Конъюгаты пептида melan-a, аналога вирусоподобной частицы - RU2351362C2
Код документа: RU2351362C2
Чертежи


Описание
В данной заявке предлагается улучшенный вариант изобретения, описанный в предварительной заявке на выдачу патента США No. 60/457348, поданной 26 марта 2003, которая, таким образом, включена здесь в виде ссылки в полном объеме.
УРОВЕНЬ ТЕХНИКИ
Область техники, к которой относится изобретение
Настоящее изобретение относится к областям вакцинологии, иммунологии и медицины. Изобретение относится к композициям и способам усиления иммунологических ответов против аналогов пептида Melan-A, которые связывают, сливают или присоединяют иным образом с вирусоподобными частицами (VLP) посредством связывания, предпочтительно посредством упаковки иммуностимулирующих веществ, в частности иммуностимулирующих нуклеиновых кислот, и еще более конкретно олигонуклеотидов, содержащих по меньшей мере одну неметилированную последовательность CpG, в VLP. Изобретение можно применять для индукции сильных и продолжительных T-клеточных ответов, особенно полезных при лечении опухолей.
Предшествующий уровень техники
Существование иммунной системы построено на двух отдельных фундаментальных основах: одна из них представляет собой специфичный, или адаптивный, иммунитет, который характеризуется относительно медленной кинетикой ответа и способностью к запоминанию; другая представляет собой неспецифичный, или врожденный, иммунитет, имеющий быструю кинетику ответа, но при отсутствии памяти.
Хорошо установлено, что введения очищенных белков отдельно обычно недостаточно, чтобы вызвать сильный иммунный ответ; изолированный антиген, как правило, должен быть введен вместе с вспомогательными веществами, называемыми адъювантами. В указанных адъювантах введенный антиген защищен от быстрой деградации, и адъювант обеспечивает длительное высвобождение низкого уровня антигена.
В отличие от изолированных белков вирусы индуцируют быстрые и эффективные иммунные ответы в отсутствие какого-либо адъюванта как с помощью, так и без помощи T-клеток (Bachmann and Zinkernagel, Ann. Rev. Immunol. 15: 235-270 (1997)). Многие вирусы имеют квазикристаллическую поверхность, на которой представлена регулярная матрица эпитопов, которые эффективно перекрестно связывают специфичные по отношению к эпитопам иммуноглобулины на B-клетках (Bachmann и Zinkernagel, Immunol. Today 17: 553-558 (1996)). Структура вируса даже связана с образованием анти-антител при аутоиммунном заболевании и является частью естественного ответа на патогены (смотри Fehr, T. , et al., J. Exp. Med. 185: 1785-1792 (1997)). Таким образом, антигены на вирусных частицах, которые организованы в виде упорядоченной и повторяющейся матрицы, являются высокоиммуногенными, так как они могут непосредственно активировать B-клетки и индуцировать формирование ответа цитотоксических T-клеток, другой ключевой ветви иммунной системы.
Вирусные частицы в качестве антигенов имеют два преимущества по сравнению с их изолированными компонентами: (1) благодаря высокоповторяющейся структуре своей поверхности они способны непосредственно активировать B-клетки, приводя к высоким титрам антител и долговременной B-клеточной памяти; и (2) вирусные частицы, но не растворимые белки, имеют возможность индуцировать ответ цитотоксических T-клеток, даже если вирусы являются неинфекционными и адъюванты отсутствуют.
Кроме того, ДНК, богатая неметилированными мотивами CG (CpG), которые присутствуют в бактериях и у большинства беспозвоночных проявляет высокую стимулирующую активность по отношению к B-клеткам, дендритным клеткам и другим APC in vitro, а также in vivo. Хотя бактериальная ДНК является иммуностимулирующей у многих видов позвоночных, индивидуальные мотивы CpG могут отличаться. В действительности мотивы CpG, которые стимулируют иммунные клетки мышей, не обязательно стимулируют иммунные клетки человека и наоборот. Кроме того, иммуностимулирующие CpG-олигодезоксинуклеотиды индуцируют сильные побочные эффекты, вызывая экстрамедуллярный гемопоэз, сопровождаемый спленомегалией и лимфаденопатией у мышей (Sparwasser et al., J. Immunol. (1999), 162: 2368-74 и пример 18).
В последнее время были достигнуты значительные успехи в методике вакцинации, и все же остается необходимость в усовершенствовании существующих методик. В частности в данной области остается необходимость в разработке новых и улучшенных вакцин, которые стимулируют сильный иммунный ответ CTL и защиту против патогенов также эффективно, как природные патогены, в отсутствие генерализованной активации APC и других клеток.
Меланомы являются агрессивными, часто метастазирующими опухолями, происходящими либо из меланоцитов, либо родственных меланоцитам клеток невуса. Меланомы составляют примерно три процента всех злокачественных опухолей кожи, и увеличение распространенности меланомы во всем мире не превосходит ни одна неоплазма за исключением рака легкого у женщин. Даже когда меланома очевидно локализована в коже, у 30% пациентов будут развиваться системные метастазы и большинство пациентов погибнет. В последнее десятилетие появились иммунотерапия и генная терапия в качестве новых и многообещающих способов лечения меланомы, например лечение пациентов с меланомой пептидом Melan-A/MART-1 с адъювантами или без них. Успех указанных методик обычно ограничен. Кроме того, в большинстве исследований не измеряли непосредственно ex vivo ответ CTL с мультимерами MHC класса I, а использовали культуры CTL и стимулировали их в течение нескольких недель перед тем, как в конце концов могли измерить специфичный по отношению к Melan-A CTL-ответ. В общем, пептиды не являются иммуногенными сами по себе и имеют очень короткий период полужизни.
Другим способом иммунотерапии является либо нагрузка дендритных клеток пептидом Melan-A/MART-1, либо трансфекция дендритных клеток Melan-A/MART-1-РНК и повторная инъекция пациентам. Недостатком указанного способа является очистка и инкубация аутологичных дендритных клеток от каждого отдельного пациента в течение нескольких дней с цитокинами in vitro. Это необходимо делать очень осторожно, так как дендритные клетки должны быть в подходящем состоянии созревания, чтобы быть иммуногенными, а не толерогенными, что может привести к тому, что T-клетки больше не будут отвечать на опухоль.
Другим способом Dudley, M. E. (Science. 2002 Oct 25; 298 (5594): 850-4) является выделение Melan-A-специфичных T-клеток из аутологичного опухолевого материала пациентов, культивирование in vitro и размножение и повторная инъекция донору. Как в указанном выше способе, специфичная вакцина должна быть получена отдельно для каждого отдельного пациента, и поэтому не является наиболее эффективной терапией.
Поэтому характеристика эффективных вакцин против меланомы важна для разработки новой методики иммунотерапии злокачественных опухолей, в частности меланомы.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение основано на обнаружении того, что отдельные аналоги пептида Melan-A человека в том случае, когда они связаны с коровой частицей, имеющей структуру с присущей ей повторяющейся организацией, и таким образом, в частности с вирусоподобными частицами (VLP) и субъединицами VLP соответственно, и указанные VLP упакованы с иммуностимулирующими веществами (ISS), такими как ДНК-олигонуклеотиды, представляют собой сильные иммуногены для индукции специфичных антител. Изобретение, кроме того, основано на обнаружении того, что иммуностимулирующие вещества, такие как ДНК-олигонуклеотиды, могут быть упакованы в VLP, которые делают их более иммуногенными. Неожиданно нуклеиновые кислоты и олигонуклеотиды соответственно, присутствующие в VLP, могут быть, в частности, заменены иммуностимулирующими веществами и ДНК-олигонуклеотидами, содержащими мотивы CpG соответственно. Неожиданно указанные упакованные иммуностимулирующие вещества, в частности иммуностимулирующие нуклеиновые кислоты, такие как неметилированные CpG-содержащие олигонуклеотиды, сохраняли свою иммуностимулирующую способность без обширной активации врожденной иммунной системы. Композиции, содержащие VLP и иммуностимулирующие вещества согласно настоящему изобретению, и в частности CpG-VLP, значительно более иммуногенны, чем их аналоги, не содержащие CpG, и индуцируют повышенные B- и T-клеточные ответы. Кроме того, T-клеточные ответы как против VLP, так и против аналогов пептида Melan-A, главным образом, направлены к типу Th1. Поэтому аналоги пептида Melan-A человека, связанные с VLP, нагруженными CpG, могут быть идеальными вакцинами для профилактической и терапевтической вакцинации против опухолей.
В первом варианте изобретение относится к композиции обычно и предпочтительно для усиления иммунного ответа у животного, содержащей вирусоподобную частицу, иммуностимулирующее вещество, предпочтительно иммуностимулирующую нуклеиновую кислоту и еще более предпочтительно неметилированный CpG-содержащий олигонуклеотид, и по меньшей мере один антиген или антигенную детерминанту, где иммуностимулирующее вещество, нуклеиновая кислота или олигонуклеотид связаны, слиты или иным образом присоединены или заключены в вирусоподобную частицу, например связаны с вирусоподобной частицей, и где указанный антиген или антигенная детерминанта связаны с указанной вирусоподобной частицей, и где указанный антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека.
В предпочтительном варианте изобретения иммуностимулирующие нуклеиновые кислоты, в частности неметилированные CpG-содержащие олигонуклеотиды, стабилизированы фосфоротиоатными модификациями фосфатного остова. В другом предпочтительном варианте иммуностимулирующие нуклеиновые кислоты, в частности неметилированные CpG-содержащие олигонуклеотиды, упакованы в VLP посредством расщепления РНК в VLP и одновременного добавления ДНК-олигонуклеотидов, содержащих предпочтительные CpG. В равной мере предпочтительном варианте VLP могут быть подвергнуты разборке перед их повторной сборкой в присутствии CpG.
В следующем предпочтительном варианте иммуностимулирующие нуклеиновые кислоты не содержат мотивов CpG, но несмотря на это проявляют иммуностимулирующие активности. Такие нуклеиновые кислоты описаны в WO 01/22972. Все последовательности, описанные в указанной заявке, таким образом, включены в виде ссылки.
В следующем предпочтительном варианте вирусоподобная частица является рекомбинантной вирусоподобной частицей. Также предпочтительная вирусоподобная частица не содержит липопротеидной оболочки. Предпочтительно рекомбинантная вирусоподобная частица содержит, или альтернативно состоит из рекомбинантных белков вируса гепатита B, вируса BK или другого вируса полиомы человека, вируса кори, вируса Синдбис, ротавируса, вируса ящура, ретровируса, вируса Норуолк или вируса папилломы человека, РНК-фагов, Qβ-фага, GA-фага, fr-фага и Ty. В конкретном варианте вирусоподобная частица содержит, или альтернативно состоит из одного или нескольких различных белков кора(капсида) вируса гепатита B (HBcAg). В следующем предпочтительном варианте вирусоподобная частица содержит рекомбинантные белки или их фрагменты РНК-фага. Предпочтительными РНК-фагами являются фаг Qβ, фаг AP 205, фаг GA, фаг fr.
В конкретном варианте антиген содержит, или альтернативно состоит из эпитопа для цитотоксической T-клетки. В родственном варианте вирусоподобная частица содержит коровый белок вируса гепатита B, и эпитоп для цитотоксических T-клеток слит с C-концом указанного корового белка вируса гепатита B. В одном варианте они слиты лейциновой связывающей последовательностью. В особенно предпочтительном варианте антиген является полипептидом, подходящим для индукции иммунного ответа против клеток злокачественной опухоли.
В другом аспекте изобретения предлагается способ усиления иммунного ответа у человека или других видов животных, включающий в себя введение животному композиции, содержащей вирусоподобную частицу, иммуностимулирующее вещество, предпочтительно иммуностимулирующую нуклеиновую кислоту, и еще более предпочтительно неметилированный CpG-содержащий олигонуклеотид, и по меньшей мере один антиген или антигенную детерминанту, где иммуностимулирующие вещество, предпочтительно нуклеиновая кислота и еще более предпочтительно олигонуклеотид, связан (например, связан, присоединен или заключен) с вирусоподобной частицей, и где указанный антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека, и где указанный аналог пептида Melan-A меланомы человека связан с указанной вирусоподобной частицей.
В еще одном варианте изобретения композицию вводят животному подкожно, внутримышечно, интраназально, интрадермально, внутривенно и непосредственно в лимфатический узел. В равной мере предпочтительном варианте усиливающую иммунитет композиция применяют местно вблизи опухоли или локального резервуара вирусной инфекции, против которой хотят провести вакцинацию.
В предпочтительном аспекте изобретения иммунный ответ представляет собой T-клеточный ответ, и T-клеточный ответ против антигена усиливают. В конкретном варианте T-клеточный ответ представляет собой ответ цитотоксических T-клеток, и ответ цитотоксических T-клеток против пептида Melan-A усиливают.
Настоящее изобретение также относится к вакцине, содержащей иммунологически эффективное количество усиливающей иммунитет композиции согласно настоящему изобретению вместе с фармацевтически приемлемым разбавителем, носителем или эксципиентом. В предпочтительном варианте вакцина, кроме того, содержит по меньшей мере один адъювант. Изобретение также относится к способу иммунизации и/или лечения животного, включающему в себя введение животному иммунологически эффективное количество предложенной вакцины.
В предпочтительном варианте изобретения, содержащие иммуностимулирующие вещество VLP, предпочтительно VLP, содержащие иммуностимулирующую нуклеиновую кислоту, еще более предпочтительно VLP, содержащие неметилированный CpG-содержащий олигонуклеотид, используют для вакцинации животных, обычно и предпочтительно человека, против меланомы или пептидов Melan-A соответственно. Модифицированные VLP обычно и предпочтительно можно использовать для вакцинации против опухолей. Вакцинация может быть в профилактических или терапевтических целях или и в тех и других целях.
Путь инъекции предпочтительно является подкожным или внутримышечным, но также можно применять CpG-содержащие VLP интрадермально, интраназально, внутривенно или непосредственно в лимфатический узел. В равной мере предпочтительном варианте CpG-содержащие связанные с аналогом пептида Melan-A или свободные VLP применяют местно вблизи опухоли или локального резервуара вирусной инфекции, против которой хотят провести вакцинацию.
Должно быть понятно, что и приведенное выше общее описание, и следующее подробное описание являются только примерными и поясняющими и предназначены для дополнительного разъяснения заявленного изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ/ФИГУР
На фигуре 1 показан анализ в SDS-ПААГ VLP Qb-Melan-A. Пептиды Melan-A связывали с VLP Qb, как описано в примере 20. Конечные продукты смешивали с буфером для образцов и разделяли в восстанавливающих условиях в 16% гелях Novex® с трис-глицином в течение 1,5 часа при 125 В. Разделенные белки красили, вымачивая гель в растворе Кумасси синего. Фоновое окрашивание удаляли промывкой геля в 50% метаноле, 8% уксусной кислоте. Маркер молекулярной массы (P 77085, New England BioLabs, Beverly, USA) использовали в качестве эталона скорости миграции Qb-Melan-A (дорожка 1). 14 мкг либо Qb отдельно (дорожка 2), либо Qb, дериватизованного SMPH (дорожка 3) наносили для сравнения с 8 мкг каждого конечного продукта: Qb-Melan-A 16-35 (дорожка 4), Qb-Melan-A 16-35 A/L (дорожка 5), Qb-Melan-A 26-35 (дорожка 6) и Qb-Melan-A 26-35 A/L (дорожка 7).
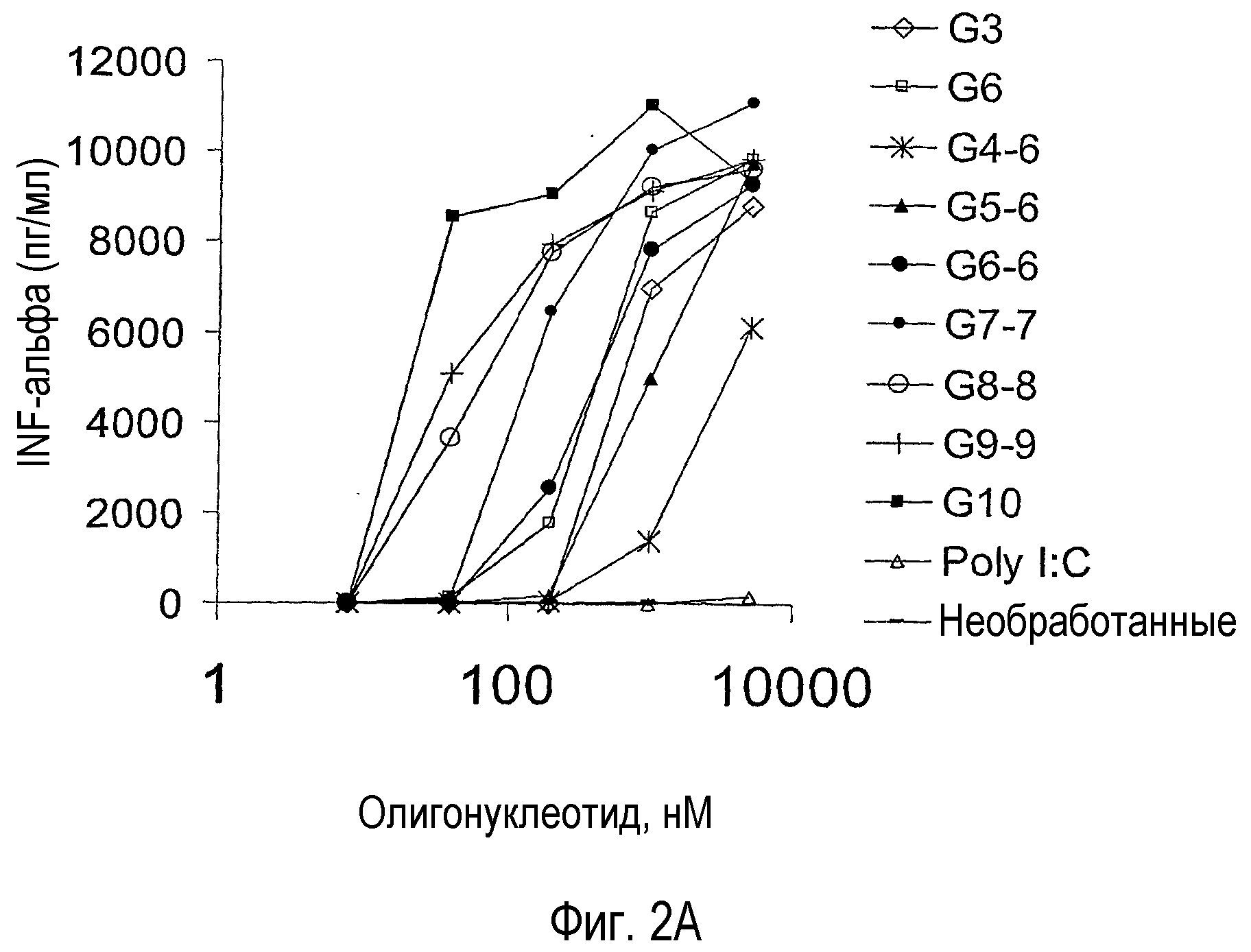
На фигуре 2A показан IFN-альфа, высвобождаемый в надосадки ISS-обработанных PBMC человека. PBMC получали из лейкоцитарной пленки и инкубировали с пятикратным разведением указанного ISS в течение 18 час. Обозначение G10 используют для олигонуклеотида G10-PO, а обозначение G3 используют для олигонуклеотида G3-6. Собирали надосадки и измеряли IFN-альфа в ELISA, используя набор антител, поставляемый PBL Biomedical Laboratories.
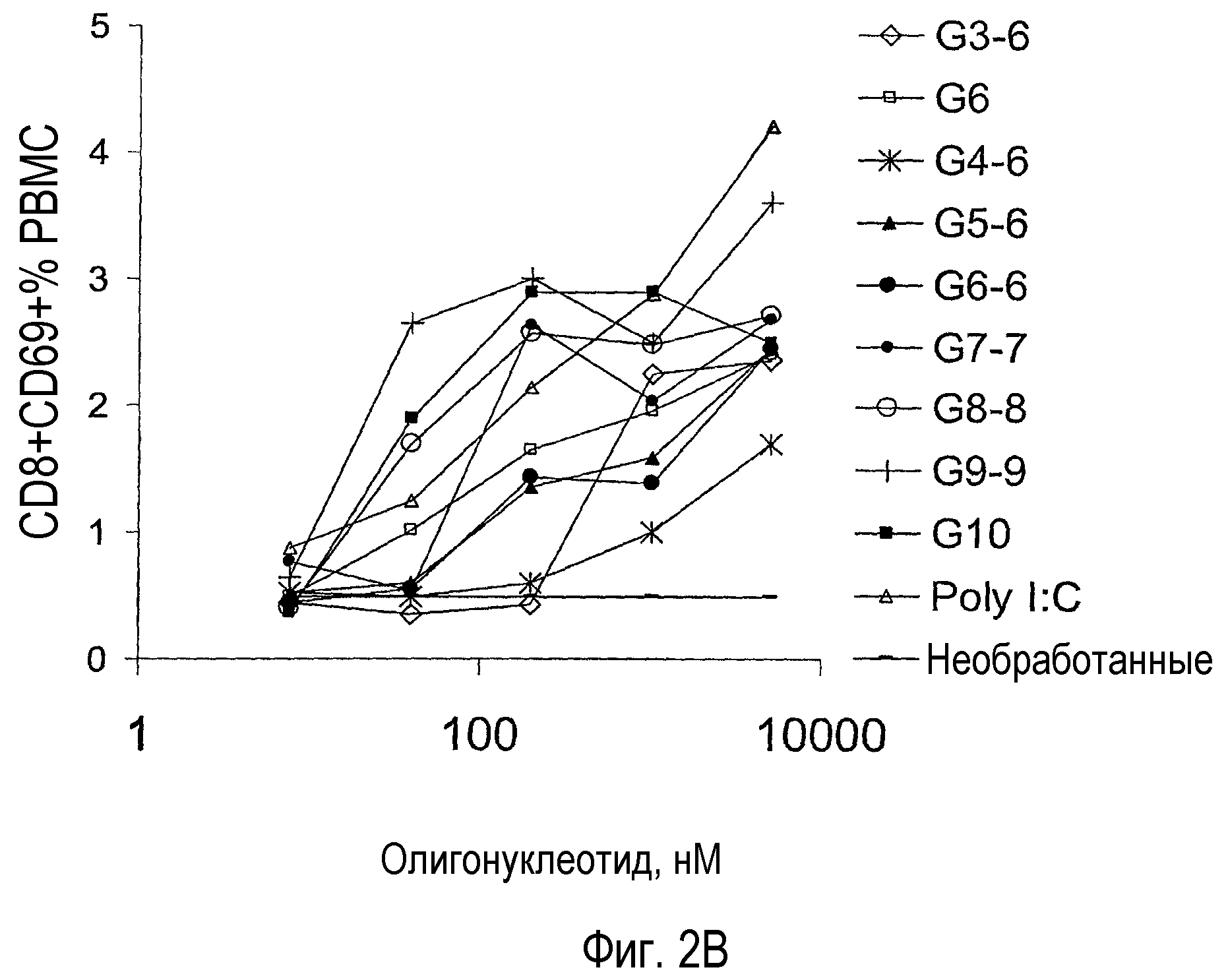
На фигуре 2B показана повышающая регуляция CD69 на PBMC CD8+ человека, обработанных ISS. PBMC получали из лейкоцитарной пленки и инкубировали с пятикратным разведением указанного ISS в течение 18 час. Клетки промывали и инкубировали с анти-CD8-ФИТЦ, анти-CD19-PE и анти-CD69-APC (все из BD PharMingen) в течение 20 мин на льду. После промывки клетки анализировали на приборе FACS Calibur, используя компьютерную программу CellQuest.
На фигуре 3 показаны титры вируса после иммунизации мышей Qbx33, упакованным с поли(I:C), G3-6 или G6. Мышей C57Bl6 иммунизировали посредством инъекции либо 100 мкг Qbx33, 100 мкг Qb VLP, упакованных с поли(I:C) и связанных с p33 (Qb-pIC-33, также называемые QbxZnxполиICxp33GGC), 90 мкг Qbx33, упакованных с G3-6 (Qbx33/G3-6), либо 90 мкг Qbx33, упакованных с G6 (Qbx33/G6). Через восемь дней мышей контрольно заражали 1,5×106 бляшкообразующих единиц вируса вакцинии, несущего эпитоп LCMV-p33. Спустя пять дней мышей забивали и собирали яичники. Готовили суспензию отдельных клеток из яичников и добавляли к клеткам BCS40 в серийных разведениях. Спустя один день слой клеток красили раствором, содержащим 50% этанол, 2% формальдегид, 0,8% NaCl и 0,5% кристаллический фиолетовый) и подсчитывали вирусные бляшки.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Если не оговорено особо, все технические и научные термины, используемые в данном описании, имеют такое же значение, которое обычно понимается специалистом в данной области, к которой относится данное изобретение. Хотя на практике или при проверке настоящего изобретения можно использовать любые способы и вещества, сходные или эквивалентные описанным в данной заявке, предпочтительном способы и вещества описаны далее.
1. Определения
Аминокислотный линкер: «аминокислотный линкер» или также называемый в данном описании просто «линкер» в используемом смысле либо связан с антигеном или антигенной детерминантой со вторым сайтом связывания или более предпочтительно уже включает в себя или содержит второй сайт связывания, обычно, но не обязательно - в виде одного аминокислотного остатка, предпочтительно остатка цистеина. Однако термин «аминокислотный линкер» в используемом в данном описании смысле не предполагает, что такой аминокислотный линкер состоит исключительно из аминокислотных остатков, хотя аминокислотный линкер, состоящий из аминокислотных остатков, является предпочтительным вариантом настоящего изобретение. Аминокислотные остатки аминокислотного линкера предпочтительно состоят из встречающихся в природе аминокислот или неприродных аминокислот, известных в данной области, всех L или всех D или их смесей. Однако аминокислотный линкер, содержащий молекулу с сульфгидрильной группой или остаток цистеина, также входит в объем изобретения. Такая молекула предпочтительно содержит остаток C1-C6-алкила-, циклоалкила (C5, C6), арила или гетероарила. Однако кроме аминокислотного линкера линкер, содержащий предпочтительно остаток C1-C6-алкила-, циклоалкила- (C5, C6), арила- или гетероарила- и лишенной какой-либо аминокислоты (аминокислот), также включен в объем изобретения. Связь между антигеном или антигенной детерминантой или необязательно вторым сайтом связывания и аминокислотным линкером предпочтительно осуществляется посредством по меньшей мере одной ковалентной связи, более предпочтительно посредством по меньшей мере одной пептидной связи.
Животное: В используемом в данном описании смысле имеется в виду, что термин «животное» включает, например, человека, овец, лошадей, коров, свиней, собак, кошек, крыс, мышей, млекопитающих, птиц, рептилий, рыб, насекомых и паукообразных.
Антитело: В используемом в данном описании смысле термин «антитело» относится к молекулам, которые способны связывать эпитоп или антигенную детерминанту. Подразумевается, что термин включает целые антитела и их антигенсвязывающие фрагменты, включая одноцепочечные антитела. Наиболее предпочтительно антитела являются антигенсвязывающими фрагментами антител человека и включают, но не ограничены указанным, Fab, Fab' и F(ab')2, Fd, одноцепочечные Fv (scFv), одноцепочечные антитела, связанные дисульфидными связями Fv (sdFv) и фрагменты, содержащие либо VL- либо VH-домен. Антитела могут быть из любого животного источника, включая птиц и млекопитающих. Предпочтительно антитела являются антителами человека, мыши, кролика, козы, морской свинки, верблюда, лошади или цыпленка. В используемом в данном описании смысле «человеческие» антитела включают антитела, имеющие аминокислотную последовательность иммуноглобулина человека, и включают антитела, выделенные из библиотек иммуноглобулинов человека или из животных, трансгенных по одному или нескольким иммуноглобулинам человека и которые не экспрессируют эндогенных иммуноглобулинов, которые, например, описаны в патенте США No. 5939598 Kucherlapati et al.
Антиген: В используемом в данном описании смысле термин «антиген» относится к молекуле, которая может быть связана антителом или рецептором T-клетки (TCR), если она презентирована молекулами MHC. Термин «антиген» в используемом в данном описании смысле также охватывает T-клеточные эпитопы. Антиген, кроме того, может узнаваться иммунной системой и/или способен индуцировать гуморальный иммунный ответ и/или клеточный иммунный ответ, приводящий к активации B- и/или T-лимфоцитов. Однако может требоваться, чтобы по меньшей мере в некоторых случаях антиген содержал или был связан с эпитопом, узнаваемым T-хелперными клетками (Th-клеточный эпитоп), и вводился в адъюванте. Антиген может иметь один или несколько эпитопов (B- и T-эпитопы). Специфичная реакция, указанная выше, предназначена для указания того, что антиген предпочтительно будет взаимодействовать, обычно высокоизбирательно, с соответствующим ему антителом или TCR и не будет взаимодействовать с множеством других антител или TCR, которые могут быть вызваны другими антигенами. Антигены в используемом в данном описании смысле также могут представлять собой смеси нескольких отдельных антигенов.
«Опухолевой антиген» в используемом в данном описании смысле является соединением, таким как пептид, связанным с опухолью или злокачественной опухолью и который способен вызывать иммунный ответ. В частности, соединение способно вызывать иммунный ответ, когда презентировано в контексте молекулы MHC. Опухолевые антигены могут быть получены из клеток злокачественной опухоли либо посредством получения неочищенных экстрактов клеток злокачественной опухоли, например, как описано в Cohen, et al., Cancer Research, 54: 1055 (1994), путем частичной очистки антигенов, рекомбинантной технологией или посредством синтеза de novo известных антигенов. К опухолевым антигенам относятся антигены, которые представляют собой антигенные части или целые полипептиды опухоли или злокачественной опухоли. Такие антигены могут быть выделены или получены рекомбинантно или любыми другими способами, известными в данной области. Злокачественные опухоли и опухоли включают, но не ограничены указанным, рак желчных путей; рак головного мозга; рак молочной железы; рак шейки матки; хориокарциному; рак ободочной кишки; эндометриальный рак; рак пищевода; рак желудка; интраэпителиальные неоплазмы; лимфомы; рак печени; рак легкого (например, мелкоклеточный и немелкоклеточный); меланома; нейробластомы; рак ротовой полости; рак яичника; рак поджелудочной железы; рак простаты; ректальный рак; саркомы; рак кожи; рак семенников; рак щитовидной железы и рак почки, а также другие карциномы и саркомы.
Антигенная детерминанта: В используемом в данном описании смысле подразумевается, что термин «антигенная детерминанта» относится к такой части антигена, которая специфично узнается либо B-, либо T-лимфоцитами. B-лимфоциты отвечают на чужеродные антигенные детерминанты продукцией антител, тогда как T-лимфоциты являются медиаторами клеточного иммунитета. Таким образом, антигенные детерминанты или эпитопы являются такими частями антигена, которые узнаются антителами или в контексте MHC рецепторами T-клеток.
Антигенпрезентирующая клетка: В используемом в данном описании смысле подразумевается, что термин «антигенпрезентирующая клетка» относится к гетерогенной популяции лейкоцитов и клеток, полученных из костного мозга, которые обладают иммуностимулирующей способностью. Например, указанные клетки, к созданию пептидов, связанных с молекулами MHC, которые могут узнаваться T-клетками. Термин является синонимом термина «вспомогательная клетка» и включает, например, клетки Лангерганса, интердигитальные клетки, B-клетки, макрофаги и дендритные клетки. В некоторых условиях эпителиальные клетки, эндотелиальные клетки и другие клетки, не происходящие из костного мозга, также могут служить в качестве антигенпрезентирующих клеток.
Ассоциация: В используемом в данном описании смысле термин «ассоциация» в применении к первому и второму сайтам связывания относится к связыванию первого и второго сайтов связывания, которое предпочтительно осуществляется посредством по меньшей мере одной непептидной связи. Природа ассоциации может быть ковалентной, ионной, гидрофобной, полярной или любой их комбинации, предпочтительно природа ассоциации является ковалентной, и снова более предпочтительно связывание осуществляется по меньшей мере посредством одной, предпочтительно одной непептидной связи. В используемом в данном описании смысле термин «ассоциация» в применении к первому и второму сайтам связывания охватывает не только прямое связывание или ассоциацию первого и второго сайта связывания, образующую композиции согласно изобретению, но также альтернативно и предпочтительно непрямую ассоциацию или связывание первого и второго сайта связывания, приводящее к композициям согласно изобретению, и при этом обычно и предпочтительно с использованием гетеробифункционального перекрестно-связывающего линкера.
Сайт связывания, первый: В используемом в данном описании смысле фраза «первый сайт связывания» относится к элементу неприродного или природного происхождения, обычно и предпочтительно входящему в вирусоподобную частицу, с которым может связываться второй сайт связывания обычно и предпочтительно входящий в аналог пептида Melan-A согласно изобретению. Первый сайт связывания может представлять собой белок, полипептид, аминокислоту, пептид, сахар, полинуклеотид, природный или синтетический полимер, вторичный метаболит или соединение (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металлов, фенилметилсульфонилфторид) или их комбинации, или их химически активную группу. Первый сайт связывания обычно и предпочтительно локализован на поверхности вирусоподобной частицы. Множество первых сайтов связывания присутствуют на поверхности вирусоподобной частицы обычно в повторяющейся конфигурации. Предпочтительно первым сайтом связывания является аминокислота или ее химически активная группа.
Сайт связывания, второй: В используемом в данном описании смысле фраза «второй сайт связывания» относится к элементу ассоциированному, обычно и предпочтительно входящему в аналог пептида Melan-A согласно изобретению, который может связываться с первым сайтом связывания, локализованным на поверхности вирусоподобной частицы. Второй сайт связывания аналога пептида Melan-A согласно изобретению может представлять собой белок, полипептид, пептид, сахар, полинуклеотид, природный или синтетический полимер, вторичный метаболит или соединение (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металлов, фенилметилсульфонилфторид) или их комбинацию, или их химически активную группу. По меньшей мере один второй сайт связывания присутствует на аналоге пептида Melan-A согласно изобретению. Термин «аналог пептида Melan-A» по меньшей мере с одним вторым сайтом связывания» относится к антигену или антигенной конструкции, содержащей по меньшей мере аналог пептида Melan-A согласно изобретению и второй сайт связывания. Однако, в частности, в случае второго сайта связывания, который имеет неприродное происхождение, например искусственно возникшего в аналоге пептида Melan-A согласно изобретению, указанные антиген или антигенные конструкции содержат «аминокислотный линкер».
Связанный: В используемом в данном описании смысле термин «связанный» относится к связыванию, которое может быть ковалентным, например, посредством химического связывания, или нековалентным, например ионные взаимодействия, гидрофобные взаимодействия, водородные связи и т.д.. Ковалентными связями могут быть, например, сложноэфирные, простые эфирные, фосфоэфирные, амидные, пептидные, имидные связи, связи углерод-сера, связи углерод-фосфор и тому подобные. Термин «связанный» является более широким и включает в себя такие термины как «соединенный», «слитый», «ассоциированный» и «присоединенный». Кроме того, по отношению к иммуностимулирующему веществу, связанному с вирусоподобной частицей, термин «связанный» также включает в себя заключение иммуностимулирующего вещества внутрь или его частичное заключение внутрь. Таким образом, по отношению к иммуностимулирующему веществу, связанному с вирусоподобной частицей, термин «связанный» является более широким и включает в себя такие термины, как «соединенный», «слитый», «заключенный», «упакованный» или «присоединенный». Например, иммуностимулирующее вещество, такое как неметилированный CpG-содержащий олигонуклеотид, может быть заключено в VLP без наличия действительного связывания, как ковалентного, так и нековалентного.
Белок(ки) оболочки: В используемом в данном описании смысле термин «белок(ки) оболочки»" относится к белку(кам) бактериофага или РНК-фага, который может быть включен в сборку капсида бактериофага или РНК-фага. Однако по отношению к специфичному генному продукту гена белка оболочки РНК-фагов используют термин «CP». Например, специфичный генный продукт гена белка оболочки РНК-фага Qβ называют «CP Qβ», в то время как «белки оболочки» бактериофага Qβ содержат «CP Qβ», а также белок A1. Капсид бактериофага Qβ, главным образом, содержит CP Qβ с минорным содержанием белка A1. Подобным образом белок оболочки VLP Qβ содержит, главным образом, CP Qβ с минорным содержанием белка A1.
Соединенный: В используемом в данном описании смысле термин «соединенный» относится к связыванию ковалентными связями или нековалентными взаимодействиями. Что касается соединения антигена с вирусоподобной частицей, термин «соединенный» предпочтительно относится к связыванию ковалентными связями. Кроме того, в отношении соединения антигена с вирусоподобной частицей термин «соединенный» предпочтительно относится к ассоциации и связыванию соответственно по меньшей мере одной пептидной связью. Любой способ, обычно используемый специалистами в данной области для соединения биологически активных веществ, можно использовать в данном изобретении.
Слияние: В используемом в данном описании смысле термин «слияние» относится к комбинации аминокислотных последовательностей разного происхождения в одной полипептидной цепи посредством комбинирования в рамке их кодирующих нуклеотидных последовательностей. Термин «слияние» особо охватывает внутренние слияния, т.е. инсерцию последовательностей другого происхождения в полипептидную цепь, кроме слиянии с одним из его концов.
CpG: В используемом в данном описании смысле термин «CpG» относится к олигонуклеотиду, который содержит по меньшей мере одну неметилированную последовательность динуклеотидов цитозин-гуанин (например, «CpG-ДНК» или ДНК, содержащая цитозин, за которым следует гуанозин, и связан фосфатной связью) и стимулирует/активирует, например, оказывает митогенное действие или индуцирует или увеличивает экспрессию цитокинов клеткой позвоночного. Например, CpG могут быть применимы для активации B-клеток, NK-клеток и антигенпрезентирующих клеток, таких как моноциты, дендритные клетки и макрофаги и T-клетки. CpG могут включать аналоги нуклеотидов, такие как аналоги, содержащие фосфотиоэфирные связи, и могут быть двунитевыми или однонитевыми. Как правило, двунитевые молекулы являются более стабильными in vivo, в то время как однонитевые молекулы обладают повышенной иммунной активностью.
Эпитоп: В используемом в данном описании смысле термин «эпитоп» относится к частям полипептида, обладающим антигенной или иммуногенной активностью у животного, предпочтительно млекопитающего и наиболее предпочтительно у человека. «Иммуногенный эпитоп» в используемом в данном описании смысле определяют как часть полипептида, которая вызывает гуморальный ответ или индуцирует T-клеточный ответ у животного, который определяют любым способом, известным в данной области (смотри, например, Geysen et al., Proc. Natl. Acad. Sci. USA 81: 3998 4002 (1983)). Термин «антигенный эпитоп» в используемом в данном описании смысле определяют как часть белка, в которой антитело может иммуноспецифично связывать свой антиген, которую определяют любым способом, хорошо известным в данной области. Иммуноспецифичное связывание исключает неспецифичное связывание, но не обязательно исключает перекрестную реактивность с другими антигенами. Антигенные эпитопы не обязательно должны быть иммуногенными. Антигенные эпитопы также могут быть T-клеточными эпитопами, и в данном случае они могут иммуноспецифично связываться рецептором T-клеток в контексте молекулы MHC.
Эпитоп может содержать 3 аминокислоты в пространственной конформации, которая является уникальной для эпитопа. Как правило, эпитоп состоит по меньшей мере примерно из 5 таких аминокислот, и более обычно состоит по меньшей мере примерно из 8-10 таких аминокислот. Если эпитоп является органической молекулой, он может быть таким небольшим, как нитрофенил. Предпочтительными эпитопами являются аналоги пептида Melan-A согласно изобретению.
Иммунный ответ: В используемом в данном описании смысле термин «иммунный ответ» относится к гуморальному иммунному ответу и/или клеточному иммунному ответу, приводящему к активации или пролиферации B- и/или T-лимфоцитов. Однако в некоторых случаях иммунные ответы могут быть низкой интенсивности и становятся выявляемыми только при использовании по меньшей мере одного вещества согласно изобретению. «Иммуногенный» относится к агенту, используемому для стимуляции иммунной системы живого организма, для того чтобы одна или несколько функций иммунной системы были повышены и направлены по отношению к иммуногенному агенту. «Иммуногенный полипептид» представляет собой полипептид, который вызывает клеточный и/или гуморальный иммунный ответ либо отдельно, либо будучи связанным с носителем в присутствии в или отсутствие адъюванта.
Иммунизация: В используемом в данном описании смысле термины «иммунизировать» или «иммунизация» или родственные термины относятся к приданию способности формировать значительный иммунный ответ (включая гуморальный и клеточный иммунитет, такой как эффекторные CTL) против антигена или эпитопа, являющегося мишенью. Указанные термины не требуют, чтобы был создан полный иммунитет, а чтобы был создан иммунный ответ, который существенно выше, чем исходный уровень. Например, млекопитающее можно считать иммунизированным против антигена-мишени, если клеточный и/или гуморальный иммунный ответ на антиген-мишень возникает после применения способов согласно изобретению.
Иммуностимулирующая нуклеиновая кислота: В используемом в данном описании смысле термин иммуностимулирующая нуклеиновая кислота относится к нуклеиновой кислоте, способной индуцировать и/или усиливать иммунный ответ. Иммуностимулирующие нуклеиновые кислоты в используемом в данном описании смысле включают рибонуклеиновые кислоты и особенно дезоксирибонуклеиновые кислоты. Предпочтительно иммуностимулирующие нуклеиновые кислоты содержат по меньшей мере один мотив CpG, например CG-динуклеотид, в котором C неметилирован. Динуклеотид CG может быть частью палиндромной последовательности или может входить в непалиндромную последовательность. Иммуностимулирующие нуклеиновые кислоты, не содержащие мотивов CpG, которые описаны выше, охватывают в качестве примера нуклеиновые кислоты, не имеющие динуклеотидов CpG, а также нуклеиновые кислоты, содержащие мотивы CG с метилированным динуклеотидом CG. Термин «иммуностимулирующая нуклеиновая кислота» в используемом в данном описании смысле следует также относить к нуклеиновым кислотам, которые содержат модифицированные основания, такие как 4-бромцитозин.
Иммуностимулирующее вещество: В используемом в данном описании смысле термин «иммуностимулирующее вещество» относится к веществу, способному индуцировать и/или усиливать иммунный ответ. Иммуностимулирующие вещества в используемом в данном описании смысле включают, но ограничены указанным, вещества, активирующие toll-подобные рецепторы, и вещества, индуцирующие секрецию цитокинов. Вещества, активирующие toll-подобные рецепторы, включают, но не ограничены указанным, иммуностимулирующие нуклеиновые кислоты, пептидогликаны, липополисахариды, липотейхоновые кислоты, соединения имидазохинолина, флагеллины, липопротеиды и иммуностимулирующие органические вещества, такие как таксол.
Термин «природный пептид Melan-A человека» или «нормальный пептид Melan-A человека» в используемом в данном описании смысле следует относить к пептиду, содержащему или альтернативно в основном состоящему или альтернативно состоящему из аминокислотной последовательности EAAGIGILTV (SEQ ID NO: 78), представляющей собой аминокислоты в положениях 26-35 нормальной последовательности белка Melan-A человека, или AAGIGILTV (SEQ ID NO: 79), представляющей собой аминокислоты в положениях 27-35 нормальной последовательности белка Melan-A человека.
Термин «аналог пептида Melan-A», или «аналог пептида Melan-A человека», или «аналог пептида Melan-A меланомы человека» в используемом в данном описании смысле следует определять как пептид, в котором аминокислотная последовательность соответствующего нормального пептида Melan-A изменена по меньшей мере одной аминокислотой или производным аминокислоты, при этом данное изменение может включать в себя замену, и/или делецию, и/или инсерцию аминокислоты или их комбинации. В предпочтительном варианте настоящего изобретения термин «аналог пептида Melan-A» в используемом в данном описании смысле следует определять как пептид, в котором аминокислотная последовательность соответствующего нормального пептида Melan-A (SEQ ID NO: 91) изменена тремя, предпочтительно двумя и еще более предпочтительно одной аминокислотой или производным аминокислоты, при этом указанное изменение может включать в себя замену, и/или делецию, и/или инсерцию аминокислоты или их комбинацию. В следующем предпочтительном варианте настоящего изобретения термин «аналог пептида Melan-A» в используемом в данном описании смысле следует определять как пептид, в котором аминокислотная последовательность соответствующего нормального пептида Melan-A изменена тремя, предпочтительно двумя и еще более предпочтительно одной аминокислотой или производным аминокислоты, где указанное изменение может включать в себя замену, и/или делецию, и/или инсерцию аминокислоты или их комбинацию, и где указанное изменение имеет место в положении 26, 27, 28 и/или 35 нормальной последовательности белка Melan-A (SEQ ID NO: 91), и где указанным изменением предпочтительно является замена аминокислоты. Термины «аналог пептида Melan-A», «аналог пептида Melan-A человека», «аналог пептида Melan-A меланомы человека» и « аналог пептида Melan-A/MART-1 меланомы человека» используют взаимозаменяемо.
Природное происхождение: В используемом в данном описании смысле термин «природное происхождение» означает, что целое или его части не синтезированы, а существуют или получены в природе.
Неприродный: В используемом в данном описании смысле термин в общем означает «не из природы», более конкретно термин означает «сделанный искусственно человеком».
Неприродное происхождение: В используемом в данном описании смысле термин «неприродное происхождение» в общем означает «синтетический» или «не из природы»; более конкретно термин означает «сделанный искусственно человеком».
Упорядоченная и повторяющаяся матрица антигенов или антигенных детерминант: В используемом в данном описании смысле термин «упорядоченная и повторяющаяся матрица антигенов или антигенных детерминант» обычно относится к повторяющемуся образцу антигена или антигенной детерминанты, характеризующемуся обычно и предпочтительно однородным пространственным распределением антигенов или антигенных детерминант по отношению к коровой частице и вирусоподобной частице соответственно. В одном варианте изобретения повторяющийся образец может представлять собой образец геометрической формы. Типичными и предпочтительными примерами подходящих упорядоченных и повторяющихся матриц антигенов или антигенных детерминант являются матрицы, которые имеют строго повторяющиеся паракристаллические порядки антигенов или антигенных детерминант, предпочтительно с промежутками от 0,5 до 30 нанометров, более предпочтительно от 3 до 15 нанометров, еще более предпочтительно от 3 до 8 нанометров.
Олигонуклеотид: В используемом в данном описании смысле термины «олигонуклеотид» или «олигомер» относятся к последовательности нуклеиновой кислоты, содержащей 2 или более нуклеотидов, как правило, по меньшей мере примерно от 6 нуклеотидов до примерно 100000 нуклеотидов, предпочтительно примерно от 6 до примерно 2000 нуклеотидов, и более предпочтительно примерно от 6 до примерно 300 нуклеотидов, еще более предпочтительно примерно от 20 до примерно 300 нуклеотидов и еще более предпочтительно примерно от 20 до примерно 100 нуклеотидов. Термины «олигонуклеотид» или «олигомер» также относятся к последовательности нуклеиновой кислоты, содержащей более чем от 100 до примерно 2000 нуклеотидов, предпочтительно более чем от 100 до примерно 1000 нуклеотидов и более предпочтительно более чем от 100 до примерно 500 нуклеотидов. «Олигонуклеотид» также, как правило, относится к любому полирибонуклеотиду или полидезоксирибонуклеотиду, которые могут представлять собой немодифициронную РНК или ДНК или модифицированную РНК или ДНК. «Олигонуклеотид» включает без ограничения одно- и двунитевую ДНК, ДНК, которая является смесью одно- и двунитевых областей, одно- и двунитевую РНК и РНК, которая является смесью одно- и двунитевых областей, гибридные молекулы, содержащие ДНК и РНК, которые могут быть однонитевыми или более обычно двунитевыми или смесью одно- и двунитевых областей. Кроме того, «олигонуклеотид» относится к трехнитевым областям, содержащим РНК или ДНК, или как РНК, так и ДНК. Кроме того, олигонуклеотид может быть синтетическим, геномным или рекомбинантным, например λ-ДНК, космидная ДНК, искусственная бактериальная хромосома, дрожжевая искусственная хромосома и нитчатый фаг, такой как M13. В очень предпочтительном варианте настоящего изобретения олигонуклеотид является синтетическим олигонуклеотидом.
Термин «олигонуклеотид» также включает ДНК или РНК, содержащие одно или несколько модифицированных оснований, и ДНК или РНК с остовом, модифицированными для стабильности или по другим причинам. Например, подходящие нуклеотидные модификации/аналоги включают пептидонуклеиновую кислоту, инозин, тритилированные основания, фосфоротиоаты, алкилфосфоротиоаты, 5-нитроиндолдезоксирибофуранозил, 5-метилдезоксицитозин и 5,6-дигидро-5,6-дигидроксидезокситимидин. Было осуществлено множество модификаций ДНК и РНК; таким образом, «олигонуклеотид» охватывает химически, ферментативно или метаболически модифицированные формы полинуклеотидов, которые обычно встречаются в природе, а также химические формы ДНК и РНК, характерные для вирусов и клеток. Другие нуклеотидные аналоги/модификации будут очевидны специалистам в данной области.
Упакованное: Термин «упакованное» в используемом в данном описании смысле относится к состоянию иммуностимулирующего вещества, предпочтительно иммуностимулирующей нуклеиновой кислоты по отношению к VLP. Термин «упакованный» в используемом в данном описании смысле включает связывание, которое может быть ковалентным, например, в результате химического связывания, или нековалентным, например ионные взаимодействия, гидрофобные взаимодействия, водородные связи и т.д.. Ковалентными связями могут быть, например, сложноэфирные, простые эфирные, фосфоэфирные, амидные, пептидные, имидные связи, связи углерод-сера, такие как тиоэфирные связи, связи углерод-фосфор и тому подобные. Термин также включает в себя заключение вещества внутри или его частичное заключение внутри. Термин «упакованный» включает в себя такие термины, как «соединенный», «заключенный» и «связанный». Например, иммуностимулирующее вещество, такое как неметилированный CpG-содержащий олигонуклеотид, может быть заключено в VLP в отсутствие действительного связывания, как ковалентного, так и нековалентного. В предпочтительных вариантах, в частности если иммуностимулирующие нуклеиновые кислоты являются иммуностимулирующими веществами, термин «упакованный» указывает, что иммуностимулирующая нуклеиновая кислота в упакованном состоянии не доступна для гидролиза ДНКазой или РНКазой. В предпочтительных вариантах иммуностимулирующая нуклеиновая кислота упакована внутри капсидов VLP, наиболее предпочтительно нековалентно.
Композиции согласно изобретению могут быть необязательно объединены с фармацевтически приемлемым носителем. Термин «фармацевтически приемлемый носитель» в используемом в данном описании смысле означает один или несколько совместимых твердых или жидких наполнителей, разбавителей или инкапсулирующих веществ, которые подходят для введения человеку или другому животному. Термин «носитель» означает органический или неорганический ингредиент, природный или синтетический, с которым объединяют активный ингредиент, чтобы облегчить применение.
Пептид: Термин «пептид» в используемом в данном описании смысле, и в частности по отношению к пептиду Melan-A меланомы человека или нормальному пептиду Melan-A человека, будет относиться к молекуле, состоящей из мономеров (аминокислот), обычно и предпочтительно линейно связанных амидными связями (также называемыми пептидными связями). Термин означает молекулярную цепь аминокислот и не относится к конкретной длине продукта.
Органическая молекула: В используемом в данном описании смысле термин «органическая молекула» относится к любому объекту природного или синтетического происхождения. В частности, термин «органическая молекула» в используемом в данном описании смысле охватывает, например, любую молекулу, являющуюся членом группы нуклеотидов, липидов, углеводов, полисахаридов, липополисахаридов, стероидов, алкалоидов, терпенов и жирных кислот либо природного, либо синтетического происхождения. В частности, термин «органическая молекула» охватывает такие молекулы, как никотин, кокаин, героин или другие фармакологически активные молекулы, входящие в состав наркотических лекарственных средств. В общем органическая молекула содержит или модифицирована так, чтобы она содержала химическую функциональную группу, обеспечивающую возможность сочетания, связывания или присоединения иным образом к вирусоподобной частице согласно изобретению.
Полипептид: В используемом в данном описании смысле термин «полипептид» относится к молекуле, состоящей из мономеров (аминокислот), линейно связанных амидными связями (также называемыми пептидными связями). Термин означает молекулярную цепь аминокислот и не относится к конкретной длине продукта. Таким образом, пептиды, олигопептиды и белки включены в определение полипептид. Подразумевается, что указанный термин также относится к постэкпрессионным модификациям полипептида, например гликозилированию, ацетилированию, фосфорилированию и тому подобному. Рекомбинантный или полученный полипептид не обязательно транслирован с соответствующей последовательности нуклеиновой кислоты. Он также может быть создан другим способом, включая химический синтез.
Вещество, которое «усиливает» иммунный ответ, относится к веществу, к которому наблюдается иммунный ответ, который выше или интенсивнее или отклоняется любым образом при добавлении вещества по сравнению с таким же иммунным ответом, измеренным без добавления вещества.
Предпочтительно вещество, которое «усиливает» иммунный ответ, относится в данном описании (i) к веществу, в случае которого частота встречаемости T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека, увеличивается по сравнению с частотой встречаемости T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека, измеренной без добавления вещества, или (ii) к веществу, в случае которого функциональность T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека, отклоняется, предпочтительно улучшается по сравнению с функциональностью T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека, измеренной без добавления вещества, или (iii) к веществу, в случае которого фенотип T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека отклоняется таким образом, что получаемые в результате T-клетки способны к повышенной пролиферации, сниженному апоптозу или более эффективному хомингу в опухолевые ткани по сравнению с пролиферацией, апоптозом и хомингом в опухолевые ткани T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека, измеренных без добавления вещества.
Частоту встречаемости T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека, измеряют посредством окрашивания комплексов MHC-класс I/пептид, таких как тетрамер или мультимер, предпочтительно посредством окрашивания тетрамера, как описано в Speiser, DE. et al. Eur. J. Immunol. 2002, Vol. 32, 731- 741, тогда как функциональность T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека, измеряют по высвобождению цитокинов, а именно внутриклеточным окрашиванием, анализом цитокином способом «захвата», Elispot, ELISA и предпочтительно с помощью Elispot, как описано в Speiser, D.E. et al. Eur. J. Immunol. 2002, Vol. 32, 731-741. Кроме того, функциональность T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека, также можно измерить посредством измерения ответа специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека цитолитических T-клеток CD8+ в анализе высвобождения хрома или европия, как описано в Valmori, D. et al. J. Immunol. 1998, 161, 6956-6962. Фенотипирование T-клеток, специфичных для Melan-A, предпочтительно специфичных для природного пептида Melan-A человека или специфичных для аналога пептида Melan-A меланомы человека, осуществляют, используя антитела против белков клеточной поверхности или внутриклеточных белков, таких как маркеры активации, маркеры дифференцировки клеток, маркеры хоминга, рецепторы хемокинов и цитокинов, костимулирующие рецепторы, рецепторы гибели, рецепторы, активирующие или ингибирующие киллеры, интегрины, по экспрессии антиапоптозных белков и отсутствию маркеров старения, и предпочтительно посредством маркеров активации клеток, как описано в Speiser, D.E. et al. Eur. J. Immunol. 2002, Vol. 32, 731-741.
Эффективное количество: В используемом в данном описании смысле термин «эффективное количество» относится к количеству, необходимому или достаточному для осуществления требуемого биологического эффекта. Эффективным количеством композиции может быть количество, которое позволяет достигать данного выбранного результата, и такое количество может обычным образом определить специалист в данной области. Например, эффективным количеством для лечения недостаточности иммунной системы может быть такое количество, которое необходимо для того, чтобы вызвать активацию иммунной системы, приводящую к развитию антиген-специфичного иммунного ответа при воздействии антигена. Термин также является синонимом «достаточного количества».
Эффективное количество для любого конкретного применения может варьировать в зависимости от таких факторов, как заболевание или состояние, подвергаемое лечению, конкретная вводимая композиция, масса субъекта и/или тяжесть заболевания или состояния. Специалист в данной области может эмпирически определить эффективное количество конкретной композиции согласно настоящему изобретению без необходимости в чрезмерном экспериментировании.
Аутоантиген: В используемом в данном описании смысле термин «аутоантиген» относится к белкам, кодируемым геномом или ДНК хозяина, и продукты, образуемые белками, или РНК, кодируемую геномом или ДНК хозяина, определяют как собственные. Предпочтительно термин «аутоантиген» в используемом в данном описании смысле относится к белкам, кодируемым геномом или ДНК человека, и продукты, образуемые белками, или РНК, кодируемую геномом или ДНК человека, определяют как собственные. Предлагаемые в изобретении композиции, фармацевтические композиции и вакцины, содержащие аутоантигены, в частности, способны нарушать толерантность к аутоантигену в случае применения у хозяина. В данном контексте «нарушение толерантности к аутоантигену» следует относить к усилению иммунного ответа, который определен в данном описании, и предпочтительно к усилению B- или T-клеточного ответа, специфичного по отношению к аутоантигену в случае применения предлагаемых в изобретении композиций, фармацевтических композиций и вакцин, содержащих аутоантиген, у хозяина. Кроме того, белки, которые являются результатом комбинирования двух или нескольких собственных молекул или которые представляют собой часть собственной молекулы, и белки, которые имеют высокую гомологию по отношению к собственным молекулам, которые определены выше (>95%, предпочтительно> 97%, более предпочтительно >99%), также могут считаться собственными.
Лечение: В используемом в данном описании смысле термины «лечение», «лечить», «леченый» или «процесс лечения» относятся к профилактике и/или терапии. В случае использования по отношению, например, к инфекционному заболеванию термин относится к профилактическому лечению, которое повышает резистентность субъекта к инфекции патогеном или, другими словами, уменьшает вероятность того, что субъект станет инфицированным патогеном или у него будут проявляться симптомы болезни, которую можно отнести к инфекции, а также к лечению после того, как субъект стал инфицированным, чтобы бороться с инфекцией, например, уменьшать или исключать инфекцию или предотвращать ухудшение.
Вакцина: В используемом в данном описании смысле термин «вакцина» относится к препарату, который содержит композицию согласно настоящему изобретению и который имеет форму, которую можно вводить животному. Как правило, вакцина содержит обычный физиологический раствор или забуференную среду на основе раствора соли, в которой суспендируют или растворяют композицию согласно настоящему изобретению. В указанной форме композицию согласно настоящему изобретению можно обычным образом использовать для профилактики, ослабления или лечения состояния иным образом. При введении в организм хозяина вакцина способна вызывать иммунный ответ, включая, но не ограничивая указанным, продукцию антител, цитокинов и/или активацию цитотоксических T-клеток, антигенпрезентирующих клеток, хелперных T-клеток, дендритных клеток и/или других клеточных ответов.
Необязательно вакцина согласно настоящему изобретению дополнительно содержит адъювант, который может составлять либо незначительную либо большую часть относительно соединения согласно настоящему изобретению. Термин «адъювант» в используемом в данном описании смысле относится к неспецифическим стимуляторам иммунного ответа или веществам, которые обеспечивает возможность создания депо в организме хозяина, которые в комбинации с вакциной согласно настоящему изобретению обеспечивают еще более повышенный иммунный ответ. Можно использовать множество адъювантов. Примеры включают неполный адъювант Фрейнда, гидроксид алюминия и модифицированный мурамилдипептид. Термин «адъювант» в используемом в данном описании смысле также относится к обычно специфичным стимуляторам иммунного ответа, которые в комбинации с вакциной согласно настоящему изобретению обеспечивают еще более повышенный и обычно специфичный иммунный ответ. Примеры включают, но не ограничены указанным, GM-CSF, IL-2, IL-12, IFNα. Другие примеры известны специалистам в данной области.
Вирусоподобная частица: В используемом в данном описании смысле термин «вирусоподобная частица» относится к структуре, аналогичной вирусной частице, но которая, как показано, не является патогенной. Обычно вирусоподобная частица согласно изобретению не несет генетической информации, кодирующей белки вирусоподобной частицы. В общем, в вирусоподобных частицах отсутствует вирусный геном, и поэтому они являются неинфекционными. Также вирусоподобные частицы часто могут быть получены в больших количествах при гетерологичной экспрессии и могут быть легко очищены. Некоторые вирусоподобные частицы могут содержать нуклеиновую кислоту, отличную от ее генома. Как указано, вирусоподобная частица согласно изобретению является нерепликативной и неинфекционной, так как в ней отсутствует весь или часть вирусного генома, в частности репликативные и инфекционные компоненты вирусного генома. Вирусоподобная частица согласно изобретению может содержать нуклеиновую кислоту, отличную от ее генома. Типичным и предпочтительным вариантом вирусоподобной частицы согласно настоящему изобретению является вирусный капсид, такой как вирусный капсид соответствующего вируса, бактериофага или РНК-фага. Термины «вирусный капсид» или «капсид», которые используют в данном описании взаимозаменяемо, относятся к макромолекулярной сборке, состоящей из белковых вирусных субъединиц. Обычно и предпочтительно вирусные белковые субъединицы собираются в вирусный капсид и капсид соответственно, имеющий структуру с присущей ему повторяющейся организацией, при этом указанная структура обычно является сферической или трубчатой. Например, капсиды РНК-фагов или HBcAg имеют сферическую форму с икосаэдрической симметрией. Термин «капсидоподобная структура» в используемом в данном описании смысле относится к макромолекулярной сборке, состоящей из вирусных белковых субъединиц, с похожей на капсид морфологией в определенном выше смысле, но с отклонениями от типичной симметричной сборки, сохраняя при этом достаточную степень упорядоченности и повторяемости.
Вирусоподобная частица бактериофага: В используемом в данном описании смысле термин «вирусоподобная частица бактериофага» относится к вирусоподобной частице, аналогичной по структуре бактериофагу, но являющаяся нерепликативной и неинфекционной, и он не содержит по меньшей мере гена или генов, кодирующих аппарат репликации бактериофага, и обычно также не содержит гена или генов, кодирующих белок или белки, ответственные за прикрепление вируса или проникновение вируса в организм хозяина. Однако указанное определение также охватывает вирусоподобные частицы бактериофагов, в которых указанный выше ген или гены все еще присутствуют, но неактивны и поэтому также приводят к нерепликативным и неинфекционным вирусоподобным частицам бактериофага.
VLP белка оболочки РНК-фага: Структуру капсида, образованную в результате самосборки 180 субъединиц белка оболочки РНК-фага и необязательно содержащую РНК хозяина, называют «VLP белка оболочки РНК-фага». Конкретным примером является VLP белка оболочки Qβ. В данном конкретном случае VLP белка оболочки Qβ может образоваться при сборке либо исключительно из субъединиц CP Qβ (SEQ ID: No 10), образованных при экспрессии гена CP Qβ, содержащего, например, стоп-кодон TAA, препятствующий экспрессии более длинного белка A1 белок посредством супрессии, смотри Kozlovska, T. M., et al., Intervirology 39: 9-15 (1996)), либо может дополнительно содержать субъединицы белка A1 (SEQ ID: No 11) в сборке капсида. Процесс считывания имеет низкую эффективность и приводит только к очень небольшому количеству белка A1 в VLP. Осуществлено большое количество примеров с разными комбинациями упакованного ISS и связанного антигена. Не наблюдали различий в эффективности связывания и упаковки, когда использовали VLP белка оболочки Qβ, собранные исключительно из субъединиц CP Qβ, или VLP белка оболочки Qβ, дополнительно содержащие субъединицы белка A1 в капсидах. Кроме того, не наблюдали различий в иммунном ответе между указанными препаратами QβVLP. Поэтому для ясности термин «QβVLP» используют в описании примеров либо для VLP белка оболочки Qβ, собранных исключительно из субъединиц CP Qβ, либо для VLP белка оболочки Qβ, дополнительно содержащих субъединицы белка A1 в капсидах.
Термин «вирусная частица» в используемом в данном описании смысле относится к морфологической форме вируса. В некоторых типах вирусов она содержит геном, окруженный белковым капсидом; другие имеют дополнительные структуры (например, оболочки, хвосты и т.д.).
Не имеющие оболочки вирусные частицы состоят из белкового капсида, который окружает вирусный геном. Вирусы, имеющие оболочки, также имеют структуру капсида, окружающую генетический материал вируса, но, кроме этого, имеют липидную бислойную оболочку, которая окружает капсид. В предпочтительном варианте изобретения VLP не содержат липопротеидной оболочки или оболочки, содержащей липопротеиды. В следующем предпочтительном варианте VLP вообще не содержат оболочки.
Формы единственного числа: Когда в данном описании используют термин «один» или форму единственного числа, он означают «по меньшей мере один» или «один или несколько», если не оговорено особо.
Как будет понятно специалистам в данной области, некоторые варианты изобретения заключаются в применении технологии рекомбинантных нуклеиновых кислот, таких как клонирование, полимеразная цепная реакция, очистка ДНК и РНК, экспрессия рекомбинантных белков в прокариотических и эукариотических клетках и т.д. Такие методики хорошо известны специалистам в данной области, и их можно легко найти в опубликованных руководствах по лабораторным способам (например, Sambrook, J. et al., eds., Molecular Cloning, a Laboratory Manual, 2nd. edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. (1989); Ausubel, F. et al., eds., Current Protocols in Molecular Biology, John H. Wiley and Sons, Inc. (1997)). Основные лабораторные методики работы с линиями культур клеток и тканей (Celis, J., ed., Cell Biology, Academic Press, 2nd edition, (1998)) и методики, основанные на антителах (Harlow, E. and Lane, D., «Antibodies: A Laboratory Manual», Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1988); Deutscher, M.P., «Guide to Protein Purification», Meth. Enzymol. 128, Academic Press San Diego (1990); Scopes, R. K., «Protein Purification Principles and Practice», 3rd ed., Springer-Verlag, New York (1994)), также в достаточной мере описаны в литературе, все публикации включены в данное описание в виде ссылки.
2. Композиции и способы усиления иммунного ответа
Заявленное изобретение относится композициям и способам усиления иммунного ответа против одного или нескольких антигенов у животного. Композиции согласно изобретению содержат, или альтернативно в основном состоят, или альтернативно состоят из вирусоподобной частицы, по меньшей мере одного иммуностимулирующего вещества, предпочтительно иммуностимулирующей нуклеиновой кислоты и еще более предпочтительно неметилированного CpG-содержащего олигонуклеотида, и по меньшей мере одного антигена или антигенной детерминанты, где иммуностимулирующее вещество, иммуностимулирующая нуклеиновая кислота или олигонуклеотид связаны с вирусоподобной частицей, и где указанный антиген или антигенная детерминанта связана с указанной вирусоподобной частицей, и где указанный антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека. Кроме того, изобретение позволяет специалисту легко конструировать такую композицию для различных терапевтических и/или профилактических целей, которые включают предотвращение и/или лечение, например, злокачественных опухолей.
Вирусоподобные частицы в контексте настоящей заявки относятся к структурам, похожим на вирусную частицу, но которые являются непатогенными. В общем в вирусоподобных частицах отсутствует вирусный геном, и поэтому они являются неинфекционными. Также вирусоподобные частицы могут быть получены в больших количествах при гетерологичной экспрессии и могут быть легко очищены.
В предпочтительном варианте вирусоподобная частица является рекомбинантной вирусоподобной частицей. Специалист в данной области может получить VLP, используя технологию рекомбинантной ДНК и кодирующие вирус последовательности, которые легко доступны для общественности. Например, кодирующая последовательность вирусного белка оболочки или кора может быть сконструирована для экспрессии в бакуловирусном экспрессирующем векторе с использованием коммерчески доступного бакуловирусного вектора под регуляторным контролем вирусного промотора с соответствующими модификациями последовательности, чтобы обеспечить функциональное связывание кодирующей последовательности с регуляторной последовательностью. Кодирующая последовательность вирусного белка оболочки или кора также может быть сконструирована для экспрессии, например, в бактериальном экспрессирующем векторе.
Примеры VLP включают, но не ограничены указанным, белки капсида вируса гепатита B, вируса кори, вируса Синдбис, ротавируса, вируса ящура, вируса Норуолк, ретровирусный белок GAG, белок p1 ретротранспозона Ty, поверхностный белок вируса гепатита B, вируса папилломы человека, вируса полиомы человека, вируса BK (BKV), РНК-фагов, Ty, fr-фага, GA-фага, фага AP 205 и в частности фага Qβ.
Как будет легко понятно специалистам в данной области, VLP согласно изобретению не ограничена какой-либо конкретной формой. Частица может быть синтезирована химически или посредством биологического процесса, который может быть природным или неприродным. В качестве примера данный тип варианта включает вирусоподобную частицу или ее рекомбинантную форму.
В более конкретном варианте VLP может содержать или альтернативно состоять из рекомбинантных полипептидов ротавируса; рекомбинантных полипептидов вируса Норуолк; рекомбинантных полипептидов альфавируса; рекомбинантных белков, которые образуют бактериальные фимбрии или подобные фимбриям структуры; рекомбинантных полипептидов вируса ящура; рекомбинантных полипептидов вируса кори, рекомбинантных полипептидов вируса Синдбис, рекомбинантных полипептидов ретровируса; рекомбинантных полипептидов вируса гепатита B (например, HBcAg); рекомбинантных полипептидов вируса табачной мозаики; рекомбинантных полипептидов вируса овечьих кошар; рекомбинантных полипептидов вируса папилломы человека; рекомбинантных полипептидов вируса полиомы и, в частности, рекомбинантных полипептидов вируса полиомы человека, и в частности, рекомбинантных полипептидов вируса BK; рекомбинантных полипептидов бактериофагов, рекомбинантных полипептидов РНК-фагов; рекомбинантных полипептидов Ty; рекомбинантных полипептидов fr-фага, рекомбинантных полипептидов GA-фага, рекомбинантных полипептидов фага AP 205, и в частности, рекомбинантных полипептидов фага Qβ. Вирусоподобная частица, кроме того, может содержать или альтернативно состоять из одного или нескольких фрагментов таких полипептидов, а также вариантов таких полипептидов. Варианты полипептидов могут иметь, например, по меньшей мере 80%, 85%, 90%, 95%, 97%, или 99% идентичность на аминокислотном уровне с их аналогами дикого типа.
В предпочтительном варианте вирусоподобная частица содержит, в основном состоит, или альтернативно состоит из рекомбинантных белков или их фрагментов РНК-фага. Предпочтительно РНК-фаг выбран из группы, состоящей из a) бактериофага Qβ; b) бактериофага R17; c) бактериофага fr; d) бактериофага GA; e) бактериофага SP; f) бактериофага MS2; g) бактериофага M11; h) бактериофага MX1; i) бактериофага NL95; k) бактериофага f2;1) бактериофага PP7; и m) бактериофага AP205.
В другом предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из рекомбинантных белков или их фрагментов РНК-бактериофага Qβ или РНК-бактериофага fr или РНК-бактериофага AP205.
В следующем предпочтительном варианте настоящего изобретения рекомбинантные белки содержат, или альтернативно в основном состоят, или альтернативно состоят из белков оболочки РНК-фагов.
Поэтому белки оболочки РНК-фагов, образующие капсиды или VLP, или фрагменты белков оболочки бактериофагов, совместимые с самосборкой в капсид или в VLP, являются следующими предпочтительными вариантами настоящего изобретения. Белки оболочки бактериофага Qβ, например, могут быть экспрессированы рекомбинантно в E. coli. Кроме того, при такой экспрессии указанные белки спонтанно образуют капсиды. Кроме того, указанные капсиды образуют структуру с присущей повторяющейся организацией.
Конкретные предпочтительные примеры белков оболочки бактериофагов, которые могут быть использованы для получения композиций согласно изобретению, включают белки оболочки РНК-бактериофагов, таких как бактериофаг Qβ (SEQ ID NO: 10; база данных PIR, No. доступа VCBPQP, относящаяся к CP Qβ, и SEQ ID NO: 11; No. доступа AAA16663, относящаяся к белку A1 Qβ), бактериофаг R17 (PIR No. доступа VCBPR7), бактериофаг fr (SEQ ID NO: 13; PIR No. доступа VCBPFR), бактериофаг GA (SEQ ID NO: 14; GenBank, No. доступа NP-040754), бактериофаг SP (GenBank, No. доступа CAA30374, относящаяся к CP SP, и No. доступа NP_695026, относящаяся к белку A1 SP), бактериофаг MS2 (PIR No. доступа VCBPM2), бактериофаг M11 (GenBank, No. доступа AAC06250), бактериофаг MX1 (GenBank, No. доступа AAC14699), бактериофаг NL95 (GenBank, No. доступа AAC14704), бактериофаг f2 (GenBank, No. доступа P03611), бактериофаг PP7 (SEQ ID NO: 19), и бактериофаг AP205 (SEQ ID NO: 31). Кроме того, в сборку капсида из белков оболочки Qβ может быть включен белок A1 бактериофага Qβ или укороченные на C-конце формы, в которых отсутствуют до 100, 150 или 180 аминокислот из C-конца. Как правило, процентное содержание белка QβA1 относительно Qβ CP при сборке капсида будет ограничено, чтобы обеспечить образование капсида. Дополнительные конкретные примеры белков оболочки бактериофагов описаны в WO02/056905 на странице 45 и 46, включенной в данное описание посредством ссылки. Дополнительные предпочтительные вирусоподобные частицы РНК-фагов, в частности Qβ согласно данному изобретению, раскрыты в WO 02/056905, описание которой включено в виде ссылки в полном объеме.
В следующем предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из рекомбинантных белков или их фрагментов РНК-фага, при этом рекомбинантные белки содержат, в основном состоят, или альтернативно состоят из мутантных белков оболочки РНК-фага, предпочтительно мутантных белков оболочки РНК-фагов, указанных выше. В другом предпочтительном варианте мутантные белки оболочки РНК-фага модифицированы удалением по меньшей мере одного остатка лизина посредством замены или добавлением по меньшей мере одного остатка лизина посредством замены; альтернативно мутантные белки оболочки РНК-фага модифицированы делецией по меньшей мере одного остатка лизина или добавлением по меньшей мере одного остатка лизина посредством инсерции. Делеция, замена или добавление по меньшей мере одного остатка лизина позволяют варьировать степень связывания, т.е. количество аналогов пептида Melan-A меланомы человека на субъединицу VLP РНК-фагов, в частности для соответствия и приспособления к требованиям для вакцины. В предпочтительном варианте настоящего изобретения в среднем по меньшей мере 1,0 аналог пептида Melan-A меланомы человека на субъединицу связан с VLP РНК-фага. Указанное значение рассчитывают в виде среднего для субъединиц или мономеров VLP РНК-фага. В следующем предпочтительном варианте настоящего изобретения по меньшей мере 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9 или по меньшей мере 2,0 аналога пептида Melan-A меланомы человека связаны с VLP РНК-фагов, которые рассчитывают в виде среднего значения связывания для всех субъединиц или мономеров VLP РНК-фага.
В другом предпочтительном варианте вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из рекомбинантных белков или их фрагментов РНК-бактериофага Qβ, при этом рекомбинантные белки содержат, или альтернативно в основном состоят, или альтернативно состоят из белков оболочки, имеющих аминокислотную последовательность SEQ ID NO: 10, или смеси белков оболочки, имеющих аминокислотные последовательности SEQ ID NO: 10 и SEQ ID NO: 11 или мутантов SEQ ID NO: 11, и при этом N-концевой метионин предпочтительно отщеплен.
В следующем предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, в основном состоит, или альтернативно состоит из рекомбинантных белков Qβ или их фрагментов, при этом рекомбинантные белки содержат, или альтернативно в основном состоят, или альтернативно состоят из мутантных белков оболочки Qβ. В другом предпочтительном варианте указанные мутантные белки оболочки модифицированы удалением по меньшей мере одного остатка лизина посредством замены или добавлением по меньшей мере одного остатка лизина посредством замены. Альтернативно указанные мутантные белки оболочки модифицированы делецией по меньшей мере одного остатка лизина или добавлением по меньшей мере одного остатка лизина посредством инсерции.
Четыре остатка лизина экспонированы на поверхности капсида из белка оболочки Qβ. Мутантны Qβ, в случае которых экспонированные остатки лизина заменяют аргининами, также могут быть использованы для настоящего изобретения. Таким образом, следующие мутантные белки оболочки Qβ и мутантные VLP Qβ могут быть использованы в практике изобретения: «Qβ-240» (Lys 13-Arg; SEQ ID NO: 20), «Qβ-243» (Asn 10-Lys; SEQ ID NO: 21), «Qβ-250» (Lys 2-Arg, Lys 13-Arg; SEQ ID NO: 22), «Qβ-251» (SEQ ID NO: 23) и «Qβ-259» (Lys 2-Arg, Lys 16-Arg; SEQ ID NO: 24). Таким образом, в следующем предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, в основном состоит, или альтернативно состоит из рекомбинантных белков мутантных белков оболочки Qβ, которые включают белки, имеющие аминокислотную последовательность, выбранную из следующей группы: a) аминокислотная последовательность SEQ ID NO: 20; b) аминокислотная последовательность SEQ ID NO: 21; c) аминокислотная последовательность SEQ ID NO: 22; d) аминокислотная последовательность SEQ ID NO: 23; и e) аминокислотная последовательность SEQ ID NO: 24. Конструирование, экспрессия и очистка указанных выше VLP и капсидов белков оболочки Qβ, мутантных белков оболочки Qβ соответственно описаны в WO 02/056905. В частности это касается примера 18 упомянутой выше заявки.
В следующем предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из рекомбинантных белков Qβ или их фрагментов, при этом рекомбинантные белки содержат, в основном состоят, или альтернативно состоят из смеси любого одного из указанных выше мутантов Qβ и соответствующего белка A1.
В следующем предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из рекомбинантных белков или их фрагментов РНК-фага AP205.
Геном AP205 состоит из генов белка созревания, белка оболочки, репликазы и двух открытых рамок считывания, не присутствующих в родственных фагах; гена лизиса и открытой рамки считывания, играющей роль в трансляции гена созревания (Klovins, J. , et al., J. Gen. Virol. 83: 1523-33 (2002)). Белок оболочки AP205 может быть экспрессирован с плазмиды pAP283-58 (SEQ ID NO: 30), которая является производной pQb10 (Kozlovska, T. M. et al., Gene 137: 133-37 (1993)) и которая содержит сайт связывания рибосомы AP205. Альтернативно белок оболочки AP205 может быть клонирован в pQb185 ниже сайта связывания рибосомы, присутствующего в векторе. Оба способа приводят к экспрессии белка и образованию капсидов, как описано в WO 04/007538, которая включена в виде ссылки в полном объеме. Векторы pQb10 и pQb185 получают из вектора pGEM, и экспрессия клонированных генов с указанных векторов регулируется промотором trp (Kozlovska, T. M. et al., Gene 137: 133-37 (1993)). Плазмида pAP283-58 (SEQ ID NO: 30) содержит предполагаемый сайт связывания рибосомы AP205 в следующей последовательности, которая расположена ниже сайта XbaI и непосредственно перед стартовым кодоном ATG белка оболочки AP205: tctagaATTTTCTGCGCACCCATCCCGGGTGGCGCCCAAAGTGAGGAAAATCACatg (основания 77-133 SEQ ID NO: 30). Вектор pQb185 содержит последовательность Шайна-Далгарно ниже сайта XbaI и выше стартового кодона (tctagaTTAACCCAACGCGTAGGAGTCAGGCCatg (SEQ ID NO: 61), последовательность Шайна-Далгарно подчеркнута).
В следующем предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из рекомбинантных белков оболочки или их фрагментов РНК-фага AP205.
Таким образом, указанный предпочтительный вариант настоящего изобретения включает в себя белки оболочки AP205, которые образуют капсиды. Такие белки экспрессируют рекомбинантно или получают из природных источников. Белки оболочки AP205, продуцированные в бактериях, спонтанно образуют капсиды, что подтверждается электронной микроскопией (ЭМ) и иммунодиффузией. Структурные особенности капсида, образованного белком оболочки AP205 (SEQ ID NO: 31), и особенности капсида, образованного белком оболочки РНК-фага AP205, почти не отличаются при наблюдении в ЭМ. VLP AP205 являются высокоиммуногенными и могут быть связаны с антигенами и/или антигенными детерминантами с образованием конструкций вакцин, экспонирующих антигены и/или антигенные детерминанты, ориентированные повторяющимся образом. Против экспонированных таким образом антигенов вырабатываются высокие титры антител, что свидетельствует о том, что связанные антигены и/или антигенные детерминанты доступны для взаимодействия с молекулами антител и являются иммуногенными.
В следующем предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из рекомбинантных мутантных белков оболочки или их фрагментов РНК-фага AP205.
Компетентные в отношении сборки мутантные формы VLP AP205, включающие белок оболочки AP205 с заменой пролина в положении аминокислоты 5 на треонин (SEQ ID NO: 32), также можно использовать в практике изобретения, и они составляют следующий предпочтительный вариант изобретения. Указанные VLP, VLP AP205, полученные из природных источников, или вирусные частицы AP205 могут быть связаны с антигенами с образованием упорядоченных повторяющихся матриц антигенов согласно данному изобретению.
Мутантный белок оболочки P5-T AP205 может быть экспрессирован с плазмиды pAP281-32 (SEQ ID NO. 33), которая получена непосредственно из pQb185 и которая содержит ген мутантного белка оболочки AP205 вместо гена белка оболочки Qβ. Векторы для экспрессии белка оболочки AP205 трансфицировали в E. coli для экспрессии белка оболочки AP205.
Способы экспрессии белка оболочки и соответственно мутантного белка оболочки, приводящей к самосборке в VLP, описаны в заявке на выдачу патента WO 04/007538, которая включена в виде ссылки в полном объеме. Подходящие штаммы E. coli включают, но не ограничены указанным, E. coli K802, JM 109, RR1. Подходящие векторы и штаммы и их комбинации можно идентифицировать в результате тестирования экспрессии белка оболочки и соответственно мутантного белка оболочки с помощью SDS-ПААГ и образования и сборки капсида сначала с помощью необязательной очистки капсидов гель-фильтрацией и затем их тестированием в анализе иммунодиффузии (тест Оухтерлони) или с помощью электронной микроскопии (Kozlovska, T. M., et al., Gene 137: 133-37 (1993)).
Белки оболочки AP205, экспрессированные с векторов pAP283-58 и pAP281-32, могут быть лишены начальной аминокислоты метионина вследствие процессинга в цитоплазме E. coli. Расщепленные, нерасщепленные формы VLP AP205 или их смеси являются следующими предпочтительными вариантами изобретения.
В следующем предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из смеси рекомбинантных белков оболочки или их фрагментов РНК-фага AP205 и рекомбинантных мутантных белков оболочки или их фрагментов РНК-фага AP205.
В следующем предпочтительном варианте настоящего изобретения вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из фрагментов рекомбинантных белков оболочки или рекомбинантных мутантных белков оболочки РНК-фага AP205.
Фрагменты рекомбинантного белка оболочки AP205, способные собираться в VLP и соответственно капсид, также применимы в практике изобретения. Указанные фрагменты могут быть образованы путем делеции либо внутри, либо на концах белка оболочки и мутантного белка оболочки соответственно. Инсерции в последовательность белка оболочки и мутантного белка оболочки или слияния антигенных последовательностей с последовательностью белка оболочки и мутантного белка оболочки, которые совместимы со сборкой в VLP, являются следующими вариантами изобретения, которые приводят к химерным белкам оболочки AP205 и частицам соответственно. Последствия инсерций, делеций и слияний с последовательностью белка оболочки и совместимо ли это со сборкой в VLP, можно определить с помощью электронной микроскопии.
Частицы, образованные белком оболочки, фрагментами белка оболочки или химерными белками оболочки AP205, описанными выше, можно выделить в чистой форме в результате комбинирования стадий фракционирования путем осаждения и стадий очистки гель-фильтрацией с использованием колонок, например, с сефарозой CL-4B, сефарозой CL-2B, сефарозой CL-6B и их комбинаций, как описано в заявке на выдачу патента WO 04/007538, которая включена в виде ссылки в полном объеме. Другие способы выделения вирусоподобных частиц известны в данной области и могут быть использованы для выделения вирусоподобных частиц (VLP) бактериофага AP205. Например, применение ультрацентрифугирования для выделения VLP дрожжевого ретротранспозона Ty описано в патенте США No. 4918166, который включен в данное описание в виде ссылки в полном объеме.
Определена кристаллическая структура нескольких РНК-бактериофагов (Golmohammadi, R. et al., Structure 4: 543-554 (1996)). Используя указанную информацию, специалист в данной области легко может идентифицировать экспонированные на поверхности остатки и модифицировать белки оболочки бактериофага так, чтобы можно было встроить один или несколько реакционноспособных аминокислотных остатков. Таким образом, специалист в данной области может легко создать и идентифицировать модифицированные формы белков оболочки бактериофага, которые можно использовать в практике изобретения. Таким образом, варианты белков, которые образуют капсиды или капсидоподобные структуры (например, белки оболочки бактериофага Qβ, бактериофага R17, бактериофага fr, бактериофага GA, бактериофага SP и бактериофага MS2), также можно использовать для получения композиций согласно данному изобретению и композиций вакцин. Следующие возможные примеры модифицированных РНК-бактериофагов, а также вариантов белков и укороченных на N- и C-концах мутантов, которые образуют капсиды или капсидоподобные структуры, а также способы получения таких композиций и композиций вакцин соответственно, которые подходят для применения в настоящем изобретении, описаны в WO 02/056905 со страницы 50, строка 33, до страницы 52, строка 29.
Таким образом, изобретение, кроме того, включает в себя композиции и композиции вакцин, полученные из белков, которые образуют капсиды или VLP, способы получения таких композиций из отдельных субъединиц белка и VLP или капсидов, способы получения указанных отдельных субъединиц белка, молекул нуклеиновых кислот, которые кодируют указанные субъединицы, и способы вакцинации и/или способы, вызывающие иммунологические ответы у индивидуумов, с использованием композиций согласно настоящему изобретению.
В другом предпочтительном варианте изобретения VLP не содержат липопротеидной оболочки или оболочки, содержащей липопротеиды. В следующем предпочтительном варианте VLP вообще не содержат оболочки.
Отсутствие липопротеидной оболочки или оболочки, содержащей липопротеиды, и, в частности, полное отсутствие оболочки приводит к более определенной вирусоподобной частице по своей структуре и композиции. Поэтому такие более определенные вирусоподобные частицы могут минимизировать побочные эффекты. Кроме того, отсутствие оболочки содержащей липопротеиды или, в частности, полное отсутствие оболочки позволяет избегать или минимизировать включения потенциально токсичных молекул и пирогенов в вирусоподобную частицу.
В одном варианте изобретение относится к композиции вакцины согласно изобретению, содержащей вирусоподобную частицу, при этом предпочтительно указанной вирусоподобной частицей является рекомбинантная вирусоподобная частица. Предпочтительно вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из рекомбинантных белков или их фрагментов РНК-фага, предпочтительно белков оболочки РНК-фагов. Альтернативно рекомбинантные белки вирусоподобной частицы композиции вакцины согласно изобретению содержат, или альтернативно в основном состоят, или альтернативно состоят из мутантных белков оболочки РНК-фагов, где РНК-фаг выбран из группы, состоящей из: (a) бактериофага Qβ; (b) бактериофага R17; (c) бактериофага fr; (d) бактериофага GA; (e) бактериофага SP; (f) бактериофага MS2; (g) бактериофага M11; (h) бактериофага MX1; (i) бактериофага NL95; (k) бактериофага f2; (l) бактериофага PP7 и (m) бактериофага AP205.
В предпочтительном варианте мутантные белки оболочки указанного РНК-фага модифицированы удалением или добавлением по меньшей мере одного остатка лизина путем замены. В другом предпочтительном варианте мутантные белки оболочки указанного РНК-фага модифицированы делецией по меньшей мере одного остатка лизина или добавлением по меньшей мере одного остатка лизина посредством инсерции. В предпочтительном варианте вирусоподобная частица содержит рекомбинантные белки или их фрагменты РНК-фага Qβ, РНК-фага fr или РНК-фага AP205.
Как указано ранее, изобретение включает в себя вирусоподобные частицы или их рекомбинантные формы. Специалистам в данной области известно, как получить такие частицы и связать с ними антигены. Следующие предпочтительные варианты настоящего изобретения описаны в разделе «Примеры».
В одном варианте вирусоподобная частица содержит, или альтернативно в основном состоит, или альтернативно состоит из рекомбинантных белков или их фрагментов вируса BK (BKV), при этом рекомбинантные белки содержат, или альтернативно в основном состоят, или альтернативно состоят из белков, имеющих аминокислотную последовательность SEQ ID NO: 12. Вирус BK (BKV) является непокрытым оболочкой двунитевым ДНК-вирусом, относящимся к подсемейству вирусов полиомы Papovaviridae. VP1 является основным белком капсида BKV. VP1 имеет 362 аминокислоты (SEQ ID NO: 12, Gene Bank доступ: AAA46882) и имеет размер 42 кД. При продуцировании в E. coli, клетках насекомых или дрожжах VP1 спонтанно образует капсидные структуры (Salunke D.M., et al, Cell 46(6):895-904 (1986); Sasnauskas, K., et al., Biol. Chem. 380(3):381-6 (1999); Sasnauskas, K., et al., 3rd International Workshop "Virus-like particles as vaccines" Berlin, September 26-29 (2001); Touze, A., et al., J Gen Virol. 82(Pt 12): 3005-9 (2001).
Капсид организован в виде 72 пентамеров VP1, образующих икосаэдрическую структуру. Капсиды имеют диаметр примерно 45 нм.
В одном варианте частицы, используемые в композициях согласно изобретению, состоят из белка капсида (кора) гепатита B (HBcAg) или фрагмента HBcAg, который был модифицирован для того, чтобы либо удалить, либо уменьшить количество свободных остатков цистеина. Zhou et al. (J. Virol. 66: 5393-5398 (1992)) показали, что HBcAg, которые были модифицированы для того, чтобы удалить имеющиеся в природных условиях остатки цистеина, сохраняют способность вступать в ассоциацию и образовывать многомерные структуры. Таким образом, коровые частицы, подходящие для применения в композициях согласно изобретению, включают коровые частицы, содержащие модифицированные HBcAg или их фрагменты, в которых один или несколько имеющихся в природных условиях остатков цистеина были либо делетированы, либо заменены другим аминокислотным остатком (например, остатком серина).
HBcAg является белком, образуемым в результате процессинга белка-предшественника корового антигена гепатита B. Идентифицирован ряд изотипов HBcAg, и их аминокислотные последовательности легко доступны специалистам в данной области. Например, белок HBcAg, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 16, имеет длину в 185 аминокислот и создается в результате процессинга белка-предшественника корового антигена гепатита B длиной 212 аминокислот. Указанный процессинг приводит к удалению 29 аминокислот из N-конца белка-предшественника корового антигена гепатита B. Подобным образом белок HBcAg, который имеет длину 185 аминокислот, образуется в результате процессинга белка-предшественника корового антигена гепатита B длиной 214 аминокислот.
В предпочтительных вариантах композиции вакцин согласно изобретению могут быть получены с использованием процессированной формы HBcAg (т.е. HBcAg, из которого была удалена N-концевая лидерная последовательность белка-предшественника корового антигена гепатита B).
Кроме того, когда HBcAg получают в условиях, при которых процессинг не будет происходить, HBcAg, как правило, будет экспрессироваться в «процессированной» форме. Например, бактериальные системы, такие как E. coli, как правило, не удаляют лидерные последовательности, также называемые «сигнальными пептидами», белков, которые обычно экспрессируются в эукариотических клетках. Следовательно, когда для получения HBcAg согласно изобретению используют систему экспрессии E. coli, направляющую экспрессию белка в цитоплазме, указанные белки, как правило, будут экспрессироваться так, чтобы в них отсутствовала N-концевая лидерная последовательность белка-предшественника корового антигена гепатита B.
Получение вирусоподобных частиц гепатита B, которые можно использовать для настоящего изобретения, описано, например, в WO 00/32227 и в указанной заявке, в частности в примерах с 17 по 19 и с 21 по 24, а также в WO 01/85208 и в ней, в частности, в примерах с 17 по 19, с 21 по 24, с 31 по 41, и в WO 02/056905. В случае последней заявки это, в частности, относится к примеру 23, 24, 31 и 51. Все три документа включены в данное описание в виде ссылки в полном объеме.
Настоящее изобретение также относится к вариантам HBcAg, которые были модифицированы для того, чтобы делетировать или заменить один или несколько дополнительных остатков цистеина. Таким образом, композиции вакцин согласно изобретению включают в себя композиции, содержащие HBcAg, в которых остатки цистеина, не присутствующие в аминокислотной последовательности, показанной в SEQ ID NO: 16, были делетированы.
В данной области хорошо известно, что свободные остатки цистеина могут быть вовлечены в ряд побочных химических реакций. Указанные побочные реакции включают дисульфидные обмены, реакцию с химическими веществами или метаболитами, которые, например, инъецированы или образованы при комбинированной терапии с другими веществами, или прямое окисление и реакцию с нуклеотидами при воздействии УФ-света. Таким образом, могут быть образованы токсичные аддукты, особенно учитывая тот факт, что HBcAg имеют устойчивую тенденцию к связыванию нуклеиновых кислот. Таким образом, токсичные аддукты могут быть представлены множеством видов, каждый из которых отдельно может присутствовать в низкой концентрации, но вместе они достигают токсичных уровней.
С точки зрения указанного выше одним их преимуществ применения в композициях вакцин HBcAg, которые были модифицированы, чтобы удалить имеющиеся в природных условиях остатки цистеина, является то, что количество сайтов, с которыми могут связываться токсичные элементы в том случае, когда связаны антигены или антигенные детерминанты, может быть уменьшено или они могут быть вообще удалены.
Идентифицирован ряд встречающихся в природе вариантов HBcAg, подходящих для применения в практике данного изобретения. Yuan et al., (J. Virol. 73: 10122-10128 (1999)), например, описывают варианты, в которых остаток изолейцина в положении, соответствующем положению 97 в SEQ ID NO: 25, заменен либо остатком лейцина, либо остатком фенилаланина. Аминокислотные последовательности ряда вариантов HBcAg, а также нескольких вариантов предшественника корового антигена гепатита B описаны в GenBank, запись AAF121240, AF121239, Х85297, Х02496, Х85305, Х85303, AF151735, Х85259, Х85286, Х85260, Х85317, Х85298, AF043593, М20706, Х85295, Х80925, Х85284, Х85275, Х72702, Х85291, Х65258, Х85302, М32138, Х85293, Х85315, U95551, Х85256, Х85316, Х85296, АВ033559, Х59795, Х85299, Х85307, Х65257, Х85311, Х85301 (SEQ ID NO:26), X85314, X85287, X85272, X85319, AB010289, X85285, AB010289, AF121242, M90520 (SEQ ID NO:27), P03153, AF110999, и M95589, описания каждой из которых включены в данную заявку в виде ссылки. Последовательности указанных выше вариантов предшественника корового антигена гепатита B дополнительно описаны в WO 01/85208 в SEQ ID NO: 89-138 в заявке WO 01/85208. Указанные варианты HBcAg отличаются по аминокислотной последовательности в ряде положений, включая аминокислотные остатки, которые соответствуют аминокислотным остаткам, распложенным в положениях 12, 13, 21, 22, 24, 29, 32, 33, 35, 38, 40, 42, 44, 45, 49, 51, 57, 58, 59, 64, 66, 67, 69, 74, 77, 80, 81, 87, 92, 93, 97, 98, 100, 103, 105, 106, 109, 113, 116, 121, 126, 130, 133, 135, 141, 147, 149, 157, 176, 178, 182 и 183 в SEQ ID NO: 28. Следующие варианты HBcAg, подходящие для применения в композициях согласно изобретению, которые могут быть дополнительно модифицированы согласно изложенному в данном описании, описаны в WO 00/198333, WO 00/177158 и WO 00/214478.
HBcAg, подходящие для применения в настоящем изобретении, могут быть получены из любого организма, при условии что они способны заключать внутри себя, быть соединенными или иным образом связанными, в частности при условии, что они способны упаковывать неметилованный CpG-содержащий олигонуклеотид и индуцировать иммунный ответ.
Как указано выше, как правило, в композициях вакцин согласно изобретению будут использованы процессированные HBcAg (т.е. те, в которых отсутствуют лидерные последовательности). Настоящее изобретение включает в себя композиции вакцин, а также способы применения таких композиций, в которых используются описанные выше варианты HBcAg.
Кроме того, в объем изобретения включены дополнительные варианты HBcAg, которые способны вступать в ассоциацию, образуя димерные или многомерные структуры. Таким образом, изобретение, кроме того, включает в себя композиции вакцин, содержащие полипептиды HBcAg, содержащие или альтернативно состоящие из аминокислотной последовательности, которая по меньшей мере на 80%, 85%, 90%, 95%, 97% или 99% идентична любой из аминокислотных последовательностей дикого типа, и формы указанных белков, которые были процессированы в соответствующих случаях, чтобы удалить N-концевую лидерную последовательность.
Имеет ли аминокислотная последовательность полипептида аминокислотную последовательность, которая, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентична одной из аминокислотных последовательностей дикого типа или ее части, можно определить стандартным способом с использованием известных компьютерных программ, таких как программа Bestfit. При использовании Bestfit или любой другой программы выравнивания последовательностей для того, чтобы определить, идентична ли конкретная последовательность, например, на 95% эталонной аминокислотной последовательности, параметры устанавливают так, чтобы процент идентичности рассчитывался по всей полной длине эталонной аминокислотной последовательности и чтобы были разрешены пробелы в гомологии до 5% от общего количества аминокислотных остатков в эталонной последовательности.
Аминокислотные последовательности указанных выше вариантов и предшественников HBcAg относительно сходны друг с другом. Таким образом, ссылка на аминокислотный остаток варианта HBcAg, расположенный в положении, которое соответствует конкретному положению в SEQ ID NO: 28, относится к аминокислотному остатку, который присутствует в данном положении в аминокислотной последовательности, показанной в SEQ ID NO: 28. Гомология между указанными вариантами HBcAg среди вирусов гепатита B, которые инфицируют млекопитающих, в большинстве случаев достаточно высока, так что специалист в данной области не будет иметь больших трудностей при просмотре обеих последовательностей, аминокислотной последовательности, показанной в SEQ ID NO: 28, и аминокислотной последовательности конкретного варианта HBcAg, и при идентификации «соответствующих» аминокислотных остатков. Кроме того, аминокислотная последовательность HBcAg, показанная в SEQ ID NO: 27, в которой представлена аминокислотная последовательность HBcAg, полученного из вируса, который инфицирует лесных сурков, имеет достаточно высокую гомологию с HBcAg, имеющим аминокислотную последовательность, показанную в SEQ ID NO: 28, так что легко увидеть, что инсерция из трех аминокислотных остатков присутствует в SEQ ID NO: 27 между остатками аминокислот 155 и 156 SEQ ID NO: 28.
Изобретение также включает в себя композиции вакцин, которые содержат варианты HBcAg вирусов гепатита B, которые инфицируют птиц, а также композиции вакцин, которые содержат фрагменты указанных вариантов HBcAg. Специалисту в данной области будет понятно, что один, два, три или более остатков цистеина, присутствующих в указанных полипептидах в природных условиях, могут быть либо заменены другим аминокислотным остатком, либо делетированы перед их включением в композиции вакцин согласно изобретению.
Как обсуждается выше, удаление свободных остатков цистеина уменьшает количество сайтов, в которых токсичные компоненты могут связываться с HBcAg, а также удаляет сайты, в которых может происходить перекрестное связывание остатков лизина и цистеина одной и той же или соседних молекул HBcAg. Таким образом, в другом варианте настоящего изобретения один или несколько остатков цистеина белка капсида вируса гепатита B либо делетированы, либо заменены другим аминокислотным остатком. Экспрессия и очистка варианта HBcAg-Lys описана в примере 24 WO 02/056905, и конструирование HBcAg, не имеющего свободных остатков цистеина и содержащих встроенный остаток лизина, описано в примере 31 WO 02/056905.
В других вариантах композиции и соответственно композиции вакцин согласно изобретению будут содержать HBcAg, из которых была удалена C-концевая область (например, аминокислотные остатки 145-185 или 150-185 SEQ ID NO: 28). Таким образом, дополнительные модифицированные HBcAg, подходящие для применения в практике настоящего изобретения, включают укороченные на C-конце мутанты. Подходящие укороченные мутанты включают HBcAg, в которых 1, 5, 10, 15, 20, 25, 30, 34, 35 аминокислот были удалены из C-конца.
HBcAg, подходящие для применения в практике настоящего изобретения, также включают укороченные на N-конце мутанты. Подходящие укороченные мутанты включают модифицированные HBcAg, в которых 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот были удалены из N-конца.
Следующие HBcAg, подходящие для применения в практике настоящего изобретения, включают укороченные на N- и C-концах мутанты. Подходящие укороченные мутанты включают HBcAg, в которых 1, 2, 5, 7, 9, 10, 12, 14, 15 и 17 аминокислот были удалены из N-конца и 1, 5, 10, 15, 20, 25, 30, 34, 35 аминокислот были удалены из C-конца.
Изобретение, кроме того, включает в себя композиции и соответственно композиции вакцин, содержащие полипептиды HBcAg, содержащие, или альтернативно в основном состоящие, или альтернативно состоящие из аминокислотных последовательностей, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны описанным выше укороченным мутантам.
В некоторых вариантах согласно изобретению в полипептид HBcAg вводят остаток лизина, чтобы опосредовать связывание аналога белка Melan-A согласно изобретению с VLP HBcAg. В предпочтительных вариантах композиции согласно изобретению получают с использованием HBcAg, содержащего или альтернативно состоящего из аминокислот 1-144 или 1-149, 1-185 SEQ ID NO: 28, который модифицирован так, чтобы аминокислоты, соответствующие положениям 79 и 80 были заменены пептидом, имеющим аминокислотную последовательность Gly-Gly-Lys-Gly-Gly (SEQ ID NO: 18), получая в результате полипептид HBcAg, имеющий последовательность, показанную в SEQ ID NO: 29). Указанные композиции особенно применимы в таких вариантах, в которых антигенная детерминанта связаны с VLP HBcAg. В следующих предпочтительных вариантах остатки цистеина в положениях 48 и 107 SEQ ID NO: 28 мутированы в серин. Изобретение, кроме того, включает в себя композиции, содержащие соответствующие полипептиды, имеющие аминокислотные последовательности, показанные в любом из упомянутых выше вариантов предшественника корового антигена гепатита B, которые также имеют указанные выше изменения аминокислот. Кроме того, в объем изобретения включены дополнительные варианты HBcAg, которые способны ассоциировать с образованием капсида или VLP и имеют указанные выше изменения аминокислот. Таким образом, изобретение, кроме того, включает в себя композиции и соответственно композиции вакцин, содержащие полипептиды HBcAg, которые содержат, или альтернативно состоят из аминокислотных последовательностей, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны любой из аминокислотных последовательностей дикого типа, и формы указанных белков, которые были процессированы в подходящих для этого случаях, чтобы удалить N-концевую лидерную последовательность, и модифицированы указанными выше изменениями.
Композиции или композиции вакцин согласно изобретению могут содержать смеси различных HBcAg. Таким образом, указанные композиции вакцин могут состоять из HBcAg, которые отличаются по аминокислотной последовательности. Например, могут быть получены композиции вакцин, содержащие HBcAg «дикого типа» и модифицированный HBcAg, в котором один или несколько аминокислотных остатков были изменены (например, делетированы, встроены или заменены). Кроме того, предпочтительными композициями вакцин согласно изобретению являются композиции, которые представляют собой высокоупорядоченную и повторяющуюся антигенную матрицу, в которой антигеном является аналог пептида Melan-A меланомы человека.
Как описано выше, изобретение частично основано на неожиданном обнаружении того, что иммуностимулирующие вещества, предпочтительно иммуностимулирующие нуклеиновые кислоты и еще более предпочтительно ДНК-олигонуклеотиды или альтернативно поли(I:C) могут быть упакованы в VLP. Неожиданно нуклеиновые кислоты, присутствующие в VLP, могут быть, в частности, заменены иммуностимулирующими веществами, предпочтительно иммуностимулирующими нуклеиновыми кислотами и еще более предпочтительно ДНК-олигонуклеотидами, содержащими мотивы CpG или поли (I:C). В качестве примера CpG-VLP являются более иммуногенными и вызывают более специфичные эффекты, чем их аналоги, не содержащие CpG, и индуцируют усиленные B- и T-клеточные ответы. Иммунный ответ против антигенов, связанных, слитых или иным образом соединенных с VLP, подобным образом усилен, так же как и иммунный ответ против самого VLP. Кроме того, T-клеточные ответы как против VLP, так и против антигенов, главным образом, направлены к типу Th1. Кроме того, упакованные нуклеиновые кислоты и CpG соответственно защищены от деградации, т.е. они являются более стабильными. Кроме того, неспецифичная активация клеток врожденной иммунной системы значительно снижена.
Врожденная иммунная система обладает способностью узнавать инвариантную молекулярную картину, общую для патогенных микроорганизмов. Недавние исследования показали, что это узнавание является ключевой стадией в индукции эффективных иммунных ответов. Основной механизм, посредством которого продукты микроорганизмов усиливают иммунные ответы, заключается в стимуляции APC, особенно дендритных клеток, к продуцированию провоспалительных цитокинов и экспрессии высоких уровней костимулирующих молекул для T-клеток. Указанные активированные дендритные клетки затем инициируют первичные T-клеточные ответы и обусловливают тип опосредованной T-клетками эффекторной функции.
Два класса нуклеиновых кислот, а именно 1) бактериальная ДНК, которая содержит иммуностимулирующие последовательности, в частности неметилированные CpG-динуклеотиды со специфичными фланкирующими основаниями (называемыми мотивами CpG), и 2) двунитевая РНК, синтезированная различными типами вирусов, являются важными представителями компонентов микроорганизмов, которые усиливают иммунные ответы. Синтетическая двунитевая (дн) РНК, такая как полиинозиновая-полицитидиловая кислота (поли I:C), способна индуцировать дендритные клетки к продукции провоспалительных цитокинов и экспрессии высоких уровней костимулирующих молекул.
В серии исследований Tokunaga и Yamamoto и соавторов показано, что бактериальная ДНК или синтетические олигодезоксинуклеотиды индуцируют PBMC человека и клетки селезенки мыши к продукции интерферона типа I (IFN) (обзор в Yamamoto et al., Springer Semin Immunopathol. 22: 11-19). Поли(I:C) исходно синтезировали в качестве сильного индуктора IFN типа I, но она также индуцирует другие цитокины, такие как IL-12.
Предпочтительная рибонуклеиновая кислота включает двунитевую РНК, представляющую собой полиинозиновую-полицитидиловую кислоту (поли I:C). Рибонуклеиновые кислоты и их модификации, а также способы их получения описаны Levy, H. B (Methods Enzymol. 1981, 78: 242-251), DeClercq, E. (Methods Enzymol. 1981, 78: 227-236) и Torrence, P.F. (Methods Enzymol, 1981; 78: 326-331) и в ссылках в указанных работах. Следующие предпочтительные рибонуклеиновые кислоты содержат полинуклеотиды инозиновой кислоты и цитидииловой кислоты, такие как поли(IC), две нити которых образуют двунитевую РНК. Рибонуклеиновые кислоты могут быть выделены из организмов. Кроме того, рибонуклеиновые кислоты также охватывают синтетические рибонуклеиновые кислоты, в частности синтетические олигонуклеотиды поли(I:C), которым была придана резистентность к нуклеазам посредством модификации фосфодиэфирного остова, в частности фосфоротиоатными модификациями. В следующем варианте рибозный остов поли(I:C) заменяют дезоксирибозой. Специалистам в данной области известны способы, как синтезировать синтетические олигонуклеотиды.
В другом предпочтительном варианте изобретения включены молекулы, которые активируют toll-подобные рецепторы (TLR). До настоящего времени известно десять toll-подобных рецепторов человека. Они активируются множеством лигандов. TLR2 активируется пептидогликанами, липопротеидами, липополисахаридами, липотейхоновой кислотой и зимозаном и активирующим макрофаги липопептидом MALP-2; TLR3 активируется двунитевой РНК, такой как поли(I:C); TLR4 активируется липополисахаридом, липотейхоновыми кислотами и таксолом и белками теплового шока, такими как белок HSP-60 и Gp96; TLR5 активируется жгутиками бактерий, особенно белком флагеллином; TLR6 активируется пептидогликанами, TLR7 активируется соединениями имихимоидома и имидазохинолина, такими как R-848, локсорибин и бропиримин, и TLR9 активируется бактериальной ДНК, в частности CpG-ДНК. Лиганды TLR1, TLR8 и TLR10 до настоящего времени не известны. Однако недавние сообщения показывают, что одни и те же рецепторы могут взаимодействовать с разными лигандами и что имеются дополнительные рецепторы. Приведенный выше список лигандов не является исчерпывающим, и специалисту в данной области известны дополнительные лиганды.
Предпочтительно неметилированный CpG-содержащий олигонуклеотид содержит последовательность:
5'X1X2CGX3X4 3',
где X1, X2, X3 и X4 означают любой нуклеотид. Кроме того, олигонуклеотид может содержать примерно от 6 до примерно 100000 нуклеотидов, предпочтительно примерно от 6 до примерно 2000 нуклеотидов, более предпочтительно примерно от 20 до примерно 2000 нуклеотидов и еще более предпочтительно содержит примерно от 20 до примерно 300 нуклеотидов. Кроме того, олигонуклеотид может содержать более чем от 100 до примерно 2000 нуклеотидов, предпочтительно более чем от 100 до примерно 1000 нуклеотидов и более предпочтительно более чем от 100 до примерно 500 нуклеотидов.
В предпочтительном варианте CpG-содержащий олигонуклеотид содержит одну или несколько фосфоротиоатных модификаций фосфатного остова. Например, CpG-содержащий олигонуклеотид, имеющий одну или несколько модификаций фосфатного остова или имеющий весь модифицированный фосфатный остов, и CpG-содержащий олигонуклеотид, в котором одна, несколько или все модификации нуклеотидного фосфатного остова являются фосфоротиоатными модификациями, включены в объем настоящего изобретения. Таким образом, в предпочтительном варианте по меньшей мере один из нуклеотидов X1, X2, X3 и X4 имеет модификацию фосфатного остова.
В следующем очень предпочтительном варианте настоящего изобретения иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом указанный неметилированный CpG-содержащий олигонуклеотид имеет последовательность нуклеиновой кислоты, выбранную из группы, состоящей из (a) GGGGACGATCGTCGGGGGG ((SEQ ID NO: 2), и обычно сокращенно называемой в данном описании G3-6), (b) GGGGGACGATCGTCGGGGGG ((SEQ ID NO: 3); и обычно сокращенно называемой в данном описании G4-6), (c) GGGGGGACGATCGTCGGGGGG ((SEQ ID NO: 4); и обычно сокращенно называемой в данном описании G5-6), (d) GGGGGGGACGATCGTCGGGGGG ((SEQ ID NO: 5); и обычно сокращенно называемой в данном описании G6-6), (e) GGGGGGGGACGATCGTCGGGGGGG ((SEQ ID NO: 6); и обычно сокращенно называемой в данном описании G7-7), (f) GGGGGGGGGACGATCGTCGGGGGGGG ((SEQ ID NO: 7); и обычно сокращенно называемой в данном описании G8-8), (g) GGGGGGGGGGACGATCGTCGGGGGGGGG ((SEQ ID NO: 8); и обычно сокращенно называемой в данном описании G9-9), (h) GGGGGGCGACGACGATCGTCGTCGGGGGGG ((SEQ ID NO: 9); и обычно сокращенно называемой в данном описании G6), (i) tccatgacgttcctgaataat ((SEQ ID NO: 34); и обычно сокращенно называемой в данном описании CyCpGpt),(j) TCCATGACGTTCCTGAATAAT ((SEQ ID NO: 35); и обычно сокращенно называемой в данном описании CyCpG), (k) tccatgacgttcctgacgtt ((SEQ ID NO: 36); и обычно сокращенно называемой в данном описании B-CpGpt), (1) TCCATGACGTTCCTGACGTT ((SEQ ID NO: 37); и обычно сокращенно называемой в данном описании B-CpG), (m) ggggtcaacgttgaggggg ((SEQ ID NO: 38); и обычно сокращенно называемой в данном описании NKCpGpt), (n) GGGGTCAACGTTGAGGGGG ((SEQ ID NO: 39); и обычно сокращенно называемой в данном описании NKCpG), (o) attattcaggaacgtcatgga ((SEQ ID NO: 40); и обычно сокращенно называемой в данном описании CyCpG-rev-pt), (p) GGGGGGGGGGGACGATCGTCGGGGGGGGGG ((SEQ ID NO: 41); и обычно сокращенно называемой в данном описании g10gacga-PO (G10-PO)), (q) gggggggggggacgatcgtcgggggggggg ((SEQ ID NO: 42); и обычно сокращенно называемой в данном описании g10gacga-PS (G10-PS)), (r) CGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGAAATGCATGTCAAAGACAGCAT ((SEQ ID NO: 43); и обычно сокращенно называемой в данном описании (CpG)20OpA), (s) TCCATGACGTTCCTGAATAATCGC GCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCG ((SEQ ID NO: 44); и обычно сокращенно называемой в данном описании Cy(CpG)20), (t) TCCATGACGTTCCTGAATAATCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGCGAAATGCATGTCAAAGACAGCAT ((SEQ ID NO: 45); и обычно сокращенно называемой в данном описании Cy(CpG)20-OpA), (u) TCCATGACGTTCCTGAATAATAAATGCATGTCAAAGACAGCAT ((SEQ ID NO: 46); и обычно сокращенно называемой в данном описании CyOpA), (v) TCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAAT ((SEQ ID NO: 47); и обычно сокращенно называемой в данном описании CyCyCy), (w) TCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTGGATGACGTTGGTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTCC ((SEQ ID NO: 48); и обычно сокращенно называемой в данном описании Cy150-1), и (x) CTAGAACTAGTGGATC CCCCGGGCTGCAGGAATTCGATTCATGACTTCCTGAATAATTCCATGACGTTGGTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAATAATTCCATGACGTTCCTGAAAATTCCAATCAAGCTTATCGATACCGTCGACC (SEQ ID NO: 49), и обычно сокращенно называемой в данном описании dsCyCpG-253 (комплементарная нить не показана). Маленькие буквы, которые показаны в перечисленных выше последовательностях от SEQ ID NO:34 до SEQ ID NO: 49, означают дезоксинуклеотиды, связанные посредством фосфоротиоатных связей, тогда как большие буквы означают дезоксинуклеотиды, связанные посредством фосфодиэфирных связей.
И в следующем очень предпочтительном варианте настоящего изобретения иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом указанный неметилированный CpG-содержащий олигонуклеотид имеет последовательность нуклеиновой кислоты GGGGGGGGGGGACGATCGTCGGGGGGGGGG ((SEQ ID NO: 41); и обычно сокращенно называемой в данном описании g10gacga-PO или G10-PO).
CpG-содержащий олигонуклеотид также может быть рекомбинантным, геномным, синтетическим, кДНК, полученным из плазмиды и одно- или двунитевым. Для применения в настоящем изобретении нуклеиновые кислоты могут быть синтезированы de novo с использованием ряда способов, хорошо известных в данной области. Например, способ на основе β-цианоэтилфосфорамидита (Beaucage, S. L., and Caruthers, M. H., Tet. Let. 22: 1859 (1981); способ на основе нуклеозид-H-фосфоната (Garegg et al., Tet. Let. 27:4051-4054 (1986); Froehler et al., Nucl. Acid. Res. 14:5399-5407 (1986); Garegg et al., Tet. Let. 27:4055-4058 (1986), Gaffney et al., Tet. Let. 29:2619-2622 (1988)). Указанные химические реакции могут быть осуществлены с использованием множества автоматизированных синтезаторов олигонуклеотидов, имеющихся на рынке. Альтернативно CpG могут быть получены в большом масштабе в плазмидах (смотри Sambrook, Т., et al., "Molecular Cloning: A Laboratory Manual," Cold Spring Harbor laboratory Press, New York, 1989), которые после введения субъекту деградируют до олигонуклеотидов. Олигонуклеотиды могут быть получены из существующих последовательностей нуклеиновых кислот (например, геномной или кДНК) с использованием известных способов, таких как способы на основе применения ферментов рестрикции, экзонуклеаз или эндонуклеаз.
Иммуностимулирующие вещества, иммуностимулирующие нуклеиновые кислоты, а также неметилированный CpG-содержащий олигонуклеотид, могут быть связаны с VLP любым способом, известным в данной области, при условии что композиция усиливает иммунный ответ у животного. Например, олигонуклеотид может быть связан либо ковалентно, либо нековалентно. Кроме того, VLP могут полностью или частично заключать внутри себя иммуностимулирующие вещества, иммуностимулирующие нуклеиновые кислоты, а также неметилированный CpG-содержащий олигонуклеотид. Предпочтительно иммуностимулирующая нуклеиновая кислота, а также неметилированный CpG-содержащий олигонуклеотид могут быть связаны с сайтом VLP, таким как сайт связывания олигонуклеотида (либо природного, либо неприродного происхождения), сайт связывания ДНК или сайт связывания РНК. В другом варианте сайт VLP содержит богатый аргинином повтор или богатый лизином повтор.
Одним из конкретных применений композиций согласно изобретению является активация дендритных клеток в целях усиления специфичного иммунного ответа против антигенов. Иммунный ответ может быть усилен с использованием методик ex vivo или in vivo. Способ ex vivo можно использовать на аутологичных или гетерологичных клетках, но предпочтительно используют на аутологичных клетках. В предпочтительных вариантах дендритные клетки выделяют из периферической крови или костного мозга, но они могут быть выделены из любого источника дендритных клеток. Манипуляции ex vivo с дендритными клетками в целях иммунотерапии злокачественной опухоли описаны в нескольких публикациях в данной области, включая Engleman, E. G., Cytotechnology 25:1 (1997); Van Schooten, W., et al., Molecular Medicine Today, June, 255 (1997); Steinman, R. M., Experimental Hematology 24:849 (1996); and Gluckman, J. C, Cytokines, Cellular and Molecular Therapy 3:187(1997).
Дендритные клетки также могут быть подвергнуты контакту с предлагаемыми в изобретении композициями с использованием способов in vivo. Чтобы это осуществить, CpG вводят в комбинации с VLP, необязательно связанными, слитыми или иным образом соединенными с антигеном, непосредственно субъекту, нуждающемуся в иммунотерапии. В некоторых вариантах предпочтительно, чтобы VLP/CpG были введены в локальную область опухоли, что можно осуществить любым способом, известным в данной области, например прямой инъекцией в опухоль.
Предпочтительный вариант настоящего изобретения относится к композиции для усиления иммунного ответа у животного, содержащей (a) вирусоподобную частицу; (b) по меньшей мере одно иммуностимулирующие вещество; и (c) по меньшей мере один антиген или антигенную детерминанту; где указанный антиген или указанная антигенная детерминанта связаны с указанной вирусоподобной частицей, и где указанный антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека, и где указанное иммуностимулирующие вещество связано с указанной вирусоподобной частицей, и где указанным иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом мотив CpG указанного неметилированного CpG-содержащего олигонуклеотида является частью палиндромной последовательности, где указанной палиндромной последовательностью является GACGATCGTC (SEQ ID NO: 1), и где указанная палиндромная последовательность фланкирована на своем 3'-конце и на своем 5'-конце более чем двумя, но менее чем 11 гуанозиновыми единицами или более предпочтительно 8-10 гуанозиновыми единицами или наиболее предпочтительно 10 гуанозиновыми единицами.
Авторы обнаружили, что предлагаемые в изобретении иммуностимулирующие вещества, т.е. неметилированные CpG-содержащие олигонуклеотиды, где мотив CpG указанных неметилированных CpG-содержащих олигонуклеотидов является частью палиндромной последовательности, где палиндромной последовательностью является GACGATCGTC (SEQ ID NO: 1) и где палиндромная последовательность фланкирована на своем 3'-конце и на своем 5'-конце более чем двумя, но менее чем 11 гуанозиновыми единицами, более предпочтительно 8-10 гуанозиновыми единицами или наиболее предпочтительно 10 гуанозиновыми единицами, в частности, эффективны при стимуляции иммунных клеток in vitro.
В предпочтительном варианте настоящего изобретения палиндромная последовательность содержит, или альтернативно в основном состоит, или альтернативно состоит из GACGATCGTC (SEQ ID NO: 1), при этом указанная палиндромная последовательность фланкирована на своем 5'-конце по меньшей мере 3 и самое большее 10 гуанозиновыми единицами, и где указанная палиндромная последовательность фланкирована на своем 3'-конце по меньшей мере 6 и самое большее 10 гуанозиновыми единицами. В другом варианте палиндромная последовательность фланкирована на своем 5'-конце по меньшей мере 3 и самое большее 10 гуанозиновыми единицами и где указанная палиндромная последовательность фланкирована на своем 3'-конце по меньшей мере 6 и самое большее 10 гуанозиновыми единицами.
В следующем очень предпочтительном варианте настоящего изобретения иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом мотив CpG указанного неметилированного CpG-содержащего олигонуклеотида является частью палиндромной последовательности, где указанный неметилированный CpG-содержащий олигонуклеотид имеет последовательность нуклеиновой кислоты, выбранную из (a) GGGGACGATCGTCGGGGGG ((SEQ ID NO: 2); и обычно сокращенно называемой в данном описании G3-6), (b) GGGGGACGATCGTCGGGGGG ((SEQ ID NO: 3); и обычно сокращенно называемой в данном описании G4-6), (c) GGGGGGACGATCGTCGGGGGG ((SEQ ID NO: 4); и обычно сокращенно называемой в данном описании G5-6), (d) GGGGGGGACGATCGTCGGGGGG ((SEQ ID NO: 5); и обычно сокращенно называемой в данном описании G6-6), (e) GGGGGGGGACGATCGTCGGGGGGG ((SEQ ID NO: 6); и обычно сокращенно называемой в данном описании G7-7), (f) GGGGGGGGGACGATCGTCGGGGGGGG ((SEQ ID NO: 7); и обычно сокращенно называемой в данном описании G8-8), (g) GGGGGGGGGGACGATCGTCGGGGGGGGG ((SEQ ID NO: 8); и обычно сокращенно называемой в данном описании G9-9), (h) GGGGGGCGACGACGATCGTCGTCGGGGGGG ((SEQ ID NO: 9); и обычно сокращенно называемой в данном описании G6), и (i) GGGGGGGGGGGACGATCGTCGGGGGGGGGG ((SEQ ID NO: 41); и обычно сокращенно называемой в данном описании G10-PO).
В следующем предпочтительном варианте настоящего изобретение иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом мотив CpG указанного неметилированного CpG-содержащего олигонуклеотида является частью палиндромной последовательности, где указанной палиндромной последовательностью является GACGATCGTC (SEQ ID NO: 1), и где указанная палиндромная последовательность фланкирована на своем 5'-конце по меньшей мере 4 и самое большее 10 гуанозиновыми единицами, и где указанная палиндромная последовательность фланкирована на своем 3'-конце по меньшей мере 6 и самое большее 10 гуанозиновыми единицами.
В другом предпочтительном варианте настоящего изобретения иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом мотив CpG указанного неметилированного CpG-содержащего олигонуклеотида является частью палиндромной последовательности, где указанный неметилированный CpG-содержащий олигонуклеотид имеет последовательность нуклеиновой кислоты, выбранную из (a) GGGGGACGATCGTCGGGGGG ((SEQ ID NO: 3); обычно называемой в данном описании G4-6), (b) GGGGGGACGATCGTCGGGGGG ((SEQ ID NO: 4); и обычно сокращенно называемой в данном описании G5-6), (c) GGGGGGGACGATCGTCGGGGGG((SEQ ID NO: 5); и обычно сокращенно называемой в данном описании G6-6), (d) GGGGGGGGACGATCGTCGGGGGGG ((SEQ ID NO: 6); и обычно сокращенно называемой в данном описании G7-7), (e) GGGGGGGGGACGATCGTCGGGGGGGG ((SEQ ID NO: 7); и обычно сокращенно называемой в данном описании G8-8), (f) GGGGGGGGGGACGATCGTCGGGGGGGGG ((SEQ ID NO: 8); и обычно сокращенно называемой в данном описании G9-9); и (g) GGGGGGGGGGGACGATCGTCGGGGGGGGGG ((SEQ ID NO: 41); и обычно сокращенно называемой в данном описании G10-PO).
В следующем предпочтительном варианте настоящего изобретения иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом мотив CpG указанного неметилированного CpG-содержащего олигонуклеотида является частью палиндромной последовательности, где указанной палиндромной последовательностью является GACGATCGTC (SEQ ID NO: 1), и где указанная палиндромная последовательность фланкирована на своем 5'-конце по меньшей мере 5 и самое большее 8 гуанозиновыми единицами, и где указанная палиндромная последовательность фланкирована на своем 3'-конце по меньшей мере 6 и самое большее 10 гуанозиновыми единицами.
Экспериментальные данные показывают, что простота упаковки предпочтительных предлагаемых в изобретении иммуностимулирующих веществ, т.е. фланкированных гуанозином, палиндромных и неметилированных CpG-содержащих олигонуклеотидов, где палиндромной последовательностью является GACGATCGTC (SEQ ID NO: 1), и где палиндромная последовательность фланкирована на своем 3'-конце и на своем 5'-конце менее чем 11 или менее чем 10 гуанозиновыми единицами, в VLP увеличивается, если палиндромные последовательности фланкированы меньшим количеством гуанозиновых единиц. Однако уменьшение количества гуанозиновых единиц, фланкирующих палиндромные последовательности, приводит к уменьшению стимулирующих клеток крови in vitro. Таким образом, платой за способность к упаковке является пониженная биологическая активность указанных предлагаемых в изобретении иммуностимулирующих веществ. Таким образом, данные предпочтительные варианты представляют собой компромисс между способностью к упаковке и биологической активностью.
В другом предпочтительном варианте настоящего изобретения иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом мотив CpG указанного неметилированного CpG-содержащего олигонуклеотида является частью палиндромной последовательности, где указанный неметилированный CpG-содержащий олигонуклеотид имеет последовательность нуклеиновой кислоты, выбранную из (a) GGGGGGACGATCGTCGGGGGG ((SEQ ID NO: 4); и обычно сокращенно называемой в данном описании G5-6), (b) GGGGGGGACGATCGTCGGGGGG ((SEQ ID NO: 5); и обычно сокращенно называемой в данном описании G6-6), (c) GGGGGGGGACGATCGTCGGGGGGG ((SEQ ID NO: 6); и обычно сокращенно называемой в данном описании G7-7), (d) GGGGGGGGGACGATCGTCGGGGGGGG ((SEQ ID NO: 7); и обычно сокращенно называемой в данном описании G8-8); и (e) GGGGGGGGGGGACGATCGTCGGGGGGGGGG ((SEQ ID NO: 41); и обычно сокращенно называемой в данном описании G10-PO).
В предпочтительном варианте настоящего изобретения иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом мотив CpG указанного неметилированного CpG-содержащего олигонуклеотида является частью палиндромной последовательности, где указанный неметилированный CpG-содержащий олигонуклеотид имеет последовательность нуклеиновой кислоты SEQ ID NO: 7, т.е. иммуностимулирующим веществом является G8-8, или SEQ ID NO: 41, т.е. G10-PO.
В очень предпочтительном варианте настоящего изобретения иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, при этом мотив CpG указанного неметилированного CpG-содержащего олигонуклеотида является частью палиндромной последовательности, где указанный неметилированный CpG-содержащие олигонуклеотид имеет последовательность нуклеиновой кислоты SEQ ID NO: 41, т.е. иммуностимулирующим веществом является G10-PO. Таким образом, в очень предпочтительном варианте настоящее изобретение относится к композиции для усиления иммунного ответа у животного, содержащей (a) вирусоподобную частицу; (b) по меньшей мере одно иммуностимулирующие вещество; и (c) по меньшей мере один антиген или антигенную детерминанту; где указанный антиген связан с указанной вирусоподобной частицей, и где указанный антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека, и где указанное иммуностимулирующие вещество связано с указанной вирусоподобной частицей, и где указанным иммуностимулирующим веществом является неметилированный CpG-содержащий олигонуклеотид, где мотив CpG указанного неметилированного CpG-содержащего олигонуклеотида является частью палиндромной последовательности, где указанной палиндромной последовательностью является GACGATCGTC (SEQ ID NO: 1), и где указанная палиндромная последовательность фланкирована на своем 3'-конце и на своем 5'- конце 10 гуанозиновыми единицами.
Как указано выше, оптимальная последовательность, используемая для упаковки в VLP, представляет собой компромисс между способностью к упаковке и биологической активностью. Принимая это во внимание, иммуностимулирующее вещество G8-8 является предпочтительным, и иммуностимулирующее вещество G10-PO является очень предпочтительным вариантом настоящего изобретения, так как они являются высокобиологически активными и при этом еще довольно хорошо подвергаются упаковке.
Предлагаемая в изобретении композиция, кроме того, содержит аналог пептида Melan-A меланомы человека согласно изобретению, связанный с вирусоподобной частицей.
В следующем предпочтительном варианте изобретения по меньшей мере один аналог пептида Melan-A слит с вирусоподобной частицей. Как указано выше, VLP обычно состоит, по меньшей мере, из одной субъединицы, собирающейся в VLP. Таким образом, снова в следующем предпочтительном варианте изобретения аналог пептида Melan-A слит по меньшей мере с одной субъединицей вирусоподобной частицы или белка, который может быть включен в VLP, с образованием химерного слияния VLP-субъединица-антиген.
Слияние аналога пептида Melan-A можно осуществить путем инсерции в последовательность субъединицы VLP или путем слияния либо с N-, либо с C-концом VLP-субъединицы или белка, который может быть включен в VLP. В дальнейшем при упоминании слитых белков пептида с субъединицей VLP подразумевают слияние либо с концами последовательности субъединицы, либо внутреннюю инсерцию пептида в последовательность субъединицы.
Слияние также можно осуществить путем инсерции последовательностей аналога пептида Melan-A в вариант субъединицы VLP, где часть последовательности субъединицы была делетирована, которые далее называют укороченными мутантами. Укороченные мутанты могут иметь N- или C-концевые или внутренние делеции части последовательности субъединицы VLP. Например, конкретный HBcAg VLP, например, с делецией аминокислотных остатков с 79 по 81, является укороченным мутантом с внутренней делецией. Слияние антигенов или антигенных детерминант либо с N-, либо с C-концом укороченных мутантов VLP-субъединиц также приводит к образованию вариантов согласно изобретению. Подобным образом слияние эпитопа с последовательностью субъединицы VLP также можно осуществить путем замены, при которой, например, для конкретного HBcAg VLP аминокислоты 79-81 заменяют чужеродным эпитопом. Таким образом, слияние, которое упоминается в дальнейшем, можно осуществить путем инсерции последовательности аналога пептида Melan-A в последовательность субъединицы VLP путем замены части последовательности субъединицы VLP аналогом пептида Melan-A, или посредством комбинирования делеции, замены или инсерций.
Химерные конструкции аналог пептида Melan-A-субъединица VLP, как правило, будут способны к самосборке в VLP. VLP, экспонирующие эпитопы, слитые с их субъединицами, также называют в данном описании химерными VLP. Как указано, вирусоподобная частица содержит или, альтернативно, состоит, по меньшей мере, из одной субъединицы VLP. В следующем варианте изобретения вирусоподобная частица содержит или, альтернативно, состоит из смеси химерных субъединиц VLP и нехимерных субъединиц VLP, т.е. субъединиц VLP, не имеющих слитого с ними антигена, образуя в результате так называемые мозаичные частицы. Это может иметь преимущества для обеспечения образования и сборки в VLP. В указанных вариантах доля химерных VLP-субъединиц может составлять 1, 2, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 95% или больше.
Могут быть добавлены фланкирующие аминокислотные остатки к любому концу последовательности пептида или эпитопа, который необходимо слить с любым из концов последовательности субъединицы VLP, или для внутренней инсерции такой пептидной последовательности в последовательность субъединицы VLP. Остатки глицина и серина являются особенно предпочтительными аминокислотами для использования во фланкирующих последовательностях, добавляемых к пептиду, который необходимо слить. Остатки глицина придают дополнительную гибкость, которая может снижать возможное дестабилизирующее действие слияния чужеродной последовательности в последовательность субъединицы VLP.
В конкретном варианте изобретения VLP является VLP корового антигена гепатита B. Описаны слитые белки либо с N-концом HBcAg (Neyrinck, S. et al., Nature Med. 5: 1157-1163 (1999)), либо инсерции в так называемую основную иммунодоминантную область (MIR) (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)), WO 01/98333), и указанные слияния являются предпочтительными вариантами изобретения. Также описаны встречающиеся в природе варианты HBcAg с делециями в MIR (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001), публикация специально включена в виде ссылки в полном объеме), и слияния с N- или C-концом, а также инсерции в положение MIR, соответствующее сайту делеции по сравнению с HBcAg дикого типа являются следующими вариантами изобретения. Также описаны слияния с C-концом (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)). Специалист в данной области легко найдет руководство по поводу того, как конструировать слитые белки, используя классические способы молекулярной биологии (Sambrook, J. et al., eds., Molecular Cloning, A Laboratory Manual, 2nd. edition, Cold Spring Habor Laboratory Press, Cold Spring Harbor, N.Y. (1989), Ho et al., Gene 77:51 (1989)). Описаны векторы и плазмиды, кодирующие HBcAg и слитые белки HBcAg и применимые для экспрессии HBcAg и слитых белков HBcAg (Pumpens, P. and Grens, E. Intervirology 44: 98-114 (2001), Neyrinck, S. et al., Nature Med. 5: 1157-1163 (1999)), которые можно использовать в практике изобретения. Важным фактором для оптимизации эффективности самосборки и экспонирования эпитопа, встраиваемого в MIR HBcAg, является выбор места инсерции, а также аминокислот, которые необходимо делетировать из последовательности HBcAg в MIR (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001); EP 421635; патент США 6231864) при инсерции, или другими словами, какие аминокислоты из HBcAg необходимо заменить новым эпитопом. Например, описана замена аминокислот HBcAg 76-80, 79-81, 79-80, 75-85 или 80-81 чужеродными эпитопами (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001); EP 0421635; патент США 6231864). HBcAg содержит длинный аргининовый хвост (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)), который не имеет существенного значения для сборки капсида и способен к связыванию нуклеиновых кислот (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)). HBcAg, либо содержащие аргининовый хвост, либо не имеющие такового, являются вариантами изобретения.
В следующем предпочтительном варианте изобретения VLP является VLP РНК-фага. Основные белки оболочки РНК-фагов спонтанно собираются в VLP при экспрессии в бактериях и, а частности, в E. coli. Конкретные примеры белков оболочки бактериофагов, которые можно использовать для получения композиций согласно изобретению, включают белки оболочки таких РНК-бактериофагов, как бактериофаг Qβ (SEQ ID NO: 10; база данных PIR, No. доступа VCBPQβ, относящаяся к CP Qβ, и SEQ ID NO: 11; No. доступа AAA16663, относящаяся к белку A1 Qβ) и бактериофаг fr (SEQ ID NO: 4; PIR, No. доступа VCBPFR).
В другом предпочтительном варианте по меньшей мере один аналог пептида Melan-A сливают с белком оболочки Qβ. Описаны конструкции слитых белков, в которых эпитопы были слиты с C-концом укороченной формы белка A1 Qβ или встроены в белок A1 (Kozlovska, T. M., et al., Intervirology, 39: 9-15 (1996)). Белок A1 образуется при супрессии в стоп-кодоне UGA и имеет длину 329 а/к или 328 а/к, если принять во внимание отщепление N-концевого метионина. Отщепление N-концевого метионина перед аланином (вторая аминокислота, кодируемая геном CP Qβ) обычно имеет место в E. coli, и так обстоит дело для N-концов белков оболочки CP Qβ. Часть гена A1 с 3'-стороны от амбер-кодона UGA кодирует удлинение CP, которое имеет длину 195 аминокислот. Инсерция, по меньшей мере, одного аналога пептида Melan-A между положениями 72 и 73 удлинения CP приводит к образованию следующих вариантов согласно изобретению (Kozlovska, T. M., et al., Intervirology 39: 9-15 (1996)). Слияние аналога пептида Melan-A с C-концом укороченного на C-конце белка A1 Qβ приводит к образованию следующих предпочтительных вариантов согласно изобретению. Например, Kozlovska et al., (Intervirology, 39: 9-15 (1996)) описали слияния белка A1 Qβ, в которых эпитоп слит с C-концом удлинения CP Qβ, укороченным в положении 19.
Как описано Kozlovska et al. (Intervirology, 39: 9-15 (1996)), сборка частиц, экспонирующих слитые эпитопы, обычно требует присутствия как слияния белок A1-белок-аналог пептида Melan-A, так и CP дикого типа, чтобы образовать мозаичную частицу. Однако варианты, содержащие вирусоподобные частицы, и таким образом, в частности, VLP белка оболочки РНК-фага Qβ, которые состоят исключительно из субъединиц VLP, имеющих, по меньшей мере, один слитый с ними аналог пептида Melan-A, также входят в объем настоящего изобретения.
Получение мозаичных частиц можно осуществить рядом способов. Kozlovska et al., Intervirolog., 39: 9-15 (1996) описывают три способа, которые можно использовать в практике изобретения. В первом способе эффективное экспонирование слитого эпитопа на VLP опосредовано экспрессией плазмиды, кодирующей слияние белка A1 Qβ, имеющего стоп-кодон UGA между CP и удлинением CP, в штамме E. coli, несущем плазмиду, кодирующую клонированную тРНК супрессора UGA, который приводит к трансляции кодона UGA в Trp (плазмида pISM3001 (Smiley B.K., et al., Gene 134: 33-40 (1993))). В другом способе стоп-кодон гена CP модифицируют в UAA и проводят котрансфекцию со второй плазмидой, экспрессирующей слияние белка A1 с антигеном. Вторая плазмида кодирует резистентность к различным антибиотикам, и начало репликации совместимо с первой плазмидой (Kozlovska, T. M., et al., Intervirology 39: 9-15 (1996)). В третьем способе CP и слияние белок A1-антиген кодируются бицистронным образом при оперативном связывании с таким промотором, как промотор Trp, как описано на фигуре 1 в Kozlovska et al., Intervirology, 39: 9-15 (1996).
В следующем варианте аналог пептида Melan-A встраивают между аминокислотами 2 и 3 (нумерация CP после расщепления, то есть в том случае, когда N-концевой метионин отщеплен) CP fr, таким образом, получая слитый белок аналог пептида Melan-A-CP fr. Описаны векторы и экспрессирующие системы для конструирования и экспрессии слитых белков CP fr, самособирающихся в VLP и применимых в практике изобретения (Pushko P. et al., Prot. Eng. 6: 883-891 (1993)). В конкретном варианте последовательность аналога пептида Melan-A встраивают в делеционный вариант CP fr после аминокислоты 2, когда остатки 3 и 4 CP fr делетированы.
Слияние эпитопов с N-концевой выступающей β-шпилькой белка оболочки РНК-фага MS-2 и последующая презентация слитого эпитопа на образованной при самосборке VLP РНК-фага MS-2 также описаны (WO 92/13081), и слияние аналога пептида Melan-A согласно изобретению путем инсерции или замены в белке оболочки РНК-фага MS-2 также попадает в объем изобретения.
В другом варианте изобретения аналоги пептида Melan-A согласно изобретению сливают с белком капсида вируса папилломы. В более конкретном варианте аналоги пептида Melan-A согласно изобретению сливают с основным белком капсида L1 вируса папилломы крупного рогатого скота типа 1 (BPV-1). Описаны векторы и экспрессирующие системы для конструирования и экспрессии слитых белков BPV-1 в системах бакуловирус/клетки насекомых (Chackerian, B. et al., Proc. Natl. Acad. Sci. USA 96: 2373-2378 (1999), WO 00/23955). Замена аминокислот 130-136 L1 BPV-1 аналогами пептида Melan-A согласно изобретению приводит к слитому белку L1 BPV-1-аналог пептида Melan-A, который является предпочтительным вариантом изобретения. Описано клонирование в бакуловирусном векторе и экспрессия в инфицированных бакуловирусом клетках Sf9, которые могут быть использованы в практике изобретения (Chackerian, B. et al., Proc. Natl. Acad. Sci. USA 96: 2373-2378 (1999), WO 00/23955). Очистку собранных частиц, экспонирующих слитый аналог пептида Melan-A согласно изобретению, можно осуществить рядом способов, таких, например, как гель-фильтрация или ультрацентрифугирование в градиенте сахарозы (Chackerian, B. et al., Proc. Natl. Acad. Sci. USA 96: 2373-2378 (1999), WO 00/23955).
В следующем варианте изобретения аналоги пептида Melan-A согласно изобретению сливают с белком Ty, способным включаться в VLP Ty. В более конкретном варианте аналоги пептида Melan-A согласно изобретению сливают с p1 или белком капсида, кодируемым геном TYA (Roth, J.F., Yeast 16: 785-795 (2000)). Дрожжевые ретротранспозоны Ty1, 2, 3 и 4 выделены из Saccharomyces cerevisiae, тогда как ретротранспозон Tf1 выделен из Schizosaccharomyces pombae.
Ретротранспозоны Ty1 и 2 родственны классу copia элементов растений и животных, тогда как Ty3 относится к семейству ретротранспозонов gypsy, которые родственны ретровирусам растений и животных. В ретротранспозоне Ty1 белок p1, также называемый Gag или капсидным белком, имеет длину 440 аминокислот. P1 расщепляется во время созревания VLP в положении 408, давая белок p2, важный компонент VLP.
Описаны слитые с p1 белки и векторы для экспрессии указанных слитых белков в дрожжах (Adams, S.E., et al., Nature 329: 68-70 (1987)). Таким образом, например, аналог пептида Melan-A согласно изобретению может быть слит с p1 путем инсерции последовательности, кодирующей аналог пептида Melan-A, в сайт BamH1 плазмиды pMA5620. Клонирование последовательностей, кодирующих чужеродные эпитопы, в векторе pMA5620 приводит к экспрессии слитых белков, содержащих аминокислоты 1-381 p1 Ty1-15, слитые C-терминально с N-концом чужеродного эпитопа. Подобным образом подразумевается, что N-концевое слияние аналогов пептида Melan-A согласно изобретению или внутренняя инсерция в последовательность p1 или замена части последовательности p1 также входит в объем изобретения. В частности, инсерция аналогов пептида Melan-A согласно изобретению в последовательность Ty между аминокислотами 30-31, 67-68, 113-114 и 132-133 p1-белка Ty (EP0677111) дает предпочтительные варианты согласно изобретению.
Следующими VLP, подходящими для слияния Melan-A или аналогов пептида Melan-A согласно изобретению, являются, например, частицы, подобные ретровирусам (WO9630523), Gag HIV2 (Kang, Y.C., et al, Biol. Chem. 380: 353-364 (1999)), вирус мозаики вигны (Taylor, K.M. et al., Biol. Chem. 380: 387-392 (1999)), VLP парвовируса VP2 (Rueda, P. et al., Virology 263: 89-99 (1999)), HBsAg (патент США 4722840, EP0020416B1).
Примерами химерных VLP, подходящими для практики изобретения, также являются VLP, описанные в Intervirology 39: 1 (1996). Следующими примерами VLP, предполагаемыми для применения в изобретении, являются: HPV-1, HPV-6, HPV-11, HPV-16, HPV-18, HPV-33, HPV-45, CRPV, COPV, GAG HIV, вирус табачной мозаики. Также получены вирусоподобные частицы SV-40, вируса полиомы, аденовируса, вируса простого герпеса, ротавируса и вируса Норуолк, и химерные VLP на основе указанных VLP также входят в объем данного изобретения.
Как указано, варианты, содержащие антигены, слитые с вирусоподобной частицей путем инсерции в последовательность вирусоподобной частицы, из которой построен мономер, также входят в объем настоящего изобретения. В некоторых случаях антигены могут быть встроены в форме мономера, образующего вирусоподобные частицы, содержащего делеции. В указанных случаях мономер, из которого построена вирусоподобная частица, может быть неспособен образовывать вирусоподобные структуры в отсутствие встроенного антигена.
В некоторых случаях можно использовать методику рекомбинантной ДНК, чтобы слить гетерологичный белок с белком VLP (Kratz, P. A., et al., Proc. Natl. Acad. Sci. USA 96: 1915 (1999)). Например, настоящее изобретение охватывает VLP рекомбинантно слитые или химически конъюгированные (включая как ковалентные, так и нековалентные конъюгации) с антигеном (или его частью, предпочтительно по меньшей мере из 10, 20 или 50 аминокислот) согласно настоящему изобретению, чтобы создать слитые белки или конъюгаты. Слияние не обязательно должно быть прямым, оно также может осуществляться через линкерные последовательности. В большинстве случаев когда эпитопы, либо слитые, конъюгированные, либо иным образом связанные с вирусоподобной частицей, используют в качестве антигенов согласно изобретению, обычно добавляют спейсерные или линкерные последовательности на одном или обоих концах эпитопов. Такие линкерные последовательности предпочтительно содержат последовательности, узнаваемые протеасомой, протеазами эндосом или другой везикулярной структурой клетки.
Одним из способов связывания является пептидная связь, при которой конъюгат может быть непрерывным полипептидом, т.е. слитым белком. В слитом белке согласно настоящему изобретению различные пептиды или полипептиды связаны в рамке друг с другом, образуя непрерывный полипептид. Таким образом, первая часть слитого белка содержит антиген или иммуноген, а вторая часть слитого белка, либо N-концевая, либо C-концевая по отношению к первой части, содержит VLP. Альтернативно внутренняя инсерция в VLP с необязательными линкерными последовательностями на обоих концах антигена также может быть использована согласно настоящему изобретению.
Когда используют HBcAg в качестве VLP, предпочтительно, чтобы антиген был связан с C-концом частицы HBcAg. Коровый антиген гепатита B (HBcAg), имеющий C-концевое слияние с ограниченным по MHC класса I пептидом p33, полученным из гликопротеида вируса лимфоцитарного хориоменингита (LCMV), может быть использован и обычно использовался в качестве модели антигена (HBcAg-p33). Белок HBc дикого типа длиной 185 аминокислот собирается в высокоструктурированные частицы, состоящие из 180 субъединиц, принимающие икосаэдрическую геометрию. Гибкость HBcAg и других VLP в отношении приема относительно крупных инсерций чужеродных последовательностей в разных положениях при сохранении способности образовывать структурированные капсиды хорошо подтверждено в литературе. Это делает HBc-VLP привлекательными кандидатами для разработки нереплицирующихся вакцин.
Гибкая линкерная последовательность (например, последовательность, содержащая полиглицин/полисерин, такая как [Gly4 Ser]2 (Huston et al., Meth. Enzymol. 203: 46-88 (1991)) может быть встроена в слитый белок между антигеном и лигандом. Также слитый белок может быть сконструирован так, чтобы он содержал «эпитопную метку», которая позволяет слитому белку связываться с антителом (например, моноклональным антителом), например, в целях мечения и очистки. Примером эпитопной метки является трипептид Glu-Glu-Phe, который узнается моноклональным антителом YL1/2.
Изобретение также относится к химерной ДНК, которая содержит последовательность, кодирующую VLP, и последовательность, кодирующую аналог пептида Melan-A. ДНК может быть экспрессирована, например, в клетках насекомых, трансформированных бакуловирусом, в дрожжах или в бактериях. Не существует ограничений в отношении систем экспрессии, большой выбор которых имеется для обычного применения. Предпочтительно используют систему, которая обеспечивает экспрессию белков в больших количествах. В общем, предпочтительными являются бактериальные системы экспрессии вследствие их эффективности. Одним из примеров бактериальной системы экспрессии, подходящей для применения в объеме настоящего изобретения, является система, описанная Clarke et al., J. Gen. Virol. 71: 1109-1117 (1990); Borisova et al., J. Virol. 67: 3696-3701 (1993); и Studier et al., Methods Enzymol. 185: 60-89 (1990). Примером подходящей дрожжевой системы экспрессии является система, описана Emr, Methods Enzymol. 185: 231-3 (1990); также подходящими являются бакуловирусные системы, которые использовали ранее для получения белков капсида. Можно использовать конститутивные или индуцируемые системы экспрессии. Посредством выбора и возможной модификации имеющихся систем экспрессии можно контролировать форму, в которой получают белок.
В конкретном варианте изобретения антиген, к которому требуется усиленный иммунный ответ, связывают, сливают или иным образом соединяют в рамке с белком капсида (кора) вируса гепатита B (HBcAg). Однако специалистам в данной области будет понятно, что в конструкции слитого белка согласно изобретению можно использовать другие вирусоподобные частицы.
В следующем предпочтительном варианте настоящего изобретения по меньшей мере один аналог пептида Melan-A связывают с вирусоподобной частицей посредством по меньшей мере одной ковалентной связи. Предпочтительно по меньшей мере один аналог пептида Melan-A связывают с вирусоподобной частицей по меньшей мере одной ковалентной связью, при этом указанная ковалентная связь является непептидной связью, приводящей к упорядоченной и повторяющейся матрице и конъюгату аналога пептида Melan-A-VLP соответственно. Указанная матрица или конъюгат аналога пептида Melan-A соответственно обычно и предпочтительно имеет повторяющуюся и упорядоченную структуру, так как по меньшей мере один аналог пептида Melan-A связан с VLP ориентированным образом. Предпочтительно 120 или больше, более предпочтительно 180 или больше, еще более предпочтительно больше 270 и еще более предпочтительно 360 или больше пептидов Melan-A согласно изобретению связывают с VLP. Образование повторяющейся и упорядоченной матрицы аналог пептида Melan-A-VLP и соответственно конъюгата обеспечивается ориентированным и направленным, а также определенным связыванием и соответственно присоединением, по меньшей мере, одного аналога пептида Melan-A к VLP, как будет понятно из нижеследующего. Кроме того, обычная присущая высокоповторяющаяся и организованная структура VLP преимущественно вносит вклад в экспонирование аналога пептида Melan-A высокоупорядоченным и повторяющемся образом, приводя к образованию высокоорганизованной и повторяющейся матрицы и соответственно конъюгата аналог пептида Melan-A-VLP.
Таким образом, предпочтительные конъюгаты и соответственно матрицы согласно изобретению отличаются от конъюгатов предшествующего уровня техники своей высокоорганизованной структурой, размерами и повторяемостью антигена на поверхности матрицы. Предпочтительный вариант данного изобретения, кроме того, позволяет осуществлять экспрессию частицы в экспрессирующем хозяине, обеспечивая правильную упаковку и сборку VLP, с которой затем связывают антиген, т.е., по меньшей мере один аналог пептида Melan-A.
В данном изобретении заявлены способы связывания или ассоциации аналога пептида Melan-A согласно изобретению с VLP. Как указано в одном аспекте данного изобретения, по меньшей мере один аналог пептида Melan-A согласно изобретению связан с VLP с помощью химического перекрестного связывания обычно и предпочтительно с использованием гетеробифункционального перекрестно сшивающего агента. В данной области известно несколько гетеробифункциональных перекрестно сшивающих агентов. В предпочтительных вариантах гетеробифункциональный перекрестно сшивающий агент содержит функциональную группу, которая может взаимодействовать с предпочтительными первыми сайтами связывания, т.е. с аминогруппой боковой цепи остатков лизина VLP или, по меньшей мере, одной субъединицы VLP, и другую функциональную группу, которая может взаимодействовать с предпочтительным вторым сайтом связывания, т.е. остатком цистеина, слитым с аналогом пептида Melan-A согласно изобретению и необязательно также сделанным доступным для реакции посредством восстановления. Первая стадия способа, обычно называемая дериватизацией, представляет собой взаимодействие VLP с перекрестно сшивающим агентом. Продуктом данной реакции является активированная VLP, также называемая активированным носителем. На второй стадии непрореагировавший перекрестно сшивающий агент удаляют, используя обычные способы, такие как гель-фильтрация или диализ. На третьей стадии осуществляют взаимодействие аналога пептида Melan-A согласно изобретению с активированной VLP, и данную стадию обычно называют стадией связывания. Непрореагировавший аналог пептида Melan-A необязательно можно удалить на четвертой стадии, например, диализом. В данной области известно несколько гетеробифункциональных перекрестно сшивающих агентов. Указанные агенты включают предпочтительные перекрестно сшивающие агенты SMPH (Pierce), сульфо-MBS, сульфо-EMCS, сульфо-GMBS, сульфо-SIAB, сульфо-SMPB, сульфо-SMCC, SVSB, SIA и другие перекрестно сшивающие агенты, доступные, например, из Pierce Chemical Company (Rockford, IL, USA) и имеющие одну функциональную группу, реакционноспособную по отношению к аминогруппам, и одну функциональную группу, реакционноспособную по отношению к остаткам цистеина. Все указанные выше перекрестно сшивающие агенты приводят к образованию тиоэфирной связи. Другой класс перекрестно сшивающих агентов, подходящих для практики изобретения, характеризуется введением дисульфидной связи между аналогом пептида Melan-A и VLP при связывании. Предпочтительные перекрестно сшивающие агенты, относящиеся к данному классу, включают, например, SPDP и сульфо-LC-SPDP (Pierce). На степень дериватизации VLP перекрестно сшивающим агентом можно влиять различными экспериментальными условиями, такими как концентрация каждого из партнеров реакции, избыток одного реагента по сравнению с другим, pH, температура и ионная сила. Степень связывания, т.е. количество антигенов или антигенных детерминант на субъединицу VLP, можно корректировать различными экспериментальными условиями, описанными выше, чтобы обеспечить соответствие требованиям вакцины.
Особенно предпочтительным способом связывания антигенов или антигенных детерминант с VLP является связывание остатка лизина на поверхности VLP с остатком цистеина на аналоге пептида Melan-A согласно изобретению. В некоторых вариантах может требоваться слияние, сочетание, присоединение или связывание аминокислотного линкера, содержащего остаток цистеина, в качестве второго сайта связывания или в качестве его части, с аналогом пептида Melan-A согласно изобретению для связывания с VLP. Такие конструкции, содержащие указанный аминокислотный линкер, также могут быть получены простым синтезом пептидов, известным в данной области.
Таким образом, в следующем предпочтительном варианте настоящего изобретения антиген или антигенная детерминанта, кроме того, содержит аминокислотный линкер, при этом предпочтительно указанный аминокислотный линкер содержит, или альтернативно состоит из второго сайта связывания.
В общем гибкие аминокислотные линкеры являются предпочтительными. Примеры аминокислотного линкера выбраны из группы, состоящей из (a) CGG; (b) N-концевого линкера гамма 1; (c) N-концевого линкера гамма 3; (d) шарнирных областей Ig; (e) N-концевых глициновых линкеров; (f) (G)kC(G)n при n=0-12 и k=0-5; (g) N-концевых глицин-сериновых линкеров; (h) (G)kC(G)m(S)l(GGGGS)n при n=0-3, k=0-5, m=0-10, l=0-2 (SEQ ID NO: 62); (i) GGC; (k) GGC-NH2; (l) C-концевого линкера гамма 1; (m) C-концевого линкера гамма 3; (n) C-концевых глициновых линкеров; (o) (G)nC(G)k при n=0-12 и k=0-5; (p) C-концевых глицин-сериновых линкеров; (q) (G)m(S)l(GGGGS)n(G)oC(G)k при n=0-3, k=0-5, m=0-10, l=0-2 и o=0-8 (SEQ ID NO: 63).
Следующими примерами аминокислотных линкеров являются шарнирная область иммуноглобулинов, глицин-сериновые линкеры (GGGGS)n (SEQ ID NO: 64) и глициновые линкеры (G)n, которые все, кроме того, содержат остаток цистеина в качестве второго сайта связывания и необязательно дополнительные остатки глицина. Обычно предпочтительными примерами указанных аминокислотных линкеров являются N-концевой гамма 1: CGDKTHTSPP (SEQ ID NO: 65); C-концевой гамма 1: DKTHTSPPCG (SEQ ID NO: 66); N-концевой гамма 3: CGGPKPSTPPGSSGGAP (SEQ ID NO: 67); C-концевой гамма 3: PKPSTPPGSSGGAPGGCG (SEQ ID NO: 68); N-концевой глициновый линкер: GCGGGG (SEQ ID NO: 69); C-концевой глициновый линкер: GGGGCG (SEQ ID NO: 70); C-концевой глицин-лизиновый линкер: GGKKGC (SEQ ID NO: 71); N-концевой глицин-лизиновый линкер: CGKKGG (SEQ ID NO: 72).
Другими аминокислотными линкерами, особенно подходящими для практического осуществления изобретения в том случае, когда гидрофобный антиген или антигенная детерминанта связаны с VLP, являются CGKKGG (SEQ ID NO: 73) или CGDEGG (SEQ ID NO: 74) для N-концевых линкеров, или GGKKGC (SEQ ID NO: 75) и GGEDGC (SEQ ID NO: 76) для C-концевых линкеров. В случае C-концевых линкеров концевой цистеин необязательно амидирован на C-конце.
Следующими линкерами, применимыми для данного изобретения, являются аминокислотные последовательности, которые обеспечивают возможность высвобождения антигенного пептида, т.е. аналога пептида Melan-A меланомы человека, из VLP. Примеры указанных линкеров описаны в Toes RE et al. J. Exp. Med. 2001 Jul. 2; 194(1): 1-12. Кроме того, можно использовать алгоритм предсказания расщеплений протеосомами PAProC (Nussbaum AK, et. al. Immunogenetics. 2001 Mar; 53 (2): 87-94) для предсказания указанной выше аминокислотной последовательности, которая обеспечивает возможность высвобождения антигенного пептида, т.е. аналога пептида Melan-A меланомы человека, из VLP.
В предпочтительных вариантах настоящего изобретения предпочтительными в качестве аминокислотных линкеров являются линкеры GGCG (SEQ ID NO: 77), GGC или GGC-NH2 («NH2» означает амидирование) на C-конце пептида или CGG на его N-конце. В общем, остатки глицина будут встроены между основной массой аминокислот и цистеином, используемым в качестве второго сайта связывания, чтобы избежать возможных стерических помех со стороны аминокислоты основной массы в реакции связывания. В наиболее предпочтительном варианте изобретения аминокислотный линкер GGC-NH2 сливают с C-концом аналога пептида Melan-A согласно изобретению.
Остаток цистеина, добавляемый к аналогу пептида Melan-A согласно изобретению, должен быть в своем восстановленном состоянии, чтобы реагировать с гетеробифункциональным перекрестно сшивающим агентом на активированной VLP, то есть должен быть доступным свободным цистеином или остатком цистеина со свободной сульфгидрильной группой. В том случае когда остаток цистеина для функционирования в качестве сайта связывания находится в окисленной форме, например, если он образует дисульфидный мостик, требуется восстановление указанного дисульфидного мостика, например, с помощью DTT, TCEP или β-меркаптоэтанола. Низкие концентрации восстанавливающего агента совместимы со связыванием, как описано в WO 02/05690, более высокие концентрации ингибируют реакцию связывания, как может быть известно специалисту в данной области, и в этом случае восстановитель должен быть удален или его концентрация должна быть снижена перед связыванием, например, посредством диализа, гель-фильтрации или обращенно-фазовой ВЭЖХ.
Связывание аналога пептида Melan-A согласно изобретению с VLP с использованием гетеробифункционального перекрестно сшивающего агента согласно описанным выше предпочтительным способам позволяет осуществлять связывание аналога пептида Melan-A согласно изобретению с VLP ориентированным образом. Другие способы связывания аналога пептида Melan-A согласно изобретению с VLP включают способы, при которых аналог пептида Melan-A согласно изобретению перекрестно сшивают с VLP, используя карбодиимид, EDC и NHS. В следующих способах аналог пептида Melan-A согласно изобретению связывают с VLP, используя гомобифункциональный перекрестно сшивающий агент, такой как глутаральдегид, DSG, BM[PEO]4, BS3, (Pierce Chemical Company, Rockford, IL, USA) или другие известные гомобифункциональные перекрестно сшивающие агенты с функциональными группами, взаимодействующими с аминогруппами или карбоксильными группами VLP.
Другие способы связывания VLP с аналогом пептида Melan-A согласно изобретению включают способы, в которых VLP биотинилируют и аналог пептида Melan-A согласно изобретению экспрессируют в виде белка, слитого со стрептавидином, или способы, при которых биотинилируют как аналог пептида Melan-A согласно изобретению, так и VLP, например, как описано в WO 00/23955. В данном случае аналог пептида Melan-A согласно изобретению сначала можно связать со стрептавидином или авидином, корректируя отношение аналога пептида Melan-A согласно изобретению к стрептавидину так, чтобы свободные сайты связывания еще были доступными для связывания VLP, которую добавляют на следующей стадии. Альтернативно все компоненты можно смешать в реакции «в одном сосуде». Другие пары лиганд-рецептор, где имеется растворимая форма рецептора и лиганда и которые способны к перекрестному сшиванию с VLP или аналогом пептида Melan-A согласно изобретению, можно использовать в качестве связывающих агентов для связывания аналога пептида Melan-A согласно изобретению с VLP. Альтернативно либо лиганд, либо рецептор могут быть слиты аналогом пептида Melan-A согласно изобретению и, таким образом, опосредовать связывание с VLP, химически связанной или слитой либо с рецептором, либо с лигандом соответственно. Слияние также можно осуществлять посредством инсерции или замены.
Как уже говорилось, в предпочтительном варианте настоящего изобретения VLP является VLP РНК-фага и в более предпочтительном варианте VLP является VLP белка оболочки РНК-фага Qβ.
Одна или несколько молекул антигена, т.е. один или несколько антигенов или антигенных детерминант могут быть связаны с одной субъединицей капсида или VLP белков оболочки РНК-фага, предпочтительно через экспонированные остатки лизина VLP РНК-фагов, если это стерически допустимо. Таким образом, особым признаком VLP белков оболочки РНК-фагов, и в частности VLP белка оболочки Qβ, является возможность связывать несколько антигенов на субъединицу. Это обеспечивает возможность образования плотной антигенной матрицы.
В предпочтительном варианте изобретения связывание и соответственно присоединение, по меньшей мере, одного аналога пептида Melan-A согласно изобретению с вирусоподобной частицей осуществляют посредством взаимодействия и соответственно ассоциации между, по меньшей мере, одним первым сайтом связывания вирусоподобной частицы и, по меньшей мере, одним вторым сайтом связывания аналога пептида Melan-A согласно изобретению.
VLP или капсиды из белка оболочки Qβ экспонируют определенное количество остатков лизина на своей поверхности с определенной топологией, при этом три остатка лизина направлены внутрь капсида и взаимодействуют с РНК, а четыре других остатка лизина экспонированы на наружную сторону капсида. Указанные определенные свойства предпочтительны для связывания антигенов с наружной стороной частицы, а не с внутренней стороны частицы, где остатки лизина взаимодействуют с РНК. VLP из других белков оболочки РНК-фагов также имеют определенное количество остатков лизина на своей поверхности и определенную топологию указанных остатков лизина.
В следующих предпочтительных вариантах настоящего изобретения первым сайтом связывания является остаток лизина и/или второй сайт связывания содержит сульфгидрильную группу или остаток цистеина. В очень предпочтительном варианте настоящего изобретения первым сайтом связывания является остаток лизина, а вторым сайтом связывания является остаток цистеина.
В очень предпочтительных вариантах изобретения аналог пептида Melan-A согласно изобретению связан через остаток цистеина с остатками лизина VLP белка оболочки РНК-фага и, в частности, VLP белка оболочки Qβ.
Другим преимуществом VLP, полученных из РНК-фагов, является высокий выход их экспрессии в бактериях, который позволяет получать большие количества материала при приемлемой стоимости.
Как указано, конъюгаты и матрицы соответственно согласно изобретению отличаются от конъюгатов предшествующего уровня техники своей высокоорганизованной структурой, размерами и повторяемостью антигена на поверхности матрицы. Кроме того, применение VLP в качестве носителей позволяет образовывать прочные антигенные матрицы и конъюгаты соответственно с изменяемой плотностью антигенов. В частности применение VLP РНК-фагов и, таким образом, в частности, применение VLP белка оболочки РНК-фага Qβ позволяет достигать очень высокой плотности эпитопа. В частности достигали плотности более 1,5 эпитопа на субъединицу при связывании пептида с VLP белка оболочки Qβ (например, пептида Aβ 1-6 человека, как описано в WO 2004/016282). Получение композиций VLP белков оболочки РНК-фагов с высокой плотностью эпитопов можно осуществлять с использованием инструкций, приведенных в данной заявке. В предпочтительном варианте изобретении, когда с VLP белка оболочки Qβ связывают аналог пептида Melan-A согласно изобретению, предпочтительно среднее количество аналога пептида Melan-A согласно изобретению на субъединицу, составляющее 0,5, 0,6, 0,7, 0,8, 0,9, 1,0, 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 2,6, 2,7, 2,8, 2,9 или более.
На аналоге пептида Melan-A согласно изобретению может присутствовать либо природный, либо неприродный второй сайт связывания, который определен в данном описании. В случае отсутствия подходящего второго сайта связывания природного происхождения на аналоге пептида Melan-A согласно изобретению необходимо сконструировать неприродный второй сайт связывания на антигене.
Как описано выше, четыре остатка лизина экспонированы на поверхности VLP белка оболочки Qβ. Обычно такие остатки дериватизируют в ходе реакции с перекрестно сшивающей молекулой. В том случае когда не все экспонированные остатки лизина могут быть связаны с антигеном, остатки лизина, которые прореагировали с перекрестно сшивающим агентом, остаются с перекрестно сшивающей молекулой, связанной с ε-аминогруппой после стадии дериватизации. Это приводит к исчезновению одного или нескольких положительных зарядов, которое может быть нежелательным для растворимости и стабильности VLP. Заменяя некоторые остатки лизина аргининами, как в заявленных мутантах белка оболочки Qβ, описанных ниже, авторы предотвратили чрезмерное исчезновение положительных зарядов, так как остатки аргинина не взаимодействуют с перекрестно сшивающим агентом. Кроме того, замена остатков лизина аргининами может приводить к более определенной антигенной матрице, так как меньше сайтов доступно для реакции с антигеном.
Таким образом, экспонированные остатки лизина заменяли аргининами в следующих мутантах белка оболочки Qβ и мутантных VLP Qβ, описанных в данной заявке: Qβ-240 (Lys13-Arg; SEQ ID NO: 20), Qβ-250 (Lys 2-Arg, Lys13-Arg; SEQ ID NO: 22) и Qβ-259 (Lys 2-Arg, Lys16-Arg; SEQ ID NO: 24). Конструкции клонировали, белки экспрессировали, VLP очищали и использовали для связывания с аналогами пептида Melan-A согласно изобретению. Также конструировали Qβ-251 (SEQ ID NO: 23), и руководство по поводу того, как экспрессировать, очищать и связывать VLP белка оболочки Qβ-251, можно найти на протяжении данной заявки.
В следующем варианте авторы предложили мутантный белок оболочки Qβ с одним дополнительным остатком лизина, подходящий для получения матрицы антигенов с еще более высокой плотностью. Данный мутантный белок оболочки Qβ, Qβ-243 (Asn 10-Lys; SEQ ID NO: 21) клонировали, белок экспрессировали и капсид или VLP выделяли и очищали, показав, что введение дополнительного остатка лизина совместимо с самосборкой субъединиц в капсид или VLP. Таким образом, матрицы и соответственно конъюгаты аналога пептида Melan-A можно получить с использованием VLP мутантов белка оболочки Qβ. Особенно предпочтительным способом связывания антигенов с VLP и, в частности, с VLP белков оболочки РНК-фага является связывание остатка лизина, присутствующего на поверхности VLP белков оболочки РНК-фага, с остатком цистеина, добавляемого к антигену. Для того чтобы остаток цистеина был эффективным в качестве второго сайта связывания, сульфгидрильная группа должна быть доступной для связывания. Таким образом, остаток цистеина должен быть в своем восстановленном состоянии, то есть должен быть доступным свободный цистеин или остаток цистеина со свободной сульфгидрильной группой. В том случае когда остаток цистеина для функционирования в качестве второго сайта связывания находится в окисленной форме, например, если он образует дисульфидный мостик, требуется восстановление указанного дисульфидного мостика, например, с помощью DTT, TCEP или β-меркаптоэтанола. Концентрация восстановителя и молярный избыток восстановителя по сравнению с антигеном должен уточняться для каждого антигена. При необходимости тестируют пределы титрования, начиная с таких низких концентраций восстановителя, как 10 мкМ или ниже, до 10-20 мМ или выше, и оценивают связывание антигена с носителем. Несмотря на то что низкие концентрации восстановителя совместимы с реакцией связывания, как описано в WO 02/05905, более высокие концентрации ингибируют реакцию связывания, как будет понятно специалисту в данной области, и в этом случае восстановитель необходимо удалить или снизить его концентрацию, например, диализом, гель-фильтрацией или обращенно-фазовой ВЭЖХ. Преимущественно pH буфера для диализа или уравновешивающего буфера ниже 7, предпочтительно 6. Совместимость буфера с низким значением pH с активностью и стабильностью антигена необходимо проверять.
Плотность эпитопа на VLP белков оболочки РНК-фага можно модулировать, выбирая перекрестно сшивающий агент и другие условия реакции. Например, перекрестно сшивающие агенты сульфо-GMBS и SMPH обычно обеспечивают достижение высокой плотности эпитопа. На дериватизацию положительно влияет высокая концентрация реагирующих веществ, и манипулирование условиями реакции можно использовать для того, чтобы контролировать количество антигенов, связанных с VLP белков оболочки РНК-фага и, в частности, с VLP белка оболочки Qβ.
Перед конструированием неприродного второго сайта связывания необходимо выбрать положение, в котором он должен быть слит, встроен или, в общем, сконструирован. Выбор положения второго сайта связывания в качестве примера может быть основан на кристаллической структуре антигена. Такая кристаллическая структура антигена может давать информацию о доступности C- или N-концов молекулы (определенной, например, исходя из их доступности для растворителя) или об экспонировании по отношению к растворителю остатков, подходящих для использования в качестве вторых сайтов связывания, таких как остатки цистеина. Экспонированные дисульфидные мостики, как в случае Fab-фрагментов, также могут быть источником второго сайта связывания, так как их, как правило, можно превратить в отдельные остатки цистеина посредством умеренного восстановления 2-меркаптоэтиламином, TCEP, β-меркаптоэтанолом или DTT. Будут выбираться умеренные условия восстановления, не влияющие на иммуногенность антигена. В общем, в том случае, когда целью иммунизации аутоантигеном является ингибирование взаимодействия данного аутоантигена с его природными лигандами, второй сайт связывания будет добавлен таким образом, чтобы он давал возможность образовывать антитела против сайта взаимодействия с природными лигандами. Таким образом, положение второго сайта связывания будет выбрано так, чтобы избежать стерических помех со стороны второго сайта связывания или любого содержащего его аминокислотного линкера. В следующих вариантах требуется гуморальный ответ, направленный на сайт, отличный от сайта взаимодействия аутоантигена с его природным лигандом. В таких вариантах второй сайт связывания может быть выбран так, чтобы он предотвращал образование антител против сайта взаимодействия аутоантигена с его природными лигандами.
Другие критерии для выбора положения второго сайта связывания включают состояние олигомеризации антигена, сайт олигомеризации, присутствие кофактора и наличие экспериментального доказательства обнаружения сайтов в структуре антигена и последовательности, в которой модификация антигена совместима с функцией аутоантигена или с образованием антител, распознающих аутоантиген.
В наиболее предпочтительных вариантах аналог пептида Melan-A согласно изобретению содержит единственный второй сайт связывания или единственный реакционноспособный сайт связывания, способный к ассоциации с первыми сайтами связывания на коровой частице и VLP или субъединицах VLP соответственно. Это обеспечивает определенное и равномерное связывание и соответственно ассоциацию, по меньшей мере, одного, но обычно нескольких, предпочтительно более 10, 20, 40, 80, 120, 150, 180, 210, 240, 270, 300, 360, 400, 450 аналогов пептида Melan-A согласно изобретению с коровой частицей и VLP соответственно. Таким образом, обеспечение одного второго сайта связывания или одного реакционноспособного сайта связывания на антигене обеспечивает один тип равномерного связывания и ассоциации соответственно, приводящий к очень высокоупорядоченной и повторяющейся матрице. Например, если связывание и ассоциацию соответственно осуществляют посредством взаимодействия лизина (в качестве первого сайта связывания) и цистеина (в качестве второго сайта связывания), то согласно данному предпочтительному варианту изобретения гарантировано, что только один остаток цистеина на антиген независимо от того, присутствует ли на антигене данный остаток цистеина природного или неприродного происхождения, способен связываться и ассоциировать соответственно с VLP и первым сайтом связывания коровой частицы соответственно.
В некоторых вариантах конструирование второго сайта связывания на аналоге пептида Melan-A согласно изобретению требует слияния аминокислотного линкера, содержащего аминокислоту, подходящую в качестве второго сайта связывания согласно описаниям данного изобретения. Таким образом, в предпочтительном варианте настоящего изобретения аминокислотный линкер связан с аналогом пептида Melan-A согласно изобретению посредством, по меньшей мере, одной ковалентной связи. Предпочтительно аминокислотный линкер содержит, или альтернативно состоит из второго сайта связывания. В следующем предпочтительном варианте аминокислотный линкер содержит сульфгидрильную группу или остаток цистеина. В другом предпочтительном варианте аминокислотным линкером является цистеин. Некоторые критерии выбора аминокислотного линкера, а также дополнительные предпочтительные варианты аминокислотного линкера согласно изобретению уже указаны выше.
В другом конкретном варианте изобретения в качестве сайта связывания выбирают остаток лизина или цистеина, который слит в рамке с HBcAg. В предпочтительном варианте антиген сливают с C-концом HBcAg через линкер из трех лейцинов.
Когда антиген или антигенная детерминанта связаны с VLP посредством остатка лизина, может быть полезно либо заменить, либо делетировать один или несколько имеющихся природных остатков лизина, а также других остатков лизина, присутствующих в вариантах HBcAg.
Во многих случаях, когда имеющиеся природные остатки лизина будут устранять, другой лизин будет введен в HBcAg в качестве сайта связывания для антигена или антигенной детерминанты. Способы инсерции такого остатка лизина известны в данной области. Остатки лизина также могут быть добавлены без удаления существующих остатков лизина.
Показано, что C-конец HBcAg управляет локализацией данного белка в ядре. (Eckhardt et al., J. Virol. 65: 575 582 (1991)). Кроме того, также предполагается, что указанная область белка придает HBcAg способность связывать нуклеиновые кислоты.
Как указано, HBcAg, подходящие для применения в практике настоящего изобретения, также включают укороченные на N-конце мутанты. Подходящие укороченные мутанты включают модифицированные HBcAg, в которых 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот были удалены из N-конца. Однако варианты вирусоподобных частиц, содержащих внутренние делеции в последовательности субъединицы, составляющей вирусоподобную частицу, также подходят согласно настоящему изобретению при условии их совместимости с упорядоченной или образующей частицу структурой вирусоподобной частицы. Например, подходящими являются внутренние делеции в последовательности HBcAg (Preikschat, P., et al., J. Gen. Virol. 80: 1777-1788 (1999)).
Следующие HBcAg, подходящие для применения в практике настоящего изобретения, включают укороченные на N- и C-концах мутанты. Подходящие укороченные мутанты включают HBcAg, в которых 1, 2, 5, 7, 9, 10, 12, 14, 15 и 17 аминокислот были удалены из N-конца и 1, 5, 10, 15, 20, 25, 30, 34, 35, 36, 37, 38, 39, 40, 41, 42 или 48 аминокислот были удалены из C-конца.
Композиции вакцин согласно изобретению могут содержать смеси различных HBcAg. Таким образом, указанные композиции вакцин могут состоять из HBcAg, которые отличаются по аминокислотной последовательности. Например, могут быть получены композиции вакцин, содержащие HBcAg «дикого типа» и модифицированный HBcAg, в котором один или несколько аминокислотных остатков были изменены (например, делетированы, встроены или заменены). Однако в большинстве применений будет использован только один тип HBcAg.
В предпочтительном варианте вирусоподобная частица содержит по меньшей мере один первый сайт связывания, и антиген или антигенная детерминанта содержит по меньшей мере один второй сайт связывания. Предпочтительно первый сайт связывания содержит или предпочтительно состоит из аминогруппы или остатка лизина. Второй сайт связывания предпочтительно выбран из группы, состоящей из (a) сайта связывания, не встречающегося в природе в случае указанного антигена или антигенной детерминанты; и (b) сайта связывания, встречающегося в природе в случае указанного антигена или антигенной детерминанты. Еще более предпочтительно второй сайт связывания содержит или предпочтительно состоит из сульфгидрильной группы или остатка цистеина. В предпочтительном варианте связывание антигена или антигенной детерминанты с вирусоподобной частицей осуществляют посредством ассоциации между первым сайтом связывания и вторым сайтом связывания, при этом предпочтительно ассоциация осуществляется посредством по меньшей мере одной непептидной связи и при этом предпочтительно антиген или антигенная детерминанта и вирусоподобная частица взаимодействуют посредством указанной ассоциации с образованием упорядоченной и повторяющейся матрицы антигена. В одном варианте первым сайтом связывания является остаток лизина, а вторым сайтом связывания является остаток цистеина. В другом варианте первым сайтом связывания является аминогруппа, а вторым сайтом связывания является сульфгидрильная группа.
В конкретном варианте изобретения антиген, и в данном случае, в частности, аналог пептида Melan-A меланомы содержит один или несколько эпитопов для цитотоксических T-клеток, Th-клеточных эпитопов или комбинацию двух эпитопов. Таким образом, в одном варианте антиген или антигенная детерминанта содержит один, два или более эпитопов для цитотоксических T-клеток. В другом варианте антиген или антигенная детерминанта содержит один, два или более Th-клеточных эпитопов. В еще одном варианте антиген или антигенная детерминанта содержит один, два или более эпитопов для цитотоксических T-клеток и один, два или более Th-клеточных эпитопов.
Природные эпитопы Melan-A/Mart-1 и, например, эпитоп 26-35 Melan-A/Mart-1 связывается с низкой аффинностью только с HLA-2 человека. Таким образом, презентация in vivo природных эпитопов и пептидов Melan-A соответственно при вакцинации может быть ограничивающим фактором. Это особенно важно, если для вакцинации используют эпитопы и пептиды Melan-A, соответственно связанные, соединенные или слитые с VLP, так как при таких условиях пептиды Melan-A загружают молекулы HLA в результате перекрестной презентации. Однако процесс перекрестной презентации не столь эффективен, как классические пути презентации антигена, и еще более важна аффинность пептида Melan-A по отношению к HLA. Таким образом, для вакцинаций, основанных на VLP, очень предпочтительно использование аналогов пептида Melan-A, которые связываются с HLA с относительно высокой аффинностью. Подобным образом, может быть полезным применение аналогов пептида Melan-A, которые с высокой аффинностью распознаются природным репертуаром T-клеток хозяина. В качестве общего правила предпочтительны эпитопы и аналоги пептида Melan-A соответственно, которые содержат заякоривающие остатки в правильных положениях, позволяющие эффективно связываться с молекулами MHC.
Таким образом, следующим аспектом настоящего изобретения и очень предпочтительным вариантом настоящего изобретение является композиция для усиления иммунного ответа у животного, содержащая (a) вирусоподобную частицу; и (b) иммуностимулирующее вещество, при этом указанное иммуностимулирующее вещество связано с указанной вирусоподобной частицей, и при этом указанная композиция, кроме того, содержит по меньшей мере один антиген или антигенную детерминанту, где указанный антиген или антигенная детерминанта связаны с указанной вирусоподобной частицей, и где указанный по меньшей мере один антиген или антигенная детерминанта содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека, и где указанный аналог пептида Melan-A меланомы человека связан с указанной вирусоподобной частицей.
В предпочтительном варианте настоящего изобретения аналог пептида Melan-A способен обеспечивать эффективное связывание с молекулами MHC. Таким образом, применение аналога пептида Melan-A в частности позволяет введение таких заякоривающих остатков, приводящее к повышенному связыванию с молекулами MHC. Введение таких заякоривающих остатков, приводящих к повышенному связыванию с молекулами MHC, в частности, полезно, если природный и соответственно нормальный пептид Melan-A не содержит таких заякоривающих остатков или не содержит такие заякоривающие остатки, которые хуже, чем заново введенный заякоривающий остаток(ки), заменяющие природный и соответственно нормальный заякоривающий остаток.
Модификация нормального пептида Melan-A человека, приводящая в результате к аналогу пептида Melan-A, и тем самым предпочтительно введение указанных заякоривающих остатков осуществляют либо (i) индуцированной мутацией (например, химической индукцией, облучением или другими способами, известными специалисту в данной области) и последующим отбором модифицированных пептидов с повышенным связыванием с MHC или (ii) отбором модифицированных пептидов с повышенным связыванием с MHC, возникающих в результате природных мутаций, возникающих на любом уровне синтеза белка, включая, но не ограничивая указанным, мутации, возникающие на уровне ДНК, транскрипционном уровне, на уровне РНК или трансляционном уровне экспрессии белка, или (iii) посредством системных или случайных аминокислотных обменов, делеций, замен или инсерций с использованием классического синтеза пептидов, известного специалисту в данной области. Идентификацию таких заякоривающих остатков обычно и предпочтительно осуществляют с использованием базы данных SYFPEITHI, как описано Rammensee et al. в Immunogenetics 50: 213-219 (1999). База данных SYFPEITHI позволяет рассчитывать эффективность связывания HLA для любого выбранного пептида, и с использованием указанной программы можно оптимизировать пептиды в отношении эффективности связывания HLA. Альтернативно идентификацию предпочтительных аналогов пептида можно осуществить в результате анализов связывания MHC-пептид, включающих без ограничения анализ на целых клетках активации T-клеток или распознавания или повышающей регуляции MHC в мутантных линиях клеток, анализ связывания MHC-тетрамер-пептид, конкурентные анализы связывания с мечеными пептидами, анализ резонанса поверхностного плазмона, все анализы известны специалисту в данной области.
В следующем предпочтительном варианте настоящего изобретения аналог пептида Melan-A характеризуется двумя, более предпочтительно одной заменой аминокислоты относительно соответствующего нормального пептида Melan-A.
В другом предпочтительном варианте настоящего изобретения аналог пептида Melan-A защищен от деградации, опосредованной протеазами или пептидазами. Применение аналогов пептида Melan-A. которые защищены от деградации протеазами или пептидазами, приводит к повышенной стабильности пептида in vivo после применения пептида у субъекта и/или к повышенной стабильности пептида при хранении в присутствии протеаз или пептидаз. Следствием такой повышенной стабильности является более эффективная и длительная презентация аналога пептида Melan-A меланомы человека на MHC и, следовательно, повышенная стимуляция специфичного T-клеточного ответа.
Предпочтительно аналог пептида Melan-A человека защищен заменой выбранных аминокислотных остатков природного пептида Melan-A человека неприродными производными аминокислот, которые приведены в качестве примеров в Blanchet et al, J. Immunol. 167: 5852-5861 (2001) и ссылках, цитированных в работе. Это позволяет преодолевать ограничение, налагаемое тем фактом, что химически модифицированные пептиды Melan-A и аналоги пептида Melan-A соответственно могут не распознаваться T-клетками также хорошо по сравнению с природным и соответственно нормальным пептидом Melan-A.
В другом предпочтительном варианте антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A/MART-1 меланомы человека, имеющего аминокислотную последовательность которая без ограничения выбрана из группы, состоящей из (a) LAGIGILTV (SEQ ID NO: 84); (b) MAGIGILTV (SEQ ID NO: 85), (c) EAMGIGILTV (SEQ ID NO: 86), (d) ELAGIGILTV (SEQ ID NO: 50), (e) EMAGIGILTV (SEQ ID NO: 87), (f) YAAGIGILTV (SEQ ID NO: 88), (g) FAAGIGILTV (SEQ ID NO: 89), (h) GHGHSYTTAE ELAGIGILTV (SEQ ID NO: 51), (i) SYTTAEELAGIGILTVILGVL (SEQ ID NO: 52), и (j) ELAGIGILTVILGVL (SEQ ID NO: 53). В очень предпочтительном варианте антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A/MART-1 меланомы человека, имеющего аминокислотную последовательность, которая без ограничения выбрана из группы, состоящей из (a) LAGIGILTV (SEQ ID NO: 84); (b) MAGIGILTV (SEQ ID NO: 85), (c) EAMGIGILTV (SEQ ID NO: 86), (d) ELAGIGILTV (SEQ ID NO: 50), (e) EMAGIGILTV (SEQ ID NO: 87), (f) YAAGIGILTV (SEQ ID NO: 88), и (g) FAAGIGILTV (SEQ ID NO: 89). Указанные аналоги пептидов, а также их синтез описаны Valmori at al., J. Immunol. 160: 1750-1758 (1998). В случае указанных аналогов пептида наблюдается сравнительно повышенное распознавание и лизис клеток-мишеней пятью разными клонами цитотоксических T-клеток, индуцированных против природного пептида меланомы.
В очень предпочтительном варианте настоящего изобретения аналог пептида Melan-A/MART-1 меланомы человека содержит, альтернативно в основном состоит, или альтернативно состоит из последовательности ELAGIGILTV (SEQ ID NO: 50). Как указано в примерах, данный предпочтительный вариант индуцирует экспансию функциональных специфичных по отношению Melan-A T-клеток CD8+ у трансгенных по HLA-A2 мышей и представляет собой хороший компромисс между связыванием HLA и распознаванием TCR (сравни Valmori et al., J. Immunol., 160: 1750-1758 (1998)).
В следующем предпочтительном варианте настоящего изобретения аналог пептида Melan-A/MART-1 меланомы человека со вторым сайтом связывания имеет аминокислотную последовательность, выбранную без ограничения из группы, состоящей из (a) CGHGHSYTTAEEAAGIGILTV (SEQ ID NO: 54), и обычно сокращенно называемой в данном описании 16-35, (b) CGHGHSYTTAEELAGIGILTV (SEQ ID NO: 55), и обычно сокращенно называемой в данном описании Melan-A 16-35 A/L, (c) CGGEAAGIGILTV (SEQ ID NO: 56), и обычно сокращенно называемой в данном описании Melan-A 26-35, (d) CGGELAGIGILTV (SEQ ID NO: 57), и обычно сокращенно называемой в данном описании Melan 26-35 A/L, (e) CSYTTAEELAGIGILTVILGVL (SEQ ID NO: 58), и обычно сокращенно называемой в данном описании Melan-A 20-40 A/L, (f) CGGELAGIGILTVILGVL (SEQ ID NO: 59), и обычно сокращенно называемой в данном описании Melan 26-40 A/L, (g) ELAGIGILTVGGC (SEQ ID NO: 60), и обычно сокращенно называемой в данном описании Melan 26-35-C A/L, (h) CSPKSLELAGIGILTV (SEQ ID NO: 92), и обычно сокращенно называемой в данном описании CSPKSL-Melan 26-35 A/L, и (i) ELAGIGILTVILGVLGGC (SEQ ID NO: 93), и обычно сокращенно называемой в данном описании Melan 26-40-C A/L. В очень предпочтительном варианте настоящего изобретения аналог пептида Melan-A/MART-1 меланомы человека со вторым сайтом связывания имеет аминокислотную последовательность, выбранную из (a) CGHGHSYTTAEELAGIGILTV (SEQ ID NO: 55), и обычно сокращенно называемой в данном описании Melan-A 16-35 A/L, (b) CGGELAGIGILTV (SEQ ID NO: 57), и обычно сокращенно называемой в данном описании Melan 26-35 A/L, (c) CSYTTAEELAGIGILTVILGVL (SEQ ID NO: 58), и обычно сокращенно называемой в данном описании Melan-A 20-40 A/L, (d) CGGELAGIGILTVILGVL (SEQ ID NO: 59), и обычно сокращенно называемой в данном описании Melan 26-40 A/L, (e) ELAGIGILTVGGC (SEQ ID NO: 60), и обычно сокращенно называемой в данном описании Melan 26-35-C A/L.
В другом очень предпочтительном варианте настоящего изобретения аналог пептида Melan-A/MART-1 меланомы человека со вторым сайтом связывания имеет аминокислотную последовательность CGHGHSYTTAEELAGIGILTV (SEQ ID NO: 55) (Melan-A 16-35 A/L). Как указано в примере 21, предлагаемая в изобретении композиция вакцины, содержащая данный очень предпочтительный вариант, т.е. вакцина QβMelan-A 16-35 A/L, процессируется дендритными клетками.
В другом варианте настоящего изобретения аналогом пептида Melan-A согласно изобретению, который связан, слит или иным образом соединен с вирусоподобной частицей, является T-клеточный эпитоп либо эпитоп для цитотоксических либо Th-клеток. В следующем предпочтительном варианте антиген представляет собой комбинацию по меньшей мере двух либо сходных, либо разных, предпочтительно разных, эпитопов, при этом по меньшей мере два эпитопа связаны непосредственно или с помощью линкерной последовательности. Указанные эпитопы предпочтительно выбраны из группы, состоящей из эпитопов для цитотоксических и Th-клеток, описанных для меланом (gp100, тирозиназа, семейство MAGE или NY-ESO-1).
Таким образом, в следующем предпочтительном варианте настоящего изобретения указанный антиген содержит, или альтернативно, кроме того, содержит эпитоп для цитотоксических T-клеток, эпитоп для Th-клеток или комбинацию по меньшей мере двух указанных эпитопов, при этом указанные по меньшей мере два эпитопа связаны непосредственно или с помощью линкерной последовательности, и при этом предпочтительно указанный эпитоп для цитотоксических T-клеток является вирусным или опухолевым эпитопом для цитотоксических T-клеток.
Предпочтительными эпитопами для цитотоксических T-клеток являются эпитопы Melan-A: (16-36 A/L)(25-36 A/L); эпитопы тирозиназы: (1-9) и (368-376) (Panelli, M. C. et. al., J. Immunol., 2000, 164, 495-504); эпитопы Gp100: (154-162), (209-217(T210M)), (280-288) и (280-288(A288V)) и (457-466) (Nielsen, M.B. et. al., 2000. J Immunol., 164 (4)., 2287-96 and Linette, G.P. et. al. J Immunol, 2000, 164, 3402-3412 и Skipper, J.C., Int. J. Cancer, 1999, 82, 669-677 and Pass, H.A., et. al., Cancer J Sci Am., 1998, 4, 316-323); эпитопы TRP2: (180-188)(288-296)(455-463) (Sun, Y. et.al., Int. J Cancer, 2000, 87 (3), 399-404 и Parkhurst, M.R. et.al., Cancer Res., 1998, 58, 4895-8901 и Harada, M., et.al., Cancer Res., 2001, 61, 1089-1094)); эпитоп NY-ESO-1: 157-165 (Chen, J. L., et. al., J. Immunol., 2000, 165, 948-955); и эпитоп MAGE-A: (248V9) (Graff-Dubois, S., et. al., 2002, J. Immunol., 169,575-580).
Предпочтительной комбинацией эпитопов для цитотоксических T-клеток является комбинация Melan-A (26-36 A/L), тирозиназы (368-376), Gp100 (209-217(T210M) и (457-466) и NY-ESO-1 (157-165).
Предпочтительными Th-клеточными эпитопами являются эпитопы Mage-3: (281-295), (141-155) и (146-160) (Kobayashi, H., et. al., Cancer Res., 2001,61, 4773-4778); эпитопы тирозиназы: (188-208), (193-203) (Kobayashi H., et. al., Immunogenetics, 1998, 47, 398-403); эпитоп GP100: (44-59) и эпитопы NY-ESO-1: (115-132), (121-138), (139-156), (119-143) и (134-148) (Zarour, H. M., et. al., Cancer Res., 2002, 62, 213-218).
Предпочтительными комбинациями эпитопов для цитотоксических и Th-клеток являются эпитопы Melan-A (1-118 A/L) (SEQ ID NO: 94); эпитопы NY-ESO-1 115-165; или комбинация Melan-A (25-36 A/L), gp100 (209-217(T210M), Mage-3(146-160) и gp100(44-59).
Также следует понимать, что мозаичная вирусоподобная частица, например вирусоподобная частица, состоящая из субъединиц, связанных с разными антигенами и эпитопами соответственно, входит в объем настоящего изобретения. Такая композиция согласно настоящему изобретению, например, может быть получена при трансформации E. coli двумя совместимыми плазмидами, кодирующими субъединицы, составляющими вирусоподобную частицу, слитые с разными антигенами и эпитопами соответственно. В данном случае мозаичная вирусоподобная частица собирается либо непосредственно в клетке, либо после лизиса клеток. Кроме того, такая предлагаемая в изобретении композиция также может быть получена связыванием смеси различных антигенов и эпитопов соответственно с изолированной вирусоподобной частицей.
Аналог пептида Melan-A согласно настоящему изобретению, и в частности указанный эпитоп или эпитопы, могут быть синтезированы или рекомбинантно экспрессированы и связаны с вирусоподобной частицей или слиты с вирусоподобной частицей с использованием технологии рекомбинантной ДНК. Примеры способов, описывающие связывание антигенов с вирусоподобными частицами, раскрыты в WO 00/32227, в WO 01/85208 и в WO 02/056905, описания которых, таким образом, включены в виде ссылки в полном объеме.
Изобретение также относится к способу получения композиции для усиления иммунного ответа у животного, содержащей VLP иммуностимулирующие вещество, предпочтительно неметилированный CpG-содержащий олигонуклеотид, связанный с VLP, который включает в себя инкубацию VLP с иммуностимулирующим веществом и соответственно олигонуклеотидом, добавление РНКазы и очистку указанной композиции. Предпочтительно способ, кроме того, включает в себя стадию связывания антигена или антигенной детерминанты с указанной вирусоподобной частицей, при этом указанный антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека. В предпочтительном варианте антиген или антигенную детерминанту связывают с вирусоподобной частицей перед инкубацией вирусоподобной частицы с иммуностимулирующим веществом. В другом предпочтительном варианте антиген или антигенную детерминанту связывают с вирусоподобной частицей после очистки композиции. В равной мере предпочтительном варианте способ включает в себя инкубацию VLP с РНКазой, добавление иммуностимулирующего вещества и соответственно олигонуклеотида и очистку композиции. Предпочтительно способ, кроме того, включает в себя стадию связывания антигена или антигенной детерминанты с указанной вирусоподобной частицей, при этом указанный антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека. В предпочтительном варианте антиген или антигенную детерминанту связывают с вирусоподобной частицей перед инкубацией вирусоподобной частицы с РНКзой. В другом предпочтительном варианте антиген или антигенную детерминанту связывают с вирусоподобной частицей после очистки композиции. В одном варианте VLP получают в бактериальной системе экспрессии. В другом варианте РНКзой является РНКаза A.
Изобретение, кроме того, относится к способу получения композиции для усиления иммунного ответа у животного, содержащей VLP, связанные с иммуностимулирующим веществом, предпочтительно с неметилированным CpG-содержащим олигонуклеотидом, который включает в себя разборку VLP, добавление иммуностимулирующего вещества и соответственно олигонуклеотида и повторную сборку VLP. Кроме того, способ может включать в себя удаление нуклеиновых кислот разобранной VLP и/или очистку композиции после повторной сборки. Предпочтительно способ, кроме того, включает в себя стадию связывания антигена или антигенной детерминанты с вирусоподобной частицей, при этом указанный антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека. В предпочтительном варианте антиген или антигенную детерминанту связывают с вирусоподобной частицей перед разборкой вирусоподобной частицы. В другом предпочтительном варианте антиген или антигенную детерминанту связывают с вирусоподобной частицей после повторной сборки вирусоподобной частицы, и предпочтительно после очистки композиции.
Изобретение также относится к композициям вакцин, которые могут быть использованы для профилактики и/или ослабления заболеваний или состояний. Композиции вакцин согласно изобретению содержат, или альтернативно состоят из иммунологически эффективного количества предлагаемой в изобретении усиливающей иммунитет композиции вместе с фармацевтически приемлемым разбавителем, носителем или эксципиентом. Вакцина необязательно также может содержать адъювант.
Таким образом, в предпочтительном варианте изобретение относится к вакцине, содержащей иммунологически эффективное количество предлагаемой в изобретении усиливающей иммунитет композиции вместе с фармацевтически приемлемым разбавителем, носителем или эксципиентом, при этом композиция содержит (a) вирусоподобную частицу; (b) по меньшей мере одно иммуностимулирующие вещество; и (c) по меньшей мере один антиген или антигенную детерминанту; где антиген или антигенная детерминанта связаны с вирусоподобной частицей, и где иммуностимулирующие вещество связано с вирусоподобной частицей, и где антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека. Предпочтительно вакцина, кроме того, содержит адъювант.
Изобретение, кроме того, относится к способам вакцинации для профилактики и/или ослабления заболеваний или состояний у животных. В одном варианте изобретение относится к вакцинам для профилактики злокачественной опухоли у широкого круга видов, особенно видов млекопитающих, таких как человек, обезьяна, корова, собака, кошка, лошадь, свинья и т.д., предпочтительно человек. Вакцины могут быть предназначены для лечения всех типов злокачественной опухоли, предпочтительно меланом.
Хорошо известно, что методики вакцинации на основе гомологичного примирования-ревакцинации белками или вирусами чаще всего безуспешны. Считается, что предсуществующие антитела при повторном обнаружении антигена мешают индукции ответа памяти. Неожиданно для авторов VLP, полученные из РНК-фага, в частности VLP, полученные из Qβ, очень эффективно индуцируют ответ T-клеток памяти CD8+ при использовании схемы гомологичное примирование-ревакцинация. В отличие от этого, как описано в примере 26, иммунизации живым вирусом вакцинии очень неэффективны в отношении индукции первичного ответа T-клеток CD8+, и гомологичная бустер-иммунизация вакцинией почти не приводит к экспансии T-клеток памяти CD8+.
Таким образом, в следующем аспекте изобретение относится к способу иммунизации или лечения животного, включающему в себя примирование T-клеточного ответа у животного введением иммунологически эффективного количества предлагаемой в изобретении вакцины. Предпочтительно способ, кроме того, включает в себя стадию повторной иммунизации для усиления иммунного ответа у животного, при этом предпочтительно повторную иммунизацию осуществляют введением иммунологически эффективного количества вакцины согласно изобретению или иммунологически эффективного количества гетерологичной вакцины, где еще более предпочтительной гетерологичной вакциной является ДНК-вакцина, вакцина на основе пептида, вакцина на основе рекомбинантного вируса или дендритных клеток.
Кроме того, в другом аспекте изобретение дополнительно относится к способу иммунизации или лечения животного, включающему в себя стадии примирования T-клеточного ответа у животного и повторную иммунизацию для усиления T-клеточного ответа у животного, при этом повторную иммунизацию осуществляют введением иммунологически эффективного количества вакцины согласно изобретению. Предпочтительно примирование осуществляют введением иммунологически эффективного количества вакцины согласно изобретению или иммунологически эффективного количества гетерологичной вакцины, где еще более предпочтительно указанной гетерологичной вакциной является ДНК-вакцина, вакцина на основе пептида, вакцина на основе рекомбинантного вируса или дендритных клеток.
Кроме того, в другом аспекте изобретение дополнительно относится к композиции, содержащей вирусоподобную частицу, по меньшей мере одно иммуностимулирующее вещество; и по меньшей мере один антиген или антигенную детерминанту; в которой указанный антиген или антигенная детерминанта связаны с указанной вирусоподобной частицей, и в которой указанное иммуностимулирующее вещество связано с указанной вирусоподобной частицей, и где указанный антиген содержит эпитоп для цитотоксических T-клеток, Th-клеточный эпитоп или комбинацию по меньшей мере двух указанных эпитопов, при этом указанные по меньшей мере два эпитопа связаны непосредственно или с помощью линкерной последовательности, и где предпочтительно указанный эпитоп для цитотоксических T-клеток является вирусным или опухолевым эпитопом для цитотоксических T-клеток.
В следующем аспекте настоящее изобретение относится к композиции, обычно и предпочтительно для усиления иммунного ответа у животного, содержащей: (a) вирусоподобную частицу; (b) иммуностимулирующее вещество; в которой указанное иммуностимулирующее вещество (b) связано с указанной вирусоподобной частицей (a); и (c) антиген, в которой указанный антиген смешан указанной вирусоподобной частицей (a), и при этом указанный антиген содержит, альтернативно в основном состоит, или альтернативно состоит из аналога пептида Melan-A меланомы человека. В используемом в данном описании смысле «смешаны» относится к комбинации двух или более веществ, ингредиентов или элементов, которые добавляют вместе, которые не объединены химически друг с другом и могут быть разделены. Способы смешивания антигенов с вирусоподобными частицами описаны в WO 04/000351, которая включена в данное описание в виде ссылки в полном объеме.
Как будет понятно специалисту в данной области, когда композиции согласно изобретению вводят животному, они могут быть в виде композиции, которая содержит соли, буферы, адъюванты или другие вещества, которые требуются для повышения эффективности композиции. Примеры веществ, подходящих для применения для приготовления фармацевтических композиций, предлагаются во многих источниках, включая Remington's Pharmaceutical Sciences (Osol, A, ed., Mack Publishing Co., (1990)).
Могут быть использованы различные адъюванты, чтобы увеличить иммунологический ответ, в зависимости от вида хозяина, и они включают без ограничения адъювант Фрейнда (полный и неполный), неорганические гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, многоатомные спирты Pluronic, полианионы, пептиды, масляные эмульсии, гемоцианины морского блюдечка «замочная скважина», динитрофенол и потенциально применимые адъюванты человека, такие как БЦЖ (бацилла Кальметта-Герена) и Corynebacterium parvum. Такие адъюванты также хорошо известны в данной области. Дополнительные адъюванты, которые можно вводить с композициями согласно изобретению, включают, но не ограничены указанным, монофосфориллипидный иммуномодулятор, AdjuVax 100a, QS 21, QS 18, CRL1005, соли алюминия, MF 59 и методика виросомных адъювантов. Адъюванты также могут содержать смесь указанных веществ.
Говорят, что композиции согласно изобретению являются «фармакологически приемлемыми», если может быть допустимо их введение человеку реципиенту. Кроме того, композиции согласно изобретению будут вводиться в «терапевтически эффективном количестве» (т.е., в количестве, которое дает требуемый физиологический эффект).
Композиции согласно изобретению можно вводить различными способами, известными в данной области. Конкретный выбранный способ конечно будет зависеть от конкретной выбранной композиции, тяжести состояния, подвергаемого лечению, и дозы, требуемой для терапевтической эффективности. Вообще говоря, способы согласно изобретению могут быть воплощены на практике с использованием любого способа введения, которое является приемлемым с медицинской точки зрения, подразумевая любой способ, который дает эффективные уровни активных соединений, не вызывая клинически неприемлемых нежелательных эффектов. Такие способы введения включают пероральное, ректальное, парентеральное, интрацистернальное, внутривагинальное, внутрибрюшинное, местное (как, например, с использованием порошков, мазей, капель или трансдермального пластыря), буккальное или в виде перорального или назального спрея. Термин «парентеральный» в используемом в данном описании смысле относится к способам введения, в которым относятся внутривенная, внутримышечная, внутрибрюшинная, внутригрудинная, подкожная и внутрисуставная инъекция и инфузия. Композицию согласно изобретению также можно инъецировать непосредственно в лимфатический узел.
Компоненты композиций для введения включают стерильные водные (например, физиологический раствор соли) или неводные растворы и суспензии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъекционные органические сложные эфиры, такие как этилолеат. Можно использовать носители или окклюзионные повязки, чтобы увеличить проницаемость кожи и усилить абсорбцию антигена.
Комбинации можно вводить либо в виде сопутствующих компонентов, например, в виде смеси, по отдельности, но попутно или одновременно; или последовательно. Вышесказанное относится к положениям, когда комбинируемые агенты вводят вместе в виде терапевтической смеси, а также способам, при которых комбинируемые агенты вводят по отдельности, но одновременно, например через отдельные капельницы одному и тому же человеку. Введение «в комбинации», кроме того, включает раздельное введение, когда одно из соединений или агентов вводят первым, а затем вводят второе.
Уровни доз зависят от способа введения, природы субъекта и количества композиции носитель/адъювант. Обычные количества находятся в диапазоне примерно от 0,1 мкг до примерно 20 мг субъекту. Предпочтительные количества составляют по меньшей мере примерно от 1 мкг до примерно 1 мг, более предпочтительно по меньшей мере примерно от 10 до примерно 400 мкг на субъекта. Многократное введение для иммунизации субъекта является предпочтительным, и протоколы являются стандартными протоколами в данной области, адаптированным для данного субъекта.
Композиции могут быть удобно представлены в дозированной лекарственной форме и могут быть приготовлены любым способом, хорошо известным в области фармации. Способы включают в себя стадию введения композиции согласно изобретению в ассоциацию с носителем, который представляет собой один или более вспомогательных ингредиентов. В общем композиции готовят посредством введения композиций согласно изобретению в однородную и тесную ассоциацию с жидким носителем, тонкодисперсным твердым носителем или обоими, и затем при необходимости приданием формы продукту.
Композиции, подходящие для перорального введения, могут быть представлены в виде дискретных единиц, таких как капсулы, таблетки или лепешки, каждая из которых содержит предварительно определяемое количество композиций согласно изобретению. Другие композиции включают суспензии в водных жидкостях или неводных жидкостях, такие как сироп, эликсир или эмульсия.
Другие системы доставки могут включать системы доставки с пролонгированным, замедленным или длительным высвобождением. Такие системы позволяют избегать многократных введений композиций согласно изобретению, описанных выше, делая их более удобными для субъекта и врача. Много типов системы доставки с высвобождением являются доступными и известны специалистам в данной области.
Другие варианты изобретения включают в себя способы получения композиций согласно изобретению и способы медицинского лечения злокачественной опухоли и аллергий с использованием указанных композиций.
Следующие аспекты и варианты настоящего изобретения станут понятными из следующих примеров и прилагаемой формулы изобретения.
Следующие примеры являются только иллюстративными и не предназначены для ограничения объема изобретения, который определен в прилагаемой формуле изобретения. Специалистам в данной области будет понятно, что могут быть осуществлены различные модификации и изменения в способах согласно настоящему изобретению, не отходя от сути и не выходя за рамки объема изобретения. Таким образом, подразумевается, что настоящее изобретение охватывает модификации и изменения данного изобретения, при условии что они входят в объем прилагаемой формулы изобретения, и их эквиваленты.
Все патенты и публикации, упоминаемые в данном описании, специально включены в виде ссылки в полном объеме.
ПРИМЕР 1
Создание VLP p33-HBcAg.
Последовательность ДНК HBcAg, содержащего пептид p33 из LCMV, приведена в SEQ ID NO: 15. VLP p33-HBcAg создавали следующим образом: клон гепатита B pEco63, содержащий полный вирусный геном вируса гепатита B, приобретали из ATCC.
Ген к кодирующий HBcAg вводили в сайт рестрикции EcoRI/HindIII экспрессирующего вектора pkk223.3 (Pharmacia) под контроль сильного промотора tac. Пептид p33 (KAVYNFATM) (SEQ ID NO: 80), полученный из вируса лимфоцитарного хориоменингита (LCMV), сливали с C-концом HBcAg(1-185) через линкер из трех лейцинов стандартными способами ПЦР. Клон E. coli K802, выбранный для хорошей экспрессии, трансфицировали плазмидой, и клетки выращивали и ресуспендировали в 5 ил лизирующего буфера (10 мМ Na2HPO4, 30 мМ NaCl, 10 мМ EDTA, 0,25% твин-20, pH 7,0). Добавляли 200 мкл раствора лизоцима (20 мг/мл). После обработки ультразвуком добавляли 4 мкл бензоназы и 10 мМ MgCl2 и суспензию инкубировали в течение 30 минут при комнатной температуре, центрифугировали в течение 15 минут при 15000 об/мин при 4°C и сохраняли надосадок.
Затем к надосадку добавляли 20% (мас./об.) (0,2 г/мл лизата) сульфат аммония. После инкубации в течение 30 минут на льду и центрифугирования в течение 15 минут при 20000 об/мин при 4°C надосадок отбрасывали, а осадок ресуспендировали в 2-3 мл PBS. 20 мл раствора в PBS наносили на колонку для гель-фильтрации с Sephacryl S-400 (Amersham Pharmacia Biotechnology AG), фракции наносили на SDS-ПААГ-гель и объединяли фракции с очищенными капсидами p33-VLP. Объединенные фракции наносили на колонку с гидроксиапатитом. Собирали проходящий поток (который содержит очищенные капсиды p33-VLP) и наносили на восстанавливающий SDS-ПААГ-гель для анализа молекулярной массы мономеров. Электронную микроскопию осуществляли согласно стандартным протоколам.
Таким образом, структуру p33-VLP оценивали электронной микроскопией и в SDS-ПААГ. Рекомбинантно полученные VLP дикого типа HBcAg (состоящие из мономеров HBcAg [а/к 1-185]) и p33-VLP наносили на колонку для гель-фильтрации с Sephacryl S-400 (Amersham Pharmacia Biotechnology AG) для очистки. Объединенные фракции наносили на колонку с гидроксиапатитом. Собирали проходящий поток (который содержит очищенные p33-VLP) и наносили на восстанавливающий SDS-ПААГ-гель для анализа молекулярной массы мономеров.
На протяжении описания термины VLP p33-HBcAg, VLP HBcAg-p33, p33-VLP и HBc33 используют взаимозаменяемо.
ПРИМЕР 2
Клонирование, экспрессия и очистка VLP GA
кДНК белка оболочки фага GA амплифицировали из фага GA обратной транскрипцией с последующей стадии ПЦР-амплификации, используя набор для синтеза первой нити кДНК RevertAid (Fermentas). кДНК разрезали ферментами NcoI и HindIII и клонировали в векторе pQβ185, предварительно разрезанном такими же ферментами, получая плазмиду 355.24, несущую кДНК GA. Последовательность встроенной кДНК проверяли секвенированием ДНК.
Плазмидой 355.24 трансформировали E. coli JM109. Экспрессию осуществляли по существу, как описано для VLP Qβ. Одну колонию инокулировали в среду LB, содержащую 20 мг/л ампициллина, в течение ночи без встряхивания. На следующий день указанный инокулят переносили в больший по размеру флакон, содержащий среду M9 с добавлением 1% казаминовых кислот, 0,2% глюкозы и 20 мг/л ампициллина, и инкубировали при встряхивании в течение 14-20 час.
VLP GA выделяли по существу, как описано для VLP Qβ. Клетки лизировали и осветленный лизат наносили на колонку с сефарозой CL-4B (Amersham Pharmacia). Элюат концентрировали осаждением сульфатом аммония и повторно хроматографировали на колонке с сефарозой CL-6B (Amersham Pharmacia). Конечная стадия представляла собой ультрацентрифугирование либо в градиенте сахарозы (20-50% мас./об.), либо в CsCl. Затем выделенные VLP диализовали против 20 мМ трис, 150 мМ NaCl, pH 8,0.
ПРИМЕР 3
Меченные флуоресцеином CpG-содержащие олигонуклеотиды могут быть упакованы в BKV-, HBcAg- и Qβ-VLP.
VLP, продуцированные в дрожжах, содержат небольшие количества РНК, которую можно легко расщепить и таким образом удалить посредством инкубации VLP с РНКазой A. Высокоактивный фермент РНКаза A имеет молекулярную массу около 14 кД и является достаточно маленьким, чтобы войти в VLP для удаления нежелательных рибонуклеиновых кислот. Рекомбинантно полученные VLP BKV (SEQ ID NO: 12) концентрировали до 1 мг/мл в буфере PBS, pH 7,2 и инкубировали в отсутствие или в присутствии РНКазы A (200 мкг/мл, Roche Diagnostics Ltd, Switzerland) в течение 3 час при 37°C. После расщепления РНКазой A к VLP BKV добавляли 75 нмоль/мл 5'-меченного флуоресцеином фосфоротиоатного CpG-FAM-олигонуклеотида (олигонуклеотид из SEQ ID NO: 34) и инкубировали в течение 3 час при 37°C. Затем VLP BKV подвергали расщеплению ДНКазой I в течение 3 час при 37°C (40 ед./мл AMPD1, Sigma, Division of Fluka AG, Switzerland) или наносили без расщепления ДНКазой I. Образцы дополняли 6-кратно концентрированным буфером для нанесения ДНК (10 мМ трис, pH 7,5, 10% об./об. глицерин, 0,4% оранжевый G) и разгоняли в течение 1 час при 65 вольтах в 0,8% нативном агарозном геле на трис-ацетатном буфере pH 7,5. Нуклеиновые кислоты выявляли при окрашивании бромидом этидия, в то время как в отсутствие бромида этидия возбуждение УФ приводит к флуоресценции флуоресцеиновой метки в CpG-FAM.
VLP BKV (15 мкг) анализировали электрофорезом в нативном 0,8% агарозном геле после контрольной инкубации или после расщепления РНКазой A и последующей инкубации с двунитевой (дн) ДНК (246 п.о.) (SEQ ID NO: 17), при окрашивании бромидом этидия или Кумасси синим. На гель наносили следующие образцы: 1: необработанные VLP BKV; 2: обработанные РНКазой A VLP BKV; 3: VLP BKV, обработанные РНКазой A и инкубированные с днДНК; дорожка M: лэддер ДНК длиной 1 т.п.о. Gene Ruler (MBI Fermentas GmbH, Heidelberg, Germany).
VLP BKV (15 мкг) анализировали электрофорезом в нативном 0,8% агарозном геле после контрольной инкубации или после расщепления РНКазой A и последующей инкубации с CpG-олигонуклеотидами (с фосфатным или фосфоротиоатным (pt) остовом) при окрашивании бромидом этидия или Кумасси синим. НА гель наносили следующие образцы: 1: исходные VLP BKV (PBS/50% глицерин); 2: необработанные VLP BKV (буфер PBS); 3: обработанные РНКазой A VLP BKV; 4: обработанные РНКазой A VLP BKV после диализа; 5: Обработанные РНКазой A VLP BKV с CpG-олигонуклеотидами; 6: обработанные РНКазой A VLP BKV c CpG(pt)-олигомерами; 7: обработанные РНКазой A VLP BKV c CpG(pt)-олигомерами после диализа; дорожка M: лэддер ДНК длиной 1 т.п.о. Gene Ruler (MBI Fermentas GmbH, Heidelberg, Germany).
Расщепление РНКазой A приводит к изменению миграции VLP, видимой на окрашенном Кумасси агарозном геле, предположительно вследствие потери отрицательных зарядов РНК. Добавление CpG-олигонуклеотида возвращает миграцию VLP BKV в прежнее состояние и в результате дает флуоресцирующую полосу с такой же миграцией, как и РНК-полоса, присутствующая в необработанных VLP. Это четко показывает, что CpG-FAM-олигонуклеотиды были упакованы в VLP.
ПРИМЕР 4
Крупные двунитевые олигонуклеотиды могут быть упакованы в VLP BKV
Чтобы ввести двунитевые (дн) нуклеотидные последовательности, к обработанным РНКазой A рекомбинантным VLP BKV (пример 3) добавляли 50 мкг/мл фрагментов (дн)ДНК (длиной 246 п.о., днДНК, SEQ ID NO: 17) и инкубировали в течение 3 час при 37°C. Образцы дополняли 6-кратно концентрированным буфером для нанесения ДНК (10 мМ трис, pH 8,0, 10% об./об. глицерин, 0,4% оранжевый G) и разгоняли в течение 1 час при 65 вольтах в 0,8% нативном агарозном геле на трис-ацетатном буфере pH 8,0. VLP BKV (15 мкг) наносили на нативный 0,8% агарозный гель для электрофореза и анализировали после контрольной инкубации или после расщепления РНКазой A и последующей инкубации с (дн)ДНК при окрашивании бромидом этидия или Кумасси синим, чтобы оценить наличие РНК/ДНК или белка. Упакованные молекулы ДНК видны в присутствии бромида этидия в виде полосы с такой же скоростью миграции, как и полоса VLP, визуализированная Кумасси синим.
Добавление (дн)ДНК восстанавливает миграцию VLP BKV до исходного состояния и в результате дает полосу ДНК с такой же скоростью миграции, как и окрашенные Кумасси синим VLP. Это четко показывает, что (дн)ДНК была упакована в VLP BKV.
ПРИМЕР 5
CpG-содержащие олигонуклеотиды могут быть упакованы в VLP BKV.
Чтобы ввести иммуностимулирующие CpG-олигонуклеотиды, к обработанным РНКазой A рекомбинантным VLP BKV (пример 3) добавляли 150 нмоль/мл CpG-олигонуклеотидов CyCpG с фосфодиэфирным остовом или CyCpGpt с фосфоротиоатным остовом и инкубировали в течение 3 час при 37°C. Препараты VLP для иммунизации мышей диализовали против большого объема (10000-кратное разведение) в течение 24 час против PBS, pH 7,2, используя диализную мембрану с отсечением по молекулярной массе 300 кД (Spectrum Medical industries Inc., Houston, USA), чтобы удалить РНКазу A и избыток CpG-олигонуклеотидов. Образцы дополняли 6-кратно концентрированным буфером для нанесения ДНК (10 мМ трис, pH 7,5, 10% об./об. глицерин, 0,4% оранжевый G) и разгоняли в течение 1 час при 65 вольтах в 0,8% нативном агарозном геле на трис-ацетатном буфере pH 7,5. VLP BKV (15 мкг) наносили на нативный 0,8% агарозный гель для электрофореза и анализировали после контрольной инкубации или после расщепления РНКазой A и последующей инкубации с CpG-олигонуклеотидами (с фосфодиэфирным или фосфоротиоатным остовом) при окрашивании бромидом этидия или Кумасси синим, чтобы оценить наличие РНК/ДНК или белка, и уменьшения количества несвязанных CpG-олигонуклеотидов после диализа. Несвязанные CpG-олигонуклеотиды видны в виде окрашенной бромидом этидия полосы с низкой молекулярной массой. Добавление CpG-олигонуклеотидов восстанавливает миграцию VLP BKV до исходного состояния и в результате дает полосу ДНК с такой же скоростью миграции, как и окрашенные Кумасси синим VLP. Это четко показывает, что CpG-олигонуклеотиды упакованы в VLP BKV.
ПРИМЕР 6
VLP, содержащие CpG-олигонуклеотиды (с фосфоротиоатной модификацией фосфатного остова), индуцируют усиленный направленный иммунный ответ Th1.
Самкам мышей BALB/c (три мыши на группу) подкожно инъецировали 10 мкг VLP BKV, содержащих фосфоротиоатный CpG-олигонуклеотид CyCpGpt (SEQ ID NO: 34). В качестве контролей мышам подкожно инъецировали либо 10 мкг обработанных РНКазой VLP BKV отдельно, либо VLP BKV, смешанных с 0,3 нмоль или 20 нмоль фсофоротиоатных CpG-олигонуклеотидов в 200 мкл PBS, pH 7,2, или оставляли необработанными. VLP BKV готовили, как описано в примере 5 и перед иммунизацией тщательно очищали от несвязанного CpG-олигонуклеотида диализом. На 14 день после иммунизации брали кровь и определяли ответ в виде антител IgG1 и IgG2a на VLP BKV (смотри таблицу 1).
Иммунизация обработанными РНКазой A VLP BKV, содержащими фосфоротиоатные CpG-олигонуклеотиды CyCpGpt приводит к пониженному титру IgG1 и повышенному титру анти-VLP BKV IgG2a по сравнению с иммунизацией таким же количеством (0,3 нмоль) CpG-олигонуклеотидов, смешанных с VLP BKV, или отдельно VLP BKV. У мышей, иммунизированных VLP BKV, смешанными с 20 нмоль фосфоротиоатного CpG-олигонуклеотида CyCpGpt, наблюдали очень низкие титры IgG1 и высокие титры IgG2a. Уменьшение титра IgG1 и увеличение титра IgG2a по сравнению с контролями свидетельствует о Th1-клеточном направленном иммунном ответе, индуцированном фосфоротиоатными CpG-олигонуклеотидами, упакованными в VLP BKV. Таблица 1 четко показывает более высокую эффективность VLP BKV, содержащих CpG-олигонуклеотиды, упакованные в частицы, по сравнению с VLP BKV, смешанными с CpG-олигонуклеотидами.
ПРИМЕР 7
Иммуностимулирующие нуклеиновые кислоты могут быть упакованы в VLP HBcAg, содержащие слитые белки с антигенами.
VLP HBcAg при продуцировании в E. coli посредством экспрессии слитого белка корового антигена гепатита B p33-HBcAg (HBc33) (смотри пример 1) или слитого белка с пептидом P1A (HBcP1A) содержат РНК, которую можно расщепить и таким образом удалить посредством инкубации VLP с РНКазой A.
Ген P1A кодирует белок, который экспрессируется линией опухолевых клеток мастоцитомы P815. Доминантный CTL-эпитоп, названный пептидом P1A, связывает MHC класса I (Ld) и комплекс распознается специфичными клонами CTL (Brandle et al., 1998, Eur. J. Immunol. 28: 4010-4019). Слияние пептида P1A-1 (LPYLGWLVF) ((SEQ ID NO: 90) с C-концом HBcAg (а/к 185, смотри пример 1) осуществляли посредством ПЦР, используя соответствующие праймеры и используя стандартные методики молекулярной биологии. Линкер из трех лейцинов клонировали между HBcAg и последовательностью пептида. Экспрессию осуществляли, как описано в примере 1. Слитый белок HBcAg с P1A, названный HBcP1A, образовывал капсиды при экспрессии в E. coli, которые можно было очистить способом, подобным способу, описанному в примере 1.
Ферментативный гидролиз РНК: Рекомбинантно полученные VLP HBcAg-p33 (HBc33) и HBcAg-P1A (HBcP1A) в концентрации 1,0 мг/мл в 1 x PBS-буфере (KCl 0,2 г/л, KH2PO4 0,2 г/л, NaCl 8 г/л, Na2HPO4 1,15 г/л) pH 7,4, инкубировали в присутствии 300 мкг/мл РНКазы A (Qiagen AG, Switzerland) в течение 3 час при 37°C в термомиксере при 650 об/мин.
Упаковка иммуностимулирующих нуклеиновых кислот: После расщепления РНК РНКазой A к VLP HBcAg-p33 добавляли 130 нмоль/мл CpG-олигонуклеотидов B-CpG, NKCpG, G10-PO (таблица 2). Подобным образом добавляли 150-мерный однонитевой Cy150-1 и 253-мерный двунитевой dsCyCpG-253, оба содержащие множественные копии мотивов CpG в концентрации 130 нмоль/мл или 1,2 нмоль/мл соответственно, и инкубировали в термомиксере в течение 3 час при 37°C. Двунитевую ДНК CyCpG-253 получали клонированием двунитевого мультимера CyCpG в сайте EcoRV pBluescript KS-. Полученную в результате плазмиду, продуцированную в E. coli XL 1-blue и выделенную с использованием набора Qiagen Endofree plasmid Giga, расщепляли эндонуклеазами рестрикции XhoI и XbaI и полученные в результате продукты рестрикции разделяли электрофорезом в агарозе. Инсерцию длиной 253 п.о. выделяли электроэлюцией и осаждением этанолом. Последовательность подтверждали секвенированием обеих нитей.
Таблица 2: Терминология и последовательности иммуностимулирующих нуклеиновых кислот, используемых в примерах.
Маленькими буквами указаны дезоксинуклеотиды, связанные посредством фосфоротиоатных связей, тогда как заглавными буквами указаны дезоксинуклеотиды, связанные посредством фосфодиэфирных связей.
Обработка ДНКазой I: Упакованный VLP HBcAg-p33 затем подвергали расщеплению ДНКазой I (5 ед./мл) в течение 3 час при 37°C (ДНКаза I, не содержащая РНКазы Fluka AG, Switzerland) и диализовали против большого объема (2×против 200-кратного объема) в течение 24 час против PBS, pH 7,4, используя диализную мембрану с пределом отсечения по молекулярной массе 300 кД (Spectrum Medical industries Inc., Houston, USA), чтобы удалить РНКазу A и избыток CpG-олигонуклеотидов.
Обработка бензоназой: Так как некоторые однонитевые олигодезоксинуклеотиды частично резистентны к обработке ДНКазой I, использовали обработку бензоназой, чтобы удалить свободные олигонуклеотиды из препарата. Добавляли 100-120 ед./мл бензоназы (Merck KGaA, Darmstadt, Germany) и 5 мМ MgCl2 и инкубировали в течение 3 час при 37°C перед диализом.
Диализ: Препараты VLP, упакованные с иммуностимулирующими нуклеиновыми кислотами, используемые в экспериментах по иммунизации мышей, диализовали против большого объема (2×против 200-кратного объема) в течение 24 час против PBS, pH 7,4, используя диализную мембрану с пределом отсечения по молекулярной массе 300 кД (Spectrum Medical Industries, Houston, US), чтобы удалить добавленные ферменты и свободные нуклеиновые кислоты.
Анализ упаковки: Высвобождение упакованных иммуностимулирующих нуклеиновых кислот: К 50 мкл раствора капсида добавляли 1 мкл протеиназы K (600 ед./мл, Roche,Mannheim, Germany), 3 мкл 10% SDS-раствора и 6 мкл 10-кратного буфера для протеиназы (0,5М NaCl, 50 мМ EDTA, 0,1М трис, pH 7,4) и затем инкубировали в течение ночи при 37°C. VLP полностью гидролизовали в данных условиях. Протеиназу K инактивировали нагреванием в течение 20 мин при 65°C. 1 мкл РНКазы A (Qiagen, 100 мкг/мл, разбавленная в 250 раз) добавляли к 25 мкл раствора капсидов. 2-30 мкг капсида смешивали с 1 объемом 2x-буфера для нанесения (1xTBE, 42% мас./об. мочевины, 12% мас./об. фикола, 0,01% бромфенолового синего), нагревали в течение 3 мин при 95°C и наносили на 10% (в случае олигонуклеотидов длиной примерно 20 нуклеотидов) или 15% (в случае> чем 40-мерных нуклеиновых кислот) полиакриламидный гель с TBE/мочевиной (Invitrogen). Альтернативно образцы наносили на 1% агарозный гель с 6x красителем для нанесения (10 мМ трис pH 7,5, 50 мМ EDTA, 10% об./об. глицерин, 0,4% оранжевый G). Гели с TBE/мочевиной красили SYBRGold, а агарозные гели красили бромидом этидия.
Олигонуклеотиды B-CpG, NKCpG и G10-PO упаковывали в HBc33. Анализ B-CpG, упакованных в VLP HBc33, проводили на 1% агарозном геле, окрашенном бромидом этидия и Кумасси синим. На гели наносили 50 мкг следующих образцов: 1) необработанные VLP HBc33; 2) VLP HBc33, обработанные РНКазой A; 3) VLP HBc33, обработанные РНКазой A и упакованные с B-CpG; 4) VLP HBc33, обработанные РНКазой A, упакованные с B-CpG и обработанные ДНКазой I; 5) VLP HBc33, обработанные РНКазой A, упакованные с B-CpG, обработанные ДНКазой I и диализованные; лэддер ДНК длиной 6,1 т.п.о. MBI Fermentas. Количество упакованного B-CpG, экстрагированного из VLP, анализировали в 1,5% агарозном геле, окрашенном бромидом этидия: На гель наносили следующие образцы: 1) 0,5 нмоль контроля B-CpG; 2) 0,5 нмоль контроля B-CpG; 3) B-CpG-олигонуклеотидное содержимое HBc33 после экстракции фенолом/хлороформом; 4) B-CpG-олигонуклеотидное содержимое HBc33 после экстракции фенолом/хлороформом и обработки РНКазой A; 5) B-CpG-олигонуклеотидное содержимое HBc33 после экстракции фенолом/хлороформом и обработки ДНКазой I; 6) пустая; 7) лэддер ДНК длиной 100 п.о. MBI Fermentas.
Анализ NKCpG, упакованного в VLP HBc33, осуществляли в 1% агарозном геле, окрашенном бромидом этидия и Кумасси синим. На гель наносили 15 мкг следующих образцов: 1) необработанные VLP HBc33; 2) VLP HBc33, обработанные РНКазой A; 3) VLP HBc33, обработанные РНКазой A и упакованные с NKCpG; 4) VLP HBc33, обработанные РНКазой A, упакованные с NKCpG, обработанные ДНКазой I и диализованные; лэддер ДНК длиной 5,1 т.п.о. MBI Fermentas. Анализ количества упакованного NKCpG, экстрагированного из VLP, анализировали в 15% гелях с TBE/мочевиной, окрашенных SYBR Gold. На гель наносили следующие образцы: 1) NKCpG-олигонуклеотидное содержимое HBc33 после расщепления протеиназой K и обработки РНКазой A; 2) 20 пмоль контрольного NKCpG; 3) 10 пмоль контрольного NKCpG; 4) 40 пмоль контрольного NKCpG.
Анализ g10gacga-PO, упакованного в VLP HBc33, осуществляли в 1% агарозном геле, окрашенном бромидом этидия и Кумасси синим. На гель наносили 15 мкг следующих образцов: 1) лэддер ДНК длиной 1 т.п.о MBI Fermentas; 2) необработанные VLP HBc33; 3) VLP HBc33, обработанные РНКазой A; 4) VLP HBc33, обработанные РНКазой A и упакованные с g10gacga-PO; 5) VLP HBc33, обработанные РНКазой A, упакованные с g10gacga-PO, обработанные бензоназой и диализованные.
Содержание РНК в VLP сильно уменьшалось после обработки РНКазой A, при этом большинство капсидов мигрировало в виде медленно мигрирующего пятна, предположительно вследствие удаления отрицательно заряженной РНК. После инкубации с избытком олигонуклеотидов капсиды содержали большее количество нуклеиновой кислоты, чем обработанные РНКазой A капсиды, и поэтому мигрировали со скоростью, сходной со скоростью необработанных капсидов. Дополнительная обработка ДНКазой I или бензоназой приводила к деградации свободных олигонуклеотидов, тогда как олигонуклеотиды, упакованные в капсиды, не разрушались, что четко свидетельствовало об упаковке олигонуклеотидов. В некоторых случаях упаковку олигонуклеотидов подтверждали при расщеплении протеиназой K после обработки ДНКазой I/бензоназой и диализа. Обнаружение того, что олигонуклеотиды, высвобождаемые из капсида при использовании описанного выше способа, были такого же размера, что и олигонуклеотид, используемый для упаковки, четко свидетельствует об упаковке олигонуклеотидов.
Крупный однонитевой олигонуклеотид Cy150-1 упаковывали в HBc33. Cy150-1 содержит 7,5 повторов CyCpG, и он был синтезирован согласно стандартным способам синтеза олигонуклеотидов (IBA, Göttingen, Germany). Анализ Cy150-1, упакованного в VLP HBc33 проводили в 1% агарозном геле, окрашенном бромидом этидия и Кумасси синим. На гель наносили 15 мкг следующих образцов: 1) лэддер ДНК длиной 1 т.п.о. MBI Fermentas; 2) необработанные VLP HBc33; 3) VLP HBc33, обработанные РНКазой A; 4) VLP HBc33, обработанные РНКазой A и упакованные с Cy150-1; 5) VLP HBc33, обработанные РНКазой A, упакованные с Cy150-1, обработанные ДНКазой I и диализованные; 6) VLP HBc33, обработанные РНКазой A, упакованные с Cy150-1, обработанные ДНКазой I и диализованные. Анализ количества упакованного Cy150-1, экстрагированного из VLP, анализировали в 10% геле с TBE/мочевиной, окрашенном SYBR Gold. На гель наносили следующие образцы: 1) 20 пмоль контрольного Cy150-1; 2) 10 пмоль контрольного Cy150-1; 3) 4 пмоль контрольного Cy150-1; 4) Cy150-1-олигонуклеотидное содержимое 4 мкг HBc33 после 3 мин при 95°C с 1 объемом буфера для образцов с TBE/мочевиной. Содержание РНК в капсиде сильно снижалось после обработки РНКазой A при этом большинство капсидов мигрировало в виде медленно мигрирующего пятна. Капсиды разбавляли 4 объемами воды и концентрировали до 1 мг/мл. После инкубации с избытком Cy150-1 капсид содержал большее количество нуклеиновой кислоты и соответственно мигрировал со скоростью, сходной со скоростью необработанных капсидов. Дополнительная обработка ДНКазой I вызвала деградацию свободные, неупакованных олигонуклеотидов, тогда как олигонуклеотиды в капсидах не разрушались. Высвобождение резистентной к ДНКазе I нуклеиновой кислоты из упакованных VLP при нагревании в течение 3 мин при 95°C в буфере для нанесения с TBE/мочевиной свидетельствовало о присутствии 150-мера.
Олигонуклеотид NKCpGpt также упаковывали в HBcP1A. Анализ NKCpGpt, упакованного в VLP HBcP1A, осуществляли в 1% агарозном геле, окрашенном бромидом этидия и Кумасси синим. На гель наносили 15 мкг следующих образцов: 1) лэддер ДНК длиной 1 т.п.о MBI Fermentas; 2) необработанные VLP HBcP1A; 3) VLP HBcP1A, обработанные РНКазой A; 4) VLP HBcP1A, обработанные РНКазой A и упакованные с NKCpGpt. Обработка РНКазой уменьшала содержание нуклеиновой кислоты и замедляла миграцию капсидов. Добавление NKCpGpt восстанавливало содержание нуклеиновой кислоты в капсидах и ускоряло миграцию.
ПРИМЕР 8
Иммуностимулирующие нуклеиновые кислоты могут быть упакованы в HBcAg-wt, связанный с антигенами.
Рекомбинантно продуцированные VLP HBcAg-wt упаковывали после связывания с пептидом p33 (CGG-KAVYNFATM) (SEQ ID NO: 81), полученные из вируса лимфоцитарного хориоменингита (LCMV). Для связывания VLP HBcAg-wt (2 мг/мл) дериватизировали 25x молярным избытком SMPH (сукцинимидил-6-[(β-малеимидопропионамидо)гексаноат], Pierce) в течение 1 час при 25°C в термомиксере. Дериватизованные VLP диализовали против Mes-буфера (2-(N-морфолино)этансульфоновая кислота), pH 7,4, в течение 2×2 час, используя диализные мембраны с пределом отсечения по молекулярной массе 10000 кД при 4°C. Затем VLP (50 мкМ) связывали с N-концевым цистеином пептида p33 (250 мкМ) в течение 2 час инкубации в термомиксере при 25°C. Образцы диализовали (предел отсечения по молекулярной массе 300000) против большого объема 1×PBS pH 7,4, чтобы удалить нежелательный свободный пептид.
Дериватизацию VLP HBcAg-wt с использованием SMPH и связывание с пептидом p33 анализировали в SDS-ПААГ. Образцы анализировали в 16% SDS-ПААГ и красили Кумасси синим. На гель наносили следующие образцы: 1) предварительно окрашенный белковый маркер NEB, широкого диапазона (№7708S), 10 мкл; 2) пептид p33; 3) VLP HBcAg-wt, дериватизованные SMPH, перед диализом; 4) VLP HBcAg-wt, дериватизованные SMPH, после диализа; 5) VLP HBcAg-wt, связанные с p33, надосадок; 6) VLP HBcAg-wt, связанные с p33, осадок. HBcAg-wt наблюдали в виде полосы белка 21 кД. Вследствие низкой молекулярной массы SMPH дериватизованный продукт только не намного больше и может быть не отличим в SDS-ПААГ. Пептид отдельно виден в виде полосы 3 кД, и связанный продукт, называемый HBx33, виден в виде интенсивной вторичной полосы примерно в области 24 кД, составляющей более 50% от общего HBcAg-wt.
Ферментативный гидролиз РНК: VLP HBx33 (0,5-1,0 мг/мл, 1×PBS-буфер, pH 7,4) в присутствии РНКазы A (300 мкг/мл, Qiagen AG, Switzerland) разбавляли 4 объемами H2O, чтобы уменьшить концентрацию соли до конечной концентрации 0,2 x PBS, и инкубировали в течение 3 час при 37°C в термомиксере при 650 об/мин.
Упаковка иммуностимулирующих нуклеиновых кислот: После расщепления РНКазой A VLP HBx33 концентрировали, используя концентрирующие устройства Millipore Microcon или Centriplus, затем добавляли 130 нмоль/мл CpG-олигонуклеотида B-CpGpt и инкубировали в термомиксере в течение 3 час при 37°C в 0,2×PBS, pH 7,4. Затем реакционные смеси подвергали расщеплению ДНКазой I (5 ед./мл) в течение 3 час при 37°C (ДНКаза I, не содержащая РНКазы, Fluka AG, Switzerland). Препараты VLP для иммунизации мышей диализовали против большого объема (2x против 200-кратного объема) в течение 24 час против PBS pH 7,4, используя диализную мембрану с пределом отсечения по молекулярной массе 300 кД (Spectrum Medical industries Inc., Houston, USA), чтобы удалить РНКазу A и избыток CpG-олигонуклеотидов. Анализ B-CpGpt, упакованного в VLP HBx33, осуществляли в 1% агарозном геле, окрашенном бромидом этидия и Кумасси синим. На гель наносили 50 мкг следующих образцов: 1) необработанные VLP HBx33; 2) VLP HBx33, обработанные РНКазой A; 3) VLP HBx33, обработанные РНКазой A и упакованные с B-CpGpt; 4) VLP HBx33, обработанные РНКазой A, упакованные с B-CpGpt и обработанные ДНКазой I; 5) VLP HBx33, обработанные РНКазой A, упакованные с B-CpGpt, обработанные ДНКазой I и диализованные; 6) лэддер ДНК длиной 1 т.п.о. MBI Fermentas. Можно видеть, что обработка РНКазой снижала содержание нуклеиновой кислоты в капсидах и замедляла их миграцию. Добавление B-CpGpt восстанавливало содержание нуклеиновой кислоты и ускоряло миграцию капсидов. ДНКаза I расщепляла только свободные олигонуклеотиды, тогда как упакованные олигонуклеотиды сохранялись в VLP также после диализа.
ПРИМЕР 9
Иммуностимулирующие нуклеиновые кислоты могут быть упакованы в VLP Qβ, связанные с антигенами.
Связывание пептидов p33 с VLP Qβ: Рекомбинантно полученные вирусоподобные частицы РНК-бактериофага Qβ (VLP Qβ) использовали необработанными или после связывания с пептидами p33, содержащими N-концевое удлинение CGG или/и C-концевое удлинение GGC (CGG-KAVYNFATM (SEQ ID NO: 81) и KAVYNFATM-GGC (SEQ ID NO: 82)). Рекомбинантно полученные VLP Qβ подвергали дериватизации 10 молярным избытком SMPH (Pierce) в течение 0,5 час при 25°C с последующим диализом против 20 мМ HEPES, 150 мМ NaCl, pH 7,2, при 4°C, чтобы удалить непрореагировавший SMPH. Пептиды добавляли в 5-кратном молярном избытке и давали возможность взаимодействовать в течение 2 час в термомиксере при 25°C в присутствии 30% ацетонитрила. Анализ связывания p33 с VLP Qβ осуществляли в SDS-ПААГ после окрашивания Кумасси синим. Наносили следующие образцы: (A) 1) предварительно окрашенный белковый маркер NEB, широкого диапазона, (№7708S), 10 мкл; 2) VLP Qβ, 14 мкг; 3) VLP Qβ, дериватизованные SMPH, после диализа; 4) VLP Qβ, связанные с CGG-p33, надосадок. (B) 1) предварительно окрашенный белковый маркер NEB, широкого диапазона, (№7708S), 10 мкл; 2) VLP Qb, 10 мкг; 3) VLP Qb, связанные с GGC-p33, надосадок. Анализ в SDS-ПААГ показал множество полос продуктов связывания, состоящих из одного, двух или трех пептидов, связанных с мономером Qβ. Для простоты продукт связывания пептида p33 и VLP Qβ в частности называют в разделе примеров Qbx33.
VLP Qβ в случае продуцирования в E. coli посредством экспрессии белка капсида бактериофага Qβ содержат РНК, которую можно расщепить и таким образом удалить при инкубации VLP с РНКазой A.
Низкая ионная сила и низкая концентрация Qβ обеспечивают возможность гидролиза РНК в VLP Qβ РНКазой A:
VLP Qβ в концентрации 1,0 мг/мл в буфере 20 мМ Hepes/150мМ NaCl (HBS) с pH 7,4 либо расщепляли, непосредственно добавляя РНКазу A (300 мкг/мл, Qiagen AG, Switzerland), либо разбавляли 4 объемами H2O до конечной концентрации 0,2×HBS и затем инкубировали с РНКазой A (60 мкг/мл, Qiagen AG, Switzerland). Инкубацию осуществляли в течение 3 час при 37°C в термомиксере при 650 об/мин. Гидролиз РНК из VLP Qb РНКазой A при низкой и высокой ионной силе анализировали в 1% агарозном геле, окрашенном бромидом этидия и Кумасси синим. На гель наносили следующие образцы: (A, B) 1) лэддер ДНК длиной 1 т.п.о. MBI Fermentas; 2) необработанные VLP Qb; 3) VLP Qb, VLP обработанные РНКазой A в 1 x HBS-буфере pH 7,2. (C, D) 1) лэддер ДНК длиной 1 т.п.о. MBI Fermentas; 2) необработанные VLP Qb; 3) VLP Qb, обработанные РНКазой A в 0,2 x HBS-буфере pH7,2. Показано, что в 1×HBS наблюдалось только очень слабое уменьшение содержания РНК, тогда как в 0,2 x HBS большая часть РНК подвергалась гидролизу. В соответствии с этим миграция капсидов не изменялась после добавления РНКазы A в 1 x HBS, тогда как миграция была более медленной после добавления РНКазы в 0,2 x HBS.
Низкая ионная сила увеличивает упаковку нуклеиновой кислоты в VLP Qβ:
После расщепления РНКазой A в 0,2 x HBS VLP Qβ концентрировали до 1 мг/мл, используя концентрирующие устройства Millipore Microcon или Centriplus, и аликвоты диализовали против 1×HBS или 0,2 x HBS. К VLP Qβ добавляли 130 нмоль/мл CpG- олигонуклеотида B-CpG и инкубировали в термомиксере в течение 3 час при 37°C. Затем VLP Qβ подвергали расщеплению бензоназой (100 ед./мл) в течение 3 час при 37°C. Образцы анализировали в 1% агарозных гелях после окрашивания бромидом этидия и Кумасси синим. На гель наносили следующие образцы: 1) необработанные VLP Qb; 2) VLP Qb, обработанные РНКазой A; 3) VLP Qb, обработанные РНКазой A и упакованные с B-CpG в 0,2×HBS-буфере, pH 7,2, и обработанные бензоназой; 4) VLP HBx33 (смотри пример 12), обработанные РНКазой A, упакованные с B-CpG в 1×HBS-буфере, pH 7,2, и обработанные бензоназой. В 1×HBS только очень небольшое количество олигонуклеотидов могло быть упаковано, тогда как в 0,2×HBS можно было выявить интенсивно окрашенную бромидом этидия полосу, которая располагалась совместно с окрашенными Кумасси синим капсидами.
Различные иммуностимулирующие нуклеиновые кислоты могут быть упакованы в VLP Qβ и Qbx33:
После расщепления РНКазой A в 0,2 x HBS VLP Qβ или VLP Qb33 концентрировали до 1 мг/мл, используя концентрирующие устройства Millipore Microcon или Centriplus, и добавляли 130 нмоль/мл CpG-олигонуклеотидов B-CpGpt, g10gacga и 253-мера dsCyCpG-253 (таблица 2) и инкубировали в термомиксере в течение 3 час при 37°C. Затем VLP Qβ или VLP Qb33 подвергали расщеплению ДНКазой I (5 ед./мл) или расщеплению бензоназой (100 ед./мл) в течение 3 час при 37°C. Образцы анализировали в 1% агарозных гелях после окрашивания бромидом этидия и Кумасси синим.
На гель наносили 50 мкг следующих образцов: 1) необработанные VLP Qb33; 2) VLP Qb33, обработанные РНКазой A; 3) VLP Qb33, обработанные РНКазой A и упакованные с B-CpGpt; 4) VLP Qb33, обработанные РНКазой A, упакованные с B-CpGpt, обработанные ДНКазой I и диализованные; 5) лэддер ДНК 1 т.п.о. MBI Fermentas. (C) показывает анализ количества упакованных олигонуклеотидов, экстрагированные из VLP, в 15% гелях с TBE/мочевиной, окрашенных SYBR Gold. НА гель наносили следующие образцы: 1) BCpGpt-олигонуклеотидное содержимое 2 мкг VLP Qb33 после расщепления протеиназой K и обработки РНКазой A; 2) 20 пмоль контрольного B-CpGpt; 3) 10 пмоль контрольного B-CpGpt; 4) 5 пмоль контрольного B-CpGpt.
На другой гель наносили 15 мкг следующих образцов: 1) лэддер ДНК длиной 1 т.п.о. MBI Fermentas; 2) необработанные VLP Qb33; 3) VLP Qb33, обработанные РНКазой A; 4) VLP Qb33, обработанные РНКазой A и упакованные с g10gacga-PO; 5) VLP Qb33, обработанные РНКазой A, упакованные с g10gacga-PO, обработанные бензоназой и диализованные.
На третий гель наносили 15 мкг следующих образцов: 1) лэддер ДНК длиной 1 т.п.о. MBI Fermentas; 2) необработанные VLP Qb33; 3) VLP Qb33, обработанные РНКазой A; 4) VLP Qb33, обработанные РНКазой A, упакованные с dsCyCpG-253 и обработанные ДНКазой I; 5) VLP Qb33, обработанные РНКазой A, упакованные с dsCyCpG-253, обработанные ДНКазой I и диализованные.
Различные нуклеиновые кислоты B-CpGpt, g10gacga и 253-мерную днДНК могли быть упакованы в Qbx33. Упакованные нуклеиновые кислоты были резистентны к расщеплению ДНКазой I и оставались упакованными во время диализа. Упаковку B-CpGpt подтверждали по высвобождению нуклеиновой кислоты при расщеплении протеиназой K с последующим электрофорезом в агарозном геле и окрашиванием бромидом этидия.
ПРИМЕР 10
Разборка - очистка и повторная сборка AP205 и упаковка иммуностимулирующих нуклеиновых кислот.
A. Разборка и повторная сборка VLP AP205 из материала, способного повторно подвергаться сборке, без добавления олигонуклеотида.
Разборка: 40 мг лиофилизованных очищенных VLP AP205 (SEQ ID: 80 или 81) перерастворяли в 4 мл 6М CuHCl и инкубировали в течение ночи при 4°C. Смесь для разборки центрифугировали при 8000 об/мин (Eppendorf 5810 R, в угловом роторе F34-6-38, используемом на всех следующих стадиях). Осадок перерастворяли в 7М мочевине, тогда как надосадок диализовали 3 дня против буфера NET (20 мМ трис-HCl, pH 7,8, с 5 мМ EDTA и 150 мМ NaCl) с 3 заменами буфера. Альтернативно диализ проводили в непрерывном режиме в течение 4 дней. Диализованный раствор центрифугировали при 8000 об/мин в течение 20 минут и осадок перерастворяли в 7М мочевине, тогда как надосадок осаждали сульфатом аммония (60% насыщение) и перерастворяли в буфере с 7М мочевиной, содержащем 10 мМ DTT. Все предыдущие осадки, перерастворенные в 7М мочевине, объединяли и осаждали сульфатом аммония (60% насыщение) и перерастворяли в буфере с 7М мочевиной, содержащем 10 мМ DTT. Вещества, перерастворенные в буфере с 7М мочевиной, содержащем 10 мМ DTT, объединяли и наносили на уравновешенную колонку с сефадексом G75 и элюировали буфером с 7М мочевиной, содержащим 10 мМ DTT со скоростью 2 мл/час. С колонки элюировался один пик. Собирали фракции по 3 мл. Фракции пика, содержащие белок оболочки AP205, объединяли и осаждали сульфатом аммония (60% насыщение). Осадок отделяли центрифугированием при 8000 об/мин в течение 20 минут. Его перерастворяли в 7М мочевине, 10 мМ DTT и наносили на короткую колонку с сефарозой 4B (1,5×27 см, сефароза 4B, 2 мл/час, 7М мочевина, 10 мМ DTT в качестве элюирующего буфера). С колонки элюировался в основном один пик с небольшим плечом. Фракции, содержащие белок оболочки AP205, идентифицировали в SDS-ПААГ и объединяли за исключением фракций плеча. При этом получали образец объемом 10,3 мл. Концентрацию белка оценивали спектрофотометрически, измеряя аликвоту белка, разбавленного для измерения в 25 раз, используя следующую формулу: (1,55×OD280-0,76×OD260) x объем. Средняя концентрация составляла 1 нмоль/мл VLP (2,6 мг/мл). Отношение оптической плотности при 280 нм по сравнению с 260 нм составляло 0,12/0,105.
Повторная сборка: к образцу добавляли 1,1 мл бета-меркаптоэтанола и следующие реакции повторной сборки проводили согласно схемам:
1 мл белка оболочки AP205 без нуклеиновой кислоты,
1 мл белка оболочки AP205, рРНК (примерно 200 единиц OD260, 10 нмоль),
9 мл белка оболочки AP205, CyCpG (370 мкл раствора 225 пмоль/мкл, т.е. 83 нмоль).
Указанные смеси диализовали 1 час против 30 мл буфера NET, содержащего 10% бета-меркаптоэтанол. Смесь, не содержащую нуклеиновых кислот, диализовали отдельно. Затем диализ проводили в непрерывном режиме и 1 л буфера NET меняли через 3 дня. Затем реакционные смеси диализовали против большого объема воды (5 замен буфера) и лиофилизовали. Их перерастворяли в воде и анализировали в электронном микроскопе (ЭМ). Все смеси содержали капсиды, свидетельствуя о том, что повторная сборка VLP AP205 не зависит от присутствия выявляемых нуклеиновых кислот, которые измеряли при электрофорезе в агарозном геле, используя окраску бромидом этидия. Способ ЭМ осуществляли следующим образом. Суспензию белков адсорбировали на покрытых углеродом-формваром сетках и красили 2% фосфорновольфрамовой кислотой (pH 6,8). Сетки исследовали в электронном микроскопе JEM100C (JEOL, Japan) при ускоряющем напряжении 80 кВ. Фоторегистрацию (негативы) осуществляли на пленке для получения электронных изображений Kodak и электронные микрофотографии получали, печатая негативы на бумаге Kodak Polymax. VLP, повторно собранные в присутствии CyCpG, очищали через колонку с сефарозой 4B (1×50 см), элюируя буфером NET (1 мл/час). Фракции анализировали, используя анализ Оухтерлони, и объединяли фракции, содержащие VLP. В результате получали образец объемом 8 мл, который обессоливали посредством диализа против воды и сушили. Выход капсида составлял 10 мг. Анализ перерастворенного вещества в 0,6% агарозном геле, окрашенном бромидом этидия, показал, что капсиды не содержали нуклеиновых кислот. Образцы реакции повторной сборки, содержащие CyCpG, взятые после стадии повторной сборки и перед диализом, анализировали в 0,6% агарозном геле, окрашенном бромидом этидия и Кумасси синим. Могла быть получена полоса, мигрирующая на той же самой высоте, что и интактная VLP AP205, и окрашиваемая как бромидом этидия, так и Кумасси синим, что свидетельствует о том, что повторно были собраны VLP AP205, содержащие олигодезоксинуклеотид. Стадии диализа в большом объеме, которые следовали после процедуры повторной сборки, вероятно, привели к диффузии олигодезоксинуклеотида наружу из VLP. Существенно, что VLP AP205 также могли подвергаться повторной сборке в отсутствие выявляемого олигодезоксинуклеотида, который измеряли электрофорезом в агарозном геле с использованием окрашивания бромидом этидия. Таким образом, олигодезоксинуклеотиды могли быть успешно связаны с VLP AP205 после исходной разборки VLP, очистки подвергнутого разборке белка оболочки от нуклеиновых кислот и последующей повторной сборки VLP в присутствии олигодезоксинуклеотида.
B. Повторная сборка VLP AP205 с использованием подвергнутого разборке материала, который повторно не собирается в отсутствие добавленного олигонуклеотида.
Разборка: 100 мг очищенных и высушенных рекомбинантных VLP AP205 использовали для разборки, как описано выше. Все стадии осуществляли по существу, как описано для разборки в части A, но с использованием 8М мочевины для растворения осадков со стадий осаждения сульфатом аммония и пропуская стадию гель-фильтрации с использованием колонки CL-4B перед повторной сборкой. Объединенные фракции с колонки с сефадексом G-75 содержали 21 мг белка, что определяли посредством спектроскопии, используя формулу, описанную в части A. Отношение оптической плотности образца при 280 нм к оптической плотности при 260 нм составляло от 0,16 до 0,125. Образец для измерения разбавляли в 50 раз.
Повторная сборка: Препарат белка, полученный в результате стадии очистки гель-фильтрацией на сефадексе G-75, осаждали сульфатом аммония при насыщении 60% и полученный в результате осадок растворяли в 2 мл 7М мочевины, 10 мМ DTT. Образец разбавляли 8 мл 10% 2-меркаптоэтанола в буфере NET и диализовали в течение 1 часа против 40 мл 10% 2-меркаптоэтанола в буфере NET. Повторную сборку инициировали добавлением 0,4 мл раствора CyCpG (109 нмоль/мл) к образцу белка в диализном мешке. Диализ обеспечивали в непрерывном режиме и буфер NET использовали в качестве элюирующего буфера. Диализ проводили в течение двух дней и после завершения указанной стадии диализа брали образец для анализа ЭМ. Диализованный раствор для повторной сборки затем диализовали против 50% об./об. глицерина в буфере NET, чтобы достичь концентрирования. Осуществляли одну замену буфера через один день диализа. Диализ проводили всего в течение трех дней.
Диализованный и концентрированный раствора для повторной сборки очищали гель-фильтрацией через колонку с сефарозой 4-B (1×60 см) со скоростью потока 1 мл/час в буфере NET. Фракции тестировали в анализе по Оухтерлони и фракции, содержащие капсиды, сушили, ресуспендировали в воде и повторно хроматографировали на колонке 4-B, уравновешенной в 20 мМ Hepes pH 7,6. Используя каждую из следующих трех формул:
1. (183×OD230 нм-75,8×OD260 нм)×объем (мл); 2. ((OD235 нм-OD280 нм)/2,51)×объем; 3. ((OD228,5 нм-OD234,5 нм)×0,37)×объем
определили количества белка в подвергнутых повторной сборке VLP, составляющие 6-26 мг.
Подвергнутые повторной сборке VLP AP205 анализировали ЭМ, как описано выше, электрофорезом в агарозном геле и SDS-ПААГ в невосстанавливающих условиях.
Анализ ЭМ подвергнутого разборке материала показывает, что обработка VLP AP205 хлоридом гуанидиния по существу разрушает сборку капсида VLP. Повторная сборка полученного после разборки материала с олигонуклеотидом давала капсиды, которые очищали и дополнительно обогащали гель-фильтрацией. Получали частицы двух размеров; на электронной микрофотографии видны частицы диаметром примерно 25 нм и более мелкие частицы. Не получали повторной сборки в отсутствие олигонуклеотидов. Нанесение повторно собранных частиц на агарозный гель для электрофореза показало, что повторно собранные частицы содержали нуклеиновые кислоты. Экстракция содержащейся нуклеиновой кислоты фенольной экстракцией и последующее нанесение на агарозный гель, окрашиваемый бромидом этидия, выявили частицы, содержащие олигонуклеотид, используемый для повторной сборки. Идентичность упакованного олигонуклеотида контролировали нанесением образца данного олигонуклеотида рядом с материалом нуклеиновой кислоты, экстрагированной из частиц. Агарозный гель, на который наносили повторно собранные VLP AP205 и ранее красили бромидом этидия, затем красили Кумасси синим, выявляя совместную миграцию олигонуклеотидного содержимого с белковым содержимым частиц, что показывает, что олигонуклеотид был упакован в частицы. На гель наносили необработанные VLP AP205, 3 образца с разным количеством VLP AP205, повторно собранные с CyCpG и очищенные, и необработанные VLP Qβ.
Нанесение повторно собранных VLP AP205 на SDS-ПААГ-гель, разгонка в отсутствие восстанавливающего агента показали, что повторно собранные частицы образовали дисульфидные мостики, как в случае необработанных VLP AP205. Кроме того, комбинация дисульфидных мостиков идентична таковой в случае необработанных частиц. На SDS-гель наносили образцы: белковый маркер, необработанный wt Qβ, повторно собранный wt Qβ, необработанные VLP AP205, повторно собранные VLP AP205. Молекулярная масса субъединицы VLP AP205 составляет 14,0 кД, тогда как молекулярная масса субъединицы Qβ составляет 14,3 кД (обе молекулярные массы рассчитаны с N-концевым метионином).
C. Связывание эпитопа p33 (последовательность: H2N-KAVYNFATMGGC-COOH со свободными N- и C-концами (SEQ ID NO: 82) с VLP AP205, повторно собранными с CyCpG
Повторно собранные VLP AP205, полученные, как описано в части B и в 20 мМ Hepes, 150 мМ NaCl, pH 7,4, в концентрации 1,4 мг/мл подвергали взаимодействию с 5-кратным избытком перекрестно-сшивающего линкера SMPH, разбавленного из 50 мМ исходного раствора в ДМСО, в течение 30 минут при 15°C. Полученные так называемые дериватизованные VLP AP205 диализовали 2×2 часа против по меньшей мере 1000-кратного объема буфера 20 мМ Hepes, 150 мМ NaCl, pH 7,4. Дериватизованные AP205 подвергали взаимодействию в концентрации 1 мг/мл либо с 2,5-кратным, либо с 5-кратным избытком пептида, разбавленного из 20 мМ исходного раствора в ДМСО, в течение 2 часов при 15°C. Затем образец мгновенно замораживали в жидком азоте для хранения.
Реакцию связывания анализировали в SDS-ПААГ. На гель наносили следующие образцы: белковый маркер; дериватизованные VLP AP205 (d); VLP AP205, связанные с 2,5-кратным избытком пептида, надосадок (s); VLP AP205, связанные с 2,5-кратным избытком пептида, осадок (p); VLP AP205 VLP, связанные с 5-кратным избытком пептида, надосадок (s); VLP AP205, связанные с 5-кратным избытком пептида, осадок (p). Результат реакции связывания показал, что более высокую степень связывания можно достичь, используя в реакции связывания 5-кратный избыток пептида, а не 2,5-кратный избыток пептида.
ПРИМЕР 11
Неферментативный гидролиз РНК-содержимого VLP и упаковка иммуностимулирующих нуклеиновых кислот.
Зависимая от ZnSO4 деградация содержимого VLP в виде нуклеиновой кислоты:
5 мг VLP Qβ (определяемых анализом по Бредфорду) в 20 мМ HEPES, pH 7,4, 150 мМ NaCl диализовали либо против 2000 мл 50 мМ трис-HCl pH 8,0, 50 мМ NaCl, 5% глицерина, 10 мМ MgCl2, либо 2000 мл 4 мМ HEPES, pH 7,4, 30 мМ NaCl в течение 2 час при 4°C в складчатых диализных трубках SnakeSkinTM (Pierce, No. в каталоге 68035). Каждый из буферов для диализа меняли один раз и диализ продолжали еще в течение 16 час при 4°C. Диализованный раствор осветляли в течение 10 минут при 14000 об/мин (Eppendorf 5417 R, в угловом роторе F45-30-11, используемом на всех последующих стадиях) и снова определяли концентрацию белка анализом по Бредфорду. VLP Qβ в 50 мМ трис-HCl pH 8,0, 50 мм NaCl, 5% глицерине, 10 мМ MgCl2 разбавляли соответствующим буфером до конечной концентрации белка 1 мг/мл, тогда как VLP Qβ в 4 мМ HEPES pH 7,4, 30 мМ NaCl разбавляли соответствующим буфером до конечной концентрации белка 0,5 мг/мл. Указанные содержащие капсиды растворы снова центрифугировали в течение 10 минут при 14 000 об/мин при 4°C. Затем надосадки инкубировали с ZnSO4, который добавляли до конечной концентрации 2,5 мМ, в течение 24 час при 60°C в термомиксере Eppendorf при 550 об/мин. Через 24 часа растворы осветляли в течение 10 минут при 14000 об/мин и осадок отбрасывали. Эффективность ZnSO4-зависимой деградации нуклеиновых кислот подтверждали электрофорезом в агарозном геле. Надосадки диализовали против 5000 мл 4 мМ HEPES pH 7,4, 30 мМ NaCl в течение 2 час при 4°C. 5000 мл буфера заменяли один раз и диализ продолжали в течение ночи при 4°C. Диализованный раствор осветляли в течение 10 минут при 14 000 об/мин и 4°C, небольшой осадок отбрасывали и концентрацию белка в надосадках определяли анализом по Бредфорду. Сходные результаты получали в случае обработки капсидов хлоридом меди/фенантролином/пероксидом водорода. Специалистам в данной области известны альтернативные неферментативные способы гидролиза РНК.
ZnSO4-обработанные VLP Qβ анализировали электрофорезом в агарозном геле: VLP Qβ , которые были очищены из E. coli и диализованы либо против буфера 1 (50 мМ трис-HCl pH 8,0, 50 мМ NaCl, 5% глицерин, 10 мМ MgCl2), либо против буфера 2 (4 мМ HEPES, pH 7,4, 30 мМ NaCl), инкубировали либо в отсутствие, либо в присутствии 2,5 мМ сульфата цинка (ZnSO4) в течение 24 час при 60°C. После указанной обработки равные количества указанных образцов (5 мкг белка) смешивали с красителем для нанесения и наносили на 0,8% агарозный гель. После разгонки гель красили бромидом этидия. Обработка VLP с использованием ZnSO4 вызывала деградацию содержимого в виде нуклеиновой кислоты, при этом ложно-обработанные контроли не подвергались влиянию.
Упаковка олигодезоксинуклеотидов в ZnSO4-обработанные VLP:
ZnSO4-обработанные и диализованные Qβ капсиды с концентрацией белка (которую определяли анализом по Бредфорду) от 0,4 мг/мл до 0,9 мг/мл (которая соответствует концентрации капсидов 159 нМ и 357,5 нМ соответственно) использовали для упаковки олигодезоксинуклеотидов. Олигодезоксинуклеотиды добавляли в 300-кратном молярном избытке к капсидам Qβ-VLP капсиды и инкубировали в течение 3 час при 37°C в термомиксере Eppendorf comfort при 550 об/мин. Через 3 час реакционную смесь центрифугировали в течение 10 минут при 14000 об/мин и 4°C. Надосадки диализовали в диализаторе Spectra/Por®CE DispoDialyzer с пределом отсечения по молекулярной массе 300000 (Spectrum, No. в каталоге 135 526) против 5000 мл 20 мМ HEPES pH 7,4, 150 мМ NaCl в течение 8 час при 4°C. 5000 мл буфера заменяли один раз и диализ продолжали в течение ночи при 4°C. Концентрацию белка в диализованных образцах определяли анализом по Бредфорду. Капсиды Qβ и содержание в них нуклеиновой кислоты анализировали как описано в примерах 7 и 9.
Упаковку олигодезоксинуклеотидов в ZnSO4-обработанные VLP анализировали электрофорезом в агарозном геле. VLP Qβ, которые были обработаны 2,5 мМ сульфатом цинка (+ ZnSO4), диализовали против 4 мМ HEPES, pH 7,4, 30 мМ NaCl и инкубировали в течение 3 часов при 37°C с избытком олигодезоксинуклеотидов (в результате диализа концентрация ZnSO4 уменьшалась на 106порядков, поэтому он указан только в скобках). После такой инкубации в присутствии олигодезоксинуклеотидов равные количества указанных образцов (5 мкг белка) смешивали с красителем для нанесения и наносили на 0,8% агарозный гель.
После разгонки гель красили бромидом этидия. Добавление олигодезоксинуклеотидов к ZnSO4-обработанным VLP Qβ могло восстанавливать электрофоретическое поведение обработанных таким образом капсидов по сравнению с необработанными капсидами Qβ, которые были очищены из E. coli.
Содержание нуклеиновой кислоты в обработанных ZnSO4и олигодезоксинуклеотидами VLP Qβ анализировали посредством расщепления бензоназой и протеиназой K и электрофорезом в полиакриламидном геле с TBE/мочевиной: олигодезоксинуклеотиды упаковывали в ZnSO4-обработанные VLP Qβ, как описано выше. 25 мкг указанных VLP расщепляли 25 мкл бензоназы (Merck, No. в каталоге 1.01694.0001) согласно инструкциям производителя. После тепловой инактивации нуклеазы (30 минут при 80°C) VLP обрабатывали протеиназой K (конечная концентрация фермента составляла 0,5 мг/мл) согласно инструкциям производителя. Через 3 час эквивалентные 2 мкг VLP Qβ, которые были расщеплены бензоназой и протеиназой K, смешивали с буфером для образцов, содержащих TBE-мочевину, и наносили на 15% полиакриламидный гель с TBE-мочевиной (Novex®, Invitrogen No. в каталоге EC6885). Капсиды, нанесенные на дорожку 2, обрабатывали 2,5 мМ ZnSO4 в присутствии буфера 1 (смотри выше), тогда как капсиды, нанесенные на дорожку 3, обрабатывали 2,5 мМ ZnSO4 в присутствии буфера 2 (смотри выше). Для качественного, а также количественного стандарта 1 пмоль, 5 пмоль и 10 пмоль олигодезоксинуклеотида, который использовали для реакции повторной сборки, наносили на тот же самый гель (дорожки 4-6). В качестве контроля капсиды Qβ, которые были очищены из E. coli, обрабатывали точно так же и анализировали на том же самом полиакриламидном геле с TBE-мочевиной (дорожка 1). После завершения разгонки гель фиксировали, уравновешивали до нейтрального pH и красили SYBR-Gold (Molecular Probes No. в каталоге S-11494). Интактные VLP Qβ (которые были выделены из E. coli) не содержали нуклеиновых кислот размера, сходного с размером нуклеиновых кислот, которые были экстрагированы из обработанных ZnSO4 и олигодезоксинуклеотидами капсидов Qβ. Кроме того, нуклеиновые кислоты, выделенные из последних VLP, мигрировали совместно с олигодезоксинуклеотидами, которые использовали в реакции повторной сборки. Полученные результате подтвердили, что используемые олигодезоксинуклеотиды были упакованы в ZnSO4-обработанные капсиды Qβ.
ПРИМЕР 12
Связывание антигенных пептидов после упаковки иммуностимулирующих нуклеиновых кислот в VLP.
Опосредованная РНКазой A и ZnSO4 деградация содержимого VLP в виде нуклеиновых кислот.
VLP Qβ обрабатывали РНКазой A, как описано в примере 9 в условиях низкой ионной силы (20 мм Hepes pH 7,4 или 4 мМ Hepes, 30 мМ NaCl, pH 7,4). Подобным образом обрабатывали другие VLP, такие как VLP, описанные в Примерах 2, 3, 7 и 10, т.е. GA, BKV, HBcAg и AP205. Альтернативно VLP Qβ и VLP AP205 обрабатывали ZnSO4 в условиях низкой ионной силы (20 мМ Hepes pH 7,4 или 4 мМ Hepes, 30 мМ NaCl pH 7,4), как описано в примере 11. VLP AP205 (1 мг/мл) либо в 20 мМ Hepes pH 7,4, либо в 20 мМ Hepes, 1 мМ трис, pH 7,4, обрабатывали в течение 48 час 2,5 мМ ZnSO4 при 50°C в термомиксере Eppendorf comfort при 550 об/мин. Образцы VLP Qβ и AP205 осветляли как описано в примере 11 и надосадки диализовали в диализной трубке с пределом отсечения по молекулярной массе 10000 Spectra/Por® (Spectrum, No. в каталоге 128 118) сначала против 2 л 20 мМ Hepes, pH 7,4, в течение 2 час при 4°C и после замены буфера в течение ночи. Образцы осветляли после диализа, как описано в примере 11, и концентрацию белка в надосадках определяли анализом по Бредфорду.
Упаковка ISS в обработанные РНКазой A и ZnSO4 VLP.
После гидролиза РНК и диализа VLP Qβ и AP205 (1-1,5 мг/мл) смешивали с 130 мкл CpG-олигонуклеотидов (NKCpG, G10-PO - сравни таблица 2; G3-6, G8-8 - сравни таблица 3; 1 мМ исходный раствор олигонуклеотида в 10 мМ трис pH 8) на мл VLP. Образцы инкубировали в течение 3 час при 37°C в термовстряхивателе при 650 об/мин. Затем образцы обрабатывали 125 ед. бензоназы/мл VLP (Merck KGaA, Darmstadt, Germany) в присутствии 2 мМ MgCl2 и инкубировали в течение 3 час при 37°C перед диализом. Образцы диализовали в диализной трубке с пределом отсечения по молекулярной массе 300000 Spectra/Por® (Spectrum, No. в каталоге 131 447) против 20 мМ Hepes, pH 7,4 в течение 2 час при 4°C и после смены буфера в течение ночи против такого же буфера. После диализа образцы осветляли, как описано в примере 11, и концентрацию белка в надосадках определяли анализом по Бредфорду.
Связывание иммуногенных пептидов с упакованными с ISS VLP.
VLP Qβ, упакованные с ISS, связывали с пептидами p33, содержащими C-концевое удлинение GGC (KAVYNFATM-GGC) (SEQ ID NO: 82), получая в результате VLP Qβ, названные VLP Qb-ISS-33. Упакованные VLP Qb в 20 мМ Hepes, pH 7,4, подвергали дериватизации с 10-кратным молярным избытком SMPH (Pierce) в течение 0,5 час при 25°C с последующими двумя стадиями диализа по 2 часа каждая против 20 мМ HEPES pH 7,4 при 4°C, чтобы удалить непрореагировавший SMPH. Пептиды добавляли в 5-кратном молярном избытке к диализованной смеси для дериватизации и давали возможность взаимодействовать в течение 2 час в термомиксере при 25°C. Образцы диализовали в диализной трубке с пределом отсечения по молекулярной массе 300000 Spectra/Por® против 20 мМ Hepes pH 7,4 в течение 2 час при 4°C и после замены буфера в течение ночи против такого же буфера. После диализа образцы осветляли, как описано в примере 11, и концентрацию белка в надосадках определяли анализом по Бредфорду. Связывание пептида p33 с Qβ анализировали в SDS-ПААГ в 16% ПААГ-гелях с трис-глицином (Novex® Invitrogen, No. в каталоге EC64952), используя буфер для образцов, содержащий 2% SDS и β-меркаптоэтанол или DTT. Упаковку анализировали в 1% агарозных гелях и после расщепления протеиназой K в гелях с TBE/мочевиной, как описано в примере 7.
VLP AP205 (1,24 мг/мл) упакованные с олигонуклеотидом G8-8, как описано выше, подвергали дериватизации и связывали с Melan-A 16-35 A/L, содержащим N-концевое удлинение C (c GHGHSYTTAE ELAGIGILTV) (SEQ ID NO: 55), получая в результате VLP AP205-G8-8-Melan-A. VLP AP205 (упакованные с G8-8) в 20 мМ Hepes pH 7,4, подвергали дериватизации с 20-кратным молярным избытком SMPH в течение 0,5 час при 25°C и затем диализовали два раза против 20 мМ HEPES, pH 7,4 при 4°C, чтобы удалить непрореагировавший SMPH. Пептид добавляли к диализованной смеси для дериватизации в 10-кратном молярном избытке и давали возможность взаимодействовать в течение 2 час в термомиксере при 25°C. Образцы диализовали в диализной трубке с пределом отсечения по молекулярной массе 10000 против 20 мМ Hepes pH 7,4 в течение 2 час при 4°C и после замены буфера в течение ночи против такого же буфера. После диализа образцы осветляли, как описано в примере 11, и концентрацию белка в надосадках определяли анализом по Бредфорду. Эффективность связывания пептида Melan-A 16-35A/L с AP205 анализировали в SDS-ПААГ в 16% ПААГ-гелях с трис-глицином. Упаковку олигонуклеотида G8-8 в AP205 анализировали в 1% агарозных гелях и после расщепления протеиназой K количество олигонуклеотида G8-8 в AP205-G8-8-Melan-A 16-35 A/L анализировали в гелях с TBE/мочевиной, как описано в примере 7.
Упаковку обработанных РНКазой A и ZnSO4 VLP Qβ с NKCpG до, а также после связывания с пептидом p33 анализировали электрофорезом в агарозном геле. VLP Qβ, содержащие олигонуклеотиды NKCpG и затем связанные с пептидом p33, названы VLP Qb-NKCpG-33. В 1% агарозном геле флуоресцирующая полоса, видимая на окрашенном бромидом этидия геле, мигрирует совместно с полосой белка, видимой на окрашенном Кумасси синим геле, свидетельствуя об упаковке. Таким образом, при упаковке VLP Qβ, обработанные как РНКазой A, так и ZnSO4, содержат олигонуклеотиды NKCpG как перед, так и после связывания с пептидом p33. Эффективность связывания пептида p33 сохраняется, о чем можно судить на основании множества продуктов связывания, видимых после анализа в SDS-ПААГ в 16% ПААГ-геле с трис-глицином в виде полос, мигрирующих медленнее, чем остальные мономерные субъединицы VLP Qβ, которые не взаимодействовали с пептидом. Эффективность упаковки можно оценить на основе анализа геля с TBE/мочевиной посредством сравнения сигнала олигонуклеотида на дорожке с упакованными Qb-NKCpG-33 с сигналом олигонуклеотидного стандарта, нанесенного на тот же самый гель. Количества упакованного NKCPG составляли от 1 до 4 нмоль/100 мкг VLP Qb-NKCpG-33.
Упаковку олигонуклеотидов G8-8 в VLP Qβ и последующее связывание с пептидом p33 анализировали электрофорезом в агарозном геле. VLP Qβ, содержащие олигонуклеотиды G8-8 и затем связанные с пептидом p33, названы VLP Qb-G8-8-33. Окрашивание бромидом этидия упакованных с G8-8 VLP Qβ можно видеть на 1% агарозном геле, окрашенном бромидом этидия. Комиграция флуоресцирующей окрашенной бромидом этидия полосы с полосой белка VLP Qβ, видимой на том же геле, затем окрашенном Кумасси синим, свидетельствует об упаковке. Эффективность связывания можно оценить анализом в SDS-ПААГ в 16% ПААГ-геле с трис-глицином, она составляет 30%. Анализ содержания G8-8 в VLP Qb-G8-8-33 осуществляли в 1% агарозном геле, при этом количество упакованного олигонуклеотида составляло примерно 1 нмоль/100 мкг VLP Qb-G8-8-33.
Упаковку олигонуклеотидов G8-8 в VLP AP205 анализировали электрофорезом в агарозном геле. Окрашивание упакованных с G8-8 VLP AP205 можно видеть на 1% агарозном геле, окрашенном бромидом этидия. Комиграция полосы белка VLP AP205, выявляемой на том же геле, затем окрашенном Кумасси синим, свидетельствует об упаковке. Эффективность связывания с пептидом Melan-A 16-35 A/L можно оценить анализом в SDS-ПААГ в 16% ПААГ-геле с трис-глицином, на котором можно видеть множественные полосы связывания, мигрирующие медленнее, чем остальные мономерные субъединицы VLP AP205, которые не взаимодействовали с пептидом. Эффективность связывания сравнивают с эффективностью связывания, полученной для VLP Qb-G8-8-33. Анализ содержания олигонуклеотида G8-8 в VLP AP205 после связывания с Melan-A 16-35 A/L можно видеть при электрофорезе в геле с TBE/мочевиной.
ПРИМЕР 13
Упаковка иммуностимулирующих фланкированных гуанозином олигонуклеотидов в VLP.
VLP Qbx33 (VLP Qβ, связанные с пептидом p33, смотри пример 9) обрабатывали РНКазой A в условиях низкой ионной силы (20 мМ Hepes pH 7,4), как описано в примере 9, чтобы гидролизовать РНК-содержимое VLP Qbx33. После диализа против 20 мМ Hepes pH 7,4, VLP Qbx33 смешивали с фланкированными гуанозином олигонуклеотидами (таблица 2: G10-PO, таблица 3: G3-6, G7-7, G8-8, G9-9 или G6, из исходного 1 мМ раствора олигонуклеотидов в 10 мМ трис pH 8) и инкубировали, как описано в примере 12. Затем VLP Qbx33 обрабатывали бензоназой и диализовали в трубке с пределом отсечения по молекулярной массе 300000. Образцы с олигонуклеотидами G7- 7, G8-8 и G9-9 диализовали против большого объема в течение 3 дней с 4 заменами буфера, чтобы удалить свободные олигонуклеотиды. Упаковку анализировали в 1% агарозных гелях и после расщепления протеиназой K в гелях с TBE/мочевиной, как описано в примере 7.
Упаковку олигонуклеотидов G3-6, G6, G8-8 в обработанные РНКазой A VLP Qbx33 анализировали электрофорезом в агарозном геле. При упаковке олигонуклеотида становится видна флуоресцирующая полоса, мигрирующая медленнее, чем эталонные необработанные VLP Qβ, в 1% агарозном геле, окрашенном бромидом этидия, что свидетельствует о присутствии олигонуклеотидов. Сигнал сохраняется после обработки бензоназой, то указывает на упаковку олигонуклеотидов в VLP Qbx33. Эффективность упаковки можно оценить на основе электрофореза в геле с TBE/мочевиной. Количество упакованного олигонуклеотида G3-6 (примерно 4 нмоль/100 мкг VLP Qbx33) намного выше, чем количество упакованного олигонуклеотида G8-8 (примерно 1 нмоль/100 мкг VLP Qbx33). Это свидетельствует о зависимости способности к упаковке от длины хвоста из гуанозиновых нуклеотидов, фланкирующего мотив CpG.
ПРИМЕР 14
Упаковка рибонуклеиновой кислоты в VLP.
Зависимая от ZnSO4 деградация содержимого VLP в виде нуклеиновой кислоты.
VLP Qβ обрабатывали ZnSO4 в условиях низкой ионной силы (20 мМ Hepes pH 7,4 или 4 мМ Hepes, 30 мМ NaCl, pH 7,4), как описано в примере 11. VLP AP205 (1 мг/мл) либо в 20 мМ Hepes pH 7,4, либо в 20 мМ Hepes, 1 мМ трис, pH 7,4, обрабатывали в течение 48 час 2,5 мМ ZnSO4 при 50°C в термомиксере Eppendorf comfort при 550 об/мин. Образцы VLP Qβ и AP205 осветляли, как в примере 11, и диализовали против 20 мМ Hepes, pH 7,4 как в примере 12.
Упаковка поли(I:C) в ZnSO4-обработанные VLP:
Иммуностимулирующую рибонуклеиновую кислоту поли(I:C), (No. в каталоге 27-4732-01, поли(I)·поли(C), Pharmacia Biotech) растворяли в PBS (Invitrogen No. в каталоге 14040) или воде до концентрации 4 мг/мл (9 мкМ). Поли(I:C) инкубировали в течение 10 минут при 60°C и затем охлаждали до 37°C. Инкубированную поли(I:C) добавляли в 10-кратном молярном избытке к ZnSO4-обработанным либо VLP Qβ, либо VLP P205 (1-1,5 мг/мл) и смеси инкубировали в течение 3 час при 37°C в термомиксере при 650 об/мин. Затем избыток свободной поли(I:C) ферментативно гидролизовали посредством инкубации с 125 ед. бензоназы на мл смеси VLP в присутствии 2 мМ MgCl2 в течение 3 час при 37°C в термомиксере при 300 об/мин. После гидролиза бензоназой образцы осветляли, как описано в примере 11, и надосадки диализовали в диализной трубке с пределом отсечения по молекулярной массе 300000 Spectra/Por® (Spectrum, No. в каталоге 131 447) против 2 л 20 мМ Hepes, pH 7,4 в течение 2 час при 4°C и после замены буфера в течение ночи против такого же буфера. После диализа образцы осветляли, как описано в примере 11, и концентрацию белка в надосадках определяли анализом по Бредфорду.
Связывание иммуногенных пептидов с VLP, упакованными с поли(I:C).
VLP Qβ (1 мг/мл), упакованные с поли(I:C), подвергали дериватизации и связывали либо с пептидом p33 (KAVYNFATM-GGC) (SEQ ID NO: 82), как описано в примере 12, либо с пептидом Melan-A (Melan-A 16-35A/L CGHGHSYTTAEELAGIGILTV) (SEQ ID NO: 55), получая в результате VLP Qb-pIC-33 и Qb-pIC-Melan-A соответственно. Для связывания с пептидом Melan-A упакованные VLP Qβ подвергали дериватизации 2,1-кратным молярным избытком SMPH (Pierce) в течение 0,5 час при 25°C с последующими двумя стадиями диализа против 20 мМ HEPES, pH 7,4 при 4°C, чтобы удалить непрореагировавший SMPH. Пептиды добавляли в 2,1-кратном молярном избытке и давали возможность взаимодействовать в течение 1,5 час в термомиксере при 25°C. Образцы диализовали в диализаторе с пределом отсечения по молекулярной массе 300000 Spectra/Por® CE Dispo против 20 мМ Hepes, pH 7,2, в течение 3 час при 4°C и после замены буфера в течение ночи против такого же буфера. После диализа образцы осветляли, как описано в примере 11, и концентрацию белка в надосадках определяли анализом по Бредфорду. Связывание пептида p33 и пептида Melan-A с Qβ анализировали в SDS-ПААГ в 16% ПААГ-гелях с трис-глицином. Упаковку анализировали в 1% агарозных гелях и после расщепления протеиназой K в гелях с TBE/мочевиной как описано в примере 7.
Упаковку поли(I:C) в ZnSO4-обработанные VLP Qβ и связывание с пептидом Melan-A, приводящие к VLP Qb-pIC-Melan-A, анализировали электрофорезом в агарозном геле. Флуоресцентный сигнал, видимый на окрашенном бромидом этидия 1% агарозном геле, свидетельствующий о присутствии нуклеиновой кислоты, мигрирует совместно с полосой белка, которая становилась видимой после окрашивания геля Кумасси синим, что свидетельствует об упаковке. Эффективность связывания пептида Melan-A оценивали анализом в SDS-ПААГ в 16% ПААГ-геле с трис-глицином. Множественные продукты связывания видны в виде полос, мигрирующих медленнее, чем мономерные субъединицы VLP Qβ, которые не взаимодействовали с пептидом. Эффективность связывания Melan-A в общем сравнима с эффективностью связывания, полученной для VLP Qb-G8-8-33 (пример 12), хотя и несколько ниже. Эффективность упаковки в Qb-pIC-Melan-A можно было оценить, используя гель с TBE/мочевиной; количество упакованной поли(I:C) в Qβ составляло примерно 25 пмоль и оставалось таким же при связывании Melan-A.
VLP AP205 (1 мг/мл), упакованные с поли(I:C), подвергали дериватизации и связывали с Melan-A 16-35 A/L, содержащим на N-конце удлинение C (cGHGHSYTTAEELAGIGILTV) (SEQ ID NO: 55), получая в результате VLP AP205-G8-8-Melan-A. VLP AP205 в 20 мМ HEPES, pH 7,4, подвергали дериватизации 20-кратным молярным избытком SMPH в течение 0,5 час при 25°C и затем диализовали два раза против 20 мМ HEPES pH 7,4 при 4°C, чтобы удалить непрореагировавший SMPH. Пептид добавляли к диализованной смеси для дериватизации в 10-кратном молярном избытке и давали возможность взаимодействовать в течение 2 час в термомиксере при 25°C. Образцы диализовали в диализной трубке с пределом отсечения по молекулярной массе 100000 против 20 мМ Hepes, pH 7,4, в течение 2 час при 4°C и после замены буфера в течение ночи против такого же буфера. После диализа образцы осветляли, как описано в примере 11, и концентрацию белка в надосадках определяли анализом по Бредфорду. Эффективность связывания пептида Melan-A 16-53 A/L с AP205 анализировали в SDS-ПААГ в 16% ПААГ-гелях с трис-глицином. Упаковку поли(I:C) анализировали в 1% агарозных гелях и после расщепления протеиназой K в гелях с TBE/мочевиной, как описано в примере 7.
Упаковку поли(I:C) в ZnSO4-обработанные VLP AP205 и продукт связывания AP205-pIC-Melan-A после связывания с Melan-A анализировали электрофорезом в агарозном геле. Эффективность связывания пептида Melan-A оценивали по появлению множества продуктов связывания, видимых в виде полос, мигрирующих медленнее, чем мономерная субъединица VLP AP205, которая не взаимодействовала с пептидом, после анализа в SDS-ПААГ при электрофорезе в 16% ПААГ-геле с трис-глицином. Эффективность упаковки можно оценить, используя TBE-гель.
ПРИМЕР 15
Упаковка иммуностимулирующих фланкированных гуанозином олигонуклеотидов в VLP HBcAg.
VLP HBcAg обрабатывали РНКазой A в условиях низкой ионной силы (20 мМ Hepes pH 7,4), как описано в примере 9, чтобы гидролизовать РНК-содержимое VLP. После диализа против 20 мМ Hepes pH 7,4, VLP смешивали с фланкированными гуанозином олигонуклеотидами (таблица 3: G3-6, G7-7, G8-8, G9-9, G10-PO или G6, исходный 1 мМ раствор в 10 мМ трис pH 8) и инкубировали, как описано в примере 12. Затем VLP обрабатывали бензоназой и диализовали в трубке с пределом отсечения по молекулярной массе 300000. Упаковку анализировали в 1% агарозных гелях и после расщепления протеиназой K в гелях с TBE/мочевиной, как описано в примере 7.
ПРИМЕР 16
Упаковка иммуностимулирующих фланкированных гуанозином олигонуклеотидов в VLP GA.
VLP GA обрабатывали РНКазой A в условиях низкой ионной силы (20 мМ Hepes pH 7,4), как описано в примере 9, чтобы гидролизовать РНК-содержимое VLP. После диализа против 20 мМ Hepes pH 7,4, VLP смешивали с фланкированными гуанозином олигонуклеотидами (таблица 3: G3-6, G7-7, G8-8, G9-9, G10-PO или G6, исходный 1 мМ раствор в 10 мМ трис pH 8) и инкубировали, как описано в примере 12. Затем VLP обрабатывали бензоназой и диализовали в трубке с пределом отсечения по молекулярной массе 300000. Упаковку анализировали в 1% агарозных гелях и после расщепления протеиназой K в гелях с TBE/мочевиной, как описано в примере 7.
ПРИМЕР 17
Упаковка рибонуклеиновой кислоты в VLP HBcAg.
VLP HBcAg обрабатывали ZnSO4 в условиях низкой ионной силы (20 мМ Hepes pH 7,4 или 4 мМ Hepes, 30 мМ NaCl, pH 7,4), как описано в примере 11, и диализовали против 20 мМ Hepes pH 7,4, как в примере 12. Поли(I:C) добавляли в 10-кратном молярном избытке к VLP HBcAg (1-1,5 мг/мл) и инкубировали в течение 3 час при 37°C в термомиксере при 650 об/мин как описано в примере 14. Затем избыток свободной поли(I:C) ферментативно гидролизовали посредством инкубации с 125 ед. бензоназы на мл смеси VLP в присутствии 2 мМ MgCl2 в течение 3 час при 37°C в термомиксере при 300 об/мин. Образцы осветляли после гидролиза бензоназой, как описано в примере 11, и диализовали, как в примере 14. После диализа образцы осветляли, как описано в примере 11, и концентрацию белка в надосадках определяли анализом по Бредфорду. VLP HBcAg (1 мг/мл), упакованные с поли(I:C) дериватизировали и связывали с Melan-A и диализовали, как в примере 14.
ПРИМЕР 18
Упаковка рибонуклеиновой кислоты в VLP GA.
VLP GA обрабатывали ZnSO4 в условиях низкой ионной силы (20 мМ Hepes pH 7,4 или 4 мМ Hepes, 30 мМ NaCl, pH 7,4), как описано в примере 11, и диализовали против 20 мМ Hepes pH 7,4 как в примере 12. Поли(I:C) добавляли в 10-кратном молярном избытке к VLP GA (1-1,5 мг/мл) и инкубировали в течение 3 час при 37°C в термомиксере при 650 об/мин, как описано в примере 14. Затем избыток свободной поли(I:C) ферментативно гидролизовали посредством инкубации с 125 ед. бензоназы на мл смеси VLP в присутствии 2 мМ MgCl2 в течение 3 час при 37°C в термомиксере при 300 об/мин. Образцы осветляли после гидролиза бензоназой как описано в примере 11 и диализовали, как в примере 14. После диализа образцы осветляли, как описано в примере 11, и концентрацию белка в надосадках определяли анализом по Бредфорду. VLP GA (1 мг/мл), упакованные с поли(I:C), дериватизировали и связывали с Melan-A и диализовали, как в примере 14.
ПРИМЕР 19
Разборка Qβ, повторная сборка и упаковка олигодезоксинуклеотидов.
Разборка и повторная сборка VLP Qβ
Разборка: 45 мг VLP Qβ (2,5 мг/мл, как определяли анализом по Бредфорду) в PBS (20 мМ фосфат, 150 мМ NaCl, pH 7,5) восстанавливали 10 мМ DTT в течение 15 мин при комнатной температуре в условиях перемешивания. Вторая инкубация в течение 15 мин при комнатной температуре в условиях перемешивания следовала после добавления хлорида магния до конечной концентрации 700 мМ, что приводило к осаждению инкапсулированной РНК клетки-хозяина и сопутствующей дезинтеграции VLP. Раствор центрифугировали 10 мин при 4000 об/мин при 4°C (Eppendorf 5810 R, в угловом роторе A-4-62, используемом на всех следующих стадиях) для того, чтобы удалить выпавшую в осадок РНК из раствора. Надосадок, содержащий высвобожденный димерный белок оболочки Qβ, использовали для стадий хроматографической очистки.
Способ двухстадийной очистки белка оболочки Qβ катионообменной хроматографией и эксклюзионной хроматографией по размеру: Надосадок после реакции разборки, содержащий димерный белок оболочки, белки клетки-хозяина и остаточную РНК клетки-хозяина, наносили на колонку FF с SP-сефарозой (xk16/20, 6 мл, Amersham Bioscience). В ходе разгонки, которую осуществляли при комнатной температуре и скорости потока 5 мл/мин, контролировали оптическую плотность при 260 нм и 280 нм. Колонку уравновешивали 20 мМ натрий-фосфатным буфером pH 7 и образец разводили 1:15 в воде, чтобы скорректировать проводимость ниже 10 мС/см для того, чтобы достичь надлежащего связывания белка оболочки с колонкой. Элюирование связанного белка оболочки осуществляли ступенчатым градиентом до 20 мМ фосфата натрия/500 мМ хлорида натрия и белок собирали в объеме фракции, составляющей примерно 25 мл. Колонку регенерировали 0,5М NaOH.
На второй стадии выделенный димер белка оболочки Qβ (элюированная фракция с катионообменной колонки) наносили (двумя потоками) на колонку HR с сефакрилом S-100 (xk26/60, 320 мл, Amersham Bioscience), уравновешенную 20 мМ фосфатом натрия/250 мМ хлоридом натрия; pH 6,5. Хроматографию осуществляли при комнатной температуре и скорости потока 2,5 мл/мин. Оптическую плотность контролировали при 260 нм и 280 нм. Собирали фракции по 5 мл. Колонку регенерировали 0,5М NaOH.
Повторная сборка при диализе: Исходный раствор очищенного димерного белка оболочки Qβ с концентрацией 2 мг/мл использовали для повторной сборки VLP Qβ в присутствии олигодезоксинуклеотида G8-8 или G10-PO. Концентрация олигодезоксинуклеотида в смеси для повторной сборки составляла 10 мкМ. Концентрация димерного белка оболочки в смеси для повторной сборки составляла 40 мкМ (примерно 1,13 мг/мл). К раствору добавляли исходные растворы мочевины и DTT, чтобы получить конечные концентрации 1М мочевины и 5 мМ DTT соответственно. Олигодезоксинуклеотид добавляли в виде последнего компонента вместе с H2O, получая конечный объем реакционной смеси для повторной сборки 3 мл. Полученный раствор диализовали при 4°C в течение 72 час против 1500 мл буфера, содержащего 20 мМ трис-HCl, 150 мМ NaCl, pH 8,0. Диализованную смесь для повторной сборки центрифугировали при 14 000 об/мин в течение 10 минут при 4°C. Незначительный осадок отбрасывали, в то время как надосадок содержал повторно собранные и упакованные VLP. Повторно собранные и упакованные VLP концентрировали центрифугирующими фильтровальными устройствами (Millipore, UFV4BCC25, 5K NMWL) до конечной концентрации белка 3 мг/мл. Концентрацию белка определяли анализом по Бредфорду.
Очистка повторно собранных и упакованных VLP эксклюзионной хроматографией по размеру: До 10 мг общего белка наносили на колонку с сефарозой CL-4B (xk16/70, Amersham Biosciences), уравновешенную 20 мМ HEPES, 150 мМ NaCl, pH 7,4. Хроматографию осуществляли при комнатной температуре и скорости потока 0,4 мл/мин. Оптическую плотность контролировали при 260 нм и 280 нм. Наблюдали два пика, которые собирали во фракциях по 0,5 мл и анализировали в SDS-ПААГ. Картину дисульфидных связей в повторно собранных и очищенных капсидах Qβ анализировали в невосстанавливающем SDS-ПААГ. 5 мкг указанных капсидов смешивали с буфером для образцов (содержащем SDS), который не содержал восстанавливающего агента, и наносили на 16% гель с трис-глицином. После завершения разгонки гель красили Кумасси синим. При сравнении с «интактными» капсиды, очищенными из E. coli, в повторно собранных VLP Qβ наблюдали такую же картину дисульфидных связей с полосами, соответствующими димеру, тримеру, тетрамеру, пентамеру и гексамеру белка оболочки Qb. Калибровка колонки с использованием интактных и высокоочищенных капсидов Qβ из E. coli показала, что кажущаяся молекулярная масса основного первого пика соответствует капсидам Qβ.
Повторная сборка при диафильтрации (оптимизированный способ): 20 мл исходного раствора очищенного белка оболочки (1,5 мг/мл) смешивали с исходным растворами мочевины, DTT, олигодезоксинуклеотида G10-PO и водой. Олигодезоксинуклеотид добавляли в последнюю очередь. Объем смеси составлял 30 мл и конечные концентрации компонентов составляли 35 мкМ димерного белка оболочки (соответствующие 1 мг/мл), 35 мкМ олигодезоксинуклеотида, 1М мочевины и 2,5 мМ DTT. Затем смесь подвергали диафильтрации против 300 мл 20 мМ фосфата натрия/250 мМ хлорида натрия, pH 7,2, в устройстве для тангенциальной фильтрации в проточном режиме при комнатной температуре, используя мембранный картридж Pellicon XL (Biomax 5K, Millipore). Общую скорость потока устанавливали 10 мл/мин и скорость прохождения фильтрата устанавливали 2,5 мл/мин. После завершения стадии диафильтрации к раствору добавляли H2O2 до конечной концентрации 7 мМ и раствор затем инкубировали при комнатной температуре в течение 60 мин, чтобы ускорить образование структурных дисульфидных связей в образованных VLP. Удаление невключенного олигодезоксинуклеотида и белка оболочки осуществляли при 2-ой диафильтрации против 600 мл 20 мМ фосфата натрия/250 мМ хлорида натрия, pH 7,2, используя мембранный картридж Pellicon XL (PLCMK 300K, Millipore).
Анализы VLP Qβ, которые были повторно собраны в присутствии олигодезоксинуклеотидов:
A) Гидродинамический размер повторно собранных капсидов: капсиды Qβ, которые были повторно собраны в присутствии олигодезоксинуклеотида G8-8, анализировали динамическим светорассеянием (DLS) и сравнивали с интактными VLP Qβ, которые были очищены из E. coli. Повторно собранные капсиды проявляли такой же гидродинамический размер (который зависит как от массы, так и от конформации), как и интактные VLP Qβ.
B) Образование дисульфидных связей в повторно собранных капсидах: Повторно собранные VLP Qβ анализировали в невосстанавливающем SDS-ПААГ и сравнивали с интактными VLP Qβ, которые были очищены из E. coli. В случае повторно собранных капсидов наблюдали картину полос с присутствием дисульфидно-связанных пентамерных и гексамерных форм белка оболочки, сходную с интактными VLP Qβ (которые описаны выше).
C) Анализ содержания нуклеиновой кислоты в VLP Qβ, которые были повторно собраны в присутствии олигодезоксинуклеотидов, посредством электрофореза в денатурирующем полиакриламидном геле с TBE-мочевиной: Повторно собранные VLP Qβ (0,4 мг/мл), содержащие олигодезоксинуклеотиды G8-8, инкубировали в течение 2 час при 37°C с 125 ед. бензоназы на мл VLP Qβ в присутствии 2 мМ MgCl2. Затем обработанные бензоназой VLP Qβ обрабатывали протеиназой K (степень очистки ПЦР, Roche Molecular Biochemicals, No. в каталоге 1964364), как описано в примере 7. Затем реакционные смеси смешивали с буфером для образцов TBE-мочевина и наносили на 15% полиакриламидный гель с TBE-мочевиной (Novex®, Invitrogen No. в каталоге EC6885). Для качественного, а также количественного стандарта 1 пмоль, 5 пмоль и 10 пмоль олигодезоксинуклеотида, который использовали для реакции повторной сборки, наносили на тот же самый гель. Указанный гель красили SYBR®-Gold (Molecular Probes No. в каталоге S-11494). Окраска SYBR®-Gold показала, что повторно собранные капсиды Qβ содержали нуклеиновую кислоту, мигрирующую совместно с олигодезоксинуклеотидами, которые были использованы в реакции повторной сборки. Вместе взятые данные о резистентности к расщеплению бензоназой представленного нуклеиновой кислотой содержимого VLP Qβ, которые были повторно собраны в присутствии олигодезоксинуклеотидов, и выделении олигодезоксинуклеотида из очищенных частиц при расщеплении протеиназой K свидетельствуют об упаковке олигодезоксинуклеотида.
ПРИМЕР 20
Связывание пептидов, полученных из антигена меланомы Melan-A, с Qb
* A/L указывает замену аланина на лизин по сравнению с исходным пептидом Melan-A дикого типа
** аминокислоты из линкерной последовательности указаны строчными буквами.
Для химического связывания остатков пептида Melan-A с VLP Qb использовали следующие способы:
Для пептида Melan-A 16-35, Melan-A 16-35 A/L и Melan-A 26-35-C A/L: 2 мл раствора 3,06 мг/мл VLP Qb в 20 мМ Hepes, pH 7,2 подвергали взаимодействию в течение 30 минут с 18,4 мкл раствора 50 мМ SMPH (сукцинимидил-6-(β-малеимидопропионамидогексаноат, Pierce) в ДМСО при 25°C на качалке. Затем реакционный раствор диализовали дважды в течение 2 часов против 2 л 20 мМ Hepes, pH 7,2 при 4°C. Затем 2 мл диализованной реакционной смеси подвергали взаимодействию с 18,4 мкл 50 мМ исходного раствора пептида (в ДМСО) в течение двух часов при 25°C на качалке. Затем реакционную смесь диализовали 2 x 2 часа против 2 литров 20 мМ Hepes, pH 7,2 при 4°C. Связанные продукты названы Qb-Melan-A 16-35 (SEQ ID NO: 54), Qb-Melan-A 16-35 A/L (SEQ ID NO: 55) и Qb-Melan-A 26-35-C A/L (SEQ ID NO: 60). Для Melan-A 26-35: 2 мл раствора 3,06 мг/мл белка капсида Qb в 20 мМ Hepes, pH 7,2, подвергали взаимодействию в течение 30 минут с 75,3 мкл раствора 50 мМ SMPH в ДМСО при 25°C на качалке. Затем реакционный раствор диализовали дважды в течение 2 часов против 2 л 20 мМ Hepes, pH 7,2 при 4°C. Затем 2 мл диализованной реакционной смеси подвергали взаимодействию с 37,7 мкл 50 мМ исходного раствора пептида (в ДМСО) в течение 4 часов при 25°C на качалке. Затем реакционную смесь диализовали 2 x 2 часа против 2 литров 20 мМ Hepes, pH 7,2 при 4°C. Связанный продукт назван Qb-Melan-A 26-35.
Для Melan-A 26-35 A/L (SEQ ID NO: 57): 2 мл раствора 3,06 мг/мл VLP Qb в 20 мМ Hepes, pH 7,2, подвергали взаимодействию в течение 30 минут с 37,7 мкл раствора 50 мМ SMPH в ДМСО при 25°C на качалке. Затем реакционный раствор диализовали дважды в течение 2 часов против 2 л 20 мМ Hepes, pH 7,2 при 4°C. Затем 2 мл диализованной реакционной смеси подвергали взаимодействию с 18,4 мкл 50 мМ исходного раствора пептида (в ДМСО) в течение 4 часов при 25°C на качалке. Затем реакционную смесь диализовали 2 x 2 часа против 2 литров 20 мМ Hepes, pH 7,2 при 4°C. Связанный продукт назван Qb-Melan-A 26-35 A/L.
Для Melan-A 20-40 A/L (SEQ ID NO: 58): 2 мл раствора 3,06 мг/мл VLP Qb в 20 мМ Hepes, pH 7,2, подвергали взаимодействию в течение 30 минут с 18,4 мкл раствора 50 мМ SMPH в ДМСО при 25°C на качалке. Затем реакционный раствор диализовали дважды в течение 2 часов против 2 л 20 мМ Hepes, pH 7,2 при 4°C. Затем 2 мл диализованной реакционной смеси подвергали взаимодействию с 184 мкл 5 мМ исходного раствора пептида (в ДМСО) в течение 4 часов при 25°C на качалке. Затем реакционную смесь диализовали 2 x 2 часа против 2 литров 20 мМ Hepes, pH 7,2 при 4°C. Связанный продукт назван Qb-Melan-A 20-40 A/L.
Для Melan-A 26-40 A/L (SEQ ID NO: 59): 2 мл раствора 3,06 мг/мл VLP Qb в 20 мМ Hepes, pH 7,2, подвергали взаимодействию в течение 30 минут с 37,7 мкл раствора 50 мМ SMPH в ДМСО при 25°C на качалке. Затем реакционный раствор диализовали дважды в течение 2 часов против 2 л 20 мМ Hepes, pH 7,2 при 4°C. Затем 2 мл диализованной реакционной смеси подвергали взаимодействию с 184 мкл 5 мМ исходного раствора пептида (в ДМСО) в течение 4 часов при 25°C на качалке. Затем реакционную смесь диализовали 2 x 2 часа против 2 литров 20 мМ Hepes, pH 7,2 при 4°C. Связанный продукт назван Qb-Melan-A 26-40 A/L.
Эффективность связывания проверяли анализом в SDS-ПААГ. На фигуре 1 показан анализ в SDS-ПААГ VLP Qb-Melan-A. Melan-A-пептиды связывали с VLP Qb. Конечные продукты смешивали с буфером для образцов и разделяли в восстанавливающих условиях в 16% геле с трис-глицином Novex® в течение 1,5 часов при 125 В. Разделенные белки красили при вымачивании геля в растворе Кумасси синего. Фоновую окраску удаляли промывкой геля в 50% метаноле, 8% уксусной кислоте. Использовали маркер молекулярной массы (P 77085, New England BioLabs, Beverly, USA) в качестве образца сравнения скорости миграции Qb-Melan-A (дорожка 1). 14 мкг либо Qb отдельно (дорожка 2), либо Qb, дериватизованного SMPH (дорожка 3) наносили для сравнения с 8 мкг конечного продукта: Qb-Melan-A 16-35 (дорожка 4), Qb-Melan-A 16-35 A/L (дорожка 5), Qb-Melan-A 26-35 (дорожка 6) и Qb-Melan-A 26-35 A/L (дорожка 7).
Пептид Melan-A 16-35 A/L содержит эпитоп для цитотоксических T-лимфоцитов (CTL) Melan-A 26-35, и Qb-Melan-A 16-35 A/L исследовали далее в отношении его иммуногенности in vitro и in vivo.
ПРИМЕР 21
VLP Qβ Melan-A 16-35 A/L процессируются дендритными клетками, и они презентируются in vitro T-клеткам.
Образованием Melan-A-специфичных T-клеток in vitro
Чтобы оценить иммуногенность вакцины Qb-Melan-A 16-35 A/L, создавали Melan-A-специфичные T-клетки in vitro. Происходящие из незрелых моноцитов дендритные клетки (DC) от здоровых добровольцев HLA-A2 получали, как описано ранее (Salusto, F. et al. 1994, J. Exp. Med. 179: 1109). DC (0,5 x 106/лунку) импульсно обрабатывали либо ложно, либо 40 мкг/мл Qb-Melan-A 16-35 A/L в течение 2 час при 37°C. Чтобы увеличить частоту Melan-A-специфичных предшественников CTL, выделяли аутологичные T-клетки CD8 из PBMC магнитной сортировкой (Miltenyi Biotech). 1 x 106/лунку T-клеток CD8 добавляли к импульсно обработанным антигеном DC. На 3 день в культуральную среду добавляли IL-2 в концентрации 10 ед./мл и увеличивали до 50 ед./мл до 12 дня.
Экспансию Melan-A-специфичных T-клеток CD8 в линиях клеток оценивали окрашиванием меченными фикоэритрином (PE) тетрамерами, полученными из HLA-A2, нагруженными пептидом Melan-A 26-35 (HLA-A2-Melan-A-PE), как описано ранее (Romero, P. et all, 1998, J. Exp. Med. 1998, 188: 1641). Клетки метили в течение 1 час при комнатной температуре, используя HLA-A2-Melan-A 26-35-PE, затем добавляли анти-CD8-ФИТЦ-антитело (BD PharMingen, San Jose, USA) на 30 мин на льду. После промывки клетки анализировали в FACS Calibur, используя компьютерную программу CellQuest. Клетки поступали в устройство прямого светорассеяния и устройство бокового светорассеяния и пропускались лимфоциты. Из указанной популяции лимфоцитов только CD8-позитивные T-клетки отбирали для дальнейших анализов. Количество Melan-A-специфичных T-клеток CD8+ рассчитывали в виде процента HLA-A2-Melan-A-позитивных клеток из лимфоцитов CD8+. Вакцина Qb-Melan-A 16-35 A/L индуцировала пролиферацию Melan-A 26-35 A/L-специфичных CTL (9,7% T-клеток CD8), свидетельствуя о том, что вакцина эффективно захватывается, процессируется до CTL-эпитопа (Melan-A 26-35 A/L) и презентируется клетками DC человека T-клеткам.
Melan-A-CTL обогащали FACS-сортировкой (FACSVantage, Becton Dickenson) и клонировали посредством лимитирующих разведений, как описано ранее (Knuth, A. 1989, PNAS, 86: 2804). Клоны CTL отбирали по позитивному окрашиванию тетрамерами HLA-A2-Melan-A-PE. CTL периодически повторно стимулировали активированными фитогемагглютинином гетерологичными облученными PBMC.
Оценка узнавания антигена клонами CTL
Осуществляли анализ высвобождения51Cr, чтобы проанализировать функциональную активность клонов Melan-A-CTL. APC инкубировали в течение 1,5 час с 10-6M пептидом Melan-A 26-35 A/L или 10-6M пептидом вируса гриппа M1 в качестве негативного контроля. APC инкубировали с51Cr, чтобы измерить неспецифичный захват радионуклидов во время импульсной обработки пептидов. После тщательной промывки для удаления остаточного антигена и радиоактивности 104/лунку APC инкубировали с разными количествами Melan-A-специфичных T-клеток в течение 5 час. Собирали надосадки и специфичный лизис Melan-A-презентирующих APC Melan-A-специфичными T-клетками рассчитывали, следуя уравнению:
% специфичного лизиса= ((имп./мин в эксперименте - имп./мин спонтанно)/(имп./мин максимально - имп./мин спонтанно)) x 100, где имп./мин в эксперименте означает счет радиоактивности, измеренный в экспериментальном образце, имп./мин спонтанно означает счет, полученный от APC, импульсно меченных51Cr, без добавления T-клеток, и имп./мин максимально означает счет, полученный от импульсно51Cr-меченных APC, лизированных 1% NP-40.
APC, нагруженные Melan-A 26-35 A/L, но не APC, нагруженные несоответствующим пептидом M1, эффективно подвергались лизису CTL (70-85% специфичный лизис), что подтверждает антигенную специфичность клонов CTL.
ПРИМЕР 22
Способность иммуностимулирующих последовательностей (ISS) активировать клетки человека in vitro
Чтобы выбрать оптимальные ISS для введения в вакцину Qb-Melan-A, тестировали серию CpG с разным количеством фланкирующих G или двунитевую РНК, такую как поли(I:C), в отношении их способности к повышающей регуляции CD69 на T-клетках CD8 человека и способности индуцировать секрецию IFN-альфа и IL-12 в PBMC человека.
PBMC человека выделяли из лейкоцитарных пленок и обрабатывали указанными ISS в среде RPMI, содержащий 10% FCS, в течение 18 час. IFN-альфа в надосадках измеряли посредством ELISA, используя набор антител, поставляемый PBL Biomedical Laboratories. PBMC красили мышиным анти-CD8 человека-ФИТЦ, мышиным анти-CD19 человека-PE и анти-CD69 человека-APC и анализировали проточной цитометрией. G10-PO была наиболее активной ISS в индукции секреции IFN-альфа (фиг. 2A). G9-9 и G8-8 были немного менее активными, чем G10- PO, хотя они индуцировали высокие уровни секреции IFN-альфа. Уменьшение количества фланкирующих G в других олигонуклеотидах приводило к более низкой секреции IFN-альфа. Поли(I:C)-обработанные PBMC не высвобождали IFN-альфа, хотя поли(I:C)-обработанные T- и B-клетки из PBMC обеспечивали повышающую регуляцию CD69 (фиг. 2B). Поли(I:C) также индуцировала секрецию IL-12 из PBMC и полученных из моноцитов DC.
Обработка PBMC G10-PO, G9-9 и G8-8 повышала количество CD69 на клеточных мембранах T-клеток CD8 почти в одинаковой степени. Уменьшение количества фланкирующих G (ниже 7) в других олигонуклеотидах снижало их способность индуцировать секрецию IFN-альфа (фиг. 2A) и способность к повышающей регуляции CD69 на T-клетках (фиг. 2B). Полученные данные показывают, что G10-PO, G9-9 и G8-8 имеют сопоставимую высокую активность на клетках человека, поэтому их можно использовать в качестве ISS в вакцине Qb-Melan-A.
ПРИМЕР 23
G10-PO увеличивает экспансию Melan-A-специфичных T-клеток in vitro.
Способность VLP Qb-Melan-A индуцировать пролиферацию in vitro Melan-A-специфичных CTL тестировали в присутствии или в отсутствие G10-PO. Полученные из незрелых моноцитов человека DC (0,5×106/лунку) импульсно обрабатывали либо пептидом Qb-Melan-A 16-35 A/L или Melan-A 26-35 A/L, либо ложно обрабатывали, как в примере 21. Полученные из моноцитов человека DC являются негативными по toll-подобному рецептору 9 (TLR-9), и поэтому они не отвечают на CpG. B-клетки и плазмацитоидные DC, присутствующие в PBMC человека, являются TLR-9-позитивными и отвечающими на обработку CpG продукцией цитокинов (IFN-альфа, IL-12). Чтобы исследовать влияние G10-PO на антигенпрезентирующую способность полученных из моноцитов DC, импульсно обработанные антигеном DC промывали и инкубировали с 1 x 106 аутологичных PBMC в присутствии или в отсутствие 2 мкМ G10-PO в течение 2 час. Аутологичные T-клетки CD8 (1×106), выделенные магнитной сортировкой, добавляли к APC и инкубировали в течение 12 дней, используя условия культивирования клеток, описанные в примере 21. Melan-A-специфичные CTL выявляли по окрашиванию тетрамером HLA-A2-Melan-A-PE и анализом проточной цитометрией. Добавление G10-PO к DC, импульсно обработанным Qb-Melan-A, увеличивало частоту специфичных CTL (от 10% до 14%), что указывает на то, что цитокинная среда, создаваемая посредством CpG-стимуляции, благоприятна для экспансии CTL in vitro.
ПРИМЕР 24
VLP Qbx33, нагруженные G3-6, G6, G10-PO или поли(I:C), индуцируют защиту против контрольного заражения p33-рекомбинантный вирус вакцинии.
Мышей B6 подкожно иммунизировали Qbx33 отдельно или нагруженным G3-6 или G6 или поли(I:C) (смотри примеры 12 и 14). Через восемь дней мышей контрольно заражали 1,5×106 БОЕ рекомбинантного вируса вакцинии, экспрессирующего антиген LCMV-p33. Через 4 дня мышей забивали и измеряли вирусные титры в яичниках, как описано ранее (Bachmann et al, Eur. J. Imunol. 1994, 24: 2228). Как изображено на фигуре 3, все мыши, получающие вакцину Qbx33, нагруженную либо G3-6 или G6, либо поли(I:C), были защищены от заражения вирусом. Напротив, у интактных мышей и мышей, иммунизированных только Qbx33, не происходило удаления вируса из яичников. Полученные данные свидетельствуют, что VLP отдельно недостаточно для индукции защитного иммунного ответа CTL, тогда как VLP, нагруженные CpG или поли(I:C) очень эффективны в примировании нативных CTL.
Подобным образом иммунизация мышей Qbx33, нагруженными G10-PO, примировала p33-специфичные CTL (6,2%±1,4% по сравнению с 0,2%±0,1% у нативных мышей), а также индуцировала защиту от заражения рекомбинантным вирусом вакцинии.
ПРИМЕР 25
VLP Qβ Melan-A 16-35 A/L процессируется и презентируется аллелем HLA-A0201 класса I MHC человека и индуцирует экспансию функциональных Melan-A-специфичных T-клеток CD8+ у HLA-A2-трансгенных мышей.
Мыши HHD экспрессируют химерную одноцепочечную молекулу класса I с β2-микроглобулином человека, ковалентно связанным с N-концом доменов α1 и α2 A2, слитых с доменом Db α3 (Firat, H. et al. 1999, Eur. J. Immunol., 29: 3112). Экспрессия трансгена HLA-A2 у указанных мышей позволяет исследовать способность VLP Qβ Melan-A 16-35 A/L процессироваться и презентироваться в виде CTL-эпитопа Melan-A 26-35 и примировать CTL in vivo. Кроме того, может быть исследовано влияние адъювантов, таких как ISS, in vivo.
Мышей HHD либо оставляли необработанными, либо иммунизировали посредством подкожной инъекции 100 мкг Qb-Melan-A 16-35 A/L или Qb-pIC-Melan-A 16-35 A/L. Через восемь дней выделяли спленоциты, ресуспендировали в буфере FACS (PBS, 2% FCS, 5 мМ EDTA, pH 8,2) и красили меченными тетрамерами HLA-A2-Melan-A-PE в течение 30 мин при комнатной температуре. На второй стадии добавляли крысиные анти-CD8 мыши-APC (BD PharMingen, San Jose, USA) и анти-Mel14-ФИТЦ (BD PharMingen, San Jose, USA) на 30 мин при 4°C. После промывки эритроциты лизировали лизирующим раствором BD (BD Biosciences, San Jose, USA) в течение 10 мин при комнатной температуре. Наконец, клетки селезенки анализировали в FACS Calibur, используя компьютерную программу CellQuest. Прежде всего клетки поступали в устройство прямого светорассеяния и устройство бокового светорассеяния, и пропускались лимфоциты. Из указанной популяции лимфоцитов только CD8-позитивные T-клетки отбирали для дальнейших анализов. Клетки, меченные HLA-A2-Melan-A-PE и Mel14-ФИТЦ, измеряли с помощью детектора FL2 и FL1 соответственно. Количество Melan-A-специфичных активированных T-клеток CD8+ рассчитывали в виде процента HLA-A2-Melan-A-позитивных, Mel14-негативных клеток от общего количества лимфоцитов CD8+.
Анализ проточной цитометрией показал, что Qb-pIC-Melan-A 16-35 A/L индуцировал неожиданно высокую экспансию Melan-A-специфичных активированных T-клеток CD8+Mel14- (2,43% и 0,73%), которая была выше по сравнению с необработанными животными (0,22% и 0,37%). Следует отметить, что эффективность вакцины значительно возрастала, только когда Qb-Melan-A нагружали поли(I:C).
Тетрамер HLA-A2-Melan-A человека, который использовали выше, не очень эффективно связывается с мышиными Melan-A-специфичными T-клетками, так как белок является химерным. Поэтому авторы предполагают намного более высокий уровень антиген-специфичных T-клеток у указанных мышей.
В сходном эксперименте авторы анализировали эффективность Qb-G10-Melan-A 16-35 A/L в примировании CTL in vivo при вакцинации пептидом, смешанным с CpG и IFA. Мышей HHD иммунизировали либо 200 мкг Q-G10-Melan-A 16-35 A/L, либо 50 мкг Melan-A 16-35 A/L, смешанным с 20 нмоль CpG и неполным адъювантом Фрейнда (IFA), или оставляли необработанными. Через восемь дней жизнеспособные лимфоциты выделяли из селезенок и красили в отношении Melan-A-специфичных T-клеток CD8+. Окрашивание осуществляли в буфере FACS в течение 1,5 час при 37°C PE-меченным тетрамером, специфичным для химерной молекулы MHC класса I HLA-A2α1α2Kbα3, нагруженной пептидом Melan-A 26-35. На второй стадии добавляли крысиные анти-CD8 мыши-APC (BD PharMingen, San Jose, USA) и анти-Mel14-ФИТЦ (BD PharMingen, San Jose, USA) на 30 мин при 4°C. Наконец, клетки селезенки анализировали в FACS Calibur, используя компьютерную программу CellQuest.
Анализ проточной цитометрией показал, что Qβ-G10-Melan-A 16-35 A/L индуцировал экспансию Melan-A-специфичных активированных T-клеток CD8+Mel14- (18,2%), которая была выше по сравнению с необработанными животными (2%) или животными, получавшими эквимолярные количества Melan-A A/L 16-35, смешанного с 20 нмоль CpG и IFA (2,0%). Следует отметить, что эффективность вакцины возрастала, только когда Qb-Melan-A нагружали G10-PO.
В сходных экспериментальных условиях иммунизация мышей HHD Qb-Melan-A 16-35 A/L или Qb-Melan-A 26-35 A/L, нагруженными G8-8 или G10 PO, индуцирует экспансию HLA-A2-Melan-A-позитивных и Mel14-негативных T-клеток CD8.
Указанные данные вместе взятые демонстрируют способность нагруженных ISS вакцин Qb-пептид очень эффективно примировать CTL против чужеродных и аутоантигенов.
ПРИМЕР 26
Qbx33, нагруженные CpG, можно использовать в режиме гомологичного, а также гетерологичного примирования-ревакцинации для индукции ответа долговременной памяти T-клеток CD8+.
Мышей иммунизировали 150 мкг Qbx33/NKCpG и через 8 дней частота встречаемости p33-специфичных T-клеток возрастала от 0,4%±0,2 % у нативных мышей до 7,5%±2,2% у иммунизированных животных, что измеряли с использованием антиген-специфичных тетрамеров MHC/пептид. Через 20 дней специфичная по отношению к пептиду популяция T-клеток CD8+ уменьшалась до 1,6%±0,7%. Вторая иммунизация указанных мышей через 30 дней после первой иммунизации с использованием 150 мкг Qbx33/NKPS повышала ответ T-клеток памяти до 8,4%±1,9% специфичных T-клеток. Указанный ответ медленно ослабевал, но мог быть снова усилен через 4 месяца после первой реиммунизации с использованием 150 мкг Qbx33/NKPS с достижением уровней T-клеток 23,8%±5,2%.
Когда 3 мышей примировали 50 мкг пептида p33, смешанного с 20 нмоль NKPS и IFA, только 0,6%±0,4% специфичных T-клеток CD8+ могли быть индуцированы вплоть до 8 дня после иммунизации. Однако полученный низкий ответ можно было эффективно усилить через 7 недель посредством реиммунизации Qbx33/NKPS до уровней 28,5%±9,8%.
Иммунизация 1 x 106 бляшкообразующих единиц рекомбинантного вируса вакцинии, экспрессирующего p33-пептид почти, не могла индуцировать T-клеточный ответ (1,1%±0,5%), но очень эффективно ответ усиливался при реиммунизации через 6 месяцев 150 мкг Qbx33/NKPS до уровней T-клеток 28,1±4,2%.
Полученные результаты показывают, что Qb, нагруженный CpG, очень эффективно усиливает любой предсуществующий T-клеточный ответ в режимах гетерологичного, а также гомологичного примирования-ревакцинации. Следует отметить, что Qb/NKPS даже может усиливать очень неэффективно примированный ответ T-клеток пептидами или рекомбинантными вирусами. Кроме того, когда сильный T-клеточный ответ вызывали с помощью Qbx33/NKPS, авторы могли усилить этот ответ, используя иммунологически эффективное количество гетерологичной вакцины, такой как пептид p33 отдельно, рекомбинантный вирус, экспрессирующий p33, или p33, слитый или связанный с VLP. В последнем случае используемая VLP не является VLP, полученной из РНК-фага Qβ, а представляет собой VLP, полученную, например, из HBcAg, или VLP, полученную из АР205.
Пример 27.
Иммунизация мышей, экспрессирующих HLA-A2, QβG10
Выступающий эпитоп HLA-A2 Melan-A был выбран для создания вакцины для иммунотерапии метастазирующей меланомы. Иммунодоминантный CD8+ Т-клеточный эпитоп содержит A-L замену в положении 27 для повышения афинности к молекуле МНС класса I человека. Кроме того, добавлена N-концевая линкерная последовательность, полученная из природного белка Melan-A, вместе с N-концевым цистеином. Полученный пептид CGHGHSYTTAEELAGIGILTV) присоединяли к вирусоподобной частице РНК-фага Qβ (дикого типа), где вирусоподобная частица была упакована вместе с безметилированным CpG-содержащим олигонуклеотидом G10 (GGGGGGGGGG GACGATCGTC GGGGGGGGGG). Полученный продукт MelQβG10 использовали для иммунизации мышей, экспрессирующих молекулу HLA-A2 человека. Мышей в количестве 2-3 на группу иммунизировали, через 8 дней собирали спленоциты и с помощью FACS измеряли Melan-A-специфические CD8+ Т-клетки, используя антигенспецифические тетрамеры МНК класса I на CD8+ Т-клеточном эпитопе ELAGIGILTV (фигура 1). Результаты показали а) правильный процессинг и презентацию эпитопа Melan-A Qb-связанного пептидного предшественника и b) эффективность вакцины MelQ(3G10 для стимуляции увеличения количества Melan-A-специфических CD8+ клеток у HLA-А2-положительного донора.
Эффективность Melan-A-специфических CD8+ клеток, индуцированных вакциной, оценивали ex vivo окрашиванием на секрецию IFNg, CD8 положительных спленоцитов после краткой повторной стимулиции HLA-A2 положительным BMDC на Melan-A-пептидном эпитопе (ELAGIGILTV) (фигура 2). Корреляция между Melan-A-тетрамером и IFNg-положительными CD8+ Т-клетками была очень близкой, что указывает на высокую эффективность MelQβG10 не только примировать меланома-специфические Т-клетки, но также проявлять эффекторные функции (уничтожать меланомные клетки), используя IFNg-секрецию в качестве заместительного маркера.
Таким образом, было доказано, что вакцина MelQβG10 in vivo на модели гуманизированных мышей является эффективной для стимуляции ответа CD8+ Т-клеток, то есть способна секретировать высокое количество IFNg, не является VLP, полученной из РНК-фага Qβ, а представляет собой VLP, полученную, например, из HBcAg, или VLP, полученную из АР205.
Пример 27.
Иммунизация мышей, экспрессирующих HLA-A2, QβG10
Выступающий эпитоп HLA-A2 Melan-A был выбран для создания вакцины для иммунотерапии метастазирующей меланомы. Иммунодоминантный CD8+ Т-клеточный эпитоп содержит A-L замену в положении 27 для повышения афинности к молекуле МНС класса I человека. Кроме того, добавлена N-концевая линкерная последовательность, полученная из природного белка Melan-A, вместе с N-концевым цистеином. Полученный пептид CGHGHSYTTAEELAGIGILTV) присоединяли к вирусоподобной частице РНК-фага Qβ (дикого типа), где вирусоподобная частица была упакована вместе с безметилированным CpG-содержащим олигонуклеотидом G10 (GGGGGGGGGG GACGATCGTC GGGGGGGGGG). Полученный продукт MelQβG10 использовали для иммунизации мышей, экспрессирующих молекулу HLA-A2 человека. Мышей в количестве 2-3 на группу иммунизировали, через 8 дней собирали спленоциты и с помощью FACS измеряли Melan-A-специфические CD8+ Т-клетки, используя антигенспецифические тетрамеры МНК класса I на CD8+ Т-клеточном эпитопе ELAGIGILTV (фигура 1). Результаты показали а) правильный процессинг и презентацию эпитопа Melan-A Qb-связанного пептидного предшественника и b) эффективность вакцины MelQβG10 для стимуляции увеличения количества Melan-A-специфических CD8+клеток у HLA-A2-положительного донора.
Эффективность Melan-A-специфических CD8+клеток, индуцированных вакциной, оценивали ex vivo окрашиванием на секрецию IFNg, CD8 положительных спленоцитов после краткой повторной стимулиции HLA-A2 положительным BMDC на Melan-A-пептидном эпитопе (ELAGIGILTV) (фигура 2). Корреляция между Melan-A-тетрамером и IFNg-положительными CD8+ Т-клетками была очень близкой, что указывает на высокую эффективность MelQβG10 не только примировать меланома-специфические Т-клетки, но также проявлять эффекторные функции (уничтожать меланомные клетки), используя IFNg-секрецию в качестве заместительного маркера.
Таким образом, было доказано, что вакцина MelQβG10 in vivo на модели гуманизированных мышей является эффективной для стимуляции ответа CD8+ Т-клеток, то есть способна секретировать высокое количество IFNg.
Реферат
Изобретение относится к областям молекулярной биологии, вирусологии, иммунологии и медицины. Композиция содержит модифицированную вирусоподобную частицу (VLP), иммуностимулирующие вещества, в частности ДНК-олигонуклеотиды, содержащие неметилированные С и G (CpG) и отдельные пептиды, полученные из Melan-A. Такие CpG-VLP значительно более иммуногенны, чем их аналоги, не содержащие CpG, и индуцируют усиленные В- и Т-клеточные ответы. Иммунный ответ против аналогов пептида Melan-A, соединенного с VLP, усиливается подобным образом, как и иммунный ответ против самой VLP. Кроме того, Т-клеточные ответы против аналогов пептида Melan-A главным образом направлены к типу Th1. Антигены, связанные с VLP, нагруженными CpG, могут быть идеальными вакцинами для профилактической или терапевтической вакцинации против аллергии, опухолей и других аутологичных молекул и хронических вирусных заболеваний. Изобретение может быть использовано в медицине. 5 н. и 47 з.п. ф-лы, 3 ил., 4 табл.
Формула
(a) вирусоподобную частицу, где указанная вирусоподобная частица представляет собой вирусоподобную частицу РНК-фага;
(b) по крайней мере одно иммуностимулирующее вещество;
(i) где указанное иммуностимулирующее вещество представляет собой неметилированный CpG-содержащий олигонуклеотид; и
(ii) где указанное иммуностимулирующее вещество связано с указанной вирусоподобной частицей; и
(c) по крайней мере один антиген или антигенную детерминанту;
(i) где указанный антиген или антигенная детерминанта связаны с указанной вирусоподобной частицей, и
(ii) где указанный антиген содержит или, альтернативно, состоит из аналога пептида Melan-A меланомы человека.
(a) LAGIGILTV (SEQ ID NO:84);
(b) MAGIGILTV (SEQ ID NO:85);
(c) EAMGIGILTV (SEQ ID NO:86);
(d) ELAGIGILTV (SEQ ID NO:50);
(e) EMAGIGILTV (SEQ ID NO:87);
(f) YAAGIGILTV (SEQ ID NO:88); и
(g) FAAGIGILTV (SEQ ID NO:89).
(a) сайта присоединения, который не является природным сайтом указанного антигена или антигенной детерминанты; и
(b) сайта присоединения, который является природным сайтом указанного антигена или антигенной детерминанты;
и где указанное связывание указанного антигена или антигенной детерминанты и указанной вирусоподобной частицы происходит посредством ассоциации указанного первого сайта присоединения и указанного второго сайта присоединения.
(a) CGHGHSYTTAEELAGIGILTV (SEQ ID NO:55);
(b) CGGELAGIGILTV (SEQ ID NO:57);
(c) CSYTTAEELAGIGILTVILGVL (SEQ ID NO:58);
(d) CGGELAGIGILTVILGVL (SEQ ID NO:59);
(e) ELAGIGILTVGGC (SEQ ID NO:60);
(f) CSPKSLELAGIGILTV (SEQ ID NO:92); и
(g) ELAGIGILTVILGVLGGC (SEQ ID NO:93).
(а) бактериофага Qβ;
(b) бактериофага R17;
(c) бактериофага fr;
(d) бактериофага GA;
(e) бактериофага SP;
(f) бактериофага MS2;
(g) бактериофага M11;
(h) бактериофага МХ1;
(i) бактериофага NL95;
(j) бактериофага f2;
(k) бактериофага РР7; и (1) бактериофага АР205.
5'X1X2CGX3X4 3'
где X1, X2, Х3 и Х4 означают любой нуклеотид.
(a) GGGGACGATCGTCGGGGGG (SEQ ID NO:2);
(b) GGGGGACGATCGTCGGGGGG (SEQ ID NO:3);
(c) GGGGGGACGATCGTCGGGGGG (SEQ ID NO:4);
(d) GGGGGGGACGATCGTCGGGGGG (SEQ ID NO:5);
(e) GGGGGGGGACGATCGTCGGGGGGG (SEQ ID NO:6);
(f) GGGGGGGGGACGATCGTCGGGGGGGG (SEQ ID NO:7);
(g) GGGGGGGGGGACGATCGTCGGGGGGGGG (SEQ ID NO:8);
(h) GGGGGGCGACGACGATCGTCGTCGGGGGGG (SEQ ID NO:9); и
(i) GGGGGGGGGGGACGATCGTCGGGGGGGGGG (SEQ ID NO:41).
Комментарии