Композиция ферментной смеси для гидролиза смеси целлюлозных и гемицеллюлозных материалов (варианты) и способы ее использования (варианты) - RU2536256C2
Код документа: RU2536256C2
Чертежи















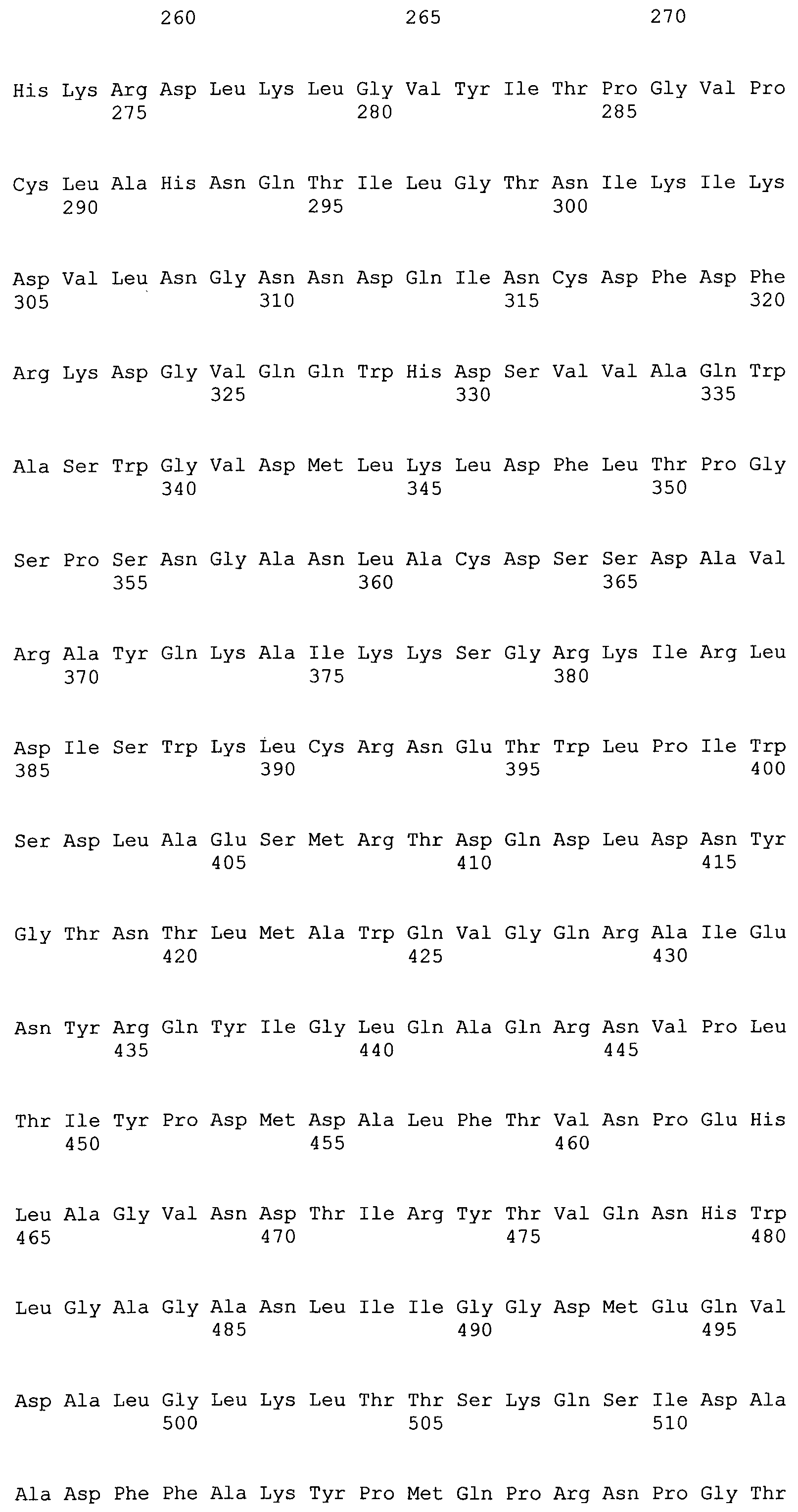


















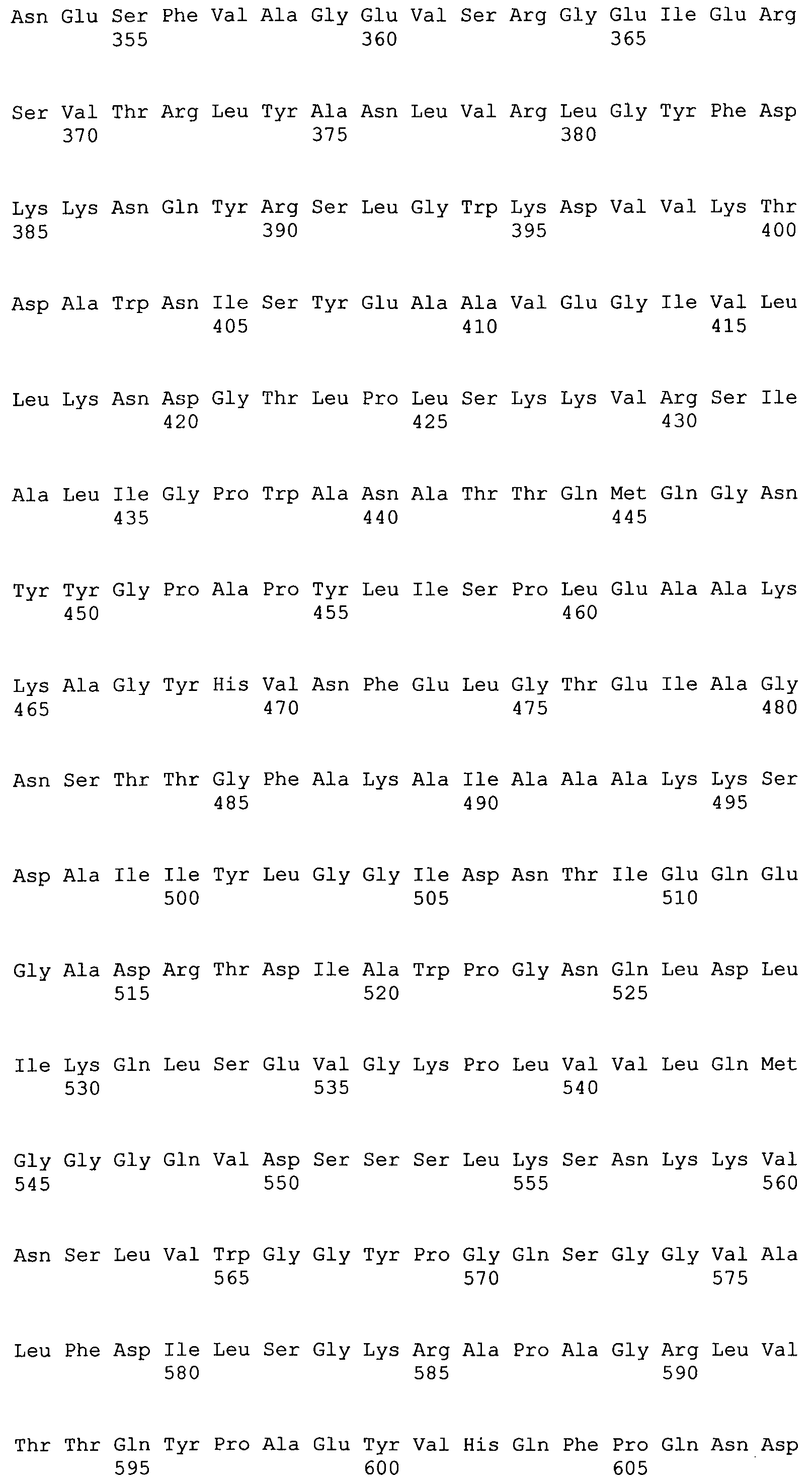

Описание
ПРИОРИТЕТ
По настоящей заявке испрашивается приоритет предварительной заявки США № 61/038520, поданной 21 марта 2008, которая включена, таким образом, в качестве ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящие композиции и способы относятся к ферментным смесям целлюлазы/гемицеллюлазы для улучшения ферментативного гидролиза целлюлозных материалов.
ПРЕДПОСЫЛКИ К СОЗДАНИЮ ИЗОБРЕТЕНИЯ
Основными компонентами биомассы являются целлюлоза и гемицеллюлоза. Целлюлоза состоит из полимеров β-1,4-связанных остатков глюкозы, которые организованы в фибриллярные структуры более высокого порядка. Гемицеллюлозы представляют собой гетерополисахариды, которые включают моносахариды, отличные от глюкозы, такие как D-ксилоза, L-арабиноза, D-манноза, D-глюкоза, D-галактоза и 4-O-метил-D-глюкуроновая кислота, связанные вместе не только гликозидными связями, а также сложноэфирными связями. Состав и структура гемицеллюлозы более сложные, чем целлюлозы, и могут варьировать качественно и количественно в различных видах древесных растений, травах и злаковых.
Целлюлоза может превращаться в сахара, такие как глюкоза, и использоваться в качестве источника энергии многочисленными микроорганизмами, в том числе бактериями, дрожжами и грибами в промышленных целях. Целлюлозные материалы также могут превращаться в сахара под действием коммерчески доступных ферментов, и полученные в результате сахара могут быть использованы в качестве исходного сырья для промышленных микроорганизмов для получения таких продуктов, как пластмассы и этанол. Однако существующие в настоящее время целлюлазные продукты в основном лишены способности гидролизовать гемицеллюлозные материалы, которые остаются не израсходованными в составах биомассы и могут препятствовать обработке и устранению биомассы.
Соответственно сохраняется потребность в разработке эффективных ферментных систем для гидролиза как целлюлозы, так и гемицеллюлозы, в том числе совместном получении или перемешивании оптимизированных наборов ферментов для превращения гемицеллюлозных олигомеров и полимеров в свободную пентозу для ферментирования. Такие оптимизированные ферментные системы требуются для улучшения эффективности и экономических показателей биомассы.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к оптимизированным биоконвертирующим ферментным смесям, способам их получения, а также к способам применения оптимизированной биоконвертирующей ферментной смеси для превращения биомассы в сахар. Биоконвертирующая ферментная смесь содержит смесь цельной целлюлазы и одной или нескольких гемицеллюлаз, выбор которых продиктован предполагаемым субстратом биомассы и условиями обработки.
В одном аспекте представлен состав ферментной смеси для гидролиза смеси целлюлозных и гемицеллюлозных материалов, содержащий:
(a) первую ферментную композицию, содержащую целлюлазу,
(b) вторую ферментную композицию, содержащую по меньшей мере одну ксиланазу, выбранную из ксиланазы GH10 или GH11, и
(c) третью ферментную композицию, содержащую по меньшей мере одну дополнительную гемицеллюлазу, которая не является ксиланазой GH10 или GH11 или не является ксиланазой GH10 или GH11, как в пункте (b),
где состав ферментной смеси обеспечивает по меньшей мере одно из следующего: (i) ускоренную конверсию глюкана или (ii) ускоренную конверсию ксилана по сравнению с эквивалентным составом ферментной смеси, лишенной по меньшей мере одной дополнительной гемицеллюлазы.
В некоторых вариантах осуществления первая ферментная композиция представляет собой смесь цельной целлюлазы из нитчатых грибов. В некоторых вариантах осуществления первая ферментная композиция представляет собой смесь цельной целлюлазы из нитчатых грибов, дополненную дополнительным количеством β-глюкозидазы.
В некоторых вариантах осуществления вторая ферментная композиция содержит ксиланазу XYN2 из Trichoderma reesei. В некоторых вариантах осуществления вторая ферментная композиция содержит ксиланазу XYN3 из Trichoderma reesei.
В некоторых вариантах осуществления по меньшей мере одна ксиланаза имеет аминокислотную последовательность, по меньшей мере на 80% идентичную аминокислотной последовательности, выбранной из SEQ ID NO: 1 или SEQ ID NO: 2. В некоторых вариантах осуществления по меньшей мере одна ксиланаза имеет аминокислотную последовательность, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или даже по меньшей мере на 99% идентичную аминокислотной последовательности, выбранной из SEQ ID NO: 1 или SEQ ID NO: 2. В конкретных вариантах осуществления по меньшей мере одна ксиланаза имеет аминокислотную последовательность, выбранную из SEQ ID NO: 1 или SEQ ID NO: 2.
В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза выбрана из группы, состоящей из гемицеллюлазы GH54, гемицеллюлазы GH62, гемицеллюлазы GH27, гемицеллюлазы GH36, гемицеллюлазы GH5, гемицеллюлазы GH74, гемицеллюлазы GH67, гемицеллюлазы GH28, гемицеллюлазы GH11, гемицеллюлазы GH10, гемицеллюлазы GH3 и гемицеллюлазы CE5.
В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза представляет собой β-ксилозидазу или арабинофуранозидазу. В конкретных вариантах осуществления β-ксилозидаза представляет собой BXL1 из Trichoderma reesei и арабинофуранозидаза представляет собой ABF1, ABF2 или ABF3 из Trichoderma reesei. В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза представляет собой сочетание β-ксилозидазы и арабинофуранозидазы.
В некоторых вариантах осуществления первая ферментная композиция представляет собой смесь цельной целлюлазы из нитчатых грибов, дополненную дополнительным количеством β-глюкозидазы, вторая ферментная композиция содержит ксиланазу, и по меньшей мере одна дополнительная гемицеллюлаза представляет собой сочетание β-ксилозидазы и арабинофуранозидазы.
В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза представляет собой гемицеллюлазу Trichoderma reesei, выбранную из группы, состоящей из α-арабинофуранозидазы I (ABF1), α-арабинофуранозидазы II (ABF2), α-арабинофуранозидазы III (ABF3), α-галактозидазы I (AGL1), α-галактозидазы II (AGL2), α-галактозидазы III (AGL3), ацетилксиланэстеразы I (AXE1), ацетилксиланэстеразы III (AXE3), эндоглюканазы VI (EG6), эндоглюканазы VIII (EG8), α-глюкуронидазы I (GLR1), β-маннаназы (MAN1), полигалактуроназы (PEC2), ксиланазы I (XYN1), ксиланазы II (XYN2), ксиланазы III (XYN3) и β-ксилозидазы (BXL1).
В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза имеет аминокислотную последовательность, по меньшей мере на 80% идентичную аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17. В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза имеет аминокислотную последовательность, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или даже по меньшей мере на 99% идентичную одной из указанных выше аминокислотных последовательностей. В конкретных вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза имеет аминокислотную последовательность, соответствующую одной из указанных выше аминокислотных последовательностей.
В другом аспекте представлен способ гидролиза смеси целлюлозных и гемицеллюлозных материалов, предусматривающий контактирование смеси целлюлозных и гемицеллюлозных материалов с:
(a) первой ферментной композицией, содержащей целлюлазу,
(b) второй ферментной композицией, содержащей по меньшей мере одну ксиланазу, выбранную из ксиланазы GH10 или GH11, и
(c) третьей ферментной композицией, содержащей по меньшей мере одну дополнительную гемицеллюлазу, которая не представляет собой ксиланазу GH10 или GH11 или не представляет собой ксиланазу GH10 или GH11, как в пункте (b),
осуществляя, таким образом, гидролиз смеси целлюлозных и гемицеллюлозных материалов,
где указанное контактирование в результате приводит по меньшей мере к одному из следующего: (i) ускорению конверсии глюкана или (ii) ускорению конверсии ксилана по сравнению с эквивалентным контактированием в отсутствие по меньшей мере одной дополнительной гемицеллюлозы.
В некоторых вариантах осуществления первая ферментная композиция представляет собой смесь цельной целлюлазы из нитчатых грибов. В некоторых вариантах осуществления первая ферментная композиция представляет собой смесь цельной целлюлазы из нитчатых грибов, дополненную дополнительным количеством β-глюкозидазы.
В некоторых вариантах осуществления вторая ферментная композиция содержит ксиланазу XYN2 из Trichoderma reesei. В некоторых вариантах осуществления вторая ферментная композиция содержит ксиланазу XYN3 из Trichoderma reesei.
В некоторых вариантах осуществления по меньшей мере одна ксиланаза имеет аминокислотную последовательность, по меньшей мере на 80% идентичную аминокислотной последовательности, выбранной из SEQ ID NO: 1 или SEQ ID NO: 2. В некоторых вариантах осуществления по меньшей мере одна ксиланаза имеет аминокислотную последовательность, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или даже по меньшей мере на 99% идентичную аминокислотной последовательности, выбранной из SEQ ID NO: 1 или SEQ ID NO: 2. В конкретных вариантах осуществления по меньшей мере одна ксиланаза имеет аминокислотную последовательность, выбранную из SEQ ID NO: 1 или SEQ ID NO: 2.
В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза выбрана из группы, состоящей из гемицеллюлазы GH54, гемицеллюлазы GH62, гемицеллюлазы GH27, гемицеллюлазы GH36, гемицеллюлазы GH5, гемицеллюлазы GH74, гемицеллюлазы GH67, гемицеллюлазы GH28, гемицеллюлазы GH11, гемицеллюлазы GH10, гемицеллюлазы GH3 и гемицеллюлазы CE5.
В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза представляет собой β-ксилозидазу или арабинофуранозидазу. В конкретных вариантах осуществления β-ксилозидаза представляет собой BXL1 из Trichoderma reesei, и арабинофуранозидаза представляет собой ABF1, ABF2 или ABF3 из Trichoderma reesei. В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза представляет собой сочетание β-ксилозидазы и арабинофуранозидазы.
В некоторых вариантах осуществления первая ферментная композиция представляет собой смесь цельной целлюлазы из нитчатых грибов, дополненную дополнительным количеством β-глюкозидазы, вторая ферментная композиция содержит ксиланазу, и по меньшей мере одна дополнительная гемицеллюлаза представляет собой сочетание β-ксилозидазы и арабинофуранозидазы.
В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза представляет собой гемицеллюлазу Trichoderma reesei, выбранную из группы, состоящей из α-арабинофуранозидазы I (ABF1), α-арабинофуранозидазы II (ABF2), α-арабинофуранозидазы III (ABF3), α-галактозидазы I (AGL1), α-галактозидазы II (AGL2), α-галактозидазы III (AGL3), ацетилксиланэстеразы I (AXE1), ацетилксиланэстеразы III (AXE3), эндоглюканазы VI (EG6), эндоглюканазы VIII (EG8), α-глюкуронидазы I (GLR1), β-маннаназы (MAN1), полигалактуроназы (PEC2), ксиланазы I (XYN1), ксиланазы II (XYN2), ксиланазы III (XYN3) и β-ксилозидазы (BXL1).
В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза имеет аминокислотную последовательность, по меньшей мере на 80% идентичную аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17. В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза имеет аминокислотную последовательность, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или даже по меньшей мере на 99% идентичную одной из указанных выше аминокислотных последовательностей. В конкретных вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза имеет аминокислотную последовательность, соответствующую одной из указанных выше аминокислотных последовательностей.
В некоторых вариантах осуществления контактирование смеси целлюлозных и гемицеллюлозных материалов с первой ферментной композицией, второй ферментной композицией и третьей ферментной композицией проводят одновременно.
В некоторых вариантах осуществления первая ферментная композиция, вторая ферментная композиция и третья ферментная композиция представлены в единой композиции ферментной смеси.
Эти и другие аспекты и варианты осуществления настоящих композиций и способов будут очевидны из следующего описания.
ПОДРОБНОЕ ОПИСАНИЕ
I. Определения
Если в настоящей заявке не определено иное, все технические и научные термины, используемые в настоящей заявке, имеют то же значение, которое обычно понимается специалистом в данной области. Заголовки, представленные в настоящей заявке, не являются ограничениями различных аспектов или варианты осуществления изобретения, описанные под одним заголовком, могут применяться к композициям и способам в целом. Как представленное выше общее описание, так и следующее подробное описание являются иллюстративными и пояснительными и не ограничивают композиции и способы, описанные в настоящей заявке. Применение единственного числа включает множественное число, если особым образом не указано иное, и применение «или» означает «и/или», если не указано иное. Термины «содержат», «содержащий», «содержит», «включают», «включающий» и «включает» не предназначены для ограничения. Все патенты и публикации, в том числе все аминокислотные и нуклеотидные последовательности, описанные в таких патентах и публикациях, на которые имеются ссылки в настоящей заявке, специально включены в качестве ссылки. Следующие термины определены для ясности:
В контексте настоящего изобретения термин «целлюлоза: относится к полисахариду, состоящему из β(1→4) связанных D-глюкозных звеньев, имеющих общую формулу (C6H10O5)n. Целлюлоза представляет собой структурный компонент первичной клеточной стенки зеленых растений, многих форм водорослей и оомицетов.
В контексте настоящего изобретения термин "целлюлаза" относится к ферменту, способному гидролизовать полимеры целлюлозы до более коротких олигомеров и/или глюкозы.
В контексте настоящего изобретения термин "композиция/препарат/смесь цельной целлюлазы" или подобный относится как к природным, так и неприродным композициям, которые включают множество целлюлаз, продуцируемых организмом, например нитчатыми грибами. Одним примером композиции цельной целлюлазы является среда, в которой культивируют нитчатые грибы, которая включает секретированные целлюлазы, например, одну или несколько целлобиогидролаз, одну или несколько эндоглюканаз и одну или несколько β-глюкозидаз в заранее определенном соотношении.
В контексте настоящего изобретения “гемицеллюлоза” представляет собой полимерный компонент растительных материалов, который содержит сахарные мономеры, отличные от глюкозы, в отличие от целлюлозы, которая содержит только глюкозу. Помимо глюкозы гемицеллюлоза может включать ксилозу, маннозу, галактозу, рамнозу и арабинозу, при этом ксилоза является наиболее распространенным сахарным мономером. Гемицеллюлозы содержат большинство из D-пентозных сахаров и иногда небольшие количества L-сахаров. Сахара в гемицеллюлозе могут быть связаны сложноэфирными связями, а также гликозидными связями. Приводимые в качестве примеров формы гемицеллюлозы включают, но не только, галактан, маннан, ксилан, арабинан, арабиноксилан, глюкоманнан, галактоманан и подобные.
В контексте настоящего изобретения термин "гемицеллюлаза" относится к ферментам, способным разрушать гемицеллюлозу на составляющие ее сахара или более короткие полимеры, и включает эндо-гидролазы, экзо-гидролазы и различные эстеразы.
В контексте настоящего изобретения термин “ксиланаза" относится к белку или полипептидному домену белка или полипептида, происходящего из микроорганизма, например гриба, бактерии или из растения или животного, и который обладает способностью катализировать расщепление ксилана в одном или нескольких различных положениях углеводного скелета ксилана, включая разветвленные ксиланы и ксилоолигосахариды. Следует отметить, что ксиланаза является разновидностью гемицеллюлазы.
В контексте настоящего изобретения «субстрат биомассы» представляет собой материал, содержащий как целлюлозу, так и гемицеллюлозу.
В контексте настоящего изобретения «природная» композиция представляет собой композицию, продуцируемую в природе или организмом, который встречается в природе.
В контексте настоящего изобретения «вариант» белка отличается от «родительского» белка, из которого он получен с помощью замены, делеции или добавления небольшого количества аминокислотных остатков, например 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более аминокислотных остатков. В некоторых случаях родительский белок представляет собой полипептиды «дикого типа», «нативные» или «природные». Варианты белков могут быть описаны как имеющие определенный процент идентичности последовательностей с родительским белком, например, по меньшей мере 80%, по меньшей мере 81%, по меньшей мере 82%, по меньшей мере 83%, по меньшей мере 84%, по меньшей мере 85%, по меньшей мере 86%, по меньшей мере 87%, по меньшей мере 88%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, даже по меньшей мере 99%, которая может быть определена с использованием подходящего программного обеспечения, известного в данной области, например, описанного в CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubel et al. (eds) 1987, Supplement 30, section 7.7.18).
Предпочтительные программы включают Vector NTI Advance™ 9.0 (Invitrogen Corp. Carlsbad, CA), GCG Pileup program, FASTA (Pearson et al. (1988) Proc. Natl, Acad. Sci USA 85:2444-2448), и BLAST (BLAST Manual, Altschul et al., Natl Cent. Biotechnol. Inf., Natl Lib. Med. (NCIB NLM NIH), Bethesda, Md., and Altschul et al. (1997) NAR 25:3389-3402). Другой предпочтительной программой выравнивания является ALIGN Plus (Scientific and Educational Software, PA), предпочтительно использующая параметры по умолчанию. Другим программным обеспечением, которое находит применение, является программа TFASTA Data Searching Program, доступная в Sequence Software Package Version 6.0 (Genetics Computer Group, University of Wisconsin, Madison, WI).
II. Биоконверсия композиций ферментных смесей и способы их применения
Целлюлоза представляет собой гомополимер ангидроцеллобиозы и, следовательно, линейный β-(1-4)-D-глюкан. Гемицеллюлозы, напротив, включают целый ряд соединений, таких как ксиланы, ксилоглюканы, арабиноксиланы и маннаны, в сложных разветвленных структурах, и со спектром заместителей. Как следствие сложного и гетерогенного состава гемицеллюлоз, в частности арабиноксиланов, ферментативное разрушение растительного материала требует действия совокупности функций как расщепления разветвленной структуры, так и деполимеризации. Дополнительно, для разрушения растительных материалов требуются ферменты, которые действуют на гемицеллюлозы, содержащие как пятиуглеродные сахара (пентозы), такие как ксилоза и арабиноза, так и шестиуглеродные сахара (гексоны), такие как манноза и глюкоза.
Ферментативный гидролиз гемицеллюлозы до ее мономеров требует участия нескольких гемицеллюлазных ферментов с различными функциями. Гемицеллюлазы можно расположить в три основные категории: эндо-ферменты, действие которых направлено на внутренние связи в полисахариде, экзо-ферменты, которые действуют поступательно с любого редуцирующего или нередуцирующего конца полисахаридной цепи, и добавочные ферменты, ацетилэстеразы и эстеразы, которые гидролизуют лигнингликозидные связи, такие как эстераза кумариновой кислоты и эстераза феруловой кислоты.
Хотя некоторые грибы продуцируют полные целлюлазные системы, которые включают экзо-целлобиогидролазы (или CBH-тип целлюлаз), эндоглюканазы (или EG-тип целлюлаз) и β-глюкозидазы (или BG-тип целлюлаз), известные целлюлазные ферменты и их смеси, обычно имеют ограниченную активность в отношении гемицеллюлозы и ограниченное значение в гидролизе растительных материалов. Биоконвертирующие композиции ферментных смесей по настоящему изобретению и способы основаны, в части, на наблюдении того, что некоторые сочетания целлюлаз и гемицеллюлаз значительно повышают эффективность гидролиза растительных материалов, главным образом, как определено мониторингом конверсии глюкана и ксилана.
Приводимая в качестве примера целлюлазная композиция, используемая для идентификации целлюлазных/гемицеллюлазных композиций, которые усиливают гидролиз глюкана и/или ксилана, представляет собой композиции цельной целлюлазы, продуцируемые нитчатыми грибами (т.е.Trichoderma reesei). Композиция включает несколько экзо-целлобиогидролаз и эндоглюканаз и дополнена дополнительной β-глюкозидазой для повышения высвобождения глюкозы. Эта композиция коммерчески доступна как ACCELLERASE 1000TM (Danisco A/S, Genencor Division, Palo Alto, CA). ACCELLERASE 1000TM содержит экзо-целлобиогидролазы (т.е. примерно 50% (масс./масс.) CBHI (CEL7A) и примерно 14% CBHII (CEL6A), эндоглюканазы (т.е. примерно 12% EGI (CEL7B) и примерно 10% EGII (CEL5A)) и β-глюкозидазу (т.е. примерно 5% BGLI (CEL3A). Также может присутствовать небольшое количество XYN2 (т.е. примерно менее 1%). Другие компоненты, которые не идентифицированы, также присутствуют в количествах примерно менее чем 1%.
Могут быть использованы другие целлюлазные композиции, в том числе другие смеси цельных целлюлаз и смеси целлюлаз, собранные из многочисленных индивидуально выделенных целлюлаз. Предпочтительные целлюлазные композиции включают по меньшей мере одну любую из экзоцеллобиогидролазы, эндоглюканазы и β-глюкозидазы. В некоторых вариантах осуществления цельную питательную среду, которая содержит многочисленные целлюлазы, получают из организма, такого как Acremonium, Aspergillus, Emericella, Fusarium, Humicola, Mucor, Myceliophthora, Neurospora, Scytalidium, Thielavia, Tolypocladium, Penicillium, или Trichoderma spp., или видов, полученных из них.
Композиция дополнительно содержит по меньшей мере одну и в некоторых случаях две, три или более гемицеллюлаз. Примеры подходящих дополнительных гемицеллюлаз включают ксиланазы, арабинофуранозидазы, ацетилксиланэстеразы, глюкуронидазы, эндогалактаназы, маннаназы, эндо- или экзо-арабиназы, экзо-галактаназы и их смеси. Примеры подходящих эндо-гемицеллюлаз включают эндо-арабиназу, эндо-арабиногалактаназу, эндо-глюканазу, эндо-маннаназу, эндо-ксиланазу и фераксанэндоксиланазу. Примеры подходящих экзо-гемицеллюлаз включают α-L-арабинозидазу, β-L-арабинозидазу, α-1,2-L-фукозидазу, α-D-галактозидазу, β-D-галактозидазу, β-D-глюкозидазу, β-D-глюкуронидазу, β-D-маннозидазу, β-D-ксилозидазу, экзо-глюкозидазу, экзо-целлобиогидролазу, экзо-маннобиогидролазу, экзо-маннаназу, экзо-ксиланазу, ксилан α-глюкуронидазу и кониферин β-глюкозидазу. Примеры подходящих эстераз включают ацетилэстеразы (ацетилксиланэстеразу, ацетилгалактанэстеразу, ацетилманнанэстеразу и ацетилксиланэстеразу) и арилэстеразы (эстеразу кумариновой кислоты и эстеразу феруловой кислоты).
Предпочтительно композиции и способы по настоящему изобретению включают по меньшей мере одну ксиланазу, которая представляет собой конкретный тип гемицеллюлазы, которая расщепляет основные цепи ксилана гемицеллюлозы. Предпочтительно ксиланаза представляет собой эндо-1,4-β-ксиланазу (E.C. 3.2.1.8). Многочисленные ксиланазы из грибных и бактериальных микроорганизмов были идентифицированы и охарактеризованы (смотри, например, патент США № 5437992; Coughlin, M. P. supra; Biely, P. et al. (1993) Proceedings of the second TRICEL symposium on Trichoderma reesei Cellulases and Other Hydrolases, Espoo 1993; Souminen, P. and Reinikainen, T. (eds)., в Foundation for Biotechnical and Industrial Fermentation Research 8:125-135). Три специфичные ксиланазы (XYN1, XYN2 и XYN3) были идентифицированы в T. reesei (Tenkanen, et al. (1992) Enzyme Microb. Technol. 14:566; Torronen, et al. (1992) Bio/Technology 10:1461; и Xu, et al. (1998) Appl. Microbiol. Biotechnol. 49:718). Четвертая ксиланаза (XYN4), выделенная из T. Reesei, описана в патентах США № 6555335 и 6768001 на имя Saloheimo et al., под названием Xylanase from Trichoderma reesei, способ ее получения и способы применения этого фермента, которые включены в настоящее описание в качестве ссылки в полном объеме.
Ксиланазы, приводимые в качестве примера для применения в настоящих композициях и способах, представляют собой XYN2 и XYN3. Подходящие варианты XYN2 и XYN3 и подходящие родственные ферменты из других организмов, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или даже по меньшей мере на 99% идентичные аминокислотной последовательности XYN2 или XYN3 (т.е. SEQ ID NO: 1 и 2 соответственно).
Помимо целлюлазной композиции и ксиланазы композиции и способы могут включать одну или несколько дополнительных гемицеллюлаз, таких как эндо-гемицеллюлаза и экзо-гемицеллюлаза, и/или эстеразы.
Подходящие эндо-гемицеллюлазы включают, но не только, эндо-1,4-β-маннозидазу (E.C. 3.2.1.78, также известную как β-манназа и β-маннаназа), которая катализирует случайный эндогидролиз 1,4,-β-D-маннозных связей в маннанах, галактоманнанах, глюкоманнанах; α-амилазу (E.C. 3.2.1.1), которая катализирует эндогидролиз 1,4-α-D-глюкозидных связей в полисахаридах, содержащих три или более 1,4-α-связанных D-глюкозных звеньев; ксилан α-1,2-глюкуронозидазу (E.C. 3.2.1.131, также известную как α-глюкуронидаза), которая катализирует гидролиз α-D-1,2-(4-O-метил)глюкуронозильных связей в основной цепи ксиланов древесины твердых пород; и эндоглюканазу (E.C. 3.2.1.4), которая катализирует эндогидролиз 1,4-β-D-глюкозидных связей в целлюлазе, лихенине и β-D-глюканах злаковых. Были идентифицированы многочисленные подтипы эндоглюканаз и являются подходящими для применения в композициях и способах, например, эндоглюканаза I, эндоглюканаза II, эндоглюканаза III, эндоглюканаза V и эндоглюканаза VI.
Подходящие экзо-гемицеллюлазы включают, но не только, α-арабинофуранозидазу, α-галактозидазу и β-ксилозидазу. α-арабинофуранозидаза, также известная как a-N-арабинофуранозидаза (E.C. 3.2.1.55), катализирует гидролиз концевых нередуцирующих остатков α-L-арабинофуранозида в α-L-арабинозидах. Может быть использован любой по меньшей мере из трех известных подтипов α арабинофуранозидазы (т.е. abf1, abf2 и abf3). α-галактозидаза (E.C. 3.2.1.22) катализирует гидролиз концевых нередуцирующих остатков α-D-галактозы в α-D-галактозидах, включая галактозные олигосахариды и галактоманнаны. Может быть использован любой из трех известных подтипов, т.е. α-галактозидаза I (agl1), α-галактозидаза II (agl2) и α-галактозидаза III (agl3). Глюкоамилаза, также известная как глюкан 1,4-α-глюкозидаза (E.C. 3.2.1.3), катализирует гидролиз концевых 1,4-связанных остатков α-D-глюкозы последовательно с нередуцирующих концов цепей с высвобождением β-D-глюкозы. β-глюкозидаза (E.C. 3.2.1.21) катализирует гидролиз концевых нередуцирующих остатков β-D-глюкозы с высвобождением β-D-глюкозы. β-ксилозидаза, также известная как ксилан 1,4-β-ксилозидаза (E.C. 3.2.1.37), катализирует гидролиз 1,4-β-D-ксиланов, для последовательного удаления остатков D-ксилозы с нередуцирующих концов. Композиции, которые включают смесь цельной целлюлазы наряду с ксиланазой и либо α-арабинофуранозидазой или β-ксилозидазой, были особенно эффективными в конверсии глюкана и/или ксилана.
Подходящие эстеразы включают, но не только, эстеразу феруловой кислоты и ацетилксиланэстеразу. Эстераза феруловой кислоты, также известная как ферулатэстераза (E.C. 3.1.1.73), катализирует гидролиз 4-гидрокси-3-метоксициннамоильной (ферулоильной) группы от этерифицированного сахара, который обычно представляет собой арабинозу в «природных» субстратах. Известные микробиологические эстеразы феруловой кислоты секретируются в культуральную среду. Любой из трех известных подтипов эстеразы феруловой кислоты (fae1, fae2 и fae3) может быть использован в композициях и способах по настоящему изобретению. Ацетилксиланэстераза I (E.C. 3.1.1.72) катализирует дезацетилирование ксиланов и ксило-олигосахаридов и также может быть использована в композициях и способах. В патентах США № 5426043 и 5681732 на имя De Graaff et al. описываются клонирование и экспрессия ацетилксиланэстераз грибного происхождения. В EP 507 369 описывается последовательность ДНК, кодирующая ацетилксиланэстеразу, выделенную из Aspergillus niger. В патенте США № 5830734 на имя Christgau et al., под названием Enzyme with acetyl esterase activity, описывается выделение целого ряда эстераз для применения в пищевой промышленности.
В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза выбрана из группы, состоящей из гемицеллюлазы GH54, гемицеллюлазы GH62, гемицеллюлазы GH27, гемицеллюлазы GH36, гемицеллюлазы GH5, гемицеллюлазы GH74, гемицеллюлазы GH67, гемицеллюлазы GH28, гемицеллюлазы GH11, гемицеллюлазы GH10, гемицеллюлазы GH3 и гемицеллюлазы CE5. В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза выбрана из группы, состоящей из α-арабинофуранозидазы I (ABF1), α-арабинофуранозидазы II (ABF2), α-арабинофуранозидазы III (ABF3), α-галактозидазы I (AGL1), α-галактозидазы II (AGL2), α-галактозидазы III (AGL3), ацетилксиланэстеразы I (AXE1), ацетилксиланэстеразы III (AXE3), эндоглюканазы VI (EG6), эндоглюканазы VIII (EG8), α-глюкуронидазы I (GLR1), β-маннаназы (MAN1), полигалактуроназы (PEC2), ксиланазы I (XYN1), ксиланазы II (XYN2), ксиланазы III (XYN3) и β-ксилозидазы (BXL1), которые могут быть из нитчатых грибов, таких как T. reesei. В некоторых вариантах осуществления по меньшей мере одна дополнительная гемицеллюлаза имеет аминокислотную последовательность по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или даже по меньшей мере на 99% идентичную аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17.
Варианты гемицеллюлаз (включая ксиланазы) могут включать замены, вставки или делеции, которые по существу не влияют на функцию или добавляют ферментам предпочтительные особенности. В некоторых вариантах осуществления замены, вставки или делеции не находятся на консервативных мотивах последовательности, а вместо этого ограничены аминокислотными последовательностями вне консервативных мотивов. Приводимые в качестве примеров замены являются консервативными заменами, которые сохраняют заряд, гидрофобность или размер боковых групп относительно родительской аминокислотной последовательности. Примеры консервативных замен представлены в следующей таблице:
Очевидно, что природные аминокислоты могут быть введены в полипептид путем изменения кодирующей последовательности нуклеиновой кислоты, кодирующей этот полипептид, тогда как неприродные аминокислоты обычно получают путем химической модификации экспрессированного полипептида.
Еще добавочные ферменты, такие как лакказа (E.C. 1.10.3.2), которые катализируют окисление ароматических соединений, и последующее восстановление кислорода до воды, также могут быть включены в биоконвертирующие композиции ферментных смесей по настоящему изобретению и способы.
В некоторых вариантах осуществления ферменты для использования в биоконвертирующих ферментных смесях по настоящему изобретению могут быть получены из одного или нескольких штаммов нитчатых грибов. Подходящие нитчатые грибы включают представителей отдела Eumycota и Oomycota, в том числе, но не только, следующие роды: Aspergillus, Acremonium, Aureobasidium, Beauveria, Cephalosporium, Ceriporiopsis, Chaetomium, Chrysosporium, Claviceps, Cochiobolus, Cryptococcus, Cyathus, Endothia, Endothia mucor, Fusarium, Gilocladium, Humicola, Magnaporthe, Myceliophthora, Myrothecium, Mucor, Neurospora, Phanerochaete, Podospora, Paecilomyces, Pyricularia, Rhizomucor, Rhizopus, Schizophylum, Stagonospora, Talaromyces, Trichoderma, Thermomyces, Thermoascus, Thielavia, Tolypocladium, Trichophyton и Trametes. В некоторых вариантах осуществления нитчатые грибы включают, но не только, следующие: A. nidulans, A. niger, A. awomari, A. aculeatus, A. kawachie.g., NRRL 3112, ATCC 22342 (NRRL 3112), ATCC 44733, ATCC 14331 и штамм UVK 143f, A. oryzae, например ATCC 11490, N. crassa, Trichoderma reesei, например NRRL 15709, ATCC 13631, 56764, 56765, 56466, 56767, и Trichoderma viride, например ATCC 32098 и 32086. В предпочтительном выполнении, нитчатым грибом являются виды Trichoderma. Особенно предпочтительными видами и штаммом для применения в настоящем изобретении является T. reesei RL-P37.
В конкретном варианте осуществления отдельный сконструированный штамм гиперэкспрессирует ферментные компоненты в желаемом соотношении так, чтобы дополнительная очистка или добавки не требовались. В альтернативном варианте осуществления биоконвертирующая ферментная смесь получена из двух или более природных или сконструированных штаммов нитчатых грибов. Желаемое соотношение ферментных компонентов может достигаться путем изменения относительного количества фермента в конечной смеси. Даже когда используют два или более продуцирующих штаммов, желаемое соотношение компонентов ферментов может достигаться путем добавления очищенного или частично очищенного фермента.
В конкретных вариантах осуществления гемицеллюлазу получают из Aspergillus aculeatus, Aspergillus awamori, Aspergillus foetidus, Aspergillus japonicus, Aspergillus nidulans, Aspergillus niger или Aspergillus oryzae. В другом аспекте цельную питательную среду получают из Fusarium bactridioides, Fusarium cerealis, Fusarium crookwellense, Fusarium culmorum, Fusarium graminearum, Fusarium graminum, Fusarium heterosporum, Fusarium negundi, Fusarium oxysporum, Fusarium reticulatum, Fusarium roseum, Fusarium sambucinum, Fusarium sarcochroum, Fusarium sporotrichioides, Fusarium sulphureum, Fusarium torulosum, Fusarium trichothecioides, Fusarium venenatum или Fusarium verticilloides. В другом аспекте гемицеллюлазный комплекс получают из Humicola insolens, Humicola lanuginosa, Mucor miehei, Myceliophthora thermophila, Neurospora crassa, Scytalidium thermophilum или Thielavia terrestris. В других вариантах осуществления гемицеллюлазу получают из Trichoderma harzianum, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma reesei, например RL-P37 [Sheir-Neiss et al. (1984) Appl. Microbiol. Biotechnology 20:46-53; патент США № 4797361; доступные в виде биологически чистой культуры из постоянной коллекции Northern Regional Research Laboratory, U.S. Department of Agriculture, Peoria, III., U.S.A. (NRRL 15709); ATCC 13631, 56764, 56466, 56767] или Trichoderma viride, например ATCC 32098 и 32086.
В некоторых вариантах осуществления входящий в состав гемицеллюлазный фермент получают путем экспрессии гена, кодирующего гемицеллюлазный фермент. Например, ксиланаза может быть секретирована во внеклеточное пространство, например, грамположительного организма, такого как Bacillus или Actinomycetes, или эукариотического организма, такого как Trichoderma, Aspergillus, Saccharomyces, или Pichia. Должно быть понятно, что в некоторых вариантах осуществления один или несколько гемицеллюлазных ферментов могут быть гиперэкспрессированы в рекомбинантном микроорганизме, относительно нативных уровней. Клетка-хозяин может быть генетически модифицирована для снижения экспрессии одного или нескольких белков, которые являются эндогенными для клетки. В одном варианте осуществления клетка может содержать один или несколько нативных генов, в частности генов, которые кодируют секретируемые белки, которые были делетированы или инактивированы. Например, один или несколько генов, кодирующих протеазу (например, ген, кодирующий аспартилпротеазу; смотри Berka et al. (1990) Gene 86:153-162 и патент США № 6509171), или гены, кодирующие целлюлазу, могут быть делетированы или инактивированы. Нуклеиновые кислоты, кодирующие гемицеллюлазу, могут присутствовать в геноме организма или перенесены в плазмиду, которая реплицируется в организме. В тех случаях, когда гемицеллюлаза экспрессируется из генома, ген и регуляторные последовательности, ассоциированные с ними, могут быть встроены в геном путем случайной или гомологичной интеграции. В некоторых случаях, например, когда требуется особенно высокий уровень экспрессии, может быть использована как случайная, так и гомологичная интеграция.
Субстрат биомассы для использования в качестве источника целлюлозы и гемицеллюлозы для гидролиза с использованием ферментных композиций и способов по настоящему изобретению может представлять собой, например, травяной материал, сельскохозяйственные остатки, остатки от лесного хозяйства, коммунально-бытовые твердые отходы, макулатуру и древесную массу и остатки бумаги, и подобное. Обычные формы субстрата биомассы включают, но не только, деревья, кустарники и травы, пшеницу, солому от пшеницы, жмых сахарного тростника, кукурузу, листовые обертки початка кукурузы, сердцевину кукурузы, в том числе волокна из сердцевин, продукты и побочные продукты перемола зерна, такого как кукуруза (в том числе влажного перемола и сухого перемола), а также коммунально-бытовые твердые отходы, макулатуру и дворовый мусор. Субстрат биомассы может быть получен из “нетронутой биомассы” (такой как деревья, кустарник, травы, плоды, цветы, зеленые культуры, древесина твердых и мягких пород), “использованной биомассы” (такой как побочные продукты сельского хозяйства, промышленные органические отходы, строительный и городской мусор, коммунально-бытовые твердые отходы и дворовый мусор) или “смешанной биомассы”, которая представляет собой смесь нетронутой и использованной биомассы. Субстрат биомассы может включать, например, древесину, древесную массу, отходы бумажного производства, отходы бумажной массы, прессованную древесину, кукурузную солому, кукурузное волокно, отходы от обработки риса, бумаги и древесной массы, древовидные или травянистые растения, плодовую мякоть, овощную мякоть, пемзу, спиртовое зерновое сырье, травы, рисовую шелуху, выжимки сахарного тростника, хлопок, джут, пеньку, лен, бамбук, лубяное волокно, манильскую пеньку, солому, сердцевины кукурузных початков, спиртовое зерновое сырье, листья, пшеничную солому, волокна кокоса, водоросли, просо прутьевидное и их смеси.
Субстрат биомассы может быть использован непосредственно или может подвергаться предварительной обработке с использованием общепринятых способов, известных в данной области. Такие предварительные обработки включают химические, физические и биологические предварительные обработки. Например, технологии предварительных физических обработок включают, без ограничения, различные типы измельчения, разрушения, обработки водяным паром/паровым взрывом, излучением и гидротермолизисом. Технологии химической предварительной обработки включают, без ограничения, разбавленную кислоту, щелочные агенты, органические растворители, аммиак, диоксид серы и pH-контролируемый гидротермолизис. Методики биологической предварительной обработки включают, без ограничения, применение лигнин-солюбилизирующих микроорганизмов.
Оптимальные уровни доз биоконвертирующей ферментной смеси и целлюлаз и гемицеллюлаз в ней, варьируют в зависимости от субстрата биомассы и используемых технологий предварительной обработки. Условия технологического процесса, такие как pH, температура и время реакции также могут влиять на скорости продукции этанола. Предпочтительно реакционноспособная композиция содержит от 0,1 до 200 мг биоконвертирующей ферментной смеси на грамм биомассы, более предпочтительно от 1 до 100 мг биоконвертирующей ферментной смеси на грамм биомассы и наиболее предпочтительно 10-50 мг биоконвертирующей ферментной смеси на грамм биомассы. Примерами количеств являются 0,1-50, 1-40, 20-40, 1-30, 2-40 и 10-20 мг биоконвертирующй ферментной смеси на грамм биомассы. Альтернативно, количество фермента может быть определено исходя из количества субстрата в системе. В таком случае реакционноспособная композиция предпочительно содержит от 0,1 до 50 мг биоконвертирующей ферментной смеси на грамм суммарных сахаридов, более предпочтительно от 1 до 30 мг биоконвертирующей ферментной смеси на грамм суммарных сахаридов и более предпочтительно от 10 до 20 мг биоконвертирующей ферментной смеси на грамм суммарных сахаридов. Альтернативно, количество фермента может быть определено исходя из количества целлюлозного субстрата в системе. В таком случае реакционноспособная композиция предпочтительно содержит от 0,2 до 100 мг биоконвертирующей ферментной смеси на грамм суммарных глюканов, более предпочтительно от 2 до 60 мг биоконвертирующей ферментной смеси на грамм суммарных глюканов и более предпочтительно от 20 до 40 мг биоконвертирующей ферментной смеси на грамм суммарных глюканов. Аналогично количество используемой биоконвертирующей ферментной смеси может быть определено по количеству гемицеллюлозы в биомассе субстрата. Соответственно реакционноспособная композиция предпочтительно содержит от 0,2 до 100 мг биоконвертирующей ферментной смеси на грамм гемицеллюлозы, более предпочтительно от 2 до 60 мг биоконвертирующей ферментной смеси на грамм гемицеллюлозы и более предпочтительно от 20 до 40 мг биоконвертирующей ферментной смеси на грамм гемицеллюлозы.
В некоторых вариантах осуществления композиция по настоящему изобретению находится в форме композиции цельной целлюлазы, усиленной гемицеллюлозой, содержащей препарат цельной целлюлазы и по меньшей мере одну гемицеллюлазу, где количество гемицеллюлазы находится в интервале от 1% до 50% от общего белка, и цельная целлюлаза находится в интервале менее чем от 99% до 50% от общего белка. Например, гемицеллюлаза может представлять 1% от общего белка, и композиция цельной целлюлазы может представлять 99% от общего белка, гемицеллюлаза может представлять 2% от общего белка, и композиция цельной целлюлазы может представлять 98% от общего белка, гемицеллюлаза может представлять 3% от общего белка, и композиция цельной целлюлазы может представлять 97% от общего белка, гемицеллюлаза может представлять 4% от общего белка, и композиция цельной целлюлазы может представлять 96% от общего белка, гемицеллюлаза может представлять 5% от общего белка, и композиция цельной целлюлазы может представлять 95% от общего белка, гемицеллюлаза может представлять 6% от общего белка, и композиция цельной целлюлазы может представлять 94% от общего белка, гемицеллюлаза может представлять 7% от общего белка, и композиция цельной целлюлазы может представлять 93% от общего белка, гемицеллюлаза может представлять 8% от общего белка, и композиция цельной целлюлазы может представлять 92% от общего белка, гемицеллюлаза может представлять 9% от общего белка, и композиция цельной целлюлазы может представлять 91% от общего белка, гемицеллюлаза может представлять 10% от общего белка, и композиция цельной целлюлазы может представлять 90% от общего белка, гемицеллюлаза может представлять 11% от общего белка, и композиция цельной целлюлазы может представлять 89% от общего белка, гемицеллюлаза может представлять 12% от общего белка, и композиция цельной целлюлазы может представлять 88% от общего белка, гемицеллюлаза может представлять 13% от общего белка, и композиция цельной целлюлазы может представлять 87% от общего белка, гемицеллюлаза может представлять 14% от общего белка, и композиция цельной целлюлазы может представлять 86% от общего белка, гемицеллюлаза может представлять 15% от общего белка, и композиция цельной целлюлазы может представлять 85% от общего белка, гемицеллюлаза может представлять 16% от общего белка, и композиция цельной целлюлазы может представлять 84% от общего белка, гемицеллюлаза может представлять 17% от общего белка, и композиция цельной целлюлазы может представлять 83% от общего белка, гемицеллюлаза может представлять 18% от общего белка, и композиция цельной целлюлазы может представлять 82% от общего белка, гемицеллюлаза может представлять 19% от общего белка, и композиция цельной целлюлазы может представлять 81% от общего белка, гемицеллюлаза может представлять 20% от общего белка, и композиция цельной целлюлазы может представлять 80% от общего белка, гемицеллюлаза может представлять 21% от общего белка, и композиция цельной целлюлазы может представлять 79% от общего белка, гемицеллюлаза может представлять 22% от общего белка, и композиция цельной целлюлазы может представлять 78% от общего белка, гемицеллюлаза может представлять 23% от общего белка, и композиция цельной целлюлазы может представлять 77% от общего белка, гемицеллюлаза может представлять 24% от общего белка, и композиция цельной целлюлазы может представлять 76% от общего белка, гемицеллюлаза может представлять 25% от общего белка, и композиция цельной целлюлазы может представлять 75% от общего белка, гемицеллюлаза может представлять 26% от общего белка, и композиция цельной целлюлазы может представлять 74% от общего белка, гемицеллюлаза может представлять 27% от общего белка, и композиция цельной целлюлазы может представлять 73% от общего белка, гемицеллюлаза может представлять 28% от общего белка, и композиция цельной целлюлазы может представлять 72% от общего белка, гемицеллюлаза может представлять 29% от общего белка, и композиция цельной целлюлазы может представлять 71% от общего белка, гемицеллюлаза может представлять 30% от общего белка, и композиция цельной целлюлазы может представлять 70% от общего белка, гемицеллюлаза может представлять 31% от общего белка, и композиция цельной целлюлазы может представлять 69% от общего белка, гемицеллюлаза может представлять 32% от общего белка, и композиция цельной целлюлазы может представлять 68% от общего белка, гемицеллюлаза может представлять 33% от общего белка, и композиция цельной целлюлазы может представлять 67% от общего белка, гемицеллюлаза может представлять 34% от общего белка, и композиция цельной целлюлазы может представлять 66% от общего белка, гемицеллюлаза может представлять 35% от общего белка, и композиция цельной целлюлазы может представлять 65% от общего белка, гемицеллюлаза может представлять 36% от общего белка, и композиция цельной целлюлазы может представлять 64% от общего белка, гемицеллюлаза может представлять 37% от общего белка, и композиция цельной целлюлазы может представлять 63% от общего белка, гемицеллюлаза может представлять 38% от общего белка, и композиция цельной целлюлазы может представлять 62% от общего белка, гемицеллюлаза может представлять 39% от общего белка, и композиция цельной целлюлазы может представлять 61% от общего белка, гемицеллюлаза может представлять 40% от общего белка, и композиция цельной целлюлазы может представлять 60% от общего белка, гемицеллюлаза может представлять 41% от общего белка, и композиция цельной целлюлазы может представлять 59% от общего белка, гемицеллюлаза может представлять 42% от общего белка, и композиция цельной целлюлазы может представлять 58% от общего белка, гемицеллюлаза может представлять 43% от общего белка, и композиция цельной целлюлазы может представлять 57% от общего белка, гемицеллюлаза может представлять 44% от общего белка, и композиция цельной целлюлазы может представлять 56% от общего белка, гемицеллюлаза может представлять 45% от общего белка, и композиция цельной целлюлазы может представлять 55% от общего белка, гемицеллюлаза может представлять 46% от общего белка, и композиция цельной целлюлазы может представлять 54% от общего белка, гемицеллюлаза может представлять 47% от общего белка, и композиция цельной целлюлазы может представлять 53% от общего белка, гемицеллюлаза может представлять 48% от общего белка, и композиция цельной целлюлазы может представлять 52% от общего белка, гемицеллюлаза может представлять 49% от общего белка, и композиция цельной целлюлазы может представлять 51% от общего белка, или гемицеллюлаза может представлять 50% от общего белка, и композиция цельной целлюлазы может представлять 50% от общего белка.
При применении композиции биоконвертирующей ферментной смеси могут быть добавлены в подходящий субстратный материал индивидуально, т.е. в виде отдельных ферментных композиций или в виде одноферментных смесей, в которых находятся все целлюлазы и гемицеллюлазы до добавления к субстрату. В тех случаях, когда целлюлазы и гемицеллюлазы представляют собой отдельные ферментные композиции, они могут быть добавлены к субстрату последовательно или одновременно. В тех случаях, когда целлюлазы и гемицеллюлазы находятся в одной смеси, их добавляют одновременно.
Другие аспекты и варианты осуществления композиций и способов могут быть еще понятнее с учетом следующих примеров, которые не следует истолковывать как ограничивающие. Специалистам в данной области будет очевидно, что многие модификации как материалов, так и способов, могут быть сделаны без отклонения от представленных сведений.
ПРИМЕРЫ
ACCELLERASE 1000™ (Danisco A/S, Genencor Division, Palo Alto, CA), полную жидкую питательную среду нейтрализованного клеточного материала, которая содержит смесь цельной целлюлазы T. reesei, дополненную β-глюкозидазой T. reesei BGLU1, использовали в качестве источника целлюлаз. MULTIFECT® Xylanase (Danisco A/S, Genencor Division, Palo Alto, CA), ксиланазу II, с высоким pI, составленный продукт, использовали в качестве источника XYN2.
Гемицеллюлазы T. reesei индивидуально гиперэкспрессировали в штамме T. reesei, в котором были делетированы гены, кодирующие CBHI, CBHII, EG1 и EG2, во избежание присутствия этих целлюлаз в полученном в результате клеточном материале (например, кондиционированной среде или “жидких питательных средах”). Гемицеллюлазы, представляющие интерес, находятся в диапазоне от <10% до 85% общего белка в этих жидких питательных средах. Во многих случаях жидкие среды использовали непосредственно; однако некоторые гемицеллюлазы дополнительно очищали для того, чтобы показать, что наблюдаемые активности не являлись результатом действия других белков, находящихся в жидкой питательной среде.
Аббревиатуры, SEQ ID NO полипептидов, и семейство Carbohydrate-Active enZymes (CAZY) и обозначения рода (где известны) конкретных ферментов приведены в таблице 1. Приведенный выше полипептид XYN2 имеет аминокислотную последовательность SEQ ID NO: 1 и представляет собой семейство ферментов GH11 Clan C. Аминокислотные последовательности незрелых полипептидов также показаны ниже.
XYN2 (SEQ ID NO: 1)
MVSFTSLLAASPPSRASCRPAAEVESVAVEKRQTIQPGTGYNNGYFYSYWNDGHGGVTYTNGPGGQFSVNWSNSGNFVGGKGWQPGTKNKVINFSGSYNPNGNSYLSVYGWSRNPLIEYYIVENFGTYNPSTGATKLGEVTSDGSVYDIYRTQRVNQPSIIGTATFYQYWSVRRNHRSSGSVNTANHFNAWAQQGLTLGTMDYQIVAVEGYFSSGSASITVS
XYN3 (SEQ ID NO: 2)
MKANVILCLLAPLVAALPTETIHLDPELAALRANLTERTADLWDRQASQSIDQLIKRKGKLYFGTATDRGLLQREKNAAIIQADLGQVTPENSMKWQSLENNQGQLNWGDADYLVNFAQQNGKSIRGHTLIWHSQLPAWVNNINNADTLRQVIRTHVSTVVGRYKGKIRAWDVVNEIFNEDGTLRSSVFSRLLGEEFVSIAFRAARDADPSARLYINDYNLDRANYGKVNGLKTYVSKWISQGVPIDGIGSQSHLSGGGGSGTLGALQQLATVPVTELAITELDIQGAPTTDYTQVVQACLSVSKCVGITVWGISDKDSWRASTNPLLFDANFNPKPAYNSIVGILQ
ABF1 (SEQ ID NO: 3)
MLSNARIIAAGCIAAGSLVAAGPCDIYSSGGTPCVAAHSTTRALFSAYTGPLYQVKRGSDGATTAISPLSSGVANAAAQDAFCAGTTCLITIIYDQSGRGNHLTQAPPGGFSGPESNGYDNLASAIGAPVTLNGQKAYGVFVSPGTGYRNNAASGTAKGDAAEGMYAVLDGTHYNGACCFDYGNAETNSRDTGNGHMEAIYFGDSTVWGTGSGKGPWIMADLENGLFSGSSPGNNAGDPSISYRFVTAAIKGQPNQWAIRGGNAASGSLSTFYSGARPQVSGYNPMSKEGAIILGIGGDNSNGAQGTFYEGVMTSGYPSDATENSVQANIVAARYAVAPLTSGPALTVGSSISLRATTACCTTRYIAHSGSTVNTQVVSSSSATALKQQASWTVRAGLANNACFSFESRDTSGSYIRHSNFGLVLNANDGSKLFAEDATFCTQAGINGQGSSIRSWSYPTRYFRHYNNTLYIASNGGVHVFDATAAFNDDVSFVVSGGFA
ABF2 (SEQ ID NO: 4)
MELKALSAVVLSFVTLVAAAPATCTLPSTYRWNSTGALASPKSGWVSLKDFSHVIYNGQHLVWGSTHDTGTIWGSMNFGLFSDWSNMATASQNKMTPGTVAPTVFYFAPKNIWVLAYQWGPTTFSYLTSSNPSSVNGWSSPQPLFSGSISGSSPLDQTVIGDSTNMYLFFAGDDGKIYRASMPIGNFPGSFGSTSTVVLSDERNNLFEAVQVYTVSGQKQYLMIVEAIGANGRYFRSFTATNLGGTWTPQATSESQPFAGKANSGATWTNDISHGDLIRSNPDQTMTIDPCNLQFLYQGRATNSGGDYGLLPYRPGLLTLQR
ABF3 (SEQ ID NO: 5)
MSPRTDRRRSGLLALGLVAASPLATAGPCDIYASGGTPCVAAHSTTRALYGAYSGPLYQVSRGSDGATTNIATLSAGGVANAAAQDSFCAGTTCLITVIYDQSGRGNHLTQAPPGGAASGPQPNGYDNLASAIGAPVRLNGQKAYGVFIAPFTGYRNNQPNGTATGDQPQGMYAIFDGTHYNTGCCFDYGNAETNSLDTGNGHMEAIYFGTGDGSGRGTGSGSGPWIMADLENGLFSGYDPINNPADPTINFRFVTAVVKGEPGQWAIRGGDATSGTLSTFYSGQRPANGYNPMSKEGAIILGIGGDNSNRAQGTFYEGVMTSGYPSDSTENAVQANLVAAKYVYDTSLMTSGPALSVGSSISLRATTSCCTNRYIAHTGATVNTQVVSSSSSTALKQQASWTVRTGLGNSACFSFESRDSPGSFIRHSNYQLMVNANDNSKLFHEDATFCPQAGLNGQGNSFRSWSYPTRYWRHFNSLGYIAANGGEHDFDTTTLFNDDVSFVVSAGFA
AGL1 (SEQ ID NO: 6)
MTPHSIDRAARPSVWSGLALLLSTAHAIVMPDGVTGKVPSLGWNSWNAYHCDIDESKFLSAAEVIVSSGLLDAGYNYVNIDDCWSMKDGRVDGHIAVNTTRFPDGIDGLAKKVHDLGLKLGIYSTAGTATCAGYPASLGYEDVDAADFADWGVDYLKYDNCNVPSDWQDEYVACAPDAVQTGPNGTCSTALEPNLAPPGYDWSTSKSAERFNAMRNALAKQSREIVLSLCIWGVADVFSWGNETGISWRMSGDISPEWGSVTHIINMNSFKMNSVGFWGHNDADILEVGNGNLTAAETRTHFALWAAMKSPLLIGTDLAQLSQENIELLKNKHLLAFNQDSVYGQPATPYKWGVNPDWTFNYTNPAEYWAGPSSKGHLVLMMNTLDHTVRKEAKWSEIPGLSAGRYEVRDVWTDKSLGCLSSYKTAVAAHDTAVILVGKKCRNW
AGL2 (SEQ ID NO: 7)
MLGAPSPRRLADVLAVTAGLVASVRAASPISVSGKSFALNGDNVSYRFHVDDDSKDLIGDHFGGPATEDGVFPPIIGPIQGWVDLIGRQRREFPDLGRGDFRTPAVHIRQAAGYTVSDFQYKSHRVVEGKPALRGLPSTFGDAGDVSTLVVHMYDNYSSVAADLTYSIFPKYDAIVRSVNITNMGKGNITIEKLASLSVDLPYEDFDMLELKGDWAREGKRLRRKVDYGSQGFGSTTGYSSHLHNPFFSLITPTTTESQGEAWGFSLVYTGSFSVEVEKGSQGLTRAAIGVNPYQLSWPLGPGETFSSPEAVAVFSTTGVGGMSRKFHNLYRKHLIKSKFATQMHPVLLNSWEGLGFDYNDTTILHLAQESADLGIKLFVLDDGWFGVKHPRVSDNAGLGDWEANPKRFPQGLPDFISDVTKLKVANSSDHLQFGLWFEPEMVNPNSTLYMEHPDWAIHAGSYPRTLTRNQLVLNVALPEVQDFIIESLSNILSNASISYVKWDNNRGIHEAPYPGLDYAYMLGLYRVFDTLSSKFPNVRWEGCASGGGRFDPGVLQYFPHIWTSDDTDAVERIAIQFGTSLVYPPSAMGAHVSAVPNGQTQRTTSIAFRAHVAMMGGSFGFELTPAEMPEDDKAQIPGIIALAEKVNPIVVKGDMWRLSLPEESNWPAALFISQDGSQAVLFYFQIRANINNAWPVLRLQGLDASAKYKIDGNQTFSGATLMNIGLQYQFNGDYDSKVVFLEKQT
AGL3 (SEQ ID NO: 8)
MSPSAAVLIPLAAAVLLRPVVGQTQCGGNLYTPGTLNFTLECYNAFQDCVAQFEANASQVDCNDGKGNLFMQQQANLGASPGSQNNDAIIAFQDIRDLCLLSGSTTATWGYSDNQWYWAAAEDACYTNDPTRTDVVKTHPAPFCIQNRDSSLPECYPQPDATPPGGPLKVIKTAKTRNGFKSSARGWNTYGVQALVNGSQVVPSFAGQSGLFYTQKFVETQCGVLARPEFKKAGYDLCSLDSGWQATTAVDQHGRIIYNTTRFNLPELASWLHKRDLKLGVYITPGVPCLAHNQTILGTNIKIKDVLNGNNDQINCDFDFRKDGVQQWHDSVVAQWASWGVDMLKLDFLTPGSPSNGANLACDSSDAVRAYQKAIKKSGRKIRLDISWKLCRNETWLPIWSDLAESMRTDQDLDNYGTNTLMAWQVGQRAIENYRQYIGLQAQRNVPLTIYPDMDALFTVNPEHLAGVNDTIRYTVQNHWLGAGANLIIGGDMEQVDALGLKLTTSKQSIDAADFFAKYPMQPRNPGTGSNAAKQLQAWIGGPSDDHEAYVLIVNYGPDLGNGGFSTKLYGKQKVTVSLKDLGISGSAWTFTDIWSGKSSRVTGSYSAWLTEGESQLLRLKRTH
AXE1 (SEQ ID NO: 9)
MPSVKETLTLLLSQAFLATGSPVDGETVVKRQCPAIHVFGARETTVSQGYGSSATVVNLVIQAHPGTTSEAIVYPACGGQASCGGISYANSVVNGTNAAAAAINNFHNSCPDTQLVLVGYSQGAQIFDNALCGGGDPGEGITNTAVPLTAGAVSAVKAAIFMGDPRNIHGLPYNVGTCTTQGFDARPAGFVCPSASKIKSYCDAADPYCCTGNDPNVHQGYGQEYGQQALAFINSQLSSGGSQPPGGGPTSTSRPTSTRTGSSPGPTQTHWGQCGGQGWTGPTQCESGTTCQVISQWYSQCL
AXE3 (SEQ ID NO: 10)
MPSIKSTVTFLLSQALLATATPMDLEKRQCPGIHVFGARETTAPPGYGSSATVVNLIINAHPGTTAEAINYPACGGQAQCGGISYANSVVAGINAVVQAVTNFHNRCPSTKLVLVGYSQGGQIMDDALCGGGDPAEGYPNTAVPLPAAAVSAIRAAIFMGDPRYVHGLAYNVGSCQAQGFAPRNVGFVCPSGNKIKSYCDASDPYCCNGNNANTHQGYGQEYGQQALAFVNSLLG
EG6 (SEQ ID NO: 11)
MKVSRVLALVLGAVIPAHAAFSWKNVKLGGGGGFVPGIIFHPKTKGVAYARTDIGGLYRLNADDSWTAVTDGIADNAGWHNWGIDAVALDPQDDQKVYAAVGMYTNSWDPSNGAIIRSSDRGATWSFTNLPFKVGGNMPGRGAGERLAVDPANSNIIYFGARSGNGLWKSTDGGVTFSKVSSFTATGTYIPDPSDSNGYNSDKQGLMWVTFDSTSSTTGGATSRIFVGTADNITASVYVSTNAGSTWSAVPGQPGKYFPHKAKLQPAEKALYLTYSDGTGPYDGTLGSVWRYDIAGGTWKDITPVSGSDLYFGFGGLGLDLQKPGTLVVASLNSWWPDAQLFRSTDSGTTWSPIWAWASYPTETYYYSISTPKAPWIKNNFIDVTSESPSDGLIKRLGWMIESLEIDPTDSNHWLYGTGMTIFGGHDLTNWDTRHNVSIQSLADGIEEFSVQDLASAPGGSELLAAVGDDNGFTFASRNDLGTSPQTVWATPTWATSTSVDYAGNSVKSVVRVGNTAGTQQVAISSDGGATWSIDYAADTSMNGGTVAYSADGDTILWSTASSGVQRSQFQGSFASVSSLPAGAVIASDKKTNSVFYAGSGSTFYVSKDTGSSFTRGPKLGSAGTIRDIAAHPTTAGTLYVSTDVGIFRSTDSGTTFGQVSTALTNTYQIALGVGSGSNWNLYAFGTGPSGARLYASGDSGASWTDIQGSQGFGSIDSTKVAGSGSTAGQVYVGTNGRGVFYAQGTVGGGTGGTSSSTKQSSSSTSSASSSTTLRSSVVSTTRASTVTSSRTSSAAGPTGSGVAGHYAQCGGIGWTGPTQCVAPYVCQKQNDYYYQCV
EG8 (SEQ ID NO: 12)
MRATSLLAAALAVAGDALAGKIKYLGVAIPGIDFGCDIDGSCPTDTSSVPLLSYKGGDGAGQMKHFAEDDGLNVFRISATWQFVLNNTVDGKLDELNWGSYNKVVNACLETGAYCMIDMHNFARYNGGIIGQGGVSDDIFVDLWVQIAKYYEDNDKIIFGLMNEPHDLDIEIWAQTCQKVVTAIRKAGATSQMILLPGTNFASVETYVSTGSAEALGKITNPDGSTDLLYFDVHKYLDINNSGSHAECTTDNVDAFNDFADWLRQNKRQAIISETGASMEPSCMTAFCAQNKAISENSDVYIGFVGWGAGSFDTSYILTLTPLGKPGNYTDNKLMNECILDQFTLDEKYRPTPTSISTAAEETATATATSDGDAPSTTKPIFREETASPTPNAVTKPSPDTSDSSDDDKDSAASMSAQGLTGTVLFTVAALGYMLVAF
GLR1 (SEQ ID NO: 13)
MVIRSLLLLLLAAIVPVFAESGIDAWLRYARLPSSATRGHLTSFPDRIVVLNASKNGPLASASSELHKGIKGILGLDLDVSSRGGKHCSTQKSIVISTLDTYQSACGKLSPKLNLKEDGYWLSTKGGSVQIIGQNERGALYGAFQYLSYLGQGDFSGKAFASNPSAPVRWSNQWDNLNAATAAHGSIERGYGGPSIFFENGLIKEDLSRVPLYGRLLASVGLNGIVINNVNADANLLNETNLQGLKRIADLFRPWGVNVGISLNFASPQVLGDLSTFDPLDDSVIKWWTDKTDRIYQLVPDLAGYLVKANSEGQPGPLTYNRTLAEGANLFAKAVQPHGGIVVFRAFVYDQLNETDWKADRANAAVDFFKSLDGQFDDNVLVQIKYGPIDFQVREPASPLFANLPKTAVSIELEVTQEYLGQQSHLVYLPPLWQTVLGFDMRYNNRQSYVRDIISGEVFGHKLGGYAGVINVGMDDTWLGSHLAMSNMFAYGRLAWNPRADSRDIVEEWTRLTFGLDRDVVSTIADMSLKSWPAYEGYSGNLGIQTLTDILYTHYGANPASQDNNGWGQWTRADSKTIGMDRTVSNGTGNAGQYPKEVAARFEHTQTTPDDLMLWFHHVPYTFRLHSGKSVIQHFYDAHYTGAATVQRFPAAWKSLKSKIDTERYNAVLYKLQYQTGHSLVWRDAITEFYRNLSSIPDQLNRVRNHPHRIEAEDMDLSGFTVVNVSPTECASKYKAIATNGTGTATTRLNVPSGKYTVAVNYYDVINGTASYDVLLNGKSLGKWKGDSETHLGHDFSTFLDCHSAIRITFEGVRISRGDKLTIRGTGNAQEQAAIDYVSILPQGVVD
MAN1 (SEQ ID NO: 14)
MMMLSKSLLSAATAASALAAVLQPVPRASSFVTISGTQFNIDGKVGYFAGTNCYWCSFLTNHADVDSTFSHISSSGLKVVRVWGFNDVNTQPSPGQIWFQKLSATGSTINTGADGLQTLDYVVQSAEQHNLKLIIPFVNNWSDYGGINAYVNAFGGNATTWYTNTAAQTQYRKYVQAVVSRYANSTAIFAWELGNEPRCNGCSTDVIVQWATSVSQYVKSLDSNHLVTLGDEGLGLSTGDGAYPYTYGEGTDFAKNVQIKSLDFGTFHLYPDSWGTNYTWGNGWIQTHAAACLAAGKPCVFEEYGAQQNPCTNEAPWQTTSLTTRGMGGDMFWQWGDTFANGAQSNSDPYTVWYNSSNWQCLVKNHVDAINGGTTTPPPVSSTTTTSSRTSSTPPPPGGSCSPLYGQCGGSGYTGPTCCAQGTCIYSNYWYSQCLNT
PEC2 (SEQ ID NO: 15)
MLKLSLFLGAVTASLCVQAHAVPPPTVTQAPKLEDRATTCTFSGSNGASSASKSQKSCATIVLSNVAVPSGVTLDLSDLNDGTTVIFEGTTTWGYKEWSGPLLQIEGNDITIQGASGAVLNPDGARWWDGQGGNGGKTKPKFFAAHDLTSSSITNLYIKNTPVQAVSVNGVNGLTITGMTIDNSAGDSGGGHNTDGFDIGSSSNVVISGAKVYNQDDCVAVNSGTNITFTGGLCSGGHGLSIGSVGGRDDNTVQTVTFSNSQVTKSANGIRIKATAGKTGTIKGVTYTGITLSSITGYGILIEQNYDGGDLHGSPTSGIPITNLVLQNISGSNGVVSSGNNIAIVCGSGACSNWTWSNVVVTGGKKYGSCQNVPSPATC
XYN1 (SEQ ID NO: 16)
MVAFSSLICALTSIASTLAMPTGLEPESSVNVTERGMYDFVLGAHNDHRRRASINYDQNYQTGGQVSYSPSNTGFSVNWNTQDDFVVGVGWTTGSSAPINFGGSFSVNSGTGLLSVYGWSTNPLVEYYIMEDNHNYPAQGTVKGTVTSDGATYTIWENTRVNEPSIQGTATFNQYISVRNSPRTSGTVTVQNHFNAWASLGLHLGQMNYQVVAVEGWGGSGSASQSVSN
BXL1 (SEQ ID NO: 17)
MVNNAALLAALSALLPTALAQNNQTYANYSAQGQPDLYPETLATLTLSFPDCEHGPLKNNLVCDSSAGYVERAQALISLFTLEELILNTQNSGPGVPRLGLPNYQVWNEALHGLDRANFATKGGQFEWATSFPMPILTTAALNRTLIHQIADIISTQARAFSNSGRYGLDVYAPNVNGFRSPLWGRGQETPGEDAFFLSSAYTYEYITGIQGGVDPEHLKVAATVKHFAGYDLENWNNQSRLGFDAIITQQDLSEYYTPQFLAAARYAKSRSLMCAYNSVNGVPSCANSFFLQTLLRESWGFPEWGYVSSDCDAVYNVFNPHDYASNQSSAAASSLRAGTDIDCGQTYPWHLNESFVAGEVSRGEIERSVTRLYANLVRLGYFDKKNQYRSLGWKDVVKTDAWNISYEAAVEGIVLLKNDGTLPLSKKVRSIALIGPWANATTQMQGNYYGPAPYLISPLEAAKKAGYHVNFELGTEIAGNSTTGFAKAIAAAKKSDAIIYLGGIDNTIEQEGADRTDIAWPGNQLDLIKQLSEVGKPLVVLQMGGGQVDSSSLKSNKKVNSLVWGGYPGQSGGVALFDILSGKRAPAGRLVTTQYPAEYVHQFPQNDMNLRPDGKSNPGQTYIWYTGKPVYEFGSGLFYTTFKETLASHPKSLKFNTSSILSAPHPGYTYSEQIPVFTFEANIKNSGKTESPYTAMLFVRTSNAGPAPYPNKWLVGFDRLADIKPGHSSKLSIPIPVSALARVDSHGNRIVYPGKYELALNTDESVKLEFELVGEEVTIENWPLEEQQIKDATPDA
Секретируемые белковые жидкие питательные среды, экспрессирующие ABF1, ABF2, ABF3, AGL1, AGL2, AGL3, AXE1, AXE3, EG6, EG8, GLR1, MAN1, PEC2, XYN1, XYN3 и BXL1, тестировали в трехкомпонентных смесях. 150 мкл AFEX-предварительно обработанной кукурузной соломы (31,7% глюкана, 19,1% ксилана, 1,83% галактана и 3,4% арабинана, исходя из сухой массы, полученной в виде либо 15,6, либо 12% взвеси сухих веществ в pH 5 50 мМ ацетат-натриевом буфере) добавляли в каждую лунку 96-луночного микротитровального планшета (все экспериментальные точки основаны на лунках в трех повторах). В одном эксперименте (показан в таблице 9) использована разбавленная предварительно обработанная аммиаком сердцевина кукурузного початка при 13,84% твердых веществ в качестве субстрата. К выбранным лункам добавляли только ACCELLERASE 1000TM (CEL) при 20 мг/г целлюлозы, ACCELLERASE 1000TM при 20 мг/г + 5 мг/г MULTIFECT® Xylanase, или ACCELLERASE 1000TM при 20 мг/г + 5 мг/г MULTIFECT® Xylanase + 1 или 5 мг/г отдельных гемицеллюлазных бульонов все в 20 мкл общего объема фермента.
Дозы фермента приводили в соответствие с суммарной целлюлозой в каждой взвеси субстрата (15,6% или 12% сухого вещества). Планшеты герметично закрывали и инкубировали со встряхиванием при 50°C в течение 72 часов. Реакции затем гасили с использованием 100 мкл 100 мМ глицина, pH 10. Эту смесь фильтровали и разбавляли дополнительно 6× (20 мкл + 100 мкл дистиллированной H2O) и анализировали на содержание сахара на колонке HPLC-Aminex HPX-87P на приборе Agilent Chem Station HPLC. Области пиков HPLC преобразовывали в концентрации сахара на основе стандартной кривой целлобиозы для целлобиозы и глюкозы или ксилозной стандартной кривой для ксилозы. Процентное преобразование на основе исходного содержания целлюлозы вычисляли для включения H2O гидролиза для каждого из трех сахарных полимеров. Также вычисляли стандартные отклонения трех повторов.
В таблице 2 и 3 представлены средние значения конверсии (± стандартное отклонение) глюканов и ксиланов для каждой ферментной смеси, как определено в двух отдельных выполнениях протокола. Эти отдельные серии проводили с двумя различными субстратными взвесями AFEX 15,6% (таблица 2) и 12% сухих веществ (таблица 3), и таким образом, содержат различные общие мг целлюлозы, хотя доза в виде мг/г целлюлозы является той же.
Добавление XYN2 было эффективным при увеличении конверсии ксилана. Шесть ферментных смесей с третьим компонентом (т.е., XYN3, AGL2, EG8, BXL1, ABF3 или PEC2) показали дополнительные преимущества относительно конверсии глюкана и/или ксилана по сравнению с целлюлазой с XYN2. Четырехкомпонентная ферментная смесь была выполнена в соответствии с описанной выше процедурой. В таблице 4 приведено среднее значение конверсии (± стандартное отклонение) глюканов и ксиланов для каждой ферментной смеси.
В другом эксперименте ACCELLERASE 1000™ смешивали с очищенной XYN2 и/или XYN3 и анализировали (таблица 5). Сочетание XYN2 и XYN3 давало более эффективную конверсию глюкана и ксилана.
В дополнительном примере XYN4, XYN5, FAE1 и новая серия ABF3 с ~50% белка, представляющего интерес (по сравнению с предыдущей серией с <10%), тестировали, как описано выше в смесях, содержащих 20 мг/г ACCELLERASE 1000™ + 5 мг/г MULTIFECT® Xylanase XYN2. Результаты представлены в Таблице 6. Добавление XYN4, XYN5 или FAE1 было эффективным в повышении конверсии глюкана и ксилана.
В другом эксперименте ACCELLERASE 1000™ смешивали с очищенной Bxl1 и XYN2 и/или XYN3 и анализировали, как описано выше. Результаты показаны в таблице 7. Некоторые сочетания ферментов были эффективны в повышении конверсии глюкана и/или ксилана.
В другом примере ABF1, ABF2 и ABF3 (ABF3 серия образцов с <10% белка, представляющего интерес), по одному, в двухкомпонентных и трехкомпонентных сочетаниях добавляли к основе 20 мг/г ACCELLERASE 1000™ + 5 мг/г очищенной XYN3 + 5 мг/г очищенной BXL1. Результаты показаны в таблице 8. Некоторые сочетания ферментов были эффективными в повышении конверсии глюкана и/или ксилана.
В другом примере 3,4 мг/г ксилана очищенной ABF1, ABF2 и/или ABF3 добавляли к 20,7 мг/г глюкана ACCELLERASE 1000™ + 5,1 мг/г ксилана каждой из очищенных XYN3 и BXL1. Результаты показаны в таблице 9. Некоторые сочетания ферментов были эффективны в повышении конверсии глюкана и/или ксилана.
Реферат
Описаны композиции ферментной смеси для гидролиза смеси целлюлозных и гемицеллюлозных материалов и способы гидролиза таких смесей. Представленные ферментные смеси включают три композиции, первая из которых содержит смесь цельной целлюлазы T.reesei, дополненную β-глюкозидазой Т. reesei BGLU1, вторая композиция состоит из по меньшей мере одной ксиланазы, выбранной из ксиланазы GH10 или GH11, и третья композиция состоит из по меньшей мере одной дополнительной гемицеллюлазы, которая представляет собой сочетание β-ксилозидазы и двух или более арабинофуранозидаз. Представленные способы предусматривают контактирование смеси целлюлозных и гемицеллюлозных материалов с указанными композициями. Охарактеризованные изобретения позволяют повысить конверсию глюкана и ксилана и могут быть использованы при превращении биомассы в сахар. 4 н. и 8 з.п. ф-лы, 10 табл.
Формула
(a) первую ферментную композицию, состоящую из полной питательной среды, которая содержит смесь цельной целлюлазы Т. reesei, дополненную β-глюкозидазой Т. reesei BGLU1, где целлюлаза составляет от 1% до 50% от общего белка в композиции ферментной смеси,
(b) вторую ферментную композицию, состоящую из по меньшей мере одной ксиланазы, выбранной из ксиланазы GH10 или GH11, и
(c) третью ферментную композицию, состоящую из по меньшей мере одной дополнительной гемицеллюлазы, где одна дополнительная гемицеллюлаза представляет собой сочетание β-ксилозидазы BXL1 из Trichoderma reesei и двух или более арабинофуранозидаз, выбранных из ABF1, ABF2 или ABF3 из Trichoderma reesei, и гемицеллюлаза составляет от 1% до 50% от общего белка в композиции ферментной смеси, где
композиция ферментной смеси обеспечивает по меньшей мере одно из следующего: (i) повышенную конверсию глюкана или (ii) повышенную конверсию ксилана по сравнению с эквивалентной композицией ферментной смеси, лишенной по меньшей мере одной дополнительной гемицеллюлазы.
(a) первую ферментную композицию, состоящую из полной питательной среды, которая содержит смесь цельной целлюлазы Т. reesei, дополненную β-глюкозидазой Т. reesei BGLU1, где целлюлаза составляет от 1% до 50% от общего белка в композиции ферментной смеси,
(b) вторую ферментную композицию, состоящую из по меньшей мере одной ксиланазы, выбранной из ксиланазы GH10 или GH11, и
(c) третью ферментную композицию, состоящую из по меньшей мере одной дополнительной гемицеллюлазы, где одна дополнительная гемицеллюлаза представляет собой сочетание β-ксилозидазы и двух или более арабинофуранозидаз, и гемицеллюлаза составляет от 1% до 50% от общего белка в композиции ферментной смеси,
где β-ксилозидаза имеет аминокислотную последовательность, имеющую по меньшей мере 90% идентичность SEQ ID NO:17, и
две или более арабинофуранозидазы выбраны из арабинофуранозидазы, которая имеет аминокислотную последовательность, имеющую по меньшей мере 90% идентичность SEQ ID NO:3, арабинофуранозидазы, которая имеет аминокислотную последовательность, имеющую по меньшей мере 90% идентичность SEQ ID NO:4, и арабинофуранозидазы, которая имеет аминокислотную последовательность, имеющую по меньшей мере 90% идентичность SEQ ID NO:5,
где композиция ферментной смеси обеспечивает по меньшей мере одно из следующего: (i) повышенную конверсию глюкана или (ii) повышенную конверсию ксилана по сравнению с эквивалентной композицией ферментной смеси, лишенной по меньшей мере одной дополнительной гемицеллюлазы.
(a) первой ферментной композицией, состоящей из полной питательной среды, которая содержит смесь цельной целлюлазы Т. reesei, дополненную β-глюкозидазой Т. reesei BGLU1, где целлюлаза составляет от 1% до 50% от общего белка в композиции ферментной смеси,
(b) второй ферментной композицией, состоящей из по меньшей мере одной ксиланазы, выбранной из ксиланазы GH10 или GH11, и
(c) третьей ферментной композицией, состоящей из по меньшей мере одной дополнительной гемицеллюлазы, где одна дополнительная гемицеллюлаза представляет собой сочетание β-ксилозидазы BXL1 из Trichoderma reesei и двух или более арабинофуранозидаз, выбранных из ABF1, ABF2 или ABF3 из Trichoderma reesei, и гемицеллюлаза составляет от 1% до 50% от общего белка в композиции ферментной смеси,
где контактирование в результате приводит по меньшей мере к одному из следующего: (i) повышенной конверсии глюкана и (ii) повышенной конверсии ксилана по сравнению с эквивалентным контактированием в отсутствие по меньшей мере одной дополнительной гемицеллюлазы.
(a) первой ферментной композицией, состоящей из полной питательной среды, которая содержит смесь цельной целлюлазы Т. reesei, дополненную β-глюкозидазой Т. reesei BGLU1, где целлюлаза составляет от 1% до 50% от общего белка в композиции ферментной смеси,
(b) второй ферментной композицией, состоящей из по меньшей мере одной ксиланазы, выбранной из ксиланазы GH10 или GH11, и
(c) третьей ферментной композицией, состоящей из по меньшей мере одной дополнительной гемицеллюлазы, где одна дополнительная гемицеллюлаза представляет собой сочетание β-ксилозидазы и двух или более арабинофуранозидаз, и гемицеллюлаза составляет от 1% до 50% от общего белка в композиции ферментной смеси,
где β-ксилозидаза имеет аминокислотную последовательность, имеющую по меньшей мере 90% идентичность SEQ ID NO:17, и
две или более арабинофуранозидазы выбраны из арабинофуранозидазы, которая имеет аминокислотную последовательность, имеющую по меньшей мере 90% идентичность SEQ ID NO:3, арабинофуранозидазы, которая имеет аминокислотную последовательность, имеющую по меньшей мере 90% идентичность SEQ ID NO:4 и арабинофуранозидазы, которая имеет аминокислотную последовательность, имеющую по меньшей мере 90% идентичность SEQ ID NO:5,
где контактирование в результате приводит по меньшей мере к одному из следующего: (i) повышенной конверсии глюкана и (ii) повышенной конверсии ксилана по сравнению с эквивалентным контактированием в отсутствие по меньшей мере одной дополнительной гемицеллюлазы.
Комментарии