Химерные флавивирусные вакцины - RU2209082C2
Код документа: RU2209082C2
Чертежи
















Описание
Настоящее изобретение касается аттенуированных вирусов, применимых в качестве вакцин против заболеваний, вызываемых флавивирусами.
Некоторые представители семейства флавивирусов представляют реальную или потенциальную угрозу здоровью населения планеты в целом. Например, японский (или комариный) энцефалит является серьезной проблемой для здоровья населения, включая миллионы жителей Дальнего Востока, подвергающихся риску этого заболевания. Вирус лихорадки денге при частоте до 100 миллионов случаев первичной лихорадки денге и более 450 тысяч случаев геморрагической лихорадки денге в мире является одним из наиболее серьезных заболеваний человека, передаваемых членистоногими переносчиками. Другие флавивирусы также обусловливают ряд эндемичных заболеваний различной природы и способны завоевывать новые регионы вследствие изменений климатических условий, динамики популяций переносчиков и нарушений естественной среды, обусловливаемых деятельностью человека. Эти флавивирусы включают, например, вирус энцефалита Сент-Луиса, который вызывает спорадическое, но серьезное острое заболевание на среднем востоке, юго-востоке и западе США; вирус западного Нила, который вызывает лихорадочное состояние, сопровождающееся острым энцефалитом, которое широко распространено в Африке, на Среднем Востоке, в бывшем СССР и в части районов Европы; вирус энцефалита долины Мюррей, который обусловливает локальную нейропатологию, распространен в Австралии; и вирус клещевого энцефалита, который распространен на территории бывшего Советского Союза и в Восточной Европе, где распространены его переносчики - иксодовые клещи, - который вызывает тяжелую форму энцефалита в этом регионе.
Вирус гепатита С (HCV) является еще одним членом семейства флавивирусов: он характеризуется организацией генома и параметрами размножения, сходными, но не идентичными по сравнению с описанными выше флавивирусами. Вирус HCV по большей части передается по парентеральному типу, он обусловливает хронический гепатит, который может перерастать в цирроз печени и гепатоклеточную карциному, а в США является ведущей причиной таких поражений печени, которые определяют необходимость ортотопической трансплантации печени.
Семейство вирусов Flaviviridae обособлено от альфавирусов (таких как WEE, VEE, EEE, SFV и т.п.) и в настоящее время насчитывает три рода - собственно флавивирусы, пестивирусы и вирусы гепатита С. Полностью сформировавшиеся зрелые вирионы флавивирусов состоят из трех структурных белков - оболочечного (Е), капсидного (С) и мембранного (М), а также семи неструктурных белков (NS1, NS2A, NS2B, NS3, NS4A, NS4B и NS5). У незрелых флавивирусов, обнаруживаемых в инфицированных клетках, выявлен премембранный белок (рrМ), который является предшественником белка М.
После связывания вирионов на рецепторах клеток-хозяев белок Е претерпевает необратимое конформационное преобразование под воздействием кислой среды эндосом, вследствие чего обусловливается связывание бислоя оболочки вириона и эндоцитозных пузырьков, в результате вирусный геном переходит в цитоплазму клетки-хозяина. Вирусы клещевого энцефалита (ТВЕ), содержащие рrМ, являются некомпетентными по такому связыванию: это определяет то, что протеолитическое расщепление предшественника рrМ является необходимым для появления компетентных по связыванию вирионов и обладающих полной инфекционностыо вирусов (Guirakhoo, et al., 1991, J. General Virol, 72, Pt. 2:333-338). Воздействием хлоридом аммония на поздних этапах вирусной репликации были получены вирусы энцефалита долины Мюррей (MVE), содержащие рrМ: было показано, что они некомпетентны по слиянию. При использовании специфичных по последовательности пептидов и моноклональных антител было показано, что пропептид рrМ взаимодействует с аминокислотами 200-327 в составе белка Е. Такое взаимодействие является необходимым для защиты белка Е от необратимой конформационной перестройки, определяемой процессом созревания в кислых пузырьках в экзоцитозном метаболическом механизме (Guirakhoo et al., 1992, Virology, 191:921-931).
Расщепление предшественника рrМ с образованием белка М происходит незадолго до выхода вирионов с участием фурино-подобной клеточной протеазы (Stadler et al., 1997, J. Virol, 71:8475-8481), что является необходимым для включения гемагглютинационной активности, фузогенной активности и инфекционности самих вирионов. Белок М отщепляется от своего полипептидного предшественника (рrМ) за консенсусной последовательностью R-X-R/K-R (где Х - произвольная аминокислота, К - лизин, R - аргинин) и включается в состав липидной вирусной оболочки вместе с белком Е.
Последовательности, по которым происходит процессинг, являются консервативными не только у флавивирусов, но также и в составе белков других, не близкородственных вирусов, таких как белки РЕ2 коронавирусов мышей, РЕ2 альфавирусов, НА вирусов гриппа и р160 ретровирусов. Расщепление полипептида-предшественника является существенным для инфекционности вируса, но не для образования его вирионов. Было показано, что в случае с вирусной химерой ТВЕ/денге-4 изменение сайта процессинга рrМ приводит к снижению нейровирулентности этой химеры (Pletnev et al., 1993, J. Virol. 67:4956-4963), что согласуется с более ранними данными о том, что эффективный процессинг предшественника рrМ является необходимым для проявления полной инфекционности (Guirakhoo et al., 1991, цит. выше; 1992, цит. выше; Heinz et al., 1994, Virology, 198:109-117). Антитела к полипептиду рrМ могут опосредовать проявление иммунитета, очевидно, благодаря нейтрализации выходящих вирионов, включающих непроцессированный рrМ. Сайт протеолитического расщепления белка РЕ2 вируса VEE (включает 4 аминокислоты) был делегирован с применением направленного мутагенеза у инфекционного клона (Smith et al., 1997, ASTMH Meet., Dec. 7-11). Делеционные мутанты реплицировались с высокой эффективностью, а белки РЕ2 инкорпорировались в состав вирионов. При анализе этого мутанта у нечеловекообразных обезьян и было показано проявление 100%-ной сероконверсии и проявление защищенности всех иммунизованных обезьян от летального исхода.
Изобретение представляет химерные живые инфекционные аттенуированные вирусы, которые включают:
(a) первый вирус желтой
лихорадки (например, штамма 17D), представляющий живой аттенуированный
вакцинный вирус, в геноме которого нуклеотидная последовательность, кодирующая белок рrМ-Е либо делетирована, либо укорочена,
либо мутирована таким образом, что функциональный белок рrМ-Е первого
флавивируса не экспрессируется; и
(b) интегрированная в геном первого флавивируса нуклеотидная последовательность,
кодирующая вирусный оболочечный компонент (белки рrМ-Е) второго,
отличающегося флавивируса, таким образом, что белок рrМ-Е второго флавивируса экспрессируется из состава измененного генома первого
флавивируса.
Таким образом, химерный вирус включает гены и кодируемые продукты, необходимые для внутриклеточного размножения, относящиеся к первому флавивирусу, и гены и кодируемые продукты оболочки второго флавивируса. Следовательно, вирус, содержащий все антигенные детерминанты, ответственные за индукцию нейтрализующих антител, в результате заражения химерным вирусом, в этом случае будет генерировать антитела только в отношении второго флавивируса.
Предпочтительным живым вирусом, предназначенным для использования в качестве первого флавивируса в составе химеры по настоящему изобретению, является вирус желтой лихорадки. По крайней мере одна из известных вакцин использует такой живой ослабленный вирус: эта вакцина известна под маркой YF17D и используется для вакцинации человека уже более 50 лет. Вакцина YF17D охарактеризована в большом числе публикаций, включая материалы Смитберна с соавт. и Фристоуна (Smithburn et al., 1956, "Yellow Fever Vaccination", World Health Org., p. 238; Freestone, 1995, In "Vaccines", 2d ed., eds Plotkin et al. , W. B. Saunders, PA). Кроме того, вирус желтой лихорадки был изучен на генетическом уровне (Rice et al. , 1985, Science, 229:726-733), а также представлялась информация о соотношении генетических и фенотипических параметров (Marchevsky et al., 1995, Amer. J. Trop. Med. Hyg., 52:75-80).
Предпочтительными флавивирусами для использования в качестве второго флавивируса в химерах по настоящему изобретению, соответственно, являющиеся источниками иммунизующего антигена, являются вирус японского энцефалита (JE), вирус лихорадки денге (DEN, например, любой из типов денге 1-4), вирус энцефалита долины Мюррей (MVE), вирус энцефалита Сент-Луиса (SLE), вирус лихорадки западного Нила (WN), вирус клещевого энцефалита (ТВЕ) и вирус гепатита С (HCV). Кроме того, флавивирусами, пригодными для использования в качестве второго флавивируса, являются вирус Куньинь, вирус центрально-европейского энцефалита, вирус весенне-летнего клещевого энцефалита, вирус повассанской лихорадки, вирус киасанурской лесной болезни и вирус омской геморрагической лихорадки. У предпочтительного химерного вируса по настоящему изобретению последовательность, кодирующая белок рrМ-Е второго флавивируса, используется для замещения последовательности, кодирующей белок рrМ-Е, в составе генома первого живого вируса желтой лихорадки. У предпочтительного химерного вируса последовательность, кодирующая белок рrМ-Е, является производной от ослабленного вирусного штамма, такого как вакцинный штамм. Также, как это будет описание далее, область рrМ в составе такого белка может нести мутацию, которая предотвращает процессинг с образованием зрелого мембранного белка.
Также настоящее изобретение представляет способы предотвращения или лечения флавивирусной инфекции у млекопитающих, таких как человек, путем введения химерного флавивируса по настоящему изобретению этому млекопитающему, использование химерных флавивирусов по настоящему изобретению при приготовлении лекарственных препаратов, предназначенных для профилактики и лечения флавивирусных инфекций, молекулы нуклеиновых кислот, кодирующих химерные флавивирусы по настоящему изобретению, и способы производства химерных флавивирусов по настоящему изобретению.
Изобретение имеет ряд преимуществ. Например, благодаря тому что химерные вирусы по настоящему изобретению являются живыми и способными к размножению, они могут быть использованы для формирования долговременного иммунитета. Поскольку эти вирусы включают в геноме репликационные гены аттенуированного вируса (например, вируса желтой лихорадки штамма 17D), то химерный вирус аттенуирован до такой степени, которая делает его безопасным при использовании в отношении человека.
Другие параметры и преимущества настоящего изобретения будут ясны из нижеследующего подробного описания, чертежей и формулы изобретения.
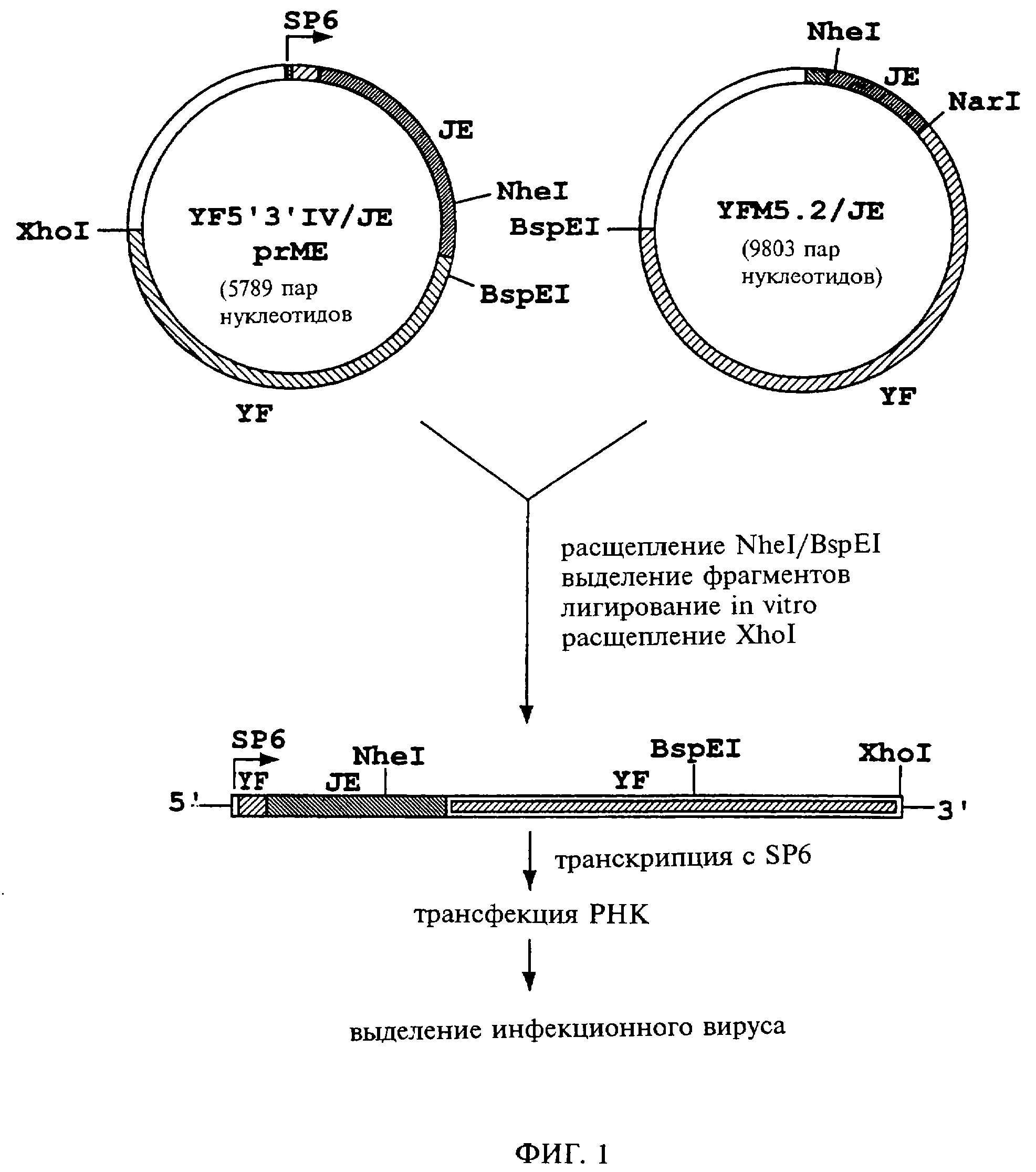
Фиг. 1 схематически представляет этапы генетических манипуляций, которые были осуществлены с целью конструирования химерного вируса по настоящему изобретению, включающего вирус желтой лихорадки и вирус японского энцефалита (YF/JE).
Фиг. 2 показывает кривые роста химерных вирусов YF/JE по настоящему изобретению в клеточных культурах, пригодных для получения человеческой вакцины.
Фиг. 3 - график, показывающий сравнение роста вирусов RMS (Research Master Seed, YF/JE SA14-14-2) и YF-Vax в клетках линии MRC-5.
Фиг.4 - график и таблица, показывающие результаты анализа нейровирулентности химерного вируса YF/JE по настоящему изобретению, проведенного на мышах.
Фиг.5 является схематическим представлением двухплазмидной системы, предназначенной для создания химерного вируса YF/DEN-2. По сути, эта стратегия соответствует той стратегии, которая описана в связи с созданием химерного вируса YF/JE.
Фиг. 6 является схематическим представлением структуры модифицированных клонов YF, сконструированных по делетированию части белка NS1 и (или) экспрессии чужеродных белков под контролем собственного сайта входа в рибосому (IRES). Фигура показывает только часть E/NS1 генома вируса. Стоп-кодон внедрен в участок, кодирующий С-конец оболочечного белка Е. Трансляция по направлению вниз инициируется в пределах межгенной открытой рамки считывания (ORF) действием фактора IRES-1, тем самым определяя экспрессию чужеродных белков (например, белков Е1 и (или) Е2 вируса HCV). Второй фактор IRES (IRES-2) контролирует инициацию трансляции неструктурного сегмента вируса желтой лихорадки, с которого экспрессируются плотно упакованные укороченные белки NS1 (например, NSldel-1, NSldel-2 или NSldel-3). Размер делеции в составе NS1 обратно пропорционален размеру ORF, соединенной с IRES-1.
Фиг.7 - график, показывающий ответ по нейтрализации антител у мышей, иммунизованных химерной вакциной YF/JE SA14 -14-2.
Настоящее изобретение представляет химерные флавивирусы, которые могут быть использованы в вакцинации против флавивирусных инфекций. Конструирование и анализ химерных флавивирусов по настоящему изобретению, таких как химер, состоящих из вируса желтой лихорадки и вируса японского энцефалита (JE), вируса лихорадки денге 1-4 (DEN 1-4), вируса энцефалита долины Мюррей (MVE), вируса энцефалита Сент-Луиса (SLE), вируса лихорадки западного Нила (WN), вируса клещевого энцефалита (ТВЕ) и вируса гепатита С (HCV) осуществляли следующим образом.
Флавивирусные белки получали путем трансляции единственной длинной открытой рамки (кодирующей, например, структурные белки - капсидный (С), предшественник мембранного (рr-М) и оболочечный (Е), а также неструктурные белки) и затем путем серии посттрансляционных процессинговых этапов протеолитического расщепления. Химерные флавивирусы по настоящему изобретению, как это обсуждалось выше, характеризуются замещением последовательностей, кодирующих белки рr-М и Е одного флавивируса, последовательностями, кодирующими белки рr-М и Е другого флавивируса. Таким образом, создание таких химерных флавивирусов включает формирование новых комбинаций капсидных и премембранных белков, с одной стороны, и оболочечных и неструктурных белков (NS1), с другой стороны, происходящих от двух разных флавивирусов. Расщепление между участками каждой из этих двух групп белков (С и рr-М, с одной стороны, и Е и NS1) происходит в ходе естественного протеолитического процессинга флавивирусных белков и нуждается в наличии сигнальных последовательностей, фланкирующих данное соединение сайтов расщепления.
Предпочтительно, чтобы в составе химерных флавивирусов по настоящему изобретению вирусные сигнальные последовательности обеспечивали химерам существенную стабильность на таком уровне, чтобы точное по месту расщепление белков между сегментами С/рr-М и E/NS1 было эффективным. Эти сигнальные последовательности, поддерживающие химеры, описаны ниже. С другой стороны, любая из многочисленных известных сигнальных последовательностей может быть использована при конструировании так, чтобы в составе химер соединять последовательности, кодирующие белки С и рr-М или Е и NS1 (см., например, von Heijne, 1983, Eur. J. Biochem., 133:17-21; von Heijne, 1985, J. Mol. Biol., 184: 99-105), или, например, используя известные регуляторные последовательности, специалист в данной области техники может сконструировать дополнительные сигнальные последовательности, которые могут быть использованы в химерах по настоящему изобретению. Обычно, например, сигнальная последовательность должна включать в качестве своего последнего остатка аминокислоту с небольшой незаряженной боковой цепью, такую как аланин, глицин, серин, цистеин, треонин или глутамин. Другие требования, предъявляемые к сигнальным последовательностям, хорошо известны в данной области техники (см., например, von Heijne, 1983, цит. выше; von Heijne, 1985, цит. выше). Также сигнальные последовательности любого вируса, входящего в состав химеры, могут быть сохранены полностью или сохранены в той степени, которая обеспечит точное расщепление.
Конструирование матриц
кДНК для создания химерного вируса YF/JE
Получение полноразмерных матриц кДНК для химер YF/JE по
настоящему изобретению, описанное ниже, основано на стратегии, сходной с той, которую ранее
использовали разработчики процесса регенерации YF17D из материала кДНК при молекулярно-генетическом изучении
процесса размножения вируса желтой лихорадки. Эта стратегия описана, например,
Несторовичем с соавт. (Nestorowicz et al., 1994, Virology, 199:114-123).
Вкратце, получение химеры YF/JE по настоящему изобретению включает следующее. Геномные последовательности вируса желтой лихорадки встраивают в состав двух плазмид (YF5'3'IV и YFM5.2), которые кодируют последовательности YF от нуклеотидов 1-2276 и 8279-10861 (YF5'3'IV) и от 1373-8704 (YFM5.2) (Rice et al., 1989, New Biologist, 1:285-296). Полноразмерные матрицы кДНК получают путем лигирования подходящих рестрикционных фрагментов, полученных на материале этих плазмид. Данный способ является наиболее пригодным с точки зрения обеспечения стабильной экспрессии последовательностей YF и получения РНК-транскриптов, обладающих высоким уровней специфической инфекционности.
Стратегия заявителей, использовавшаяся ими при конструировании химер, включает замещение последовательностей вируса YF в составе плазмид YF5'3'IV и YFM5.2 соответствующими последовательностями из генома JE от начала белка рrМ (478-й нуклеотид; 128-я аминокислота) через сайт расщепления E/NS1 (2452-й нуклеотид; 817-я аминокислота). В дополнение к клонированию кДНК JE были необходимы некоторые этапы, обеспечивающие встраивание или уничтожение рестрикционных сайтов в состав последовательностей и YF, и JE, нужных для осуществления лигирования in vitro. Структура матрицы, предназначенной для регенерирования химерного вируса YF(C)/JE (рrМ-Е), показана на фиг.4. С применением сходной стратегии была сконструирована вторая химера, кодирующая полный структурный компонент (С-рrМ-Е) вируса японского энцефалита.
Молекулярное клонирование структурного компонента вируса JE
Клоны аутотентичных
генов, кодирующих структурные белки вируса японского энцефалита, были
получены на материале штамма JE SA14-14-2 (живой аттенуированный вакцинный штамм вируса JE), потому что биологические
характеристики и молекулярные параметры этого штамма хорошо изучены
(см., например, Eckels et al. , 1988, Vaccine, 6:513-518; вирусный штамм JE SA14-14-2 доступен из Центра по контролю за
заболеваниями в Форт-Коллинзе, Колорадо и из Арбовирусного
исследовательского центра Йельского университета, Нью-Хейвен, Коннектикут, США, которые являются официальными в США референсными центрами
изучения арбовирусов Всемирной организации здравоохранения).
Был получен вирусный штамм JE SA14-14-2, находящийся на культивационом уровне PDK-5: его пересеивали в клетки LLC-MK2
с целью получения большого количества вируса, необходимого
для клонирования кДНК. Использованная заявителями стратегия включала клонирование структурного сегмента в двух частях, которые перекрываются
по рестрикционному сайту Nhel (1125-й нуклеотид генома JE),
который затем может быть использован для осуществления лигирования in vitro.
Пулы РНК экстрагировали из монослойных культур инфицированных клеток LLC-MK2 и синтез первой цепи антисмысловой кДНК проводили с использованием обратной транскриптазы с антисмысловым праймером (нуклеотиды 2456-2471 нуклеотидной последовательности генома JE), несущего сгруппированные рестрикционные сайты XbaI и NarI, необходимые, соответственно, для клонирования сначала в состав вектора pBluescript-II KS(+) и затем в состав плазмиды YFM5.2 (NarI). После синтеза первой цепи кДНК проводили амплификацию с помощью ПЦР последовательности JE в нуклеотидах 1108-2471, используя те же антисмысловой праймер и смысловой праймер (нуклеотиды 1108-1130 нуклеотидной последовательности генома JE), включающие сгруппированные рестрикционные сайты XbaI и NsiI, необходимые для клонирования в составе, соответственно, pBluescript и YFM5.2 (NarI). Последовательности JE были верифицированы путем рестрикционного расщепления и прямого нуклеотидного секвенирования. Нуклеотидная последовательность генома JE в нуклеотидах 1-1130 была получена с помощью ПЦР-амплификации кДНК негативной цепи JE с использованием антисмыслового праймера, соответствующего нуклеотидам 1116-1130 генома JE, и смыслового праймера, соответствующего нуклеотидам 1-18 генома JE: оба они содержат сайты рестрикции EcoRI. Полученные ПЦР-фрагменты клонировали в состав pBluescript, а последовательности JE верифицировали путем прямого нуклеотидного секвенирования. Все указанное вместе представляет клонирование последовательности генома JE в нуклеотидах 1-2471 (аминокислоты 1-792).
Конструирование производных вариантов YF5'3'IV/JE
и YFM5.2/JE
Для включения С-концевой части
оболочечного белка JE в сайт расщепления YF E/NS1, уникальный рестрикционный сайт Narl был внесен с состав плазмиды YFM5.2, с использованием
олигонуклеотид-направляемого мутагенеза сигнальной
последовательности в пределах сайта расщепления E/NS1 (нуклеотиды YF 2447-2452; аминокислоты 816-817) с получением YFM5.2 (NarI). Транскрипты,
считанные с таких матриц, несущих такое изменение,
тестировали по их инфекционности: они проявили специфическую инфекционность, сходную с родительскими матрицами (приблизительно 100 бляшкообразующих
единиц на 250 нг транскриgта). Последовательность
JE в нуклеотидах 1108-2471 была субклонирована из состава нескольких независимо полученных ПЦР-клонов pBluescript/JE в состав плазмиды YFM5.2 (NarI) с
использованием уникальных рестрикционных сайтов
NsiI и NarI. Клоны YF5'3'IV/JE, включающие 5'-нетранслируемый сегмент вируса желтой лихорадки (нуклеотиды 1-118), соседствующий с участком кодирования
prM-E JE, были получены с помощью
ПЦР-амплификации.
Для получения последовательностей, включающих соединение капсидного кода YF и белка prM JE, антисмысловой праймер, охватывающий этот сегмент, был использован наряду со смысловым праймером, соответствующим нуклеотидам 6625-6639 в составе YF5'3'IV, с целью формирования ПЦР-фрагментов, которые затем использовали в качестве антисмысловых праймеров для ПЦР в сочетании со смысловыми праймерами, комплементарными последовательности вектора pBluescript, расположенными выше сайта рестрикции EcoRI, с целью амплификации последовательности JE (кодируемой в обратной ориентации в составе вектора pBluescript) от 477-го нуклеотида (N-конец белка prM) через имеющийся внутри сайт рестрикции NheI до 1125-го нуклеотида. Получаемые в результате ПНР-фрагменты встраивали в плазмиду YF5'3'IV с использованием рестрикционных сайтов NotI и EcoRI. Эта конструкция включает промотор SP6, предваряющий 5'-нетранслируемый сегмент YF, и далее такую последовательность: коды YF (С) JE (рrМ-Е), также включая рестрикционный сайт NheI (1125-й нуклеотид JE), необходимый для лигирования in vitro.
Конструирование YFM5.2
и YF5'3'IV,
включающих рестрикционные сайты, необходимые для лигирования in vitro
С целью использования рестрикционного сайта NheI в пределах последовательности, кодирующей оболочечный белок
JE, в
качестве сайта лигирования in vitro, избыточный сайт NheI в составе плазмиды YFM5.2 (5459-й нуклеотид) был удален. Это обеспечивалось несмысловой мутацией последовательности YF по нуклеотиду
5461
(Т-->С: аминокислотный смысл - аланин, 1820-я аминокислота). Этот сайт включали в состав YFM5.2 путем лигирования подходящих рестрикционных фрагментов, а в состав YFM5.3(NarI)/JE вносили
путем
замены фрагмента NsiI/NarI, кодирующего химерную последовательность YF/JE.
Для создания уникального 3'-рестрикционного сайта, предназначенного для лигирования in vitro, сайт BspEl был сформирован так, чтобы располагаться ниже сайта AatII, в норме используемого для создания полноразмерных матриц на материале плазмид YF5'3'IV и YFM5.2. (В последовательности, кодирующей структурные компоненты вируса JE, присутствуют множественные рестрикционные сайты AatII, что не позволяет использовать их для целей лигирования in vitro). Сайт рестрикции BspEl создавали путем несмыслового мутирования 8581-го нуклеотида YF (A-->С: аминокислотный смысл - серин, 2860-я аминокислота) и вносили в состав YFM5.2 путем замены подходящих рестрикционных фрагментов. Уникальный рестрикционный сайт вносили в состав YFM5.2/JE путем замены фрагмента XbaI/SphI, а в состав плазмиды YF5'3'IV/JE(рrМ-Е) путем 3-компонентного лигирования подходящих рестрикционных фрагментов, происходящих из этих исходных плазмид и из производной плазмиды YFM5.2 (BspI) с делетированной последовательностью YF, находящейся между сайтами EcoRI в нуклеотидах 1 и 6912.
Обмен
кДНК JE-Nakayama в состав химерных плазмид YF/JE
Из-за неочевидности того, способен ли правильно функционировать полученный с помощью ПЦР структурный сегмент JE SA14-14-2 в
контексте последовательностей химерного вируса, заявители использовали кДНК клона, представляющего штамм Nakayama вируса японского энцефалита, который был детально исследован в экспериментах по
экспрессии и по его способности индуцировать выработку иммунитета к нему (см., например, McIda et al., 1987, Virology, 158:348-360; штамм JE-Nakayama доступен из Центра по контролю за заболеваниями в
Форт-Коллинзе, Колорадо и из Арбовирусного исследовательского центра Йельского университета, Нью-Хейвен, Коннектикут, США). кДНК JE-Nakayama встраивали в состав химерных плазмид YF/JE с
использованием
имеющихся рестрикционных сайтов (HindIII к PvuII и BpmI к MunI) с целью замещения полного участка, кодирующего рrМ-Е в составе двухплазмидной системы, за исключением единственной
аминокислоты в 49-м
положении (серин), которую оставляли интактной с целью использования рестрикционного сайта NheI для лигирования in vitro. Полный сегмент JE в клоне Nakayama был секвенирован с
целью проверки
аутентичности замещающей кДНК (табл. 2).
Создание полноразмерных матриц кДНК, РНК-трансфекция и получение инфекционных вирусов
Процедуры создания
полноразмерных кДНК-матриц в
принципе соответствуют тому, что описано Райсом с соавт. (Rice et al., 1989, New Biologist, 1: 285-296) (см. фиг.1). В случае химерных матриц плазмиды YF5'3'IV/JE(рrМ-Е)
и YFM5.2/JE расщепляют
рестриктазами NheI/BspEI и лигирование in vitro проводят с использованием 50 нг очищенных фрагментов в присутствие ДНК-лигазы фага Т4. Продукты лигирования линеаризуют с
помощью XhoI для обеспечения
свободной транскрипции. Транкрипты с промотора SP6 синтезируют с использованием 50 нг очищенной матрицы (количественный контроль осуществляют включением меченного3H-УТФ) и целостность РНК
проверяют с использованием электрофореза в неденатурирующем агарозном геле. Выход колеблется в пределах 5-10 мкг РНК на 1 реакцию при использовании данной процедуры,
причем большинство материала
представлено полноразмерными транскриптами. Трансфекцию РНК-транскриптов в присутствие катионных липосом осуществляют так, как это описано Райсом с соавт. (цит. выше) для
YF17D. В исходных
экспериментах были использованы клетки LLC-MK2 для трансфекции и количественного определения вируса, на материале которых заявители определяли пригодность этих клеток для
размножения вируса
и бляшкообразования тестированием родительских штаммов YF и JE. В табл. 1 показаны типичные результаты трансфекционных экспериментов с использованием липофектина (Gibco/BRL) в
качестве
трансфекционного носителя. Также клеточные линии Vero были использованы для получения инфекционных популяций вирусов, оценки меченых белков и нейтрализационных тестов.
Секвенирование
нуклеотидной последовательности химерных кДНК-матриц
Для идентификации точных последовательностей оболочечного белка штамма SA14-14-2 и Nakayama проводили анализ
нуклеотидной
последовательности плазмид, включающих химерную кДНК YF/JE, в отношении клонов JE-сегмента. Различия между нуклеотидными последовательностями у этих конструкций в сравнении с
опубликованными
последовательностями (McAda et al., цит. выше) показаны в табл. 2.
Структурная и биологическая характеристика химерных вирусов YF/JE
Геномная структура
химерных вирусов YF/JE,
полученных в экспериментах по трансфекции, была проверена с использованием метода полимеразной цепной реакции с ревертированием в отношении вирусной РНК, выделенной из
монослоев инфицированных клеток.
Эти эксперименты проводят с целью предотвращения вероятности того, что популяции вирусов были загрязнены в ходе процедуры
трансфекции. Для этих экспериментов вирусы
первого пассажа были использованы
для инициирования цикла заражения с целью предотвращения любых артефактов, которые могут быть вызваны присутствием остаточной трансфицированной вирусной РНК. Общие
пулы РНК, экстрагированные из клеток,
инфицированных химерами либо YF/JE(prM-E)-SA14-14-2 либо YF/JE(prM-E)-Nakayama, подвергали ПЦР с ревертированием, используя специфичные для YF и JE
праймеры, которые позволяют выделять
весь структурный сегмент в виде двух ПЦР-продуктов длиной примерно 1000 нуклеотидов. Эти продукты затем анализировали путем рестриктазного расщепления по
предполагаемым сайтам, имеющимся в составе
последовательностей JE штаммов SA14-4-14-2 и Nakayama и обеспечивающим дифференцирование этих вирусов. С использованием такого подхода была
продемонстрирована химерная природа вирусной РНК,
а для выделенных вирусов было подтверждено наличие соответствующих рестрикционных сайтов. Затем была верифицирована реальная граница С-рrМ, которая
оказалась интактной на уровне данной
последовательности - было применено секвенирование участка соединения в составе химеры YF/JE С-рrМ.
Присутствие оболочечного белка JE в этих двух химерах было проверено с помощью иммунопреципитации антисывороткой, специфичной в отношении JE, и в нейтрализационном тесте по проявлению бляшек с использованием антисывороток, специфичных в отношении YF и JE. Иммунопреципитация35S-меченых экстрактов клеток LLC-MK2, инфицированных данными химерами, с использованием моноклонального антитела к белку Е вируса JE, показала, что оболочечный белок Е вируса JE может быть выделен в виде белка с молекулярной массой 55 кДа, в то время как та же антисыворотка не способна иммунопреципитировать какой-либо белок из состава клеток, инфицированных вирусом желтой лихорадки. Обе гипериммунные сыворотки к YF и JE проявили перекрестную реактивность по отношению к двум оболочечным белкам, однако при этом в ряде повторностей воспроизводится размерная дифференцированность двух белков: у вируса YF - 53 кДа, негликозилирован; у вируса JE 55 кДа, гликозилирован. Использование моноклональных антител к YF в условиях иммунопреципитации оказалось неэффективным: следовательно, специфичность зависела в данном анализе от моноклональных антител к JE. Нейтрализационный тест по снижению числа бляшек (PRNT) был осуществлен в отношении химерных вирусов и "чистых" вирусов YF и JE SA14-14-2 с использованием гипериммунной асцитной жидкости (АТСС), специфичной в отношении YF и JE, и очищенного иммуноглобулина-G, специфичного в отношении YF (моноклональное антитело 2 Е10). Для этих антисывороток были выявлены существенные различия в их титре, необходимом для 50%-снижения числа бляшек при тестировании химер в сравнении с контрольными ("чистыми") вирусами по данному эксперименту (табл. 3). Таким образом, необходимые для нейтрализации эпитопы экспрессируются инфекционными химерными вирусами YF/JE.
Параметры роста в клеточной культуре
Способность химер к росту была исследована по количественным параметрам в линиях клеток, происходящих как от приматов,
так и от комаров. На фиг.2 показаны
кумулятивные кривые роста химер в клетках линии LLC-MK2 после множественного заражения с низкой плотностью (0,5 бляшкообразующей единицы на клетку). В
этом эксперименте для сравнения были
использованы вирусы YF5.2IV (клонированный дериват) и JE SA14-14-2 (неклонированный). Оба химерных вируса достигали максимального выхода вирусов на
уровне, приблизительно на 1 логарифм выше,
чем выход любого из родительских вирусов. В случае с химерой YF/JE SA14-14-2 пик выхода вируса был отмечен на 12 часов позднее, чем тот же
показатель у химеры YF/JE Nakayama
(соответственно, 50 и 38 часов). Химера YF/JE Nakayama проявляла существенно больший цитопатический эффект в сравнении с химерой YF/JE SA14-14-2 в той же
клеточной линии. Сходный
эксперимент был проведен с клетками С 6/36 при множественном заражении с низкой плотностью (0,5 бляшкообразующей единицы на 1 клетку). На фиг.2 также показана динамика роста
вирусов в этой линии
клеток беспозвоночного. Сходные уровни выхода вирусов были отмечены во все моменты тестирования в данном эксперименте, что дополнительно подтверждает заключение о том, что у
химерных вирусов
эффективность размножения нисколько не нарушена.
Сравнение динамики роста RMS (YF/JE SA14-14-2) и вакцины YF17D в клетках MRC-5
Эксперимент был
проведен с целью
оценки способности предполагаемой вакцины размножаться в клеточной линии, приемлемой для вакцин, предназначенных для человека. Коммерческая вакцина против желтой лихорадки YF17D
(YF-Vax®) была получена от Connaught Laboratories, Swiftwater, PA. Клетки MRC-5 (диплодные клетки эмбриональных легких человека) были приобретены в АТСС (171-CCL, Batch F-14308:
18-й пассаж): их
выращивали в культуральной среде ЕМЕМ с добавлением 2 мМ L-глутамина, солевого раствора Эрла, вносимого для доведения до 1,5 г/л по бикарбонату натрия, 0,1 мМ заменимых аминокислот и
10% фетальной
сыворотки телят.
Для сравнения кинетики роста RMS (Research Master Seed, YF/JE SA14-14-2) и YF-Vax® клетки выращивали до 90%-ной конфлюэнтности и проводили их заражение RMS или YF-Vax® с плотностью (MOI) 0,1 б.о.е. С учетом того, что клетки MRC-5 обычно растут медленно, эти клетки выдерживали после инфицирования в течение 10 дней. Образцы замораживали на 7-10 дней и их инфекционность определяли в тесте на формирование бляшек с использованием клеток Vero.
Вакцина YF-Vax® и химера YF/JE вырастали в клетках MRC-5 до умеренных титров (фиг.3). Пиковый титр составил примерно 4,7 log10 б.о.е. для YF-Vax® (он достигался на 2-й день), а для RMS был ненамного ниже - 4,5 log10 б.о.е. на 6-й день.
Тестирование нейровирулентности у нормальных взрослых мышей
Свойства вирулентности химеры YF/JE
SA14-14-2 были проанализированы
с использованием молодых взрослых мышей: применена интрацеребральная инокуляция. Группы по 10 мышей (самцы и самки линии ICR в возрасте 4 недель - по 5
особей каждого пола в каждой группе) были
инокулированы 10 тысячами бляшкообразующими единицами химеры YF/JE SA14-14-2, YF5.2IV или JE SA14-14-2 и наблюдали ежедневно в течение
3 недель. Результаты этих экспериментов
проиллюстрированы на фиг.4. Мыши, зараженные родительским штаммом YF5.2IV, погибали спустя примерно одну неделю после заражения. Смертность или признаки
заболевания не были обнаружены у мышей,
зараженных штаммом японского энцефалита JE SA14-14-2 или химерой. Инокуляты, использовавшиеся в этих экспериментах, были оттитрованы в момент
заражения, а подгруппы выживающих мышей
тестировали на присутствие нейтрализующих антител, что было необходимо для подтверждения того, что заражение имело место. При сравнении тестированных особей
отмечено, что титры антител против JE
SA14-14-2 были сходными с таковыми, которые были выявлены у животных, которые были заражены либо этим штаммом, либо химерным вирусом.
Результаты дополнительных экспериментов, связанных с изучением нейровирулентности химеры YF/JE SA14-14-2 в отношении мышей, проиллюстрированы в табл. 4. В этих экспериментах все мыши, зараженные вирусом желтой лихорадки YF5.2.IV, погибли в течение 7-8 дней. Напротив, ни одна из мышей, зараженных химерным вирусом YF/JE SA14-14-2, не погибла в течение двух недель после заражения.
Результаты экспериментов, посвященных изучению нейровирулентности и патогенности химер YF/JE, проиллюстрированы в табл. 5. В этих экспериментах химерные вирусы использовались для заражения 3-недельных мышей при дозах, варьировавшихся от 10 тысяч до 1 миллиона бляшкообразующих единиц при их внутрибрюшинном введении. Ни одна из мышей, зараженных химерами YF/JE Nakayama или YF/JE SA14-14-2, не погибла в течение 3 недель после заражения: это указывает на то, что эти вирусы не способны вызывать заболевание после периферической инокуляции. У мышей, зараженных химерой YF/JE SA14-14-2, формировались нейтрализующие антитела против вируса японского энцефалита JE (фиг. 7).
Конструирование матриц кДНК, необходимых
для создания химерных вирусов желтой лихорадки/лихорадки
денге (YF/DEN)
Конструирование химерных вирусов желтой лихорадки/лихорадки денге (YF/DEN), описанное ниже, было осуществлено, в
принципе, таким же образом, что и конструирование химерных
вирусов YF/JE, описанное выше. Другие флавивирусные химеры могут быть созданы с помощью аналогичной стратегии с использованием естественных
или искусственно встроенных рестрикционных сайтов и,
например, олигонуклеотидных праймеров, которые показаны в табл. 6.
Конструирование химерного вируса YF/DEN
Хотя некоторые
молекулярные клоны вирусов лихорадки денге были
сформированы ранее, сохраняется проблема обеспечения стабильности фрагментов вирусной кДНК, внесенных в состав плазмидных систем, а также эффективности
размножения полученного вируса. Для эксперимента
заявители выбрали клон DEN-2, сформированный Д-ром Питером Райтом (Dept. Microbiol, Monash Univ., Clayton, Австралия), потому что эта система является
относительно эффективной с точки зрения
регенерирования вируса, а также важно то, что она основана на двухплазмидной системе, сходной с той, которая используется в методологии заявителей. Полная
последовательность этого клона DEN-2 является
доступной и, соответственно, облегчает конструирование химерных матриц YF/DEN, поскольку при этом требуется осуществить лишь незначительные модификации
клона YF. Необходимые для этого шаги являются
следующими.
Аналогично двухплазмидной системе, использовавшейся в отношении вирусов YF5.2IV и YF/JE, система YF/DEN использует уникальный рестрикционный сайт в составе последовательности, кодирующей оболочечный белок Е вируса DEN-2 в качестве точки разрыва для внесения структурного сегмента (рrМ-Е) в состав двух плазмид, которые в данном тексте обозначены как YF5'3'IV/DEN (prM-E') и YFM5.2/DEN (Е'-Е) (см. фиг. 5). Два рестрикционных сайта, необходимые для лигирования in vitro химерной матрицы, маркируются рестриктазами AatII и SphI. Реципиентной плазмидой для 3'-сегмента последовательности гена белка-Е DEN является плазмида YFM5.2 (Narl[+]SphI[-]). Эта плазмида включает рестрикционный сайт NarI в составе участка соединения E/NS1, который использовали для встраивания С-концевой части белка Е вируса JE. Он был далее модифицирован путем элиминации дополнительного рестрикционного сайта SphI в составе последовательности, кодирующей белок NS5, - применен метод направленного несмыслового мутагенеза. Это позволяет встраивать последовательность DEN-2 от уникального сайта SphI до сайта NarI с помощью простейшего направленного клонирования. Подходящий фрагмент кДНК DEN-2 был сформирован с помощью ПЦР на материале производного от DEN-2 клона MON310, предоставленного Д-ром Райтом. Праймеры для ПЦР включали 5'-праймер, фланкирующий рестрикционный сайт SphI, и 3'-праймер, гомологичный нуклеотидам в составе DEN-2, расположенным сразу вверх от сигнального сайта в соединении E/NS1 и замещающим сигнальный сайт за счет замен, которые создают новый такой сайт, но при этом еще и вносят рестрикционный сайт NarI. Полученный в результате ПЦР-фрагмент, состоящий из 1170 нуклеотидов, затем был встроен в состав плазмиды YFM5.2 (NarI[+]SphI[-]).
Сегмент 5' в составе клона DEN-2, включающий последовательности, которые кодируют рrМ и N-часть белка Е, был сконструирован в составе плазмиды YF5'3'IV с использованием химерного праймера для ПЦР. Химерный праймер, включающий 3'-конец антисмысловой последовательности белка С вируса YF и 5'-конец гена белка рrМ DEN-2, был использован наряду со смысловым праймером, фланкирующим промотор SP6 из состава плазмиды YF5'3'IV, с целью создания ПЦР-продукта, состоящего из 771 нуклеотида, характеризующегося 20-парным удлинением, представляющим последовательность рrМ вируса DEN-2. Этот ПЦР-продукт затем был использован для праймирования плазмиды DEN-2 наряду с 3'-праймером, представляющим нуклеотиды 1501-1522 последовательности DEN-2 и фланкирующим рестрикционный сайт SphI, с целью получения конечного ПЦР-продукта, состоящего из 1800 нуклеотидов, включающего последовательность вируса желтой лихорадки YF от рестрикционного сайта NotI через промотор SP6, 5'-нетранслируемый сегмент YF и последовательность, кодирующую белок С, смежную с последовательностью, кодирующей ргМ-Е1522 DEN-2. ПЦР-продукт был лигирован в состав плазмиды YF5'3'IV с использованием рестрикционных сайтов NotI и SphI с получением плазмиды YF5'3'IV/DEN(prM-E).
Конструирование химерных матриц для других флавивирусов
Процедуры
создания полноразмерных кДНК-матриц, кодирующих химерные вирусы YF/MVE, YF/SLE, YF/WN и YF/TBE, сходны с теми процедурами, которые были описаны выше для системы YF/DEN-2. Табл. 6 показывает
особенности стратегии создания химерных вирусов, основанных на вакцинном штамме YF17D. Также показаны уникальные рестрикционные сайты, использовавшиеся для лигирования in vitro, и химерные праймеры
для конструирования гибридных белков С/рrМ и E/NS1. Источники кДНК для указанных гетерологичных вирусов легко доступны (MVE: Dalgarno et al., 1986, J. Mol. Biol, 187:309-323; SLE: Trent et al., 1987,
Virology, 156:293-304; TBE: Mandl et al., 1988, Virology, 166: 197-205; вирус денге-1: Mason et al., 1987, Virology, 161:262-267; вирус денге-2: Deubel et al., 1986, Virology, 155:365-377; вирус
денге-3: Hahn et al. , 1988, Virology, 162:167-180; вирус денге-4: Zhao et al., 1986, Virology, 155:77-88).
Альтернативным подходом к конструированию дополнительных химерных вирусов является создание гибридного белка С/рrМ путем лигирования по "тупым концам" ПЦР-производных рестрикционных фрагментов, характеризующихся концами, которые встречаются в месте этой гибридизации, и 5'- и 3'-концами, которые фланкируют рестрикционные сайты, подходящие для встраивания в плазмиду YF5'3'IV или в какую-либо промежуточную плазмиду, такую как pBS-KS(+). Подходы к использованию химерного олигонуклеотида либо лигированию по "тупым концам" должны варьироваться в зависимости от доступности уникальных рестрикционных сайтов в составе последовательности, кодирующей оболочечный белок в составе генома конкретного вируса.
Конструирование вирусов YF, кодирующих антигены HCV
Поскольку структурные белки Е1 и Е2 вируса HCV (гепатита С) негомологичны
структурным белкам описанных выше флавивирусов, стратегия экспрессии этих белков включает встраивание в пределы того участка генома, который не является "жизненно необходимым", таким образом, чтобы
все эти белки затем экспрессировались вместе с белками вируса желтой лихорадки в процессе размножения вируса в инфицированных клетках. Участком, являющимся мишенью для встраивания последовательностей,
кодирующих эти белки, является последовательность, кодирующая N-концевую часть белка NS1, поскольку наличие полного белка NS1 не является строго обязательным для размножения вирусов. С учетом
имеющейся проблемы со стабильностью генома YF в присутствии гетерологичной последовательности, превышающей нормальный размер вирусного генома (приблизительно 10 тысяч нуклеотидов), может быть
использована описанная ниже "детекционная стратегия". Кроме того, делетирование гена NS1 может давать преимущества химерным флавивирусным системам "YF/флавивирус", описанным выше, благодаря тому, что
частичная делеция этого белка может предотвращать выработку иммунитета к YF, основанного на антителах к белку NS1, что, таким образом, снимает проблемы вирусного иммунитета тогда, когда необходимо
вводить данному пациенту более одной химерной вакцины или когда ранее была введена вакцина против желтой лихорадки или же ее введение понадобится в будущем.
Данная стратегия включает формирование серии внутрирамочных делеций в пределах последовательности, кодирующей белок NS1, находящейся в составе плазмиды YFM5.2, в сочетании с конструированием стоп-кодона по окончании кодирующей рамки белка Е, а также серии из двух IRES (т.н. "внутренние сайты присоединения к рибосоме"). Один из сайтов IRES расположен сразу вниз от стоп-кодона и обеспечивает экспрессиию открытой рамки на участке между кодирующими последовательностями Е и NS1. Второй сайт IRES инициирует трансляцию укороченного белка NS1, что обеспечивает экспрессию оставшегося полипредшественника неструктурных белков. Эти производные варианты тестируют на восстановление инфекционного вируса, а конструкцию с самой большой делецией используют для встраивания чужеродных последовательностей (например, генов белков HCV) по их первому сайту IRES. Такая частная конструкция также может служить основой для определения того, будет ли делеция гена NS1 подавлять вектор-специфичный иммунитет в контексте химерных конструкций "YF/флавивирус", экспрессирующих рrМ-Е, таким способом, который был описан выше.
Встраивание нуклеотидов, кодирующих белки вируса HCV E1, E2 и (или)
Е1+Е2, ограничено размером делеции,
по которой вирус-реципиент "толерантен" к утрате белка NS1. С учетом этого с целью усиления стабильности модифицируемого клона YF могут быть использованы
укороченные антигены вируса HCV. Белки HCV
конструируют таким образом, чтобы N-концевая
сигнальная последовательность располагалась сразу вслед за сайтом IRES, а стоп-кодон - непосредственно
на С-конце. Такая конструкция будет
направлять белки HCV в эндоплазматический ретикулум с последующей секрецией из клетки. Схематически стратегия получения таких конструкций показана на фиг.6. Для
такого конструирования могут быть
использованы плазмиды, кодирующие белки HCV генотипа I, например плазмиды с HCV, полученные от Д-ра Чарльза Раиса из Вашингтонского университета (Grakoui et al.,
1993, J. Virol, 67: 1385-1395),
который экспрессировал этот сегмент вируса в процессирующих системах и в полноразмерном клоне HCV, способном к репликации.
Делеционные мутанты по
расщеплению рrМ, как предполагаемые
ослабленные вакцинные флавивирусы
Дополнительные химерные вирусы, включенные в настоящее изобретение, несут мутации, которые предотвращают расщепление
(процессинг) предшественника рrМ, такие
как мутации, затрагивающие сайт расщепления рrМ. Например, сайт расщепления рrМ у представляющих интерес инфекционных флавивирусных клонов, таких как вирусов
денге, ТВЕ, SLE и другие, может быть
мутирован с применением метода направленного (сайт-специфичного) мутагенеза. Любая из или все аминокислоты в составе сайта расщепления, которые были названы выше,
могут быть делетированы или замещены.
Фрагмент нуклеиновой кислоты, включающий мутированные гены рrМ-Е, может быть затем встроен в вектор вируса желтой лихорадки с использованием описанных выше
методов. Делетирование в гене рrМ может быть
проведено наряду с или без сопровождающих ослабляющих мутаций, таких как, например, мутаций в гене белка Е, перед встраиванием в состав генома вируса
желтой лихорадки. Эти мутанты обладают
преимуществами по сравнению с монозамещенными мутантами в качестве предполагаемых вакцин, потому что они обеспечивают практически полную невозможность реверсии
делетированных последовательностей и
восстановления вирулентности.
Следующие химерные флавивирусы по настоящему изобретению были внесены в Американскую Коллекцию типовых культур (АТСС), находящуюся в Роквилле (штат Мэриленд, США) в соответствии с Будапештским договором с датой включения 6 января 1998 года: химерный вирус "желтой лихорадки 17D/денге типа 2" (YF/DEN-2; депозитарный АТСС VR-2593) и химерный вирус "желтой лихорадки 17D/японского энцефалита" SA14-14-2 (YF/JE А1.3; депозитарный АТСС VR-2594).
Другие варианты
Другие
варианты также охватываются приведенной ниже
формулой изобретения. Например, гены других имеющих медицинское значение флавивирусов, кодирующие белок рrМ-Е, могут быть встроены в вакцинный вирус
желтой лихорадки с целью получения вакцин против
других важных с медицинской точки зрения флавивирусов (см., например, Monath et al., 1995, "Flaviviruses", In "Virology", ed. by Fields,
Raven-Lippincott, New York, Vol. I, 961-1034).
Примерами других флавивирусов, гены из геномов которых могут быть встроены в химерные векторы по настоящему изобретению, являются вирусы лихорадки Куньиня, центрально-европейского энцефалита, весенне-летнего клещевого энцефалита, повассанской лихорадки, киасанурской лесной болезни и омской геморрагической лихорадки. Кроме того, гены даже более филогенетически отдаленных вирусов могут быть встроены в вакцинный вирус желтой лихорадки в процессе конструирования новых вакцин.
Получение и использование вакцин
Вакцины по настоящему изобретению вводят в количестве и с
использованием тех способов, которые могут быть легко определены специалистами в данной области техники. Вакцины могут быть приготовлены и
введены, например, таким же образом, как и вакцина против
желтой лихорадки 17D, например, в виде осветленной суспензии инфицированной ткани куриного эмбриона или в виде жидкости, выделяемой из
клеточных культур, инфицированных химерным вирусом желтой
лихорадки. Таким образом, живой ослабленный вакцинный вирус используется при приготовлении лекарственного препарата в виде стерильного
водного раствора, содержащего от 100 до 1 миллиона инфекционных
единиц (например, бляшкообразующих единиц или инфекционных доз для тканевых культур) в дозах от 0,1 до 1,0 мл, предназначенного для,
например, внутримышечного, подкожного или внутрикожного введения.
Кроме того, поскольку флавивирусы могут быть способны к заражению человека через слизистые оболочки, например, через слизистые рта
(Gresikova et al., 1988, "Tick-borne encephalitis". In "Arboviruses:
Ecology & Epidemiology", ed. by Monath, CRC Press, Boca Raton, FL, Vol. IV, 177-203), то вакцинный вирус может быть введен
по пути через слизистые, с целью достижения защитного иммунного
ответа. Вакцина может быть введена в качестве первичного профилактического средства взрослым или детям при наличии риска заражения
флавивирусной инфекцией. Также вакцины могут быть использованы в
качестве вторичных средств для лечения зараженных флавивирусами пациентов путем стимулирования иммунного ответа на флавивирусную
инфекцию.
Может быть желательным использовать векторную систему вакцины против желтой лихорадки, предназначенной для иммунизации реципиента против одного из вирусов (например, вируса японского энцефалита) и для последующей реиммунизации того же объекта против второго или третьего вируса с применением различающихся химерных конструкций. Существенное преимущество химерной системы вируса желтой лихорадки состоит в том, что вектор не приводит к выработке сильного иммунитета к самому себе. Также предварительный иммунитет к желтой лихорадке не является препятствием для использования химерной вакцины в качестве вектора для экспрессии гетерологичных генов. Эти преимущества обусловливаются удалением части гена белка Е вируса желтой лихорадки, который кодирует нейтрализующие (защитные) антигены к желтой лихорадке, и замещением его другим гетерологичным геном, который не обеспечивает перекрестного защитного эффекта против желтой лихорадки. Хотя неструктурные белки вируса YF17D могут играть роль в защите, например, за счет формирования антител к NS1, который вовлечен в опосредованный комплемент-зависимыми антителами лизис инфицированных клеток (Schlesinger et al., 1985, J. Immunol, 135: 2805-2809), или за счет индуцирования Т-клеточного ответа на белок NS3 или на другие вирусные белки, то практически исключено, чтобы эти иммунные ответы предотвращали бы способность живой вирусной вакцины стимулировать выработку нейтрализирующих антител. Это заключение подтверждается теми фактами, что: 1) пациенты, которые были ранее инфицированы вирусом японского энцефалита, реагируют на вакцинацию штаммом YF17D сходным образом с теми пациентами, у которых предварительного заражения вирусом JE не было, и 2) пациенты, которые ранее получали вакцину YF17D, реагируют на повторную вакцинацию повышением титров нейтрализующих антител (Sweet et al., 1962, Amer. J. Trop. Med. Hyg. , 11:562-569). Таким образом, химерный вектор может быть использован в популяциях, являющихся иммунными к желтой лихорадке благодаря первичному естественному иммунитету или вакцинации, а также может быть использован повторно, или для иммунизации одновременно или последовательно наряду с некоторыми другими отличающимися конструкциями, включая химеры вируса желтой лихорадки со встроенными вирусами, например, японского энцефалита, энцефалита Сент-Луиса или лихорадки западного Нила.
Для практической вакцинации могут быть использованы адъюванты, которые хорошо известны специалистам в данной области техники. Адъюванты, которые могут быть использованы для усиления иммуногенности химерных вакцин, включают, например, липосомные препараты, синтетические адъюванты, такие как сапонины (например, QS21), мурамил-дипептид, монофосфорный липид-А или полифосфазин. Хотя эти адъюванты обычно используются для усиления иммунных ответов на инактивированные (убитые) вакцины, они также могут быть использованы и для живых вакцин. В случае применения химерных вакцин через слизистые, например, через слизистые рта, применимыми адъювантами могут быть специальные адъюванты для слизистых оболочек, такие как теплоустойчивый токсин E.coli (LT) или мутантные производные LT. Кроме того, гены, кодирующие цитокины, обладающие активностью адъювантов, могут быть встроены в векторы вируса желтой лихорадки. Таким образом, гены, кодирующие цитокины, такие как GM-CSF (колониестимулирующий фактор гранулоцитов/макрофагов), IL-2, IL-12, IL-13 или IL-5 (интерлейкины), могут быть встроены вместе с гетерологичными флавивирусными генами с целью получения вакцины, которая бы обусловила усиление иммунных ответов, или с целью модулирования иммунитета, более специфически направленного в отношении клеточных, гуморальных или слизистых ответов. В дополнение к применению вакцин, как понятно специалисту в данной области техники, векторы по настоящему изобретению могут быть использованы в методах генотерапии с целью внесения "терапевтических генных продуктов" в клетки пациента. В этих методах гены, кодирующие "терапевтические генные продукты", встраиваются в состав векторов, например, вместо гена, кодирующего белок рrМ-Е.
Дополнительное преимущество векторной системы вируса желтой лихорадки состоит в том, что репликаты флавивируса находятся в цитоплазме клеток, поэтому стратегия размножения вируса не связана интеграцией вирусного генома в геном клетки-хозяина (Chambers et al., 1990, "Flavivirus genome organization, expression and replication", Ann. Rev. Microbiol, 44:649-688), что является важным фактором безопасности.
Все цитированные ссылки включены в данный текст в своей полноте.
Реферат
Изобретение относится к медицине и касается химерного живого инфекционного аттенуированного вируса, который применяется для создания химерных флавивирусных вакцин. Изобретение включает химерный живой инфекционный аттенуированный вирус, содержащий вирус желтой лихорадки, у которого нуклеотидная последовательность, кодирующая белок prМ-Е, делетирована, укорочена или мутирована таким образом, что функциональный белок prМ-Е не экспрессируется, но при этом в геном вируса желтой лихорадки интегрирована нуклеотидная последовательность, кодирующая белок prМ-Е второго, отличающегося, флавивируса таким образом, что белок prМ-Е второго флавивируса экспрессируется, а также молекулу нуклеиновой кислоты, кодирующую этот вирус. Преимущество изобретения заключается в создании химерных вирусов, которые могут применяться в качестве живых аттенуированных вакцин. 3 с. и 15 з.п. ф-лы, 7 ил., 6 табл.
Формула
28.02.1997 по пп.1-5, 7-10, 11, 13-17;
15.01.1998 по пп.4, 10 и 16: в качестве второго вируса представлен вирус клещевого энцефалита;
15.01.1998 по пп.6, 12 и 18.
Комментарии