокислотной группе желаемого пептида, закрепляется на смоляном носителе и далее формируется пептидная цепь, начинающаяся с нанесенной на смолу C-терминальной аминокислоты. Отдельные аминокислоты вводятся последовательно до тех пор, пока не получается желаемая аминокислотная последовательность. Как возможный вариант могут быть получены небольшие пептидные фрагменты, которые могут быть введены в пептидную цепь в желаемом порядке. Пептидная цепь остается скрепленной со смолой в процессе синтеза, и по завершении построения цепи данный пептид отщепляется от смолы.
Пептидная цепь закрепляется с полистирольной смолой посредством сложноэфирной связи, образующейся между карбоксильной группой C-терминального звена и специфической метиленовой группой, присутствующей в смоляной матрице, являющейся точкой такого скрепления.
Аминокислоты связываются сопряженными связями хорошо известными в данной области приемами образования пептидных связей.
Один такой прием заключается к конверсии аминокислоты в производное, которое придает карбоксильной группе более высокую чувствительность к реакции со свободной N-терминальной аминогруппой пептидного фермента. Так, например, аминокислота может быть превращена в смешанный ангидрид путем реакции защищенной аминокислоты с этилхлорформатом, фенилхлорформатом, вторбутилхлорформатом или изобутилхлорформатом. Как возможный вариант аминокислота может быть превращена в активный сложный эфир, такой как 1,4,5-трихлорфениловый сложный эфир, пентахлорфениловый сложный эфир, пара-нитрофениловый сложный эфир, N-оксисукцинимидный сложный эфир или сложный эфир, образуемый из l-оксибензотриазола.
Другой прием сопряжения заключает в себе использование подходящего агента, такого как N,N'-дихлоргексилкарбодиимид (ДСС) или N,N'-диизопропилкарбодиимид (DIC). Другими подходящими сопрягающими агентами являются соединения, которые уже известны для специалистов в данной области (см. работу Schroder и Lubke "Пептиды", Academic Press, 1965 г. Глава III).
Следует иметь в виду, что α-аминогруппа каждой аминокислоты, используемой в синтезе пептидов, должна быть защищена в ходе реакции сопряжения для предотвращения побочных реакций, в которых принимает участие α-аминовая функциональная группа. Следует также иметь в виду, что некоторые аминокислоты содержат боковые цепи, являющиеся функциональными группами (например, сульфгидрил, ε-амино, β- и γ - карбоксил, имидазол, гуанидино и гидрокси), и что такие функциональные группы также должны быть защищены в начальном и в последующих этапах реакции сопряжения. Подходящие защитные группы известны в данной области (см. , например, публикацию "Защитные группы в органической химии". M.McOmie. Ред. Plenum Press, N 4, 1973, и патент США N 4617149).
При выборе типа защитной группы следует соблюдать определенные условия. Защитная для α -амино группа должна придавать функциональной α - аминогруппе инертность в условиях, имеющих место в реакции сопряжения, должна быть легко удаляемой после реакции сопряжения в таких условиях, в которых не удаляются защитные группы боковой цепи, не должна изменять структуру пептидного фрагмента и должна исключать возможность рацемизации при активации непосредственно перед реакцией сопряжения. Защитная группа боковой цепи должна придавать этой функциональной боковой цепи инертность в условиях, используемых при протекании реакции сопряжения, должна быть устойчивой в условиях, имеющих место при удалении защитной для α - амино группы, и должна легко удаляться по завершении построения желаемой аминокислотной последовательности в таких условиях реакции, которые не изменяют структуру пептидной цепи.
Для специалистов в данной области должно быть ясно, что защитные группы, которые уже известны как группы, применяемые для синтеза пептидов, будут по разному активны к агентам, используемым для их удаления. Так, например, некоторые защитные группы, такие как трифенилметил и 2-(пара-бифенил)изопропилоксикарбонил, очень неустойчивы и могут расщепляться в слабо кислотных условиях. Другие защитные группы, такие как трет-бутилоксикарбонил, трет-аминоксикарбонил, адаматнилоксикарбонил и пара-метоксибензилоксикарбонил, менее устойчивы и требуют использования умеренно сильных кислот, таких как трифторуксусная кислота, соляная кислота или трифтор бора в уксусной кислоте, для их удаления. Некоторые защитные группы, такие как бензилоксикарбонил, галоилбензилоксикарбонил, пара-нитробензилоксикарбонил, циклоалкилоксикарбонил и изопропилоксикарбонил, еще менее устойчивы и для их удаления требуется использование еще более сильных кислот, таких как фтористоводородная кислота, бромистоводородная кислота или трифторацетат бора в трифторуксусной кислоте.
По завершении желаемого построения последовательности пептида защищенный пептид должен быть отщиплен от смоляного носителя и все защищающие группы должны быть удалены. Реакция отщепления и удаление защитных групп могут осуществляться одновременно или поэтапно. Когда смоляной носитель представляет собой хлорметилированную полистироловую смолу, то связь, скрепляющая пептид со смолой, является сложноэфирной связью, образуемой между свободной карбоксильной группой C-терминального звена и одной из многих хлорметильных групп, присутствующих в смоляной матрице.
Следует иметь в виду, что указанная скрепляющая связь может быть расщеплена посредством реагентов, которые, как известно, способны разрывать сложноэфирную связь и проникать в смоляную матрицу. Особенно легко осуществимым способом является обработка жидким безводным фтористым водородом. Этот реагент не только отщепляет пептид от смолы, но и удаляет все защитные группы. Следовательно, использование этого реагента может обеспечить получение полностью лишенного защиты пептида. Когда желательно удаление пептида без удаления защитных групп, то защищенный пептид - смола может подвергаться метанолизу, в результате чего получается защищенный пептид, в котором C - терминальная карбоксильная группа подвергнута метилированию. Этот сложный метиловый эфир затем может быть гидролизован в слабо щелочных условиях, и в результате будет получен свободный C-терминальный карбоксил. Защитные группы пептидной цепи могут быть удалены путем обработки сильной кислотой, такой как жидкая фтористоводородная кислота. Особенно интересный способ метанолиза описывается в работе G.Moore и др. Пептиды. Труды 5-го Американского симпозиума по пептидам. M.Goodman и J.Meinhofer, ред. John Wiley, N 9., 1977 г., с. 518-521, где замещенный пептид-смола обрабатывается метанолом и цианидом калия в присутствии простого кроун-эфира.
Следующий способ отщепления защищенного пептида от смолы заключается а аминолизе или обработке гидразином. При желании образуемый C-терминальный амид или гидразид может быть гидролизован до свободного C-терминального карбоксила, и защитные группы могут быть удалены общепринятым образом.
Следует также иметь в виду, что защитные группы, находящиеся у N-терминальной α-аминогруппы, удаляются предпочтительно либо до, либо одновременно с отщеплением защищенного пептида от смоляного носителя.
A- и B-цепи аналогов инсулина, отвечающие данному изобретению, также могут быть получены методом рекомбинантной ДНК. При этом получается нуклеотидная последовательность, кодирующая желаемый пептид A- или B- цепи, с использованием общепринятой технологии для такого синтеза. Эти методы обычно заключают в себе получение олигонуклеотидов, кодирующих как фрагменты желаемой кодирующей последовательности, так и их полную последовательность. Олигонуклеотиды обеспечивают перекрытие одного фрагмента кодирующей последовательности двумя фрагментами дополняющей последовательности и наоборот. Эти олигонуклеотиды конъюгируют и связываются с образованием в конечном итоге желаемой генной последовательности.
Эта последовательность вставляется в клонирующий вектор в точке, которая допускает возможность выражения пептидного продукта, который он кодирует. Подходящий клонирующий вектор заключает в себе, по меньшей мере, часть контрольной последовательности выражения гена.
Цепи A и B аналогов инсулина согласно изобретению могут быть также получены посредством исходной молекулы проинсулинового типа с использованием методов рекомбинантной ДНК (см. Frank и др. Пептиды: Синтез - Структура - Функции. Труды 7-го Американского симпозиума по пептидам. Ред. D.Rich и E. Gross (1981 г.).
Однако этап комбинирования отдельных получаемых цепей A и B может осуществляться способом, описанным в работе Chance и др., Пептиды: Синтез, структура и функции. Труды 7-го Американского симпозиума по пептидам, 1981 г.
Для иллюстрации данного изобретения даются нижеследующие примеры, которые, однако, не служат для ограничения объема изобретения.
Пример 1. Человеческий инсулин Pro (B29), Lys (B28).
A. Получение образованной от рекомбинанты A-цепи.
A-цепь человеческого инсулина получается методом рекомбинантной ДНК путем химического синтеза гена, кодирующего A-цепь, и его выражения в E.Coli. Ген для A-цепи синтезируется из различных тринуклеотидов, синтетические фрагменты, составляющие по длине от дека до пентадекануклеотидов, посредством метода получения блок-фосфотриэфира. Этот ген имеет однониточный когезионный конец для ограничительных эндонуклеаз Eco RI и Bam HI. Вставка его в подходящий выраженный вектор, содержащий β-галактозный ген (β-gae), приводит к химерной плазмиде, содержащей A-цепь, связанную с геном β-gae через метиониновый кодон. Для достижения более высоких уровней выражения используется предпочтительно ген триптофан синтетазы (trp LE') в качестве промотора, а не β -gae-ген. Химерная плазмида трансформируется в E.Coli, приводящей в результате к выражению исходного белка (белка предшественника), то есть β -gae-met A-цепи (или trp LE'-met A-цепи, когда используется система промотора trp LE'). Обработка исходного белка цианогенбромидом приводит к расщеплению метиониновой связи с получением после очистки A-цепи человеческого инсулина. Окислительный сульфитолиз приводит к образованию S-сульфонированной A-цепи, которая используется для комбинации с B-цепью (S-сульфонат), как описано ниже.
Подробное описание химического синтеза генов для A-человеческого инсулина дается в публикации Crea и др. Proc. Natl., Acad. Sci., США, 75, 57655769, 1978. Для получения полных сведений о выражении в E.Coli химически синтезированного гена для A-цепи человеческого инсулина см. публикацию Goeddel и др. Proc. Natl. Acad. Sci., США, 76, 101-110, 1979.
B. Получение аналога B-цепи [Lys (B28), Pro (B29)].
Для получения сырой пептидиловой смолы используется синтезатор пептида Applied Biosystems 430A (включая модификацию A4). Используется 0,5 ммоль исходной твердофазной смолы (t-BOC-Thr (BZl) OCH2 Pam смолы) (0,76 ммоль/г•0,658 г). Все аминокислоты являются защищенными BOC, и, кроме глутаминовой кислоты и гистидина, все аминокислоты используются непосредственно в том виде, как они получены (то есть в упаковках, содержащих примерно от 2 ммоль защищенной аминокислоты, поставляемых фирмой Applied Biosystems Inc). Глутаминовая кислота и гистидин поставляются фирмой Peptides International Corporation, и они заключаются в такие упаковки, что каждая содержит примерно 2 ммоль желаемой защищенной аминокислоты. После сушки сырой пептидиловой смолы (в вакууме при комнатной температуре в течение ночи) определяется ее вес и сопоставляется с начальным весом для гарантии нужного привеса. Небольшая часть образца подвергается аминокислотному анализу для гарантии того, что желаемые аминокислоты вводятся в точно заданных количествах.
Пептид отщепляется от пептидиловой смолы, и защита с боковой цепи удаляется в результате перемешивания примерно в течение 1 ч при 0oC в растворе 10 частей (об./вес.) HF (содержащего 5% об./об. этилмеркаптана и 5% об./об. метакрезола) с 1 частью пептидиловой смолы. После вакуумной отгонки большей части HF пептид осаждается в простом этиловом эфире. После нескольких промывок простым этиловым эфиром с последующей вакуумной фильтрацией пептид растворяется примерно в 200 мл 7 М деионизированной воды, содержащей 0,1 моль Трис, 0,1 моль Na2SO3 и 0,01 моль Na2S4O6. Величина pH раствора доводится до 8,5 посредством 5 н. NaOH, и этот раствор интенсивно перемешивается в течение ночи при температуре 4oC.
Образующийся S-сульфонированный пептидный раствор вводится в колонку размерами 5 • 215 см, наполненную Sephadex G-25 (тонкоизмельченным) при комнатной температуре. Образец элюируется 50 мМ бикарбонатом аммония с расходом его 20 мл/мин при комнатной температуре. Выходящий поток анализируется (при 276 нм). Фракции по 20 мл собираются, и получают объем желаемых фракций, который затем очищается дополнительно методом жидкостной хроматографии высокого разрешения (HPLC), как описано ниже.
Слитные желаемые фракции нагнетаются в колонку 2,5 • 30 см с Iu Pont C-8, 9-12 мкм (колонка HPLC), и осуществляется элюирование с использованием линейного ингредиента с увеличивающейся концентрацией ацетонитрила в 100 мл бикарбоната аммония при комнатной температуре (2,6 мл/мин). Выходящий из колонки поток анализируется (280 нм). Фракции по 25 мл собираются. Выбранные фракции HPLC подвергаются анализу для определения того, какие фракции остаются. Желаемые фракции собираются, лиофилизируются и используются в последующей комбинации с A-цепью, полученной, как описано выше.
C. Получение человеческого инсульта Pro (B 29), Lys (B 28).
Комбинация цепи A и B осуществляется согласно процедуре, описанной в публикации Chance и др., см. выше. 700 мг S-сульфоната A-цепи, образованного от рекомбинантной ДНК, и 140 мг синтетического S-сульфоната B-цепи Lys (B28), Pro (B29) (оба получены, как описано выше) каждый растворяются в 70 мл и 14 мл соответственно 0,1 М буферного раствора глицина и величина pH каждого доводится до 10,5 посредством 4 н. NaOH, затем они охлаждают до 5oC. Приготавливается 6 мл раствора дитиотреитола (ДТТ) концентрацией 10,5 мг/мл в 0,1 М глициновом буферном растворе при комнатной температуре, величина pH этого раствора доводится до 10,5 посредством 5 н. NaOH, затем этот раствор охлаждается до 5oC.
Эти растворы A-цепи и B-цепи смешиваются друг с другом, после чего в эту смесь быстро вводится 5,21 мл раствора ДТТ (SH/SSO3 = 0,90). Этот реакционный раствор перемешивается при температуре 5oC в открытых склянках 200-миллилитровых пробирках для центрифуги в течение 2,5 ч при 5oC. Вводится 45 мл ледяной уксусной кислоты, и раствор выстаивается при температуре 5oC в течение ночи.
Полученная смесь с осадком центрифугируется в течение 20 мин со скоростью 2000 об/мин при температуре 5oC. Поверхностный слой смешивается с 1 М уксусной кислотой, промывается от твердых частиц, вводится в колонку 5 • 200 см с Sephadex G-50 (сверхтонкого) в 1 М уксусной кислоте при 5oC и элюируется при ускорении силы тяжести. Двадцатиминутные фракции элюирования собираются в течение 3 дн. Эти фракции анализируются (при 276 нм), и некоторые образцы анализируются методами аналитической HPLC. Фракции, содержащие последовательность Lys (B 28), Pro (B 29) аналога инсулина, сливаются и лиофилизируются, давая 125-миллиграммовый образец. Этот образец дополнительно очищается методом жидкостной хроматографии высокого разрешения (HPLC) с обратимой фазой (с использованием колонки 2,12 • 25 см с Iu Pont C-8 при элюировании при комнатной температуре со скоростью 2,6 мл/мин с использованием линейного градиента с увеличивающейся концентрацией ацетонитрила в 0,1 М NaH2 PO4, pH 2,2). Выходящий поток анализируется (при 276 нм). Выбранные фракции анализируются аналитическим методом HPLC. Желаемые фракции собираются и дополнительно очищаются с использованием pH 7 HPLC, как описано ниже.
Слитный продукт от HPLC низкого pH разбавляется примерно в два раза в ледяной бане 0,1 М (NH4)2HPO4. Величина pH доводится до 7 посредством холодной 3 н. NaOH в ледяной бане. Этот образец вводится в ту же колонку и элюируется из нее в тех же условиях, что и при приготовлении образца с низким pH, с той разницей, что элюирующий буферный раствор в данном случае представляет собой 0,1 М (pH 7) (NH4)2HPO4/ацетонитрил.
Слитный продукт от HPLC с pH 7 быстро охлаждается в ледяной бане и в два раза разбавляется 0,1%-ным водным раствором трифторуксусной кислоты (TFA). Вводится 1 н. HCl (холодная, образец в ледяной бане) для снижения величины pH до 3. Образец вводится в колонку Vydae C4 или, как возможный вариант, в колонку с Iu Pont C-8 HPLV (2,12 • 25 см), и осуществляется элюирование линейным градиентом с повышающейся концентрацией ацетонитрила в 0,1%-ном водном растворе TFA. Выходящий из колонки продукт анализируется (при 214 или 276 нм). Желаемые фракции сливаются и лиофилизируются, давая образец в количестве 41 мг желаемого аналога со степенью чистоты более 97 %, достигаемой путем HPLC с обратимой фазой.
Пример 2. Человеческий инсулин Pro (B29), Lys (B28).
Второй способ получения человеческого инсулина Pro (B29), Lys (B28) заключается в использовании ферментного полусинтеза (обратимый протеолиз) для комбинирования des- октапептидного инсулина (A1-21-B1-22) с синтетическим октапептидом. des-Октапептидный инсулин получается в результате триптического переваривания естественного свиного или человеческого инсулина, как описано в работе Bromer и Chance "Получение и свойства les-октапептид-инсулина", Biochem. Biophys Acta 133, 219-223, (1967 г). Синтетический октапептид Gly-Phe-Phe-Tyr-Tyr-Lys-Pro-Thr получается методом автоматического твердофазного синтеза так, как описано выше.
les-октапептидный инсулин (435 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Lys-Pro-Thr (465 мг) смешиваются в 15 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть 0,25 М буферного раствора Трис-ацетата, pH 7,3. Пептиды полностью растворяются путем нагревания раствора на горячей плите. Затем раствор инкубируется при 37oC и добавляется 90 мг свиного трипсина. Раствор время от времени перемешивается в течение 90 мин при температуре 37oC. Реакция прекращается путем смешивания раствора с 135 мл 0,05 н. HCl.
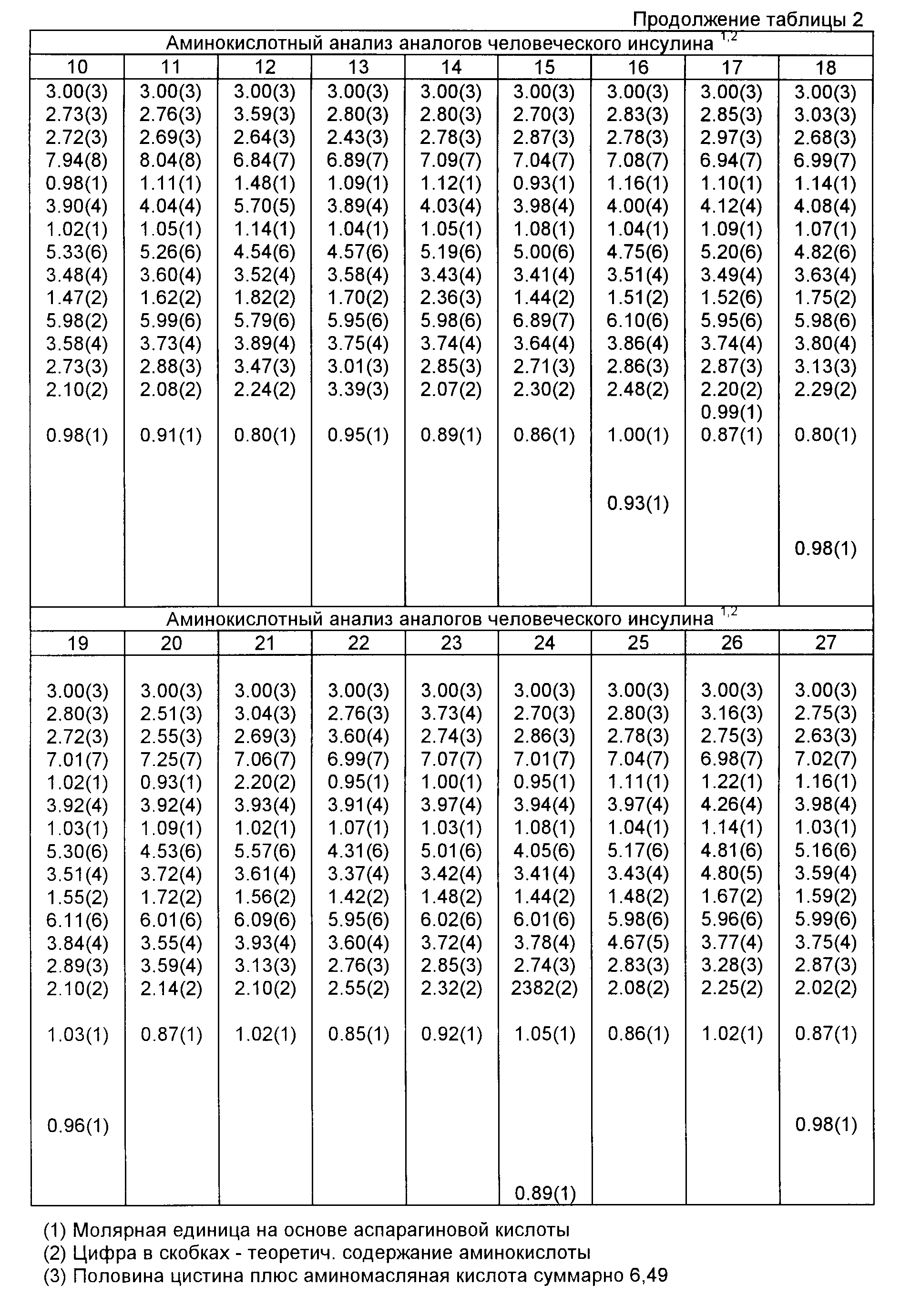
Указанный в названии аналог инсулина очищается путем ввода подкисленного раствора, содержащего этот аналог, в колонну HPLC размерами 2,5 • 25 см, наполненную C-8 Zorbax, и элюирования линейным градиентом с повышающейся концентрацией ацетонитрила в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2,2. Выходящий на колонки продукт анализируется (при 276 нм). Желаемые фракции сливаются, в два раза разбавляются водой и вводится в колонку HPLC размерами 1 •25 см с C-8 Ultrasphere. Этот аналог элюируется линейным градиентом с увеличивающейся концентрацией ацетонитрила в 0,5%-ном водном растворе TFA. Выходящий из колонки продукт анализируется (при 276 нм). Желаемые фракции снова сливаются и лиофилизируются, давая 125 мг очищенного аналога. Данная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5809,2 (теоретически: 5808,7).
Осуществляется анализ методом быстрой атомной бомбардировки - масс-спектроскопии с использованием масс-спектрометра с двойной фокусировкой VG-AB-25E, с разрешением примерно 1500. Аналоги человеческого инсулина растворяются в смеси глицерина и триглицерина с содержанием щавелевой кислоты. Для калибровки измерительного прибора, который осуществляет магнитное сканирование в диапазоне от m/Z 5300 до m/Z 6500, используется иодид цезия. Полученные данные представлены как среднемассовое значение +1.
Пример 3. Человеческий инсулин Pro (B29), Aba (B28).
Свиной des-октапептидный инсулин (384 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Aba-Pro-Thr (362 мг) смешиваются в 13 мл раствора, содержащего диметилсульфоксид, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при температуре 37oC. Вводится свиной трипсин (75 г). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается в результате ввода этой смеси в 137 мл 0,05 н. HCl. Весь этот раствора вводится в колонку размерами 21 • 250 мм с C-8 Zorbax, и продукты элюируются при небольшом ацетонитриловом градиенте в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Соответствующие фракции, которые определены методом аналитической HPLC, сливаются, в два раза разбавляются водой и вводятся в колонку 25 • 300 мм с C-18 Vydae. Желаемый белок элюируется из колонки ацетонитрильным градиентом в 0,05%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются с получением выхода 59 мг. Указанная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5765,7 (теоретически: 5765,6).
Пример 4. Человеческий инсулин Pro (B 29), Ala (B28).
Свиной des-октапептидный инсулин (290 мг) и систематический октапептид Gly-Phe-Phe-Tyr-Thr-Ala-Pro-Thr (310 мг) смешиваются в 10 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3. Вводится свиной трипсин (60 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 60 мин при 37oC. В этот момент времени реакция прекращается за счет ввода смеси в 90 мл 0,05 н. HCl. Весь раствор вводится в колонку размерами 21 • 250 мм с C-8 Zorbax, и продукты элюируются в небольшом ацетонитрильном градиенте в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Соответствующие фракции, которые определены методом аналитической HPLC, сливаются, в два раза разбавляются водой и вводятся в колонку размерами 10•250 мм с C-8 Ultrasphere. Желаемый белок элюируется из колонки с использованием ацетонитрильного градиента в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются с получением выходы 43 мг. Указанная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5752,3 (теоретически: 5751,6).
Пример 5. Человеческий инсулин Pro (B29), Arg (B28).
Свиной des-октапептидный инсулин (290 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Arg-Pro-Thr (310 нг) смешивают в 10 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной инсулин (60 мг). Раствор смешивают и время от времени тщательно перемешивается в течение 60 мин при 37oC.
В этот момент времени реакция прекращается в результате ввода этой смеси в 90 мл 0,05 н. HCl. Весь раствор вводится в колонку размерами 21•250 мм с C-8, и продукты элюируются в небольшом ацетонитрильном градиенте в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Соответствующие фракции, которые определены аналитической HPLC, сливаются, в два раза разбавляются водой и вводятся в колонку 10•250 мм с C-8 Ultrasphere. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, и получается 103 мг продукта. Указанная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектрометрическим анализом (MS). MS: 5836,1 (теоретически: 5836,7).
Пример 6. Человеческий инсулин Pro (B29), Asn (B28).
Свиной des-октапептидный инсулин (409 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Asn-Pro-Thr (398 мг) смешиваются в 14 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1, 4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,9, при 37oC. Вводят свиной трипсин (81 мг). Раствор смешивают и время от времени тщательно перемешивают в течение 120 мин при 37oC. В этот момент времени реакция прекращается за счет ввода смеси в 136 мл 0,5 н. HCl. Весь раствор вливается в колонку 21•250 мм с C-8 Zorbax, и продукты элюируются в небольшом ацетонитрильном градиенте в буферном растворе 0,2 М одноосновного фосфата натрия, pH 2.
Соответствующие фракции, которые определены посредством аналитической HPLC, сливаются, в два раза разбавляются водой и вводятся в колонку размерами 25•300 мкм с C-18 Vydae. Желаемый белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается 56 мг продукта. Указанная структура подтверждается аминокислотным аналогом (табл. 1) и масс-спектроскопией (MS). MS: 5794,7 (теоретически: 5794,6).
Пример 7. Человеческий инсулин Pro (B29), Asp (B28).
Свиной des-октапептидный инсулин (400 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Asp-Pro-Thr (388 мг) смешиваются в 13 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3 при 37o C. Вводится свиной трипсин (78 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин. при 37oC.
Реакция прекращается в данный момент времени путем ввода этой смеси в 137 мл 0,05 н. HCl. Весь раствор вводится в колонку 21•250 мм с C-8 Zorbax, и продукты элюируются небольшим ацетонитрильным градиентом в буферном растворе 0,1 М одноосновного фосфата натрия pH 2.
Соответствующие фракции, которые определены аналитической HPLC, сливаются, разбавляются в четыре раза водой и вводятся в колонку размерами 25• 300 мм с C-8 Vydae. Желаемый белок элюируется из колонки ацетонитрильным градиентом в 0,1%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливают и лиофилизируются, в результате получается 85 мг продукта. Указанная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5795,7 (теоретически: 5795,6).
Пример 8. Человеческий инсулин Pro (B29), Asp (B10), Lys (B28).
A. Получение B-цепи человеческого инсулина Pro (B29), Asp (B10), Lys (B28).
Для получения сырой пептидной смолы используется пептидный синтезатор Applied Biosystems 430 A (включая модификацию 1,4). Используется 0,5 ммоль исходной твердофазной смолы (t-BOC-Thr (BZI) OCH2Pam смолы) (0,72 ммоль/г • 0,705 г). Все используемые аминокислоты представляют собой защищенные BOC, за исключением глутаминовой кислоты, аспарагиновой кислоты и гистидина, все аминокислоты используются в том виде, как они получены (то есть в упаковках, поставляемых фирмой Applied Biosystems Inc.; каждая упаковка содержит примерно 2 ммоль защищенной аминокислоты). Глутаминовая кислота, аспарагиновая кислота и гистидин являются промышленно получаемыми кислотами, они заключаются в упаковки таким образом, что каждая упаковка содержит примерно 2 ммоль желаемой защищенной аминокислоты. Сырая пептидиловая смола высушивается в вакууме при комнатной температуре в течение ночи и ее вес сопоставляется с весом исходной смолы для гарантии нужного привеса. Небольшая часть образца подвергается аминокислотному анализу для гарантии того, что аминокислоты вводятся в правильных пропорциях.
Пептид отщепляется от пептидиловой смолы, и защита боковой цепи удаляется в результате перемешивания в течение примерно 1 ч при 0oC в растворе 10 частей (об. /вес. ) HF (содержащего 5% об./об. пара-тиокрезола и 5% об./об. метакрезола) с 1 частью пептидиловой смолы. После удаления наибольшей части HF при вакуумной отгонке пептид осаждается в простом этиловом эфире. После нескольких промывок простым этиловым эфиром с последующей вакуумной фильтрацией пептид растворяется примерно в 120 мл 8 М гуанидина в HCl, pH 11, содержащего 0,1 моль Трис, 35 мг/мл Na2SO3 и 25 мг/мл Na2S4O9. Величина pH раствора доводится до 8,8 посредством 5 н. NaOH, и раствор интенсивно перемешивается в течение 3 ч при комнатной температуре.
Полученный S-сульфонированный пептидный раствор вводится в колонку 5•215 см с Sephadex G-25 при комнатной температуре. Образец элюируется со скоростью 21 мл/мин при комнатной температуре с использованием 50 ммоль бикарбоната аммония. Выходящий из колонки продукт анализируется (при 276 нм). Фракции по 25 мл собираются и сливаются, и собранный продукт из желаемых фракций дополнительно очищается путем жидкостной хроматографии высокого разрешения (HPLC), как описано ниже.
Слитный продукт, состоящий из желаемых фракций, вводится в хроматографическую колонку 2,5•30 см с Iu Pont C-8, 9-12 мкм, и элюирование осуществляется с использованием линейного градиента с увеличивающейся концентрацией ацетонитрила в 100 ммоль бикарбоната аммония при комнатной температуре (2,6 мл/мин). Выходящий из колонки продукт анализируется при 280 нм. Фракции (по 25 мл) собираются и сливаются. Отобранные фракции подвергаются анализу методом аналитической HPLC для определения того, какие фракции сохраняются. Желаемые фракции сливаются, лиофилизируются и используются в указанной ниже комбинации с A-цепью.
B. Комбинация B-цепи человеческого инсулина Pro (B29), Lys (B28), Asp (B10) с A-цепью человеческого инсулина.
Комбинация A- и B-цепей осуществляется согласно процедуре, описанной в работе Chance и др., см. выше.
Два грамма S-сульфоната A-цепи, образованного от рекомбинантной ДНК, и 400 мг синтетического S-сульфоната B-цепи Pro (B29), Asp (B10), Lys (B28) каждого растворяются в 200 мл и 40 мл соответственно 0,1 М буферного раствора глицерина при комнатной температуре, величина pH каждого доводится до 10,5 посредством 5 н. NaOH и затем каждый охлаждается до 5oC. Приготавливается раствор дитиотретиола (ДТТ) концентрацией 15,5 мг/мл в 0,1 М буферном растворе глицина при комнатной температуре, величина pH доводится до 10,5 посредством 5 н. NaOH, и затем раствор охлаждается до 5oC.
Растворы A-цепи и B-цепи смешиваются, и затем в эту смесь быстро вводится 15,9 мл раствора ДТТ (su/SSO=1) . Реакционный раствор перемешивается при 4oC в открытых 200-миллилитровых стеклянных пробирках для центрифугирования в течение 19,6 ч при 4oC. Вводится ледяная уксусная кислота (129 мл), и раствор выстаивается при 4oC в течение 1 ч.
Полученная смесь с выпавшим осадком центрифугируется в течение 30 мин со скоростью 2000 об/мин при 4oC. Поверхностный слой смешивается с 292 мл милли-Q-воды и 73 мл ацетонитрила, и эта смесь дополнительно очищается посредством HPLC с обратимой фазой (с использованием колонки 2,5 • 30 см с Vydae C18 с элюированием при комнатной температуре со скоростью 2,6 мл/мин с использованием линейного ингредиента с увеличивающейся концентрацией ацетонитрила в 0,1 М NaH2PO4, pH 2,1). Выходящий из колонки продукт анализируется при 280 нм. Выбранные фракции анализируются методом аналитической HPLC, и желаемые фракции сливаются и разбавляются в два раза 0,1%-ным водным раствором трифторуксусной кислоты (TFA), затем слитый продукт вводится в хроматографическую колонку (0,1 • 25 см) с Ultrasphere, и осуществляется элюирование линейным градиентом с увеличивающейся концентрацией ацетонитрила в 0,1%-ном водном растворе TFA. Выходящий из колонки продукт анализируется при 280 нм. Выбранные фракции анализируются методом аналитической HPLC, и желаемые фракции сливаются и повторно очищаются с использованием той же колонки и в тех же условиях, что указаны выше, при несколько отличающемся градиенте ацетонитрила. Соответствующие фракции сливаются и лиофилизируются, в результате получается 65 мг аналога инсулина со степенью чистоты более чем 93% в результате HPLC с обратимой фазой. Указанная структура подтверждается аминокислотным анализом (табл. 1) и методом масс-спектроскопии (MS). MS: 5786,1 (теоретически: 5786,7).
Пример 9. Человеческий инулин Pro (B29), Cya (B28).
Осуществляется окисление надмуравьиной кислоты с целью превращения цистеина откапептида в форму цистеиновой кислоты. Синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Lys-Pro-Thr (363 мг) растворяется в 36 мл свежеприготовленной надмуравьиной кислоты в охлажденной льдом колбе, и раствор осторожно перемешивается в течение 1 ч. Окисленный материал разбавляется в десять раз водой и лиофилизируется. В данном полусинтезе используется этот лиофилизат.
Свиной des-октапептидный инсулин (222 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Cya-Pro-Thr (225 мг) смешивают в 18 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (45 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается в результате ввода этой смеси в 242 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 см с C-8 Zorbax, и продукты элюируются слабым ацетонитрильным градиентом в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Подходящие фракции, которые определены методом аналитической HРLC, сливаются, разбавляются в два раза водой и вводятся в колонку 25 • 300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки с использованием ацетонитрильного градиента в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается выход продукта 16 мг. Указанная структура продукта подтверждается аминокислотным анализом (табл. 1) и методом масс-спектроскопии (MS). MS: 5831,5 (теоретически: 5831,7).
Пример 10. Человеческий инсулин Pro (B29), Gln (B28).
Свиной des-октапептидный инсулин (290 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Gln-Pro-Thr (310 мг) смешиваются в 10 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (60 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 60 мин при 37oC.
В этот момент времени реакция прекращается в результате ввода смеси в 90 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zordax, и продукты элюируются с использованием слабого ацетонитрильного градиента в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Подходящие фракции, которые определены путем аналитической HPLC, сливаются, разбавляются в два раза водой, вводятся в колонку размерами 10 • 250 мм с C-8 Ultraphase. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, получается выход продукта 87 мг. Указанная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5809,4 (теоретически: 5808, 6).
Пример 11. Человеческий инсулин Pro (B29), Glu (B28).
Свиной des-октапептидный инсулин (402 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Glu-Pro-Thr (398 мг) смешиваются в 14 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, при 37oC. Вводится свиной трипсин (80 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается путем ввода этой смеси в 136 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются в слабом ацетонитрильном градиенте в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Подходящие фракции, которые определены путем аналитической HPLC, сливаются, разбавляются в четыре раза водой и вводятся в колонку 25 • 300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5 %-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается выход продукта 59 мг. Данная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5809,6 (теоретически: 5809,6).
Пример 12. Человеческий инсулин Pro (B29), Gly (B28).
Свиной des-октапептидный инсулин (412 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Gly-Pro-Thr (376 мг) смешиваются в 13 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1, 4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3. Вводят свиной трипсин (79 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 180 мин при 37oC.
В этот момент времени реакция прекращается путем ввода смеси в 147 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются слабым ацетонитрильным градиентом в 0,1 М буферном растворе одноосновного фосфата натрия, pH 2.
Подходящие фракции, которые определены аналитической HPLC, сливаются, разбавляются в четыре раза водой и вводятся в колонку 25 • 300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,1%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается 11 мг продукта. Данная структура продукта подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5737,2 (теоретически: 6537,6).
Пример 13. Человеческий инсулин Pro (B29), His (B28).
Свиной des-октапептидный инсулин (400 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-His-Pro-Thr (398 мг) смешиваются в 13 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (79 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается путем ввода смеси в 237 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zordax, и продукты элюируются в слабом ацетонитрильном градиенте в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Подходящие фракции, которые определены аналитической HPLC, сливаются, разбавляются в четыре раза водой, вводятся в колонку 25 • 300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,1%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается выход продукта 79 мг. Данная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5816,9 (теоретически: 5817,7).
Пример 14. Человеческий инсулин Pro (B29), Ile (B28).
Свиной des-октапептидный инсулин (409 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Ile-Pro-The (398 мг) смешиваются в 13 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводят свиной трипсин (81 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается путем ввода смеси в 136 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются в слабом ацетонитрильном градиенте в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Подходящие фракции, определяемые аналитической HPLC, сливаются, разбавляются в два раза водой и вводятся в колонку 25 • 300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0, 5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный инсулиновый аналог, сливаются и лиофилизируются, в результате получается выход продукта 57 мг. Данная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5793,7 (теоретически: 5793,7).
Пример 15. Человеческий инсулин Pro (B29), Leu(B28).
Свиной des-октапептидный инсулин (418 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Leu-Pro-Thr (410 мг) смешиваются в 14 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (83 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37o C.
В этот момент реакция прекращается путем ввода этой смеси в 136 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются в слабом ацетонитрильном градиенте в 0,1 М буферном растворе одноосновного сульфата натрия, pH 2.
Соответствующие фракции, определяемые аналитической HPLC, сливаются, разбавляются в четыре раза водой и вводятся в колонку 25 • 300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается выход продукта 74 мг. Структура этого продукта подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5793,8 (теоретически: 5793,7).
Пример 16. Человеческий инсулин Pro (B29), Nle (B28).
Свиной des-октапептидный инсулин (290 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Nle-Pro-Thr (310 мг) смешиваются в 10 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводят свиной трипсин (60 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 60 мин при 37oC.
В этот момент реакция прекращается путем ввода смеси в 90 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются в слабом ацетонитрильном градиенте в 0,1 М буферном растворе одноосновного фосфата натрия, pH 2. Подходящие фракции, которые определены аналитической HPLC, сливаются, разбавляются в два раза водой и вводятся в колонку 10 • 259 мм с C-8 Ultraphere. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие инсулиновый аналог, сливаются и лиофилизируются, в результате получается выход продукта 54 мг. Указанная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5794,6 (теоретически: 5793,7).
Пример 17. Человеческий инсулин D-Lys (B29).
Свиной des-октапептидный инсулин (392 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Pro-D-Lys-Thr (387 мг) смешиваются в 23,5 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводят свиной трипсин (78 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент реакция прекращается путем ввода смеси в 136,5 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются в слабом ацетонитрильном градиенте в 0,1 М буферном растворе одноосновного сульфата натрия, pH 2.
Подходящие фракции, определяемые аналитической HPLC, сливаются, разбавляются в четыре раза водой и вводятся в колонку 25 • 300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается 94 кг продукта. Данная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5809,0 (теоретически: 5808,7).
Пример 18. Человеческий инсулин Pro (B29), Met (B28).
Свиной des-октапептидный инсулин (350 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Met-Pro-Thr (366 мг) смешиваются в 12 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7, при 37o C. Вводится свиной трипсин (71 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В данный момент времени реакция прекращается путем ввода смеси в 118 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются в слабом ацетонитрильном градиенте в 0,1 М буферном растворе одноосновного фосфата натрия, pH 2.
Подходящие фракции, определяемые аналитической HPLC, сливаются, разбавляются в четыре раза водой и вводятся в колонку 25 • 300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,1%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается 72 мг продукта. Указанная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5811,0 (теоретически: 5811,7).
Пример 19. Человеческий инсулин Pro (B29), Orn (B28).
Свиной des-октапептидный инсулин (290 мг) и синтетический октапептид Gly-Phe-Phe-Thr-Orn-Pro-Thr (310 мг) смешиваются в 10 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (60 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 90 мин при 37oC.
В этот момент времени реакция прекращается путем ввода данной смеси в 90 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются в слабом ацетонитрильном градиенте в 0,1 М буферном растворе одноосновного фосфата натрия, pH 2.
Подходящие фракции, определяемые аналитической HPLC, сливаются, разбавляются в четыре раза водой и вводятся в колонку 10 • 250 мм с C-8 Ultraphase. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный инсулиновый аналог, сливаются и лиофилизируются, в результате получается 89 мг продукта. Структура продукта подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5795,2 (теоретически: 5794,7).
Пример 20. Человеческий инсулин Pro (B29), Phe (B28).
Свиной des-октапептидный инсулин (290 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Phe-Pro-Thr (310 мг) смешивают в 10 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (60 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 80 мин при 37o C.
В этот момент времени реакция прекращается путем ввода смеси в 90 мл 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются в слабом ацетонитрильном градиенте в 0,1 М буферном растворе одноосновного фосфата натрия, pH 2.
Подходящие фракции, определяемые аналитической HPLC, сливаются, разбавляются в четыре раза
водой и вводятся в колонку 25 • 300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,1%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог
инсулина, сливаются и лиофилизируются, в результате получается 17 мг продукта. Структура продукта подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5827,9 (теоретически:
5827,7)
Пример 21. Человеческий инсулин Pro (B29).
Свиной des-октапептидный инсулин (339 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Pro-Pro-Thr (363 мг) смешивают в 9 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3 при 37oC. Вводится свиной трипсин (70 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 80 мин при 37oC.
Данная реакция в этот момент времени прекращается путем ввода смеси в 108 мл 0,05 н. HCl. Весь раствор вводится в колонку 10 • 250 мм с C-8 Zorbax, и продукты элюируются слабым ацетонитрильным градиентом в 0,1 М буферном растворе одноосновоного фосфата натрия, pH 2.
Подходящие фракции, которые определены аналитической HPLC, сливаются, разбавляются два раза водой и вводятся в колонку 10 • 250 мм с C-8 Ultrasphere. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается 97 мг продукта. Структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5778,6 (теоретически: 5777,6).
Пример 22. Человеческий инсулин Pro (B29), Ser (B28).
Свиной des-октапептидный инсулин (412 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Ser-Pro-Thr (390 мг) смешиваются в 13 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1, 4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (80 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В данный момент времени реакция прекращается путем ввода смеси в 137 мг 0,05 н. HCl. Весь раствор вводится в колонку 21 • 250 мм с C-8 Zorbax, и продукты элюируются в слабом ацетонитрильном градиенте в 0,1 М буферном растворе одноосновного фосфата натрия, pH 2.
Подходящие фракции, определяемые аналитической HPLC, сливаются, разбавляются в два раза водой и вводятся в колонку 25х300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, собираются и лиофилизируются с получением выхода продукта 37 мг. Структура продукта подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5768,1 (теоретически: 5767,6).
Пример 23. Человеческий инсулин Pro (B29), Thr (B28).
Свиной des-октапептидный инсулин (437 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Thr-Pro-Thr (420 мг) смешиваются в 14,5 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводят свиной трипсин (86 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается путем ввода смеси в 135,5 мл 0,05 н. HCl. Весь раствор вводится в колонку 21х250 мм с C-8 Lorbax, и продукты элюируются небольшим ацетонитрильным градиентом в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Подходящие фракции, определяемые аналитически HPLC, сливаются, в два раза разбавляются водой и вводятся в колонку 25х300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, получается выход продукта 78 мг. Данная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5781,9 (теоретически: 5781,6).
Пример 24. Человеческий инсулин Pro, (B29), Trp (B28).
Свиной des-октапептидный инсулин (310 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Thr-Trp-Pro-Thr (325 мг) смешиваются в 10,5 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,28 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (64 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается путем ввода смеси в 140 мл 0,05 н. HCl. Весь раствор вводится в колонку 21х250 мм с C-8 Lorbax, и продукты элюируются слабым ацетонитрильным градиентом в 0,1М буферном растворе одноосновного фосфата натрия, pH 2.
Подходящие фракции, которые определены аналитической HPLC, сливаются, в два раза разбавляются водой и вводятся в колонку размерами 25х300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный инсулин, сливаются и лиофилизируются, в результате получается выход продукта 47 мг. Данная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5866,2 (теоретически 5866,7).
Пример 25. Человеческий инсулин Pro (B29), Tyr (B28).
Свиной des-октапептидный инсулин (391 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Thr-Tyr-Pro-Thr (400 мг) смешиваются в 13 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0, 25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (79 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается путем ввода смеси в 137 мл 0,05 н. HCl. Весь раствор вводится в колонку размерами 21х250 мм с C-8 Zorbax, и продукты элюируются слабым ацетонитрильным градиентом в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Подходящие фракции, которые определены аналитической HPLC, сливаются, разбавляются в два раза водой и вводятся в колонку размерами 25х300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается выход продукта 30 мг. Данная структура подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5843,7 (теоретически 5843,7).
Пример 26. Человеческий инсулин Pro (B29), Tyr (B28).
Свиной des-октапептидный инсулин (400 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Tyr-Pro-Thr (383 мг) смешиваются в 12 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1,4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (78 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается путем ввода смеси в 238 мл 0,05 н. HCl. Весь раствор вводится в колонку 21х250 мм с C-8 Zorbax, и продукты элюируются слабым ацетонитрильным градиентом в 0,1 М буферном растворе одноосновного фосфата натрия, pH 2.
Подходящие фракции, которые определены аналитической HPLC, сливаются, разбавляются в четыре раза водой и вводятся в колонку размерами 25х300 с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,1%-ной трифторуксусной кислоте. Фракции, содержащие очищенный аналог инсулина, сливаются и лиофилизируются, в результате получается выход продукта 74 мг. Структура продукта подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5780,0 (теоретически: 5799,6).
Пример 27. Человеческий инсулин Pro (B29), Nva (B28).
Свиной des-октапептидный инсулин (292 мг) и синтетический октапептид Gly-Phe-Phe-Tyr-Thr-Nva-Pro-Thr (279 мг) смешиваются в 10 мл раствора, содержащего 1 часть диметилсульфоксида, 2 части 1, 4-бутандиола и 1 часть буферного раствора 0,25 М Трис, pH 7,3, при 37oC. Вводится свиной трипсин (57 мг). Раствор тщательно смешивается и время от времени перемешивается в течение 120 мин при 37oC.
В этот момент времени реакция прекращается путем добавления смеси в 240 мл 0,05 н. HCl. Весь раствор вводится в колонку 21х250 мм с C-8 Zorbax, и продукты элюируются слабым ацетонитрильным градиентом в буферном растворе 0,1 М одноосновного фосфата натрия, pH 2.
Подходящие фракции, которые определены аналитической HPLC, сливаются, разбавляются в два раза водой и вводятся в колонку размерами 25х300 мм с C-18 Vydae. Обессоленный белок элюируется из колонки ацетонитрильным градиентом в 0,5%-ной трифторуксусной кислоте. Фракции, содержащие очищенный инсулиновый аналог, сливаются и лиофилизируются, в результате достигается выход продукта 51 мг. Структура продукта подтверждается аминокислотным анализом (табл. 1) и масс-спектроскопией (MS). MS: 5870,0 (теоретически: 5779,6).
Пример 28. Осуществляя процедуры так, как описано в данной заявке, получают нижеследующие дополнительные аналоги
инсулина:
(a) Человеческий инсулин Asp(B1), Lys(B28), Pro(B29);
(b) Человеческий инсулин des(Phe-B1), Lys(B28), Pro(29);
(c) Человеческий инсулин des(Phe-B1), Asp(B10),
Lys(B28), Pro(B29);
(d) Человеческий инсулин des(Phe-B1, Val-B2), Lys(B28), Pro(B29);
(e) Человеческий инсулин des(Phe-B1, Val-B2), Asp(B10), Lys(B28), Pro(B29);
(f)
Человеческий инсулин Gly(A21), Asp(B10), Lys(B28), Pro(B29);
(g) Человеческий инсулин Ala(A21), Asp(B10), Lys(B28), Pro(B29);
(h) Человеческий инсулин des(Thr-B30), Lys(B28),
Pro(B29);
(i) Человеческий инсулин Asp(B10), Arg(B28), Pro(B29);
(j) Человеческий инсулин Ala(A21), Arg(B28), Pro(B29);
(k) Человеческий инсулин Asp(B1), Arg(B28),
Pro(B29).
Пример 29. Человеческий инсулин Pro (B29), Lys (B28).
A. Построение плазмиды pCP126.
1. Выделение плазмиды pKC283.
Лиофилы E.Coli K12 BE1201/pKC283 получены из Северной Региональной Научно-исследовательской Лаборатории, Пеория, Иллинойс 61604, где они имеют официальный номер NRRL B-15830. Эти лиофилы декантируются в пробирки, содержащие 10 мл среды LB (10 г Бакто-триптона, 5 г Бакто-дрожжевого экстракта и 10 г NaCl на литр; pH доводится до 7,5), и инкубируются в течение 2 ч при 32oC, в течение этого времени культуры доводятся до концентрации 50 мкг/мл в ампициллине и затем инкубируются при 32oC в течение ночи. Клетки E.Coli K12 BE1201 / pKC283 выращивают при 32oC, поскольку они содержат температурочувствительный подавляющий ген, интегрированный в клеточную ДНК. При использовании в процедурах выделения плазмиды (как описано в нижеследующих примерах) клеток, которые содержат подавляющий ген лямбда p дикого типа или не содержат промотора лямбда p, температура инкубации составляет 37oC.
Небольшая часть выращенной за ночь культуры помещается в чашки с LB-агаром (среда LB с 15 г/л Бакто-агара), содержащие 50 мкг/мл ампициллина, так, чтобы получился один единственный изолят колонки E.Coli K12 BE1201/pKC283. Эта единственная полученная колония инокулируется в 10 мл среды LB, содержащей 50 мкг/мл ампициллина, и инкубируется в течение ночи при 32oC при интенсивном встряхивании. 10 мл ночной культуры инокулируется в 500 мл среды LB, содержащей 50 мкг/мл ампициллина, и инкубируется при 32oC с интенсивным встряхиванием до тех пор, пока культура не достигает стационарной фазы.
Принята нижеследующая процедура, описанная в работе Maniatis и др., 1982 г., Moulular Cloning (Cold Spring Harbor Laboratory).
Клетки собираются путем центрифугирования с ускорением 4000 g в течение 10 мин при 4oC, и поверхностный слой удаляется. Клеточный осадок промывается в 100 мл охлажденного льдом буферного раствора STE (0,1 моль NaCl, 10 ммоль Трис-HCl, pH 7,8, и 1 ммоль ЭДТА). После промывки клеточный осадок повторно суспензируется в 10 мл раствора 1 (50 ммоль глюкозы, 25 ммоль Трис-HCl, pH 8, и 10 ммоль ЭДТА), содержащего 5 мг/мл лизозима, и оставляется при комнатной температуре в течение 10 мин. Затем в обработанные лизозимом клетки вводится 20 мл раствора 2 (2 н. NaOH и 1% SDS), и раствор подвергается осторожному инверсионному перемешиванию. Смесь инкубируется на льду в течение 10 мин. 15 мл охлажденного льдом 5 М ацетата калия, pH 4,8, вводятся в лизированную клеточную смесь, и раствор подвергается инверсионному перемешиванию. Приготавливается 5 М раствор ацетата калия путем ввода 11,5 мл ледяной уксусной кислоты в 28,5 мл воды и 60 мл 5 М ацетата калия, полученный раствор является 3 М в отношении концентрации калия и 5 М в отношении концентрации ацетата.
Лизированная клеточная смесь центрифугируется в центрифуге Бекмана SW 27 (или его эквивалента) со скоростью 20000 об/мин в течение 20 мин при температуре 4oC. Клеточная ДНК и клеточные продукты распада образуют осадок на дне пробирки. Извлекается примерно 36 мл поверхностного слоя и вводится 0,6 объема изопропанола, осуществляется перемешивание, и полученный раствор выдерживается при комнатной температуре в течение 15 мин. Плазмидная ДНК извлекается путем центрифугирования с ускорением 12000 g в течение 3 мин при комнатной температуре. Поверхностный слой декантируется, и осадок ДНК промывается 70%-ным этанолом при комнатной температуре. Затем этанольная промывка декантируется, и осадок высушивается в эксикаторе. Этот осадок снова суспендируется в 8 мл буферного раствора TE (10 мл Трис-HCl, pH 8, и 1 мл этилендиаминтетраацетата (ЭДТА).
В раствор ДНК вводится 8 г CaCl2, вводится примерно 0,8 мл раствора этилиденбромида в воде на каждые 10 мл раствора CaCl2-ДНК. Конечная плотность этого раствора составляет примерно 1,55 г/мл, и концентрация этилиденбромида составляет примерно 600 мкг/мл. Данный раствор подается в пробирку для центрифугирования типа Бекмана 50, наполняется сверху парафиновым маслом, уплотняется и центрифугируется со скоростью 45000 об/мин в течение 24 ч при 20oC. После центрифугирования в обычном свете наблюдаются два слоя ДНК. После удаления крышки с пробирки нижний слой ДНК удаляется посредством шприца с гиподермической иглой #21, вставляемой в пробирку для центрифугирования сбоку.
Этиленбромид удаляется путем нескольких экстракций l-бутанолом, насыщенным водой. CaCl2 удаляется путем диализа с TE-буфером. После экстракций буферным раствором фенола, а затем хлороформом ДНК осаждается, промывается 70%-ным этанолом и высушивается. Получается примерно 1 мг плазмиды pKC283, и она выдерживается при 4oC в буферном растворе ТЕ концентрацией примерно 1 мкг/мкл. Ограничительная точка и функциональная карта плазмиды pKC283 представлены на фиг. 2.
2. Построение плазмиды pKC283PX.
Примерно 10 мкл ДНК плазмиды pKC283, приготовленной как описано в примере 1, смешивается с 20 мкл 10X среднесолевого ограничительного буфера (500 ммоль NaCl, 100 ммоль Трис-HCl, pH 7,5, 100 ммоль MgCl2 и 10 ммоль ДТТ), 20 мкл BSA (1 мг/мл), 5 мкл ограничительного фермента PVU11 (≈50 ед., как определено в лаборатории Bethesda Research Laboratories (BEL), из которой все эти ограничительные ферменты получены) и 145 мкл воды, и эта реакционная смесь инкубируется при 37oC в течение 2 ч. Реакции ограничительного фермента, описанные в данной заявке, завершаются обычным образом путем экстракции фенолом, а затем хлороформом, после чего происходит осаждение ДНК, который промывается этанолом и снова суспензируется в буферном растворе ТЕ. После завершения выпаривания с Pvu, как описано выше, вываренная с PvuII плазмидная pKC283 ДНК осаждается и затем снова суспензируется в 5 мкл буферного раствора ТЕ.
Примерно 500 пмоль связывающих звеньев XhoI (5'-CCTCGAGG-3') киназируются в смеси, содержащей 10 мкл 5X киназного буферного раствора (300 ммоль Трис-HCl, pH 7,8, 50 ммоль MgCl2 и 25 ммоль ДТТ), 5 мкл 5 М ATP, 24 мкл H2O, 0,5 мкл полинуклеотидокиназы Т4 (примерно 2,5 единиц, как определено в P-L Biochemicals), 5 мкл 1 мг/мл BSA и 5 мкл 10 мМ спермидина, путем инкубирования этой смеси при 37oC в течение 30 мин.
Примерно 12,5 мкл киназированных связующих звеньев вводятся в 5 мкл вываренных с PvuII ДНК плазмиды pKC283, и затем 2 мкл 10X буферного раствора лигазы (300 ммоль Трис-HCl, pH 7,6, 100 ммоль MgCl2 и 50 ммоль ДТТ), 2,5 мкл BSA концентрацией 1 мг/мл, 7 мкл 5 мМ ATP, 2,5 мкл лигазы Т4 (ДНК примерно 2,5 ед., как определено P-L Biochemicals), 2,5 мкл 10 мМ спермидина и 3 мкл воды вводятся в указанную ДНК. Полученная реакционная смесь сшивки инкубируется при 4oC в течение ночи. После реакции сшивки состав реакционной смеси регулируется таким образом, чтобы получился высокосолевой буферный раствор (0,1 моль NaCl, 0,05 моль Трис-HCl, pH 7,5, 10,0 ммоль MgCl2 и 1 ммоль ДТТ). В смесь вводится примерно 10 мкл (100 ед.) ограничительного фермента XhoI, и полученная реакционная смесь инкубируется при 37oC в течение 2 ч.
Реакция завершается, и вываренная с XhoI ДНК осаждается, повторно суспензируется и сшивается, как описано выше, с той разницей, что никаких связывающих звеньев XhoI не вводится в смесь сшивки. Сшитая ДНК состоит из желаемой плазмиды pKC283PX. Ограничительная точка и функциональная карта плазмиды pKC283PX представлены на фиг. 3.
3. Построение E.Coli K12 MO ( λ+ )/pKC283PX.
E. Coli K12 MO ( λ+ ) может быть получена из Северных Региональных Научных Лабораторий в лиофилизированной форме под официальным номером NRRL B-15993. E. Coli K12 MO включает подавляющий ген лямбда PL данного типа, так что транскрипции от гибридного промотора pL-Ipp, отвечающего данному изобретению, не происходит в клетках E.Coli K12 MO ( λ+ ). Осуществляется выделение лиофилов и перестроенных одиночных колоний MO ( λ+ ), и приготавливается 10 мл ночной культуры клеток MO ( λ+ ) в основном согласно процедуре, описанной в примере 29 A1, с той разницей, что температура инкубации составляет 37oC и в среде роста не используется никакого ампициллина.
50 мкл ночной культуры используется для инкуляции 5 мл среды LB, которая также содержит 2 ммоль MgSO4 и 10 ммоль MgCl2. Данная культура инкубируется при 37oC в течение ночи при интенсивном встряхивании. На следующее утро культура разбавляется до 200 мл средой LB, содержащей 10 ммоль MgSO4 и 10 ммоль MgCl2. Разбавленная культура инкубируется при 37oC при интенсивном встряхивании до тех пор, пока поглощение при 550 мм (А550) не составит примерно 0,5, что показывает клеточную плотность примерно 1•10 клеток/мл. Эта культура охлаждается в течение 10 мин в ледяной водяной бане, и затем клетки извлекаются путем центрифугирования при ускорении 4000 g в течение 10 мин при 4oC. Клеточный осадок на дне пробирки снова суспензируется в 100 мл холодного 10 мМ MgSO4 и затем тотчас же повторно гранулируется путем центрифугирования. Клеточный осадок снова суспензируется в 100 мл 30 мМ CaCl2 и инкубируется во льду в течение 20 мин.
Эти клетки снова собираются путем центрифугирования и снова суспензируются в 10 мл 30 мМ CaCl2. Аликвота этих клеток, составляющая пол-мл, вводится в сшитую ДНК, приготовленную как в примере 29 A2, приготавливается ДНК концентрацией 30 ммоль в CaCl2. Смесь клетки - ДНК инкубируется во льду в течение 1 ч, нагревается при 42oC в течение 90 с и затем охлаждается на льду в течение 2 мин. Смесь клетки - ДНК разбавляется в 10 мл среды LB и 125-миллилитровых колбах и инкубируется при 37oC в течение 1 ч. Аликвоты в количестве 100 мкл высеваются в чашках с LB-агаром, содержащих ампициллин, и инкубируются при 37oC до тех пор, пока не появляются колонии.
Эти колонии индивидуально культивируются, и ДНК плазмиды отдельных колоний подвергаются анализу на ограничительный фермент и гель-электрофорезу. Выделение ДНК плазмиды осуществляется в небольших количествах согласно процедуре примера 29 A1, но этап градиента CsCl не осуществляется до тех пор, пока интифицируются E.Coli K12 MO (λ+) pKC283PX трансформанты. Ограничительная точка и функциональная карта плазмиды pKC283PX представлены на фиг. 3.
4. Построение E.Coli K12 MO (λ+) /pKC283-L.
10 мкг ДНК плазмиды pKC283PX, полученной согласно процедуре примера 29 A1, растворяются в 20 мкл 10X высокосолевого буферного раствора, 20 мкл 1 мг/мл BSA, 5 мкл (≈50 ед.) ограничительного фермента BgeII, 5 мкл (≈50 ед.) ограничительного фермента XhoI и 150 мкл воды, и полученная реакционная смесь инкубируется при 37oC в течение 2 ч. Реакция прекращается и после осаждения вываренной с BgeII - XhoI ДНК эта ДНК снова суспензируется в 5 мкл буферного раствора TE.
Синтезируется и киназируется связывающее звено ДНК с однониточными концами ДНК, характерными для расщепления ограничительного фермента BgeII и XhoI. Это связующее звено киназируется согласно процедуре примера 29 A2. Связывающее звено ДНК имеет структуру, приведенную на фиг. 23.
Приведенное связующее звено синтезируется из однониточных деоксилигонуклеотидов согласно уже хорошо известной в данной области процедуре. Эти однониточные деоксилигонуклеотиды могут быть синтезированы посредством выпускаемых промышленностью инструментов, таких как синтезатор ДНК 380 A, выпускаемых в продажу фирмой Applid Biosystems 1850 Lincoln Centre Irive, Foster City (A 94404), который используется в химии фосфорамидитов. Известны также и другие процедуры синтеза ДНК. Обычный модифицированный метод фосфотриэфира для синтеза однониточной ДНК описывается в работе Itakura и др., 1977, Science, 198 : 1056 и Crea и др., 1978, Proc. Nat. Acad. Sci. USA, 75 : 5765. Кроме того, особенно предпочтительный способ синтеза ДНК описывается в работе Hsiung и др., 1983, Nucleic Acid Resеarch, 11, 3227; и в работе Narang и др., 1980, Methods in Enzymology, 68 : 90.
Это связующее звено и вываренная с BgeII - XhoI плазмида pKC283PX сшиваются в основном согласно процедуре примера 29 A2. Сшитая ДНК образует желаемую плазмиду pKC283-L. Ограничительная точка и функциональная карта плазмиды pKC283-L представлены на фиг. 4. ДНК плазмиды pKC283-L используется для трансформации E. Coli K12 MO (λ+), и полученные трансформанты E.Coli E12 MO (λ+)/pKC283-L идентифицируются в основном согласно процедуре примера 29 A3.
5. Построение E.Coli K12 MO (λ+) / pKC283-LB.
Примерно 10 мкг ДНК плазмиды pKC283-L, полученной согласно процедуре примера 29 A1, растворяются в 20 мкл 10X высокосолевого буферного раствора, 20 мкл 1 мг/мл BSA, 5 мкл (≈50 ед.) ограничительного фермента XhoI и 155 мкл H2O, и полученная реакционная смесь инкубируется при 37oC в течение 2 ч. Вываренная с XhoI ДНК плазмиды pKC283-L затем осаждается из реакционной смеси путем ввода трех объемов 95%-ного этанола и 1/10 объема 3 М ацетата натрия, инкубации в бане сухой лед - этанол в течение 5 мин и центрифугирования. Полученный осадок ДНК промывается 70%-ным этанолом, высушивается и повторно суспензируется в 2 мкл 10X Ник-трансляционного буферного раствора (0,5 моль Трис-HCl, pH 7,2, 0,1 моль MgSO4 и 1 ммоль ДТТ), 1 мкл раствора 2 мМ в каждом из деоксинуклеотидотрифосфатов, 15 мкл H2O, 1 мкл (≈6 ед., как определено P-L Biochemical) фрагмента Кленова, который является большим фрагментом 1 ДНК полимеразы E. Coli, и 1 мкл 1 мг/мл BSA. Полученная реакционная смесь инкубируется при 25oC в течение 30 мин, реакция прекращается в результате инкубирования раствора при 70oC в течение 5 мин.
Связывающие звенья Bam HI (5'- CCGGATCCCG-/3') киназируются и сшиваются с вываренной с XhoI, обработанной фрагментом Кленова ДНК плазмиды pKC283-L в основном согласно процедуре примера 29 A2. После реакции сшивки ДНК вываривается примерно с 100 ед. Bam AI в течение ≈2 ч при 37oC в высокосолевом буферном растворе. После вываривания с Bam AI ДНК подготавливается для сшивки согласно процедуре примера 29 A2.
Ограничительный фрагмент ≈5,9 кб Bam HI замыкается в кольцо путем сшивки и трансформируется в E.Coli K12 MO (λ+) согласно процедуре примера 29 A2 и 29 A3. Трансформанты K12 MO (λ+) / pKC283-LB идентифицируются, и затем получается плазмида pKC283-LB согласно процедуре примера 29 AI. Ограничительная точка и функциональная карта плазмиды pKC283-LB представлены на фиг. 5.
6. Построение E.Coli K12 MO (λ+) / pL32.
Примерно 10 мкг плазмиды pKC283PX вываривается с ограничительным фрагментом SalI в высокосолевом буферном растворе, обрабатывается фрагментом Кленова и сшивается со связывающими звеньями Eco RI (5'-GAGGAATTCCTC-3') в основном согласно процедуре примера 29 A5, с той разницей, что используются исходная плазмида, ограничительные фрагменты и связывающие звенья. После вываривания с ограничительным фрагментом Eco RI, который образует в результате фрагмент ≈2,1 Lb от ДНК, ограничительный фрагмент ≈4 кб Eco RI замыкает кольцо путем сшивки с получением плазмиды pKC283pPS. Эта сшитая ДНК используется для трансформации E.Coli K12 MO (λ+) в основном согласно процедуре примера 29 A3. После идентификации трансформант E. Coli K12 MO (λ+) / pKC283pPS ДНК плазмиды pKC283PRS обрабатывается согласно процедуре примера 29 A1. Ограничительная точка и функциональная карта плазмиды pKC283PRS представлены на фиг. 6.
Примерно 10 мкл плазмиды pKC283PRS вываривается в 200 мкл высокосолевого буферного раствора примерно с 50 ед. каждого из ограничительных фрагментов PstI и SphI. После инкубирования реакционной смеси при 37oC в течение примерно 2 ч реакционная смесь подвергается электрофорезу на 0,6% гель-агарозе с низкой температурой гелеобразования (фирма Отделение Marine Colloids Division, Rockland, Maine 04841) в течение 2 - 3 ч при ≈130 B и ≈75 мА в Трис-ацетатном буферном растворе.
Этот гель окрашивается в разбавленном растворе этилиденбромида, и слой ДНК, составляющий ограничительный фрагмент ≈0,85 кб PstI - SphI, который визуально обнаруживается в длинноволновых ультрафиолетовых лучах, отсекается от данного геля в небольшом сегменте. Объем этого сегмента определяется по весу и плотности, и в пробирку, содержащую данный сегмент, вводится равный объем 10 ммоль Трис-HCl, pH 7,6. Затем этот сегмент расплавляется путем инкубации при 72oC. Получается примерно 1 мкг ограничительного фрагмента ≈0,85 кб PstI - SphI плазмиды pKC283PRS а объеме примерно 100 мкл. Аналогичным образом плазмида pKC283-LB вываривается с ограничительными ферментами PstI и SphI, полученный ограничительный фрагмент ≈3 кб изолируется путем электрофореза на агарозном геле и обрабатывается для подготовки к сшивке.
Ограничительный фрагмент 0,85 кб PstI - SphI плазмиды pKC283PRS сшивается с ограничительным фрагментом ≈0,3 кб PstI - SphI плазмиды pKC283-LB согласно процедуре примера 29 A2. Сшитая ДНК составляет желаемую плазмиду pL32. Ограничительная точка и функциональная карта плазмиды pL32 представлены на фиг. 7. Плазмида pL32 трансформируется в клетки E.Coli K12 MO (λ+) в основном согласно процедуре примера 29 A3. ДНК плазмиды pL32 приготавливается из трансформанта E.Coli K12 MO (λ+) / pL32 согласно процедуре, описанной в примере 29 A1. Анализ ДНК плазмиды pL32 продемонстрировал, что более чем одно связующее звено Eco RI прикрепляется к обработанным фрагментом Кленова концам SalI плазмиды pKC283PX. Присутствие более чем одного связывающего звена Eco RI не влияет на полезность плазмиды pL32 или производных плазмиды pL32 и может быть определено по присутствию ограничительной точки XhoI, которая образуется независимо от того, когда сшиваются оба связующих звена Eco RI вместе друг с другом. Как возможный вариант, плазмида pL32 может быть построена путем эксцизии фрагмента SalI-Eco RI и сшивки согласно первому разделу данного примера с описанием плазмиды pKC283-LB.
7. Построение E.Coli K12 MO (λ+) / pL47.
E. Coli K12 RV308/pNM789 может быть получена из Северной региональной научно-исследовательской лаборатории в лиофилизированной форме под номером NRRL B-18216. Ограничительная точка и функциональная карта pNM789 представлены на фиг. 8. Плазмидная ДНК экстрагируется из данной культуры согласно описанию примера 1, с той разницей, что температура инкубирования составляет 37oC. 10 мкг pNM789 суспензируются в 200 мкл буферного раствора PvuII (50 ммоль Трис-HCl, pH 7,5, 60 ммоль NaCl и 6 ммоль MgCl2). Вводится 1 ед. PvuII, и реакционная смесь инкубируется в течение 5 мин при 37oC. Данный фермент инактивируется путем нагрева в течение 10 мин при 65oC. Затем вводятся 30 мкл 10X буферного раствора Bam HI (200 ммоль Трис-HCl (pH 8), 1 моль NaCl и 70 ммоль MgCl2), 70 мкл H2O и 10 ед. BamHI, и реакционная смесь инкубируется в течение 1 ч. при 37oC. После этого вводятся 5 ед. щелочной фосфотазы и осуществляется инкубирование в течение 1 ч при 65oC. Фрагменты ДНК отделяются на 1%-ном агарозном геле, и осуществляется очистка фрагмента ДНК (фиг. 9) размером в единичный фрагментный отрезок.
Связывающее звено ДНК с отсеченным концом и с Bam HI-концом синтезируется в основном согласно процедуре примера 29 A4. Это связывающее звено (показано на фиг. 9) имеет структуру, представленную на фиг. 24.
Это связывающее звено киназируется и сшивается в вываренную Bam HI - PvuII плазмиду pNM789 в основном согласно процедуре примера 29 A2. Эта смесь сшивки используется как трансформация клеток E.Coli K12 RV308, и выделение плазмиды осуществляется с использованием данных трансформант согласно описанию примера 29 A3. Выбираются некоторые плазмиды, которые содержат фрагмент PvuII (494 hp) подходящего размера и фрагмент XbaI - Bam HI (628 hp). Последовательность по меньшей мере двух из них определяется из последовательности от точки Bam HI к уникальной точке SmaI, и выбирается один клон с желаемой последовательностью. Эта промежуточная плазмида обозначается как плазмида 120. Схематическое изображение данной процедуры, ограничительная точка и функциональная карта плазмиды 120 представлены на фиг. 9.
Для выделения кодирующей EK-BGH ДНК примерно 10 мкг плазмиды 120 вываривается в 200 мкл высокосолевого буферного раствора, содержащего примерно 50 ед. каждого из ограничительных ферментов XbaI и Bam HI. Продукты вываривания отделяются посредством электрофореза на агарозном геле, и ограничительный фермент ≈ 0,6 кб XbaI-Bam HI, который кодирует EK-BGH, выделяется и препарируется для сшивки согласно процедуре, описанной в примере 29 A6.
Плазмида pL32 также вываривается с ограничительными ферментами XbaI и Bam HI, и ограничительный фрагмент ≈ 3,9 кб извлекается и приготавливается для сшивки. Ограничительный фрагмент ≈ 3,9 кб XbaI-Bam HI плазмиды pL32 сшивается с ограничительным фрагментом 0,6 кб XbaI-Bam HI плазмиды 120 в основном согласно процедуре примера 29 A2, и в результате получается плазмида pL47. Ограничительная точка и функциональная карта плазмиды pL47 представлены на фиг. 10. Плазмида pL47 трансформируется в E.Coli K12 MO (λ+) в основном согласно процедуре примера 29 A3, и идентифицируются трансформанты E. Coli K12 MO (λ+) / pL47. ДНК плазмиды pL47 получается от данных трансформант в основном согласно процедурам примера 29 A1.
8. Построение E.Coli К12 P308 /pPR12AR1.
Плазмида pPR12 заключает в себе чувствительный к температуре подавляющий pL - ген с 1857, и плазмида pBR322 заключает ген, придающий стойкость к тетрациклину. Плазмида pPR12 описана и заявлена в патенте США N 4436815, выпущенном 13 марта 1984 года. Ограничительная точка и функциональная карта плазмиды pPR12 представлены на фиг. 11.
Примерно 10 мкг плазмиды pPR12 вывариваются примерно с 50 ед. ограничительного фермента Eco RI в 200 мкл высокосолевого буферного раствора при 37oC в течение 2 ч. Вываренная с Eco RI ДНК плазмиды pPR12 осаждается и обрабатывается фрагментом Кленова согласно процедуре примерна 29 A5. После протекания реакции Кленова вываренная Eco RI, обработанная фрагментом Кленова ДНК плазмиды pPR12, снова замыкается в кольцо путем сшивки согласно процедуре примера 29 A2. Эта сшитая ДНК, которая образует желаемую плазмиду pPR12AR1, используется для трансформации E.Coli K12 RV308 в основном согласно процедуре примера 29 A3, с той разницей, что этот выбор основан на стойкости к тетрациклину (5 мкг/мл), а не к ампициллину. E.Coli К12 RV308 имеет официальный номер NRRL B-15624. После идентификации трансформант E.Coli K12 RV308/pPR12АR1 из данных трансформант получается ДНК плазмиды pPR12AR1 согласно процедуре примера 29 A11.
Примерно 10 мкг плазмиды pPR12AR1 вывариваются примерно с 50 ед. ограничительного фермента AvaI в 200 мкл среднесолевого буферного раствора при 37oC в течение 2 ч.
Вываренная с AvaI ДНК плазмиды pPR12AR1 осаждается и обрабатывается фрагментом Кленова согласно процедуре примера 29 A5. После реакции с фрагментом Кленова вываренная с AvaI, обработанная фрагментом Кленова ДНК плазмиды pPR12AR1, сшивается со связывающими звеньями Eco RI (5'-GAGGAATTCCTG-3') в основном согласно процедуре примера 29 A2.
После сшивки связывающим звеном ДНК осаждается и затем повторно суспензируется примерно в 200 мкл высокосолевого буферного раствора, содержащего примерно 50 ед. ограничительного фермента Eco RI. Полученная реакционная смесь инкубируется при 37oC в течение примерно 2 ч. После вываривания с Eco RI реакционная смесь помещается на агарозный гель, и ограничительный фрагмент ≈ 5,1 кб Eco RI очищается в основном согласно процедуре примера 29 A6. Ограничительный фрагмент ≈ 5,1 кб Eco RI снова замыкается в кольцо путем сшивки согласно процедуре примера 29 A2. Эта сшитая ДНК образует желаемую плазмиду pPR12AR1. Эта ДНК плазмиды pPR12AR1 трансформируется в E.Coli K12 RV308 согласно процедуре примере 29 A3, с той разницей, что выбор основывается на стойкости к тетрациклину, а не к ампициллину.
После идентификации трансформант E.Coli K12 RV308/pPR12AR1 получается ДНК плазмиды pPR12AR1 в основном согласно процедуре примера 29 A1. Ограничительная точка и функциональная карта плазмиды pPR12AR1 представлены на фиг. 12.
9. Построение E.Coli K12 RV308/pL110.
Примерно 10 мг ДНК плазмиды pPR12AR1 суспензируются в примерно 200 мл высокосолевого буферного раствора, содержащего примерно 50 ед. каждого из ограничительных ферментов PstI и Eco RI, и реакционная смесь вываривания инкубируется при 37oC в течение примерно 2 ч. Затем эта реакционная смесь помещается на агарозный гель, и ограничительный фрагмент ≈ 2,9 кб PstI - Eco RI плазмиды pPR12AR1 выделяется и подготавливается для сшивки в основном согласно процедуре примера 29 A6.
Примерно 10 мкг плазмиды pL47 вывариваются с ограничительными ферментами PstI и Bam HI в 200 мкл высокосолевого буферного раствора при 37oC в течение 2 ч. Вываренная с PstI - Bam HI ДНК помещается на агарозный гель, и выделяется ограничительный фрагмент ≈ 2,7 кб PstI-Bam HI, который является источником репликации с частью стойкого к ампициллину гена и который подготавливается для сшивки согласно процедуре примере 29 A6. В реакции разделения примерно 10 мкг ДНК плазмиды pL47 вывариваются с ограничительными фрагментами Eco RI и Bam HI в 200 мкл высокосолевого буферного раствора при 37oC в течение 2 ч, и выделяется ограничительный фрагмент 1,03 кб PstI- Bam HI, который заключает в себя новую транскрипционную и трансляционную активирующую последовательность и кодирующую EK-BGH ДНК, и этот фрагмент подготавливается для сшивки согласно процедуре примера 29 A6. Примерно 2 мкг полученного ограничительного фрагмента ≈ 1,03 кб Eco RI - Bam HI используются для построения плазмиды pL110. Ограничительные фрагменты ≈ 2,7 кб PstI-Bam HI и 1,03 кб Eco RI - Bam HI плазмиды pL47 сшиваются с ограничительным фрагментом ≈ 2,9 кб PstI - Eco RI плазмиды pPR12AR1 для построения плазмиды pL110, и сшитая ДНК используется для трансформации E.Coli K12 RV308 в основном согласно процедуре примеров 29 A2 и 29 A3, с той разницей, что основой для выбора трансформант является стойкость к тетрациклину, а не к ампициллину. В кодирующей зоне EK-BGH присутствуют две распознавательные точки ограничительного фрагмента PstI, которые не отражаются на ограничительной точке и на функциональных картах на фигурах данной заявки. Ограничительная точка и функциональная карта плазмиды pL110 представлены на фиг. 13.
10. Построение E.Coli K12 RV308 /pL110C.
а. Построение E.Coli K12 RV308 /pL110A.
Примерно 1 мкг ДНК плазмиды pL110 вываривается с ограничительным ферментом NdeI в 20 мкл общего объема, содержащего 2 мкл 10X высокосолевого буферного раствора (1 моль NaCl, 0,50 моль Трис-HCl, pH 7,5, 0,10 моль MgCl2 и 10 ммоль дитиотреитола) и 3 ед. фермента NdeI, в течение 1 ч при 37oC. Реакционная смесь экстрагируется фенолом /хлороформом, и ДНК осаждается этанолом. Вываренная с NdeI ДНК плазмиды pL110 растворяется в 50 мкл 1X буферного раствора Кленова (40 ммоль K2HPO4, pH 7,5, 6,6 ммоль MgCl2, 1 ммоль 2-меркаптоэтанола, 33 мкмоль dATP, 33 мкмоль dGTP и 33 мкмоль TTP). 2 мкл (≈ 10 ед. , New England Biolabs) крупных фрагментов ДНК полимеразы 1 E. Coli, известных как фрагменты Кленова, вводятся в данный раствор и смешиваются с ДНК, и полученная реакционная смесь инкубируется при 16oC в течение 1 ч. Реакция завершается путем экстрактации фенолом, и ДНК очищается обычными приемами. Вываренная с NdeI, обработанная фрагментом Кленова ДНК затем сшивается с Т4 ДНК лигазой при 4oC в течение 16 ч. Полученная ДНК используется для обычной трансформации штамма E. Coli K12 RV308 (NRRL B-15624). Трансформанты отбираются на чашках с L-агаром, содержащих 100 мкг/мл ампициллина, и плазмиды извлекаются из стойких колоний путем быстрой щелочной экстракции, как описано в работе Birnboim и Doly. Отбирается плазмида (pL110A на фиг. 14) с отсутствием точки NdeI.
b. Построение фага pL110 B путем точечно-специфического мутагенеза.
Осуществление процедуры удаления точки Bam HI в гене, сообщающем стойкость к тетрациклину путем точечно-специфического мутагенеза, представлено с правой стороны фиг. 14.
b (i) Построение фага m13 Tc3.
Плазмида pL110 служит в качестве источника, придающего стойкость к тетрациклину гена. Примерно 50 мкг плазмиды pL110 в 50 мкл буферного раствора ТЕ вводятся в 25 мкл буферного раствора 10X Hind III и 170 мкл H2O. Примерно 5 мкл (≈ 50 ед. ) ограничительного фермента Hind III вводятся в раствор ДНК плазмиды pL110, и полученная реакционная смесь инкубируется при 37oC в течение 2 ч. Примерно 13 мкл 2 М Трис-HCl, pH 7,4, и 5 мкл (≈ 50 ед.) ограничительного фермента Eco RI вводятся в вываренную с HindIII ДНК плазмиды pL110, и реакционная смесь инкубируется в течение более 2 ч при 37oC. Реакция прекращается в результате экстракции реакционной смеси насыщенным TE-фенолом, фенол удаляется путем экстракций хлороформом. Вываренная с Eco RI-HindIII ДНК плазмиды pL110 затем извлекается путем осаждения и центрифугирования, помещается на 1%-ный агарозный гель, и крупный ограничительный фермент ≈ 4,3 кб Eco RI -HindIII выделяется и очищается.
Примерно 5 мкг фага m13 mp18 (New England Biolabs) растворяются в 50 мкл буферного раствора TE, и затем выпариваются HindIII и Eco RI, как описано выше.
Отсеченная HindIII - Eco RI ДНК фага m 13 mp18 очищается, как описано для pL110, с той разницей, что осуществляются выделение и очистка ограничительного фрагмента ≈ 7,25 кб.
Примерно 100 нг фрагмента ≈ 4,3 кб HindIII - Eco RI празмиды pL110 смешиваются с примерно 100 нг фрагмента ≈ 7,25 кб HindIII - Eco RI фага m13 mp18, 2 мкл 10X буферного раствора лигазы, 1 мкл (≈ 100 ед.) T4 ДНК лигазы и 14 мкл H2O. Реакционная смесь сшивки инкубируется при 15oC в течение 1,5 ч, сшитая ДНК составляет ДНК желаемого фага m13 Tc3. Ограничительная точка и функциональная карта фага m13 Tc3 представлены на фиг. 14.
1 мл ночной культуры E. Coli K12 JM109 (E. Coli K12 JM101 получена из New England Biolabs, может использоваться вместо E. Coli K12 JM109) используется для инокулирования 50 мл питательного бульона L (50 мл), и полученная культура инкубируется при 37oC с аэрацией до тех пор, пока OD660 (оптическая плотность) не будет составлять от 0,3 до 0,4. Клетки повторно суспензируются в 25 мл и 10 ммоль NaCl, инкубируются на льду в течение 10 мин и извлекаются путем центрифугирования. Эти клетки помещаются в 1,25 мл 0,75 мМ CaCl2, 200 мкл аликвота этих клеток удаляется, вводится в 10 мкл сшитой ДНК, полученной, как описано выше, и инкубируется на льду в течение ≈ 40 мин. Затем смесь клетки - ДНК инкубируется при 92oC в течение 2 мин, и различные аликвоты (1,10 и 100 мкл) удаляются и вводятся в 3 мл верхнего агара (L-бульон с 0,5%-ным агаром, поддерживаемым в расплавленном состоянии при 45oC), который содержит также 50 мкл 2% X-Gal, 50 мкл 100 мМ IPTG и 200 мкл E. Coli K12 JM109 в фазе логарифмического роста. Затем верхняя клеточная часть агаровой смеси высевается в чашках с L-агаром, содержащих 40 мкг/мл X-Gal (5-бромо-4-хлоро-3-индолил-D-тиогалактозид) и 0,1 ммоль IPTG (изопропил- β -D-тиогалактозид), и чашки инкубируются в течение ночи при 37oC.
На следующее утро несколько прозрачных, но не синих колоний используются для инокулирования 2 мл L-буольна, и образующиеся в результате культуры инкубируются при 37oC с аэрацией в течение 2 ч. Отсутствие синего окрашивания показывает, что происходит желаемая вставка ДНК. Затем данные культуры центрифугируются, и 200 мкл полученного поверхностного слоя вводятся в 10 мл культур (OD550 = 0,5) E. Coli K12 JM109, выращенных при 37oC с аэрацией. Эти культуры инкубируются еще в течение 30 мин при 37oC, затем эти клетки выпадают в осадок в результате центрифугирования и используются для получения репликаторной формы рекомбинантного фага, который они содержат. Из этих клеток выделяется двухниточная репликаторная форма фаговой ДНК с использованием процедур (в меньших объемах), описанных в примере 1. Транфсформанты, содержащие фаговую m13 Tc3 ДНК, идентифицируются путем анализа на ограничительный фрагмент их фаговой ДНК.
b (ii) Получение однониточной ДНК фага m13 Tc3.
1,5 мл ночной культуры Coli K12 JM109/m13 Tc3 центрифугируются, и 100 мкл верхнего слоя, содержащего фаг m13 Tc3, используются для инокуляции 25 мл культуры Coli K12 JM109 с оптической плотностью OD660 примерно 0,4 - 0,5. Данная культура инкубируется примерно в течение 6 ч при 37oC с аэрацией, и затем культура центрифугируется, образованный верхний слой примерно 20 мл переносится в новую пробирку. Примерно 2 мл раствора, содержащего 20% полиэтиленгликоля (PEC) 6000 и 14,6% NaCl, вводятся в поверхностный слой, который затем инкубируется на льду в течение 20 мин.
Поверхностный слой центрифугируется в течение 25 мин со скоростью 7000 об/мин, и образующийся осадок, который содержит однониточную ДНК фага m13 Tc3, снова суспендируется в 500 мкл буферного раствора TE. Раствор ДНК двукратно экстрагируется насыщенным TE-фенолом и двукратно - хлороформом. Затем однониточная ДНК осаждается с использованием NaOAc и этанола и центрифугируется. Полученный осадок промывается 70%-ным этанолом, высушивается и затем растворяется в 60 мкл H2O.
b (iii) Мутагенез.
Фрагмент однониточной ДНК, используемый в процессе мутагенеза, синтезируется в автоматическом синтезаторе ДНК. Этот фрагмент имеет последовательность 5'-CCCGTCCTCTGGATACTCTACGCCGA-3' и является гомологом зоны, окружающей точку Bam HI (5'-GGATCC-3'), в придающем стойкость к тетрациклину гене от плазмиды pBR322, с той разницей что A-группа, вторая от 5'-конца (или третья от 3'-конца), является C в плазмиде pBR322. Такое видоизменение не меняет аминокислотный состав белка, придающего стойкость к тетрациклину, но исключает точку Bam HI.
Примерно 10 пмоль мутагенной затравки и универсальной затравки M 13 (Bethesda Research Laboratories (BRL) P.O.Box 6009, Гатесбург, MD 20760) обрабатываются по отдельности 10 ед. (BRL) полинуклеотидокиназы T4 в 20 мкл буферного раствора киназы 1X (60 ммоль Трис-HCl, pH 7,8, 15 ммоль 2-меркаптоэтанола, 10 ммоль MgCl2 и 0,41 мкмоль АТР) в течение 30 мин при 37oC. Обработанные киназой ДНК используются в описанной ниже процедуре мутагенеза.
Реакция ренатурации (анилирования) осуществляется путем смешивания друг с другом 300 нг (1,2 мкл) однониточного фага m13 Tc3, 1 пмоль (2 мкл) универсальной затравки, 1 пмоль (2 мкл) мутагенной затравки, 2 мкл 10X ренатурирующего буферного раствора (100 ммоль Трис-HCl, pH 7,5, 1 ммоль ЭДТА и 500 ммоль NaCl) и 12,8 мкл H2O. Реакционная смесь инкубируется при 80oC в течение 2 мин, при температуре 50oC в течение 5 мин, а затем охлаждается до комнатной температуры.
Реакция удлинения цепи осуществляется путем ввода 5 мкл 10X буферного раствора вытягивания (или удлинения) (500 ммоль Трис-HCl, pH 8, 1 ммоль ЭДТА и 120 ммоль MgCl2), 5 мкл 2 мМ dATP, 1 мкл раствора 6 мМ каждого из dGTP, TTP и dCTP, 1 мкл (≈ 2 ед., Pharmacia, P-L Biochemicals, 800 Centennial Avenue Piseataway, NJ 08854) фермента Кленова, 1 мкл (100 ед.) T4 ДНК лигазы и 17 мкл H2O в смесь ренатурированной ДНК. Реакционная смесь удлинения цепи инкубируется при комнатной температуре в течение 1 ч, затем при 37oC в течение 2,5 ч и затем в течение ночи при 4oC.
Реакция прекращается путем осуществления двух экстракций насыщенным TE-фенолом, после которых осуществляются две экстракции CHCl3. ДНК осаждается этанолом и ацетатом натрия (NaOAc). ДНК извлекается путем центрифугирования и снова суспензируется в 50 мкл H2O, и затем 6 мкл 10X буферного раствора SI вводятся в раствор ДНК.
Этот раствор ДНК распределяется поровну в трех пробирках. В две из этих пробирок вводится примерно 200 ед. (Miles Laboratories) нуклеазы 1. Одна реакционная смесь SI инкубируется при комнатной температуре в течение 5 мин, другая в течение 10 мин. Реакции прекращаются в результате экстракции реакционной смеси два раза насыщенным TE-фенолом. После фенольных экстракций осуществляются две экстракции хлороформом, затем из данной реакционной смеси осаждается ДНК ацетатом натрия (NaOAc) и этанолом. Необработанный образец ДНК служит в качестве отрицательного контрольного образца. Обработанные образцы сохраняются изолированными друг от друга в ходе последующей процедуры, но результаты получаются одинаковыми.
Выпавшие осадки ДНК повторно суспензируются в 20 мкл H2O, и используется 10 мкл полученного раствора для трансформации E. Coli K12 JM109 (E. Coli K12 JM101 также может быть использован) согласно процедуре, осуществляемой для построения фага m13 Тс3, с той разницей, что в чашки не вводится ни IPTG, ни X-Gal. Двухниточная репликаторная форма ДНК от примерно 48 колоний выделяется, как описано выше, и отбирается на присутствие ограничительной точки Bam HI. Изоляты без точки Bam HI дополнительно отбираются с получением однониточной ДНК, как описано выше.
Аминокислотная последовательность однониточной ДНК определяется с использованием определения дидеокси последовательности (J.H. Smith, Методы в энзимологии, 65 : 560 - 580). Желаемый изолят обозначается как pL110B (фиг. 14).
с. Построение плазмиды pL110C.
Примерно 50 мкг репликаторной формы фага pL110B ДНК вывариваются в 250 мкг 1X буферного раствора NheI (50 ммоль NaCl, 6 ммоль Трис-HCl, pH 7,5, 6 ммоль MgCl2 и 6 ммоль β -меркаптоэтанола), содержащего ≈ 50 ед. ограничительного фермента NheI, при 37oC в течение 2 ч. 5 мкл 5 M NaCl затем вводятся в вываренную с NheI фаговую pL110B ДНК с последующим вводом 5 мкл (≈ 50 ед.) ограничительного фермента Sal. Вываривание продолжается в течение 2 ч при 37oC. Затем желаемый фрагмент ≈ 422 вр NheI-SalI, содержащий мутированную зону гена, придающего стойкость к тетрациклину, выделяется из акриламидного геля согласно известным стандартным процедурам. ДНК плазмиды pL110A вываривается с NheI и SalI в идентичных условиях, с той разницей, что вместо плазмиды pL110B используется плазмида pL110A. Ограничительный фрагмент ≈ 6,1 кб NheI-SalI плазмиды pL110A очищается от агарозы.
Построение желаемой плазмиды pL110C осуществляется путем сшивки друг с другом по 100 нг каждого из NheI-SalI фрагментов pL110A (≈ 6,1 кб) и pL110B (≈ 422 вр) путем общеизвестных процедур. Ограничительная точка и функциональная карта плазмиды pL110C представлены на фиг. 15. Желаемая плазмида pL110C сообщает стойкость к тетрациклину для 10 мкг/мл тетрациклина в E. Сoli, но в сообщающем стойкость к тетрациклину гене отсутствует точка Bam HI.
11. Построение плазмиды pCZRIII.
Плазмида pL110C содержит единственную ограничительную точку ClaI, которая удаляется при протекании нижеследующих реакций. Примерно 1 мкг плазмиды pL110C вываривается с ClaI согласно процедуре примера 29 A2, с той разницей, что используются ограничительный фермент ClaI и 10X буферный раствор ClaI (500 ммоль NaCl, 100 ммоль Трис-HCl, pH 7,9 и 100 ммоль MgCl2). Вываренная с ClaI ДНК затем обрабатывается фрагментом Кленова согласно описанию примера 29 A5, с той разницей, что вводится лишь dCTP, а не все четыре dNTР.
Затем ДНК осаждается и повторно суспензируется в 50 мкл буферного раствора бобовой нуклеазы Mung (50 ммоль ацетата натрия, pH 5, 30 ммоль NaCl и 1 ммоль ZnSO4 ). 1 ед. бобовой нуклеазы Mung (промышленно выпускается New England Biolabs) вводится в эту суспензию, и реакционная смесь инкубируется при 30oC в течение 30 мин. Затем пробирка помещается в лед, и NaCl вводится в концентрации 0,2 моль, после чего смесь экстрагируется фенолом (хлорофоромом), осаждается этанолом и повторно суспензируется в 10 ммоль Трис-HCl (pH 8). Затем ДНК подвергается самосшивке и трансформируется в клетки E. Сoli согласно описанию примеров 29 A3 и 29 A4. Полученная плазмида представляет собой вываренную плазмиду pCZR111.
12. Построение плазмиды pCZR126.
Примерно 26 мкг плазмиды pCZR111 вывариваются с XbaI указанным ниже образом. Буферный раствор 10X XbaI состоит из 600 ммоль Трис-HCl, 100 ммоль MgCl2, 1 моль NaCl и 10 ммоль 2-меркаптоэтанола, pH 7,5 (при 37oC), 50 мкл буферного раствора 10X XbaI, 15 мкл XbaI (10 ед/мкл) и 185 мкл H2O вводятся в 250 мкл воды, содержащей примерно 25 мкл плазмиды pL110. Вываривание осуществляется при 37oC в течение 1 ч. pL110, вываренная с XbaI, затем экстрагируется в феноле, вводится 1/10 объема этанола, смесь инкубируется в бане сухой лед-этанол в течение 5 мин и затем центрифугируется. Осажденная ДНК повторно суспензируется в 50 мкл H2O. Вываренная с XbaI плазмида pCZR111, вываривается в Bam HI следующим образом. 0,2 мкл Bam HI (10 ед/мкл), 10 мкл буферного раствора Bam HI (100 ммоль Трис-HCl, 50 ммоль MgCl2, 1 моль NaCl и 10 ммоль 2-меркаптоэтанола, pH 8, при 37oC) и 90 мкл H2O вводятся в 50 мкл pL110, вываренного с XbaI, полученного как указано выше. Вываривание осуществляется в течение 5 мин при 37oC. Вываренная pCZR111 экстрагируется в феноле, вводится 1/10 объема CH3COONa, после чего вводятся 3 объема этанола. Осажденная ДНК снова суспензируется в 50 мкл 10 мМ Трис, 1 ммоль ЭДТА, pH 8, буферном растворе.
pCZR111, вываренная с XbaI и Bam HI, затем помещается на агарозный гель, и выделяется слой ДНК примерно 5,8 кб. Плазмида pCZR126S получается путем сшивки фрагмента ≈ 5,8 кб pCZR111 со связывающими звеньями XbaI и NdeI и синтетическим геном, кодирующим EK-бычий гормон роста, который содержит точку NdeI у его 5'-конца и точку Bam HI у его 3'-конца. Последовательность XbaI до NdeI получается с использованием метода построения стандартной олигонуклеотидной последовательности, и она заключает в себе последовательность, приведенную на фиг. 25.
Указанная последовательность построена путем химического синтеза обеих нитей с последующим смешиванием с целью гибридизации. Кодирующий EK-bGH-ген построен из 16 химически синтезированных сегментов однониточной ДНК, заключающих в себе от 71 до 83 нуклеотидов, которые совместно друг с другом составляют дополняющие нити всего гена. Синтез осуществляется в устройстве Applied Biosystems (ABS) и включает последовательность, приведенную на фиг. 26 и 27.
Построение плазмиды pCZR126S осуществляется путем сшивки нижеследующих компонентов: ≈ 0,28 мкг фрагмента 5,8 кб, полученного из плазмиды pL110 после полного вываривания с XbaI и частичного вываривания c Bam HI в общем объеме 2 мкл, ≈ 0,18 мкг синтетического гена, кодирующего производное фактора бычьего роста, который имеет конец 5', соответствующий точке XbaI, и конец 3', соответствующий точке Bam HI в общей объеме 2,5 мкл, 8,75 пмоль химически синтезированного XbaI - NdeI связывающего звена в 1 мкл. Эти компоненты плазмиды вводятся в 8 мкл 5X буферного раствора сшивки: 250 ммоль Трис-HCl, 50 ммоль MgCl2, 5 ммоль АТР, 5 ммоль ДТТ, 25% об./об. полиэтиленгликоля 8000, pH 7,6, 2 мкл лигазы и 16,5 мкл H2O. Эта смесь сшивки инкубируется в течение ночи при 16oC. Далее замкнутая в кольцо плазмида pCZR126S используется для трансформации клеток E. Coli RV308 согласно способу примера 29 A3. Ограничительная точка и функциональная карта плазмиды pCZR126S представлены на фиг.16.
B. Построение выраженной плазмиды человеческого проинсулина pRB172
1. Построение плазмиды pRB145.
Ген человеческого проинсулина сначала синтезируется обычным образом и клонируется в плазмиду PuC18 (промышленно выпускаемая BRL). Этот ген синтезируется стандартными приемами и заключает в себе последовательность, представленную на фиг. 28.
Один из клонов, имеющий правильную последовательность, выбирается для продуцирования очищенной от хлорида цезия ДНК. Данная плазмида извлекается путем стандартной процедуры (см., например, публикацию "Молекулярное клонирование. Лабораторное пособие, 1982 г., ed. Maniatis T.; Fritsh E.F. u Sambrook, J., Cold Spring Heorbour Laboratory Publucations, Нью Йорк). Примерно 6 мкл (29 мкг) этой ДНК плазмиды вводятся в 20 мкл 10X буферного раствора NdeI (150 ммоль NaCl, 10 ммоль Трис-HCl, pH 7,8, 6 ммоль MgCl2, 6 ммоль 2-меркаптоэтанола, 100 мкг/мл BSA), 5 мкл ограничительного фермента Nde I (≈ 40 ед. ) и 169 мкл H2O. После перемешивания реакционная смесь инкубируется при 37oC в течение 2 ч. ДНК осаждается путем приготовления 0,3 моль NaOAc, ввода трех объемов этанола, перемешивания, охлаждения до -70oC и центрифугирования. Осажденная ДНК промывается 70%-ным этанолом (1 мл), высушивается и растворяется в 20 мкл 10X Bam HI буфера (150 ммоль NaCl, 6 ммоль Трис-HCl, pH 7,9, 6 ммоль MgCl2, 100 мкг/мл BSA), 2 мкл ограничительного фермента Bam HI (40 ед. ) и 178 мкл H2O. После слабого перемешивания реакционная смесь инкубируется при 37oC в течение 2 ч. ДНК снова осаждается тремя объемами этанола, как определено выше, и подвергается электрофорезу на 1%-ном расплавленном агарозном геле (FMC, морская агароза). Желаемый фрагмент ДНК, соответствующий примерно 270 вр, срезается с геля, и затем ДНК извлекается путем расплавления агарозы и пропускания через колонку Elutip-d (Schlicher and Schuell, Kune N.H) согласно процедуре, рекомендуемой продавцом. После осаждения и сушки ДНК выдерживается в 30 мкл 10 мМ Трис-HCl, pH 8.
Примерно 15 мкг плазмиды pCZR126S (из примера 29 A12) суспензируются в 20 мкл 10X буферного раствора NdeI, 5 мкл ограничительного фермента NdeI (40 ед. ) и H2O (175 мкл), слабо перемешиваются, и смесь инкубируется при 37oC в течение 2 ч. После инкубации ДНК осаждается тремя объемами этанола, как и выше, высушивается и затем повторно суспензируется в 20 мкл 10X буферного раствора Bam HI, 2 мкл ограничительного фермента Bam HI (40 ед.) и 178 мкл воды. После слабого перемешивания реакционная смесь инкубируется при 37oC в течение 2 ч. ДНК снова осаждается тремя объемами этанола, и осуществляется электрофорез на 1%-ном низкоплавком агарозном геле. Наиболее крупный фрагмент, соответствующий векторной ДНК, срезается с этого геля, и ДНК извлекается путем осуществления процедуры в колонке Elutip-d, как описано выше. После осаждения и сушки векторная ДНК выдерживается в 35 мкл 10 мМ Трис-HCl, pH 8.
Примерно 2,5 мкл векторной ДНК смешиваются с 12 мкл генного фрагмента очищенного человеческого инсулина, 4 мкл 10 мМ АТР, 0,5 мкл и 1 моль дитиотреитола, 5 мкл 10X буферного раствора лигазы (50 ммоль Трис-HCl, pH 7,6, и 100 ммоль MgCl2), 26 мкл воды и 0,5 мкл T4 ДНК лигазы (Pharmacia, 3,5 ед.). Эта реакционная смесь инкубируется при 4oC в течение 16 ч. Сшитая смесь разбавляется 50 мкл 10 мМ Трис-HCl (pH 7,6) и 3 мкл 1 М CaCl2 и затем последовательно трансформируется в E. Coli K12 RV308 согласно описанию примера 29 A3. Клетки высеваются на чашках ТУ, содержащих 5 мкг/мл тетрациклина, и затем инкубируются в течение ночи при 32oC.
Плазмиды от 3 мл культур выделяются из стойких к тетрациклину колоний путем быстрой щелочной экстракции, как описано в публикации "Молекулярное клонирование". Лабораторное пособие, 1982 г., ред. Moniatis T., Fritsh E.F. и Sambrook J. Cold Spring Harbour Pablication, Нью Йорк, с. 368-369. Присутствие нужного генного фрагмента человеческого проинсулина определяется путем процедуры миниотбора с использованием электрофореза на полиакриламидном геле для анализа вываренного с XbaI/Bam HI фрагмента. Плазмиды с правильными по размеру вставками (примерно 316 вр) отбираются для амплификации и очистки. Плазмида, содержащая ген человеческого проинсулина, представляет собой pRB145. Ограничительная точка и функциональная карта плазмиды pRB145 представлены на фиг. 18.
2. Построение плазмиды pRB164A.
Примерно 30 мкг плазмиды pRB145 суспензируются в 20 мкл 10X буферного раствора NdeI, 5 мкл ограничительного фермента NdeI (New England Biolаbs, 40 ед. ) и 175 мкл H2O с осторожным перемешиванием, и смесь инкубируется при 37oC в течение 1 ч. Затем в реакционную смесь вводится 2 мкл ограничительного фермента Bam HI (New England Biolabs, 40 ед.), и осуществляется инкубирование при 37oC в течение еще 2 ч. ДНК осаждается тремя объемами этанола и 0,3 моль NaOAc и подвергается электрофорезу на 1%-ном низкоплавком агарозном геле. Более мелкий ограничительный фрагмент (примерно 270 вр) NdeI/Bam HI, кодирующий ген человеческого проинсулина, срезается с геля, и ДНК извлекается путем прохождения через колонку Elutip-d, как описано выше. После осаждения и сушки ДНК выдерживается в 30 мкл 10 мМ Трис, pH 8.
Затем к этой ДНК (30 мкл) добавляется 20 мкл 10X буферного раствора AvaII (50 ммоль NaCl, 6 ммоль Трис-HCl, pH 8, 10 ммоль MgCl2, 6 ммоль 2-меркаптоэтанола, 100 мкг/мл BSA), 5 мкл ограничительного фермента AvaII (New England Biolabs, 20 ед.) и 175 мкл H2O. После осторожного перемешивания данная реакционная смесь инкубируется при 37oC в течение 2 ч. ДНК осаждается тремя объемами этанола и 3 ммоль NaOAc (20 мкл) и затем подвергается электрофорезу на 1,2%-ном низкоплавком агарозном геле. Более крупный ограничительный фрагмент AvaII/Bam HI (примерно 156 вp) срезается с геля, и затем ДНК извлекается путем прохождения через колонку Elutip-d, как описано выше. После осаждения и сушки ДНК выдерживается в 30 мкл 10 мМ Трис, pH 8.
ДНК (≈ 115 вp), соответствующая ограничительному фрагменту NdeI/AvaII гена человеческого проинсулина, получается путем синтеза. Первый этап заключается в синтезе четырех однониточных деоксирибоолигонуклеотидов посредством синтезатора ДНК Applied Biosystems модели 380B. Нуклеотидные последовательности этих четырех олигонуклеотидов представлены на фиг. 29.
После очистки каждого олигонуклеотида путем электрофореза на полиакриламидном геле олигонуклеотиды два и три фосфорилируются согласно описанию публикаций Brown E.L. Belagaje R. Kyan M.J. и Khorana H.G. (1979 г.), Методы в энзимологии. Ред. Wu R., Academic Precc, Нью Йорк, 68, 109-151. Фосфорилированные олигонуклеотиды два и три (≈715 пмоль каждого) затем смешиваются с олигонуклеотидами один и два (≈ 860 пмоль), ренатурируются и сшиваются в буферном растворе (200 мкл), содержащем 50 ммоль Трис-HCl (pH 7,6), 10 ммоль MgCl2 , 10 ммоль ДТТ, 59 мкмоль АТР и 20 ед. Т4 ДНК лигазы (Pharmacia) в течение 16 ч при 4oC. Продукт сшивки очищается на 15%-ном полиакриламидном геле. ДНК извлекается из гелевого слоя путем электрофореза с последующим обессоливанием в колонке Sephadex C-50. Выход желаемого сшитого продукта составляет 485 пмоль.
Примерно 100 пмоль этой ДНК фосфорилируется в буферном растворе (50 мкл), содержащем 50 ммоль Трис (pH 7,6), 10 ммоль MgCl2, 10 ммоль ДТТ и АТР, как описано в публикации Brown E.L. и др., 1979 г. Методы в энзимологии, 68, 109-151. После фильтрации через колонку Sephadex C-50 ДНК выдерживается в 50 мкл 10 мМ Трис, pH 8.
Примерно 2,5 мкл векторной ДНК (вываренной в NdeI-Bam HI pСZR126) смешиваются с 18 мкл ограничительного фрагмента AvaIII/Bam HI от плазмиды pPRB145 и 10 мкл (10 пмоль) синтетического связывающего звена NdeI/AvaII в буферном растворе (50 мкл), содержащем 50 ммоль Трис (pH 7,6), 10 ммоль MgCl2, 10 ммоль ДТТ, 800 мкл АТР и 3,5 ед. Т4 ДНК лигазы. Реакционная смесь инкубируется при 4oC в течение ночи и затем трансформируется в E.Coli K12 RV308 согласно процедуре примера 29 A3. Желаемая трансформанта E. Coli K12 RV308/pRB164A идентифицируется путем анализа ее плазмидной ДНК. Нужная трансформанта выращивается при 30oC в среде T4, содержащей 5 мгк/мл тетрациклина, до достижения ОD550 примерно 0,2 (ранняя логарифмическая фаза роста) и затем при 42oC в течение 3-3,5 ч для индуцирования синтеза человеческого проинсулина. Клетки выпадают в осадок и лизируются путем добавления буфера (0,125 моль Трис -HCl, pH 6,8, 2% SDS, 30% глицерина, 1 моль 2-меркаптоэтанола, 6 моль мочевины). Образцы нагреваются до 97oC в течение 2 мин до помещения их на акриламидный гель. Слои легко визуально просматриваются путем окрашивания красителем Кумаси Брильянтовым синим. Для количественного определения процента клеточного белка используется сканирующий гелевый денситометр. Ограничительная точка и функциональная карта плазмиды pPR16 4A представлены на фиг. 19.
3. Построение плазмиды pRB172.
Примерно 25 мкг плазмиды pRB145 суспензируются в 15 мкл 10X буферного раствора DraIII (200 ммоль NaCl, 50 ммоль Трис-HCl, pH 7,5, 1 ммоль ДTT, 10 мкг/мл BSA), 6 мкл ограничительного фермента DraIII (Bochringer Mаnnheim, 27 ед. ) и 129 мкл H2O, осторожно перемешиваются и инкубируются при 37oC в течение 6 ч. После инкубации ДНК осаждается, высушивается и повторно суспензируется в 10 мкл 10X буфера XbaI (150 ммоль NaCl, 6 ммоль Трис-HCl, pH 7,9, 6 ммоль MgCl2, 7 ммоль 2-меркаптоэтанола, 100 мкг/мл BSA), 3 мкл ограничительного фермента XbaI (Bochringer Mannheim, 36 ед.) и 77 мкл H2O. Реакционная смесь осторожно перемешивается и инкубируется при 37oC в течение 4 ч. ДНК снова осаждается тремя объемами этанола и ацетата натрия, подвергается электрофорезу на 1%-ном низкоплавком агарозном геле. Нижний слой, соответствующий примерно 85 вp ограничительного фермента XbaI/DraIII гена человеческого проинсулина, срезается с геля, и ДНК извлекается путем осуществления процедуры в колонке Elutip-d. После осаждения и сушки ДНК выдерживается в 30 мкл 10 мМ Трис, pH 8.
Примерно 15 мкг плазмиды pRB164A отсекаются с ограничительным ферментом DraIII и XbaI согласно описанной выше процедуре. Верхний слой, соответствующий векторному фрагменту XbaI/DraIII, выделяется из агарозного геля путем осуществления процедуры в колонке Elutip-d. После осаждения и сушки ДНК выдерживается в 30 мкл 10 мМ Трис, pH 8.
Примерно 5 мкл векторной ДНК pRB16 4A, вываренной с XbaI/DraIII, смешиваются с 5 мкл ограничительного фрагмента XbaI/DraIII от плазмиды pBR145 с буферном растворе (50 мкл), содержащем 50 ммоль Трис-HCl, pH 7,6, 10 ммоль MgCl2 , 10 ммоль ДTT, 800 ммоль АТР и 6 ед. T4ДНК лигазы. Реакционная смесь инкубируется при 4oC в течение ночи и используется для трансформации клеток E. Coli K12 RV308, которые превращаются в компетентные клетки путем стандартной обработки CaCl2.
Трансформанты выбираются на ТУ агаровых чашках, содержащих 5 мкг/мл тетрациклина. Плазмиды выделяются из стойких к тетрациклину колоний и анализируются путем вываривания с ограничительными нуклеазами XbaI и Bam HI. Выстраивается аминокислотная последовательность ДНК от положительных клонов с использованием системы построения последовательности (U.S.Biochemicals). Плазмиды с желаемой правильной последовательностью отбираются для амплификации и очистки. Таким образом изолируется трансформанта E.Coli K12 RV308/pRB172. Выражение и аккумуляция человеческого проинсулина, заключающего в себе данную плазмиду, анализируются путем визуализации всего клеточного белка с последующим электрофоретическим отделением в 15%-ной полиакриламидной матрице. Ограничительная точка и функциональная карта плазмиды pRB172 представлены на фиг. 20.
C. Построение Escherichia Coli RV308/pRB173 и pRB174.
Примерно 20 мкг плазмиды pRB172 или pRB145 суспензируются в 20 мкл 10X буфера XbaI (150 ммоль NaCl, 6 ммоль Трис-HCl, pH 7,9, 6 ммоль MgCl2, 7 ммоль 2-меркаптоэтанола, 100 мкг/мл BSA), вводятся 2 мкл ограничительного фермента XbaI (Bochringer Mannheim, 24 ед.), смесь осторожно перемешивается и инкубируется при 37oC в течение 4 ч. После инкубирования ДНК осаждается, высушивается и снова суспензируется в 10 мкл 10X буферного раствора Bam HI (100 ммоль NaCl, 10 ммоль Трис-HCl, pH 8, 5 ммоль MgCl2, 1 ммоль 2-меркаптоэтанола, 100 мкг/мл BSA), 5 мкл ограничительного фермента Bam HI (Bochringer Mannheim, 40 ед. ) и 75 мкл H2O. Реакционная смесь осторожно перемешивается и инкубируется при 37oC в течение 4 ч. ДНК снова осаждается этанолом и ацетатом натрия и подвергается электрофорезу на 1%-ном низкоплавком агарозном геле. Ограничительный фермент XbaI / Bam HI примерно 320 вp, соответствующий гену человеческого проинсулина, извлекается из геля путем осуществления процедуры в колонке Elutip-d. После осаждения и сушки ДНК снова суспензируется в 20 мкл 10X буферного раствора XmaI (25 ммоль NaCl, 10 ммоль Трис-HCl, pH 7,5, 10 ммоль MgCl2, 10 ммоль 2-меркаптоэтанола, 10 мкг/мл BSA), 10 мкл ограничительного фермента XmaI (New England Biolabs, 10 ед. ) и 170 мкл воды. Реакционная смесь осторожно перемешивается и инкубируется при 37oC в течение 4 ч. После инкубирования ДНК осаждается этанолом и ацетатом натрия и подвергается электрофорезу на 1%-ном низкоплавком агарозном геле. Более крупная ДНК, соответствующая ограничительному фрагменту XbaI/XmaI примерно 200 вp, срезается с геля, и эта ДНК выделяется путем осуществления процедуры в колонке Elutip-d, как описано выше. Эта ДНК выдерживается в 30 мкл 10 мМ Трис, pH 8.
ДНК на 51 вp, соответствующая ограничительному фрагменту Xma/Bam HI гена человеческого проинсулина, выравнивается в последовательность, которая определена ниже. Два однониточных деоксирибоолигонуклеотида, приведенных на фиг. 30, синтезируются с помощью синтезатора ДНК, Applied Biosystems (модели 380B), и очищаются путем электрофореза на полиакриламидном геле в присутствии 7 моль мочевины. После изоляции оба нуклеотида фосфорилируются согласно процедуре в описании Brown E.L., Belagaje R.; Ryan M.J. и Khorana H.G. (1979 г.). Методы энзимологии. Ред. Wu R.Academic Press, N.J., 68, 109-151, и ренатурируются с образованием желаемого связывающего звена.
Примерно 19 мкг плазмиды pCZR126S (из примера 29 A12) сусупензируются в 20 мкл 10X буферного раствора XbaI (150 ммоль NaCl, 6 ммоль Трис-HCl, pH 7,9, 6 ммоль MgCl2, 7 ммоль 2-меркаптоэтанола, 100 мкг/мл BSA), 2 мкг органического фермента XbaI (Bochringer Mannheim, 24 ед.) и 178 мкл воды. Реакционная смесь осторожно перемешивается и инкубируется при 37oC в течение 2 ч. После этого инкубирования вводятся 4 мкл 5 M NaCl и 2 мкл ограничительного фермента Bam HI (New England Biolabs, 20 ед.), и инкубирование продолжается еще в течение 2 ч при 37oC. Затем ДНК осаждается эталоном и ацетатом натрия стандартным приемом и подвергается электрофорезу на 1%-ном низкоплавком агарозном геле. Верхний слой, соответствующий векторной ДНК XbaI/Bam HI, выделяется путем осуществления процедуры в колонке Elutip-d. После осаждения и сушки ДНК выдерживается в 30 мкл 10 мМ Трис, pH 8.
Примерно 5 мкл векторной ДНК pCZR126S, вываренной с XbaI/Bam HI, смешиваются с 10 мкл ограничительного фрагмента XbaI/XmaI от pRB145 или pRB172 и 4 мкл киназированного связующего XmaI/Bam HI, приготовленного, как указано выше, в буферном растворе (50 мкл), содержащем 50 мкмоль Трис (pH 7,6), 10 ммоль MgCl2, 10 ммоль ДTT, 800 мкмоль АТР и 6 ед. T4 ДНК лигазы. Реакционная смесь инкубируется при 4oC в течение ночи и используется для трансформации штамма E.Сoli K12 RV308 согласно процедуре примера 29 A3. Желаемые трансформанты E. Сoli K12 RB308/pRB173 (если они образованы от векторного фрагмента pRB172) и E.Сoli K12 RV308/pRB174 (если они образованы от векторного фрагмента pRB145) идентифицируются путем анализа их плазмидной ДНК и белка до и после индукции при 42oC. Ограничительная точка и функциональная карта плазмиды pRB173 представлены на фиг. 21.
Примерно 200 нг ДНК плазмиды от мини-препаративной обработки трансформируются в клетки E. Сoli 201, и трансформанты отбираются на чашках с 2X ТУ-агаром, содержащих 15 мкг/мл тетрациклина. Белки, выраженные этими трансформантами, анализируются, и желаемые трансформанты подтверждаются по продуцированию человеческого инсулина. Таким образом выделяется E.Сoli L201 pRB173 и E.Сoli L201/pRB174.
D. Построение Escherichia Сoli K12 RV308/pRB175, pRB176, pRB177 и pRB178.
Примерно 12 мкг плазмиды pRB172 суспензируются в 15 мкл 10X буфера NdeI (100 ммоль NaCl, 50 ммоль Трис-HCl, pH 7, 5, 10 ммоль MgCl2, 1 ммоль ДTT, 100 мкг/мл BSA), 2,5 мкл ограничительного фермента NdeI (20 ед.) и 133 мкл воды. Реакционная смесь осторожно перемешивается и инкубируется при 37o C в течение ночи. После инкубирования ДНК осаждается этанолом и ацетатом натрия стандартными приемами, высушивается и повторно суспензируется в 15 мкл 10X буфера DraIII (50 ммоль Трис-HCl, pH 7,5, 20 ммоль NaCl, 10 ммоль MgCl2, 1 ммоль ДTT), 5 мкл ограничительного фермента DraIII (20 ед.) и 130 мкл H2O. После осторожного перемешивания реакционная смесь инкубируется при 37oC в течение ночи. ДНК снова осаждается этанолом и натрийацетатом, как указано выше, и подвергается электрофорезу на 15%-ном низкоплавком агарозном геле. Желаемый крупный ограничительный фрагмент срезается с геля, и ДНК извлекается путем осуществления процедуры в колонке Elutip-d. ДНК выдерживается в 30 мкл 10 мМ Трис-HCl, pH 8.
Связывающие звенья ДНК, представленные на фиг. 31, химически синтезируются с помощью синтезатора Applied Biosystems (модели 380B).
После очистки каждого олигонуклеотида путем электрофореза на акриламидном геле они фосфорилируются согласно описанной выше процедуре и ренатурируются для ускорения сшивки и построения ДНК фрагментов, кодирующих аналог гена человеческого проинсулина. Связывающие звенья от (i) до (iv) используются для построения плазмид pRB175, pRB176, pRB177 и pRB178 соответственно.
Примерно 3 мкл фрагмента ДНК вектора pRB172, вываренного с NdeI/DraIII, смешиваются с 4 мкл каждого из киназированных связывающих звеньев (i-iv) в буферном растворе (50 мкл), содержащем 50 мкмоль Трис-HCl, pH 7,6, 10 ммоль MgCl2, 10 ммоль ДТT, 800 мкмоль АТР и 6 ед. Т4 ДНК лигазы. Реакционная смесь инкубируется при 4oC в течение ночи, и эта смесь используется для трансформации клеток E.Сoli K12 RV308 согласно процедуре примера 29 A3. Желаемые трансформанты E. Сoli K12 RV308/pRB175, E.Соli K12 RN308/pRB176, E.Сoli K12 RV308/pRB177 и E.Сoli K12 RV308/pRB178 идентифицируются путем анализа их ДНК плазмиды и продуцирования белка до и после индукции клеток при 42oC. Ограничительная точка и функциональная карта плазмиды pRB175 представлены на фиг. 22.
Аналогии человеческого проинсулина приведены в табл. 1.
E. Исходная культура E.Coli K12 RV308/pRB172 приготавливается в форме перманентной исходной культуры следующим образом.
Выделенные колонии высеваются на нижеследующих фенотипных чашках: L-агар + 5 мкг/мл Tc (тетрациклина), L-агар + Sm (стрептомицинсульфат), агаровая среда M-9 (солевая среда, содержащая MgSO4, тиамин, HYC ACE и глюкозу) и агаровая среда M-9, содержащая глюкозу и не содержащая лактозу. Одна чашка с L-агаром/Tc инкубируется при 42oC, остальные чашки инкубируются при 30oC в течение 24 ч. Фенотипно правильная колония, отобранная с чашки L-агар/Tc при 30oC, используется для инокулирования в колбе, содержащей 50 мл L-питательного бульона (с содержанием триптона, дрожжевого экстракта, NaCl и глюкозы) плюс 5 мкг/мл среды Tc. Эта колба инкубируется при 30oC со встряхиванием в течение 7 ч. Материал в колбе (1,2 мл) распределяется в бис-замораживающие ампулы и замораживается в парофазном жидком азоте. Затем материал оттаивает и используется в количестве 1 мл для инокуляции в колбе, содержащей 50 мл L-бульона плюс 5 мкг/мл Tc. Эта колба инкубируется при 30oC со встряхиванием в течение 4 ч. Две колбы, содержащие 500 мл среды BC-7 (содержит MgSO4, тиамин, глюкозу, Tc и изотопные соли), инокулируются с 1 мл каждой из указанных выше сред, и эти колбы инкубируются со взбалтыванием в течение 9 ч при 30oC.
150-литровый ферментер, содержащий 80 л среды (содержит лимонную кислоту, сульфат железа-аммония, K2HPO4, NH4Cl, NaH2PO4, CaCl2, MgSO4, глюкозу и изотопные неорганические соли), инокулируется в 1 л питательного бульона BC-7. Ферментер функционирует при 30oC до тех пор, пока скорость выделения двуокиси углерода не достигнет значения 1 ммоль/л/мин. В этот момент 50 л питательного бульона вводится в ферментер для получения клеточного инокулума.
2500-литровый ферментер, содержащий 1500 л той же среды, что используется в 150-литровом ферментере, функционирует при 30oC до тех пор, пока скорость выделения двуокиси углерода
не достигнет значения 1 ммоль/л/мин, и затем температура повышается до 40oC. Функционирование ферментера продолжается еще в течение 8 ч. Затем питательный бульон инактивируется нагревом при
61oC в течение 7 мин. Из инактивированного нагреванием бульона получаются нижеследующие выводы:
выход клеток 13,1 г/л на сухой вес; эффективность продукта 273 мкг DES-формилата/мл
питательного бульона; удельная активность (например, г продукта/г клеток) 2,1%; процент формилированного продукта 13%.
Бульон брожения подается в сборник из нержавеющей стали, в котором температура поддерживается от 2 до 8oC. Затем весь питательный бульон центрифугируется или фильтруется для сбора клеток E.Coli так, чтобы выход твердых продуктов в расчете на сухой вес составлял 15-20%. Твердый продукт, составляющий собранные клетки, снова суспензируется в технологической воде. Затем клетки разрываются с использованием гомогенизации под высоким давлением или других подходящих методов так, чтобы лизис клеток составлял 90-99%. Гомогенизированная суспензия разбавляется до 5% сухой массы технологической водой. Величина pH разбавленной разорванной клеточной суспензии доводится до значения в пределах от 7 до 10 путем ввода гидрата окиси натрия. Инклюзионные образования (гранулы) отделяются от клеточных осколков путем дифференциального центрифугирования. Полученный концентрат содержит от 15 до 20% твердых веществ.
Эти инклюзионные образования (гранулы) солюбилизируются при щелочном значении pH в присутствии цистеина и мочевины. Эти солюбилизированные гранулы подвергаются катионообменной хроматографии на текучей сефарозе SP для отделения смешанной дисульфидной формы проинсулинового аналога от общего объема гранулированного материала.
Метионил-тиразоновое удлинение аминового конца аналога проинсулина удаляется путем инкубирования с катепсином C дипентидиламинопептидазой. Реакция катепсинового расщепления завершается путем сульфитолиза тетратионатом калия и цистеином при pH 8,8.
После вытеснения растворителем через Sephadex C-25 полученный S-сульфонат проинсулинового аналога подвергается перегибу путем инкубации при величине pH, равной 10,6, в присутствии цистеина. Процесс перегиба подавляется путем доведения величины pH до 2,4. Проинсулиновый аналог с должным перегибом отделяется от материала с неправильным перегибом, а также от загрязнений, оставшихся после этапа расщепления катепсина C, путем хроматографического разделения с гидрофобным взаимодействием на смоле SP20SS. Это разделение происходит с использованием ацетонового градиента, так что органический растворитель должен быть удален из основного потока, путем выпаривания перед дальнейшей операцией.
После выпаривания органического растворителя проинсулиновый агент с правильным перегибом инкубируется с трипсином и карбоксипептидазой B. Эти ферменты эксцизируют C-пептид аналога проинсулина и образуют человеческий инсулин Lys (B28), Pro (B29). Этот аналог инсулина затем дополнительно очищается путем катионообменной хроматографии на S-сефарозе с последующей хроматографией высокого разрешения с обратимой фазой на смоле C-8 (10 мкм). Ацетонитрил удаляется из основного потока обратимой фазы путем вытеснения растворителем на Sephadex C-25, и полученный продукт концентрируется с использованием спирально намотанной системы ультрафильтрации. Концентрированный продукт подвергается хроматографии исключения на смоле Sephadex C-50 и лиофилизируется до сухого состояния.
В табл. 2 представлены данные аминокислотного анализа соединения, отвечающие изложенным выше примерам.
Физиологические эффекты аналогов инсулина, отвечающих настоящему изобретению, показаны в нижеследующей испытательной системе в условиях in vivo.
В качестве испытательных животных используются обычные самцы крыс Sprague Dawley, которые получены из Charles River Laboratories. Полученные крысы имеют вес в диапазоне от 160 до 180 г, и их выдерживают в течение 1 нед в помещении при 75oF (24oC) с регулярным световым циклом (с включенным светом с 7 вечера до 7 утра и с выключенным светом с 7 утра до 7 вечера). Рацион крыс составляет капуста Пурина 5001 ad libitum. Крысы, используемые для каждого испытания, подвергаются голоданию в течение примерно 16 дн до их использования. При первом использовании их вес составляет примерно 200 г. При достижении веса примерно 275 г (в период за 3 нед) животных уже больше не используют. Одна группа из 10 крыс-самцов используется ежедневно для каждого испытываемого соединения (то есть биосинтезированный человеческий инсулин, свиной инсулин и аналог человеческого инсулина). В течение недели каждая группа используется лишь однократно. Десять крыс были разделены на две группы по пять крыс в каждой. Одна группа служит в качестве контрольной, и в нее вводится лишь носитель путем подкожной инъекции. Другая группа, состоящая из пяти крыс, подвергается обработке испытываемым соединением. Белки растворяются в 0,05 н. HCl (pH 1,6), и в результате получается исходный раствор концентрацией 100 мкг на мл. Из него получают ряд разбавленных растворов в обычном солевом растворе, который вводится путем подкожной инъекции в организм крыс. 100 мкл образца крови берут их хвостовой вены у каждой крысы при нулевом моменте времени и через 30 мин, 1 ч, 2 ч, 3 ч и 4 ч после еды. Глюкозу определяют колориметрическим способом оксидазы глюкозы (Sigma Chemical Co). Для каждой крысы рассчитывают изменение содержания глюкозы в крови от нулевого момента времени, и конечные результаты выражены как среднее изменение процента ± SEM для экспериментальной группы с корректировкой на среднее изменение для контрольной группы для того же дня.
Кривая доза - реакция была построена по данным испытания с 4-7 различными концентрациями испытываемого соединения с использованием максимальной реакции в заданный момент времени (обычно 1 или 2 ч после ввода белка). Из данной кривой определяют значение ED50, как значение вводимой подкожной дозы белка, которая дает половину максимальной гипогликемической реакции. Результаты представлены в табл. 3.
Как указывалось выше, аналоги инсулина, отвечающие данному изобретению, имеют пониженную способность к димеризации или, иными словами, способность к самоассоциированию высокомолекулярных форм. Так, при вводе одного или нескольких указанных аналогов достигается очень быстрое инициирование активности. Аналоги инсулина, отвечающие данному изобретению, эффективны при лечении гипергликемии при вводе в организм пациента, нуждающегося в этом лечении, эффективного количества аналога инсулина формулы (I). Под используемым термином "эффективное количество" имеется в виду один или несколько аналогов инсулина, отвечающих данному изобретению, которые необходимы для снижения или поддержания уровня сахара в крови, либо терапевтически, либо профилактически. Это количество обычно может находиться в пределах примерно от 10 до 60 ед. или более в день (или примерно от 0,3 до 2 мг, что составляет примерно 29 ед. на миллиграмм). Однако следует иметь в виду, что количество аналога (аналогов) инсулина, которое фактически вводится в организм, должно определяться лечищим врачом в соответствии с обстоятельствами, включающими состояние заболевания (то есть причину, вызвавшую гипергликемию), тип используемого аналога инсулина, который выбран для ввода в организм, выбранный парентеральный способ ввода, возраст пациента, вес и реакцию каждого пациента и остроту симптомов пациента. Следовательно, указанные выше пределы дозы никак не могут являться ограничением объема данного изобретения.
Аналоги инсулина, отвечающие данному изобретению, вводятся в организм пациента, нуждающегося в этом лечении (то есть пациента, страдающего от гипергликемии), с использованием фармацевтических композиций, содержащих эффективное количество по меньшей мере одного аналога инсулина формулы (I) в композиции с одним или несколькими фармацевтически пригодными эксципиентами или носителями. Для этих целей фармацевтические композиции могут быть приготовлены таким образом, что будут содержать примерно 100 ед. на мл или аналогичные концентрации с содержанием эффективного количества аналога (аналогов) инсулина. Эти композиции обычно (хотя и необязательно) парентеральны по своей природе и могут быть приготовлены любым способом с применением обычных эксципиентов или носителей для вводимых парентерально продуктов, которые уже известны (см. , например, публикацию Remington's Pharmaceutical Sciences, 17-е изд. , Mack Publishing Company, Easton PA, USA, 1985). Так, например, дозированные формы для парентерального ввода могут быть приготовлены путем суспензирования или растворения желаемого количества по меньшей мере одного аналога инсулина формулы (I) в нетоксичном жидком носителе, пригодном для инъекции, таком как водная среда и стерилизующая суспензия или раствор. Как возможный вариант, измеренное количество соединения может быть помещено в ампулу, и ампула и ее содержимое стерилизуются и запаиваются. Ампула или носитель могут быть подготовлены для целей смешивания до ввода в организм. В фармацевтических композициях, пригодных для парентерального ввода в организм, используются разбавители, эксципиенты и носители, такие как вода и смешиваемые с водой органические растворители, такие как глицерин, сезамовое масло, арахисовое масло, водный пропиленгликоль, N,N'-диметилформамид и т.д. Примерами таких фармацевтических композиций являются стерильные изотонические водносолевые растворы аналогов инсулина формулы (I), которые могут быть приготовлены в форме буферного раствора с фармацевтически пригодным буфером и которые свободны от пирогенных веществ. Кроме того, вводимая парентерально фармацевтическая композиция может содержать предохраняющие средства, такие как мета-крезол, и другие вещества для регулирования величины pH конечного продукта, такие как гидрат окиси натрия или соляная кислота.
Аналоги инсулина, отвечающие данному изобретению, могут быть приготовлены также в форме фармацевтических композиций, пригодных для ввода через нос. Такие композиции описываются подробно в Европейской патентной заявке N 2200383. Такие композиции приготавливаются вместе с одним или несколькими фармацевтически пригодными разбавителями, фармацевтически приемлемым количеством соли щелочного металла, соли аммония или свободной кислоты свободного от цинка инсулина, и при желании усиливающим абсорбцию количеством по меньшей мере одного усиливающего абсорбцию агента, выбранного из группы, включающей олеиновую кислоту или ее сложный эфир, жидкую форму сложного эфира сорбита и жирной кислоты, жидкую форму полиоксиэтиленового производного сложного эфира сорбита и жирной кислоты и жидкую форму сополимера оксиполиоксиэтилен-полиоксипропилен-полиоксиэтилен.
Для демонстрации эффективности внутриносового ввода аналога человеческого инсулина Lys (B28), Pro (B29) в сопоставлении с человеческим натриевым инсулином приводится нижеследующее исследование. Самцы коротконогих гончих собак весом от 10 до 12 кг поддерживаются в прекрасном физическом состоянии до и в ходе данного исследования. Собаки выдерживаются в режиме голодания в течение 16 ч, но они имеют доступ к воде для гарантии предотвращения непредвиденных колебаний содержания инсулина. В течение всего исследования собак анестезируют пентабарбитолом натрия, и температура их тела сохраняется посредством нагревательных подушек для сведения к минимуму колебания содержания глюкозы. Испытательные образцы человеческого инсулина (то есть Lys (B28), Pro (B29) и человеческого натрийинсулина) растворяются в дистиллированной воде, и величина pH раствора доводится до 7,5 посредством гидрата окиси натрия. Конечная концентрация инсулина составляет 70 ед. на мл. Человеческий инсулин Lys (B28), Pro (B29) дозой 0,8 ед. на кг вводится в каждую из восьми собак, и человеческий натрийинсулин вводится дозой 0,8 ед. на кг в каждую из четырех собак. Все дозы вводятся через нос путем ввода струи через ноздрю с помощью дозировочного распылителя и модифицированного носового аппликатора. Образцы крови отбираются из шейной вены за 30, 15 и 0 мин до ввода и через 10, 20, 30, 45, 60, 90, 120, 180 и 240 мин после ввода для измерения снижения глюкозы в крови. Результаты данного исследования приводятся в табл. 4.
При осуществлении описанной выше методики сопоставляется профиль снижения глюкозы крови при внутриносовом вводе человеческого инсулина Lys (B28), Pro (B29) с профилем снижения при внутривенном и подкожном вводе аналога. Для внутривенного ввода раствор инсулина, содержащий аналог человеческого инсулина Lys (B28), Pro (B29), вводится путем инъекции ударной дозы (0,1 ед. на кг) в подкожную вену ног каждой из четырех собак. Образцы крови берутся за 30, 15 и 0 мин до ввода и через 5, 10, 15, 20, 30, 45, 60, 90, 120, 180 и 240 мин после ввода. Для подкожного ввода раствор инсулина, содержащий аналог человеческого инсулина Lys (B28), Pro (B29), вводится под эпидермис (0,2 ед. на кг) другой боковой стороны живота каждой из шести собак. Образцы крови берутся таким же образом, как в случае носового ввода, как описано выше. Результаты данного сопоставительного исследования приводятся в табл. 5.
Реферат
Использование: в медицинской практике для снижения уровня глюкозы в крови. Сущность изобретения: аналоги инсулина, модифицированные в положении 29 его цепи и при желании в других положениях, имеют модифицированные физико-химические и фармокинетические свойства. 6 з.п. ф-лы, 5 табл., 31 ил.

Формула
или его фармацевтически приемлемая соль, отличающийся тем, что А21 является аспарагином; В1 является фенилаланином; В2 является валином; В3 является аспарагином; В9 является серином; В10 является гистидином или аспарагиновой кислотой; В27 является треонином; В28 является любой природной L-аминокислотой, В29 является L-пролином; В30 является треонином.
Комментарии