Получение терминально дифференцированных дофаминергических нейронов из эмбриональных стволовых клеток человека - RU2345133C2
Код документа: RU2345133C2
Чертежи





Описание
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
1. ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к улучшенному способу получения терминально дифференцированных нейрональных клеток, таких как дофаминергические или серотонинергические нейроны, из плюрипотентных эмбриональных стволовых клеток, таких как человеческие эмбриональные стволовые клетки. Дофаминергические и серотонинергические нейроны, полученные в соответствии с настоящим изобретением, могут служить в качестве превосходного источника для клеточно-заместительной терапии при нейродегенеративных нарушениях и нейрональных заболеваний.
2. ОПИСАНИЕ ПРЕДШЕСТВУЮЩЕГО УРОВНЯ ТЕХНИКИ
Нейродегенеративные нарушения и неврологические заболевания, такие как болезнь Паркинсона, болезнь Альцгеймера и шизофрения, являются деструктивными заболеваниями, которые становятся все более распространенными в нашем обществе. Многие из этих неврологических заболеваний связаны с дофаминергическими или серотонинергическими нейронами. Дофаминергические нейроны находятся в вентральной или вентролатеральной области среднего мозга и контролируют постуральные рефлексы, движение и вознаграждение, ассоциированное с поведением. Эти нейроны иннервируют различные структуры переднего мозга, и их разрушение или анормальная функция связаны с болезнью Паркинсона, шизофренией или склонностью к наркотикам (Hynes et al., 1995, Cell 80:95-101). Серотонинергические нейроны концентрируются в вентральной или вентролатеральной области заднего мозга и иннервируют различные части центральной нервной системы, включая кору головного мозга, лимбическую систему и спинной мозг. Эти нейроны контролируют уровень сознания, пробуждения, особенностей поведения, потребление пищи, и их анормальная функция связана с агрессией, депрессией и шизофренией (Jacobs и Gelperin, 1981, Serotonin Neurotransmission and Behavior. The MIT Press, Cambridge, Mass.). Серотонинергическая дисфункция также может играть роль в патофизиологии различных психиатрических, неврологических и других заболеваний, например, умственной депрессии (Asberg et al., 1986, J. Clin. Psychiatry 47:23-35), суицида (Lester, 1995, Pharmocopsychiatry 28(2):45-50) и вспыльчивого агрессивного поведения (Brown et al., J. Clin. Psychiatry, 1990, 54:31-41; Eichelman, 1990, Annu. Rev. Med. 41:149-158).
Болезнь Паркинсона является прогрессирующим неврологическим заболеванием, вызванным разрушением нервных клеток (нейронов) в области мозга, которая контролирует движения. Это разрушение вызывает недостаток сигнального агента (нейромедиатор) в мозге, известного как дофамин, что вызывает нарушение двигательной активности, которое характеризует это заболевание. Исследования патологии показывают, что потеря дофаминергических нейронов в черном веществе способствуют возникновению болезни Паркинсона. Например, билатеральное поражение нигростриатального пути (эфферентной связи черного вещества с полосатым телом) приводит у экспериментальных животных к синдрому, очень сходному с наблюдаемой моторной дисфункцией при болезни Паркинсона: тремор в покое, ригидность, акинез и постуральные отклонения. Следствием билатерального повреждения нигростриатального пути, вызванные 6-гидроксидопамином (OHDA), является значительная акинезия, адипсия, афагия и сенсорная нечувствительность у грызунов (Ungerstedt, 1971, U. Acta Physiol. Scand. Suppl. 367:95-121; Yirek and Sladek, 1990, Annu. Rev. Neurosci. 13:415-440).
При паркинсонизме изменение статуса дофаминергических рецепторов может зависеть от стадии прогрессирования заболевания. Признаком паркинсонизма является значительное уменьшение дофамина во всех компонентах базального ганглия (Hornykiewicz, 1988, Mt. Sinai J. Med. 55:11-20). Когда дофамин исчерпан, возникает дисфункция различных других областей мозга, таких как таламус, бледный шар и субталамическое ядро. Поскольку эти области посылают сигналы в другие области мозга, дисфункция этой небольшой области может привести к обширному мозговому нарушению.
Распространение болезни Паркинсона широко варьирует от 82 больных на 100000 человек в Японии и 108 больных на 100000 человек в Великобритании до практически 1% (предположительно 1 миллион) населения Северной Америки. В Индии величина распространения болезни Паркинсона составляет 14 больных на 100000 человек в Северной Индии, 27 больных на 100000 человек в Южной Индии, 16 больных на 100000 человек в Восточной Индии и 363 больных на 100000 человек в общине Парси в Западной Индии. В настоящий момент болезнь Паркинсона считается неизлечимой, но доступен широкий набор медикаментов, которые облегчают симптоматику болезни Паркинсона, к ним относятся леводопа, бромокриптин, перголид, селегилин, антихолинергик и амантадин. Хотя эти лекарства могут облегчать симптомы болезни Паркинсона, зачастую они обладают побочными эффектами. Более того, эти лекарства не оказывают влияния ни на саму болезнь, ни на разрушение нейронов, а только ослабляют симптомы, при этом зачастую со временем положительный эффект ослабляется. Некоторые пациенты становятся менее чувствительными к препаратам, тогда как другие становятся гиперчувствительными и у них развивается дискинезия.
Такие неудовлетворительные результаты привели к разработке других стратегий для лечения этого заболевания, таких как терапия агонистами дофарецепторов и хирургические подходы, которые включают паллидотомию, глубокую стимуляцию мозга (ГСМ) в районе бледного шара и попытки прервать сетевые аномалии с помощью разрушения гиперактивных областей мозга или размещением ГСМ электродов для успокоения этой области. Хотя эти и другие виды хирургии для пациентов с болезнью Паркинсона привели к некоторым положительным результатам, долгосрочные эффекты таких хирургических операций пока не известны. Эти подходы к лечению также имеют некоторые ограничения и побочные эффекты.
Другой стратегией, которая в настоящий момент рассматривается для этого неизлечимого заболевания, является генная терапия. Открытие молекулярной основы неврологического заболевания и достижения в системах переноса генов, позволили целенаправленно и глобально доставлять терапевтические гены при большом количестве расстройств центральной нервной системы. Но генная терапия имеет определенные ограничения, такие как стабильность и регуляция трансгенной экспрессии, и безопасность как вектора, так и экспрессируемых трансгенов (Costantini et al., 2000, Gene Therapy 7:93-109). Векторы, обычно используемые для генной терапии, включают, но ими не ограничиваются вирусом простого герпеса 1 типа (HSV-1) (During et al., 1994, Science 266:1399-1403), аденоассоциированный вирусный вектор (AAV) (During et al., 1998, Gene Therapy 5:820-827), ретровирусы, гибридный вектор HSV/вирус Эпштейна-Барра (HSV/EVB), и HSV/AAV гибридный вектор. Было показано, что подход лечения, связанный с одним геном, может быть эффективным на моделях болезни Паркинсона на животных. Инкапсулированная, генетически сгенерированная клеточная линия выделяла нейропротективную молекулу - ген глиального нейротрофического фактора (GDNF), и лентивирусный вектор, кодирующий ген GDNF, улучшал выживание трансплантата и дифференцировку, таким образом ускоряя восстановление поведения у модельных животных (Zurn et al., 2001, Brain Res Rev. 36:222-229; Date et al., 2001, Cell Transplant 10:397-401). Также было показано, что генная терапия с использованием нейрональных стволовых клеток также эффективна для экспрессии терапевтических доз GDNF in vivo (Akerud et al., 2001. J. Neurosci. 21:8108-8118).
Клеточная имплантация является другим подходом к лечению, который возможно поможет заменить нервные клетки, потерянные при болезни Паркинсона, а также других нейродегенеративных расстройств и нейрональных заболеваний. Благодаря клиническим исследованиям с трансплантацией эмбриональной ткани, находящихся все еще в процессе реализации, были разработаны методы имплантации клеток в мозге и показана возможность этого подхода, а также многообещающие результаты по крайней мере для некоторых пациентов. Также были предприняты некоторые попытки трансплантации предшественниц дофаминергических нервных клеток прямо в полосатое тело пациентов с болезнью Паркинсона, и было показано, что трансплантация человеческих эмбриональных или зародышевых дофаминергических нейронов приводило к положительному эффекту у пациентов с болезнь Паркинсона (Freed et al., 2001, N. Engl. J. Med. 344:710- 719). На основании этих данных было сделано предположение, что для полного восстановления необходима анатомическая замена путей, а не эктопическое помещение трансплантата (Winkler et al., 2000, Prog. Brain Res. 127:233-265). Кроме того, для трансплантации эмбрионального черного вещества и получения клинически достоверного улучшения у пациентов необходимы эмбриональные ткани человека по крайней мере от 5-10 утробных плодов, что связано с огромными этическими, юридическими проблемами и проблемами безопасности. Таким образом, в настоящее время остро необходимы альтернативные источники нейрональных клеток, таких как дофаминергических нейронов для лечения нейродегенеративных расстройств и нейрональных заболеваний.
Недавно было обнаружено, что в мозге взрослого человека имеется возобновляемый источник нервных стволовых клеток. Нервные стволовые клетки со способностью возобновлять сами себя и образовывать все типы клеток мозга являются потенциально неограниченным источником клеток мозга, продуцирующих дофамин, обеспечивая, такие образом, полностью новый терапевтический подход к нейродегенеративным расстройствам и неврологическим заболеваниям (Eriksson et al., 1998, Nature Medicine 4:1313-1317). Сообщалось, что культуры нервных стволовых клеток, полученных из переднего мозга человеческого эмбриона, могут быть увеличены до десяти миллионов раз in vitro. Эти взрослые нервные стволовые клетки были трансплантированы во взрослых крыс, обладающих ярко выраженной моделью болезни Паркинсона. Клетки в этих животных моделях выживали вплоть до года после трансплантации, дифференцировались в нейроны и были способны уменьшить нарушение моторики у некоторых экспериментальных животных (Svendsen et al., 1997, Exp. Neurol. 148:135-146). К сожалению, взрослые нервные стволовые клетки имеют ограниченную продолжительность жизни в культуре ткани (Kukekov et al., 1999, Exp. Neurol. 156:333-344).
Одним жизнеспособным альтернативным источником дофаминергических нейронов и других нейронов, которые могут быть использованы для лечения различных нейродегенеративных расстройств и нейрональных заболеваний, являются плюрипотентные эмбриональные стволовые (ЭС) клетки, в особенности человеческие ЭС клетки. ЭС клетки могут пролиферировать бесконечно в недифференцированном состоянии и являются плюрипотентными, что означает, что они способны дифференцироваться практически во все типы клеток, присутствующих в организме. Так как ЭС клетки способны становиться практически всеми специализированными клетками тела, они вероятно могут использоваться для получения замещающих клеток для большого множества тканей и органов, таких как сердце, поджелудочная железа, нервная ткань, мускулы, хрящ и тому подобное. ЭС клетки могут быть получены из внутренней клеточной массы (ВКМ) бластоциста, являющегося стадией эмбрионального развития, которая происходит до имплантации. Человеческие ЭС клетки могут быть получены из человеческого бластоциста на ранних стадиях эмбрионального развития, длящегося от 4- до 7-го дня после оплодотворения. ЭС клетки, полученные из ВКМ, можно культивировать in vitro и при подходящих условиях пролиферировать бесконечно.
ЭС клеточная линия была успешно получена для некоторых видов, включая мышей (Evans et al., 1981, Nature 292:154-156), крыс (Iannaccone et al., 1994, Dev. Biol., 163:288-292), свиней (Evans et al., 1990, Theriogenology 33:125-128; Notarianni et al., 1990, J. Reprod. Fertil. Suppl. 41:51-6), овец и коз (Meinecke-Tillmann и Meinecke, 1996, J. Animal Breeding and Genetics 113:413-426; Notarianni et al., 1991, J. Reprod. Fertil. Suppl. 43:255-60), кроликов (Giles et al., 1993, Mol. Reprod. Dev. 36:130-138; Graves et al., 1993, Mol. Reprod. Dev. 36:424-433), норок (Sukoyan et al., Mol. Reprod. Dev. 1992, 33:418-431), хомяков (Doetschman et al., 1988, Dev. Biol. 127:224-227), домашних птиц (Pain et al., 1996, Development 122(8):2339-48), приматов (патент США № 5843780) и человека (Thomson et al., 1998, Science 282:1145-1147; Reubinoff et al., 2000, Nature Biotech. 18:399-403). Как и другие ЭС клетки млекопитающих, человеческие ЭС клетки дифференцируются и образуют ткани всех трех зародышевых листков при введении иммунодефицитным мышам, обеспечивая их плюрипотентность. Опубликованные данные показывают, что человеческие ЭС клетки, поддерживаемые в культуре в течение более одного года, сохраняли свою плюрипотентность, способность самовозобновляться и нормальный кариотип (Thomson et. al., 1995, PNAS 92:7844-7848).
Исследования показали, что ЭС клетки могут дифференцироваться в нейрональные клетки-предшественницы (Zhang et al., 2001, Nature Biotech. 19:1129-33; WO 01/88104; США № 09/872,183, 09/888,309, 10/157,288; WO 03/000868; каждый конкретно приведенный здесь в качестве ссылки). Эти клетки затем могут дополнительно дифференцироваться в дофаминергические нейроны (Rolletschek et al., 2001, Mech. Dev. 105:93-104). Первоначальным шагом в дифференцировке ЭС клеток может быть образование эмбриоидных тел, например, 1 мкМ ретиноидной кислоты способствует нейрональной дифференцировке в эмбриоидные тела (Bain et al., 1995, Dev. Biol. 168:342-357). Тогда как ретиноидная кислота может быть использована для получения нервных клеток, ретиноидная кислота является сильным тератогеном. В некоторых отчетах была опубликована информация о том, что ЭС клетки могут дифференцироваться в дофаминергические нейроны благодаря индуцирующей активности стромальных клеток (SIDA) (Kawasaki et al., 2000, Neuron 28:1-20), путем экспрессии гена ядерного рецептора 1 (Nurr-1) (Kim et al., 2002, Nature 418:50-56) или путем трансплантации недифференцированных ЭС клеток непосредственно в мышиную модель (Bjorklund et al., 2002, Proc. Natl Acad. Sci. 99:2344-2349). Lee et al. (2000, Nat. Biotechnol. 18:675-79) сообщил о методе дифференцировки ЭС клеток в клетки-предшественницы нейронов и в дофаминергические и серотонинергические нейроны in vitro. Все эти эксперименты, однако, были проведены с использованием ЭС клеток мышей, и в протоколах выход дифференцировки дофаминергических нейронов изменялся в пределах от 5 до 50%. Около 20% ЭС клеток мышей развивались в дофаминергические нейроны в исследовании Lee et al. (WO 01/83715) и 5-50% в исследовании Studer et al. (WO 02/086073). Были получены данные о том, что несмотря на то, что дофаминергические нейроны также дифференцировались из ЭС клеток человека, выход дофаминергических нейронов составлял лишь 5-7% от всей клеточной популяции (WO 03/000868).
Полагают, что при болезни Паркинсона особенно удобным клиническим подходом может быть трансплантация клеток, поскольку болезнь характеризуется селективной и постепенной потерей дофаминергических нейронов в субстанции нигра среднего мозга. Потеря дофаминпродуцирующих нейронов в этой конкретной области мозга приводит к патологической активации нервных клеток, что приводит к неспособности пациентов контролировать или направлять движения. Но для клеточно-заместительной терапии необходимо большое количество дофаминергических нейронов. Поэтому необходимы альтернативные способы более эффективного получения дофаминергических нейронов из ЭС клеток человека, которые позволят как ускорить доступность этого лечения болезни Паркинсона, так и увеличить вероятность успешного лечения. Кроме того, эти дофаминергические нейроны могут быть использованы in vitro для определения веществ, которые предотвратят или уменьшат смертность клеток мозга, продуцирующих дофамин при нейродегенеративных расстройствах и нейрональных заболеваниях.
КРАТКАЯ СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к улучшенным способам получения нейрональных клеток-предшественниц, а также терминально дифференцированных нейронов из плюрипотентных стволовых клеток, таких как эмбриональные стволовые клетки человека. Например, в настоящем описании показано, что популяция эмбриональных стволовых клеток человека может дифференцироваться с высоким количественным выходом в нейроны позитивные для тирозингидроксилазы (ТН), которая является специфическим маркером для дофаминергических нейронов (например, по крайней мере около 60%). В настоящем описании также показано, что популяция эмбриональных стволовых клеток человека может дифференцироваться с высоким количественным выходом в серотонинергические нейроны. Процент дофаминергических и серотонинергических нейронов, полученных в соответствии со способами настоящего изобретения, выше по сравнению со способами, описанными ранее. Способы, описанные здесь, могут также использоваться для получения клеток с фенотипическими характеристиками холинергических и сенсорных нейронов, а также астроцитов и олигодендроцитов из эмбриональных стволовых клеток человека.
Благодаря настоящему изобретению можно получить дифференцированную клеточную популяцию в культуре in vitro путем дифференцирующихся плюрипотентных стволовых клеток приматов, где по крайней мере 60% дифференцированных клеток являются дофаминергическими нейронами, экспрессируют тирозингидроксилазу, или являются дофаминергическими нейронами, которые экспрессируют тирозингидроксилазу. В другом варианте осуществления по крайней мере около 30, 40, 50, 70, 80, 90, 95 или 99% дифференцированных клеток являются дофаминергическими нейронами. Благодаря настоящему изобретению можно получить также дифференцированную клеточную популяцию в культуре in vitro путем дифференцировки плюрипотентных стволовых клеток приматов, где по крайней мере 30% дифференцированных клеток являются серотонинергическими нейронами. В другом варианте осуществления по крайней мере около 40, 50, 60, 70, 80, 90, 95 или 99% дифференцированных клеток являются серотонинергическими нейронами. В предпочтительном варианте осуществления плюрипотентные стволовые клетки приматов, дифференцированные в клетки-предшественницы нейронов или нейроны, являются эмбриональными стволовыми клетками человека.
В настоящем описании представлены также способы получения дифференцированных нейрональных клеточных популяций из плюрипотентных стволовых клеток приматов, предусматривающие следующие стадии:
(а) увеличение культуры плюрипотентных стволовых клеток приматов;
(b) культивирование плюрипотентных стволовых клеток для селекции нейрональных нестин-положительных клеток-предшественниц;
(с) сортировку нестин-положительных нейрональных клеток-предшественниц для увеличения NCAM-положительных клеток;
(d) дифференцировку нестин-положительных, NCAM-положительных клеток для получения дифференцированной нейрональной клеточной популяции путем культивирования клеток в среде для дифференцировки.
В предпочтительном варианте осуществления плюрипотентные стволовые клетки приматов являются человеческими эмбриональными стволовыми клетками. В другом предпочтительном варианте осуществления плюрипотентные стволовые клетки, использованные в приведенных выше способах, предпочтительно получают с помощью способа лазерного разрушения.
В другом варианте осуществления вышеуказанные способы предусматривают также стадию культивирования плюрипотентных стволовых клеток в стадии (b) с образованием эмбриоидных тел. Предпочтительно, эти эмбриоидные тела культивируют в условиях, которые выбраны для нейрональных нестин-положительных клеток-предшественниц, например, путем культивирования плюрипотентных стволовых клеток или эмбриоидных тел в среде без сыворотки. В предпочтительном варианте осуществления средой, не содержащей сыворотку, является безсывороточная среда ITSFn, которая предпочтительно включает один или несколько растворимых факторов, выбранных из группы, включающей инсулин, селенит натрия, основной фактор роста фибробластов, трансферрин и фибронектин. В предпочтительном варианте осуществления на основании этих способов можно получить нейрональные клетки-предшественницы, которые предпочтительно включают по крайней мере около 60-75% нестин-положительных клеток, более предпочтительно около 80-90% нестин-положительных клеток и наиболее предпочтительно около 95-99% нестин-положительных клеток.
Нестин-положительные нейрональные клетки-предшественницы затем могут быть отсортированы с получением обогащения NCAM-положительными клетками, используя подходящие иммунологические методы, такие как сортировка иммуномечением и флуоресцентная сортировка, например, магнитоактивируемая клеточная сортировка (MACS), адсорбция на твердой фазе, флуоресцентно-активируемая клеточная сортировка (FACS), проточная иммуноцитохимия для клеточных маркеров или проточный цитометрический анализ. В предпочтительном варианте осуществления эти методы дают нестин-положительные клетки, которые предпочтительно включают по крайней мере около 40-70% NCAM-положительных клеток, более предпочтительно около 50-60% NCAM-положительных клеток и наиболее предпочтительно около 80-99% NCAM-положительных клеток. В конкретном варианте осуществления указанные выше способы также предусматривают стадию роста нестин-положительных, NCAM-положительных нейрональных клеток-предшественниц стадии (с) в среде для роста предпочтительно в течение 6-10 дней. Предпочтительно, чтобы среда для роста содержала один или несколько растворимых факторов, выбранных из группы, включающей инсулин, селенит натрия, трансферрин, ламинит, путресцин, прогестерон, основной фактор роста фибробластов (bFGF), эпидермальный фактор роста (EGF), "звуковой ежик" (SHH), фактор роста фибробластов-8 (FGF-8) и мозговой нейротрофический фактор (BDNF).
Для удвоения или большего увеличения популяции предпочтительно, чтобы нестин-положительные, NCAM-положительные нейрональные клетки-предшественницы росли в культуре и их периодически пассировали. Эти клетки также могут быть заморожены в жидком азоте. NCAM-положительные нейрональные клетки-предшественницы предпочтительно выращивать в среде для дифференцировки в течение 30-50 дней, как указано в четвертой стадии (d) указанных выше способов. В предпочтительном варианте осуществления среда для дифференцировки включает нейробазальную среду с добавлением эмбриональной сыворотки теленка, В27, аскорбиновую кислоту и N-ацетилцистеин. В другом предпочтительном варианте осуществления среда для дифференцировки может содержать TGF-β3, или интерлейкин-1β, или оба этих факта. Предпочтительно среда для дифференцировки также включает один или несколько агентов для дифференцировки выбранных из группы, включающей аскорбиновую кислоту, N-ацетилцистеин, глиальный нейротрофический фактор (GDNF), дибутирил-цикло АМР (db-cAMP), нейротрофический мозговой фактор (BDNF), неутурин, белок "звуковой ежик" (SHH), фактор роста фибробластов-8 (FGF-8).
В предпочтительных вариантах осуществления способы, описанные выше, используются для получения дифференцированной нейрональной клеточной популяции, которая предпочтительно содержит около 40-60% дофаминергических нейронов, более предпочтительно около 70-80% дофаминергических нейронов и наиболее предпочтительно около 90-99% дофаминергических нейронов. В определенном варианте осуществления эти способы применяют для получения дифференцированных нейрональных клеточных популяций, которые предпочтительно включают около 20-50% серотонинергических нейронов, более предпочтительно около 30-70% серотонинергических нейронов и наиболее предпочтительно около 60-99% серотонинергических нейронов. В другом варианте осуществления эти способы применяют для получения дифференцированной нейрональной клеточной популяции, которая предпочтительно включает около 15-40% олигодендроцитов, более предпочтительно около 25-50% олигодендроцитов и наиболее предпочтительно около 60-99% олигодендроцитов.
Настоящее изобретение относится к способам получения дофаминергических нейронов из нейрональных клеток-предшественниц, предусматривающим обогащение нейрональных клеток-предшественниц нестин-положительными клетками, дифференцировку нестин-положительных клеток для получения дофаминергических нейронов путем культивирования клеток в присутствии TGF-β3, или интерлейкина-1β, или обоих этих факторов. Предпочтительно по крайней мере около 40-99% нестин-положительных клеток дифференцируются в дофаминергические нейроны в соответствии с этими методами. В другом варианте осуществления эти способы также предусматривают обогащение нейрональных клеток-предшественниц NCAM-положительными клетками, и эти нестин-положительные, NCAM-положительные клетки предпочтительно дифференцируются с получением дофаминергических нейронов (например, по крайней мере около 60-99% клеток дифференцируются в дофаминергические нейроны). Настоящее изобретение также относится к способам получения нейронов из нейрональных клеток-предшественниц, которые предусматривают обогащение нейрональных клеток-предшественниц клетками, которые являются положительными по нестину и NCAM, и дифференцировку нестин-положительных, NCAM-положительных клеток с получением серотонинергических нейронов путем культивирования клеток в присутствии TGF-β3, или интерлейкина-1β, или обоих этих факторов. Предпочтительно по крайней мере около 30-99% нестин-положительных, NCAM-положительных клеток дифференцируется в серотонинергические нейроны в соответствии с этими способами.
Настоящее изобретение также относится к способам лечения индивидуумов с нейродегенеративными нарушениями или неврологическими заболеваниями путем введения индивидуумам дифференцированных нервных клеток, полученных из плюрипотентных стволовых клеток приматов, как описано здесь. Например, популяция дифференцированных нейрональных клеток может быть получена следующим образом:
(а) увеличением культуры плюрипотентных стволовых клеток приматов;
(b) культивированием плюрипотентных стволовых клеток с отбором нейрональных клеток-предшественниц, которые положительны по нестину;
(с) сортировкой нестин-положительных нейрональных клеток-предшественниц с обогащением NCAM-положительными клетками;
(d) дифференцировкой нестин-положительных, NCAM-положительных клеток с получением дифференцированной популяции нервных клеток путем культивирования клеток в среде для дифференцировки;
(е) введением терапевтически эффективных количеств популяции дифференцированных нервных клеток в центральную нервную систему пациента.
Предпочтительно среда для дифференцировки включает TGF-β3, или интерлейкин-1, или оба этих фактора. В предпочтительном варианте осуществления, индивидуумом является пациент, более предпочтительно пациентом является человек, и плюрипотентные стволовые клетки приматов являются эмбриональными стволовыми клетками человека. Предпочтительно, человеческие эмбриональные стволовые клетки гистосовместимы с пациентом, например, используемые плюрипотентные стволовые клетки содержат геном, по существу идентичный геному пациента. В конкретном варианте осуществления дофаминергические, серотонинергические, холинергические или сенсорные нейроны или же астроциты или олигодендроциты выделяют из дифференцированной популяции нервных клеток и вводят пациенту. Эти клетки, включая дифференцированную популяцию нервных клеток, могут вводиться индивидууму для лечения различных нейродегенеративных расстройств или неврологических заболеваний, включая, но не ими ограничиваясь, болезни ЦНС, болезнь Паркинсона, болезнь Альцгеймера, хорею Гентингтона, повреждение спинного мозга, боковой амиотрофический склероз (БАС), эпилепсию, инсульт и ишемию. Предпочтительно, клетки вводятся путем трансплантации, например трансплантации желаемых клеток в мозг индивидуума.
В другом варианте осуществления дифференцированные нервные клетки, полученные из плюрипотентной стволовой клетки, как описано здесь, могут быть использованы для скрининга соединений, например, небольших молекул и лекарств, для определения их влияния на дифференцированные нервные клетки или активность этих клеток. Соединения могут быть также оценены на их токсичность в отношении нервных клеток или на способность их модулировать. Соединение может быть оценено путем добавления соединения к популяции дифференцированных нервных клеток и сравнением их выживания, морфологии, фенотипа, функциональной активности или других характеристик клеток по сравнению с дифференцированными нервными клетками, культивируемыми в тех же условиях, но не обработанными соединением. Например, соединения могут быть проскринированы для определения изменения синтеза нейромедиаторов, высвобождения или захвата клетками.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Следующие чертежи составляют часть настоящего описания и приведены здесь для дополнительной демонстрации некоторых аспектов настоящего изобретения, которое может быть лучше понято при отсылке на один или несколько из этих рисунков в сочетании с подробным описанием конкретных вариантов осуществлений, представленных здесь.
На фигуре 1 показаны нейрональные клетки-предшественницы, полученные из эмбриональной стволовой клетки человека, которые положительны по нестину: на (А) показаны нервные клетки-предшественницы, которые являются иммунореактивными с маркерами нестина; на (B) показана фазово-контрастная фотография нестин-положительных клеток, которые росли в среде без добавления сыворотки в присутствии выбранных факторов роста.
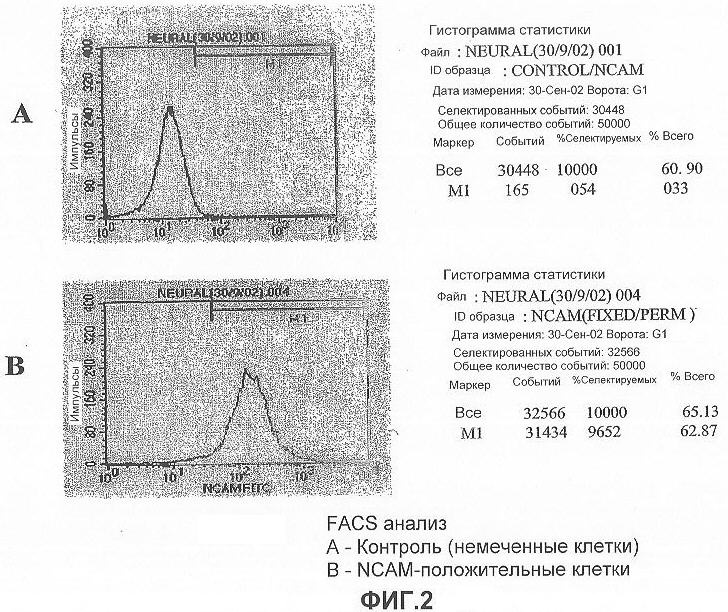
На фигуре 2 показан FACS анализ нестин-положительных клеток, полученных из эмбриональных стволовых клеток человека и меченных NCAM-FITC: на (A) представлен анализ немеченых клеток, которые были обработаны антикроличьими антителами, меченными FITC; на (B) показан анализ клеток, обработанных первичными антителами к NCAM, и меченных антикроличьими антителами с FITC (вторичное антитело). В этом исследовании около 50-60% нестин-положительных клеток были иммуноположительными для NCAM.
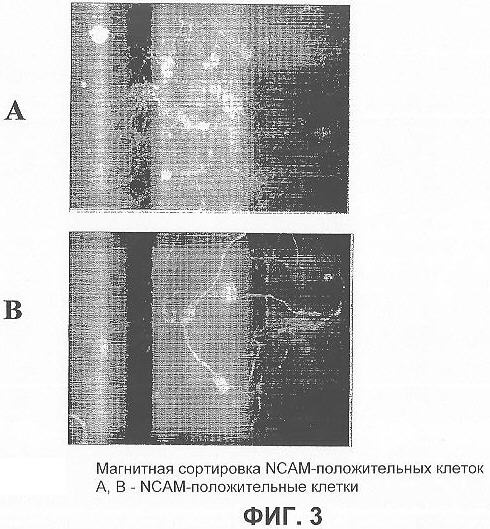
На фигурах 3А и 3В представлены флуоресцентные микрофотографии NCAM-положительных клеток, полученных из эмбриональных стволовых клеток человека, которые были отсортированы с использованием MACS и пересажены на чашку с культурой ткани.
На фигурах 4А и 4В представлены флуоресцентные микрофотографии нейронов, меченных MAP-2 и β-тубулином, полученных из эмбриональных стволовых клеток человека.
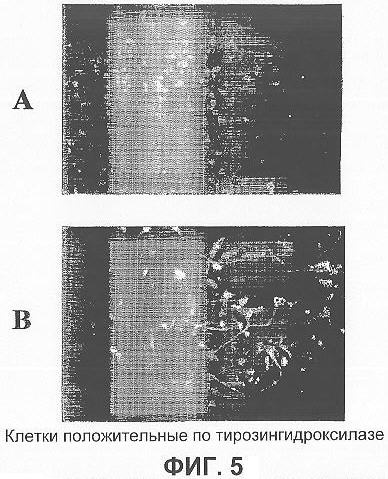
На фигуре 5 показано наличие нейронов, положительных для тирозингидроксилазы (TH): (А) иммунофлуоресцентный анализ показал, что около 60% нейронов были положительными для TH в обогащенной c использованием MACS популяции NCAM-положительных клеток; (В) иммунофлуоресцентный анализ показал, что около 40% нейронов были положительными для TH после размножения и дифференцировки несортированных нестин-положительных клеток.
На фигуре 6 представлена характерная флуоресцентная микрофотография олигодендроцитов. Иммунофлуоресцентный анализ показал, что приблизительно 25-30% выделенных нестин-положительных клеток окрашивались как олигодендроциты.
На фигуре 7 представлена флуоресцентная микрофотография нервных клеток, экспрессирующих нейромедиатор серотонин. Предположительно 30% нестин-положительных клеток, а также предположительно 20% NCAM-положительных, были иммунореактивными с серотонином.
На фигуре 8 представлена флуоресцентная микрофотография дофаминергических нейронов иммуномеченных TH (зеленый) и другим специфическим для нейронов антигеном (красный): солокализация ТН с (А) β-тубулином; (В) МАР-2; (С) Nurr1 и (D) DAT.
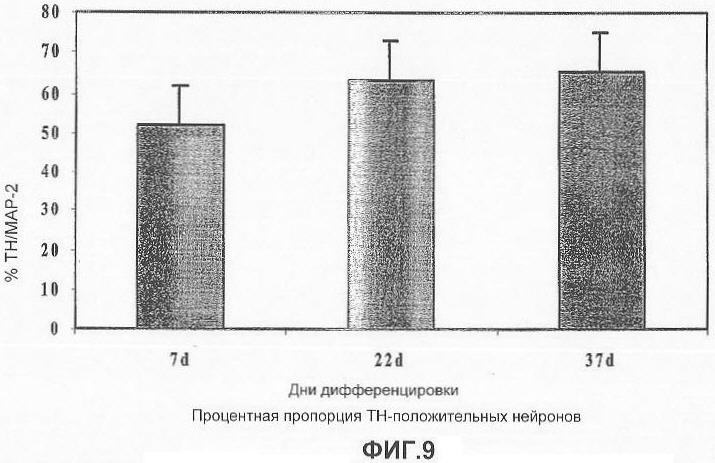
На фигуре 9 представлена столбчатая диаграмма, показывающая процент МАР-2 положительных нейронов, положительных в отношении ТН (дофаминергические нейроны): как видно, процент нейронов положительных по ТН увеличивается на протяжении 7, 22 и 37 дня дальнейшей дифференцировки.
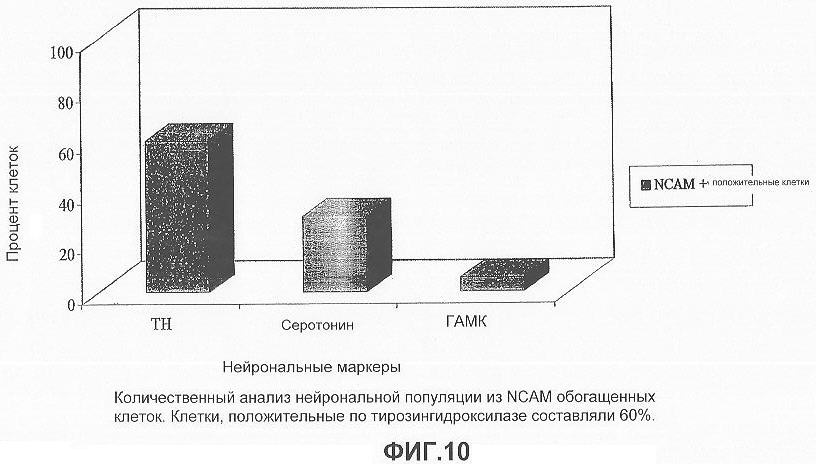
На фигуре 10 представлена столбчатая диаграмма, показывающая количественный анализ различных нейрональных клеточных популяций в NCAM-положительных обогащенных клетках: приблизительно 60% NCAM-положительных клеток были также иммуноположительными к ТН; приблизительно 30% были иммуноположительными к серотонину и приблизительно 15% были иммуноположительными по ГАМК и глутамату.
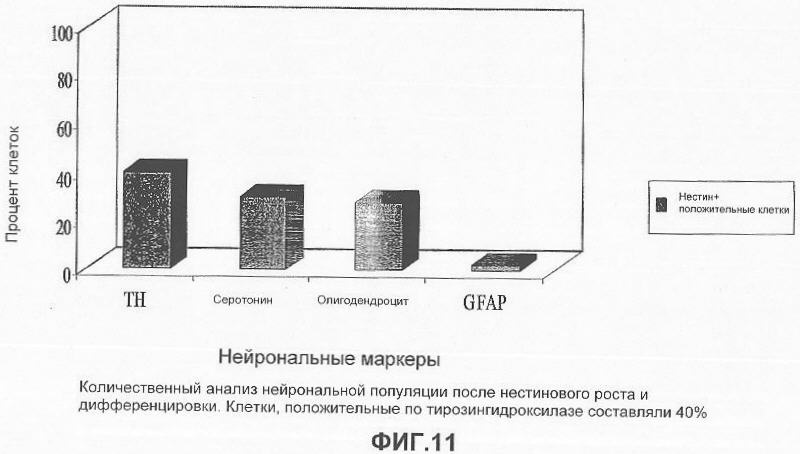
На фигуре 11 представлена столбчатая диаграмма, показывающая количественный анализ различных нейрональных клеточных популяций после роста и дифференцировки нестин-положительных клеток, анализируемых с помощью иммунофлуоресценции: приблизительно 40% нестин-положительных клеток были иммуноположительными к ТН; приблизительно 30% были иммуноположительными к серотонину; приблизительно 28% были иммуноположительными к олигодендроцитам и приблизительно 2% были иммуноположительными к глиальному фибриллярному кислому белку (GFAP, маркер для астроцитов).
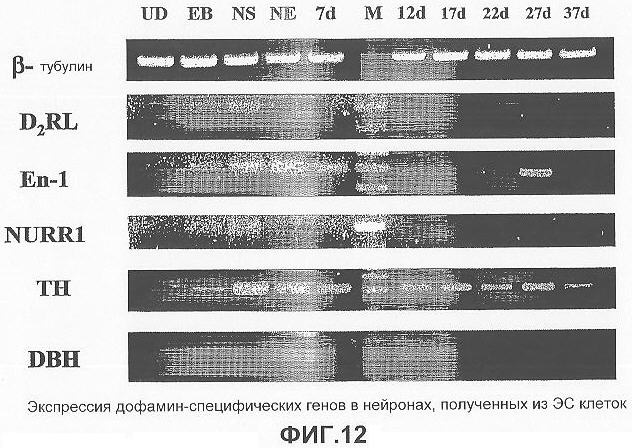
На фигуре 12 представлен профиль генетической экспрессии эмбриональных стволовых клеток человека в течение терминальной дифференцировки этих клеток in vitro в условиях, описанных в примере 1: UD = недифференцированные; EB = эмбриоидные тела; NS = нестин-положительные клетки; NE = нестин увеличенные клетки; и оставшееся время указывает число дней культивирования клеток в нейробазальной среде и выбранных факторов роста, как описано в примере 1. Факторы транскрипции, специфичные для дофаминергических нейронов, таких как Nurr1, En-1, D2RL, экспрессируются в течение ранних стадий дифференцировки. Экспрессия гена ТН, специфичного для дофаминергического нейрона, наблюдалась на всех стадиях, за исключением недифференцированных стволовых клеток. Отсутствие любой экспрессии DBH подтверждает фенотип этих клеток (среднего мозга), характерный для среднего мозга.
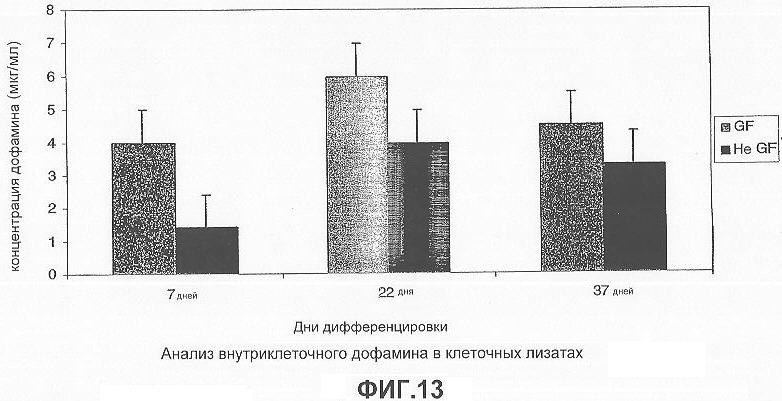
На фигуре 13 показан внутриклеточный уровень дофамина в клеточных лизатах, определенный с помощью ОФ-ВЭЖХ на 7, 22 и 37 день дифференцировки. В присутствии факторов роста уровень дофамина был значительно выше (4-6 мкг/мл), чем в их отсутствие (1-4 мкг/мл). Дофамин не был обнаружен в клетках, обработанных MPTP, на 7, 22 дни, тогда как на 37 день уровень дофамина был пониженным.
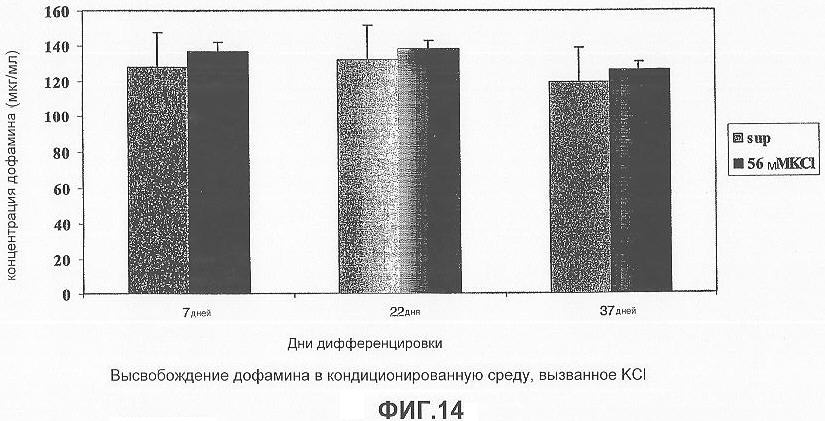
На фигуре 14 представлена столбчатая диаграмма, демонстрирующая, что KCl вызывает высвобождение дофамина в культуре дифференцированных клеток в кондиционированной среде после 7, 22 и 37 дня дифференцировки. Клетки были стимулированы 56 мМ KCl в течение 15 минут с индукцией секреции дофамина, супернатант культуры стабилизировали 7,5% ортофосфорной кислотой и пиросульфитом натрия перед проведением анализа ОФ-ВЭЖХ.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам эффективного получения клеток нейрональной линии, которые дифференцируются из плюрипотентной стволовой клетки. Клетки, полученные здесь, включают, но ими не ограничиваются, клетками с фенотипическими признаками нейрональных клеток-предшественниц, дофаминергических, серотонинергических, холинергических и сенсорных нейронов, а также астроцитов и олигодендроцитов. Клетки, полученные здесь, идентифицируют с помощью фенотипических признаков, морфологических признаков и/или клеточных маркеров, которые хорошо известны специалисту в данной области. Как используется здесь, термин «нейрональная клетка-предшественница» является взаимозаменяемым с термином «клетка-предшественница нейрона» и относится к клетке, которая может образовывать предков, которые являются либо нейрональными клетками, такими как нейрональные предшественницы или нейроны, либо глиальными клетками, такими как глиальные предшественницы, астроциты или олигодендроциты. Способы, описанные здесь, предусматривают культивирование клеток в сочетании с растворимыми факторами и условиями культивирования, которые способствуют дифференцировке клеток в клетки нейрональной линии. Кроме того, для еще большего обогащения желаемыми клетками нейронального типа могут быть использованы методы физического разделения или манипуляции.
Эти предшественницы и дифференцированные нейрональные клетки могут использоваться для различных применений, включая терапевтическое и экспериментальное применение, а также при разработке лекарств in vitro и скрининга, такого как скрининг соединений, являющихся токсичными для клетки или способных модулировать функцию нейрональных клеток. Получение предшественниц и дифференцированных нейрональных клеток, таких как дофаминергических или серотонинергических нейронов, а также других специализированных нейрональных клеточных типов из плюрипотентной стволовой клетки, является, по сути, неограниченным источником этих клеток с огромным возможным положительным эффектом для людей, страдающих разрушающими нейродегенеративными нарушениями и неврологическими заболеваниями. Предшественник и дифференцированные нервные клетки, описанные здесь, являются предками клеточной популяции, из которой они были получены, и поэтому в значительной степени будут иметь тот же геном, что и родительская популяция, включая родительскую популяцию, которая была генетически изменена, трансформирована или трансфицирована.
Один из вариантов осуществления настоящего изобретения относится к улучшенному способу получения нейронов из плюрипотентной стволовой клетки, предпочтительно эмбриональной стволовой (ЭС) клетки приматов или эмбриональной зародышевой (ЭЗ) клетки приматов, которые обладают свойствами нейронов среднего мозга, таких как дофаминергических нейронов. Другой вариант осуществления относится к улучшенному способу получения нейронов из плюрипотентной стволовой клетки, предпочтительно ЭС клетки приматов или ЭЗ клетки приматов, которые обладают свойствами нейронов заднего мозга, например, свойствами серотонинергических нейронов. ЭС клетка или ЭЗ клетка приматов, которая может быть использована в этих способах, наиболее предпочтительно, является ЭС клеткой или ЭЗ клеткой человека. Эти нейроны получают из плюрипотентной стволовой клетки путем культивирования этих клеток в присутствии некоторых растворимых факторов и условий внешней среды.
Как используется здесь, термин «дофаминергические нейроны» относится к нейрональным клеткам, которые экспрессируют тирозингидроксилазу (ТН), фермент, лимитирующий синтез дофамина. Предпочтительно, дофаминергические нейроны секретируют нейромедиатор дофамин и либо не экспрессируют его, либо экспрессируют дофамин-β-гидроксилазу в небольшом количестве. In vivo дофаминергические нейроны иннервируют полосатое тело, лимбическую систему и неокортекс и расположены в вентральной части среднего мозга вместе с нейронами других классов, включая моторные нейроны. Более точно дофаминергические нейроны располагаются в черной субстанции (substantia nigra) среднего мозга и контролируют поступальные рефлексы, движение и участвуют в системе поощрения (центр удовольствия). Потеря нормально функционирующих дофаминергических нейронов приводит к болезни Паркинсона, и их патологическая функция связана с шизофренией и наркоманией. Как используется здесь «серотонинергические нейроны» относятся к нейронам, которые секретируют нейромедиатор серотонин (5-гидрокситриптамин). Серотонинергические нейроны обычно имеют медленный, ритмический характер возбуждения и in vivo концентрируются в вентральной и вентролатеральных областях заднего мозга и иннервируют многие части центральной нервной системы, включая кору головного мозга, лимбическую систему и спинной мозг. Эти нейроны контролируют степень сознания, активность ЦНС, поведение и аппетит. Патологическая функция серотонинергических нейронов связана с агрессией, депрессией (включая суицидальное поведение) и шизофренией.
Настоящее изобретение относится к улучшенным способам дифференцировки плюрипотентных стволовых клеток в нейрональные клетки-предшественницы, а также в дифференцированную популяцию нервных клеток, обладающих фенотипическими, молекулярными и/или клеточными свойствами, сходными с клетками нейрональной линии. Плюрипотентные стволовые клетки могут быть ЭС клетками человека, и дифференцированные нервные клетки могут быть дофаминергическими нейронами или серотонинергическими нейронами. Настоящее изобретение также относится к клеткам и клеточным популяциям, полученным указанными способами. В конкретных вариантах осуществления описанные способы предусматривают следующие стадии:
1. Выделение популяции плюрипотентных стволовых клеток; предпочтительно плюрипотентные стволовые клетки являются ЭС клетками человека, полученными путем новой методики лазерного разрушения.
2. Увеличение количества плюрипотентных стволовых клеток для обеспечения достаточного начального материала.
3. Культивирование плюрипотентных стволовых клеток в суспензии с получением эмбриоидных тел.
4. Эмбриоидные тела наносят на субстрат и инкубируют в среде без сыворотки, которая является селективной для нейрональных нестин-положительных клеток-предшественниц.
5. Нестин-положительные клетки сортируют и выделяют и обогащают популяцию NCAM-положительными клетками.
6. Увеличение количества нестин-положительных и/или NCAM-положительных нейрональных клеток-предшественниц в среде для роста, которая содержит растворимые факторы, относящиеся к нервной системе.
7. Нестин-положительные и/или NCAM-положительные нейрональные клетки-предшественницы дифференцируются в зрелые нейроны в нейробазальной среде, которая содержит сочетание растворимых факторов, относящихся к нервной системе.
Источники плюрипотентных стволовых клеток
Способы дифференцировки клеток нейрональной линии из плюрипотентных стволовых клеток, описанные здесь, предусматривают использование специальных условий культивирования, которые направляют дифференцировку значительной части плюрипотентных стволовых клеток по определенному нейрональному клеточному пути. Плюрипотентные стволовые клетки получают из пре-эмбриональных, эмбриональных или зародышевых тканей в любое время после оплодотворения, и эти клетки при подходящих условиях способны дифференцироваться в несколько различных клеточных типов, которые являются источниками всех трех зародышевых листов (эндодермы, мезодермы и эктодермы). Клетки нейрональной линии также могут быть получены из стволовых клеток, выделенных из зародышевой или взрослой ткани, которые способны дифференцироваться или быть перепрограммированными в клетки нейрональной линии. Плюрипотентные стволовые клетки включают, но ими не ограничиваются, ЭС клетки и ЭЗ клетки млекопитающих, предпочтительно ЭС клетки и ЭЗ клетки примата или человека. Предпочтительно недифференцированные плюрипотентные стволовые клетки способны неограниченно делиться и пролиферироваться в культуре. Как используется здесь, термин «дифференцировка» относится к процессу, посредством которого недифференцированные плюрипотентные стволовые клетки или клетки-предшественницы приобретают более специализированный путь. Например, дифференцированные клетки облают фенотипом, который характерен для определенного типа клеток или тканей.
В предпочтительном варианте осуществления ЭС клетки и линии ЭС клеток, используемых здесь, получают из внутренней клеточной массы бластоцисты. Бластоцисты могут быть выделены из восстановленных in vivo оплодотворенных преимплантационных эмбрионов или при оплодотворении in vitro (IVF), например, из эмбрионов, оплодотворенных обычным проникновением сперматозоидов в яйцо, внутриклеточной инъекции спермы или переносом ооплазмы. Человеческие бластоцисты получают от пар или доноров, которые добровольно предоставили свои эмбрионы. Эти эмбрионы используются для исследовательских целей после получения письменного и добровольного согласия этих пар или доноров. С другой стороны, бластоциста может быть получена путем переноса соматической клетки или клеточного ядра в безъядерный ооцит человеческого или нечеловеческого происхождения, который затем стимулируют для развития до стадии бластоцисты. Используемые бластоцисты также могут быть заморожены или могут быть получены из замороженных на ранних стадиях эмбрионов и затем продолжать развитие до стадии бластоцисты эмбриона. Развитие как бластоцисты, так и внутренней клеточной массы будет варьировать в зависимости от видов и хорошо известно специалистам.
ЭС клетки приматов или человека могут быть получены из бластоцисты, используя стандартные иммунохирургические методики, описанные в патенте США №5843780 и 6200806 Thomson et al. (Science 282:1145-1147, 1998) и Reubinoff et al. (Nature Biotech. 18:399-403, 2000), каждая конкретно приведена здесь в качестве ссылки. Хотя в описанных способах могут использоваться ЭС клетки, полученные любым известным специалисту способом, предпочтительно использовать ЭС клетки человека, полученные уникальным методом лазерного разрушения (патент США №10/798790, конкретно приведенный здесь в качестве ссылки). Коротко, в этом методе клетки выделяют из внутренней клеточной массы бластоцисты путем лазерного разрушения части zona pellucida и трофобласта бластоцисты, которое образует отверстие или дырку в бластоцисте, через которую могут быть аспирированы клетки внутренней клеточной массы. Эти клетки затем дополнительно могут быть культивированы с получением ЭС клеточных линий. Эта технология обладает преимуществом, поскольку позволяет выделять клетки внутренней клеточной массы без проведения обычных трудоемких методик иммунохирургии. Кроме того, линии ЭС клеток, полученные с использованием этой технологии, в частности, линии ЭС клеток человека, могут быть выделены без каких-либо антител и сыворотки, полученных из продуцируемых животных, что минимизируют риск переноса микроорганизмов животных в линию ЭС клеток. В еще одном варианте осуществления используют ЭЗ клетки человека, полученные из первоначальных зародышевых клеток, находящихся в зародышевом материале человека (патент США № 6090622, и Shamblott et al., 1998, Proc. Natl. Acad. Sci. USA. 95:13726-13731, каждая приведена здесь в качестве ссылки).
Предпочтительно, линии ЭС клеток могут поддерживаться в культуре в недифференцированном состоянии в течение продолжительного периода времени, например, в течении более одного года, при сохранении нормального диплиодного кариотипа. ЭС клетки человека могут быть морфологически идентифицированы по высокому показателю отношения ядра и цитоплазмы, заметным нуклеолям и образованию компактных колоний, зачастую с отчетливым клеточным краем и колониями, которые, как правило, более плоские, чем ЭС клетки мышей. Желательно, чтобы ЭС клетки человека были также иммунореактивными с маркерами для плюрипотентных ЭС клеток человека, например, SSEA-3, SSEA-4, GCTM-2 антигеном и TRA 1-60, как описано Thomson et al. (1998), Reubinoff et al (2000), Buehr и Mclaren (1993), каждая приведена здесь в качестве ссылки. Предпочтительно, ЭС клетки человека также экспрессируют щелочную фосфатазу, а также OCT-4. В другом варианте осуществления ЭС клетки человека способны образовывать эмбриональные тела при неадгезивных условиях культивирования (патент США № 6602711, приведенный здесь в качестве ссылки). Эти эмбриоидные тела могут быть использованы для получения дифференцированных производных зародышевых листов эндодермы, мезодермы и эктодермы, а также желаемых линий клеток.
Плюрипотентные стволовые клетки, в особенности ЭС и ЭЗ клетки человека, можно репродуцировать продолжительное время в условиях культивирования, при которых клетки поддерживаются в по существу недифференцированном состоянии. ЭС клетки должны находиться с подходящей клеточной плотностью, и их регулярно следует диссоциировать и пересевать, часто меняя культуральную среду для предотвращения их от дифференцировки. Для обычных методов, относящихся к клеточной культуре и культивированию ЭС клеток, специалист может обратиться за соответствующей информацией в стандартные справочники и обзоры, например: E. J. Robertson, "Teratocarcinomas and embryonic stem cells: A practical approach" ed., IRL Press Ltd. 1987; Hu и Aunins, 1997, Curr. Opin. Biotechnol. 8(2):148-53; Kitano, 1991, Biotechnology 17:73-106; Spier, 1991, Curr. Opin. Biotechnol. 2:375-79; Birch и Arathoon, 1990, Bioprocess Technol. 10:251-70; Xu et al., 2001, Nat. Biotechnol. 19 (10):971-4; и Lebkowski et al., 2001, Cancer J. 7 Suppl. 2:S83-93; каждая приведена здесь в качестве ссылки.
Обычно ЭС клетки культивируют в ЭС среде для ЭС клеток на слое питающих клеток. Слои питающих клеток состоят из клеток одного тканевого типа, которые сокультивируют с ЭС клетками, и обеспечивают окружающие условия, в которых ЭС клетки могут расти без существенной дифференцировки. Методы для культивирования ЭС клеток на слое питающих клеток хорошо известны специалистам в этой области (патент США № 5843780 и 6200806, WO 99/20741, 09/530346 и 09/849022, WO 01/51616, каждая конкретно приведена здесь в качестве ссылки). Слой питающих клеток предпочтительно уменьшает, ингибирует или предотвращает дифференцировку ЭС клеток. Слои питающих клеток обычно представляют собой слой эмбриональных фибробластов, полученных либо у человека, либо мыши, например, эмбриональные фибробласты мыши, эмбриональные фибробласты человека, фибробластоподобные клетки человека или клетки мезенхимы, полученные из эмбриональных стволовых клеток человека или клеток STO.
ЭС клетки культивируют в присутствии среды для ЭС клеток, которая уменьшает, ингибирует или предотвращает дифференцировку ЭС клеток. Предпочтительно ЭС среда для ЭС клеток, используемая для культивирования ЭС клеток, содержит питательную сыворотку, например, сыворотку или раствор, основанный на сыворотке, который содержит питательные вещества, достаточные для поддержания роста и жизнеспособности ЭС клеток. Питательная сыворотка может быть животной сывороткой, такой как эмбриональная бычья сыворотка (FBS) или эмбриональная телячья сыворотка (FCS) (патент США № 5453357, 5670372 и 5690296, которая приведена здесь в качестве ссылки). Среда для клеток ЭС также может быть без сыворотки (WO 98/30679, WO 01/66697, США № 09/522030, каждая приведена здесь в качестве ссылки). Примером подходящей ЭС среды для ЭС клеток с сывороткой для культивирования ЭС клеток является измененная среда Дульбекко-Игл (DMEM), без пирувата натрия, с высоким содержанием глюкозы (70-90%) (GIBCO), содержащая FBS или FCS (10-30%), β-меркаптоэтанол (0,1 мМ), заменимые аминокислоты (1%) и L-глутамин 2 мМ, 4 нг/мл основной фактор роста фибробластов (bFGF), 50 Ед/мл пенициллина, 50 мкг/мл стрептомицина и 1000 Ед/мл фактора ингибирования лейкемии (LIF). Примером подходящей ЭС среды без сыворотки для культивирования ЭС клеток является 80% «нокаут» модифицированная среда Дульбекко-Игл (DMEM) (GIBCO), 20% "нокаут" SR (безсывороточная замена, GIBCO), β-меркаптоэтанол (0,1 мМ), заменимые аминокислоты (1%) и L-глутамин 1 мМ.
Клетки ЭС также могут быть культивированы в культуральной среде без питательных клеток. Способы культивирования ЭС клеток в среде без питательных клеток хорошо известны специалистам (патентная публикация США 2002/0022268, WO 03/020920, США № 10/235094, каждая конкретно приведена здесь в качестве ссылки). ЭС клетки в среде без питательных клеток предпочтительно растут на подходящем культуральном субстрате, например, на внеклеточном матриксе, таком как Matrigel® (Becton Dickenson) или ламинине. В культуре без питательных клеток также предпочтительно использовать кондиционированную среду для поддержания роста ЭС клеток. Кондиционированную среду получают культивированием первой популяции либо эмбриональных фибробластов мышей, либо клеток эмбриональных фибробластов человека в среде в течение достаточного периода времени для получения «кондиционированной» среды, которая будет поддерживать культивирование ЭС клеток без значительной дифференцировки. С другой стороны, в культуре без питательных клеток можно комбинировать внеклеточный матрикс с оптимальной средой, которая добавляется свежей к культуре без предварительного кондиционирования клетками другого типа (патентная публикация США № 2003/0017589, конкретно приведена здесь в качестве ссылки).
Получение нейрональных клеток-предшественниц
Количество выделенных плюрипотентных стволовых клеток может быть увеличено и затем подвергнуто культивированию в условиях, которые вызывают их дифференцировку в нейрональные клетки-предшественницы. Для прохождения плюрипотентных стволовых клеток по нейрональному пути дифференцировки, клетки культивируют в соответствии с протоколом дифференцировки приведенным здесь. Плюрипотентные стволовые клетки культивируют на подходящем субстрате в питательной среде для дифференцировки, которая содержит агенты дифференцировки, такие как растворимые факторы и факторы роста. Подходящие субстраты включают, но ими не ограничиваются, твердую поверхность, положительно заряженную, например, поли-L-лизином или полиорнитином, субстраты, покрытые внеклеточными компонентами матрикса, например, фибронектином, ламинином или Matrigel® или их сочетанием. Предпочтительными питательными средами дифференцировки являются среды, которые поддерживают пролиферацию, дифференцировку и выживание желаемых клеток нейрональных типов и могут включать один или несколько подходящих агентов дифференцировки. Как используется здесь, термин «фактор роста» относится к белкам, которые связываются с рецепторами на поверхности клеток и активируют клеточную пролиферацию и дифференцировку. Подходящие растворимые факторы включают, но ими не ограничиваются, нейротрофины, митогены, факторы стволовых клеток, факторы роста, факторы дифференцировки (например, факторы суперсемейства TGF-β), агонисты факторов суперсемейства TGF-β, нейротрофные факторы, антиоксиданты, нейромедиаторы и факторы, необходимые для выживания. Многие растворимые факторы являются многофункциональными и стимулируют клеточное деление большого количества клеток различных типов, тогда как другие специфичны к определенному типу клеток.
Подходящие агенты дифференцировки, которые специфично стимулируют дифференцировку клеток нейронального типа, включают, но ими не ограничиваются, прогестерон, путресцин, ламинин, инсулин, селенит натрия, трансферрин, неуртурин, "звуковой ежик" (SHH), ноггин, фоллистатин, эпидермальный фактор роста (EGF), фактор роста фибробластов любого типа (например, FGF-4, FGF-8, основной фактор роста фибробластов (bFGF)), фактор роста и дифференцировки 5 (GDF-5), члены нейротрофического семейства (фактор роста нервов (NGF), нейротрофин 3 (NT-3), нейротрофин 4 (NT-4), мозговой нейротрофический фактор (BDNF), трансформирующий фактор роста α (TGF-α), трансформирующий фактор роста бета-3 (TGF-β3), фактор роста тромбоцитов (PDGF-AA), инсулиноподобный фактор роста (IGF-1), морфогенные белки кости (BMP-2, BMP-4), глиальный нейротрофический фактор (GDNF), ретиноивая кислота (RA), мидкин, аскорбиновая кислота, дибутирил-сАМР, дофамин и лиганды к рецепторам, которые образуют комплекс с gp130 (например, LIF, CNTF, SCF, IL-11 и IL-6). Питательная среда для дифференцировки может содержать также добавки, которые помогают поддерживать культуру для нейрональных клеток, например, добавки N2 и В27 (Gibco).
Сначала плюрипотентные стволовые клетки индуцируют для образования эмбриоидных тел. Эмбриоидные тела непосредственно помещают на подходящий субстрат с или без компонентов внеклеточного матрикса, такими как фибронектин или ламинин, и культивируют в подходящей питательной среде для дифференцировки, стимулируя дифференцировку в нейрональные клетки-предшественниц, такие как нестин-положительные нейрональные клетки-предшественницы. Нестин является клеточным маркером, характерным для нейрональных клеток-предшественниц. В другом варианте осуществления плюрипотентные стволовые клетки сначала агрегируют в гетерогенную клеточную популяцию, образуя эмбриоидные тела, например, культивируя плюрипотентные стволовые клетки в суспензии. Эти клетки можно культивировать в питательной среде с или без сыворотки, а также с одним или несколькими агентами дифференцировки, указанными выше, для стимуляции дифференцировки клеток в эмбриоидных телах. Как используется здесь, термин «эмбриоидные тела» относится к агрегации дифференцированных клеток, полученных, когда плюрипотентные стволовые клетки растут в суспензионной культуре, или переросли в культуру монослоя. Эмбриоидные тела также могут содержать недифференцированные клетки в клеточных агрегатах. Предпочтительно, такие агрегированные клетки окружены примитивной эндодермой. Эмбриоидные тела обычно содержат клетки, полученные из всех трех зародышевых листов, эктодермы, мезодермы и эндодермы. В зрелых эмбриоидных телах человека возможно распознать клетки, несущие маркеры различных типов клеток, таких как нейрональные клетки, гематопоэтические клетки, клетки печени и клетки сердечной мышцы. Некоторые клетки зрелых эмбриоидных тела функционально могут вести себя как дифференцированные клетки. Например, активные клетки сердечной мышцы могут вызывать пульсацию эмбриоидных тел. Предпочтительно, дифференцировка плюрипотентных стволовых клеток находится под контролем и клетки специфических типов могли бы получаться для терапевтических целей.
Эмбриоидные тела культивируют до достижения достаточного размера или желаемой степени дифференцировки, например, после 3-10 дней культивирования, и затем пересаживают на субстрат. Предпочтительно, субстрат покрыт компонентами внеклеточного матрикса, включая, но ими не ограничиваясь, поли-L-лизин, поли-L-орнитин, ламинин, коллаген, фибронектин, Matrige® или их сочетаниями. Предпочтительно, эмбриоидные тела помещают непосредственно на субстрат без диспергирования клеток. Эмбриоидные тела затем культивируют в условиях, способствующих дальнейшей дифференцировке клеток на чашке, например в среде ITSFn (селекция по нестину) определенного состава без сыворотки, которая селективна для нестин-положительных клеток. Нестин является белком промежуточных филаментов, который экспрессируется в нейроэпителии. Предпочтительно, селекцию нестин-положительных клеток проводят в течение 5-16 дней. Предпочтительно, средой ITSFn, используемой для увелечения количества нестин-положительных клеток, является DMEM:F-12, содержащая один или несколько факторов роста, выбранных из группы, включающей прогестерон, путресцин, ламинин, инсулин, селенит натрия, трансферрин, bFGF, SHH, EGF, FGF-2, FGF-8 и BDNF.
Эту гетерогенную клеточную популяцию, которая включает нестин-положительные клетки, последовательно увеличивают в размере или сортируют для обогащения NCAM-положительными адгезивными нейрональными клетками (NCAM). NCAM является клеточным маркером, характерным для нервных клеток. NCAM-положительные клетки могут быть отсортированы сразу после селекции нестин-положительных клеток или после размножения нестин-положительных клеток в культуре. В определенных вариантах осуществления клетки сортируют на NCAM-положительные клетки путем контактирования клеток с антителом или лигандом, который связывается с NCAM, с последующим отделением специфически узнанных клеток, используя подходящую иммунологическую методику, такую как иммунное мечение или флуоресцентная сортировка, например, адсорбция на твердой фазе, флуоресцентно-активируемая клеточная сортировка (FACS), проточная иммуноцитохимия для маркеров клеточной поверхности, проточный цитометрический анализ или магнитная клеточная сортировка (MACS). Другие методы выделения NCAM-положительных клеток включают, но ими не ограничиваются, дифференцированное рассеивание, иммуноспецифический лизис загрязняющих клеток или методики сбора клеток, хорошо известные специалисту в данной области. В предпочтительном варианте осуществления сортировка нестин-положительных клеток, например, с помощью MACS, обогащает популяцию нестин-положительных жизнеспособных клеток, экспрессирующих NCAM примерно на 40-70%, предпочтительно примерно на 60-80%, более предпочтительно примерно на 85-90% и наиболее предпочтительно примерно на 95-99%.
В одном из вариантов осуществления эмбриоидные тела образуются из ЭС клеток человека путем культивирования клеток в бактериологической плашке в подходящей среде в отсутствии питательных клеток. Предпочтительно, ЭС клетки человека диссоциируют на кластеры и затем помещают на неадгезивные чашки для облегчения развития эмбриоидных тел. Подходящая среда предпочтительно содержит DMEM с высоким уровнем глюкозы и дополнена 10-20% FCS. Среда также может содержать другие добавки, такие как 0,1 мМ 2-меркаптоэтанол, 2 мМ L-глутамин, 50 Ед/мл пенициллина и 50 мкг/мл стрептомицина. Предпочтительно, эмбриоидные тела оставляют расти в течение 4-8 дней, и затем эмбриоидные тела пересевались в культуральную плашку, покрытую 0,1% желатином в среде ITSFn без сыворотки для селекции нестин-положительных клеток. Предпочтительно, среда ITSFn без сыворотки содержит базальную среду DMEM:F-12 (1:1) или среду IMDM, содержащую факторы роста инсулин, селенит натрия, трансферрин и фибронектин, которые селективны в отношении нестин-положительных клеток.
В предпочтительном варианте осуществления нестин-положительные клетки сортируют с помошью MACS для выделения NCAM-положительных клеток. Количество NCAM-положительных клеток затем увеличивается в среде, которая помогает повысить процент нейрональных клеток-предшественниц и дополнительно индуцирует эти клетки для дальнейшей фенотипической дифференцировки. Предпочтительно, NCAM-положительные клетки культивируют на субстрате, предварительно покрытом компонентами внеклеточного матрикса, такими как поли-1-лизин, поли-1-орнитин, ламинин, коллаген, фибронектин, Matrigel® или их сочетания. Предварительное покрытие культуральных плашек внеклеточным матриксом обеспечивает лучшую адгезию и пролиферацию в среде для роста, а также дает лучшие результаты дифференцировки дофаминергических нейронов. Клетки культивируют в среде для роста, например, в среде DMEM/F12, содержащей один или несколько факторов роста, выбранных из группы, включающей прогестерон, путресцин, ламинин, инсулин, селенит натрия, трансферрин, bFGF, SHH, EGF, FGF-2, FGF-8, BDNF, PDGF, IGF-1, CTNF и NT-3. Не привязываясь к какому-либу конкретному механизму, полагают, что эти различные факторы, присутствующие в среде для роста, способствуют общему увеличению процента нейрональных клеток и дополнительно стимулируют нейрональные клетки-предшественницы среднего мозга для приобретения фенотипа дофаминергических клеток. В предпочтительном варианте осуществления нестин-положительные, NCAM-положительные клетки оставляют пролиферировать в течение 3-10 дней. Эти клетки также могут быть последовательно пассированы по крайней мере для троекратного увеличения популяции и заморожены для дальнейшего использования без какой-либо потери возможности к дифференцировки.
Дифференцировка дофаминергических и серотонинергических нейронов
Нейрональные клетки-предшественницы, полученные в соответствии с методами, описанными здесь, могут быть затем дифференцированы с большим процентом в зрелые нейроны, например дофаминергические нейроны и серотонинергические нейроны. Нейрональные клетки-предшественницы могут быть также затем дифференцированы в астроциты и олигодендроциты. Предпочтительно, нестин-положительные и/или NCAM-положительные нейрональные клетки-предшественницы оставляют расти в течение 5-60 дней в нейробазальной среде, которая способствует дифференцировке клеток-предшественниц в терминально дифференцированные нервные клетки или зрелые нейроны. В предпочтительном варианте осуществления нейробазальная среда (Gibco) содержит 10% FBS или FCS, B27 и один или несколько факторов роста, выбранных из группы, включающей интерлейкин-1β, дибутирил циклический АМФ (db-AMP), глиальный нейротрофический фактор (GDNF), трансформирующий фактор роста бета 3 (TGF-β3), трансформирующий фактор роста α (TGFα), неуртурин, SHH, аскорбиновую кислоту, BDNF, FGF-2, FGF-8, N-ацетилцистеин, с-kit лиганд, ретиновую кислоту, NT-3, BMP-2 и BMP-4. В предпочтительном варианте осуществления нейробазальная среда включает один или несколько следующих факторов: интерлейкин-1β, db-cAMP, GDNF, TGF-3, неуртурин, SHH, аскорбиновая кислота, BDNF, FGF-8 и N-ацетилцистеин. Кроме того, дифференцировка может быть облегчена посредством удаления некоторых или всех факторов, которые способствуют дифференцировке, пролиферации или тому, или другому нейрональных клеток-предшественниц.
Предпочтительно, высокий процент нейрональных клеток-предшественниц дифференцируется в дофаминергические нейроны, серотонинергические нейроны или олигодендроциты, например по крайней мере около 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 99% клеток. Кроме того, нервные клетки одного типа, например, дофаминергические нейроны, могут быть дополнительно очищены от популяции дифференцированных нервных клеток методами, хорошо известными специалистам, таким как иммунное мечение и флуоресцентная сортировка, например, адсорбция на твердой фазе, FACS, MACS и тому подобное. В одном из предпочтительных вариантов осуществления популяция NCAM-положительных клеток, обогащенных иммуносорбцией, растет и дифференцируется в подходящей среде, что приводит к большому проценту (около 60%) дофаминергических нейронов.
Использование нейрональных клеток-предшественниц и дифференцированных нервных клеток
Нейрональные клетки-предшественницы и дифференцированные нервные клетки, описанные здесь (например, зрелые нейроны, астроциты и олигодендроциты), могут быть использованы для различных применений, таких как терапевтическое применение, а также для оценки и скрининга in vitro различных соединений, таких как лекарства небольшой молекулярной массы, и их влияния на эти клетки. Клетки также могут использоваться для получения экспрессионных библиотек кДНК для анализа экспрессионных моделей этих клеток, а также для получения моноклональных или поликлональных антител, которые специфичны в отношении маркеров, определенных использованных клеток, с использованием технологии, которая хорошо известна специалистам в этой области. Эти клетки можно также использовать в терапии пациентов, страдающих изнуряющими нейродегенеративными расстройствами и неврологическими заболеваниями.
Настоящее изобретение относится к использованию нейрональных клеток-предшественниц и дифференцированных нервных клеток, описанных здесь, для восстановления функции центральной нервной системы (ЦНС) у людей, которым такая терапия необходима. Например, эти клетки могут быть использованы в лечебных целях путем трансплантации их непосредственно в паренхимную или интратекальную области ЦНС в зависимости от заболевания или условий лечения. Эти клетки могут быть использованы для лечения острых и хронических повреждений нервной системы, а также изнуряющих нейродегенеративных расстройств и неврологических заболеваний, таких как болезнь Паркинсона, болезнь Альцгеймера, хорея Гентингтона, повреждение спинного мозга, боковой амиотрофический склероз (БАС), эпилепсия, инсульт, ишемия и тому подобное.
В одном из вариантов осуществления настоящее изобретение относится к способам лечения нейродегенеративных расстройств или неврологических заболеваний, характеризующихся вырождением или разрушением дофаминергических нейронов, посредством введения или трансплантации терапевтически эффективных количеств дофаминергических нейронов, полученных из плюрипотентных стволовых клеток, предпочтительно из плюрипотентных стволовых клеток человека. В другом варианте осуществления настоящее изобретение относится к способам лечения нейродегенеративных расстройств и неврологических заболеваний, характеризующихся вырождением или разрушением серотонинергических нейронов, посредством трансплантации терапевтически эффективных количеств серотонинергических нейронов, полученных из плюрипотентных стволовых клеток, предпочтительно из плюрипотентных стволовых клеток человека. Предпочтительно, люди, страдающие нейродегенеративными расстройствами или неврологическими заболеваниями, получают лечение введением пациенту терапевтически эффективного количества нейрональных клеток-предшественниц и дифференцированных нервных клеток по настоящему изобретению. Как используется здесь, «терапевтически эффективное количество» клеток является количеством, достаточным для остановки или улучшения физиологических эффектов у индивидуума, вызванных потерей, повреждением или вырождением дифференцированных нервных клеток, таких так зрелые нейроны (например, дофаминергические и серотонинергические нейроны), астроциты и олигодендроциты.
Используемые терапевтически эффективные количества клеток будут зависеть от потребностей субъекта, его возраста, физиологического состояния и здоровья, необходимого терапевтического эффекта, размера области ткани, на которую направлено лечение, распространенности патологии и выбранного пути доставки. Например, лечение расстройства с поражением большой области мозга, возможно, потребует большее количество клеток для достижения терапевтического эффекта по сравнению с меньшей мишеневой областью. Клетки также могут быть введены в более чем одну область этой мишеневой ткани, с большим количеством небольших трансплантатов с низкой дозой клеток. Клетки по настоящему изобретению могут быть полностью диссоциированы перед трансплантацией таким образом, чтобы образовалась клеточная суспензия, или почти полностью диссоциированы перед трансплантацией таким образом, чтобы образовались небольшие агрегаты клеток. Клетки могут вводить способом, который позволяет им имплантироваться или мигрировать в нужное место ткани и реконструировать или регенерировать недостаточную функцию области.
Подходящее количество клеток, которое может быть введено для достижения терапевтического эффекта, может составлять примерно от 100 до примерно 1000000 нейронов, предпочтительно от примерно 500 до примерно 500000 нейронов или примерно от 1000 до примерно 100000 нейронов. Терапевтические концентрации нервных клеток, вводимых субъекту, также могут изменяться в пределах от примерно 10, 100, 500, 1000, 5000, 10000, 15000, 20000, 25000, 30000, 35000, 40000, 45000, 50000, 60000, 70000, 80000, 90000, 100000, 150000, 200000, 250000, 300000, 350000, 400000, 450000 до примерно 500000 клеток на микролитр фармацевтически доступного носителя. Диапазон концентрации клеток в носителе включает, например, 100-50000, 1000-10000, 5000-25000, 15000-45000, 20000-50000, 55000-200000, 100000-40000, 150000-50000 клеток/мкл и т.д. Количество клеток, введенных в область трансплантации, также влияет на терапевтическую эффективность.
Для терапевтического применения зачастую предпочтительно, чтобы популяция предшественниц или дифференцированных нервных клеток не содержала недифференцированных плюрипотентных стволовых клеток. Одной из стратегий для удаления плюрипотентных стволовых клеток из терапевтического препарата является трансфекция клеток вектором, который несет ген, который предпочтительно экспрессируется в недифференцированных клетках, экспрессия которого позволяет провести селекцию плюрипотентных стволовых клеток. Подходящими промоторами, которые предпочтительно экспрессируются в недифференцированных клетках, являются промотор обратной транскриптазы теломеразы (TERT) и промотор ОСТ-4. Ген, экспрессируемый в векторе, может быть, например, литическим для клеток, таким как токсин, или может быть выбрана обратная селекция с использованием внешнего агента.
Способность образовывать дофаминергические нейроны и серотонинергические нейроны из плюрипотентных стволовых клеток, как описано здесь, имеет большое клиническое значение для лечения методом трансплантации различных нейродегенеративных расстройств и неврологических заболеваний. Например, дофаминергические нейроны могут быть использованы для лечения нейродегенеративных расстройств и неврологических заболеваний, которые характеризуются палогией регуляции постуральных рефлексов, движения и поведения, с системой поощрения (центр удовольствия), например, болезнь Паркинсона, шизофрения и наркомания, а также поражение вследствие травмы или других заболеваний, которые приводят к симптомам болезни Паркинсона, таким как тремор в покое, ригидность, акинезия и патология постуральных рефлексов, такие как акинезия, адипсия, афагия и сенсорное нераспознавание. Кроме того, серотонинергические нейроны могут быть использованы для лечения нейродегенеративных расстройств и неврологических заболеваний, которые характеризуются патологией в регуляции приема пищи, секреции гормонов, ответа на стресс, боли и иммунной функции, сексуальной активности, сердечно-сосудистой функции и регуляции температуры, например, различные психиатрические, неврологические и другие заболевания, например, умственная депрессия, склонность к суициду, вспыльчивое агрессивное поведение, обессивно-компульсивное поведение и анорексия/булимия и шизофрения.
В другом варианте осуществления настоящее изобретение относится к одновременному введению одного или нескольких нейрональных факторов выживания с нейрональными клетками-предшественницами и дифференцированными нервными клетками по настоящему изобретению, полученных из плюрипотентных стволовых клеток для лечения нейродегенеративных расстройств или неврологических заболеваний. Нейрональный(е) фактор(ы) выживания могут быть введены перед, вместе, в сочетании с или после введения желаемых клеток. Как используется здесь, «нейрональный фактор выживания» является веществом, которое позволяет нейронам (либо in vitro или in vivo), которые контактируют с фактором, выживать в течение периода времени большего, чем могло бы произойти в отсутствие фактора. Нейрональные факторы выживания, которые могут быть использованы в терапевтическом варианте осуществления настоящего изобретения, включают, но ими не ограничиваются, глиальный нейротрофический фактор (GDNF), фактор роста нервов (NGF), цилиальный нейротрофический фактор (CNTF), мозговой нейротрофический фактор (BDNF), нейротофин-3 (NT-3), нейротофин-4 (NT-4), FGF, IL-1β, TNFα, инсулиноподобный фактор роста (IGF-1, IGF-2), трансформирующий фактор роста бета (TGF-β, TGF-β1).
Известно, что GDNF обладает трофической активностью для вентральных мезэнцефалических дофаминергических нейронов эмбрионального среднего мозга (Lin et al., 1993, Science 260:1130-1132; Lin et al., 1994, J. Neurochem. 63:758-768). Было также показано, что рекомбинантный человеческий GDNF индуцирует прорастание дофаминергических нитей in vivo (Hudson et al., 1993, Soc. Neurosci. Absir. 19:652), увеличивает оборот дофамина в черной субстанции крыс (Miller et al., 1994, Soc. Neitrosci. Abstr. 20:535-7), защищает нейроны от разрушения 6-OHDA и увеличивает рост и образование фибрилл эмбриональных трансплантатов черной ткани крыс in oculo (Stromberg et al., 1993, Exp. Neurol. 124:401-412). BDNF является трофическим фактором для периферических сенсорных нейронов, дофаминергических нейронов и ретинального ганглия (Henderson et al., 1993, Restor. Neurol. Neurosci. 5:15-28), и было показано, что он предотвращает происходящую в норме клеточную смерть in vitro и in vivo (Hofer и Barde, 1988, Nature 331:161-262).
Как используется здесь, термин «лечение» относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам. То есть лечение необходимо как для тех, кто страдает нейродегенеративным расстройством или неврологическим заболеванием, так и для тех, у кого нейродегенеративное расстройство или неврологическое заболевание следует предотвратить. Методы по настоящему изобретению могут быть использованы для лечения любых животных, которым необходимо такое лечение, включая, но ими не ограничиваясь, человека, приматов и домашний скот, фермерских животных, домашних или спортивных животных, таких как собаки, лошади, кошки, овцы, свиньи, коровы и тому подобное. «Расстройство» является любым состоянием, при котором будет наблюдаться улучшение в ходе лечения предшественницами нейрональных клеток, дифференцированными нервными клетками или обоими типами клеток настоящего изобретения. Примерами расстройств, при которых будет наблюдаться улучшение после введения дофаминергических нейронов, являются такие, которые связаны с неправильными постуральными рефлексами, движением и поведением, системой поощрения (цент удовольствия), такие как болезнь Паркинсона, шизофрения и наркомания. Примерами расстройств, которые можно лечить введением серотонинергических нейронов, являются такие расстройства, которые характеризуются аномальным сознанием, проявляется в пробуждении, поведении, аппетите, включая, но ограничиваясь агрессией, депрессией (включая суицидальное поведение), шизофренией и анорексией/булимией. Другими расстройствами, которые также можно лечить клетками настоящего изобретения, являются болезнь Альцгеймера, хорея Гентингтона и болезнь Гиршспрунга.
Способы по настоящему изобретению могут быть преимущественно осуществлены посредством прямой трансплантации нейрональных клеток-предшественниц или дифференцированных нервных клеток настоящего изобретения в поврежденную область. Способы нейрональной трансплантации и клеточной культуры хорошо известны специалистам в этой области, например, патент США № 5514552; Yurek и Sladek, 1990, Annu. Rev. Neurosci. 13:415-440; Rosenthal, 1998, Neuron 20:169-172; Vescovi et al., 1999, J. Neurotrauma 16 (8):689-93; Vescovi et al., 1999, Exp. Neuro. 156 (1):71-83; Brustle et al., 1999, Science 285:754-56; которые приведены здесь в качестве ссылки. В одном из вариантов осуществления дофаминергические нейроны по настоящему изобретению могут быть имплантированы в черную субстанцию или в полосатое тело пациента, страдающего болезнью Паркинсона. Могут быть введены только клетки или в сочетании с другими факторами, например, нейрональным фактором выживания, и могут быть введены с фармацевтически приемлемым носителем. В идеале такой носитель мог бы увеличивать стабильность и облегчать доставку клеток.
Настоящее изобретение также относится к фармацевтической композиции, содержащей клетки, которые могут быть введены с использованием подходящего носителя, такого как липосомы, микрочастицы или микрокапсулы. Клетки настоящего изобретения могут быть также доставлены в форме фармацевтической композиции, включающей изотонический эксипиент, полученной в условиях, достаточно стерильных для введения человеку. Основные принципы формирования медицинского состава клеточной композиции описаны в Cell Therapy: Stem Cell Transplantation, Gene Therapy, и Cellular Immunotherapy, G. Morstyn & W. Sheridan eds, Cambrigge University Press, 1996, и Hematopoietic Stem Cell Therapy, E. Ball, J. Lister & P. Law, Churchill Livingstone, 2000, которые приведены здесь в качестве ссылки. Кроме того, может быть желательно вводить фармацевтическую композицию, содержащую нейрональный фактор выживания вблизи области, нуждающейся в лечении, которая может быть достигнута, например, местным введением во время операции, инъекцией, с помощью катерера или пересаживанием, где такой имплантат может быть с порами, без пор или из гелеобразного материала, включая мембраны, такие как силастическая мембрана или волокна.
Нейрональные клетки-предшественницы и дифференцированные нервные клетки по настоящему изобретению могут быть трансплантированы в субъект либо в виде по существу гомогенной, почти гомогенной или гетерогенной клеточной популяции. Значительная гомогенность клеточной популяции включает больше 75% клеток одного типа, таких как дофаминергических или серотонинергических нейронов, более предпочтительно больше 90% и наиболее предпочтительно больше 95-99%. Гетерогенная клеточная популяция может состоять из двух или нескольких типов клеток, смешанных в одной клеточной популяции, например, дофаминергические нейроны, серотонинергические нейроны, клетки Шванна, олигодендроциты, астроциты, ГАМК нейроны и глиальные клетки. Клетки также могут быть генетически измененными методами, хорошо известными специалистам в данной области, используемыми для экспрессии или высвобождения трофических факторов, факторов роста, нейрональных факторов выживания или других терапевтических соединений в поврежденной области мозга, центральной нервной системы, периферической нервной системы или других тканей. Использование сочетаний промоторов и клеточных типов для экспрессии белка общеизвестны специалистам в области молекулярной биологии, например, см. Sambrook, et al. 1989, Molecular Cloning: A Laboratory Manual 2nd Ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, конкретно приведена здесь в качестве ссылки.
Для осуществления экспрессии трофических факторов, факторов роста, нейрональных факторов выживания или других терапевтических соединений в нейрональных клетках-предшественницах и дифференцированных нервных клеток по настоящему изобретению, подходящие регуляторные элементы могут быть получены из различных источников и могут быть легко выбраны специалистом в данной области. Примеры регуляторных элементов включают транскрипционный промотор, энхансер и последовательность для связывания РНК-полимеразы, а также последовательность связывания рибосомы, включая инициирующий сигнал трансляции. Также в рекомбинантную молекулу могут быть введены другие добавочные генетические элементы, такие как селективные маркеры. Рекомбинантная молекула может быть введена в плюрипотентную стволовую клетку, или нейрональную клетку-предшественницу, или в дифференцированную нервную клетку, полученную из плюрипотентной стволовой клетки, с помощью векторов доставки in vitro или методов in vivo. Примеры способов доставки включают ретровирусные векторы, аденовирусные векторы, ДНК вирусные векторы, липосомы, физические методы, такие как микроинъекция и трансфекция посредством электропорации или осаждение с помощью фосфата кальция, или другими известными методами для трансфекции и получения рекомбинантных клеток. Генетически измененные клетки могут быть инкапсулированы в микросферы и имплантированы в или близлежащую область больной или поврежденной ткани. Используемые методики хорошо известны специалистам и могут быть найдены, например, в Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, New York, N.Y., 1997, приведена здесь в качестве ссылки.
Клинические эксперименты, включающие нейрональную трансплантацию людям, страдающим болезнью Паркинсона, проводили от основополагающих научных исследований с использованием различных моделей паркинсонизма у животных в качестве реципиентов либо зародышевых эмбриональных нервных клеток или трансплантатов паранейрональных тканей в поврежденную область мозга. Эксперимент на животных с трансплантатами зародышевых дофаминергических нервных клеток дал основание полагать, что такие трансплантаты могут уменьшить дофаминергический дефицит и восстановить моторную функцию у животных с разрушенной в эксперименте нигростриальной дофаминергической системой. Например, мезэнцефалические клетки мышей, взятые из 14-16-дневного эмбриона, были с успехом использованы при трансплантации в качестве гомотрансплантатов или гетеротрансплантатов без иммуносупрессивного лечения по сравнению с клетками, взятыми у более поздних беременных доноров (Yurek и Sladek, 1990, Annu. Rev. Neurosci. 13:415-440). Интересно, что этот возраст соответствуют возрасту беременности, при котором дофаминергические нейроны подвергаются заключительным стадиям клеточного деления (Lauder и Bloom, 1974, J. Comp. Neurol. 155:469-82). Твердые трансплантаты эмбриональной мезэнцефалической ткани могут также быть диссоциированы в клеточную суспензию для трансплантации. Обычная вероятность выживания такой суспензии, однако, ограничена примерно 10% пересаженных дофаминергических клеток (Brudin et al., 1987, Ann. N.Y. Acad. Sci. 495:473-96). Поскольку эмбриональная мезэнцефалическая ткань не является чистым источником дофаминергических клеток, только около 0,1-1,0% всех жизнеспособных клеток являются выжившими дофаминергическими клетками, предпочтительно, любой трансплантат диссоциированных мезэнцефалических клеток содержит минимум от 100000 до 150000 жизнеспособных клеток для эффективного компенсирования недостатка дофамина.
Предпочтительно, трансплантационная терапия по настоящему изобретению также включает некоторые средства хранения и сохранения нейрональных клеток-предшественниц и дифференцированных нервных клеток для использования при трансплантации, например, хранение в течение длительного времени при криоконсервации или в течение небольшого времени в среде для хранения. Эмбриональную мезэнцефалическую ткань успешно хранили в жидком азоте в течение 70 дней и пересаживали в виде гомотрансплантата грызунам (Collier et al., Progress in Brain Research, Vol. 78, New York, Elsevier (1988), pp. 631-36, которые приведены здесь в качестве ссылки) и приматам (Collier et al., 1987, Brain Res. 436:363-66, которые приведены здесь в качестве ссылки). Было также показано, что эмбриональные мезэнцефалические клетки могут быть успешно культивированы после криоконсервации. Мезэнцефалическая ткань можно также хранить в течение короткого промежутка времени (2-5 дней) в среде для хранения при 4°С и затем пересаживать, при этом объем выжившей части трансплантата сходен с объемом для свежей ткани (Sauer et al., 1989, Restor. Neurol. Neurosci. (Suppl.:3.sup.rd Int. Symp. Neural Tranplan.):56, которые приведены здесь в качестве ссылки).
Размещение трансплантата и степень, с которой трансплантат реиннервирует полосатое тело, являются главными факторами для функционального восстановления мезостриальной дофаминергической системы. Было показано, что у односторонне дофаминергических денервированных животных кортикальное размещение дофаминергического трансплантата уменьшает моторную асимметрию, но оказывает небольшой эффект на сенсорное игнорирование (Bjorklund et al., 1980, Brain Res. 199:307-33; Dunnett et al., 1981, Brain Res. 215:147-61). С другой стороны, трансплантат черной субстанции, помещенный вблизи латерального полосатого тела у билатерально дофаминергических денервированных животных, эффективно восстанавливает сенсорные повреждения (Dunnett et al., 1983, Acta Physiol. Scan. Suppl. 522:39-47). Более того, прекращение акинезии у билатерально разрушенных животных наблюдалось только тогда, когда прилежащее ядро реиннервировалось дофаминергическим трансплантатом (Nadaud et al., 1984, Brain Res. 304:137-41).
Несмотря на то, что место трансплантации для дофаминергического трансплантата наиболее часто было вблизи вентрикулярной области, из-за благоприятного окружения цереброспинальной жидкости (CSF) в этой области для обеспечения выживания трансплантата, степень дофаминергической дегенерации при болезни Паркинсона более значительно была в скорлупе, чем в хвостатом ядре. Уровень дофамина в скорлупе был зачастую на 10-15% меньше, чем в хвостатом ядре (Bemhiemer et al., J. Neurol. Sci. 20:415-55; Nyberg et al., 1983, Neurochem. Pathol. 1:93-202). Кроме того, каузальная часть скорлупы более значительно обеднена дофаминергическими нейронами, чем передняя часть скорлупы (Kish et al., 1986, Ann. Neurol. 20:26-31). Из двух компонентов полосатого ядра (хвостатое ядро и скорлупа) скорлупа получает большинство моторных сигналов посредством кортико-таламическо-путаментного пути (DeLong & Georgopoulos, 1983, Handbook of Physiology, Section I: The Nervous System, Vol.2, ed. Brookhard, Mountcastle, Geiger, pp. 1017-61, Bethesda, Md.: Am. Physiol. Soc.). Поэтому скорлупа может быть более благоприятным местом для дофаминергической трансплантации, направленной на восстановление моторного расстройства, связанного с болезнью Паркинсона.
Нейрональные клетки-предшественницы и дифференцированные нервные клетки, описанные здесь, могут быть также использованы для поиска соединений, таких как фармацевтические соединения, растворители, небольшие молекулы, пептиды или полинуклеотиды, а также факторов окружающей среды, таких как условия культивирования или манипуляции, которые затрагивают фенотип или свойства этих клеток. Кроме того, эти клетки могут быть использованы для оценки возможных факторов роста или факторов дифференцировки. Например, возможное фармацевтическое соединение может быть добавлено к нейрональным клеткам-предшественницам или зрелым нейронам, либо одно, либо в сочетании с другими лекарствами, и любые изменения в морфологии, фенотипе или функциональной активности в этих клетках можно оценить и измерить.
Кроме того, нейрональные клетки-предшественницы и дифференцированные нервные клетки, описанные здесь, могут быть в дальнейшем модифицированы на любой стадии дифференцировки. Например, эти клетки могут быть генетически модифицированы для однократной или множественных генетических изменений, либо в переходной, либо в стабильной стадии. Генетические изменения этих клеток могут быть необходимы по многим причинам, таким как модификация клеток для генной терапии или замены тканей для пересадки или имплантации. Клетки по настоящему изобретению могут быть генетически модифицированы посредством введения экспрессионных вектор селективных маркеров под контролем нейрального специфического промотора, которые хорошо известны специалистам. Эти клетки также могут быть модифицированы на любой стадии для экспрессии определенных маркеров или генов, которые могут быть использованы для дальнейшей очистки дифференцированных клеток, полученных из плюрипотентных стволовых клеток, или с другой стороны, для индукции дифференцировки по определенной клеточной линии. Эти клетки могут быть модифицированы для уменьшения или предотвращения иммунного отторжения после трансплантации, например, гистосовместимость с предназначенным реципиентом.
Для увеличения репликативной способности клеток, полученных с использованием настоящего изобретения, эти клетки могут теломеризированы посредством из генетического изменения подходящим вектором, таким образом, чтобы они экспрессировали каталитический компонент теломеразы (TERT). Используемая последовательность TERT может быть получена у человека или мыши (WO 98/14592 и WO 99/27113, приведено здесь в качестве ссылки), а также других видов млекопитающих. С другой стороны, транскрипция эндогенного гена TERT может быть повышена. Способы, используемые для генетической модификации клеток, хорошо известны специалистам в данной области. В этих способах используют различные методики молекулярной биологии, многие из которых, в целом, описаны в Sambrook, et al. 1989, Molecular Cloning: A Laboratory Manual 2nd Ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, которое приведено здесь в качестве ссылки.
Следующие примеры включены для демонстрации предпочтительного варианта осуществления изобретения. Специалистам будет понятно, что способы, представленные в примерах, которые следуют за представленными методами, были разработаны с целью повышения эффективности в практике настоящего изобретения и, таким образом, могут рассматриваться как предпочтительные варианты осуществления на практике. Однако специалистам понятно, что в свете настоящего изобретения могут быть сделаны другие изменения в конкретных вариантах осуществления, которые известны, но позволяют получить сходный или аналогичный результат, не выходя за рамки области изобретения и не изменяя его сущности.
Пример 1
В нижеследующем примере авторы настоящего изобретения показывают получение функциональных дофаминергических и серотонинергических нейронов из эмбриональных стволовых клеток человека.
1) Эмбриональные стволовые клетки человека
Сначала ЭС клетки человека выделяли из внутренней массы бластоцисты человека. Плюрипотентные ЭС клетки человека получали из бластоцист человека после получения согласия пациентов для экспериментального использования бластоцист. Линию ЭС клеток человека, используемую здесь, получали из внутренней клеточной массы бластоцисты человека с использованием нового метода лазерного разрушения, как описано в патенте США №10/798790, которое приведено здесь в качестве ссылки. Кратко, выделяли бластоцист человека с zona pellucida, троходермой и внутренней клеточной массой, и 1,48 мкм не-контактный диодный лазер использовался для создания отверстия в zona pellucida и троходерме бластоциста человека. Затем выделяли клетки внутренней клеточной массы с использованием аспираторной пипетки, введенной через отверстие. Затем эти клетки культивировали на 0,1% желатинизированной чашке с обработанными митомуцином-С питающими клетками мыши в ЭС среде, и образовывалась клеточная масса, полученная из внутренней клеточной массы. ЭС среда для клеток состояла из модифицированной среды Дульбекко-Игл (DMEM) или "нокаут" DMEM (Gibco), содержащая 10-20% пригодной для ЭС клеток фетальную сыворотку теленка (FCS) (Hyclone) или "нокаутная" сыворотка с заменителем сыворотки (Gibco), 1% раствора заменимых аминокислот MEM, 2 мМ L-глутамина, 0,1 мМ (5-мерхаптоэтанола, 4 мкг/мл основного фактора роста фибробластов (bFGF) (Sigma), 50 Ед/мл пенициллина и 50 мкг/мл стрептомицина. Эту массу, полученную из внутренней клеточной массы, диссоциировали, высевали на слой мышиных питающих клеток в ЭС среде и использовали для получения линий ЭС клеток человека.
Полученные ЭС клетки человека морфологически имеют высокое отношение ядро/цитоплазма и были охарактеризованы клеточными маркерами, такими как SSEA-1, SSEA-3, SSEA-4, TRA-1-60, TRA-1-81, ОСТ-4, щелочная фосфатаза, теломераза, кариотипирование, CD-ЗС, cripto-1, GCNF, c-kit и CD-90.
2) Культивирование и размножение эмбриональных стволовых клеток человека
ЭС клетки человека культивировали и пролиферировали при стандартных условиях роста из замороженного основного раствора. Недифференцированные ЭС клетки человека из замороженной пробирки ресуспендировали в ЭС среде с LIF. Среда для ЭС клеток состояла из модифицированной среды Дульбекко-Игл (DMEM), содержащей 20% пригодной для ЭС клеток фетальной сыворотки теленка (FCS) (Hyclone)), 1% раствор заменимых аминокислот (NEAA), 2 мМ L-глутамина, 0,1 мМ β-меркаптоэтанола, 50 Ед/мл пенициллина и 1000 Ед/мл LIF. Клетки осаждали и высаживали на слои мышиных питающих клеток на покрытой желатиной пластиковой культуральной плашке с использованием ЭС среды. Среда для ЭС клеток включала DMEM с высоким содержанием глюкозы, содержащая 10-20% FCS, 2 мМ L-глутамина, 1% раствор МЕМ заменимых аминокислот, 0,1 мМ β-меркаптоэтанола, 50 Ед/мл пенициллина, 50 мкг/мл стрептомицина, 1000 Ед/мл LIF и 4 мкг/мл bFGF в соответствии с методом, описанным Thomson et al., 1998, Science 282:1145-1147; Shamblott et al., 1998. Proc. Natl. Acad. Sci. USA. 95: 13726-13731; и Reubinoff et al., 2000. Nature Biotech. 18:399-403.
ЭС клетки человека размножили посредством культивирования и регулярных высеваний ЭС клеток для ингибирования дифференцировки. Вначале ЭС клетки пересадили промывкой клеток фосфатным буфером Dulbecco без Са++, Mg++ в растворе салина в течение 10 секунд и затем обработали клетки 0,05% трипсином в течение 5 секунд. После 5 секунд активность трипсина ингибировали добавлением ЭС среды, содержащей сыворотку. Затем клетки соскребали и разрушали на маленькие кластеры. Кластеры затем помещали на две 0,1% 100 мм чашки Петри, покрытые желатином и питающими клетками в ЭС среде с LIF и bFGF, как описано выше. Клетки размножались в течение 4-6 дней.
3) Образование эмбриоидных тел
После пролиферации и роста недифференцированных ЭС клеток человека их культивировали для образования эмбриоидных тел. Вначале ЭС клетки диссоциировали с использованием 0,05% трипсина-ЭДТА, затем клетки соскабливали и разрушали на небольшие кластеры. Затем эти кластеры высевали на бактериологическую чашку с плотностью примерно 1·105 клеток/мл в отсутствии питающих клеток. Используемые бактериологические чашки не обладали адгезивной поверхностью, что предотвращало прикрепление, и таким образом стимулировались дифференцировка ЭС клеток и образование эмбриоидных тел. Эти клетки культивировали в виде суспензионной культуры в ЭС среде без LIF и bFGF. ЭС среда, использованная для культивирования этих клеток, была DMEM с высоким содержанием глюкозы или "нокаут" DMEM, в котором содержались 10-20% FCS или "нокаутная" сывороточная замена, а также другие добавки, такие как β-меркаптоэтанол, L-глутамин и антибиотики. К ЭС среде bFGF не добавляли. Эмбриональные тела росли в течение 4-8 дней. В течение этого времени ЭС среду меняли каждые 2 дня с помощью метода седиментации. Этот седиментационный метод проводили помещением суспензии агрегатов в центрифужную пробирку, агрегаты осаждали на дно пробирки, удаляли среду и заменяли ее свежей средой. Затем агрегаты в свежей среде возвращали в культуральные плашки. В конце 4-8 дней эмбриоидные тела собирали и откручивали при низкой скорости и ресуспендировали в среде для ЭС клеток. Примерно 20-30 эмбриоидных тел затем высевали на тканевую культуру, покрытую 0,1% желатины в ЭС среде без LIF, и инкубировали в течение 24 часов.
4) Селекция и рост нестин-положительных нейрональных клеток-предшественниц
После 24 часов нестин-положительные клетки (нейрональные клетки-предшественницы) собирали посредством замены ЭС среды ITSFn (нестиновая селекция) без сывороточной определенной среды. ITSFn среда, состоящая из DMEM:F12 среды (Gibco), содержащая факторы роста инсулин (2-25 мкг/мл) (Sigma), селенит натрия (10-50 нМ) (Sigma), трансферрин (1-10 мкг/мл) (Gibco) и фибронектин (1-5 мкг/мл). Эта среда позволяет отобрать нестин-положительные клетки в течение 6-10 дней, обычно 8-9 дней, с заменой ITSFn среды каждые 2 дня. После полной селекции нейрональные клетки-предшественницы были охарактеризованы на экспрессию нестина с использованием иммунофлуоресцентного метода, который показал, что примерно 95% клеток экспрессируют нестин (Фигура 1А).
Нестин-положительные клетки затем размножали или сортировали с использованием магнитной клеточной сортировки (MACS) для обогащения положительными клетками на адгезивные молекулы нервных клеток (NCAM).
5) Сортировка NCAM-положительных клеток с помощью магнитной сортировки
Затем нестин-положительные клетки выделяли и размножали или подвергали магнитной клеточной сортировке (MACS) для обогащения положительными клетками на адгезивные молекулы нервных клеток (NCAM). NCAM является специфичным для нейронов клеточным маркером. Для сортировки нестин-положительных клеток посредством MACS с использованием NCAM нестин-положительные клетки вначале собирали из чашки с культурой ткани инкубированием клеток с 0,05% трипсин-ЭДТА. Затем диссоциированные живые клетки промывали PBS и окрашивали антителом на клеточный маркер NCAM (Chemicon) в течение 30 минут. Затем клетки инкубировали с козлиными антикроличьими IgG MACS микрошариками (Miltenyi Biotec) в течение 15 минут. Клетки подвергали магнитной сортировке после осторожной промывки PBS. Для сортировки клеток суспензия с помеченными магнитами клетками (предположительно 2·108 клеток) пипетировали в MACS MS Разделительной Колонке (Miltenyi Biotec), клетки протекали через колонку и эффлюент (негативная фракция) собирали. Позитивную фракцию с NCAM-положительными клетками собирали затем с помощью элюции с PBS.
До иммуносортировки нестин-положительные клетки проанализировали с помощью FACS и около 50-60% нестин-положительных клеток экспрессировали нейрональный клеточный маркер NCAM (Фигура 2А и 2В). После иммуносортировки с помощью MACS популяцию жизнеспособных нестин-положительных клеток, экспрессирующую NCAM, обогащали примерно до 80-85% (Фигура 3А и 3В).
6) Рост NCAM-положительных нейрональных клеток-предшественниц
Выделенные NCAM-положительные клетки размножили вначале посредством диссоциации клеток в 0,05% трипсин-ЭДТА и затем высевали клетки на плашку, покрытую поли-L-орнитин/ламинином, содержащую среду для размножения. Культивирование NCAM-положительных клеток, таким образом, позволяет лучшей адгезии и пролиферации в среде для роста. Среда для роста включала DMEM:F12, инсулин (10-100 мкг/мл), селенит натрия (10-50 нМ), трансферрин (1-10 мг/мл), путресцин (50-200 мкМ), прогестерон (5-40 нМ) и ламинин (10-50 мкг/мл). Два или более фактора роста, такие как bFGF (10-50 нг/мл), EGF (10-50 нг/мл), BDNF (50-200 нг/мл), FGF-8 (50-200 нг/мл) и/или SHH (200-400 нг/мл), также включали в среду для роста без сыворотки. Эти различные факторы, присутствующие в среде для роста, способствуют общему увеличению процента нейрональных клеток и еще более способствуют приобретению нейрональными клетками-предшественницами среднего мозга дофаминергического фенотипа. Среду для роста заменяли каждые 2 дня и нестин-положительные клетки, NCAM-положительные клетки пролиферировали в течение 6-10 дней (Фигура 1В).
При таких условиях культивирования примерно 50-60% клеток усиливали нейритные процессы и окрашивались положительно по NCAM, что доказывает их способность образовывать нейроны (Фигура 3). Эти клетки также были иммунореактивными к раннему нейрональному маркеру β-тубулину (Фигура 4В). Эти нейрональные клетки-предшественницы затем последовательно пассировали, и ученые наблюдали, что эти нейрональные клетки-предшественницы можно пассировать до 10 раз, и они все еще сохраняли способность дифференцироваться в дофаминергические нейроны, без заметного влияния на потенциал этих клеток дифференцироваться в зрелые дофаминергические нейроны.
7) Дифференцирование и созревание дофаминергических нейронов
Развитие дофаминергических нейронов in vivo зависит от регуляторных действий внеклеточных сигнальных молекул. Эти молекулы активируют каскад факторов транскрипции с экспрессией в зависимости от происхождения. Эти транскрипционные факторы ответственны за увеличение специфического набора генов, вовлеченных в развитие дофаминергического фенотипа. Для стимулирования превращения выросших нейрональных клеток-предшественниц, полученных с использованием приведенного выше протокола, в функциональные дофаминергические нейроны факторы роста bFGF и EGF, во-первых, были удалены из среды с клетками. Затем дифференцировка нейрональных клеток-предшественниц в дофаминергические нейроны была индуцирована посредством культивирования клеток в нейробазальной среде в течение 30-50 дней. Нейробазальная среда включала нейробазальную среду А (Gibco), FCS (10-20%) (HYCLONE) и B27 (2-10%) (Gibco), а также различные факторы роста. Факторами роста, включенными в эту среду, были интерлейкин-1β (IL-lβ) (1-2 мкг/мл) (Sigma), аскорбиновая кислота (50-150 мМ) (Sigma), N-ацетилцистеин (50-150 нМ) (ICN), db-cAMP (500-1000 мкМ) (Sigma), GDNF (1-5 мкг/мл) (Sigma), TGF-β3 (1-5 мкг/мл) (Sigma), нейртурин (100-500 мкг/мл) (Chemicon), BDNF (50-200 нг/мл) (Sigma), FGF-8 (50-200 нг/мл) (R&D Systems) и/или SHH (200-400 нг/мл) (R&D Systems). Нейробазальную среду простерилизовали с помощью фильтрации с 0,22 мкмиллипоры шприцовым фильтром.
Тирозингидроксилаза (ТН) является ферментом, лимитирующим синтез дофамина. Факторы роста некоторых линий способствуют индукции синтеза ТН. Добавление этих факторов в среду приводит к увеличению процента клеток, принимающих дофаминергический фенотип. Эти факторы добавляли в различные временные точки в течение нейрональной дифференцировки, как указано в Таблице 1. IL-1β, которое присутствует в нейробазальной среде, по-видимому, играет ключевую роль в дифференцировке нейрональных предшественниц в дофаминергические нейроны. Хотя какого-то определенного механизма нет, предполагают, что IL-1β увеличивает количество ТН-положительных нейронов, а также ответ клеток на сигнальные молекулы в среде. Использование этих факторов роста комбинировали с заменой нейробазальной среды каждые 3 дня во время культивирования ткани в течение 30-50 дней.
Коктейль факторов роста в нейробазальной среде, добавленных в различные дни в течение дифференцировки нейрональных клеток-предшественниц, приведены в Таблице 1.
Когда ЭС клетки индуцируются для дифференцировки, используя нейробазальную среду, ЭС клетки развиваются в различные типы нейрональных клеток, включая дофаминергические нейроны, серотонинергические нейроны и олигодендроциты (Фигуры 5, 6 и 7). Тогда как процент клеток, которые дифференцируются в дофаминергические нейроны, остается относительно небольшим в стандартных условиях культивирования, добавление в среду факторов роста, как показано в Таблице 1, приводит к увеличению процента получающихся дофаминергических нейронов. Например, более 60% ЭС клеток дифференцируется в клетки, которые являлись положительными для ТН, - специфический маркер для дофаминергических нейронов.
8) Характеризация дофаминергических нейронов
Как общая морфология клеток, так и фенотип, определенный с помощью иммунофлуоресценции, были оценены для дифференцированных типов нейрональных клеток, полученных в соответствии с настоящим изобретением. Иммунофлуоресцентный анализ проводили на стадии роста нейрональных предшественниц (рост NCAM положительных клеток) и на различных других временных точках дифференцировки, используя стандартные методы, хорошо известные специалистам. Вначале выделенные клетки подращивали в 2-луночной плашке, предварительно покрытой внеклеточным матриксом, промывали PBS и фиксировали в течение 10 минут 4% параформальдегидом при комнатной температуре. Затем увеличивали проницаемость клеточной мембраны клеток 0,2% Тритоном Х-100 в течение 5 минут, блокировали 1% бычьим сывороточным альбумином (БСА)/PBS в течение 2 часов и инкубировали с главным антителом (разбавление антитела было проведено в 1% БСА/TBS) в течение ночи при 4°С. Клетки окрашивали следующими главными антителами: ранний нейрональный маркер β-тубулин, NCAM, нейрофиламент, поздний нейрональный маркер Белок, ассоциированный с микротубулином 2 (MAP-2), поверхностный антиген А2В5 нейронной клетки, Nurr-1, тирозингидроксилаза, дофаминовый транспортер DAT, дофамин β-гидроксилаза (DBH), маркер астроцитов глиального фибриллярного кислого белка (GFAP), ГАМК, олигодендроцит, серотонин и синаптофизин, все были получены в CHEMICON. Наконец, клетки инкубировали с FITC мечеными вторичными антителами. После каждого из указанных шагов клетки промыли три раза PBS.
Плашки рассматривали под флуоресцентным микроскопом для оценки иммуноположительных областей. Иммунофлуоресцентный анализ показал, что большой процент дифференцированных клеток были иммунореактивными к специфическому маркеру нейронов NCAM (Фигура 3), МАР-2 (Фигура 4А) и β-тубулину (Фигура 4В). Экспрессия этих ключевых антигенов увеличивается с увеличением времени инкубации в среде для дифференцировки. Иммунофлуоресцентный анализ также показал, что меньший процент клеток экспрессировал серотонин (Фигура 7), а также не нейрональный маркер глиального фибриллярного кислого белка (GFAP), который присутствует в астроцитах, и ГАМК и глутамата, которые присутствуют в олигодендроцитах (Фигура 6).
Дифференцированные клетки также проанализировали с помощью двойного мечения клеток главными антителами к ТН (зеленые), а также к МАР-2, DAT, Nurr-1, β-тубулину, FITC-меченными вторичными антителами (Техасский красный), соответственно (Фигура 8). Этот анализ показал, что часть МАР-2 положительных клеток, экспрессирующих ТН, была больше. Клеточная плотность дофаминергических нейронов оценили подсчетом количества ТН-положительных клеток и МАР-2 положительных клеток на область в случайно выбранной области с использованием 40х объективной линзы. Процентная часть ТН положительных клеток была вычислена (Фигура 9). ТН положительные нейроны также экспрессировали Nurr-1 и DAT, но не было со-локализации ТН с DBH, который подтверждает дофаминергический фенотип клеток. Кроме того, образование синапсов определяли с помощью иммуноокрашивания с синаптофизином.
Иммунофлуоресцентный анализ также показал, что процент сортированных NCAM-положительных клеток, экспрессирующих ТН, составляет приблизительно 60%, тогда как около 30% экспрессируют серотонин (Фигура 10). Кроме того, около 40% нестин-положительных клеток окрашивались положительно на ТН, около 30% окрашивались положительно на серотонин, и около 28% окрашивались положительно на олигодендроцит (Фигура 11). Иммунологические маркеры, используемые на различных стадиях дифференцировки, показаны в Таблице 3 и указаны фенотипические характеристики недифференцированных и дифференцированных ЭС клеток.
9) Профиль генной экспрессии
Анализировали также профиль генной экспрессии клеток, собранных на разных стадиях дифференцировки. Клетки собрали для анализа с помощью Полимеразной Цепной Реакции с Обратной Транскриптазой (RT-PCR) с каждой из следующих стадий описанного метода: недифференцированные ЭС клетки, эмбриоидные тела, нестин-положительные нейрональные клетки-предшественницы, NCAM-положительные клетки и дифференцированные клетки, выделенные после 12, 17, 22, 27 и 37 дня дифференцировки в нейробазальной среде. После сбора клеток они были осажены, и из них выделили тотальную клеточную РНК с использованием набора RNeasy Qiagen. Выделенную РНК хранили при -20°С.
кДНК синтезировалась из выделенной тотальной РНК с использованием обратной транскриптазы вируса Moloney Leukemia superscript II и олиго (dT)12-18. Синтезированную с помощью реакции с обратной транскриптазой кДНК использовали для ПЦР амплификации с различными наборами специфических праймеров для определения, какие гены экспрессировались в собранных клетках. ПЦР проводилась с использованием кДНК в качестве матрицы и Taq полимеразы, соединенной с платиной в стандартных условиях для ПЦР, которые хорошо известны специалистам. Общими циклическими параметрами, используемыми для амплификации ДНК продуктов, были следующие:
1. денатурация матрицы кДНК при 94°С в течение 30 секунд;
2. отжиг праймеров при 55-56°С в течение 1 минуты, в зависимости от используемых праймеров; и
3. инкубация реакции при 72°С в течение 1 минуты и повтор шагов 1-3 (циклы) от 25 до 40 раз.
После проведения ПЦР продукты разделяли в 1,5% агарозном геле с помощью электрофореза с маркерами размера ДНК. Экспрессию ТН, D2RL, DBH, En-1, Nurr-1 и β-тубулина анализировали с помощью RT-PCR с использованием праймеров, приведенных в Таблице 4.
Выше приведенный анализ с помощью RT-PCR показал, что экспрессия специфических для дофаминергического фенотипа гена - ТН наблюдалась после нейрональной дифференцировки ЭС клеток до терминальной дифференцировки в дофаминергические нейроны (Фигура 12). Как и ожидалось, β-тубулин - повсеместно экспрессируемый ген, наблюдался во всех образцах, тогда как DHB не экспрессировался ни в одном образце.
10) Определение дофамина на ВЭЖХ с обращенной фазой
Одной определенной характеристикой дофаминергического нейрона является продукция дофамина. Поэтому функциональная способность дофаминергических нейронов, полученных из эмбриональных стволовых клеток, продуцировать дофамин была оценена с помощью прямого измерения внутриклеточного уровня дофамина с использованием ВЭЖХ с обращенной фазой (ОФ-ВЭЖХ). Концентрацию дофамина, определенную в каждом образце, определяли с помощью сравнения со стандартным раствором дофамина, наносимого на колонку сразу до и после каждого эксперимента.
Для начала клетки собирали на различных стадиях описанного метода: недифференцированные ЭС клетки, эмбриоидные тела, нестин-положительные нейрональные клетки-предшественницы, NCAM-положительные клетки и дифференцированные клетки, выделенные после 7, 22 и 37 дня дифференцировки в нейробазальной среде. До сбора клетки, дифференцированные в нейробазальной среде, вначале стимулировали для индукции секреции дофамина посредством добавления 56 мМ KCl в HBSS в течение 15 минут. Приблизительно 5·106 клеток трипсинизировали и собирали с помощью центрифугирования. Затем клетки озвучили с помощью ультразвука в холодной 1N перхлорной кислоте с антиоксидантами (0,2 г/л пиросульфита натрия) и центрифугировали при 15000 об/мин 20 минут при 4°С. Супернатант отбирали и хранили при -70°С для последующего определения внутриклеточной концентрации дофамина с помощью ОФ-ВЭЖХ. Уровень дофамина определяли в клеточном лизате и культуральном супернантанте (48 часов после последней смены среды). Культуральный супернантант немедленно стабилизировали 7,5% ортофосфорной кислотой и пиросульфитом натрия.
Клеточный лизат анализировали на ОФ-ВЭЖХ с ранних стадий описанного метода, таких как недифференцированные ЭС клетки, эмбриоидные тела, нестин-положительные нейрональные клетки-предшественницы и NCAM-положительные клетки не содержали каких-либо детектируемых уровней дофамина. Однако после первой недели дифференцировки в нейробазальной среде все клеточные лизаты, анализируемые на ОФ-ВЭЖХ, содержали дофамин. Уровень внутриклеточного дофамина значительно вырос с увеличением количества дофаминергических нейронов в клеточном лизате и терминированных зрелых за время в нейробазальной среде для дифференцировки. Внутриклеточный уровень дофамина был даже выше в клеточной культуре, которая была обработана факторами роста по сравнению с необработанной культурой в указанные временные точки, как показано на Фигуре 13. Дальним подтверждением продукции дофамина этими клетками в культуре было показано после инкубации клеточной культуры с N-метил-4-фенил 1,2,3,6-тетрагидропиридин гидрохлоридом (МРТР) - нейротоксином, который поражает дофаминергические нейроны. В клеточных культурах, инкубированных с МРТР, дофамин в клеточном лизате не был детектирован с помощью ОФ-ВЭЖХ. С другой стороны, высвобождение дофамина в кондиционированную среду было увеличено с помощью стимуляции 56 мМ KCl (Фигура 14). Добавление набора факторов роста в нейробазальную среду для дифференцировки также увеличивало внутриклеточный уровень дофамина в клеточном лизате (Таблица 5).
Изобретатели также показали возможное применение дофаминергических нейронов, полученных из эмбриональных стволовых клеток человека (чЭС), для терапевтического использования при болезни Паркинсона (БП). Была исследована in vivo функциональность данных нейрональных клеток на модели болезни Паркинсона у крыс.
Модель болезни Паркинсона у крыс
Все исследования на животных были выполнены в соответствии с CPCSEA (Committee for purpose of control and supervision of experimental animals) и были одобрены Institutional Animals Ethics Committee. Модель болезни Паркинсона у крыс была создана согласно Ravindran and Rao, 2006(27). Для создания модели болезни Паркинсона у животных крысам путем инъекции вводили 6-OHDA и наблюдали в течение 4-6 недель. Крысы, получавшие 6-OHDA и проявляющие ипсилатеральное вращательное поведение, которые двигались по кругу более 20 раз в минуту в одном и том же направлении, были отобраны как крысы с индуцированной болезнью Паркинсона, которых использовали для дальнейшего исследования. Крысы, получавшие 6-OHDA, представляют собой наиболее часто используемую модель паркинсонизма, вероятно, потому, что введение 6-OHDA обычно способствует асимметричным и количественно измеримым патологиям двигательного поведения. Большинство крыс, получавших 6-OHDA, проявляли ипсилатеральное движение по кругу, тогда как крысы, получавшие физиологический раствор, не проявляли подобного поведения.
Трансплантация нейрональных клеток
Для эффективности исследования БП-индуцированных крыс, у которых наблюдалась значительная ипсилатеральная вращательная ответная реакция, распределяли на экспериментальную и контрольную группы. В экспериментальной группе в субстанцию нигра каждой крысы вводили приблизительно 1,2 миллиона нейрональных клеток, используя шприц с размером иглы 28 со скоростью 2 мл/мин. Трансплантацию клеток осуществляли, используя механический микроинъектор (Stoelting Co., USA). Группу контроля подвергли той же самой процедуре, за исключением того, что животным вводили 10 мл среды N2. Каждой крысе делали инъекцию 15 мг/кг циклоспорина A (Sigma) за 24 часа перед и в течение до 1 недели после трансплантации клеток для предотвращения отторжения нейрональных клеток. Животных, трансплантированных клетками, умерщвляли и выделяли мозг для гистологических исследований на 8, 16 и 24 неделях.
Исследования поведения
На 4, 8, 16 и 24 неделях после трансплантации оценивали тяжесть заболевания и степень восстановления в экспериментальной и контрольной группах. Исследования поведения включали (а) перемещение передней конечности, (b) вращение, индуцированное апоморфином. Исследование перемещения передней части конечностей позволяет оценить способность крысы совершать направленные движения передней конечностью в ответ на сенсорный стимул. Результаты выражали как число перемещений передней конечностью в пораженной половине тела по сравнению с нормой и БП-контролем. Вращения, индуцированные апоморфином, отслеживали в течение одного часа. Все исследования поведения оценивал независимый наблюдатель "вслепую".
Иммуногистохимический анализ БП мозга
На 8, 16 и 24 неделях после трансплантации крысам, которым инъецировали NPC и БП контрольных крыс, умертвили. Крыс анестезировали кетамином (50 мг/кг интраперитонеально) и валиумом (30 мг/кг, интраперитонеально), а затем перфузировали физиологическим раствором, а затем 4% параформальдегидом. Мозги удалили и уравновешивали в 20% сахарозе в течение ночи при комнатной температуре. Делали серию парафиновых срезов толщиной 10 мкм, используя микротом, и помещали на стекла, покрытые поли-L-лизином. Парафиновые срезы мозга депарафинировали, а неспецифические центры связывания блокировали 1% бычьим сывороточным альбумином в PBS. Срезы затем окрашивали антителами к ядрам клеток человека, МАР-2, NeuN, TH и β-тубулину. (Подробное описание антител дано в таблице 1.) После трехразовой промывки PBS срезы инкубировали с вторичными антителами, меченными флуоресцином (FITC), в течение 2 часов. Срезы контрастно окрашивали 1 мкг/мл DAPI (Sigma) и оценивали под флуоресцентным микроскопом (Nikon Eclipse Е600) для определения иммуноположительной области. Аналогично было сделано двойное мечение TH/NeuN, ТН/β-тубулин.
Исследование эффективности in vivo
Затем авторы оценивали выживаемость, дифференцировку и функцию нейрональных клеток, полученных из эмбриональных стволовых клеток человека (чЭСК), после трансплантации в субстанцию нигра крыс, страдающих паркинсонизмом. Через шесть недель после поражения нейросферы, полученные из нейрональных клеток, дифференцированных из чЭСК, трансплантировали в субстанцию нигра крыс. Через 12 недель у посттрансплантированных животных было отмечено значительное улучшение в восстановлении поведения, которое продолжалось до шести месяцев (фигура 15а). Было отмечено значительное уменьшение вращений, индуцированных апоморфином (р<0,05), после клеточной инъекции по сравнению с контрольными животными. Перемещение передней части конечностей дает более наглядный показатель моторных нарушений, которые аналогичны нарушениям, обнаруженным при болезни Паркинсона у человека. Данное исследование было проведено на 4, 8, 12 и 24 неделях. На 4 и 8 неделях трансплантированные крысы не совершали никаких движений на пораженной половине тела, а незначительные улучшения наблюдали на 12 неделе. На 24 неделе у трансплантированных животных наблюдали значительное (р<0,05) улучшение моторной координации (фигура 15b). Гистология трансплантированной области показала, что трансплантированные клетки выжили в субстанции нигра, а также дифференцировались на дофаминергические нейроны in vivo no данным гистологических исследований. Для подтверждения, что трансплантированные клетки были человеческого происхождения, было проведено мечение антителами к ядрам клеток человека (фигура 17b) и ТН (фигура 17с). Трансплантированные клетки также были иммунореактивны к TH/NeuN (фигура 17d), ТН/р-тубулин (фигура 17е), ТН/МАР2 (фигура 17f).
На основании настоящего исследования было показано, что дофаминергические нейрональные клетки, дифференцированные из чЭСК, выжили и вызвали улучшения в поведении уже через 5 недель после трансплантации животным с моделью болезни Паркинсона. Кроме того, была подтверждена функциональная способность NPC за счет уменьшения вращений, индуцированных апоморфином. Иммуногистохимический анализ трансплантированной области выявил экспрессию ТН-положительных клеток на 24 неделе после трансплантации и нейроспецифических маркеров, таких как β-тубулин, МАР2 и NeuN. Двойное иммунное мечение ТН и Hu Nu показало, что TH-IR клетки в трансплантате имели человеческую природу.
Для достижения успешного результата терапии, основанной на клетке чЭС, одним из основных условий является то, что популяция терапевтических клеток не имеет онкогенную природу, так как недифференцированные чЭСК, как известно, дают тератомы in vivo. Ранее сообщалось, что использование мышиных, эмбриональных стволовых клеток, полученных из DA нейронов, для трансплантации крыс на модели БП показало образование тератомы в мозге одного из животных. Это обусловлено присутствием небольшой популяции недифференцированных чЭС клеток даже после дифференцировки в нейрон индуцированной среде в течение более 21 дня (Nishimura et al). В настоящем исследовании трансплантация нейрональных клеток, полученных из клеток чЭС, не привела к образованию опухоли ни у SCID мышей, ни в мозге крыс с БП, по данным гистологических анализов, через шесть месяцев после трансплантации.
Результаты in vivo функциональности NPC в моделях БП подтверждают, что нейрональные клетки, полученные из чЭСК, могут использоваться в терапии болезни Паркинсона у человека.
Все материалы и методы, описанные и заявленные здесь, могут быть сделаны и использованы без сложных экспериментов в свете настоящего изобретения. Хотя композиции и способы настоящего изобретения были описаны здесь в виде предпочтительного осуществления, для специалиста будет очевидным, что могут быть применены различные варианты к составу и/или способам и в шагах, или последовательности шагов способов, описанных здесь, без отхода от концепции, духа и области изобретения. В особенности, будет очевидным, что определенные агенты, которые химически или физиологически связаны, могут быть заменены агентами, описанными здесь, при этом такие же или сходные результаты могли бы быть достигнуты. Все эти сходные замены и модификации очевидны для специалисто, и считается, что они находятся в духе области и концепции изобретения, как определено прилагаемой формулой изобретения.
Реферат
Изобретение относится к биотехнологии. Описана дифференцированная клеточная популяция в культуре in vitro, применяемая для лечения нейродегенеративных расстройств, полученная путем дифференцировки плюрипотентных стволовых клеток приматов и селекции нестин-положительных клеток и NCAM-положительных клеток путем культивирования клеток в присутствии TGF-β3, или интерлейкина-1β, или обоих этих факторов и N-ацетилцистеина, где по крайней мере 60% дифференцированных клеток являются дофаминергическими нейронами или где 85% или более клеток представляют собой селектированную популяцию нестин-положительных клеток, и 80% или более клеток являются обогащенной популяцией, которая экспрессирует NCAM, и 60% или более дифферецированных клеток экспрессируют тирозингидроксилазу. Раскрыт способ получения описанной дифференцированной клеточной популяции. Предложены способы получения дофаминергических нейронов и серотонинергических нейронов. Также предложен способ лечения пациентов с нейродегенеративными расстройствами и неврологическими заболеваниями. Настоящее изобретение позволяет получать нейрональные клетки-предшественницы и терминально дифференцированные клетки в больших количествах, и таким образом они могут служить прекрасным источником для клеточно-заместительной терапии при неврологических расстройствах, таких как болезнь Паркинсона. 6 н. и 38 з.п. ф-лы, 17 ил., 5 табл.
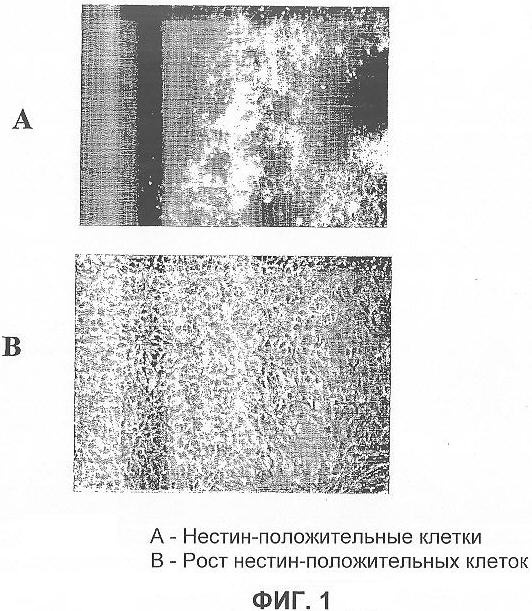
Комментарии