Рекомбинантные клетки дрожжей, продуцирующие полимолочную кислоту, и их использования - RU2758138C2
Код документа: RU2758138C2
Чертежи







Описание
Настоящее изобретение относится к рекомбинантным клеткам дрожжей, обладающим способностью продуцировать полимолочную кислоту, и к их использованиям. Настоящее изобретение дополнительно относится к способу получения полимолочной кислоты с использованием таких рекомбинантных клеток дрожжей и более конкретно гомополимеров полимолочной кислоты, а также к ферментативным бульонам или бесклеточным экстрактам указанных рекомбинантных клеток дрожжей, содержащим полимолочную кислоту.
ПРЕДПОСЫЛКИ
Полимолочная кислота (PLA) представляет собой биоразрушаемый сложный полиэфир, который можно извлекать из возобновляемых ресурсов, таких как кукуруза, рис или другие растения, продуцирующие сахара или крахмал. PLA можно легко обрабатывать посредством экструзии, инжекции, формования, формирования пленок и т. д., что делает возможным широкий диапазон применений, в частности, для краткосрочных использований (т. е. пищевая упаковка, мешки, пленки и т. п.). Кроме того, в силу нетоксичности мономеров молочной кислоты, PLA представляет собой один из наиболее перспективных биополимеров для медицинских применений. Из-за их превосходной биосовместимости и механических свойств, PLA и их сополимеры получают широкое использование в технологии культивирования тканей для восстановления функции нарушенных тканей, в системах доставки лекарственных средств и в различных медицинских имплантах.
Полимеры молочной кислоты можно синтезировать с помощью различных процессов с тем, чтобы получать продукты с богатым спектром химических и механических свойств. Более конкретно, PLA преимущественно синтезируют с помощью двух способов: поликонденсация молочной кислоты (LA), которую осуществляют общей массой или в растворе; или полимеризация с раскрытием кольца лактида (циклического димера молочной кислоты), для которой необходимы катализаторы. Прямую поликонденсацию молочной кислоты общей массой не применяют в большом масштабе по причине конкурирующей реакции образования лактида и одновременно протекающего процесса разложения. Поликонденсация молочной кислоты в растворе дает PLA с молекулярными массами в диапазоне от десятков до нескольких сотен тысяч г/моль. До сих пор синтез PLA из лактида является наиболее эффективным способом синтеза в промышленности. Однако использование металлических катализаторов, катионных катализаторов и/или органических катализаторов может влиять на качество конечного продукта, некоторые остатки катализаторов встраиваются в полимер. Кроме того, рацемизация части лактидов во время процесса может вести к гетерополимерам, содержащим как L-молочную кислоту, так и D-молочную кислоту. Вдобавок эта химическая полимеризация требует подвода больших энергий (тепла), что налагает дополнительное экономическое и экологическое обременение.
В последнее время разработаны альтернативные биологические процессы, в которых прокариотические клетки, такие как бактерии, сконструированы для получения микробных сложных полиэфиров на основе LA. При этом биологическом получении используют преимущество ферментативной активности синтазы полигидроксиалканоата (PHA), что ведет к продуцированию сложных полиэфиров на основе LA. Более точно, разработан путь для утилизации лактил-КоА в качестве субстрата для продуцирования сложных полиэфиров на основе LA в бактериях. Однако основной полимер, получаемый с помощью этого биологического процесса, представляет собой сополимер, состоящий из LA и других мономеров PHA (оксикислоты или гидроксиалканоат, такой как 3-гидроксибутират). Получаемые сополимеры, такие как P(3HB-co-LA) представляют ограниченный механический и промышленный интерес. Кроме того, полимеры PLA, создаваемые с помощью этого способа, аморфны и имеют низкую молекулярную массу (меньше чем 30000 г/моль). Такая PLA проявляет низкие механические свойства и не может быть легко обработана в таких промышленных применениях, как литье под давлением, горячее формование или экструзия. Соответственно, такой полимер представляет слабый промышленный интерес.
Настоящее изобретение описывает новые биологические способы и микроорганизмы для получения PLA. Изобретение делает возможным эффективное получение гомополимеров PLA с высокой молекулярной массой в большом масштабе.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к клеткам дрожжей, сконструированным для того, чтобы продуцировать PLA, и к их использованиям. Более конкретно, изобретение относится к рекомбинантным клеткам дрожжей, которые демонстрируют как активность лактил-КоА-синтазы, так и активность лактил-КоА-полимеразы, и, необязательно, измененный метаболизм молочной кислоты (LA), ведущий к сниженному потреблению LA в качестве источника углерода. Следовательно, изобретение описывает способность указанных выше клеток дрожжей продуцировать высокие количества PLA и что продуцируемая PLA может быть в форме гомополимеров с высокой молекулярной массой. Компартментализация клеток дрожжей и их мембраносвязанных органелл представляет особый интерес для получения гомополимеров PLA, по существу лишенных других гидроксиалканоатных мономеров. Эти клетки, таким образом, делают возможным создание усовершенствованных способов получения PLA с высокими механическими свойствами и обширной промышленной полезностью.
Следовательно, цель изобретения состоит в том, чтобы предоставить рекомбинантные клетки дрожжей, содержащие ген, кодирующий белок, обладающий активностью лактил-КоА-синтазы, и ген, кодирующий белок, обладающий активностью лактил-КоА-полимеразы, указанные рекомбинантные клетки обладают способностью продуцировать полимолочную кислоту (PLA).
В конкретном варианте осуществления белок, демонстрирующий активность лактил-КоА-синтазы, представляет собой ацил-КоА-трансферазу, более предпочтительно пропионил-КоА-трансферазу (Pctp). В другом варианте осуществления белок, демонстрирующий активность лактил-КоА-синтазы, представляет собой лигазу, такую как ацил-КоА-лигаза.
Предпочтительно белок, демонстрирующий активность лактил-КоА-полимеразы, представляет собой синтазу полигидроксиалканоата (PHA).
В предпочтительном варианте осуществления изобретение относится к рекомбинантным клеткам дрожжей, экспрессирующим по меньшей мере ген, кодирующий белок, обладающий активностью лактил-КоА-синтазы, и ген, кодирующий белок, обладающий активностью лактил-КоА-полимеразы, и в которых инактивируют активность оксидоредуктазы молочной кислоты дрожжей.
В конкретном варианте осуществления изобретение относится к рекомбинантным клеткам дрожжей, экспрессирующим по меньшей мере ген, кодирующий белок, обладающий активностью лактил-КоА-синтазы, и ген, кодирующий белок, обладающий активностью лактил-КоА-полимеразы, и в которых инактивируют активность оксидоредуктазы молочной кислоты дрожжей, и в которых удалены ацил-КоА-оксидазы.
В конкретном варианте осуществления клетка дрожжей относится к роду Yarrowia, предпочтительно Yarrowia lipolytica.
Изобретение также относится к способу получения PLA, который включает
- культивирование рекомбинантных клеток дрожжей по изобретению в присутствии молочной кислоты; и необязательно
- извлечение полученной PLA.
Дополнительная цель изобретения относится к композиции, содержащей бесклеточный экстракт рекомбинантных клеток дрожжей по изобретению.
Дополнительная цель изобретения относится к композиции, содержащей высушенные рекомбинантные клетки дрожжей по изобретению.
Изобретение дополнительно относится к использованию рекомбинантных клеток дрожжей по изобретению для получения PLA и к способу получения PLA и ее использованиям.
Изобретение, в частности, адаптировано для получения гомополимолочной кислоты, в частности поли-D-молочной кислоты (PDLA). Кроме того, описанный способ, в частности, благоприятен для получения высокомолекулярного полимера, такого как PLA, и более конкретно PDLA, с усредненной молекулярной массой (Mw) выше 40000 г/моль, например, приблизительно 80000 г/моль.
ОПИСАНИЕ ФИГУР
Фиг. 1: схематическое представление ферментативного получения полимолочной кислоты с использованием молочной кислоты и лактил-КоА в качестве субстратов; КоА-SH: кофермент A; ACS: ацил-КоА-синтаза.
Фиг. 2: изображение различных метаболических путей, ведущих к получению или потреблению ацетил-КоА у Y. lipolytica и подлежащих модификации; PDH: пируватдегидрогеназа; OAA: оксалоуксусная кислота; DGA1, DGA2: ацил-КоА:диацилглицеринацилтрансферазы; LRO1: фосфолипид:диацилглицеринацитрансфераза.
Фиг. 3: схематическое представление различных метаболических путей, которые являются субъектами экспрессии в Yarrowia lipolytica, для увеличения цитозольного запаса ацетил-КоА. GAPN: глицеральдегид-3P-дегидрогеназа; PDH: пируватдегидрогеназа; PTA: киназа фосфотрансацетилаза; ACS: ацил-КоА-синтетаза; ACK: ацетаткиназа; XPK: фосфокетолаза; PPP: пентозофосфатный путь.
Фиг. 4: кривая роста контрольного штамма JMY2341 (фиг. 4A) и штамма ThYl_434 с нокаутом дегидрогеназы молочной кислоты YlDLD1 (фиг. 4B) в средах, содержащих лактат или глюкозу (положительный контроль). Штамм с нарушенной YlDLD1 не способен расти на D-лактате и только поддерживает половину своего потенциала к росту (OD600нм) на эквимолярной рацемической смеси DL-лактата.
Фиг. 5: кривая роста контрольного штамма JMY2341 (фиг. 5A) и штамма ThYl_436 с нокаутом оксидоредуктазы молочной кислоты YlCYB21 (фиг. 5B) на L-лактате. Штамм с нокаутом YlCYB21 не способен расти на L-молочной кислоте.
Фиг. 6: схематическая иллюстрация последовательных геномных модификаций, выполняемых в штамме W29 Y. lipolytica дикого типа, в соответствии с конкретным вариантом осуществления.
Фиг. 7: ЯМР спектры для: (1) экстрагированной по Сокслету липидной фракции из высушенной биомассы контрольного штамма ThYl_964 и (2) экстрагированной по Сокслету липидной фракции из высушенной биомассы штамма ThYl_976, экспрессирующего как активность лактил-КоА-синтазы, так и активность лактил-КоА-полимеразы. Сигналы между 5,3 и 5,2 м. д. и между 1,7 и 1,5 м. д. соответствуют фракции PLA, продуцируемой рекомбинантными клетками дрожжей; (3) коммерческой PLA, используемой в качестве контроля.
Фиг. 8: эволюция биомассы и потребление субстратов L-лактата, D-лактата и глюкозы во время роста рекомбинантного штамма дрожжей ThYl_976.
Фиг. 9: графическое представление получения PLA из рекомбинантных клеток дрожжей ThYl_976 во время культивирования.
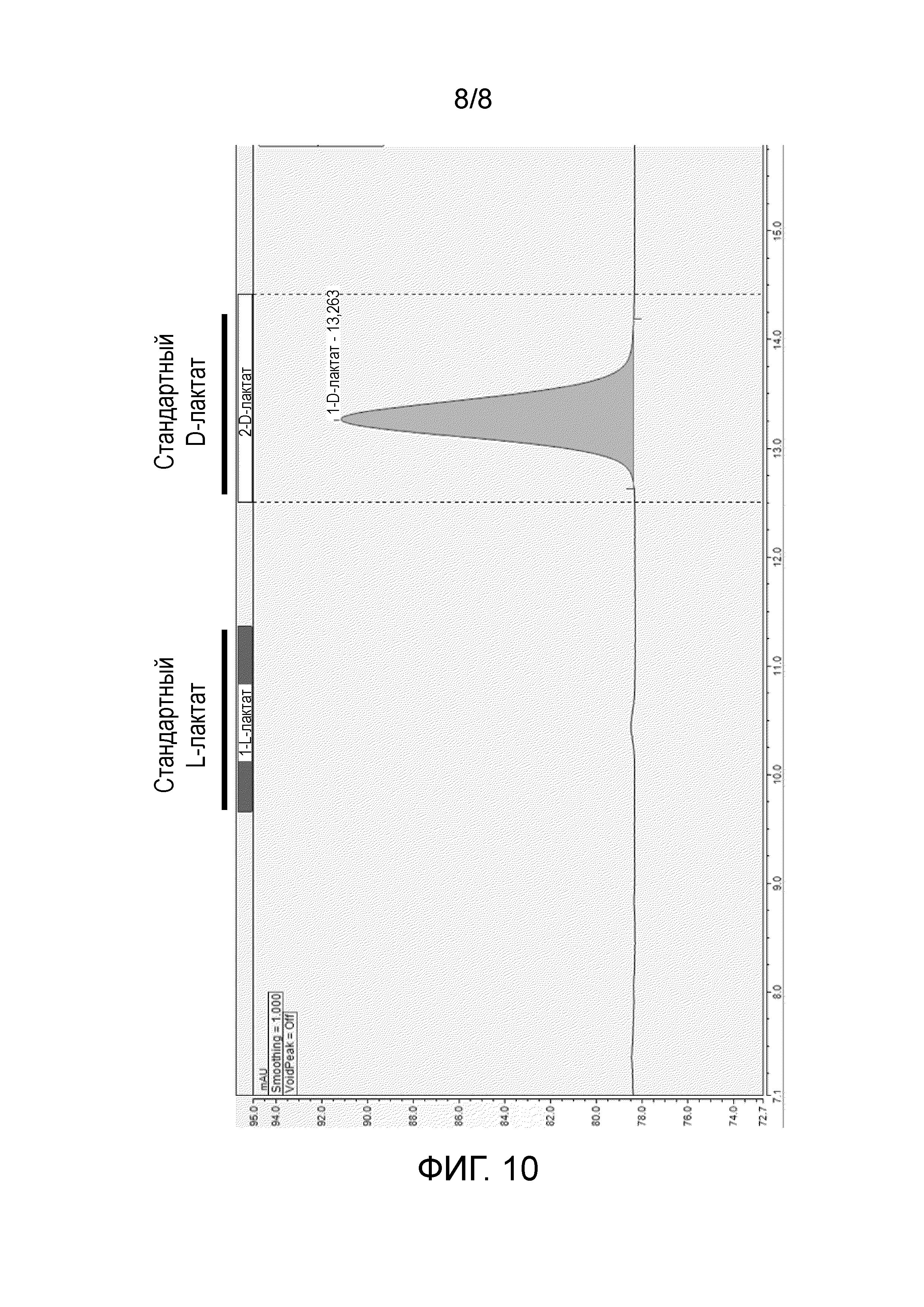
Фиг. 10: HPLC хроматограмма, полученная после полного гидролиза PLA, экстрагированной из штамма ThYl_976, и показывающая, что полученная PLA представляет собой гомополимер, состоящий исключительно из мономеров D-молочной кислоты.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новым рекомбинантным эукариотическим микроорганизмам, сконструированным для получения полимолочной кислоты. Более конкретно, изобретение относится к рекомбинантным клеткам дрожжей, сконструированным авторами изобретения, которые делают возможным крупномасштабное получение высокомолекулярных (Mw) гомополимеров PLA, обладающих примечательными механическими и физическими свойствами. Более конкретно, авторы изобретения обнаружили, что использование эукариотической клетки делает возможным получение гомополимеров PLA, не содержащих гидроксиалканоаты. В действительности, дрожжи не имеют свободных 3-гидроксиалканоатов (3HA), помимо одного в форме КоА, присутствующей в пероксисоме. Соединения 3HA образуются в форме ACP (ацил-белок-переносчик) в качестве временного промежуточного соединения во время биосинтеза жирных кислот в ограниченных реакционных камерах дрожжевой синтазы жирных кислот I типа и, таким образом, не могут быть высвобождены в цитозоль и использованы при продуцировании полимера. Указанное продуцирование 3HA-КоА, происходящее во время второй стадии процесса β-окисления, можно устранять посредством делеции генов, кодирующих белки, отвечающие за первую или вторую стадии, если продуцирование гомополимеров PLA происходит в пероксисомах. Тогда, используя клетки дрожжей, возможно продуцировать PLA, не содержащие другой гидроксиалканоат, продуцируя полимер в компартменте, не содержащем их (таком как цитозоль или митохондрии), или удаляя гены, отвечающие за их синтез в пероксисомах. Кроме того, видоизменение органелл дрожжей, таких как пероксисомы и митохондрии, в качестве продуцирующего компартмента может позволять обходить пересечения метаболических путей между сконструированным путем PLA и эндогенными путями. Дополнительное преимущество использования дрожжей, например, рода Yarrowia, состоит в их устойчивости к кислой среде, поскольку подача мономера может вызывать снижение pH. Что более важно, такая кислая среда значительно повышает захват мономера и существенно снижает риск нежелательной контаминации культуры. Кроме того, некоторые дрожжи, такие как Yarrowia lipolytica, являются GRAS микроорганизмами (общепризнанными в качестве безопасных). Они не продуцируют какие-либо эндотоксины. PLA можно очищать без какой-либо контаминации липополисахаридами, которые встречаются у бактерий и вызывают иммуногенные реакции. Гены, которые добавляют для получения штамма, представляющего интерес, встраивают в геномную хромосому дрожжей и, таким образом, более стабильны, чем экспрессия через плазмиды, как практикуют у бактерий. Изобретение, таким образом, предоставляет конкурентные альтернативные способы получения PLA, которые являются менее дорогостоящими и более эффективными, чем химические процессы.
Определения
Настоящее раскрытие лучше всего понимать, обратившись к следующим определениям.
«Полимер» относится к химическому соединению или смеси соединений, структура которых состоит из множества звеньев, связанных ковалентными химическими связями. В рамках контекста изобретения, термин «полимер» включает полимеры, содержащие повторяющееся звено одного типа (т. е. гомополимеры) или звенья различных типов (т. е. блок-сополимеры и статистические сополимеры). Более конкретно, в контексте изобретения термин «гомополимер PLA» относится к полимеру, состоящему только из звеньев L- или D-молочной кислоты и не содержащему какие-либо другие звенья (такие как гидроксиалканоат или тому подобное). Соответственно, «гомополимеры PLA» обозначают поли-L-молочную кислоту (PLLA) или поли-D-молочную кислоту (PDLA). «Гетерополимеры PLA» обозначают поли-DL-молочную кислоту (PDLLA).
В контексте изобретения термины «лактат» и «молочная кислота» используют взаимозаменяемо, чтобы отослать к органическому соединению с формулой CH3CH(OH)CO2H или его ионизированной форме. В отсутствие дополнительных указаний, эти термины обозначают без различий L-молочную кислоту, D-молочную кислоту и их смесь. Аналогичным образом, термины «полилактат» и «полимолочная кислота» используют взаимозаменяемо, чтобы отослать к сложному полиэфиру, состоящему по существу из звеньев молочной кислоты. В отсутствие дополнительных указаний эти термины обозначают без различий PLLA, PDLA и PDLLA.
Термины «нуклеиновая кислота», «нуклеиновая последовательность», «полинуклеотид», «олигонуклеотид» и «нуклеотидная последовательность» обозначают молекулу или последовательность дезоксирибонуклеотидов и/или рибонуклеотидов.
«Ген» обозначает любую молекулу нуклеиновой кислоты, содержащую открытую рамку считывания, кодирующую белок. Ген может содержать дополнительные последовательности, например, такие как промотор, терминатор транскрипции, сигнальный пептид или интрон.
Термин «рекомбинантный» относится к конструкции нуклеиновой кислоты, вектору, полипептиду или клетке, полученным с помощью генетической инженерии.
В настоящем описании «рекомбинантная клетка дрожжей» относится к клетке дрожжей, которая генетически модифицирована, наиболее часто с помощью инсерции последовательности нуклеиновой кислоты или звена, которое не существует в природе в существующей клетке дрожжей, и/или посредством инактивации нативной последовательности нуклеиновой кислоты или звена. Указанные последовательность нуклеиновой кислоты или звено могут быть вставлены или инактивированы в указанной клетке дрожжей или ее предке, используя технологию рекомбинантных ДНК или случайный мутагенез.
В контексте изобретения, термины «вариант» или «мутант» используют для обозначения функционального белка, содержащего по меньшей мере одну аминокислотную модификацию или изменение, т. е. замену, инсерцию и/или делецию, в одном или больше (например, нескольких) положениях по сравнению с исходным белком (т. е. в него вводят мутацию по меньшей мере в одном аминокислотном положении), который представляет желаемые свойства. Варианты можно получать различными способами, хорошо известными в данной области. В частности, примеры способов изменения ДНК последовательности, кодирующей белок дикого типа, включают, но не ограничиваясь этим, сайт-специфический мутагенез, случайный мутагенез и конструирование синтетических олигонуклеотидов. Термин «экспрессия», как используют в настоящем описании, относится к любой стадии, вовлеченной в образование полипептида, включая, но без ограничения этим, транскрипцию, посттранскрипционную модификацию, трансляцию, посттрансляционную модификацию и секрецию.
В контексте изобретения «геном клетки дрожжей» обозначает весь генетический материал, содержащийся в указанной клетке дрожжей, в том числе внехромосомный генетический материал, содержащийся, например, в плазмидах, эписомах, синтетических хромосомах и т. д.
Термины «гетерологичная последовательность» или «гетерологичный белок» обозначают последовательность или белок, которые не существуют в природном состоянии у рассматриваемого микроорганизма. Наоборот, термины «гомологичная последовательность» и «эндогенная последовательность» или «гомологичный белок» и «эндогенный белок» обозначают последовательность или белок, которые существуют в природном состоянии у рассматриваемого микроорганизма.
Аналогичным образом, термин «эндогенная активность» обозначает активность, которая присутствует у рассматриваемого микроорганизма в природном состоянии. Наоборот, термины «экзогенная активность» обозначает активность, которая не присутствует у рассматриваемого микроорганизма в природном состоянии.
Здесь термины «пептид», «полипептид» и «белок» используют взаимозаменяемо, и они относятся к цепочке аминокислот, связанных пептидными связями, независимо от числа аминокислот, образующих указанную цепочку. Аминокислоты в настоящем описании представлены с помощью их однобуквенного или трехбуквенного кода в соответствии со следующей номенклатурой: A: аланин (Ala); C: цистеин (Cys); D: аспарагиновая кислота (Asp); E: глутаминовая кислота (Glu); F: фенилаланин (Phe); G: глицин (Gly); H: гистидин (His); I: изолейцин (Ile); K: лизин (Lys); L: лейцин (Leu); M: метионин (Met); N: аспарагин (Asn); P: пролин (Pro); Q: глутамин (Gln); R: аргинин (Arg); S: серин (Ser); T: треонин (Thr); V: валин (Val); W: триптофан (Trp) и Y: тирозин (Tyr).
Как используют здесь, термины «инактивированный», «инактивация», «измененный» или «дефектный» в отношении данной активности или белка указывают на снижение или ослабление уровня указанной активности или белка в клетке дрожжей. Такое снижение обычно составляет приблизительно 20%, более предпочтительно 30%, по сравнению с белком дикого типа в клетке дрожжей. Снижение может быть более существенным (например, выше 50%, 60%, 70%, 80% или больше) или полным (т. е. нокаут). В соответствии с изобретением, такую инактивацию можно получать различными способами, выполняемыми на уровне ДНК, мРНК или белка, чтобы ингибировать экспрессию соответствующего гена (например, транскрипцию или трансляцию), или активности белка. На уровне ДНК или мРНК, инактивацию можно выполнять, например, посредством делеции, инсерции и/или замены одного или нескольких нуклеотидов, сайт-специфического мутагенеза, мутагенеза с этилметансульфонатом (EMS), направленных индуцированных локальных повреждений генома (TILLING), способов нокаута или сайленсинга генов, например, с использованием РНК-интерференции, антисмысловых, аптамеров и т. п. Другой конкретный подход представляет собой инактивацию гена посредством инсерции чужеродной последовательности, например, через транспозонный мутагенез с использованием мобильных генетических элементов, называемых транспозонами, которые могут иметь природное или искусственное происхождение. Белок также можно делать дефектным посредством изменения его активности, или посредством изменения структуры белка или, например, посредством экспрессии в клетке лиганда белка или его ингибитора. Предпочтительные способы инактивации влияют на экспрессию и ведут к отсутствию продуцирования функционального белка в клетке дрожжей. В частности, дефектный ген получают посредством делеции, мутации, инсерции и/или замены одного или нескольких нуклеотидов.
Биологический синтез полимолочной кислоты
Биологическое получение PLA может происходить в две стадии. Первая состоит в активации молочной кислоты до лактил-КоА с использованием донора КоА. В конкретном варианте осуществления, проиллюстрированном на фиг. 1, ацетил-КоА используют в качестве донора КоА. Для этой стадии необходима лактил-КоА-синтаза, такая как КоА-трансфераза, такая как пропионил-КоА-трансфераза (Pctp). В другом варианте осуществления можно использовать лигазу, такую как ацил-КоА-лигаза, которая связывает КоА с молочной кислотой для того, чтобы формировать лактил-КоА.
Вторая стадия представляет собой полимеризацию лактил-КоА до PLA с помощью лактил-КоА-полимеразы, такой как PHA-синтаза.
Для того чтобы сделать возможным биологическое получение PLA в клетках дрожжей, авторы изобретения сконструировали клетки дрожжей, которые демонстрируют как активность лактил-КоА-синтазы, так и активность лактил-КоА-полимеразы.
В соответствии с изобретением, дрожжи могут естественным путем проявлять одну из этих активностей и могут быть генетически модифицированы, чтобы иметь вторую активность. Например, клетки дрожжей исходно проявляют активность лактил-КоА-синтазы, и белок, обладающий активностью лактил-КоА-полимеразы, генетически вводили в клетку дрожжей или ее предков, или наоборот. Альтернативно, дрожжи исходно не обладают обеими активностями, и генетически модифицированы ради этих двух активностей. Альтернативно, или кроме того, дрожжи исходно проявляют по меньшей мере одну из этих активностей и модифицированы с тем, чтобы содействовать и/или усиливать экспрессию вовлеченного гена(ов).
В соответствии с изобретением, геном клетки дрожжей или ее предка модифицирован посредством введения по меньшей мере одной нуклеиновой последовательности, кодирующей по меньшей мере один фермент, участвующий в пути продуцирования PLA, или его биологически активный фрагмент.
В конкретном варианте осуществления изобретение относится к рекомбинантным Yarrowia, обычно рекомбинантным Y. lipolytica, которые проявляют эндогенную активность лактил-КоА-синтазы и которые дополнительно сконструированы для экспрессии гетерологичного белка, обладающего активностью лактил-КоА-полимеразы.
В другом варианте осуществления изобретение относится к рекомбинантным Yarrowia, обычно рекомбинантным Y. Lipolytica, которые сконструированы для экспрессии как гетерологичного белка, обладающего активностью лактил-КоА-полимеразы, так и гетерологичного белка, обладающего активностью лактил-КоА-синтазы.
В конкретном варианте осуществления белок, обладающий активностью лактил-КоА-синтазы, представляет собой КоА-трансферазу. В частности, белок, обладающий активностью КоА-трансферазы, представляет собой пропионил-КоА-трансферазу (EC:2.8.3.1), которая, как известно, способна активировать молочную кислоту до лактил-КоА.
Например, нуклеиновую кислоту, кодирующую пропионил-КоА-трансферазу (Pctp), обладающую аминокислотной последовательностью, как изложено в SEQ ID № 1, SEQ ID № 2, SEQ ID № 3, SEQ ID № 4 или SEQ ID № 16, или ее функциональные варианты, обладающие активностью лактил-КоА-синтазы, вводили в рекомбинантную клетку дрожжей или ее предков.
SEQ ID № 1/Pctp Clostridium propionicum (номер доступа Q9L3F7/CAB77207.1)
MRKVPIITADEAAKLIKDGDTVTTSGFVGNAIPEALDRAVEKRFLETGEPKNITYVYCGSQGNRDGRGAEHFAHEGLLKRYIAGHWATVPALGKMAMENKMEAYNVSQGALCHLFRDIASHKPGVFTKVGIGTFIDPRNGGGKVNDITKEDIVELVEIKGQEYLFYPAFPIHVALIRGTYADESGNITFEKEVAPLEGTSVCQAVKNSGGIVVVQVERVVKAGTLDPRHVKVPGIYVDYVVVADPEDHQQSLDCEYDPALSGEHRRPEVVGEPLPLSAKKVIGRRGAIELEKDVAVNLGVGAPEYVASVADEEGIVDFMTLTAESGAIGGVPAGGVRFGASYNADALIDQGYQFDYYDGGGLDLCYLGLAECDEKGNINVSRFGPRIAGCGGFINITQNTPKVFFCGTFTAGGLKVKIEDGKVIIVQEGKQKKFLKAVEQITFNGDVALANKQQVTYITERCVFLLKEDGLHLSEIAPGIDLQTQILDVMDFAPIIDRDANGQIKLMDAALFAEGLMGLKEMKS
В конкретном варианте осуществления используют нуклеиновую кислоту, кодирующую вариант Pctp с SEQ ID № 1, содержащий предпочтительно по меньшей мере одну из мутаций, выбранных из V193A и A243T, и обладающий активностью лактил-КоА-синтазы.
SEQ ID № 2/Pctp Aspergillus nidulans (номер доступа Q5B0U7/AN5833.2)
MTHPQQAVHAASLQNPEAFWSHHAQQLHWHKKPSRAIGRSTKTLASGASHESWSWFPDGEISTTYNCVDRHVLNGNGDNVAIIWDSAVTGKKEKYTYRQLLDEVEVLAGVLREEGVKKGDVVIIYMPMIPAALIGALAVARLGAIHAAVFGGFAAKSLAQRIEAARPRAILTASCGIEGAKGPIAYRPLVEGAIEASSFKPEKVLIWQRDQLRWNNPDKLGGQRNWNRLVKSARMRGIRAEPVPVRSTDGLYIIYTSGTTGLPKGVVREAGGHAVGLSLSIKYLFDIHGPGDTMFCASDIGWVVGHSYILYAPLLVGATTVLFEGKPVGTPDAGTFWRVVAEHKANVLFTAPTALRAIRKEDPDNKHFEKVAGDNNLRHLRALFLAGERSEPSIVRAYQDLLTKHAARGALVVDNWWSSESGSPISGLALRSAVGRVPPRSDEYDVAPLAIRPGSAGLPMPGFDVRVVDDEGNEVAQGTMGNIVMATPLAPTAFTRLFNDDERFYKGYLKRFGGRWLDTGDAGMIDQDGYIHVMSRSDDIINVAAHRFSTGQGSIEQAILSHPAIGEASVVGIPDALKGHLPFAFITLKQSGGNSPARPSAELFNSVNRLVREQIGAIASLGGMIQGQGMIPKTRSGKTLRRVLRELVENGARGEFEKEVAVPPTVEDRGVVEVAREKVREYFESQSGSPKAKL
SEQ ID № 3/Pctp Escherichia coli (номер доступа A0A0F6C4Y4/AFJ29290.1)
MKPVKPPRINGRVPVLSAQEAVNYIPDEATLCVLGAGGGILEATTLITALADKYKQTQTPRNLSIISPTGLGDRADRGISPLAQEGLVKWALCGHWGQSPRISDLAEQNKIIAYNYPQGVLTQTLRAAAAHQPGIISDIGIGTFVDPRQQGGKLNEVTKEDLIKLVEFDNKEYLYYKAIAPDIAFIRATTCDSEGYATFEDEVMYLDALVIAQAVHNNGGIVMMQVQKMVKKATLHPKSVRIPGYLVDIVVVDPDQSQLYGGAPVNRFISGDFTLDDSTKLSLPLNQRKLVARRALFEMRKGAVGNVGVGIADGIGLVAREEGCADDFILTVETGPIGGITSQGIAFGANVNTRAILDMTSQFDFYHGGGLDVCYLSFAEVDQHGNVGVHKFNGKIMGTGGFIDISATSKKIIFCGTLTAGSLKTEIADGKLNIVQEGRVKKFIRELPEITFSGKIALERGLDVRYITERAVFTLKEDGLHLIEIAPGVDLQKDILDKMDFTPVISPELKLMDERLFIDAAMGFVLPEAAH
SEQ ID № 4/Pctp Ralstonia eutropha (номер доступа Q0K874/CAJ93797.1)
MKVITAREAAALVQDGWTVASAGFVGAGHAEAVTEALEQRFLQSGLPRDLTLVYSAGQGDRGARGVNHFGNAGMTASIVGGHWRSATRLATLAMAEQCEGYNLPQGVLTHLYRAIAGGKPGVMTKIGLHTFVDPRTAQDARYHGGAVNERARQAIAEGKACWVDAVDFRGDEYLFYPSFPIHCALIRCTAADARGNLSTHREAFHHELLAMAQAAHNSGGIVIAQVESLVDHHEILQAIHVPGILVDYVVVCDNPANHQMTFAESYNPAYVTPWQGEAAVAEAEAAPVAAGPLDARTIVQRRAVMELARRAPRVVNLGVGMPAAVGMLAHQAGLDGFTLTVEAGPIGGTPADGLSFGASAYPEAVVDQPAQFDFYEGGGIDLAILGLAELDGHGNVNVSKFGEGEGASIAGVGGFINITQSARAVVFMGTLTAGGLEVRAGDGGLQIVREGRVKKIVPEVSHLSFNGPYVASLGIPVLYITERAVFEMRAGADGEARLTLVEIAPGVDLQRDVLDQCSTPIAVAQDLREMDARLFQAGPLHL
SEQ ID № 16/Pctp Yarrowia lipolytica (номер доступа XP_505057.1)
SEDHPAIHPPSEFKDNHPHFGGPHLDCLQDYHQLHKESIEDPKAFWKKMANELISWSTPFETVRSGGFEHGDVAWFPEGQLNASYNCVDRHAFANPDKPAIIFEADEPGQGRIVTYGELLRQVSQVAATLRSFGVQKGDTVAVYLPMIPEAIVTLLAITRIGAVHSVIFAGFSSGSLRDRINDAKSKVVVTTDASMRGGKTIDTKKIVDEALRDCPSVTHTLVFRRAGVENLAWTEGRDFWWHEEVVKHRPYLAPVPVASEDPIFLLYTSGSTGTPKGLAHATGGYLLGAALTAKYVFDIHGDDKLFTAGDVGWITGHTYVLYGPLMLGATTVVFEGTPAYPSFSRYWDIVDDHKITHFYVAPTALRLLKRAGTHHIKHDLSSLRTLGSVGEPIAPDVWQWYNDNIGRGKAHICDTYWQTETGSHIIAPMAGVTPTKPGSASLPVFGIDPVIIDPVSGEELKGNNVEGVLALRSPWPSMARTVWNTHERYMETYLRPYPGYYFTGDGAARDNDGFYWIRGRVDDVVNVSGHRLSTAEIEAALIEHAQVSESAVVGVHDDLTGQAVNAFVALKNPVEDVDALRKELVVQVRKTIGPFAAPKNVIIVDDLPKTRSGKIMRRILRKVLAGEEDQLGDISTLANPDVVQTIIEVVHSLKK
В другом варианте осуществления белок, обладающий активностью лактил-КоА-синтазы, представляет собой КоА-лигазу (EC:6.2.1), предпочтительно ацил-КоА-лигазу.
В конкретном варианте осуществления белок, обладающий активностью лактил-КоА-синтазы, экспрессируют в цитозоле. В другом варианте осуществления белок, обладающий активностью лактил-КоА-синтазы, экспрессируют в субклеточной органелле, такой как пероксисома или митохондрии. С этой целью, нуклеиновую кислоту, кодирующую соответствующую направляющую последовательность, можно добавлять на 5'- или 3'-конец последовательности, представляющей интерес (см. далее).
В соответствии с изобретением, рекомбинантная клетка дрожжей может экспрессировать гетерологичный белок, обладающий активностью лактил-КоА-полимеразы.
В конкретном варианте осуществления белок, обладающий активностью лактил-КоА-полимеразы, является стереоспецифическим. Благоприятно, белок имеет активность D-лактил-КоА-полимеразы, которая ведет исключительно к продуцированию PDLA. В другом варианте осуществления белок имеет L-активность лактил-КоА-полимеразы, которая ведет исключительно к продуцированию PLLA.
В конкретном варианте осуществления лактил-КоА-полимераза обладает аффинностью к ацил-КоА с малой длиной цепи, который содержит от 3 до 7 углеродов, предпочтительно от 3 до 5 углеродов, более предпочтительно 3 углерода, более предпочтительно молочную кислоту.
В конкретном варианте осуществления белок, обладающий активностью лактил-КоА-полимеразы, экспрессируют в цитозоле. В другом варианте осуществления белок, обладающий активностью лактил-КоА-полимеразы, экспрессируют в субклеточной органелле, такой как пероксисома или митохондрии. С этой целью, нуклеиновую кислоту, кодирующую соответствующую направляющую последовательность, можно добавлять на 5'- или 3'-конец последовательности, представляющей интерес (см. далее).
Предпочтительно, рекомбинантная клетка дрожжей экспрессирует гетерологичную PHA-синтазу (PhaCp) в качестве лактил-КоА-полимеразы.
В конкретном варианте осуществления PHA-синтаза представляет собой PHA-синтазу II класса из Pseudomonas aeruginosa PAO1 (PaPhaCp).
Например, PHA-синтаза имеет аминокислотную последовательность, как изложено в SEQ ID № 5, или ее вариант, имеющий активность лактил-КоА-полимеразы.
SEQ ID № 5/PhaC1p из P. aeruginosa PAO1 (номер доступа G3XCV5/NP_253743.1)
MSQKNNNELPKQAAENTLNLNPVIGIRGKDLLTSARMVLLQAVRQPLHSARHVAHFSLELKNVLLGQSELRPGDDDRRFSDPAWSQNPLYKRYMQTYLAWRKELHSWISHSDLSPQDISRGQFVINLLTEAMSPTNSLSNPAAVKRFFETGGKSLLDGLGHLAKDLVNNGGMPSQVDMDAFEVGKNLATTEGAVVFRNDVLELIQYRPITESVHERPLLVVPPQINKFYVFDLSPDKSLARFCLRNGVQTFIVSWRNPTKSQREWGLTTYIEALKEAIEVVLSITGSKDLNLLGACSGGITTATLVGHYVASGEKKVNAFTQLVSVLDFELNTQVALFADEKTLEAAKRRSYQSGVLEGKDMAKVFAWMRPNDLIWNYWVNNYLLGNQPPAFDILYWNNDTTRLPAALHGEFVELFKSNPLNRPGALEVSGTPIDLKQVTCDFYCVAGLNDHITPWESCYKSARLLGGKCEFILSNSGHIQSILNPPGNPKARFMTNPELPAEPKAWLEQAGKHADSWWLHWQQWLAERSGKTRKAPASLGNKTYPAGEAAPGTYVHER
Альтернативно, используют нуклеиновую кислоту, кодирующую вариант PHA-синтазы из P. aeruginosa PAO1 (SEQ ID № 5), содержащий предпочтительно по меньшей мере одну из мутаций, выбранных из E130D; S325T; S477R; S477F; Q481M; Q481K; S482G; L484V и A547V, и обладающий активностью лактил-КоА-полимеразы.
В конкретном варианте осуществления рекомбинантная клетка дрожжей экспрессирует PHA-синтазу с тремя следующими мутациями в SEQ ID № 5: E130D; S477F; Q481K.
В другом варианте осуществления рекомбинантная клетка дрожжей экспрессирует PHA-синтазу с четырьмя следующими мутациями в SEQ ID № 5: E130D; S325T; S477R; Q481M (SEQ ID № 11 или генная последовательность SEQ ID № 15).
Альтернативно или кроме того, можно использовать нуклеиновую кислоту, кодирующую функциональный мутант PHA-синтазы I класса (EC:2.3.1.B3), такой как мутант PHA-синтазы из R. eutropha, или нуклеиновую кислоту, кодирующую функциональный мутант PHA-синтазы III класса (EC:2.3.1.B4), такой как мутант PHA-синтазы из Chromatium vinosum (например, SEQ ID № 6 или SEQ ID № 7).
SEQ ID № 6/PhaC1p из R. eutropha H16 (номер доступа P23608/CAJ92572.1)
MATGKGAAASTQEGKSQPFKVTPGPFDPATWLEWSRQWQGTEGNGHAAASGIPGLDALAGVKIAPAQLGDIQQRYMKDFSALWQAMAEGKAEATGPLHDRRFAGDAWRTNLPYRFAAAFYLLNARALTELADAVEADAKTRQRIRFAISQWVDAMSPANFLATNPEAQRLLIESGGESLRAGVRNMMEDLTRGKISQTDESAFEVGRNVAVTEGAVVFENEYFQLLQYKPLTDKVHARPLLMVPPCINKYYILDLQPESSLVRHVVEQGHTVFLVSWRNPDASMAGSTWDDYIEHAAIRAIEVARDISGQDKINVLGFCVGGTIVSTALAVLAARGEHPAASVTLLTTLLDFADTGILDVFVDEGHVQLREATLGGGAGAPCALLRGLELANTFSFLRPNDLVWNYVVDNYLKGNTPVPFDLLFWNGDATNLPGPWYCWYLRHTYLQNELKVPGKLTVCGVPVDLASIDVPTYIYGSREDHIVPWTAAYASTALLANKLRFVLGASGHIAGVINPPAKNKRSHWTNDALPESPQQWLAGAIEHHGSWWPDWTAWLAGQAGAKRAAPANYGNARYRAIEPAPGRYVKAKA
SEQ ID № 7/PhaCp of C. vinosum (номер доступа Q402A9)
MFPIDIRPDKLTQEMLDYSRKLGQGMENLLNAEAIDTGVSPKQAVYSEDKLVLYRYDRPEGAPEAQPVPLLIVYALVNRPYMTDIQEDRSTIKGLLATGQDVYLIDWGYPDQADRALTLDDYINGYIDRCVDYLREAHGVDKVNLLGICQGGAFSLMYSALHPDKVRNLVTMVTPVDFKTPDNLLSAWVQNVDIDLAVDTMGNIPGELLNWTFLSLKPFSLTGQKYVNMVDLLDDPDKVKNFLRMEKWIFDSPDQAGETFRQFIKDFYQNNGFLNGGVVLGGQEVDLKDITCPVLNIFALQDHLVPPDASRALKGLTSSPDYTELAFPGGHIGIYVSGKAQKEVTPAIGKWLNER
В конкретном варианте осуществления несколько копий нуклеиновой последовательности, кодирующей по меньшей мере один фермент, вовлеченный в путь получения PLA, вводят в рекомбинантные дрожжи, в частности, по меньшей мере две копии.
В конкретном варианте осуществления в рекомбинантные дрожжи вводят по меньшей мере две копии нуклеиновой последовательности, кодирующей активность лактил-КоА-синтазы, предпочтительно по меньшей мере две копии нуклеиновой последовательности, кодирующей пропионил-КоА-трансферазу.
Альтернативно или кроме того, в рекомбинантные дрожжи вводят по меньшей мере две копии нуклеиновой последовательности, кодирующей активность лактил-КоА-полимеразы, предпочтительно по меньшей мере две копии нуклеиновой последовательности, кодирующей PHA-синтазу.
В предпочтительном варианте осуществления в рекомбинантные дрожжи вводят по меньшей мере две копии нуклеиновой последовательности, кодирующей пропионил-КоА-трансферазу, и по меньшей мере две копии нуклеиновой последовательности, кодирующей PHA-синтазу.
Благоприятно, нуклеиновая последовательность(и), введенная в рекомбинантные дрожжи, находится (находятся) под управлением индуцибельного или конститутивного промотора. В частности, промоторы различной силы используют для того, чтобы управлять экспрессией по меньшей мере одного фермента. В качестве примера, промоторы TEF и 4UAS-TEF используют для того, чтобы управлять экспрессией нуклеиновых последовательностей. Предпочтительно, нуклеиновая последовательность, кодирующая пропионил-КоА-трансферазу, и нуклеиновая последовательность, кодирующая PHA-синтазу, находятся под управлением промотора 4UAS-TEF.
Следовательно, цель изобретения состоит в том, чтобы предоставить рекомбинантную клетку дрожжей, предпочтительно рекомбинантную Yarrowia, обычно рекомбинантную Y. Lipolytica, которую сконструировали для экспрессии как по меньшей мере одной Pctp, имеющей аминокислотную последовательность, как изложено в SEQ ID № 1, SEQ ID № 2, SEQ ID № 3, SEQ ID № 4, SEQ ID № 16, или ее варианта, так и по меньшей мере одной PhaCp, имеющей аминокислотную последовательность, как изложено в SEQ ID № 5, SEQ ID № 6, SEQ ID № 7 или SEQ ID № 11, или ее варианта.
Направление белков в субклеточные органеллы
В конкретном примере продуцирование PLA может быть ограничено конкретными субклеточными органеллами рекомбинантной клетки дрожжей, такими как митохондрии или пероксисома, вместо цитозоля. Такое ограниченное продуцирование можно использовать, чтобы содействовать продуцированию PLA с высокой молекулярной массой.
Тогда активности, необходимые для продуцирования PLA, и ассоциированные белки следует направлять в субклеточную органеллу, представляющую интерес. С этой целью, конкретные последовательности, позволяющие направлять белок, можно добавлять на 5'- или 3'-конец нуклеиновой кислоты, кодирующей белки, представляющие интерес.
В конкретном варианте осуществления продуцирование PLA может быть ограничено пероксисомой, посредством использования пероксисомальной направленной последовательности (PTS) с белками, которые не экспрессируются или не направляются естественным путем в пероксисомы.
Например, можно использовать нуклеиновую кислоту, кодирующую PTS, как изложено в SEQ ID № 8 (PTS1), которая соответствует 14 последовательным аминокислотам из изоцитратдегидрогеназы (YALI0C16885p/P41555 - Icl1p изоцитратлиаза из Y. lipolytica). В другом варианте осуществления PTS может соответствовать трипептиду, такому как SKL или AKL или любая последовательность, известная специалисту в данной области. Предпочтительно, PTS располагают на C-конце белка, представляющего интерес. В другом варианте осуществления последовательность PTS находится на N-конце тиолазы Y. lipolytica (YALI0E18568p/QO5493 - Pot1p 3-кетоацил-КоА-тиолаза), которая отщепляется при попадании в пероксисому (SEQ ID № 9 - PTS2). PTS2 предпочтительно располагают на N-конце белка, представляющего интерес. Говоря в общем, можно использовать любую PTS, известную специалисту в данной области.
SEQ ID № 8: MGAGVTEDQFKSKL
SEQ ID № 9: MDRLNNLATQLEQNPA
Следовательно, цель изобретения состоит в том, чтобы предоставить рекомбинантную клетку дрожжей, предпочтительно рекомбинантные Yarrowia, обычно рекомбинантные Y. Lipolytica, которую сконструировали для экспрессии по меньшей мере одной Pctp, имеющей аминокислотную последовательность, как изложено в SEQ ID № 1, SEQ ID № 2, SEQ ID № 3, SEQ ID № 4, SEQ ID № 16, или ее варианта, в цитозоле и по меньшей мере одной PhaCp, имеющей аминокислотную последовательность, как изложено в SEQ ID № 5, SEQ ID № 6, SEQ ID № 7 или SEQ ID № 11, или ее варианта в пероксисоме. В конкретном варианте осуществления аминокислотная последовательность PhaCp дополнительно содержит на ее C-конце аминокислотную последовательность, изложенную в SEQ ID № 8, или на ее N-конце аминокислотную последовательность, изложенную в SEQ ID № 9.
Альтернативно, продуцирование PLA может быть ограничено митохондриями, и более конкретно митохондриальным матриксом или митохондриальным внутримембранным пространством, посредством прикрепления митохондриальной направленной последовательности (MTS) к белкам, которые не экспрессируются или не направляются естественным путем в митохондрии. В конкретном варианте осуществления можно использовать нуклеиновую кислоту, кодирующую MTS, как изложено в SEQ ID № 10, которая соответствует 26 последовательным аминокислотам яблочного фермента (YALI0E18634p/Q6C5F0 - Mae1p). Эту MTS предпочтительно добавляют на N-конец белка, представляющего интерес. Говоря в общем, можно использовать любую MTS, известную специалисту в данной области.
SEQ ID № 10: MLRLRTMRPTQTSVRAALGPTAAARN
Метаболизм молочной кислоты
В соответствии с изобретением, способность клетки дрожжей потреблять молочную кислоту в качестве источника углерода можно ослаблять или ингибировать. Соответственно, рекомбинантная клетка дрожжей будет способна лишь полимеризовать LA для получения PLA. Это может представлять особый интерес при использовании Y. lipolytica или Saccharomyces cerevisiae, которые обычно потребляют молочную кислоту в качестве источника углерода.
С этой целью, клетку дрожжей можно модифицировать для того, чтобы инактивировать ее путь расщепления молочной кислоты. Например, инактивируют по меньшей мере одну эндогенную оксидоредуктазу молочной кислоты, отвечающую за превращение молочной кислоты в пируват.
В контексте изобретения, выражение «оксидоредуктаза молочной кислоты» используют для обозначения любого фермента, способного продуцировать пируват из лактата, и наоборот (EC:1.1.1.27, EC:1.1.1.28, EC:1.1.2.3, EC:1.1.2.4). Более конкретно, это выражение охватывает лактатдегидрогеназу, феррицитохром, флавоцитохром и цитохромовую оксидоредуктазу.
Предпочтительно инактивируют дегидрогеназу D-молочной кислоты, которая превращает D-молочную кислоту в пируват. В конкретном варианте осуществления можно сохранять оксидоредуктазу цитохром b2 или c эндогенной L-молочной кислоты с тем, чтобы дрожжи сохраняли способность расщеплять L-молочную кислоту. D-молочная кислота (D-LA) остается доступной для синтеза PLA. Такая рекомбинантная клетка дрожжей благоприятно может быть способна продуцировать PDLA.
Альтернативно, инактивируют оксидоредуктазу эндогенной L-молочной кислоты, тогда как оксидоредуктазу эндогенной D-молочной кислоты сохраняют с тем, чтобы дрожжи оставались способными расщеплять D-молочную кислоту. L-молочная кислота (L-LA) остается доступной для синтеза PLA. Такая рекомбинантная клетка дрожжей благоприятно может быть способна продуцировать PLLA.
Альтернативно, можно инактивировать обе оксидоредуктазы эндогенной D-молочной кислоты и L-молочной кислоты с тем, чтобы и D-молочная кислота (D-LA) и L-молочная кислота (L-LA) оставались доступными для синтеза PLA. Такая рекомбинантная клетка дрожжей благоприятно может быть способна продуцировать PDLLA.
В конкретном варианте осуществления оксидоредуктаза D-молочной кислоты представляет собой дегидрогеназу D-молочной кислоты (DLD), предпочтительно из семейств белков GL3C0735 или GL3C0514, например, Dldl1p, Dld2p и Dld3p. В другом конкретном варианте осуществления оксидоредуктаза L-молочной кислоты представляет собой цитохромовую оксидоредуктазу (CYB), предпочтительно из семейства белков GL3C0472, например, Cyb2p, Cyb21p и Cyb22p.
Далее в таблице 1 и таблице 2 перечислены четыре предполагаемые оксидоредуктазы молочной кислоты из Y. lipolytica и S. cerevisiae, соответственно, и их предсказанная локализация.
Таблица 1. Предполагаемые оксидоредуктазы молочной кислоты из Y. lipolytica
Таблица 2. Предполагаемые оксидоредуктазы молочной кислоты из S. cerevisiae.
В конкретном варианте осуществления изобретение относится к рекомбинантным Y. Lipolytica, в которых инактивирован YlDld1p.
В другом конкретном варианте осуществления изобретение относится к рекомбинантным Y. lipolytica, в которых инактивирован YlCyb21p.
Следовательно, цель изобретения состоит в том, чтобы предоставить рекомбинантные Y. Lipolytica, которые сконструированы для экспрессии по меньшей мере одной Pctp, имеющей аминокислотную последовательность, как изложено в SEQ ID № 1, SEQ ID № 2, SEQ ID № 3, SEQ ID № 4, SEQ ID № 16, или ее варианта, в цитозоле, по меньшей мере одной PhaCp, имеющей аминокислотную последовательность, как изложено в SEQ ID № 5, SEQ ID № 6, SEQ ID № 7 или SEQ ID № 11, или ее варианта, и дополнительно содержащей на ее C-конце аминокислотную последовательность, изложенную в SEQ ID № 8, или на ее N-конце аминокислотную последовательность, изложенную в SEQ ID № 9, чтобы направлять в пероксисому, и которые дополнительно сконструированы для того, чтобы инактивировать лактатдегидрогеназу (YlDld1p и/или YlCyb21p).
Альтернативно, авторы изобретения обнаружили, что активность D-лактатоксидоредуктазы не наблюдают при культивировании рекомбинантной клетки дрожжей в среде, не содержащей конкретные аминокислоты. Например, Y. lipolytica можно культивировать в средах для культивирования, не содержащих Ala, Arg, Asn, Gln, Ile, Leu, Lys, Phe, Pro, Thr и Val, чтобы избегать экспрессии YlDld1p.
Альтернативно, возможно ингибировать или снижать потребление молочной кислоты посредством предоставления других подходящих источников углерода для клетки дрожжей.
Для того чтобы улучшать транспорт молочной кислоты в клетки дрожжей или чтобы снижать ее экспорт из клеток дрожжей, клетки можно частично инактивировать для экспорта или усовершенствовать для импорта, предпочтительно транспорта/экспорта D-лактата.
Например, захват молочной кислоты клетками можно увеличивать посредством чрезмерной экспрессии или чрезмерной активации импортеров лактата, таких как специфические мембранные переносчики. Альтернативно или кроме того, экспорт молочной кислоты из клеток можно по меньшей мере уменьшать посредством инактивации или делеции экспортеров молочной кислоты.
В одном из вариантов осуществления рекомбинантная клетка дрожжей демонстрирует экзогенную активность рацемазы молочной кислоты. Например, рацемаза превращает L-лактат в D-лактат или наоборот. В конкретном варианте осуществления рекомбинантная клетка дрожжей содержит рацемазу из Lactobacillus plantarum (LpLarAp - EC:5.2.2.1 - M4KDH2/AGE37852).
Модифицированный цитозольный метаболизм ацетил-КоА
В соответствии с изобретением, рекомбинантная клетка дрожжей демонстрирует активность лактил-КоА-синтазы для продуцирования лактил-КоА из молочной кислоты. В конкретном варианте осуществления активность лактил-КоА-синтазы осуществляют с использованием Pctp, которая использует ацетил-КоА в качестве донора КоА.
В этом контексте, продуцированию PLA в цитозоле можно содействовать посредством увеличения доступности ацетил-КоА в цитозоле рекомбинантных клеток дрожжей. С этой целью можно рассмотреть три альтернативных или кумулятивных пути: перенаправление части ацетил-КоА (в основном, содержащегося в митохондриях клеток дрожжей) из митохондрий в цитозоль; изменение метаболических путей, в которых используют ацетил-КоА в цитозоле; введение новых метаболических путей для того, чтобы увеличивать цитозольное продуцирование ацетил-КоА (фиг. 2 и фиг. 3).
Некоторые дрожжи, такие как Y. lipolytica, могут использовать цитозольный ацетил-КоА для синтеза триацилглицерина (TAG). В таком случае, продуцирование TAG непосредственно конкурирует с продуцированием PLA. Таким образом, в соответствии с предпочтительным вариантом осуществления, рекомбинантную клетку дрожжей можно модифицировать для того, чтобы инактивировать путь синтеза TAG. Например, в Y. Lipolytica можно инактивировать гены YlDGA1 (YALI0E32769g/XM_504700.1) и/или YlDGA2 (YALI0F06578g/XM_505086.1) и/или YlLRO1 (YALI0E16797g/XM_504038.1), кодирующие эндогенные диацилглицеринтрансферазые (Beopoulos et al., 2008) (фиг. 2, точка 3).
Кроме того, некоторые дрожжи, такие как Y. lipolytica, также имеют ген, кодирующий цитратсинтазу (EC:2.3.3.16), которая выполняет обратную реакцию относительной той, которую катализирует АТФ-цитратлиаза. Такая реакция потребляет цитозольный ацетил-КоА и непосредственно конкурирует с продуцированием PLA. Соответственно, в другом предпочтительном варианте осуществления в рекомбинантной клетке дрожжей инактивируют активность цитратсинтазы (фиг. 2, точка 4).
Кроме того, в некоторых клетках дрожжей, таких как Y. lipolytica, основная часть запаса цитозольного ацетил-КоА поступает из превращения цитрата, получаемого через цикл Кребса, в ацетил-КоА посредством АТФ-цитратлиазы (EC:2.3.3.8). Механизм трансмембранного транспорта сопрягает импорт малата с экспортом цитрата из митохондрий. Соответственно, в конкретном варианте осуществления рекомбинантную клетку дрожжей по изобретению культивируют в конкретных условиях, которые индуцируют физиологическое состояние, благоприятное для экспорта цитрата в цитозоль (см. фиг. 2, точка 1). Например, среду для культивирования по меньшей мере временно ограничивают в конкретном питательном веществе(ах), таком как азот, фосфат и/или кислород.
Альтернативно или кроме того, можно инактивировать или ослаблять активность малатсинтазы (EC:2.3.3.9), которая забирает часть митохондриального ацетил-КоА из продуцирования цитрата в качестве подлежащего использованию для продуцирования малата (путь глиоксилатного шунта) (фиг. 2, точка 2).
В конкретном варианте осуществления ген, кодирующий цитратсинтазу, и/или ген, кодирующий малатсинтазу, инактивируют в рекомбинантной клетке дрожжей по изобретению или ее предке.
В другом варианте осуществления рекомбинантная клетка дрожжей демонстрирует инактивированную активность ацил-КоА:диацилглицеринацилтрансферазы (DGAT1/2, DGA1/2) и/или фосфолипид:диацилглицеринацилтрансферазы (PDAT, LRO1).
Как проиллюстрировано на фиг. 1, реакция, катализируемая с помощью Pctp, продуцирует ацетат в дополнение к лактил-КоА. В конкретном варианте осуществления, чтобы увеличивать цитозольное продуцирование ацетил-КоА, гетерологичный ген, кодирующий ацетил-КоА-синтазу, также называемый Acsp (EC:6.2.1), можно вводить в рекомбинантную клетку дрожжей для того, чтобы превращать ацетат обратно в ацетил-КоА. Альтернативно, эндогенный ген, кодирующий ацетил-КоА-синтазу, можно чрезмерно экспрессировать. Эта реакция может быть АДФ- или АМФ-зависимой, в соответствии с рассматриваемым ферментом (фиг. 3, точка 5). Соответственно, в конкретном варианте осуществления рекомбинантная клетка дрожжей дополнительно содержит ген, кодирующий ацетил-КоА-синтазу.
Аналогичным образом, гетерологичную пируватдегидрогеназу можно вводить в рекомбинантную клетку дрожжей для превращения гликолитического пирувата в ацетил-КоА в цитозоле (фиг. 3, точка 6). Предпочтительно используют фермент с низкой чувствительностью к высокому соотношению NADH/NAD+, такой как EfPdhp (NP_815074.1, NP_815075.1, NP_815076.1, NP_815077.1) из Enterococus faecalis или любой другой фермент, известный специалисту в данной области. Соотношение NADH/NAD+ в цитозоле также можно модулировать посредством введения гена, кодирующего нефосфорилирующую глицеральдегид-3P-дегидрогеназу, которая позволяет создавать NADPH во время гликолиза вместо NADH (фиг. 3, точка 6). Соответственно, в конкретном варианте осуществления рекомбинантная клетка дрожжей дополнительно содержит ген, кодирующий пируватдегидрогеназу, указанную пируватдегидрогеназу экспрессируют по меньшей мере в цитозоле рекомбинантной клетки.
Альтернативно или кроме того, можно вводить альтернативный путь для цитозольного продуцирования ацетил-КоА в клетке дрожжей, посредством гетерологичной экспрессии фосфокетолазы (EC:4.1.2.22). Фосфокетолаза катализирует образование ацетил-P и эритрозо-4P из фруктозо-6P (Meile et al., 2001). В этом метаболическом пути, ацетил-P превращают в ацетил-КоА под действием фосфотрансацетилазы (EC:2.3.1.8) или посредством комбинированного действия ацетаткиназы (EC:2.7.2.1) и ацетил-КоА-синтазы. Эритрозо-4P подвергается углеродной перегруппировке под действием активностей транскетолазы и трансальдолазы, обе от природы экспрессированы в пентозофосфатном пути Y. Lipolytica, и, таким образом, становится способной вступать в гликолиз. Кроме того, введение гена, кодирующего гетерологичную фруктозо-1-6-бифосфатазу (EC:3.1.1.11), делает возможной полную рециркуляцию эритрозо-4P, которая дает возможность превращать 100% углерода, присутствующего в субстрате, в ацетил-КоА без образования CO2 (фиг. 3, точка 7).
Рекомбинантная клетка дрожжей
Цель изобретения состоит в том, чтобы предоставить рекомбинантную клетку дрожжей, проявляющую активность лактил-КоА-синтазы, предпочтительно активность КоА-трансферазы и активность лактил-КоА-полимеразы, и которая способна продуцировать PLA.
Благоприятно, выбранная клетка дрожжей исходно способна продуцировать и накапливать липиды в высокой концентрации. В конкретном варианте осуществления клетку дрожжей выбирают из GRAS дрожжей (в целом рассматриваемых как безопасные). Кроме того, выбранные дрожжи благоприятно способны расти при низком pH.
В соответствии с изобретением, клетка дрожжей предпочтительно относится к роду Yarrowia, Saccharomyces, Kluyveromyces, Candida, Pichia, Schizosaccharomyces, Zygosaccharomyces, Hansenula, Trichosporon, Yamadazyma, Cryptococcus, Lipomyces, Rhodosporidium, Rhodotorula, Geotrichum, Kloeckera, Schwanniomyces, Brettanomyces, Debaryomyces или Issatchenkia.
В конкретном варианте осуществления клетка дрожжей представляет собой Yarrowia, выбранную из группы, состоящей из Yarrowia bubula, Yarrowia deformans, Yarrowia lipolytica, Yarrowia yakushimensis, Yarrowia galli, Yarrowia oslonensis, Yarrowia hollandica, Yarrowia phangngensis, Yarrowia alimentaria и Yarrowia porcina, предпочтительно Yarrowia lipolytica.
Следовательно, цель изобретения состоит в том, чтобы предоставить рекомбинантные Y. Lipolytica, экспрессирующие гетерологичную Pctp и гетерологичную PhaCp.
В конкретном варианте осуществления рекомбинантные Yarrowia lipolytica экспрессируют Pctp, имеющую аминокислотную последовательность, как изложено в SEQ ID № 1, SEQ ID № 2, SEQ ID № 3, SEQ ID № 4, или SEQ ID № 16, или ее функциональные варианты, обладающие активностью лактил-КоА-синтазы
В конкретном варианте осуществления рекомбинантные Y. lipolytica экспрессируют PhaCp, имеющую аминокислотную последовательность, как изложено в SEQ ID № 5, или ее функциональный вариант, обладающие активностью лактил-КоА-полимеразы, которые содержат предпочтительно по меньшей мере одну мутацию, выбранную из E130D, S325T, S477F, S477R, Q481K Q481M, S482G, L484V и A547V, или как изложено в SEQ ID № 6 или SEQ ID № 7.
Благоприятно, активность оксидоредуктазы молочной кислоты рекомбинантных Y. lipolytica инактивирована. В конкретном варианте осуществления инактивирован по меньшей мере YlDld1p, чтобы супрессировать или по меньшей мере ослаблять путь расщепления D-молочной кислоты.
В конкретном варианте осуществления рекомбинантные Y. lipolytica экспрессируют четверной вариант PhaC1p из P. aeruginosa (SEQ ID № 11) и функциональный вариант Pctp из C. propionicum (SEQ ID № 1) с мутацией V193A.
SEQ ID № 11/PhaC1p из P. aeruginosa PA01 с мутациями E130D, S325T, S477R и Q481M
MSQKNNNELPKQAAENTLNLNPVIGIRGKDLLTSARMVLLQAVRQPLHSARHVAHFSLELKNVLLGQSELRPGDDDRRFSDPAWSQNPLYKRYMQTYLAWRKELHSWISHSDLSPQDISRGQFVINLLTDAMSPTNSLSNPAAVKRFFETGGKSLLDGLGHLAKDLVNNGGMPSQVDMDAFEVGKNLATTEGAVVFRNDVLELIQYRPITESVHERPLLVVPPQINKFYVFDLSPDKSLARFCLRNGVQTFIVSWRNPTKSQREWGLTTYIEALKEAIEVVLSITGSKDLNLLGACSGGITTATLVGHYVASGEKKVNAFTQLVTVLDFELNTQVALFADEKTLEAAKRRSYQSGVLEGKDMAKVFAWMRPNDLIWNYWVNNYLLGNQPPAFDILYWNNDTTRLPAALHGEFVELFKSNPLNRPGALEVSGTPIDLKQVTCDFYCVAGLNDHITPWESCYKSARLLGGKCEFILSNRGHIMSILNPPGNPKARFMTNPELPAEPKAWLEQAGKHADSWWLHWQQWLAERSGKTRKAPASLGNKTYPAGEAAPGTYVHER
Как раскрыто выше, если необходимо, целиком или частично гетерологичные последовательности, экспрессируемые в рекомбинантной клетке дрожжей, могут содержать PTS или MTS для того, чтобы направлять в пероксисому или митохондрии клетки. В случае, когда направленный сигнальный пептид уже присутствует в последовательности, его можно удалять и/или заменять на белок для направления в желаемый компартмент. В частности, рекомбинантная клетка дрожжей экспрессирует гетерологичную Pctp, направленную в цитозоль, и гетерологичную PhaCp, направленную в пероксисому.
В конкретном варианте осуществления в рекомбинантной клетке дрожжей удаляют диацилглицеринтрансферазы для того, чтобы избегать продуцирования TAG (триацилглицерин). Соответственно, настоящее изобретение предусматривает рекомбинантные Yarrowia, в которых удаляли по меньшей мере один из генов, выбранных из YlLRO1, YlDGA1 и YlDGA2.
В другом конкретном варианте осуществления в рекомбинантной клетке дрожжей удалены 6 ацил-КоА-оксидаз (YlPOX1: YALI0E32835g/XM_504703.1, YlPOX2: YALI0F10857g/XM_505264.1, YlPOX3: YALI0D24750g/XM_503244.1, YlPOX4: YALI0E27654g/XM_504475.1, YlPOX5: YALI0C23859g/XM_502199.1, YlPOX6: YALI0E06567g/XM_503632.1), что предотвращает β-окисление (генотип: pox1-6Δ) для того, чтобы избегать встраивания 3-гидрокси-ацил-КоА, образуемого при β-окислении. Эту делецию можно использовать для продуцирования гомополимера PLA в пероксисоме.
Следовательно, цель изобретения состоит в том, чтобы предоставить рекомбинантные Y. Lipolytica, которые сконструированы для экспрессии по меньшей мере одной Pctp, имеющей аминокислотную последовательность, как изложено в SEQ ID № 1, SEQ ID № 2, SEQ ID № 3, SEQ ID № 4, SEQ ID № 16, или ее варианта, в цитозоле, по меньшей мере одной PhaCp, имеющей аминокислотную последовательность, как изложено в SEQ ID № 5, SEQ ID № 6, SEQ ID № 7 или SEQ ID № 11, или ее варианта, и дополнительно содержащей на ее C-конце аминокислотной последовательности, изложенной в SEQ ID № 8, или на ее N-конце аминокислотной последовательности, изложенной в SEQ ID № 9, для направления в пероксисому, и которые дополнительно сконструированы для того, чтобы инактивировать как лактатдегидрогеназу (YlDld1p и/или YlCyb21p), так и ацил-КоА-оксидазы (YlPOX1, YlPOX2, YlPOX3, YlPOX4, YlPOX5, YlPOX6).
В конкретном варианте осуществления рекомбинантная клетка дрожжей дополнительно может экспрессировать амфифильные белки, способные формировать микрокомпартменты, которые инкапсулируют и, тем самым, изолируют образованную PLA. Например, рекомбинантную клетку дрожжей дополнительно можно конструировать для того, чтобы экспрессировать фазины, которые собираются в микрокомпартменты, инкапсулирующие образованную PLA.
В конкретном варианте осуществления рекомбинантная клетка дрожжей экспрессирует может экспрессировать белок фазин, как изложено в SEQ № 17 или SEQ № 18. Эти белки могут не содержать пептидный сигнал, PTS или MTS, чтобы направляться в цитозоль, пероксисомы или митохондрии, соответственно. Например, можно использовать PTS, как изложено в SEQ ID № 8 или SEQ ID № 9. Например, можно использовать MTS, как изложено в SEQ ID № 10.
SEQ ID № 17/PhaIp из Pseudomonas putida (Q5QBP4)
MAKVTVKKKDDALGTLGEVRGYARKIWLAGIGAYARVGQEGSDYFQELVKAGEGVEKRGKKRIDKELDAANNQIDEAAEEVSRVRGEVEIQLDKIEKAFDARVGRALNRLGIPSKHDVEALSIKLEQLHELLERVAHKP
SEQ ID № 18/phaP1p из Rastonia eutropha H16 (QOKBV4)
MILTPEQVAAAQKANLETLFGLTTKAFEGVEKLVELNLQVVKTSFAEGVDNAKKALSAKDAQELLAIQAAAVQPVAEKTLAYTRHLYEIASETQSEFTKVAEAQLAEGSKNVQALVENLAKNAPAGSESTVAIVKSAISAANNAYESVQKATKQAVEIAETNFQAAATAATKAAQQASATARTATAKKTTAA
Способ получения полимолочной кислоты (PLA) с использованием рекомбинантных клеток дрожжей
Изобретение дополнительно относится к способу получения PLA посредством культивирования рекомбинантных клеток дрожжей по изобретению.
В соответствии с изобретением, PLA можно получать посредством культивирования таких рекомбинантных клеток дрожжей в присутствии молочной кислоты.
В конкретном варианте осуществления рекомбинантные клетки дрожжей сначала культивируют в ферментативном бульоне, не содержащем молочную кислоту. Например, такой ферментативный бульон содержит дешевый источник углерода, такой как пшеничные отруби или рафинированные сахара, такие как глюкоза, галактоза, ксилоза, сахароза, глицерин и т. д., в качестве источника углерода. Затем в ферментативном бульоне задействуют молочную кислоту для того, чтобы сделать возможным продуцирование PLA. Эту молочную кислоту можно получать с помощью микроорганизма, такого как лактобактерии, непосредственно в том же ферментативном бульоне. Альтернативно, молочную кислоту можно получать с помощью такого микроорганизма, как лактобактерии, в другом ферментативном бульоне, который соединен или не соединен с ферментативным бульоном для продуцирования PLA.
Благоприятно, ферментативный бульон содержит по меньшей мере D-LA для того, чтобы продуцировать PDLA.
В конкретном варианте осуществления дегидрогеназу D-молочной кислоты инактивировали в рекомбинантной клетке дрожжей, дополнительно содержащей PDLA-синтазу, чтобы содействовать продуцированию PDLA.
В конкретном варианте осуществления pH в среде управляют для того, чтобы увеличивать захват клеткой молочной кислоты из среды. Предпочтительно, pH поддерживают между 3 и 7, более предпочтительно между 3 и 4.
Благоприятно, способ по изобретению ведет к продуцированию по меньшей мере 0,01 г PLA/г сухих клеток, по меньшей мере 0,02 г, 0,03 г, 0,04 г, 0,05 г, 0,1 г, 0,2 г, 0,5 г или больше PLA/г сухих клеток. Предпочтительно, способ по изобретению ведет к продуцированию по меньшей мере 1 г PLA/г сухих клеток.
В конкретном варианте осуществления способ по изобретению включает стадию извлечения PLA из клеток и необязательно стадию очистки экстрагированной PLA.
PLA можно экстрагировать из клеток дрожжей любым способом, известным специалисту.
В конкретном варианте осуществления клетки лизируют перед извлечением PLA. В частности, клетки можно лизировать биологически, посредством их контакта с ферментом, способным расщеплять их клеточные стенки (например, зимолиазой). Альтернативно, клетки дрожжей можно лизировать механически, разрушая их клеточные стенки, например, с использованием вортекса со стеклянными шариками, или лизировать химически, например, посредством осмотического шока или обработки щелочью.
PLA можно экстрагировать непосредственно из не лизированных клеток или после лизиса с использованием процесса экстракции растворителем, такого как экстрагирование по Сокслету (Yamada et al., 2011), мацерация в растворителе (т. е. хлороформе, дихлорметане, ацетоне и т. д.), фильтрование и преципитация или ультрацентрифугирование и т. д. После экстрагирования и солюбилизации очистку PLA можно выполнять с использованием способа преципитации в соответствии с Matsumoto et al., (2005) или любого другого способа, известного специалисту в данной области.
В соответствии с изобретением, извлекаемая PLA благоприятно имеет молекулярную массу (Mw) больше чем 40000 г/моль, 50000 г/моль, 60000 г/моль, 70000 г/моль или 80000 г/моль.
Изобретение дополнительно предусматривает ферментативный бульон, содержащий такие рекомбинантные клетки дрожжей, и/или бесклеточный экстракт таких рекомбинантных клеток дрожжей, выбранный из супернатанта клеток, клеточного детрита и экстракта клеточных стенок. Изобретение дополнительно относится к высушенным рекомбинантным клеткам дрожжей, содержащим PLA в их цитозоле и/или пероксисоме и/или митохондриях. В соответствии с изобретением, такой ферментативный бульон и/или высушенные клетки можно использовать непосредственно в качестве сырья для получения пластмассовых продуктов или тому подобного. Например, высушенные клетки по изобретению можно вводить непосредственно в экструдер, необязательно с другими сложными полиэфирами, в том числе другими PLA, чтобы предоставлять PLA-содержащий материал и/или PLA-содержащие пластмассовые продукты.
Говоря в общем, настоящее изобретение предлагает использование рекомбинантных клеток дрожжей, как описано выше, для получения PLA, предпочтительно PDLA. Интересно, что получаемая PLA демонстрирует высокую молекулярную массу (Mw), в целом больше 40000 г/моль, 50000 г/моль, 60000 г/моль, 70000 г/моль или 80000 г/моль.
Следовательно, цель настоящего изобретения состоит в том, чтобы предоставить PLA, более конкретно PDLA, имеющую молекулярную массу (Mw) больше 40000 г/моль, 50000 г/моль, 60000 г/моль, 70000 г/моль или 80000 г/моль.
Дополнительные аспекты и преимущества по изобретению раскрыты в следующих примерах, которые следует рассматривать в качестве иллюстративных и не ограничивающих объем этой заявки.
ПРИМЕРЫ
Пример 1: конструирование штаммов дрожжей, не способных ассимилировать молочную кислоту
Конструирование нарушенных штаммов с последующим удалением маркера
Разрушающие кассеты создавали посредством ПЦР амплификации на геномной ДНК Y. lipolytica. Это заключается в первой амплификации промоторной области (P) и области терминации (T) гена, подлежащего удалению. Праймеры разрабатывали, как описано у Fickers и сотрудников (2003) и/или как описано у Beopoulos et al. (2008 и 2011). После вторичной амплификации, состоящей в слиянии фрагментов P и T, получаемую кассету PT вставляли в вектор PCR4RBlunt-TOPO из Life Technologies (Carlsbad, California). Затем ауксотрофный маркер URA3ex или LEU2ex вставляли в вектор PT через специфический сайт клонирования IsceI, чтобы создавать соответствующую плазмиду (PUT или PLT, соответственно). Разрушающую кассету PUT или PLT вводили в Y. lipolytica посредством трансформации способом с ацетатом лития (Barth et al. 1996). Трансформанты отбирали на достаточной минимальной среде. Верификационные праймеры, кодирующие конкретные последовательности, присутствующие в ауксотрофном маркере и в гене, использовали для того, чтобы верифицировать разрушение гена посредством ПЦР амплификации геномных локусов. Спасение маркера осуществляли после трансформации с использованием репликативной плазмиды pUB-CreI, как описано у Fickers и сотрудников (2003).
Эффект разрушения YlDLD1, оказываемый на рост на D-лактате
Рост штамма с нокаутом YlDLD1 (ThYl_434 - MATA ura3-302 leu2-270 xpr2-322, ku70Δ, zeta, DLD1::URA3, LEU2ex-) (SEQ ID № 12: ген YlDLD1) сравнивали с ростом штамма-хозяина (JMY2341 - MATA ura3-302 leu2-270 xpr2-322, KU70::URA3ex, zeta-LEU2ex-) на среде YNB (1,7 г/л YNB, 5 г/л NH4Cl, 50 мМ фосфатный буфер, pH 6,8), содержащей лейцин 400 мг/л и различные источники углерода: глюкоза 10 г/л или DL-лактат 10 г/л или D-лактат 10 г/л или L-лактат 10 г/л (фиг. 4A и 4B). Рост на D-, L- и DL-лактате достигает той же OD600нм у штамма-хозяина, тогда как рост нокаутного штамма на D-лактате полностью прекращался и рост на DL-лактате достигал половины от OD600нм при росте на L-лактате, что подсказывает, что нокаутный штамм не потребляет D-лактат и его рост обусловлен потреблением L-лактата.
Затем можно модифицировать штамм с нарушенной YlDLD1 в соответствии с изобретением для того, чтобы продуцировать PLA, более предпочтительно PDLA.
Эффект делеции YlCYB21, оказываемый на рост на L-лактате
Рост штамма с нокаутом YlCYB21 (ThYl_436 - MATA ura3-302 leu2-270 xpr2-322, ku70Δ, zeta, CYB21::LEU2ex, URA3ex-) (SEQ ID № 13: ген YlCYB21) сравнивали с ростом штамма-хозяина (JMY2341) на среде YNB, содержащей L-лактат 10 г/л (фиг. 5A и 5B). Тогда как JMY2341 растет на L-лактате, который полностью расходовался после 72 часов, нарушенный штамм не потребляет L-лактат и поэтому не растет.
Затем штамм с нокаутом YlCYB21 можно модифицировать в соответствии с изобретением для того, чтобы продуцировать PLA, более предпочтительно PLLA.
Пример 2: конструирование штаммов дрожжей, способных продуцировать PLA
Клонирование и экспрессия генов под управлением конститутивного промотора TEF
Гены амплифицировали посредством ПЦР и помещали под управление конститутивного промотора TEF дрожжей (pTEF). Затем кодирующие генные последовательности вставляли между сайтами BamHI-AvrII экспрессирующего вектора JMP62-pTEF, содержащего селективный маркер URA3ex или LEU2ex. Затем плазмиды расщепляли рестрикционным ферментом NotI и высвобожденный содержащий кодирующий ген фрагмент использовали для трансформации штаммов способом с ацетатом лития (Barth et al. 1996). Трансформанты отбирали по соответствующей им ауксотрофии на достаточной минимальной среде. Спасение маркера осуществляли после трансформации репликативной плазмидой pUB-CreI, как описано у Fickers и сотрудников (2003).
Штамм дрожжей
Используемый штамм Y. lipolytica получали из штамма JMY2159, описанного у Beopoulos et al., 2014 (QPF). Ген YlDLD1 (SEQ ID № 12) удаляли для получения штамма ThYl_967 с использованием протокола, описанного в примере 1 и на фиг. 6. Вариант Pctp из C. propionicum (SEQ ID № 1) с мутацией V193A (CpPctp opt V193A - ген: SEQ ID № 14) и вариант PHA-синтазы (PhaC1p opt E130D S325T S477R Q481M - SEQ ID № 11) из P. aeruginosa PAO1 с мутациями E130D; S325T; S477R; Q481M (ген: SEQ ID № 15) экспрессировали под управлением того же промотора TEF. Вариант Pctp экспрессировали в цитозоле и вариант PhaC1p направляли в пероксисомы. Наконец, генотип штамма ThYl_976, используемого в этом примере, представлял собой: MATA ura3-302 leu2-270 xpr2-322, pox1-6Δ, dga1Δ, lro1Δ, dga2Δ, fad2Δ, dld1Δ, pTEF-PaPHAC1 opt E130D, S325T, S477R, Q481M, perox-URA3ex, pTEF-CpPCT opt V193A cyto-LEU2ex, как показано на фиг. 6.
Условия культивирования
Рекомбинантные клетки дрожжей ThYl_976 сначала культивировали на богатой среде (дрожжевой экстракт 10 г/л, пептон 10 г/л, глюкоза 10 г/л) при 28°C в течение ночи. Затем клетки собирали посредством центрифугирования для того, чтобы удалять среду, и ресуспендировали в контролируемой среде с L-молочной кислотой в качестве единственного источника углерода и D-молочной кислоте в качестве синтона для продуцирования PDLA (YNB 1,7 г/л, казаминокислоты 2 г/л, NH4Cl 5 г/л, DL-молочная кислота 20 г/л, фосфатный буфер 50 мМ) при начальной OD600нм приблизительно 10. L-молочную кислоту добавляли по необходимости. Альтернативно, для усиленного накопления полимера, клетки ресуспендировали в контролируемой среде с глюкозой и L-молочной кислотой в качестве источника углерода и D-молочной кислотой в качестве синтона для продуцирования PDLA (YNB 1,7 г/л, казаминокислоты 2 г/л, NH4Cl 5 г/л, глюкоза 40 г/л, L-молочная кислота 15 г/л, D-молочная кислота 5 г/л, фосфатный буфер 50 мМ) при начальной OD600нм приблизительно 0,5. Культуры выращивали при 28°C со встряхиванием 100 об./мин в течение 150 часов и затем в указанное время клетки собирали посредством центрифугирования, промывали два раза водой и клеточный осадок хранили при -80°C до последующего анализа. Среды для культивирования фильтровали через 0,4 мкм фильтр и хранили при -20°C до анализа (см. фиг. 9).
Экстрагирование полимера
Приблизительно 1,5 г лиофилизированных клеток ресуспендировали в 15 мл 100 мМ Tris, pH 8, зимолиазе 0,5 мг/мл и инкубировали при 25°C в течение ночи. Клеточные суспензии замораживали при -80°C и/или сублимировали посредством лиофилизации до экстрагирования полимера.
Полученный полимер экстрагировали с использованием аппарата Сокслета и хлороформа. Использовали приблизительно 1,5 г высушенных клеток и камеру аппарата Сокслета заполняли 10 раз прежде, чем собирали растворитель и экстрагированные материалы.
Альтернативно, после экстрагирования, PLA можно преципитировать посредством добавления гексана (или циклогексана) в смесь.
Анализ
Концентрацию глюкозы определяли с использованием анализатора YSI 2900 (System C industrie, St Paul Trois Chateaux, France).
Концентрацию молочной кислоты определяли посредством высокоэффективной жидкостной хроматографии (HPLC) с использованием системы Thermo Fisher Scientific (Courtaboeuf, France), оборудованной UV-детектором на 254 нм и колонкой Phenomenex (Chirex 3126 (D)-penicillamine 150×4,6 мм, Le Pecq, France), используя 2 мМ CuSO4, 15% метанол (об./об.) в качестве подвижной фазы на 1 мл/мин.
Композицию полимера определяли посредством ЯМР на спектрометре Bruker Avance II 500. Клеточные экстракты тщательно сушили перед разведением в CDCl3, содержащим 1% TMS (внутренний стандарт), и переносили в 5 мм ЯМР пробирки. ЯМР спектры регистрировали при 298K. Каждый ЯМР спектр получали с использованием угла отклонения вектора возбуждения 30° с радиочастотным полем 29,7 кГц, задержкой релаксации 10 с и 2 пробными сканированиями. Для каждого эксперимента осуществляли 16 сканирований с задержкой повторения 6,5 с. Концентрации PLA определяли посредством интегрирования специфического квадруплетного сигнала на 4,19 м. д.
Средневзвешенную молекулярную массу (Mw) и дисперсность полимера определяли посредством гельпроникающей хроматографии (GPC) при 20°C с использованием системы Shimadzu (Marne la Vallée, France), оборудованной детекторами Wyatt (MALLS, Dawn Heleos-II, 18 углов и рефрактометр на 22°C, Optilab T-rEX, Toulouse, France) с двумя колонками Agilent (PLGel 5 мкм MIXED-C 300×7,5 мм). Используемый растворитель для элюирования представлял собой дихлорметан. Образцы ресуспендировали в дихлорметане и фильтровали через 0,4 мкм фильтр. Молекулярную массу Mw и дисперсность вычисляли с использованием значения dn/dc, предварительно определяемого и равного 0,0296.
Очистка PLA и анализ композиции
После экстрагирования PLA разводили в хлороформе и очищали посредством преципитации с добавлением 10 объемов гексана (по материалам Yamada et al., 2011). Затем осадитель собирали посредством фильтрования на PTFE мембране. Полный гидролиз полимера осуществляли при 180°C и 100 бар в течение 50 минут, в соответствии с Faisal et al. (2007), чтобы идентифицировать изомер молочной кислоты, который составлял полимер.
Результаты
В первый раз культуры осуществляли в DL-лактате, используя L-молочную кислоту в качестве единственного источника углерода. Рост дрожжей измеряли по OD на 600 нм и концентрации субстратов измеряли с течением времени. После 120 часов культивирования клетки собирали и обрабатывали как для экстракции PLA. Синтез PLA демонстрировали с использованием ЯМР на основании PLA-специфических сигналов и количественно определяли с использованием внутреннего стандарта (TMS) (фиг. 7). Данные ЯМР демонстрируют, что рекомбинантный штамм ThYl_976, проявляющий как активность КоА-трансферазы, так и активность лактил-КоА-полимеразы, продуцирует PLA, тогда как штамм, который не проявляет указанные выше активности (ThYl_964, см. фиг. 6), нет (спектр 1 против 2 на фиг. 7). Спектр также показывает, что основные контаминанты при экстрагировании по Сокслету представляют собой жирные кислоты, в первую очередь состоящие из олеиновой кислоты, которые извлекают в обоих экстрактах в схожих пропорциях. Спектр 3 показывает специфические сигналы, получаемые с использованием коммерческого PLA. Анализ показывает, что синтезированный полимер представляет собой гомополимер, содержащий 100% мономера молочной кислоты. Молекулярную массу Mw и дисперсность PLA определяли количественно с использованием GPC. Mw полученной PLA достигала 43800 г/моль после 120 часов культивирования с дисперсностью 1,4. Когда культивировали в DL-лактате с L-молочной кислотой, используемой в качестве единственного источника углерода, рекомбинантные клетки накапливали 1,9% (г/г) PLA на протяжении культивирования с конечной концентрацией в биомассе приблизительно 2,5 г/л.
Чтобы усовершенствовать накопление PLA и продуцирование биомассы, тестировали другие источники углерода. И то и другое можно значительно усовершенствовать с использованием глюкозы в качестве дополнительного источника углерода. При культивировании с DL-лактатом и глюкозой, с использованием L-молочной кислоты и глюкозы в качестве источника углерода, концентрацию субстратов отслеживали и восполняли при необходимости. Также измеряли концентрацию D-молочной кислоты, которая медленно снижалась с течением времени, указывая на то, что ее используют для продуцирования полимера PLA (фиг. 8).
Продуцирование PLA начиналось во время экспоненциальной фазы (первые 48 часов) с производительностью 0,21 мг/г/ч. Затем производительность возрастала до 0,33 мг/г/ч во время стационарной фазы. После 150 часов культивирования общая биомасса составляла приблизительно 10,5 г сухой клеточной массы на литр и общее продуцирование PLA достигало 0,5 г/л. Тогда накопление PLA составляло 4,25% (г/г) (фиг. 9).
После экстрагирования, PLA преципитировали и ее полный гидролиз осуществляли при 180°C, 100 бар в течение 50 минут. Затем определяли композицию PLA посредством HPLC анализа на хиральной колонке. Этот анализ показывает, что PLA состояла на 100% из D-молочной кислоты (фиг. 10).
Пример 3: усовершенствование продуцирования PLA посредством добавления двух копий генов Pctp opt V193A или PhaC1 opt E130D S325T S477R Q481M
Конструирование штаммов дрожжей
Используемый штамм Y. lipolytica получали из штамма ThYl_976, описанного в примере 2 и экспрессирующего одну копию варианта Pctp из C. propionicum (SEQ ID № 1) с мутацией V193A (CpPctp opt V193A) и одну копию варианта PHA-синтазы (PhaC1p opt E130D S325T S477R Q481M) из P. aeruginosa PAO1 с мутациями E130D; S325T; S477R; Q481M (SEQ ID № 11). Вторую копию гена, кодирующего вариант Pctp из C. propionicum (SEQ ID № 1) с мутацией V193A (CpPctp opt V193A), и/или вторую копию варианта PHA-синтазы (PhaC1p opt E130D S325T S477R Q481M) из P. aeruginosa PAO1 с мутациями E130D; S325T; S477R; Q481M (ген SEQ ID № 15) экспрессировали в штамме ThYl_976 при том же направлении, что и первую копию (цитозоль или пероксисома).
Различные штаммы, используемые в этом примере, создавали с одной или двумя копиями гена Pctp и одной или двумя копиями гена PhaC1p под управлением промотора TEF. Генотипы штаммов из этого примера описаны в таблице 3.
Таблица 3: название и генотип штаммов с одной или двумя копиям генов, представляющих интерес
Условия культивирования, экстрагирование и анализ полимера
Рекомбинантные клетки дрожжей сначала культивировали на богатой среде (дрожжевой экстракт 10 г/л, пептон 10 г/л, глюкоза 10 г/л) при 28°C в течение ночи. Затем клетки ресуспендировали в контролируемой среде с глюкозой и L-молочной кислотой в качестве источника углерода и D-молочной кислотой в качестве синтона для продуцирования PDLA (YNB 1,7 г/л, казаминокислоты 2 г/л, NH4Cl 5 г/л, глюкоза 40 г/л, DL-молочная кислота 10 г/л, фосфатный буфер 50 мМ) при начальной OD600нм приблизительно 0,5. Культуры выращивали при 28°C со встряхиванием 100 об./мин в течение 5 суток и затем клетки собирали посредством центрифугирования, промывали два раза водой и клеточный осадок хранили при -80°C до последующего анализа. Среды для культивирования фильтровали через 0,4 мкм фильтр и хранили при -20°C до анализа.
Экстрагирование полимера и ЯМР анализ выполняли, как описано в примере 2.
Результаты
После 5 суток культивирования собирали клетки и экстрагировали PLA. Накопление PLA (которое определяли с помощью количественного ЯМР) в ThYl_1086 было улучшено приблизительно на 42%, когда две копии обоих генов вводили в геном, по сравнению с контрольным штаммом ThYl_976.
Пример 4: усовершенствование продуцирования PLA с использованием сильного и конститутивного промотора (4UAS-TEF)
Клонирование и экспрессия генов под управлением сильного и конститутивного промотора 4UAS-TEF и конструирование штамма дрожжей
Гены амплифицировали посредством ПЦР и помещали под управление сильного и конститутивного промотора 4UAS-TEF дрожжей. Клонирование и экспрессию осуществляли с использованием того же приема, который описан в примере 2.
Используемый штамм Y. lipolytica получали из штамма ThYl_967, описанного в примере 2. Вариант Pctp из C. propionicum (SEQ ID № 1) с мутацией V193A (CpPctp opt V193A) экспрессировали в цитозоле и вариант PHA-синтазы (PhaC1p opt E130D S325T S477R Q481M - SEQ ID № 11) из P. aeruginosa PAO1 с мутациями E130D; S325T; S477R; Q481M (ген SEQ ID № 15) экспрессировали в пероксисоме. Для обоих генов использовали промотор TEF и 4UAS-TEF. Различные штаммы, используемые в этом примере, создавали с одним или двумя генами под управлением промотора TEF или промотора 4UAS-TEF. Генотипы штамма, используемого в этом примере, описаны в таблице 4.
Таблица 4: название и генотип штаммов с генами под управлением различных промоторов
Условия культивирования, экстрагирование и анализ полимера
Использовали те же условия культивирования, которые описаны в примере 3. Экстрагирование и анализ полимера осуществляли также, как описано в примере 2.
Результаты
После 5 суток культивирования, собирали клетки и экстрагировали PLA. Результаты количественного ЯМР описаны в таблице 5. Накопление PLA было усовершенствовано приблизительно на 28%, когда ген Pct находился под управлением сильного промотора 4UAS-TEF и ген PhaC1 под управлением промотора TEF (таблица 5, ThYl_1174 vs ThYl_976). Кроме того, накопление PLA было увеличено еще больше приблизительно на 46%, когда оба гена находились под управлением сильного промотора 4UAS-TEF (таблица 5, ThYl_1156 против ThYl_976).
Таблица 5: накопление PLA, определяемое с помощью количественной ЯМР
ЛИТЕРАТУРА
Barth G, Gaillardin C (1996) Yarrowia lipolytica. In: Wolf K (ed) Non conventional yeasts in biotechnology, vol 1. Springer-Verlag, Berlin, Heidelberg, New York, pp 313-388
Beopoulos A, Mrozova Z, Thevenieau F, Le Dall MT, Hapala I, Papanikolaou S, Chardot T, Nicaud JM (2008) Control of lipid accumulation in the yeast yarrowia lipolytica. Appl Environ Microbiol 74 (24):7779-7789
Beopoulos A, Nicaud JM, Gaillardin C (2011) An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl Microbiol Biotechnol 90 (4):1193-1206
Beopoulous A., Verbeke J., Bordes F., Guicherd M., Bressy M., Marty A. and Nicaud J.-M. (2014) Metabolic engineering for ricinoleic acid production in the yeast Yarrowia lipolitica. Appl. Microbiol. Biotechnol. 98(1):25-262
Faisal M., Saeki T., Tsuji T., Daimon H. and Fujie K. (2007) Depolymerization of poly(L-lactic acid) under hydrothermal conditions. Asian J. of Chem. 19:1714-1722Fickers P, Le Dall MT, Gaillardin C, Thonart P, Nicaud JM (2003) New disruption cassettes for rapid gene disruption and marker rescue in the yeast Yarrowia lipolytica. J Microbiol Methods 55 (3):727-737
Matsumoto K, Takase K, Aoki E, Doi Y and Taguchi S (2005) Synergistic Effects of Glu130Asp Substitution in the Type II Poly-hydroxyalkanoate (PHA) Synthase: Enhancement of PHA Production and Alteration of Polymer Molecular Weight. Biomacromolecules 2005, 6, 99-104
Meile, L., Rohr, L.M., Geissmann, T.A., Herensperger, M., and Teuber, M. (2001). Characterization of the D-Xylulose 5-Phosphate/D-Fructose 6-Phosphate Phosphoketolase Gene (xfp) from Bifidobacterium lactis. J. Bacteriol. 183:2929-2936
Yamada M, Matsumoto K, Uramoto S, Motohashi R, Abe H and Taguchi S (2011) Lactate fraction dependent mechanical properties of semitransparent poly(lactate-co-3-hydroxybutyrate)s produced by control of lactyl-CoA monomer fluxes in recombinant Escherichia coli. Journal of Biotechnology 154:255-260
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CARBIOS
<120> РЕКОМБИНАНТНЫЕ КЛЕТКИ ДРОЖЖЕЙ, ПРОДУЦИРУЮЩИЕ ПОЛИМОЛОЧНУЮ
КИСЛОТУ, И ИХ ИСПОЛЬЗОВАНИЯ
<130> B2161PC
<160> 18
<170> PatentIn версии 3.5
<210> 1
<211> 524
<212> Белок
<213> Clostridium propionicum
<400> 1
Met Arg Lys Val Pro Ile Ile Thr Ala Asp Glu Ala Ala Lys Leu Ile
1 5 10 15
Lys Asp Gly Asp Thr Val Thr Thr Ser Gly Phe Val Gly Asn Ala Ile
20 25 30
Pro Glu Ala Leu Asp Arg Ala Val Glu Lys Arg Phe Leu Glu Thr Gly
35 40 45
Glu Pro Lys Asn Ile Thr Tyr Val Tyr Cys Gly Ser Gln Gly Asn Arg
50 55 60
Asp Gly Arg Gly Ala Glu His Phe Ala His Glu Gly Leu Leu Lys Arg
65 70 75 80
Tyr Ile Ala Gly His Trp Ala Thr Val Pro Ala Leu Gly Lys Met Ala
85 90 95
Met Glu Asn Lys Met Glu Ala Tyr Asn Val Ser Gln Gly Ala Leu Cys
100 105 110
His Leu Phe Arg Asp Ile Ala Ser His Lys Pro Gly Val Phe Thr Lys
115 120 125
Val Gly Ile Gly Thr Phe Ile Asp Pro Arg Asn Gly Gly Gly Lys Val
130 135 140
Asn Asp Ile Thr Lys Glu Asp Ile Val Glu Leu Val Glu Ile Lys Gly
145 150 155 160
Gln Glu Tyr Leu Phe Tyr Pro Ala Phe Pro Ile His Val Ala Leu Ile
165 170 175
Arg Gly Thr Tyr Ala Asp Glu Ser Gly Asn Ile Thr Phe Glu Lys Glu
180 185 190
Val Ala Pro Leu Glu Gly Thr Ser Val Cys Gln Ala Val Lys Asn Ser
195 200 205
Gly Gly Ile Val Val Val Gln Val Glu Arg Val Val Lys Ala Gly Thr
210 215 220
Leu Asp Pro Arg His Val Lys Val Pro Gly Ile Tyr Val Asp Tyr Val
225 230 235 240
Val Val Ala Asp Pro Glu Asp His Gln Gln Ser Leu Asp Cys Glu Tyr
245 250 255
Asp Pro Ala Leu Ser Gly Glu His Arg Arg Pro Glu Val Val Gly Glu
260 265 270
Pro Leu Pro Leu Ser Ala Lys Lys Val Ile Gly Arg Arg Gly Ala Ile
275 280 285
Glu Leu Glu Lys Asp Val Ala Val Asn Leu Gly Val Gly Ala Pro Glu
290 295 300
Tyr Val Ala Ser Val Ala Asp Glu Glu Gly Ile Val Asp Phe Met Thr
305 310 315 320
Leu Thr Ala Glu Ser Gly Ala Ile Gly Gly Val Pro Ala Gly Gly Val
325 330 335
Arg Phe Gly Ala Ser Tyr Asn Ala Asp Ala Leu Ile Asp Gln Gly Tyr
340 345 350
Gln Phe Asp Tyr Tyr Asp Gly Gly Gly Leu Asp Leu Cys Tyr Leu Gly
355 360 365
Leu Ala Glu Cys Asp Glu Lys Gly Asn Ile Asn Val Ser Arg Phe Gly
370 375 380
Pro Arg Ile Ala Gly Cys Gly Gly Phe Ile Asn Ile Thr Gln Asn Thr
385 390 395 400
Pro Lys Val Phe Phe Cys Gly Thr Phe Thr Ala Gly Gly Leu Lys Val
405 410 415
Lys Ile Glu Asp Gly Lys Val Ile Ile Val Gln Glu Gly Lys Gln Lys
420 425 430
Lys Phe Leu Lys Ala Val Glu Gln Ile Thr Phe Asn Gly Asp Val Ala
435 440 445
Leu Ala Asn Lys Gln Gln Val Thr Tyr Ile Thr Glu Arg Cys Val Phe
450 455 460
Leu Leu Lys Glu Asp Gly Leu His Leu Ser Glu Ile Ala Pro Gly Ile
465 470 475 480
Asp Leu Gln Thr Gln Ile Leu Asp Val Met Asp Phe Ala Pro Ile Ile
485 490 495
Asp Arg Asp Ala Asn Gly Gln Ile Lys Leu Met Asp Ala Ala Leu Phe
500 505 510
Ala Glu Gly Leu Met Gly Leu Lys Glu Met Lys Ser
515 520
<210> 2
<211> 694
<212> Белок
<213> Aspergillus nidulans
<400> 2
Met Thr His Pro Gln Gln Ala Val His Ala Ala Ser Leu Gln Asn Pro
1 5 10 15
Glu Ala Phe Trp Ser His His Ala Gln Gln Leu His Trp His Lys Lys
20 25 30
Pro Ser Arg Ala Ile Gly Arg Ser Thr Lys Thr Leu Ala Ser Gly Ala
35 40 45
Ser His Glu Ser Trp Ser Trp Phe Pro Asp Gly Glu Ile Ser Thr Thr
50 55 60
Tyr Asn Cys Val Asp Arg His Val Leu Asn Gly Asn Gly Asp Asn Val
65 70 75 80
Ala Ile Ile Trp Asp Ser Ala Val Thr Gly Lys Lys Glu Lys Tyr Thr
85 90 95
Tyr Arg Gln Leu Leu Asp Glu Val Glu Val Leu Ala Gly Val Leu Arg
100 105 110
Glu Glu Gly Val Lys Lys Gly Asp Val Val Ile Ile Tyr Met Pro Met
115 120 125
Ile Pro Ala Ala Leu Ile Gly Ala Leu Ala Val Ala Arg Leu Gly Ala
130 135 140
Ile His Ala Ala Val Phe Gly Gly Phe Ala Ala Lys Ser Leu Ala Gln
145 150 155 160
Arg Ile Glu Ala Ala Arg Pro Arg Ala Ile Leu Thr Ala Ser Cys Gly
165 170 175
Ile Glu Gly Ala Lys Gly Pro Ile Ala Tyr Arg Pro Leu Val Glu Gly
180 185 190
Ala Ile Glu Ala Ser Ser Phe Lys Pro Glu Lys Val Leu Ile Trp Gln
195 200 205
Arg Asp Gln Leu Arg Trp Asn Asn Pro Asp Lys Leu Gly Gly Gln Arg
210 215 220
Asn Trp Asn Arg Leu Val Lys Ser Ala Arg Met Arg Gly Ile Arg Ala
225 230 235 240
Glu Pro Val Pro Val Arg Ser Thr Asp Gly Leu Tyr Ile Ile Tyr Thr
245 250 255
Ser Gly Thr Thr Gly Leu Pro Lys Gly Val Val Arg Glu Ala Gly Gly
260 265 270
His Ala Val Gly Leu Ser Leu Ser Ile Lys Tyr Leu Phe Asp Ile His
275 280 285
Gly Pro Gly Asp Thr Met Phe Cys Ala Ser Asp Ile Gly Trp Val Val
290 295 300
Gly His Ser Tyr Ile Leu Tyr Ala Pro Leu Leu Val Gly Ala Thr Thr
305 310 315 320
Val Leu Phe Glu Gly Lys Pro Val Gly Thr Pro Asp Ala Gly Thr Phe
325 330 335
Trp Arg Val Val Ala Glu His Lys Ala Asn Val Leu Phe Thr Ala Pro
340 345 350
Thr Ala Leu Arg Ala Ile Arg Lys Glu Asp Pro Asp Asn Lys His Phe
355 360 365
Glu Lys Val Ala Gly Asp Asn Asn Leu Arg His Leu Arg Ala Leu Phe
370 375 380
Leu Ala Gly Glu Arg Ser Glu Pro Ser Ile Val Arg Ala Tyr Gln Asp
385 390 395 400
Leu Leu Thr Lys His Ala Ala Arg Gly Ala Leu Val Val Asp Asn Trp
405 410 415
Trp Ser Ser Glu Ser Gly Ser Pro Ile Ser Gly Leu Ala Leu Arg Ser
420 425 430
Ala Val Gly Arg Val Pro Pro Arg Ser Asp Glu Tyr Asp Val Ala Pro
435 440 445
Leu Ala Ile Arg Pro Gly Ser Ala Gly Leu Pro Met Pro Gly Phe Asp
450 455 460
Val Arg Val Val Asp Asp Glu Gly Asn Glu Val Ala Gln Gly Thr Met
465 470 475 480
Gly Asn Ile Val Met Ala Thr Pro Leu Ala Pro Thr Ala Phe Thr Arg
485 490 495
Leu Phe Asn Asp Asp Glu Arg Phe Tyr Lys Gly Tyr Leu Lys Arg Phe
500 505 510
Gly Gly Arg Trp Leu Asp Thr Gly Asp Ala Gly Met Ile Asp Gln Asp
515 520 525
Gly Tyr Ile His Val Met Ser Arg Ser Asp Asp Ile Ile Asn Val Ala
530 535 540
Ala His Arg Phe Ser Thr Gly Gln Gly Ser Ile Glu Gln Ala Ile Leu
545 550 555 560
Ser His Pro Ala Ile Gly Glu Ala Ser Val Val Gly Ile Pro Asp Ala
565 570 575
Leu Lys Gly His Leu Pro Phe Ala Phe Ile Thr Leu Lys Gln Ser Gly
580 585 590
Gly Asn Ser Pro Ala Arg Pro Ser Ala Glu Leu Phe Asn Ser Val Asn
595 600 605
Arg Leu Val Arg Glu Gln Ile Gly Ala Ile Ala Ser Leu Gly Gly Met
610 615 620
Ile Gln Gly Gln Gly Met Ile Pro Lys Thr Arg Ser Gly Lys Thr Leu
625 630 635 640
Arg Arg Val Leu Arg Glu Leu Val Glu Asn Gly Ala Arg Gly Glu Phe
645 650 655
Glu Lys Glu Val Ala Val Pro Pro Thr Val Glu Asp Arg Gly Val Val
660 665 670
Glu Val Ala Arg Glu Lys Val Arg Glu Tyr Phe Glu Ser Gln Ser Gly
675 680 685
Ser Pro Lys Ala Lys Leu
690
<210> 3
<211> 531
<212> Белок
<213> Escherichia coli
<400> 3
Met Lys Pro Val Lys Pro Pro Arg Ile Asn Gly Arg Val Pro Val Leu
1 5 10 15
Ser Ala Gln Glu Ala Val Asn Tyr Ile Pro Asp Glu Ala Thr Leu Cys
20 25 30
Val Leu Gly Ala Gly Gly Gly Ile Leu Glu Ala Thr Thr Leu Ile Thr
35 40 45
Ala Leu Ala Asp Lys Tyr Lys Gln Thr Gln Thr Pro Arg Asn Leu Ser
50 55 60
Ile Ile Ser Pro Thr Gly Leu Gly Asp Arg Ala Asp Arg Gly Ile Ser
65 70 75 80
Pro Leu Ala Gln Glu Gly Leu Val Lys Trp Ala Leu Cys Gly His Trp
85 90 95
Gly Gln Ser Pro Arg Ile Ser Asp Leu Ala Glu Gln Asn Lys Ile Ile
100 105 110
Ala Tyr Asn Tyr Pro Gln Gly Val Leu Thr Gln Thr Leu Arg Ala Ala
115 120 125
Ala Ala His Gln Pro Gly Ile Ile Ser Asp Ile Gly Ile Gly Thr Phe
130 135 140
Val Asp Pro Arg Gln Gln Gly Gly Lys Leu Asn Glu Val Thr Lys Glu
145 150 155 160
Asp Leu Ile Lys Leu Val Glu Phe Asp Asn Lys Glu Tyr Leu Tyr Tyr
165 170 175
Lys Ala Ile Ala Pro Asp Ile Ala Phe Ile Arg Ala Thr Thr Cys Asp
180 185 190
Ser Glu Gly Tyr Ala Thr Phe Glu Asp Glu Val Met Tyr Leu Asp Ala
195 200 205
Leu Val Ile Ala Gln Ala Val His Asn Asn Gly Gly Ile Val Met Met
210 215 220
Gln Val Gln Lys Met Val Lys Lys Ala Thr Leu His Pro Lys Ser Val
225 230 235 240
Arg Ile Pro Gly Tyr Leu Val Asp Ile Val Val Val Asp Pro Asp Gln
245 250 255
Ser Gln Leu Tyr Gly Gly Ala Pro Val Asn Arg Phe Ile Ser Gly Asp
260 265 270
Phe Thr Leu Asp Asp Ser Thr Lys Leu Ser Leu Pro Leu Asn Gln Arg
275 280 285
Lys Leu Val Ala Arg Arg Ala Leu Phe Glu Met Arg Lys Gly Ala Val
290 295 300
Gly Asn Val Gly Val Gly Ile Ala Asp Gly Ile Gly Leu Val Ala Arg
305 310 315 320
Glu Glu Gly Cys Ala Asp Asp Phe Ile Leu Thr Val Glu Thr Gly Pro
325 330 335
Ile Gly Gly Ile Thr Ser Gln Gly Ile Ala Phe Gly Ala Asn Val Asn
340 345 350
Thr Arg Ala Ile Leu Asp Met Thr Ser Gln Phe Asp Phe Tyr His Gly
355 360 365
Gly Gly Leu Asp Val Cys Tyr Leu Ser Phe Ala Glu Val Asp Gln His
370 375 380
Gly Asn Val Gly Val His Lys Phe Asn Gly Lys Ile Met Gly Thr Gly
385 390 395 400
Gly Phe Ile Asp Ile Ser Ala Thr Ser Lys Lys Ile Ile Phe Cys Gly
405 410 415
Thr Leu Thr Ala Gly Ser Leu Lys Thr Glu Ile Ala Asp Gly Lys Leu
420 425 430
Asn Ile Val Gln Glu Gly Arg Val Lys Lys Phe Ile Arg Glu Leu Pro
435 440 445
Glu Ile Thr Phe Ser Gly Lys Ile Ala Leu Glu Arg Gly Leu Asp Val
450 455 460
Arg Tyr Ile Thr Glu Arg Ala Val Phe Thr Leu Lys Glu Asp Gly Leu
465 470 475 480
His Leu Ile Glu Ile Ala Pro Gly Val Asp Leu Gln Lys Asp Ile Leu
485 490 495
Asp Lys Met Asp Phe Thr Pro Val Ile Ser Pro Glu Leu Lys Leu Met
500 505 510
Asp Glu Arg Leu Phe Ile Asp Ala Ala Met Gly Phe Val Leu Pro Glu
515 520 525
Ala Ala His
530
<210> 4
<211> 542
<212> Белок
<213> Ralstonia. eutropha
<400> 4
Met Lys Val Ile Thr Ala Arg Glu Ala Ala Ala Leu Val Gln Asp Gly
1 5 10 15
Trp Thr Val Ala Ser Ala Gly Phe Val Gly Ala Gly His Ala Glu Ala
20 25 30
Val Thr Glu Ala Leu Glu Gln Arg Phe Leu Gln Ser Gly Leu Pro Arg
35 40 45
Asp Leu Thr Leu Val Tyr Ser Ala Gly Gln Gly Asp Arg Gly Ala Arg
50 55 60
Gly Val Asn His Phe Gly Asn Ala Gly Met Thr Ala Ser Ile Val Gly
65 70 75 80
Gly His Trp Arg Ser Ala Thr Arg Leu Ala Thr Leu Ala Met Ala Glu
85 90 95
Gln Cys Glu Gly Tyr Asn Leu Pro Gln Gly Val Leu Thr His Leu Tyr
100 105 110
Arg Ala Ile Ala Gly Gly Lys Pro Gly Val Met Thr Lys Ile Gly Leu
115 120 125
His Thr Phe Val Asp Pro Arg Thr Ala Gln Asp Ala Arg Tyr His Gly
130 135 140
Gly Ala Val Asn Glu Arg Ala Arg Gln Ala Ile Ala Glu Gly Lys Ala
145 150 155 160
Cys Trp Val Asp Ala Val Asp Phe Arg Gly Asp Glu Tyr Leu Phe Tyr
165 170 175
Pro Ser Phe Pro Ile His Cys Ala Leu Ile Arg Cys Thr Ala Ala Asp
180 185 190
Ala Arg Gly Asn Leu Ser Thr His Arg Glu Ala Phe His His Glu Leu
195 200 205
Leu Ala Met Ala Gln Ala Ala His Asn Ser Gly Gly Ile Val Ile Ala
210 215 220
Gln Val Glu Ser Leu Val Asp His His Glu Ile Leu Gln Ala Ile His
225 230 235 240
Val Pro Gly Ile Leu Val Asp Tyr Val Val Val Cys Asp Asn Pro Ala
245 250 255
Asn His Gln Met Thr Phe Ala Glu Ser Tyr Asn Pro Ala Tyr Val Thr
260 265 270
Pro Trp Gln Gly Glu Ala Ala Val Ala Glu Ala Glu Ala Ala Pro Val
275 280 285
Ala Ala Gly Pro Leu Asp Ala Arg Thr Ile Val Gln Arg Arg Ala Val
290 295 300
Met Glu Leu Ala Arg Arg Ala Pro Arg Val Val Asn Leu Gly Val Gly
305 310 315 320
Met Pro Ala Ala Val Gly Met Leu Ala His Gln Ala Gly Leu Asp Gly
325 330 335
Phe Thr Leu Thr Val Glu Ala Gly Pro Ile Gly Gly Thr Pro Ala Asp
340 345 350
Gly Leu Ser Phe Gly Ala Ser Ala Tyr Pro Glu Ala Val Val Asp Gln
355 360 365
Pro Ala Gln Phe Asp Phe Tyr Glu Gly Gly Gly Ile Asp Leu Ala Ile
370 375 380
Leu Gly Leu Ala Glu Leu Asp Gly His Gly Asn Val Asn Val Ser Lys
385 390 395 400
Phe Gly Glu Gly Glu Gly Ala Ser Ile Ala Gly Val Gly Gly Phe Ile
405 410 415
Asn Ile Thr Gln Ser Ala Arg Ala Val Val Phe Met Gly Thr Leu Thr
420 425 430
Ala Gly Gly Leu Glu Val Arg Ala Gly Asp Gly Gly Leu Gln Ile Val
435 440 445
Arg Glu Gly Arg Val Lys Lys Ile Val Pro Glu Val Ser His Leu Ser
450 455 460
Phe Asn Gly Pro Tyr Val Ala Ser Leu Gly Ile Pro Val Leu Tyr Ile
465 470 475 480
Thr Glu Arg Ala Val Phe Glu Met Arg Ala Gly Ala Asp Gly Glu Ala
485 490 495
Arg Leu Thr Leu Val Glu Ile Ala Pro Gly Val Asp Leu Gln Arg Asp
500 505 510
Val Leu Asp Gln Cys Ser Thr Pro Ile Ala Val Ala Gln Asp Leu Arg
515 520 525
Glu Met Asp Ala Arg Leu Phe Gln Ala Gly Pro Leu His Leu
530 535 540
<210> 5
<211> 559
<212> Белок
<213> Pseudomonas aeruginosa PAO1
<400> 5
Met Ser Gln Lys Asn Asn Asn Glu Leu Pro Lys Gln Ala Ala Glu Asn
1 5 10 15
Thr Leu Asn Leu Asn Pro Val Ile Gly Ile Arg Gly Lys Asp Leu Leu
20 25 30
Thr Ser Ala Arg Met Val Leu Leu Gln Ala Val Arg Gln Pro Leu His
35 40 45
Ser Ala Arg His Val Ala His Phe Ser Leu Glu Leu Lys Asn Val Leu
50 55 60
Leu Gly Gln Ser Glu Leu Arg Pro Gly Asp Asp Asp Arg Arg Phe Ser
65 70 75 80
Asp Pro Ala Trp Ser Gln Asn Pro Leu Tyr Lys Arg Tyr Met Gln Thr
85 90 95
Tyr Leu Ala Trp Arg Lys Glu Leu His Ser Trp Ile Ser His Ser Asp
100 105 110
Leu Ser Pro Gln Asp Ile Ser Arg Gly Gln Phe Val Ile Asn Leu Leu
115 120 125
Thr Glu Ala Met Ser Pro Thr Asn Ser Leu Ser Asn Pro Ala Ala Val
130 135 140
Lys Arg Phe Phe Glu Thr Gly Gly Lys Ser Leu Leu Asp Gly Leu Gly
145 150 155 160
His Leu Ala Lys Asp Leu Val Asn Asn Gly Gly Met Pro Ser Gln Val
165 170 175
Asp Met Asp Ala Phe Glu Val Gly Lys Asn Leu Ala Thr Thr Glu Gly
180 185 190
Ala Val Val Phe Arg Asn Asp Val Leu Glu Leu Ile Gln Tyr Arg Pro
195 200 205
Ile Thr Glu Ser Val His Glu Arg Pro Leu Leu Val Val Pro Pro Gln
210 215 220
Ile Asn Lys Phe Tyr Val Phe Asp Leu Ser Pro Asp Lys Ser Leu Ala
225 230 235 240
Arg Phe Cys Leu Arg Asn Gly Val Gln Thr Phe Ile Val Ser Trp Arg
245 250 255
Asn Pro Thr Lys Ser Gln Arg Glu Trp Gly Leu Thr Thr Tyr Ile Glu
260 265 270
Ala Leu Lys Glu Ala Ile Glu Val Val Leu Ser Ile Thr Gly Ser Lys
275 280 285
Asp Leu Asn Leu Leu Gly Ala Cys Ser Gly Gly Ile Thr Thr Ala Thr
290 295 300
Leu Val Gly His Tyr Val Ala Ser Gly Glu Lys Lys Val Asn Ala Phe
305 310 315 320
Thr Gln Leu Val Ser Val Leu Asp Phe Glu Leu Asn Thr Gln Val Ala
325 330 335
Leu Phe Ala Asp Glu Lys Thr Leu Glu Ala Ala Lys Arg Arg Ser Tyr
340 345 350
Gln Ser Gly Val Leu Glu Gly Lys Asp Met Ala Lys Val Phe Ala Trp
355 360 365
Met Arg Pro Asn Asp Leu Ile Trp Asn Tyr Trp Val Asn Asn Tyr Leu
370 375 380
Leu Gly Asn Gln Pro Pro Ala Phe Asp Ile Leu Tyr Trp Asn Asn Asp
385 390 395 400
Thr Thr Arg Leu Pro Ala Ala Leu His Gly Glu Phe Val Glu Leu Phe
405 410 415
Lys Ser Asn Pro Leu Asn Arg Pro Gly Ala Leu Glu Val Ser Gly Thr
420 425 430
Pro Ile Asp Leu Lys Gln Val Thr Cys Asp Phe Tyr Cys Val Ala Gly
435 440 445
Leu Asn Asp His Ile Thr Pro Trp Glu Ser Cys Tyr Lys Ser Ala Arg
450 455 460
Leu Leu Gly Gly Lys Cys Glu Phe Ile Leu Ser Asn Ser Gly His Ile
465 470 475 480
Gln Ser Ile Leu Asn Pro Pro Gly Asn Pro Lys Ala Arg Phe Met Thr
485 490 495
Asn Pro Glu Leu Pro Ala Glu Pro Lys Ala Trp Leu Glu Gln Ala Gly
500 505 510
Lys His Ala Asp Ser Trp Trp Leu His Trp Gln Gln Trp Leu Ala Glu
515 520 525
Arg Ser Gly Lys Thr Arg Lys Ala Pro Ala Ser Leu Gly Asn Lys Thr
530 535 540
Tyr Pro Ala Gly Glu Ala Ala Pro Gly Thr Tyr Val His Glu Arg
545 550 555
<210> 6
<211> 589
<212> Белок
<213> Ralstonia. eutropha H16
<400> 6
Met Ala Thr Gly Lys Gly Ala Ala Ala Ser Thr Gln Glu Gly Lys Ser
1 5 10 15
Gln Pro Phe Lys Val Thr Pro Gly Pro Phe Asp Pro Ala Thr Trp Leu
20 25 30
Glu Trp Ser Arg Gln Trp Gln Gly Thr Glu Gly Asn Gly His Ala Ala
35 40 45
Ala Ser Gly Ile Pro Gly Leu Asp Ala Leu Ala Gly Val Lys Ile Ala
50 55 60
Pro Ala Gln Leu Gly Asp Ile Gln Gln Arg Tyr Met Lys Asp Phe Ser
65 70 75 80
Ala Leu Trp Gln Ala Met Ala Glu Gly Lys Ala Glu Ala Thr Gly Pro
85 90 95
Leu His Asp Arg Arg Phe Ala Gly Asp Ala Trp Arg Thr Asn Leu Pro
100 105 110
Tyr Arg Phe Ala Ala Ala Phe Tyr Leu Leu Asn Ala Arg Ala Leu Thr
115 120 125
Glu Leu Ala Asp Ala Val Glu Ala Asp Ala Lys Thr Arg Gln Arg Ile
130 135 140
Arg Phe Ala Ile Ser Gln Trp Val Asp Ala Met Ser Pro Ala Asn Phe
145 150 155 160
Leu Ala Thr Asn Pro Glu Ala Gln Arg Leu Leu Ile Glu Ser Gly Gly
165 170 175
Glu Ser Leu Arg Ala Gly Val Arg Asn Met Met Glu Asp Leu Thr Arg
180 185 190
Gly Lys Ile Ser Gln Thr Asp Glu Ser Ala Phe Glu Val Gly Arg Asn
195 200 205
Val Ala Val Thr Glu Gly Ala Val Val Phe Glu Asn Glu Tyr Phe Gln
210 215 220
Leu Leu Gln Tyr Lys Pro Leu Thr Asp Lys Val His Ala Arg Pro Leu
225 230 235 240
Leu Met Val Pro Pro Cys Ile Asn Lys Tyr Tyr Ile Leu Asp Leu Gln
245 250 255
Pro Glu Ser Ser Leu Val Arg His Val Val Glu Gln Gly His Thr Val
260 265 270
Phe Leu Val Ser Trp Arg Asn Pro Asp Ala Ser Met Ala Gly Ser Thr
275 280 285
Trp Asp Asp Tyr Ile Glu His Ala Ala Ile Arg Ala Ile Glu Val Ala
290 295 300
Arg Asp Ile Ser Gly Gln Asp Lys Ile Asn Val Leu Gly Phe Cys Val
305 310 315 320
Gly Gly Thr Ile Val Ser Thr Ala Leu Ala Val Leu Ala Ala Arg Gly
325 330 335
Glu His Pro Ala Ala Ser Val Thr Leu Leu Thr Thr Leu Leu Asp Phe
340 345 350
Ala Asp Thr Gly Ile Leu Asp Val Phe Val Asp Glu Gly His Val Gln
355 360 365
Leu Arg Glu Ala Thr Leu Gly Gly Gly Ala Gly Ala Pro Cys Ala Leu
370 375 380
Leu Arg Gly Leu Glu Leu Ala Asn Thr Phe Ser Phe Leu Arg Pro Asn
385 390 395 400
Asp Leu Val Trp Asn Tyr Val Val Asp Asn Tyr Leu Lys Gly Asn Thr
405 410 415
Pro Val Pro Phe Asp Leu Leu Phe Trp Asn Gly Asp Ala Thr Asn Leu
420 425 430
Pro Gly Pro Trp Tyr Cys Trp Tyr Leu Arg His Thr Tyr Leu Gln Asn
435 440 445
Glu Leu Lys Val Pro Gly Lys Leu Thr Val Cys Gly Val Pro Val Asp
450 455 460
Leu Ala Ser Ile Asp Val Pro Thr Tyr Ile Tyr Gly Ser Arg Glu Asp
465 470 475 480
His Ile Val Pro Trp Thr Ala Ala Tyr Ala Ser Thr Ala Leu Leu Ala
485 490 495
Asn Lys Leu Arg Phe Val Leu Gly Ala Ser Gly His Ile Ala Gly Val
500 505 510
Ile Asn Pro Pro Ala Lys Asn Lys Arg Ser His Trp Thr Asn Asp Ala
515 520 525
Leu Pro Glu Ser Pro Gln Gln Trp Leu Ala Gly Ala Ile Glu His His
530 535 540
Gly Ser Trp Trp Pro Asp Trp Thr Ala Trp Leu Ala Gly Gln Ala Gly
545 550 555 560
Ala Lys Arg Ala Ala Pro Ala Asn Tyr Gly Asn Ala Arg Tyr Arg Ala
565 570 575
Ile Glu Pro Ala Pro Gly Arg Tyr Val Lys Ala Lys Ala
580 585
<210> 7
<211> 355
<212> Белок
<213> Chromatium vinosum
<400> 7
Met Phe Pro Ile Asp Ile Arg Pro Asp Lys Leu Thr Gln Glu Met Leu
1 5 10 15
Asp Tyr Ser Arg Lys Leu Gly Gln Gly Met Glu Asn Leu Leu Asn Ala
20 25 30
Glu Ala Ile Asp Thr Gly Val Ser Pro Lys Gln Ala Val Tyr Ser Glu
35 40 45
Asp Lys Leu Val Leu Tyr Arg Tyr Asp Arg Pro Glu Gly Ala Pro Glu
50 55 60
Ala Gln Pro Val Pro Leu Leu Ile Val Tyr Ala Leu Val Asn Arg Pro
65 70 75 80
Tyr Met Thr Asp Ile Gln Glu Asp Arg Ser Thr Ile Lys Gly Leu Leu
85 90 95
Ala Thr Gly Gln Asp Val Tyr Leu Ile Asp Trp Gly Tyr Pro Asp Gln
100 105 110
Ala Asp Arg Ala Leu Thr Leu Asp Asp Tyr Ile Asn Gly Tyr Ile Asp
115 120 125
Arg Cys Val Asp Tyr Leu Arg Glu Ala His Gly Val Asp Lys Val Asn
130 135 140
Leu Leu Gly Ile Cys Gln Gly Gly Ala Phe Ser Leu Met Tyr Ser Ala
145 150 155 160
Leu His Pro Asp Lys Val Arg Asn Leu Val Thr Met Val Thr Pro Val
165 170 175
Asp Phe Lys Thr Pro Asp Asn Leu Leu Ser Ala Trp Val Gln Asn Val
180 185 190
Asp Ile Asp Leu Ala Val Asp Thr Met Gly Asn Ile Pro Gly Glu Leu
195 200 205
Leu Asn Trp Thr Phe Leu Ser Leu Lys Pro Phe Ser Leu Thr Gly Gln
210 215 220
Lys Tyr Val Asn Met Val Asp Leu Leu Asp Asp Pro Asp Lys Val Lys
225 230 235 240
Asn Phe Leu Arg Met Glu Lys Trp Ile Phe Asp Ser Pro Asp Gln Ala
245 250 255
Gly Glu Thr Phe Arg Gln Phe Ile Lys Asp Phe Tyr Gln Asn Asn Gly
260 265 270
Phe Leu Asn Gly Gly Val Val Leu Gly Gly Gln Glu Val Asp Leu Lys
275 280 285
Asp Ile Thr Cys Pro Val Leu Asn Ile Phe Ala Leu Gln Asp His Leu
290 295 300
Val Pro Pro Asp Ala Ser Arg Ala Leu Lys Gly Leu Thr Ser Ser Pro
305 310 315 320
Asp Tyr Thr Glu Leu Ala Phe Pro Gly Gly His Ile Gly Ile Tyr Val
325 330 335
Ser Gly Lys Ala Gln Lys Glu Val Thr Pro Ala Ile Gly Lys Trp Leu
340 345 350
Asn Glu Arg
355
<210> 8
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> PTS1
<400> 8
Met Gly Ala Gly Val Thr Glu Asp Gln Phe Lys Ser Lys Leu
1 5 10
<210> 9
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> PTS2
<400> 9
Met Asp Arg Leu Asn Asn Leu Ala Thr Gln Leu Glu Gln Asn Pro Ala
1 5 10 15
<210> 10
<211> 26
<212> Белок
<213> Искусственная последовательность
<220>
<223> MTS
<400> 10
Met Leu Arg Leu Arg Thr Met Arg Pro Thr Gln Thr Ser Val Arg Ala
1 5 10 15
Ala Leu Gly Pro Thr Ala Ala Ala Arg Asn
20 25
<210> 11
<211> 559
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутант PhaCp из P. aeruginosa PA01
<400> 11
Met Ser Gln Lys Asn Asn Asn Glu Leu Pro Lys Gln Ala Ala Glu Asn
1 5 10 15
Thr Leu Asn Leu Asn Pro Val Ile Gly Ile Arg Gly Lys Asp Leu Leu
20 25 30
Thr Ser Ala Arg Met Val Leu Leu Gln Ala Val Arg Gln Pro Leu His
35 40 45
Ser Ala Arg His Val Ala His Phe Ser Leu Glu Leu Lys Asn Val Leu
50 55 60
Leu Gly Gln Ser Glu Leu Arg Pro Gly Asp Asp Asp Arg Arg Phe Ser
65 70 75 80
Asp Pro Ala Trp Ser Gln Asn Pro Leu Tyr Lys Arg Tyr Met Gln Thr
85 90 95
Tyr Leu Ala Trp Arg Lys Glu Leu His Ser Trp Ile Ser His Ser Asp
100 105 110
Leu Ser Pro Gln Asp Ile Ser Arg Gly Gln Phe Val Ile Asn Leu Leu
115 120 125
Thr Asp Ala Met Ser Pro Thr Asn Ser Leu Ser Asn Pro Ala Ala Val
130 135 140
Lys Arg Phe Phe Glu Thr Gly Gly Lys Ser Leu Leu Asp Gly Leu Gly
145 150 155 160
His Leu Ala Lys Asp Leu Val Asn Asn Gly Gly Met Pro Ser Gln Val
165 170 175
Asp Met Asp Ala Phe Glu Val Gly Lys Asn Leu Ala Thr Thr Glu Gly
180 185 190
Ala Val Val Phe Arg Asn Asp Val Leu Glu Leu Ile Gln Tyr Arg Pro
195 200 205
Ile Thr Glu Ser Val His Glu Arg Pro Leu Leu Val Val Pro Pro Gln
210 215 220
Ile Asn Lys Phe Tyr Val Phe Asp Leu Ser Pro Asp Lys Ser Leu Ala
225 230 235 240
Arg Phe Cys Leu Arg Asn Gly Val Gln Thr Phe Ile Val Ser Trp Arg
245 250 255
Asn Pro Thr Lys Ser Gln Arg Glu Trp Gly Leu Thr Thr Tyr Ile Glu
260 265 270
Ala Leu Lys Glu Ala Ile Glu Val Val Leu Ser Ile Thr Gly Ser Lys
275 280 285
Asp Leu Asn Leu Leu Gly Ala Cys Ser Gly Gly Ile Thr Thr Ala Thr
290 295 300
Leu Val Gly His Tyr Val Ala Ser Gly Glu Lys Lys Val Asn Ala Phe
305 310 315 320
Thr Gln Leu Val Thr Val Leu Asp Phe Glu Leu Asn Thr Gln Val Ala
325 330 335
Leu Phe Ala Asp Glu Lys Thr Leu Glu Ala Ala Lys Arg Arg Ser Tyr
340 345 350
Gln Ser Gly Val Leu Glu Gly Lys Asp Met Ala Lys Val Phe Ala Trp
355 360 365
Met Arg Pro Asn Asp Leu Ile Trp Asn Tyr Trp Val Asn Asn Tyr Leu
370 375 380
Leu Gly Asn Gln Pro Pro Ala Phe Asp Ile Leu Tyr Trp Asn Asn Asp
385 390 395 400
Thr Thr Arg Leu Pro Ala Ala Leu His Gly Glu Phe Val Glu Leu Phe
405 410 415
Lys Ser Asn Pro Leu Asn Arg Pro Gly Ala Leu Glu Val Ser Gly Thr
420 425 430
Pro Ile Asp Leu Lys Gln Val Thr Cys Asp Phe Tyr Cys Val Ala Gly
435 440 445
Leu Asn Asp His Ile Thr Pro Trp Glu Ser Cys Tyr Lys Ser Ala Arg
450 455 460
Leu Leu Gly Gly Lys Cys Glu Phe Ile Leu Ser Asn Arg Gly His Ile
465 470 475 480
Met Ser Ile Leu Asn Pro Pro Gly Asn Pro Lys Ala Arg Phe Met Thr
485 490 495
Asn Pro Glu Leu Pro Ala Glu Pro Lys Ala Trp Leu Glu Gln Ala Gly
500 505 510
Lys His Ala Asp Ser Trp Trp Leu His Trp Gln Gln Trp Leu Ala Glu
515 520 525
Arg Ser Gly Lys Thr Arg Lys Ala Pro Ala Ser Leu Gly Asn Lys Thr
530 535 540
Tyr Pro Ala Gly Glu Ala Ala Pro Gly Thr Tyr Val His Glu Arg
545 550 555
<210> 12
<211> 1737
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ген YlDLD1
<400> 12
atgaacagaa tctcacgagc cgcccagagc gtctctcgaa ttgcctccag gcaattggct 60
cgaagtaact tctccacaca ggcaagatca tcccagccat ctatcagctg gggtgctgtt 120
gcaggagccg ctggtgtcgc tgcgggagtg acctacttca ttaccaagtc cgacaagaac 180
gctgtggccc ttaagaagga agatgaggat gttggccgag gaatccagaa ggtcctcgat 240
ggtgcctctc tcgacacgga cttctctcac aagcccaagt acggaggtga ggaggagttc 300
aagaaggccc tgccggaatt tatcaaggct atcggtgagg aatacgtttc taccgacgag 360
gaggatatcc aattccatgg ctggtccaac gtttcttctt ccaacctcga cactctgccc 420
tttggtgttc tgtaccccaa gtctactgag gaggtttctg ccattgccaa gatctgccac 480
aagcacaagc ttcccatggt cggctactcc ggaggtactt ctctggaggg ccacctttct 540
gccgcctacg gaggtgtctg catcgacttc tccaacatga acaaaattat tgctattaga 600
cccgatgata tggatgccac tgtccagccc tctgttggct gggttgatct gaacaacgag 660
attctcaagg agggccatcg cctgtttctg gctgttgatc ccggcccaac agcacaggtg 720
ggaggcatgg ttgccaactc ttgttctgga accaactgtg ttaagtacgg acccatgcga 780
gatcacgttg ttaatctcac tgttgtcctg gcggacggta ccattctcaa gacccggcag 840
cgacctcgaa agacctctgc tggatacaac ctcaaccatc tattcgctgg aaccgaggga 900
acacttggtc tgatcaccga aatcaccgtt aagctccagc ccatccccaa tgtcacctct 960
gtcgccgtcc aacagtttcc gaccgtccac gccgcttgca agactgctat ggatatcctc 1020
aaggagggta tccccatggc tgctcttgaa cttatggatg atcagcacat gatctgggtc 1080
aacaactccg gttacgccaa gagaaagtgg gaagagaagc ccgccctgtt catgaagttc 1140
gccggtgcct ccgaggaaac tgtcgccgag caggtgaagg ttgctaagga gaaggctgcc 1200
aagtacacag actctccctt gcatttcgga agagatcagg aggaacagga tgagctgtgg 1260
tctgcccgaa aaaacgcgct ttacctcgct cttgctgccg agaaggacgg tatgaaggca 1320
tggaccactg atgttgctgt tcccctttct cagcttcctg atattgtcat gaaggcaaag 1380
cagagcatca ctgatgctgg tcttcttgga ggagttctgg gccacattgg tgacggaaac 1440
taccatgcca tcatgcttta cactcccgag caggccgata ttgtcaccga cgtcgtccat 1500
aagatggtcg accagggtct ggctgccgag ggaacctgca ctggtgagca cggtgttgga 1560
ttcggaaaga tcgaggggct tcttcacgag gttggtcctg tctctctcaa cctgatgcga 1620
accatcaagc tgtctcttga tccccttgag ctacttaacc ccggcaagat cttcactgac 1680
gatgccattc agcagggtct caagactctg gataacaaca aggctggtaa gacttaa 1737
<210> 13
<211> 1149
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ген YlCYB21
<400> 13
atgcctatca cccagaagat tctgtcgctc agcgatctgg aggccgcggc catgccctac 60
gccgaaaagg cgcctcgaga ctactgggaa accggatcca acgatctgct gactctggca 120
gagaaccaaa acgccttcaa ctacctcaag atccgggccc gagccatgcg gggcgtcggc 180
accattgaca tctcgcccaa ggtggagctg tttggccgaa agttccgtgc ccccatcggt 240
gtggccccct cagcctacca ccagatggcg gatgattccg gcgagtgtgg cacagctgcg 300
gcttgtcagg cccgaaactg gcccatggga ctttcctctt tctccaacaa gcctctggag 360
gaagtccgag aggctggacc tgatgcagct ctcttcttcc agctctacgt gttcaagaac 420
aagaagacgt cagagaacct ggtcaagaag gccgaaaagg ccggcttcaa ggccattgcg 480
ctgaccgtcg acactcccta cctcggaaac cgatacgccg acgtgcgaaa caacttcaag 540
ctgccatcgc atctctctgc ccgaaacttc gagggcacca ccgaccagcc tattgacaac 600
gccgccgagg ccgactcgtg ggcccgaaag atcttcaacg gcgaggagtg tccccccgac 660
gccaacgtgg tcgaccccga tatcaactgg gccgaaacca tcccttggct gcgatccatc 720
accaacatgc agatctgggt caagggagtc gtgactgccg aggacaccca tgccgccatt 780
gaggccggcg tggacggaat ctgggtctcc aaccacggtg gccgacagct ggactccggt 840
ctggccacca ttgacgctct ccccgaggtg gtcgaggccg ctgctggccg agtccccatc 900
cacattgacg gaggcatccg acgaggagga gacgtgttca aatgccttgc tctgggcgcc 960
gatttcgtgt ggctgggccg acctgccatc tggggtctca agtacgacgg ccaggccggc 1020
gtggagctca tggagcagat catcgaggac gatcttaagc tcaccatggc tctggctggc 1080
accaagaccg tggccgagat caaccgaagc tgtctggttc gaatcggacc cgccatcgtc 1140
aagctttag 1149
<210> 14
<211> 1575
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ген, кодирующий цитоплазматический CpPCT opt V193A
<400> 14
atgcgaaagg ttcccatcat cactgctgac gaggctgcca agctcatcaa ggacggagat 60
accgttacta cttcgggttt tgtcggaaac gctatccctg aggctctgga ccgagctgtc 120
gagaagcgat tcctcgagac cggcgagcct aagaacatta cttacgttta ctgtggatct 180
cagggtaacc gagacggacg aggtgctgag cactttgccc atgagggcct gctcaagcga 240
tacattgctg gacactgggc caccgttccc gctctgggaa agatggccat ggagaacaag 300
atggaggctt acaacgtgtc ccagggagcc ctgtgccacc tcttccgaga catcgcctcg 360
cataagcccg gtgttttcac caaggtcggc atcggaactt ttattgaccc tcgaaacggc 420
ggcggcaagg tcaacgacat caccaaggaa gacattgttg agctggtgga gattaagggc 480
caggagtacc tcttctaccc cgcctttcct atccacgtgg ctctgattcg aggaacctac 540
gccgacgagt ccggtaacat cacttttgag aaggaagccg ctcccctcga gggaacctct 600
gtctgtcagg ctgttaagaa ctccggcgga attgtggtcg ttcaggtcga gcgagtggtc 660
aaggccggaa ctctggaccc ccgacatgtc aaggttcctg gtatctacgt ggattacgtt 720
gtggtcgctg accccgagga tcaccagcag tcgctggact gcgagtacga tcccgccctc 780
tctggcgagc atcgacgacc tgaggttgtg ggagagcccc tgcctctctc ggctaagaag 840
gtcatcggcc gacgaggagc cattgagctg gagaaggacg tggctgtcaa cctcggtgtg 900
ggagctcctg agtacgtggc ttctgttgct gacgaggaag gcatcgtcga tttcatgacc 960
ctgactgccg agtctggagc tattggtggc gtgcctgctg gaggtgtccg attcggtgcc 1020
tcctacaacg ccgacgctct gattgatcag ggctaccagt ttgactacta cgatggcgga 1080
ggtctggacc tctgttacct gggtctcgct gagtgcgatg agaagggcaa catcaacgtg 1140
tcccgattcg gtccccgaat tgccggctgt ggcggcttca tcaacattac ccagaacact 1200
cctaaggttt tcttttgcgg caccttcact gctggtggcc tgaaggtgaa gatcgaggac 1260
ggcaaggtca tcattgtcca ggaaggcaag cagaagaagt tcctgaaggc cgtcgagcag 1320
atcaccttta acggagacgt tgccctcgct aacaagcagc aggtgaccta cattactgag 1380
cgatgtgtct tcctgctcaa ggaagacggt ctgcacctct ctgagattgc tcctggcatt 1440
gatctgcaga cccagatcct cgacgtgatg gattttgctc ctatcattga ccgagatgcc 1500
aacggccaga ttaagctgat ggatgctgct ctgtttgctg agggtctgat gggcctgaag 1560
gagatgaagt cttaa 1575
<210> 15
<211> 1680
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ген, кодирующий PaPhaCp из P. aeruginosa с мутациями
E130D; S325T; S477R; Q481M
<400> 15
atgtctcaga agaacaacaa cgagctgccc aagcaggccg ccgagaacac cctgaacctg 60
aaccccgtga tcggcatccg aggcaaggac ctgctgacct ctgcccgaat ggtgctgctg 120
caggccgtgc gacagcccct gcactccgcc cgacacgtgg cccacttctc gctcgagctg 180
aagaacgtgc tgctgggcca gtctgagctg cgacccggcg acgacgaccg acgattctct 240
gaccccgcct ggtcccagaa ccccctgtac aagcgataca tgcagaccta cctggcctgg 300
cgaaaggagc tgcactcttg gatctctcac tctgacctgt ctccccagga catctctcga 360
ggccagttcg tgatcaacct gctcaccgac gctatgtctc ccaccaactc cctgtctaac 420
cccgctgccg tgaagcgatt cttcgagact ggcggcaagt ctctgctgga cggcctgggc 480
cacctggcta aggacctggt gaacaacggc ggaatgccct ctcaggtgga catggacgcc 540
ttcgaggtgg gcaagaacct ggccaccacc gagggcgccg tggtgttccg aaacgacgtc 600
ctcgagctga tccagtaccg acccatcacc gagtctgtgc acgagcgacc cctgctggtg 660
gtgccccccc agatcaacaa gttctacgtg ttcgacctgt cccccgacaa gtccctggcc 720
cgattctgcc tgcgaaacgg cgtgcagacc ttcatcgtgt cttggcgaaa ccccaccaag 780
tctcagcgag agtggggcct gaccacctac atcgaggccc tgaaggaggc catcgaggtc 840
gtcctgtcta tcaccggctc gaaggacctg aacctcctgg gcgcctgctc tggcggcatc 900
accaccgcca ccctggtggg ccactacgtg gcctctggcg agaagaaggt gaacgccttc 960
acccagctgg tgaccgtgct ggactttgag ctgaacaccc aggtcgccct gttcgccgac 1020
gagaagaccc tcgaggccgc caagcgacga tcttaccagt ctggcgtgct cgagggcaag 1080
gacatggcca aggtgttcgc ctggatgcga cccaacgacc tgatctggaa ctactgggtc 1140
aacaactacc tgctgggcaa ccagccccct gccttcgaca tcctgtactg gaacaacgac 1200
accacccgac tgcccgctgc cctgcacggc gagttcgtcg agctgttcaa gtccaacccc 1260
ctgaaccgac ccggtgctct cgaggtgtct ggcaccccca tcgacctgaa gcaggtcacc 1320
tgtgacttct actgcgtggc cggcctgaac gaccacatca ccccctggga gtcttgctac 1380
aagtctgccc gactgctggg cggaaagtgc gagttcatcc tgtccaaccg aggccacatc 1440
atgtctatcc tgaaccctcc cggcaacccc aaggctcgat tcatgaccaa ccccgagctg 1500
cctgccgagc ccaaggcctg gctcgagcag gctggcaagc acgccgactc ttggtggctg 1560
cactggcagc agtggctggc cgagcgatct ggcaagaccc gaaaggctcc cgcctctctg 1620
ggcaacaaga cctaccccgc tggcgaggcc gctcccggca cctacgtcca cgagcgataa 1680
<210> 16
<211> 656
<212> Белок
<213> Yarrowia lipolytica
<400> 16
Ser Glu Asp His Pro Ala Ile His Pro Pro Ser Glu Phe Lys Asp Asn
1 5 10 15
His Pro His Phe Gly Gly Pro His Leu Asp Cys Leu Gln Asp Tyr His
20 25 30
Gln Leu His Lys Glu Ser Ile Glu Asp Pro Lys Ala Phe Trp Lys Lys
35 40 45
Met Ala Asn Glu Leu Ile Ser Trp Ser Thr Pro Phe Glu Thr Val Arg
50 55 60
Ser Gly Gly Phe Glu His Gly Asp Val Ala Trp Phe Pro Glu Gly Gln
65 70 75 80
Leu Asn Ala Ser Tyr Asn Cys Val Asp Arg His Ala Phe Ala Asn Pro
85 90 95
Asp Lys Pro Ala Ile Ile Phe Glu Ala Asp Glu Pro Gly Gln Gly Arg
100 105 110
Ile Val Thr Tyr Gly Glu Leu Leu Arg Gln Val Ser Gln Val Ala Ala
115 120 125
Thr Leu Arg Ser Phe Gly Val Gln Lys Gly Asp Thr Val Ala Val Tyr
130 135 140
Leu Pro Met Ile Pro Glu Ala Ile Val Thr Leu Leu Ala Ile Thr Arg
145 150 155 160
Ile Gly Ala Val His Ser Val Ile Phe Ala Gly Phe Ser Ser Gly Ser
165 170 175
Leu Arg Asp Arg Ile Asn Asp Ala Lys Ser Lys Val Val Val Thr Thr
180 185 190
Asp Ala Ser Met Arg Gly Gly Lys Thr Ile Asp Thr Lys Lys Ile Val
195 200 205
Asp Glu Ala Leu Arg Asp Cys Pro Ser Val Thr His Thr Leu Val Phe
210 215 220
Arg Arg Ala Gly Val Glu Asn Leu Ala Trp Thr Glu Gly Arg Asp Phe
225 230 235 240
Trp Trp His Glu Glu Val Val Lys His Arg Pro Tyr Leu Ala Pro Val
245 250 255
Pro Val Ala Ser Glu Asp Pro Ile Phe Leu Leu Tyr Thr Ser Gly Ser
260 265 270
Thr Gly Thr Pro Lys Gly Leu Ala His Ala Thr Gly Gly Tyr Leu Leu
275 280 285
Gly Ala Ala Leu Thr Ala Lys Tyr Val Phe Asp Ile His Gly Asp Asp
290 295 300
Lys Leu Phe Thr Ala Gly Asp Val Gly Trp Ile Thr Gly His Thr Tyr
305 310 315 320
Val Leu Tyr Gly Pro Leu Met Leu Gly Ala Thr Thr Val Val Phe Glu
325 330 335
Gly Thr Pro Ala Tyr Pro Ser Phe Ser Arg Tyr Trp Asp Ile Val Asp
340 345 350
Asp His Lys Ile Thr His Phe Tyr Val Ala Pro Thr Ala Leu Arg Leu
355 360 365
Leu Lys Arg Ala Gly Thr His His Ile Lys His Asp Leu Ser Ser Leu
370 375 380
Arg Thr Leu Gly Ser Val Gly Glu Pro Ile Ala Pro Asp Val Trp Gln
385 390 395 400
Trp Tyr Asn Asp Asn Ile Gly Arg Gly Lys Ala His Ile Cys Asp Thr
405 410 415
Tyr Trp Gln Thr Glu Thr Gly Ser His Ile Ile Ala Pro Met Ala Gly
420 425 430
Val Thr Pro Thr Lys Pro Gly Ser Ala Ser Leu Pro Val Phe Gly Ile
435 440 445
Asp Pro Val Ile Ile Asp Pro Val Ser Gly Glu Glu Leu Lys Gly Asn
450 455 460
Asn Val Glu Gly Val Leu Ala Leu Arg Ser Pro Trp Pro Ser Met Ala
465 470 475 480
Arg Thr Val Trp Asn Thr His Glu Arg Tyr Met Glu Thr Tyr Leu Arg
485 490 495
Pro Tyr Pro Gly Tyr Tyr Phe Thr Gly Asp Gly Ala Ala Arg Asp Asn
500 505 510
Asp Gly Phe Tyr Trp Ile Arg Gly Arg Val Asp Asp Val Val Asn Val
515 520 525
Ser Gly His Arg Leu Ser Thr Ala Glu Ile Glu Ala Ala Leu Ile Glu
530 535 540
His Ala Gln Val Ser Glu Ser Ala Val Val Gly Val His Asp Asp Leu
545 550 555 560
Thr Gly Gln Ala Val Asn Ala Phe Val Ala Leu Lys Asn Pro Val Glu
565 570 575
Asp Val Asp Ala Leu Arg Lys Glu Leu Val Val Gln Val Arg Lys Thr
580 585 590
Ile Gly Pro Phe Ala Ala Pro Lys Asn Val Ile Ile Val Asp Asp Leu
595 600 605
Pro Lys Thr Arg Ser Gly Lys Ile Met Arg Arg Ile Leu Arg Lys Val
610 615 620
Leu Ala Gly Glu Glu Asp Gln Leu Gly Asp Ile Ser Thr Leu Ala Asn
625 630 635 640
Pro Asp Val Val Gln Thr Ile Ile Glu Val Val His Ser Leu Lys Lys
645 650 655
<210> 17
<211> 139
<212> Белок
<213> Pseudomonas putida
<400> 17
Met Ala Lys Val Thr Val Lys Lys Lys Asp Asp Ala Leu Gly Thr Leu
1 5 10 15
Gly Glu Val Arg Gly Tyr Ala Arg Lys Ile Trp Leu Ala Gly Ile Gly
20 25 30
Ala Tyr Ala Arg Val Gly Gln Glu Gly Ser Asp Tyr Phe Gln Glu Leu
35 40 45
Val Lys Ala Gly Glu Gly Val Glu Lys Arg Gly Lys Lys Arg Ile Asp
50 55 60
Lys Glu Leu Asp Ala Ala Asn Asn Gln Ile Asp Glu Ala Ala Glu Glu
65 70 75 80
Val Ser Arg Val Arg Gly Glu Val Glu Ile Gln Leu Asp Lys Ile Glu
85 90 95
Lys Ala Phe Asp Ala Arg Val Gly Arg Ala Leu Asn Arg Leu Gly Ile
100 105 110
Pro Ser Lys His Asp Val Glu Ala Leu Ser Ile Lys Leu Glu Gln Leu
115 120 125
His Glu Leu Leu Glu Arg Val Ala His Lys Pro
130 135
<210> 18
<211> 192
<212> Белок
<213> Искусственная последовательность
<220>
<223> phaP1p из Rastonia eutropha H16 (QOKBV4)
<400> 18
Met Ile Leu Thr Pro Glu Gln Val Ala Ala Ala Gln Lys Ala Asn Leu
1 5 10 15
Glu Thr Leu Phe Gly Leu Thr Thr Lys Ala Phe Glu Gly Val Glu Lys
20 25 30
Leu Val Glu Leu Asn Leu Gln Val Val Lys Thr Ser Phe Ala Glu Gly
35 40 45
Val Asp Asn Ala Lys Lys Ala Leu Ser Ala Lys Asp Ala Gln Glu Leu
50 55 60
Leu Ala Ile Gln Ala Ala Ala Val Gln Pro Val Ala Glu Lys Thr Leu
65 70 75 80
Ala Tyr Thr Arg His Leu Tyr Glu Ile Ala Ser Glu Thr Gln Ser Glu
85 90 95
Phe Thr Lys Val Ala Glu Ala Gln Leu Ala Glu Gly Ser Lys Asn Val
100 105 110
Gln Ala Leu Val Glu Asn Leu Ala Lys Asn Ala Pro Ala Gly Ser Glu
115 120 125
Ser Thr Val Ala Ile Val Lys Ser Ala Ile Ser Ala Ala Asn Asn Ala
130 135 140
Tyr Glu Ser Val Gln Lys Ala Thr Lys Gln Ala Val Glu Ile Ala Glu
145 150 155 160
Thr Asn Phe Gln Ala Ala Ala Thr Ala Ala Thr Lys Ala Ala Gln Gln
165 170 175
Ala Ser Ala Thr Ala Arg Thr Ala Thr Ala Lys Lys Thr Thr Ala Ala
180 185 190
<---
Реферат
Группа изобретений относится к рекомбинантной клетке дрожжей, подходящей для получения полимолочной кислоты (PLA), и ее применению. Предложена рекомбинантная клетка дрожжей, подходящая для получения PLA, содержащая и экспрессирующая ген, кодирующий лактил-КоА-синтазу, и ген, кодирующий синтазу полигидроксиалканоата (PHA) (PhaCp), дополнительно содержащую пероксисомальную направленную последовательность (PTS). При этом лактил-КоА-синтазу выбирают из группы, состоящей из КоА-трансферазы, пропионил-КоА-трансферазы (Pctp) или КоА-лигазы. Также в клетке инактивируют по меньшей мере одну активность оксидоредуктазы молочной кислоты, предпочтительно инактивируют ген, кодирующий дегидрогеназу молочной кислоты, более предпочтительно ген, кодирующий дегидрогеназу D-молочной кислоты. Рекомбинантная клетка дрожжей сконструирована для экспрессии лактил-КоА-синтазы в цитозоле и для экспрессии PHA-синтазы в пероксисоме указанной клетки. Предложены также способ получения PLA с использованием указанной клетки, ферментативный бульон для получения PLA, содержащий указанную клетку. Группа изобретений обеспечивает продуцирование высоких количеств PLA. 4 н. и 9 з.п. ф-лы, 10 ил., 5 табл., 4 пр.
Комментарии