Препараты, осуществляющие рнк-интерференцию, для модуляции гена gst-pi - RU2719185C2
Код документа: RU2719185C2
Чертежи












Описание
ОБЛАСТЬ ТЕХНИКИ
[0001] Настоящее изобретение относится к области биофармацевтических и терапевтических препаратов, составленных из молекул на основе нуклеиновых кислот. Более конкретно, настоящее изобретение относится к соединениям и композициям, осуществляющим РНК-интерференцию (RNAi), для модуляции экспрессии GST-π человека, и их применению.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Эта заявка включает список последовательностей представленных в электронном виде в качестве файла ASCII, созданного 20 декабря 2015 года, названного ND5123385WO_SL.txt, размер которого составляет 100000 байтов, и является, таким образом, полностью включенным в качестве ссылки.
УРОВЕНЬ ТЕХНИКИ, ПРЕДШЕСТВУЮЩИЙ ИЗОБРЕТЕНИЮ
[0003] Было обнаружено, что различные раковые ткани человека коррелируют с наличием мутированного гена KRAS. В некоторых случаях, в тканях также присутствует повышенный уровень экспрессии глутатион S-трансферазы Pi (GST-π). (Miyanishi et al., Gastroenterology, 2001, Vol. 121:865-874, реферат) Например, повышенные уровни GST-π в сыворотке наблюдали у пациентов с различными злокачественными новообразованиями желудочно-кишечного тракта. (Niitsu et al., Cancer, 1989, Vol.63, No. 2, pp. 317-323, реферат)
[0004] GST-π является членом семейства ферментов GST, которые играют роль в детоксикации посредством катализирования конъюгации гидрофобных и электрофильных соединений с восстановленным глутатионом. Экспрессия GST-π может быть понижена in vitro с применением siRNA. (Niitsu et al., US 2014/0315975 A1). Однако, есть много недостатков существующих siRNA препаратов, такие как недостаточная активность, побочные эффекты, недостаточная стабильность в сыворотке и недостаток активности или эффективности in vivo.
[0005] Существует неотложная необходимость в композициях и способах для модуляции экспрессии генов, связанных с раком. Конкретно, для терапевтических препаратов, основанных на ингибировании экспрессии GST-π, требуются высоко активные и стабильные siRNA последовательности и структуры, которые могут уменьшить побочные эффекты.
[0006] Что действительно необходимо, это siRNA последовательности, соединения и структуры для модуляции экспрессии GST-π, с применением для лечения таких заболеваний, как злокачественные опухоли.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0007] Настоящее изобретение относится к соединениям, композициям и способам для модуляции экспрессии GST-π человека с применением РНК-интерференции.
[0008] В некоторых вариантах осуществления настоящее изобретение относится к молекулам для сайленсинга гена GST-π на основе РНК-интерференции.
[0009] В дополнительных вариантах осуществления структуры, молекулы и композиции по настоящему изобретению можно использовать в способах для предотвращения или лечения заболеваний, или облегчения симптомов состояний или расстройств, связанных с GST-π, включая злокачественную опухоль.
[0010] Варианты осуществления по настоящему изобретению включают следующие:
[0011] Молекула нуклеиновой кислоты для ингибирования экспрессии GST-π, содержащая смысловую цепь и антисмысловую цепь, где цепи образуют дуплексную область. Молекулы нуклеиновой кислоты могут быть siRNA молекулами для ингибирования экспрессии GST-π, и могут содержать один или более нуклеотидов, которые являются модифицированными или химически модифицированными.
[0012] В некоторых вариантах осуществления нуклеиновая кислота молекулы siRNA для ингибирования экспрессии GST-π может включать 2′-дезокси нуклеотиды, 2′-O-алкил замещенные нуклеотиды, 2′-дезокси-2′-фтор замещенные нуклеотиды, или любое их сочетание. В определенных вариантах осуществления 2′-дезокси нуклеотиды могут находиться в затравочной области молекулы siRNA. В определенных аспектах молекулы siRNA для ингибирования экспрессии GST-π могут иметь дезоксинуклеотиды в множестве позиций в антисмысловой цепи.
[0013] Молекулы нуклеиновой кислоты по настоящему изобретению могут успешно ингибировать экспрессию GST-π mRNA с IC50 менее, чем 300 пМ. В определенных вариантах осуществления молекулы нуклеиновой кислоты могут ингибировать экспрессию уровней GST-π mRNA, по меньшей мере, на 25% in vivo, посредством однократного введения молекул. В некоторых вариантах осуществления молекулы нуклеиновой кислоты могут иметь сниженную побочную активность сопровождающей цепи, или сниженную, по меньшей мере, в 50 раз или, по меньшей мере, в 100 раз.
[0014] Варианты осуществления по настоящему изобретению дополнительно представляют фармацевтические композиции, содержащие молекулы siRNA и фармацевтически приемлемый носитель. В некоторых вариантах осуществления носитель может быть липидной молекулой или липосомой. Это изобретение включает векторы или клетки, содержащие молекулы нуклеиновой кислоты.
[0015] Также в настоящем изобретении рассмотрены способы лечения заболевания, связанного с экспрессией GST-π, посредством введения индивидууму при необходимости композиции, содержащей siRNA, где заболеванием является злокачественная опухоль, рак, рак, вызванный клетками, экспрессирующими мутированный KRAS, саркома или карцинома.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
[0016] ФИГ. 1: На фиг. 1 показано значительное уменьшение ортотопических раковых опухолей легких in vivo посредством siRNA по настоящему изобретению, направленных на GST-π. GST-π siRNA вводили в липосомальном составе в дозе 2 мг/кг бестимусным голым мышам, имеющим A549 ортотопические раковые опухоли легких. Массу окончательной первичной опухоли измеряли при вскрытии для лечебных групп и контрольных групп, получающих носитель. GST-π siRNA демонстрировала значительную эффективность при ингибировании раковых опухолей легких в шестинедельном исследовании. Как показано на фиг. 1, после 43 дней GST-π siRNA демонстрировала заметное успешное ингибирование опухоли с окончательным средним весом первичной опухоли, значительно сниженным в 2.8 раз по сравнению с контролем.
[0017] ФИГ. 2: На фиг. 2 показана эффективность ингибирования опухоли in vivo для GST-π siRNA. Ксенотрансплантационную модель рака с применением клеток A549 использовали при относительно низкой дозе siRNA в 0.75 мг/кг. GST-π siRNA продемонстрировала успешное ингибирование опухоли в течение нескольких дней. После 36 дней GST-π siRNA продемонстрировала заметно успешное ингибирование опухоли с окончательнымисредними объемами опухолей значительно уменьшенными приблизительно в 2 раза по сравнению с контролем.
[0018] ФИГ. 3: На фиг. 3 показана эффективность ингибирования опухоли in vivo для GST-π siRNA в конечной точке на фиг. 2. GST-π siRNA продемонстрировала успешное ингибирование со средними массами опухолей, уменьшенными более чем в 2 раза.
[0019] ФИГ. 4: На фиг. 4 показано, что GST-π siRNA по настоящему изобретению существенно увеличивает смерть злокачественных клеток посредством апоптоза in vitro. GST-π siRNA вызвала повышение экспрессии PUMA, биомаркера апоптоза, который связан с потерей жизнеспособности клеток. На фиг. 4 экспрессия PUMA была существенно повышена через 2-6 дней после трансфекции GST-π siRNA.
[0020] ФИГ. 5: На фиг. 5 показано, что GST-π siRNA по настоящему изобретению обеспечивала сокрушительную эффективность в отношении A549 ксенотрансплантированных опухолей in vivo. Дозозависимый нокдаун GST-π mRNA наблюдали у самок бестимусных мышей (nu/nu) (Charles River) с применением siRNA, мишенью которой является GST-π. Как показано на фиг. 5, при дозе 4 мг/кг значительное уменьшение приблизительно на 40% GST-π mRNA было детектировано через 24 часа после инъекции.
[0021] ФИГ. 6: На фиг. 6 показано, что GST-π siRNA по настоящему изобретению ингибировал ксенотрансплантированные опухоли рака поджелудочной железы in vivo. GST-π siRNA обеспечивал сайленсинг гена in vivo при введении в липосомальном составе в ксенотрансплантированные опухоли рака поджелудочной железы в самках бестимусных голых мышей, в возрасте от 6 до 8 недель. Как показано на фиг. 6, эффект дозы получали с применением доз в диапазоне от 0.375 мг/кг до 3 мг/кг siRNA, мишенью которых является GST-π. GST-π siRNA продемонстрировал успешное ингибирование опухоли в течение нескольких дней после введения, объем опухоли снижался приблизительно в 2 раза в конечной точке.
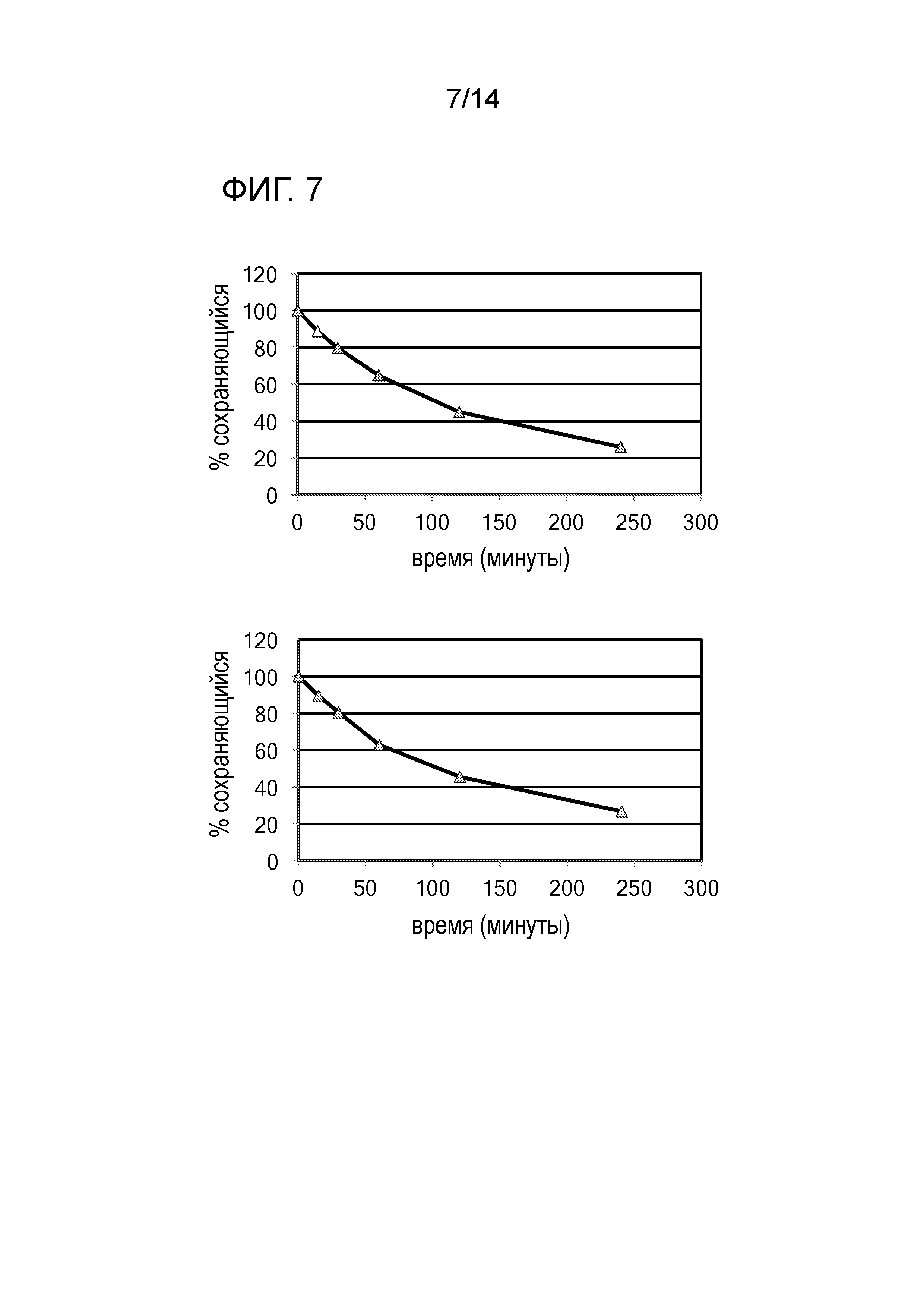
[0022] ФИГ. 7: На фиг. 7 показано, что GST-π siRNA по настоящему изобретению продемонстрировал повышенную стабильность в сыворотке. Как показано на фиг. 7, время полужизни (t½) в сыворотке для обеих смысловой цепи (фиг. 7, сверху) и антисмысловой цепи (фиг. 7, снизу) GST-π siRNA составляло приблизительно 100 минут.
[0023] ФИГ. 8: На фиг. 8 показано, что GST-π siRNA по настоящему изобретению продемонстрировала повышенную стабильность в составе в плазме. На фиг. 8 показана инкубация липосомального состава GST-π siRNA в 50% сыворотки человека в PBS, и детекция оставшейся siRNA в различные моменты времени. Как показано на фиг. 8, время полужизни (t½) в плазме состава GST-π siRNA было значительно дольше, чем 100 часов.
[0024] ФИГ. 9: На фиг. 9 показан in vitro нокдаун для направляющей цепи GST-π siRNA. Как показано на фиг. 9, нокдаун направляющей цепи GST-π siRNA был приблизительно экспоненциальным, по сравнению с контролем с применением последовательности со случайной перестановкой, которая не демонстрировала никакого эффекта.
[0025] ФИГ. 10: На фиг. 10 показан in vitro нокдаун для сопровождающей цепи GST-π siRNA на фиг. 9. Как показано на фиг. 10, нецелевой нокдаун сопровождающей цепи для GST-π siRNA был значительно снижен, без существенного эффекта.
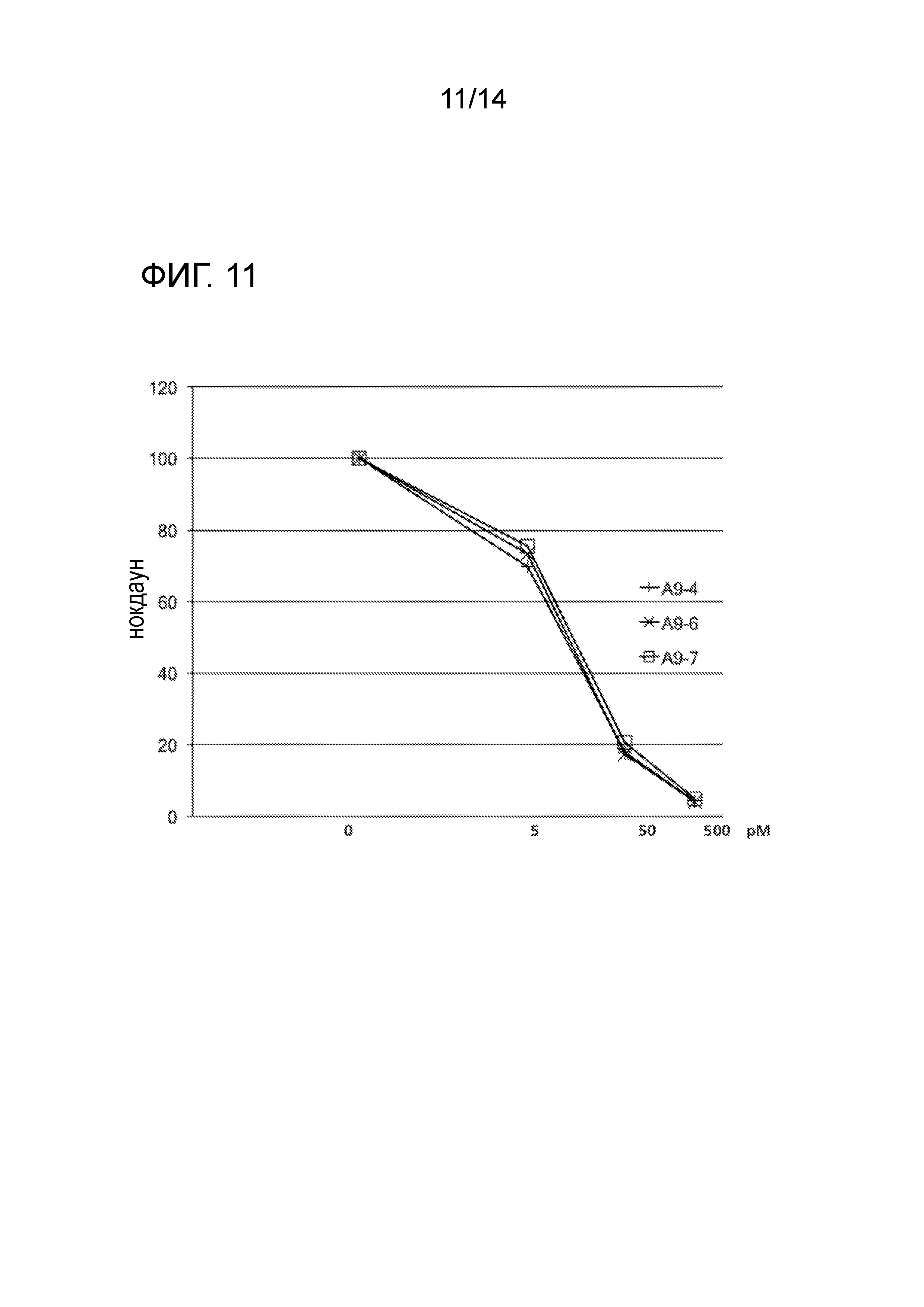
[0026] ФИГ. 11: На фиг. 11 показан in vitro нокдаун для направляющих цепей нескольких высоко активных GST-π siRNAs. Как показано на фиг. 11, активности нокдауна направляющей цепи для GST-π siRNAs были приблизительно экспоненциальными.
[0027] ФИГ. 12: На фиг. 12 показан in vitro нокдаун для сопровождающей цепи для GST-π siRNAs на фиг. 11. Как показано на фиг. 12, активности нецелевого нокдауна сопровождающей цепи для GST-π siRNAs были значительно уменьшены ниже приблизительно 500 пМ.
[0028] ФИГ. 13: На фиг. 13 показан in vitro нокдаун для направляющей цепи высоко активной GST-π siRNA. Как показано на фиг. 13, активность нокдауна направляющей цепи для GST-π siRNA была приблизительно экспоненциальной.
[0029] ФИГ. 14: На фиг. 14 показан in vitro нокдаун для сопровождающей цепи для GST-π siRNA на фиг. 13. Как показано на фиг. 14, активность нецелевого нокдауна сопровождающей цепи для GST-π siRNA была значительно снижена.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0030] Настоящее изобретение относится к соединениям, композициям и способам, касающимся терапевтических препаратов на основе нуклеиновой кислоты для модуляции экспрессии GST-π.
[0031] В некоторых вариантах осуществления настоящее изобретение относится к молекулам, активным в отношении РНК-интерференции, а также структурам и композициям, которые могут подавлять экспрессию GST-π.
[0032] Структуры и композиции по данному изобретению можно использовать при предотвращении или лечении различных заболеваний, таких как злокачественная опухоль.
[0033] В дополнительных вариантах осуществления настоящее изобретение относится к композициям для доставки и захвата одной или более терапевтических RNAi молекул по настоящему изобретению, а также способов их применения. Композиции на основе RNA по настоящему изобретению можно использовать в способах для предотвращения или лечения злокачественных опухолей, таких как раковые заболевания.
[0034] Терапевтические композиции по настоящему изобретению включают молекулы нуклеиновой кислоты, которые являются активными при РНК-интерференции. Терапевтические молекулы нуклеиновой кислоты могут быть направлены на GSTP1 (GST-π) для сайленсинга гена.
[0035] В различных вариантах осуществления настоящее изобретение относится к диапазону молекул, которые могут быть активными в качестве малой интерферирующей РНК (siRNA), и могут регулировать или подавлять экспрессию гена GST-π.
[0036] siRNAs по настоящему изобретению можно использовать для предотвращения или лечения злокачественных опухолей.
[0037] Варианты осуществления по настоящему изобретению дополнительно представляют носитель, состав, или состав липидных наночастиц для доставки siRNAs по изобретению индивидуумам при необходимости предотвращения или лечения злокачественной опухоли. Это изобретение дополнительно рассматривает способы введения siRNAs в качестве терапевтических препаратов млекопитающим.
[0038] Терапевтические молекулы и композиции по настоящему изобретению можно использовать для РНК-интерференции, направленной на предотвращение или лечение заболевания, связанного с GST-π, посредством введения соединения или композиции индивидууму при необходимости.
[0039] В способах по настоящему изобретению могут быть использованы соединения по изобретению для предотвращения или лечения злокачественной опухоли.
[0040] В некоторых аспектах злокачественная опухоль может присутствовать при различных заболеваниях, например, раковые опухоли с высокой экспрессией GST-π, раковые опухоли, вызванные клетками, экспрессирующими мутированный KRAS, саркома, фибросаркома, злокачественная фиброзная гистиоцитома, липосаркома, рабдомиосаркома, лейомиосаркома, ангиосаркома, Саркома Капоши, лимфангиосаркома, синовиальная саркома, хондросаркома, остеосаркома и карциномы.
[0041] В определенных аспектах в способах по настоящему изобретению могут быть использованы соединения по изобретению для предотвращения или лечения злокачественных опухолей и раковых опухолей в любом органе или ткани, включая, например, опухоль головного мозга, рак головы и шеи, рак молочной железы, рак легких, рак пищевода, рак желудка, рак двенадцатиперстной кишки, колоректальный рак, рак печени, рак поджелудочной железы, рак желчного пузыря, рак желчного протока, рак почки, рак уретры, рак мочевого пузыря, рак предстательной железы, рак яичка, рак матки, рак яичника, рак кожи, лейкемия, злокачественная лимфома, эпителиальные злокачественные опухоли, и неэпителиальные злокачественные опухоли.
[0042] В определенных вариантах осуществления комбинацию терапевтических молекул по настоящему изобретению можно использовать для подавления или ингибирования экспрессии гена GST-π.
[0043] Настоящее изобретение относится к диапазону молекул RNAi, где каждая молекула имеет полинуклеотидную смысловую цепь и полинуклеотидную антисмысловую цепь; каждая цепь молекулы состоит из от 15 до 30 нуклеотидов в длину; смежная область из от 15 до 30 нуклеотидов антисмысловой цепи является комплементарной последовательности mRNA, кодирующей GST-π; и, по меньшей мере, часть смысловой цепи является комплементарной, по меньшей мере, части антисмысловой цепи, и молекула составляет от 15 до 30 нуклеотидов в длину.
[0044] Молекула RNAi по настоящему изобретению может иметь смежную область от 15 до 30 нуклеотидов антисмысловой цепи, которая является комплементарной последовательности mRNA, кодирующей GST-π, которая расположена в дуплексной области молекулы.
[0045] В некоторых вариантах осуществления молекула RNAi может иметь смежную область от 15 до 30 нуклеотидов антисмысловой цепи, которая является комплементарной последовательности mRNA, кодирующей GST-π.
[0046] Варианты осуществления по настоящему изобретению могут дополнительно представлять способы для предотвращения, лечения или облегчения одного или более симптомов злокачественной опухоли, или снижения риска развития злокачественной опухоли, или отсрочки начала злокачественной опухоли у млекопитающего при необходимости.
[0047] GST-π и молекулы RNAi
[0048] Последовательность нуклеиновой кислоты примерной мишени mRNA глутатион S-трансферазы pi человека (GST-π человека) описана в GenBank номер доступа NM_000852.3 (hGSTP1), и составляет 986 нуклеотидов в длину.
[0049] Специалист в данной области поймет, что сообщаемая последовательность меняться в течение времени и включать любые необходимые изменения в молекулах нуклеиновой кислоты в данном документе, таким образом.
[0050] Варианты осуществления по настоящему изобретению могут предоставлять композиции и способы для подавления экспрессии гена GST-π с применением малых молекул нуклеиновой кислоты. Примеры молекул нуклеиновой кислоты включают молекулы, активные в отношении РНК-интерференции (RNAi молекулы), малые интерферирующие РНК (siRNA), двухцепочечную РНК (dsRNA), микро-RNA (miRNA), и короткую шпилечную RNA (кшРНК), а также DNA-направленные RNAs (ddRNA), Piwi-взаимодействующие RNAs (piRNA), и связанные с повторами siRNAs (rasiRNA). Такие молекулы способны к опосредованию РНК-интерференции против экспрессии гена GST-π.
[0051] Композиция и способы, описываемые в настоящем документе также можно использовать при лечении различных видов злокачественных опухолей у индивидуума.
[0052] Молекулы нуклеиновой кислоты и способы по настоящему изобретению можно использовать для подавления экспрессии генов, которые кодируют GST-π.
[0053] Композиции и способы по настоящему изобретению могут включать одну или более молекул нуклеиновой кислоты, которая, независимо или в комбинации, может модулировать или регулировать экспрессию белка GST-π и/или генов, кодирующих белки GST-π, белков и/или генов, кодирующих GST-π, связанные с поддержанием и/или развитие заболеваний, состояний или расстройств, связанных с GST-π, таких как злокачественная опухоль.
[0054] Композиции и способы по настоящему изобретению описаны с отсылкой к примерным последовательностям GST-π. Специалист в данной области поймет, что различные аспекты и варианты осуществления по изобретению направлены на любые, связанные с GST-π гены, последовательности, или варианты, такие как гомологичные гены и варианты транскриптов, и полиморфизмы, включая однонуклеотидный полиморфизм (SNP), связанный с любыми генами GST-π.
[0055] В некоторых вариантах осуществления композиции и способы по настоящему изобретению могут предоставить молекулу двухцепочечной малой интерферирующей нуклеиновой кислоты (siRNA), которая подавляет экспрессию гена GST-π, например, GST-π человека.
[0056] Молекула RNAi по настоящему изобретению может быть направлена на GST-π и любые гомологичные последовательности, например, с применением комплементарных последовательностей или посредством включения неканонических пар оснований, например, пар оснований ошибочного спаривания и/или неоднозначных пар оснований, которые могут представлять дополнительные последовательности-мишени.
[0057] В случаях, когда идентифицировано ошибочное спаривание, неканонические пары оснований, например, пары оснований ошибочного спаривания и/или неоднозначные основания можно использовать для создания молекул нуклеиновой кислоты, мишенью которых является больше чем одна генная последовательность.
[0058] Например, неканонические пары оснований, такие как пары оснований UU и CC, можно использовать для создания молекул нуклеиновой кислоты, которые способны к направленному воздействию на последовательности для различающихся GST-π мишеней, которые имеют гомологичную последовательность. Таким образом, молекула RNAi может быть направлена на нуклеотидную последовательность, которая является консервативной между гомологичными генами, и единичную RNAi молекулу можно использовать для ингибирования экспрессии более, чем одного гена.
[0059] В некоторых аспектах композиции и способы по настоящему изобретению включают молекулы RNAi, которые являются активными против GST-π mRNA, где молекула RNAi включает последовательность, комплементарную любой mRNA, кодирующей GST-π последовательность.
[0060] В некоторых вариантах осуществления молекула RNAi по данному изобретению может обладать активностью против GST-π RNA, где молекула RNAi включает последовательность, комплементарную RNA, имеющей вариантную GST-π кодирующую последовательность, например, мутантный GST-π ген, который, как известно в данной области, связан со злокачественной опухолью.
[0061] В дополнительных вариантах осуществления молекула RNAi по настоящему изобретению может включать нуклеотидную последовательность, которая может взаимодействовать с нуклеотидной последовательностью гена GST-π и опосредованно подавлять экспрессию гена GST-π.
[0062] Молекулы нуклеиновой кислоты для ингибирования экспрессии GST-π могут иметь смысловую цепь и антисмысловую цепь, где цепи образуют дуплексную область. Молекулы нуклеиновой кислоты могут иметь один или более нуклеотидов в дуплексной области, являющиеся модифицированными или химически модифицированными, включая такие модификации как известные в данной области. Любой нуклеотид в «липком» конце siRNA может также быть модифицированным или химически модифицированным.
[0063] В некоторых вариантах осуществления предпочтительными модифицированными или химически модифицированными нуклеотидами являются 2′-дезоксинуклеотиды. В дополнительных вариантах осуществления модифицированные или химически модифицированные нуклеотиды могут включать 2′-O-алкил замещенные нуклеотиды, 2′-дезокси-2′-фтор замещенные нуклеотиды, фосфотиоат нуклеотиды, закрытые нуклеотиды, или любое их сочетание.
[0064] В определенных вариантах осуществления предпочтительная структура может иметь антисмысловую цепь, содержащую дезоксинуклеотиды в множестве позиций, множество позиций, являющихся одной из следующих: каждая из позиций 4, 6 и 8, от 5ʹ конца антисмысловой цепи; каждая из позиций 3, 5 и 7, от 5ʹ конца антисмысловой цепи; каждая из позиций 1, 3, 5 и 7, от 5ʹ конца антисмысловой цепи; каждая из позиций 3-8, от 5ʹ конца антисмысловой цепи; и каждая из позиций 5-8, от 5ʹ конца антисмысловой цепи. Любую из этих структур можно комбинировать с одним или более 2′-дезокси-2′-фтор замещенными нуклеотидами в дуплексной области.
[0065] Молекулы нуклеиновой кислоты по настоящему изобретению могут ингибировать экспрессию GST-π mRNA с эффективной IC50 менее чем приблизительно 300 пМ, или менее чем приблизительно 200 пМ, или менее чем приблизительно 100 пМ, или менее чем приблизительно 50 пМ.
[0066] Дополнительно, молекулы нуклеиновой кислоты могут ингибировать уровни экспрессии GST-π mRNA, по меньшей мере, на 25% in vivo, при однократном введении.
[0067] В настоящем изобретении рассмотрены фармацевтические композиции, которые могут содержать одну или более siRNAs, как описано в настоящем документе, в комбинации с фармацевтически приемлемым носителем. Можно использовать любой подходящий носитель, включая носители, известные в данной области, а также липидные молекулы, наночастицы или липосомы, каждая из которых может инкапсулировать молекулы siRNA.
[0068] Настоящее изобретение относится к способам лечения заболевания, связанного с экспрессией GST-π, способы которого включают введение индивидууму при необходимости композиции, содержащей одну или более siRNAs. Заболевания, подвергающиеся лечению, могут включать злокачественную опухоль, рак, рак, вызванный клетками, экспрессирующими мутированный KRAS, саркому, и карциному, наряду с другими.
[0069] Примеры молекул RNAi, по настоящему изобретению направленных на GST-π mRNA, представлены в таблице 1.
Таблица 1: Последовательности молекулы RNAi для GST-π
[0070] Ключ для таблицы 1: заглавные A, G, C и U относятся к рибо-A, рибо-G, рибо-C и рибо-U, соответственно. Строчные буквы a, u, g, c, t относятся к 2ʹ-дезокси-A, 2ʹ-дезокси-U, 2ʹ-дезокси-G, 2ʹ-дезокси-C и дезокситимидину, соответственно.
[0071] Примеры молекул RNAi по настоящему изобретению, направленных на GST-π mRNA, представлены в таблице 2.
Таблица 2: Последовательности молекулы RNAi для GST-π
[0072] Ключ для таблицы 2: Заглавные A, G, C и U относятся к рибо-A, рибо-G, рибо-C и рибо-U, соответственно. Строчные буквы a, u, g, c, t относятся к 2ʹ-дезокси-A, 2ʹ-дезокси-U, 2ʹ-дезокси-G, 2ʹ-дезокси-C, и дезокситимидин (dT=T=t) соответственно. Подчеркивание относится к 2ʹ-OMe-замещенным, например, U. Строчная буква f относится к 2ʹ-дезокси-2ʹ-фтор замещению, например, fU является 2ʹ-дезокси-2ʹ-фтор-U. N является A, C, G, U, U, a, c, g, u, t, или модифицированным, интвертированным или химически модифицированным нуклеотидом. Знак ʺsʺ представляет фосфотиоатную связь.
[0073] Примеры молекул RNAi по настоящему изобретению, направленные на GST-π mRNA, представлены в таблице 3.
Таблица 3: Последовательности молекулы RNAi для GST-π
[0074] Ключ для таблицы 3: Заглавные A, G, C и U относятся к рибо-A, рибо-G, рибо-C и рибо-U, соответственно. Строчные буквы a, u, g, c, t относятся к 2ʹ-дезокси-A, 2ʹ-дезокси-U, 2ʹ-дезокси-G, 2ʹ-дезокси-C и дезокситимидину (dT=T=t), соответственно. Подчеркивание относится к 2ʹ-OMe-замещенному, например, U. Строчная буква f относится к 2ʹ-дезокси-2ʹ-фтор замещению, например, fU является 2ʹ-дезокси-2ʹ-фтор-U. N является A, C, G, U, U, a, c, g, u, t, или модифицированным, интвертированным или химически модифицированным нуклеотидом.
[0075] Примеры молекул RNAi по настоящему изобретению, направленных на GST-π mRNA, представлены в таблице 4.
Таблица 4: Последовательности молекул RNAi для GST-π
[0076] Ключ для таблицы 4: Заглавные A, G, C и U относятся к рибо-A, рибо-G, рибо-C и рибо-U, соответственно. Строчные буквы a, u, g, c, t относятся к 2ʹ-дезокси-A, 2ʹ-дезокси-U, 2ʹ-дезокси-G, 2ʹ-дезокси-C и дезокситимидину (dT=T=t), соответственно. Подчеркивание относится к 2ʹ-OMe-замещенному, например, U. Строчная буква f относится к 2ʹ-дезокси-2ʹ-фтор замещению, например, fU является 2ʹ-дезокси-2ʹ-фтор-U. N является A, C, G, U, U, a, c, g, u, t, или модифицированным, интвертированным или химически модифицированным нуклеотидом.
[0077] Примеры молекул RNAi, по настоящему изобретению направленных на GST-π mRNA, представлены в таблице 5.
Таблица 5: Последовательности молекул RNAi для GST-π
[0078] Ключ для таблицы 5: Заглавные A, G, C и U относятся к рибо-A, рибо-G, рибо-C и рибо-U, соответственно. Строчные буквы a, u, g, c, t относятся к 2ʹ-дезокси-A, 2ʹ-дезокси-U, 2ʹ-дезокси-G, 2ʹ-дезокси-C и дезокситимидину (dT=T=t), соответственно. Подчеркивание относится к 2ʹ-OMe-замещенному, например, U. Строчная буква f относится к 2ʹ-дезокси-2ʹ-фтор замещению, например, fU является 2ʹ-дезокси-2ʹ-фтор-U. N является A, C, G, U, U, a, c, g, u, t, или модифицированным, интвертированным или химически модифицированным нуклеотидом.
[0079] Примеры молекул RNAi, по настоящему изобретению направленных на GST-π mRNA, представлены в таблице 6.
Таблица 6: Последовательности молекул RNAi для GST-π
[0080] Ключ для таблицы 6: Заглавные A, G, C и U относятся к рибо-A, рибо-G, рибо-C и рибо-U, соответственно. Строчные буквы a, u, g, c, t относятся к 2ʹ-дезокси-A, 2ʹ-дезокси-U, 2ʹ-дезокси-G, 2ʹ-дезокси-C и дезокситимидину (dT=T=t), соответственно. Подчеркивание относится к 2ʹ-OMe-замещенному, например, U. Строчная буква f относится к 2ʹ-дезокси-2ʹ-фтор замещению, например, fU является 2ʹ-дезокси-2ʹ-фтор-U. N является A, C, G, U, U, a, c, g, u, t, или модифицированным, интвертированным или химически модифицированным нуклеотидом.
[0081] В некоторых вариантах осуществления настоящее изобретение относится к диапазону молекул нуклеиновой кислоты, где: a) молекула имеет полинуклеотидную смысловую цепь и полинуклеотидную антисмысловую цепь; b) каждая цепь молекулы составляет от 15 до 30 нуклеотидов в длину; c) смежная область от 15 до 30 нуклеотидов антисмысловой цепи является комплементарной последовательности mRNA, кодирующей GST-π; d) по меньшей мере, часть смысловой цепи является комплементарной, по меньшей мере, части антисмысловой цепи, и молекула имеет дуплексную область от 15 до 30 нуклеотидов в длину.
[0082] В некоторых вариантах осуществления молекула нуклеиновой кислоты может иметь смежную область от 15 до 30 нуклеотидов антисмысловой цепи, которая является комплементарной последовательности mRNA, кодирующей GST-π, расположенную в дуплексной области молекулы.
[0083] В дополнительных вариантах осуществления молекула нуклеиновой кислоты может иметь смежную область от 15 до 30 нуклеотидов антисмысловой цепи, которая является комплементарной последовательности mRNA, кодирующей GST-π.
[0084] В определенных вариантах осуществления каждая цепь молекулы нуклеиновой кислоты может образовывать от 18 до 22 нуклеотидов в длину. Дуплексная область молекулы нуклеиновой кислоты может составлять 19 нуклеотидов в длину.
[0085] В качестве альтернативы, молекула нуклеиновой кислоты может иметь полинуклеотидную смысловую цепь и полинуклеотидную антисмысловую цепь, которые соединены в качестве единой цепи, и образуют дуплексную область, присоединенную на одном конце посредством петли.
[0086] Некоторые варианты осуществления молекулы нуклеиновой кислоты по данному изобретению могут иметь тупой конец. В определенных вариантах осуществления молекула нуклеиновой кислоты может иметь один или более 3ʹ «липких» концов.
[0087] Настоящее изобретение относится к диапазону молекул нуклеиновой кислоты, которые являются молекулами RNAi, активными в отношении сайленсинга гена. Молекулами нуклеиновой кислоты по изобретению могут быть dsRNA, siRNA, микро-RNA, или кшРНК, активные в отношении сайленсинга гена, а также DNA-направленная RNA (ddRNA), Piwi-взаимодействующая RNA (piRNA), или ассоциированная с повторами siRNA (rasiRNA). Молекулы нуклеиновой кислоты могут быть активными в отношении ингибирования экспрессии GST-π.
[0088] Варианты осуществления по настоящему изобретению дополнительно представляют молекулы нуклеиновой кислоты, имеющие IC50 для нокдауна GST-π менее чем 100 пМ.
[0089] Дополнительные варианты осуществления по настоящему изобретению представляют молекулы нуклеиновой кислоты, имеющие IC50 для нокдауна GST-π менее чем 50 пМ.
[0090] Это изобретение дополнительно рассматривает композиции, содержащие одну или более молекул нуклеиновой кислоты по изобретению наряду с фармацевтически приемлемым носителем. В определенных вариантах осуществления носителем может быть липидная молекула или липосома.
[0091] Соединения и композиции по настоящему изобретению пригодны в способах для предотвращения или лечения заболевания, связанного с GST-π, посредством введения соединения или композиции индивидууму при необходимости.
[0092] В способах по настоящему изобретению могут быть использованы соединения по изобретению для предотвращения или лечения злокачественной опухоли. Злокачественная опухоль может присутствовать при различных заболеваниях, например, раковые опухоли, связанные с экспрессией GST-π, раковые опухолями, вызванные клетками экспрессирующими мутированный KRAS, саркомы, фибросаркома, злокачественная фиброзная гистиоцитома, липосаркома, рабдомиосаркома, лейомиосаркома, ангиосаркома, саркома Капоши, лимфангиосаркома, синовиальная саркома, хондросаркома, остеосаркома, карциномы, опухоль головного мозга, рак головы и шеи, рак молочной железы, рак легких, рак пищевода, рак желудка, рак двенадцатиперстной кишки, рак слепой кишки, колоректальный рак, рак прямой кишки, рак печени, рак поджелудочной железы, рак желчного пузыря, рак желчного протока, рак анального отверстия, рак почки, рак уретры, рак мочевого пузыря, рак предстательной железы, рак яичка, рак матки, рак яичника, рак кожи, лейкемия, злокачественная лимфома, эпителиальные злокачественные опухоли и неэпителиальные злокачественные опухоли.
[0093] Модифицированные и химически модифицированные siRNAs
[0094] Варианты осуществления по настоящему изобретению охватывают молекулы siRNA, которые являются модифицированными или химически модифицированными для того, чтобы предоставить улучшенные качества для терапевтического применения, такие как повышенная активность и эффективность сайленсинга гена. Настоящее изобретение относится к модифицированным или химически модифицированным молекулам siRNA, которые могут обладать повышенной стабильностью в сыворотке, а также сниженными нецелевыми эффектами без потери активности и эффективности молекул siRNA в отношении генной модуляции и сайленсинга генов. В некоторых аспектах настоящее изобретение относится к siRNAs, имеющим модификации или химические модификации в различных комбинациях, которые усиливают стабильность и эффективность siRNA.
[0095] В некоторых вариантах осуществления молекулы siRNA по настоящему изобретению могут иметь нецелевую активность сопровождающей цепи, сниженную, по меньшей мере, в 10 раз, или, по меньшей мере, в 20 раз, или, по меньшей мере, в 30 раз, или, по меньшей мере, в 50 раз, или, по меньшей мере, в 100 раз.
[0096] Как применяют в настоящем документе, термины модифицированный и химически модифицированный относятся к изменениям, произведенным в структуре природного нуклеотида или структуре нуклеиновой кислоты siRNA, которая охватывает siRNAs, имеющие один или более аналогов нуклеотидов, измененных нуклеотидов, нестандартных нуклеотидов, неприродных нуклеотидов и их сочетаний.
[0097] В некоторых вариантах осуществления число модифицированных или химически модифицированных структур в siRNA может включать все из структурных компонентов и/или все нуклеотиды молекулы siRNA.
[0098] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие модификацию группы сахара нуклеотида, модификацию нуклеинового основания нуклеотида, модификацию скелета или связи нуклеиновой кислоты, модификацию структуры нуклеотида или нуклеотидов на конце цепи siRNA, и их сочетания.
[0099] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие модификацию заместителя в 2' углероде сахара.
[00100] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имещие модификацию на 5' конце, 3' конце или на обоих концах цепи.
[00101] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие модификации, которые приводят к несоответствию комплементарности между цепями.
[00102] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие 5'-пропиламиновый конец, a 5'-фосфорилированный конец, 3'-пуромициновый конец или 3'-биотиновую концевую группу.
[00103] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие 2'-фтор замещенный рибонуклеотид, 2'-OMe замещенный рибонуклеотид, 2'-дезоксирибонуклеотид, 2'-амино замещенный рибонуклеотид, 2'-тиозамещенный рибонуклеотид.
[00104] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие один или более 5-галоуридинов, 5-галоцитидинов, 5-метилцитидинов, риботимидинов, 2-аминопуринов, 2,6-диаминопуринов, 4-тиоуридинов или 5-аминоаллилуридинов.
[00105] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие одну или более фосфотиоатных групп.
[00106] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие один или более 2'-фтор замещенных рибонуклеотидов, 2'-фторуридинов, 2'-фторцитидинов, 2'-дезоксирибонуклеотидов, 2'-дезоксиаденозинов или 2'-дезоксигуанозинов.
[00107] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие одну или более фосфотиоатных связей.
[00108] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие одну или более алкилендиольных связей, оксиалкилтиольных связей или оксикарбонилокси связей.
[00109] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие одну или более дезоксиосновных групп, инозинов, N3-метил-уридинов, N6,N6-диметиладенозинов, псевдоуридинов, пуриновых рибонуклеозидов и рибавиринов.
[00110] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие одну или более 3ʹ или 5ʹ интвертированных концевых групп.
[00111] Примеры модифицированных и химически модифицированных siRNAs включают siRNAs, имеющие один или более 5-(2-амино)пропилуридинов, 5-бромуридинов, аденозинов, 8-бромгуанозинов, 7-деаза-аденозинов или N6-метиладенозин.
[00112] Способы для модуляции GST-π и лечения злокачественной опухоли
[00113] Варианты осуществления по настоящему изобретению могут предоставить RNAi молекулы, которые можно использовать для подавления или ингибирования экспрессии GST-π и/или GST-π белков.
[00114] В некоторых вариантах осуществления молекулу RNAi по настоящему изобретению можно использовать для подавления или ингибирования экспрессии GST-π и/или GST-π белков, возникающих на основе полиморфизма гаплотипов GST-π, который может быть связан с заболеванием или с таким состоянием, как злокачественная опухоль.
[00115] Мониторинг уровней белка или GST-π mRNA можно использовать, чтобы охарактеризовать сайленсинг гена, и чтобы определить эффективность соединений и композиций по настоящему изобретению.
[00116] RNAi молекулы по данному изобретению можно использовать индивидуально или в комбинации с другими siRNAs для модуляции экспрессии одного или более генов.
[00117] RNAi молекулы по данному изобретению можно использовать индивидуально или в комбинации, или в сочетании с другими известными препаратами для предотвращения или лечения заболеваний, или облегчения симптомов состояний или заболеваний, связанных с GST-π, включая злокачественную опухоль.
[00118] RNAi молекулы по настоящему изобретению можно использовать для модуляции или ингибирования экспрессии GST-π сиквенс-специфичным образом.
[00119] RNAi молекулы по данному изобретению могут включать направляющую цепь, для которой ряд смежных нуклеотидов, по меньшей мере, частично комплементарны GST-π mRNA.
[00120] В определенных аспектах злокачественную опухоль можно лечить посредством РНК-интерференции с использованием молекулы RNAi по настоящему изобретению.
[00121] Лечение злокачественной опухоли может быть охарактеризовано с применением подходящих клеточных моделей, а также ex vivo или in vivo моделей на животных.
[00122] Лечение злокачественной опухоли может быть охарактеризовано посредством определения уровня GST-π mRNA или уровня белка GST-π в клетках пораженной ткани.
[00123] Лечение злокачественной опухоли может быть охарактеризовано посредством неинвазивного медицинского исследования пораженного органа или ткани.
[00124] Варианты осуществления по настоящему изобретению могут включать способы для предотвращения, лечения или облегчения симптомов заболевания или состояния, связанного с GST-π, у нуждающегося в этом индивидуума.
[00125] В некоторых вариантах осуществления способы для предотвращения, лечения или облегчения симптомов злокачественной опухоли у индивидуума могут включать введение индивидууму молекулы RNAi по настоящему изобретению для модуляции экспрессии гена GST-π у индивидуума или организма.
[00126] В некоторых вариантах осуществления это изобретение рассматривает способы для подавления экспрессии гена GST-π в клетке или организме посредством контакта клетки или организма с молекулой RNAi по настоящему изобретению.
[00127] Варианты осуществления по настоящему изобретению охватывают молекулы siRNA из таблиц 1-6, которые являются модифицированными или химически модифицированными в соответствии с вышеприведенными примерами.
[00128] РНК-интерференция
[00129] РНК-интерференция (RNAi) относится к сиквенс-специфичному посттранскрипционному сайленсингу генов у животных посредством малой интерферирующей РНК (siRNAs). Смотри, например, Zamore et al., Клетка, 2000, Vol. 101, pp. 25-33; Fire et al., Nature, 1998, Vol. 391, pp. 806811; Sharp, Genes & Development, 1999, Vol. 13, pp. 139-141.
[00130] RNAi ответ в клетках может быть запущен посредством двухцепочечной RNA (dsRNA), хотя механизм еще не полностью понят. Определенные dsRNAs в клетках могут подвергаться действию дайсер фермента, фермента рибонуклеаза III. Смотри, например, Zamore et al., Клетка, 2000, Vol. 101, pp. 25-33; Hammond et al., Nature, 2000, Vol. 404, pp. 293-296. Дайсер может преобразовывать dsRNA в более короткие куски dsRNA, которые являются siRNAs.
[00131] В основном, siRNAs могут составлять приблизительно от 21 до приблизительно 23 нуклеотидов в длину и включают дуплексную область из пар оснований приблизительно 19 нуклеотидов в длину.
[00132] RNAi задействует эндонуклеазный комплекс, известный как РНК индуцируемый комплекс сайленсинга (RISC). siRNA имеет антисмысловую или направляющую цепь, которая заходит в RISC комплекс и опосредует расщепление одноцепочечной RNA мишени, имеющей последовательность, комплементарную антисмысловой цепи siRNA дуплекса. Другая цепь siRNA является сопровождающей цепью. Расщепление RNA мишени происходит в середине области, комплементарной антисмысловой цепи siRNA дуплекса. Смотри, например, Elbashir et al., Genes & Development, 2001, Vol. 15, pp. 188-200.
[00133] Как применяют в настоящем документе, термин ʺсмысловая цепьʺ относится к нуклеотидной последовательности молекулы siRNA, которая является частично или полностью комплементарной, по меньшей мере, части соответствующей антисмысловой цепи молекулы siRNA. Смысловая цепь молекулы siRNA может включать последовательность нуклеиновой кислоты, имеющую гомологию с последовательностью нуклеиновой кислоты, являющейся мишенью.
[00134] Как применяют в настоящем документе, термин ʺантисмысловая цепьʺ относится к нуклеотидной последовательности молекулы siRNA, которая является частично или полностью комплементарной, по меньшей мере, части последовательности нуклеиновой кислоты, являющейся мишенью. Антисмысловая цепь молекулы siRNA может включать последовательность нуклеиновой кислоты, которая является комплементарной, по меньшей мере, части соответствующей смысловой цепи молекулы siRNA.
[00135] RNAi молекулы могут подавлять или блокировать экспрессию гена, опосредуя РНК-интерференцию сиквенс-специфичным образом. Смотри, например, Zamore et al., Клетка, 2000, Vol. 101, pp. 25-33; Elbashir et al., Nature, 2001, Vol. 411, pp. 494-498; Kreutzer et al., WO2000/044895; Zernicka-Goetz et al., WO2001/36646; Fire et al., WO1999/032619; Plaetinck et al., WO2000/01846; Mello et al., WO2001/029058.
[00136] Как применяют в настоящем документе, термины ʺингибироватьʺ, ʺподавлятьʺ или ʺснижатьʺ в отношении экспрессии гена означает, что экспрессия гена или уровень mRNA молекул, кодирующих один или более белков, или активность одного или более кодируемых белков уменьшена ниже уровня, наблюдаемого в отсутствие молекулы RNAi или siRNA по настоящему изобретению. Например, уровень экспрессии, уровень mRNA или уровень активности кодируемого белка может быть снижен, по меньшей мере, на 1%, или, по меньшей мере, на 10%, или, по меньшей мере, на 20%, или, по меньшей мере, на 50%, или, по меньшей мере, на 90% или более от уровня, наблюдаемого в отсутствие молекулы RNAi или siRNA по настоящему изобретению.
[00137] RNAi молекулы также можно использовать для подавления экспрессии гена вируса, и, таким образом, воздействовать на репликацию вируса.
[00138] RNAi молекулы могут быть сделаны из отдельных полинуклеотидных цепей: смысловой цепи или сопровождающей цепи, и антисмысловой цепи или направляющей цепи. Направляющая и сопровождающая цепи являются, по меньшей мере, частично комплементарными. Направляющая цепь и сопровождающая цепь могут образовывать дуплексную область, имеющую приблизительно от 15 до приблизительно 49 пар оснований.
[00139] В некоторых вариантах осуществления дуплексная область siRNA может иметь 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, или 49 пар оснований.
[00140] В определенных вариантах осуществления молекула RNAi может быть активной в RISC комплексе, с длиной дуплексной области, активной для RISC.
[00141] В дополнительных вариантах осуществления молекула RNAi может быть активной в качестве субстрата дайсер фермента, предназначенного для превращения в молекулу RNAi, которая может быть активной в RISC комплексе.
[00142] В некоторых аспектах молекула RNAi может иметь части комплементарной направляющей и сопровождающей последовательности на противоположных концах длинной молекулы, таким образом, что молекула может образовать дуплексную область с частями комплементарной последовательности, и цепи связаны на одном конце дуплексной области посредством или нуклеотида, или ненуклеотидных линкеров. Например, расположение «шпилька», или расположение «ствол» и расположение «петля». Взаимодействия линкеров с цепями могут быть ковалентными связями или нековалентными взаимодействиями.
[00143] Молекула RNAi по данному изобретению может включать нуклеотид, ненуклеотид, или смешанный нуклеотидный/ненуклеотидный линкер, который соединяет смысловую область нуклеиновой кислоты с антисмысловой областью нуклеиновой кислоты. Нуклеотидный линкер может быть линкером ≧2 нуклеотидов в длину, например приблизительно 3, 4, 5, 6, 7, 8, 9 или 10 нуклеотидов в длину. Нуклеотидный линкер может быть аптамером на основе нуклеиновой кислоты. Под ʺаптамеромʺ или ʺаптамером на основе нуклеиновой кислотыʺ, как применяют в настоящем документе, подразумевают молекулу нуклеиновой кислоты, которая связывается специфическим образом с молекулой-мишенью, где молекула нуклеиновой кислоты имеет последовательность, которая включает последовательность, распознаваемую молекулой-мишенью в ее естественном окружении. Альтернативным образом, аптамером может быть молекула нуклеиновой кислоты, которая связывается с молекулой-мишенью, где молекула-мишень не связывается с нуклеиновой кислотой в естественном окружении. Например, аптамер можно использовать для связывания с лиганд-связывающим доменом белка, таким образом, предотвращая взаимодействие природного лиганда с белком. Смотри, например, Gold et al., Annu Rev Biochem, 1995, Vol. 64, pp. 763-797; Brody et al., J. Biotechnol., 2000, Vol. 74, pp. 5-13; Hermann et al., Science, 2000, Vol. 287, pp. 820-825.
[00144] Примеры ненуклеотидного линкера включают нуклеотид с удаленным азотистым основанием, простой полиэфир, полиамин, полиамид, пептид, углевод, липид, полиуглеводород или другие полимерные соединения, например, полиэтиленгликоли, такие как имеющие от 2 до 100 единиц этиленгликоля. Некоторые примеры описаны Seela et al., Nucleic acids Research, 1987, Vol. 15, pp. 3113-3129; Cload et al., J. Am. Chem. Soc., 1991, Vol. 113, pp. 6324-6326; Jaeschke et al., Tetrahedron Lett., 1993, Vol. 34, pp. 301; Arnold et al., WO1989/002439; Usman et al., WO1995/006731; Dudycz et al., WO1995/011910, и Ferentz et al., J. Am. Chem. Soc., 1991, Vol. 113, pp. 4000-4002.
[00145] Молекула RNAi может иметь один или более «липких» концов из дуплексной области. «Липкие» концы, которые являются неспаренными основаниями, одноцепочечными областями, могут иметь от одного до восьми нуклеотидов в длину, или длиннее. «Липкий» конец может быть 3′-концевым «липким» концом, где 3′-конец цепи имеет одноцепочечную область от одного до восьми нуклеотидов. «Липкий» конец может быть 5′-концевым «липким» концом, где 5′-конец цепи имеет одноцепочечную область от одного до восьми нуклеотидов.
[00146] «Липкие» концы молекулы RNAi могут иметь ту же длину или могут быть различной длины.
[00147] Молекула RNAi может иметь один или более тупых концов, в которых концы дуплексной области без «липкого» конца, и цепи являются спаренными с концом дуплексной области.
[00148] Молекула RNAi по данному изобретению может иметь один или более тупых концов или может иметь один или более «липких» концов, или может иметь комбинацию тупой конец и «липкий» конец.
[00149] 5′-конец цепи молекулы RNAi может находиться на тупом конце или может находиться на «липком» конце. 3′-конец цепи молекулы RNAi может находиться на тупом конце или может находиться на «липком» конце.
[00150] 5′-конец цепи молекулы RNAi может находиться на тупом конце, в то время как 3′-конец находится на «липком» конце. 3′-конец цепи молекулы RNAi может находиться на тупом конце, в то время как 5′-конец находится на «липком» конце.
[00151] В некоторых вариантах осуществления оба конца молекулы RNAi являются тупыми концами.
[00152] В дополнительных вариантах осуществления оба конца молекулы RNAi имеют «липкий» конец.
[00153] «Липкие» концы на 5′- и 3′-концах могут быть различной длины.
[00154] В определенных вариантах осуществления молекула RNAi может иметь тупой конец, где 5′-конец антисмысловой цепи и 3′-конец смысловой цепи не имеют каких-либо «липких» концевых нуклеотидов.
[00155] В дополнительных вариантах осуществления молекула RNAi может иметь тупой конец, где 3′-конец антисмысловой цепи и 5′-конец смысловой цепи не имеют каких-либо «липких» концевых нуклеотидов.
[00156] Молекула RNAi может иметь несоответствия спаривания оснований в дуплексной области.
[00157] Любой нуклеотид в «липком» конце молекулы RNAi может быть дезоксирибонуклеотидом или рибонуклеотидом.
[00158] Один или более дезоксирибонуклеотидов могут находиться на 5′-конце, где 3′-конец другой цепи молекулы RNAi может не иметь «липкого» конца или может не иметь дезоксирибонуклеотидный «липкий» конец.
[00159] Один или более дезоксирибонуклеотидов могут находиться на 3′-конце, где 5′-конец другой цепи молекулы RNAi может не иметь «липкий» конец, или может не иметь дезоксирибонуклеотидный «липкий» конец.
[00160] В некоторых вариантах осуществления один или более, или все из нуклеотидов «липкого» конца молекулы RNAi могут быть 2′-дезоксирибонуклеотидами.
[00161] Дайсер субстратные молекулы RNAi
[00162] В некоторых аспектах молекула RNAi может иметь длину, подходящую в качестве дайсер субстратата, который может быть подвергнут обработке для образования RISC активной молекулы RNAi. Смотри, например, Rossi et al., US2005/0244858.
[00163] Двухцепочечная RNA (dsRNA), которая является дайсер субстратом, может иметь длину, достаточную для того, чтобы подвергаться обработке дайсер ферментом для образования активной молекулы RNAi, и может дополнительно иметь одно или более из следующих качеств: (i) дайсер субстратная dsRNA может быть асимметричной, например, имеющей 3′ «липкий» конец на антисмысловой цепи, и (ii) дайсер субстратная dsRNA может иметь модифицированный 3′ конец на смысловой цепи, чтобы направлять ориентацию дайсер связывания и обработки dsRNA до активной молекулы RNAi.
[00164] В определенных вариантах осуществления самая длинная цепь в дайсер субстратной dsRNA может составлять 24-30 нуклеотидов в длину.
[00165] Дайсер субстратная dsRNA может быть симметричной или асимметричной.
[00166] В некоторых вариантах осуществления дайсер субстратная dsRNA может иметь смысловую цепь длиной 22-28 нуклеотидов и антисмысловую цепь длиной 24-30 нуклеотидов.
[00167] В определенных вариантах осуществления дайсер субстратная dsRNA может иметь «липкий» конец на 3′ конце антисмысловой цепи.
[00168] В дополнительных вариантах осуществления дайсер субстратная dsRNA может иметь смысловую цепь 25 нуклеотидов в длину и антисмысловую цепь 27 нуклеотидов в длину, с 3′-«липким» концом длиной в 2 основания. «Липкий» конец может иметь 1, 2 или 3 нуклеотида в длину. Смысловая цепь может также иметь 5′ фосфат.
[00169] Асимметричная дайсер субстратная dsRNA может иметь два дезоксирибонуклеотида на 3′-конце смысловой цепи вместо двух из рибонуклеотидов.
[00170] Смысловая цепь дайсер субстратной dsRNA может составлять от приблизительно 22 до приблизительно 30, или от приблизительно 22 до приблизительно 28; или от приблизительно 24 до приблизительно 30; или от приблизительно 25 до приблизительно 30; или от приблизительно 26 до приблизительно 30; или от приблизительно 26 до 29; или от приблизительно 27 до приблизительно 28 нуклеотидов в длину.
[00171] Смысловая цепь дайсер субстратной dsRNA может иметь 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидов в длину.
[00172] В определенных вариантах осуществления дайсер субстратная dsRNA может иметь смысловую и антисмысловую цепи, которые составляют, по меньшей мере, приблизительно 25 нуклеотидов в длину, и не длиннее, чем приблизительно 30 нуклеотидов в длину.
[00173] В определенных вариантах осуществления дайсер субстратная dsRNA может иметь смысловую и антисмысловую цепи, которые составляют от 26 до 29 нуклеотидов в длину.
[00174] В определенных вариантах осуществления дайсер субстратная dsRNA может иметь смысловую и антисмысловую цепи, которые составляют 27 нуклеотидов в длину.
[00175] Смысловая и антисмысловая цепи дайсер субстратной dsRNA могут иметь одинаковую длину при наличии тупых концов, или различную длину при наличии «липких» концов, или могут иметь тупой конец и «липкий» конец.
[00176] Дайсер субстратная dsRNA может иметь дуплексную область 19, 20, 21, 22, 23, 24, 25, 26 или 27 нуклеотидов в длину.
[00177] Антисмысловая цепь дайсер субстратной dsRNA может иметь любую последовательность, которая ренатурирует, по меньшей мере, часть последовательности смысловой цепи в биологическом состоянии, как, например, внутри цитоплазмы эукариотической клетки.
[00178] Дайсер субстрат со смысловой и антисмысловой цепью может быть связан посредством третьей структуры, такой как линкерная группа или линкерный олигонуклеотид. Линкер соединяет две цепи dsRNA, например, таким образом, что «шпилька» образуется при ренатурации.
[00179] Смысловая и антисмысловая цепи дайсер субстрата являются в основном комплементарными, но могут иметь несоответствия при спаривании оснований.
[00180] В некоторых вариантах осуществления дайсер субстратная dsRNA может быть асимметричной, таким образом, что смысловая цепь имеет 22-28 нуклеотидов и антисмысловая цепь имеет 24-30 нуклеотидов.
[00181] Область одной из цепей, конкретно антисмысловой цепи, дайсер субстратной dsRNA может иметь последовательность длиной, по меньшей мере, 19 нуклеотидов, где эти нуклеотиды находятся в 21-нуклеотидной области, прилегающей к 3′ концу антисмысловой цепи, и являются в достаточной степени комплементарными нуклеотидной последовательности RNA, синтезированной на основе таргетного гена.
[00182] Антисмысловая цепь дайсер субстратной dsRNA может иметь от 1 до 9 рибонуклеотидов на 5′-конце, при длине 22-28 нуклеотидов. Если антисмысловая цепь имеет длину 21 нуклеотид, тогда 1-7 рибонуклеотидов или 2-5 рибонуклеотидов или 4 рибонуклеотида можно добавлять на 3′-конце. Добавленные рибонуклеотиды могут иметь любую последовательность.
[00183] Смысловая цепь дайсер субстратной dsRNA может иметь 24-30 нуклеотидов. Смысловая цепь может быть по существу комплементарной антисмысловой цепи, чтобы ренатурировать с антисмысловой цепью в биологическом окружении.
[00184] Способы применения молекул RNAi
[00185] Молекулы нуклеиновой кислоты и RNAi молекулы по настоящему изобретению могут быть доставлены к клетке или ткани посредством прямого воздействия молекул, или с применением молекул, комбинированных с носителем или растворителем.
[00186] Молекулы нуклеиновой кислоты и RNAi молекулы по настоящему изобретению могут быть доставлены или введены в клетку, ткань, орган или индивидуума посредством прямого воздействия молекул с применением носителя или растворителя, или любой другой системой для доставки, которая действует для помощи, ускорения или облегчения входа внутрь клетки, например, вирусных последовательностей, вирусного материала или липидных или липосомальных составов.
[00187] Молекулы нуклеиновой кислоты и RNAi молекулы по настоящему изобретению могут быть объединены с катионными липидами, упакованы внутрь липосом или иным образом доставлены к клеткам-мишеням или тканям. Нуклеиновая кислота или комплексы с нуклеиновой кислотой могут быть доставлены местно в соответствующие ткани ex vivo или in vivo посредством прямого нанесения на кожу, трансдермального нанесения или посредством инъекции.
[00188] Системы доставки могут включать, например, водные и неводные гели, кремы, эмульсии, микроэмульсии, липосомы, мази, водные и неводные растворы, лосьоны, аэрозоли, углеводородные основы и порошки, и могут содержать эксципиенты, такие как солюбилизаторы и усилители проницаемости.
[00189] Композиции и способы по данному изобретению могут включать экспрессирующий вектор, который включает последовательность нуклеиновой кислоты, кодирующую, по меньшей мере, одну молекулу RNAi по настоящему изобретению таким образом, который обеспечивает экспрессию молекулы нуклеиновой кислоты.
[00190] Молекулы нуклеиновой кислоты и RNAi молекулы по настоящему изобретению могут экспрессироваться из единиц транскрипции, вставленных в DNA или RN Векторы. Рекомбинантные векторы могут быть DNA плазмидами или вирусными векторами. Можно использовать вирусные векторы, которые обеспечивают транзиентную экспрессию молекул нуклеиновой кислоты.
[00191] Например, вектор может содержать последовательности, кодирующие обе цепи молекулы RNAi дуплекса, или единичную молекулу нуклеиновой кислоты, которая является самокомплементарной и, таким образом, образует молекулу RNAi. Экспрессирующий вектор может включать последовательность нуклеиновой кислоты, кодирующую две или более молекул нуклеиновой кислоты.
[00192] Молекула нуклеиновой кислоты может экспрессироваться внутри клетки из эукариотических промоторов. Специалисты в данной области понимают, что любая нуклеиновая кислота может экспрессироваться в эукариотических клетках из соответствующего DNA/RN Вектора.
[00193] В некоторых аспектах вирусную конструкцию можно использовать для введения экспрессирующей конструкции внутрь клетки с целью транскрипции dsRNA конструкции, кодируемой посредством экспрессирующей конструкции.
[00194] Липидные составы можно вводить животным посредством внутривенной, внутримышечной или интраперитонеальной инъекции, или перорально или посредством ингаляции или другими способами, которые известны в данной области.
[00195] Для введения олигонуклеотидов известны и можно использовать фармацевтически приемлемые составы.
[00196] Примерный протокол для in vitro нокдауна
[00197] За один день до трансфекции клетки высевали в 96-луночный планшет в количестве 2×103 клеток на лунку с 100 μl DMEM (HyClone Cat. # SH30243.01), содержащей 10% FBS, и культивировали при 37˚C в инкубаторе, содержащем увлажненную атмосферу 5% CO2 в воздухе. До трансфекции среду заменяли на 90 μl Opti-MEM I Reduced Serum Medium (Life Technologies Cat. # 31985-070), содержащую 2% FBS. Затем, 0.2 μl липофектамина RNAiMax (Life Technologies Cat. # 13778-100) смешивали с 4.8 μl Opti-MEM I в течение 5 минут при комнатной температуре. Далее, 1 μl siRNA смешивали с 4 μl Opti-MEM I и комбинировали с LF2000 раствором, и аккуратно перемешивали без встряхивания. После 5 минут при комнатной температуре смесь инкубировали в течение дополнительных 10 минут при комнатной температуре, чтобы обеспечить образование комплексов RNA-RNAiMax. Дополнительно, 10 μl комплексов RNA-RNAiMax добавляли в лунку, и планшет аккуратно встряхивали руками. Клетки инкубировали при 37˚C в инкубаторе, содержащем увлажненную атмосферу 5% CO2 в воздухе в течение 2 часов. Среду меняли на свежую Opti-MEM I Reduced Serum Medium, содержащую 2% FBS. Через 24 часа после трансфекции клетки отмывали с применением ледяного PBS однократно. Клетки лизировали с применением 50 μl Клетка-to-Ct Lysis Buffer (Life Technologies Cat. # 4391851 C) в течение 5-30 минут при комнатной температуре. Добавляли 5 μl останавливающего раствора и инкубировали в течение 2 минут при комнатной температуре. Уровень mRNA измеряли посредством RT-qPCR с TAQMAN незамедлительно. Образцы могли быть заморожены при -80°C и проанализированы позднее.
[00198] Примерный протокол для стабильности в сыворотке
[00199] 0.2 мг/мл siRNA инкубировали с 10% сыворотки человека при 37 °C. В определенные моменты времени (0, 5, 15 и 30 минут) 200 μl образца разделяли на аликвоты и экстрагировали с применением 200 μl экстрагента (хлороформ: фенол: изоамиловый спирт =24:25:1). Образец перемешивали на центрифуге «вортексировали» и центрифугировали при 13,000 rpm в течение 10 минут при RT, затем раствор верхнего слоя переносили и фильтровали его с примением 0.45 мкм фильтра. Фильтрат переносили в 300 μl ВЭЖХ инъекционную трубку. Для LCMS подвижная фаза представляла MPA: 100 mM HFIP+7 mM TEA в H2O, MPB: 50% метанол+50% ацетонитрил. Колонка: Waters Acquity OST 2.1×50 мм, 1.7 мкм.
[00200] ПРИМЕРЫ
[00201] Пример 1: Было выявлено, что siRNAs по настоящему изобретению, мишенью которых является GST-π, являются активными для сайленсинга гена in vitro. Было выявлено, что дозозависимое действие GST-π siRNAs в отношении нокдауна гена демонстрирует IC50 ниже приблизительно 250 пикомолярной (пМ), и до 1 пМ.
[00202] Чтобы определить эффективность нокдауна siRNA, проводили in vitro трансфекцию в клеточной линии A549. Дозозависимый нокдаун для GST-π mRNA наблюдали с применением siRNAs из таблицы 1, как показано на таблице 7.
Таблица 7: Дозозависимый нокдаун для GST-π mRNA в клеточной линии A549
[00203] Как показано в таблице 7, действие GST-π siRNAs из таблицы 1 было в диапазоне 17-235 пМ, который является подходящим для многих применений, включая применение в качестве лекарственного средства для применения in vivo.
[00204] Пример 2: Структура GST-π siRNAs по настоящему изобретению, имеющая дезоксинуклеотиды, расположенные в затравочной области антисмысловой цепи siRNA, обеспечивала непредвиденное и эффективное повышение активности в отношении нокдауна гена in vitro.
[00205] In vitro трансфекцию проводили в клеточной линии A549 для определения эффективности нокдауна для GST-π siRNAs на основе структуры BU2ʹ (SEQ ID NO:131 и 157). Дозозависимый нокдаун GST-π mRNA наблюдали с применением GST-π siRNAs на основе структуры BU2ʹ, как показано в таблице 8.
Таблица 8: дозозависимый нокдаун GST-π mRNA в клеточной линии A549 для GST-π siRNAs на основе структуры BU2ʹ
[00206] Как показано на таблице 8, активности GST-π siRNAs на основе структуры BU2ʹ, имеющей три дезоксинуклеотида в затравочной области антисмысловой цепи к удивлению и неожиданно повышались до 6 раз по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00207] Эти данные демонстрируют, что GST-π siRNAs, имеющие структуру с тремя дезоксинуклеотидами, расположенными в положениях 3, 5 и 7 или в положениях 4, 6 и 8 в затравочной области антисмысловой цепи, к удивлению обеспечивали повышенную активность нокдауна гена по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00208] Активности, показанные в таблице 8 для GST-π siRNAs, имеющих три дезоксинуклеотида в затравочной области антисмысловой цепи, находились в диапазоне от 5 до 8 пМ, которая является исключительно подходящей для многих применений, включая применение в качестве лекарственного средства in vivo.
[00209] Пример 3: Структура GST-π siRNAs по настоящему изобретению, имеющих дезоксинуклеотиды, расположенные в затравочной области антисмысловой цепи siRNA, обеспечивала непредвиденное и эффективное повышение активности нокдауна гена in vitro.
[00210] In vitro трансфекцию проводили в клеточной линии A549, чтобы определить эффективность нокдауна для GST-π siRNAs на основе структуры A9ʹ (SEQ ID NO:183 и 195). Дозозависимый нокдаун GST-π mRNA наблюдали с применением GST-π siRNAs на основе структуры A9ʹ, как показано в таблице 9.
Таблица 9: дозозависимый нокдаун GST-π mRNA в клеточной линии A549 для GST-π siRNAs на основе структуры A9ʹ
[00211] Как показано в таблице 9, активности GST-π siRNAs на основе структуры A9ʹ, имеющих от трех до шести дезоксинуклеотидов в затравочной области антисмысловой цепи, к удивлению были повышены до 24 раз по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00212] Эти данные демонстрируют, что GST-π siRNAs, имеющие структуру с от трех до шести дезоксинуклеотидов, расположенных в положениях 4, 6 и 8 или в положениях 1, 3, 5 и 7, или в положениях 3-8, или в положениях 5-8, или в положениях 3, 5 и 7 в затравочной области антисмысловой цепи, обеспечивали непредвиденное повышение активности нокдауна гена по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00213] Активность, показанная в таблице 9 для GST-π siRNAs, имеющих от трех до шести дезоксинуклеотидов в затравочной области антисмысловой цепи, находилась в диапазоне от 1 до 15 пМ, которая является исключительно подходящей для многих применений, включая применение в качестве лекарственного средства in vivo.
[00214] Пример 4: структура GST-π siRNAs, имеющих дезоксинуклеотиды, расположенные в затравочной области антисмысловой цепи siRNA, обеспечивала непредвиденное и эффективное повышение активности нокдауна гена in vitro.
[00215] In vitro трансфекцию проводили в клеточной линии A549 для определения эффективности нокдауна для GST-π siRNAs на основе структуры B13ʹ (SEQ ID NO:207 и 222). Дозозависимый нокдаун GST-π mRNA наблюдали с применением GST-π siRNAs на основе структуры B13ʹ, как показано в таблице 10.
Таблица 10: дозозависимый нокдаун GST-π mRNA в клеточной линии A549 для GST-π siRNAs на основе структуры B13ʹ
[00216] Как показано в таблице 10, активность GST-π siRNA на основе структуры B13ʹ, имеющей три дезоксинуклеотида в затравочной области антисмысловой цепи, была неожиданно повышена по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00217] Эти данные демонстрируют, что GST-π siRNAs, имеющие структуру с тремя дезоксинуклеотидами, расположенными в положениях 4, 6 и 8 в затравочной области антисмысловой цепи, обеспечивали неожиданно повышенную активность нокдауна гена по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00218] Активность, показанная в таблице 10 для GST-π siRNAs, имеющих три дезоксинуклеотида в затравочной области антисмысловой цепи, находилась в пикомолярном диапазоне при 11 пМ, которая является исключительно подходящей для многих применений, включая применения в качестве лекарственного средства in vivo.
[00219] Пример 5: Структура GST-π siRNAs, имеющих дезоксинуклеотиды, расположенные в затравочной области антисмысловой цепи siRNA, обеспечивала неожиданно и успешно повышенную активность нокдауна гена in vitro.
[00220] In vitro трансфекцию проводили в клеточной линии A549 для определения эффективности нокдауна для GST-π siRNAs на основе структуры B4ʹ (SEQ ID NO:261 и 273). Дозозависимый нокдаун GST-π mRNA наблюдали с применением GST-π siRNAs на основе структуры B4ʹ, как показано в таблице 11.
Таблица 11: Дозозависимый нокдаун GST-π mRNA в клеточной линии A549 для GST-π siRNAs на основе структуры B4ʹ
[00221] Как показано в таблице 11, активности GST-π siRNAs на основе структуры B4ʹ, имеющие шесть дезоксинуклеотидов в затравочной области антисмысловой цепи, были неожиданно повышены более чем в два раза по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00222] Эти данные демонстрируют, что GST-π siRNAs, имеющие структуру с шестью дезоксинуклеотидами, расположенными в положениях 3-8 в затравочной области антисмысловой цепи, к удивлению обеспечивали повышенную активность нокдауна гена по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00223] Активность, показанная в таблице 11 для GST-π siRNA, имеющей шесть дезоксинуклеотидов в затравочной области антисмысловой цепи, находилась в пикомолярном диапазоне при 113 пМ, которая является исключительно подходящей для многих применений, включая применение в качестве лекарственного средства in vivo.
[00224] Пример 6: Cтруктура GST-π siRNAs, имеющих дезоксинуклеотиды, расположенные в затравочной области антисмысловой цепи siRNA, обеспечивала непредвиденное и эффективное повышение активности нокдауна гена in vitro.
[00225] In vitro трансфекцию проводили в клеточной линии A549, чтобы определить эффективность нокдауна для GST-π siRNAs на основе структуры B2ʹ (SEQ ID NO:237 и 249). Дозозависимый нокдаун GST-π mRNA наблюдали с применением GST-π siRNAs на основе структуры B2ʹ, как показано в таблице 12.
Таблица 12: Дозозависимый нокдаун GST-π mRNA в клеточной линии A549 для GST-π siRNAs на основе структуры B2ʹ
[00226] Как показано в таблице 12, активности GST-π siRNAs на основе структуры B2ʹ, имеющих от трех до четырех дезоксинуклеотидов в затравочной области антисмысловой цепи, к удивлению были повышены до 4 раз по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00227] Эти данные демонстрируют, что GST-π siRNAs, имеющие структуру с от трех до четырех дезоксинуклеотидов, расположенных в положениях 5-8, или в положениях 1, 3, 5 и 7, или в положениях 3, 5 и 7 в затравочной области антисмысловой цепи, обеспечивали непредвиденное повышение активности нокдауна гена по сравнению с GST-π siRNA без дезоксинуклеотидов в дуплексной области.
[00228] Активности, представленные в таблице 12 для GST-π siRNAs, имеющих от трех до четырех дезоксинуклеотидов в затравочной области антисмысловой цепи, находились в диапазоне 30-100 пМ, которая является исключительно подходящей для многих применений, включая применение в качестве лекарственного средства in vivo.
[00229] Пример 7: Структура GST-π siRNAs, содержащих один или более 2ʹ-дезокси-2ʹ-фтор замещенных нуклеотидов, обеспечивала непредвиденное повышение активности нокдауна гена in vitro.
[00230] In vitro трансфекцию проводили в клеточной линии A549, чтобы определить эффективность нокдауна для GST-π siRNAs на основе структуры BU2ʹ (SEQ ID NO:131 и 157). Дозозависимый нокдаун GST-π mRNA наблюдали с применением GST-π siRNAs на основе структуры BU2ʹ, как показано в таблице 13.
Таблица 13: Дозозависимый нокдаун GST-π mRNA в клеточной линии A549 для GST-π siRNAs на основе структуры BU2ʹ
[00231] Как показано в таблице 13, активности GST-π siRNAs на основе структуры BU2ʹ, имеющих один или более 2ʹ-F дезоксинуклеотидов, к удивлению были повышены до 10 раз по сравнению с GST-π siRNA без 2ʹ-F дезоксинуклеотидов.
[00232] Эти данные демонстрируют, что GST-π siRNAs, имеющие структуру с одним или более 2ʹ-F дезоксинуклеотидами, обеспечивали непредвиденное повышение активности нокдауна гена по сравнению с GST-π siRNA без 2ʹ-F дезоксинуклеотида.
[00233] Активности, представленные в таблице 13 для GST-π siRNAs, имеющих один или более 2ʹ-F дезоксинуклеотидов, находились в диапазоне от 3 до 13 пМ, который является исключительно подходящим для многих применений, включая применение в качестве лекарственного средства in vivo.
[00234] Пример 8: Структура GST-π siRNAs, содержащих один или более 2ʹ-дезокси-2ʹ-фтор замещенных нуклеотидов, обеспечивала непредвиденное повышение активности нокдауна гена in vitro.
[00235] In vitro трансфекцию проводили в клеточной линии A549, чтобы определить эффективность нокдауна для GST-π siRNAs на основе структуры B13ʹ (SEQ ID NO:207 и 222). Дозозависимый нокдаун GST-π mRNA наблюдали с применением GST-π siRNAs на основе структуры B13ʹ, как показано в таблице 14.
Таблица 14: Дозозависимый нокдаун GST-π mRNA в клеточной линии A549 для GST-π siRNAs на основе структуры B13ʹ
[00236] Как показано в таблице 14, активность GST-π siRNA на основе структуры B13ʹ, имеющей три 2ʹ-F дезоксинуклеотида, расположенных в положениях на «нелипких» концах, была к удивлению повышена приблизительно в 3 раза, по сравнению с GST-π siRNA без 2ʹ-F дезоксинуклеотидов.
[00237] Эти данные демонстрируют, что GST-π siRNAs, имеющие структуру с одним или более 2ʹ-F дезоксинуклеотидами, обеспечивали непредвиденное повышение активности нокдауна гена по сравнению с GST-π siRNA без 2ʹ-F дезоксинуклеотида.
[00238] Активность, представленная в таблице 14 для GST-π siRNAs, имеющих один или более 2ʹ-F дезоксинуклеотидов, находилась в пикомолярном диапазоне на 6 пМ, что является исключительно подходящим для многих применений, включая применение в качестве лекарственного средства in vivo.
[00239] Пример 9: Ортотопическая модель A549 рака легких в мыши. GST-π siRNAs по настоящему изобретению могут демонстрировать значительное уменьшение ортотопических раковых опухолей легких in vivo. В данном примере GST-π siRNA обеспечивала активность нокдауна гена in vivo при введении в липосомальном составе в ортотопические раковые опухоли легких у бестимусных голых мышей.
[00240] В основном, ортотопическая модель опухоли может демонстрировать непосредственную клиническую значимость эффективности и силы препарата, а также улучшенную прогностическую возможность. В ортотопической модели опухоли опухолевые клетки имплантируют непосредственно внутрь того же органа, из которого произошли клетки.
[00241] Противоопухолевую эффективность siRNA состава против рака легких человека A549 оценивали посредством сравнения окончательного веса первичной опухоли, измеренного при некропсии, между лечебной группой и контрольной группой, получающей носитель.
[00242] На фиг. 1 продемонстрировано ингибирование опухоли ортотопического ракого легких in vivo для GST-π siRNA на основе структуры BU2 (SEQ ID NO:61 и 126). Ортотопическая модель A549 рака легких в мыши была использована при относительно низкой дозе 2 мг/кг siRNA, направленной на GST-π.
[00243] GST-π siRNA демонстрировала значительную и непредвиденно успешную эффективность ингибирования опухоли в этом шестинедельном исследовании. Как показано на фиг. 1, после 43 дней, GST-π siRNA демонстрировала значительно успешную эффективность ингибирования опухоли с окончательными средними размерами опухоли, значительно уменьшенными в 2,8 раза по сравнению с контролем.
[00244] Для этого исследования применяли 5-6 недельных самцов мышей NCr nu/nu. Экспериментальных животных содержали в HEPA фильтруемом окружении во время экспериментального периода. siRNA составы хранили при 4℃ C перед применением, и нагревали до комнатной температуры 10 минут до инъекции в мышь.
[00245] Для этой ортотопической модели A549 рака легких в день хирургической ортотопической имплантации (SOI) собирали опухоли из подкожного очага у животных, несущих A549 опухолевый ксенотрансплантат, и помещали в среду RPMI-1640. Некротизированные ткани удаляли, и жизнеспособные ткани разрезали на кусочки 1.5-2 мм3. Животные получали анестезию посредством ингаляции изофлурана, и операционное поле стерилизовали с применением йода и спирта. Поперечный разрез приблизительно 1.5 см длиной делали в левой части грудной стенки мыши с применением хирургических ножниц. Межреберный разрез делали между третьим и четвертым ребром и обнажали левое легкое. Один фрагмент A549 опухоли трансплантировали на поверхность легкого с применением хирургической нити 8-0 (нейлон). Грудную клетку закрывали с применением хирургической нити 6-0 (шелк). Легкое снова надували посредством интраторакальной пункции с использованием 3 cc шприца с иглой 25 G X 1 ½, чтобы вывести наружу воздух, оставшийся в грудной полости. Грудную стенку закрывали с применением хирургической шелковой нити 6-0. Все процедуры операции, описанные выше, проводили с применением микроскопа с 7x увеличением в HEPA фильтруемых вытяжных шкафах с ламинарным потоком.
[00246] Через три дня после имплантации опухоли мышей, несущих модели опухолей, случайным образом делили на группы по десять мышей в группе. Для группы, представляющей интерес, лечение десяти мышей начинали через три дня после имплантации опухоли.
[00247] Для группы, представляющей интерес, состав представлял собой липосомальную композицию (ионизируемый липид:холестерин:DOPE:DOPC:DPPE-PEG-2K:DSPE-PEG-2K). В липосомах была инкапсулирована GST-π siRNA.
[00248] В конечной точке исследования экспериментальных мышей умерщвляли через сорок два дня после начала лечения. Первичные опухоли вырезали и взвешивали на электронных весах для последующего анализа.
[00249] Для оценки токсичности соединения среднюю массу тела мышей в лечебной и контрольной группах содержали в пределах нормального диапазона в течение всего экспериментального периода. У мышей не наблюдали других симптомов токсичности.
[00250] Пример 10: GST-π siRNAs по настоящему изобретению демонстрировали существенное уменьшение раковых опухолевых ксенотрансплантатов in vivo. GST-π siRNAs обеспечивали силу нокдауна гена in vivo при введении в липосомальном составе в раковые опухолевые ксенотрансплантаты.
[00251] На фиг. 2 показана эффективность ингибирования опухоли для GST-π siRNA (SEQ ID NO:156 и 182). Ксенотрансплантационную модель рака использовали в относительно низкой дозе при 0.75 мг/кг siRNA, направленной на GST-π.
[00252] GST-π siRNA демонстрировала значительную и непредвиденно успешную эффективность ингибирования опухоли внутри в течение нескольких дней после введения. После 36 дней GST-π siRNA продемонстрировала существенную успешную эффективность ингибирования опухоли с объемом опухоли, уменьшенным в 2 раза по сравнению с контролем.
[00253] Как показано на фиг. 3, GST-π siRNA продемонстрировала значительную и непредвиденно успешную эффективность ингибирования опухоли в день конечной точки исследования. Конкретно, вес опухоли был снижен более, чем в 2 раза.
[00254] GST-π siRNA вводили в двух инъекциях (день 1 и 15) липосомального состава, содержащего композицию (ионизируемый липид: холестерин: DOPE: DOPC: DPPE-PEG-2K) (25:30:20:20:5).
[00255] Для ксенотрансплантационной модели рака A549 клеточную линию получали из ATCC. Клетки поддерживали в среде для культивирования с добавлением 10% эмбриональной телячьей сыворотки и 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина. Клетки рассаживали за 48 часов до введения, так что клетки находились в фазе логарифмического роста при снятии. Клетки слегка трипсинизировали с применением трипсина-EDTA и снимали с тканевой культуры. Число жизнеспособных клеток подсчитывали и определяли в гемоцитометре в присутствии трипанового синего (подсчитывали только жизнеспособные клетки). Клетки ресуспендировали до концентрации 5 × 107/мл в среде без сыворотки. Затем клеточную суспензию тщательно смешивали с оттаявшим на льду BD матригелем в соотношении 1:1 для инъекции.
[00256] Mыши были бестимусными голыми самками мышей (nu/nu) с иммунодефицитом из Charles River Laboratory, 6-8 недельного возраста, 7-8 мышей в группе.
[00257] Для получения модели опухоли каждой мыши подкожно вводили в правый бок 0,1 мл инокулята 2,5 × 106 клеток A549 с использованием 25 G иглы и шприца, один инокулят на мышь. Мыши не получали анестезии для инокуляции.
[00258] Для измерений объема опухоли и рандомизации размер опухоли измеряли с точностью до 0,1 мм. Объемы опухолей рассчитывали с использованием формулы: объем опухоли=длина x ширина2/2. Как только установленная опухоль достигала приблизительно 120-175 мм3, средний объем опухоли составлял приблизительно 150 мм3, мышей распределяли в различные получающую носитель контрольную и лечебную группы, таким образом, что средние объемы опухолей в лечебных группах находились в пределах 10% от среднего объема опухоли в контрольной группе, получающей носитель, в идеальном случае, CV% объема опухоли составлял менее, чем 25%. В тот же день тестируемые препараты и контрольный носитель вводили в соответствии с режимом дозирования. Объемы опухолей мониторировали три раза в течение недели 1, дважды в течение оставшихся недель, включая день завершения исследования.
[00259] Для введения дозы в деь получения дозировки тестируемые препараты извлекали из холодильника -80°C и оттаивали на льду. До ввдения в шприцы флакон, содержащий состав, переворачивали в руках несколько раз. Все тестируемые препараты дозировали по 0.75 мг/кг посредством IV, q2w X 2, при 10 мл/кг.
[00260] Для массы тела мышей взвешивали с точностью до 0,1 г. Массы тела мониторировали и записывали ежедневно в течение 7 дней после дозирования первой дозы. Массы тела мониторировали и записывали дважды в неделю, в течение оставшихся недель, включая день завершения исследования.
[00261] Для сбора опухолей на 28 день после введения первой дозировки измеряли объем опухоли, и опухоль рассекали для измерения веса и хранили в течение PD биомаркерного исследования. Вес опухоли записывали.
[00262] Пример 11: GST-π siRNAs по настоящему изобретению демонстрировали повышенную смерть посредством апоптоза среди злокачественных клеток in vitro. GST-π siRNAs обеспечивали GST-π нокдаун, который приводил к повышению экспрессии PUMA, биомаркера апоптоза и связанного с потерей жизнеспособности клеток.
[00263] GST-π siRNA SEQ ID NO:156 и 182, которые содержали комбинацию дезоксинуклеотидов в затравочной области, 2ʹ-F замещенного дезоксинуклеотида и 2ʹ-OMe замещенных рибонуклеотидов, обеспечивали непредвиденное усиление апоптоза злокачественных клеток.
[00264] Уровень экспрессии PUMA для GST-π siRNA SEQ ID NO:156 и 182 измеряли, как показано на фиг. 4. На фиг. 4, экспрессия PUMA была существенным образом повышена в течение 2-4 дней после трансфекции GST-π siRNA.
[00265] Эти данные демонстрируют, что структура GST-π siRNAs, содержащих комбинацию дезоксинуклеотидов в затравочной области, 2ʹ-F замещенного дезоксинуклеотида и 2ʹ-OMe замещенных рибонуклеотидов, обеспечивала непредвиденное усиление апоптоза злокачественных клеток.
[00266] Протокол для биомаркера PUMA был следующим. За один день до трансфекции клетки высевали на 96-луночный планшет при 2×103 клеток на лунку с 100 мкл DMEM (HyClone Cat. # SH30243.01), содержащей 10% FBS, и культивировали в 37˚C инкубаторе с увлажненной атмосферой 5% CO2 в воздухе. На следующий день, до трансфекции среду заменяли 90 мкл среды Opti-MEM I Reduced Serum Medium (Life Technologies Cat. # 31985-070), содержащей 2% FBS. Затем, 0,2 мкл липофектамина RNAiMAX (Life Technologies Cat. #13778-100) смешивали с 4,8 мкл Opti-MEM I в течение 5 минут при комнатной температуре. 1 мкл GST-π siRNA (концентрация стокового раствора 1 мкM) смешивали c 4 мкл Opti-MEM I и комбинировали с раствором RNAiMAX, а затем аккуратно перемешивали. Смесь инкубировали в течение 10 минут при комнатной температуре, чтобы обеспечить образование комплексов RNA-RNAiMAX. 10 мкл комплексов RNA-RNAiMAX добавляли на лунку до конечной концентрации siRNA 10 нМ. Клетки инкубировали в течение 2 часов и среду меняли на свежую среду Opti-MEM I Reduced Serum Medium, содержащую 2% FBS. В течение 1, 2, 3, 4 и 6 дней после трансфекции клетки отмывали ледяным PBS однократно, а затем лизировали с применением 50 мкл Клетка-to-Ct Lysis Buffer (Life Technologies Cat. # 4391851 C) в течение 5-30 минут при комнатной температуре. Добавляли 5 мкл останавливающего раствора и инкубировали в течение 2 минут при комнатной температуре. Уровни PUMA (BBC3, Cat# Hs00248075, Life Technologies) mRNA измеряли посредством qPCR с применением TAQMAN.
[00267] Пример 12: GST-π siRNAs по настоящему изобретению могут демонстрировать значительное уменьшение раковой ксенотрансплантированной опухоли in vivo. GST-π siRNAs могут обеспечивать способность нокдауна гена in vivo при введении в липосомальном составе в раковую ксенотрансплантированную опухоль.
[00268] На фиг. 5 показана эффективность ингибирования опухоли для GST-π siRNA (SEQ ID NO:61 и 126). Дозозависимый нокдаун GST-π mRNA наблюдали in vivo с применением siRNA, направленной на GST-π. Ксенотрансплантационную модель рака использовали с siRNA, направленной на GST-π.
[00269] GST-π siRNA демонстрировала значительную и непредвиденно успешную эффективность ингибирования опухоли в течение нескольких дней после введения. Как показано на фиг. 5, лечение с применением GST-π siRNA, приводило к значительному уменьшению экспрессии GST-π mRNA через 4 дня после инъекции в липидном составе. При наиболее высокой дозе 4мг/кг детектировали значительное уменьшение приблизительно на 40% через 24 часа после инъекции.
[00270] GST-π siRNA вводили в однократной инъекции 10 мл/кг липосомального состава, содержащего композицию (ионизируемый липид:холестерин: DOPE: DOPC: DPPE-PEG-2K) (25:30:20:20:5).
[00271] Для ксенотрансплантационной модели рака, клеточную линию A549 получали из ATCC. Клетки поддерживали в RPMI-1640 с добавлением 10% эмбриональной телячьей сыворотки и 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина. Клетки разреживали за 48 часов до инокуляции таким образом, чтобы клетки находились в логарифмической фазе роста при снятии. Клетки аккуратно трипсинизировали с применением трипсина-EDTA и собирали из тканевой культуры. Число жизнеспособных клеток подсчитывали и определяли в гемоцитометре в присутствии трипанового синего (подсчитывали только жизнеспособные клетки). Клетки ресуспендировали до концентрации 4 × 107/мл в среде RPMI бе сыворотки. Затем клеточную суспензию тщательно перемешивали с растопленным на льду BD матригелем в соотношении 1:1 для инъекции.
[00272] Мыши были самками бестимусных голых (nu/nu) иммунодефицитных мышей из Charles River Laboratory, 6-8 недельного возраста, по 3 мыши в группе.
[00273] Для получения модели опухоли каждой мыши подкожно инокулировали в правый бок 0,1 мл 2 × 106 клеток A549 с применением 25 G иглы и шприца, один инокулят на мышь. Мышам не давали анестезию для инокуляции.
[00274] Для измерения объема опухоли и рандомизации размер опухоли измеряли с точностью до 0,1 мм. Объемы опухолей рассчитывали с использованием формулы: объем опухоли=длина x ширина2/2. Объемы опухолей мониторировали дважды в неделю. Как только опухоль достигала приблизительно 350-600 мм3, мышей распределяли в группы с варьирующими временными точками. В тот же день тестируемые препараты вводили в соответствии с режимом дозирования.
[00275] Для введения дозировки в тот день, когда образовавшиеся опухоли достигали приблизительно 350-600 мм3, тестируемые препараты извлекали из 4°C холодильника. До введения в шприцы флакон, содержащий состав, несколько раз переворачивали руками, чтобы получить гомогенный раствор.
[00276] Для массы тела, мышей взвешивали с точностью до 0,1 г. Массы тела мониторировали и записывали дважды в неделю, в течение оставшихся недель, включая день завершения исследования.
[00277] Для сбора опухолей животных умерщвляли посредством передозировки CO2 и опухоли рассекали на 0, 24, 48, 72, 96 (необязательно) и 168 часы после получения дозы. Опухоли сначали взвешивали в мокром состоянии, а затем разделяли на три части для KD, распределения и биомаркерного анализа. Образцы мгновенно замораживали в жидком азоте и хранили при -80°C до готовности к дальнейшей обработке.
[00278] Пример 13: GST-π siRNAs по настоящему изобретению ингибировали ксенотрансплантированные опухоли рака поджелудочной железы in vivo. GST-π siRNAs обеспечивали способность нокдауна гена in vivo при введении в липосомальном составе в ксенотрансплантированные опухоли рака поджелудочной железы.
[00279] В этой ксенотрансплантационной модели каждой мыши подкожно инокулировали в правый бок 0,1 мл инокулята 2.5 × 106 клеток PANC-1. Применяли самок бестимусных голых мышей, в возрасте от 6 до 8 недель из Charles River. Размер опухоли измеряли с точностью до 0,1 мм. Как только образовавшиеся опухоли достигали приблизительно 150-250 мм3 (средний объем опухоли приблизительно 200 мм3), мышей распределяли в различные контрольную группу, получающую носитель, и лечебную группу, так чтобы средние объемы опухолей в лечебных группах находились в пределах 10% от среднего объема опухоли в контрольной группе, получающей носитель. В тот же день тестируемые препараты и контрольный носитель вводили в соответствии с режимом дозирования. Объемы опухолей мониторировали три раза в неделю 1, дважды в течение оставшихся недель, включая день завершения исследования.
[00280] На фиг. 6 показана эффективность ингибирования опухоли для GST-π siRNA (SEQ ID NO:61 и 126). Как показано на фиг. 6, эффект дозы получали с применением доз в диапазоне от 0.375 мг/кг до 3 мг/кг siRNA, направленной на GST-π. GST-π siRNA продемонстрировала значительную и непредвиденно успешную эффективность ингибирования опухоли в течение нескольких дней после введения. Таким образом, GST-π siRNA продемонстрировала значительную и непредвиденно успешную эффективность ингибирования опухоли в конечной точке.
[00281] GST-π siRNAs вводили в липосомальном составе, содержащем композицию (ионизируемый липид: холестерин: DOPE: DOPC: DPPE-PEG-2K) (25:30:20:20:5).
[00282] Пример 14: GST-π siRNAs по настоящему изобретению демонстрировал повышенную стабильность в сыворотке.
[00283] На фиг. 7 показана инкубация в сыворотке человека и детекция оставшейся siRNA в различные моменты времени посредством HPLS/LCMS. Как показано на фиг. 7, время полужизни (t½) в сыворотке для обеих смысловой цепи (фиг. 7, top) и антисмысловой цепи (фиг. 7, снизу) GST-π siRNA (SEQ ID NO:61 и 126) составляла приблизительно 100 минут.
[00284] Пример 15: GST-π siRNAs по настоящему изобретению демонстрировала повышенную стабильность в составе в плазме.
[00285] На фиг. 8 показана инкубация состава в плазме и детекция оставшейся siRNA в различные моменты времени. Как показано на фиг. 8, время полужизни (t½) в плазме состава GST-π siRNA (SEQ ID NO:61 и 126) была значительно дольше, чем 100 часов.
[00286] GST-π siRNA получали в липосомальном составе, содержащем композицию (ионизируемый липид: холестерин: DOPE: DOPC: DPPE-PEG-2K) (25:30:20:20:5). Z-средний размер для липосомальных наночастиц составлял 40.0 нм, и siRNA была инкапсулирована на 91%.
[00287] Состав инкубировали в 50% сыворотке человека в PBS в течение 40 минут, 1,5 часов, 3 часов, 24 часов и 96 часов. Количество GST-π siRNA определяли посредством анализа на основе ELISA.
[00288] Пример 16: GST-π siRNAs по настоящему изобретению демонстрировали сниженные нецелевые эффекты сопровождающей цепи.
[00289] Для GST-π siRNA (SEQ ID NO:156 и 182) на фиг. 9 показано, что in vitro нокдаун для направляющей цепи был приблизительно экспоненциальным по сравнению с контролем с последовательностью со случайной перестановкой, которая не демонстрировала никакого эффекта. IC50 этой siRNA измеряли при 5 пМ. На фиг. 10 показан in vitro нокдаун для сопровождающей цепи той же GST-π siRNA. Как показано на фиг. 10, нецелевой нокдаун сопровождающей цепи для GST-π siRNA был значительно снижен, более чем в 100 раз.
[00290] Для GST-π siRNAs (SEQ ID NO:187 и 199), (SEQ ID NO:189 и 201) и (SEQ ID NO:190 и 202) на фиг. 11 показано, что in vitro нокдауны для направляющих цепей были приблизительно экспоненциальными. IC50 этих siRNAs измеряли при 6, 7 и 5 пМ, соответственно. Как показано на фиг. 12, in vitro нокдауны для сопровождающей цепи этих GST-π siRNAs были значительно уменьшены, по меньшей мере, в 10 раз. Все эти GST-π siRNAs имели дезоксинуклеотиды в затравочной области дуплексной области, без других модификаций в дуплексной области.
[00291] Для GST-π siRNAs (SEQ ID NO:217 и 232), на фиг. 13 показано, что in vitro нокдаун для направляющей цепи высоко активной GST-π siRNA был приблизительно экспоненциальным. IC50 этой siRNA измеряли при 11 пМ. Как показано на фиг. 14, in vitro нокдаун для сопровождающей цепи этой GST-π siRNA был значительно снижен более, чем в 100 раз. Эта GST-π siRNA содержала дезоксинуклеотиды в затравочной области дуплексной области, без других модификаций в дуплексной области.
[00292] Нецелевые эффекты определяли с использованием экспрессионной репортерной плазмиды psiCHECK-2, которая кодирует ген Renilla luciferase. (Dual-Luciferase Reporter Assay System, Promega, Cat#:E1960). Концентрация siRNA составляла обычно 50 пМ. Протокол: день 1, клетки HeLa рассевали от 5 до 7.5×103/100мкл/лунку. День 2, котрансфекция с клеточной конфлюэнтностью приблизительно 80%. День 3, клетки собирали для измерения активности люциферазы. Люциферазную активность измеряли с применением Promega's Luciferase Assay System (E4550), по протоколу производителя.
[00293] Вектор psiCHECK-2 обеспечивал мониторинг изменений экспрессии таргетного гена, слитого с репортерным геном Renilla luciferase. siRNA конструкции клонировали в множественный клонирующую область, и вектор котрансфецировали с siRNA в HeLa клетки. Если специфическая siRNA связывается с мРНК-мишенью и инициирует RNAi процесс, слитая Renilla luciferase: конструкция mRNA расщепляется и затем деградирует, снижая сигнал Renilla luciferase.
[00294] Например, плазмидные вставки siRNAs с BU2ʹ структурой были следующими:
[00295] PsiCHECK-2 (F) плазмидная вставка:
SEQ ID NO.: 285
ctcgag gggcaacTGAAGCCTTTTGAGACCCTGcTgTcccag gcggccgc
[00296] PsiCHECK-2 (R) плазмидная вставка:
SEQ ID NO.: 286
ctcgag cTgggacagCAGGGTCTCAAAAGGCTTCagTTgccc gcggccgc
[00297] Пример 17: GST-π siRNAs по настоящему изобретению демонстрировали преимущественно сниженные miRNA-подобные нецелевые эффекты, которые являются зависящим от затравки, нежелательным, нецелевым сайленсингом гена.
[00298] Было выявлено, что для GST-π siRNAs (SEQ ID NO:156 и 182), (SEQ ID NO:187 и 199), (SEQ ID NO:189 и 201), (SEQ ID NO:190 и 202) и (SEQ ID NO:217 и 232) мимикрирующая miRNA с нецелевой активностью была по существу пренебрежима слабой. Зависящий от затравки, нежелательный, нецелевой сайленсинг генов для этих GST-π siRNAs был, по меньшей мере, в от 10 раз до 100 раз меньше, чем целевая активность направляющей цепи.
[00299] Для тестирования miRNA-связанных нецелевых эффектов в область, соответствующую 3'UTR люциферазной mRNA, были введены от одного до четырех повторов, подходящих к затравке таргетной последовательности, комплементарной всей области, содержащей затравку, положения 1-8 5ʹ конца антисмысловой цепи, но не в оставшуюся не затравочную область, положения 9-21, чтобы определить эффективность зависящих от затравки, нежелательных нецелевых эффектов. Плазмидные вставки применяли, чтобы имитировать miRNA с полным совпадением в затравочной области и несовпадениях (петлях) в незатравочной области.
[00300] Например, плазмидные вставки для siRNAs с BU2ʹ структурой были следующими:
[00301] PsiCHECK-2 (Fmi1) плазмидная вставка:
SEQ ID NO.: 287
ctcgag gggcaacTCTACGCAAAACAGACCCTGcTgTcccag gcggccgc
[00302] PsiCHECK-2 (Fmi2) плазмидная вставка:
SEQ ID NO.: 288
ctcgag gggcaacTCTACGCAAAACAGACCCTGcT CTACGCAAAACAGACCCTGcT gTcccag gcggccgc
[00303] PsiCHECK-2 (Fmi3) плазмидная вставка:
SEQ ID NO.: 289
ctcgag gggcaacTCTACGCAAAACAGACCCTGcT CTACGCAAAACAGACCCTGcT CTACGCAAAACAGACCCTGcT gTcccag gcggccgc
[00304] PsiCHECK-2 (Fmi4) плазмидная вставка:
SEQ ID NO.: 290
ctcgag gggcaacTCTACGCAAAACAGACCCTGcT CTACGCAAAACAGACCCTGcT CTACGCAAAACAGACCCTGcT CTACGCAAAACAGACCCTGcT gTcccag gcggccgc
[00305] Варианты осуществления, описываемые в настоящем документе, не являются ограничивающими и специалист в данной области с легкостью может оценить, что специфические комбинации модификаций, описываемых в настоящем документе, могут быть протетстированы без излишнего экспериментирования с целью идентификации молекул нуклеиновой кислоты с улучшенной RNAi активностью.
[00306] Все публикации, патенты и литература конкретно упоминаемые в данном документе, включены в качестве ссылок полностью для всех целей.
[00307] Следует понимать, что данное изобретение не ограничено конкретной описанной методологией, протоколами, материалами и реагентами, так как они могут варьировать. Также следует понимать, что терминология, использованная здесь, предназначена только для описания конкретных вариантов осуществления, и не предназначена ограничивать объем настоящего изобретения. Специалисту в данной области очевидно, что можно варьировать замещения и модификации, описываемые в настоящем документе, без отклонения от объема и духа описания изобретения, и что эти варианты осуществления находятся в объеме этого описания и приложенной формулы изобретения.
[00308] Следует отметить, что, как применяют в настоящем документе и в прилагаемой формуле изобретения, формы единственного числа "a", "an", и "the" включают отсылки к множественному числу, если из контекста явно не следует иначе. Кроме того, термины "a" (или "an"), "один или более" и "по меньшей мере, один" можно использовать взаимозаменяемо в данном документе. Также следует отметить, что термины ʺсодержитʺ, "содержащий", ʺвключающийʺ, "включая", и "имеющий" можно использовать взаимозаменяемо, и следует читать экспансивно и без ограничения.
[00309] Перечисление диапазонов величин в данном документе предназанчено исключительно, чтобы служить в качестве сокращенного метода отсылки индивидуально к каждому отдельному значению, входящему в диапазон, если не указано иначе в данном докменте, и каждая отдельная величина включена в описание изобретения, как если бы они были индивидуально перечислены в настоящем документе. Для групп Маркуша, специалисты в данной области поймут, что описание включает индивидуальных членов, а также подгруппы членов группы Маркуша.
[00310] Без дополнительной обработки, полагают, что специалист в данной области может на основе приведенного выше описания изобетения использовать настоящее изобретение в его полном размере. Следующие специфические варианты осуществления по этой причине следует толковать исключительно как иллюстративные, и не ограничивающие оставшуюся часть изобретения каким-либо образом.
[00311] Все признаки, раскрытые в данном описании изобретения, можно комбинировать в любой комбинации. Каждый признак, раскрытый в данном описании, может быть замещен альтернативным признаком, служащим для той же, эквивалентной или подобной цели.
Реферат
Изобретение относится к биотехнологии. Описана siRNA для ингибирования экспрессии GST-π, содержащая смысловую цепь и антисмысловую цепь, где цепи образуют дуплексную область, и где антисмысловая цепь представляет собой SEQ ID NO:157, и смысловая цепь представляет собой SEQ ID NO:131, где длина каждой цепи составляет 30 нуклеотидов или менее. Также описана композиция, содержащая указанную siRNA. Раскрыт способ лечения заболевания, связанного с экспрессией GST-π. Также раскрыта siRNA для ингибирования экспрессии GST-π, содержащая смысловую цепь и антисмысловую цепь, где цепи образуют дуплексную область, и где антисмысловая цепь представляет собой SEQ ID NO:195, и смысловая цепь представляет собой SEQ ID NO:183, где длина каждой цепи составляет 30 нуклеотидов или менее. Раскрыты соответствующие фармацевтические композиции. Изобретение можно использовать в способах для предотвращения или лечения заболевания, такого как злокачественная опухоль. 16 н. и 49 з.п. ф-лы, 14 ил., 14 табл., 17 пр.
Комментарии