Рекомбинантный белок уриказы млекопитающего для конъюгирования с пэг, молекула нуклеиновой кислоты,кодирующая его, вектор для его экспрессии, способего получения и применение - RU2290439C2
Код документа: RU2290439C2
Чертежи













Описание
Данная заявка заявляет эффект предварительной заявки Соединенных Штатов с номером 60/095 489, поданной 6 августа 1998 года, все содержание которой включено здесь в качестве ссылки.
Описанное здесь изобретение было выполнено с поддержкой Правительства Соединенных Штатов по гранту №DK48529, присужденному Национальным институтом здравоохранения. Правительство имеет определенные права в отношении данного изобретения.
Данное изобретение относится, в общем, к белкам уратоксидазы (уриказы) и кодирующим их молекулам "нуклеиновых кислот. Конкретно, данное изобретение относится к уриказным белкам, которые, в частности, применимы, например, в качестве промежуточных продуктов для получения улучшенных модифицированных уриказных белков с пониженной иммуногенностыо и увеличенной биодоступностью. Предпочтительные модифицированные уриказные белки данного изобретения включают в себя уриказные белки, ковалентно связанные с поли(этиленгликолями) или поли(этиленоксидами). Таким образом, данное изобретение представляет уриказные белки, антитела, которые специфически связываются с этими белками, молекулы нуклеиновых кислот, кодирующие уриказные белки и их полезные фрагменты, векторы, содержащие эти молекулы нуклеиновых кислот, клетки-хозяева, содержащие эти векторы, и способы применения и получения этих уриказных белков и молекул нуклеиновых кислот.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Подагра является наиболее обычным воспалительным заболеванием суставов у людей в возрасте более 40 лет (Roubenoff 1990). Болезненный подагрический артрит появляется, когда повышенный уровень мочевой кислоты в крови (гиперурицемия или гиперурикемия) приводит к эпизодическому образованию микроскопических кристаллов моногидрата урата мононатрия в суставах. С течением времени, хроническая гиперурицемия может также приводить к деструктивным кристаллическим отложениям урата (подагрическим узлам) вокруг суставов, в мягких тканях и в некоторых органах (Hershfield 1996). Мочевая кислота имеет ограниченную растворимость в моче и при повышенной экскреции (гиперурикозурии) может вызывать образование почечных камней (уриколитиаз). У пациентов с некоторыми злокачественными заболеваниями, в частности с лейкозом и лимфомой, заметная гиперурицемия и гиперурикозурия (обусловленные усиленным оборотом и лизисом опухолевых клеток во время химиотерапии) создают серьезный риск острой, обструктивной почечной недостаточности (Sandberg et al. 1956; Gold and Fritz 1957; Cohen et al. 1980; Jones et al. 1990). Серьезные гиперурицемия и подагра связаны с дисфункцией почек вследствие различных причин, в том числе вследствие терапии с использованием циклоспорина для предупреждения отторжения органа при аллотрансплантации (West et al. 1987; Venkataseshan et al. 1990; Ann et al. 1992; Delaney et al. 1992; George and Mandell 1995).
Гиперурицемия может быть результатом как повышенного образования урата, так и пониженной секреции урата (Hershfield and Seegmiller 1976; Kelley et al. 1989; Becker and Roessler 1995). Умеренная гиперурицемия может контролироваться диетой, но сильно выраженная и связанная с серьезными клиническими последствиями гиперурицемия требует лечения лекарственными средствами, либо агентом, способствующим выведению мочевой кислоты, который усиливает экскрецию мочевой кислоты (неэффективным при снижении функции почек), либо ингибитором ксантиноксидазы аллопуринолом, который блокирует образование уратов. Аллопуринол является главной средством терапии в случае пациентов с подагрическими узлами, почечной недостаточностью, лейкозом и некоторыми наследственными нарушениями. Лечение гиперурицемии является обычно эффективным и хорошо переносимым. Однако некоторые пациенты с обезображивающей, лишающей трудоспособности подагрой с отложением солей трудно поддаются всем общепринятым способам терапии (Becker 1988; Fam 1990; Rosenthal and Ryan 1995). Кроме того, у ˜2% пациентов, принимающих аллопуринол, развиваются аллергические реакции и тяжелый синдром гиперсенсибилизации имеет место в ˜0,4% (Singer and Wallace 1986; Arellano and Sacristan 1993). Этот часто опасный для жизни синдром может вызвать острую почечную и печеночную недостаточность и серьезное кожное повреждение (токсический эпидермальный некролиз, эксфолиативный дерматит, мультиформную (экссудативную) эритему, синдром Стивенса-Джонсона). Аллопуринол нарушает также метаболизм азатиоприна и 6-меркаптопурина, лекарственных средств, применяемых в лечении лейкоза и для предотвращения отторжения органа-аллотрансплантата, состояний, при которых имеет место заметная гиперурицемия, и может вызывать тяжелую подагру или создавать угрозу для почечной функции.
В конечном счете, гиперурицемия является результатом мутационной инактивации гена уратоксидазы (уриказы) человека во время эволюции (Wu et al. 1989; Wu et al. 1992). Активная уриказа в пероксисомах печени большинства приматов, не являющихся человеком, и других млекопитающих превращает урат в аллантоин (+СО2 и H2O2), который является в 80-100 раз более растворимым, чем мочевая кислота, и более эффективно выводится почками. Парентерально вводимую уриказу, полученную из Aspergillus flavus (Uricozyme®, Clin-Midy, Paris) использовали для лечения тяжелой гиперурицемии, связанной с химиотерапией лейкоза, на протяжении 20 лет во Франции и Италии (London and Hudson 1957; Kissel et al. 1968; Brogard et al. 1972; Kissel et al. 1972; Potaux et al. 1975; Zittoun et al. 1976; Brogard et al. 1978; Masera et al. 1982), и использовали в недавних клинических испытаниях на пациентах с лейкозом в Соединенных Штатах (Pui et al. 1997). Уриказа имеет более быстрое наступление действия, чем аллопуринол (Masera et al. 1982; Pui et al. 1997). У пациентов с подагрой инфузии уриказы могут прерывать острые приступы и уменьшать размер подагрических узлов (Kissel et al. 1968; Potaux et al. 1975; Brogard et al. 1978).
Хотя и эффективная для лечения острой гиперурицемии во время кратковременного курса химиотерапии, ежедневная инфузия уриказы A. flavus была бы серьезным недостатком для лечения рецидивирующей подагры или подагры с отложениями солей. Кроме того, эффективность уриказы A. flavus быстро снижается в пациентах, которые образуют антитела против уриказы (Kissel et al. 1968; Brogard et al. 1978; Escudier et al. 1984; Mourad et al. 1984; Sibony et al. 1984). Имели место серьезные аллергические реакции, в том числе анафилаксия (Donadio et al. 1981; Montagnac and Schillinger 1990; Pui et al. 1997). Для более продолжительной терапии явно необходим более длительно действующий, менее иммуногенный препарат уриказы.
Один подход для секвестрации (изоляции) экзогенных ферментов от протеаз и иммунной системы включает в себя ковалентное присоединение инертного, нетоксичного полимера, монометоксиполиэтиленгликоля (ПЭГ) к поверхности белков (Harris and Zaiipsky 1997). Сначала было показано, что ПЭГ с молекулярным весом (Mr) ˜1000 - >10000 пролонгирует время циркулирования в кровотоке и снижает иммуногенность некоторых чужеродных белков в животных (Abuchowski et al. 1877а; Abuchowski et al. 1977b; Davis et al. 1981a; Abuchowski et al. 1984; Davis et al. 1991). В 1990 году бычья аденозин-деаминаза (ADA), модифицированная ПЭГ с Mr 5000 (PEG-ADA, ADAGEN®, производимая Enzon, Inc.), стала первым ПЭГилированным (ПЭГ-модифицированным) белком, одобренным Управлением по контролю за продуктами и лекарствами Соединенных Штатов (FDA), для лечения тяжелого заболевания комбинированного иммуннодефицита, вызываемого недостаточностью ADA (Hershfield et al. 1987). Опыт последних 12 лет показал, что антитела против ADA могут быть обнаружены при помощи чувствительного анализа ELISA в большинстве пациентов во время продолжительной терапии с использованием PEG-ADA, но не было обнаружено аллергических реакций или реакций гиперсенсибилизации; ускоренный клиренс PEG-ADA имел место в небольшом числе образующих антитела против ADA пациентов, но обычно это было временным эффектом (Chaffee et al. 1992; Hershfield 1997). Должно быть понятно, что иммунная функция пациентов с ADA-недостаточностью обычно не становится нормальной во время лечения с использованием PEG-ADA (Hershfield 1995; Hershfield and Mitchell 1995). Таким образом, иммуногенность может быть более важной проблемой в разработке ПЭГилированного фермента для продолжительного лечения пациентов с нормальной иммунной функцией.
Специалисту с обычной квалификацией в данной области будет понятно, что иммуногенность обозначает индукцию иммунного ответа введенным препаратом антигена (такого как ПЭГ-модифицированный белок или немодифицированный белок), тогда как антигенность обозначает реакцию антигена с предсуществующими антителами. В целом, антигенность и иммуногенность называют иммунореактивностью. В прежних исследованиях ПЭГ-уриказы иммунореактивность оценивали различными способами, в том числе: реакцией in vitro ПЭГ-уриказы с предварительно образованными антителами; измерениями синтеза индуцированных антител и ускоренными скоростями клиренса после повторяемых введений.
Было показано, что ПЭГилирование снижает иммуногенность и пролонгирует время циркуляции грибковой и свиной уриказ у животных (Chen et al. 1981; Savoca et al. 1984; Tsuji et al. 1985; Veronese et al. 1997). ПЭГ-модифицированная уриказа Candida быстро снижала содержание урата в сыворотке до не детектируемых уровней у 5 нормоурицемических людей-добровольцев (Davis et al. 1981b). ПЭГилированную уриказу Arthrobacter, производимую Enzon, Inc., использовали для лечения имеющего повышенную чувствительность к аллопуринолу пациента с лимфомой, который обнаруживал почечную недостаточность и явную гиперурицемию из этических соображений (Chua et al. 1988; Greenberg and Hershfield 1989). Четыре внутримышечные инъекции вводили на протяжении приблизительно двух недель. Во время этого короткого периода гиперурицемия устранялась и антитела против уриказы не могли быть детектированы при помощи ELISA в плазме пациента. Дальнейшее применение и клиническая разработка этого препарата не были продолжены.
К настоящему времени, не была разработана ни одна форма уриказы или ПЭГ-уриказы, которая имеет время существования в кровообращении подходящей длительности и достаточно пониженную иммуногенность для безопасного и надежного применения в длительной терапии. Целью данного изобретения является обеспечение улучшенной формы уриказы, которая, в сочетании с ПЭГилированием, может удовлетворять этим требованиям. Данное изобретение заключается в получении уникального производного рекомбинантной уриказы млекопитающего, которая была модифицирована мутацией таким способом, который, как было показано, повышает способность ПЭГилирования маскировать потенциально иммуногенные эпитопы.
Сущность изобретения
Основной целью данного изобретения является представление новых уриказных белков и кодирующих их последовательностей нуклеиновых кислот.
Другой целью данного изобретения является представление способа очистки рекомбинантно полученных уриказных белков, таких как описанные здесь.
Следующей целью данного изобретения является представление способа уменьшения количества мочевой кислоты в жидкости тела млекопитающего путем введения композиции, содержащей уриказный белок данного изобретения, млекопитающему.
Еще одной целью данного изобретения является представление антител к описанным здесь уриказным белкам.
Следующей целью данного изобретения является представление векторов и клеток-хозяев, содержащих описанные здесь последовательности нуклеиновых кислот, и способов их применения для получения кодируемых ими уриказных белков.
Данное изобретение представляет уриказные белки, которые могут быть использованы для получения по существу неиммуногенной ПЭГ-уриказы, которая сохраняет всю или почти всю уриколитическую активность немодифицированного фермента. Уриколитическая активность выражена здесь в Международных Единицах (ME) на мг белка, где одна ME уриказной активности определена как количество фермента, которое потребляет один микромоль мочевой кислоты в минуту.
Данное изобретение представляет рекомбинантный уриказный белок вида млекопитающего, который был модифицирован для встраивания одного или нескольких остатков лизина. Термин "рекомбинантный белок" в применении здесь относится к любому искусственно полученному белку и рекомбинантный белок отличается от природно продуцируемых белков (т.е. белков, которые продуцируются в тканях животного, который имеет только природный ген для представляющего интерес специфического белка). Термин "белок" включает в себя пептиды и аминокислотные последовательности. Рекомбинантный уриказный белок данного изобретения может быть химерой или гибридом из двух или более белков, пептидов или аминокислотных последовательностей млекопитающего. В одном варианте, данное изобретение может быть использовано для получения рекомбинантного уриказного белка видов млекопитающих, причем этот белок был модифицирован для увеличения числа лизинов до такого количества, при котором, после ПЭГилирования этого рекомбинантного уриказного белка, ПЭГилированный уриказный продукт является по существу таким же ферментативно активным, что и немодифицированная уриказа, и этот ПЭГилированный уриказный продукт не является неприемлемо иммуногенным. Также представлены укороченные формы уриказ данного изобретения, в которых могут отсутствовать амино- и/или карбокси-концевые участки уриказы. Предпочтительно уриказа не является укороченной до такой степени, что удаляются лизины.
Специалисту в данной области будет понятно, что конъюгированный комплекс уриказаноситель не должен содержать так много связей, чтобы существенно уменьшать ферментативную активность уриказы, или слишком мало связей, так что он остается неприемлемо иммуногенным. Предпочтительно, этот конъюгат будет сохранять, по меньшей мере, приблизительно 70-90% уриколитической активности немодифицированного белка уриказы, являясь более стабильным, так что он сохраняет его ферментативную активность при хранении, в плазме и/или сыворотке млекопитающего при физиологической температуре, в сравнении с немодифицированным белком уриказы. Сохранение, по меньшей мере, приблизительно 80-85% уриколитической активности было бы приемлемым. Кроме того, в предпочтительном варианте, этот конъюгат обеспечивает существенно уменьшенную иммуногенность и/или иммунореактивность, в сравнении с немодифицированным белком уриказы. В одном варианте, данное изобретение представляет уриказный белок, описанный здесь, который может быть модифицирован присоединением к нетоксичному, неиммуногенному, фармацевтически приемлемому носителю, такому как ПЭГ, ковалентной связью с, по меньшей мере, 1 из лизинов, содержащихся в этом уриказном белке. Альтернативно, уриказный белок модифицирован ковалентным присоединением к носителю через менее чем приблизительно 10 лизинов, его аминокислотной последовательности. В качестве альтернативных вариантов, предлагается присоединение к 2, 3, 4, 5, 6, 7, 8 или 9 из этих лизинов.
Уриказный белок данного изобретения является рекомбинантной молекулой, которая включает в себя сегменты белков уриказы печени свиньи и павиана. Представлена также модифицированная последовательность павиана. В одном варианте, данное изобретение представляет химерную уриказу свиньи-павиана (РВС-уриказу (SEQ ID NO:2)), которая включает аминокислоты (аа) 1-225 свиной уриказы (SEQ ID NO:7) и аминокислоты (аа) 226-304 уриказы павиана (SEQ ID NO:6) (см. также последовательность на фигуре 5). В другом варианте, данное изобретение представляет химерную уриказу свиньи-павиана (PKS-уриказу), которая включает аминокислоты (аа) 1-288 свиной уриказы и аминокислоты (аа) 289-304 уриказы павиана (SEQ ID NO:4). Укороченные производные РВС и PKS также рассматриваются. Предпочтительными укороченными формами являются белки РВС и PKS, укороченные таким образом, что у них делегированы 6 амино-концевых аминокислот или 3-карбокси-концевые аминокислоты или и те и другие. Последовательности, представленные в SEQ ID NO:8 (амино-укороченная РВС), SEQ ID NO:9 (карбокси-укороченная РВС), SEQ ID NO:10 (амино-укороченная PKS) и SEQ ID NO:11 (карбокси-укороченная PKS), приведены в качестве примера. Каждая из РВС-уриказы, PKS-уриказы и их укороченных форм имеет на один-четыре лизина больше, чем обнаружено в других уриказах млекопитающих, которые были клонированы.
Данное изобретение представляет молекулы (последовательности) нуклеиновых кислот (ДНК и РНК), в том числе выделенные, очищенные и/или клонированные формы молекул нуклеиновых кислот, которые кодируют описанные здесь уриказные белки и укороченные белки. Предпочтительные варианты показаны в SEQ ID NO:1 (РВС-уриказа) и SEQ ID NO:3 (PKS-уриказа).
Данное изобретение представляет также векторы (экспрессирующие и клонирующие), включающие эти молекулы нуклеиновых кислот.
Кроме того, данное изобретение представляет клетки-хозяева, содержащие эти векторы.
Представлены также антитела, которые специфически связываются с уриказными белками данного изобретения. Антитела к амино-части свиной уриказы и антитела к карбокси-части уриказы павиана, при совместном использовании, должны быть применимы в детектировании РВС или других подобных химерных белков. Предпочтительно, антитело к амино-части химерной уриказы не должно узнавать амино-часть уриказы павиана, и, подобным образом, антитело к карбокси-части химерной уриказы не должно узнавать карбокси-часть свиной уриказы. Более предпочтительно, представлены антитела, которые специфически связываются с РВС или PKS, но не связываются с нативными белками, такими как уриказы свиньи и/или павиана.
В другом варианте, данное изобретение может быть использовано для получения фармацевтической композиции для снижения количества мочевой кислоты в жидкостях тела, таких как моча и/или сыворотка или плазма, содержащей, по меньшей мере, один из уриказных белков или конъюгатов уриказы, описанных здесь, и фармацевтически приемлемый носитель, разбавитель или наполнитель.
Данное изобретение может быть также использовано в способе для уменьшения количества мочевой кислоты в жидкостях тела млекопитающего. Этот способ предусматривает введение млекопитающему эффективного снижающего мочевую кислоту количества композиции, содержащей уриказный белок или конъюгат уриказы данного изобретения и разбавитель, носитель или наполнитель, который предпочтительно является фармацевтически приемлемым носителем, разбавителем или наполнителем.
Млекопитающим, которого подвергают лечению, предпочтительно является человек.
Путь введения может быть, например, внутривенный, внутрикожный, подкожный, внутримышечный или внутрибрюшинный. Повышенные уровни мочевой кислоты могут быть в крови или моче и могут быть связаны с подагрой, отложением солей, почечной недостаточностью, трансплантацией органов или злокачественным заболеванием.
В другом варианте, данное изобретение представляет способ выделения и/или очистки уриказы из раствора уриказы, содержащего, например, клеточный или субклеточный дебрис, из, например, рекомбинантного способа получения. Предпочтительно, способ очистки использует вследствие ограниченной растворимости уриказы млекопитающих при низком рН (Conley et al. 1979), путем промывания неочищенного рекомбинантного экстракта при рН приблизительно 7-8,5 для удаления большей части белков, которые растворимы в этом низком диапазоне рН, после чего активную уриказу растворяют в буфере, предпочтительно натрий-карбонатном буфере, при рН приблизительно 10-11, предпочтительно около 10,2. Затем растворенную активную уриказу можно нанести на анионообменную колонку, такую как колонка Q-Sepharose, которую промывают низкосолевым-высокосолевым градиентом в буфере при рН приблизительно 8,5, после чего очищенную уриказу получают элюцией градиентом хлорида натрия в натрий-карбонатном буфере при рН приблизительно 10-11, предпочтительно около 10,2. Фермент может быть очищен дополнительно гель-фильтрационной хроматографией при рН приблизительно 10-11. На этой стадии фермент может быть очищен дополнительно путем снижения рН до приблизительно 8,5 или ниже для селективного осаждения уриказы, но не более растворимых примесей. Затем после промывания при низком рН (7-8) уриказу растворяют при рН около 10,2. Затем препарат уриказы может быть подвергнут анализам с использованием способов, известных в области получения фармацевтических препаратов, таких как, например, жидкостная хроматография высокого разрешения (ВЭЖХ), другие хроматографические способы, светорассеяние, центрифугирование и/или гель-электрофорез.
Краткое описание рисунков
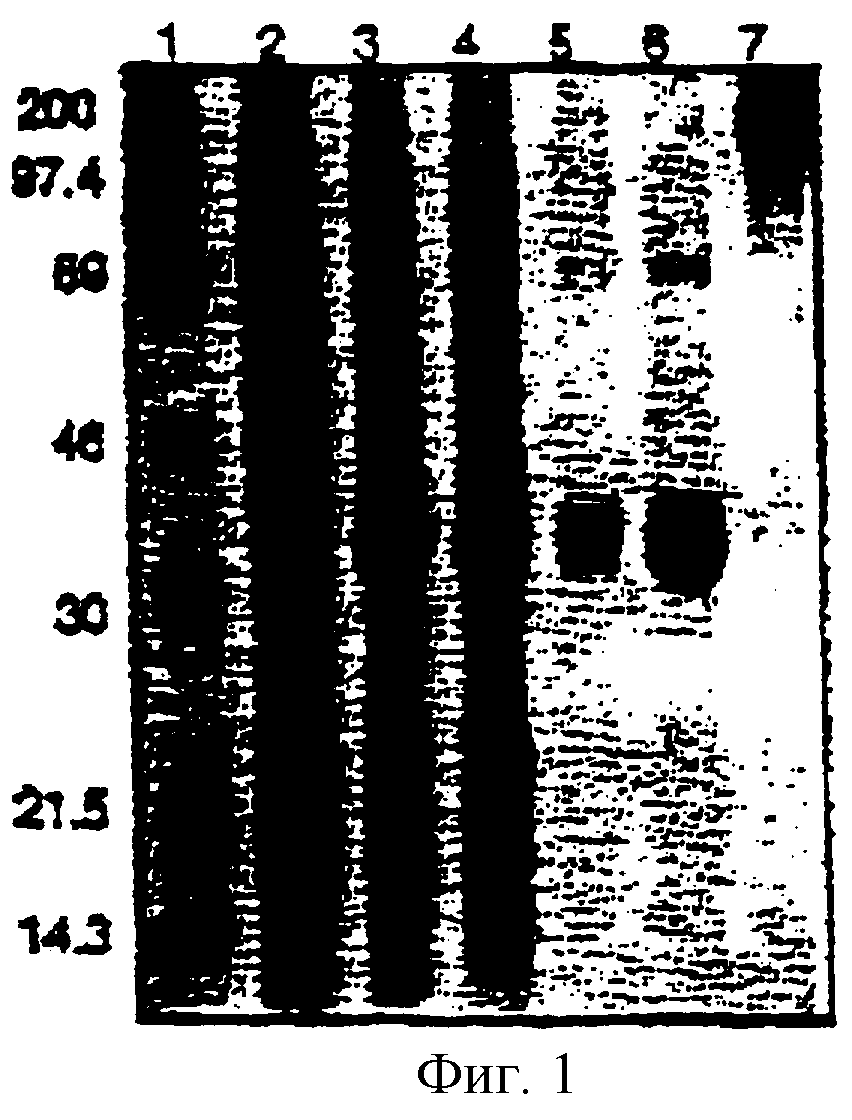
Фигура 1. Анализ при помощи гель-электрофореза в ПААГ с ДСН-меркаптоэтанолом (12% гель).
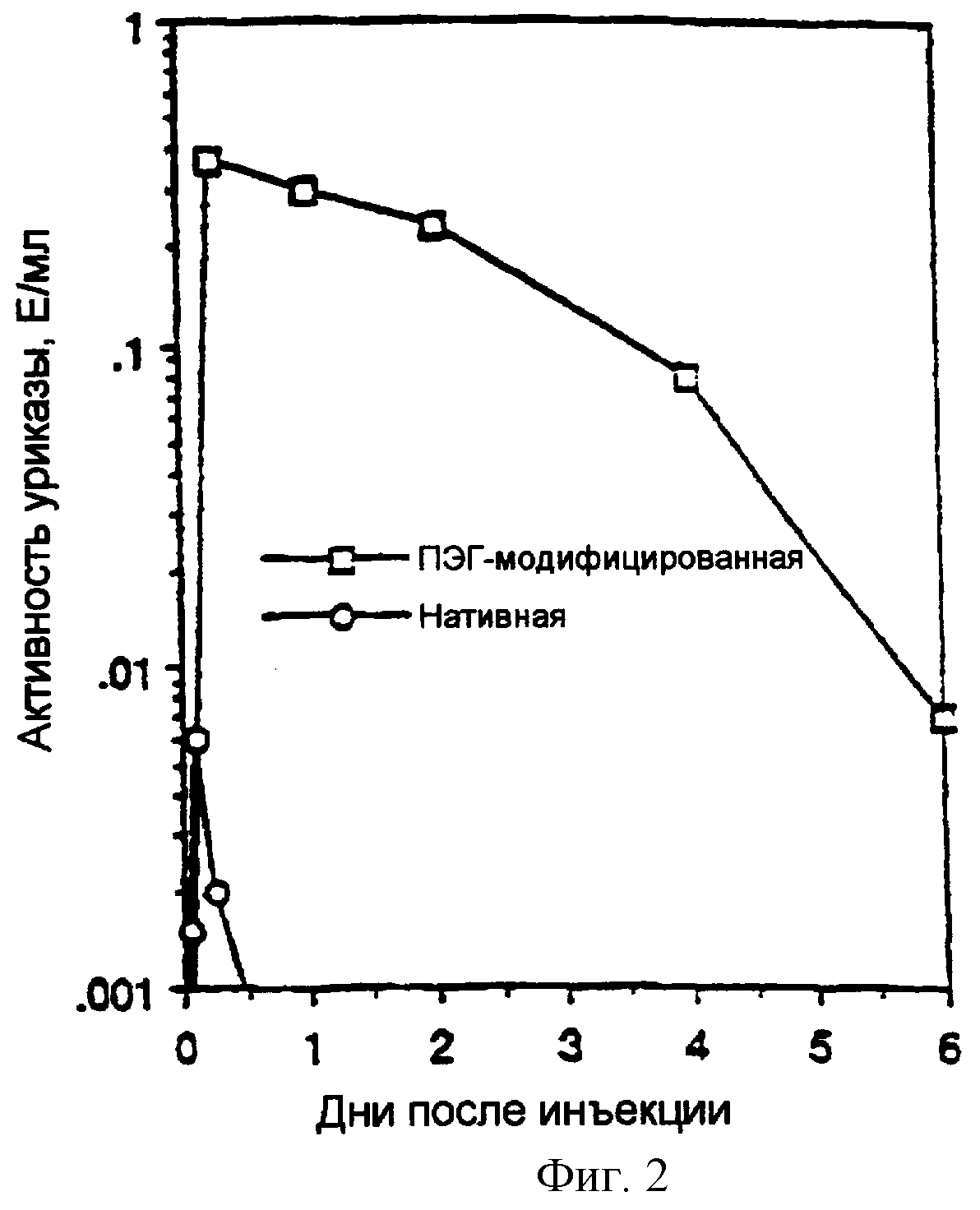
Фигура 2. Время циркуляции нативной и ПЭГилированной РВС-уриказы.
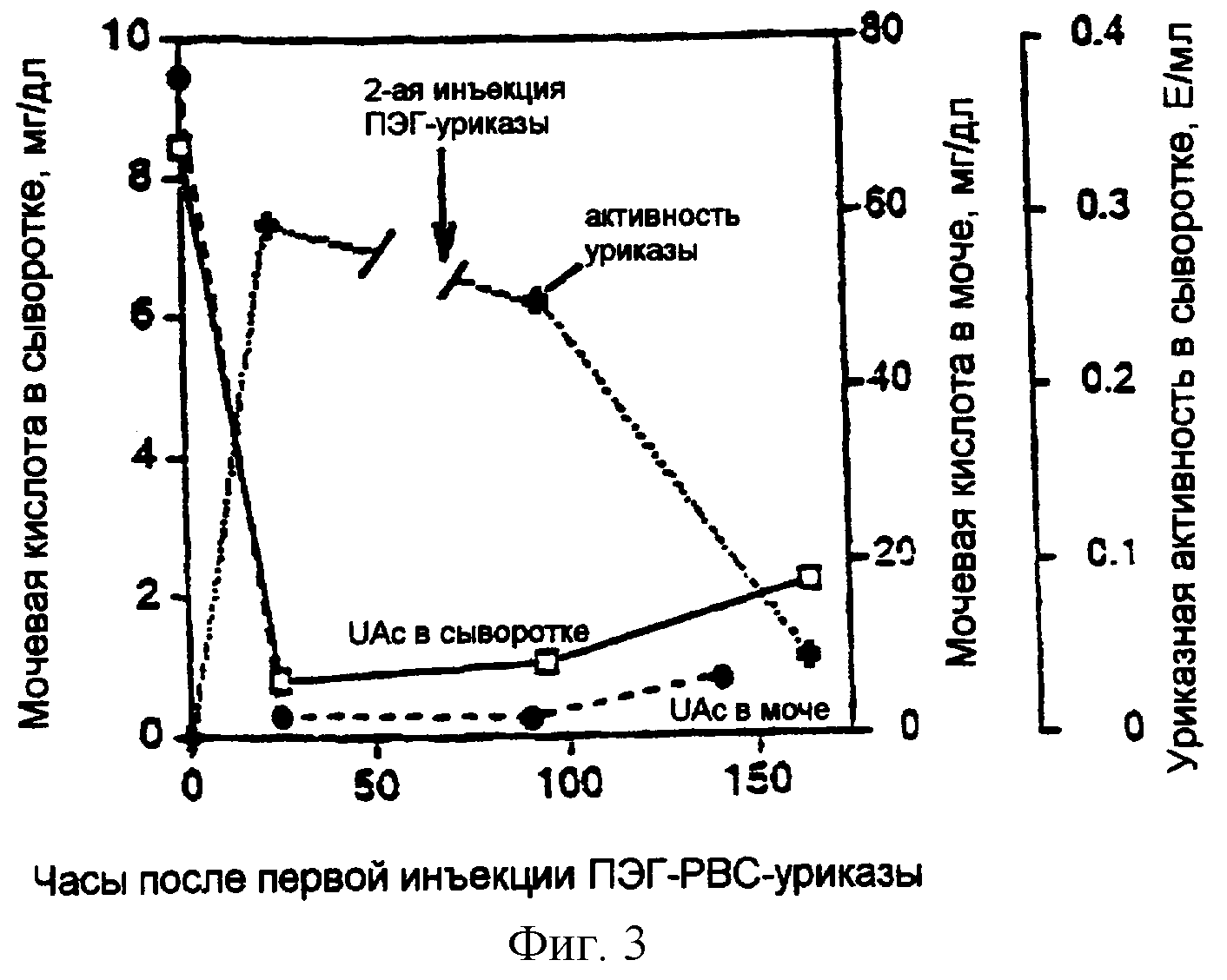
Фигура 3. Взаимосвязь активности уриказы в сыворотке и концентраций мочевой кислоты в сыворотке и моче.
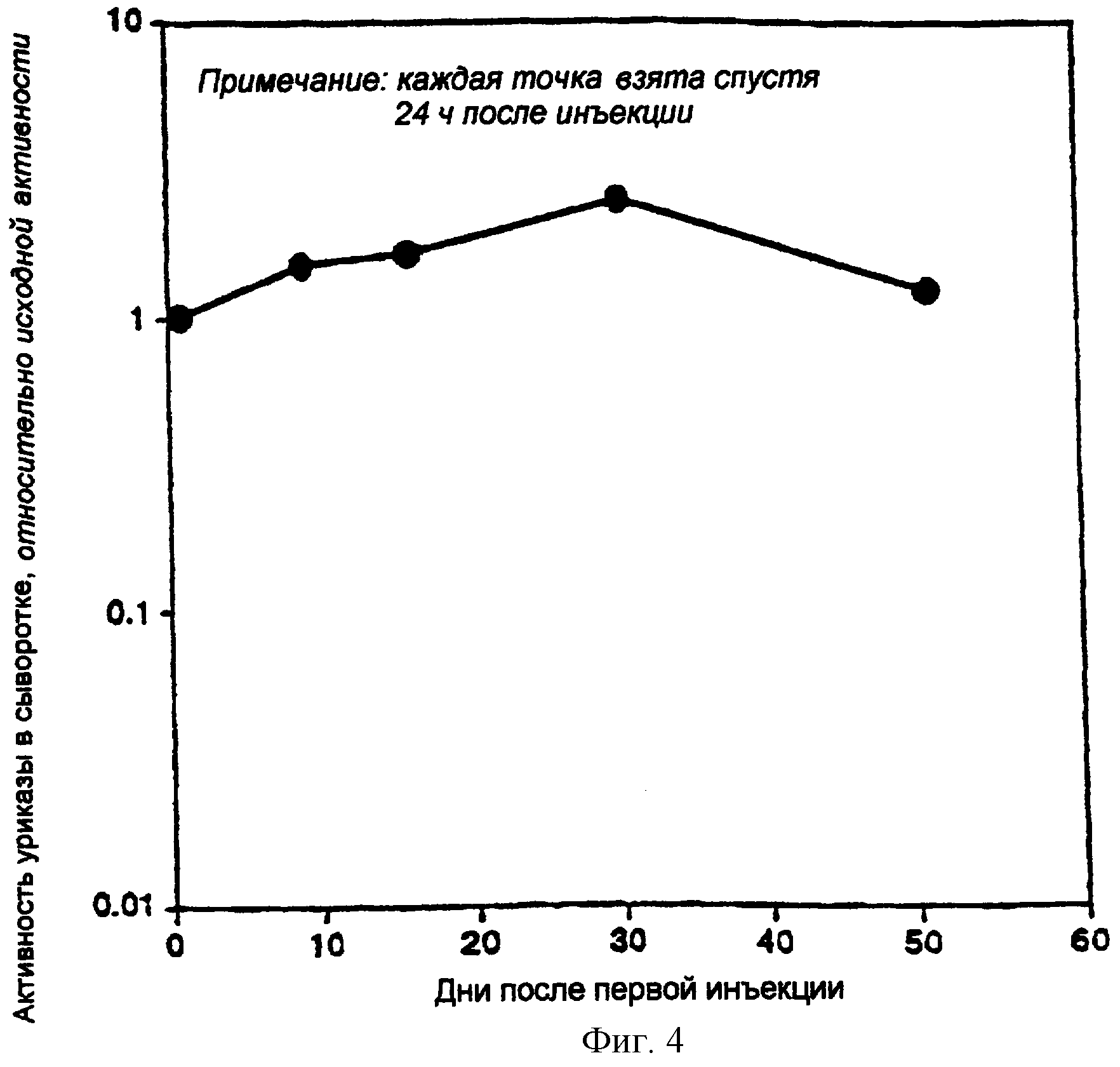
Фигура 4. Поддержание уровня циркуляции уриказной активности (измеренного в сыворотке) после повторяемой инъекции.
Фигура 5 показывает расшифрованные аминокислотные последовательности химерной уриказы свиньи-павиана (РВС-уриказы) (SEQ ID NO:2) и свиной уриказы, содержащей мутации R291K и T301S (PKS-уриказы) (SEQ ID NO:4), в сравнении с последовательностями свиной уриказы (SEQ ID NO:7) и уриказы павиана (SEQ ID NO:6).
Фигура 6. Сравнение аминокислотных последовательностей PKS-уриказы (SEQ ID NO:4) и свиной уриказы (SEQ ID NO:7).
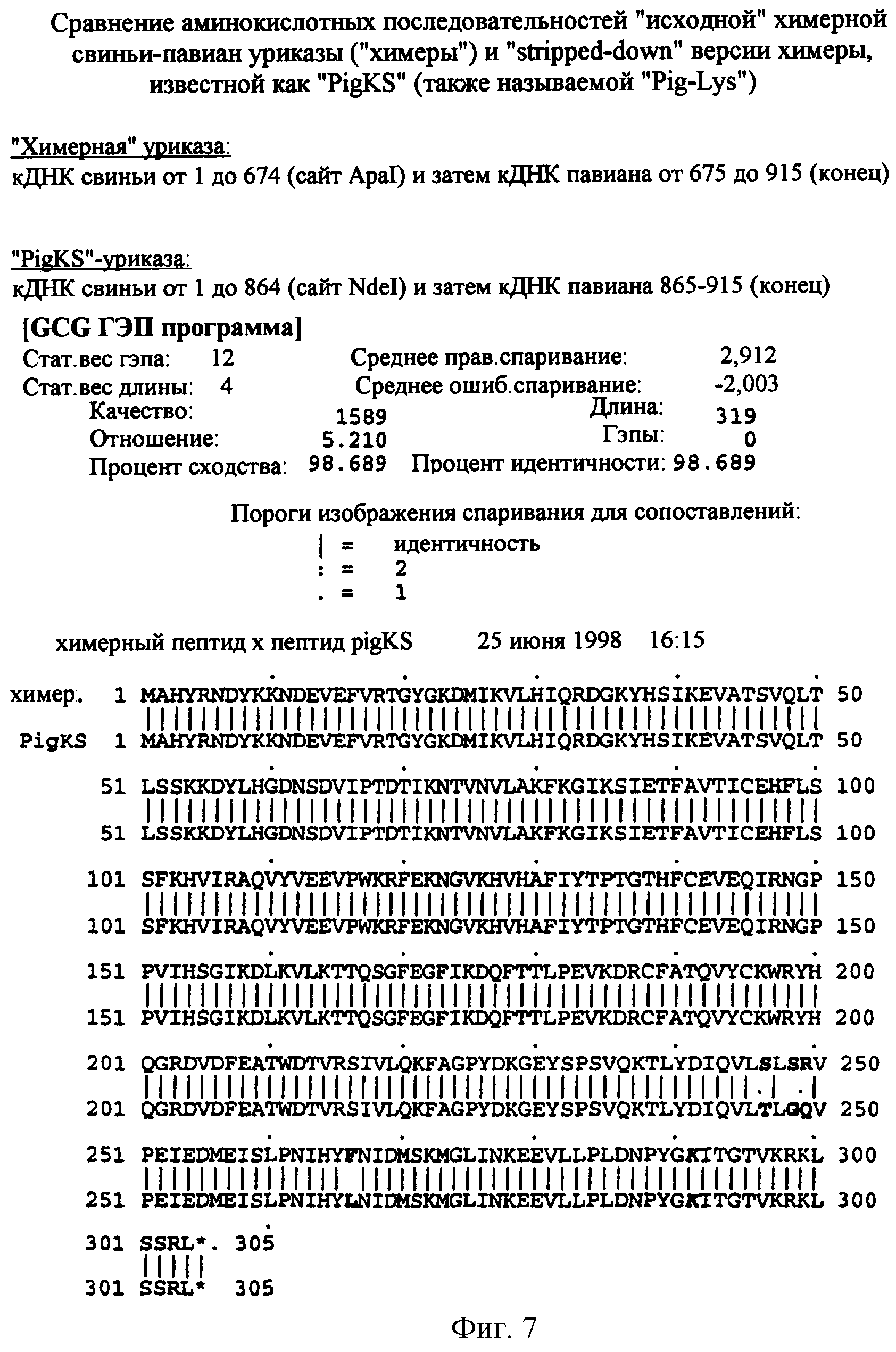
Фигура 7. Сравнение аминокислотных последовательностей РВС (SEQ ID NO:2) и PKS (SEQ ID NO:4).
Фигура 8. Сравнение аминокислотных последовательностей РВС (SEQ ID NO:2) и свиной (SEQ ID NO:7) уриказы.
Фигура 9. Сравнение аминокислотной последовательности свиной уриказы (SEQ ID NO:7) и D3H (SEQ ID NO:5).
Фигура 10. Сравнение аминокислотных последовательностей РВС (SEQ ID NO:2) и D3H (SEQ ID NO:5).
Фигура 11-1 и 11-2. Сравнение Bestfit (программное обеспечение GCG) (сравнение с оптимальным сопоставлением) кодирующих последовательностей кДНК PKS (SEQ ID NO:3) и свиной (SEQ ID NO:12) уриказы.
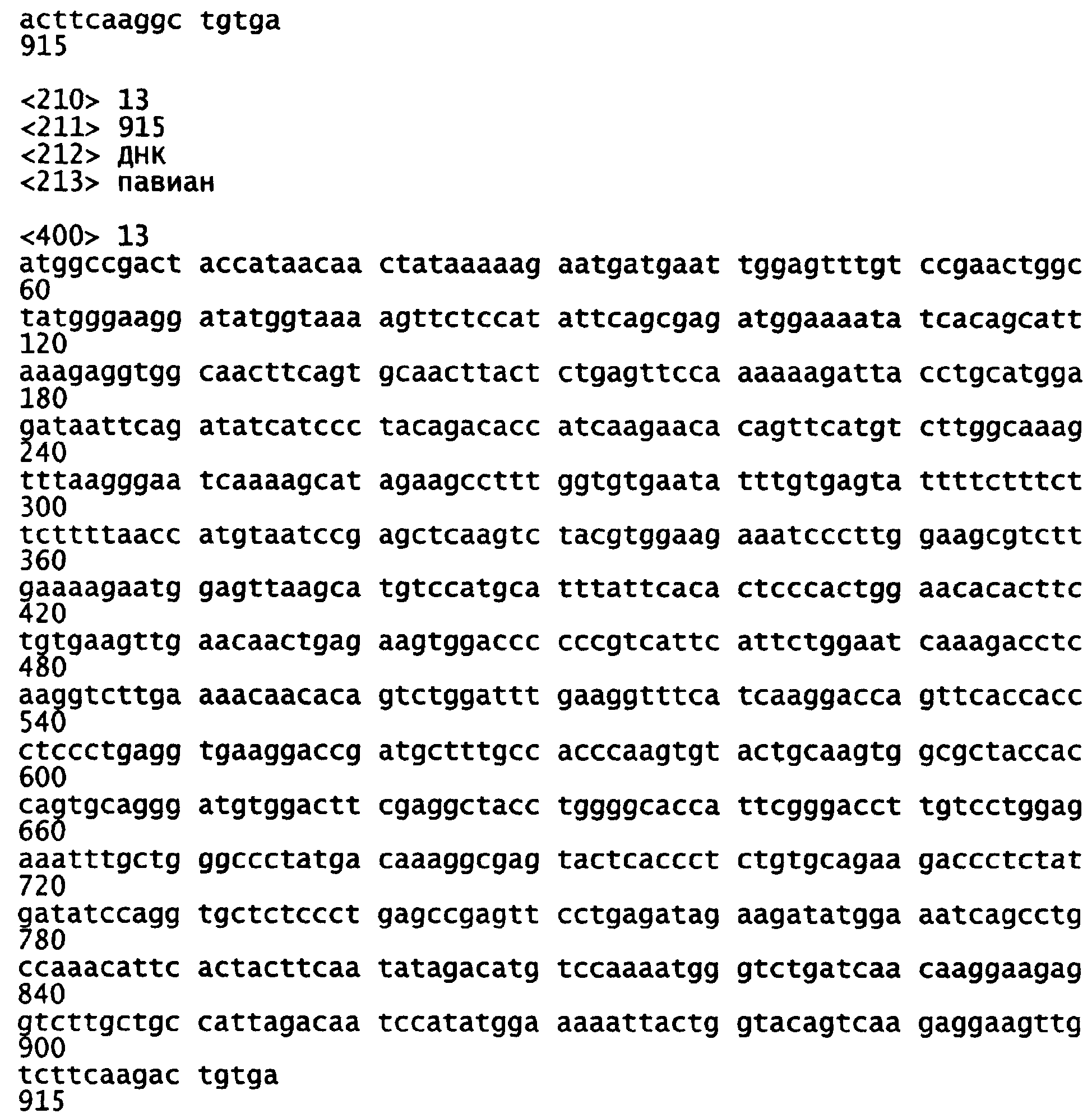
Фигура 12-1 и 12-2. Сравнение Bestfit (программное обеспечение GCG) кодирующих последовательностей кДНК PKS (SEQ ID NO:3) и уриказы павиана (SEQ ID NO:13).
Фигура 13-1 и 13-2. Сравнение Bestfit (программное обеспечение GCG) кодирующих последовательностей кДНК РВС (SEQ ID NO:1) и свиной (SEQ ID NO:12) уриказы.
Фигура 14-1 и 14-2. Сравнение Bestfit (программное обеспечение GCG) кодирующих последовательностей кДНК РВС (SEQ ID NO:1) и уриказы павиана (SEQ ID NO:13).
Подробное описание изобретения
Данное изобретение представляет уриказные белки, которые являются ценными промежуточными продуктами для улучшенных уриказных конъюгатов водорастворимых полимеров, предпочтительно поли(этиленгликолей) или поли(этиленоксидов), с уриказами. Под уриказой здесь подразумевают индивидуальные субъединицы, а также нативный тетрамер, если нет иных указаний.
Хотя в человеке не продуцируется активный фермент, мРНК-транскрипты уриказы были амплифицированы из РНК печени человека (Wu et al. 1992). Теоретически возможно, что некоторые транскрипты уриказы человека транслируются; даже если эти пептидные продукты были не полноразмерными или были нестабильными, они могли бы быть процессированы антигенпредставляющими клетками и играть роль в определении иммунологического ответа на экзогенную уриказу, используемую для лечения. Теоретически могут быть возможными реконструкция и экспрессия кДНК уриказы человека путем устранения двух известных нонсенс-мутаций (бессмысленных мутаций). Однако, в отсутствие давления отбора, очень вероятно, что неблагоприятные миссенс-мутации (мутации с изменением смысла) накапливались в этом гене человека в течение миллионов лет с введения первой нонсенс-мутации (Wu et al. 1989; Wu et al. 1992). Идентификация и "коррекция" всех мутаций для получения максимальной каталитической активности и стабильности белка была бы очень трудной.
Авторы данного изобретения понимали, что существует высокая степень гомологии (сходства) между расшифрованной аминокислотной последовательностью уриказы человека и аминокислотными последовательностями уриказ свиньи (около 86%) и павиана (около 92%) (см., фигуры 6-14, например, измерение сходства), тогда как гомология (сходство) между уриказами человека и A. flavus составляет <40% (Lee et al. 1988; Ready et а1. 1988; Wu et al. 1989; Legoux et а1. 1992; Wu et al. 1992). Данное изобретение представляет полученные рекомбинантно химерные уриказные белки из двух различных млекопитающих, которые были сконструированы для того, чтобы они были менее иммунореактивными для человека, чем более отдаленно родственный грибковый или бактериальный фермент. Ожидается, что применение производного уриказы млекопитающих является более приемлемым для пациентов и их врачей.
Опыт показал, что активированные ПЭГ, такие, которые были использованы для получения PEG-ADA и для модификации других белков, присоединяются через первичные аминогруппы амино-концевого остатка (когда они присутствуют и являются неблокированными) и эпсилон-аминогруппы лизинов. Эта стратегия применима, поскольку могут быть использованы средние условия реакции и поскольку положительно заряженные лизины имеют тенденцию к локализации на поверхности белков. Последнее является важным, так как для любого терапевтического белка желательные эффекты ПЭГилирования будут зависеть отчасти от характеристик ПЭГ-полимера (например, массы, разветвленной или неразветвленной структуры и т.д.), а также от числа и распределения сайтов присоединения ПЭГ белка относительно эпитопов и структурных элементов, которые определяют функцию и клиренс данного белка. Была придумана стратегия для усиления способности ПЭГилирования "маскировать" эпитопы и понижать иммуногенность путем полуселективного введения новых остатков лизина для потенциального присоединения ПЭГ (Hershfield et al. 1991). Эта стратегия использует мутагенез для замены выбранных кодонов аргинина кодонами лизина, замены, которая сохраняет положительный заряд и оказывает минимальное действие на предсказанные компьютером показатели поверхностной вероятности и антигенности (применимые, когда известна только одна аминокислотная последовательность).
В качестве экспериментального испытания этой стратегии, использовали рекомбинантную пуриннуклеозидфосфорилазу Е. coli (EPNP) (Hershfield et al. 1991). Вводили замены Arg на Lys в 3 сайтах, увеличивая число лизинов на субъединицу с 14 до 17, без изменения каталитической активности. Очищенный тройной мутант сохранял полную активность после модификации ˜70% доступных NH2 -групп избытком дисукцинил-ПЭГ5000. Титрование реакционноспособных аминогрупп до и после ПЭГилирования предполагало, что тройной мутант мог акцептировать больше на одну цепь ПЭГ на субъединицу, чем фермент дикого типа. ПЭГилирование увеличивало время циркуляции как EPNP-фермента дикого типа, так и мутантного EPNP-фермента в мышах с ˜4 часов до >6 дней. После серии внутрибрюшинных инъекций с интервалами одна неделя/две недели, у всех мышей, обработанных немодифицированными EPNP, и у 10 из 16 мышей (60%), инъецированных ПЭГилированной EPNP дикого типа, развивались высокие уровни антител против EPNP и заметно снижалось время циркуляции. В противоположность этому, только у 2 из 12 мышей (17%), обработанных мутантным ПЭГ-EPNP, развился быстрый клиренс; низкие уровни антител в этих мышах не коррелировали с временем циркуляции. Таким образом, эта стратегия была успешной в существенном уменьшении иммуногенности, даже хотя только 1 из 3 новых лизинов становился модифицированным после обработки активированным ПЭГ.
Каждая из субъединиц уриказы павиана и свиньи состоит из 304 аминокислот, 29 из которых (т.е. 1 приблизительно на 10 остатков) представляют собой лизины. Первоначальные попытки введения 2 замен Arg на Lys в клонированную кДНК уриказы павиана, а также замены кодон Lys на Glu в положении 208, которое соответствует Lys в гене уриказы человека, привели к экспрессированному мутантному белку павиана, который имел значительно уменьшенную уриказную каталитическую активность. Из этого эксперимента видно, что способность сохранять уриказную ферментативную активность после мутации аргинина на лизин ДНК-последовательности млекопитающих была непредсказуемой.
Впоследствии было обнаружено, что аминокислотный остаток 291 в уриказе павиана является лизином, а соответствующий остаток в уриказе свиньи является аргинином. Сайт рестрикции Apal, присутствующий в обеих кДНК, использовали для конструирования химерной уриказы, в которой первые 225 аминокислот произведены из кДНК свиньи, а карбокси-концевые 79 аминокислот произведены из кДНК павиана. Полученная химерная уриказа свиньи-павиана (РВС) (SEQ ID NO:2) имеет 30 лизинов, на один лизин больше, чем любой «исходный» фермент. Дополнительным признаком РВС-уриказы является то, что ее «павиан»-часть отличается от уриказы человека в 4 из 79 аминокислотных остатков, тогда как уриказы свиньи и человека различаются в 10 остатках в том же самом участке. Впоследствии была сконструирована модифицированная версия РВС, которая сохраняет лишний лизин в положении 291, а в других отношениях отличается от уриказы свиньи только заменой треонина серином в остатке 301 (уриказа "pigKS" (SEQ ID NO:4)). В связи с результатами, описанными в предыдущем абзаце, где несколько других инсерций лизинов были неблагоприятными для активности, было неожиданным, что химерные уриказы РВС и PKS были такими же активными в сравнении с немутированной нативной уриказой свиньи и приблизительно более чем в 4 раза, более активными, чем немутированная нативная уриказа павиана.
Данное изобретение представляет рекомбинантную химерную уриказу свиньи-павиана, составленную из частей последовательностей уриказы печени свиньи и павиана. Один пример такой химерной уриказы содержит первые 225 аминокислот из последовательности уриказы свиньи (SEQ ID NO:7) и последние 79 аминокислот из последовательности уриказы павиана (SEQ ID NO:6) (уриказа свиньи-павиана или уриказа РВС; фигура 6 и SEQ ID NO:2). Другой пример такой химерной уриказы содержит первые 288 аминокислот из последовательности уриказы свиньи (SEQ ID NO:7) и последние 16 аминокислот из последовательности уриказы павиана (SEQ ID NO:6). Поскольку эта последняя последовательность отличается от последовательности свиньи только в двух положениях, имея лизин (К) вместо аргинина в остатке 291 и серин (S) вместо треонина в остатке 301, этот мутант называют pig-K-S или уриказой PKS.
Представлены также векторы (экспрессирующие и клонирующие), включающие молекулы нуклеиновых кислот, кодирующие белки данного изобретения. Предпочтительные векторы представлены здесь в качестве примера. Специалисту в данной области будет понятно, что молекулы нуклеиновых кислот могут быть встроены в экспрессирующий вектор, такой как плазмида, в правильной ориентации и точной рамке считывания для экспрессии. Если необходимо, нуклеиновая кислота (ДНК) может быть связана с подходящими транскрипционными и трансляционными регуляторными нуклеотидными последовательностями, узнаваемыми желательным хозяином, хотя такие регуляторные элементы обычно доступны в экспрессирующих векторах, используемых и известных в данной области. Затем вектор может быть введен в клетки-хозяева при помощи стандартных способов. Обычно не все из клеток-хозяев будут трансформированы этим вектором. Таким образом, может быть необходимым отбор трансформированных клеток-хозяев. Один из таких способов отбора, известный из уровня техники, включает встраивание в экспрессирующий вектор ДНК-последовательности с любыми необходимыми регуляторными элементами, которая кодирует селектируемый маркерный признак в трансформированной клетке, такой как устойчивость к антибиотику. Альтернативно, ген для такого селектируемого признака может находиться в другом векторе, который используют для котрансформации целевых клеток-хозяев. Векторы могут также включать в себя подходящий промотор, такой как прокариотический промотор, способный к экспрессии (транскрипции и трансляции) этой ДНК в бактериальной клетке-хозяине, такой как Е. coli, трансформированной этим вектором. Многие экспрессионные системы доступны и известны в данной области, в том числе бактериальные системы (например, Е. coli и Bacillus subtilis), дрожжи (например, Saccharomyces cerevisiae), нитчатые грибы (например, Aspergillus), клетки растений, клетки животных и клетки насекомых.
Подходящие векторы могут включать в себя прокариотический репликон, такой как ColEI ori, для размножения, например, в прокариотах. Типичными прокариотическими векторными плазмидами являются pUC18, pUC19, pUC322 и pBR329, доступные из Biorad Laboratories (Richmond, CA) и рТсr99А и pKK223-3, доступные из Pharmacia (Piscataway, NJ). Типичной векторной плазмидой клеток млекопитающих является pSVL, доступная из Pharmacia (Piscataway, NJ). Этот вектор использует поздний промотор SV40 для запуска экспрессии клонированных генов, причем наивысший уровень экспрессии обнаружен в Т-антигенпродуцирующих клетках, таких как клетки COS-1. Примером индуцируемого экспрессирующего вектора млекопитающих является pMSG, также доступный из Pharmacia. Этот вектор использует индуцируемый глюкокортикоидом промотор длинного концевого повтора вируса опухоли молочной железы мыши для запуска экспрессии клонированного гена. Применимыми дрожжевыми плазмидными векторами являются pRS403-406 и pRS413-416 и они обычно доступны из Stratagene Cloning Systems (La Jolla, CA). Плазмиды pRS403, pRS404, pRS405 и pRS406 являются дрожжевыми интегрирующими плазмидами (Yip) и включают дрожжевые селектируемые маркеры HIS3, TRP1, LEU2 и URA3. Плазмиды pRS413-416 являются дрожжевыми центромерными плазмидами (Ycp).
Кроме того, данное изобретение представляет клетки-хозяева, содержащие эти векторы. Предпочтительные клетки-хозяева включают в себя приведенные здесь в качестве примеров и описанные здесь клетки-хозяева.
Уриказные белки данного изобретения могут быть конъюгированы через биологически стабильную, нетоксичную, ковалентную связь с относительно небольшим числом нитей ПЭГ для улучшения биологического периода полужизни и растворимости этих белков и уменьшения их иммунореактивности. Такие связи могут включать в себя уретановые (карбаматные) связи, вторичные аминные связи и амидные связи. Различные активированные ПЭГ, пригодные для такой конъюгации, коммерчески доступны из Shearwater Polymers, Huntsville, AL.
Данное изобретение может быть также использовано для получения фармацевтических композиций уриказных белков в виде конъюгатов. Эти конъюгаты являются по существу неиммуногенными и сохраняют, по меньшей мере, 70%, предпочтительно 80% и более предпочтительно, по меньшей мере, около 90% или более уриколитической активности немодифицированного фермента. Водорастворимые полимеры, пригодные для применения в данном изобретении, включают линейные и разветвленные поли(этиленгликоли) или поли(этиленоксиды), все обычно известные как ПЭГ. Один пример разветвленных ПЭГ является предметом патента США 5643575.
В одном варианте данного изобретения среднее число лизинов, встроенных на субъединицу уриказы, находится между 1 и 10. В предпочтительном варианте, число дополнительных лизинов на субъединицу уриказы находится между 2 и 8. Должно быть понятно, что число дополнительных лизинов не должно быть таким большим, чтобы наносить ущерб каталитической активности уриказы. Молекулы ПЭГ конъюгата предпочтительно конъюгированы через лизины уриказного белка, более предпочтительно через не встречающийся природно лизин или лизины, которые были введены в участок сконструированного белка, который природно не содержит лизин в этом участке.
Данное изобретение представляет способ увеличения доступных не причиняющих вреда сайтов присоединения ПЭГ к уриказному белку, в котором нативный уриказный белок мутируют таким образом, чтобы ввести в него, по меньшей мере, один остаток лизина. Предпочтительно этот способ включает замену аргининов лизинами.
ПЭГ-уриказные конъюгаты, использующие данное изобретение, применимы для снижения уровней (т.е. уменьшения количества) мочевой кислоты в крови и/или моче млекопитающих, предпочтительно людей, и могут быть, следовательно, использованы для лечения повышенных уровней мочевой кислоты, связанных с состояниями, включающими в себя подагру, отложения солей, почечную недостаточность, трансплантацию органов и злокачественное заболевание.
ПЭГ-уриказные конъюгаты могут быть введены в млекопитающее, имеющее избыточные уровни мочевой кислоты, любым из путей введения, в том числе пероральным, при помощи клизмы или суппозитория, внутривенным, подкожным, внутримышечным и внутрибрюшинным путями. Patton, JS, et al. (1992) Adv Drug Delivery Rev 8:179-228.
Эффективная доза ПЭГ-уриказы будет зависеть от уровня мочевой кислоты и размера индивидуума. В одном варианте этого аспекта данного изобретения ПЭГ-уриказу вводят в фармацевтически приемлемом наполнителе или разбавителе в количестве в диапазоне от 10 мкг до приблизительно 1 г. В предпочтительном варианте, вводимое количество находится между приблизительно 100 мкг и 500 мг. Более предпочтительно, конъюгированную уриказу вводят в количестве между 1 мг и 100 мг, например 5 мг, 20 мг или 50 мг. Массы, приводимые для дозовых количеств этих вариантов, относятся к количеству белка в конъюгате.
Фармацевтические композиции, содержащие ПЭГ-уриказу могут быть получены общепринятыми способами, например, как описано в Remington's Pharmaceutical Sciences (1985) Easton, PA: Mack Publishing Co. Подходящие наполнители для получения инъекционных растворов включают в себя, например, забуференный фосфатом солевой раствор, содержащий лактат раствор Рингера, воду, полиолы и глицерин. Фармацевтические композиции для парентерального введения содержат фармацевтически приемлемые стерильные водные или неводные жидкости, дисперсии, суспензии или эмульсии, а также стерильные порошки для реконструирования в стерильные инъекционные растворы или дисперсии непосредственно перед использованием. Эти композиции могут содержать дополнительные компоненты, такие как, например, консерванты, растворители, стабилизаторы, увлажняющие агенты, эмульгаторы, буферы, антиоксиданты и разбавители.
ПЭГ-уриказа может быть также представлена в виде композиций с замедленным высвобождением для имплантации в индивидуум для непрерывной регуляции уровней мочевой кислоты в крови и моче. Например, полимолочная кислота, полигликолевая кислота, регенерированный коллаген, поли-L-лизин, альгинат натрия, геллановая (gellan) камедь, хитозан, ага-роза, многослойные липосомы и многие другие общепринятые депо-композиции содержат биоразлагаемые или биодеградируемые материалы, которые могут быть приготовлены с биологически активными композициями. Эти материалы, при имплантации или инъекции, постепенно разрушаются и высвобождают активный материал в окружающую ткань. Например, одним способом инкапсулирования ПЭГ-уриказы является способ, описанный в патенте США №5653974, который включен здесь в качестве ссылки. Применение биоразлагаемых, биодеградируемых и других депо-композиций ясно предполагается в данном изобретении. Применение инфузионных насосов и систем инкапсулирования в матриксе для доставки ПЭГ-уриказы также находится в объеме данного изобретения. ПЭГ-уриказа может быть выгодно заключена в мицеллы или липосомы. Технология инкапсулирования в липосомах хорошо известна в данной области. См., например, Lasic, D, et al., (Eds.) (1995) Stealth Liposomes, Boca Raton, FL: CRC Press.
Фармацевтические композиции ПЭГ-уриказы, описанные здесь, будут уменьшать необходимость в гемодиализе в случае пациентов высокого риска индуцированной уратом почечной недостаточности, например, реципиентов трансплантируемых органов (см. Venkataseshan, VS. et al., (1990) Nephron 56:317-321) и пациентов с некоторыми злокачественными заболеваниями. У пациентов с большими накоплениями кристаллического урата (подагрическими узлами) такие фармацевтические композиции будут улучшать качество жизни более быстро, чем применимые в настоящее время способы лечения.
Следующие ниже примеры, которые не должны рассматриваться как ограничивающие каким-либо образом данное изобретение, иллюстрируют различные описанные выше аспекты.
ПРИМЕР 1
А. Конструирование кДНК РВС, PKS и родственных уриказ
Стандартные способы и инструкции по применению, в тех случаях, когда они прилагались изготовителем реагентов, использовали для получения общей клеточной РНК, для ПЦР-амплификации (патенты США №№4683195 и 4683202, 4965188 и 5075216) кДНК уратоксидаз и для клонирования и секвенирования этих кДНК (Erlich 1989; Sambrook et al. 1989; Ausubel 1998). ПЦР-праймеры для уратоксидаз свиньи и павиана (таблица 1) были сконструированы на основе опубликованных кодирующих последовательностей (Wu et al. 1989) и с использованием программы программного обеспечения PRIME (Genetics Computer Group, Inc.).
Последовательности рестрикционных ферментов (маленькие буквы), введенные на концах этих праймеров, представляют собой: смысловой (свинья и павиан) EcoRI и Ncol; антисмысловой (свинья) Ncol, HindIII, Xbal; антисмысловой (павиан) Ncol. В случае смыслового праймера павиана, третий кодон GAC (аспарагиновая кислота), присутствующий в уратоксидазе павиана (Wu et al. 1992), был заменен САС (гистидин), кодоном, который присутствует в этом положении в кодирующей последовательности псевдогена уратоксидазы человека (Wu et al. 1992). По этой причине рекомбинантная уратоксидаза павиана, полученная с использованием этих праймеров, была названа уратоксидазой павиана D3H.
Общую клеточную РНК из печени свиньи и печени павиана обратно транскрибировали с использованием набора для 1-й цепи (Pharmacia Biotech Inc. Piscataway, NJ). ПЦР-амплификацию с использованием ДНК-полимеразы Taq (Gibco BRL, Life Technologies, Gaithersburg, MD) выполняли в термоциклере (Ericomp, San Diego, CA) с программой [30 с, 95°С; 30 с, 55°С; 60 с, 70°С], 20 циклов, затем [30 с, 95°С; 60 с, 70°С] 10 циклов. ПЦР-продукты уратоксидазы расщепляли EcoRI и HindIII и клонировали в pUC18 (свинья) и также клонировали непосредственно (свинья и павиан D3H) с использованием системы клонирования ТА (Invitrogen, Carlsbad, CA). кДНК-клоны трансформировали в штамм Е. coli XLl-Blue (Stratagene, La Jolla, CA). Получали плазмидную ДНК, содержащую клонированные кДНК уриказы и встроенную последовательность кДНК анализировали при помощи стандартного дидезокси-способа. Клоны, которые имели опубликованные кодирующие ДНК-последовательности уратоксидазы (за исключением замены D3H в уратоксидазе павиана, описанной в таблице 1), конструировали и проверяли в серии последовательных стадий при помощи стандартной методологии рекомбинантных ДНК.
кДНК свиньи и павиана D3H, содержащие полноразмерные кодирующие последовательности, вводили в экспрессирующие векторы рЕТ (Novagen, Madison, WI) следующим образом. кДНК уриказы павиана D3H вырезали из плазмиды ТА рестрикционными ферментами Ncol и BamHI и затем субклонировали в сайты клонирования Ncol и BamHI экспрессионных плазмид pET3d и pET9d. Полноразмерную кДНК уриказы свиньи вырезали из клона плазмиды pUC рестрикционными ферментами EcoRI и HindIII и субклонировали в сайты EcoRI и HindIII pET28b. Кодирующий участок кДНК свиньи также вводили в сайты Ncol и Вlр1 экспрессионной плазмиды pET9d после вырезания из сайтов Ncol и BlpI pET28b.
Химерную кДНК свинья-павиан (РВС) конструировали вырезанием NcoI-ApaI-фрагмента рестрикции 624 п.н. кДНК уриказы павиана D3H из клона pET3d-D3H павиана и затем заменяли этот сегмент D3H павиана соответствующим NcoI-ApaI-фрагментом рестрикции 624 п.н. кДНК свиньи. Полученная кДНК уратоксидазы РВС состоит из кодонов 1-225 уратоксидазы свиньи, соединенных в рамке считывания с кодонами 226-304 уратоксидазы павиана.
кДНК уратоксидазы pig-KS (PigKS) конструировали вырезанием NcoI-NdeI-фрагмента рестрикции 864 п.н. кДНК уриказы павиана D3H из клона pET3d-D3H павиана и затем заменяли этот сегмент D3H павиана соответствующим NcoI-NdeI-фрагментом рестрикции 864 п.н. кДНК свиньи. Полученная кДНК уратоксидазы PKS состоит из кодонов 1-228 уратоксидазы свиньи, соединенных в рамке считывания с кодонами 289-304 уратоксидазы павиана.
Аминокислотные последовательности уратоксидаз павиана D3H, свиньи, РВС и PKS показаны на фигуре 5 и в СПИСКЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ. Стандартные способы использовали для получения исходных растворов в 15%-ном глицерине каждого из этих трансформантов и их хранили при -70°С. При экспрессии каждого из этих трансформантов и выделении рекомбинантных ферментов (таблица 2) уриказа свиньи, химерная уриказа РВС и уриказа PigKS имели очень сходную специфическую активность, которая была приблизительно в 4-5 раз выше, чем специфическая активность рекомбинантной уриказы павиана. Этот порядок подтверждали в нескольких других экспериментах. Специфическая активность уриказы РВС, полученной несколькими различными процедурами, варьировала в 2-2,5-кратном диапазоне.
Трансформанты Е. coli BL21(DE3)pLysS 4 конструкций кДНК уриказы-рЕТ, показанных в таблице 2, высевали на LB-агар, содержащий селективные антибиотики (карбенициллин и хлорамфеникол для pET3d (pigKS); канамицин и хлорамфеникол для pET9d (РВС, свинья, павиан)), как рекомендовано в руководстве рЕТ System Manual (Novagen, Madison, WI). 5-мл-культуры (LB плюс антибиотики) инокулировали одиночными колониями трансформантов и выращивали в течение 3 часов при 37°С. Затем аликвоты 0,1 мл переносили в 100 мл LB-среды, содержащей селективные антибиотики и 0,1% лактозу (для индукции экспрессии уриказы). После роста в течение ночи при 37°С бактериальные клетки из аликвот 0,5 мл этих культур экстрагировали в буфер для нанесения для электрофореза в ДСН-ПААГ и анализировали при помощи электрофореза в ПААГ с ДСН-меркаптоэтанолом; этот анализ установил, что сравнимые уровни уриказного белка экспрессировались в каждой из 4 культур (результаты не показаны). Остальные клетки из каждой культуры (100 мл) собирали центрифугированием и промывали в ЗФР. Затем клетки ресуспендировали в 25 мл забуференного фосфатом солевого раствора, рН 7,4 (ЗФР), содержащего 1 мМ ингибитор протеаз AEBSF (Calbiochem, San Diego, CA) и затем лизировали на льду в измельчителе бактериальных клеток (Microfluidics, Boston MA). Нерастворимый материал (включающий в себя уриказу) осаждали центрифугированием (20190 × g, 4°С, 15 мин.). Осадки промывали дважды 10 мл ЗФР и затем экстрагировали в течение ночи при 4°С 2 мл 1 М Na2СО3, рН 10,2. Экстракты разбавляли до 10 мл водой и затем центрифугировали (20190 × g, 4°С, 15 мин.). Затем определяли уриказную активность и концентрацию белка.
ПРИМЕР 2
Экспрессия и выделение рекомбинантной уриказы РВС (получение в ферментере на 4 л)
Трансформант уриказы рЕТЗо-РВС высевали из глицеринового исходного раствора на чашку с LB-агаром, содержащим карбенициллин и хлорамфеникол, как рекомендовано в руководстве Novagen pET System Manual. 200 мл инокулята, полученного из одиночной колонии, получали в жидкой среде LB, содержащей антибиотик, на ротационном шейкере (250 об/мин) при 37°С с использованием процедур, рекомендованных в руководсте pET System Manual, для максимизации сохранения плазмиды pET. При OD525 2,4, клетки из этих 200 мл-культур собирали центрифугированием и ресуспендировали в 50 мл свежей среды. Эту суспензию переносили в ферментер для более высокой плотности, содержащий 4 литра карбенициллин- и хлорам-фениколсодержащей среды SLBH (состав среды SLBH и конструкция и эксплуатация ферментера описаны в (Sadler et al. 1974)). После 20 часов выращивания в О2 при 32°С (OD525=19) добавляли изопропилтиогалактозид (IPTG) до 0,4 мМ для индукции образования уриказы. Спустя еще 6 часов (OD525=37) бактериальные клетки собирали центрифугированием (10410 × g, 10 минут, 4°С), промывали один раз ЗФР и хранили в замороженном виде при -20°С.
Бактериальные клетки (189 г) ресуспендировали в 200 мл ЗФР и лизировали при охлаждении в бане со льдом и солью обработкой ультразвуком (Heat Systems Sonicator XL, Probe model CL, Farmingdale, NY) в течение 4×40-секундных импульсов при 100% интенсивности, с 1-минутным интервалом между импульсами. ЗФР-нерастворимый материал (который включает в себя уриказу) осаждали центрифугированием (10410 × g, 10 минут, 4°С) и затем промывали 5 раз 200 мл ЗФР. Уриказу в ЗФР-нерастворимом осадке экстрагировали в 80 мл 1 М Na2CO3, pH 10,2, содержащего 1 мМ фенилметилсульфонилфторид (ФМСФ) и 130 мкг/мл апротинина. Нерастворимый дебрис удаляли центрифугированием (20190 × g, 2 часа, 4°С). Все последующие стадии очистки проводили при 4°С (результаты суммированы в таблице 3).
Экстракт pH 10,2 разбавляли до 1800 мл 1 мМ ФМСФ (для снижения концентрации Na2СО3 до 0,075 М). Этот раствор наносили на колонку (2,6×9 см) свежеприготовленной Q-Sepharose (Pharmacia Biotech, Inc., Piscataway, NJ), которая была уравновешена 0,075 М Na2СО3, pH 10,2. После нанесения, колонку промывали последовательно 1) 0,075 М Na2СО3, pH 10,2, пока поглощение A280 эффлюента не достигало фона; 2) 10 мМ NaHCO3, pH 8,5, пока pH эффлюента не падал до 8,5; 3) 50 мл 10 мМ NaHCO3, pH 8,5, 0,15 М NaCl; 4) 100 мл градиента 0,15 М NaCl - 1,5 М NaCl в 10 мМ NaHCO3, pH 8,5; 5) 150 мл 10 мМ NaHCO3, pH 8,5, 1,5 М NaCl; 6) 10 мМ NaHCO3, pH 8,5; 7) 0,1 М Na2CO3 , pH 11, пока рН эффлюента не повышался до 11. Наконец, уриказу элюировали 500 мл градиента от 0 до 0,6 М NaCl в 0,1 М Na2CO3, pH 11. Активность элюировалась в двух пиках абсорбции А280, которые собирали по отдельности (Фракция А и Фракция В, таблица 3). Затем уриказу в каждом из этих пулов осаждали снижением рН до 7,1 медленным добавлением 1 М уксусной кислоты с последующим центрифугированием (7000 × g, 10 минут). Полученный осадок растворяли в 50 мл 1 М Na2СО3, рН 10,2 и хранили при 4°С.
ПРИМЕР 3
Получение небольшой партии и ПЭГилирование рекомбинантной РВС-уриказы
Этот пример показывает, что очищенная рекомбинантная РВС-уриказа может быть использована для получения ПЭГилированной уриказы. В этой реакции все субъединицы уриказы были модифицированы (фигура 1, дорожка 7) с сохранением приблизительно 60% каталитической активности (таблица 4).
А. Низкомасштабные экспрессия и выделение РВС-уриказы (таблица 4, фигура 1)
Четырехлитровую культуру Е. coli BL21(DE3)pLysS, трансформированную кДНК рЕТ3d-РВС, инкубировали на ротационном шейкере (250 об/мин) при 37°С. При 0,7 OD525 культуру индуцировали IPTG (0,4 мМ, 6 часов). Клетки собирали и замораживали при -20° С. Эти клетки (15,3 г) разрушали замораживанием и оттаиванием и экстрагировали 1 М Na2СО3, рН 10,2, 1 мМ ФМСФ. После центрифугирования (12000 × g, 10 минут, 4°С) супернатант (85 мл) разбавляли 1:10 водой и затем хроматографировали на Q-сефарозе способом, сходным с описанным в примере 1. Объединенную уриказную активность из этой стадии концентрировали ультрафильтрацией под давлением с использованием мембраны РМЗО (Amicon, Beverly, MA). Концентрат хроматографировали на колонке (2,5×100 см) сефакрил S-200 (Pharmacia Biotech, Inc., Piscataway, NJ), которая была уравновешена и осуществляла разделение в 0,1 М Na2СО3, рН 10,2. Фракции, содержащие уриказную активность, объединяли и концентрировали ультрафильтрацией под давлением, как описано выше.
В. ПЭГилирование
100 мг концентрированной сефакрил S-200-РВС-уриказы (5 мг/мл, 2,9 мкмоль фермента; 84,1 мкмоль лизина) в 0,1 М Na2 CO3, pH 10,2 подвергали реакции с 2-кратным избытком (моль ПЭГ:моль лизинов уриказы) активированной формы ПЭГ при 4°С в течение 60 минут. ПЭГилированную уриказу высвобождали из непрореагировавшего или гидролизованного ПЭГ тангенциальной проточной диафильграцией. В этой стадии реакцию разбавляли 1:10 в 0,1 М Na2СО3, pH 10,2 и диафиль тровали против 3,5 объемов 0,1 М Na2СО3, pH 10,2, затем против 3,5 объемов 0,05 М фосфата натрия, 0,15 М NaCl, pH 7,2. Стерилизованный фильтрованием фермент был стабильным при 4°С в течение по меньшей мере одного месяца.
Фигура 1 показывает анализ при помощи электрофореза в ПААГ с ДСН-меркаптоэтанолом (12% гель) фракций, полученных во время очистки и ПЭГилирования рекомбинантной химерной (свинья-павиан, РВС) уриказы. Дорожки: 1 = MW-маркеры; 2 = ДСН-экстракт неиндуцированных рЕТ3d-РВС-кДНК-трансформированных клеток (Е. coli BL21(DE3)pLysS); 3 = ДСН-экстракт IPTG-индуцированных рЕТ-РВС-кДНК-трансформированных клеток; 4 = неочищенный экстракт (см. таблицу 5); 5 = концентрированный пул Q-сефароз-уриказы; 6 = концентрированный пул сефакрил S-200-уриказы; 7 = ПЭГилированая сефакрил S-200-рекомбинантная РВС-уриказа.
Результаты, представленные в таблице 4, показывают, что очищенная РВС-уриказа могла быть модифицирована с сохранением приблизительно 60% каталитической активности. В этой реакции ПЭГилирования все субъединицы уриказы были модифицированы (фигура 1, дорожка 7). В непредставленных исследованиях, ПЭГилированный фермент имел одинаковые кинетические свойства с немодифицированной РВС-уриказой (Км 10-20 мкМ). Важно, что модифицированный фермент был гораздо более растворимым, чем немодифицированный фермент, при физиологическом рН (>5 мг/мл в ЗФР против <1 мг/мл). ПЭГилированный фермент мог быть также лиофилизирован и затем реконструирован в ЗФР, рН 7,2, с минимальной потерей активности. В других экспериментах, авторы сравнили активности этого препарата ПЭГ-РВС-уриказы с клиническим препаратом уриказы A. flavus. При рН 8,6 в боратном буфере, фермент A. flavus имел в 10-14 раз более высокую Vmax и в 2 раза более высокую Км. Однако, в ЗФР, рН 1,2, фермент ПЭГ-РВС и немодифицированный грибковый фермент отличались по уриказной активности менее чем в 2 раза.
ПРИМЕР 4
Время циркуляции немодифицированной и ПЭГилированной
РВС-уриказы у мышей
Фигура 2 показывает время циркуляции нативной и ПЭГилированной РВС-уриказы. Группы мышей (3 для каждой временной точки) инъецировали внутрибрюшинно (IP) 1 единицу нативной (кружки) или ПЭГ-модифицированной (квадраты) рекомбинантной РВС-уриказы (препаратом, описанным в примере 3). В указанных временных точках кровь получали из групп из трех мышей для измерения уриказной активности в сыворотке. ПЭГилированная уриказа (описанная в примере 3) имела период полужизни циркуляции приблизительно 48 часов, против <2 часов для немодифицированного фермента (фиг.2).
ПРИМЕР 5
Эффективность ПЭГилированной уриказы изобретения Фигура 3 показывает связь уриказной активности в сыворотке с концентрацией мочевой кислоты в сыворотке и в моче. В этом эксперименте, гомозиготная уриказа-недостаточная мышь с нокаутом гена (Wu et al. 1994) получала две инъекции, при 0 и 72 часах, 0,4 ME рекомбинантной РВС-уриказы, которая была ПЭГилирована. Уриказа-недостаточную мышь с нокаутом гена использовали в этих экспериментах потому, что, в отличие от нормальных мышей, которые имеют уриказу, эти мыши с генным нокаутом, как и люди, имеют высокие уровни мочевой кислоты в их крови и жидкостях тела и выделяют высокие уровни мочевой кислоты в их моче. Эти высокие уровни мочевой кислоты вызывают серьезное поврежедение почек этих мышей, которое часто является смертельным (Wu et al. 1994).
Эксперимент, показанный на фигуре 3, демонстрирует, что внутрибрюшинные инъекции ПЭГилированного препарата рекомбинантной РВС-уриказы приводили к увеличению уриказной активности в сыворотке, которое сопровождалось заметным снижением концентраций мочевой кислоты в сыворотке и в моче в уриказа-недостаточной мыши.
ПРИМЕР 6
Отсутствие иммуногенности комплекса конструкция-носитель
ПЭГилированную рекомбинантную РВС-уриказу инъецировали повторяемым образом в гомозиготных уриказа-недостаточных мышей без индукции ускоренного клиренса, в соответствии с отсутствием значимой иммуногенности. Это подтверждали при помощи ELISA. Фигура 4 показывает сохранение уровней циркуляции уриказной активности (измеренной в сыворотке) после повторяемой инъекции. ПЭГилированную РВС-уриказу вводили внутрибрюшинной инъекцией с интервалами 6-10 дней. Уриказную активность в сыворотке определяли через 24 часа после инъекции.
ПРИМЕР 7
Ковалентное связывание с мутационно введенным лизином
ПЭГилирование очищенной рекомбинантной РВС-уриказы должно приводить к присоединению ПЭГ к новому лизину (остатку 291). В этом эксперименте препарат РВС-уриказы мог быть модифицирован ПЭГилированием (присоединением ПЭГ). Способами, известными в данной области, можно определить, был ли пептид, содержащий новый лизин (остаток 291), модифицирован ПЭГилированием.
ССЫЛКИ
Abuchowski A, Kazo GM, Verhoest CR, Jr., van Es T, Kafkewitz D, Nucci ML, Viau AT et al (1984) Cancer therapy with modified enzymes. I. Antitumor properties of polyethylene glycol-asparaginase conjugates. Cancer Biochern Biophys 7:175-186.
Abuchowski A, McCoy JR, Palczuk NC, van Es T, Davis FF (1977a) Effect of attachment of polyethylene glycol on immunogenicity and circulating life of bovine liver catalase. J Biol Chem 252:3582-3586.
Abuchowski A, van Es T, Palczuk NC, Davis FF (1977b) Alteration of immunological properties of bovine serum albumin by covalent attachment of polyethylene glycol. J Biol Chem 252:3578-3581.
Ausubel FM (1998) Current Protocols in Molecular Biology John Wiley & Sons.
Ahn KJ, Kim YS, Lee HC, Park K, Huh KB (1992) Cyclosporine-induced hyperuricemia after renal transplant: Clinical characteristics and mechanisms. Transplantation Proceedings 24:1391-1392.
Arellano F, Sacristan JA (1993) Allopurinol hypersensitivity syndrome: A review. Ann Pharmacother 27:337-343.
Becker MA (1988) Clinical aspects of monosodium urate monohydrate crystal deposition disease (gout). Rheumatic Disease Clinics of North America 14:377-394.
Becker MA, Roessler BJ (1995) Hyperuricemia and gout. In: Scriver CR, Beaudet AL, Sly WS, Valle D (eds) The Metabolic and Molecular Bases of Inherited Disease, 7 th ed. McGraw-Hill, New York, pp 1655-1677.
Brogard JM, Coumaros D. Frankhauser J, Stahl A, Stahl J (1972) Enzymatic uricolysis: A study of the effect of a fungal urate-oxydase. Eur J Clin Biol Res 17:890-895.
Brogard JM, Stahl A, Stahl J (1978) Enzymatic uricolysis and its use in therapy. In: Kelley WN, Arnold WJ, Weiner IM (eds) Uric Acid, Springer-Verlag, New Yolk, pp 515-524.
Chaffee S, Mary A, Stiehm ER, Girault D, Fischer A, Hershfield MS (1992) IgG. antibody response to polyethylene glycol-modified adenosine deaminase (PEG-ADA) in patients with adenosine deaminase deficiency. J Clin Invest 89:1643-1651.
Chen RHL, Abuchowski A, van Es T, Palczuk NC, Davis FF (1981) Properties of two urate oxidases modified by the covalent attachment of poly(ethylene glycol). Biochim Biophys Acia 660:293-298.
Chua CC, Greenberg ML, Viau AT, Nucci M, Brenckman WD, Jr., Hershfield MS (1988) Use of polyethylene glycol-modified unease (PEG-uricase) to treat hyperuricemia in a patient with non-Hodgkin lymphoma. Ann Int Med 109:114-117.
Cohen LF, Balow JE, Magrath IT, Poplack DG, Ziegler JL (1980) Acute tumor lysis syndrome: A review of 37 patients with Burkitt's lymphoma. Am J Med 64:468-491.
Conley TG, Priest DG (1979) Purification of uricase from mammalian tissue. Preparative Biochemistry 9:197-203.
Davis FF, Kazo GM, Nucci ML, Abuchowski A (1991) Reduction of immunogenicity and extension of circulating life of peptides and proteins. In: Lee VHL (eds) Peptide and Protein Drug Delivery, Marcel Dekkar, New York, pp 831-864.
Davis S, Abuchowski A, Park YK, Davis FF (1981a) Alteration of the circulating life and antigenic properties of bovine adenosine deaminase in mice by attachment of polyethylene glycol. Clin Exp Immunol 46:649-652.
Davis S, Park YK, Abuchowski A, Davis FF (1981b) Hypouricaemic effect of polyethylene glycol modified urate oxidase. Lancet 1:281-283.
Delaney V, Sumrani N, Daskalakis P, Hong JH, Sommer BG (1992) Hyperuricemia and gout in renal allograft recipients. Transplantation Proceedings 24:1773-1774.
Donadio O, Errera. J, Navarro M, Izarn P (1981) Anaphylaxis-like manifestations after intravenous injection of urate oxidase in an asthmatic child with acute leukemia (letter). Nouv Presse Med 10:711-712.
Erlich HA (1989) PCR Technology. Principles and applications for DNA amplification Stockton Press, New York.
Escudier B, Leclercq B, Tandonnet F, Nitenberg G (1984) Hyperuricemia resistant to urate oxidase. Efficacy of high doses (letter). Presse Med 13:1340.
Fam AG (1990) Strategies and controversies in the treatment of gout and hyperuricaemia. Balliere's Clinical Rheumatology 4:177-192.
George T, Mandell BF (1995) Gout in the transplant patient J Clin Rheumatol 1:328-334.
Gold GL, Fritz BD (1957) Hyperuricemia associated with the treatment of leukemia. Ann Int Med 47:428-434.
Greenberg ML, Hershfield MS (1989) A radiochemical-high-performance liquid chromatographic assay for urate oxidase in human plasma. Anal Biochem 176:290-293.
Harris JM, Zalipsky S (Ed.) (1997) Poly(ethylene glycol) Chemistry and Biological Applications ACS, Washington, DC.
Hershfield MS. (1997) Biochemistry and immunology of poly(ethylene glycol)-modified adenosine deaminase (PEG-ADA). In: Harris JM, Zalipsky S (eds) Poly(ethylene glycol) Chemistry and Biological Applications, ACS, Washington, DC, pp 145-154.
Hershfield MS (1995) PEG-ADA replacement therapy for adenosine deaminase deficiency: An update after 8.5 years. Clin Immunol Immunopathol 76:S228-S232.
Hershfield MS (1996) Gout and uric acid metabolism. In: Bennett JC, Plum F (eds) Cecil Textbook of Medicine, XX ed. WB Saunders, New York, pp 1508-1515.
Hershfield MS, Buckley RH, Greenberg ML. Melton AL, Schiff R, Hatem C, Kurtzberg J et al (1987) Treatment of adenosine desminase deficiency with polyethylene glycol-modified adenosine deaminase. N Engl J Med 316:589-596.
Hershfield MS, Chaffee S. Koro-Johnscn L. Mary A, Smith AA, Short SA (1991) Use of site-directed mutagenesis to enhance the epitope shielding effect of covalent modification of proteins with polyethylene glycol. Proc Nail Acad Sci USA 88:7185-7189.
Hershfield MS, Mitchell BS (1995) Immunodeficiency diseases caused by adenosine deaminase deficiency and purine nucleoside phosphorylase deficiency. In: Scriver CR, Beaudet AL, Sly WS, Valle D (eds) The Metabolic and Molecular Bases of Inherited Disease. 7 th ed. McGraw-Hill, New York. ppl725-1768.
Hershfield MS, Seegmiller JE (1976) Gout and the regulation of purine biosynthesis. In: Quagliariello E (eds) Horizons in Biochemistry and Biophysics, Addison-Wesley, Reading, MA, pp 134-162.
Jones DP, Stapleton FB, Kalwinsky D, McKay CP, Kellie SJ, Pui CH (1990) Renal dysfunction and hyperuricemia at presentation and relapse of acute lymphoblastic leukemia. Med Pediatr Oncol 18:283-286.
Kelley WN, Fox IH, Pallela TD (1989) Gout and related disorders of purine metabolism. In: Kelley WN, Harris ED, Ruddy S, Sledge CO (eds) Textbook of Rheumatology. 3rd ed. WB Saunders, Philadelphia, pp 1395-1448.
Kissel P, Lamarche M, Royer R (1968) Modification of uricaemia and the excretion of uric acid nitrogen by an enzyme of fungal origin. Nature 217:72-74.
Kissel P, Schmitt J, Streiff F, Makuary G, Schmidt C, Toussain P (1972) L'urate oxydase: son intéret dans la prévention des hyperuricemies therapeutiques en hematologie. Ann Med Nancy 11:519-535.
Lee CC, Wu X, Gibbs RA, Cook RG, Muzny DM, Caskey CT (1988) Generation of cDNA directed by amino acid sequence: Cloning of urate oxidase. Science 239:1288-1291.
Legoux R, Delpech B, Dumont X, Guillemot JC, Ramond P, Shire D, Caput D et al (1992) Cloning and expression in Escherichia coli of the gene encoding Aspergillus flavus urate oxidase. J Biol Chem 267:8565-8570.
London M, Hudson PM (1957) Uricolytic activity of purified uricase in two human beings. Science 125:937-938.
Masera G, Jankovic M, Zurlo MG, Locasciulli A, Rossi MR, Uderzo C, Recchia M (1982) Urate-oxidase prophylaxis of uric acid-induced renal damage in childhood leukemia. J Pediatr 100:152-155.
Montagnac R, Schillinger F (1990) Anaphylactic complication tied to intravenous injection of urate oxidase. Nephrologie 11:259.
Mourad G, Cristol JP, Chong G, Andary M, Mion С (1984) Role of precipitating anti-urate oxidase antibodies in unite oxidase-resistant hyperuricemia (letter). Presse Med 13:2585.
Potaux L, Aparicio M, Maurel C, Ruedas ME, Martin-Dupont CL (1975) Uricolytic therapy. Value of urate oxidase in the treatment of hyperuricemias. Nouv Presse Med 4:1109-1112.
Priest DG, Pins OM (1972) Reaction intermediate effects on (he spectrophotometric uricase assay. Analytical Biochemistry 50:195-205.
Put C-H, Relling MV, Lascombes F, Harrison PL, Scruxiano A, Mondesir J-M, Riberio RC et al (1997) Urate oxidase in prevention and treatment of hyperuricemia associated with lymphoid malignancies. Lsukemia 11:1813-1816.
Reddy PG, Nemali MR, Reddy MK, Reddy MN, Yuan PM, Yuen S, Laffler TG et al (1988) Isolation and sequence determination of a cDNA clone for rat peroxisomal urate oxidase: Liver-specific expression in the rat. Proc Nad Acad Sci USA 85:9081-9085.
Rosenthal AK, Ryan LM (1995) Treatment of refractory crystal-associated arthritis. Rheum Dis Clin North Amer 21:151-161.
Roubenoff R (1990) Gout and hyperuricemia. Rheumatic Disease Clinics of North America 16:539-550.
Sadler JR, Miwa J, Maas P, Smith Т (1974) growth of high density bacterial cultures; a simple device. Laboratory Practice 23:632-643.
Sambrook J, Fritsch EF, Maniatis Т (1989) Molecular cloning. A laboratory manual 2 nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, Pages.
Sandberg AA, Cartwright GB, Wintrobe MM (1956) Studies on leukemia. I. Uric acid excretion. Blood 11:154-166.
Savoca KV, Davis FF, Palczuk NC (1984) Induction of tolerance in mice by uricase and monomethoxypolyethylene glycol-modified uricase. Int Arch Allergy Appl Immunol 75:58-67.
Sibony G, North ML, Bergerat IP, Lang JM, Oberling F (1984) Hyperuricemia resistant to urare oxidase. Role of anti-serum urate oxidase precipitating antibodies (letter). Presse Med 13:443.
Singer JZ, Wallace SL (1986) The allopurinol hypersensitivity syndrome. Unnecessary morbidity and mortality. Arthritis Rheum 29:32-87.
Tsuji J, Hirose K, Kasahara E, Naitoh M, Yamamoto I (1985) Studies on the antigenicity of the polyethylene glycol-modified uricase. Int J Immunopharmacol 7:725-730.
Venkataseshan VS, Feingold R, Dikman S, Churg J (1990) Acute hyperuricemic nephropathy and renal failure after transplantation. Nephron 56:317-321.
Veronese FM, Caliceti P, Schiavon O (1997) New synthetic polymers for enzyme and liposome modification. In: Harris JM, Zalipsky S (eds) Poly(ethylene glycol) Chemistry and Biological Applications, ACS, Washington, DC, pp.182-192.
West C, Carpenter BJ, Hakala TR (1987) The incidence of gout in renal transplant recipients. Am J Kidney Dis 10:369-371.
Wu X, Lee CC, Muzny DM, Caskey CT (1989) Urate oxidase: Primary structure and evolutionary implications. Proc Natl Acad Sci USA 86:9412-9416.
Wu X, Muzny DM, Lee CC, Caskey CT (1992) Two independent mutational events in the loss of urate oxidase. J Mol Evol 34:78-84.
Wu X, Wakamiya M, Vaishnav S, Geske R, Montgomery CM, Jr., Jones P, Bradley A et al (1994) Hyperuricemia and urate nephropathy in urate oxidase-deficient mice. Proc Natl Acad Sci USA 91:742-746.
Zittoun R, Dauchy F, Teillaud C, Barthelemy M, Bouchard P (1976) Le traitement des hyperuricemies en hematologie par l'urate-oxydase et l'allopurinol. Ann Med Interne 127:479-482.
Все цитированные выше документы включены здесь в качестве ссылки в их полном виде.

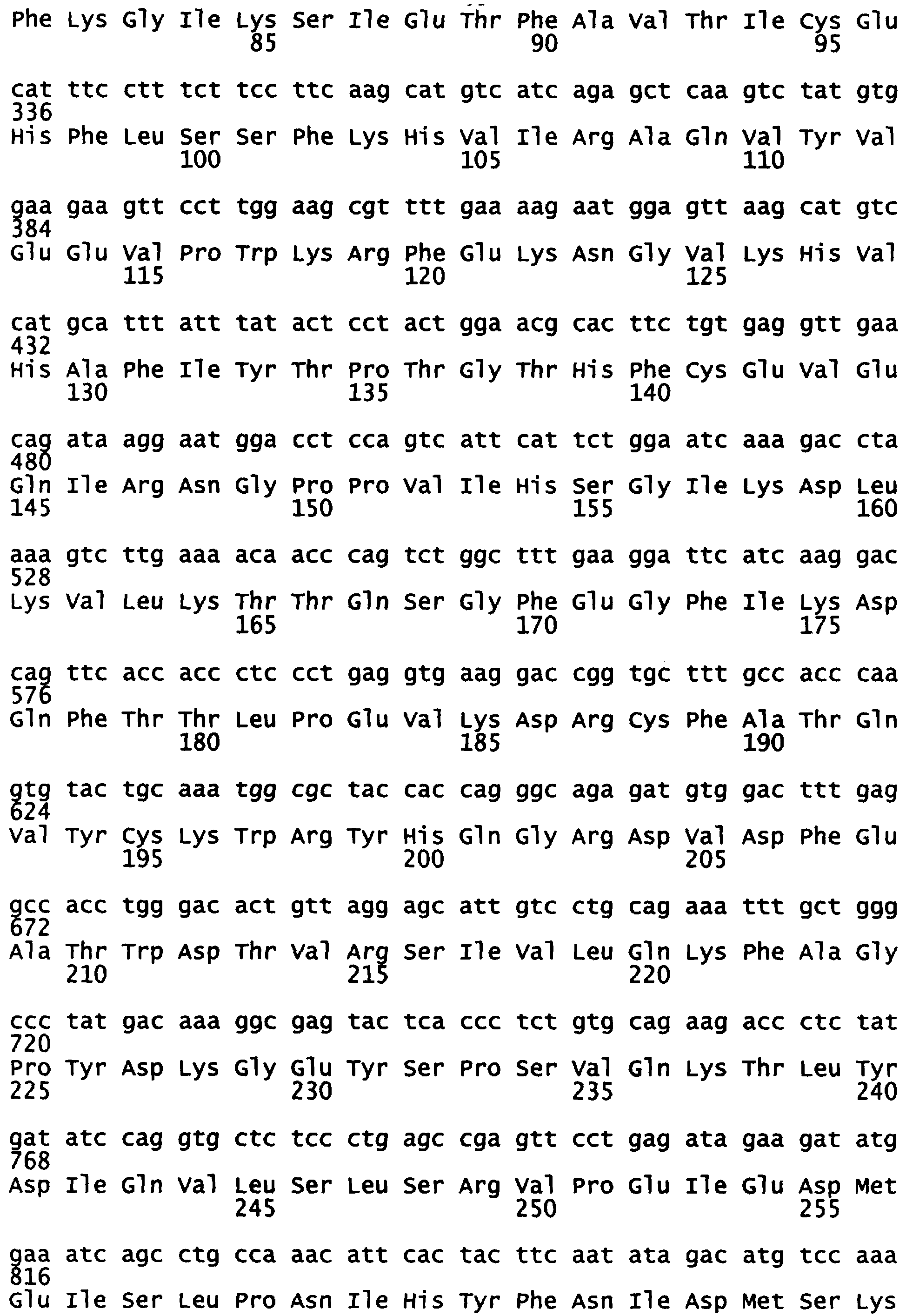


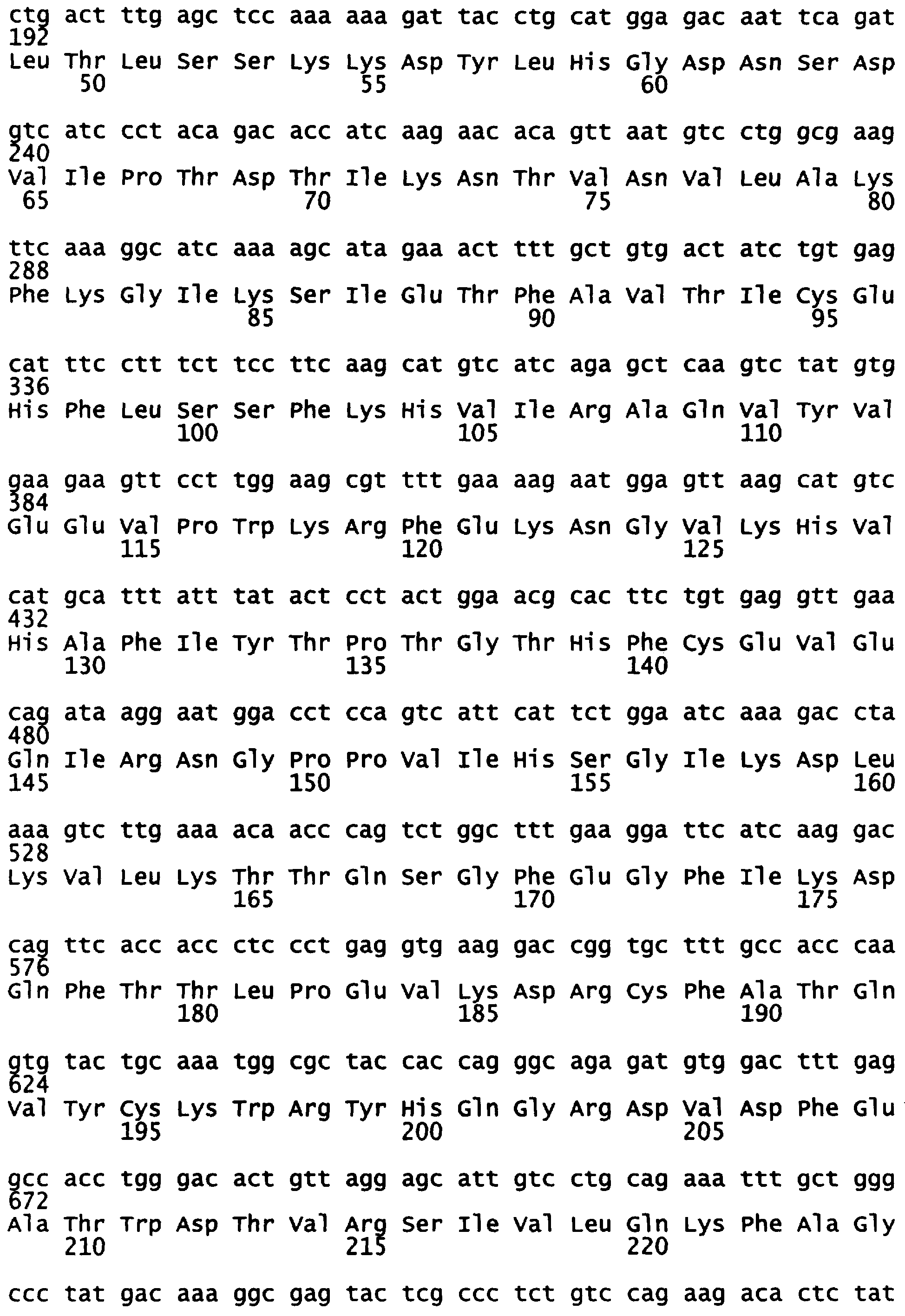


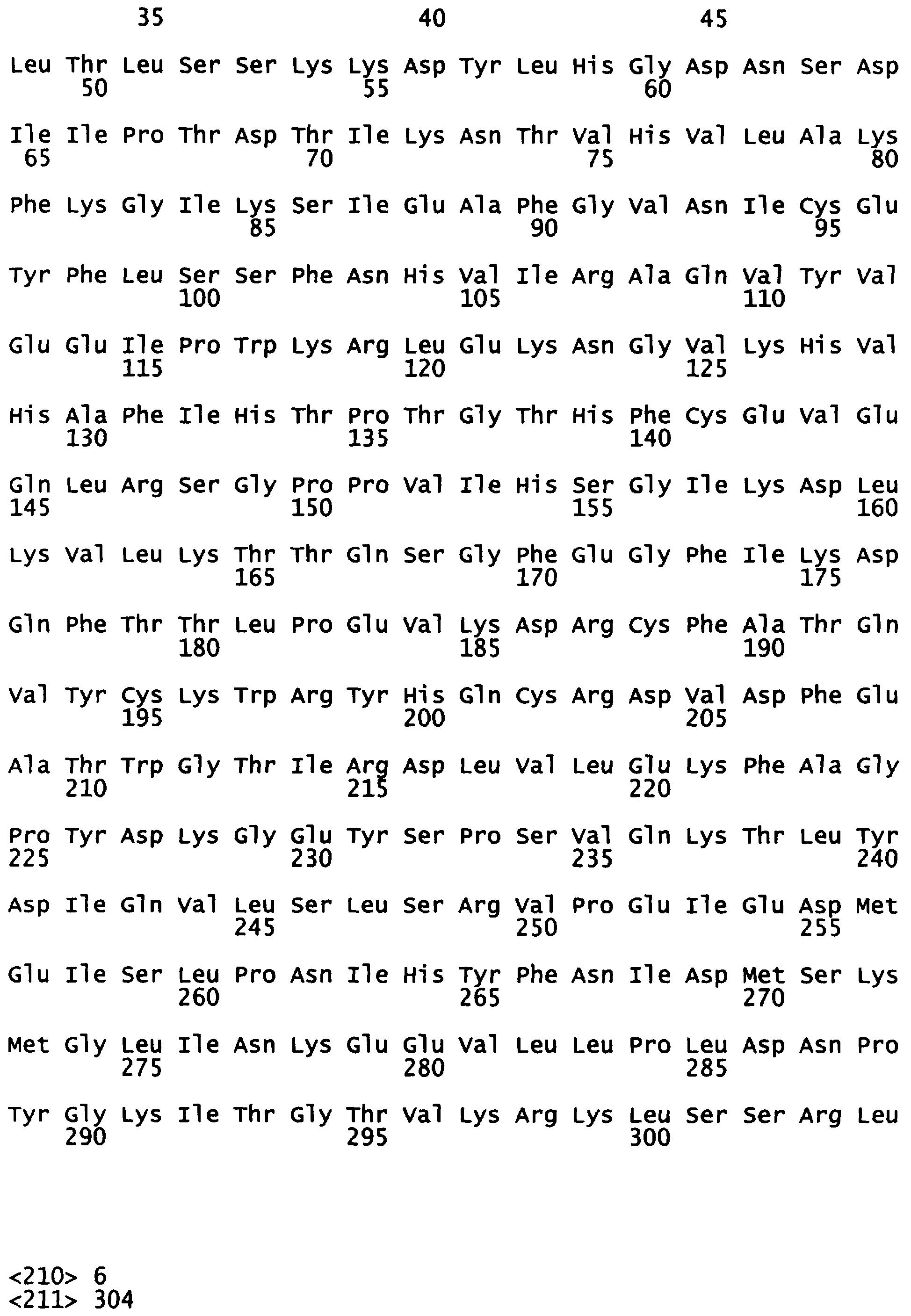
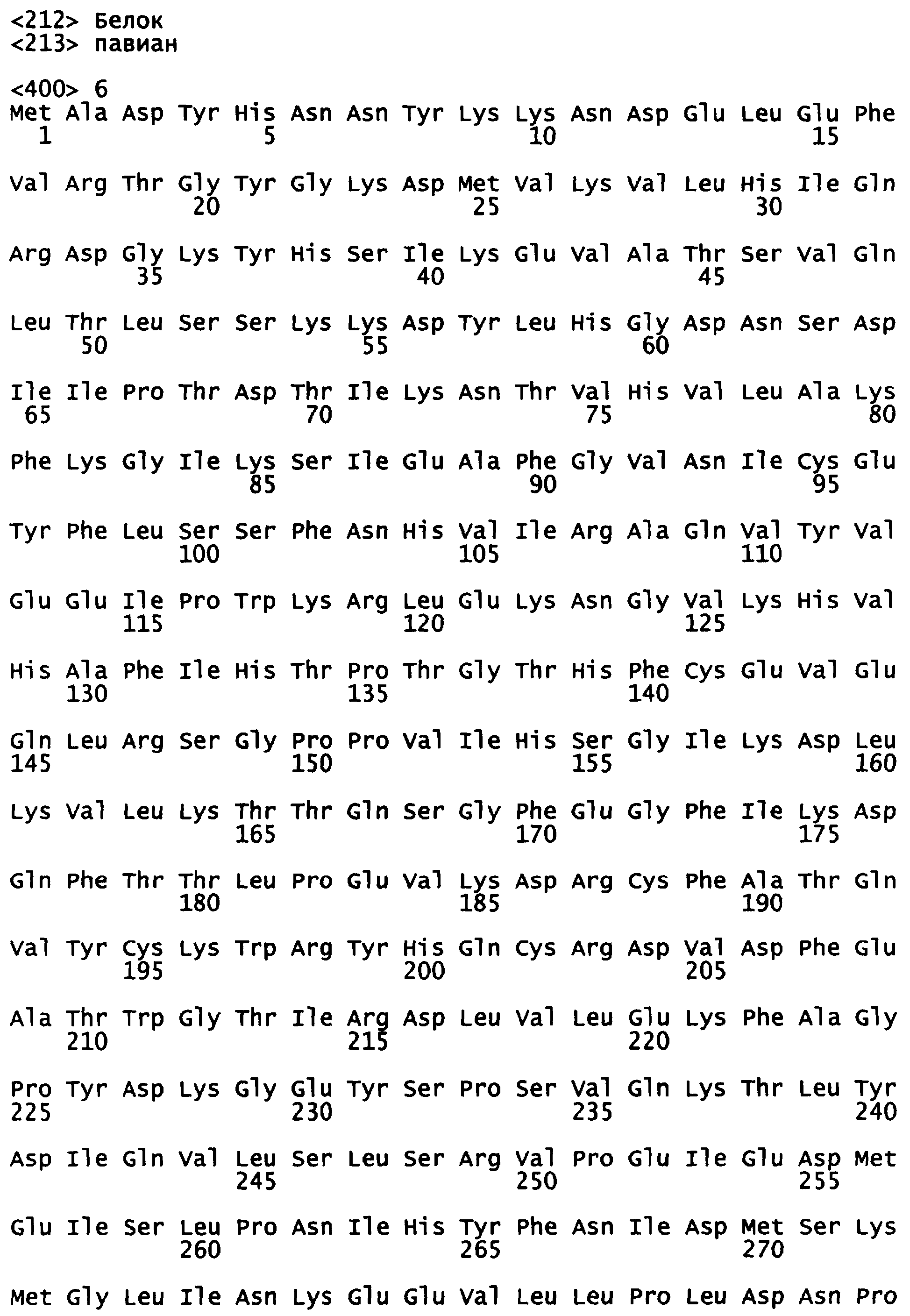



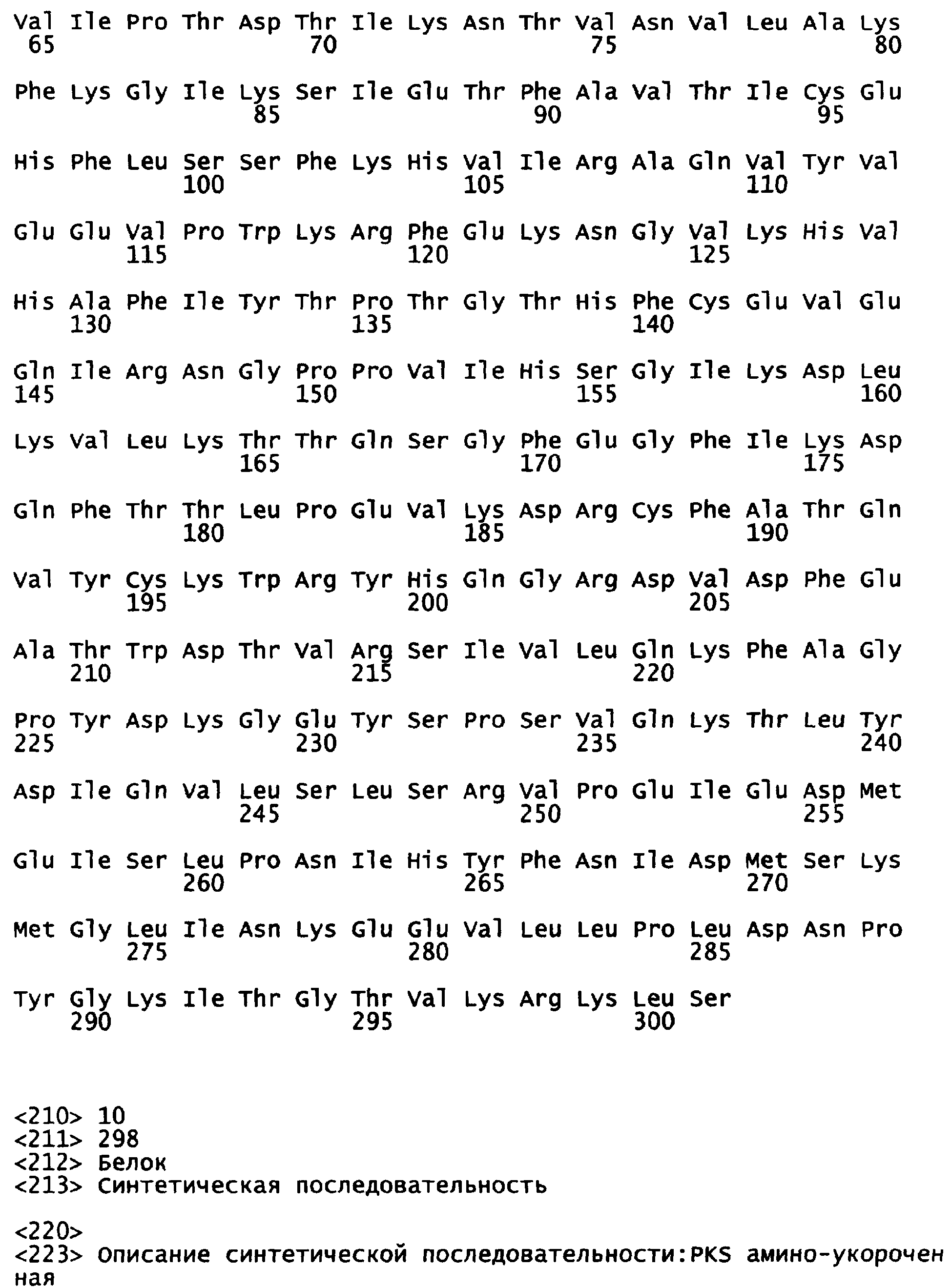
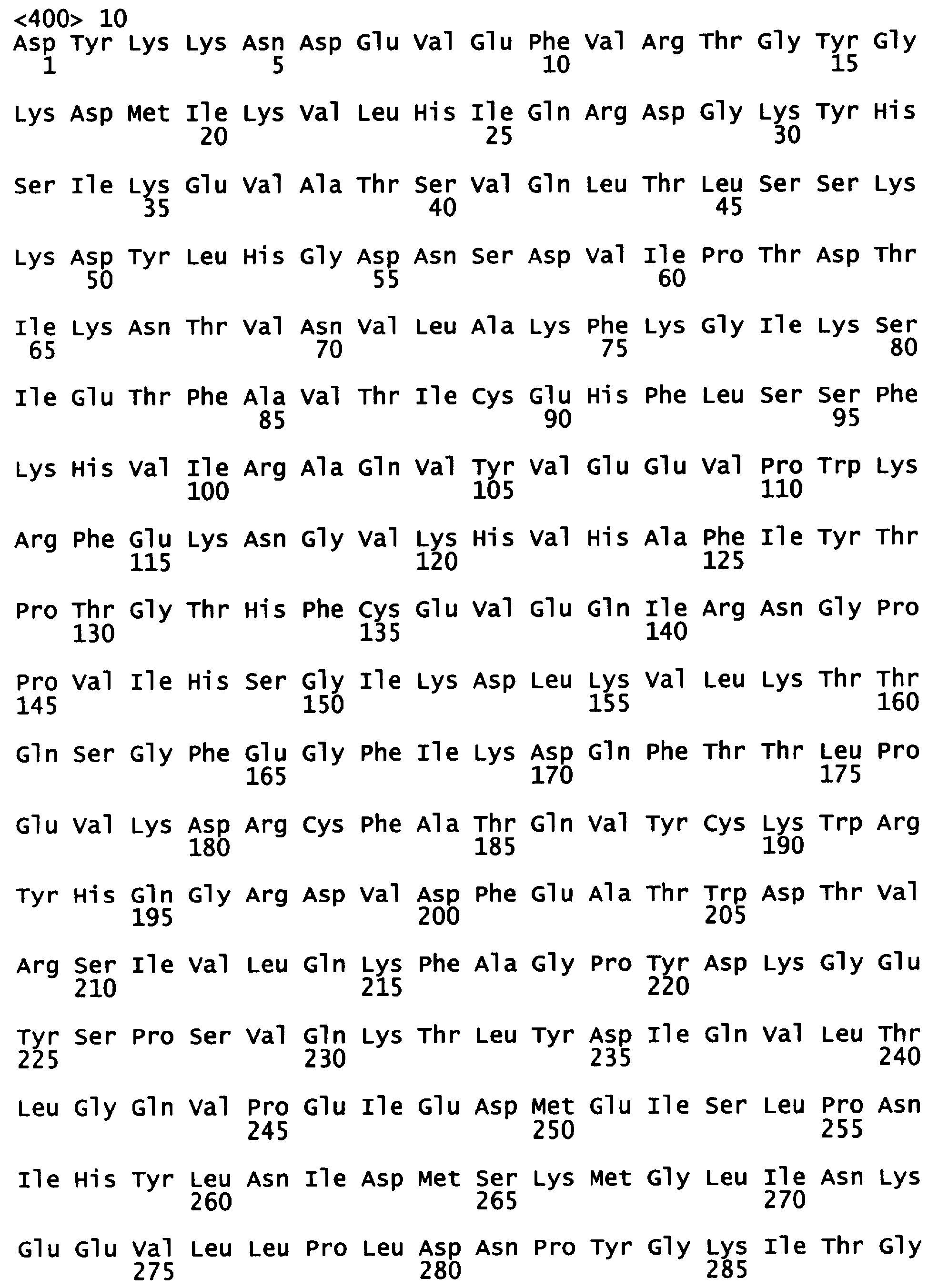


Реферат
Изобретение относится к области биотехнологии, в частности генетической инженерии, и может быть использовано в медицине. В результате увеличения числа доступных сайтов связывания ПЭГ в молекуле уриказы млекопитающего, достигаеиого путем замены, по меньшей мере, одного фрагмента аминокислотной последовательности природного фермента млекопитающего, не содержащего остатка лизина, гомологичным фрагментом уриказы млекопитающего другого вида, содержащим остаток лизина, и скрининга вариантов уриказы по показателям активности и иммуногенности после конъюгирования с ПЭГ получены новые химерные ферментные белки, в частности формы, состоящие из фрагментов уриказы свиньи и уриказы павиана (РВС и PKS). Предлагаемые варианты уриказы отличаются тем, что после ПЭГ - илирования сохраняют по существу такую же уриколитическую активность, что и немодифицированный фермент, и являются по существу неиммуногенными. Описаны способы получения новых химерных белков уриказы с помощью технологии рекомбинантных ДНК и необходимые для их осуществления векторные конструкции и системы экспрессии. Применение изобретения обеспечивает получение промежуточных продуктов для производства медицинских препаратов с низкой иммуногенностью и повышенной биодоступностью. 5 с. и 11 з.п. ф-лы, 4 табл., 14 ил.
Комментарии