Рекомбинантные вирусоподобные частицы (vlp) с использованием протеина группового антигена (gag) вируса бычьего иммунодефицита - RU2757723C2
Код документа: RU2757723C2
Чертежи






























Описание
Настоящая заявка испрашивает приоритет и преимущество предварительной патентной заявки США № 62/188,084, поданной 2 июля 2015 года, содержание которой полностью включено в настоящую заявку посредством ссылки для любых целей и передано патентовладельцу по настоящей заявке.
Предмет настоящей заявки был разработан частично с поддержкой правительства США по гранту номер AI111532-02, предоставленному Национальным институтом по изучению аллергии и инфекционных заболеваний в Национальных институтах здравоохранения и гранту номер 2011-33610-30433, предоставленному Национальным институтом продовольствия и сельского хозяйства при Министерстве сельского хозяйства Соединенных Штатов. Правительство США может иметь определенные права в отношении предмета заявки.
Область техники, к которой относится изобретение
Система или платформа на основе вакцины из вирусоподобных частиц (virus like particle, VLP) для стимулирования иммунных ответов на один или несколько различных патогенов и способы создания и использования новой системы или платформы.
Уровень техники
Ранее были предложены различные вакцинные системы или платформы. Так как данные вакцинные системы или платформы не являются оптимальными, в этой области существует потребность в усовершенствованных системах или платформах, включая системы или платформы, которые могут действовать против нескольких патогенов.
Раскрытие изобретения
В настоящей заявке описана новая VLP, состоящая из протеина группового антигена вируса бычьего иммунодефицита («Bgag»), экспрессирующая или ко-экспрессирующая один или несколько различных целевых белков-патогенов. В некоторых вариантах осуществления целевой белок-патоген представляет собой трансмембранный белок, локализованный на мембране VLP. В некоторых вариантах осуществления целевой белок-патоген выбран из одного или нескольких различных вирусных патогенов, включая ортомиксовирусы, филовирусы, коронавирусы и ретровирусы.
В некоторых вариантах осуществления, где целевыми вирусами гриппа являются вирусы гриппа A, трансмембранные белки состоят из одного или нескольких разных подтипов гемагглютинина HA 1-18 («H1» - «H18») и/или подтипов нейраминидазы NA 1-11 («N1» - «N11»). Например, в некоторых вариантах трансмембранные белки включают в себя четыре разных подтипа HA и один подтип NA. В другом примере, в некоторых других вариантах, трансмембранные белки состоят из трех разных подтипов HA и одного подтипа NA, и так далее. В качестве дополнительного примера, в некоторых вариантах четырьмя HA, ко-экспрессирующие в новых VLP Bgag, являются H5, H7, H9 и H10.
В некоторых вариантах осуществления один или несколько различных трансмембранных белков гриппа A, экспрессированных в новых VLP Bgag, могут связываться только с клетками-хозяевами или птиц, или человека, тогда как в некоторых других вариантах один или несколько различных трансмембранных белков связываются с клетками-хозяевами как птиц, так и человека.
В некоторых вариантах осуществления трансмембранные белки гриппа A, ко-эспрессированные в новых VLP Bgag, могут образовывать гомотримеры, тогда как в некоторых других вариантах трансмембранные белки образуют гетеротримеры или смесь гомотримеров и гетеротримеров.
В любой из VLP, описанной в настоящей заявке, целевой белок-патоген может быть генетически модифицирован. Например, в некоторых вариантах вариабельные участки целевого белка-патогена могут быть удалены генетически. В качестве еще одного примера, в некоторых вариантах С-конец целевого белка-патогена может быть модифицирован для увеличения эффективности связывания с Bgag. В качестве дополнительного примера в некоторых вариантах осуществления целевым белком-патогеном может быть химера. Одним из примеров такой химеры является химера, полученная генетической модификацией С-конца целевого гликопротеина Эбола с включением трансмембранного и/или С-концевого участка HA гриппа. Bgag также может быть генетически модифицирован. Например, в некоторых вариантах осуществления Bgag модифицирован для увеличения эффективности связывания с целевым белком-патогеном.
В некоторых вариантах осуществления новые VLP Bgag могут защищать или вызывать иммунный ответ на один или несколько различных патогенов у пациента. В некоторых вариантах осуществления новые VLP Bgag могут защищать или вызывать иммунный ответ у пациента на один или несколько различных типов и/или подтипов вирусов, включая различные типы и подтипы гриппа. В некоторых вариантах осуществления новые VLP Bgag могут защищать или вызывать иммунный ответ у пациента на подтипы гриппа, содержащие H5, H7, H9 и/или H10.
В некоторых вариантах осуществления новые VLP Bgag имеют диаметр более приблизительно 100 нм, предпочтительно от приблизительно 150 нм до приблизительно 200 нм, например, приблизительно 155 нм, приблизительно 160 нм, приблизительно 165 нм, приблизительно 170 нм, приблизительно 175 нм, приблизительно 180 нм, приблизительно 185 нм, приблизительно 190 нм, приблизительно 195 нм, их комбинации и тому подобное.
В некоторых вариантах осуществления каждая VLP Bgag содержит в среднем более 375 шипов белка на мембране VLP, предпочтительно от приблизительно 375 до приблизительно 800 шипов и, более предпочтительно, приблизительно 800 шипов. В соответствии с другими вариантами осуществления количество шипов может составлять приблизительно 400, приблизительно 450, приблизительно 500, приблизительно 550, приблизительно 600, приблизительно 650, приблизительно 700, приблизительно 750, их комбинации и тому подобное.
В некоторых вариантах осуществления каждая VLP Bgag имеет большую площадь поверхности, предпочтительно больше, чем у патогена, из которого произведен один из целевых белков-патогенов.
Также описана новая вакцина, содержащая VLP Bgag, которая может защищать или вызывать иммунный ответ у пациента на один или несколько различных патогенов. В некоторых вариантах осуществления новая вакцина, содержащая VLP Bgag, может защищать или вызывать иммунный ответ у пациента на один или несколько различных типов и/или подтипов вирусов, включая различные подтипы вируса гриппа. В некоторых вариантах осуществления новая вакцина, содержащая VLP Bgag, может защищать или вызывать иммунный ответ у пациента на подтипы гриппа, содержащие H5, H7, H9 и/или H10.
Также описан новый ДНК-вектор для создания новых VLP Bgag. В некоторых вариантах осуществления ДНК-вектор экспрессирован в вирусе-носителе для образования новых VLP Bgag, и предпочтительно, чтобы вирусом-носителем был рекомбинантный бакуловирус («rBV»). В некоторых вариантах осуществления rBV, содержащий ДНК-вектор, экспрессирован в эукариотических клетках, предпочтительно клетках Spodoptera frugiperda («Sf9»). В некоторых вариантах осуществления каждый ген целевого белка-патогена контролируется индивидуальным промотором и, предпочтительно, отдельным промотором гена полиэдрина.
Также описан способ получения новых VLP Bgag или вакцин с использованием ДНК-вектора, содержащего ген Bgag и по меньшей мере один ген из одного или нескольких различных целевых патогенов, при этом все они организованы в тандеме, и каждый контролируется промотором, предпочтительно промотором гена полиэдрина; вирусом-носителем предпочтительно является rBV; а эукариотной клеткой предпочтительно является Sf9.
Также описан способ различения пациентов, вакцинированных VLP Bgag, и невакцинированных пациентов.
краткое описание чертежей
На Фиг. 1 показаны схематические диаграммы конструкций VLP Bgag и конструкций M1-VLP, экспрессирующих гены гриппа.
На Фиг. 2 показан анализ гемагглютинации VLP Bgag или M1-VLP, полученных из конструкций, показанных на Фиг. 1, с использованием эритроцитов индейки.
На Фиг. 3 показан вестерн-блот VLP гриппа, созданный из конструкций, показанных на Фиг. 1, окрашенный специфическим моноклональным антителом H1.
На Фиг. 4 показан профиль белка VLP гриппа, созданный на основе конструкций, показанных на Фиг. 1, с использованием геля для электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE), окрашенного кумасси голубым.
На Фиг. 5 показана ферментативная активность NA в VLP гриппа, созданных из конструкций, показанных на Фиг. 1, измеренная в БОЕ.
На Фиг. 6 показана электронная микрофотография VLP Bgag, на которой показаны белки гриппа NA и H1.
На Фиг. 7 показана электронная микрофотография частиц VLP М1, на которой показаны белки гриппа NA и H1.
На Фиг. 8 показана схематическая диаграмма VLP Bgag, экспрессирующей гены гриппа NA и H10.
На Фиг. 9 показан анализ гемагглютинации VLP Bgag H10 гриппа, полученных из конструкций, показанных на Фиг. 8.
На Фиг. 10 показан вестерн-блот VLP Bgag вируса гриппа H10, созданных из конструкции, показанной на Фиг. 8, и четырехподтипной VLP Bgag H5/7/9/10, созданной из конструкции, показанной на Фиг. 13, окрашенный антителами к H10N7, H5N1, H7N9 и H9N2.
На Фиг. 11 показан гель SDS-PAGE с VLP гриппа Bgag H10, созданной из конструкции, показанной на Фиг. 8, и четырехподтипной VLP Bgag H5/7/9/10, созданной из конструкции, показанной на Фиг. 13.
На Фиг. 12 показана электронная микрофотография VLP Bgag H10, созданных из конструкции, показанной на Фиг. 8.
На Фиг. 13 показана схематическая диаграмма VLP Bgag, экспрессирующей гены гриппа НА, H9, H5, H7 и H10.
На Фиг. 14 показан анализ гемагглютинации четырехподтипной VLP Bgag H5/7/9/10, полученных из конструкции, показанной на Фиг. 13.
На Фиг. 15 показана ферментативная активность NA в частицах VLP гриппа Bgag H10, созданных из конструкции, показанной на Фиг. 8, и четырехподтипных VLP Bgag H5/7/9/10, созданных из конструкции, показанной на Фиг. 13, измеренная в БОЕ.
На Фиг. 16 показана электронная микрофотография четырехподтипных VLP Bgag H5/7/9/10, полученных из конструкции, показанной на Фиг. 13.
На Фиг. 17 показана исходная белковая последовательность гликопротеина (GP) Эболы, производная из штамма Mayinga («EboMay GP»).
На Фиг. 18 показана белковая последовательность химерного EboMay GP с трансмембранным доменом HA гриппа и C-концом («EboMay GP-TMCT»). Последовательность из гриппа НА подчеркнута.
На Фиг. 19 показан вестерн-блот EcoMay-GP-TMCT IECC Fxn1, дополнительно очищенного на градиенте сахарозы путем ультрацентрифугирования и окрашенного с использованием антисыворотки к Эболе.
На Фиг. 20 показан белковый профиль пиковых фракций градиента сахарозы IECC Fxn 1 и IECC Fxn 3 на геле SDS-PAGE, окрашенном кумасси голубым.
На Фиг. 21 показана электронная микрофотография VLP Bgag EboMay GP-TMCT.
На Фиг. 22 показан анализ гемагглютинации со сравнением стабильности четырехподтипного Bgag от пассажа 1 («Р1») до пассажа 5 («Р5»).
На Фиг. 23 показана ко-локализация подтипов HA в четырехподтипной VLP Bgag H5/7/9/10 с использованием вестерн-блота и SDS-PAGE.
На Фиг. 24 показан профиль распределения подтипов HA в четырехподтипной VLP Bgag H5/7/9/10 с использованием денситометрии SDS-PAGE и полуколичественного вестерн-блота.
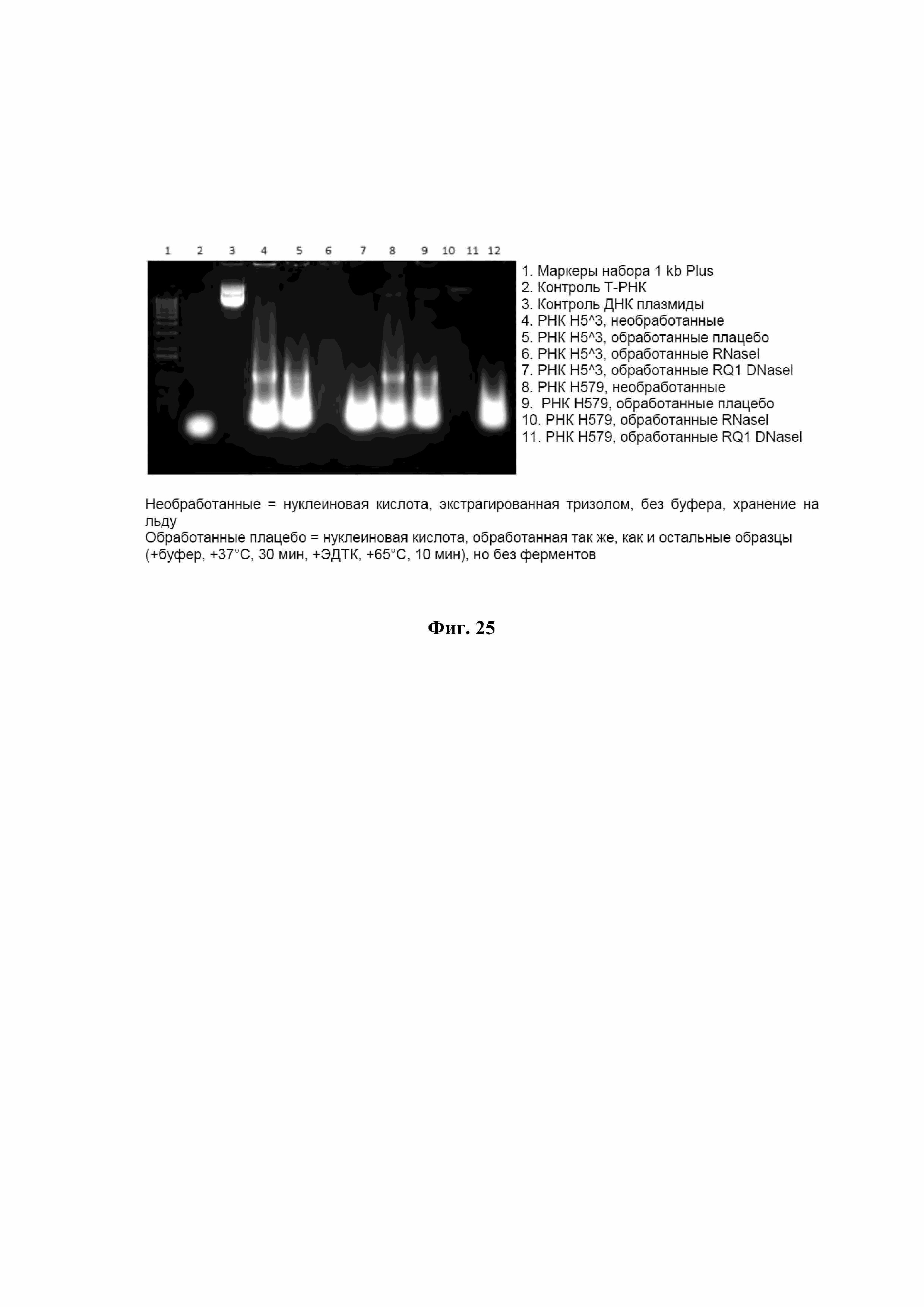
На Фиг. 25 показано присутствие РНК в VLP Bgag с использованием электрофореза в агарозном геле.
На Фиг. 26 показано присутствие РНК в VLP Bgag с использованием электрофореза в агарозном геле при более высоком разрешении, с указанием разделения нуклеиновых кислот.
На Фиг. 27 показан профиль ДНК и РНК VLP Bgag в различных условиях испытаний.
На Фиг. 28 показаны получение и характеристика VLP Bgag одинарного подтипа H10 в панели A и четырехподтипной VLP Bgag H5/7/9/10 в панели B.
На Фиг. 29 показаны косвенные иммунофлуоресцентные анализы клеток Sf9, инфицированных рекомбинантным бакуловирусом (rBV), экспрессирующим гены H5, H7, H9, H10, NA и Bgag.
На Фиг. 30 показан профиль иммуногенности хорьков, иммунизированных с помощью VLP Bgag одинарного подтипа H10 или четырехподтипной VLP Bgag H5/7/9/10.
На Фиг. 31 обобщен титр антител к ингибированию гемагглютинации (ИГ) HA у хорьков в сыворотках хорьков после иммунизации, перед проверочным заражением.
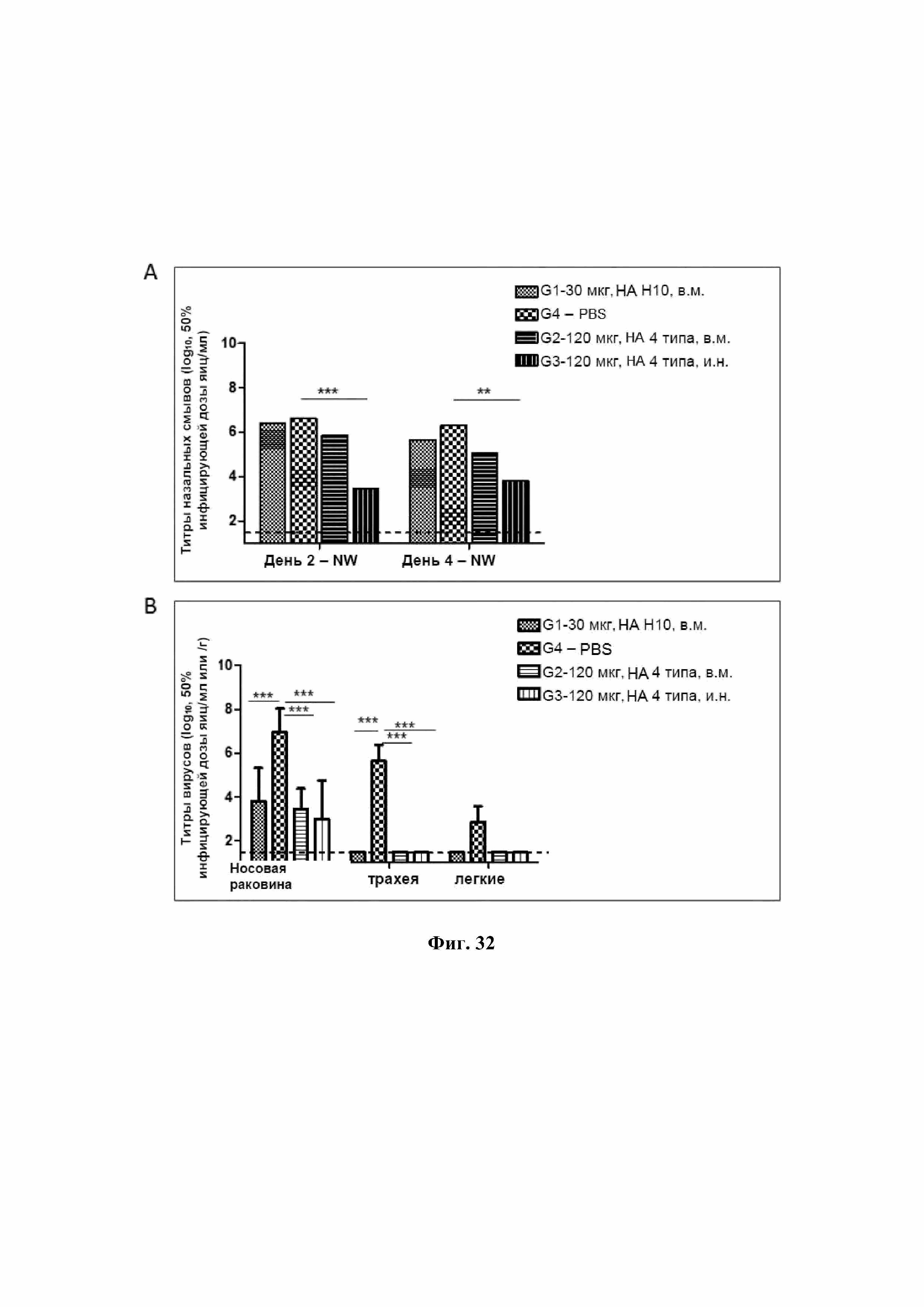
На Фиг. 32 обобщены титры реплицирующего вируса у иммунизированных хорьков в день 2 или день 4 после экспериментального заражения.
На Фиг. 33 показано получение и характеристика VLP Bgag H555 с тремя кладами, содержащей белки HA H5 из трех клад вирусов HPAI H5N1.
На Фиг. 34 показана характеристика очищенных VLP с Bgag H555 с тремя кладами.
На Фиг. 35 показана реакция антител птиц, вакцинированных VLP Bgag H555 против клады 2.3.4.4 GyrF H5N8 (группа 1), клады 2.1.3 Ck/Египет H5N1 (группа 2) и клады 2.2.1 Ck/WJ (группа 3).
На Фиг. 36 показаны графики выживания Каплана-Мейера для защиты цыплят, вакцинированных VLP H555, после экспериментального заражения HPAI H5N1 или H5N8.
На Фиг. 37 показаны индивидуальные титры ингибирования гемагглютинации (log2) и стандартная ошибка в течение 5 недель после вакцинации для предварительно экспериментально зараженных групп птиц против вируса, используемого для экспериментального заражения.
На Фиг. 38 показаны индивидуальные титры ингибирования гемагглютинации (log2) и стандартная ошибка в течение 2 недель после экспериментального заражения групп птиц, вакцинированных к вирусу, используемому для экспериментального заражения.
На Фиг. 39 показаны титры вируса из мазков со слизистой оболочки рта и из клоаки на 2 и 4 день после экспериментального заражения вирусом A/кречет/Вашингтон/2014 H5N8 (клада 2.3.4.4).
На Фиг. 40 показаны титры вируса из оральных и клоакальных мазков на 2 и 4 день после экспериментального заражения вирусом A/курица/Египет/2010 H5N1 (клада 2.2.1).
На Фиг. 41 показаны титры вируса из оральных и клоакальных мазков на 2 и 4 день после экспериментального заражения вирусом A/курица/Западнояванский Субанг/2007 H5N1 (клада 2.1.3).
Осуществление изобретения
Если не указано иное, в настоящем описании используются обычные химические, биохимические, иммунологические, молекулярно биологические, иммунологические и фармакологических методики и термины, которые имеют свое обычное значение для специалистов в этой области (если иное не определено/не описано в настоящем документе). Все публикации, ссылки, патенты и патентные заявки, упомянутые в настоящей заявке, настоящим полностью включены посредством ссылки.
При использовании в настоящем описании и прилагаемой формуле изобретения, применяются следующие общие правила. Формы в единственном числе включают в себя формы во множественном числе, если по контексту прямо не требуется иное. Также применяются общие правила обозначений генов и белков. Гены выделены курсивом или подчеркнуты (например, Bgag или Bgag), но белки указаны стандартным шрифтом, а не курсивом или подчеркнутым шрифтом (например, Bgag). Также применяются общие правила обозначений классификации организмов. Названия отрядов, семейств, родов и видов выделены курсивом.
При использовании в настоящем описании следующие термины имеют указанное значение. Термин «приблизительно» имеет обычное и простое значение «примерно» в том смысле, как его понял бы специалист в данной области. Термин «состоять из», «состоящий из», «содержать», «содержащий», «включать», «включая», «включать, помимо прочего», или «характеризуется», является включительным или открытым и не исключает дополнительных, неуказанных элементов.
При использовании в настоящей заявке следующие термины имеют указанное значение.
«СПИД» означает синдром приобретенного иммунодефицита.
«Bgag» означает протеин группового антигена (gag) вируса бычьего иммунодефицита.
«VLP Bgag» означает VLP на основе Bgag, т.е. вирусоподобную частицу с протеином Bgag во внутреннем ядре.
«БВИ» означает бычий вирус иммунодефицита.
«CDC» означает Центр по контролю и профилактике заболеваний.
«DIVA» означает дифференциацию инфицированных от вакцинированных животных.
«EboMay GP» означает гликопротеин Эбола, присутствующий в штамме Mayinga, пример которого приведен на Фиг. 17.
«EboMay GP-TMCT» означает химерный EboMay GP с трансмембранным доменом HA и C-концом гриппа, пример которого приведен на Фиг. 18.
«ENV» означает белок оболочки ВИЧ.
«Fxns» означает фракционирования.
«H1» - «H18» означает типы HA гриппа 1-18.
«HA» означает гемагглютинин трансмембранного гликопротеина гриппа.
«ВИЧ» означает вирус иммунодефицита человека.
Вирус «HK09» означает вирус H9N2 гриппа A/ Гонконг/ 33982/2009.
«HPAI» означает высокопатогенный птичий грипп.
«IECC Fxn 1» означает фракцию 1 VLP EboMay GP-TMC из пиковой фракции, полученной на колонке ионообменной хроматографии.
«IECC Fxn 3» означает фракцию 3 VLP EboMay GP-TMC из пиковой фракции, полученной на колонке ионообменной хроматографии.
«и.н.» означает интраназально, как указано в отношении способа введения вакцин.
«в.м.» означает внутримышечно, как указано в отношении способа введения вакцин.
Вирус «IN/5» означает вирус H5N1 гриппа A/Индонезия/5/2005.
Вирус «JX/13» означает вирус H10N8 гриппа A/Цзянси/IPB13a/2013.
«М1» означает матрицу 1 белка внутреннего ядра гриппа.
«VLP M1» означает VLP на основе M1, т.е. вирусоподобную частицу с белком M1 во внутреннем ядре.
«MERS» означает ближневосточный респираторный синдром.
«МЗ» означает множественность заражения.
«N1» - «N11» означает типы 1-11 гриппа NA.
«NA» означает трансмембранный гликопротеин гриппа нейраминидазу.
«p.c.» означает после заражения, как указано в отношении периода после экспериментального заражения пациента целевым патогеном (патогенами).
«PBS» означает физиологический раствор с фосфатным буфером.
«PEDV» означает вирус эпизоотической диареи свиней.
Вирус «PR8» означает вирус H1N1 гриппа A/Пуэрто-Рико/8/1934.
«rBV» означает рекомбинантный бакуловирус.
«RFU» означает относительные единицы флуоресценции.
«SARS» означает тяжелый острый респираторный синдром.
«Rasmussen» означает клетку Spodoptera frugiperda.
Вирус «SH/13» означает вирус H7N9 гриппа A/ Шанхай/2/2013.
«Пациент» по настоящему изобретению предпочтительно является животным, например, мышью, хорьком, курицей, свиньей и т.д., и предпочтительно является млекопитающим или птицей, а наиболее предпочтительно - человеком.
«Целевой белок-патоген» включает в себя любой белок или пептид любого патогена. Патогеном может быть вирус, бактерия, прион, эукариот, грибок или любой иной микроорганизм. Патогеном также может быть паразит, например, паразитический простейший. Патогеном также может быть многоклеточный организм, включая человека. Примером целевого белка-патогена человека является белковый или пептидный маркер рака.
Вирус «VN/04» означает вирус H5N1 гриппа A/Вьетнам/1203/2004.
«VLP» означает частицу, подобную рекомбинантному вирусу.
«ВОЗ» означает Всемирную организацию здравоохранения;
Вакцины против патогенов
Вакцинация является наиболее эффективной стратегией профилактики или минимизации/лечения эпидемий, вызываемых патогенами. Например, эпидемия сезонного гриппа серьезно поражает 5 (пять) миллионов человек по всему миру. Относительно недавно вспышки Эбола в Западной Африке искалечили десятки тысяч людей на континенте.
В отличие от лекарственной терапии, в рамках которой осуществляют лечение пациентов, инфицированных патогеном, вакцины защищают пациентов от будущих инфекций посредством стимулирования устойчивого иммунного ответа у пациента. Традиционные разработки вакцин были узкими и специфическими, сосредотачиваясь на выявлении иммунных реакций против одного патогена. Тем не менее, наблюдается растущая потребность в разработке поливалентных вакцин широкого спектра действия, способных вызывать устойчивые иммунные ответы против нескольких патогенов. В настоящем документе изобретателями впервые описана новая вакцинная система или платформа, предлагающая иммунную защиту от одного или нескольких различных патогенов.
Вирусы гриппа являются распространенным вирусным патогеном, который может быть смертелен для людей и является серьезной угрозой для общественного здравоохранения (Morens and Fauci, 2012; Palese, 2006; Yen and Webster, 2009). Вирус гриппа представляет собой оболочечный вирус, содержащий восемь сегментированных двухнитевых отрицательно-полярных геномов РНК и относится к семейству Orthomyxovirideae. Существуют вирусы гриппа типа A, B и C, из них Всемирная организация здравоохранения («ВОЗ») и Центр по контролю и профилактике заболеваний («CDC») пристально отслеживают потенциальные эпидемические и пандемические вспышки вируса гриппа A и, в частности, птичьего гриппа A.
В вирусе гриппа A, помимо прочего, присутствуют два трансмембранных гликопротеина: гемагглютинин («HA») и нейраминидаза («NA»), а также белок («M1») белковой матрицы 1 внутреннего ядра. У вируса гриппа А известно 18 типов HA и 11 типов NA, в результате чего существует в общей сложности 198 различных комбинаций подтипов гриппа А. Считается, что большинство этих подтипов гриппа А могут инфицировать птиц, при этом только некоторые могут инфицировать человека. Так как подтипы птичьего гриппа способны к рекомбинации и частым генетическим изменениям (Morens and Fauci, 2012; Palese, 2004), известно, что несколько подтипов вируса птичьего гриппа A начинали пересекать барьер между человеком и птицами.
Пересечение барьера между человеком и птицами вызвало озабоченность по всему миру в связи с тем, что высокопатогенные вирусы птичьего гриппа («HPAI») вызовут пандемию (Kang et al., 2009; Morens and Fauci, 2012), во многом аналогичную пандемии H1N1 в 1918 году, которая унесла от 40 до 100 миллионов жизней по всему миру. В настоящее время примером вируса HPAI является грипп H5N1, тем не менее существуют и другие. Они включают в себя, например, вирус гриппа H7N9, вызвавший вспышку заболевания в 2013 года и унесший жизни многих людей (Chen et al., 2015; Gao et al., 2013), вирус гриппа H9N2 с птичьим происхождением, но который, как известно, является патогеном человека (Blanco et al., 2013; Pushko et al., 2005; Yen and Webster, 2009), и вирус гриппа H10N8, который, как было недавно обнаружено, приобрел способность инфицировать человека (Garcia-Sastre and Schmolke, 2014; To et al., 2014).
В настоящее время существуют вакцины против сезонного гриппа. Однако они состоят из вирусов H1N1, H3N2 и гриппа B, отдельно выращиваемых в оплодотворенных куриных яйцах и затем смешиваемых в форме трехвалентной вакцины. Производство вакцины с использованием яиц имеет существенные ограничения. Это необеспечение защиты широкого спектра действия; неэффективность против новых вирусов HPAI; неэффективное производство; низкий выход; и проблемы с увеличением производства. Так как многие патогены, например, грипп, характеризуются ежегодным антигенным дрейфом, необходимо постоянно дорабатывать и изменять вакцину в ответ на изменения циркулирующих вирусов сезонных заболеваний. Кроме того, так как каждую из трех одновалентных вакцин против гриппа необходимо готовить отдельно, производство смешанной трехвалентной вакцины значительно увеличивает стоимость производства вакцины, увеличивает время реакции на угрозу гриппа и увеличивает вероятность побочных реакций после введения. Соответственно, существует настоятельная необходимость в разработке новых вакцинных систем широкого спектра действия.
Вирус Эбола является патогеном 4-й группы риска, характеризуется чрезвычайно высоким уровнем смертности и за последние годы уничтожил население многих западноафриканских стран. Вирус Эбола представляет собой оболочечный вирус, содержащий однонитевую отрицательно-полярную РНК, и относится к семейству Filoviridae. Вирус Эбола содержит трансмембранный гликопротеин («GP»), а также белки внутреннего ядра. В настоящее время нет вакцин против Эбола, одобренных для общего применения у людей.
Коронавирус Ближневосточного респираторного синдрома («MERS») представляет собой новый вирус, выявленный в 2012 году, и, как и коронавирус тяжелого острого респираторного синдрома (SARS), он относится к семейству Coronaviridae. Коронавирус MERS представляет собой оболочечный вирус, содержащий однонитевую положительно-полярную РНК. Также, аналогично другим коронавирусам, например, вирусу эпизоотической диареи свиней («PEDV»), коронавирус MERS содержит трансмембранные гликопротеины. В настоящее время о коронавирусе MERS известно очень мало.
Как широко известно, вирус иммунодефицита человека («ВИЧ») вызывает синдром приобретенного иммунодефицита («СПИД»). ВИЧ представляет собой оболочечный вирус, содержащий однонитевую положительно-полярную РНК, которая относится к семейству Retroviridae. Целью в исследованиях вакцин стал белок оболочки ВИЧ («ENV»). Тем не менее, в настоящее время нет вакцин против ВИЧ, одобренных для клинического использования у человека.
Список патогенов, опасных для животных и людей, длинен и продолжает увеличиваться. Наблюдается насущная потребность в разработке устойчивой вакцинной системы/платформы широкого спектра действия, способной вызывать устойчивые иммунные ответы против нескольких патогенов. В настоящей заявке изобретателями впервые описана новая вакцинная платформа, в которой используется новая VLP на основе Bgag для вызова специфического иммунного ответа или иммунного ответа широкого спектра действия против одного или нескольких (например, множества) различных патогенов.
Использование VLP в качестве кандидатной вакцины
Рекомбинантные вирусоподобные частицы («VLP»), являются перспективными кандидатными вакцинами. Они высоко иммуногенны, морфологически и антигенно подобны нативным вирусным частицам, но неспособны к репликации. Иммунозащитное действие VLP было показано в доклинических и клинических испытаниях (Pushko et al., 2011). Кроме того, в отличие от живых или инактивированных вакцин, VLP не подразумевают производство целевого патогена, но могут вызывать устойчивые иммунные ответы против целевого патогена при помощи антигенов целевого патогена, экспрессированных и присутствующих в VLP. Соответственно, VLP являются безопасными и эффективными кандидатными вакцинами для многих патогенов, включая грипп (Kushnir et al., 2012; Pushko et al., 2013).
В последнее время было показано, что VLP являются перспективными вакцинами против птичьего гриппа (Bright et al., 2007; Galarza et al., 2005; Kang et al., 2009; Perrone et al., 2009; Pushko et al., 2005; Quan et al., Ross et al., 2009). Нами было показано, например, что VLP, состоящие из белков гриппа HA, NA и M1 (Pushko et al., 2005), вызвали высокоэффективные защитные иммунные ответы, которые в некоторых случаях превосходили иммунные ответы, вызванные традиционными противогриппозными вакцинами (Bright et al., 2007; Pushko et al., 2007). Наблюдаемая высокая иммуногенность противогриппозных вакцин VLP была отнесена на счет организации белка HA в виде регулярных структур, напоминающих структуры вируса гриппа, что способствует активации иммунной системы хозяина (Kang et al., 2009; Pushko et al., 2013).
В отличие от технологии на основе яиц, VLP производят в клеточной культуре и конструируют с использованием способов молекулярной биологии. Тем не менее, даже в настоящее время многие кандидатные вакцины на основе VLP, например, производимые на основе яиц трехвалентные вакцины против сезонного гриппа, по-прежнему разрабатываются как VLP, специфичные к определенному штамму, которые синтезируют и производят индивидуально и затем смешивают или соединяют. Кроме того, многие из этих VLP были получены с использованием гомологичных белков HA, NA и M1; т.е., HA, NA и M1, полученных из одного вируса (Perrone et al., 2009; Pushko et al., 2010; Pushko et al., 2005; Pushko et al., 2007). Этот подход проблематичен, так как очень часто данные о последовательностях новых вирусов не всегда легкодоступны. Например, на ранних стадиях разработки новых противогриппозных вакцин, как правило, недоступна последовательность М1 новых вирусов гриппа. Вследствие этого особо проблематична разработка вакцин с использованием VLP с гомологичными белками HA, NA и M1 против, например, новых возникающих вирусов.
В ответ на это некоторые исследователи разработали VLP с использованием HA и NA, полученных из целевого вируса гриппа, и M1, полученного из другого вируса гриппа (Liu et al., 2015). Другие также пытались заменить M1 белком gag вируса лейкемии мышей (Haynes, 2009; Haynes et al., 2009) или белком gag вируса иммунодефицита обезьян/человека (Guo et al., 2003). Недавно нами были созданы и описаны несколько различных VLP M1 с NA и тремя различными HA, каждый из которых был получен из другого подтипа гриппа (Pushko et al., 2011; Tretyakova et al., 2013). Также посредством электронной микроскопии нами было показано, что все три подтипа HA ко-локализуются на одной VLP M1 (Pushko et al., 2011; Tretyakova et al., 2013). Кроме того, нами показано, что VLP М1 вызывает высоко защитные иммунные ответы против всех трех штаммов гриппа и является эффективной трехвалентной противогриппозной вакциной (Pushko et al., 2011; Tretyakova et al., 2013). Другими словами, вакцина VLP M1, производная из единственной VLP, может защитить от всех трех вирусов гриппа.
Тем не менее, VLP M1 не является идеальным кандидатом для вакцин и, в частности, вакцин для человека. Например, белки M1 некоторых вирусов гриппа экспрессированы на более низких уровнях. Кроме того, как рассмотрено выше, часто для новых вирусов гриппа не всегда доступна последовательность M1 или M1. Более того, большинство людей сталкиваются с M1; таким образом, существует вероятность того, что у людей уже существует иммунитет хозяина против VLP, экспрессирующей M1. Соответственно, это значительно ухудшило бы эффективность VLP M1 в плане стимулирования специфических иммунных ответов у человека.
Новая VLP Bgag, не содержащая М1
Изобретателями впервые получена и описана в настоящей заявке VLP, не содержащая М1, c использованием белка gag бычьего вируса иммунодефицита («БВИ»), обозначенного в настоящей заявке как VLP Bgag, который экспрессирует один или несколько различных целевых белков-патогенов (см. пример 1). В новой VLP, описанной в настоящей заявке, в качестве белка внутреннего ядра использован Bgag. БВИ, относящийся к роду Lentivirus семейства вирусов Retroviridae, ранее использовали в рамках исследований по разработке вакцин, и ранее было описано использование векторов БВИ (Luo, 2012). Тем не менее, до настоящего раскрытия не было описано использование VLP Bgag для экспрессии или ко-экспрессии одного или нескольких различных целевых белков-патогенов, например, различных белков гриппа; использование VLP Bgag в качестве кандидатной вакцины; или использование Bgag или Bgag в качестве диагностического инструмента.
Как описано в настоящем документе, изобретатели, среди прочего, впервые получили VLP Bgag, экспрессирующие и представляющие один или несколько различных целевых белков-патогенов (см. пример 1 и примеры 5-12); впервые показано, что VLP Bgag может экспрессировать и представлять один или несколько различных функциональных белков гриппа, в том числе одновременно ко-экспрессировать и со-представлять функциональные HA, относящиеся к четырем различным подтипам гриппа (см. примеры 5-7 и примеры 11-12); впервые показано, что VLP Bgag может экспрессировать и представлять негриппозные белки-патогены (см. пример 9); и впервые показано, что VLP Bgag может экспрессировать и представлять генетически модифицированные химерные белки-патогены (см. пример 10). Кроме того, изобретатели впервые описали способ получения новой специфической (одновалентной) вакцины или вакцины широкого спектра действия (многовалентной) на основе VLP Bgag и опосредованной VLP Bgag, (см. примеры 1-3 и примеры 5-12). Дополнительно изобретатели описали преимущества, связанные с использованием VLP Bgag, включая ее преимущества в качестве улучшенной вакцины. Кроме того, дополнительно изобретатели описали преимущества использования VLP Bgag, включая использование в качестве кандидатной вакцины широкого спектра действия. Кроме того, авторы изобретения описали несколько диагностических способов с использованием белка Bgag в связи с системой на основе VLP Bgag.
VLP Bgag могут быть использованы в отношении широкого спектра целевого белка-патогена
Типы целевых белков-патогенов, которые могут быть экспрессированы и представлены в платформе на основе VLP Bgag, очень разнообразны. Например, такой белок-патоген может включать в себя любой пептид или белок из любого вируса, бактерии, приона, эукариота, гриба, паразита или любого иного одноклеточного или многоклеточного организма. Более того, при введении пациенту VLP Bgag, экспрессирующей один или несколько из этих целевых белков-патогенов, VLP Bgag может вызывать иммунный ответ против одного или нескольких из этих патогенов. Соответственно, систему или платформу на основе VLP Bgag можно использовать, в частности, как противовирусную вакцину, противобактериальную вакцину, противоприонную вакцину, противогрибковую вакцину и даже противопаразитную вакцину для лечения, ингибирования и/или профилактики патогенного воздействия этих патогенов. При использовании принципов и способов, описанных в настоящем изобретении, также можно использовать систему или платформу на основе VLP Bgag в качестве вакцины против одного или нескольких различных вирусов, бактерий, прионов, эукариот, грибов, паразитов, любого другого микро- или макроорганизма, их комбинаций и тому подобного; их преимущество состоит в том, что в зависимости от уязвимости или чувствительности пациента может быть предложена индивидуально подобранная вакцина или терапия. Например, пациенту, чувствительному к двум определенным вирусам, одной определенной бактерии и одному определенному грибу, можно вводить индивидуально подобранную вакцину для ингибирования/профилактики и/или лечения проявления действия вирусов, бактерий и грибков.
Новая платформа VLP Bgag также может быть использована для экспрессирования одного или нескольких маркеров рака. Например, в качестве целевого белка-патогена могут быть использованы один или несколько пептидов или белков - маркеров рака. В качестве вакцины VLP Bgag можно использовать для усиления иммунной системы субъекта и улучшения наблюдения за развитием раковых клеток для эффективного ингибирования/профилактики, лечения или борьбы с раком.
VLP Bgag, содержащая целевые белки-патогены, может вызывать иммунный ответ и может быть использована в качестве вакцины. Например, VLP Bgag, содержащие целевые белки-патогены, могут быть введены инъекционно или иным образом пациенту с целью вызова иммунного ответа против целевых белков-патогенов. Затем с помощью стандартного анализа определяют иммунный ответ вакцинированного пациента на целевой патоген. Например, в течение от приблизительно 2 до приблизительно 4 недель производят забор крови вакцинированного пациента и антитело против целевого патогена определяют с помощью иммуносорбентного ферментного анализа, иммунофлуоресцентного анализа антител или других анализов на выявление антител и сравнивают с профилем антител невакцинированных пациентов. Присутствие антитела к целевому патогену у вакцинированного пациента означает, что VLP Bgag вызывает иммунный ответ у пациента и, следовательно, является иммуногенным в живом организме. Стандартные исследования экспериментальных заражений также могут показать эффективность VLP Bgag в плане защиты пациента от целевого патогена.
VLP Bgag с консенсусным целевым белком-патогеном или консервативными эпитопами
В некоторых вариантах осуществления целевой белок-патоген VLP Bgag может быть разработан таким образом, чтобы расширять иммунную защиту. Например, целевой белок-патоген может быть получен из консенсусной последовательности HA, определенной с помощью передовых методов генетического анализа (Denis et al., 2008; Ebrahimi et al.; Pica and Palese, 2013; Rao et al., 2010; Schotsaert et al., 2009; Wang and Palese, 2009; Wei et al., 2010). В качестве еще одного примера, целевой белок-патоген может быть получен из консервативных эпитопов гриппа, например, внеклеточной части белка с ионным каналом М2 гриппа, определенной передовыми методами генетического анализа (Denis et al., 2008; Ebrahimi et al.; Pica and Palese, 2013; Rao et al., 2010; Schotsaert et al., 2009; Wang and Palese, 2009; Wei et al., 2010). VLP Bgag гриппа, полученная в результате этих дизайнов, может обеспечить иммунную защиту широкого спектра действия для множества или даже всех подтипов гриппа.
VLP Bgag с генетически модифицированными целевыми белками-патогенами
Изобретателями показано, что генетически модифицированный целевой белок-патоген может быть экспрессирован и представлен в VLP Bgag (см. пример 8 и пример 10). Например, в некоторых вариантах осуществления часть целевого белка-патогена экспрессирована и представлена в VLP Bgag. В некоторых вариантах осуществления в некоторых целевых белках-патогенах могут быть удалены вариабельные участки. Например, VLP Bgag может быть создан с использованием «безголового» HA, в котором удалены самые вариабельные эпитопы HA, ответственные за нейтрализацию вируса. Преимущество такого подхода состоит в том, что «безголовый» HA, представленный в VLP Bgag, может вызывать иммунный ответ на эпитопы HA, которые обычно скрыты в стандартной VLP, содержащей полноразмерный HA. В качестве еще одного примера, VLP Bgag может быть создана с использованием «стеблевой» области HA. Иммунный ответ на безголовый HA или стеблевой HA может привести к созданию вакцины с широкой защитой или универсальной вакцины против гриппа, способной защищать от нескольких штаммов и подтипов вируса гриппа. В этих вариантах VLP Bgag может представлять в основном консервативные области целевого белка-патогена в целях стимулирования иммунного ответа широкого спектра.
Трансмембранные белки из целевых патогенов могут служить прекрасными целевыми белками-патогенами и могут быть экспрессированы и представлены в VLP Bgag. Тем не менее, цитоплазматический белок и пептид, а также белок и пептид, выделенный из целевых патогенов, также могут служить в качестве целевых белков-патогенов при применении генной инженерии. Например, цитоплазматический и/или секретируемый белок или пептиды вируса, бактерии, приона, эукариота, гриба, паразита, паразитарного простейшего или белок, или пептиды человеческого происхождения (например, маркеры рака) могут быть сконструированы в трансмембранный белок, который может быть представлен в VLP Bgag. Одним таким примером является генетическое конструирование трансмембраного домена HA гриппа на таких не-трансмембранных белках и/или пептидах с помощью стандартных методов или способов генетической инженерии, по существу описанных в настоящем документе, для создания химерного EboMay GP-TMCT. Например, трансмембранный домен HA гриппа может быть добавлен к C-концу простат-специфического антигена, белка-маркера рака, который обычно секретируется клетками предстательной железы. Используя принципы и способы, по существу описанные в настоящей заявке, генетически конструированный белок простат-специфического антигена может быть экспрессирован и представлен в VLP Bgag и использован у пациентов для ингибирования, профилактики, лечения, отслеживания и/или контроля прогрессирования рака.
В некоторых вариантах осуществления генетическая модификация включает в себя модификацию целевого белка-патогена для увеличения эффективности связывания с Bgag (см. пример 10). Например, модификации могут быть выполнены на С-конце целевого белка-патогена для оптимизации эффективности связывания с Bgag.
В некоторых других вариантах осуществления генетическая модификация подразумевает создание химерного целевого белка-патогена. Одной такой химерой является белок EboMay GP-TMCT (см. Фиг. 17 и 18). Как описано в примере 10 и в иных местах настоящего описания, химерный белок GP-TMCT EboMay создают путем замены С-конца целевого гликопротеина Эбола на трансмембранную и/или С-концевую область HA гриппа.
В некоторых других вариантах осуществления целевой белок-патоген может быть рационально сконструирован и/или переконструирован для улучшения экспрессии и представления в VLP Bgag. Например, было показано, что рациональное конструирование белка оболочки ВИЧ может увеличить представление белка в 10 раз (Wang et al., 2007).
Во всех случаях, белок Bgag также можно генетически модифицировать и улучшить, используя те же принципы, по существу описанные выше. Например, определенные области Bgag могут быть модифицированы для увеличения эффективности связывания с целевым белком-патогеном.
Аналогично другим VLP Bgag, VLP Bgag, содержащий генетически сконструированные целевые белки-патогены, может вызывать иммунный ответ и служить в качестве вакцины, например, с использованием принципов и способов, описанных в настоящем документе.
Новая VLP Bgag может экспрессировать и представлять один или несколько различных типов целевых белков-патогенов
Изобретателями создана и описана новая система на основе VLP Bgag для экспрессии/ко-экспрессии и представления/сопредставления одного или нескольких различных типов целевых белков-патогенов (см. примеры 5-12). В некоторых вариантах осуществления целевой белок-патоген локализован на мембране VLP (см. Фиг. 6, 7, 12, 16). Различные белки-патогены, независимо от того, являются ли они генетически модифицированными или нет, могут быть использованы в настоящей новой системе на основе VLP Bgag. Два примера включают в себя трансмембранные белки гриппа А (см. примеры 5-8 и примеры 11-12) и гликопротеин Эбола (см. примеры 9-10).
Изобретателями также было показано, что целевые белки-патогены могут быть представлены VLP Bgag, а также VLP M1. Например, изобретателями было показано, что белки HA и NA гриппа представлены в VLP Bgag, также, как и в VLP M1 (см. Фиг. 2-7). Изобретателями также было показано, что представленные целевые белки-патогены являются функциональными. Например, было показано, что белки HA и NA на VLP Bgag проявляют функциональную гемагглютинацию и ферментативную активность NA (см. Фиг. 2, 5, 9, 14 и 15).
В некоторых вариантах осуществления новые VLP Bgag имеют диаметр более приблизительно 100 нм, и до 200 нм, предпочтительно от приблизительно 120 нм до приблизительно 200 нм, более предпочтительно от приблизительно 150 нм до приблизительно 200 нм, более предпочтительно от приблизительно 160 нм до приблизительно 200 нм, более предпочтительно от приблизительно 170 нм до приблизительно 200 нм, более предпочтительно от приблизительно 180 нм до приблизительно 200 нм, более предпочтительно от приблизительно 190 нм до приблизительно 200 нм. VLP Bgag значительно крупнее, чем некоторые естественные патогены. Например, средний диаметр вируса гриппа составляет приблизительно 100 нм; средний диаметр ВИЧ составляет около 120 нм; и средний диаметр коронавируса SARS составляет около 80-90 нм. VLP Bgag также значительно больше, чем VLP M1. Например, средний диаметр VLP Bgag гриппа составляет от приблизительно 150 до приблизительно 180 нм, при этом средний диаметр VLP M1 гриппа составляет от приблизительно 120 нм до приблизительно 150 нм. Несмотря на более крупный размер, VLP Bgag имеют общую морфологию, аналогичную VLP M1, а также вирусу гриппа (см. Фиг. 6 и 7). Более крупная VLP Bgag имеет большие преимущества, в частности, как кандидатная вакцина, так как каждая VLP может представлять существенно больше целевых белков-патогенов, чем естественный патоген или VLP M1.
Новая VLP Bgag является универсальной системой, которая может представлять один или несколько различных типов целевых белков-патогенов (см. примеры 5-12). В некоторых вариантах осуществления VLP Bgag содержит целевой белок-патоген, выбранный из одного патогена (см. примеры 5-10), при этом в некоторых других вариантах VLP Bgag содержит несколько (например, множество) целевых белков-патогенов, выбранных из нескольких типов или подтипов патогена (см. примеры 11-12). В некоторых вариантах осуществления одним из выбранных патогенов является вирус. В некоторых вариантах осуществления флаконный патоген выбирают из комбинации ортомиксовирусов, предпочтительно вируса гриппа, более предпочтительно, вируса гриппа А; филовирусов, предпочтительно вирусов Эбола; коронавирусов, предпочтительно вирусов MERS; и ретровируса, предпочтительно ВИЧ.
В некоторых вариантах осуществления VLP Bgag могут защищать или вызывать иммунный ответ на один или несколько различных патогенов у пациента. Таким образом, вакцины, полученные с помощью одновалентной VLP Bgag, могут обеспечивать устойчивую специфическую к определенному патогену иммунную защиту, при этом вакцины, полученные с использованием многовалентных VLP Bgag, могут обеспечивать многовалентную иммунную защиту широкого спектра действия. Важно отметить, что защита широкого спектра действия обеспечивается одной многовалентной VLP Bgag, а не смешиванием различных одновалентных вакцин. Тем не менее, безусловно, для обеспечения более широкого иммунного охвата можно дополнительно смешать и/или добавить другие многовалентные VLP Bgag. Соответственно, многовалентная VLP Bgag может быть использована для обеспечения иммунной защиты широкого спектра действия против различных целевых патогенов и является важным дополнением к стратегиям подготовки к пандемии.
Новая VLP Bgag может экспрессировать и представлять один или несколько различных подтипов целевых белков-патогенов
Изобретателями было показано, что система или платформа на основе VLP Bgag является платформой для представления (размещения) одного или нескольких различных подтипов целевых белков-патогенов (см. примеры 5-12). В некоторых вариантах осуществления VLP Bgag одного подтипа изобретателями было показано, что могут быть получены VLP Bgag, содержащие функциональные трансмембранные белки HA и NA гриппа, включая PR8 H1 и нового гриппа H10 (см. пример 5 и пример 6). До настоящего изобретения Bgag не использовали для производства VLP гриппа, и было неизвестно, что Bgag может быть использован для получения VLP одного подтипа. В некоторых вариантах осуществления VLP Bgag нескольких подтипов изобретателями впервые была создана и описана VLP Bgag нескольких подтипов, т.е., VLP, которая одновременно ко-экспрессирует и сопредставляет различные подтипы целевого белка-патогена, вследствие чего она может обеспечивать одновременную иммунную защиту широкого спектра действия против нескольких различных подтипов целевого вируса (см. пример 11 и пример 12). До настоящего изобретения не было известно, что Bgag может быть использован для получения VLP нескольких подтипов.
В некоторых вариантах осуществления VLP Bgag могут защищать или вызывать иммунный ответ у пациента на один или несколько различных подтипов вирусов. Таким образом, вакцины, полученные с VLP Bgag одного подтипа, могут обеспечивать устойчивую специфическую к определенному подтипу иммунную защиту, при этом вакцины, полученные с VLP Bgag нескольких подтипов, могут обеспечивать иммунную защиту широкого спектра действия от нескольких подтипов. Аналогично многовалентным VLP Bgag, защита широкого спектра действия обеспечивается одной VLP Bgag нескольких подтипов, а не смешиванием различных вакцин от одного подтипа. Тем не менее, безусловно, для обеспечения более широкого иммунного охвата можно смешать и/или добавить различные VLP Bgag нескольких подтипов. Вместе со структурами многовалентной VLP Bgag системы на основе VLP Bgag, которые могут быть использованы для вызова иммунной защиты широкого спектра действия против различных типов и подтипов патогенов, являются важным дополнением к стратегиям готовности к пандемии.
В качестве примера, новая VLP Bgag может защищать или вызывать иммунный ответ у пациента на один или несколько (например, множество) различных подтипов вирусов, включая различные подтипы вирусов гриппа. В некоторых вариантах осуществления трансмембранные белки гриппа A, ко-экспрессированные в новых VLP Bgag, могут связываться с клетками-хозяевами только птиц или человека, при этом в некоторых других вариантах трансмембранные белки связываются с клетками-хозяевами как птиц, так и человека. В некоторых вариантах осуществления новая VLP Bgag состоит из одного или нескольких различных типов HA 1-18 («Г1» - «Г18») и типов NA 1-11 («Н1» - «Н11») в качестве ее целевого белка-патогена (белков-патогенов), и экспрессирует и представляет белки. В соответствии с некоторыми другими вариантами осуществления новые VLP Bgag могут защищать или вызывать иммунный ответ у пациента на один или несколько различных подтипов гриппа, например, подтипов H5, H7, H9 и H10.
Используя данную новую платформу, изобретателями впервые создана и описана четырехподтипная VLP Bgag (см., например, Фиг. 10-11, 13-16 и пример 11). В некоторых вариантах осуществления четырехподтипная VLP Bgag ко-экспрессировала NA гриппа, полученный из подтипов гриппа 5, 7, 9 и 10. Благодаря этому четырехподтипная VLP может одновременно экспрессировать и представлять белки H5, H7, H9 и H10 птичьего гриппа на поверхности VLP для обеспечения одновременной иммунной защиты от всех четырех подтипов вируса гриппа. В вариантах также может быть ко-экспрессирован и сопредставлен (или не экспрессирован и не представлен) NA одного и того же или разных типов. В некоторых вариантах осуществления NA из таких четырехподтипных VLP Bgag локализованы с образованием гомотримеров, при этом в некоторых других вариантах осуществления HA образуют гетеротримеры или смесь гомотримеров и гетеротримеров. В некоторых вариантах осуществления VLP Bgag ко-экспрессирует и сопредставляет три различных типа HA и один тип NA.
Вирусы подтипов H5, H7, H9 и H10 птичьего гриппа были идентифицированы как патогены, опасные с точки зрения возникновения пандемий (Belser et al., 2008; Garcia-Sastre and Schmolke, 2014; Palese, 2004; Pappas et al., 2007; WHO, 2013). В частности, вирусы VN/04 (H5N1) и HK/09 (H9N2) включены в список кандидатных вакцин, рекомендованных Всемирной организацией здравоохранения (ВОЗ) в 2012 году для обеспечения готовности к пандемии (ВОЗ, 2012a). Для повышения готовности к пандемии были одобрены инактивированные вакцины против H5N1 (O'Neill and Donis, 2009), включая противогриппозные вакцины, полученные из клеточных культур. Тем не менее, хотя сообщалось о наличии перспективных экспериментальных вакцин (Chen et al., 2014; Kong et al., 2015; Smith et al., 2013; Tretyakova et al., 2013; Wohlbold et al., 2015), в настоящее время нет одобренных человеческих вакцин для подтипов H7, H9 или H10 (ВОЗ, 2012a).
Новая VLP Bgag, экспрессирующая один или несколько различных целевых белков-патогенов, является отличной вакцинной платформой, так как она может вызывать устойчивый иммунный ответ против одного или нескольких различных целевых патогенов. Например, инъекционное или иное введение хорькам VLP Bgag H10, содержащей гемагглютинин вируса H10N1, вызывает образование у хорьков устойчивых нейтрализующих антител к Н10 (см. Фиг. 30 и 31). Аналогичным образом инъекционное или иное введение хорькам VLP Bgag H5/H7/H9/10 одновременно вызывает образование у хорьков устойчивых нейтрализующих антител к H5, H7, H9 и H10 (см. Фиг. 30 и 31). Кроме того, обнаружено, что полученные нейтрализующие антитела могут нейтрализовать вирусы из разных клад одного подтипа, что обеспечивает возможность создания нейтрализующих антител, обеспечивающих перекрестную защиту. Например, нейтрализующие антитела к H5, индуцированные с помощью четырехподтипной VLP Bgag H5/H7/H9/10, могут приводить к образованию нейтрализующих антител к вирусам клады 1, клады 0, клады 1.1.2, клады 2.2 и клады 2.2.2 H5N1 (см. Фиг. 31).
Новая VLP Bgag, экспрессирующая один или несколько различных целевых белков-патогенов, является отличной вакцинной платформой, так как она может обеспечить иммунную защиту вакцинированного пациента от одного или нескольких различных целевых патогенов. Например, инъекционное или иное введение хорькам VLP Bgag H10, содержащей гемагглютинин вируса H10N1, может защитить хорьков при последующих экспериментальных заражениях живым вирусом гриппа H10N1, о чем можно судить по значительному снижению реплицирования титров вируса, например, в носовой раковине и трахее (см. Фиг. 32). В качестве еще одного примера, инъекционное или иное введение хорькам четырехподтипной VLP Bgag H5/H7/H9/10 также защищает хорьков при последующих экспериментальных заражениях живым вирусом гриппа H10N1, о чем можно судить по значительному снижению реплицирования титров вируса, например, в носовой раковине и трахее (см. Фиг. 32).
Новая VLP Bgag является отличной кандидатной вакциной широкого спектра действия, которая может обеспечить специфическую иммунную защиту по меньшей мере от четырех вирусных подтипов, штаммов или клад. Как описано в настоящем раскрытии, новая кандидатная вакцина от нескольких подтипов или кандидатная вакцина от четырех подтипов, опосредуемая настоящей новой четырехподтипной VLP Bgag H5/H7/H9/10, обладает высокой эффективностью, в том числе для использования у людей. В отличие от традиционных смешанных вакцин, рассмотренных выше, иммунная защита широкого спектра действия, обеспечиваемая с помощью платформы на основе VLP Bgag, может быть достигнута без смешивания отдельных вакцин.
Новая VLP Bgag может экспрессировать и представлять одну или несколько различных клад одного подтипа целевых белков-патогенов
Кроме экспрессии и представления целевых патогенных белков, которые относятся к различным типам и подтипам, система на основе VLP Bgag также может быть использована для экспрессии и представления целевых белков-патогенов, относящихся к различным кладам одного подтипа. Например, VLP Bgag может экспрессировать грипп H5, относящийся к трем различным кладам, например, кладе 2.3.4.4 H5 (A/курица / Германия/ 2014), кладе 2.1.3 H5 (A/курица/ Западная Ява/ Субанг/29/2007) и кладе 2.2.1 H5 (A/курица Египет/ 121/2012). Полученная VLP Bgag гриппа H555 экспрессирует функциональный HA (см. Фиг. 33) и имеет морфологию, общую с другими VLP Bgag (см. Фиг. 34).
Полученная VLP Bgag H555 гриппа также демонстрирует сильную иммуногенность против гриппа H5. Например, после вакцинации цыплят VLP Bgag H555 вакцинированные цыплята демонстрируют сильную иммуногенность и наличие антител к H5, как показано стандартным анализом ингибирования гемагглютинина (см. Фиг. 35, 37 и 38). Кроме того, полученная VLP Bgag гриппа H555 одновременно защищает вакцинированных пациентов при экспериментальных заражениях из всех трех клад. Например, цыплята, вакцинированные VLP Bgag H555 в день 1 и день 3, а затем подвергнутые экспериментальному заражению в день 35 вирусом клады 2.3.4.4, вирусом клады 2.1.3 или вирусом клады 2.2.1, выжили все после экспериментального заражения (см. Фиг. 36). Наоборот, цыплята, вакцинированные плацебо, умерли в течение 6 дней после экспериментального заражения. Исследования иммуногенности также показывают, что цыплята, вакцинированные VLP Bgag H555, имели высокие титры антител против всех трех клад H5 при измерении через 2 недели после экспериментального заражения. Эффективность вакцины также показана уменьшением выделения вируса в ротовой полости и клоаке у вакцинированных пациентов. Например, титры вирусов, присутствующие в мазках со слизистой ротовой полости и клоаки на второй день и на четвертый день после экспериментального заражения у цыплят, вакцинированных VLP Bgag H555, значительно ниже, чем у невакцинированных цыплят, независимо от того, относится ли используемый для заражения вирус к кладе 2.3.4.4, кладе 2.1.3 или кладе 2.2.1 (см. Фиг. 39-41).
Способ получения VLP Bgag, экспрессирующей один или несколько различных целевых белков-патогенов.
Изобретателями также разработаны новые способы эффективного получения VLP Bgag, экспрессирующих и представляющих один или несколько различных целевых белков-патогенов (см. примеры 1-3 и примеры 5-12). В некоторых вариантах осуществления ДНК-вектор экспрессирован в вирусе-носителе для создания новой VLP Bgag, и предпочтительно, чтобы вирусом-носителем был рекомбинантный бакуловирус («rBV»). В некоторых вариантах осуществления rBV, содержащий ДНК-вектор, экспрессирован в эукариотических клетках, предпочтительнее, клетках Spodoptera frugiperda («Sf9»). В некоторых вариантах осуществления каждый ген целевого белка-патогена контролируется индивидуальным промотором и, предпочтительнее, отдельным промотором полиэдрина. В некоторых вариантах осуществления настоящий способ используют для получения VLP Bgag, экспрессирующих и представляющих несколько различных целевых белков-патогенов. В некоторых других вариантах осуществления способ используют для получения VLP Bgag, экспрессирующих и представляющих белки-патогены нескольких подтипов. В соответствии с некоторыми другими вариантами осуществления способ включает в себя использование ДНК-вектора, содержащего ген Bgag и ген (гены) из одного или нескольких различных типов целевых патогенов, при этом все они организованы последовательно и каждый контролируется промотором, предпочтительнее, промотором полиэдрина; вирус-носитель, предпочтительно rBV; и эукариотную клетку, предпочтительнее Sf9, для получения VLP Bgag, экспрессирующей один или несколько различных целевых белков-патогенов.
Усовершенствованный способ производства вакцин
VLP, экспрессирующие один или несколько различных целевых белков-патогенов, таких как описанные в настоящей заявке четырехподтипные VLP Bgag, превосходят традиционные способы производства вакцин, так как в них не используют яйца, и они могут быть получены в рамках одного производственного цикла без необходимости смешивания вакцин, при этом они эффективно стимулируют иммунитет широкого спектра действия против нескольких целевых белков-патогенов. В одном варианте способ включает в себя клонирование генов HA, NA и Bgag гриппа в один вектор rBV для ко-экспрессии белков для получения VLP Bgag. Преимущество использования одного вектора rBV состоит в том, что это приводит к ко-экспрессии нескольких генов в инфицированной клетке Sf9, что обеспечивает более рациональную методологию и модель, которые можно легко расширять. Кроме того, этот способ позволяет клонировать штамм-специфический антиген целевых белков гриппа в готовый вектор переноса rBV, содержащий стандартные гены Bgag и NA, что дополнительно упрощает процесс производства вакцин против сезонного гриппа. Например, описанная в настоящей заявке VLP Bgag четырех подтипов является превосходной кандидатной вакциной для штаммов сезонного гриппа по сравнению с используемыми в настоящее время способами смешивания трех вакцин, получаемых с использованием яиц. В совокупности, эти превосходные модель и методология облегчают подготовку векторов и ускоряют производство VLP и получение вакцин.
Кроме того, описанная в настоящей заявке новая VLP Bgag является более надежной системой производства вакцин в случае пандемии или появления новых вирусов. Например, пандемический грипп или высоко вирулентный новый вирус гриппа может привести к серьезным нагрузкам на разработку и производство традиционных вакцин. Пандемия или новый вирус гриппа могут вызывать эпизоотию видов сельскохозяйственной птицы. Таким образом, помимо угрозы здоровью человека пандемия или новый вирус гриппа могут серьезно угрожать поставкам здоровых цыплят и яиц, что приводит к нехватке яиц, пригодных для производства вакцин. Так как производство вакцин против негриппозных патогенов, таких как желтая лихорадка, инфекционный паротит и корь, также в значительной степени зависит от яиц, эпизоотия будет создавать дополнительную нагрузку на производство и доступность таких производимых на основе яиц вакцин, что создаст еще большее напряжение в области здравоохранения.
Как минимум, по данным причинам новая моно- или многовалентная VLP Bgag, описанная в настоящей заявке, является важной вакцинной платформой во время и в преддверии таких угрожающих ситуаций. Кроме того, платформа на основе VLP Bgag может обеспечить быструю и надежную защиту широкого спектра действия для людей и/или животных от опасных пандемических, эпизоотических или новых патогенов. В случае вспышки заболеваемости, вызванной вирусами птичьего гриппа H5, H7, H9 и/или H10, описанная в настоящей заявке вакцина против гриппа четырех подтипов H5/H7/H9/H10 будет ценной первой линией защиты, особенно если недоступны иные вакцины.
VLP Bgag в качестве превосходной кандидатной вакцины
В дополнение к преимуществам, описанным в других местах в спецификации, новая VLP Bgag также является превосходной кандидатной вакциной и, в частности, превосходной кандидатной вакциной для человека. Одна из причин состоит в том, что белок Bgag внутреннего ядра не имеет значительной гомологии с патогенами человека. БВИ не является патогентом человека, и, таким образом, у людей, как правило, нет ранее существовавшего иммунитета против белков из БВИ, например, Bgag. Анализ последовательности Bgag при помощи программного обеспечения NCBI BLASTP с параметрами по умолчанию показал, что самое близкое сходство составляло лишь приблизительно 29% генетического сходства с вирусом иммунодефицита кошек и приблизительно 26% генетического сходства с вирусом инфекционной анемии лошадей. Кроме того, изобретателями не было обнаружено сходства между БВИ и человеческими белками ретровирусными gag, включая белки gag ВИЧ. Таким образом, VLP, опосредованные Bgag, имеют значительные преимущества как кандидатная вакцина для человека, поскольку она позволяет обойти ранее существовавший иммунитет хозяина и может вызывать более сильные и более устойчивые иммунные ответы, чем другие VLP, например, VLP M1.
В некоторых вариантах осуществления каждая VLP Bgag содержит большую площадь поверхности, предпочтительно больше, чем у патогена, из которого получен один из целевых белков. В соответствии с настоящим описанием, VLP Bgag с более крупным размером превосходит VLP M1 меньшего размера. Более значительная площадь поверхности может быть особенно важна для VLP Bgag, представляющих несколько целевых белков, например, VLP Bgag H5/7/9/10, описанных в примере 11, в которых несколько подтипов HA ко-локализованы на одной оболочке VLP. Изобретателями в настоящей заявке показано, что новые VLP Bgag имеют средний диаметр от приблизительно 150 нм до приблизительно 180 нм по сравнению со средним значением от приблизительно 120 нм до приблизительно 150 нм для VLP M1 (см. Фиг. 6-7). Этот увеличенный размер VLP создает значительно большую площадь поверхности для большего количества целевых белков-патогенов, представленных VLP Bgag, по сравнению с VLP M1. Например, больше тримеров HA может локализоваться и быть представлено в VLP Bgag, чем в VLP M1 (см. Фиг. 6-7).
В некоторых вариантах осуществления каждый VLP Bgag содержит в среднем более приблизительно 375 шипов белка на мембране VLP, предпочтительно от приблизительно 375 шипов до приблизительно 800 шипов, более предпочтительно от приблизительно 475 шипов до приблизительно 800 шипов, более предпочтительно от приблизительно 575 шипов до приблизительно 800 шипов, более предпочтительно от приблизительно 675 шипов до приблизительно 800 шипов, более предпочтительно от приблизительно 775 шипов до приблизительно 800 шипов и, более предпочтительно, приблизительно 800 шипов. Было подсчитано, что сферический вирион со средним диаметром 120 нм, по оценкам, содержит приблизительно 375 шипов (Harris et al., 2006). Исходя из того, что тримеры распределены равномерно, VLP с диаметром приблизительно 180 нм может вмещать приблизительно 800 шипов, что более чем в два раза превышает количество шипов, наблюдаемое в VLP с диаметром всего 120 нм. Таким образом, VLP Bgag как кандидатная вакцина превосходит VLP M1, так как она может иметь более высокую эффективность при стимулировании иммунного ответа хозяина и может вызвать более устойчивый иммунный ответ хозяина.
Использование гена Bgag или белка Bgag в качестве диагностического инструмента
В некоторых вариантах осуществления ген Bgag или белок Bgag используют в качестве диагностического инструмента. Одна из проблем используемых в настоящее время вакцин и других вакцин на основе VLP, например, вакцины VLP M1, состоит в дифференциации вакцинированных пациентов от невакцинированных пациентов. По всей вероятности, для системы VLP Bgag это не будет проблемой. Например, как описано в настоящем документе, в отличие от M1, люди и некоторые животные обычно не подвергаются воздействию гена Bgag или белка Bgag. Соответственно, медицинский работник может использовать ген Bgag или белок Bgag для определения, вводили ли ранее пациенту вакцину на основе VLP Bgag. Эта диагностическая способность была бы невозможна, например, в вакцинах, опосредуемых VLP M1, вследствие повсеместного распространения гена M1 и белка M1 у населения и в определенной популяции животных. Это также может быть особенно важно в применении в ветеринарии, например, когда важно дифференцировать инфицированных от вакцинированных животных («DIVA») (Rahn et al., 2015; Suarez, 2012).
VLP Bgag с целевыми нуклеиновыми кислотами
Нуклеиновая кислота также может быть встроена в VLP Bgag для стимулирования или усиления иммунного ответа целевого объекта. Полученные авторами изобретения последние данные показывают, что в VLP Bgag может быть встроена РНК (см. Фиг. 25-27). Сборка VLP Bgag выполняется при отсутствие инфекционного РНК БВИ и белка оболочки БВИ. Таким образом, присутствующая РНК, обнаруженная в VLP, вероятно, получена из клеток Sf9. Не ограничиваясь конкретной теорией, мы считаем, что РНК, присутствующая в структуре VLP Bgag, может индуцировать или усиливать иммунный ответ. Исходя из той теории, что РНК может быть инкапсулирована во время производства VLP Bgag, считается, что инкапсулированная РНК является иммунным модулятором и может усилить иммунный ответ, присутствие РНК в VLP Bgag может индуцировать или усиливать иммунный ответ пациента. В качестве еще одного примера, Coffman et. al (2010) предполагают, что РНК может быть естественным агонистом толл-подобного рецептора TLL7 и обладает иммуностимулирующими свойствами, которые могут работать как адъювант через активацию врожденного иммунитета, включая TLR7. Насколько известно авторам изобретения, получение VLP Bgag, содержащего этот тип РНК в структуре VLP Bgag, и наличие и эффект этого типа РНК в VLP Bgag не были известны и/или не описаны.
Примеры
Пример 1
Генерация плазмиды рекомбинантного вектора переноса для получения VLP Bgag и VLP M1
Были получены последовательности белков в белке внутреннего ядра и в целевых белках-патогенах. Судя по указанным последовательностям белков гены кодон-оптимизированы для осуществления высокоуровневой экспрессии в клетках Sf9 (Life Technologies, Карлсбад, Калифорния) и синтезируются биохимически (Genscript, Пискатауэй, Нью-Джерси). Для генерации VLP гены белка Bgag или M1 внутреннего ядра и гены одного или нескольких различных целевых белков-патогенов, например, HA и NA гриппа, клонируют совместно в плазмиду вектора переноса бакуловируса, так что каждый ген находится внутри собственной транскрипционной кассеты, которая включала в себя промотор полиэдрина выше по потоку от каждого гена. На Фиг. 1 показаны четыре иллюстративных схематических диаграммы. Конструкция клонируется в рекомбинантный бакуловирус («rBV») в клетках Spodoptera frugiperda («Sf9»). Гены белка внутреннего ядра, NA и HA, кодон-оптимизированы для осуществления высокоуровневой экспрессии в клетках насекомых и клонируются в rBV совместно, как показано. Каждая конструкция содержит набор промоторов полиэдрина, которые в первой конструкции для наглядности представлены в виде стрелок. Процесс по существу соответствует процессу, описанному в работе (Pushko et al., 2005), которая включена в настоящую заявку посредством ссылки. В приведенных примерах все гены HA представляют собой HA1 («H1») и получены из одного вируса гриппа A/Пуэрто-Рико/8/1934 (H1N1) («PR8») (показан в виде черных квадратов). Тем не менее, это не обязательно так. Аналогично генам NA, гены HA могут быть выбраны из того же самого вируса PR8 (показан в виде черных квадратов) или другого вируса, например, вируса гриппа A/Индонезия/5/2005 (H5N1) («IN/5») (показан в виде серых квадратов). Создают конструкции, содержащие белок gag БВИ («Bgag») или M1 внутреннего ядра. VLP M1 используют в качестве контроля. В приведенных примерах ген M1 (показан пунктирными линиями) выбран из вируса IN/5.
Экспрессия и функциональные профили белков VLP HA и NA гриппа показаны на Фиг. 2-5.
VLP гриппа, созданные из каждой из четырех конструкций, показанных на Фиг. 1, представляют функциональные белки HA, как показано в анализе гемагглютинации с использованием эритроцитов индейки (см. Фиг. 2). Для инфицирования клеток Sf9 используют rBV, содержащий каждую из четырех конструкций, показанных на Фиг. 1. VLP гриппа очищают от клеток Sf9 в день 3 после инфицирования и выполняют анализ гемагглютинации. Эритроциты индейки серийно разводят с интервалами в 2 раза, начиная с разведения 1:56. Очищенные VLP добавляют в каждую лунку. Самая правая линия содержала PBS и была использована как отрицательный контроль. В данной реакции функциональный HA из VLP связывается с эритроцитами индейки с образованием решетки эритроцитов, которая не осаждается, тем самым создается диффузный внешний вид в лунке. В отрицательном контроле отсутствует функциональный HA, и решетка эритроцитов не образуется. Соответственно, эритроциты осаждаются из раствора и выглядят как точка в центре лунки. Результаты этого анализа гемагглютинации показывают, что VLP, созданные из всех четырех конструкций, экспрессируют функциональный HA, этот HA локализуется на мембране VLP, и эти HA представлены на поверхности VLP.
VLP гриппа, созданные из каждой из конструкций, показанных на Фиг. 1, экспрессируют белки H1, как показано в вестерн-блоте с использованием моноклонального антитела, специфического к H1 (см. Фиг. 3). В крайнем правом ряду обозначены маркеры в килодальтонах. Локализация белков H1 обозначена стрелкой.
VLP гриппа, созданные из конструкций 1-2, показанных на Фиг. 1, содержат белок Bgag внутреннего ядра, при этом VLP гриппа, созданные из конструкций 3-4, показанных на Фиг. 1, содержат белок M1 внутреннего ядра (см. Фиг. 4). VLP из каждой из четырех конструкций загружают в гель для электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия и окрашивают кумасси для оценки профиля белка VLP. В крайнем правом ряду обозначены маркеры белка в килодальтонах. Локализация HA, белков Bgag и M1 обозначена стрелкой.
VLP гриппа, созданные из каждой из конструкций, показанных на Фиг. 1, содержат функциональный NA (см. Фиг. 5). VLP из каждой из конструкций, показанных на Фиг. 1, серийно разводят и оценивают ферментативную активность NA посредством флуоресцентного анализа, включающего NA-Fluor и N-ацетил-нейраминную кислоту метил-умбеллиферона. Ферментативную активность NA измеряют в относительных единицах флуоресценции («RFU»). Конструкции VLP показаны следующим образом: конструкция 1 - закрашенный квадрат, конструкция 2 - пустой ромб, конструкция 3 - пустой круг и конструкция 4 - сплошная линия. Отрицательный контроль в форме PBS показан в виде залитых треугольников. Линия нормализации показана пунктирной линией. Результаты этого флуоресцентного анализа показывают, что все четыре конструкции экспрессируют функциональную NA.
VLP состоят из белка Bgag внутреннего ядра, который имеет ту же морфологию, как и вирус гриппа, при рассмотрении под просвечивающим электронным микроскопом (см. Фиг. 6). VLP окрашивают 1% фосфорновольфрамовой кислотой. Полоска маркирует 100 нм.
VLP состоят из белка M1 внутреннего ядра, который имеет ту же морфологию, как и вирус гриппа, при рассмотрении под просвечивающим электронным микроскопом в тех же условиях, что и на Фиг. 6 (см. Фиг. 7).
Примеры источника белковых последовательностей следующие.
gag R-29 БВИ: GenBank, номер доступа AAA42763.
M1 гриппа IN/05: GenBank, номер доступа ABI36004.
HA гриппа PR8: GenBank, номера доступа ABP64731.
HA гриппа VN/04: GenBank, номера доступа AAW80717.
HA гриппа SH/13: GenBank, номера доступа YP_009118475.
HA гриппа HK/09: GenBank, номера доступа AGO17847.
HA гриппа JX/13: GenBank, номер доступа AHK10762.
NA гриппа PR8: GenBank, номер доступа ABD77678.
NA гриппа IN/05: GenBank, номер доступа ABW06107.
GP Эбола EboMay: последовательность приведена на Фиг. 17.
GP-TMCT Эбола EboMay: последовательность приведена на Фиг. 18.
Примеры источника последовательностей генов:
Последовательности генов HA гриппа были произведены из вирусов PR8, VN/04, SH/13, HK/09 и JX/13.
Пример 2
Генерация плазмиды рекомбинантного вектора переноса для получения четырехподтипных VLP Bgag
Белковые последовательности Bgag и четырех различных целевых белков-патогенов кодон-оптимизировали, а гены соответствующих белков клонировали, как в примере 1, в единую плазмиду вектора переноса бакуловируса. Например, четыре полноразмерных гена различных HA пандемических гриппов, а также NA гриппа IN/05 клонировали в плазмиду вектора переноса, по существу, как показано на Фиг. 13.
Пример 3
Генерация rBV для получения VLP Bgag и VLP M1
Бакмиды, содержащие полноразмерную инфекционную ДНК бакуловируса с рекомбинантными генами для VLP Bgag, выделяли из DH10Bac E.coli с использованием системы экспрессии бакуловируса Bac-to-Bac (Life Technologies, Карслбад, Калифорния) и использовали их для трансфицирования клеток Sf9 для получения rBV. Затем препараты rBV очищали методом бляшек. Титры препаратов rBV определяли с помощью стандартного анализа бляшек в клетках Sf9.
Клетки Sf9 содержали в виде суспензионных культур в среде SF900II-SFM, не содержащей сыворотку насекомых (Life Technologies, Карслбад, Калифорния) при 27°C. Для производства VLP использовали клетки Sf9 при 2×106 клеток/мл и инфицировали множественными инфекциями («MOI») 3,0 в течение 72 часов с rBV, экспрессирующим целевые гены. VLP собирали из супернатанта питательной среды, осветляли фильтрованием через мембрану с размером пор 0,2 мкм, далее концентрировали и очищали при помощи ступенчатого градиента сахарозы 20% (вес/об.) в физиологическом растворе с фосфатным буфером («PBS»). Альтернативно, VLP сначала очищали ионообменной хроматографией, как описано в другой работе (Liu et al., 2015), а потом очищали ультрацентрифугированием.
Пример 4
Оценка VLP
SDS-PAGE выполняли в 4-12% полиакриламидных гелях ((Life Technologies, Карслбад, Калифорния) с последующим окрашиванием красителем GelCode Blue Pierce, Рокфорд, Иллинойс).
Вестерн-блоты выполняли с использованием специфических первичных антител с последующей обработкой козьего IgG против хорьков, конъюгированного с щелочной фосфатазой (H+L). Примеры используемых первичных антител: мышиное моноклональное антитело (MAb) IgG1 к HA клады 1 вирусов гриппа A H1N1 IT-003-001M14, клон 15B7; мышиное моноклональное антитело IgG2a к H5 (H5N1) IT-003-005M6, клон 268D8; мышиное моноклональное антитело IgG1 к HA (H7N9) (A/Шанхай/1/2013) IT-003-0073M1, клон 9B12; мышиное моноклональное антитело IgG1 к H9 (A/Гонконг/33982/2009) (H9N2) IT-003-0094M5, клон 17D8; и кроличье поликлональное антитело к H10 (A/голубокрылый чирок/Луизиана/Sg-00073/07 (H10N7)) IT-003-034 (Immune Technology, Нью-Йорк, Нью-Йорк).
Концентрацию белка VLP определяли флуорометрическим способом Qubit 2.0 (Life Technologies).
Функциональные анализы гемагглютинации в общем проводили следующим образом. VLP серийно разводили с 2-кратным увеличением в объеме 50 мкл в 96-луночном планшете. К каждому разведению VLP добавляли 50 мкл 1% рабочего раствора эритроцитов индейки; смеси VLP и эритроцитов осторожно перемешивали и планшет инкубировали при комнатной температуре в течение 30-60 минут перед исследованием. Отрицательные результаты гемагглютинации выглядели как точки в центре лунок. Титр рассчитывали как самый высокий коэффициент разведения, который дал положительное показание. Положительный результат реакции гемагглютинации показывает, что VLP экспрессируют функциональный HA, этот HA локализуется на мембране VLP, и эти HA представлены на поверхности VLP.
Функциональные анализы фермента NA гриппа в общем проводили следующим образом. Функциональную ферментативную активность нейраминидазы определяли с использованием анализа NA на основе флуоресценции (NA-Fluor, Life Technologies) с N-ацетилнеураминовой кислотой метил-умбеллиферона (MUNANA; Sigma, Сент-Луис, Миссури) в качестве подложки согласно инструкциям производителя. Разбавитель (физиологический раствор или PBS) использовали в качестве отрицательного контроля.
Просвечивающую электронную микроскопию выполняли абсорбцией образцов VLP на свежеразряженные медные сетки с покрытием углеродным парлодием с размером ячейки 400 меш (Poly-Sciences, Уоррингтон, Пенсильвания). Сетки промывали буфером, содержащим 20 мМ Трис, рН 7,4 и 120 мМ KCl и отрицательно окрашивали 1% фосфорновольфрамовой кислотой, затем высушивали аспирацией. VLP визуализировали на просвечивающем электронном микроскопе Hitachi H-7600 (Hitachi High Technologies America, Шомберг, Иллинойс), работающем при напряжении 80 кВ и осуществляли их цифровой захват камерой на ПЗС с разрешением 1k × 1k (Advanced Microscopy Techniques Corp., Данверс, Массачусетс).
Генетическую стабильность rBV и стабильность экспрессии VLP Bgag можно продемонстрировать стандартными методами, например, серийным пассажем rBV в клетку-носитель, и можно оценить экспрессию VLP, содержащих целевой белок. Например, можно продемонстрировать, что экспрессия четырехподтипной VLP Bgag гриппа может быть стабильной, с помощью первоначального пассажа rBV, содержащего VLP Bgag гриппа четырех подтипов, в клетки Sf9, при МЗ 0,01 пять раз, затем экспрессией VLP и измерения наличия функционального HA в функциональной реакции гемагглютинации (см. Фиг. 22). Как показано на Фиг. 22, анализ гемагглютинации четырехподтипной VLP Bgag, полученных в клетках Sf9, инфицированных rBV при пассажах P1-P5. VLP Bgag собирают из супернатантов клеток Sf9, инфицированных rBV, фильтровали через мембрану с размером пор 0,22 мкм, концентрировали в 100 раз ультрацентрифугированием и ресуспендировали в PBS. Анализ гемагглютинации проводили с использованием эритроцитов индейки начиная с разведения VLP 1:128. В качестве отрицательного контроля, маркированного как NC, вместо VLP использовали PBS.
Колокализацию целевых белков-патогенов в VLP Bgag можно продемонстрировать стандартными способами, например, иммунопреципитацией или иммуноэлектронной микроскопией. Например, можно показать, что H9, H10 и H7 колокализуются с H5 на VLP Bgag гриппа H5/H7/H9/H10 за счет первой иммунопреципитации VLP Bgag гриппа H5/H7/H9/H10 с использованием магнитных микроносителей SureBeads (BioRad, Геркулес, Калифорния) и антитела к H5. Сначала можно выполнить связывание SureBeads с антителом к H5 при +20°C в течение 1 часа. Любое несвязанное антитело к H5 можно вымыть. Затем VLP Bgag инкубировали с SureBeads с антителом к H5. Любые VLP Bgag, не связанные с SureBeads, вымывали. Захваченные VLP Bgag можно элюировать буфером по Laemmli при +70°C в течение 10 минут, затем анализировать на SDS-PAGE и/или вестерн-блоте с использованием антител к H7, H9 и/или H10 для подтверждения колокализации целевых белков HA (см. Фиг. 23). В качестве еще одного примера, колокализация также может быть продемонстрирована иммуноэлектронной микроскопией, в частности, описанной в работе (Pushko et al., 2011).
Как показано на Фиг. 23, панель a, различные подтипы HA колокализуются с теми же четырехподтипными VLP H5/7/9/10, как продемонстрировано при иммунопреципитации с использованием H5-специфического моноклонального антитела с последующим вестерн-блотом с использованием моноклональных антител, специфичных к H5, H7 и H9, и кроличьей антисыворотки, специфичной к H10. Образцы 1-5 представляют собой rH5, rH7, rH9, rH10 и антигены четырехподтипной VLP Bgag, соответственно. C представляют собой отрицательный контроль иммунопреципитации, который является буфером PBST.
Количество каждого типа целевого белка-патогена, присутствующего в VLP Bgag, также может быть определено стандартными методами, например, полуколичественным вестерн-блотом. Например, количество H5, H7, H9 и H10, присутствующих в VLP Bgag гриппа H5/H7/H9/H10, может быть измерено сравнением четырехподтипных VLP Bgag с очищенными стандартами белка H5, H7, H9, H9 с использованием флуориметрического метода Qubit 2.0 (Life Technologies). Концентрацию очищенного стандарта белка HA можно измерить сравнением его денситометрии на окрашенном SDS-PAGE со стандартом BSA с известной концентрацией. Может быть сгенерирована линейная стандартная кривая для стандарта BSA с использованием программного обеспечения NIH ImageJ, и может быть определено количество очищенного стандарта белка HA. Для определения концентрации каждого из белков HA в четырехподтипных VLP Bgag может быть выполнен полуколичественный вестерн-блот, где интенсивность полосы четырехподтипных VLP Bgag измеряют денситометрией и сравнивают со стандартом HA с известной концентрацией (см. Фиг. 23 и 24).
Как показано на Фиг. 23, панель b, распределение подтипов HA на VLP Bgag может быть измерено полуколичественным вестерн-блотом. Как показано на верхней панели b, rH5, rH7, rH9 и rH10 одноподтипной VLP Bgag и четырехподтипной VLP Bgag H5/7/9/10 определяют SDS-PAGE с окрашиванием кумасси голубым, а количественно определяют денситометрией. Полоса 54 кДа в дорожке 5 представляет собой белок Bgag, окрашенный кумасси голубым. В нижних панелях выполнены вестерн-блоты с указанными антителами. Содержание каждого подтипа HA в VLP определяют с использованием вестерн-блота сравнением интенсивности полосы в дорожке VLP с интенсивностью полосы известного количества соответствующего эталонного антигена rHA. Образцы 1-7 представляют собой одноподтипной VLP Bgag rH5, rH7, rH9 и четырехподтипной VLP Bag H5/7/9/10, 0,2 мг/мл альбумина бычьей сыворотки (5 мкл) и 0,1 мг/мл альбумина бычьей сыворотки (5 мкл), соответственно. M представляют собой набор молекулярных весов белков See Blue Plus2 (Life Technologies).
На основе полуколичественного вестерн-блота определяют распределение HA в четырехподтипных VLP Bgag (см. Фиг. 24).
Пример 5
Получение VLP Bgag PR8 H1
VLP готовят с использованием белков Bgag и PR8 гриппа в системе экспрессии рекомбинантного бакуловируса (rBV), опосредованной клетками Sf9, по существу, как описано в примерах 1-3. H1 гриппа PR8 клонировали в rBV вместе с полноразмерными генами Bgag и NA, по существу, как показано на Фиг. 1. Ген NA получен из вируса гриппа PR8 или IN/05. Для целей сравнения изобретатели также генерировали rBV с геном M1 вместо гена Bgag. В совокупности, получали четыре вектора rBV, по существу, как показано на Фиг. 1. Каждый rBV содержал три гена тандемным образом, причем каждый ген в своей собственной кассете экспрессии контролировался собственным промотором полиэдрина. Клетки Sf9 инфицировали rBV с МЗ 3 и инкубировали в течение 72 часов для обеспечения экспрессии VLP. VLP собирали из питательной среды инфицированных клеток и концентрировали и частично очищали ультрацентрифугированием. На Фиг. 2-7 показана оценка VLP с использованием способа, по существу описанного в примере 4.
Все четыре препарата VLP демонстрировали активность HA с эритроцитами индейки с приблизительно одинаковыми титрами, как показано на Фиг. 2. Экспрессию PR8 H1 ожидаемых 63,44 килодальтон (кДа) (565 аминокислотных остатков) детектировали с использованием антитела, специфического к H1, с помощью вестерн-блота, как показано на Фиг. 3. В соответствии с предыдущими наблюдениями (Perrone et al., 2009; Pushko et al., 2011; Pushko et al., 2005) PR8 H1 экспрессирован как полноразмерный полипептид HA0, без переработки в HA1 и HA2, детектированных вестерн-блотом. На окрашенном геле SDS-PAGE обнаруживают полосы белков HA, gag и M1 с их ожидаемой молекулярной массой около 63 кДа, 54 кДа и 28 кДа соответственно, как показано на Фиг. 4.
NA гриппа не обнаружена c помощью вестерн-блота или SDS-PAGE, что соответствует данным предшествующих исследований. Тем не менее, VLP демонстрировали ферментную активность NA в функциональном анализе NA, как показано на Фиг. 5, что подтверждает наличие функционального белка NA в VLP. Активность фермента NA была выше в препаратах VLP, содержащих NA IN/5, по сравнению с VLP, содержащими NA PR8. NA IN/5 и NA PR8 относятся к подтипу N1 и обе содержат 83% идентичных аминокислотных остатков. Несмотря на различия в активности NA, изобретатели наблюдали приблизительно одинаковую экспрессию HA, как показано на Фиг. 2-4.
Наконец, электронно-микроскопическое исследование подтвердило наличие оболочечных частиц в препаратах VLP (Фиг. 6-7). Диаметр VLP Bgag составляет приблизительно 150-200 нм, при этом диаметр VLP M1 составляет приблизительно 120-150 нм.
В совокупности эти результаты подтвердили образование VLP Bgag, экспрессирующих и представляющих белки HA и NA.
Пример 6
Получение VLP Bgag JX/13 H10
Изобретателями подготовлены VLP Bgag, содержащие новые штаммы гриппа, например, вирус птичьего происхождения JX/13 (H10N8), при помощи способов и принципов, по существу аналогичных описанным в примерах 1-3 и примере 5. В 2013 году вирус гриппа JX/13 (H10N8) вызвал заражение человека и был выявлен как патоген, способный вызывать пандемию (Garcia-Sastre and Schmolke, 2014; To et al., 2014). Вирус гриппа JX/13 (H10N8) первоначально был выделен у пожилого пациента в Китае, умершего в результате инфекции (Garcia-Sastre and Schmolke, 2014; To et al., 2014).
rBV сконфигурирован для экспрессии трех генов: гена H10, полученного из вируса гриппа JX/13, гена NA и гена Bgag. На Фиг. 1 показано примерное схематическое представление конструкции rBV, экспрессирующей VLP гриппа, содержащей HA 10 («H10») в клетках Sf9. Гены белка внутреннего ядра Bgag, NA и НА кодон-оптимизированы для осуществления высокоуровневой экспрессии в клетках насекомых и клонируются в rBV совместно, как показано. Каждая конструкция содержит набор промоторов полиэдрина, показанных стрелками. В приведенных примерах ген HA получен из вируса A/Цзянси/IPB13a/2013 (H10N8) («JX/13») (показан в виде черных квадратов), а ген NA получен из вируса IN/5 (показан в виде серых квадратов). Полученный rBV используют для инфицирования клеток Sf9 для получения VLP JX/13 H10. VLP, содержащие H10, NA и Bgag, собирали из 2 л питательной среды Sf9 и очищали ионообменной хроматографией. Морфология VLP гриппа, экспрессия и функциональные профили белков HA и НА гриппа показаны на Фиг. 9-12 и 15 с использованием способа, по существу описанного в примере 4.
VLP гриппа, созданные из каждой из конструкций, показанных на Фиг. 8, содержат функциональные белки HA, как показано в анализе гемагглютинации. Эритроциты индейки серийно разводили с интервалом в 2 раза, начиная с разведения 1:210. VLP Bgag H10 демонстрирует функциональную активность HA с титром 1:8192 или 1:213 на 50 мкл 3,5 мг/мл общего содержания белка. Результаты этого анализа гемагглютинации показывают, что VLP, созданные из конструкций, показанных на Фиг. 8, экспрессируют функциональный HA, этот HA локализуется на мембране VLP, и эти HA представлены на поверхности VLP.
VLP гриппа, созданные из конструкции H10, показанной на Фиг. 8, и четырех подтипов H5/7/9/10, показанных на Фиг. 13, экспрессируют соответствующие белки HA, как показано вестерн-блотом (см. Фиг. 10). VLP H10 (дорожка 1), четырехподтипные VLP H5/7/9/10 (дорожка 2), контроль VLP H5 (дорожка 3) окрашивают кроличьим антителом к H10N7, моноклональным антителом к H5N1, моноклональным антителом к H7N9 и моноклональным антителом к H9N2. Отмечены маркеры в килодальтонах. Локализация белков HA обозначена стрелкой. Результаты показывают, что VLP Bgag H10 реагирует с антителами, специфическими к H10, в вестерн-блотах (см. Фиг.10, дорожка 1), но не реагирует перекрестно с антителами, специфическими к H5, H7 или H9 (см. Фиг. 10) Полосы как HA, так и Bgag могут быть обнаружены окрашенным SDS-PAGE.
Также подтверждена функциональная активность фермента NA (см. Фиг. 15). Полосы белков H10 и Bgag также обнаруживают окрашенным SDS-PAGE (см. Фиг. 11, дорожка 1).
VLP H10, созданные из конструкции, показанной на Фиг. 8, имеют морфологию, аналогичную морфологии вируса гриппа, при рассмотрении под просвечивающим электронным микроскопом в условиях, описанных на Фиг. 6 (см. Фиг. 12). Согласно измерениям, диаметр оболочечных VLP H10 составляет приблизительно 150-180 нм.
Пример 7
Получение VLP Bgag только с HA или только с NA
Получение и оценка VLP Bgag также могут быть выполнены только с использованием только HA или только NA в качестве целевого белка-патогена с использованием способов и принципов, по существу описанных в примерах 1-6.
Пример 8
Получение VLP Bgag с генетически модифицированными HA и/или NA
Получение и оценка VLP Bgag также могут быть выполнены с использованием генетически модифицированных HA и/или NA с использованием способов и принципов, по существу описанных в примерах 1-7.
Используя аналогичный принцип и подход, могут быть получены VLP Bgag с использованием генетически модифицированного NA. Например, VLP Bgag могут быть сконструированы таким образом, что будут включать в себя только частичные последовательности NA.
Пример 9
Получение VLP Bgag с негриппозными целевыми белками-патогенами.
Получение и оценка VLP Bgag также могут быть выполнены с использованием целевого белка других негриппозных патогенов с использованием способов и принципов, по существу описанных в примерах 1-8. Например, VLP Bgag могут быть созданы с использованием гликопротеина вируса Эбола (ГП), например, GP EboMay, последовательность белков которого описана на Фиг. 17. Такие VLP могут индуцировать иммунный ответ на гликопротеин Эбола и могут быть использованы в качестве вакцины для предотвращения заражения вирусом Эбола.
В качестве дополнительного примера, при применении тех же способов и принципов VLP Bgag можно создать с использованием целевых белков из других филовирусов. Полученная VLP Bgag очень полезна в качестве пан-филовирусной вакцины.
В качестве дополнительного примера, VLP Bgag также может быть создана с использованием гликопротеина шипов MERS. Полученная VLP Bgag очень полезна в качестве вакцины для профилактики инфекций MERS. В качестве дополнительного примера, VLP Bgag также может быть создана с использованием целевых белков иных коронавирусов. Полученная VLP Bgag очень полезна в качестве вакцины от коронавирусов.
В качестве дополнительного примера, VLP Bgag также могут быть созданы с использованием других антигенов из оболочечных и безоболочечных вирусов. Например, VLP Bgag также могут быть созданы с использованием гликопротеина оболочки ВИЧ. Полученная VLP Bgag очень полезна в качестве вакцины против ВИЧ-инфекций. VLP Bgag также могут быть созданы с использованием целевых гликопротеинов из других ретровирусов для получения вакцин против ретровируса.
Пример 10
Получение VLP Bgag с химерными целевыми белками-патогенами
Получение и оценка VLP Bgag также могут быть выполнены с использованием генетически модифицированных целевых белков-патогенов для оптимизации взаимодействий с Bgag с использованием способов и принципов, по существу описанных в примерах 1-9. Например, гликопротеины целевых оболочечных вирусов могут быть модифицированы с образованием химерных гликопротеинов. Изобретателями подготовлен пример такого химерного целевого белка-патогена генетической модификацией GP EboMay таким образом, чтобы он содержал трансмембранный домен и С-конец EboMay TMCT белка HA вируса гриппа, как показано на Фиг. 18. Последовательность белков трансмембранного домена и C-конца гриппа HA выделена подчеркиванием. Очистка и оценка VLP показаны на Фиг. 19-21 с использованием определенного способа, по существу описанного в примере 4.
VLP EboMay GP-TMCT экспрессируют соответствующий белок Эбола, как показано вестерн-блотом (см. Фиг. 19). Фракцию 1 VLP EboMay GP-TMC из пиковой фракции, полученной из колонки ионообменной хроматографии («IECC Fxn 1») загружали на градиент сахарозы и дополнительно очищают ультрацентрифугированием. Четырнадцать объединенных фракций («Fxns») из фракций 1-28 градиента сахарозы окрашивали антисывороткой к Эбола.
VLP EboMay GP-TMCT экспрессировали Bgag и соответствующий белок Эбола, как показано гелем SDS-PAGE, окрашенным кумасси (см. Фиг. 20). Дорожка 1 представляет собой объединенный пиковый градиент сахарозы Fxns 9-10 от IECC Fxn1. GP Эбола и Bgag отмечены стрелками на самом дальнем левом углу геля. Результат показывает, что в очищенных VLP присутствуют как GP Эбола, так и Bgag, потому что они совместно очищают под дорожкой 1. Дорожка 2 представляет собой объединенный пиковый градиент сахарозы Fxns 7-8 от IECC Fxn 3. Белки бакуловируса GP64, P39 и P10 отмечены стрелками в левой части геля. Результат показывает, что IECC Fxn 3 содержит главным образом бакуловирусное загрязнение и что GP Эбола, содержащий VLP Bgag, может быть очищен от бакуловирусного загрязнителя.
Морфология VLP EboMay GP-TMCT включает в себя общую сферическую оболочку, как видно при рассмотрении под просвечивающим электронным микроскопом (см. Фиг. 21). Полоска маркирует 500 нм.
Подобно VLP Bgag, экспрессирующей другие целевые белки-патогены, VLP EboMay GP VLP и VLP EboMay GP-TMCT Bgag могут индуцировать иммунный ответ на эпитопы GP Эбола и могут служить в качестве вакцины для профилактики вирусных инфекций Эбола. Например, VLP Bgag EboMay GP и VLP EboMay GP-TMCT Bgag инъекционно или иным образом вводят пациенту с целью стимулирования иммунного ответа у пациента. Затем стандартными анализами определяют иммунный ответ вакцинированного пациента на вирус Эбола. Например, в течение 2-4 недель производят забор крови вакцинированного пациента и антитело к Эбола (или, более конкретно, антитело к GP Эбола) определяют с помощью иммуносорбентного ферментного анализа, иммунофлуоресцентного анализа антител или других анализов на выявление антител и сравнивают с профилем антитела невакцинированных пациентов. Присутствие антитела к Эбола у вакцинированного пациента означает, что VLP Bgag вызывал иммунный ответ у пациента и, следовательно, является иммуногенным в живом организме. Эффективность VLP Bgag также могут продемонстрировать стандартные исследования экспериментальных заражений. Например, сероположительным вакцинированным пациентам проводят экспериментальное заражение патогенным вирусом Эбола в лаборатории с высоким уровнем защиты для оценки эффективности вакцины VLP Bgag в профилактике Эбола, вызывающего заболевание. Выживание вакцинированных пациентов после экспериментального заражения является показателем защитной эффективности вакцин VLP Bgag.
Пример 11
Получение четырехподтипных VLP Bgag H5/7/9/10
Изобретатели подготовили и оценили первые четырехподтипные VLP Bgag с использованием способов и принципов, по существу описанных в примерах 1-10. Изобретатели использовали белки HA, полученные из четырех вирусов птичьего происхождения, например, подтипов H5N1, H7N9, H9N2 и H10N8. Вирус HPAI H5N1, VN/04 клады 1 первоначально выделили у смертельно больного пациента (Maines et al., 2005). Вирус H7N9, SH/13, выделен у госпитализированного пациента с неизлечимым заболеванием (Gao et al., 2013). Вирус H9N2, HK/09 клады G1, первоначально выделен из носоглоточного аспирата взрослой пациентки (Cheng, 2010). Вирус гриппа JX/13 (H10N8) первоначально был выделен у пожилого пациента в Китае, умершего в результате инфекции (Garcia-Sastre and Schmolke, 2014; To et al., 2014).
Для получения четырехподтипных VLP получали плазмиду вектора переноса rBV в порядке, описанном в примере 2, для коэкспрессии генов HA из генов гриппа VN/05, SH/13, HK/09 и JX/13, генов NA из IN/05 и гена Bgag. Гены H5, H7, H9 и H10 кодируют полипептиды длиной 568 aa, 560 aa, 560 aa и 561 aa с предсказанными средними молекулярными массами 64,5 кДа, 62,1 кДа, 62,9 кДа и 62,3 кДа соответственно. Длина NA и Bgag составляет 449 аа и 476 аа, предсказанные молекулярные массы составляют 49,1 кДа и 53,5 кДа соответственно.
На Фиг. 13 показано примерное схематическое представление конструкции rBV, экспрессирующей четырехподтипные VLP Bgag гриппа H5/7/9/10 в клетках Sf9. Гены белка внутреннего ядра Bgag, NA и HA, кодон-оптимизированы для осуществления высокоуровневой экспрессии в клетках насекомых и клонируются в rBV совместно, как показано. Каждая конструкция содержит набор промоторов полиэдрина, показанных стрелками. В приведенных примерах ген H10 получен из вируса JX/13, ген HA 9 («H9») получен из вируса A/Гонконг/33982/2009(H9N2) («HK09»), ген HA 5 («H5») получен из вируса A(H5N1)A/Вьетнам/1203/2004(H5N1) («VN/04»), ген HA 7 («H7») получен из вируса A/Шанхай/2/2013(H7N9) («SH/13»), ген NA получен из вируса IN/5. Все гены HA показаны в виде черных квадратов, а ген NA показан в виде серых квадратов.
Для получения VLP клетки Sf9 инфицируют rBV с МЗ 3 для обеспечения экспрессии генов H5, H7, H9, H10, НА и gag, и VLP собирают из супернатанта клеточной культуры в день 3 после инфицирования. Питательную среду инфицированных клеток Sf9 осветляют центрифугированием и фильтрованием через мембрану с размером пор 0,2 мкм. Затем VLP очищают ионообменной хроматографией.
Морфология VLP гриппа, экспрессия и функциональные профили белков HA и NA гриппа показаны на Фиг. 10-11, 14-16 с помощью способа, по существу описанного в примере 4.
VLP гриппа, созданные из конструкции, показанной на Фиг. 13, содержат функциональные белки HA, как показано в анализе гемагглютинации (см. Фиг. 14). Функциональная способность VLP H5/H7/H9/H10 агглютинировать эритроциты индейки подтверждена анализом гемагглютинации. В этом анализе разбавления VLP смешивали с эритроцитами индейки, рекомендованными ВОЗ в качестве реагента для диагностики гриппа животных и эпиднадзора за гриппом животных (ВОЗ, 2012b). В частности, эритроциты индейки серийно разводили с интервалом в 2 раза, начиная с разведения 1:210. VLP Bgag H5/7/9/10 гриппа имели титр 1:8192 или 1:213 на 50 мкл 3,5 мг/мл общего содержания белка. Результаты этого анализа гемагглютинации показывают, что VLP, созданные из конструкций, показанных на Фиг. 13, экспрессируют функциональный HA, этот HA локализуется на мембране VLP, и эти HA представлены на поверхности VLP.
Присутствие Bgag и белков HA подтверждено электрофорезом в полиакриламидном геле в присутствии додецилсульфата натрия и вестерн-блота с использованием антител, специфичных к каждому HA, как показано на Фиг. 10-11, дорожка 2. При помощи вестерн-блота NA не обнаружен, что указывает на низкий уровень перекрестной реактивности с антисыворотками, используемыми в вестерн-блоте (см. Фиг. 10).
На Фиг. 11 показано, что VLP гриппа, созданные из конструкции H10, показанной на Фиг. 8, и четырехподтипные VLP H5/7/9/10, показанные на Фиг. 13, экспрессируют HA и белок Bgag внутреннего ядра. VLP из каждой из двух конструкций загружали в SDS-PAGE и окрашивали кумасси для оценки профиля белка VLP. В крайнем правом ряду обозначены маркеры белка в килодальтонах. Локализации HA указаны стрелкой, а локализации Bgag указаны звездочкой. Эти HA в VLP представляют нерасщепленные полипептиды HA0 длиной приблизительно 62-64 кДа. Полосу длиной приблизительно 55 кДа, соответствующую белку Bgag, обнаруживают с помощью SDS-PAGE, как показано на Фиг. 11, дорожка 2.
На Фиг. 15 показано, что VLP гриппа, созданные из конструкции H10, показанной на Фиг. 8, и четырехподтипная конструкция H5/7/9/10, показанная на Фиг. 13, содержат функциональную NA. VLP из каждой из этих конструкций серийно разводят и оценивают ферментативную активность NA с использованием флуоресцентного анализа NA гриппа. Конструкции VLP показаны следующим образом: VLP H10 показаны залитыми квадратами, а четырехподтипные VLP H5/7/9/10 показаны залитыми треугольниками. Отрицательный контроль в форме PBS показан в виде залитых кругов. Результаты этого флуоресцентного анализа показывают, что обе конструкции экспрессируют функциональную NA. Присутствие функциональной NA в VLP четырех подтипов подтверждено на Фиг. 15.
В совокупности эти результаты позволили предположить, что все белки HA, белки NA и Bgag присутствуют в четырехподтипных VLP, продуцируемых клетками Sf9, и совместно очищены ультрацентрифугированием или хроматографией.
Четырехподтипные VLP H5/7/9/10, созданные из конструкции, показанной на Фиг. 13, имеют морфологию, аналогичную морфологии вируса гриппа, при рассмотрении под просвечивающим электронным микроскопом при использовании способа, по существу описанного в примере 4 (см. Фиг. 16). Четырехподтипные VLP H5/H7/H9/H10 идентифицированы как в целом сферические, похожие на грипп, плеоморфные, оболочечные частицы с диаметром приблизительно 150-200 нм и содержащие типичные шипы HA гриппа, выступающие из оболочки VLP, как показано на Фиг. 16.
Пример 12
Получение других четырехподтипных и множественноподтипных VLP Bgag
Получение и оценку других четырехподтипных и множественноподтипных VLP Bgag можно выполнить с использованием способов и принципов, по существу описанных в примерах 1-11. Например, могут быть получены множественноподтипных VLP Bgag, содержащие H1, H2, H3 и другие подтипы.
Кроме того, с использованием способов и принципов, по существу описанных в примерах 1-11, также могут быть получены и оценены множественноподтипные VLP Bgag, содержащие HA гриппа B.
Кроме того, с использованием способов и принципов, по существу описанных в примерах 1-11, могут быть получены и оценены многовариантные VLP Bgag, содержащие целевые варианты белка того же самого подтипа вируса. Например, различные варианты (клады) вируса H5N1 могут быть совместно встроены в VLP Bgag. Например, нами получены многовариантные VLP Bgag, в которых три варианта HA H5N1, применимые для человеческой вакцины, были ко-экспрессированы в VLP. HA H5N1 HA, полученные нами, например, могут быть получены из вирусов HPAI H5N1 A/Вьетнам/ 203/2004, A/Египет/ 3300-NAMRU3/2008 и A/Хубэй/1/2010, все они рекомендованы ВОЗ для разработки вакцины против H5N1. Например, мы также получили многовариантные VLP Bgag, в которых три варианта HA H5N1, применимые в ветеринарии, были ко-экспрессированы в VLP.
Пример 13
Получение вакцины на основе VLP Bgag H10
VLP Bgag H10 получали для вакцинации с использованием H10 из вируса H10N8 птичьего происхождения JX/13, как по существу описано в примере 6. Вирус гриппа JX/13 вызвал фатальную человеческую инфекцию у пожилого пациента в Китае в 2013 году и был идентифицирован как патоген, способный вызвать пандемию. Вектор rBV сконфигурирован таким образом, чтобы ко-экспрессировать три гена: H10, NA и Bgag, по существу, как описано в примерах 1-5. Полученный rBV использовали для инфицирования клеток Sf9 для получения одноподтипных VLP Bgag H10, показанных, например, на Фиг. 28, панель А.
VLP Bgag, полученные из клеток Sf9, затем очищали, как описано, например, в примере 3. Фильтрат, содержащий VLP Bgag, мигрирует через мембрану и становится фракцией фильтрата, при этом частицы вектора rBV отделялись мембраной. Это подтверждено анионообменной хроматографией. Например, анионообменный хроматограф показывает, что VLP Bgag отделены от rBV и присутствуют во фракции фильтрата (см. Фиг. 28, панель A). Перед вакцинацией VLP Bgag могут быть введены в PBS.
В соответствии с предыдущими наблюдениями HA в VLP H10 Bgag экспрессируется как HA0, длиной приблизительно 60-64 кДа и без детектируемой переработки HA1 и HA2 (см. Фиг. 28, панель A). Также обнаружены заметные полосы частиц длиной приблизительно 55 кДа, что соответствует ожидаемым размерам NA и Bgag. Функциональная активность фермента NA по существу подтверждена, как показано в примере 4. Аналогично другим VLP Bgag, VLP Bgag H10 выглядят как оболочечные плеоморфные частицы диаметром приблизительно 100-180 нм под просвечивающим электронным микроскопом (см. Фиг. 28, панель А).
Три партии VLP Bgag H10 получали и описывали, как показано в таблице 1. Все партии VLP Bgag H10 демонстрировали гемагглюцинационную активность с титром 8192:65536 на 50 мкл VLP Bgag с концентрацией приблизительно 3,5 мг/мл общего содержания белка. Условия изготовления или испытаний оказывали влияние на вариации между различными партиями.
Таблица 1. Получение VLP Bgag PR8 H10
* Определено с использованием флуорометра Qubit 2.0 (Thermo).
** Определено SDS-PAGE и денситометрией.
*** Определено анализом HA с использованием эритроцитов индейки.
Пример 14
Получение вакцины четырехподтипных VLP Bgag H5/7/9/10
Четырехподтипные VLP Bgag H5/7/9/10 также получали и очищали для вакцинации в порядке, по существу описанном в примере 11. Перед вакцинацией VLP Bgag могут быть введены в PBS.
Экспрессию генов H5, H7, H9 и H10 измеряли косвенным иммунофлуоресцентным анализом. Например, при использовании антител, специфичных к подтипу H5, H7, H9 и H10, косвенный иммунофлуоресцентный анализ показывает, что присутствует экспрессия каждого подтипа (см. Фиг. 29).
Четырехподтипные VLP Bgag H5/7/9/10 очищали и анализировали анионообменной хроматографией (см. Фиг. 28, панель B). На анионообменной хроматограмме видно, что четырехподтипные VLP Bgag H5/7/9/10 отделены от rBV и присутствуют во фракции фильтрата.
Аналогично VLP Bgag H10, HA в четырехподтипных VLP Bgag H5/7/9/10 представляют собой нерасщепленные HA0 с размером приблизительно от 62 до 64 кДа и соответствуют размерам 64,5 кДа, 62,1 кДа, 62,9 кДа и 62,3 кДа, предсказанным генам H5, H7, H9 и H10, соответственно (см. Фиг. 28, панель B). Как показано на Фиг. 28, панель B, заметная полоса на 55 кДа соответствовала ожидаемой молекулярной массе NA и Bgag. Функциональная активность фермента NA по существу подтверждена, как показано в примере 4. Аналогично другим VLP Bgag, четырехподтипные VLP Bgag H5/7/9/10 выглядят как оболочечные плеоморфные частицы с диаметром приблизительно 150-200 нм под просвечивающим электронным микроскопом (см. Фиг. 28, панель В). Ко-локализация подтипов и количественное определение каждого подтипа в четырехподтипных VLP H5/H7/H9/H10 также подтверждено, как описано в примере 4.
Как показано в таблице 2, получали и описывали три партии четырехподтипных VLP Bgag H5/7/9/10. Все партии четырехподтипных VLP Bgag H5/7/9/10 имеют гемагглютинационную активность, измеряемую функциональным анализом гемагглютинации, по существу описанным в примере 4.
Таблица 2. Получение четырехподтипных VLP Bgag H5/7/9/10
* Определено с использованием флуорометра Qubit 2.0 (Thermo).
** Определено SDS-PAGE и денситометрией.
*** Определено анализа HA с использованием эритроцитов индейки.
Пример 15
Косвенный иммунофлуоресцентный анализ
Могут быть выполнены косвенные иммунофлуоресцентные анализы. Например, аликвоты 0,3 мл клеток Sf9, инфицированных rBV, высевали в восьмилуночные предметные стекла Nunc LabTek. Через 72 часа инкубации при +28°C клетки Sf9 фиксировали холодным ацетоном и выполняли косвенную иммунофлуоресцентную микроскопию с использованием специфической антисыворотки к H5, H7, H9 и H10. Примерами использованных антисывороток могут быть антисыворотки, использованные для вестерн-блота, по существу как описано в примере 4. Клетки, экспрессирующие H5, H7, H9 и H10, визуализируют с использованием конъюгированного с флуоресцинизотиоцианатом козьего антимышиного IgG (H+L) (KPL, Гейтерсберг, Мэриленд). Гистологическую среду, содержащую пропидиум-иодидный ядерный контрастный краситель, использовали в иммунофлуоресцентном анализе для визуализации клеточных ядер.
Как показано на Фиг. 29, клетки Sf9, инфицированные рекомбинантным бакуловирусом (rBV) с четырехподтипными VLP Bgag H5/7/9/10, продуцируют белки H5, H7, H9, H10, НА и Bgag. Аликвоты инфицированных клеток (верхняя панель) и неинфицированных контролей (нижняя панель) высевали в слайд-камеры и инкубировали в течение 48 часов. Монослои клеток фиксировали холодным ацетоном и зондировали первичными антителами, специфичными к антигенам H5, H7, H9 или H10, как указано. После инкубации монослои зондировали соответствующими видоспецифическими антителами (зелеными) с метками флуоресцеина изотиоцианата (FITC), чтобы визуализировать экспрессированные антигены HA. Клеточные ядра окрашивали в красный цвет с помощью гистологической среды, содержащей иодид пропидия.
Пример 16
Анализ ингибирования гемагглютинации
Может быть выполнен анализ ингибирования гемагглютинации. Например, все сыворотки обрабатывали ферментом, разрушающим рецептор, из холерного вибриона (Denka Seiken, Токио, Япония), затем выполняли анализ ингибирования гемагглютинации с использованием свежеприготовленного 0,5% раствора эритроцитов индейки или 1% раствора эритроцитов лошади с 4 единицами HA каждого указанного антигена.
Гомологичные антигены гриппа, которые используют для определения титров HI, включают полноразмерные гомологичные рекомбинантные антигены, rH5, полученные из A/Вьетнам/1203/2004 (H5N1); rH9 из A/Гонконг/33982/2009 (H9N2); rH7 из A/Аньхой/1/2013 (H7N9), который имеет аминокислотную последовательность, идентичную A/Шанхай/2/2013 (H7N9)); и rH10 из A/Цзянси-Дунху/346/2013 (H10N8). Данные эталонные антигены экспрессировали с использованием экспрессирующей бакуловирусной векторной системы (BEVS) и выделяли и очищали из клеток Sf9 по стандартному способу.
Гетерологичные антигены гриппа также использовали для определения титров ингибирования гемагглютинации. Они включают в себя, например, инактивированные бета-пропиолактоном вирусы A/чирок/Египет/12908-NAMRU3/2005 (H10N1); A/Калифорния/07/2009 NYMC X-197A (H1N1); A/ Швейцария/9715293/2013 (H3N2); и вирусы H5N1, инактивированные бета-пропиолактоном A/Гонконг/156/1997 (клада 0), A/Камбоджа/ X012331/2013 (клада 1.1.2), A/Индонезия/05/2005 (клада 2.1.3.2), A/горный гусь/Цинхай/1A/2005:PR8, (клада 2.2) и A/Бангладеш/3233/2011 (клада 2.2.2).
Пример 17
Вакцинация пациентов посредством VLP Bgag
В качестве вакцины VLP Bgag могут быть введены пациентам, например, взрослым самцам-хорькам Фитча (от 4 до 5 месяцев, фермы Triple F, Сейр, Пенсильвания), с использованием стандартных способов. Например, одной группе из четырех животных VLP Bgag H10 (30 мкг общего HA) может быть инокулирована три раза (недели 0, 4 и 22) внутримышечно («в.м.»). Две группы из пяти хорьков вакцинировали четырехподтипными VLP Bgag либо внутримышечно, либо интраназально («и.н.») (120 мкг общего содержания HA, содержащие 30 мкг каждого подтипа HA). Контрольная группа из четырех хорьков получила PBS в виде плацебо. Перед первоначальной вакцинацией серологический отрицательный статус хорьков подтвержден с помощью анализа ингибирования гемагглютинации для циркулирующих в настоящее время вирусов гриппа. У всех хорьков проводили забор крови в недели 4, 8, 12, 16 и 26 для сбора сыворотки для оценки титров специфических антител ингибированием гемагглютинации. Также у животных проводили забор крови после экспериментального заражения вирусом гриппа, таким как вирус А/чирок/Египет/12908-NAMRU3/2005 (H10N1).
Пример 18
Иммуногенность вакцин на основе VLP Bgag, экспрессирующей один или несколько различных целевых белков-патогенов
VLP Bgag, экспрессирующие H10, и VLP Bgag, экспрессирующие H5, H7, H9 и H10, ввели в PBS для вакцинации. Хорькам, вакцинированным VLP Bgag H10, вводили дозу 30 мкг HA. Хорькам, вакцинированным четырехподтипными VLP Bgag H5/7/9/10, вводили дозу 120 мкг HA. Так как H5, H7, H9 и H10 присутствуют в четырехподтипных VLP Bgag H5/7/9/10 в объеме приблизительно 20-30%, доза 120 мкг соответствует приблизительно 30 мкг каждого подтипа. Этот режим введения соответствует режиму вакцинации против H5N1. Например, для вакцины от A/Вьетнам/1203/04 (H5N1) здоровым взрослым ввели 2мя внутримышечными инъекциями 2 дозы по 45 мкг и 90 мкг, вследствие чего было обнаружено потенциально защитное нейтрализующее антитело (Treanor JJ et al., 2006).
Два способа вакцинации, внутримышечно и интраназально, сравнивают для вакцины на основе четырехподтипной VLP Bgag. Режим вакцинации включает в себя три вакцинации (первичная вакцинация и две стимулирующие вакцинации на недели 0, 4 и 22). Измеряли титры ингибирования гемагглютинации по сравнению с контрольными антигенами гомологичных рекомбинантных HA rH5, rH7, rH9 и rH10. В сыворотке вакцинированных плацебо хорьков отсутствуют антитела, нейтрализующие HA, к любому из тестируемых HA (см. Фиг. 30). И наоборот, VLP Bgag H10 вызывали образование антитела, нейтрализующего HA, только против rH10, при этом четырехподтипные VLP Bgag H5/7/9/10 вызывают образование антитела, нейтрализующего HA, к антигенам H5, H7, H9 и H10 (см. Фиг. 30)
После первичной вакцинации VLP Bgag H10 внутримышечно, антитела, нейтрализующие HA, становятся детектируемыми, и их количество увеличивается после первой стимулирующей вакцинации. Затем титры антител, нейтрализующих HA, постепенно уменьшались до момента второй стимулирующей вакцинации. Так как используется относительно небольшое количество хорьков (4-5 на группу), наблюдается значительная вариативность ответов, особенно в образцах при интраназальном введении.
Сыворотка пациентов, которым ввели VLP Bgag H10 внутримышечно, может иметь сопоставимые титры антител, нейтрализующих HA, с пациентами, которым вводили VLP Bgag четырех подтипов (см. Фиг. 30), при этом титры нейтрализующих антител ниже у пациентов, вакцинированных интраназально четырехподтипными VLP Bgag.
В группах, вакцинированных четырехподтипными VLP Bgag H5/7/9/10, самые низкие титры нейтрализующих антител были обнаружены у антител к rH5, в которых по определяемым титрам нейтрализующего антитела необходимы были, по крайней мере, две вакцинации. Это соответствует относительно низкой иммуногенности подтипа H5. И наоборот, титры нейтрализующих антител к rH7, rH9 и rH10 могут быть обнаружены после одной вакцинации в обеих группах с внутримышечным и интраназальным введением, они увеличиваются после стимулирующей вакцинации, а затем постепенно уменьшаются до момента второй стимулирующей вакцинации (см. Фиг. 30).
Как показано на Фиг. 30, обе разновидности: одноподтипной VLP H10 и четырехподтипных VLP H5/H7/H9/H10 - вызывают устойчивый иммунный ответ у хорьков. Хорьков вакцинируют на неделях 0, 4 и 22 (обозначены стрелками). Группы вакцинации: G1 - группа хорьков, инокулированных внутримышечно три раза (недели 0, 4 и 22) VLP H10 Bgag (30 мкг общего HA); G4 - группа, вакцинированная плацебо, которая получает PBS в качестве плацебо; G2 и G3 -две группы хорьков, вакцинированных внутримышечно или интраназально, соответственно, четырехподтипными VLP (120 мкг общего HA). Кровь собирали на неделях 0, 4, 8, 12, 16 и 26 и сыворотку анализировали исследованием ингибирования гемагглютинации с использованием гомологичных контрольных антигенов rH5, rH7, rH9 и rH10. Полоски над данными наблюдений показывают стандартные отклонения. Пунктирная горизонтальная линия означает предел обнаружения.
Для исследования перекрестно-защитных возможностей вакцинаций на основе VLP, сыворотки перед экспериментальным заражением тестировали исследованием ингибирования гемагглютинина с несколькими изолятами гриппа, включая подтипы H1, H3, H5 и H10. Никакого детектируемого перекрестно-защитного нейтрализующего антитела к вирусам H1 и H3 в сыворотке хорьков, вакцинированных либо VLP H10, либо четырехподтипными VLP после либо внутримышечной или интраназальной вакцинации, не обнаружено. Тем не менее, перекрестно-реактивное нейтрализующее антитело обнаружено при тестировании с несколькими дивергирующими изолятами/кладами вирусов H5N1, а также с вирусом H10N1 (см. Фиг. 30).
Как показано на Фиг. 31, сравнивают профиль нейтрализующего антитела к HA перед исследованиями экспериментального заражения. G1-4 соответствуют указанным группам, и, по существу, соответствуют группам на Фиг. 30. Антитела, нейтрализующие HA, показаны на оси Y, используемые антигены HA показаны на оси X. Антигены HA включают в себя: rH5, полученный из A/Вьетнам/1203/2004 (клада 1 H5N1); rH9 из A/Гонконг/33982/2009 (H9N2); rH7 из A/Аньхой/1/2013 (H7N9); и rH10 из A/Цзянси-Дунху/346/2013 (H10N8). Также проанализированы вирусы, инактивированные бета-пропиолактоном, включая H10* A/чирок/Египет/12908-NAMRU3/2005 (H10N1), H1 A/Калифорния/07/2009 NYMC X-197A (H1N1), H3 A/Швейцария/9715293/2013 (H3N2). Также проанализированы несколько вирусов H5N1, включая A/Гонконг/156/1997 (клада 0), A/Камбоджа/X012331/2013 (клада 1.1.2), A/Индонезия/05/2005 (клада 2.1.3.2), A/горный гусь/Цинхай/1A/2005:PR8, (клада 2.2), и A/Бангладеш/3233/2011 (клада 2.2.2). Полоски над данными наблюдений показывают стандартные отклонения. Пунктирная горизонтальная линия означает предел обнаружения.
Пример 19
Защита вакцинированных пациентов при исследованиях экспериментального заражения
Хорькам выполняли обезболивание с помощью внутримышечной инъекции коктейля кетамин-ксилазин-атропин и проводили экспериментальное заражение интраназально на неделе 27 посредством 106 50% инфицирующих доз яиц вируса гриппа A/чирок/Египет/12908-NAMRU3/2005 (H10N1) в общем объеме 1 мл (500 мкл на ноздрю), разведенного в PBS. Экспериментальное заражение вирусом может быть выполнено на неделе 27 - через пять недель после последней вакцинации. После экспериментального заражения H10N1 проводили ежедневный контроль состояния хорьков в течение четырех дней в целях отслеживания изменений массы тела и температуры, а также клинических симптомов болезни. Образцы назальных смывов собирали в день 2 и 4 после экспериментального заражения («p.c.»). На день 4 после экспериментального заражения животных умерщвляли и осуществляли забор носовых раковин, трахеи и легких. Титры вируса в образцах из нижнего и верхнего дыхательных путей определяли в яйцах. Статистическую значимость различий титров вирусов между вакцинированными пациентами и контрольными пациентами, получавшими PBS, определяли двухфакторным статистическим анализом ANOVA.
После вакцинации либо VLP Bgag H10, либо четырехподтипными VLP Bgag хорьков экспериментально заражали живым целевым патогеном, например, вирусом гриппа H10N1. Гетерологичное экспериментальное заражение может быть выполнено, например, с использованием вируса A/чирок/Египет/12908-NAMRU3/2005 (H10N1), так как вирус географически и эволюционно отличается от антигена вируса H10 A/Цзянси/IPB13a/2013 (H10N8), присутствующего в вакцинах VLP Bgag, введенных хорькам. Перекрестно-реактивные титры подтверждены исследованием ингибирования гемагглютинации для вируса H10N1 (см. Фиг. 31). Экспериментальное заражение вирусом H10N1 не вызывало летального или тяжелого заболевания у хорьков до дня 4 после экспериментального заражения, но вакцины защищали вакцинированную группу от вируса H10N1, как продемонстрировано снижением титров вирусов в назальных смывах и в тканях верхнего (носовые раковины и трахеи) и нижнего (легкие) отделов дыхательных путей после экспериментального заражения (см. Фиг. 32). Например, в назальных смывах в день 2 или день 4 после экспериментального заражения максимальный титр обнаружен у хорьков, получивших плацебо-вакцину, при этом минимальные титры наблюдались у хорьков, вакцинированных интраназально четырехподтипными VLP Bgag (см. Фиг. 32, панель A). Хорьки, вакцинированные внутримышечно четырехподтипными VLP Bgag, также показывали значительное уменьшение титров вирусов по сравнению с группой, получившей плацебо (см. Фиг. 32, панель A). Это позволяет предположить потенциальную роль местных иммунных свойств слизистых оболочек в защите с помощью вакцин на основе четырехподтипных VLP Bgag.
Уменьшение титров реплицирующего вируса еще более выражено в носовых раковинах и тканях трахеи и тканях легких (см. Фиг. 32, панель B). В тканях трахеи и легких титры вирусов снижаются ниже обнаруживаемых уровней у всех вакцинированных хорьков, которые получали либо VLP Bgag H10, либо четырехподтипные VLP Bgag H5/7/9/10, вводимых любым способом. Титры вирусов в тканях носовой раковины также значительно снижены у всех вакцинированных животных, что демонстрирует защитную эффективность этих вакцин на основе VLP против гетерологичного экспериментального заражения H10N1.
Как показано на Фиг. 32, панель А, титры реплицирующегося вируса в день 2 и день 4 после экспериментального заражения в назальных смывах показывают, что вакцины на основе четырехподтипной VLP Bgag в значительной степени защищают пациентов при последующих экспериментальных заражениях. Как показано на Фиг. 32, панель B, титры реплицирующегося вируса в день 2 и день 4 после экспериментального заражения в тканях носовой раковины, трахеи и легких показывают, что вакцины на основе VLP Bgag защищают пациентов при последующих экспериментальных заражениях. Статистически значимые различия обозначены звездочками.
Пример 20
Получение VLP Bgag H555 трех клад
Клетки Sf9 содержат в виде суспензионных культур в среде SF900II-SFM, не содержащей сыворотку насекомых (Life Technologies, Карслбад, Калифорния) при +27°C ± 2°C. Для производства VLP Bgag H555 2 литра клеток Sf9 при концентрации 2 × 106 клеток/мл инфицировали множественными инфекциями («МЗ») 3,0 с rBV, экспрессирующим три указанных гена: H5, а также NA и гены gag БВИ. VLP Bgag собирали через 72 часа после инфицирования из надосадочной жидкости питательной среды, стерильно фильтровали с использованием фильтра с размером 0,2 мкм и концентрировали, используя фильтрацию тангенциальным потоком (TFF, отсечение по молекулярной массе 500000) перед очисткой. VLP Bgag очищали ионообменной хроматографией с последующим ультрацентрифугированием. Анионообменную хроматографию использовали для удаления основных примесей, таких как бакуловирус и ДНК клетки-хозяина, с использованием стандартных способов. Затем выполняли ультрацентрифугирование для очистки и концентрации VLP Bgag с использованием стандартных способов. Препараты VLP Bgag исследовали и хранилипри температуре от +2°C до +8°C в буфере PBS до вакцинации. Исследование VLP Bgag, включая SDS-PAGE и вестерн-блот, выполняли в основном, как описано в настоящем документе.
Пример 21
Иммуногенность и эффективность защиты VLP Bgag H555 трех клад у цыплят
Вакцинация и экспериментальные заражения вирусами. Вакцина на основе VLP Bgag H555 выполнена с использованием коммерческого адъюванта (SEPPIC, Montanide 70/30, Ферфилд, Нью-Джерси), содержащего 1536 единиц HA на дозу VLP H555. Так как все три гена H5 экспрессированы из идентичных кассет экспрессии в клетках Sf9, ожидается, что клады H5 присутствуют на аналогичных уровнях в VLP H555. Цыплят, свободных от специфической патогенной микрофлоры (SPF) (n = 30), подкожно вакцинировали 0,2 мл VLP Bgag H555 в возрасте день 1 и 0,5 мл в возрасте день 21. Контрольные птицы могут получить PBS. Птиц произвольно распределяли в группы по 10 и выполняли экспериментальное заражение интраназально (106 50% инфицирующих доз яиц на птицу) с использованием одного из следующих изолятов HPAI в день 35: GyrF H5N8 (клада 2.3.4.4); Ck/WJ H5N1 (клада 2.1.3) или Ck/Египет H5N1 (клада 2.2.1). После экспериментальных заражений состояние птиц ежедневно контролировали на предмет наличия клинических симптомов, сыворотку брали в день 0 и день 14 после экспериментального заражения, и мазки со слизистой оболочки рта брали для измерения пролиферации вируса.
Для определения выделения вируса орофарингеальные и клоачные мазки собирали в стерильной среде с бульоном с сердечно-мозговым экстрактом и хранили в замороженном виде при температуре -70°C. Вирусную РНК экстрагировали с использованием реагента Trizol LS (Invitrogen, Карлсбад, Калифорния) и набора для выделения вирусной РНК MagMAX AI/ND (Ambion, Остин, Техас). Количественный анализ ПЦР с обратной транскрипцией в режиме реального времени (qRRT-PCR) выполняли с использованием стандартных способов. Кратко: qRRT-PCR, целью которой является ген гриппа M, проводили с использованием одноэтаптого набора ПЦР в режиме реального времени (Ambion) AgPath-ID и системы ПЦР в режиме реального времени ABI 7500 Fast (Applied Biosystems, Карлсбад, Калифорния). Для количественного определения вируса строили стандартную кривую с вирусной РНК, экстрагированной из оттитрованного вируса экспериментального заражения: GyrF H5N8, Ck/WJ H5N1 или Ck/Египет H5N1. Результаты представлены в виде эквивалентов 50% инфицирующей доз яиц/мл, нижний предел обнаружения составляет 100,9 50% инфицирующих доз яиц/мл для образцов от цыплят.
Статистический анализ. Кривые выживания Каплана-Мейера генерировали с помощью Prism 5 (GraphPad Co., Сан-Диего, Калифорния). Логранговый критерий Мантель-Кокса использовали для сравнения кривых выживаемости между экспериментальными группами (Prism 5). Статистические различия в средней квадратической и стандартной ошибках между титрами ингибирования гемагглютинина анализировали с использованием дисперсионного анализа (Prism 5). Строчные буквы обозначают статистическую значимость между сравниваемыми группами. Т-критерий Стьюдента используют для попарного сравнения титров вируса из мазков со слизистой оболочки рта и клоаки (Prism 5). Все статистические испытания выполняли с использованием р < 0,05.
После вакцинации с VLP Bgag и до экспериментального заражения уровни антител к вакцинному антигену определяли анализом ингибирования гемагглютинина. Как показано на Фиг. 35, все группы вакцинированных птиц показывали высокие титры антител, нейтрализующих NA (> 9 log2), через две недели после второго применения вакцины VLP Bgag, когда в анализе использовали гомологичный вакцинный антиген.
Получивших плацебо и вакцинированных птиц экспериментально заражали одним из трех вирусов HPAI через две недели после вакцинации. Титры антител, нейтрализующих HA, до и после экспериментального заражения к вирусам, использованным для экспериментального заражения, показаны на Фиг. 36 и 38, соответственно. Перед экспериментальным заражением выявлены высокие титры антител, нейтрализующих HA. Сопоставимые титры антител, нейтрализующих HA, к каждому штамму вируса показывают наличие трех клад H5 в VLP Bgag. Титры антител, нейтрализующих HA, увеличились после экспериментального заражения (см. Фиг. 38).
Вакцинальную защиту измеряют выживаемостью и уменьшением выделения вируса у цыплят после экспериментального заражения тремя вирусами H5 (см. Фиг. 36 и 39-41). Все птицы в группах вакцин 1, 2 или 3, соответствующих экспериментальному заражению посредством GyrF (H5N8), Ck/Египет (H5N1) и Ck/WJ (H5N1), соответственно, выжили после экспериментального заражения (см. Фиг. 36). Никаких клинических симптомов заболевания не наблюдалось у какой-либо из птиц, подвергшихся экспериментальному заражению после вакцинации. И наоборот, все птицы, вакцинированные плацебо, умерли после экспериментального заражения со средним временем смерти 4, 3 и 3 дня в группах экспериментального заражения посредством GyrF, Ck/Египет, и Ck/WJ, соответственно. Таким образом, VLP Bgag обеспечивает полную защиту от каждого из трех изолятов гриппа, используемых для экспериментального заражения, включая вирус H5N8. Все птицы, вакцинированные плацебо, умерли на 5 дней после экспериментального заражения, что означает значительный уровень защиты у птиц, вакцинированных VLP Bgag.
Выделение вируса значительно уменьшается по сравнению с тремя испытанными изолятами H5 (см. Фиг. 39-41). После экспериментального заражения вирусом с 106 50% инфицирующих доз яиц или PFU в каждом эксперименте по экспериментальному заражению все три вируса H5N1 эффективно реплицируются в верхней части дыхательных путей неиммунизированных контрольных цыплят. Для сравнения, иммунизированные птицы демонстрируют значительное снижение нагрузки вируса H5N1 в каждый день после экспериментального заражения. Аналогичные данные наблюдались в клоачных мазках (см. Фиг. 39-41).
Соответственно, иммунизация с помощью VLP Bgag H555 с тремя кладами обеспечивает значительную защиту от летального гриппа после экспериментального заражения двумя различными изолятами вируса H5N1 и вируса H5N8 HPAI.
Список литературы
Следующие публикации, ссылки, патенты и патентные заявки настоящим полностью включены посредством ссылки.
Belser, J.A., Blixt, O., Chen, L.M., Pappas, C., Maines, T.R., Van Hoeven, N., Donis, R., Busch, J., McBride, R., Paulson, J.C., Katz, J.M., Tumpey, T.M., 2008. Современные вирусы H7 гриппа Северной Америки обладают специфичностью к рецепторам человека: последствия для переносимости вирусов. Труды Национальной академии наук Соединенных Штатов Америки 105, 7558-7563.
Blanco, J.C., Pletneva, L.M., Wan, H., Araya, Y., Angel, M., Oue, R.O., Sutton, T.C., Perez, D.R., 2013. Характеристика и восприимчивость рецепторов хлопковых хомяков к штаммам вирусов птичьего гриппа и гриппа, вызвавшего пандемию в 2009 года. Journal of virology 87, 2036-2045.
Boulay, F., Doms, R.W., Webster, R.G., Helenius, A., 1988. Посттрансляционная олигомеризация и совместная активация кислоты смешанными тримерами гемагглютинина гриппа. The Journal of cell biology 106, 629-639.
Bright, R.A., Carter, D.M., Daniluk, S., Toapanta, F.R., Ahmad, A., Gavrilov, V., Massare, M., Pushko, P., Mytle, N., Rowe, T., Smith, G., Ross, T.M., 2007. Частицы, подобные вирусам гриппа, вызывают более широкие иммунные ответы, чем цельно-вирионный инактивированный вирус гриппа или рекомбинантный гемагглютинин. Vaccine 25, 3871-3878.
Chao, C.C., 1992. Делеция одной аминокислоты на N-конце гемагглютинина вируса гриппа вызывает неправильное сворачивание и блокирует экзоцитоз молекулы в клетках млекопитающих. The Journal of biological chemistry 267, 2142-2148.
Chen, F., Li, J., Sun, B., Zhang, H., Zhang, R., Yuan, J., Ou, X., Ye, W., Chen, J., Liu, Y., Huang, Y., 2015. Выделение и характеристический анализ нового штамма H7N9 вируса птичьего гриппа A у пациента с симптомами, аналогичными гриппу, в Китае. Int J Infect Dis.
Chen, Z., Baz, M., Lu, J., Paskel, M., Santos, C., Subbarao, K., Jin, H., Matsuoka, Y., 2014. Разработка высокопродуктивной живой аттенуированной вакцины против вируса гриппа H7N9, которая обеспечивает защиту от гомологичных и гетерологичных вирусов H7 дикого типа у хорьков. J. Virol. 88, 7016-7023.
Cheng, P.K.C.a.L., W.L., 2010. Молекулярная характеристика H9N2, выделенного в Гонконге в период с 2008 по 2009 год, Неопубликованные материалы, Гонконг, GenBank № доступа CY055137.
Denis, J., Acosta-Ramirez, E., Zhao, Y., Hamelin, M.E., Koukavica, I., Baz, M., Abed, Y., Savard, C., Pare, C., Lopez Macias, C., Boivin, G., Leclerc, D., 2008. Разработка универсальной вакцины против гриппа A на основе пептида M2e, слитого с вакцинной платформой против вируса папайи-мозаики (PapMV). Vaccine 26, 3395-3403.
Ebrahimi, S.M., Tebianian, M., Aghaiypour, K., Nili, H., Mirjalili, A. Прокариотическая экспрессия и характеристика гена М2 вируса птичьего гриппа A в качестве кандидата на универсальную рекомбинантную вакцину против подтипов гриппа А; в частности, H5N1 и H9N2. Molecular biology reports 37, 2909-2914.
Galarza, J.M., Latham, T., Cupo, A., 2005. Вакцина на основе вирусоподобных частиц (VLP) обеспечила полную защиту от экспериментального заражения летальным вирусом гриппа. Viral Immunol. 18, 244-251.
Gao, R., Cao, B., Hu, Y., Feng, Z., Wang, D., Hu, W., Chen, J., Jie, Z., Qiu, H., Xu, K., Xu, X., Lu, H., Zhu, W., Gao, Z., Xiang, N., Shen, Y., He, Z., Gu, Y., Zhang, Z., Yang, Y., Zhao, X., Zhou, L., Li, X., Zou, S., Zhang, Y., Li, X., Yang, L., Guo, J., Dong, J., Li, Q., Dong, L., Zhu, Y., Bai, T., Wang, S., Hao, P., Yang, W., Zhang, Y., Han, J., Yu, H., Li, D., Gao, G.F., Wu, G., Wang, Y., Yuan, Z., Shu, Y., 2013. Инфицирование человека новым вирусом гриппа A (H7N9) птичьего происхождения. N Engl J Med 368, 1888-1897.
Garcia-Sastre, A., Schmolke, M., 2014. Птичий грипп A H10N8 - вирус на пороге? Lancet 383, 676-677.
Guo, L., Lu, X., Kang, S.M., Chen, C., Compans, R.W., Yao, Q., 2003. Усовершенствование иммунных ответов слизистой оболочки частицами, подобными химерным вирусам гриппа HA вирусу иммунодефицита обезьян. Virology 313, 502-513.
Harris, A., Cardone, G., Winkler, D.C., Heymann, J.B., Brecher, M., White, J.M., Steven, A.C., 2006. Плейоморфность вируса гриппа, исследованная посредством криоэлектронной томографии. Proc Natl Acad Sci U S A 103, 19123-19127.
Haynes, J.R., 2009. Вакцины на основе частиц, подобных вирусу гриппа. Expert Rev Vaccines 8, 435-445.
Haynes, J.R., Dokken, L., Wiley, J.A., Cawthon, A.G., Bigger, J., Harmsen, A.G., Richardson, C., 2009. Вакцины на основе частиц, подобных вирусу псевдотипированного Gag гриппа обеспечивает широкую защиту от экспериментального заражения высокопатогенным птичьим гриппом. Vaccine 27, 530-541.
Kang, S.M., Pushko, P., Bright, R.A., Smith, G., Compans, R.W., 2009. Частицы, подобные вирусу гриппа, как пандемические вакцины. Curr. Top. Microbiol. Immunol. 333, 269-289.
Coffman, R.L., Sher, A., Seder, R.A., 2010. Вакцины-адъюванты: заставить работать врожденный иммунитет. Immunity 33, 492-503.
Kong, H., Zhang, Q., Gu, C., Shi, J., Deng, G., Ma, S., Liu, J., Chen, P., Guan, Y., Jiang, Y., Chen, H., 2015. Живая аттенуированная вакцина предотвращает репликацию и передачу вируса H7N9 у млекопитающих. Sci Rep 5, 11233.
Kushnir, N., Streatfield, S.J., Yusibov, V., 2012. Вирусоподобные частицы как высокоэффективная вакцинная платформа: разнообразие целевых объектов и производственных систем и достижения в клинических исследованиях. Vaccine 31, 58-83.
Liu, Y.V., Massare, M.J., Pearce, M.B., Sun, X., Belser, J.A., Maines, T.R., Creager, H.M., Glenn, G.M., Pushko, P., Smith, G.E., Tumpey, T.M., 2015. Частицы, подобные рекомбинантному вирусу, вызывают защитный иммунитет от инфекции вируса птичьего гриппа A (H7N9) у хорьков. Vaccine.
Luo, T., Berkowitz, R., Kaleko, M., 2012. Векторы на основе бычьего вируса иммунодефицита (БВИ) в: Office, U.P.a.T. (Ed.). Novartis AG, USA.
Maines, T.R., Lu, X.H., Erb, S.M., Edwards, L., Guarner, J., Greer, P.W., Nguyen, D.C., Szretter, K.J., Chen, L.M., Thawatsupha, P., Chittaganpitch, M., Waicharoen, S., Nguyen, D.T., Nguyen, T., Nguyen, H.H., Kim, J.H., Hoang, L.T., Kang, C., Phuong, L.S., Lim, W., Zaki, S., Donis, R.O., Cox, N.J., Katz, J.M., Tumpey, T.M., 2005. Вирусы птичьего гриппа (H5N1), выделенные у людей в Азии в 2004 году, проявляют повышенную вирулентность у млекопитающих.
Morens, D.M., Fauci, A.S., 2012. Новые инфекционные заболевания в 2012 году: 20 лет после доклада института медицины. mBio 3.
O'Neill, E., Donis, R.O., 2009. Генерация и характеристика вирусов кандидатных вакцин для предпандемических вакцин против гриппа. Current topics in microbiology and immunology 333, 83-108.
Palese, P., 2004. Грипп: старые и новые угрозы. Nat. Med. 10, S82-87.
Palese, P., 2006. Улучшение вакцин против гриппа? Emerg Infect Dis 12, 61-65.
Pappas, C., Matsuoka, Y., Swayne, D.E., Donis, R.O., 2007. Разработка и оценка кандидатной вакцины против вируса гриппа H7N2 для готовности к пандемии. Clin Vaccine Immunol 14, 1425-1432.
Perrone, L.A., Ahmad, A., Veguilla, V., Lu, X., Smith, G., Katz, J.M., Pushko, P., Tumpey, T.M., 2009. Интраназальная вакцинация посредством частиц, подобных вируса гриппа 1918 года, защищает мышей и хорьков от экспериментального заражения летальным вирусом гриппа 1918 года и H5N1. J. Virol. 83, 5726-5734.
Pica, N., Palese, P., 2013. К универсальной вакцине против вируса гриппа: перспективы и проблемы. Annual review of medicine 64, 189-202.
Pushko, P., Kort, T., Nathan, M., Pearce, M.B., Smith, G., Tumpey, T.M., 2010. Вакцина на основе частиц, подобных рекомбинантному вирусу H1N1, вызывает защитный иммунитет у хорьков против вируса пандемического гриппа H1N1 2009 года. Vaccine 28, 4771-4776.
Pushko, P., Pearce, M.B., Ahmad, A., Tretyakova, I., Smith, G., Belser, J.A., Tumpey, T.M., 2011. Частицы, подобные вирусу гриппа, могут принимать несколько подтипов гемагглютинина и защищать от нескольких типов и подтипов гриппа. Vaccine 29, 5911-5918.
Pushko, P., Pumpens, P., Grens, E., 2013. Разработка технологии вирусоподобных частиц от небольших высокосимметричных до крупных сложных структур вирусоподобных частиц. Intervirology 56, 141-165.
Pushko, P., Tumpey, T.M., Bu, F., Knell, J., Robinson, R., Smith, G., 2005. Частицы, подобные вирусу гриппа, содержащие белки HA, NA и M1 вируса гриппа H9N2, индуцируют защитные иммунные ответы у мышей BALB/c. Vaccine 23, 5751-5759.
Pushko, P., Tumpey, T.M., Van Hoeven, N., Belser, J.A., Robinson, R., Nathan, M., Smith, G., Wright, D.C., Bright, R.A., 2007. Оценка частиц, подобных вирусам гриппа, и адъюванта Novasome в качестве кандидатной вакцины для птичьего гриппа. Vaccine 25, 4283-4290.
Quan, F.S., Vunnava, A., Compans, R.W., Kang, S.M. Вакцина на основе вирусоподобных частиц защищает от вируса пандемического гриппа H1N1 2009 года у мышей. PloS one 5, e9161.
Rahn, J., Hoffmann, D., Harder, T.C., Beer, M., 2015. Вакцины против вирусов гриппа A у домашней птицы и свиней: состояние и перспективы разработки. Vaccine 33, 2414-2424.
Rasmussen, L., Battles, J.K, Ennis, W.H, Nagashima, K., Gonda, M.A., 1990. Исследование вирусоподобных частиц, произведенных при помощи рекомбинантного бакуловируса, содержащего ген gag вируса, подобного вирусу иммунодефицита крупного рогатого скота. Virology 178, 435-451.
Rao, S.S., Kong, W.P., Wei, C.J., Van Hoeven, N., Gorres, J.P., Nason, M., Andersen, H., Tumpey, T.M., Nabel, G.J., 2010. Сравнительная эффективность вакцинации на основе гемагглютинина, нуклеопротеина и гена белка матрицы 2 против гриппа H5N1 у мышей и хорьков. PloS one 5, e9812.
Ross, T.M., Mahmood, K., Crevar, C.J., Schneider-Ohrum, K., Heaton, P.M., Bright, R.A., 2009. A Трехвалентная вакцина на основе вирусоподобных частиц, вызывает защитные иммунные ответы против штаммов сезонного гриппа у мышей и хорьков. PloS one 4, e6032.
Schotsaert, M., De Filette, M., Fiers, W., Saelens, X., 2009. Универсальные вакцины против гриппа A на основе эктодомена M2: доклинические и клинические исследования. Expert review of vaccines 8, 499-508.
Smith, G.E., Flyer, D.C., Raghunandan, R., Liu, Y., Wei, Z., Wu, Y., Kpamegan, E., Courbron, D., Fries, L.F., 3rd, Glenn, G.M., 2013. Разработка вакцины на основе вирусоподобных частиц гриппа H7N9 (VLP): аутологичная защита A/Аньхой/1/2013 (H7N9) и гетерологичная перекрестная защита A/курица/Халиско/CPA1/2012 (H7N3) у вакцинированных мышей, экспериментально зараженных вирусом H7N9. Vaccine 31, 4305-4313.
Suarez, D.L., 2012. Стратегии вакцинации DIVA против вируса птичьего гриппа. Avian Dis. 56, 836-844.
To, K.K., Tsang, A.K., Chan, J.F., Cheng, V.C., Chen, H., Yuen, K.Y., 2014. Возникновение в Китае заболевания человека вследствие птичьего гриппа A (H10N8) - повод для беспокойства? J. Infect. 68, 205-215.
Treanor JJ, Campbell JD, Zangwill KM, Rowe T, Wolff M. Безопасность и иммуногенность инактивированной субвирионной вакцины против гриппа A (H5N1). N Engl J Med. 2006 Mar 30;354(13):1343-51.
Tretyakova, I., Pearce, M.B., Florese, R., Tumpey, T.M., Pushko, P., 2013. Интраназальная вакцинация гемагглютининами H5, H7 и H9, ко-локализованными в вирусоподобной частице, защищает хорьков от нескольких вирусов птичьего гриппа. Virology 442, 67-73.
Wang, B.Z., Liu, W., Kang, S.M., Alam, M., Huang, C., Ye, L., Sun, Y., Li, Y., Kothe, D.L., Pushko, P., Dokland, T., Haynes, B.F., Smith, G., Hahn, B.H., Compans, R.W., 2007. Включение высоких уровней оболочечных гликопротеинов химерных вирусов иммунодефицита человека в вирусоподобные частицы.
Wang, B.Z., Xu, R., Quan, F.S., Kang, S.M., Wang, L., Compans, R.W., 2010. Интраназальная иммунизация с VLP гриппа, включающих в себя флагеллин, закрепленный на мембране, вызывает сильную гетеросубтипическую защиту. PLoS One 5, e13972.
Wang, T.T., Palese, P., 2009. Универсальные эпитопы гемагглютининов вируса гриппа? Nature structural & molecular biology 16, 233-234.
Wei, C.J., Boyington, J.C., McTamney, P.M., Kong, W.P., Pearce, M.B., Xu, L., Andersen, H., Rao, S., Tumpey, T.M., Yang, Z.Y., Nabel, G.J., 2010. Стимулирование образования широко нейтрализующих антител к гриппу H1N1 вакцинацией. Science (New York, N.Y 329, 1060-1064.
ВОЗ, 2012a. Антигенная и генетическая характеристика вирусов гриппа A (H5N1) и гриппа A (H9N2) и вирусов кандидатной вакцины, разработанных для потенциального использования в вакцинах для человека.
ВОЗ, 2012b. Руководство ВОЗ по диагностике и эпиднадзору за гриппом животных, WHO/CDS/CSR/NCS/2002.5 Ред. 1.
ВОЗ, 2013. Антигенная и генетическая характеристика вирусов гриппа A(H5N1), A(H7N3), A(H9N2) вариантных вирусов гриппа и вирусов кандидатной вакцины, разработанных для потенциального использования в вакцинах для человека.
Wohlbold, T.J., Hirsh, A., Krammer, F., 2015. Вакцинный штамм вируса гриппа H10N8 и модель экспериментального заражения мышей, основанная на изоляте человеческого гриппа A/Цзянси-Донгху/ 346/13. Vaccine 33, 1102-1106.
Yen, H.L., Webster, R.G., 2009. Пандемический грипп как современная угроза. Current topics in microbiology and immunology 333, 3-24.
Реферат
Изобретение относится к биотехнологии, а именно к вакцине для стимулирования иммунного ответа на целевые патогены, содержащей вирусоподобную частицу (VLP) на основе протеина группового антигена вируса бычьего иммунодефицита (Bgag), где VLP Bgag содержит один или несколько различных целевых белков-патогенов. Получена плазмида вектора переноса для получения вакцины, которая содержит ген gag вируса бычьего иммунодефицита ("ген Bgag"), один или несколько генов, кодирующих один или несколько различных целевых белков-патогенов, и один или несколько промоторов, причем указанные один или несколько генов, кодирующих целевые белки-патогены, содержат ген, кодирующий трансмембранный белок. Заявлен способ предотвращения или замедления вызванного целевым патогеном заболевания у животного путем введения эффективного количества вакцины. Заявлен способ получения вакцины для стимулирования иммунного ответа на целевые патогены, включающий в себя следующие этапы: клонирование в плазмиду вектора переноса указанного гена Bgag и указанного гена, кодирующего целевой белок-патоген, причем каждый ген контролируется промотором; приготовление вируса-носителя с использованием указанной плазмиды вектора переноса; инфицирование эукариотической клетки указанным вирусом-носителем для создания VLP Bgag; смешивание эффективного количества полученной VLP Bgag с одним или несколькими адъювантами, пригодными для введения вакцины. Заявлен способ отличения пациентов, которым была ранее введена вакцина. Способ включает этапы измерения, присутствует ли ген Bgag или протеин Bgag у пациента, и соотнесения предшествующей вакцинации с присутствием гена Bgag или протеина Bgag. Изобретение позволяет получить вакцину, содержащую рекомбинантные вирусоподобные частицы (VLP) протеина группового антигена вируса бычьего иммунодефицита белка («Bgag»), содержащие один или несколько различных типов целевых белков-патогенов. 5 н. и 37 з.п. ф-лы, 2 табл., 41 ил., 21 пр.
Формула
Документы, цитированные в отчёте о поиске
Способы получения и применения мультипотентных клеточных популяций
Комментарии