Вакцина против цитомегаловируса и способ ее получения - RU2491340C2
Код документа: RU2491340C2
Чертежи




Описание
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области разработки вакцин. А именно, настоящее изобретение относится к способам увеличения диверсификации вакцин против цитомегаловируса за счет выбора клеточного типа, в котором реплицируется этот вирус, и применению цитомегаловируса, полученного этими способами, в разработке композиций вакцин.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Различные публикации, включая патенты, опубликованные заявки, технические и научные статьи цитируются на протяжении всего данного описания. Каждая из цитируемых публикаций включена в данный документ путем ссылки во всей полноте. Полные цитаты публикаций, указанных номерами ссылок в скобках или не цитируемых полностью в рамках данного описания, приведены в конце данного описания.
Цитомегаловирус представляет собой герпесвирус, в классификации отнесенный к членам бета-подсемейства герпесвирусов. Согласно Центру по контролю и профилактике инфекционных заболеваний цитомегаловирусная инфекция распространена у человека практически повсеместно, при этом по расчетам в США этим вирусом инфицировано 40-80% населения. Вирус распространяется преимущественно через биологические жидкости и часто передается от беременных женщин плоду или новорожденному. У большинства людей цитомегаловирусная инфекция протекает в латентной форме, хотя активация вируса может проявляться высокой лихорадкой, ознобами, повышенной утомляемостью, головной болью, тошнотой и спленомегалией.
Хотя большинство случаев цитомегаловирусных инфекций у человека протекают бессимптомно, у лиц с незрелой иммунной системой или иммунодефицитом, таких как новорожденные, ВИЧ-положительные, пациенты, перенесшие аллотрансплантацию, и онкологические пациенты, цитомегаловирусные инфекции могут вызывать проблемы. Цитомегаловирусная инфекция у таких лиц может вызывать тяжелые осложнения, включая пневмонию, гепатит, энцефалит, колит, увеит, ретинит, слепоту и нейропатию среди других пагубных последствий. Кроме того, цитомегаловирус является ведущей причиной врожденных аномалий. В настоящее время терапевтической или профилактической вакцины против цитомегаловирусной инфекции не существует.
Проникновение герпесвирусов в клетки представляет собой сложный процесс, который запускается адсорбцией и связыванием рецептора с последующим слиянием оболочки вируса с клеточной мембраной. Слияние происходит либо на плазматической мембране, либо на эндосомальной мембране. Например, вирус Эпштейна-Барр (ВЭБ) проникает преимущественно в В-лимфоциты путем эндоцитоза, опосредованного рецептором (1, 2), Однако он инфицирует эпителиальные клетки или трансформированные В-лимфоциты путем слияния оболочки вируса с плазматической мембраной (1). Вирус простого герпеса сливается с плазматической мембраной некоторых клеточных типов, а у других проникает путем эндоцитоза (3-6). Человеческий цитомегаловирус инфицирует различные клеточные типы in vivo, включая эпителиальные клетки, эндотелиальные клетки и фибробласты (7). Он сливается с плазматическими мембранами фибробластов (8), но в клетки пигментного эпителия сетчатки и эндотелиальные клетки пупочной вены он проникает путем эндоцитоза (9, 10).
Механизм, с помощью которого герпесвирусы «выбирают» путь проникновения, остается неясным. Принято считать, что пути проникновения в основном определяются клеткой хозяина, но имеются и данные о роли гликопротеинов вириона, определяющих тропизм (11). Вирионы ВЭБ содержат два gH-комплекса, gH/gL и gH/gL/gp42 (12, 13), которые обладают взаимоисключающими функциями (11). Слияние с плазматической мембраной В-лимфоцитов опосредовано gH/gL/gp42 (14-16). Но проникновение в эпителиальные клетки запускается gH/gL (11, 12, 17). Клеточный тип, в котором реплицируется ВЭБ, может менять его тропизм. Вирионы ВЭБ - производные В-лимфоцитов - содержат меньше gH-gL-gp42, чем вирионы - производные эпителиальных клеток. В итоге вирус, генерируемый В-лимфоцитами, является более инфекционным для эпителиальных клеток, а вирус - производное эпителиальных клеток - обладает тропизмом к В-лимфоцитам (18).
Человеческий цитомегаловирус также кодирует два gH/gL-комплекса: gH/gL/gO и gH/gL/pUL128/pUL130/ pUL131 (19, 20). gO-содержащий комплекс достаточен для инфицирования фибробластов, в то время как pUL128/pUL130/pUL131-содержащий комплекс необходим для инфицирования эндотелиальных и эпителиальных клеток (19-21). Лабораторный штамм AD 169 содержит только gH/gL/gO-комплекс в вирионах (19). Отсутствие второго gH/gL-комплекса обусловливает потерю тропизма к эпителиальным и эндотелиальным клеткам у лабораторных штаммов человеческого цитомегаловируса (19-22).
Имеется потребность в разнообразии и диверсификации вакцины против цитомегаловируса, а также в эффективных средствах контроля распространения и активации вируса, особенно у лиц с иммунодефицитом и беременных женщин. Настоящее изобретение отвечает этой потребности.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Один аспект настоящего изобретения относится к способу получения вакцин против цитомегаловируса. Этот способ включает культивирование вируса или изолятов цитомегаловируса в клеточных культурах отдельных клеточных типов с получением таким образом цитомегаловируса, кондиционированного клеточным типом, и получение вакцины против цитомегаловируса из цитомегаловируса, кондиционированного клеточным типом. В некоторых вариантах штамм или изолят цитомегаловируса является штаммом или изолятом человеческого цитомегаловируса. Для данного способа пригодны разнообразные клеточные типы, включая, но не ограничиваясь перечисленным, эпителиальные клетки, эндотелиальные клетки, фибробласты, нейроны, гладкомышечные клетки, макрофаги, дендритные клетки и стромальные клетки. В конкретном варианте выбранный клеточный тип представляет собой эпителиальную клетку.
Вышеупомянутый способ может дополнительно включать получение цитомегаловируса, кондиционированного клеточным типом или двумя или больше различными выбранными клеточными типами и комбинирование этих цитомегаловирусов для получения вакцины против цитомегаловируса. Иначе или дополнительно данный способ включает две или больше культуры клеток, содержащих выбранный клеточный тип или два или больше различных клеточных типа, и комбинирование всех полученных цитомегаловирусов для получения вакцины против цитомегаловируса.
В некоторых вариантах данный способ включает получение живой аттенуированной вакцины против цитомегаловируса. В других вариантах он включает получение инактивированной или убитой вакцины против цитомегаловируса. Еще в других вариантах он включает получение комбинированных вакцин, содержащих один или несколько живых аттенуированных вирусов, инактивированных вирусов и другие иммуногенные компоненты, например, иммуногенные белки и пептиды цитомегаловируса и им подобные.
Вакцины против цитомегаловируса, которые получают вышеупомянутыми способами, также входят в объем настоящего изобретения.
Другой аспект настоящего изобретения относится к набору для осуществления способов настоящего изобретения. Такие наборы обычно включают упаковку, в которой содержатся один или несколько штаммов или клинических изолятов цитомегаловируса, клеточных культур одного или нескольких выбранных клеточных типов и инструкции по использованию клеточных культур и штаммов или изолятов цитомегаловируса для получения кондиционированного клеточным типом цитомегаловируса для применения в вакцине против цитомегаловируса.
Другой аспект настоящего изобретения включают композицию вакцин, включающую популяцию цитомегаловируса или компоненты вирионов данной популяции, смешанные с соответствующим фармацевтическим носителем или адъювантом, где популяцию цитомегаловируса выделяют из клеточной культуры выбранного клеточного типа. В одном варианте выбранный клеточный тип представляет собой эпителиальный клеточный тип. В одном варианте композиция вакцины включает человеческий цитомегаловирус.
В различных вариантах композиции вакцин популяция цитомегаловируса, выделенная из культуры эпителиальных клеток, характеризуется одним или несколькими особенностями в инфицированных впоследствии клетках хозяина, включая, но не ограничиваясь перечисленным: а) проникновение в клетки хозяина путем слияния с плазматическими мембранами клеток хозяина; б) более выражение слияние клеток хозяина, опосредованное вирионом, по сравнению с эквивалентной популяцией цитомегаловируса, выделенного из культуры фибробластов; в) ускоренную репликацию вируса в клетках хозяина по сравнению с эквивалентной популяцией цитомегаловируса, выделенного из культуры фибробластов; г) индукцию клеточного ответа, включающего изменение экспрессии не менее чем в 2,5 раза генах, число которых примерно на две трети меньше чем при ответе, вызванном эквивалентной популяцией цитомегаловируса, выделенного из культуры фибробластов через 10 часа после инфицирования; или д) индукцию клеточного ответа, включающую изменение экспрессии одного или нескольких генов, как показано в таблице 2 и таблице 4 данного документа, при этом последние представлены в Банке генов под входящими номерами; АК094860, NM_145023, NM-133492, NM_001039580. NM_001004301, NM-001034, А1369525, АК123066, NM_005345, NM_020731, BC071797, NM-003414, NM_000800, NM_138467, AK090803, AL133118, NM-00165, BG001037, NM_024861, NM-001043, NM-016239, NM-001018084, NM-001037442, NM-017600, NM_022097, NM-175868, NM_032266, NM_003841, NM_005039, NM_14505 1, NM_004294, AW856073, NM_024050, AF085968, NM-080927, NM_022115, AK056703, NM_000808, NM-012377, NM_006793, NM_031466, NM_005185, NM-139173, BX360933, NM-016125, NM_002104, NM-032188, NM_004185, NM-004843 или NM-173550.
В некоторых вариантах композиция вакцины включает популяцию цитомегаловируса или компоненты его вириона, выделенные из клеточной культуры одного или нескольких выбранных клеточных типов. Например, популяция цитомегаловируса может быть выделена из эпителиальных клеток и клеток другого клеточного типа, такого как клеточный тип фибробластов. В других вариантах популяция цитомегаловируса включает два или несколько штаммов или клинических изолятов цитомегаловируса, выращенного в выбранном клеточном типе. Некоторые варианты могут включать множество штаммов или клинических изолятов цитомегаловируса, выращенного в клеточных культурах множества различных клеточных типов.
В одном варианте композиция вакцины включает живую аттенуированную вакцину против цитомегаловируса. В другом варианте она включает инактивированную вакцину против цитомегаловируса. Еще в других вариантах композиция вакцины может быть комбинацией вакцин, включающей один или несколько штаммов живого аттенуированного вируса или его компонентов, инактивированный вирус или его компоненты и (или) другие иммуногенные пептиды или белки цитомегаловируса.
Другой аспект настоящего изобретения включает способ иммунизации субъекта против цитомегаловируса, включающий введение данному субъекту композиции вакцины против цитомегаловируса, произведенной вышеупомянутыми способами и (или) включающими вышеупомянутые свойства. В одном варианте субъектом, которого предстоит иммунизировать, является человек.
Другие особенности и преимущества настоящего изобретения будут понятны из рисунков, подробного описания и примеров, приведенных ниже.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фиг.1. Кинетика экспрессии IE1 человеческого цитомегаловируса в клетках ARPE-19. А) Инфицированные клетки (0,1 БОЕ/клетку) смешали в указанные сроки и окрасили на IE1 (зеленый цвет на цветной фотографии, светло-серый цвет на черно-белой фотографии), Sp100 (красный цвет на цветной фотографии, очень темный цвет на черно-белой фотографии) и ДНК (синий цвет на цветной фотографии, темно-серый цвет на черно-белой фотографии). Б) В различные сроки после инфекции (0,1 БОЕ/клетку) подсчитали процент IE1 - экспрессирующих клеток; результаты показаны на диаграмме.
Фиг.2. Электронно-микроскопический анализ проникновения человеческого цитомегаловируса в клетки ARPE-19. Частицы epiBADrUL131 или fibroBADrUL131 (50 БОЕ/клетку) связывали с клетками при 4°С, а затем оставляли при 37°С на 15 мин, чтобы обеспечить интернализацию. Представлены репрезентативные рисунки.
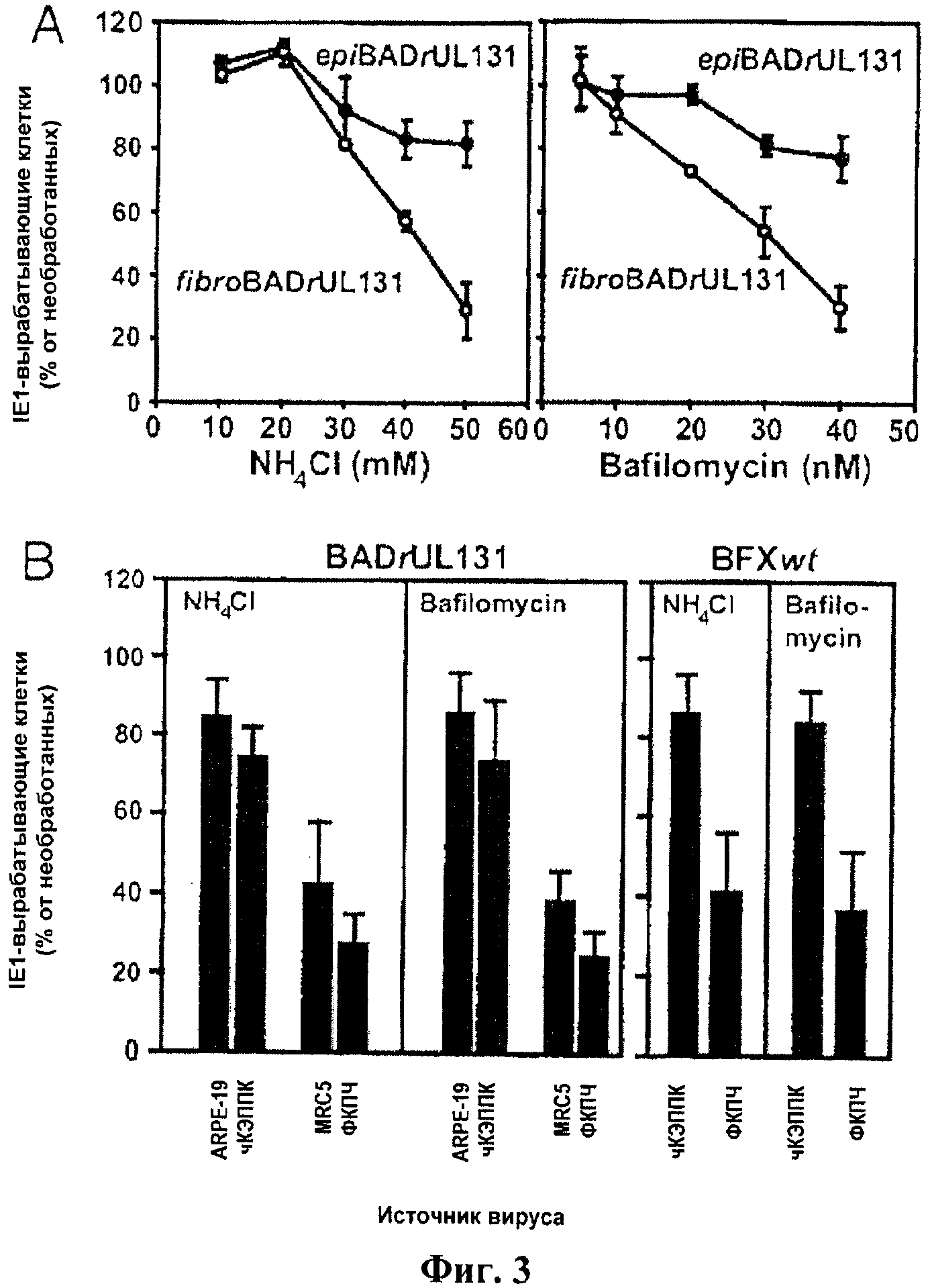
Фиг.3. Влияние ингибиторов окисления эндосом и источника вириона на проникновение человеческого цитомегаловируса в клетки ARPE-19. Эксперименты проводили в трех копиях и подсчитывали число положительных клеток после обработки лекарственным средством в сравнении с необработанными культурами. А) Клетки предварительно обрабатывали NH4Cl или бафломицином А (БФА) в течение 1 ч, инокулировали epiBADrUL131 или fibroBADrUL131 (1 БОЕ/клетку) и через 16 ч окрашивали на IE1. Б) Клетки предварительно обрабатывали 50 моль NH4Cl или 40 мМоль БФА в течение 1 ч, инокулировали BADrUL131 (0,1 БОЕ/клетку) или FIXwt (0,01 БОЕ/клетку) и через 16 ч окрашивали на IE1.
Фиг.4. Слияние клеток ARPE-19, индуцированное вирусом - производным эпителиальных клеток. А) Клетки инокулировали epiBADrUL131 или fibroBADrUL131 (20 БОЕ/клетку) и затем держали в среде, содержащей 200 мкг/мл БФА. Через 16 ч после инфицирования получали фазовоконтрастные изображения. Б) Смесь репортерных и эффекторных клеток инфицировали epiBADrUL131 или fibroBADrUL131 (20 БОЕ/клетку) при 4°С в течение 1 ч. Затем культуру держали при 37°С в течение 6 ч, после чего измеряли относительную активность люциферазы.
Фиг.5. Влияние pUL130-специфичного нейтрализующего антитела на инфекцию и проникновение человеческого цитомегаловируса. А) Вирусы - производные эпителиальных клеток или фибробластов - инкубировали с различными концентрациями антитела к pUL130 и определяли остаточную инфективность. Б) Частицы вирусов - производных эпителиальных клеток или фибробластов - предварительно обрабатывали антителом к pUL130 в конечной концентрации 20 мкг/мл или фосфатным буфером, а затем адсорбировали на клетках ARPE-19 при 4°С в течение 1 ч. Клетки дважды отмывали холодным фосфатным буфером и экстрагировали вирусную ДНК, связанную с клетками, чтобы определить относительное число частиц, связанных в клетками. Или клетки оставляли при 37°С на 2 ч, чтобы обеспечить проникновение вируса. Вирионы, не проникшие в клетки, удаляли с помощью обработки ЭДТК-трипсином. Интернализованную вирусную ДНК впоследствии измеряли с помощью ПЦР в режиме реального времени.
Фиг.6. Модуляция транскритома ARPE-19 человеческим цитомегаловирусом, полученным в эпителиальных клетках и фибробластах. А) Диаграмма Венна отображает распределение генов, регулируемых различным образом, через 6 ч или 10 ч после инфицирования epiBADrUL131 или fibroBADrU1131 (3 БОЕ/клетку) по сравнению с симуляцией инфекции. Б) Изменения относительных концентраций РНК, измеренных с помощью ПЦР в режиме реального времени. Исследованные гены представляют собой ген гидрометилбилансинтазы (HMBS, NM_000190), ген 1, связанный с патогенезом GLI (глиома) (GIiPR, NM_006851), ген, связанный с пентраксином, быстро индуцируемый ИД-1 бета (РТХ3, NM_002852), ген 2'-5'-олигоаденилатсинтетазы 3 (OAS3, NM_006187), ген белка 44, индуцируемого интерфероном (IFI44, NM_006417), гомолог В вирусного онкогена, связанного с ретикулоэндотелиозом, ген-энхансер ядерного фактора легкой цепи полипептида 3 в В-лимфоцитах (relB, NM_006509) и АТФ-связывающую кассету, подсемейство С (CFTR/MRP), член 3 (MRP3, NM_003786).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В тексте данного описания и формулы изобретения используются различные термины, связанные со способами и другими аспектами настоящего изобретения. Если не указано иначе, все технические и научные термины, использующиеся в данном документе, имеют то же значение, что обычно понимает специалист в данной области техники, к которой относится настоящее изобретение. Хотя на практике для тестирования настоящего изобретения можно использовать любые способы и материалы, схожие или эквивалентные тем, что описаны в данном документе, предпочтительны материалы и способы, описанные в данном документе. В описании настоящего изобретения и формуле изобретения будет использоваться следующая терминология. Следует понимать, что терминология, использующаяся в данном документе, предназначена только для описания определенных вариантов и не налагает каких-либо ограничений.
Определения:
Согласно использованию в данном описании и приложенной формуле изобретения существительные в единственном числе включают и соответствующее множественное число, если содержание документа ясно не указывает иначе. Таким образом, например, термин «клетка» включает комбинацию двух или более клеток и так далее.
«Примерно», согласно использованию в данном документе, при употреблении в отношении измеряемой величины, такой как количество, длительность или так далее, означает колебания ±20%, или ±10%, или более предпочтительно ±5%, и даже более предпочтительно ±1%, и еще более предпочтительно ±0,1% от указанной величины, так как такие колебания приемлемы для осуществления описанных способов.
Термины «амплификация», «репликация» и «культивирование» в данном документе используются взаимозаменяемо и означают общий процесс введения вируса в клеточные культуры или инфицирование клеток вирусов в условиях, обеспечивающих репликацию вируса в клетках в соответствии с методиками, хорошо известными вирусологам и медико-биологам. В частности, эти термины используются в данном документе, для обозначения этапа способа, являющего предметом изобретения, при котором цитомегаловирус «кондиционируют» путем культивирования в выбранном клеточном типе в качестве этапа, предшествующего использованию кондиционированного цитомегаловируса в получении вакцины.
«Биомолекулы» включают белки, полипептиды, нуклеиновые кислоты, липиды, полисахариды и все фрагменты, аналоги, гомологи, конъюгаты и производные перечисленных соединений.
«Клеточная культура» обычно означает клетки, взятые из живого организма и выращенные при контролируемых условиях («в культуре» или «культивированные»). «Первичная клеточная культура» представляет собой культуру клеток, тканей или органов, взятых непосредственно из организма(ов) до получения первой субкультуры. «Клеточная линия» означает культуру клеток, образованную путем получения одной или нескольких субкультур из первичной клеточной культуры.
«Кодирующий район» гена состоит из остатков нуклеотидов в кодирующей цепи гена и нуклеотидов некодирующей цепи гена, гомологичных или комплементарных соответственно кодирующему району молекулы мРНК, которая получается при транскрипции этого гена.
«Кодирующий район» молекулы мРНК также состоит из остатков нуклеотидов молекулы мРНК, комплементарных району антикодона молекул транспортной РНК в ходе трансляция молекулы мРНК, или кодирующих стоп-кодом. Таким образом, кодирующий район может включать остатки нуклеотидов, соответствующие аминокислотным остаткам, которые отсутствуют в зрелом белке, кодируемом этой молекулой мРНК (например, аминокислотные остатки в последовательности экспорта сигнала белка).
Термины «кондиционированный вирус», «кондиционированный клеточным типом вирус», «кондиционированный цитомегаловирус» или «кондиционированный клеточным типом цитомегаловирус» означает цитомегаловирус, который был культивирован к выбранном клеточном типе до его применения в получении вакцины, согласно способам, описанным в данном документе. Эти термины употребляются как аналоги термина «кондиционированная среда», который описывает питательную среду, в которой выращен определенный клеточный тип или клеточная линия, после чего этот тип или линия были удалены, и которая содержит компоненты или факторы, вырабатывающиеся клетками, что таким образом меняет функциональные свойства среды. В рамках настоящей заявки термин «кондиционированный вирус» также означает вирус, который культивировали в выбранном клеточном типе и затем отделили от этих клеток, где этот вирус впоследствии проявляет одно или несколько измененных функциональных свойств в результате культивирования в этом клеточном типе.
«Кодирующий» означает естественное свойство определенных последовательностей нуклеотидов в полинуклеотиде, таком как ген, кДНК или мРНК, служить матрицей для синтеза других полимеров и макромолекул в биологических процессах, при этом «кодирующий» имеет определенную последовательность нуклеотидов (т.е. рРНК, тРНК или мРНК) или определенную последовательность аминокислот и биологические свойства, обусловленные этой последовательностью. Таким образом, ген кодирует белок, если транскрипция и трансляция мРНК, соответствующей данному геному, приводит к образованию белка в клетке или другой биологической системе. И кодирующую цепь, нуклеотидная последовательность, которой идентична последовательности мРНК и обычно приводится в перечислении последовательностей, и некодирующую цепь, использующуюся как матрица для транскрипции гена или кДНК, можно назвать кодирующей белок или другие продукты данного гена или кДНК. Если не указано иначе, «нуклеотидная последовательность, кодирующая аминокислотную последовательность» включает все нуклеотидные последовательности, являющиеся вырожденными вариантами друг друга и кодирующими одну и ту же аминокислотную последовательность. Нуклеотидные последовательности, которые кодируют белки и РНК, могут включать интроны.
«Эффективное количество» или «терапевтически эффективное количество» употребляются в данном документе взаимозаменяемо, и эти термины означают количество соединения, состава, материала или композиции согласно описанию в данном документе, эффективное для достижения определенного биологического результата. Такие результаты могут включать, но не ограничиваться перечисленным, торможение вирусной инфекции согласно определению каким-либо методом, приемлемым для данной области техники.
Согласно употреблению в данном документе, «эндогенный» означает любой материал из или вырабатывающийся в организме, клетке, ткани или системе. «Экзогенный» означает любой материал, введенный извне или вырабатывающийся вне организма, клетки, ткани или системы.
Термин «экспрессия» согласно употреблению в данном документе означает транскрипцию и (или) трансляцию определенной нуклеотидной последовательности, запускающуюся промотором.
Согласно употреблению в данном документе «иммунизация» или «вакцинация» взаимозаменяемы и применяются в отношении профилактической иммунизации или вакцинации. «Терапевтическая вакцинация» означает вакцинацию пациента с цитомегаловирусной инфекцией.
«Выделенный» означает измененный или извлеченный из естественного состояния. Например, нуклеиновую кислоту или пептид, присутствующий естественным образом у живого животного, не «выделяют», но ту же самую нуклеиновую кислоту или пептид, частично или полностью отделенный от сопутствующих материалов в его естественном состоянии, «выделяют». Выделенная нуклеиновая кислота или белок могут существовать в значительно очищенной форме или могут существовать в окружении, отличающемся от естественного, таком как, например, клетка хозяина. Если в данном документе не указано иначе, белки, комплексы вирионов, антитела и другие биологические молекулы, относящиеся к предмету настоящего изобретения, являются выделенными или могут быть выделены.
Термин «пациент», «субъект», «особь» и так далее употребляются в данном документе взаимозаменяемо и означают какое-либо животное или его клетки, in vitro или in situ, которые можно инфицировать цитомегаловирусом. В некоторых вариантах, не налагающих ограничений, пациент, субъект или особь является человеком.
«Парентеральное» введение иммуногенной композиции или композиция вакцин включает, например, подкожное (п/к), внутривенное (в/в), внутримышечное (в/м) или внутригрудинное (в/г) введение или пути введения.
Термин «полинуклеотид» согласно употреблению в данном документе определен как цепь нуклеотидов. Кроме того, нуклеиновые кислоты - это полимеры нуклеотидов. Таким образом, нуклеиновые кислоты и полинуклеотиды согласно употреблению в данном документе являются взаимозаменяемыми. Специалист в данной области техники знает, что нуклеиновые кислоты являются полинуклеотидами, которые можно гидролизовать на мономерные «нуклеотиды». Мономерные нуклеотиды можно гидролизовать до нуклеозидов. Согласно употреблению в данном документе полинуклеотиды включают, но не ограничиваются перечисленным, все последовательности нуклеиновых кислот, которые получают любым путем, известным в данной области техники, включая, но не ограничиваясь перечисленным, рекомбинантные методики, например, клонирование последовательности нуклеиновой кислоты из рекомбинантной библиотеки или клеточного генома с использованием методик клонирования или амплификации и им подобным и путем синтеза.
Согласно употреблению в данном документе термины «пептид», «полипептид» и «белок» используются взаимозаменяемо и означают соединение, состоящее из аминокислотных остатков, ковалентно связанных пептидными связями. Белок или пептид должны содержать по крайней мере две аминокислоты, и на максимальное число аминокислот, которые могут входить в белковую или пептидную последовательность, не налагается никаких ограничений. Полипептиды включают любой пептид или белок, состоящий из двух или более аминокислот, соединенных друг с другом пептидными связями. Согласно употреблению в данном документе этот термин означает и короткие цепочки, которые также обычно называют пептидами, олигопептидами и олигомерами в данной области техники, например, и длинные цепи, которые в данной области техники обычно называют белками, к которым относится множество типов. «Полипептиды» включают, например, биологически активные варианты полипептидов, модифицированные полипептиды, производные, аналоги, гибридные белки, среди прочих. Полипептиды включают естественные пептиды, рекомбинантные пептиды, синтетические пептиды и комбинации перечисленных.
«Фармацевтически приемлемый» означает те свойства и (или) вещества, которые приемлемы для пациента с фармацевтической и (или) токсикологической точки зрения и для фармацевтического химика, занимающего производством, с физической и (или) химической точки зрения в отношении композиции, формы выпуска, стабильности, приемлемости для пациента и биодоступности. «Фармацевтически приемлемый носитель» означает среду, которая не влияет на эффективность или биологическую активность активного ингредиента(ов) и не является токсичным для хозяина, которому его вводят.
Термин «разовая упаковка» означает, что компоненты набора физически связаны в или с одной или несколькими емкостями и считаются единицей для производителя, дистрибьютора, продавца или потребителя. Емкости включают, но не ограничиваются перечисленным, пакеты, коробки, флаконы, упаковки в целлофановой оболочке, штемпелированные или прикрепленные другим путем компоненты или комбинации перечисленного. «Разовая упаковка» может также включать виртуальные компоненты. Например, набор может содержать краткие физические инструкции, имеющиеся в физической упаковке, и инструкции по доступу к более подробным инструкциям из виртуальной среды, такой как, например, Интернет-сайт.
Термин «терапевтический» согласно употреблению в данном документе означает лечение и (или) профилактику. Терапевтический эффект получают путем избежания, отсрочки, подавления, ремиссии или излечения патологического состояния, связанного с цитомегаловирусной инфекцией.
Термин «лечение» согласно употреблению в контексте настоящего изобретения означает терапевтическое вмешательство, а также профилактические или сдерживающие меры в отношении заболевания или нарушения. Таким образом, например, термин «лечение» включает введение какого-либо агента до начала или после начала какого-либо заболевания или нарушения, что тем самым предупреждает или устраняет все симптомы заболевания или нарушения. В качестве другого примера введение этого агента после появления клинических проявлений заболевания для устранения симптомов включает «лечение» этого заболевания. Оно включает, например, предупреждение распространения цитомегаловируса в неинфицированных клетках организма. Фраза «подавление цитомегаловирусной инфекции» иногда используется в данном документе и означает метод, который включает снижение уровня инфекции у пациента цитомегаловирусом, как определяют средствами, известными врачам.
Описание:
Цитомегаловирус инфицирует множество клеточных типов in vivo, включая эпителиальные клетки, эндотелиальные клетки и фибробласты. Как суммировано выше в материале о предпосылках, в различных исследованиях сообщается, что вирус сливается с плазматическими мембранами фибробластов, но проникает в клетки пигментного эпителия сетчатки и эндотелиальные клетки пупочной вены путем эндоцитоза. Из-за относительной простоты культивирования цитомегаловируса в культуре фибробластов по сравнению с культурами эпителиальных или эндотелиальных клеток исследования, такие как суммированные выше, проведены с использованием штаммов цитомегаловируса, культивированного в фибробластах. Также культура фибробластов - типичный наилучший клеточный тип для культивирования цитомегаловируса для клинических целей, таких как получение аттенуированных штаммов вируса для вакцин.
На сегодняшний день в соответствии с настоящим изобретением установлено, что клеточный тип, в котором образуются частицы цитомегаловируса, существенно влияет на поведение последующих циклов инфекции. Так, например, хотя выше указано, что цитомегаловирус проникает в эпителиальные клетки путем эндоцитоза, авторы настоящего изобретения продемонстрировали этот путь проникновения цитомегаловируса, культивированного в фибробластах, но не цитомегаловируса, выращенного в культуре эпителиальных клеток. Цитомегаловирус, выращенный в эпителиальных клетках, проникает в эпителиальные клетки преимущественно путем слияния с плазматической мембраной. Такой особый путь проникновения влечет ряд физиологических последствий: он влияет на кинетику, с которой распространяется инфекция, и значительно влияет на клеточный ответ на инфекцию. Например, вирус, выращенный в эпителиальных клетках, вызывает резко сниженный клеточный ответ по сравнению с клетками, инфицированными вирусом, выращенным в фибробластах. Многие клеточные антивирусные гены, экспрессирующиеся после инфекции вирусом, выращенным в фибробластах, не экспрессируются после инфекции, вызванной вирусом, выращенным в эпителиальных клетках. В результате цитомегаловирус, выращенный в эпителиальных клетках, должен теоретически вести себя иначе, чем вакцина, произведенная из цитомегаловируса, выращенного в фибробластах, что предлагает новый и неожиданный источник диверсификации в получении вакцины против цитомегаловируса. Аналогично культивирование цитомегаловируса в других клеточных типах, таких как эндотелиальные клетки или специализированные клеточные типы, которые цитомегаловирус способен инфицировать (например, нейроны, другие клетки центральной или периферической нервной системы, гладкомышечные клетки, гепатоциты, стромальные клетки, макрофаги или дендритные клетки), должно давать дополнительные новые источники диверсификации в получении вакцины против цитомегаловируса.
Итак, один аспект настоящего изобретения относится к способам получения вакцин против цитомегаловируса, где используется вариабельность, связанная с выбором клеточного типа, в котором предстоит культивировать вирус. Второй аспект относится к набору для осуществления вышеописанных способов. Другой аспект настоящего изобретения относится к композиции вакцин для профилактики или лечения цитомегаловирусной инфекции и способам иммунизации особи с использованием таких композиций. Различные варианты этих аспектов настоящего изобретения изложены ниже.
Способы получения вакцин против цитомегаловируса:
Эти способы согласно одному аспекту настоящего изобретения включают: 1) получение штамма или изолята цитомегаловируса; 2) культивирование этого штамма или изолята в клеточной культуре выбранного клеточного типа, и 3) извлечение вирионов цитомегаловируса, полученного путем выращивания в этом клеточном типе (называемом в данном документе как «кондиционированный клеточным типом цитомегаловирус») для использования в получении вакцины против цитомегаловируса.
Клеточный тип, выбранный для культивирования цитомегаловируса до его использования в получении вакцины, может быть любой клеточной линией, восприимчивой к цитомегаловирусной инфекции и продуцирующей какое-либо количество вирусных частиц. Эти вирусные частицы могут быть высокоинфекционными в некоторых анализах, или эти частицы могут проявлять низкую инфективность или не проявлять инфективность вовсе во многих анализах. Подходящие клеточные типы включают, но не ограничиваются перечисленным: 1) линии эпителиальных клеток, такие как ARPE-19, которая приведена в данном документе в качестве примера, и другие линии пигментного эпителия сетчатки, например, линия эпителиальных клеток К-1034 (Ando, Y.. et al. 1997, Arch. Virol. 142(8): 1645-1658); HCMC - производное нормальной слизистой толстой кишки человека (Smith, JD, 1986, J Virol. 60(2): 583-588); клетки кишечного эпителия Сасо-2 (Esclatine. A., et al.. 2000, J. of Virol. 74 (1): 513-51); SW480, HCT1 16, HeLa, H 1299 и MCF-7 (в отношении последних пяти см. Wang. D. & Т. Shenk, 2005. J.Virol. 79: 10330); 2) линии эндотелиальных клеток, такие как НМЕС-I - линия эндотелиальных клеток микрососудов человека, иммортализованных с помощью большого Т-антигена вируса SV-40 (Guetta. Е.. et al., 2001, Cardiovascular Research 50: 538-546); HUVEC и LMVEC (в отношении последних- двух см. Wang. D. & Т. Shenk, 2005, J. Virol. 79: 10330); 3) нейроны, такие как SK-N-SH, SK-N-AS и IMR-32 (см. Wang, D. & Т. Shenk, 2005. J. Virol. 79: 10330), а также первичные эпителиальные, эндотелиальные, гладкомышечные клетки, макрофаги и дендритные клетки - производные различных тканей и органов.
Любой цитомегаловирус или комбинация цитомегаловирусов, приемлемая для разработки в качестве вакцины, пригодны для использования как источник цитомегаловируса для данного способа постольку, поскольку их можно культивировать по крайней мере в одном выбранном клеточном типе. В одном варианте цитомегаловирус является человеческим цитомегаловирусом, или изолятом, которые был выделен ранее и охарактеризован, или новым изолятом человеческого цитомегаловируса, или вирусом, подобным человеческого цитомегаловирусу. В другом варианте цитомегаловирус происходит от другого примата, включая, но не ограничиваясь перечисленным, шимпанзе (Davison, AJ et al. 2003, J. Gen. Virol. 84: 17-28) и макак-резус (Hansen. SG et al. 2003. J. Virol. 77:6620-36: Rivailler. P et al. 2006. J.Virol. 80:4179-82). Этот цитомегаловирус может быть немодифицированным вирусом из выбранного источника, или он может быть химерным вирусом, полученным путем генетической модификации элементов двух или больше различных штаммов или изолятов цитомегаловируса.
Методики получения химерных вирусов известны в данной области техники. В этом отношении по крайней мере шесть штаммов человеческого цитомегаловируса клонированы как инфекционные бактериальные искусственные хромосомы (БИХ) и секвенированы (Murphy. Е et al. 2003. Proc. Natl. Acad. Sci. USA 100: 14976-14981. Последовательности БИХ имеются в Банке генов под входящими номерами АС146999 (лабораторный штамм AD169, из которого получен вариант BADrUL131, описанный в данном документе); АС146851 (лабораторный штамм Towne); АС146904 (клинический изолят РН); АС146905 (подобный клиническому изоляту Toledo); АС 146906 (клинический изолят TR); и АС146907 (клинический изолят FIX). По крайней мере два штамма человеческого цитомегаловируса секвенированы без предварительного клонирования в БИХ и имеются в Банке генов под входящими номерами ВК000394 (лабораторный штамм AD 169) и AY446894 (клинический изолят Merlin). Весь геном штамма цитомегаловирус шимпанзе имеется в Банке генов под входящим номером AF480884. Также имеется геномная последовательность двух штаммов цитомегаловируса макак-резус (входящие номера AY186194 и DQ205516). Используя идеи, изложенные в настоящей заявке, специалист в данном области техники сможет использовать любую из вышеупомянутых последовательностей, или любую другую общедоступную последовательность цитомегаловируса для получения химерных цитомегаловирусов или цитомегаловируса, измененного каким-либо генетическим путем.
В соответствии с настоящим изобретением продемонстрировано, что лабораторные, штаммы цитомегаловируса, которые были многократно перевиты в фибробластах, можно успешно кондиционировать путем культивирования в выбранной клеточной линии. Например, как описано в примере в данном документе, BADrUL131 - БИХ-клон многократного перевитого штамма человеческого цитомегаловируса AD169, в котором изменен UL1310RF - введен путем электропорации в культуру фибробластов крайней плоти человека, и полученный вирус был амплифицирован однократно в линии эпителиальных клеток ARPE-19. Таким образом, различные варианты настоящего изобретения включают использование цитомегаловируса (или геномов цитомегаловируса), перевитых в клеточных типах, отличающихся от клеточного типа, выбранного для этапа кондиционирования. Например, штамм цитомегаловируса может быть многократно перевит в фибробластах, затем амплифицирован в эпителиальных клетках и впоследствии использован для получения вакцины. Понятно, что этот цитомегаловирус может быть амплифицирован и (или) культивирован на протяжении одного или нескольких циклов в выбранном клеточном типе.
В предпочтительных вариантах способы настоящего изобретения используются для получения живого аттенуированного цитомегаловируса для применения в качестве вакцины. Методики аттенуирования вирусов известны в данной области техники. Предпочтительно, чтобы аттенуированный цитомегаловирус проявлял сниженную способность к инфективности, и (или) патогенности, включая латентное действие и активацию, но оставался способным вызывать иммунный ответ, который оказывает терапевтический эффект или защищает от цитомегаловирусной инфекции. Примеры аттенуированных штаммов цитомегаловирусов включают, но не ограничиваются перечисленным, лабораторные штаммы, такие как AD169 и Towne, которые реплицируются практически только в фибробластах. Такие аттенуированные штаммы, которые создают генно-инженерными методами при необходимости, если требуется получить нужный поверхностный белок или белковые комплексы для соответствующего тропизма, можно культивировать в эпителиальных клетках или фибробластах и впоследствии в эпителиальных клетках, как обсуждено выше, для использования в композиции вакцин настоящего изобретения.
Для аттенуирования цитомегаловируса можно использовать серийный пассаж в клеточных культурах, особенно в фибробластах. Многократный пассаж инфицированных вирусом клеток хозяина проводят in vitro до тех пор, пока не будет достигнуто нужное аттенуирование вируса. Пассаж можно проводить при определенных параметрах внешней среды, таких как регулируемая температура, pH или влажность, чтобы выбрать вирусы со сниженной инфективностью или патогенностью. Если используется такая методика аттенуирования, серийно перевитый вирус затем амплифицируют в выбранном клеточном типе на протяжении одного или нескольких пассажей для получения цитомегаловируса, предназначенного для применения в композиции вакцин настоящего изобретения.
Для аттенуирования вируса также можно использовать мутагенез. Например, вирионы цитомегаловируса можно подвергнуть воздействию ультрафиолетового или ионизирующего излучения или химических мутагенов в соответствии с методиками, известными в данной области техники. Кроме их использования для получения химерных вирусов, рекомбинантные методики также можно применять для получения вирионов аттенуированного цитомегаловируса. Например, для получения штаммов со сниженной инфективностью, патогенностью или латентностью можно использовать такие методики, как сайт-специфический мутагенез, замена гена или выключение гена. Пример модификации цитомегаловируса путем мутагенеза с выключением гена изложен в патенте WO/2007/038316, где описаны цитомегаловирусы с геномами с делецией одного или нескольких генов, обусловливающих латентность, что проявляется в измененной способности проникать в клетку или поддерживать латентное состояние.
В других вариантах цитомегаловирусы, выделенные из выбранных клеточных культур, являются инактивированными или убитыми и используются в композиция вакцин. Методики инактивации или получения убитых вирусов, например, с помощью таких химических соединений как формалин, хорошо известны в данной области техники. Специалисту в данной области техники будет понятно, что этот убитый или инактивированный цитомегаловирус будет состоять из всех или большой доли компонентов вирусной частицы так, что в композиции вакцины за счет амплификации в выбранном клеточном типе будет поддерживаться диверсификация.
Способы настоящего изобретения можно использовать для создания комбинаций цитомегаловирусов, культивирующихся в различных выбранных клеточных типах, таким образом обеспечивая дополнительную степень диверсификации производящимся вакцинам. В одном варианте для инфицирования одной или нескольких различных линий клеточных культур различных клеточных типов, например, клеток эпителия сетчатки и эндотелиальных клеток, используется один изолят или штамм цитомегаловируса. Цитомегаловирус, полученный путем амплификации в соответствующих клеточных типах, затем комбинируют для использования в одной вакцине. В другом варианте, два или больше различных клинических изолята или штамма цитомегаловируса используют для инфицирования одной выбранной клеточной линии, и популяцию из множества штаммов или изолятов цитомегаловируса, полученную путем амплификации в этом клеточном типе, используют для получения вакцины. Еще в одном варианте для инфицирования двух и больше различных линий клеточных культур различных клеточных типов используются множественные изоляты или штаммы, и популяцию цитомегаловирусов, полученную путем амплификации в соответствующих клеточных типах, комбинируют для использования в этой вакцине.
Другой аспект настоящего изобретения относится к наборам для получения материалов вакцины против цитомегаловируса согласно вышеописанным способам. Эти наборы помещены в отдельные емкости в единице упаковки или в отдельных емкостях в виртуальной упаковке, согласно пути применения и компонентам набора, порциям клеточных линий одного или нескольких выбранных клеточных типов, а также одного или нескольких изолятов или штаммов цитомегаловируса, или векторов, несущих геномы таких штаммов цитомегаловируса, для введения и амплификации в выбранных культурах клеточных линий. Такие наборы также обычно содержат инструкции, или ссылки на инструкции, как осуществлять различные этапы данного способа. По желанию заказчика наборы также могут включать питательную среду и другие реагенты, пригодные для культивирования клеток и других манипуляций с вирусом.
Композиции вакцины и способы применения:
Другой аспект настоящего изобретения относится к иммуногенной композиции (именуемой в данном документе взаимозаменяемо композицией вакцины), состоящей из популяции цитомегаловируса или компонентов его вириона, смешанной с соответствующим фармацевтическим носителем или адъювантом, где цитомегаловирус получают путем культивирования в выбранном клеточном типе, например, культуре эпителиальных клеток. Как упомянуто выше, вакцины против цитомегаловируса прежде производили с использованием цитомегаловируса, культивированного в фибробластах. Однако в соответствии с настоящим изобретением продемонстрировано, что культивирование в эпителиальных клетках дает вирус, который отличается от вируса, культивированного в фибробластах, во многих отношениях. Вирус, полученный в эпителиальных клетках, преимущественно сливается с плазматической мембраной, в то время как вирус, культивированный в фибробластах, проникает путем эндоцитоза, опосредованного рецептором. Кроме того, вирионы, полученные в эпителиальных клетках, обладают более выраженной активностью в отношении слияния клеток хозяина без репликации вируса, чем частицы, полученные в фибробластах, что влияет на кинетику инфекции. Кроме того, две разновидности вируса запускают разные клеточные сигнальные ответы, о чем свидетельствуют различные изменения профиля транскрипции в инфицированных эпителиальных клетках.
В частности, цитомегаловирус, полученный путем культивирования в эпителиальных клетках, обладает одним или несколькими из следующих свойств по сравнению с эквивалентным штаммом или изолятом вируса, полученного путем культивирования в фибробластах. Во-первых, как упомянуто выше, их можно отличить по проникновению клетки хозяина путем слияния с плазматическими мембранами клетки хозяина. Цитомегаловирус, полученный в эпителиальных клетках, также вызывает более выраженное слияние клеток хозяина, опосредованное вирионом, по сравнению с эквивалентной популяцией цитомегаловируса, выделенной из культуры фибробластов, а также более быструю репликацию в клетках хозяина по сравнению с эквивалентной популяцией цитомегаловируса, выделенной из культуры фибробластов. Кроме того, этот вирус вызывает менее выраженный клеточный ответ по сравнению с эквивалентным цитомегаловирусом, выращенным в фибробластах. Через 10 ч после инфекции примерно на две трети меньше генов (-50 против -150 генов) проявляет 2,5-кратное или больше изменение уровня экспрессии. Кроме того, цитомегаловирус, выращенный в эпителиальных клетках, можно охарактеризовать определенным профилем генов хозяина, чья экспрессия после инфекции изменяется (увеличивается или уменьшается). Такие профили экспрессии генов приведены в примере и могут включать изменение экспрессии одного или нескольких генов, представленных в Банке генов под входящими номерами: АК094860, NM_145023, NM-133492, NM_001039580. NM_001004301, NM-001034, А1369525, АК123066, NM_005345, NM_020731, BC071797, NM-003414, NM_000800, NM_138467, AK090803, AL133118, NM-00165, BG001037, NM_024861, NM-001043, NM-016239, NM-001018084, NM-001037442, NM-017600, NM_022097, NM-175868, NM_032266, NM_003841, NM_005039, NM_04505 1, NM_004294, AW856073, NM_024050, AF085968, NM-080927, NM_022115, AK056703, NM_000808, NM-012377, NM_006793, NM_031466, NM_005185, NM-139173, BX360933, NM-016125, NM_002104, NM-032188, NM_004185, NM-004843 или NM-173550.
В этом, аспекте, как и в предшествующих аспектах настоящего изобретения, цитомегаловирус или комбинация цитомегаловирусов, пригодных для разработки в качестве вакцины, подходят для использования в качестве источника вышеупомянутой популяции цитомегаловируса постольку, поскольку их можно вырастить по крайней мере в одной линии эпителиальных клеток или другом выбранном клеточном типе. В одном варианте цитомегаловирус является человеческим цитомегаловирусом или вирусом, подобным человеческому цитомегаловирусу. В другом варианте цитомегаловирус происходит от другого примата, включая, но не ограничиваясь перечисленным, шимпанзе и макак-резус, как описано выше. Цитомегаловирус может быть немодифицрованным из выбранного источника, или он может быть химерным вирусом, полученным путем генетической модификации или комбинации элементов двух или больше различных штаммов или изолятов цитомегаловирусов, как описано выше.
В предпочтительных вариантах композиция вакцин включает живой аттенуированной цитомегаловирус, который можно получить способами, описанными выше, все из которых известны специалисту в данной области техники. В других вариантах цитомегаловирус, выделенный из выбранных клеточных культур, является инактивированным или убитым и используются в композиции вакцины.
Композиция вакцины может включать комбинации различных штаммов или изолятов цитомегаловируса, который можно культивировать в одной культуре эпителиальных клеток или в нескольких различных культурах эпителиальных клеток, или в клетках другого клеточного типа, для обеспечения дополнительной диверсификации.
Кроме того, живой аттенуированный цитомегаловирус можно комбинировать с убитым или инактивированным цитомегаловирусом, или с иммуногенными компонентами цитомегаловируса для получения комбинации вакцины, например, живого аттенуированного цитомегаловируса, комбинированного с убитым высокой температурой цитомегаловирусом, или комбинированного с материалом для субъединицы вакцин, или комбинации всех трех типов материалов. Примеры иммуногенных полипептидов цитомегаловируса и комплексов, пригодных для субъединицы вакцин, описаны в WO 2007/146024 под названием «Комплекс поверхностных белков цитомегаловируса для использования в вакцинах и в качестве мишени для лекарственных средств».
Кроме того, композиция вакцины может включать один или несколько адъювантов. Адъювантом может быть любое вещество, усиливающее иммунный ответ на антигены вакцины. Примеры адъювантов, пригодных для использования в настоящем изобретении, включают, но не ограничиваются перечисленным, адъювант Фройнда, неполный адъювант Фройнда, сапонин, сурфактанты, такие как гексадециламин, октадециламин, лизолецитин, деметилдийодактадецил-аммония бромид, N,N-диоктадецил-N'-N-бис(2-гидроксиэтилпропандиамин), метоксигекса-децил-глицерин, плюроновые полиолы, полианионы, такие как пиран, диэтиламиноэтил (ДЭАЭ) декстран, декстран-сульфат, полибрен, полиинозиновая-полицитидиловая кислота, полиакриловая кислота, карбопол, этиленмалеиновая кислота, гидроксид алюминия, и алюминий-фосфатные пептиды, масляные или углеводородные эмульсии и им подобные.
Вакцины можно выпускать в виде водных растворов, таких как водные или спиртовые, или в физиологически совместимых буферных растворах, таких как раствор Хэнкса, раствор Рингера или физиологический раствор, включая фосфатный буфер. Также вакцины можно выпускать в виде твердых форм, которые предназначены для превращения непосредственно перед применением в жидкие формы, пригодные для введения субъекту, например, путем разведения соответствующим наполнителем, таким как стерильная вода, солевой раствор или спирт.
Композиции вакцин также можно выпускать с использованием наполнителей, обеспечивающих пролонгированное высвобождение, или в виде депо-форм. Такие пролонгированные формы можно вводить путем имплантации (например, подкожно или внутримышечно) или путем внутримышечной инъекции. Так, например, вакцины можно выпускать вместе с соответствующими полимерными или гидрофобными материалами (например, в виде эмульсии в соответствующем масле) или ионообменными смолами, или в виде трудно растворимых производных, например, в виде трудно растворимых солей. Липосомы и эмульсии можно использовать в качестве носителей, пригодных для применения с гидрофобными формами. Наполнители пролонгированного действия в зависимости от своей химической природы могут высвобождать антигены на протяжении периода от нескольких часов до нескольких суток и до нескольких недель или нескольких месяцев.
Композиции вакцины могут также включать один или несколько антиоксидантов. Примеры восстанавливающих агентов включают меркаптопропионилглицин, N-ацетилцистеин, β-меркаптоэтиламин, глутатион, аскорбиновую кислоту и ее соли, сульфит или метабисульфит натрия и подобные соединения. В дополнительные антиоксиданты также могут быть включены натуральные антиоксиданты, такие как витамин Е, С, лютеин, ксантин, бета-каротин и минералы, такие как цинк и селен.
Композиции вакцин могут также включать дополнительные вещества, действующие как стабилизаторы, консерванты, буферы, увлажняющие агенты, эмульсифицирующие агенты, диспергирующие агенты и моносахариды, полисахариды и соли для контроля осмотического равновесия. Эти вакцины могут также дополнительно включать молекулы иммуностимуляторов для усиления эффективности вакцины. Такие молекулы могут усиливать иммунный ответ, могут вызывать воспаление и могут быть каким-либо лимфокином или цитокином. Примеры цитокинов включают, но не ограничиваются перечисленным, интерлейкин (ИЛ) 1, ИЛ-2, ИЛ-3, ИЛ-4, ИЛ-12, ИЛ-13, колониестимулирующий фактор гранулоцитов-макрофагов (КСФГМ), макрофагальный воспалительный фактор и им подобные.
Вакцины можно выпускать для и вводить путем инфузий или инъекций (внутривенно, внутриартериально, внутримышечно, внутрикожно, подкожно, интратекально, интрадуоденально, интраперитонеально и тому подобное). Вакцины также можно вводить интраназально, вагинально, ректально, внутрь, местно, для введения на слизистую щеки, на слизистую или трансдермально.
Эффективную дозу антигена для лечения цитомегаловирусной инфекции можно определить эмпирически, с помощью методик, хорошо известных в данной области техники. Эффективная доза вакцины может зависеть от ряда показателей, включая, но не ограничиваясь перечисленным, размер, рост, вес, возраст, пол, общее состояние здоровья субъекта, тип формы выпуска, путь или методику введения, является ли вирус активным или латентным, страдает ли пациент вторичными инфекциями или другими сопутствующими заболеваниями.
Схемы применения вакцин тоже могут быть основаны на вышеупомянутых факторах. Вакцинацию можно проводить в любое время в течение жизни субъекта, от развития плода до взрослого состояния включительно. Для полной защиты может требоваться дополнительное введение, или бустеры. Чтобы определить, достигнут ли достаточный уровень иммунной защиты, у пациента после вакцинации можно следить за сероконверсией и титром антител.
Следующий пример приведен для более подробного описания настоящего изобретения. Он предназначен для иллюстрации настоящего изобретения, но не налагает каких-либо ограничений.
ПРИМЕР
Человеческий цитомегаловирус использует два разных пути для проникновения в клетки пигментного эпителия сетчатки
Экспериментальные результаты, описанные в данном примере, демонстрируют, что человеческий цитомегаловирус, полученный в двух различных клеточных типах, проникает в эпителиальные клетки различными путями. Вирионы, образовавшиеся в эпителиальных клетках, проникают преимущественно путем слияния с плазматической мембраной, в то время как вирионы из фибробластов проникают путем pH-зависимого эндоцитоза. Эти две разновидности вируса вызывают существенно различающиеся клеточные ответы.
Материалы и методы
Биологические реагенты. Фибробласты крайней плоти человека (ФКПЧ) при пассаже от 10 до 15 раз держали в среде с 10% сывороткой новорожденного теленка. Фибробласты легких человеческого плода MRC-5 и клетки пигментного эпителия сетчатки ARPE-19 (Американская коллекция типов культур) при пассаже от 24 до 34 раз держали в среде с 10% сыворотки плода коровы. Клетки эпителия проксимальных почечных канальцев человека (чКЭППК) (Cambrex) выращивали в среде с 10% сыворотки плода коровы и использовали при пассаже от 4 до 5.
BADwt является производным БИХ-клона штамма человеческого цитомегаловируса AD169; BADrUL131 (19, 21) является производным BADwt, в котором изменен UL1310RF; BFXwt является производным БИХ-клона клинического изолята человеческого цитомегаловируса VR1814. Вирусы готовили путем электропорацйи ДНК БИХ в ФКПЧ, и полученный вирус амплифицировали однократно в клетках ARPE-19 или ФКПЧ, если не указано иначе. Вирионы, не содержащие клеток, частично очищали путем центрифугирования через сорбитоловую прокладку и вновь взвешивали в бессывороточной среде. Титры вируса определяли с помощью анализа на плашке на клетках MRC-5. Нейтрализацию BADrUL131 оценивали с помощью анализа с восстановлением на плашке (19) с использованием очищенного моноклонального антитела к pUL130 (19).
Моноклональное антитело к IE1 IB12 описано ранее (21). Кроличье поликлональное антитело к Sp100 (Chemicon) использовали для визуализации ND10.
Электронная микроскопия. Клетки ARPE-19 подвергали воздействию вируса при 4°С в течение 1 ч, не связанный вирус удаляли двукратным промыванием холодным фосфатным буфером, добавляли питательную среду на 15 мин (37°С), клетки промывали фосфатным буфером, фиксировали и готовили для электронной микроскопии и просматривали под микроскопом FEI Tecnai-T12 при 80 кВ.
Анализ зависимости инфекции от окисления эндосом. Клетки ARPE-19 предварительно обрабатывали NH4G или бафломицином А1 (БФА) Sigma) в течение 1 ч при 37°С, после чего через 16 ч инфицировали в присутствии ингибитора, культуры фиксировали в 2% формальдегиде и пермеабилизировали 0,1% Triton Х-100. IE1 идентифицировали с помощью иммунофлуоресценции с использованием моноклонального антитела IB12 (21) и дополнительного антитела, конъюгированного с Alexa 546, и окрашивали ядра с помощью 4,6-диамидино-2-фенилиндол дигидрохлорида. Торможение рассчитывали в виде процента экспрессии IE1 в обработанных клетках относительно необработанных.
Анализ активности слияния белков вириона. Для анализа слияния клеток без репликации вируса клетки ARPE-19 выращивали до 90% слияния и инфицировали. Через 1 ч при 37°С инокулюм удаляли, и добавляли среду, содержащую 200 мкг/мл фосфономуравьиной кислоты (ФМК), для торможения синтеза вирусной ДНК. Слияние отслеживали визуально, оценивая образование синцития.
Анализ с репортером люциферазой адаптировали для количественного анализа активности слияния вирионов. Репортерные и эффекторные клетки ARPE-19 готовили путем электропорации (90-95% эффективность) с использованием плазмиды, несущей ген люциферазы, регулируемый промотором Т7, и плазмиды с полимеразой pcDNA3-17 соответственно. К 24 ч после инфицирования клетки смешивали в соотношении 1:1 и инкубировали при 37°С на протяжении еще 16 ч. Затем на смешанную популяцию воздействовали вирионами человеческого цитомегаловируса при 4°С в течение 1 ч, после чего монослой отмывали дважды холодным фосфатным буфером, а затем добавляли буферные растворы (фосфатный буфер с 10 мМоль 2-(N-морфолино)этансульфоновой кислоты и 10 мМоль HEPES) до конечного pH в диапазоне от 4,5 до 8. Через 3 мин при 37°С буферные растворы удаляли и добавляли обычную питательную среду. Через 6 ч после инфицирования клетки лизировали и измеряли активность люциферазы с использованием люциферазной репортерной системы (Promega).
Анализ клеточных транскрипционных ответов. Слившиеся клетки ARPE-19 держали на бессывороточной среде на протяжении 24 ч, после чего проводили симуляцию инфицирования и настоящее инфицирование. Всю РНК экстрагировали через 6 или - 10 ч после инфицирования с использованием Тризола (Invitrogen) и очищали с помощью колонки RNeasy (Qiagen). Образцы РНК амплифицровали и метили (цианин-3) с использованием набора Agilent для линейной амплификации флуоресценции при низком содержании РНК. Для учета вариабельности чипов метили (цианин-5) контрольную РНК (Clontech) и когибридизовали с зондами, приготовленными из клеток, подвергнутых симуляции инфицирования или инфицированных человеческим цитомегаловирусом. Гибридизацию проводили в двух копиях с наборами олигонуклеотидов Aligent human 44К. Наборы сканировали с использованием сканера Agilent с разрешением 5 микрон и изображения анализировали с помощью программы Agilent Feature Extraction для определения интенсивностей сигналов флуоресценции гибридизованных пятен и вычитания фона. Для нормализации и количественного определения относительных изменений РНК использовали программу Agilent GeneSpring GX.
Результаты
Вирионы, полученные в фибробластах, активируют немедленную-раннюю экспрессию генов в клетках ARPE-19 с кинетикой более медленной, чем вирионы, полученные в эпителиальных клетках. Штамм человеческого цитомегаловируса AD169 (BADwt) плохо реплицируется в эпителиальных клетках ARPE-19 из-за мутации в гене UL131 (10, 21). Репарация мутации в AD169 с получением BADrUL131 восстанавливает тропизм к эпителиальным клеткам (21), что обеспечивает выработку гликопротеинового комплекса вириона gH/gL/pUL128/pUL 130/pUL131, который требуется для успешного проникновения в эти клетки (19, 20).
BADrUL131, выращенный в эпителиальных клетках ARPE-19 (epiBADrUL131), запускает свою программу экспрессии генов в эпителиальных клетках быстрее, чем BADrUL131, выращенный в фибробластах КПЧ (fibroBADrUL131) (фиг.1А). Когда клетки ARPE-19 инфицировали epiBADrUL131, 17% клеток вырабатывали измеряемый белок IE1 через 6 ч после инфицирования. Выработка IE1 сопровождалась разрушением ND10 в ядре. В то же время инфицирование fibroBADrUL131 приводило к выработке IE1 только в 2,8% клеток ARPE-19 через 6 ч после инфицирования. Однако число вырабатывающих IE1 клеток со временем увеличивалось. Статистически значимых различий в проценте клеток ARPE-19, вырабатывающих IE1, через 24 ч после инфицирования вирусом, выращенным в двух клеточных типах, не отмечалось (фиг.1 В).
Вирионы, полученные из ФКПЧ, в отличие от вирионов, полученных в клетках ARPE-19, проникают в клетки ARPE-19 разными путями. Электронная микроскопия проникновения вируса проведена с тем, чтобы определить, являются ли различия кинетики накопления IE1 в случае вируса, выращенного в клетках ARPE-19, и вируса, выращенного в ФКПЧ, следствием некого события, предшествующего началу экспрессии вирусных генов. Клетки ARPE-19, инкубированные с epiBADrUL131 или fibroBADrUL131, оставляли при 4°С, чтобы произошло прикрепление вируса к клеточной поверхности, а затем культуры помещали на 15 мин при 37°С, чтобы обеспечить интернализацию до подготовки к электронной микроскопии. В каждом образце исследовали 40-50 клеток, при этом по крайней мере в 90% клеток визуализировали целые вирионы или капсиды. Число вирусных частиц в каждой клетке колебалось от 2 до 8, при этом большинство клеток содержали 2-3 частицы.
В клетках ARPE-19, инфицированных epiBADrUL131, вирионы обнаруживались почти исключительно на клеточной поверхности, при этом 97% вирионов находились на апикальной мембране. Некоторые частицы располагались близко к клеткам, но на этом срезе признаков контакта не выявлено (фиг.2А, блок а), а другие визуализированы в процессе слияния с плазматической мембраной (фиг.2А, блоки b и с). Капсиды рядом с внутренней поверхностью мембраны наблюдали редко; на самом деле обнаружено только два примера (фиг.2А, блоки d и е). Внутри клеток не обнаружено ни одного вириона в оболочке. Этот результат указывает, что epiBADrUL131 проникает в клетки ARPE-19 путем слияния с плазматической мембраной. И наоборот, клетки, инфицированные fibroBADrUL131, содержали вирионы на клеточной мембране (65% от общего числа) и внутри клетки в везикулах (35% от общего числа) (фиг.2 В). Частицы внутри пузырьков были в оболочке, что указывало на их путь проникновения - эндоцитоз.
Также исследовано проникновение клинического изолята BFXwt, выращенного в фибробластах. Этот клинический изолят накапливался в везикулах в клетках ARPE-19 (фиг.2С), что свидетельствовало в пользу BADrUL131 в качестве модели проникновения в клетку клинического изолята человеческого цитомегаловируса.
Инфицирование клеток ARPE-19 вирусом, выращенным в фибробластах, но не в эпителиальных клетках, зависит от pH. Многие вирусы, которые проникают в клетку путем эндоцитоза (1, 4, 10), требуют окисления эндосом для того, чтобы оболочка вириона слилась с эндосомальной мембраной и высвободила капсид в цитоплазму. NH4Cl, который забуферивает эндосомальный pH, и бафломицин А1 (БФА), который блокирует эндосомальный АТФазный протонный насос, исследованы на предмет их влияния на инфицирование клеток ARPE-19. После предварительной обработки тем или другим агентом клетки инфицировали и культивировали в среде, содержащей лекарственное средство, на протяжении дополнительных 16 ч. Успешное инфицирование оценивали количественно по числу IE1-положительных клеток. В соответствии с вышеописанным ультраструктурным анализом предварительная обработка тем или иным агентов оказывала лишь умеренный эффект на инфицирование epiBADrUL131 (фиг.3А). В то же время оба агента тормозили выработку IE1 после инфицирования fibroBADrUL131 доза-зависимым образом, что указывало на зависимость проникновения вируса, выращенного в фибробластах, от окисления эндосом. Тот факт, что эти агенты слабо влияли на проникновение epiBADrUL131, указывает, что торможение fibroBADrUL131 не является результатом токсического действия.
Затем было исследовано, проявляет ли вирус, выращенный в других типах эпителиальных клеток и фибробластах, те же самые свойства, что и вирионы, полученные в клетках ARPE-19 и ФКПЧ. Для инфицирования клеток ARPE-19 после обработки NH4Cl или БФА использовали готовые вирусы из эпителиальных клеток чКЭППК и фибробластов MRC-5, и в этих клетках эффект ингибиторов оказался точно таким же, как в случае вируса, выращенного в клетках ARPE-19 или ФКПЧ (фиг.3В, левый блок). Таким образом, BADrUL131, полученные в двух различных типах фибробластов, оказались гораздо более чувствительными к ингибиторами, чем вирус, выращенный в двух различных линиях эпителиальных клеток.
Также исследовано влияние pH эндосом на проникновение клинического изолята BFXwt в клетки ARPE-19 (фиг.3В, правый блок). NH4Cl или БФА существенно снижали число IE1 - положительных клеток ARPE-19, полученных с использованием BFXwt, выращенного в фибробластах, но после инфицирования BFXwt, выращенного в эпителиальных клетках, отмечено лишь слабое торможение.
Вирионы, полученные в эпителиальных клетках, обладают более выраженной активностью слияния, чем вирионы, полученные в фибробластах. Как и в случае других герпесвирусов, клинические изоляты стимулируют слияние клеток между собой, что можно выявить уже через 3-5 ч после инфицирования. Быстрое образование синцития без синтеза новых белков вирусной оболочки указывает, что этот процесс запускается слиянием клеток без репликации вируса, когда вирионы в оболочке прямо стимулируют слияние клеток-мишеней. Поскольку BADrUL131, выращенные в эпителиальных клетках против фибробластов, проникают в эпителиальные клетки разными путями, исследована возможность, что эти будут проявлять различную активность слияния клеток без репликации вируса.
Клетки ARPE-19, подвергнутые симуляции инфицирования, не проявляли признаков образования синцития (фиг.4А), и синцитий редко обнаруживался после инфицирования fibroBADrUL131 (фиг.4В). Наоборот, после инфицирования epiBADrUL13I, слияние клеток между собой выявлялось уже через 6 ч после инфицирования, и 20-30% ядер агрегировались в синцитии к 24 ч после инфицирования (фиг.4С). Клетки обрабатывали БФА, который блокирует прогрессирование до поздней стадии инфекции, поэтому слияние, должно быть, являлось результатом воздействия частиц epiBADrUL131 и не вновь образованных белков вириона.
Анализ с репортером люциферазой использовали для количественной оценки активности слияния вирусных частиц, а также влияния pH на слияния клеток без репликации вируса. Репортерные и эффекторные клетки получали плазмиду, содержащую ген люциферазы, контролируемый промотором Т7, или плазмиду с экспрессией РНК-полимеразы Т7. Два производных клеток ARPE-19 смешивали и по выработке люциферазы количественно оценивали слияние клеток, зависимое от инфекции. epiBADrUL131 неизменно вызывал более высокую активность слияния, чем fibroBADrUL131 (фиг.4Г). При pH 7-8 активность fibroBADrUL131 была в 3 раза ниже, чем активность epiBADrUL131. Когда клетки обрабатывали буферными растворами с низким pH после адсорбции вируса, обе разновидности вируса вызывали умеренное усиление слияния. BADwt в этом анализе слияние не вызывал.
Путь проникновения не влияет на клеточный тропизм человеческого цитомегаловируса. Как обсуждалось выше, существует прецедент в отношении герпесвируса, когда отмечается склонность вируса проникать в определенный клеточный тип в зависимости от клеток, в которых выращен инфекционный вирус. Это явление отличается от вышеописанного, т.е. когда две разновидности человеческого цитомегаловируса из двух разных клеточных типов проникают в эпителиальные клетки путем разных механизмов. Тем не менее, нельзя было полностью исключить, что различные механизмы проникновения могут влиять на эффективность репликации и выход вируса, что проявляется в эффекте тропизма. По этой причине проведены эксперименты, чтобы выяснить, влияет ж путь проникновения человеческого цитомегаловируса на бляшкообразование в эпителиальных клетках и фибробластах (табл.1). Исходный BADrUL131 получен в клетках ARPE-19, чКЭППК, ФКПЧ или MRC-5 и с ним проведен анализ на бляшкообразование на клетках ARPE-19 or MRC-5 (табл.1). Хотя на клетках ARPE-19 получено несколько больше бляшек, чем на клетках MRC-5, ни вирус, выращенный в эпителиальных клетках, ни вирус, выращенный в фибробластах, не проявил предпочтения в отношении бляшкообразования на каком-либо клеточном типе.
Антитело, специфичное к pUL130, блокирует инфицирование ARPE-19 вируса, выращенного в эпителиальных клетках, и вируса, выращенного в фибробластах. Антитело, специфичное к pUL130, которое нейтрализует инфекцию эпителиальных клеток, вызванную человеческим цитомегаловирусом (19), оказалось способно блокировать инфекцию ARPE-19 при любом пути проникновения (фиг.5А). Оно тормозило инфекцию, вызванную обоими вирусами, доза-зависимым путем, хотя epiBADrUL131 был несколько более чувствительным к нейтрализации, чем fibroBADrUL131. Способность этого антитела тормозить оба пути проникновения дополнительно свидетельствует в пользу вывода о том, что pUL130 - содержащий комплекс функционирует вне зависимости от того, происходит ли слияние с плазматической или эндосомальной мембраной.
Раньше уже сообщалось, что комплекс gH/gL/pUL128/pUL130/pUL131 необязателен для интернализации человеческого цитомегаловируса эндотелиальным и или эпителиальными клетками, так как в лабораторном штамме, в котором этот комплекс отсутствует, успешно подвергается эндоцитозу (10). Однако последующее слияние с эндосомальной мембраной и выход в цитоплазму требуют присутствия этого комплекса. В соответствии с этими более ранними данными антитело к pUL130 не блокировало связывание или интернализацию epiBADrUL131, fibroBADrUL131 или BADwt в анализе с клетками ARPE-19 (фиг.5В). Однако общее количество интернализованного вируса, выращенного в фибробластах, было меньше, чем количество вируса, выращенного в эпителиальных клетках. Это может отражать сниженную скорость интернализации, что может соответствовать отсрочке в начале выработки IE1 вирусом, выращенным в фибробластах (фиг.1).
epiBADrUL131 и fibroBADrUL131 индуцируют различные транскрипционные ответы в клетках ARPE-19. Как и многие другие вирусы, человеческий цитомегаловирус модулирует клеточную передачу сигнала в ходе проникновения. Одно последствие измененной передачи сигнала заключается в резком изменении клеточного транскриптома, что в большой мере обусловлено контактом гликопротеинов вириона в клетками хозяина.
Соответственно, исследовано влияние двух путей проникновения на транскрипционный ответ клеток ARPE-19. В клетках симулировали инфекцию или инфицировали их epiBADrUL131 или fibroBADrUL131 и через 6 или 10 ч очищали всю РНК. Относительные концентрации РНК анализировали с помощью микронаборов и идентифицировали РНК инфицированных клеток, чья концентрация в 2,5 раза превышала количество в клетках, подвергнутых симуляции инфицирования (табл.2-5). Распределение РНК с повышенной или сниженной экспрессией отображено на диаграммах Венна на фиг.6А.



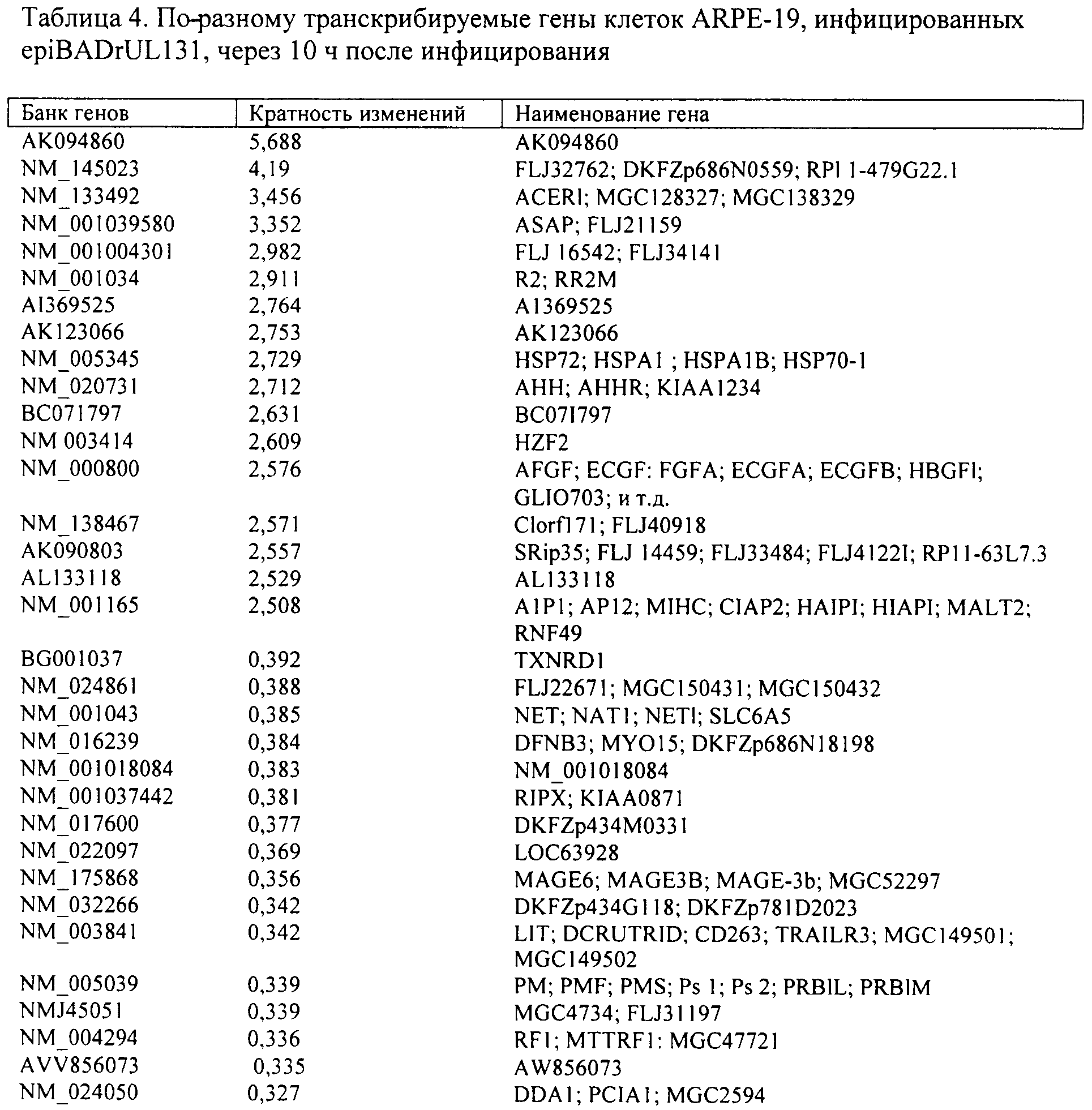





Мишени в микронаборах, которые гибридизовались с меченой РНК клеток ARPE-19, инфицированных fibroBADrUL131, сравнили с клетками, где симулировали инфицирование; перечислены зонды, где различия уровней составили 2,5 раза и больше. Приведены обозначения в Банке генов, кратность изменений и наименования генов.
Через 6 ч после инфицирования epiBADrULBl концентрации 47 РНК изменились по сравнению с клетками с симуляцией инфекции, и 121 РНК изменились в клетках, инфицированных fibroBADrUL131, по сравнению с клетками с симуляцией инфекции. Набор измененных РНК значительно отличался у двух этих вирусов; только 19 РНК изменились после инфицирования либо epiBADrUL131, либо fibroBADrUL131. Однако возможны несколько случаев, когда ген изменяется под воздействием одного вируса в 2,5 раза и больше, а другой вирус вызывает более умеренные изменения, не превышающие точку отсечения, изучение данных обнаружило, что это встречается нечасто. Через 10 ч после инфицирования число РНК в клетках хозяина под воздействием epiBADrUL13l изменилось лишь немного (50 РНК), в то время как после инфицирования fibroBADrUL131 отмечалось более существенное увеличение (153 РНК). В более поздние сроки число РНК, измененных обоими вирусами, увеличивалось в незначительной степени (28 РНК). Результаты анализов с микронаборами подтверждены с помощью ПЦР в режиме реального времени для одной РНК, которая не изменилась, и для шести РНК, измененных за счет инфекции (фиг.6 В).
Для дальнейшего сравнения изменений концентрации РНК под воздействием fibroBADrUL131 против epiBADrUL13l результаты, полученные с микронаборами, профильтровали по перечню генов, состоящего из четырех групп Онтологии генов: взаимодействие хозяин-возбудитель (GO:0030383), межклеточное взаимодействие (GO:0007154), жизненный цикл вируса (GO:0016032) и межклеточная передача сигнала (GO:0007267). Почти одна треть РНК (70 из 222), которые изменились больше чем в 2,5 раза под воздействием fibroBADrUL131, присутствовали в объединенной группе (табл.6). При этом только 86 РНК, индуцированных epiBADrUL131, обнаружены в этих четырех группах Онтологии генов. Эти две разновидности вируса существенно различались по транскрипционном ответу на инфицирование эпителиальных клеток.


Обсуждение
Эпителиальные клетки ARPE-19 можно инфицировать человеческим цитомегаловирусом двумя путями: слияния с плазматической мембраной или эндоцитоза с последующим слиянием с эндосомальной мембраной. Оба пути проникновения запускают продуктивную инфекцию. Путь проникновения зависит от клеточного типа, в котором был культивирован вирус. Человеческий цитомегаловирус из эпителиальных клеток проникает в клетки первым путем, а вирус, культивированный в фибробластах, проникает вторым путем. Этот вывод следует из ультраструктурного анализа и разной чувствительности инфекции к агентам, блокирующим окисление эндсом. Тот факт, что вирус, выращенный в эпителиальных клетках, вызывает слияние клеток без репликации вируса в большей мере, чем вирус, выращенный в фибробластах, дополнительно свидетельствует в пользу точки зрения о том, что эти две разновидности вируса взаимодействуют с клетками ARPE-19 двумя кардинально отличающимися путями. Важно, что оба пути проникновения требуют функции pUL130, так как антитело к pUL130 нейтрализовало инфицирование вирусом, полученный тем или иным путем. Комплекс gH/gL/pUL128/pUL130/pUL131 действует на плазматической мембране клеток ARPE-19, если инфицирующий вирус выращен в эпителиальных клетках, и на эндосомальной мембране, если вирус выращен в фибробластах. Нейтрализованный вирус в эндосоме не способен «сбежать» и, видимо, следует той же судьбе, что и AD169, в котором отсутствует pUL130-содержащий комплекс и который накапливается в эндосом ах эпителиальных клеток без запуска продуктивной инфекции (10).
Вирус, выращенный в фибробластах, вызывает накопление белка IE1 в клетках ARPE-19 после некоторой задержки по сравнению с вирусом, выращенным в эпителиальных клетках, и это предполагает, что в некотором смысле проникновение путем эндоцитоза происходит медленнее, чем проникновение путем слияния с плазматической мембраной. Многие вирионы видны в эндосомах, но капсиды в цитоплазме не видны после проникновения вируса, выращенного в фибробластах, и капсиды редко находят в цитоплазме клеток, инфицированных вирусом, который был выращен в эпителиальных клетках. Очевидно, вирионы задерживаются на некоторое время в эндосомах, но как только капсид освобождается от оболочки и достигает цитоплазмы, он быстро разрушается.
Как различаются вирионы человеческого цитомегаловируса, полученного в двух клеточных типах? Видимо, проявлением различий служат различия активности в отношении слияния клеток без репликации вируса. Не только epiBADrUL131 более эффективно индуцируют слияние клеток, чем fibroBADrUL131, но и низкий pH усиливает активность обоих разновидностей вируса. Без каких-либо намерений быть связанными или ограниченным любым объяснением механизма, возможно, слияние мембран требует пороговой активности слияния. Способность антитела к pUL130 нейтрализовать обе разновидности вируса указывает, что они обе зависят от комплекса gH/gL/pUL128/pUL130/pUL131 при слиянии, поэтому спланированы эксперименты, предназначенные для проверки гипотезы о том, что вирусы содержат разные количества этого комплекса. Проведены анализы нескольких его компонентов и обнаружено, что несколько большее соотношение (примерно 2-кратное) gH/gL/pUL128/pUL130/pUL131 к gH/gL/gO имелось в частицах epiBADrUL131 по сравнению с частицами fibroBADrUL131. Уровни gB, рр28 и рр65 у обоих разновидностей вируса были схожи.
Существует прецедент у ВЭБ в отношении продукции вируса с различными количествами gH-комплекса: частицы, продуцированные В-лимфоцитами, имеют дефицит gH/gL/gp42 (18). Однако могут участвовать и другие факторы. Возможно, меняется компонент комплекса, который не исследован в анализах. Или же соотношение gH-комплекса и одного или нескольких дополнительных гликопротеиновых комплексов вириона может влиять на активность слияния. Наконец, возможно, что некий неидентифицированный клеточный белок, поступающий из вирионов, когда они образуются в эпителиальных клетках или фибробластах, изменяет этот комплекс.
Имеются ли физиологические последствия двух путей проникновения? epiBADrUL131 и fibroBADrUL131 вызывают значительно различающиеся клеточные транскрипционные открыты после инфицирования клеток ARPE-19. Если предположить, что различия обусловлены вирионами или вирионами в совокупности с определенными связанными с ними клеточными факторами, эксперимент с микронаборами демонстрирует поразительные отличия транскрипционного ответа на инфекцию. Эндоцитоз тесно связан с регуляцией передачи сигнала молекулами клеточной поверхности. В итоге, возможно, что вирус влияет на передачу сигнала и клеточный транскриптом по разному, если он проникает путем слияния с плазматической мембраной или же путем эндоцитоза. Различия в передаче сигнала, вероятно, влекут за собой физиологические последствия, которые не выявляются в клеточных культурах, такие как влияние на распространение вируса, уход от иммунного ответа или вирулентность.
Литература
1. Miller. N. & Hutt-Fletcher, L.М. (1992) J Virol 66, 3409-14.
2. Nemerow. G.R. & Cooper. N.R. (1984) Virology 132, 186-98.
3. Nicola. A.V.. Hou. J.. Major, E.O. & Straus, S.E. (2005) J Virol 79, 7609-16.
4. Nicola, A.V., McEvoy, A.M. & Straus, S.E. (2003) J Virol 77, 5324-32.
5. Milne. R.S., Nicola. A.V., Whitbeck, J.C. Eiscnberg. R.J. & Cohen. G.H. (2005) J Virol 79S 6655-63.
6. Wittels. M. & Spear. P.G. (1991) Virus Res 18, 271-90.
7. Plachter. В.. Sinzger, С & Jahn, G. (f 996) Adv Virus Res 46, 195-261.
8. Compton, Т.. Nepomuceno. R.R. & Nowlin, D.M. (1992) Virology 191, 387-95.
9. Bodaghi. В., Goureau. O., Zipeto, D., Laurent, L., Virelizier. J.L. & Michelson. S. (1999) JImmunol 162, 957-64.
10. Ryckman, B.J.. Jarvis. M.A.. Drummond, D.D., Nelson. J.A. & Johnson. D.C. (2006) J Virol 80, 710-22.
11. Wang. X.. Kenyon. W.J.. Li, Q.. Mullberg, J, & Hutt-Fletcher, L.M. (1998) J Virol 72, 5552-8.
12. Li, Q., Turk. S.M. & Hutt-Flelcher, L.M. (1995) 7 Virol 69, 3987-94.
13. Hutt-Fletcher, L.M. & Lake. С.M - (2001) Curr Top Microbiol hmmmol 258, 51-64.
14. Haan. K.M. Kwok. W, W.. Longnecker. R, & Speck, P. (2000) J Virol 1A, 2451-4.
15. Haan. К.M. & Longnecker. R. (2000) Proc Nat! A cad Sci USA 97, 9252-7.
16. Li, Q.. Spriggs, M.K.. Kovats. S.. Turk, S. ML Comeau. M.R.. Nepom. B. & Hutt-Fletcher, L.M. (1997) J Virol 71, 4657-62,
17. Wang. X. & Hutt-Fletcher, L. M. (1998) J Virol 72, 158-63.
18. Borza. С M. & Hutt-Fletcher. L.M. (2002) Nat Med 8, 594-9.
19. Wang. D. & Shenk. T. (2005) ProcNatlAcadSci USA 102, 18153-8.
20. Adler. В.. Scrivano. L.. Ruzcics, Z.. Rupp, В., Sinzger. C. & Kosziiiowski. U. (2006) J Gen Virol 87, 2451-2460.
21. Wang, D. & Shenk, T. (2005) J Virol 79, 10330-8.
22. Hahn. G., Revello, M.G.. Patrone, ML. Percivalle. E.. Campanini. G.. Sarasini, A., Wagner, M., Gallina, A., Milanesi, G., Koszinowski, U., Baldanti, F. & Gerna. G. (2004) J Virol 78, 10023-33.
Настоящее изобретение не ограничивается вариантами, изложенными в описании и приведенными в качестве примеров, но оно включает изменения и модификации, находящиеся в объеме формулы изобретения, приведенной в приложении.
Реферат
Изобретение относится к области вакцинологии и клеточной биологии. Предложен способ получения вакцины против цитомегаловируса за счет выбора клеточного типа, в котором реилицируется этот вирус, вакцина и способ иммунизации особи против ЦМВ. 4 н.п. и 15 з.п. ф-лы, 6 ил, 6 табл., 1 пр.
Формула
а) штамм или изолят цитомегаловируса (ЦМВ) пассируют в культуре клеток первого выбранного клеточного типа; и
б) амплифицируют штамм или изолят ЦМВ в клетках второго выбранного клеточного типа, получая тем самым ЦМВ, кондиционированный клеточным типом, причем клетки первого и второго типа являются различными; и
в) из цитомегаловируса, кондиционированного клеточным типом, получают вакцину против цитомегаловируса.
Комментарии