Способ лечения онкологических заболеваний - RU2387456C1
Код документа: RU2387456C1
Чертежи
Описание
Изобретение относится к медицине и может быть использовано при лечении пациентов препаратами фрагментированной ДНК человека в инъекционной форме или в форме микроклизм в нуклеолитически стабильном комплексе с гистоноподобным белком протамином, использующимся как лейкостимулирующее в схемах лечения онкологических заболеваний и как противораковое средство.
Известен способ лечения пациентов с использованием препарата, обладающего противоопухолевым, антитоксическим и радиопротекторным действием, представляющим собой фрагментированный до 200-2000 пар нуклеотидов нуклеопротеидный комплекс с содержанием, мас.%: двухспиральной ДНК 60-80 и белков ядерного матрикса 20-40, при получении которого используют видоспецифическое биологическое сырье - плаценту человека [RU 2234323, А61К 48/00, 2004, 27.05.2003. Препарат, обладающий противоопухолевым, антитоксическим и радиопротекторным действием].
Недостатком способа является относительно низкая эффективность лечения.
Наиболее близким по своей сущности к предлагаемому является способ лечения онкологических заболеваний, основанный на введении в организм пациента фрагментированной ДНК, в качестве которой используют гомологичную ДНК, имеющую биологически активный размер, составляющий полный геном физиологически и генетически здорового донора, при этом ДНК вводят в дозе, создающей такую концентрацию ДНК в плазме крови, которая равна или превышает концентрацию собственно ДНК плазмы крови, но не превышает максимально допустимую концентрацию, равную 30 мкг/мл [RU 2313349, A61K 48/00, 2006, 16.01.2006. Способ лечения онкологических заболеваний].
В этом способе фрагментированная ДНК не защищена от действия нуклеаз крови человека и в клетки доставляются преимущественно короткие фрагменты ДНК. Для доставки более крупных фрагментов экзогенной ДНК используют максимально допустимые дозы фрагментированной ДНК (приближающейся к 1500 нг/мл). Только в этом случае в клетку доставляется некоторое количество полноразмерных фрагментов ДНК, которые в состоянии репарировать зоны повреждения, связанные с утратой одного нуклеотида или фрагмента хромосомной ДНК.
В этом проявляется недостаточная эффективность способа лечения, поскольку даже при максимально возможных концентрациях терапевтической ДНК в крови, равной 1500 нг/мл, не на все клетки, где произошло выпадение генетического материала, хватит сохранивших свой полный размер фрагментов экзогенной ДНК.
Все это снижает эффективность лечения лейкопений, вызванных применением цитостатических препаратов, и эффективность лечения пациентов с онкологическими заболеваниями, поскольку действующее начало - фрагментированная ДНК - подвергается незамедлительной нуклеолитической деградации нуклеазами плазмы крови, и для достижения терапевтического эффекта приходится применять максимально допустимые концентрации препарата.
Требуемый результат заключается в повышении эффективности лечения пациентов, проходящих программную химиотерапию, и пациентов с онкологическими заболеваниями.
Требуемый результат достигается тем, что по способу лечения пациентов с лейкопениями, вызванными программной химиотерапией, и пациентов, больных онкологическими заболеваниями, основанном на введении в организм фрагментированной ДНК человека, фрагментированную ДНК человека вводят в комплексе с негистоновым белком протамина.
Кроме того, требуемый результат достигается тем, что введение в организм фрагментированной ДНК человека в комплексе с негистоновым белком протамина производят парентерально или микроклизмированием.
Кроме того, требуемый результат достигается тем, что соотношение фрагментированная ДНК/негистоновый белок протамин составляет 1/0,8.
Кроме того, требуемый результат достигается тем, что введение фрагментированной ДНК человека в комплексе с негистоновым белком протамином производят при концентрации фрагментированной ДНК человека, равной 25-1000 нг/мл.
Воздействие фрагментированной ДНК человека в комплексе с негистоновым белком протамином при концентрации 25 нг/мл означает, что это в 1000 раз меньше предельно допустимой концентрации, равной 30 мкг/мл, и в 10 раз меньше концентрации фрагментированной ДНК, не ассоциированной с протамином, при которой впервые появляется эффект воздействия.
В более широком плане задачей настоящего изобретения является разработка экспериментально подтвержденного способа совместного воздействия препарата фрагментированной ДНК человека и негистонового белка протамина при лечении пациентов с лейкопениями, вызванными химиотерапиями, применяемыми в схемах лечения онкозаболеваний, и пациентов с онкологическими заболеваниями, когда фрагменты терапевтической ДНК защищаются комплексом протамина от действия нуклеаз крови. Стабилизация фрагментов фрагментированной ДНК человека, в качестве которой может быть использован препарат «Панаген», негистоновым белком протамином расширяет его терапевтические возможности, выражающиеся в том, что при осуществлении гомологичной рекомбинации происходит захват более протяженных участков генома, что повышает вероятность попадания в зону замещения мутаций, приведших к сбою в пролиферативной активности у неотрансформированных клеток, или дефектов в геноме СКК, сформированных в результате жесткой химиотерапии или гамма-облучения.
В современной литературе отсутствуют указания на предлагаемый способ лечения пациентов с лейкопениями, вызванными программными химиотерапиями, и пациентов с онкозаболеваниями, при котором используется комплекс фрагментированной ДНК человека с негистоновым белком протамином.
Следовательно, предложение отвечает критериям новизны и изобретательского уровня.
Ниже приводятся теоретические и экспериментальные данные, подтверждающие, что изобретение отвечает и критерию практической (промышленной) применимости.
На чертежах представлены:
на фиг.1 - электрофоретическая характеристика экзогенной терапевтической ДНК плаценты человека, используемой для инъекций в организм опытных мышей, где 1 - маркер молекулярного веса BssT1I гидролизат фага λ, цифрами слева от блока отмечены фрагменты соответствующего молекулярного веса в т.п.н.;
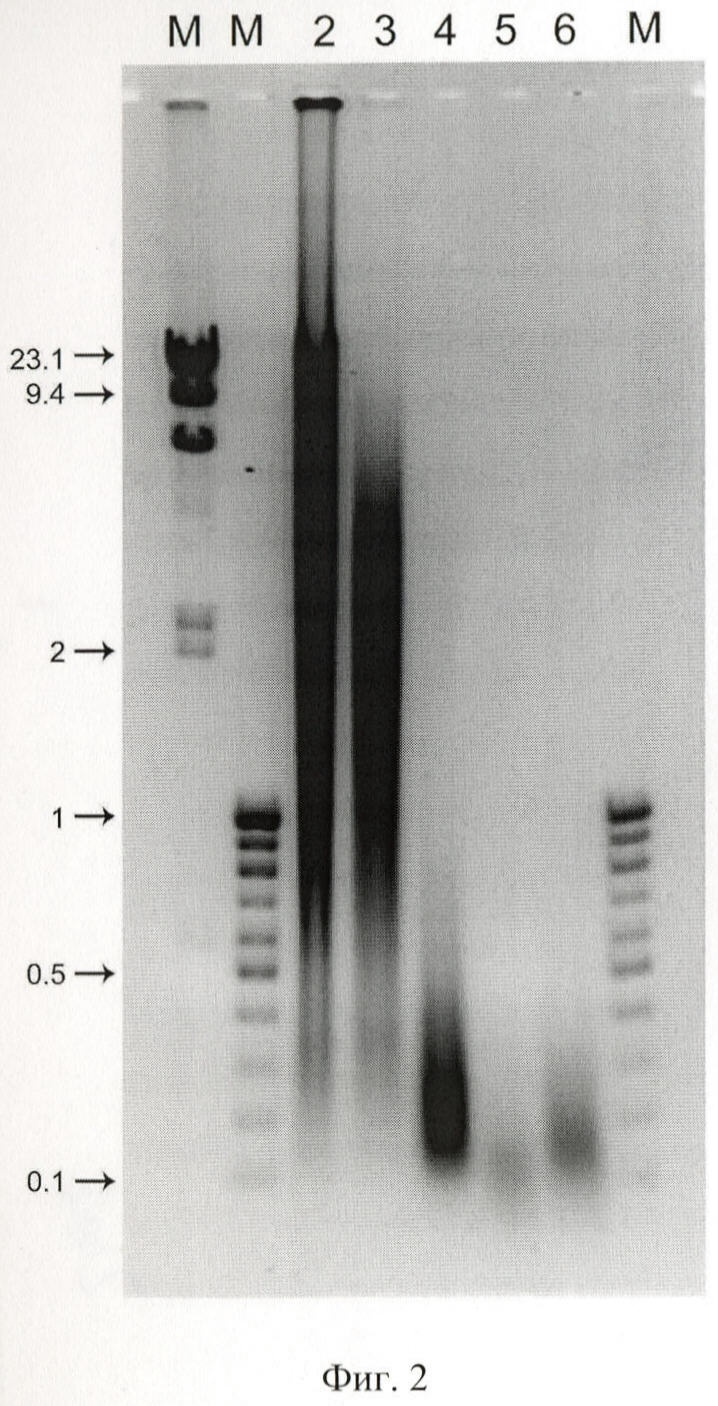
на фиг.2 - нуклеазная активность сыворотки крови мыши, где М - маркеры молекулярных весов, цифрами указаны размеры соответствующих фрагментов в т.п.н.; 2 - исходная ДНК человека, фрагментированная гидродинамическим способом до фрагментов размером от 200 до >6000 п.о.; 3 - ДНК человека, инкубированная с 1% сывороткой крови мыши, 4 - с 10% сывороткой крови мыши, 5 - с 50% и 6 - с 97% сывороткой крови мыши;
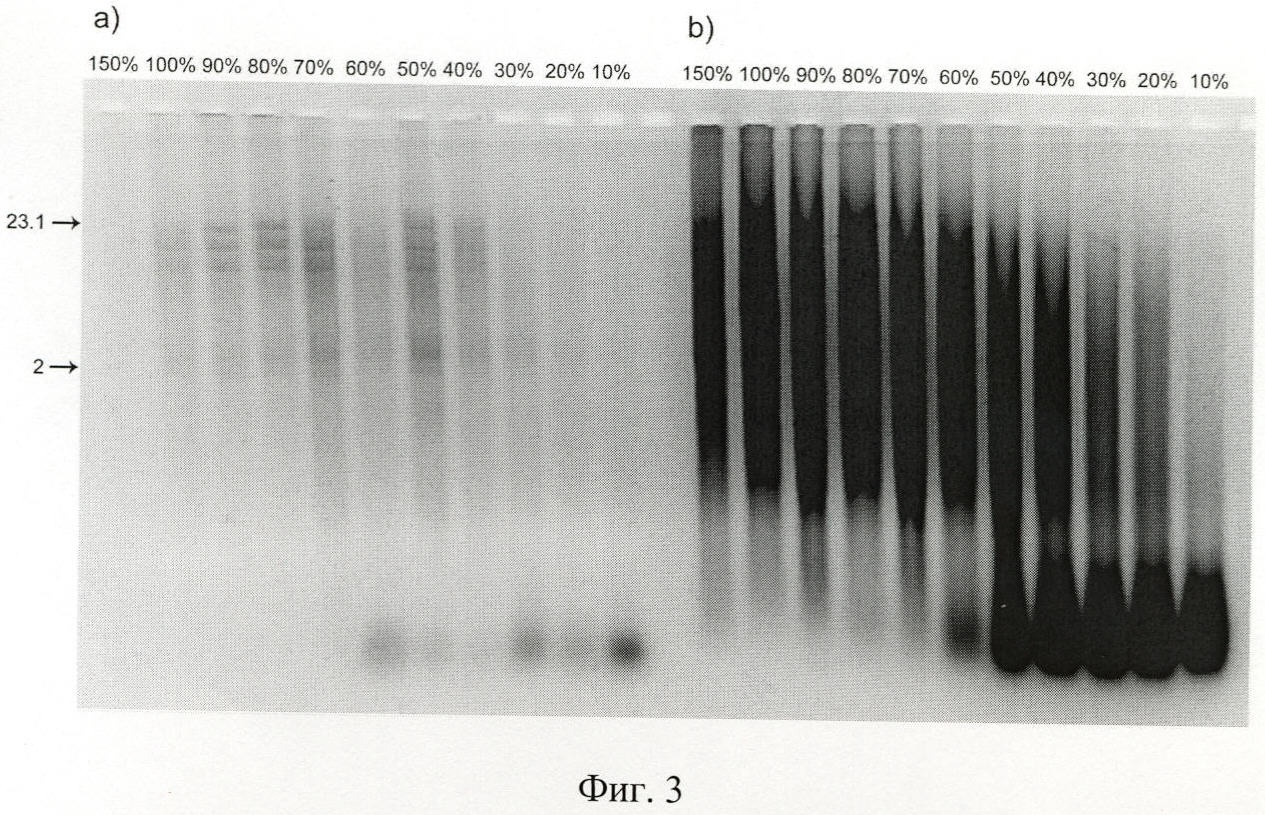
на фиг.3 - защита ДНК, ассоциированной с различным (по массе) количеством протамина, от действия нуклеаз при инкубации в 10% сыворотке крови мыши, где а) - Hind III гидролизат ДНК фага λ, b) - фрагментированная ДНК человека. Сверху геля указано содержание протамина в комплексе по отношению к ДНК в %. Слева отмечены фрагменты соответствующего молекулярного веса в т.п.н.;
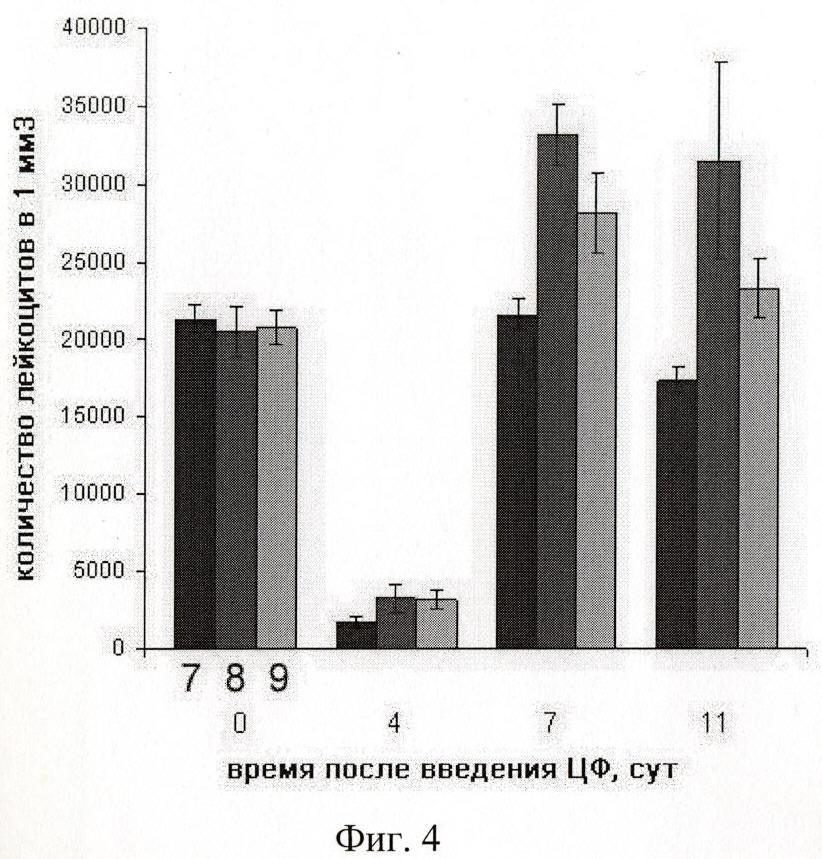
на фиг.4 - сравнение лейкостимулирующей активности экзогенной ДНК человека (8) и ДНК человека, ассоциированной с протамином (9), введенной мышам после воздействия цитостатика циклофосфана; 7 - контрольная группа мышей, без воздействия ДНК;
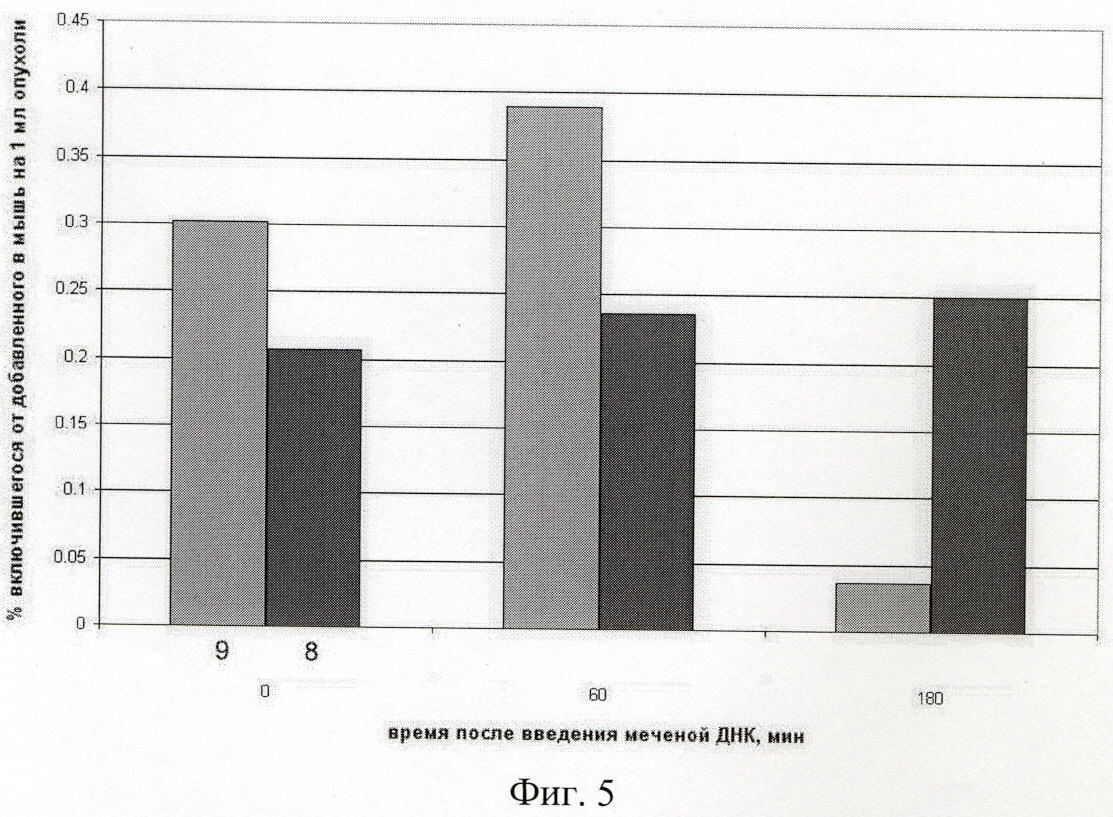
на фиг.5 - количество меченого материала (9 - ДНК, ассоциированной с протамином, или 8 - незащищенной ДНК), находящееся в опухоли в различное время после инъекции;
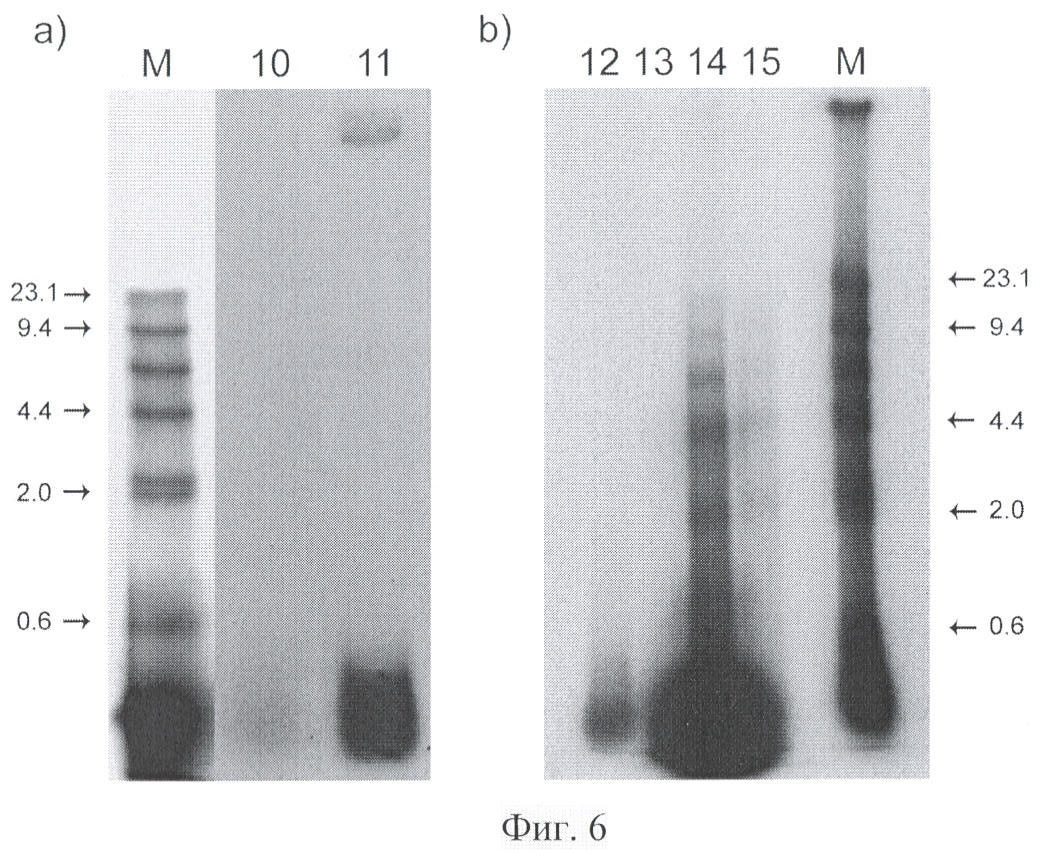
на фиг.6-a) электрофореграмма меченой λ Hind III ДНК после ее внутривенного введения мышам и выделения межхромосомной фракции клеток опухоли мышей в солидной (10) и асцитной (11) форме; b) электрофореграмма меченой λ Hind III ДНК, свободный (12, 13) и связанной с протамином (14, 15), выделенной из крови мышей через 5 мин (12 и 14) и 20 мин (13 и 15) после ее внутривенного введения. М - маркер молекулярного веса, Hind III гидролизат ДНК фага λ, цифрами отмечены фрагменты соответствующего молекулярного веса в т.п.н. ДНК доставляется в опухоль в деградированном виде. Незащищенная ДНК уже через 5 мин деградирует под воздействием нуклеаз крови. ДНК, ассоциированная с протамином, присутствует в крови через 20 мин в высокомолекулярной форме;
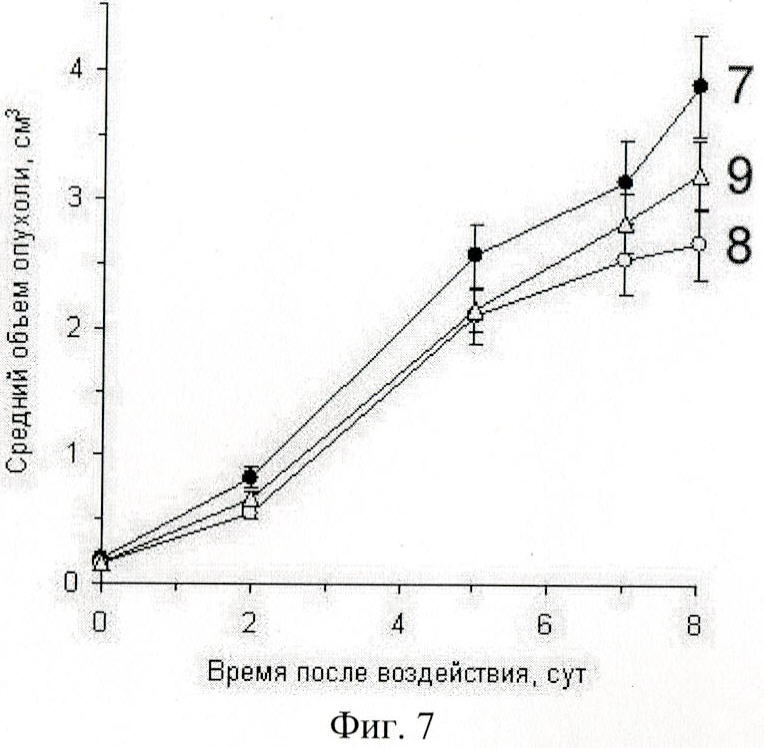
на фиг.7 - действие на рост экспериментальной опухоли мыши экзогенной ДНК в незащищенной форме (8) и ассоциированной с протамином (9);
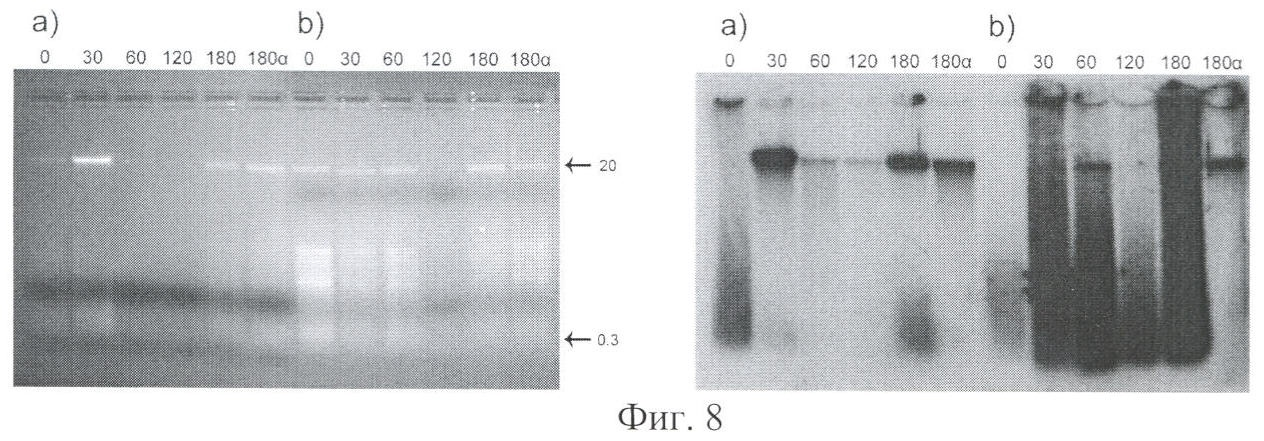
на фиг.8 - морфологические отличия фрагментированной экзогенной ДНК, доставляемой в межхромосомное пространство клеток MCF-7 в комплексе с протамином (b) и без протамина (а). Слева - электрофоретическая характеристика в 0,7% агарозном геле экзогенной ДНК, обнаруживаемой в межхромосомном пространстве ядра клеток MCF-7 после их культивирования в среде, содержащей экзогенную ДНК в концентрациях: для незащищенной ДНК 100 мкг/мл, для защищенной протамином ДНК 25 мкг/мл. Справа - рентгенограмма агарозного блока после высушивания. Цифры над блоками указывают время инкубации (мин) культуры клеток с меченной α32Р экстраклеточной ДНК человека и α-P32dATP (180α); цифрами слева от блока отмечены фрагменты соответствующего молекулярного веса в т.п.н.;
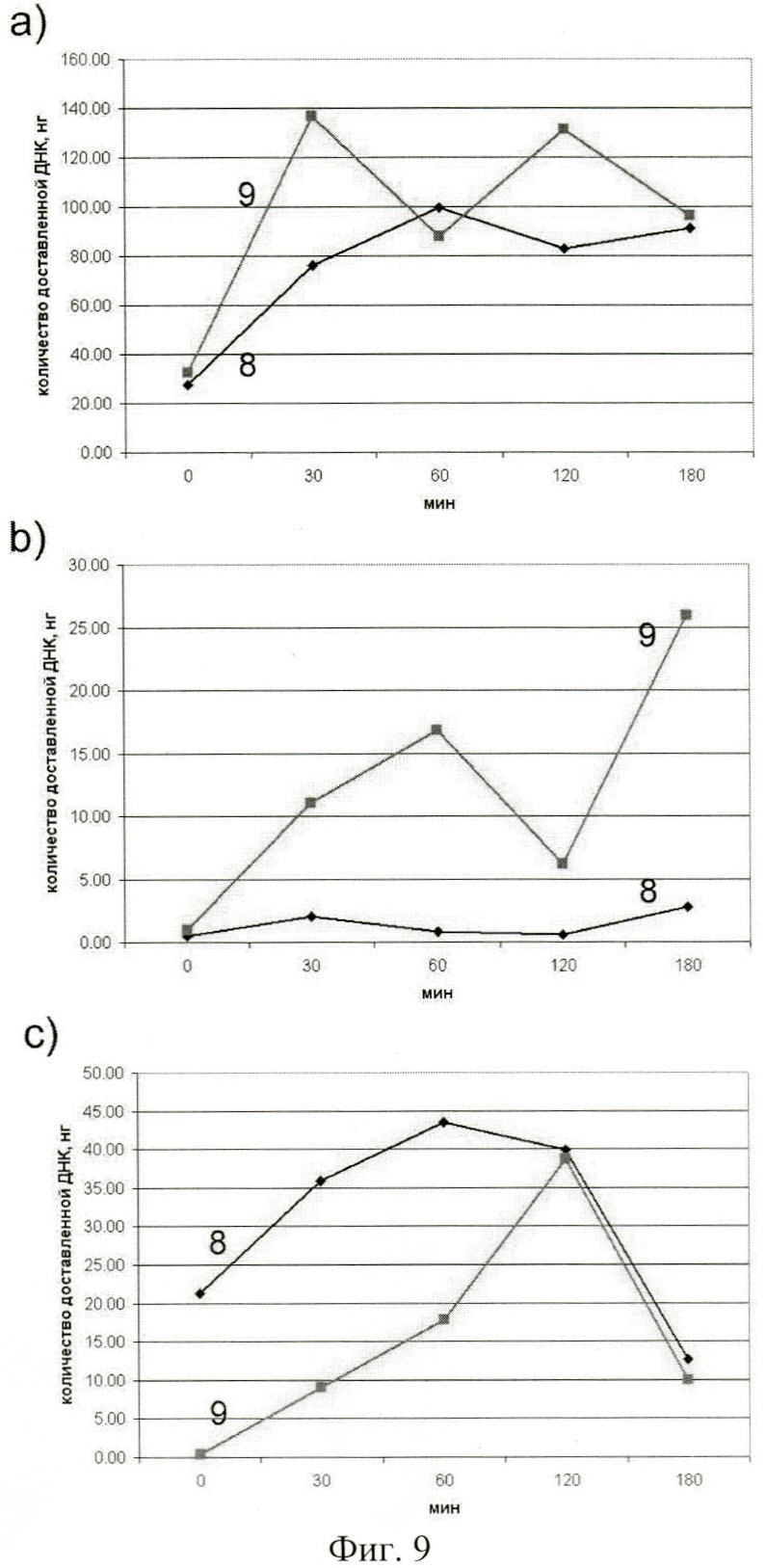
на фиг.9 - динамика доставки фрагментов экзогенной ДНК, ассоциированных с протамином (9) и незащищенных протамином (8), в цитоплазму (а) и межхромосомное пространство (b) ядер клеток MCF-7. Динамика появления меченого материала во фракции хроматина ядер клеток MCF-7 (с);
на фиг.10 - анализ индукции апоптоза клеток аденокарциномы молочной железы человека MCF-7, инкубированных в присутствии 100 мкг/мл незащищенной протамином фрагментированной ДНК плаценты человека (18), 25 мкг/мл ДНК человека, ассоциированной с протамином (19), в отсутствие ДНК (17). Культуры (17, 18, 19) индуцировали к апоптозу ФНО-α в концентрации 100 мкг/мл в течение 24 часов в среде, содержащей 10 мкг/мл циклогексимида. (16) - ДНК, выделенная из необработанных клеток MCF-7. М - маркер молекулярного веса, цифрами слева от блока отмечены фрагменты соответствующего молекулярного веса в п.н. В обоих случаях (18 и 19) в присутствии в культурной среде экзогенной ДНК индуцируется апоптоз и формируется лестница фрагментов ДНК, характеризующая апоптотическую деградацию ядерного хроматина;
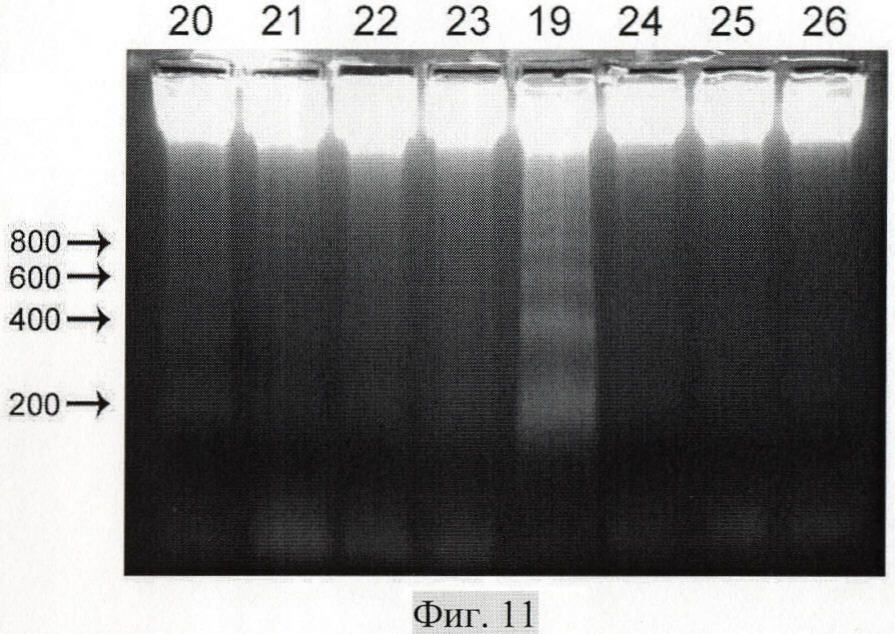
на фиг.11 - анализ индукции апоптоза клеток аденокарциномы молочной железы человека MCF-7, инкубированных в присутствии незащищенной протамином фрагментированной ДНК плаценты человека в концентрациях 25 мкг/мл (20), 2.5 мкг/мл (21), 0.25 мкг/мл (22) и 0.025 мкг/мл (23) и ДНК человека, ассоциированной с протамином в концентрациях 25 мкг/мл (19), 2.5 мкг/мл (24), 0.25 мкг/мл (25) и 0.025 мкг/мл (26); цифрами слева от блока отмечены фрагменты соответствующего молекулярного веса в п.н.
Предложенный способ лечения пациентов с лейкопениями, вызванными программными химиотерапиями в схемах лечения онкологических заболеваний, (лейкостимулирующее средство) и пациентов с онкологическими заболеваниями (противораковое средство) реализуется следующим образом.
Препарат фрагментированной ДНК человека, используемый в качестве монопрепарата как лейкостимулирующее и как противораковое средство в инъекционной форме или в форме микроклизм, смешивается непосредственно перед введением в организм с раствором негистонового белка протамина в соотношении 1 часть ДНК / 0.8 частей протамина для формирования нуклеолитически стабильного комплекса.
Эффективная концентрация ДНК в защищенном протамином комплексе может быть существенно ниже, чем фрагментированная ДНК в незащищенной форме (минимально в 4 раза), причем лейкостимулирующее и противораковое действие препарата фрагментированной ДНК человека остается без изменений.
Кроме того, при стабилизации фрагментов ДНК (препарат «Панаген») негистоновым белком протамином расширяются его терапевтические возможности, выражающиеся в том, что при осуществлении гомологичной рекомбинации происходит захват более протяженных участков генома, что повышает вероятность попадания в зону замещения мутаций, приведших к сбою в пролиферативной активности у неотрансформированных клеток, или дефектов в геноме СКК, сформированных в результате жесткой химиотерапии или гамма-облучения.
Пример реализации способа.
Препарат фрагментированной ДНК, используемый как лейкостимулирующее или противораковое средство, вводят в организм пациента в виде инъекций или микроклизм 2-6 раз в сутки по 5 мг активного вещества (фрагментированной ДНК человека). Непосредственно перед ведением в одном флаконе смешиваются препарат фрагментированной ДНК 5 мг (в растворе) и 4 мг белка протамина (в растворе). Нуклеолитически стабильный комплекс формируется немедленно (несколько секунд) при комнатной температуре. После смешения и формирования комплекса препарат вводится в организм пациента.
Для подтверждения того, что предложенный способ позволяет достигнуть требуемого результата, приведем его теоретическое и экспериментальное обоснование.
В наших ранних работах [Yakubov LA, Popova NA, Nikolin VP, Semenov DV, Bogachev SS, Os'kina IN. Extracellular genomic DNA protects mice against radiation and chemical mutagens. Genome Biol. 2003; 5: 3; Khegai II, Bogachev SS, Os'kina IN, Popova NA, Semenov DV, Shurdov MA, Yakubov LA. Changes in the symptoms of hypothalamic diabetes insipidus after treatment with homologous exogenous DNA. Dokl Biol Sci. 2004 May-Jun; 396: 233-235; Николин В.П., Попова Н.А., Себелева Т.Е., Стрункин Д.Н., Рогачев В.А., Семенов Д.В., Богачев С.С., Якубов Л.А., Шурдов М.А. Влияние экзогенной ДНК на рост экспериментальных опухолей. Вопросы онкологии. 2006. Т.52, № 1, с.66-69; Николин В.П., Попова Н.А., Себелева Т.Е., Стрункин Д.Н., Рогачев В.А., Семенов Д.В., Богачев С.С., Якубов Л.А., Шурдов М.А. Влияние экзогенной ДНК на восстановление лейкопоэза и противоопухолевый эффект циклофосфана. Вопросы онкологии. 2006. Т.52, №3, с.336-340; Rogachev VA, Likhacheva A, Vratskikh O, Mechetina LV, Sebeleva ТЕ, Bogachev SS, Yakubov LA, Shurdov MA. Qualitative and quantitative characteristics of the extracellular DNA delivered to the nucleus of a living cell. Cancer Cell Int. 2006 Oct 11; 6:23; Likhacheva AS, Nikolin VP, Popova NA, Dubatolova TD, Strunkin DN, Rogachev VA, Sebeleva ТЕ, Erofeev IS, Bogachev SS, Yakubov LA, Shurdov MA. Integration of human DNA fragments into the cell genomes of certain tissues from adult mice treated with cytostatic cyclophosphamide in combination with human DNA. Gene Ther Mol Biol. 2007; 11: 185-202; Likhacheva AS, Nikolin VP, Popova NA, Rogachev VA, Prokhorovich MA, Sebeleva ТЕ, Bogachev SS, Shurdov MA. Exogenous DNA can be captured by stem cells and be involved in their rescue from death after lethal-dose γ-radiation. Gene Ther Mol Biol. 2007; 11: 305-314; Yakubov LA, Rogachev VA, Likhacheva AC, Bogachev SS, Sebeleva ТЕ, Shilov AG, Baiborodin SI, Petrova NA, Mechetina LV, Shurdov MA, Wickstrom E. Natural human gene correction by small extracellular genomic DNA fragments. Cell Cycle. 2007 Jul; 6(18): 2293-2301] мы описали плейотропное воздействие фрагментированной экзогенной ДНК на живую клетку. Были выделены в отдельные группы несколько отличающихся по механизму типов ее воздействия, которые проявлялись как следующие описанные явления.
1. Лейкостимуляция при миелосупрессии, вызванной цитостатическим препаратом циклофосфаном.
2. Радиопротекторый и радиотерапевтический эффект.
3. Ингибирующее воздействие на рост экспериментальных опухолей мышей.
4. Участие в репарации мутантного аллеля гена каспазы 3 клеток аденокарциномы молочной железы человека MCF-7.
5. Способность интегрировать в реципиентный геном (фрагменты ДНК человека интегрируют в геном мыши) в ходе репарации межцепочечных сшивок, вызванных действием цитостатика циклофосфана.
В проведенных экспериментах при анализе указанных явлений использовалось большое количество экзогенной ДНК, что гарантировало ее участие в молекулярных процессах, определяющих указанные явления.
В настоящей работе мы использовали гистоноподобный белок протамин для защиты фрагментированной экзогенной ДНК от действия нуклеаз плазмы крови и культуральной среды во время роста культуры клеток. Как следует из результатов проведенных экспериментов, способность фрагментированной экзогенной ДНК, ассоциированной с протамином, проявлять лейкостимулирующее действие, ингибировать рост экспериментальной опухоли мыши и принимать участие в репарации мутантного аллеля гена каспазы 3 клеток аденокарциномы молочной железы человека MCF-7 сохраняется. Фрагменты экзогенной ДНК, защищенной протамином, не деградируют как в организме, так и в культуральной среде. Это позволяет использовать существенно меньшее количество экзогенной ДНК для определения указанных типов ее плейотропного действия.
Используемые методики.
Исследования проводились на мышах-самцах линии СВА разводки вивария Института цитологии и генетики СО РАН. Мышей содержали в пластиковых клетках по 10 особей в каждой со свободным доступом к пище и воде.
Препараты человеческой ДНК получали из плацент здоровых рожениц. Для выделения ДНК использовали безфенольный метод, который позволяет получить полноценный геном, сохраняя те фрагменты ДНК, которые in vivo прочно ассоциированы с белками ядерного матрикса (скафолда). Фрагментация ДНК осуществлялась ультразвуковым дезинтегратором при частоте 22 КГц, в результате чего получали смесь фрагментов ДНК размером от 200 до >6000 п.н. (фиг.1). Полученные препараты ДНК хранили в морозильной камере при ~18°С.
Для формирования комплекса с ДНК использовали раствор протамина сульфата 1% или 10 мкг/мкл для инъекций (Ферейн, Москва) и ДНК бактериофага λ, гидролизованную Hind III («Медиген», Новосибирск), или ДНК плаценты человека, фрагментированную до размеров от 200 до>6000 п.о.
При исследовании нуклеазной активности сыворотки крови мышей после совместной инкубации ДНК и сыворотки крови мышей добавляли протеиназу К и инкубировали 1 ч при 65°С. После экстракции фенол/хлороформом ДНК переосаждали 0,6 V изопропанола в присутствии линейного акриламида и проводили электрофорез в агарозных блоках.
В экспериментах использовали циклофосфан (ЦФ) производства ОАО «Биохимик» (г.Саранск). ЦФ перед применением растворяли в физрастворе и вводили внутрибрюшинно в дозе 200 мкг/кг веса животного.
Кровь для подсчета лейкоцитов брали из кончика хвоста индивидуально у каждой мыши непосредственно перед введением и на 4, 7 и 11 сутки после введения циклофосфана. Каждая экспериментальная группа состояла из 4 мышей. Кровь разводили в 20 раз в 3% растворе уксусной кислоты и в камере Горяева подсчитывали количество лейкоцитов в 1 мм3. Достоверность различий между данными определяли, используя t-критерий Стъюдента.
Исследование влияния ДНК на противоопухолевый эффект циклофосфана проводили на перевиваемой мышиной лимфосаркоме LS и ее субштамме RLS. LS была получена сотрудником ИЦиГ В.И.Калединым индукцией нитрозометилмочевиной у мыши линии СВА, переведена в асцитную форму и поддерживается на мышах этой линии. Эта опухоль является высокочувствительной к индукции апоптоза циклофосфаном и некоторыми другими алкилирующими агентами. Опухоль RLS получена из LS путем последовательных внутримышечных перевивок ее рецидивов, возникавших после воздействия возрастающими дозами циклофосфана, вследствие чего она приобрела перекрестную резистентность к индукции апоптоза и противоопухолевому действию алкилирующих препаратов. Противоопухолевый эффект того или иного воздействия оценивали по торможению роста опухоли и увеличению продолжительности жизни опухоленосителей. Достоверность различий между данными определяли, используя t-критерий Стъюдента.
Культуру клеток аденокарциномы молочной железы человека MCF-7 культивировали в среде RPMI 1640 с 10 мМ L-глутамина и 50 мкг/мл стрептомицина "Sigma" (США) при 37°С в присутствии 5% эмбриональной бычьей сыворотки (ФБС) «Биолот» (Россия) в атмосфере с 5% СО2 до плотности 0.7×107 клеток на 4 лунки в 24-луночном планшете.
Для приготовления ДНК и предшественника 10 мкг ДНК плаценты человека, фрагментированной до размеров 200-6000 п.о., метили ник-трансляцией в присутствии фрагмента Кленова, трех холодных и одного горячего dNTP. Для удаления невключившихся предшественников ДНК переосаждали 2 раза изопропанолом согласно процедуре, описанной у Гловера [Клонирование ДНК. Методы. Пер. с англ. под ред. Д.Гловера. М.: Мир, 1988. 538 с]. ДНК разводили в соответствующем объеме дистиллированной воды. ДНК и протамин смешивали в пропорции 1:1 по массе.
Образцы ДНК добавляли в каждую лунку планшета (в объеме, не превышающем 20 мкл на лунку). В каждой лунке находилось 25-35 мл среды. Аликвоту, 1 мкл из рабочего объема отбирали на счет радиоактивности.
В культуральную среду, в лунку, где культивировались клетки, добавляли меченую ДНК, обработанную, как описано выше. Клетки инкубировали с ДНК при 37°С, в течение необходимого времени в воздушном термостате. Время инкубации было выбрано: 0' (меченую ДНК добавляли в среду, содержащегося на льду титратора, сразу после этого среду отбирали и добавляли Тритон Х-100), 30'; 60'; 120'; 180'.
По завершении времени инкубации планшет помещали на лед. Среду количественно отбирали и немедленно добавляли Тритон Х-100 в буфере А [Roberts DB. Drosophila: A Practical Approach. IRL Press, Oxford, 1986] (буфер А содержал 2 мМ CaCl2, 0.5% Тритона Х-100). В каждую лунку добавляли 250 мкл лизирующего буфера. Клетки оставляли с лизирующим буфером на льду в течение 10'. Лизированные клетки ресуспендировали несколько раз и наносили на 1 мл градиента 10% сахарозы на буфере А, содержащем 2 мМ CaCl2. Центрифугирование проводилось в конических пробирках (25 мл) в бакет-роторе на центрифуге К23 при 600g (2000 об/мин) в течение 20 минут. Супернатант, представляющий цитоплазматичекую фракцию, отбирали в отдельную пробирку и осаждали изопропанолом. Осадок ядер промывали дважды 250 мкл лизирующего буфера (буфер А, содержащий 2 мМ CaCl2, 0.5% Тритона Х-100). После каждой промывки препарат центрифугировали 10' в бакет-роторе на центрифуге К23 при 600g (2000 об/мин). Ядра ресуспендировали в 500 мкл буфера А, содержащего 2 мМ CaCl2, анализировали цитологически и переносили в пробирки от ротора JA-21 центрифуги J2-21. К суспензии ядер добавляли до 2 М NaCl и до 1% SDS. Лизат ядер инкубировали при 65°С 15' до полного просветления реакционной смеси без какого либо встряхивания. Лизат ядер центрифугировали при 21000 об/мин (52000 g) в течение 20 минут при 35°С. Супернатант, представляющий ядерный сок или межхромосомный материал, отбирали желтым носиком досуха и переносили в другую пробирку. К супернатанту добавляли 1:10 объема NaAc и осаждали 0.6 объема изопропанола. Количество меченого материала определяли стандартным способом в эппендорфовских пробирках, опущенных в счетные флаконы на счетчике Rac Betta. Образцы фракционировали в 0.7% агарозном геле. После проведения электрофореза агарозный блок высушивали на подложке под струей горячего воздуха. Гель экспонировали с рентгеновской пленкой в течение ночи или в зависимости от количества меченого материала.
Для анализа фрагментации ДНК при апоптозе клетки MCF-7 инкубировали 24 ч при 37°С в присутствии 0,1 мкг/мл ФНО-α и 10 мкг/мл циклогексимида в среде RPMI 1640, содержавшей 10 мМ L-глутамина, 50 мкг/мл стрептомицина и 5% ЭБС. Для выделения ДНК к ресуспендированным в воде клеткам добавляли SDS до 1%, ЭДТА до 20 mM и PrK до 200 мкг/мл и инкубировали при 60°С в течение 1 ч. ДНК осаждали изопропанолом, растворяли в воде и вновь добавляли ЭДТА до 20 mM и PrK до 200 мкг/мл и инкубировали при 60°С в течение 1 ч. Образцы анализировали в 2% агарозном геле электрофорезом.
Рассмотрим лейкостимулирующее действие фрагментов экзогенной ДНК, защищенных протамином, при экспериментально вызванной ЦФ миелосупрессии мышей.
Остановимся на динамике деградации экзогенной ДНК, ассоциированной с протамином и без него при воздействии нуклеаз сыворотки крови.
Любая незащищенная ДНК, попавшая в кровь, практически мгновенно подвергается воздействию нуклеаз, что приводит к ее деградации до фрагментов размером, кратным 1-2 нуклеосомным мономерам и составляющим ~200 п.о. [Черепанова А.В., Тамкович С.Н., Власов В.В., Лактионов П.П. Активность дезоксирибонуклеаз крови в норме и при патологии. Биомедицинская химия. 2007. Т.53, №5, с.488-496]. Для достижения плейотропного [Николин В.П., Попова Н.А., Себелева Т.Е., Стрункин Д.Н., Рогачев В.А., Семенов Д.В., Богачев С.С., Якубов Л.А., Шурдов М.А. Влияние экзогенной ДНК на рост экспериментальных опухолей. Вопросы онкологии. 2006. Т.52, №1, с.66-69; Николин В.П., Попова Н.А., Себелева Т.Е., Стрункин Д.Н., Рогачев В.А., Семенов Д.В., Богачев С.С., Якубов Л.А., Шурдов М.А. Влияние экзогенной ДНК на восстановление лейкопоэза и противоопухолевый эффект циклофосфана. Вопросы онкологии. 2006. Т.52, №3, с.336-340; Likhacheva AS, Nikolin VP, Popova NA, Dubatolova TD, Strunkin DN, Rogachev VA, Sebeleva ТЕ, Erofeev IS, Bogachev SS, Yakubov LA, Shurdov MA. Integration of human DNA fragments into the cell genomes of certain tissues from adult mice treated with cytostatic cyclophosphamide in combination with human DNA. Gene Ther Mol Biol. 2007; 11: 185-202; Likhacheva AS, Nikolin VP, Popova NA, Rogachev VA, Prokhorovich MA, Sebeleva ТЕ, Bogachev SS, Shurdov MA. Exogenous DNA can be captured by stem cells and be involved in their rescue from death after lethal-dose γ-radiation. Gene Ther Mol Biol. 2007; 11: 305-314; Yakubov LA, Rogachev VA, Likhacheva AC, Bogachev SS, Sebeleva ТЕ, Shilov AG, Baiborodin SI, Petrova NA, Mechetina LV, Shurdov MA, Wickstrom E. Natural human gene correction by small extracellular genomic DNA fragments. Cell Cycle. 2007 Jul; 6 (18): 2293-2301] терапевтического эффекта, связанного с размером вводимой ДНК, участвующей затем в различных типах репаративной гомологичной рекомбинации, количество экзогенной ДНК, вводимой в организм, составляло суммарно от 0,3 до 6,25 мг на животное, что многократно превышало физиологическую норму.
Первоначально мы оценили нуклеазную активность сыворотки крови. В качестве субстрата использовалась геномная ДНК человека, выделенная из плацент здоровых рожениц, фрагментированная до размеров от 200 до >6000 п.н. Была исследована динамика деградации ДНК человека в сыворотке мышиной крови в концентрациях 1, 10, 50 и 97%. Смесь инкубировали 10 мин при 37°С.
Как видно из полученных результатов, уже через 10 мин инкубации в 1% сыворотке крови заметна деградация исходной ДНК. При всех остальных концентрациях сыворотки, находящейся в реакционной смеси, исходная ДНК деградирует до фрагментов размером ~200 п.о. (фиг.2).
Для предотвращения деградации в настоящее время активно разрабатываются различные методы доставки ДНК в клетку, например применение ДНК-связывающих белков в сочетании с лигандами, избирательно взаимодействующими с рецепторами на поверхности клеток-мишеней, опосредующими эндоцитоз. Так же для защиты ДНК от действия нуклеаз крови используются вирусные векторы, липосомы и др. [Анисимова В.А. Адресная доставка ДНК-вакцин. 8-ая Международная конференция "Вакцина против ВИЧ". 2000].
Мы предположили, что комплекс ДНК с протамином позволит защитить ДНК от действия нуклеаз крови и при этом не снизит лейкостимулирующей активности препарата экзогенной ДНК, охарактеризованной в нашей предыдущей работе [Николин В.П., Попова Н.А., Себелева Т.Е., Стрункин Д.Н., Рогачев В.А., Семенов Д.В., Богачев С.С., Якубов Л.А., Шурдов М.А. Влияние экзогенной ДНК на восстановление лейкопоэза и противоопухолевый эффект циклофосфана. Вопросы онкологии. 2006. Т.52, № 3, с.336-340].
Протамин - низкомолекулярный ядерный белок, молекулярная масса 4-12 кДа. Для протамина характерно высокое содержание щелочных аминокислот, особенно аргинина (70-80%), что обусловливает основные свойства протамина. Он хорошо растворим в воде, кислой и нейтральной среде, осаждается щелочами, не денатурирует при нагревании. У многих животных протамин наряду с гистонами содержится в сперматозоидах, а у некоторых (например, у рыб) полностью замещает гистоны. В ядрах клеток протамин, подобно гистонам, ассоциирован с дезоксирибонуклеиновыми кислотами в нуклеопротамины. Методом рентгеноструктурного анализа показано, что цепочка протамина обматывается как третья нить вокруг двойной спирали ДНК. Присутствие протамина защищает ДНК от действия нуклеаз и придает хроматину компактную форму. Протамин образуют соли с кислотами и комплексы с кислыми белками (малорастворимый комплекс протамина с инсулином используют в медицинской практике для продления срока действия последнего) [Brewer L, Corzett M, Lau EY, Balhorn R. Dynamics of protamine 1 binding to single DNA molecules. J Biol Chem. 2003 Oct 24; 278 (43):42403-42408; Vilfan ID, Conwell CC, Hud NV. Formation of native-like mammalian sperm cell chromatin with folded bull protamine. J Biol Chem. 2004 May 7; 279 (19): 20088-20095].
Для выяснения соотношения ДНК и протамина, при котором молекулы ДНК полностью защищаются от деградации, λ Hind III ДНК и ДНК человека, связанные с протамином в различных соотношениях, инкубировали в 10% сыворотке крови мыши.
Первоначально для формирования комплекса протамин/ДНК 2 мкг ДНК λ Hind III и 15 мкг ДНК человека инкубировали несколько минут с различным (от 10 до 150% по массе) количеством протамина. Далее комплексы протамин/ДНК, полученные для различных соотношений контрагентов, инкубировали 60 минут в 10% сыворотке крови мыши. Как следует из полученных данных, протамин в соотношении 80% к ДНК по массе практически абсолютно защищает ДНК от деградации (фиг.3).
Остановимся на стимуляции восстановления лейкопоэза экзогенной ДНК, ассоциированной с протамином, в соотношении протамин/ДНК 0.8/1.0.
В данной работе были проведены эксперименты, в которых для инъекций мышам использовали ДНК, связанную с протамином. Исследовалась стимуляция восстановления лейкопоэза после введения мышам алкилирующего цитостатика циклофосфана, угнетающего кроветворение.
Циклофосфамид (ЦФ) - один из наиболее широко применяемых препаратов в онкологической практике. Противоопухолевый эффект ЦФ в настоящее время связывают с его генотоксическим алкилирующим производным фосфорамид мустардом (ФМ), который спонтанно образуется из альдофосфамида, производного 4-гидрокси-ЦФ. ФМ образует аддукты с пуриновыми основаниями молекулы ДНК, в особенности соседствующими с остатками гуанина, и приводит к возникновению перекрестных сшивок в двойной цепи молекулы ДНК [Yu LJ, Drewes P, Gustafsson К, Brain EG, Hecht JE, Waxman DJ. In vivo modulation of alternative pathways of P-450-catalyzed cyclophosphamide metabolism: impact on pharmacokinetics and antitumor activity. J Pharmacol Exp Ther. 1999 Mar; 288 (3): 928-937; De Silva IU, McHugh PJ, Clingen PH, Hartley JA. Defining the roles of nucleotide excision repair and recombination in the repair of DNA interstrand cross-links in mammalian cells. Mol Cell Biol. 2000 Nov; 20 (21): 7980-7990; Karle P, Renner M, Salmons B, Günzburg WH. Necrotic, rather than apoptotic, cell death caused by cytochrome P450-activated ifosfamide. Cancer Gene Ther. 2001 Mar; 8 (3): 220-230].
Циклофосфан индуцирует образование межцепочечных сшивок в активно делящихся клетках, в результате чего гибнут не только опухолевые клетки, но и, например, клетки крови. В результате на организменном уровне наблюдается лейкопения, степень выраженности которой зависит от дозы циклофосфана.
Мышам-самцам СВА в/б был введен циклофосфан по 200 мг/кг веса. Для подсчета количества лейкоцитов за 30 мин до этого у всех мышей брали кровь из кончика хвоста по 10 мкл. Аналогично забор крови проводили на 4, 7 и 11 сутки после введения ЦФ. Одна группа мышей служила контролем (физиологический раствор), а двум другим следующие 3 дня после введения ЦФ внутрибрюшинно вводили по 50 мкг ДНК человека - одной группе, другой - ДНК человека в комплексе с протамином.
Как демонстрируют результаты эксперимента (фиг.4), на 4 сутки, несмотря на полное подавление активности белого ростка крови, лейкоцитов сохраняется в 2 раза больше, чем в контрольной группе.
На 7 сутки после введения ЦФ количество лейкоцитов у мышей, которым водили ДНК человека или ДНК человека, связанную с протамином, достоверно выше, чем в контрольной группе, и даже превышает исходный уровень.
На 11 сутки наблюдается незначительное естественное снижение числа лейкоцитов, однако, по-прежнему восстановление лейкопоэза в группах мышей, получавших дополнительно инъекции экзогенной ДНК, более эффективное, чем в контроле.
Такое свойство экзогенной ДНК стимулировать лейкопоэз связано с несколькими механизмами ее воздействия на клетку-предшественницу лейкоцитов.
1. Экзогенные фрагменты ДНК доставляются в межхромосомное пространство активно пролиферирующих клеток, включая всех предшественников лейкопоэза [Гистология, цитология и эмбриология. Под ред. Афанасьева Ю.И., Кузнецова С.Л., Юриной Н.А. М.: Медицина, 2004. 768 с], и репарация межцепочечных сшивок (МЦС), возникших при воздействии ЦФ, происходит с их участием. На завершающем этапе репарации МЦС требуется гомологичная рекомбинация, для которой донорными последовательностями в обычных условиях служат гомологичные участки сестринской хроматиды или гомологичной хромосомы. При множественных МЦС процесс репарации может быть затруднен стерическими проблемами, возникающими при поиске гомологичных последовательностей и спаривании цепей. Расположенные в непосредственной близости с репарируемыми МЦС экзогенные фрагменты, имеющие гомологию с участками репарации, могут быть использованы в качестве донорных последовательностей и могут принимать участие в гомологичной рекомбинации, необходимой для завершения репарационного процесса. При таком участии экзогенной ДНК экстрахромосомальной локализации обслуживается большее количество репарируемых повреждений, что безусловно должно отразится на жизнеспособности прогениторных клеток. Факт интеграции экзогенной ДНК в ходе репарации МЦС был описан в работе [Likhacheva AS, Nikolin VP, Popova NA, Dubatolova TD, Strunkin DN, Rogachev VA, Sebeleva ТЕ, Erofeev IS, Bogachev SS, Yakubov LA, Shurdov MA. Integration of human DNA fragments into the cell genomes of certain tissues from adult mice treated with cytostatic cyclophosphamide in combination with human DNA. Gene Ther Mol Biol. 2007; 11: 185-202].
2. Следующим фактором, влияющим на выживаемость созревающих клеток-предшественников лейкоцитов после воздействия ЦФ и обработки экзогенной ДНК, является описанный в литературе феномен «вскрытия генома», свойственный для созревающих прогениторных клеток-предшественников лейкоцитов или СК, вступивших на путь терминальной дифференцировки. Это явление связано с возникновением многочисленных функциональных одноцепочечных разрывов хроматина, что предполагает вовлечение генетического материала в реорганизацию, которая является обязательной и неотъемлемой частью дифференцировки и изменения паттерна экспрессии генов [Farzaneh F, Zalin R, Brill D, Shall S. DNA strand breaks and ADP-ribosyl transferase activation during cell differentiation. Nature. 1982 Nov 25; 300 (5890): 362-366; Johnstone АР, Williams GT. Role of DNA breaks and ADP-ribosyl transferase activity in eukaryotic differentiation demonstrated in human lymphocytes. Nature. 1982 Nov 25; 300 (5890): 368-370; Vatolin SY, Okhapkina EV, Matveeva NM, Shilov AG, Baiborodin SI, Philimonenko VV, Zhdanova NS, Serov OL. Scheduled perturbation in DNA during in vitro differentiation of mouse embryo-derived cells. Mol Reprod Dev. 1997 May; 47 (1): 1-10]. Всплеск никирования ДНК совпадает с первым-третьим митозом дифференцирующихся клеток и определяет резкое увеличение (в 5-6 раз) частоты сестринских хроматидных обменов. Предполагаемый механизм для такого рода обменов в геноме - это гомологичная рекомбинация [Vatolin SY, Okhapkina EV, Matveeva NM, Shilov AG, Baiborodin SI, Philimonenko VV, Zhdanova NS, Serov OL. Scheduled perturbation in DNA during in vitro differentiation of mouse embryo-derived cells. Mol Reprod Dev. 1997 May; 47 (1): 1-10]. По-видимому, каждая стадия созревания лейкоцитов характеризуется реорганизацией хроматина и активацией нового ансамбля экспрессирующихся генов. Предположительно механизм, определяющий повышенную выживаемость прогениторных клеток после обработки ЦФ и экзогенной ДНК, состоит в следующем. Клетка-предшественник лейкоцитов после прошедшей репарации МЦС в присутствии экзогенной ДНК экстрахромосомальной локализации или без нее сохраняет определенное количество дефектов хроматина, возникших в результате аберраций при формировании и реализации интермедиатов. Покоящаяся, спасенная клетка с оставшимися дефектами хроматина после репарации МЦС приступает к делению. Если при этом в клетке не начинается реорганизация хроматина, связанная с переходом клетки в следующую стадию созревания, то она неминуемо гибнет вследствие дефектного митоза. Если деление сопровождается одновременным переходом клетки в следующую стадию созревания, характеризующуюся реорганизацией хроматина и появлением множественных одноцепочечных разрывов, то присутствие в ядре фрагментов экзогенной ДНК позволяет клетке исправить оставшиеся дефекты и полностью восстановить свою жизнеспособность. По-видимому, существует некая согласованность между системами контроля прогрессии клеточного цикла и рекомбинационно-репарационными системами клетки. Предполагается, что контроль checkpoint систем на время появления функциональных разрывов генома прекращен или каким-то образом изменен. Это позволяет фрагментам экзогенной ДНК остаться незамеченными системами контроля и быть вовлеченными в репаративно-рекомбинационные процессы, связанные с восстановлением целостности хроматина после произошедшей реорганизации генома в качестве донорных гомологичных последовательностей.
3. Присутствие экзогенной ДНК экстрахромосомальной локализации в межхромосомном пространстве ядра стимулирует непрерывную пролиферацию прогениторных клеток-предшественников лейкоцитов на всех стадиях созревания. Предполагаемый механизм такой стимуляции заключается в следующем. В результате попадания в покоящуюся клетку экзогенных фрагментов ДНК происходит активация систем checkpoint, контролирующих прогрессию клеточного цикла [MacDougall CA, Byun TS, Van С, Yee MC, Cimprich KA. The structural determinants of checkpoint activation. Genes Dev. 2007 Apr 15; 21 (8): 898-903; Yoo HY, Jeong SY, Dunphy WG. Site-specific phosphorylation of a checkpoint mediator protein controls its responses to different DNA structures. Genes Dev. 2006 Apr 1; 20 (7): 772-783; Yoo HY, Shevchenko A, Shevchenko A, Dunphy WG. Mcm2 is a direct substrate of ATM and ATR during DNA damage and DNA replication checkpoint responses. J Biol Chem. 2004 Dec 17; 279 (51): 53353-53364; Zou L. Single - and double-stranded DNA: building a trigger of ATR-mediated DNA damage response. Genes Dev. 2007 Apr 15; 21 (8): 879-885]. При этом включаются определенные программы, позиционирующие клетку как арестованную на соответствующей стадии клеточного цикла. После активации таких программ клетка уже не в состоянии блокировать каскад событий, связанных с такой активацией, и покоящаяся до этого клетка начинает делиться. При этом, если фрагменты экзогенной ДНК каждый раз попадают во внутренние компартменты клетки-предшественницы, то она будет без отдыха делиться до того времени, пока стрессовый фактор не будет удален из ее окружения.
4. И последнее. Фрагменты экзогенной экстраклеточной ДНК действуют на все стадии созревания предшественников терминально дифференцированных клеток крови во всех зонах костного мозга и тимуса, в 3 из которых происходит формирование Т-лимфоцитов, а в 4 идет созревание В-лимфоцитов. На каждой из стадий происходит определенное количество делений клеток соответствующего клона, приводящее к увеличению популяции этих клеток. После воздействия цитостатика клеткам всех стадий зрелости требуется время для завершения репаративных процессов и продолжения созревания до выхода терминально дифференцированных клеток в кровь. В том случае, когда репарация межцепочечных сшивок проходит без участия экзогенной ДНК, основная масса предшественников на всех стадиях созревания погибает. И восстановление количества форменных элементов крови начинается с выжившей СКК. При этом требуется время и силы организма для прохождения всех стадий созревания и всех групп предшественников форменных элементов крови. Воздействие экзогенной ДНК спасает популяции клеток на всех стадиях зрелости. Мы полагаем, что именно этим фактом объясняется сильный и быстро развивающийся лейкостимулирующий эффект при воздействии кросслинкирующего цитостатика и терапии экзогенной ДНК.
В проделанной работе показано, что после совместного введения мышам алкилирующего цитостатика циклофосфана и препарата фрагментированной экзогенной ДНК, связанной с протамином - белком, который защищает ДНК от действия нуклеаз крови, наблюдается эффективное восстановление лейкопоэза.
Полученные данные свидетельствуют о том, что применение комплексов ДНК с протамином позволяет защитить ДНК от действия нуклеаз крови без изменения терапевтической активности препарата ДНК.
Связывание ДНК с протамином позволяет уменьшить количества вводимой ДНК и сохранить исходный размер фрагментов, что способствует вовлечению более продолжительных участков хроматина в репаративную гомологичную рекомбинацию, в конечном счете, увеличивая шанс клетки избавиться от возникших многочисленных повреждений.
Рассмотрим действие экзогенной ДНК, ассоциированной с протамином, на экспериментальные опухоли мыши.
Остановимся на анализе некоторые количественных и качественных характеристиках экзогенной ДНК (незащищенной протамином и ассоциированной с ним), доставляемой в экспериментальные опухоли.
Для оценки количества поглощенной опухолью ДНК и морфологии ДНК, доставленной в межхромосомное пространство клеток солидной и асцитной формы опухоли RLS, были проведены инъекции либо ДНК фага лябда, гидролизованного Hind III, либо ДНК человека, фрагментированной до размеров 200-6000 п.о., мышам с перевитыми экспериментальными опухолями. Как следует из полученных данных, в опухоль доставляется незначительный процент от инъецированной ДНК. При пересчете на п.о. приблизительно 1 фрагмент размером около 1.3 т.п.о. может присутствовать в одной клетке опухоли при инъекции до 5 мкг экзогенной ДНК (фиг.5, таблицы 1-3).
Морфология меченой ДНК, выделяемой из плазмы крови мышей, свидетельствует о том, что в кровяном русле происходит практически мгновенная деградация исходной ДНК (фиг.6b, 12). В плазме крови мышей опухоленосителей ДНК также подвергается деградации. В деградированной форме экзогенная ДНК доставляется во внутренние компартменты раковой клетки (Фиг.6а).
После ассоциации с протамином инъецированная ДНК защищается от действия нуклеаз крови и через 20 мин в крови все еще можно обнаружить высокомолекулярные фрагменты меченой ДНК (фиг.6b, 15).
Остановимся на действии фрагментов экзогенной ДНК, ассоциированной с протамином, на рост экспериментальной опухоли RLS.
Как следует из данных, полученных в первой части исследования, в опухоль доставляется крайне незначительное количество ДНК. Это позволяет считать, что эффект ингибирования роста опухоли, показанный в наших ранних работах [Yakubov LA, Popova NA, Nikolin VP, Semenov DV, Bogachev SS, Os'kina IN. Extracellular genomic DNA protects mice against radiation and chemical mutagens. Genome Biol. 2003; 5: 3; Николин В.П., Попова Н.А., Себелева Т.Е., Стрункин Д.Н., Рогачев В.А., Семенов Д.В., Богачев С.С., Якубов Л.А., Шурдов М.А. Влияние экзогенной ДНК на рост экспериментальных опухолей. Вопросы онкологии. 2006. Т.52, № 1, с.66-69] и в настоящем исследовании, не связан с прямым воздействием фрагментов экзогенной ДНК на геном, как это описано для раковых клеток MCF-7, растущих в культуре [Yakubov LA, Rogachev VA, Likhacheva AC, Bogachev SS, Sebeleva ТЕ, Shilov AG, Baiborodin SI, Petrova NA, Mechetina LV, Shurdov MA, Wickstrom E. Natural human gene correction by small extracellular genomic DNA fragments. Cell Cycle. 2007 Jul; 6 (18): 2293-2301]. Более вероятно воздействие фрагментов экзогенной ДНК через определенные звенья иммунной системы.
Известно, что фрагменты экзогенной ДНК или олигонуклеотиды, обогащенные GC мотивами активируют созревание ДК и индуцируют иммунный ответ по Т-клеточному типу. По-видимому, даже очень короткие фрагменты ДНК, обладающие определенной структурой, способны воздействовать на антигенпрезентирующие дендритные клетки и активировать Т-клеточный иммунный ответ [Krieg AM. CpG motifs: the active ingredient in bacterial extracts? Nat Med. 2003 Jul; 9 (7): 831-835; Olishevsky S, Kozak V, Yanish Y. Immunostimulatory CpG DNA in cancer vaccinotherapy. Exp Oncol. 2003; 25: 85-92]. Мы полагаем, что именно по такому пути идет ингибирование роста опухоли, наблюдаемое при инъекции мышам с перевитыми опухолями, фрагментированной экзогенной ДНК. В случае прямого действия на геном с использованием гомологичной рекомбинации как механизма замещения онколокусов на здоровые аллели предполагается, что только длинные фрагменты (не менее 1 т.п.о.) могут осуществлять такого рода рекомбинацию (двойной реципрокный обмен концевых гомологии) [Orr-Weaver TL, Szostak JW, Rothstein RJ. Yeast transformation: a model system for the study of recombination. Proc Natl Acad Sci USA. 1981 Oct; 78 (10): 6354-6358; Kucherlapati RS, Eves EM, Song KY, Morse BS, Smithies O. Homologous recombination between plasmids in mammalian cells can be enhanced by treatment of input DNA. Proc Natl Acad Sci USA. 1984 May; 81 (10): 3153-3157; Deng С, Capecchi MR. Reexamination of gene targeting frequency as a function of the extent of homology between the targeting vector and the target locus. Mol Cell Biol. 1992 Aug; 12 (8): 3365-3371; Thomas KR, Deng C, Capecchi MR. High-fidelity gene targeting in embryonic stem cells by using sequence replacement vectors. Mol Cell Biol. 1992 Jul; 12 (7): 2919-2923; Hastings PJ, McGill C, Shafer B, Strathern JN. Ends-in vs. ends-out recombination in yeast. Genetics. 1993 Dec; 135 (4): 973-980; Langston LD, Symington LS. Gene targeting in yeast is initiated by two independent strand invasions. Proc Natl Acad Sci USA. 2004 Oct 26; 101 (43): 15392-15397]. По-видимому, в этом случае существует прямая зависимость, выражающаяся в том, что чем длиннее фрагмент, участвующий в ГР, то тем больше вероятность замещения онколокуса на не мутантный аллель и изменения ракового статуса клетки. Если предположить, что оба указанных механизма используются одновременно для подавления роста раковых клеток, то длина доставляемых фрагментов является важным фактором для такого ингибирования.
В наших работах для терапии экспериментальных раков мы использовали большое количество однократно вводимой ДНК [Yakubov LA, Popova NA, Nikolin VP, Semenov DV, Bogachev SS, Os'kina IN. Extracellular genomic DNA protects mice against radiation and chemical mutagens. Genome Biol. 2003; 5: 3; Николин В.П., Попова Н.А., Себелева Т.Е., Стрункин Д.Н., Рогачев В.А., Семенов Д.В., Богачев С.С., Якубов Л.А., Шурдов М.А. Влияние экзогенной ДНК на восстановление лейкопоэза и противоопухолевый эффект циклофосфана. Вопросы онкологии. 2006. Т.52, № 3, с.336-340]. Это было связано с быстрой ее деградацией в кровяном русле и, как следствие, невозможностью попадания в опухоль протяженных фрагментов ДНК, которые, как предполагается, наряду с индукцией иммунного ответа, могут замещать неотрансформированные гомологичные участки генома и изменять генетику раковой клетки. Ассоциация фрагментов ДНК с протамином защищает фрагменты от нуклеазной деградации. При этом высокомолекулярные фрагменты достигают межхромосомного пространства ядер раковых клеток и могут быть использованы клеткой в качестве субстрата для гомологичной рекомбинации.
Тем не менее мы установили, что при количестве вводимой ДНК, равном суммарно 120 мкг на мышь, в клетки опухоли будет доставлено не более нескольких фрагментов размером 1 т.п.о. (см. предыдущий раздел). Этого количества, скорее всего, недостаточно для того, что бы повлиять на генетику раковой клетки. Мы полагаем, что действие на опухоль в данном случае происходило через активацию определенных звеньев иммунной системы. Важным оказался тот факт, что ассоциированная с протамином и сохранившая в связи с этим свой размер экзогенная ДНК ингибирует рост опухоли в той же степени, что и незащищенная протамином ДНК (фиг.7).
То есть основной механизм ингибирования роста опухоли при ассоциации терапевтической ДНК с протамином сохраняется. При этом появляется возможность использования второго механизма воздействия на опухоль, когда протяженный фрагмент ДНК, защищенный протамином, попадая внутрь раковой клетки, вступает в гомологичный обмен с геномной ДНК, что при определенных условиях может повлиять на генетику раковой клетки.
Рассмотрим интернализацию в клетках MCF-7 экзогенных фрагментов ДНК, защищенными протамином и индукцию ими апоптоза.
Остановимся на сравнительных морфологических особенностях фрагментов экзогенной ДНК, интернализованных в основных клеточных компартментах клеток MCF-7 из культуральной среды в незащищенном виде и ассоциированных с протамином.
На фиг.8 представлены результаты электрофореза ДНК экстрахромосомальной фракции, выделенной из ядер клеток MCF-7.
Как следует из представленной электрофореграммы образцов ДНК, при ассоциации добавляемой в среду экзогенной ДНК с протамином, в межхромосомном пространстве ядер клеток обнаруживается шлейф меченой ДНК, простирающийся от низкомолекулярной фракции до высокомолекулярной, что может свидетельствовать о доставке в ядерное пространство всего спектра (200-6000 п.о.) фрагментов из находящихся в культуральной среде. Высокомолекулярный фрагмент в части эксперимента с незащищенной ДНК, выявляемый в межхромосомной фракции, представляет собой результат лигирования коротких деградированных фрагментов, доставленных в ядерное пространство [Rogachev VA, Likhacheva A, Vratskikh О, Mechetina LV, Sebeleva ТЕ, Bogachev SS, Yakubov LA, Shurdov MA. Qualitative and quantitative characteristics of the extracellular DNA delivered to the nucleus of a living cell. Cancer Cell Int. 2006 Oct 11; 6:23]. Также на электрофореграмме образцов ДНК, выделенных из культуральной среды после различных времен инкубации, при ее ассоциации с протамином в среде присутствует высокомолекулярная форма ДНК (данные не приводятся). При этом в образцах с ДНК, добавленной в среду в форме, неассоциированной с протамином, она полостью деградирует [Rogachev VA, Likhacheva A, Vratskikh О, Mechetina LV, Sebeleva ТЕ, Bogachev SS, Yakubov LA, Shurdov MA. Qualitative and quantitative characteristics of the extracellular DNA delivered to the nucleus of a living cell. Cancer Cell Int. 2006 Oct 11; 6:23].
Динамика доставки фрагментов экзогенной ДНК во внутренние компартменты клетки представлена в таблице 4 и на фиг.9.
Можно отметить несколько особенностей интернализации экзогенной фрагментированной ДНК, защищенной протамином.
1. Прослеживается два пика интернализации фрагментов экзогенной ДНК в цитоплазме в точках 30' и 120'.
2. В межхромосомное пространство фракции эти пики смещены к точкам 60' и 180'.
3. В межхромосомное пространство доставляется практически в 10 раз большее количество экзогенной ДНК, защищенной протамином, по сравнению с незащищенной ДНК.
4. Динамика появления меченого материала во фракции хроматина значительно отличается в случае ДНК, ассоциированной с протамином, и без него.
Во фракции хроматина при использовании незащищенной экзогенной ДНК меченый материал обнаруживается в значительном количестве уже в нулевой точке. В точках 30-120' количество меченого материала незначительно возрастает и выходит на плато. В точке 180' количество меченого материала во фракции хроматина резко падает. В случае защищенной протамином ДНК процесс имеет явный накопительный характер. С практически отсутствия в нулевой точке меченый материал накапливается и в точке 120' достигает своего максимального значения, совпадающего с образцом ДНК без протамина. Затем количество меченого материала резко падает до тех же самых значений, что и в образце ДНК, используемой без ассоциации с протамином.
Обнаруженную разницу в количестве доставляемого материала ДНК в межхромосомную фракцию можно объяснить тем, что рецепторы, связывающие ДНК на поверхности клетки, обладают большей специфичностью к ДНК в форме хроматина, чем к незащищенной ДНК. Два пика в количестве меченого материала, выявляемых в цитоплазме 30' и 120' и межхромосомной фракции 60' и 180', могут свидетельствовать о том, что количество рецепторов конечное, что они вместе с ДНК интернализируются в клеточных компартментах, что оборот рецепторов происходит в течение ориентировочно одного часа и что проникновение меченых фрагментов из цитоплазмы во внутреннее пространство ядра занимает порядка 30 минут. Различия в динамике появления меченого материала во фракции хроматина можно объяснить тем, что при использовании незащищенной ДНК проникшие в ядерное пространство меченые фрагменты быстро подвергаются гидролизу и меченые предшественники участвуют в процессе репликации. В случае использования ДНК в защищенной протамином форме, несмотря на то, что в межхромосомном пространстве скапливается большое количество фрагментов экзогенной ДНК, сохраняющееся на протяжении всего анализируемого времени 0-180', в хроматине обнаруживается последовательно увеличивающееся количество меченого материала. Можно полагать, что в такой динамике могут быть задействованы два процесса, которые могут действовать по отдельности или накладываться один на другой. Один из них, аналогичный описанному для незащищенной ДНК и связанный с деградацией меченых фрагментов и использованием меченых предшественников в синтетических процессах, проходящих в клетке. Также можно предполагать, что фрагменты экзогенной ДНК, защищенные протамином и депонированные в межхромосомном пространстве, постепенно рекомбинируют с гомологичными участками хромом, что и приводит к постепенному накоплению метки в хроматине.
Остановимся на анализе индукции апоптоза фрагментами экзогенной ДНК, незащищенной протамином и в комплексе с ним.
Мы провели эксперименты по индукции апоптоза клеток MCF-7, обработанных ДНК, защищенной протамином в концентрации 25 мкг/мл и незащищенной ДНК в концентрации 100 мкг/мл. Как следует из фиг.10, ДНК, защищенная протамином, в такой же степени индуцирует апоптоз, как и незащищенная ДНК.
Этот факт может свидетельствовать о том, что защищенная ДНК также способна вступать в репаративную гомологичную рекомбинацию, исправляя мутантный аллель, при этом ее эффективное количество в культуральной среде может быть существенно ниже.
Кроме проведенного анализа индукции апоптоза фрагментами ДНК, защищенной протамином, в сравнении с незащищенной ДНК, мы провели эксперименты, в которых концентрация защищенной ДНК в культуральной среде была уменьшена до таковой, определяемой для плазмы крови высших эукариот 30-100 нг/мл [Anker P, Mulcahy H, Chen XQ, Stroun M. Detection of circulating tumour DNA in the blood (plasma/serum) of cancer patients. Cancer Metastasis Rev. 1999; 18 (1):65-73].
В культуру клеток MCF-7 добавлялась незащищенная ДНК и ДНК, связанная с протамином, в концентрациях 25, 2.5, 0.25 и 0.025 мкг/мл среды. Количество клеток для всех экспериментальных точек было одинаковым (107). Через 17 дней постоянной инкубации клеток с ДНК культуры клеток индуцировали к апоптозу, выделяли ДНК и смотрели появление нуклеосомной фрагментации хроматина (фиг.11).
Было обнаружено, что концентрация ДНК, связанной с протамином, 25 нг/мл среды уже способна приводить к появлению нуклеосомной фрагментации хроматина.
Протамин, защищая ДНК от действия нуклеаз, способствует тому, что большее количество ДНК попадает в клетку в недеградированном виде и соответственно оказывается способным участвовать в рекомбинационном процессе. Отсюда можно предположить, что большее количество клеток восстанавливает ген каспазы 3, и при индукции апоптоза мы можем наблюдать более ярко выраженную нуклеосомную фрагментацию хроматина.
Таким образом, связывание с протамином обеспечивает молекулам ДНК устойчивость к действию нуклеаз, находящихся в культуральной среде. В результате этого фрагменты экзогенной ДНК способны достигать основных клеточных компартментов в исходной форме. При культивировании клеток MCF-7 совместно с ДНК, ассоциированной с протамином, наблюдается восстановление гена каспазы 3, что фенотипически проявляется в появлении нуклеосомной деградации хроматина при индукции апоптоза. Протамин, защищая ДНК от действия нуклеаз, обеспечивает попадание молекул экзогенной ДНК в клетку в недеградированном виде, что делает возможным участие их в рекомбинационном процессе. При связывании ДНК с протамином появляется возможность снизить концентрацию ДНК, не повлияв на эффективность действия экзогенной ДНК.
Таким образом, благодаря усовершенстованию известного способа достигается требуемый результат, заключающийся в повышении эффективности лечения пациентов, проходящих программную химиотерапию, и пациентов с онкологическими заболеваниями. Результаты проделанного исследования позволяют сказать, что защита фрагментированной экзогенной ДНК гистоноподобным белком протамином не влияет на способность фрагментированной экзогенной ДНК проявлять лейкостимулирующее действие, ингибировать рост экспериментальной опухоли мыши и принимать участие в репарации мутантного аллеля гена каспазы 3 клеток аденокарциномы молочной железы человека MCF-7. Ассоциация фрагментированной экзогенной ДНК с протамином позволяет сохранить исходный размер терапевтических фрагментов и существенно уменьшить эффективную концентрацию экзогенной ДНК в околоклеточных жидкостях.
Реферат
Изобретение относится к медицине, а именно к онкологии, и может быть использовано при лечении опухолей. Способ включает введение пациенту фрагментированной ДНК человека в комплексе с негистоновым белком протамином, при этом длина фрагментов ДНК человека составляет 200-6000 пар оснований и соотношение фрагментированная ДНК человека/негистоновый белок протамин составляет 1/0,8. Комплекс вводят в такой дозе, чтобы концентрация фрагментированной ДНК человека в крови составляла 25-1000 нг/мл. Использование изобретения позволяет лечить опухоли за счет замещения онколокуса фрагментом ДНК на немутантный аллель с формированием нуклеотически стабильного комплекса. 1 з.п. ф-лы, 4 табл., 11 ил.
Комментарии