Субъединичный вакцинный препарат респираторно-синцитиального вируса - RU2243234C2
Код документа: RU2243234C2
Чертежи

Описание
Настоящее изобретение относится к области иммунологии и касается, в частности, вакцинных препаратов против респираторно-синцитиальной вирусной инфекции.
Данная заявка является частичным продолжением одновременно рассматриваемой Патентной Заявки Соединенных Штатов №08/679060 от 12 июля 1996.
Респираторно-синцитиальный вирус человека (RS-вирус) является основной причиной инфекций нижних дыхательных путей у младенцев и маленьких детей (ссылки 1-3 - список ссылок приведен в конце описания, а кроме того, каждая ссылка из списка включена при цитировании). В мировом масштабе ежегодно в результате 65 миллионов случаев инфекционных заболеваний происходит 160000 смертей (ссылка 4). Из-за пневмонии и бронхиолитов, вызванных RS-вирусом, только в США за один год могут нуждаться в госпитализации 100000 детей (ссылки 5, 6). В США обеспечение стационарного и амбулаторного ухода за детьми с RS-вирусными инфекционными заболеваниями ежегодно обходится в сумму более 340 миллионов долларов (ссылка 7). Тяжелое заболевание нижних дыхательных путей вследствие RS-вирусной инфекции преимущественно развивается у младенцев 2-6 месячного возраста (ссылка 8).
Каждый год в США примерно 4000 младенцев умирают от осложнений, являющихся результатом тяжелых заболеваний нижних дыхательных путей вследствие инфицирования RS-вирусом и вирусом парагриппа типа 3 (Parainfluenza type 3 virus=PIV-3). Консультационные комитеты по вакцинам Всемирной организации здравоохранения (WHO) и Национального Института аллергических и инфекционных заболеваний (NIAID) по разработке вакцины приравнивают вторичный RS-вирус только к вирусу иммунодефицита человека (HIV).
Структура и состав RS-вируса (RSV) освещены и подробно описаны в сборнике “Fields Virology” Fields B.N. и др., Raven Press, N.Y. (1996), в частности, в Главе 44, стр. 1313-1351 “Respiratory Syncytial Virus”, Collins P. McIntosh К. и Chanock R.M. (ссылка 9).
Двумя основными защитными антигенами RSV являются гликопротеины оболочки вируса: гликопротеин слияния (F) и гликопротеин прикрепления (G)(ссылка 10). Белок F синтезируют как молекулу-предшественник с молекулярной массой около 68 кД (F0), которая протеолитически расщепляется на полипептидные фрагменты: дисульфид-связанный F1 (примерно 48 кД) и F2 (примерно 20 кД) (ссылка 11). Белок G (примерно 33 кД) сильно O-гликозилируют, получая гликопротеин со средней молекулярной массой примерно 90 кД (ссылка 12). Два широких подтипа RS-вируса обозначают А и В (ссылка 13). Основные антигенные различия между этими двумя подтипами обнаружены в гликопротеине G, тогда как гликопротеин F более консервативен (ссылки 1, 14).
Показано, что кроме реакции антител, генерированной гликопротеинами F и G, цитотоксические Т-лимфоциты человека, продуцируемые RSV-инфекцией, распознают RSV-белок F, матричный белок М, нуклеопротеин N, малый гидрофобный белок SH и неструктурный белок 1b (ссылка 15).
Не имеется безопасной и эффективной RSV-вакцины, и она крайне необходима. Подходы к разработке RS-вирусных вакцин включают инактивацию вируса формалином (ссылка 16), выделение адаптированного к холоду и/или чувствительных к температуре мутантных вирусов (ссылка 17) и очищенных гликопротеинов F или G (ссылки 18, 19, 20). Результаты клинических исследований показывают, что обе вакцины (живая ослабленная и инактивированная формалином) не являются адекватно действующими защитными вакцинами против RS-вирусной инфекции (ссылки 21-23). Неожиданные проблемы с ослабленными адаптированными к холоду и/или чувствительными к температуре мутагенами вируса, введенными через нос, включают клиническую заболеваемость, генетическую нестабильность и чрезмерное ослабление (ссылки 24-26). Живая RS-вирусная вакцина, введенная подкожно, также неэффективна (ссылка 27). Инактивированные RS-вирусные вакцины обычно получают, используя в качестве инактивирующего агента формальдегид. Murphy и др. (ссылка 28) приводят данные по иммунной реакции у младенцев и детей, привитых формалин-инактивированным RS-вирусом. Младенцы (2-6-месячного возраста) дают высокий титр антител на гликопротеин F, но имеют слабую реакцию на белок G. Дети старшего возраста (7-40 месяцев) дают титры F и G антител, сравнимые с соответствующими значениями у детей, инфицированных RS-вирусом. Однако и младенцы, и дети дают меньший уровень нейтрализующих антител, чем дети сравнимого возраста с естественным RS-вирусным инфекционным заболеванием. Неустойчивая иммунная реакция с высоким титром антител на основные иммуногенные белки RS-вируса F (слияния) и G(белок прикрепления), но низким титром нейтрализующих антител может быть частично вследствие альтераций важных эпитопов в гликопротеинах F и G при обработке формалином. Кроме того, у некоторых младенцев, которые принимали формалин-инактивированную RS-вирусную вакцину, после воздействия естественного RS-вируса развивалось более серьезное заболевание нижних дыхательных путей, чем у непривитых детей (ссылки 22, 23). Следовательно, считалось, что формалин-инактивированные RS-вирусные вакцины непригодны для использования человеком.
На хлопковых хомяках, привитых формалин-инактивированным RS-вирусом, наблюдают также доказательство искаженной иммунной реакции (ссылка 29). Кроме того, оценка вакцины формалин-инактивированного RS-вируса на хлопковых хомяках также показала, что после введения живого вируса у привитых животных развивается повышенная легочная гистопатология (ссылка 30).
Механизм усиления заболевания, вызванного вакцинными препаратами формалин-инактивированного RS-вируса, все еще подлежит изучению и является основным препятствием в разработке эффективной RS-вирусной вакцины. Усиление может происходить частично вследствие действия формалина на гликопротеины F и G. К тому же предполагают не-RS-вирус-специфический механизм усиления заболевания, когда свой вклад в обостряющееся заболевание частично может вносить иммунологическая реакция на присутствующие в вакцинном препарате примесные клеточные или сывороточные компоненты (ссылка 31). Действительно, мыши и хлопковые хомяки, которых прививают лизатом клеток НЕр-2 и которым вводят RS-вирус, выращенный на клетках НЕр-2, дают усиленную легочную воспалительную реакцию.
Кроме того, гликопротеины RS-вируса, очищенные способом иммуноаффинной хроматографии с элюированием при кислом рН, являются иммуногенными и защитными, но также вызывают иммуностимуляцию у хлопковых хомяков (ссылки 29, 32).
Ясно, что сохраняется необходимость в иммуногенных препаратах, включая вакцины, которые не только эффективны в создании защиты против заболевания, вызванного RSV, но также не дают нежелательных побочных эффектов, таких как иммуностимуляция. Существует также потребность в антигенах для диагностики RSV-инфекций и иммуногеннах для выработки антител (включая моноклональные антитела), которые специфически распознают RSV-белки, для использования, например, в диагностике заболевания, вызванного RS-вирусом.
Настоящее изобретение обеспечивает способ получения респираторно-синцитиального вируса (RSV) на клеточной линии для производства вакцин, например, клетках VERO, MRC5 или WI38, выделение вируса из образующихся в биологическом реакторе продуктов, экстракцию белков F, G и М из выделенного вируса и одновременную очистку белков F, G и М без стадий иммунного подобия или сродства к лентил-лектину или к конканавалину А. В частности, процедура сродства к лектину, описываемая, например, в патенте WO 91/00104 (США 07/7736949 от 28 июня 1990), закрепленном за представителем данной заявки, приведенной здесь в виде ссылки, может вести к выщелачиванию (leaching) лиганда в продукт.
Кроме того, в данной заявке впервые представлен способ совместного выделения и очистки белков F, G и М RS-вируса, а также иммуногенные композиции, включающие смеси совместно очищенных RSV-белков.
Совместно выделенные и очищенные RSV-белки F, G и М не пирогенны, не являются иммуностимуляторами и по существу не содержат сывороточных и клеточных примесей. Выделенные и очищенные белки иммуногенны, свободны от каких-либо инфекционных RSV и других случайных агентов.
В соответствии с этим один аспект настоящего изобретения обеспечивает смесь очищенных белков: белка слияния (F), белка прикрепления (G) и матричного белка (М), респираторно-синцитиального вируса (RSV).
Белок слияния (F) может состоять из многомерных составных белков (F), которые могут включать (при проведении анализа в невосстановительных условиях) гетеродимеры с молекулярной массой примерно 70 kDa и димерные и тримерные формы.
Белок прикрепления (G) может включать (при проведении анализа в невосстановительных условиях) олигомерный белок G, белок G с молекулярной массой примерно 95 кД и белок G с молекулярной массой примерно 55 кД.
Матричный белок (М) может включать (при проведении анализа в невосстановительных условиях) белок с молекулярной массой примерно от 28 до 34 кД.
Представленная здесь смесь белков (при проведении ДСН-электрофореза в полиакриламидном геле в восстановительных условиях (SDS-PAGE)) может включать белок слияния (F), включающий F1 с молекулярной массой примерно 48 кД и F2 с молекулярной массой примерно 23 кД, белок прикрепления (G), включающий белок G с молекулярной массой примерно 95 кД и белок G с молекулярной массой примерно 55 кД, и матричный белок (М), включающий белок М с молекулярной массой примерно 31 кД.
Смесь, которая обеспечивает соответствие с данным аспектом изобретения, может включать белки F, G и М в следующих относительных долях:
F примерно от 35 до 70 весовых %;
G примерно от 5 до 30 весовых %;
М примерно от 10 до 40 весовых %.
При проведении анализа способом электрофореза в полиакриламидном геле с ДСН (додецилсульфатом натрия) (SDS-PAGE) в восстановительных условиях и способом денситометрического сканирования после окрашивания серебром отношение F1 с молекулярной массой примерно 48 кД к F2 с молекулярной массой примерно 23 кД в данной смеси может составлять примерно от 1:1 до 2:1. Смесь белков F, G и М может иметь чистоту, по меньшей мере, около 75%, предпочтительно, по меньшей мере, около 85%.
Смесь, представленная здесь согласно данному аспекту изобретения, которая соответствует применяемому здесь способу выделения, описываемому ниже, свободна от моноклональных антител и свободна от лентил-лектина (lentil lectin) и конканавалина А.
RSV-белки, обеспеченные в представленной здесь смеси белков, обычно практически не денатурированы в мягких условиях получения и могут включать RSV-белки от одного или обоих подтипов RSV А и RSV В.
В соответствии с предпочтительным вариантом данного изобретения обеспечивают совместно выделенную и очищенную смесь неденатурированных белков респираторно-синцитиального вируса (RSV), состоящую по существу из белка слияния (F), белка прикрепления (G) и матричного белка (М) респираторно-синцитиального вируса (RSV), причем смесь свободна от лентил-лектинов, включая конканавалин А, и от моноклональных антител.
В соответствии с другим аспектом настоящего изобретения обеспечен иммуногенный препарат, включающий иммуноэффективное количество представленных в нем смесей.
Представленные здесь иммуногенные композиции можно составить в виде вакцины, содержащей белки F, G и М, для введения in vivo носителю, который может быть приматом, в частности человеком, с целью создания защиты против заболевания, вызванного RSV.
Иммуногенные композиции данного изобретения можно получить в виде микрочастиц, капсул, комплексов с иммуностимулирующими свойствами (ISCOM) или липосом. Иммуногенные композиции могут дополнительно включать, по меньшей мере, один отличающийся иммуногенный или иммуностимулирующий материал, которым может быть, по меньшей мере, один адъювант или, по меньшей мере, один иммуномодулятор, такой как цитокины, включая ILK.
По меньшей мере, один адъювант можно выбрать из группы, состоящей из фосфата алюминия, гидроксида алюминия, QS21, Quil А или их производных и компонентов, фосфата кальция, гидроксида кальция, гидроксида цинка, аналога гликолипида, октодецилового эфира аминокислоты, мурамил-дипептида, полифосфазена, липопротеина, ISCOM-матрицы, DC-Chol, DDA и других адъювантов и бактериальных токсинов, их компонентов и производных, описываемых, например, в USSN 08/258228 от 10 июня 1994, закрепленном за представителем данной заявки, описание которой включено в данное описание в виде ссылки (WO 95/34323). В конкретных обстоятельствах требуются адъюванты, которые индуцируют Th1-ответ.
Можно получить представленные здесь иммуногенные композиции, включающие, по меньшей мере, один подходящий дополнительный иммуноген, который может включать белок вируса парагриппа человека (PIV) из PIV-1, PIV-2 и/или PIV-3, такой как PIV-белки F и HN. Однако в композиции в виде поливалентных (комбинированных) вакцин можно также включить и другие иммуногены, такие как, например, из Chlamydia, полиовируса (polio), гепатита В, токсоида дифтерии, токсоида столбняка, гриппа, гемофильных бактерий (haemophilus), В. pertussis, пневмококков, микобактерий, гепатита А и Moraxella.
Еще один аспект настоящего изобретения обеспечивает способ генерирования иммунного ответа у носителя при введении ему иммуноэффективного количества представленной здесь иммуногенной композиции. Предпочтительно получать иммуногенную композицию в виде вакцины для введения носителю in vivo, и такое введение носителю, включая человека, создает защиту против заболевания, вызванного RSV. Иммунная реакция может быть гуморальной или клеточно-опосредованной.
Настоящее изобретение представляет собой еще один аспект - способ получения вакцины для защиты от заболевания, вызванного респираторно-синцитиальной вирусной (RSV) инфекцией, включающий введение представленной здесь иммуногенной композиции подопытному носителю для определения вводимого количества и частоты введения с целью создания защиты против заболевания, вызванного RSV; и составление иммуногенной композиции в виде, подходящем для введения пациенту, в соответствии с определенным количеством и частотой введения. Изучаемым носителем может быть человек.
Еще один аспект данного изобретения обеспечивает способ определения наличия в образце антител, специфически реагирующих с белком F, G или М, респираторно-синцитиального вируса (RSV), включающий следующие стадии:
(а) взаимодействие данного образца с представленной здесь смесью для получения комплексов, включающих белок респираторно-синцитиального вируса и любое из указанных антител, присутствующих в образце и специфически с ним реагирующих;
(б) определение образования комплексов.
Еще один аспект данного изобретения обеспечивает способ определения наличия в образце белка F, G или М респираторно-синцитиального вируса (RSV), включающий следующие стадии:
(а) иммунизацию субъекта посредством представленной здесь иммуногенной композиции для получения антител, специфически реагирующих с RSV-белками;
(б) взаимодействие данного образца с антителами для получения комплексов, включающих присутствующий в образце RSV-белок и белок-специфические антитела;
(в) определение образования комплексов.
Еще один аспект данного изобретения обеспечивает диагностический набор для определения наличия в образце антител, специфически реагирующих с RSV-белком F, G или М респираторно-синцитиального вируса, включающий:
(а) представленную здесь смесь;
(б) средства для взаимодействия данной смеси с образцом с целью получения комплексов, включающих белок респираторно-синцитиального вируса и любое из указанных антител, присутствующих в образце; и
(в) средства для определения образования комплексов.
Еще один аспект данного изобретения обеспечивает способ получения моноклональных антител, специфических для белков F, G или М респираторно-синцитиального вируса (RSV), включающий:
(а) введение представленной здесь иммуногенной композиции, по меньшей мере, одной мыши для получения, по меньшей мере, одной иммунизированной мыши;
(б) выделение В-лимфоцитов, по меньшей мере, из одной иммунизированной мыши;
(в) слияние В-лимфоцитов, по меньшей мере, от одной иммунизированной мыши с клетками миеломы с получением гибридом;
(г) клонирование гибридом, которые продуцируют селективные антитела против RSV-белка;
(д) культивирование клонов, продуцирующих селективные антитела против RSV-белка; и
(е) выделение антител против RSV-белка из селективных культур.
Еще один аспект данного изобретения обеспечивает способ получения совместно выделяемой и совместно очищаемой смеси белков респираторно-синцитиального вируса, который включает выращивание RSV на клетках в культуральной среде, выделение выращенного вируса из культуральной среды, солюбилизацию, по меньшей мере, белков F, G и М из выделенного вируса и совместное выделение и очистку солюбилизованных RSV-белков.
Совместное выделение и очистку можно проводить, загружая солюбилизованные белки на ионообменный носитель, предпочтительно носитель из фосфата кальция, в частности носитель из гидроксиапатита, и проводя совместное селективное элюирование белков F, G и М с ионообменного носителя. Выращенный вирус можно сначала промыть мочевиной для удаления примесей, не удаляя в существенной степени белки F, G и М.
Достижения настоящего изобретения включают:
- совместно выделенные и совместно очищенные смеси белков F, G и М RSV;
- иммуногенные композиции, содержащие такие белки;
- способы выделения такого белка,
- диагностические наборы для идентификации RSV и инфицированных им носителей.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
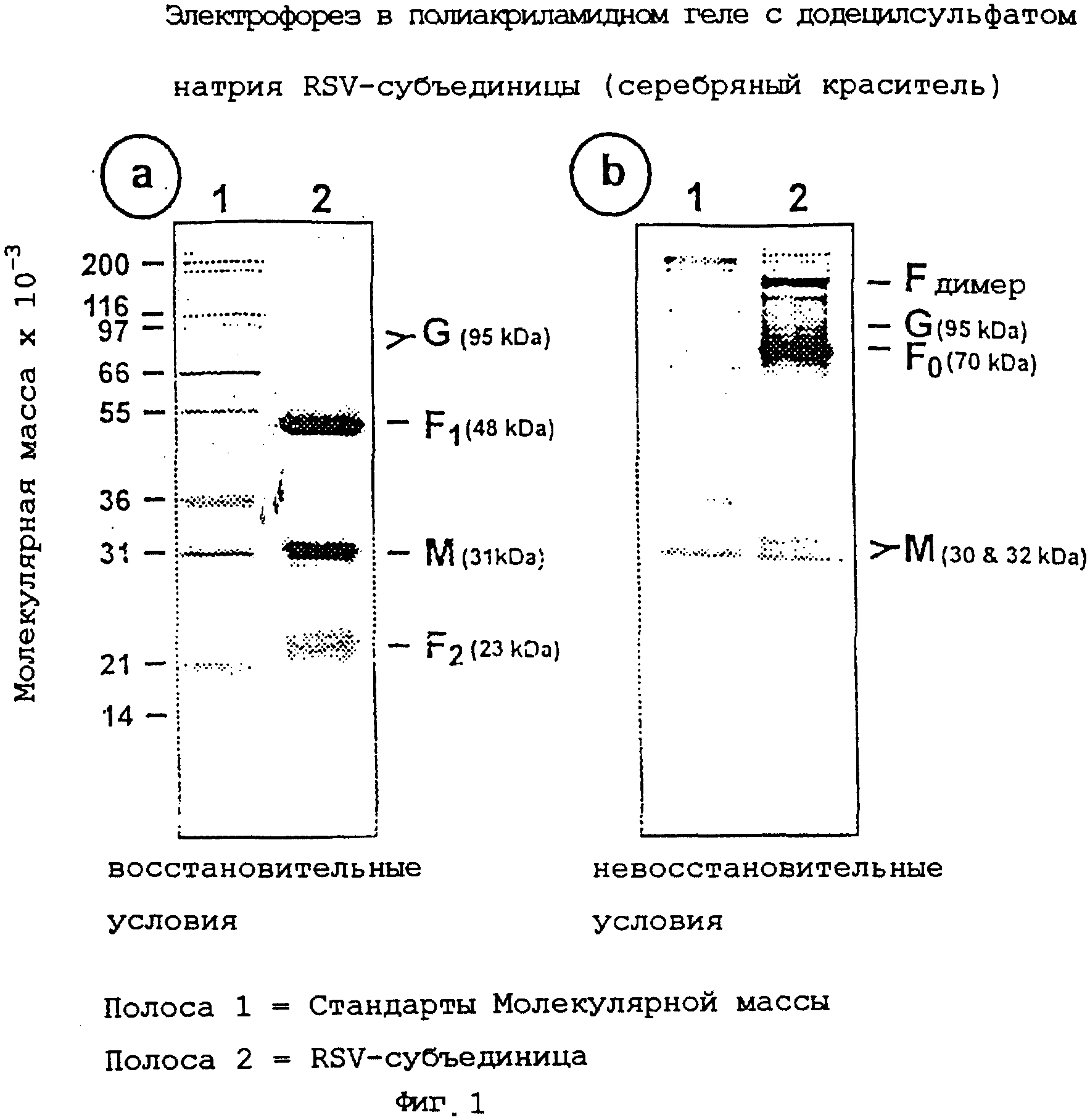
На фиг.1, содержащей изображения а и b, показан анализ способом электрофореза в полиакриламидном геле с додецилсульфатом натрия (ЭФ в ДСН-ПААГ) очищенного препарата субъединицы RSV А с использованием акриламидных гелей, окрашенных серебром в восстановительных (изображение а) и невосстановительных (изображение b) условиях.
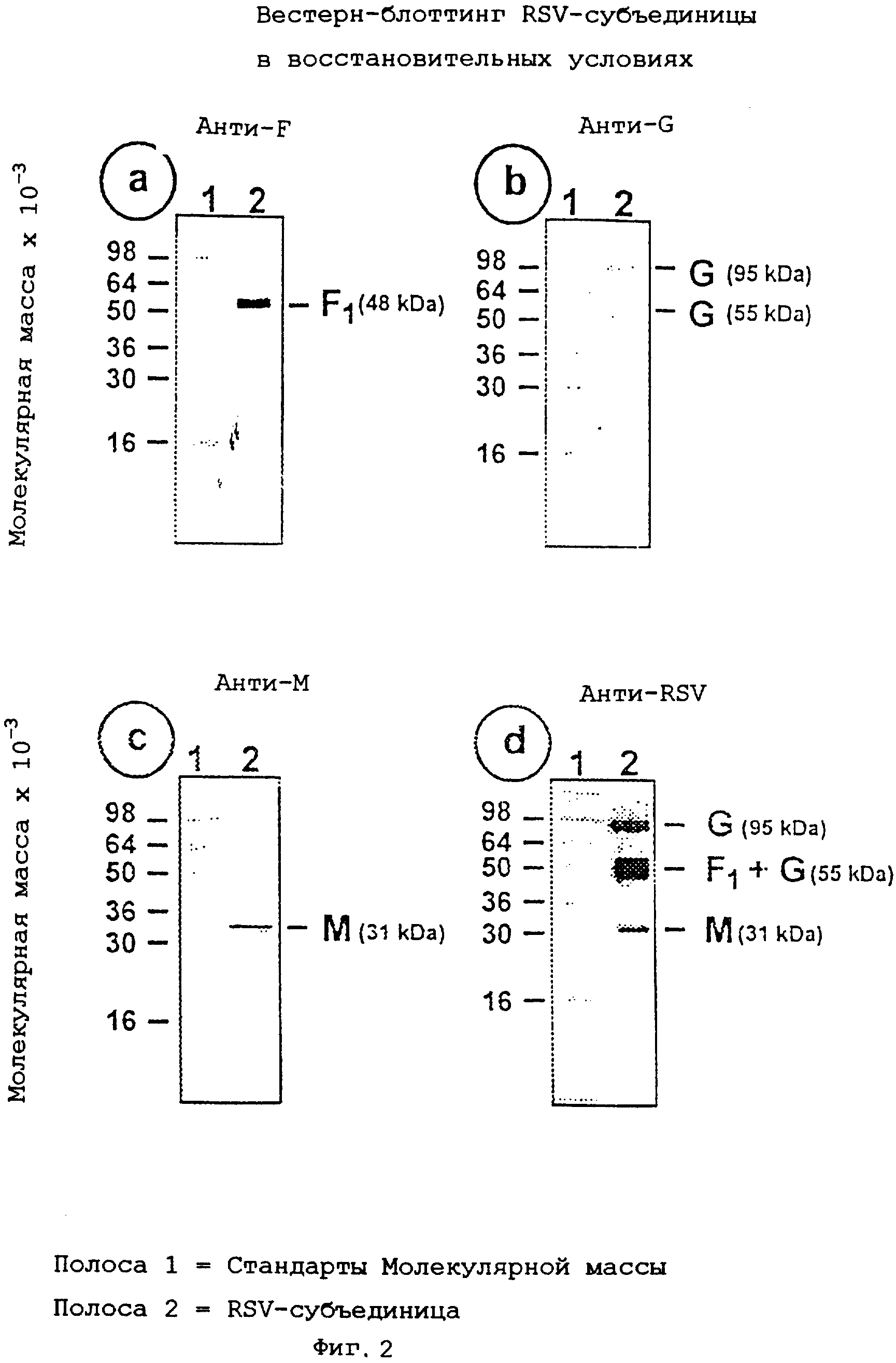
На фиг.2, содержащей изображения a, b, c и d, показан Вестерн-блоттинг очищенного препарата субъединицы RSV А в восстановительных условиях.
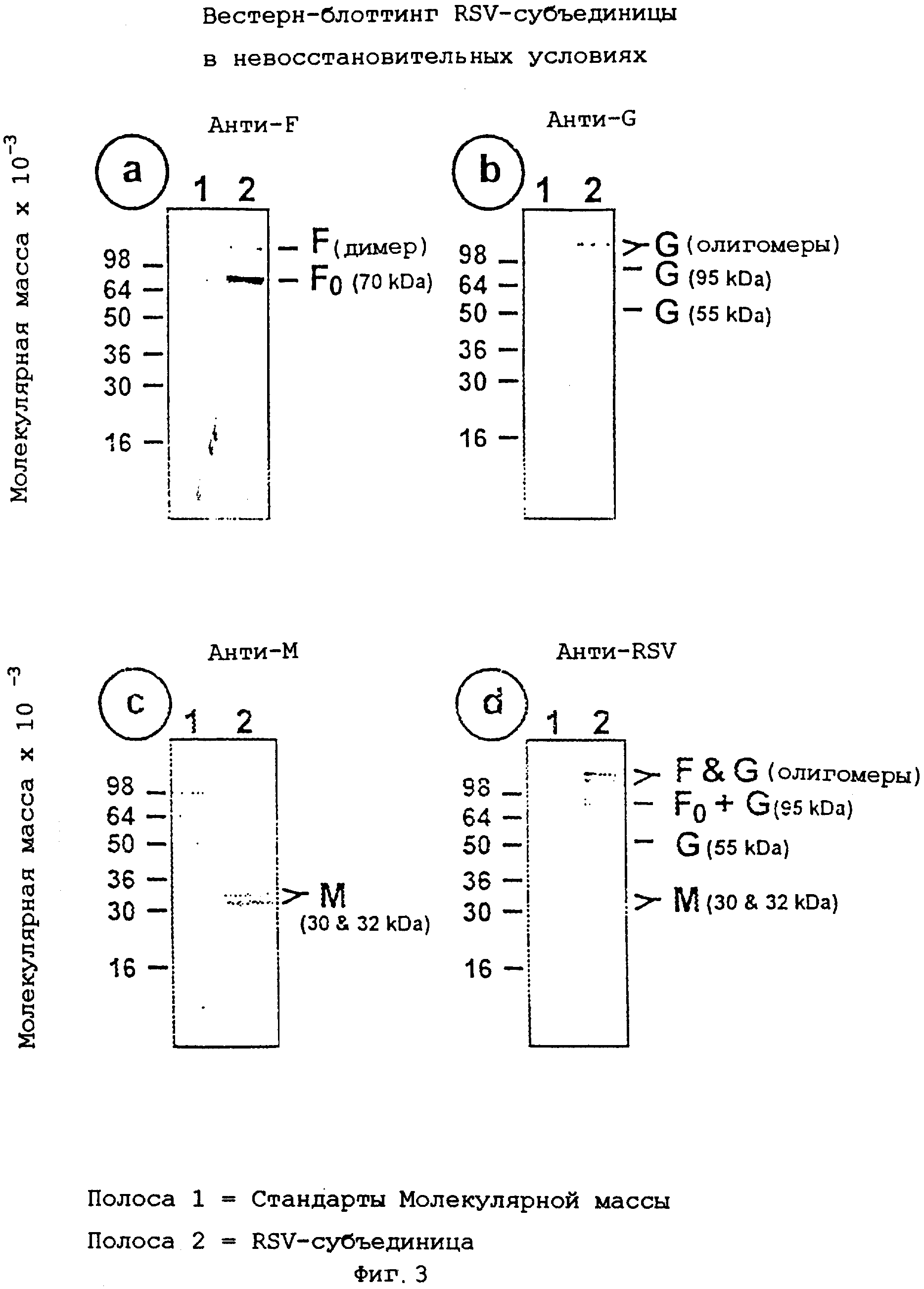
На фиг.3, содержащей изображения а, b, с и d, показан Вестерн-блоттинг очищенного препарата субъединицы RSV в невосстановительных условиях.
На фиг.4, содержащей изображения а, b, с и d, показан анализ способом электрофореза в полиакриламидном геле с додецилсульфатом натрия (ЭФ в ДСН-ПААГ) очищенного препарата субъединицы RSV В, с использованием акриламидных гелей, окрашенных серебром в восстановительных условиях.
ОСНОВНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Как обсуждалось выше, настоящее изобретение обеспечивает белки F, G и М RS-вируса, совместно выделенные из RS-вируса и очищенные. Вирус выращивают на клеточной линии для производства вакцин, такой как клетки VERO и диплоидные клетки человека, такие как MRC5 или WI38, и выращенный вирус собирают. Ферментацию можно проводить в присутствии эмбриональной бычьей сыворотки (FBS) и трипсина.
Вирусный сбор фильтруют и затем концентрируют, обычно применяя способ тангенциальной проточной ультрафильтрации и мембрану с необходимым порогом молекулярной массы, и проводят диафильтрацию. Концентрат вирусного сбора можно отцентрифугировать и супернатант выбросить. Остаток после центрифугирования можно сначала промыть буфером, содержащим мочевину, для удаления растворимых примесей, оставляя белки F, G и М практически нетронутыми, и затем повторно центрифугировать. Затем остаток после этого центрифугирования экстрагируют детергентом для солюбилизации из него белков F, G и М. Такую экстракцию детергентом можно проводить путем повторного суспендирования сухого остатка до объема исходного концентрата сбора в экстракционном буфере, содержащем детергент, такой как неионный детергент, включая TRITON® X-100, неионный детергент, который представляет собой отктадиенилфенол(этиленгликоль)10. Другие детергенты включают октилглюкозид и Меgа-детергенты.
После центрифугирования с целью удаления нерастворимых белков экстракт белков F, G и М чистят хроматографическими способами. Сначала можно нанести экстракт на ионообменный хроматографический носитель, позволяя белкам F, G и М связаться с носителем, тогда как примесям дают протечь сквозь колонку. Ионообменным хроматографическим носителем может быть любой необходимый хроматографический материал, в частности носитель из фосфата кальция, конкретно гидроксиапатит, хотя можно применять и другие материалы, такие как DEAE, ТМАЕ и другие.
Связанные белки F, G и М затем совместно элюируют из колонки подходящим элюентом. Полученные совместно очищенные белки F, G и М можно дополнительно обработать для повышения степени их чистоты.
Используемые здесь очищенные белки F, G и М могут быть в виде гомо- и гетероолигомеров, включая F:G гетеродимеры и димеры, тетрамеры и образцы более высокого порядка. В препаратах RSV-белков, полученных этим способом, не выявляют присутствия какого-либо побочного агента, гемадсорбирующего агента или живого вируса.
Группам хлопковых хомяков проводили внутримышечную иммунизацию представленными здесь препаратами в комбинации с alum или Iscomatrix™ в качестве адъюванта.
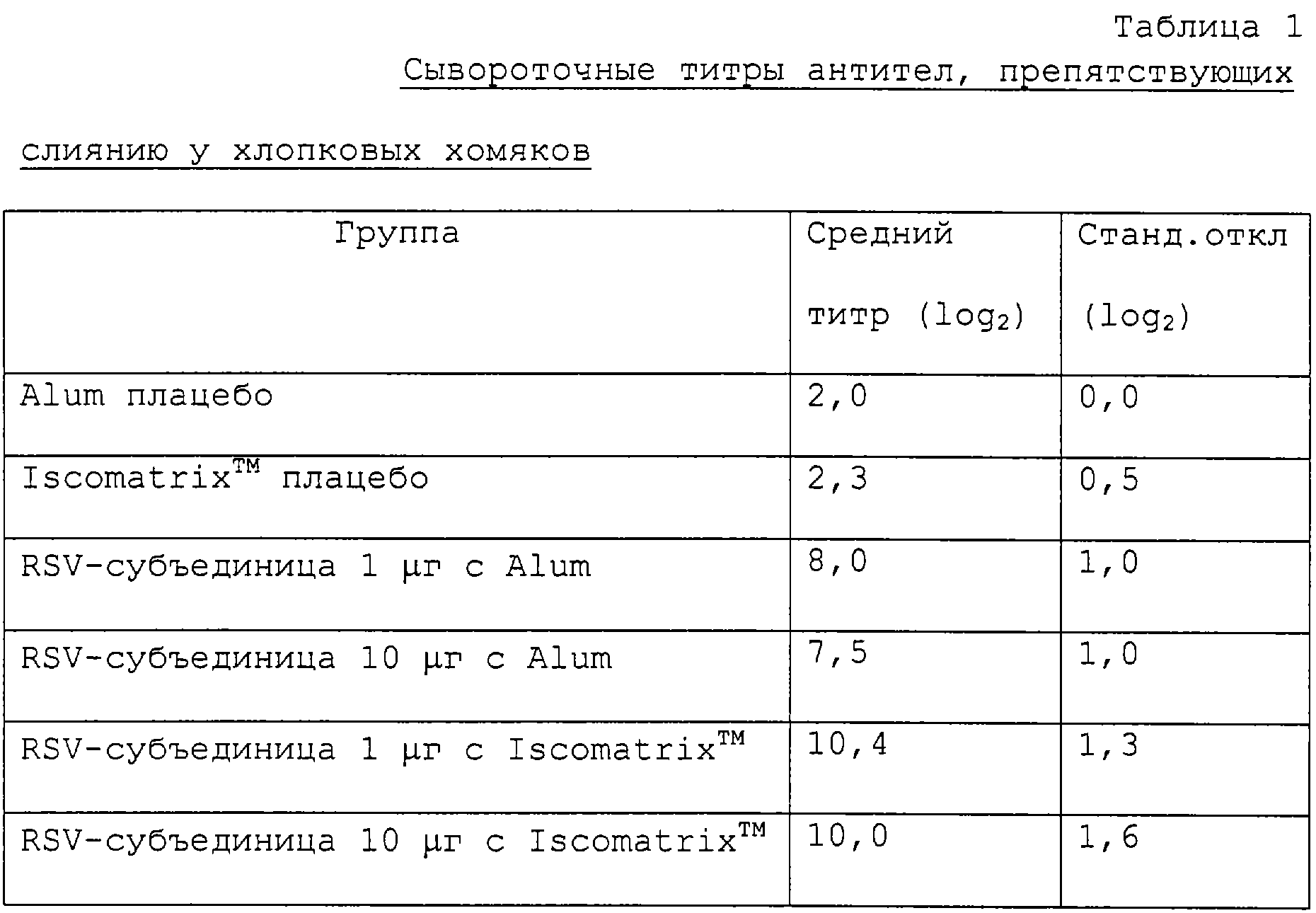
Как показано ниже в табл.1 и 2, получены устойчивые титры антител, препятствующих слиянию, и нейтрализующих антител. Как показано ниже в табл.3 и 4, получена полная защита против вирусной инфекции верхних и нижних дыхательных путей.
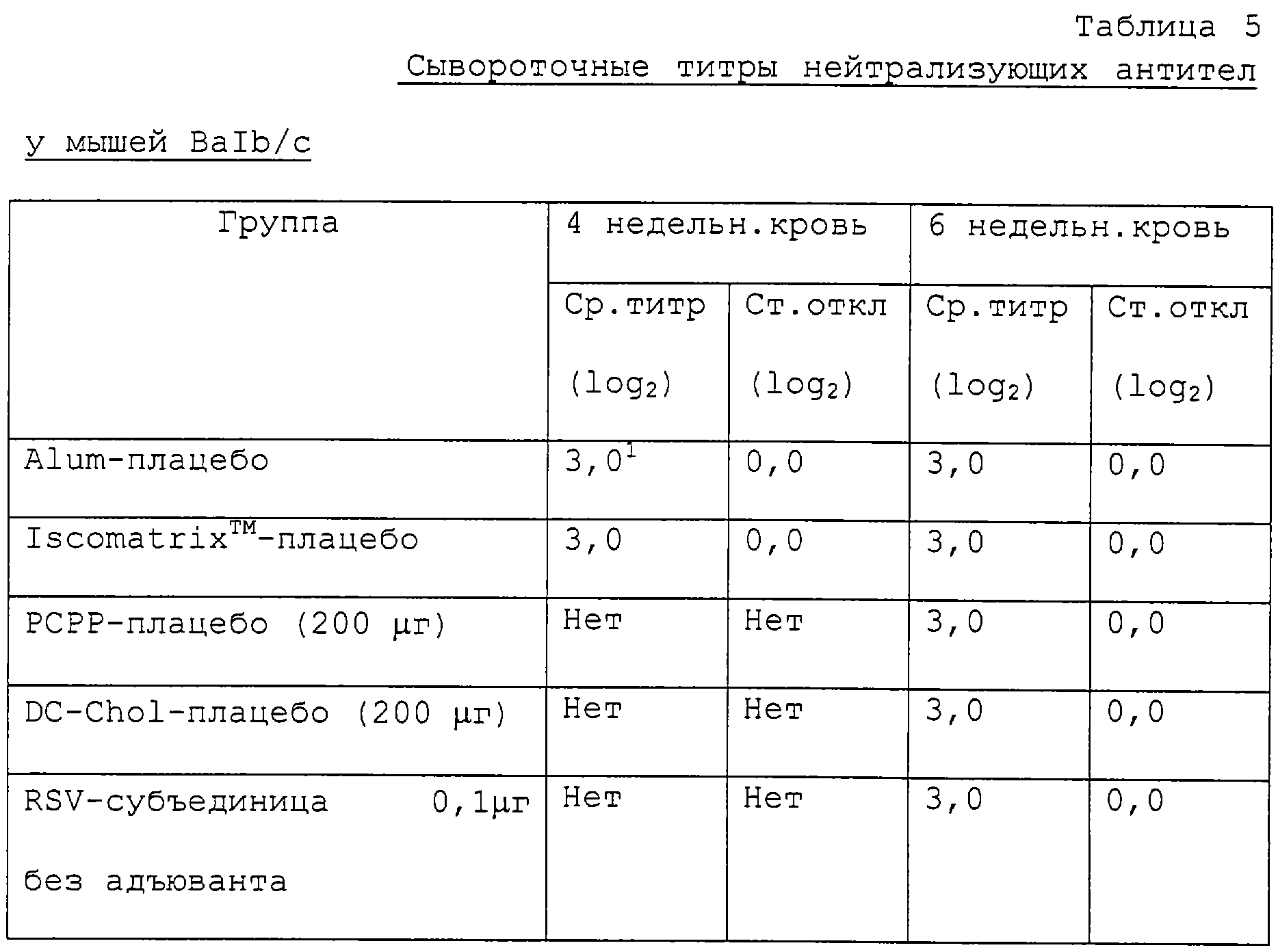
Кроме того, группам мышей проводили внутримышечную иммунизацию представленными здесь препаратами в комбинации с alum, Iscomatrix™, полифосфазеном и DC-chol в качестве адъюванта. Как показано ниже в табл.5 и 6, получены устойчивые сывороточные титры нейтрализующих антител и анти-F-антител. К тому же получена полная защита против вирусной инфекции, что показано отсутствием вируса в легочных гомогенатах (табл.7).
Группам обезьян также проводили иммунизацию представленными здесь препаратами в комбинации с alum или Iscomatrix™ в качестве адъюванта. Как показано в табл.8 и 9, получены устойчивые сывороточные титры нейтрализующих антител и титры анти-F-антител.
Полученные данные по иммунизации животных показывают, что, применяя мягкую экстракцию детергентом основных RSV-белков из вируса и мягкое элюирование белков солевым раствором с ионообменного носителя, получают совместно очищенные смеси RSV-белков F, G и М, которые способны вызывать иммунную реакцию у подопытных модельных животных, что создает защиту против RSV.
Данное изобретение распространяется на смеси белков F, G и М из респираторно-синцитиального вируса для использования в качестве фармацевтического вещества - активного ингредиента - в вакцине против заболевания, вызванного инфицированием респираторно-синцитиальным вирусом.
Еще один аспект данного изобретения обеспечивает применение белков F, G и М из респираторно-синцитиального вируса с целью получения вакцинной композиции для иммунизации против заболевания, вызванного инфицированием респираторно-синцитиальным вирусом.
Специалисту ясно, что различные варианты настоящего изобретения имеют много применений в областях вакцинации, диагностики и лечения респираторно-синцитиальных вирусных инфекций и получения иммунологических агентов.
Ниже приведено дополнительное неограничительное обсуждение таких результатов.
1. Получение и применение вакцины
Как описано в данной работе, из смесей, включающих иммуногенные белки F, G и М RS-вируса, можно получать иммуногенные композиции, подходящие для использования в качестве вакцин. Иммуногенная композиция усиливает иммунную реакцию, которая продуцирует антитела, включая анти-RSV-антитела, включающие анти-F, анти-G и анти-М-антитела. Такие антитела могут быть антителами, нейтрализующими вирусы, и/или антителами, препятствующими слиянию.
Иммуногенные композиции, включая вакцины, можно получить в виде препаратов для инъекций, жидких растворов, суспензий или эмульсий. Активный иммуногенный ингредиент или ингредиенты можно смешать с фармацевтически пригодными совместимыми наполнителями. Такие наполнители могут включать воду, солевой раствор, декстрозу, глицерин, этанол и их комбинации. Иммуногенные композиции и вакцины могут дополнительно содержать вспомогательные вещества, такие как увлажнители или эмульгаторы, рН-буферные агенты или адъюванты для усиления их эффективности. Иммуногенные композиции и вакцины можно вводить парентерально, посредством подкожных, внутрикожных или внутримышечных инъекций. По-другому, иммуногенные композиции, составленные в соответствии с настоящим изобретением, можно получить и доставить способом, вызывающим иммунную реакцию на поверхностях слизистых оболочек. Таким образом, иммуногенные композиции можно наносить на поверхности слизистых оболочек, например, через нос или оральным способом (внутрижелудочно). Могут потребоваться другие модели введения, включая суппозитории и препараты для орального применения. Связующие и носители для суппозиториев могут включать, например, полиалкенгликоли или триглицериды. Такие суппозитории можно получить из смесей, содержащих активный иммуногенный ингредиент (ингредиенты) в количестве примерно от 0,5 до 10%, предпочтительно примерно от 1 до 2%. Препараты для орального применения могут включать обычно применяемые носители, такие как фармацевтические марки сахарина, целлюлозы и карбоната магния. Эти композиции могут иметь вид растворов, суспензий, таблеток, пилюль, капсул, препаратов с замедленным выделением или порошков и содержать примерно от 1 до 95% активного ингредиента (ингредиентов), предпочтительно примерно от 20 до 75%.
Иммуногенные препараты и вакцины вводят способом, совместимым с дозированной препаративной формой, и в таком количестве, которое является терапевтически эффективным, иммуногенным и защитным. Принимаемое количество зависит от подлежащего лечению субъекта, включая, например, способность индивидуальной иммунной системы синтезировать антитела и, если требуется, давать клеточно-опосредованную иммунную реакцию. Точные количества подлежащего введению активного ингредиента зависят от заключения лечащего врача. Однако специалист может легко определить подходящие диапазоны доз, которые могут составлять количество порядка от микрограммов до миллиграммов активного ингредиента (ингредиентов) на вакцину. Подходящие схемы для начального приема и повторной иммунизации также различны, но могут включать первое введение с последующей повторной иммунизацией. Передозировка может также зависеть от способа введения и будет меняться в соответствии с размером носителя.
Концентрация белка-активного ингредиента в иммуногенной композиции, соответствующей данному изобретению, обычно составляет примерно от 1 до 95%. Вакцина, содержащая антигенный материал только одного патогена, является моновалентной вакциной. Вакцины, содержащие антигенный материал нескольких патогенов, являются комбинированными вакцинами и также принадлежат настоящему изобретению. Такие комбинированные вакцины содержат, например, материал от разных патогенов или от разных штаммов одного патогена или от комбинаций различных патогенов. В настоящем изобретении, как отмечено выше, белки F, G и М вируса RSV А и RSV В объединены в одну мультивалентную иммуногенную композицию, которая также может содержать другие иммуногены.
Иммуногенность может быть значительно усилена в случае, если антигены вводятся вместе с адъювантами. Адъюванты усиливают иммуногенность антигена, при этом необязательно должны быть сами иммуногенными. Адъюванты могут действовать, удерживая антиген локально вблизи сайта применения для получения длительного эффекта, облегчающего медленное и постепенное высвобождение антигена и поставки его клеткам иммунной системы. Адъюванты могут также привлекать клетки иммунной системы к антигенному депо и стимулировать такие клетки для получения иммунного ответа.
Иммуностимуляторы или адъюванты используют в течение многих лет для улучшения иммунных реакций носителя, например, на вакцины. Внутренние адъюванты, такие как липополисахариды, обычно являются компонентами применяемых в вакцинах убитых или аттенюированных бактерий. Внешние адъюванты являются иммуномодуляторами, которые включают для усиления иммунных реакций носителя. Таким образом, адъюванты определяют как вещества, усиливающие иммунную реакцию на парентерально доставленные антигены. Однако некоторые из этих адъювантов токсичны и могут вызывать нежелательные побочные эффекты, делая их непригодными для применения людьми и для многих животных. Действительно, в человеческих и ветеринарных вакцинах обычно используют в качестве адъюванта только гидроксид алюминия и фосфат алюминия (общее обозначение alum). Эффективность alum в повышении реакций антител на токсоиды дифтерии и столбняка доказана, и в вакцине HbsAg используют в качестве адъюванта alum. При том, что полезность alum доказана для многих применений, она имеет ограничения. Например, alum неэффективны для вакцинации гриппа и обычно не вызывают клеточно-опосредованную иммунную реакцию. Антитела, вызванные антигенами с alum-адъювантами, в основном имеют IgGl-изотип у мышей, что может быть неоптимальным для защиты посредством некоторых вакцинных агентов.
Широкий диапазон внешних адъювантов может вызывать сильные иммунные реакции на антигены. Они включают сапонины, образующие комплекс с мембранными белковыми антигенами (иммуностимулирующие комплексы), Pluronic-полимеры с минеральным маслом, убитые микобактерии в минеральном масле, неполный адъювант Фрейнда, бактериальные продукты, такие как мирамил дипептид (MDP) и липополисахариды (LPS), а также липид А и липосомы.
Для эффективного индуцирования гуморальных иммунных реакций (HIR) или клеточно-опосредованного иммунитета (CMI) иммуногены часто эмульгируют в адъювантах. Многие адъюванты токсичны и вызывают гранулемы, острые и хронические воспаления (полный адъювант Фрейнда, FCA), цитолиз (сапонины и Pluronic-полимеры) и повышение температуры тела, артрит и предшествующий увейт (LPS и MDP). Хотя FCA является превосходным адъювантом и широко используется в исследованиях, из-за своей токсичности он не имеет лицензии на применение в человеческих или ветеринарных вакцинах.
2. Иммунологические исследования
К тому же RSV-белки F, G и М настоящего изобретения полезны в качестве иммуногенов для генерации антител, в качестве антигенов в иммунологических исследованиях, включая твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (RIA) и другие исследования неферментного связывания антител или известные в практике способы детектирования антител. В ELISA-исследованиях белки F, G и М или смесь белков иммобилизуют на выбранной поверхности, например поверхности, способной связывать белки, такой как ячейки полистиренового планшета для микротитрования. После промывания с целью удаления неполностью адсорбированного материала можно связать с выбранной поверхностью неспецифический белок, такой как раствор бычьего сывороточного альбумина (BSA), который, как известно, антигенно нейтрален в отношении тестируемого образца. Это позволяет блокировать места неспецифической адсорбции на иммобилизующей поверхности и снизить таким образом основание, вызванное неспецифическим связыванием белков в антисыворотках на поверхности.
Затем иммобилизующую поверхность приводят в контакт с подлежащим исследованию образцом, таким как клинический или биологический материал, способом, приводящим к образованию иммунного комплекса (антиген/антитело). Способ может включать разбавление образца разбавителями, такими как растворы BSA, бычьего гамма-глобулина (BGG) и/или фосфатный буферный солевой раствор (PBS)/Tween. Затем образец инкубируют примерно от 2 до 4 ч при температурах порядка 25-37°С. После инкубации контактирующую с образцом поверхность промывают для удаления не включенного в иммунный комплекс материала. Процедура промывания может включать промывание раствором, таким как (PBS)/Tween или боратный буфер. После образования специфических иммунных комплексов между тестируемым образцом и связанным белком и последующего промывания можно определить возможность образования и даже количество иммунного комплекса, воздействуя на иммунный комплекс вторым антителом, специфичным на первое антитело. Если тестируемый образец имеет человеческую природу, второе антитело является антителом со специфичностью на человеческий иммуноглобулин и в основном IgG. Для обеспечения способов детектирования второе антитело может иметь ассоциированную активность, такую как ферментативная активность, которая будет генерировать, например, развитие цвета при инкубации с подходящим хромогенным субстратом. Затем можно провести количественное определение посредством измерения степени развития цвета, используя, например, спектрофотометр.
ПРИМЕРЫ
Приведенное выше раскрытие описывает настоящее изобретение. Более полного понимания можно достичь, обратившись к следующим специфическим примерам. Эти примеры приведены только в целях иллюстрации и не предназначены для ограничения области изобретения. В зависимости от обстоятельств или способа применения предполагаются изменения в форме и эквивалентная замена. Несмотря на то что здесь использованы специфические термины, такие термины предназначены для описания, а не с целью ограничения.
Способы определения дозы 50% инфицирования культуры ткани (ТСID50/мл), титр с образованием бляшек (plaque titre) и титр нейтрализующих антител, описываемые в данном описании без подробностей, достаточно широко представлены в научной литературе и хорошо известны специалистам. Концентрации белков определяют методом с применением бицинхониновой кислоты (ВСА), как описано в работе Pierce Manual (23220, 23225; Pierce Chemical company, США), включенной здесь в виде ссылки.
Для клеточных культур и роста вируса применяли культуральные среды: CMRL 1969 и среду Дульбекко, модифицированную по способу Искова (IMDM). Используемые в данном исследовании клетки являются клетками почек Африканской зеленой мартышки для производства вакцин (VERO lot М6), полученными из Института Merieux. Использованный здесь RS-вирус является RS-вирусом подтипа А (штаммы Long и А2), полученным из культуральных культур из Америки (АТСС), клиническим штаммом подтипа А и клиническим штаммом RSV подтипа В от Baylor College of Medicine.
Пример 1
Этот пример иллюстрирует производство RSV на клеточной линии млекопитающего на гранулах микроносителя в управляемом биологическом реакторе на 150 л.
Клетки почек Африканской зеленой мартышки для производства вакцин (VERO) при концентрации 105 клеток/мл добавляют к 60 л питательной среды CMRL 1969 (рН 7,2) в биологическом реакторе на 150 л, содержащем 360 г гранул микроносителя Cytodex-1, и перемешивают в течение 2 ч. Добавляют еще 60 л CMRL 1969, получая общий объем 120 л. Добавляют эмбриональную бычью сыворотку до конечной концентрации 3,5%. Добавляют глюкозу до конечной концентрации 3 г/л и L-глутамин до конечной концентрации 0,6 г/л. Регулируют количество растворенного кислорода (40%), рН (7,2), перемешивание (36 об/мин) и температуру (37°С). Контролируют рост клеток, содержание глюкозы, лактата и глутамина. На 4 день культуральную среду из биологического реактора осушают, добавляют 100 л питательной среды Е199 (без эмбриональной бычьей сыворотки) и перемешивают в течение 10 минут. Биологический реактор снова осушают и заполняют 120 л Е199.
Добавляют RSV-инокулят RS-вируса подтипа А при множественности заражения (M.O.I.) 0,001 и затем культуру сохраняют в течение 3 дней, после чего одну треть или половину питательной среды сливают и заменяют свежей средой. На 6-й день после инфицирования перемешивание прекращают и дают гранулам отстояться. Вирусную культуральную жидкость сливают и перед дальнейшей обработкой фильтруют через 20 μм фильтр, затем через 3 μм фильтр.
Осветленный вирусный сбор концентрируют в 75-150 раз способом тангенциальной проточной ультрафильтрации с 300 NnWL-мембранами и проводят диафильтрацию с использованием фосфатного буферного соляного раствора, содержащего 10%-ный глицерин. Вирусный концентрат до следующей очистки хранят в замороженном виде при -70°С.
Пример 2
Этот пример иллюстрирует способ выделения RSV-субъединицы из вирусного концентрата RSV подтипа А.
Раствор 50% полиэтиленгликоля-8000 добавляют к аликвоте вирусного концентрата, полученного, как описано в примере 1, получая конечную концентрацию 6%. После перемешивания при комнатной температуре в течение одного часа смесь центрифугируют в роторе Sorvall SS-34 при 4°С и 15000 об/мин в течение 30 минут. Вирусный осадок суспендируют в 1 мМ фосфате натрия (рН 6,8), 2 М мочевине, 0,15 М NaCl, перемешивают в течение 1 ч при комнатной температуре и затем повторно центрифугируют в течение 30 мин в роторе Sorvall SS-34 при 15000 об/мин и 4°С. Затем вирусный осадок суспендируют в 1 мМ фосфате натрия (рН 6,8), 50 мМ NaCl, 1% Triton Х-100 и перемешивают в течение 30 минут при комнатной температуре. Нерастворимый вирусный остаток удаляют посредством центрифугирования в роторе Sorvall SS-34 при 4°С и 15000 об/мин в течение 30 мин. Растворимый белковый супернатант вносят в колонку с керамическим гидроксиапатитом (тип II, Bio-Rad Laboratories) и затем промывают колонку пятикратным объемом (от объема колонки) 1 мМ фосфата натрия (рН 6,8), 50 мМ NaCl, 0,02% Triton X-100. Элюируя колонку десятикратным объемом (от объема колонки) 1 мМ фосфата натрия (рН 6,8), 400 мМ NaCl, 0,02% Triton X-100, получают композицию RSV-субъединицы из RSV подтипа А, содержащую F, G и М белки.
Пример 3
Этот пример иллюстрирует анализ препарата RSV-субъединицы, полученного из RSV подтипа А путем электрофореза на полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) и иммуноблоттинга.
Композицию RSV-субъединицы, полученную, как описано в примере 2, анализируют путем SDS-PAGE, используя 12,5% акриламидные гели. Электрофорез образцов осуществляют в присутствии или в отсутствие 2-меркаптоэтанола (восстановительный агент). Гели окрашивают серебряным красителем для регистрации вирусных белков (фиг.1, изображения а и b). Готовят иммуноблоты репликативных гелей и зондируют их мышиным моноклональным антителом (mAb 5353С75) на гликопротеин F (фиг.2, изображение а и фиг.3, изображение а), или мышиным моноклональным антителом (mAb 131-2G) на гликопротеин G (фиг.2, изображение b и фиг.3, изображение b), или антисывороткой морской свинки (gp178) против RSV-белка М (пептидная последовательность: LKSKNMLTTVKDLTMKTLNPTHDIIALCEFEN - SEQ ID №:1) (фиг.2, изображение с и фиг.3, изображение с), или козьей антисывороткой (Virostat #0605) против всего RSV (фиг.2, изображение d и фиг.3, изображение d). Денситометрический анализ окрашенного серебром геля препарата RSV-субъединицы после электрофореза в восстановительных условиях показывает следующее композиционное распределение белков:
гликопротеин G (95 кД)=10%;
гликопротеин F1 (48 кД)=30%;
белок М (31 кД)=23%;
гликопротеин F2 (23 кД)=19%.
Гликопротеин F в невосстановительных условиях мигрирует как гетеродимер с молекулярной массой примерно 70 кД (F0), а также в более высоких олигомерных формах (димеры и тримеры) (фиг.3, изображение а).
Пример 4
Этот пример иллюстрирует иммуногенность препарата RSV-субъединицы на хлопковых хомяках.
Группам из пяти хлопковых хомяков проводят внутримышечную иммунизацию (0,1 мл) в дни 0 и 28 при дозе 1 μг или 10 μг препарата RSV-субъединицы, полученного, как описано в примере 2, и приготовленного либо с 1,5 мг/дозу alum, либо с 5 μг/дозу Iscomatrix™ (Iscotec, Sweden). На 41 день берут образцы крови и определяют титры антител, препятствующих слиянию, и титры нейтрализующих антител. На 43 день хомякам вводят через нос RSV и через четыре дня умерщвляют. Собирают лаваж легких и носоглотки и определяют RSV-титры.
Получают устойчивые титры антител, препятствующих слиянию, и титры нейтрализующих антител, как показано ниже в табл.1 и 2. Кроме того, получают полную защиту против вирусной инфекции, за исключением одного хомяка, в верхних и нижних дыхательных путях (табл.3 и 4 ниже).
Пример 5
Этот пример иллюстрирует иммуногенность препарата RSV-субъединицы на мышах.
Группам из шести мышей BALB/c проводят внутримышечную иммунизацию (0,1 мл) в дни 0 и 28 при разных дозах препарата RSV-субъединицы, полученного, как описано в примере 2, и приготовленного с 1,5 мг/дозу alum, или 10 μг/дозу Iscomatrix™, или 200 μг/дозу полифосфазена (РСРР), или 200 μг/дозу DC-chol. Разные исследованные препараты представлены ниже в табл.5, 6 и 7. На 28 и 42 дни берут образцы крови и определяют титры нейтрализующих антител и анти-F антител. На 44 день мышам вводят RSV и через четыре дня умерщвляют. Легкие удаляют и гомогенизируют для определения титров вируса. Определяют устойчивые титры нейтрализующих антител и титры анти-F антител, приведенные ниже в табл.5 и 6. Кроме того, получают полную защиту против вирусной инфекции, что показано отсутствием вируса в гомогенатах легких и лаваже носа (табл.7 ниже).
Пример 6
Этот пример иллюстрирует иммуногенность препарата RSV-субъединицы на Африканских зеленых мартышках.
Группам из четырех мартышек проводят внутримышечную иммунизацию (0,5 мл) в дни 0 и 21 при дозе 100 μг/дозу препарата RSV-субъединицы, полученного, как описано в примере 2, и приготовленного либо с 1,5 мг/дозу alum, либо с 50 μг/дозу Iscomatrix™. На 21, 35 и 49 дни берут образцы крови и определяют титры нейтрализующих антител и анти-F-антител. Получают устойчивые титры, приведенные ниже в табл.8 и 9.
Пример 7
Этот пример дополнительно иллюстрирует получение RSV на линии клеток млекопитающего или микрогранулах в регулируемом биологическом ферментере на 150 л.
Клетки почек Африканской зеленой мартышки для производства вакцин (VERO cells) добавляют к 150 л питательной среды Дульбекко, модифицированной по способу Искова (IMDM), содержащей 3,5% фетальную коровью сыворотку (рН 7,2), до конечной концентрации 2× 105 клеток/мл (диапазон от 1,5×105 до 3,5×105 клеток/мл) в биологическом реакторе на 150 л, содержащем 450 г гранул микроносителя Cytodex-1 (3 г/л). После инокуляции клеток регулируют количество растворенного кислорода (40% насыщение воздуха (диапазон от 25 до 40%), рН (7,1×0,2)), перемешивание (36±2 об/мин) и температуру (37± 0,5°С). Контролируют начальное присоединение клеток к гранулам, ежедневный рост клеток (определяют количество клеток) и содержание в питательной среде глюкозы и лактата. Через 3-4 дня после начала роста клеток, когда концентрация клеток составляет от 1,5 до 106 до 2,0×106 клеток/мл, осуществляют инфицирование культуры VERO-клеток. Перемешивание прекращают, дают гранулам отстояться в течение 60 мин и сливают культуральную жидкость из биореактора, используя дренажную линию, помещенную приблизительно на 3 см выше установившегося объема гранул. Добавляют 75 л IMDM без эмбриональной бычьей сыворотки (среда для промывания) и смесь перемешивают в течение 10 мин при 36 об/мин. Перемешивание прекращают и дают гранулам отстояться в течение 30 мин. Удаляют среду для промывания, используя дренажную линию, и затем наполняют реактор до 75 л (половина объема) питательной средой IMDM без эмбриональной бычьей сыворотки.
Для инфицирования добавляют RSV-инокулят RS-вируса подтипа В при множественности заражения (multiplicity of infection=M.O.I.) 0,001 и в течение 2 ч при перемешивании (36 об/мин) проводят адсорбцию вируса на клетки при половинном объеме. Затем в биореактор добавляют 75 л IMDM до конечного объема 150 л. После инфицирования регулируют количество растворенного кислорода (40% насыщение воздуха (диапазон от 10 до 40%)), рН (7,25±0,1), перемешивание (36±2 об/мин) и температуру (37±0,5°С). После инфицирования ежедневно контролируют рост клеток (определение количества клеток), содержание в питательной среде глюкозы и лактата, антигены RSV F и G, и RSV-инфективность. На 3 день после инфицирования перемешивание прекращают, дают гранулам отстояться в течение 60 мин, через дренажную линию удаляют 75 л (50%) питательной среды и заменяют ее свежей питательной средой. Через 8 дней после инфицирования (диапазон от 7 до 9 дней), когда наблюдают полный вирус-индуцированный цитопатический эффект (то есть клетки отсоединяются от гранул микроносителя, и культура больше не поглощает кислород), перемешивание прекращают и дают гранулам микроносителя отстояться в течение 60 мин. Удаляют вирус-содержащую культуральную жидкость из биореактора и переносят в сосуд для хранения. Добавляют в биореактор 75 л IMDM без эмбриональной бычьей сыворотки и перемешивают в течение 30 мин при 75 об/мин. Гранулам микроносителя дают отстояться в течение 30 мин, удаляют из биореактора жидкость для промывания и объединяют с собранным материалом в сосуде для хранения.
Собранный материал концентрируют приблизительно в 20 раз способом тангенциальной проточной ультрафильтрации (вирус-содержащий материал удерживается мембраной), используя ультрафильтрационную мембрану на 500 или 1000 килодальтон (К) или, по-другому, 0,45 μМ микрофильтрационную мембрану, до конечного объема 10 л. Концентрированный материал подвергают диафильтрации с 10 объемами солевого раствора с фосфатным буфером (рН 7,2). Вирусный концентрат после диафильтрации до дальнейшей очистки хранят в замороженном виде при -70°С.
Пример 8
Этот пример иллюстрирует способ выделения RSV-субъединицы из вирусного концентрата RSV подтипа В.
Вирусный концентрат, полученный, как описано в примере 7, центрифугируют в роторе Sorvall SS-34 при 4°С и 15000 об/мин в течение 30 мин. Затем вирусный осадок суспендируют в 1 мМ фосфате натрия (рН 6,8), 300 мМ NaCl, 2% Triton X-100 и перемешивают в течение 1 ч при комнатной температуре. Нерастворимый вирусный остаток удаляют посредством центрифугирования в роторе Sorvall SS-34 при 4°С и 15000 об/мин в течение 30 мин. Растворимый белковый супернатант вносят в колонку с керамическим гидроксиапатитом (тип I, Bio-Rad Laboratories) и затем промывают колонку десятикратным объемом (от объема колонки) 1 мМ фосфата натрия (рН 6,8), 10 мМ NaCl, 0,02% Triton X-100. Элюируя колонку десятикратным объемом (от объема колонки) 1 мМ фосфата натрия (рН 6,8), 600 мМ NaCl, 0,02% Triton X-100, получают композицию RSV-субъединицы, содержащую F, G и М белки. В некоторых случаях композицию RSV-субъединицы дополнительно чистят, сначала разбавляя элюат из первой колонки с керамическим гидроксиапатитом, снижая концентрацию NaCl до 400 мМ и затем внося разбавленную субъединицу в колонку из керамического гидроксиапатита (тип II, Bio-Rad Laboratories). Посредством пропускания композиции RSV-субъединицы через эту колонку выделяют ее из RSV подтипа В.
Пример 9
Этот пример иллюстрирует анализ препарата RSV-субъединицы, полученного из RSV подтипа В, способом электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE).
Композицию RSV-субъединицы, полученную, как описано в примере 8, анализируют способом SDS-PAGE, используя 15% акриламидного геля. Образец был подвергнут электрофорезу в присутствии 2-меркаптоэтанола (восстановительный агент). Гель окрашивают серебряным красителем для регистрации вирусных белков (фиг.4). Денситометрический анализ окрашенного серебром геля препарата RSV-субъединицы в восстановительных условиях показывает следующее композиционное распределение белков:
гликопротеин G (95 кД)=21%;
гликопротеин F1 (48 кД)=19%;
белок М (31 кД)=22%;
гликопротеин F2 (23 кД)=20%.
КРАТКОЕ СОДЕРЖАНИЕ ИЗОБРЕТЕНИЯ
Говоря кратко, настоящее изобретение обеспечивает совместно выделенную и совместно очищенную смесь белков F, G и М RS-вируса, которая способна создать защиту против RSV на моделях инфицированных подопытных животных. Внутри области данного изобретения возможны модификации.










Список ссылок
1. Glezen, W.P., Paredes, A. Allison, J.E., Taber, L.K. and Frank, A.L. (1981). J.Pediatr. 96, 708-715.
2. Chanock, R.M., Parrot, R.H., Connors, M., Collins, P.L. and Murphy, B.R. (1992) Pediatrics 90, 137-142.
3. Martin, A.J. Gardiner, P.S. and McQuillin, J. (1978). Lancel ii, 1035-1038.
4. Robbins, A., and Freeman, P. (1988) Sci. Am. 259, 126-133.
5. Glezen, W.P., Taber, L.H., Frank, A.L. and Kasel, J.A. (1986) Am. J. Dis. Child. 140, 143-146.
6. Katz, S.L. New vaccine development establishing priorities. Vol. 1. Washington: National Academic Press. (1985) pp. 397-409.
7. Wertz. G.W., Sullender, W.M. (1992) Biotech. 20, 151-176.
8. McIntosh, K. and Chanock, R.M. (1990) in Fields Virology (Fields, B.M., and Knipe, D.M. eds.) pp.1045-1075, Raven Press, Ltd., New York.
9. Collins, P., McIntosh, K., and Chanock, R.M. in "Fields Virology" ed. by Fields, B.N., Knipe, D.M., and Howley, P.M., Lippincott-Raven Press, New York, (1996) pp. 1313-1351.
10. Walsh, E.E., Hall, C.B., Briselli, M., Brandiss, M.W. and Schlesinger, J.J. (1987) J. Infect. Dis. 155, 1198-1204.
11. Walsh, E.E., Hruska, J. (1983) J. Virol. 47, 171-177.
12. Levine, S., Kleiber-France, R., and Paradiso, P.R. (1987) J. Gen. Virol. 69, 2521-2524.
13. Anderson, L.J. Hierholzer, J.C., Tsou, C., Hendry, R.M., Fernie, B.F., Stone, Y. and McIntosh, K. (1985), J.Infect. Dis. 151, 626-633.
14. Johnson, P.R., Olmsted, R.A., Prince, G.A., Murphy, B.R., Alling, D.W., Walsh, E.E. and Collins, P.L. (1987) J. Virol. 61 (10), 3163-3166.
15. Cherrie, A.H., Anderson, К., Wertz, G.W., and Openshaw, P.J.M. (1992) J. Virology 66, 2102-2110.
16. Kim, H.W., Canchola, J.G., Brandt, C.D., Pyles, G.. Chanock, R.M. Jensen, K., and Parrott, r.h. (1969) Amer. J. Epidemiology 89, 422-434.
17. Firedewald, W.T., Forsyth, B.R., Smith, C.B., Gharpure, M.S., and Chanock, R.M. (1968) JAMA 204, 690-694.
18. Walsh, E.E., Brandriss, M.W., Schlesinger, J. J. (1985) J. Gen. Virol. 66, 409-415.
19. Walsh, E.E., Schlesinger, J.J. and Brandriss, M.W. (1984) J. Gen. Virol. 65, 761-766.
20. Routledge, E.G., Willcocks, M.M., Samson, A.C.R., Morgan, L., Scott, R., Anderson, J.J., and Toms, G.L. (1988) J. Gen. Virology 69, 293-303.
21. Fulginiti, V.A., Eller, J.J., Sieber, O.F., Joyner, I.W., Minamitani, M. and Meiklejohn, G. (1969) Am J. Epidemiol. 89 (4), 435-448.
22. Chin, J., Magoffin, R.L., Shearer, L.A., Schieble, J. H. and Lennette, E.H. (1969) Am. J. Epidemiol. 89 (4), 449-463.
23. Kapikian, A.Z., Mitchell, R.H., Chanock, R.M., Shvedoff, R.A. and Stewart, C.E. (1969) Am. J. Epidemiol. 89 (4), 405-421.
24. Kirm, H.W., Arrobio, J.O., Pyles, G., Brandt, C.D. Camargo, E., Chanock, R.M. and Parrott, R.H. (1971) Pediatrics 48, 745-755.
25. Wright, P.F., Belshe, R.B., Kirn, H.W., Van Voris, L.P. and Chanock, r.M. (1982) Infect. Immun. 37 (1), 397-400.
26. Wright, P.F., Chinozaki, Т. and Fleet, W. (1976) J. Pediatr. 88, 931-936.
27. Belshe, R.B., Van Voris, P. and Mufson, M.A. (1982) J. Infect. Dis. 145, 311-319.
28. Murphy, B.R., Prince, G.A., Walsh, E.E., Kim, H.W., Parrott, R.H., Hemming V.G., Rodriguez, W.J., and Chanock, R.M.. J. Clin. Microbiol. (1986), 24(2), 197-202.
29. Connors, M., Collins, P.L., Firestone, C.Y., Sotnikov, A.V., Waitze, A., Davis, A.R., Hung, P.P., Chanock, R.M., Murphy, b. (1992) Vaccine, 10, 475-484.
30. Prince, G.A., Jenson, A.В., Hemming, V.G., Murphy, E R., Walsh, E.E., Horswood, R.L. and Chanock, R.L. (1986b) J. Virol. 57 (3), 721-728.
31. Piedra, P.A., Camussi, F. and Ogra, P.L. (1989) J. Gen. Virol. 70, 325-333.
32. Walsh, E.E., Hall, С.В., Briselli, M., Brandiss, M.W. and Schlesinger, J.J. (1987) J. Infect. Dis. 155 (6), 1198-1204.
Реферат
Изобретение относится к области биотехнологии. Белок слияния (F), белок прикрепления (G) и матричный белок (М) респираторно-синцитиального вируса (RSV) выделяют из респираторно-синцитиального вируса путем экстракции белков слабым детергентом из концентрированного вируса и чистят, загружая белок в колонку с гидроксиапатитным или другим носителем и элюируя белок с применением мягкой солевой обработки. Белки F, G и М, приготовленные в виде иммуногенных композиций, безопасны и высоко иммуногенны. 3 с. и 15 з.п. ф-лы, 9 табл., 4 ил.
Комментарии