Способ получения человеческого иммунного интерферона - RU2107728C1
Код документа: RU2107728C1
Чертежи

Описание
Изобретение относится к технологии рекомбинантных ДНК, к средствам и способам использования такой технологии при раскрытии последовательности ДНК и определяемой ею последовательности аминокислот для иммунного интерферона человека, его получения, а также к полученным при этом различным продуктам и их использованию.
Более конкретно настоящее изобретение относится к выделению и идентификации последовательностей ДНК, кодирующих иммунный интерферон человека и к построению рекомбинантного вектора экспрессии, содержащего эти последовательности ДНК, оперативно связанные с последовательностями промотора, и к осуществлению экспрессии с полученными таким образом векторами. В другом аспекте настоящее изобретение относится к системам культуры хозяина, таким как различные микроорганизмы и культуры клеток позвоночных, трансформированные векторами экспрессии и таким образом направленные на экспрессию указанных ранее последовательностей ДНК. Еще в одном аспекте настоящее изобретение относится к средствам и способам превращения конечных продуктов такой экспрессии в новые объекты, такие как фармацевтические композиции, пригодные для лечения и профилактики людей.
В предпочтительных вариантах настоящее изобретение предлагает конкретные векторы экспрессии, которые имеют такие последовательности, что иммунный интерферон человека продуцируется и выделяется из клеток хозяина в зрелом виде. Кроме того, настоящее изобретение относится к различным процессам, используемым для получения таких последовательностей ДНК, векторов экспрессии, систем культур хозяина и конечных продуктов и их производных, а также к конкретным и связанным с ними условиям.
Настоящее изобретение частично вытекает из открытия последовательности ДНК и установленной аминокислотной последовательности, кодирующей иммунный интерферон человека. Кроме того, настоящее изобретение предоставляет информацию о последовательности 3'- и 5' - боковых последовательностей гена иммунного интерферона человека, что облегчает связывание его in vitro в векторы экспрессии. В частности, установлен 5'-ДНК сегмент, кодирующий предлагаемый эндогенный сигнальный полипептид, который непосредственно предшествует аминокислотной последовательности предполагаемого зрелого иммунного интерферона человека. Эти открытия облегчает разработку средств и способов получения с помощью рекомбинантной технологии ДНК достаточного количества иммунного интерферона человека, что обеспечило, в свою очередь, возможность определения его биохимических свойств и биоактивности.
Публикации и другие материалы, использованные для освещения предпосылок изобретения, а также в ряде случаев для освещения дополнительных подробностей, касающихся его применения, включены посредством ссылок, для удобства пронумерованы в нижеследующем тексте и соответствующим образом расположены в прилагаемом списке библиографии.
Предпосылки изобретения
A. Иммунный интерферон человека
Интерфероны человека можно классифицировать на три группы в зависимости от
различной антигенности и биологических и биохимических свойств.
Первую группу составляет семейство лейкоцитарных интерферонов ( α -интерферон, Le IF или IFN-α ), которые обычно вырабатываются, в основном, соответствующими клетками крови человека под действием вирусов. Эти интерфероны были получены микробиологическим способом и была обнаружена их биологическая активность (1, 2, 3). Их биологические свойства определили их использование в клиниках в качестве терапевтических агентов при лечении вирусных инфекций и злокачественных состояний (4).
Во второй группе находится фибробластный интерферон человека ( β- интерферон, FIF или IFN-β ), вырабатываемый обычно фибробластами под действием вирусов, который также был получен микробиологическим способом, и было обнаружено, что он демонстрирует широкий спектр биологических активностей (5). Клинические испытания также указывают на его потенциальную терапевтическую ценность. Лейкоцитарные и фибробластные интерфероны имеют очень заметное сходство в их биологических свойствах, несмотря на тот факт, что степень гомологии на аминокислотном уровне относительно низка. Кроме того, обе группы интерферонов содержат от 165 до 166 аминокислот и являются кислотными стабильными белками.
Иммунный интерферон человека ( γ- -интерферон, IIF или IFN-γ ), который составляет предмет настоящего изобретения в противоположность α- и β - интерферонам, pH 2 лабилен, его получают главным образом митогенной индукцией лимфоцитов и он также совершенно отличен по антигенным свойствам. До недавнего времени иммунный интерферон человека можно быть получать лишь в очень незначительных количествах, что, естественно, затрудняло его характеристику. Недавно было сообщено о гораздо более высокой, но все еще частичной очистке иммунного интерферона человека (6). Как сообщалось, это соединение было получено из культур лимфоцитов, стимулированных сочетанием фитогемагглютинина и сложного эфира форбола, и было очищено при помощи последовательных хроматографических разделений. В результате этой процедуры получили продукт с молекулярной массой 58000.
Иммунный интерферон человека получали в очень малых количествах трансляций мРНК в осциты, проявляющие характеристики интерферонной активности иммунного интерферона человека, причем выражалась надежда, что кДНК иммунного интерферона можно будет синтезировать и клонировать (7).
Полученное до сих пор количество иммунного интерферона явно недостаточно для проведения не вызывающих сомнений экспериментов по характеристике и определению биологических свойств очищенной компоненты. Однако при исследованиях in vitro, проведенных с неочищенными препаратами, также как и in vivo - экспериментах с препаратами γ- интерферона крыс, предполагалось, что основной функцией иммунного интерферона может быть функция иммунорегулирующего агента (8 и 9). Иммунный интерферон обладает не только противовирусной и противоклеточной активностью, общей для всех интерферонов человека, но и потенцирующим действием на эти активности α- и β -интерферонов (10). Кроме того, антипролиферативное действие γ -интерферона на опухолевые клетки in vitro, как сообщается, приблизительно в 10-100 раз выше нежели действие других классов интерферонов (8, 11, 12). Этот результат вместе с его выраженной иммунорегуляторной ролью (8 и 9) предлагает гораздо более выраженную противоопухолевую способность для IFN-γ, чем для ΙFN-α и IFN-β . Действительно в экспериментах in vivo с мышами и крысами IFN-γ препараты демонстрируют заметное превосходство по сравнению со стимулированными против вируса интерферонами в плане противоопухолевой активности против остеогенной саркомы (13).
Все эти исследования до настоящего изобретения приходилось выполнять на существенно загрязненных препаратах из-за их чрезвычайно малой доступности. Однако они однозначно подтверждали очень важные биологические функции иммунного интерферона. Иммунный интерферон обладает не только основной противовирусной активностью, но также, вероятно, сильной иммунорегуляторной и противоопухолевой активностью, что явно определяет его как потенциально многообещающий клинический объект.
Было явно, что применение технологии рекомбинантной ДНК должно быть наиболее эффективным путем получения необходимых больших количеств иммунного интерферона человека. Независимо от того, будут ли полученные таким образом вещества включать гликозилирование, которое рассматривают как характеристику природного, полученного от человека материала, они будут, по-видимому, демонстрировать биоактивность, определяющих их клиническое применение при лечении широкого ряда вирусных заболеваний, новообразований и состояний с подавленным иммунитетом.
B.
Технология рекомбинантной ДНК
Технология рекомбинантной ДНК достигла зрелости.
Молекулярные биологи способны достаточно легко рекомбинировать различные последовательности ДНК, создавая новые виды ДНК, способные продуцировать значительные количества экзогенных белковых продуктов в трансформированных микробах. В основном средства и способы для in vitro связывания различных фрагментов ДНК с тупыми или "липкими" концами разработаны, с их помощью получают потенциальные векторы экспрессии, пригодные для трансформации конкретных микроорганизмов, за счет чего регулируют направленный синтез нужных экзогенных продуктов. Однако для отдельных продуктов этот путь остается достаточно трудным, и наука не достигла еще той стадии, на которой можно гарантировать успех.
Плазмида, нехромосомная петля двунитевой ДНК, найденная в бактериях и других микробах, и часто в виде множества копий на клетку остается основным элементом рекомбинантной ДНК технологии. В информацию, закодированную в ДНК плазмиды, включена информация, необходимая для репродуцирования плазмиды в дочерних клетках (то есть, источник репликации) и обычно одна или более фенотипических характеристик селекции, таких как устойчивость к антибиотикам для бактерий, которая позволяет клонам клетки хозяина, содержащим интересующую плазмиду, быть узнанными и предпочтительно расти на селективной среде. Польза плазмид состоит в том факте, что их можно специфически расщеплять той или другой рестикционной эндонуклеазой или "рестрикционным ферментом", каждый из которых узнает различные участки ДНК плазмиды. После этого гетерологичные гены или генные фрагменты можно включать в плазмиду путем присоединения концами к участку расщепления или к реконструированным концам, прилежащим к участку расщепления. Так получают так называемые репликабельные векторы экспрессии. Рекомбинацию ДНК осуществляют вне клетки, однако получаемый "рекомбинантный" репликабельный вектор экспрессии или плазмиду можно ввести в клетки способом, известным, как трансформация, с получением больших количеств рекомбинантных векторов в результате роста трансформанта. Более того, если ген соответствующим образом ориентирован по отношению к участку плазмиды, который управляет транскрипцией и трансляцией кодирующей ДНК, полученный вектор экспрессии можно использовать для реального получения полипептидной последовательности, кодируемой встроенным геном, т.е. для процесса, который носит название экспрессии.
Экспрессию инициируют в области, известной как промотор, который распознается и связывается РНК полимеразой. На транскрипционной фазе экспрессии ДНК раскручивается, открывая его как химическую матрицу для инициированного синтеза информационной РНК с последовательности ДНК. Информационная РНК, в свою очередь, транслируется в полипептид с последовательностью аминокислот, закодированной мРНК. Каждая аминокислота закодирована нуклеотидным триплетом или "кодоном", которые все вместе составляют "структурный ген", то есть ту часть, которая кодирует аминокислотную последовательность экспрессированного полипептидного продукта. Трансляции инициируется "старт" - сигналом (обычно ATG, который в полученной информационной РНК становится AUG). Так называемые "стоп" - кодоны определяют конец трансляции и соответственно присоединения следующих аминокислотных единиц. Целевой продукт можно получить, лизируя в случае необходимости клетку хозяина и отделяя продукт путем соответствующих методов очистки от остальных белков.
На практике применение технологии рекомбинантной ДНК может обеспечить экспрессию полностью гетерологичных полипептидов (так называемая прямая экспрессия) или в другом варианте - гетерологичных полипептидов, присоединенных к участку аминокислотной последовательности гомологичного полипептида. В последнем случае целевой биоактивный продукт иногда остается неактивным в слитом гомологично-гетерологичном полипептиде до тех пор, пока он не будет отщеплен во внеклеточное окружение (см. опубликованный патент Великобритании N 2007676A и American Scientist 68, 664 (1980).
C. Технология клеточной культуры.
Искусство культур клеток или тканей для изучения генетики и физиологии клеток хорошо разработано. Известны устройства и способы поддержания перманентных линий клеток, полученных последовательной серией переносов из изомированных нормальных клеток. Для применения в исследованиях такие клеточные линии поддерживают на твердых подложках в жидкой среде или выращивают в суспензии, содержащей поддерживающие питательные вещества. Получение более крупных партий препаратов сводится лишь к механическим проблемам. Более подробное описание предпосылок изобретения можно найти в Microbiology and Edition, Harpes and Row Reblishers, Inc., Hagerstown, Maryland (1973), особенно на стр. 1122, и далее Scientific American 245, 66 и далее (1981), каждая из которых включена здесь в качестве ссылки.
Настоящее изобретение основано на открытии, что технологию рекомбинантной ДНК можно с успехом использовать для получения иммунного интерферона человека, предпочтительно в непосредственной форме и в количествах, достаточных для инициирования и проведения тестов на животных и в клиниках, что необходимо перед выходом на рынок. Этот продукт пригоден для использования во всех его формах для профилактики и лечения вирусных инфекций, злокачественных новообразований и состояний с подавленной или дефектной иммунной системой. Его варианты включают различные олигомерные формы, которые могут включать гликозилирование. Этот продукт получают в перестроенных генетически трансформированных микроорганизмах или в трансформированных системах клеточных культур.
В используемом здесь контексте термин "клетка-трансформант" относится к клетке, в которую ведена ДНК, причем указанная ДНК является продуктом экзогенной ДНК-рекомбинации, и к потомству любой такой клетки, которое сохраняет введенную таким путем ДНК.
Так, теперь стало возможно получать и выделять иммунный интерферон человека более эффективно, чем было возможно ранее. Одним из существенных факторов настоящего изобретения в его наиболее предпочтительном варианте является осуществление возможности генетически направить микроорганизм или клеточную культуру на продуцирование иммунного интерферона человека в достаточных для выделения количествах, секретированных клетках хозяина в зрелой форме.
Настоящее изобретение включает полученный таким образом иммунный интерферон человека, средства и способы его получения. Далее настоящее изобретение направлено на способные к репликации векторы экспрессии ДНК, хранящие последовательности генов, кодирующих иммунный интерферон человека в доступной для экспрессии форме.
Настоящее изобретение направлено также на штаммы микроорганизмов или клеточные культуры, трансформированные векторами экспрессии, описанными ранее, и на микробные или клеточные культуры таких трансформированных штаммов или культур, способные продуцировать иммунный интерферон человека. Еще в одном аспекте настоящее изобретение направлено на различные процессы, пригодные для получения указанных последовательностей гена иммунного интерферона, векторов экспрессии ДНК, штаммов микроорганизмов и клеточных культур и на их конкретные варианты. Кроме того, настоящее изобретение направлено на получение ферментационных культур указанных микроорганизмов и клеточных культур.
Настоящее изобретение направлено также на получение иммунного интерферона человека как продукта прямой экспрессии секретированного клетками хозяина в зрелой форме. Это достижение может использовать ген, кодирующий последовательность зрелого иммунного интерферона человека, плюс 5'-фланкирующую ДНК, кодирующую сигнальный полипептид. Считают, что сигнальный полипептид служит для транспорта молекулы к стенке клетки организма хозяина, где он отщепляется во время процесса секреции зрелого человека. Этот вариант делает возможным выделение и очистку целевого зрелого иммунного интерферона, не обращаясь к включению процедуры, предназначенной для удаления примесей внутриклеточного белка хозяина или клеточных осколков.
Встречающееся в тексте выражение "зрелый иммунный интерферон человека" означает продукт микробной или клеточной культуры иммунного интерферона человека, не содержащий сигнального пептида или последовательности препептида, который обязательно сопровождает трансляцию мРНК иммунного интерферона человека.
Первый рекомбинантный иммунный интерферон человека, полученный в соответствии с настоящим изобретением, имеет метионин в качестве своей первой аминокислоты (представлен в результате включения кодона стартового сигнала ATG перед структурным геном) или в том случае, если метионин внутри - или внеклеточно отщеплен, имеет в качестве нормальной первой аминокислоты цистеин. Зрелый иммунный интерферон человека можно также получить в виде конъюгата с белком, отличным от обычного сигнального полипептида, причем конъюгата, который может быть специфически расщеплен внутри или вне клетки (см. публикацию патента Великобритании N 2007676A). И, наконец, зрелый иммунный интерферон человека можно получить прямой экспрессией без необходимости отщепления каких-либо посторонних излишних полипептидов. Это особенно важно в тех случаях, когда данный хозяин не способен удалять или удаляет недостаточно эффективно сигнальный пептид, а вектор экспрессии предназначен экспрессировать зрелый интерферон человека вместе с его сигнальным пептидом. Полученный таким образом зрелый иммунный интерферон человека выделяют и очищают до уровня, удовлетворяющего требованиям, необходимым для применения при лечении вирусных заболеваний, злокачественных новообразований и состояний с подавленным или недостаточным иммунитетом.
Иммунный интерферон человека был получен следующим образом.
1. Ткани человека, например, ткань селезенки человека или периферические лимфоциты крови культивировали с митогенами для стимулирования продукции иммунного интерферона.
2. Осадок клеток из такой клеточной культуры экстрагировали в присутствии ингибитора рибонуклеазы с целью всей цитоплазмической РНК.
3. На олиго-dТ колонке выделили тотальную информационную РНК (мРНК) в полиаденилированной форме. Эту РНК расфракционировали по размерам, используя градиент плотности сахарозы и гель электрофорез в присутствии кислоты-мочевины.
4. Соответствующую РНК (от 12S до 18S) превратили в соответствующую однонитевую комплементарную ДНК (мДНК), из которой получили двунитевую кДНК. После поли - dC удлинения ее включили в вектор так, чтобы плазмида имела один или более фенотипических маркеров.
5. Полученные таким образом векторы использовали для трансформации бактериальных клеток с получением библиотеки колоний. Меченные радиоизотопами кДНК, полученные как из индуцированных, так и из неиндуцированных РНК, выделенных как описано ранее, использовали для раздельного определения дубликатных библиотек колоний. Затем избыток кДНК удаляли и колонии экспонировали на рентгеновской пленке для идентификации индуцированных клонов кДНК.
6. Из индуцированных клонов кДНК выделили соответствующую плазмидную ДНК и определили в ней последовательность оснований.
7. Секвенированную ДНК подготовили in vitro для включения в соответствующий вектор экспрессии, который использовали для трансформации подходящей клетки хозяина, который, в свою очередь, дали возможность расти в культуре и экспрессировать целевой иммунный интерферон человека.
8. В некоторых системах клеток хозяина, будучи включена в вектор экспрессии так, чтобы быть экспрессированной вместе с сигнальным пептидом, зрелая форма иммунного интерферона человека секретируется в среду клеточной культуры, что облегчает выделение и методы очистки.
Описание предпочтительных вариантов изобретения.
A. Система клеточных культур/векторы клеточных культур.
Размножение клеток позвоночных в культуре (культура ткани) стало обычной процедурой за последние годы (см. Tissue Culture Academic Riess Kruse and Paterson ends, 1973). В данном случае использовали COS - 7 линию фибробластов почки обезьян в качестве хозяина для получения иммунного интерферона (25a). Однако подробно описанные здесь опыты можно проводить на любой линии клеток, которая способна к репликации и экспрессии совместимого вектора, например, W138, BHK, 3T3, CHO, VERO и линий клеток HeLa. Кроме того, необходимо, чтобы вектор экспрессии имел сайт инициации репликации и промотор, расположенный перед геном, подлежащим экспрессии, вкупе с необходимыми также участками связывания рибосомы, участками сплайсинга РНК, участками полиаденилирования и транскрипционными терминаторами. Хотя здесь были использованы эти важные элементы SV40, следует иметь в виду, что изобретение, хотя оно и описано здесь с точки зрения его предпочтительного варианта, не следует рассматривать как ограниченное лишь этими последовательностями. Так, например, могут быть использованы источники репликации других вирусных (например, Polyoma, Adeno, VSV, BPV и т.д.) векторов, а также клеточные источники репликации ДНК, которые могут функционировать в неинтегрированном состоянии.
B. Экспрессия в культуре клеток млекопитающих.
Стратегия синтеза иммунного интерферона в культуре клеток млекопитающих основана на разработке вектора, способного как к автономной репликации, так и к экспрессии чужого гена под контролем гетерологичного транскрипционного фрагмента. Репликация этого вектора в культуре ткани обеспечивалась за счет стимуляции инициатора репликации ДНК (происходящего из вируса SV 40) и стимуляции вспомогательной функции (T антиген) путем введения вектора в линию клеток, эндогенно экспрессирующих этот антиген (23 и 29). Поздний промотор вируса SV 40 предшествует структурному гену интерферона и обеспечивает транскрипцию гена.
Вектор, который использовали для получения экспрессии γ , состоял из последовательностей pBR 322, которая обеспечивает маркер, пригодный для отбора в E. coli (устойчивой к ампициллину), а также инициатор репликации ДНК. Эти последовательности были получены из плазмиды pML - 1 (28) и представляют область, содержащую ECo RI и Bam HI рестрикционные сайты. SV 40 инициатор получен в составе фрагмента PVu II - Hind III размером 342 п.о., включающего эту область (30 и 31) (причем оба конца превращены в концы Eco RI). Эти последовательности, кроме того, что содержат вирусный инициатор репликации ДНК, кодируют промотор как для ранней, так и для поздней транскрипционной единицы, ориентация участка инициации из SV - 40 была такова, что промотор для поздней единицы транскрипции был расположен проксимально по соотношению к гену, кодирующему интерферон.
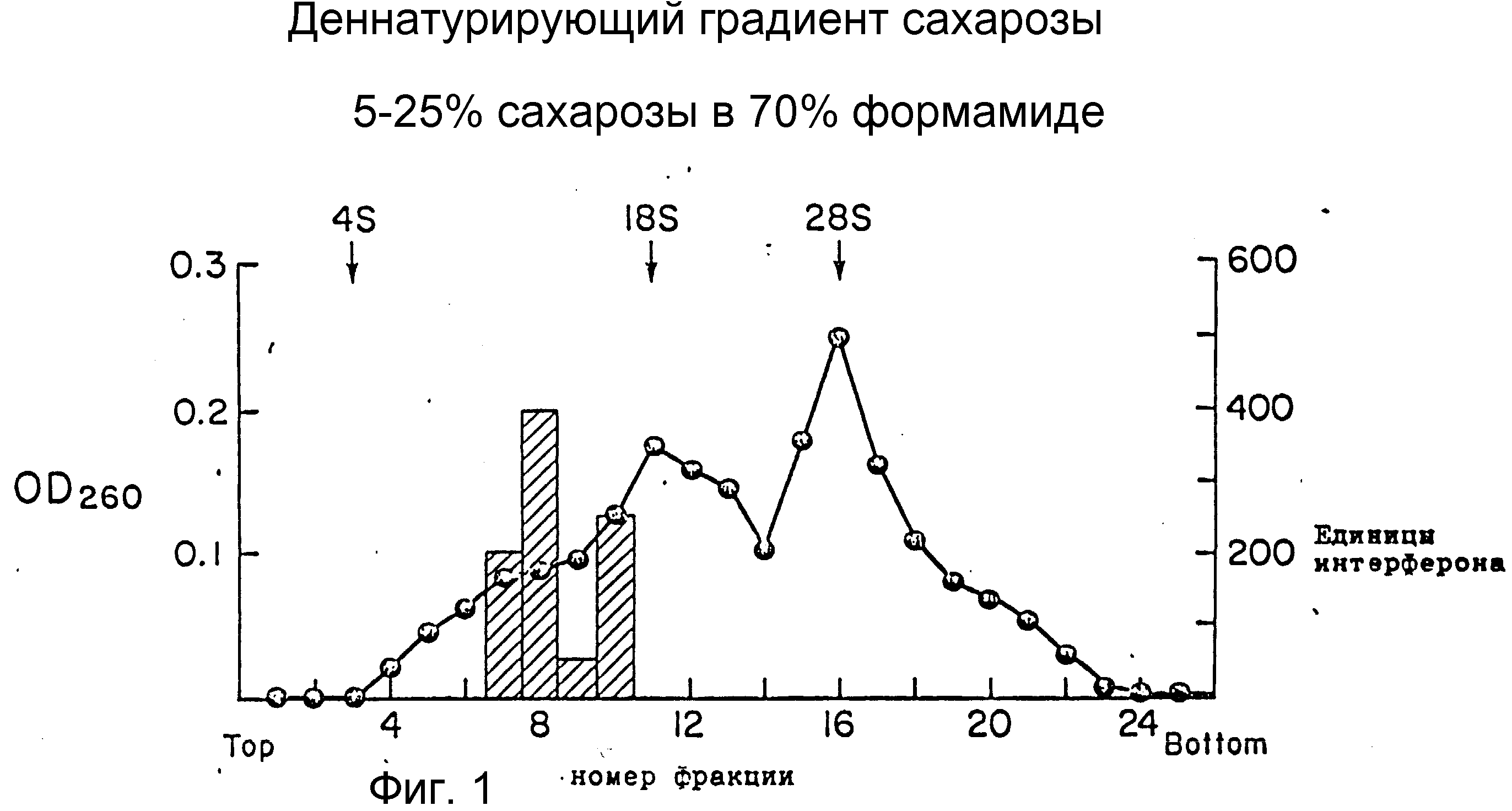
На фиг. 1 изображено градиентное сахарозное центрифугирование поли(А)+ РНК стимулированных лимфоцитов периферической крови. Наблюдается два максимума активности интерферона (показано заштрихованными прямоугольниками) с размерами 12S и 16S. Расположение маркеров рРНК (центрифугированных независимо) помечено над контуром поглощения.
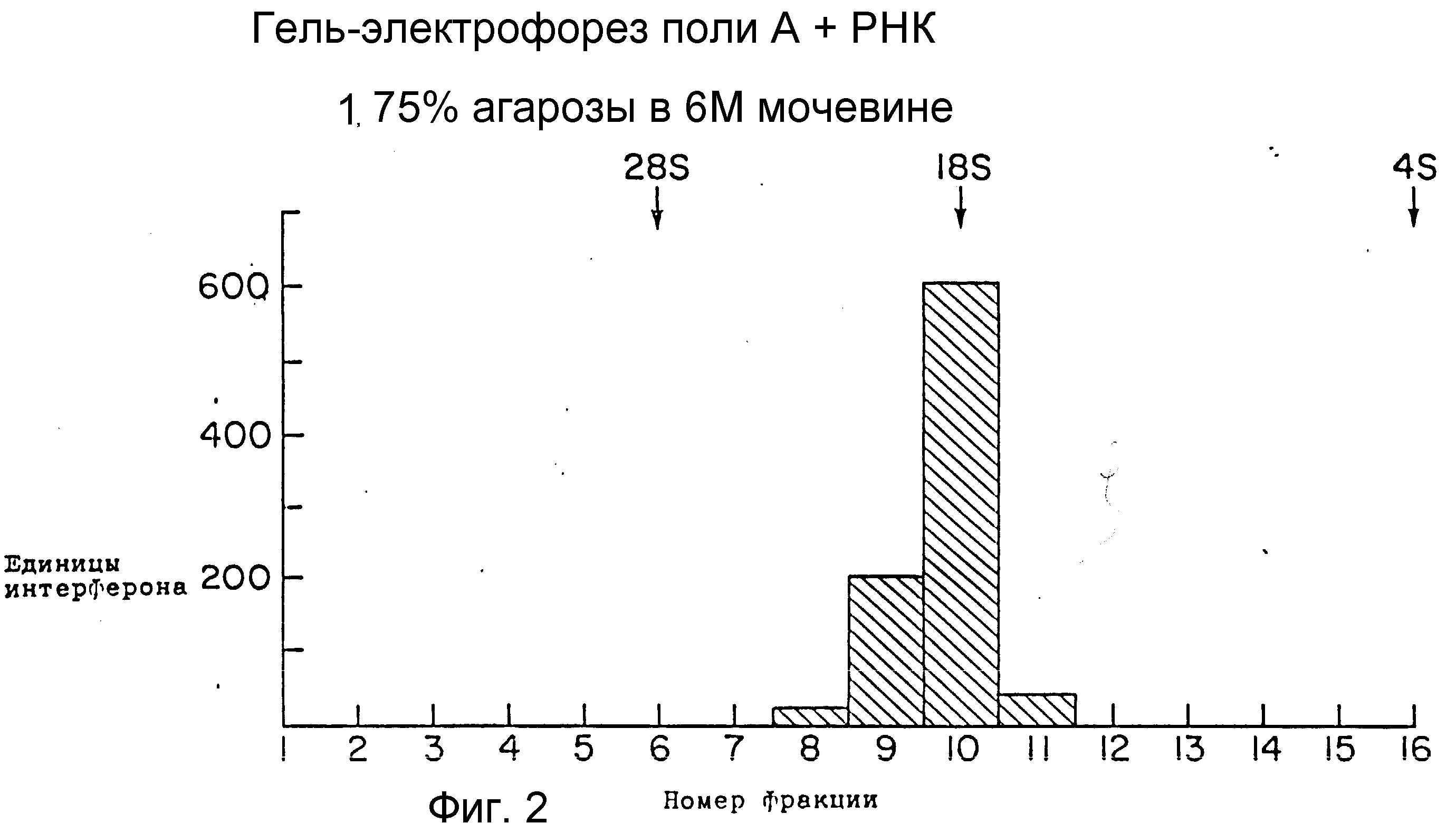
На фиг. 2 изображен электрофорез поли (A)+ РНК стимулированных PBL на кислота-мочевина-агарозе. Наблюдается только один максимум активности, который мигрирует совместно с 18S РНК. Положение рибосомных маркеров РНК, которые были подвергнуты электрофорезу на прилежащей дорожке и проявлены окрашиванием этидиум бромидом, помечено выше контура активности.
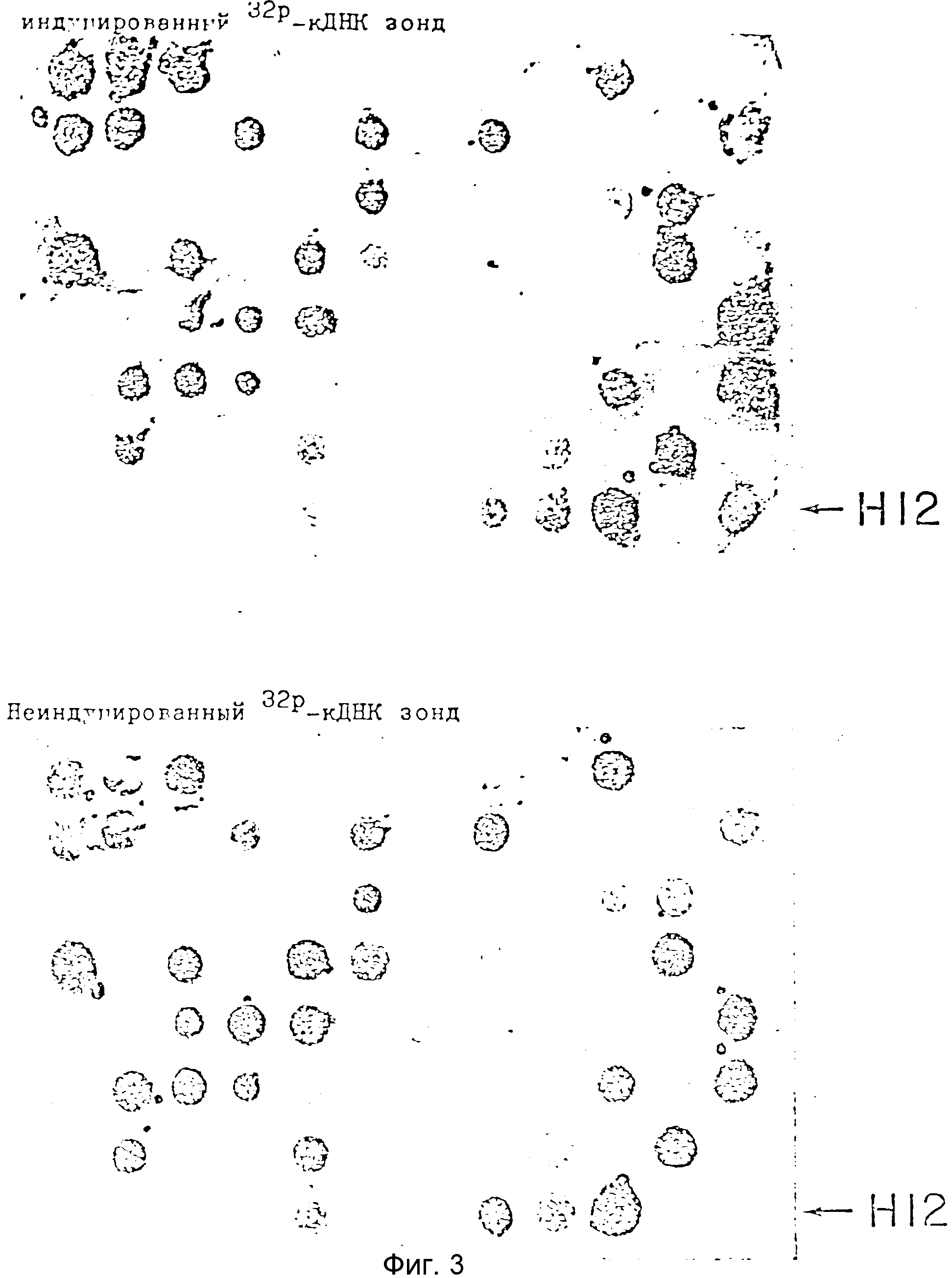
На фиг. 3 изображены картины гибридизации 96 колонии с индуцированными и неиндуцированными32P меченными кДНК пробами. 96 индивидуальных трансформантов выращивали на пластине для микротитрования, реплика была помещена на две нитроцеллюлозные мембраны, а затем фильтры гибридизовали с32P-кДНК образцами, полученными либо из индуцированных мРНК (вверху), либо из мРНК, выделенных из неиндуцированных культур pBL (неиндуцированные, внизу ). Фильтры промывали для удаления негибридизованных РНК, а затем экспонировали на пленке для рентгеновских лучей. Эта серия фильтров представляет 86 таких серий (8300 независимых колоний). Примером индуцированного клона является помеченный H12.
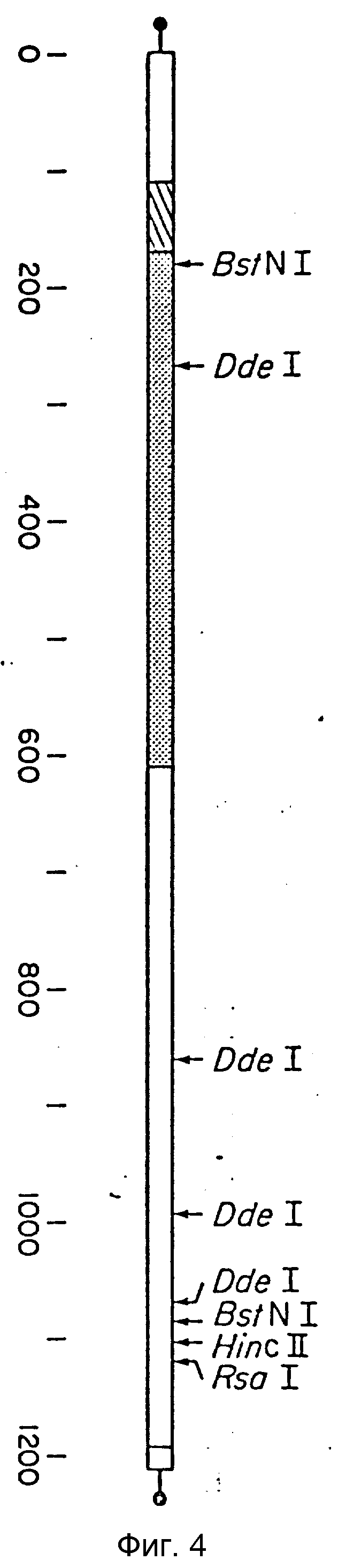
Фиг. 4 является рестрикционной картой вставки кДНК клона 69. Вставка кДНК связана с сайтами PstI (точки с обоих концов) и олиго-dC-dG "хвостами" (прямые линии). Число и размеры фрагментов, полученных расщеплением рестрикционными нуклеазами, были установлены с помощью электрофореза 6% акриламидном геле. Положения участков были подтверждены секвенированием (представлено на фиг. 5). Кодирующий участок самой большой открытой рамки считывания обведен, заштрихованный участок представляет 20 остатков последовательности сигнального пептида, тогда как участок с точечным пунктиром представляет последовательность зрелого IEN (46 аминокислот); 5' - конец мРНК находится слева, а 3' конец находится справа.
На фиг. 5 представлена нуклеотидная последовательность вставки кДНК плазмиды р69. Представлена также "выведенная" последовательность аминокислот наиболее длинного открытого участка считывания. Предполагаемая сигнальная последовательность представлена остатками, помеченными от S1 до S20.
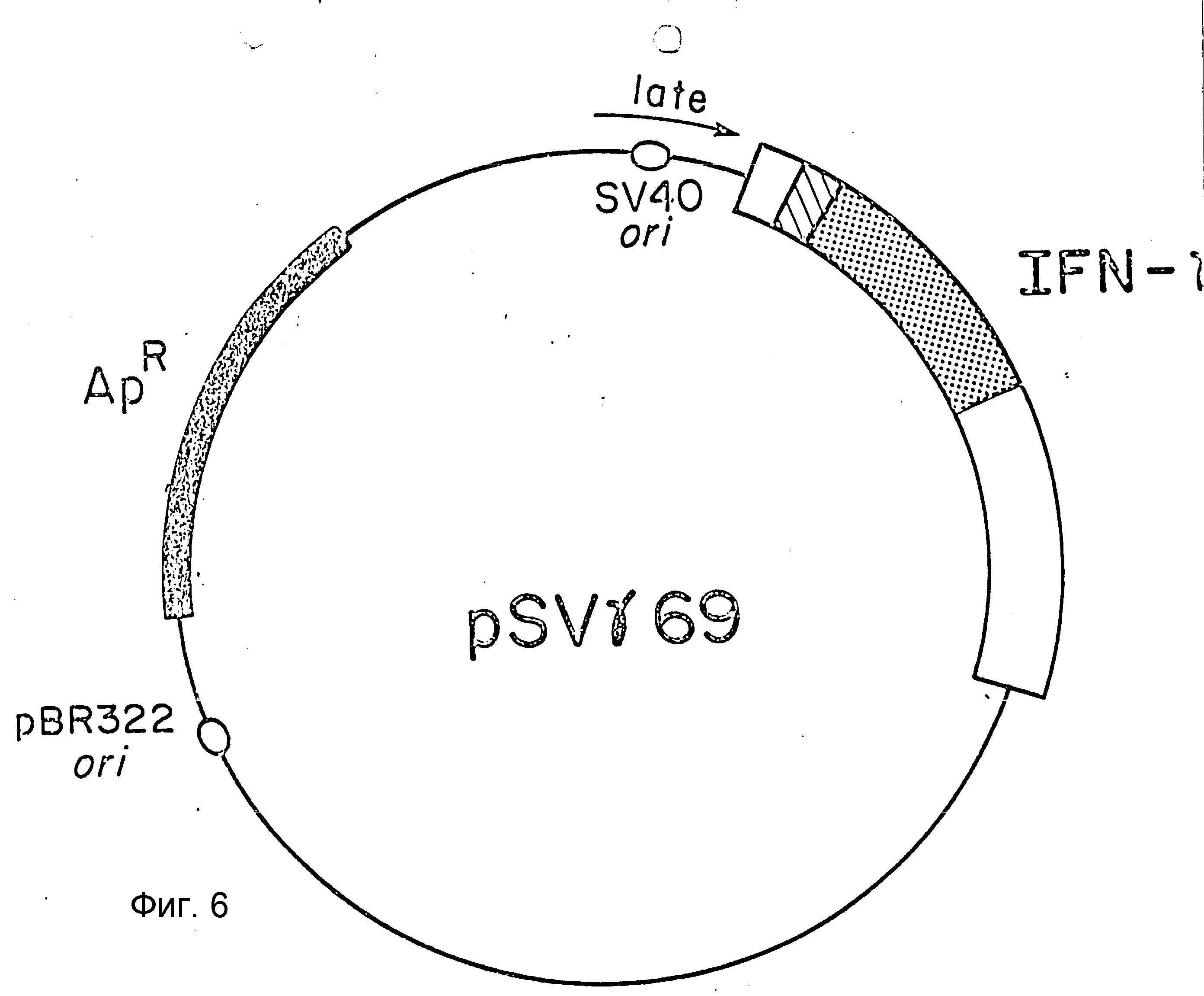
На фиг. 6 приведена схема плазмиды pSVγ69, использованной для экспрессии IFN-γ в клетках обезьяны.
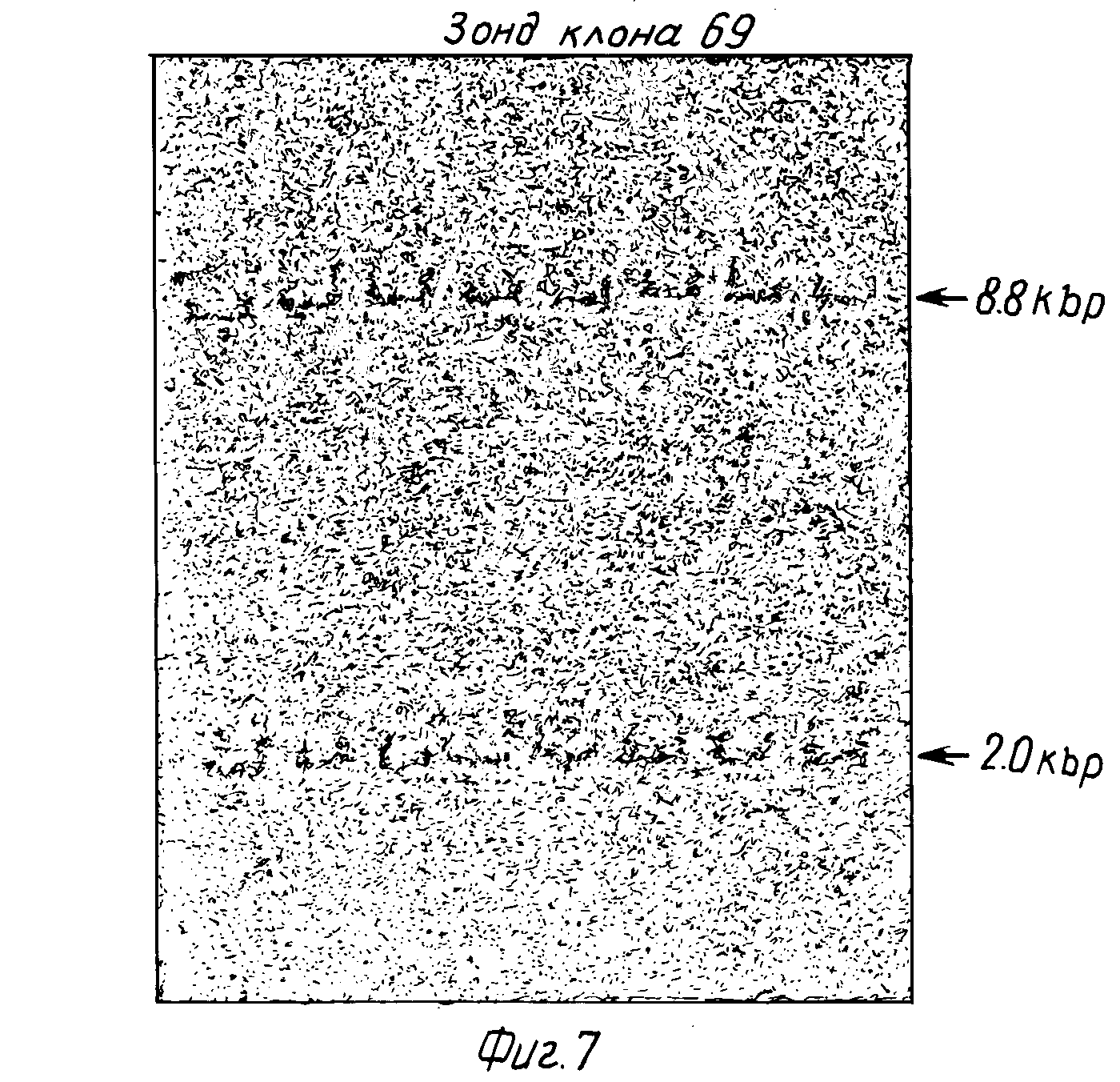
Фиг. 7 изображает Саузерн-гибридизацию 8 различных переваренных Eco RI человеческих геномных ДНК, гибридизованных с32P-меченным DdeI- фрагментом 600 п.о. из кДНК-вставки p69. Два Eco RI-фрагмента явно гибридизованы с зондом в каждом образце ДНК.
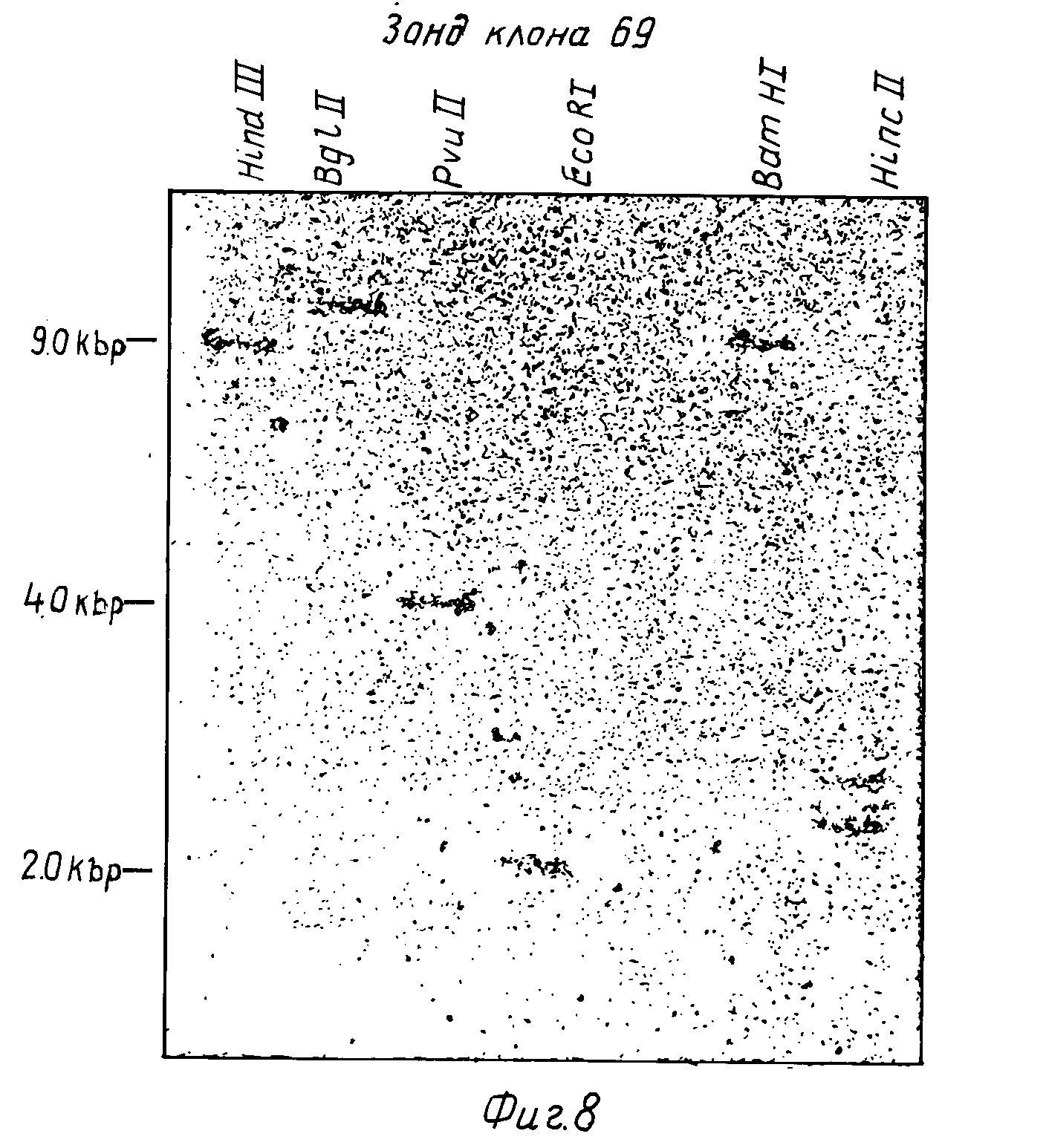
На фиг. 8 представлена Саузерн-гибридизация человеческой геномной ДНК, переваренной 6 различными рестрикционными эндонуклеазами, гибридизованной с32P-меченным зондом из p69.
А. Источник IFN-γ мРНК
Лимфоциты периферической
крови (PBL) были получены от доноров (людей) с помощью лейкофореза. Далее PBL были очищены градиентным центрифугированием в смеси фиколл-гепарин, а затем их культивировали при концентрации 5•
106 кл/мл в RPMI 164 с 1 % L-глутамина, 25 мМ HEPS и 1 % раствора пенициллин-стрептомицин (Gibco Jrand gsland, Ny). Эти клетки индуцировали для получения IFN-γ митогенным
стафилококковым энтеротоксином B (N мкг/мл) и культивировали в течение 34-48 ч при 37oC в 5 % CO. К культуре PBL добавили дезацетилтимозин -α- (0,1 мкг/мл) для повышения
относительного выхода IFN-γ активности.
В. Выделение информационной РНК.
Тотальную РНК из культур PBL экстрагировали, в основном, в соответствии с сообщением Bergen, S.L. et al. (33). Клетки выделяли центрифугированием, а затем повторно суспендировали в 10 мМ NaCl, 10мМ TrCS-HCl (pH 7,5), 1,5мМ MgCl2 и 10 мМ рибонуклеозид-ванадильного комплекса. Клетки лизировали, добавляя NP-40 (конечная концентрация 1%), и ядра выделяли центрифугированием.
Надосадочная жидкость содержала тотальную РНК, которую очистили далее многократными экстракциями фенолом и хлороформом. Водную фазу довели до 0,2М NaCl, а затем все РНК осадили, добавив два объема этанола. РНК неиндуцированных (нестимулированных) культур выделили таким же способом. Для очистки мРНК от остальных видов РНК использовали олиго-dТ целлюлозную хроматографию (34). Типичные выходы из 1-2 л культивированных PBL составляли 5-10 мг тотальной РНК и 50-200 мкг поли(А)+ PHK.
C.Фракционирование мРНК по размерам.
Для фракционирования препаратов мРНК использовали два способа. Эти способы использовали независимо ( а не вместе) и каждый из них приводил к значительному обогащению IFN - γ мРНК.
Для фракционирования мРНК использовали сахарозное градиентное центрифугирование в присутствии денатуранта формамида. Градиенты от 5 до 25 % сахарозы в 70 %-ном формамиде (32) центрифугировали при 154000•g в течение 19 ч при 20oC. Последовательные фракции (0,5 мл) выделяли затем с верхней части градиента, осаждали этанолом, а затем аликвоты вводили в социты Xenopus laevis для трансляции мРНК (35). Спустя 24 ч при комнатной температуре инкубационную среду стандартным методом ингибирования цитопатического эффекта исследовали на противовирусную активность, используя при этом Vesicular Stomatitis вирус (штамм Indiana) или вирус Eucepholomycarditis на клетках WISH (амнион человека) по описанию Стюарта (36), за исключением того, что образцы инкубировали с клетками в течение 24 ч (вместо 4) перед заражением вирусом. Соответственно наблюдали два пика активности для РНК, фракционированных в сахарозном градиенте (фиг. 1). Один пик седиментировал с рассчитанным размером 12S и содержал 100-400 ед/мл антивирусной активности (по сравнению со стандартом IFN-α ) на мкг введенной РНК.
Другой пик активности седиментировал с размером 16S и содержал около половины активности более медленно седиментированного пика. Каждый из этих пиков активности, по-видимому, связан с IFN-γ , так как для тех же фракций, исследовавшихся на линии бычьих клеток (MDBK), которые не защищены человеческим IFN-γ , не наблюдалось никакой активности. Как активность IFN-α , так и активность IFN-β , можно легко определить, исследуя МДВК (5).
Фракционирование мРНК (260 мкг) проводили также электрофорезом через кислотно-мочевинные агарозные гели. Вязкий агарозный гель (37 и 38) состоял из 1,75 % агарозы. О,025 М цитрата натрия, pH 3,8 и 6М мочевины. Электрофорез проводили в течение 7 ч при 25 мА и 4oC. Затем гель разрезали лезвием бритвы. Отдельные ломтики расплавляли при 70oC, после чего дважды экстрагировали фенолом и один раз хлороформом. Фракции осаждали этанолом, последовательно анализировали на предмет содержания мРНК IFN-γ введением в социты Xenopus laevis и затем проводили противовирусный анализ. Для фракционированных в геле образцов наблюдали только один пик активности (фиг. 2). Этот пик выходит вместе с 18S и имеет активность 600 ед/мл на микрограмм введенной РНК. Эта активность также, по-видимому, специфична для IFN-γ , так как не защищает клетки МДВК. Расхождение размеров, наблюдавшееся на сахарозных градиентах (12S и 16S) и кислотно-мочевинных гелях (18S) можно объяснить тем, что эти независимые методы фракционирования проводились в неодинаковых условиях полного денатурирования.
Д. Получение библиотеки колоний,
содержащих последовательности IFN-γ.
3 мг фракционированных в геле мРНК использовали для получения двунитевой кДНК по стандартным методикам ( 26 и 39)); кДНК фракционировали по размерам на 6% полиакриламидном геле. Фракции двух размеров электроэлюировали, 800 - 1500 п.о. (138 ng) и > 1500 п.о.(204 ng). Порции 35 ng каждого размера кДНК удлинили дезокси C-остатками, используя терминальную дезоксинуклеотидтрансферазу (40), и отжигали с 300 ng плазмиды pBK 322 (41), которая была аналогично сшита с дезокси-остатками в сайте Pst I(40). Каждую ренатурированную смесь трансформировали затем в E.coli К12 штамм 294. Получили приблизительно 8000 трансформантов с кДНК 800-1500 п.о. и 400 трансформантов кДНК > 1500 п.о.
Е. Выделение из библиотеки колоний,
индуцированных кДНК.
Полученные колонии отдельно инокулировали в лунки пластинок для микротитрования, содержащих LB (58) + 5 мгк/мл тетрациклина и хранившихся при - 20oC после добавления ЖДМСО до 7 %. Две копии библиотеки колоний выращивали на нитроцеллюлозных фильтрах и ДНК из каждой колонии фиксировали на фильтре по методу Gruushtein - Hogness (42).
32P - меченные кДНК зонды приготовили, используя гельфракционированные мРНК размером 18S из индуцированных и неиндуцированных культур РВL. В качестве праймеров использовали олиго-d Т 12-18; применялись условия реакции, описанные ранее (I). Фильтры, содержащие 8000 трансформантов с размером кДНК 600-1500 п.о. и 400 трансформантов с размером кДНК более 1500 п.о. гибридизовали с 20•106 срм индуцированных30P -кДНК. Дублирующий набор фильтров гибридизовали с 20•106 срм неиндуцированных32P- кДНК. Гибридизацию проводили в течение 16 ч, используя условия, описанные Fritsch et al (43). Фильтры тщательно промыли (43), а затем экспонировали на Кодаковской пленке XR-5 для рентгеновских лучей с помощью Du Pont Lighitning-Plus. интенсифицирующих экранов в течение 16-48 ч. Сравнивали картину гибридизации для каждой колонии с двумя пробами.
Приблизительно 40 % колоний явно были гибридизованы с обоими зондами, тогда как приблизительно 50 % колоний не подверглись гибридизации ни с одним зондом (см. фиг. 3). 124 колонии были гибридизованы заметно с индивидуальным зондом, но недетектируемо или очень слабо - с неиндуцированным зондом. Эти колонии индивидуально инокулировали в лунки пластинок для микротитрования, вырастили и перенесли на нитроцеллюлозные фильтры, а затем гибридизовали с теми же двумя пробами, как описано выше. Плазмидная ДНК, выделенная из каждой из этих колоний быстрым способом (44), была также связана с нитроцеллюлозными фильтрами и гибридизована (45) со стимулированными зондами. ДНК из 22 колоний, гибридизовавшееся только с индукционными зондами, были названы "индуцированными" колониями.
F. Характеристики индуцированных колоний.
Плазмидную ДНК получили из 5 индуцированных колоний (46) и использовали для того, чтобы охарактеризовать кДНК вставки. Рестрикционное мечение пяти индуцированных плазмид (р67, р68, р69, р70 и р71) показало, что четыре из них имеют аналогичные рестрикционные карты. Эти четыре плазмиды (р67, р69, р71 и р72) имеют каждая четыре Dde участка, 2 Hinf 1 участка и один Rsa 1 участок во вставке кДНК. Пятая плазмида (р68) содержит обычный Dde 1 фрагмент и, по-видимому, является коротким кДНК-клоном, относящимся к остальным четырем. Гомологичность, предполагаемая на основании картрирования с помощью рестрикционных нуклеаз, была подтверждена гибридизацией. Приготовили32P-меченную ДНК пробу (47) из DdeI фрагмента размером 600 п.о. плазмиды р67 и использовали для гибридизации (42) с остальными индуцированными колониями. Все пять картрированных рестрикционных нуклеазой колоний перекрестно гибридизовались с этим зондом, как и 17 других колоний из 124, выбранных при скрининге. Длину вставки кДНК в каждой из этих перекрестно гибридизирующихся плазмид определяли по перевариванию PstI и с помощью гельэлектрофореза. Клон с самой длинной кДНК-вставкой, по-видимому, является клоном 69 с размером вставки 1200-1400 п.о. Эту ДНК использовали во всех дальнейших экспериментах, ее рестрикционная карта приведена на фиг. 4.
Вставка кДНК в р69, как было показано, является IFN-γ кДНК по полученным в трех независимых системах экспрессии продуктам, проявлявшим противовирусную активность, как описано более подробно infra.
G. Анализ последовательной вставки кДНК р69.
Полная нуклеотидная последовательность плазмидной p69 кДНК-вставки была определена методом дидеоксинуклеотидного обрыва цепи (48) после субклонирования фрагментов в M 13 вектор m 7 (49) и химическим методом Максама и Гилберта (52). Наиболее длинная открытая рамка считывания кодирует белок из 166 аминокислот, представленный на фиг. 5. Первый остаток кодирует первый метиониновый кодон, включенный в 5'-конец кДНК. Первые 20 остатков у аминоконца, вероятно, служат сигнальной последовательностью для секреции остальных 146 аминокислот. Эту предполагаемую сигнальную последовательность с другими известными сигнальными последовательностями объединяют, например, размеры и гидрофобность. Кроме того, четыре аминокислоты, найденные у предполагаемой отщепляемой последовательности (ser-leu-glu-cys) были идентичны с четырьмя остатками, найденными в точке отщепления нескольких лейкоцитных интерферонов (LeIF B, C, D, F и H (2)). Закодированная зрелая аминокислотная последовательность из 146 аминокислот (именуемая в дальнейшем "рекомбинантный человеческий иммуноинтерферон") имеет молекулярную массу 17140.
Имеются два потенциальных положения гликозилирования (50) в закодированной белковой последовательности, у аминокислот от 28 до 30 (asn-gly-thr) и аминокислот от 100 до 102 ( asn-tyr-ser). Существование этих положений согласуется с наблюдавшимся гликозилированием человеческого IFN-γ (6 и 51). Кроме того, единственные два цистеиновых остатка (положения 1 и 3) стерически слишком близки, чтобы образовывать дисульфидный мостик, что согласуется с наблюдавшейся стабильностью IFN-γ в присутствии таких восстанавливающих агентов, как IFN-β - меркаптоэтанол (51). Выведенная зрелая аминокислотная последовательность, в общем, является основной, имея в сумме 30 лизиновых, аргининовых и гистидиновых остатков и всего 19 остатков аспарагиновой и глутаминовой кислот.
Структура мРНК IFN-γ , установленная из ДНК последовательности плазмиды р69, заметно отличается от IFN-α (1,2) или IFN-β (5) мРНК. Так, кодирующий участок IFN-γ короче, хотя 5' нетранслируемый и 3'-нетранслируемый участки гораздо длиннее, чем в IFN-ΙFN-α и IFN-β .
H. Структура кодирующей последовательности гена IFN-γ .
Структура гена, кодирующего IFN-γ , анализировали гибридизацией. В этой процедуре (54) 5 мкг высокомолекулярной человеческой ДНК (полученной по методу 55) переваривают до завершения с различными рестрикционными эндонуклеазами, проводят электрофорез на 1,0 % агарозном геле (56) и переносят на нитроцеллюлозный фильтр (54).32P-меченную пробу ДНК приготавливают (47) из DdeI фрагмента размером 600 п.о. кДНК-вставки р69 и гибридизуют (43) с ДНК пятном на фильтре. 107 импульсов в минуту пробы гибридизовали в течение 16 ч, а затем промывали как описано (43). Восемь геномных ДНК образцов от различных доноров (людей) переваривали Eco RI и гибридизовали с р6932P-меченным зондом. Как представлено на фиг. 7, наблюдается два четких сигнала гибридизации с размерами 8,8 m.п.о. и 2,0 m.п.о., что установлено путем сравнения подвижностей с Hind III переваренной λ-ДНК . Это могло быть результатом наличия или двух генов IFN-γ , или одного гена, расщепленного по Eco RI сайту. Так как р69 кДНК не содержит Eco RI сайтов, для объяснения наличия его в гене придется допустить промежуточную последовательность (интрон) с внутренним участком Eco RI. Для того, чтобы сделать различие между этими двумя возможностями, провели еще одну гибридизацию саузерна с тем же зондом и пятью другими эндонуклеазными перевариваниями одной человеческой ДНК (фиг. 8). По два гибридизуемых фрагмента ДНК наблюдали для других эндонуклеазных перевариваний, PVUII - 6,7 т.п.о. и 4,0 и Hinc II (2,5 т.п.о. и 2,2 т.п.о.). Однако три остальные картины эндонуклеазного переваривания дают только один гибридизующийся ДНК-фрагмент: Hind III (9,0 т.п.о.), Bdl II (11,5 т.п.о.) и BamHI (9,5 т. п. о. ) Два IFN-γ гена должны быть связаны необычно коротким расстоянием (менее 9,0 квр), чтобы оказаться в одном и том же Hihd III фрагменте. Этот результат предполагает, что только один гомологичный IFN-γ ген (в отличие от многих связанных с IFN-α генов) присутствует в человеческой геномной ДНК и что этот ген разделен одним или более интронов, содержащих Eco RI, PVU II и Hind II сайты. Это предположение было подтверждено гибридизацией32P-меченного (47) фрагмента, полученного из 3'-нетранслируемого участка кДНК из р69 (130 п.о. Dde I фрагмент от 860 положения до 990 положения на фиг. 5) с Eco RI переваром человеческой геномной ДНК. Только 2,0 т.п.о. Eco RI фрагмент гибридизуется с этим зондом, указывая на то, что этот фрагмент содержит 3'-нетранслированные последовательности, тогда как 3,8 т.п.о. Eco RI фрагмент содержит 5'-последовательности. Структура гена IFN-γ (один ген по крайней мере с одним интроном) существенно отличается от IFN-α (множество генов (2) без интронов (56)) или IFN-β (один ген без интронов (57)).
J. Конструирование вектора клеточной культуры pSVγ69.
Фрагмент из 342 пар оснований Hind III- PVU III, включающий инициатор SV 40, превратили во фрагмент, связанный с Eco R I рестрикционным сайтом. Hind III сайт превратили добавлением синтетического олигомера (5d AGCTGAATTC) и PVU II сайт превратили сшиванием по тупому концу в Eco RI сайт, дополнив его с использованием полимеразы I (фрагмент Кленова). Полученный Eco RI фрагмент вставили в Eсо RI сайт pML - (28). Плазмиду с поздним промотором SV 40, ориентированным в сторону от ampR гена, далее модифицировали, удалив Eco R I - сайт, ближайший к ampR гену pML - 1 (27).
Был выделен фрагмент размером 1023 пар оснований HpaI - BglII из клонированной HBV ДНК (60), и HpaI сайт вируса гепатита B (HBV) превратили в Eco RI сайт с синтетическим олигомером (5'dGGGAATTCGC). Этот Eco RI-Bgl II фрагмент непосредственно клонировали в Eco RI - BamH I сайты плазмиды, описанной ранее и несущей инициатор SV 40.
В оставшийся Eco R I - сайт вставили кодирующую IFN-γ последовательность на Pst I фрагменте р69 из 1250 п.о. после конверсии PstI концов в Eco R I концы. Выделили те клоны, в которых поздний промотор SV 40 предшествовал гену IFN-γ . Полученная плазмида pSVγ69 (фиг. 6) была затем введена в клетки культур тканей (29), в частности COS-7 клетки, с использованием ДЕАЕ-декстранметодики (61), модифицированной таким образом, что трансфекцию в присутствии ДЕАЕ-декстрана проводили в течение 8 ч. Клеточную среду меняли каждые 2-3 дн. Ежедневно отбирали 200 мкл на интерфероновый биоанализ. Типичные выходы составили 50-100 ед/мл в образцах, проанализированных три или четыре дня спустя после трансфекции.
Анализ показал, что продукт экспрессии не имеет cys-tyr-cys-N-концевой части рекомбинантного человеческого иммуноинтерферона (ср. фиг. 5), указывая на то, что явление отщепления сигнального пептида прошло по связи cys-GLN (аминокислоты 3 и 4 на фиг. 5) и что зрелый полипептит фактически состоит из 143 аминокислот (geз-cys-Tyr-cys-иммунный интерферон).
J. Частичная очистка рекомбинантного человеческого geз-cys-Tyr-cys-иммунного интерферона.
Для получения больших количеств человеческого интерферона IFN-γ , выделяемого обезьяньими клетками, свежие монослои COS-7 клеток в десяти 10 см пластинах были трансфецированы в общем 30 мкг рDL 1 3 в 110 мл ДЕАЕ-декстрана (200 мкг/мл ДЕАЕ декстран 500000 MW; 0,05 М трис pH 7,5 в ДМЕМ). Спустя 16 ч при 37oC, пластины промыли дважды ДМЕМ. 15 мл свежей ДМЕМ с добавлением 10 % f.b.s., 2 мМ глутамина, 50 мкг/мл пенициллина G и 50 мг/мл стрептомицина добавили затем на каждую пластину. Среду заменили освобожденной от сыворотки ДМЕM. Свежую, свободную от сыворотки среду добавляли затем ежедневно. Собранную среду хранили при 4oC до тех пор, пока не использовали для анализа, или связывали с CPG. Было обнаружено, что фракции, собранные спустя 3 и 4 дн после трансфекции, сохраняли практически всю активность.
0,5 г CPG (контролируемое пористое стекло Electonucleonics CPG 350 размер в мешках 120/200) добавляли к 100 мл клеточной надосадочной жидкости, и полученную смесь перемешивали в течение 3 ч при 4oC. После недолгого центрирования в beuch top центрифуге осевшие шарики набили в колонку и тщательно промыли буфером 20 мМ NaPO4, 1M NaCl, 0,1 % β -меркаптоэтанол, pH 7,2. Затем активность элюировали тем же самым буфером, содержащим 30 % этиленгликоля с последующим элюированием вышеуказанным буфером, содержащим 50 % этиленгликоль. Практически вся активность связана с CPG. 75 % элюированной активности нашли во фракциях, элюированных с 30 % этиленгликолем. Эти фракции собрали и разбавили 20 мМ NaPO4 1M NaCl, pH 7,2 до финальной концентрации 10 % этиленгликоля и непосредственно вводили в 10 мл колонку Con A Sepharose (Pharmacia). После тщательной промывки 20 мМ NaPO4 - 1M NaCl, pH 7,2 активности элюировали 20 мМ NaPO4 - 1M NaCl - 0,2 M α -метил-Д-маннозидом. Существенное количество активности (55 %) не связано с этим лектином, 45 % активности элюировано α -метил-Д-маннозидом.
К. Фармацевтические композиции.
Соединения настоящего изобретения можно включить в соответствии с известными способами в фармацевтически приемлемые композиции, в которых человеческий иммунный интерферон соединяют в смеси с фармацевтически приемлемым носителем. Подходящие носители и их композиции описаны в Pemington's Pharmaceutical Science, которая включена в виде ссылки. Такие композиции должны содержать эффективное количество белка интерферона в соответствии с настоящим изобретением вместе с подходящим количеством носителя для получения фармацевтически приемлемой композиции, подходящей для эффективного введения больному.
Парэнтеральный прием.
Человеческий иммунный интерферон настоящего изобретения можно парэнтерально вводить пациенту, для которого необходимо противоопухолевое или противовирусное лечение, а также тем, кто находится в иммуноподавленном состоянии. Дозы и частота приема могут быть аналогичными тем, которые обычно используют в клинических исследованиях на других человеческих интерферонах; то есть около (1-10)•106 единиц ежедневно, а в случае материалов с чистотой выше 1 %, по-видимому, вплоть 50•106 ед. ежедневно. Дозы IFN-γ можно значительно повысить для достижения большего эффекта, что связано с практическим отсутствием человеческих белков, отличных от IFN-γ , которые при использовании материалов, полученных от людей, могут привести к обратному действию.
В качестве одного примера подходящей дозировки для применяемого здесь практически гомогенного IFN-γ в парэнтеральной форме, можно указать 3 мг. IFN-γ специфической активности, скажем 2•108 ед/мг, можно растворить в 25 мл 5 н. сывороточного альбумина (человека) USP, раствор пропустить через бактериологический фильтр и отфильтрованный раствор асептически разделить на 100 ампул, каждая из которых содержит 6•106 ед. чистого интерферона, пригодного для парэнтерального введения. Ампулы предпочтительно хранить на холоде (-20oC) перед употреблением.
Данные биоисследований.
1. Характеристика противовирусной активности.
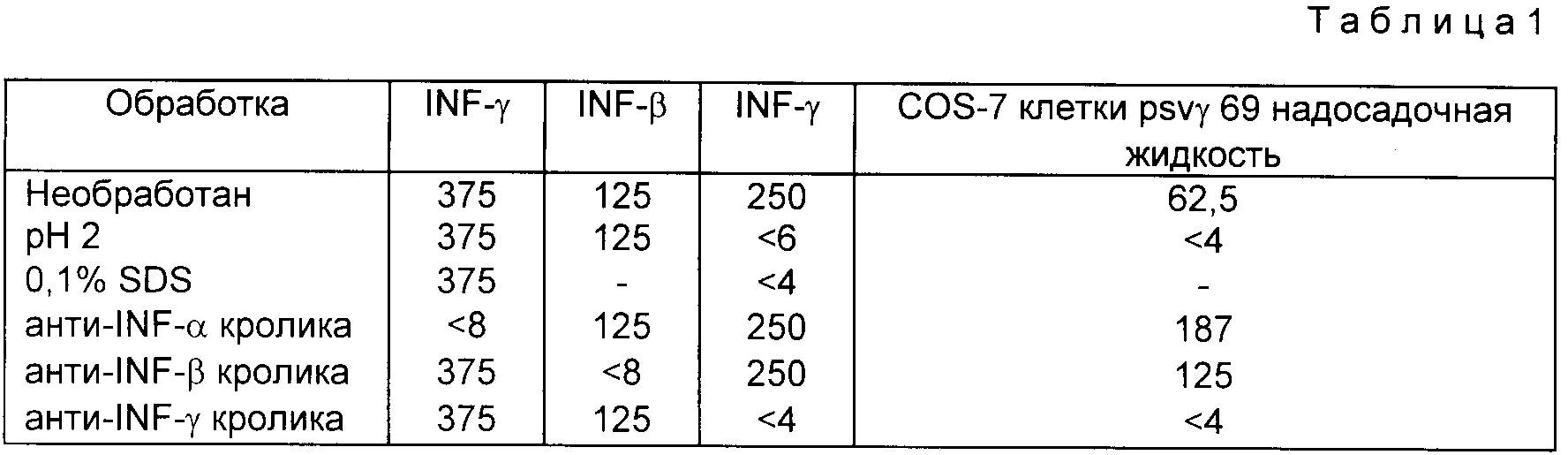
Для нейтрализации антител образцы разбавляли, в случае необходимости, до концентрации 500 - 1000 ед/мл, добавляя PBS - BSA. Равные объемы образца инкубировали в течение 2-12 ч при 4oC с рядом разбавлений кроличьих античеловеческих лейкоцитов, фибробластов или антисыворотки иммунного интерферона. Анти-IFN-α и β получили из национального института аллергических и инфекционных заболеваний. Анти-IFN-γ приготовили, используя аустентичный IFN-γ (5-20 % чистоты), очищенный из стимулированных лимфоцитов периферической крови. Образцы цетрифугировали в течение 3 мин при 1200 xg за 3 мин перед исследованием. Для проверки pH 2 - стабильности образцы доводили до pH 2, добавляя 1 н. HCl, инкубировали в течение 2-12 ч при 4oC и нейтрализовали добавлением 1 н. NaOH перед исследованием. Для тестирования натрийдодецилсульфатной (SDS) чувствительности образцы инкубировали с равным объемом 0,2 % SDS в течение 2-12 ч при 4oC непосредственно перед анализом.
Характеристики IFN-γ полученного в COS - 7
клетках даны в таблице
Эта таблица показывает характеристическое поведение IFN-α,β,γ - стандартов после различных обработок. Активность интерферона, полученного
COS-7/psvγ69, кислотно-чувствительная, SDS чувствительная и нейтрализуется антисывороткой иммунного интерферона. Она не нейтрализуется антителами к IFN-α или β . Эти данные
подтверждают то, что продукты, полученные в этой системе, являются иммуноинтерферонами и что к ДНК-вставка плазмиды p69 кодирует IFN-γ .
Источники информации.
1. Goeddel et al., Nature 287, 411 (1930).
2. Goeddel et al., Nature 290. 20(1981).
3. Yelverton et al., Nucleic Aceds Research 9. 731 (1981).
4. Gutterman et al., Annals of Int. Med. 93, 399 (1980).
5. Goeddel et al., Nucleic Acids Reseach 8, 4057 (1980).
6. Yip et al., Proc. Natl. Acad. Sci. (USA) 78, 1601 (1981).
7. Taniguchi et al., Proc. Natl. Acad. Sci. (USA) 78, 3469 (1981).
8. Bloom, Nature 289, 593 (1980).
9. Sonnenfeld et al., Cellular Immunol. 40, 285 (1978).
10. Fleishmann et al., Infection and Immunity 26, 248 (1979).
11. Blalock et al., Cellular Immunology 49, 390 (1980).
12. Rudin et al., Proc. Natl. Acad. Sci. (USA) 77, 5928 (1980).
13. Crane et al., J. Natl. Cancer Inst. 61, 871 (1978).
14. Stinchcomb et al., Nature 282, 39 (1979).
15. Kingsman et al., Gene 7, 141 (1979).
16. Tschumper el al.,. Gene 10, 157 (1980).
17. Mortimer et al., Microbiological Reviews 44, 519 (198 ).
18. Miozzari et al., Journal of Bacteriology 134, 48 (1978).
19. Jones, Genetics 85, 23 (1977).
20. Hitzeman, et al., J. Biol. Chem. 255, 12073 (1980).
21. Hess et al., J. Adv. Enzyme Regul. 7, 149 (1968).
22. Holland et al.. Biochemistry 17, 4900 (1978).
23. Bostian et al., Proc. Natl. Acad. Sci. (USA) 77, 4504 (1980).
24. The Molecular Biology of Yeast (Aug 11-18, 1981), Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
25. Chambon, Ann. Rev. Biochemistry, 44, 613 (1975).
25a. Gluzman, Cell 23. 175 (1981).
26. Goeddel et al., Nature 281, 544 (1979).
27. Itakura et al., Science 198, 1056 (1977).
28. Lusky et al., Nature 293, 79 (1981).
29. Gluzman et al. , Cold Spring Harbor Symp. Quant. Biol. 44, 293 (1980).
30. Fiers et al., Nature 273, 113 (1978).
31. Reddy et al., Science 200, 494 (1978).
32. Boedtker et al. , Prog. in Nucleic Acids Res. Mol. Biol. 19, 253 (1976).
33. Berger et al., Biochemistry 18, 5143 (1979).
34. Aviv et al., Proc. Natl. Acad. Sci. USA 69, 1408 (1972).
35. Gurdon et al., J. Molec. Biol. 80, 539 (1975).
36. Stewart, The Interferon System. Springer, New fork, p. 13-26 (1979).
37. Lehrach et al., Biochemistry 16, 4743 (1977).
38. Lynch el al., Virology 98, 251 (1979).
39. Wickens et al., J. Biol. Chem. 253, 2483 (1978).
40. Chang et al., Nature 275, 617 (1978).
41. Bolivar et al., Gene 2, 95 (1977).
42. Grunstein et al., Proc. Natl. Acad. Sci. U.S.A. 72, 3961 (1975).
43. Fritsch et al., Cell 19, 959 (1980).
44. Birnboim et al., Nucleic Acids Res. 7, 1513 (1979).
45. Kafatos et al., Nucleic Acids Res. 7, 1541 (1979).
46. Clewel et al., Biochemistry 9, 4428 (1970).
47. Taylor et al., Biochim. Biophys. Acta 442, 324 (1976).
48. Smith, Methods Enzymol. 61, 560 (1980).
49. Messing et al., Nucleic Acids Res. 9, 309 (1981).
50. Winzler, Hormonal Proteins and Peptides (ed. Li) Academic Press, New York, p. 1 (1973).
51. Mathan et al., Nature 292, 842 (1981).
52. Maxam et al., Methods in Enzymol. 65, 490 (1980).
53. Crea et al., Proc. Natl. Acad. Sci. (USA) 75, 5765 (1978).
54. Southern, J. Molec. Biol. 98, 503 (1975).
55. Blin et al., Nucleic Acids Res. 3, 2303 (1976).
56. Lawn et al., Science 212, 1159 (1981).
57. Lawn et al., Nucleic Acids Res. 9, 1045 (1981).
58. Miller, Experiments in Molecular Genetics, p. 431-3, Cold Spring Harbor Lab., Cold Spring Harbor, New York (1972).
59. Beggs, Nature 275, 104 (1978).
60. Valenzuela et al., Animal Virus Genetics (ed. Fields, Jaenisch and Fox) p. 57, Academic Press, New York (1980).
61. McCuthan et al., J. Natl. Cancer Inst. 41, 351 (1968).
Реферат
Использование: биотехнология, медицинская промышленность. Сущность изобретения: конструируют рекомбинантную плазмиду pSVγ69, включающую фрагмент ДНК-кодирующий предшественник иммунного интерферона человека; трансформируют полученным рекомбинантным вектором линию клеток СОS-7, отбирают трансформированные клетки, которые культивируют в условиях, обеспечивающих накопление целевого продукта, и выделяют зрелую форму γ интерферона без N-концевых Сys-Туr-Сys (des-Суs-Тyr- Сysγинтерферон). 8 ил.
Комментарии