Модификация рекомбинантного аденовируса иммуногенными эпитопами белка circumsporozoite плазмодия - RU2585228C2
Код документа: RU2585228C2
Чертежи
















































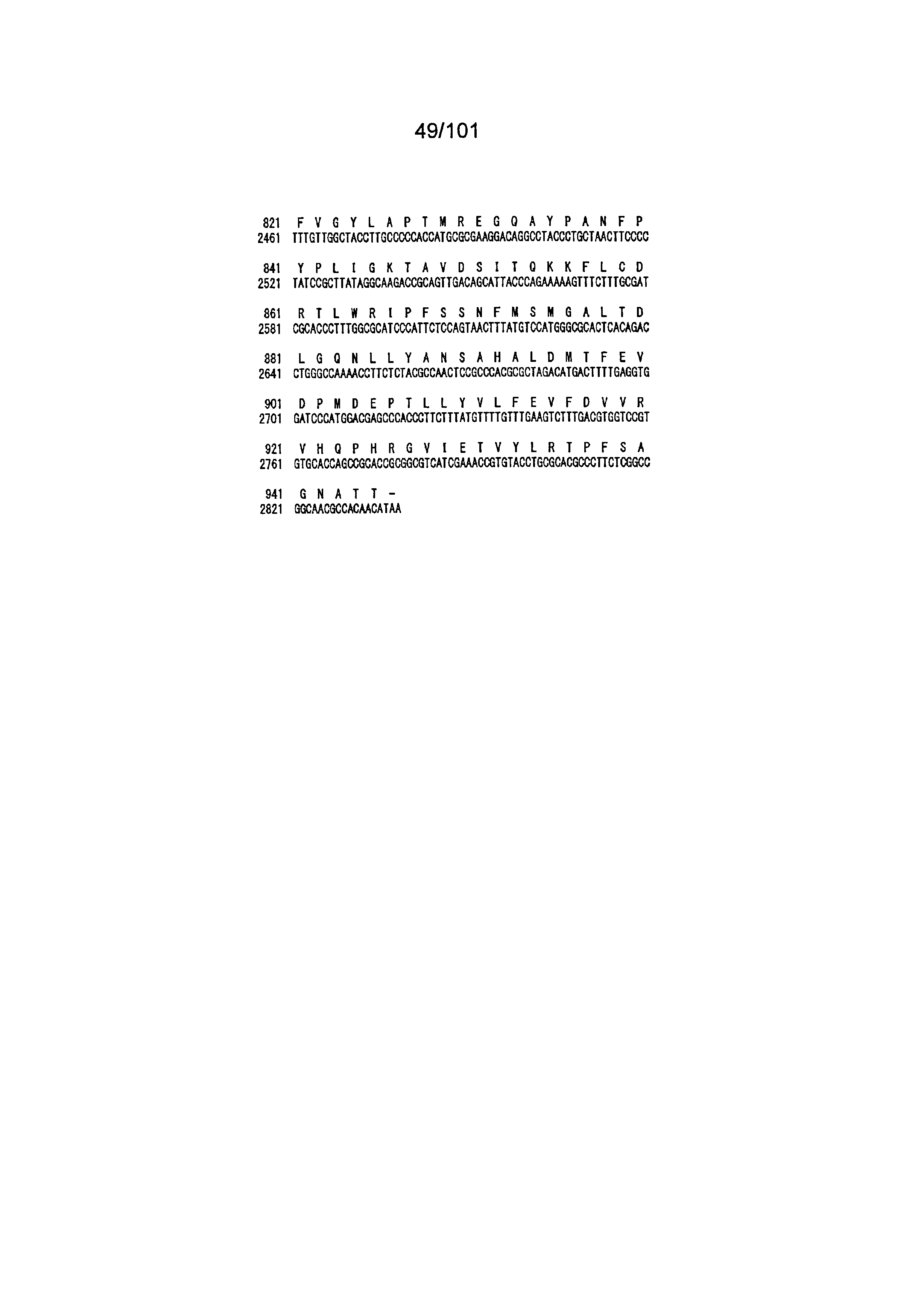


























































































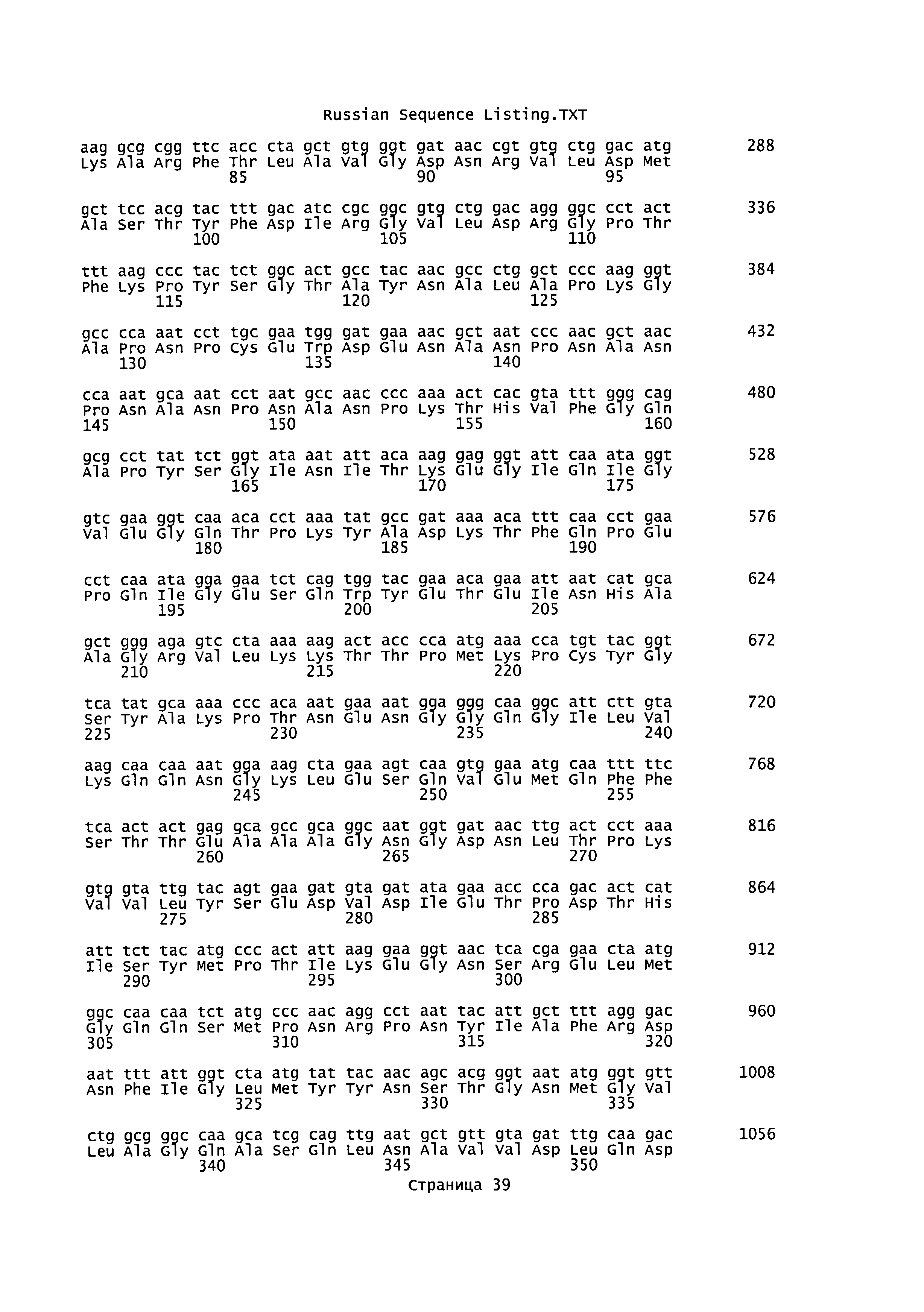






















































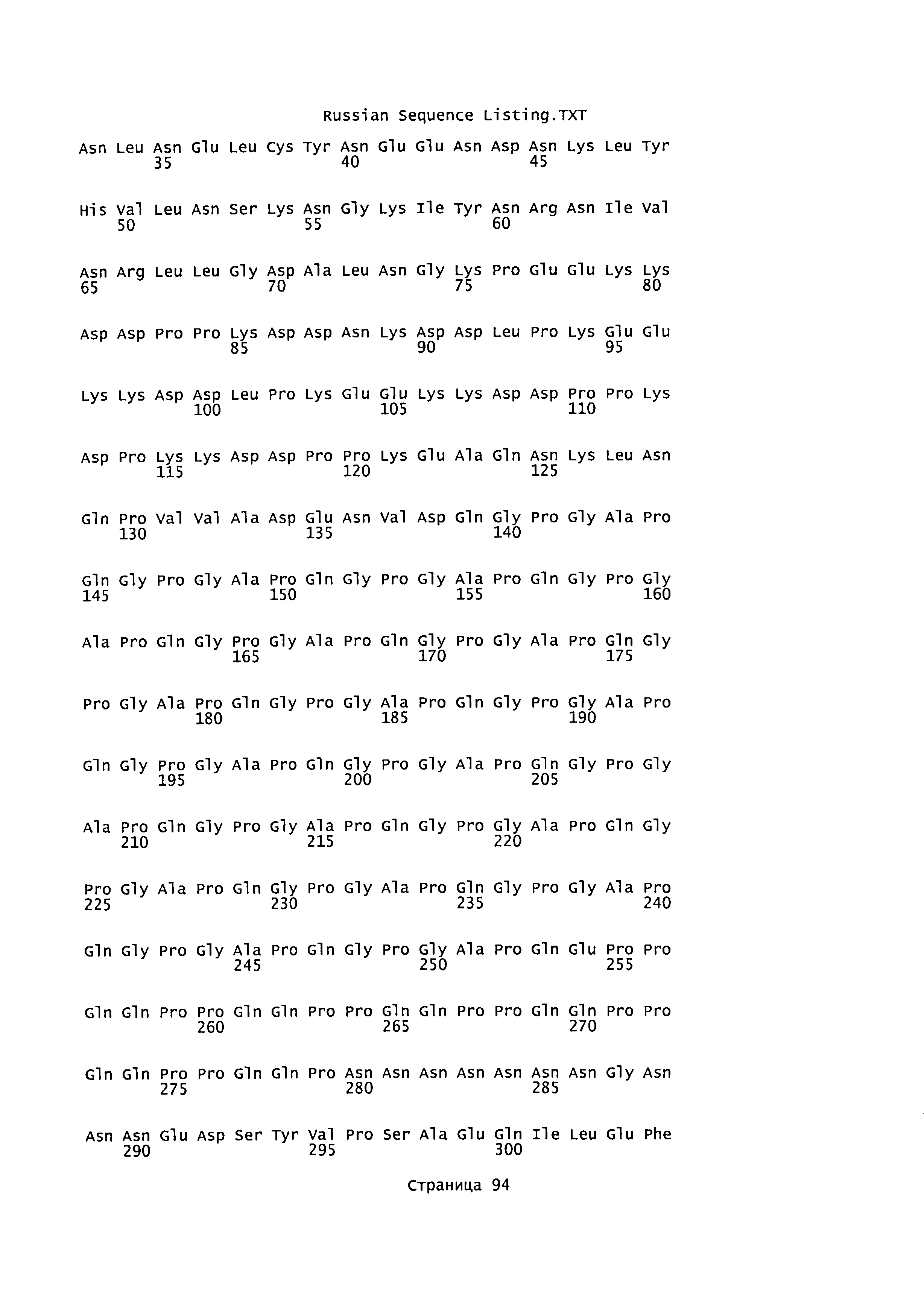

























































































Описание
ПРИОРИТЕТ
[0001] Настоящая заявка испрашивает приоритет и представляет собой частичное продолжение международной заявки № PCT/US09/054212 от 18 августа 2009 года, которая полностью включена, как если бы она была полностью приведена в настоящем описании.
ЗАЯВЛЕНИЕ ОБ ИССЛЕДОВАНИИ, ФИНАНСИРУЕМОМ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА
[0002] Настоящее изобретение было сделано при поддержке государства в рамках Гранта №1R01AI081510-01A1 Национального института аллергии и инфекционных заболеваний (NIAID), учреждения, которое входит в состав Национальных институтов здоровья. Государство обладает определенными правами на настоящее изобретение.
ОБЛАСТЬ ТЕХНИКИ
[0003] Настоящее изобретение относится к области медицины и биотехнологии. Более конкретно, изобретение относится к применению векторов на основе аденовирусов с модифицированным капсидом для индукции эффективного иммунного ответа на антиген малярийных плазмодиев, такой как белок circumsporozoite плазмодия, который пригоден для получения вакцин от малярии.
УРОВЕНЬ ТЕХНИКИ
[0004] Малярия - тяжелое заболевание, которое входит в число наиболее распространенных инфекций в тропических областях. Ежегодно инфицируется приблизительно 300-500 миллионов людей, и относительно высок процент заболеваемости и смертности. Протекание с осложнениями и смертность особенно характерны для молодых людей и взрослых, попадающих в эндемические области малярии и ранее не переносивших малярию. По оценкам Всемирной Организации Здравоохранения (ВОЗ) 2-3 миллиона детей ежегодно умирает от малярии в одной только Африке. Широкое распространение и возрастающая во многих странах частота возникновения малярии, вызываемой резистентными к лекарственным препаратам плазмодиями (Plasmodium falciparum, а в последнее время также Plasmodium vivax) и устойчивыми к инсектицидам переносчиками (малярийный комар), указывают на необходимость разработки новых способов борьбы с данным заболеванием (Nussenzweig and Long 1994).
[0005] Малярийные плазмодии отличаются сложным жизненным циклом, который включает преэритроцитарные, эритроцитарные и половые формы паразитов; их жизненный цикл является потенциальной мишенью для разработки вакцин от малярии. Преэритроцитарные и эритроцитарные формы обнаруживаются в организме хозяина, а половые формы существуют в организме переносчика инфекции. Было показано, что вакцинация живыми, ослабленными облучением спорозоитами (IrSp) вызывает стерильную защиту (например, полная резистентность в отношении инфицирования плазмодиями) у мышей (Nussenzweig et al. 1967), приматов, не являющихся человеком (Gwadz et al. 1979), и у людей (Clyde et al. 1973, Edelman et al. 1993). Защита, обеспечиваемая IrSp, обусловлена нейтрализацией спорозоитов за счет развития как гуморального (В-лимфоциты), так и клеточного (Т-лимфоциты) иммунного ответа (Tsuji et al. 2001). Хотя вакцинация IrSp являет собой привлекательное решение, единственный способ получения спорозоитов - это вскрытие слюнных желез комара, и к настоящему времени технология выращивания большого количества спорозоитов in vitro не известна. Следовательно, требуется альтернативный вектор для вакцинации, который может обеспечивать такой же сильный защитный иммунитет против малярии.
[0006] Одной из перспективных мишеней для создания такого вектора вакцины является белок circumsporozoite (CS), который экспрессируется на поверхности спорозоита. Эффективные нейтрализующие антитела направлены против иммунодоминантных, видоспецифичных повторяющихся доменов белка circumsporozoite (CS). У Plasmodium falciparum (паразит, вызывающий малярию у человека), повторы (NANP)n являются консервативными в изолятах из всех регионов мира. Этот центральный повтор содержит множественные повторы эпитопов, распознаваемых В-лимфоцитами, и, соответственно, белок CS может вызывать сильный гуморальный иммунный ответ, запуская реакцию В-лимфоцитов (Tsuji et al. 2001). В области С-конца белка CS имеется несколько эпитопов, распознаваемых Т-лимфоцитами, которые могут вызывать сильный клеточный иммунный ответ (Tsuji et al. 2001). Гуморальный ответ (антитела) может приводить к уничтожению паразитов в результате взаимодействия и нейтрализации инфицирующей способности спорозоитов (внеклеточного плазмодия) до попадания в гепатоцит, а клеточный ответ (ответ Т-лимфоцитов) может быть направлен на EEF (внутриклеточные формы плазмодия) за счет секреции интерферона-гамма. Указанные виды иммунного ответа предотвращают созревание и быстрое деление EEF, в ходе которых образуются тысячи спорозоитов, возвращающихся в кровь и инфицирующих эритроциты, вызываемое заболевание, определяемое как малярия.
[0007] Было показано, что одна из вакцин от малярии, которая в настоящее время проходит клинические испытания и представляет собой химерный белок RTS, S «GlaxoSmithKline», составленный из поверхностного антигена вируса гепатита В и части белка circumsporozoite плазмодия falciparum (PfCSP) в форме вирусоподобной частицы (Заявка на Международный патент № РСТ/ЕР 1992/002591 для компании «SmithKline Beecham Biologicals S.A.» от 11 ноября 1992 г.), уменьшает инфицирование малярией в клинических исследованиях (Alonso et al. 2004, Alonso et al. 2005, Bejon et al. 2008). RTS, S вызывает развитие анти-PfCSP гуморального иммунного ответа, но относительно слабый PfCSP-специфичный клеточный ответ (CD8+) (Kester et al. 2008), что может послужить причиной относительно слабой защиты, обеспечиваемой RTS, S. Напротив, вакцины от малярии на основе аденовируса могут вызывать защитный клеточный иммунный ответ (Заявка на Международный патент № РСТ/ЕР2003/051019, от 16 декабря 2003 г., Rodrigues et al. 1997). Однако в настоящее время существуют два препятствия, ограничивающие применение системы на основе аденовируса в качестве вакцины против малярии: (1) неспособность вызывать эффективный гуморальный ответ на трансгенный продукт, и (2) предсуществующий иммунитет на аденовирусы, в частности, на аденовирус серотипа 5, который снижает иммуногенность вакцины на основе аденовируса.
[0008] Один из подходов, которые недавно были взяты на вооружение работах по повышению вызываемого аденовирусом иммунного ответа, состоит во встраивании эпитопа антигена, распознаваемого В-лимфоцитами (например, эпитопа бактерии или вируса) в белки капсида аденовируса, такие как гексон, белок нити, пентон и pIX (Worgall et al. 2005, McConnell et al. 2006, Krause et al. 2006, Worgall et al. 2007).
[0009] Кроме того, чтобы обойти проблему предсуществующего иммунитета на аденовирус серотипа 5 (Ad5), в качестве платформы для вакцины исследовали аденовирусы других, менее распространенных серотипов, такие как 11, 35, 26, 48, 49 и 50, и было показано, что они вызывают иммунный ответ на трансген, несмотря на наличие анти-Ad5 иммунитета (Заявка на Международный патент № РСТ/ЕР2005/055183 Crucell Holland B.V., от 12 октября 2005 г., Abbink et al. 2007). Также, чтобы обойти проблему предсуществующего анти-Ad5-иммунитета была разработана замена гексона Ad5, который является целевым белком капсида для нейтрализующих антител, на гексоны других серотипов (Wu et al. 2002, Roberts et al. 2006).
[0010] Однако сведений об улучшенном векторе на основе аденовируса, который позволял бы преодолеть оба указанных препятствия одновременно при применении вектора на основе аденовируса для получения вакцины против малярии, в настоящее время нет. В связи с высоким распространением серотипа Ad5 высоко в областях, эндемических по малярии (Ophorst et al. 2006), возникает потребность в противомалярийной вакцине на основе аденовируса, которая вызывает защитный гуморальный и клеточный иммунный ответ, даже при наличии предсуществующего иммунитета к аденовирусу.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0011] Настоящее описание относится к различным модификациям белков аденовируса, направленным на увеличение иммунного ответа на трансген рекомбинантной аденовирусной вакцины и на преодоление предсуществующего иммунитета к аденовирусу.
[0012] Более конкретно, один вариант реализации относится к рекомбинантному аденовирусу, полученному из плазмидного вектора на основе рекомбинантного аденовируса, причем плазмидный вектор на основе рекомбинантного аденовируса содержит последовательность нуклеотидов, кодирующую (i) белок circumsporozoite плазмодия или его антигенную часть, функционально связанную с гетерологичным промотором; и (ii) модифицированный белок капсида или сердцевины, причем иммуногенный эпитоп circumsporozoite плазмодия встроен по меньшей мере в часть одного белка капсида и/или сердцевины или замещает указанную часть.
[0013] Согласно некоторым вариантам реализации белок circumsporozoite плазмодия также включает белок circumsporozoite Plasmodium falciparum или Plasmoclium yoelii. Белок circumsporozoite также может включать белок с оптимизацией кодонов circumsporozoite Plasmodium falciparum или Plasmodium yoelii, и согласно некоторым аспектам может кодироваться последовательностью нуклеотидов SEQ ID NO:2 или SEQ ID NO:1, соответственно.
[0014] Согласно другим вариантам реализации иммуногенный эпитоп также включает эпитоп circumsporozoite плазмодия, распознаваемый В-лимфоцитами. Эпитоп, распознаваемый В-лимфоцитами, может быть встроен в модифицированный белок капсида, и согласно некоторым аспектам белок капсида может включать гипервариабельный участок гексона (HVR). HVR может также включать HVR1 или HVR5, в которых часть HVR1 или HVR5 заменена эпитопом, распознаваемым В-лимфоцитами. Согласно другим аспектам белок капсида может также включать белок нити капсида, в котором эпитоп, распознаваемый В-лимфоцитами, встроен в белок нити. Согласно некоторым аспектам эпитоп, распознаваемый В-лимфоцитами, представляет собой белок circumsporozoite Plasmodium falciparum, в котором эпитоп, распознаваемый В-лимфоцитами, представляет собой последовательность повтора, например, (NANP)n (SEQ ID NO:60), причем последовательностью повтора может представлять собой (NANP)4, (NANP)6, (NANP)8, (NANP)10, (NANP)12, (NANP)14, (NANP)16, (NANP)18, (NANP)20, (NANP)22 или (NANP)28. Согласно другому аспекту эпитоп, распознаваемый В-лимфоцитами, представляет собой эпитоп белка circumsporozoite Plasmodium yoelii, в котором эпитоп, распознаваемый В-лимфоцитами, представляет собой последовательность повтора, например, (QGPGAP)n (SEQ ID NO:59), причем последовательностью повтора может представлять собой (QGPGAP)3, (QGPGAP)4, (QGPGAP)5, (QGPGAP)6, (QGPGAP)7, (QGPGAP)8, (QGPGAP)9, (QGPGAP)11, или (QGPGAP)12.
[0015] Согласно другим вариантам реализации иммуногенный эпитоп также включает эпитоп circumsporozoite плазмодия, распознаваемый CD4+ или CD8+Т-лимфоцитами. Эпитоп, распознаваемый CD4+ или CD8+Т-лимфоцитами, может быть встроен в модифицированный белок капсида или сердцевины. Согласно некоторым аспектам белок капсида может включать гипервариабельный участок гексона (HVR). HVR может также содержать HVR1, в котором часть HVR1 замещена эпитопом, распознаваемым CD4+ или CD8+Т-лимфоцитами. Согласно другим аспектам белок капсида может также включать белок pVII, и эпитоп, распознаваемый CD4+ или CD8+Т-лимфоцитами, введен в белок pVII. Согласно некоторым аспектам эпитоп, распознаваемый CD4+Т-лимфоцитами, представляет собой белок circumsporozoite Plasmodium falciparum, причем эпитоп, распознаваемый CD4+Т-лимфоцитами, представляет собой EYLNKIQNSLSTEWSPCSVT (SEQ ID NO:62). Согласно другим аспектам эпитоп, распознаваемый CD4+Т-лимфоцитами, представляет собой эпитоп белка circumsporozoite Plasmodium yoelii, причем эпитоп, распознаваемый CD4+Т-лимфоцитами, представляет собой YNRNIVNRLLGDALNGKPEEK (SEQ ID NO:61).
[0016] Другие варианты реализации относятся к фармацевтической композиции или композиции вакцины против малярии, включающей рекомбинантный аденовирус в соответствии с вариантами реализации, перечисленными выше. Дополнительные варианты реализации включают способ лечения, профилактики или диагностики малярии, включающий введение терапевтического количества фармацевтической композиции или композиции вакцины против малярии в соответствии с вариантами реализации, перечисленными выше.
[0017] Согласно еще одному варианту реализации предложен способ лечения, включающий проведение первичной-поддерживающей вакцинации, при которой субъекту вводят серию возрастающих доз или одинаковых доз через заданные периоды времени. Периоды времени могут быть любой продолжительности, достаточной для того, чтобы вызвать гуморальный и/или клеточный иммунный ответ. Например, как описано ниже, период времени может составлять 3 недели, но не ограничивается этим.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
[0018] На Фиг.1 приведена структурная схема рекомбинантного аденовируса с модифицированным капсидом в соответствии с вариантами реализации настоящего изобретения.
[0019] На Фиг.2 приведена структурная схема, иллюстрирующая конструкцию ДНК HVR1-модифицированного аденовируса, составляющей рекомбинантный плазмидный вектор на основе HVR1-модифицированного аденовируса.
[0020] На Фиг.3 приведена структурная схема, иллюстрирующая конструкцию ДНК HVR5-модифицированного аденовируса, составляющей плазмидный вектор на основе рекомбинантного HVR5-модифицированного аденовируса.
[0021] На Фиг.4 приведена структурная схема, иллюстрирующая конструкцию ДНК модифицированного по белку нити аденовируса, составляющей плазмидный вектор на основе модифицированного по белку нити рекомбинантного аденовируса.
[0022] На Фиг.5 приведена структурная схема, иллюстрирующая конструкцию ДНК модифицированного по HVR1 и белку нити аденовируса, составляющей плазмидный вектор на основе модифицированного по HVR1 и белку нити аденовируса.
[0023] На Фиг.6 приведена структурная схема, иллюстрирующая конструкцию ДНК модифицированного по белку нити и pVII аденовируса, составляющей плазмидный вектор на основе модифицированного по белку нити и pVII аденовируса.
[0024] На Фиг.7 приведена структурная схема, иллюстрирующая конструкцию ДНК HVR1- и pVII-модифицированного аденовируса, составляющей плазмидный вектор на основе HVR1- и pVII-модифицированного аденовируса.
[0025] На Фиг.8 приведена структурная схема, иллюстрирующая конструкцию ДНК модифицированного по HVR1, белку нити и pVII аденовируса, составляющей плазмидный вектор на основе модифицированного по HVR1, белку нити и pVII аденовируса.
[0026] На Фиг.9 приведена последовательность нуклеиновой кислоты белка circumsporozoite Plasmodium yoelii с оптимизированными кодонами (PyCS, SEQ ID NO:1) и соответствующая последовательность аминокислот (SEQ ID NO:30).
[0027] На Фиг.10 приведена последовательность нуклеиновой кислоты белка circumsporozoite Plasmodium falciparum с оптимизированными кодонами (PfCSP, SEQ ID NO:2) и соответствующая последовательность аминокислот (SEQ ID NO:43).
[0028] На Фиг.11 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего три повтора последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=3) в HVR1(SEQ ID NO:3, нуклеиновая кислота; SEQ ID NO:31, аминокислоты). Встроенная последовательность (QGPGAP)3 подчеркнута.
[0029] На Фиг.12 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего четыре повтора последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=4) в HVR1(SEQ ID NO:4, нуклеиновая кислота; SEQ ID NO:32, аминокислоты). Встроенная последовательность (QGPGAP)4 подчеркнута.
[0030] На Фиг.13 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего пять повторов последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=5) в HVR1(SEQ ID NO:5, нуклеиновая кислота; SEQ ID NO:33, аминокислоты). Встроенная последовательность (QGPGAP)5 подчеркнута.
[0031] На Фиг.14 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего шесть повторов последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=6) в HVR1(SEQ ID NO:6, нуклеиновая кислота; SEQ ID NO:34, аминокислоты). Встроенная последовательность (QGPGAP)6 подчеркнута.
[0032] На Фиг.15 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего семь повторов последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=7) в HVR1(SEQ ID NO:7, нуклеиновая кислота; SEQ ID NO:35, аминокислоты). Встроенная последовательность (QGPGAP)7 подчеркнута.
[0033] На Фиг.16 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего восемь повторов последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=8) в HVR1(SEQ ID NO:8, нуклеиновая кислота; SEQ ID NO:36, аминокислоты). Встроенная последовательность (QGPGAP)8 подчеркнута.
[0034] На Фиг.17 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего девять повторов последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=9) в HVR1(SEQ ID NO:9, нуклеиновая кислота; SEQ ID NO:37, аминокислоты). Встроенная последовательность (QGPGAP)9 подчеркнута.
[0035] На Фиг.18 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего одиннадцать повторов последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=11) в HVR1(SEQ ID NO:10, нуклеиновая кислота; SEQ ID NO:38, аминокислоты). Встроенная последовательность (QGPGAP)11 подчеркнута.
[0036] На Фиг.19 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего двенадцать повторов последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=12) в HVR1(SEQ ID NO:11, нуклеиновая кислота; SEQ ID NO:39, аминокислоты). Встроенная последовательность (QGPGAP)12 подчеркнута.
[0037] На Фиг.20 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего четыре повтора последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=4) в HVR1(SEQ ID NO:12, нуклеиновая кислота; SEQ ID NO:44, аминокислоты). Встроенная последовательность (NANP)4 подчеркнута.
[0038] На Фиг.21 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего шесть повторов последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=6) в HVR1(SEQ ID NO:13, нуклеиновая кислота; SEQ ID NO:45, аминокислоты). Встроенная последовательность (NANP)6 подчеркнута.
[0039] На Фиг.22 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего восемь повторов последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=8) в HVR1(SEQ ID NO:14, нуклеиновая кислота; SEQ ID NO:46, аминокислоты). Встроенная последовательность (NANP)8 подчеркнута.
[0040] На Фиг.23 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего десять повторов последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=10) в HVR1(SEQ ID NO:15, нуклеиновая кислота; SEQ ID NO:47, аминокислоты). Встроенная последовательность (NANP)10 подчеркнута.
[0041] На Фиг.24 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего двенадцать повторов последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=12) в HVR1 (SEQ ID NO:16, нуклеиновая кислота; SEQ ID NO:48, аминокислоты). Встроенная последовательность (NANP)12 подчеркнута.
[0042] На Фиг.25 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего четырнадцать повторов последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=14) в HVR1 (SEQ ID NO:17, нуклеиновая кислота; SEQ ID NO:49, аминокислоты). Встроенная последовательность (NANP)14 подчеркнута.
[0043] На Фиг.26 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего шестнадцать повторов последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=16) в HVR1 (SEQ ID NO:18, нуклеиновая кислота; SEQ ID NO:50, аминокислоты). Встроенная последовательность (NANP)16 подчеркнута.
[0044] На Фиг.27 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего восемнадцать повторов последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=18) в HVR1 (SEQ ID NO:19, нуклеиновая кислота; SEQ ID NO:51, аминокислоты). Встроенная последовательность (NANP)18 подчеркнута.
[0045] На Фиг.28 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего двадцать повторов последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=20) в HVR1 (SEQ ID NO:20, нуклеиновая кислота; SEQ ID NO:52, аминокислоты). Встроенная последовательность (NANP)20 подчеркнута.
[0046] На Фиг.29 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего двадцать два повтора последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=22) в HVR1 (SEQ ID NO:21, нуклеиновая кислота; SEQ ID NO:53, аминокислоты). Встроенная последовательность (NANP)22 подчеркнута.
[0047] На Фиг.30 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего двадцать восемь повторов последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=28) в HVR1 (SEQ ID NO:22, нуклеиновая кислота; SEQ ID NO:54, аминокислоты). Встроенная последовательность (NANP)28 подчеркнута.
[0048] На Фиг.31 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного гексона, содержащего три повтора последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=3) в HVR5 (SEQ ID NO:23, нуклеиновая кислота; SEQ ID NO:40, аминокислоты). Встроенная последовательность (QGPGAP)3 подчеркнута.
[0049] На Фиг.32 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного белка нити, содержащего три повтора последовательности эпитопа PyCS, распознаваемого В-лимфоцитами (QGPGAP)n, (SEQ ID NO:59; n=3) в последовательности белка нити (SEQ ID NO:24, нуклеиновая кислота; SEQ ID NO:41, аминокислоты). Встроенная последовательность (QGPGAP)3 подчеркнута.
[0050] На Фиг.33 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного белка нити, содержащего четыре повтора последовательности эпитопа PfCSP, распознаваемого В-лимфоцитами (NANP)n, (SEQ ID NO:60; n=4) в последовательности белка нити (SEQ ID NO:25, нуклеиновая кислота; SEQ ID NO:55, аминокислоты). Встроенная последовательность (NANP)4 подчеркнута.
[0051] На Фиг.34 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного белка pVII, содержащего последовательность эпитопа PyCS, распознаваемого CD4+-лимфоцитами, YNRNIVNRLLGDALNGKPEEK, (SEQ ID NO:61) в N-конце pVII (SEQ ID NO:26, нуклеиновая кислота; SEQ ID NO:42, аминокислоты). Встроенная последовательность YNRNIVNRLLGDALNGKPEEK подчеркнута.
[0052] На Фиг.35 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного белка pVII, содержащего последовательность эпитопа PfCSP, распознаваемого CD4+-лимфоцитами, EYLNKIQNSLSTEWSPCSVT, (SEQ ID NO:62), в С-конце pVII (pVII-1; SEQ ID NO:27, нуклеиновая кислота; SEQ ID NO:56, аминокислоты). Встроенная последовательность EYLNKIQNSLSTEWSPCSVT подчеркнута.
[0053] На Фиг.36 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного белка pVII, содержащего последовательность эпитопа PfCSP, распознаваемого CD4+-лимфоцитами, EYLNKIQNSLSTEWSPCSVT, (SEQ ID NO:62), перед первым клеточным сигналом внутриядерной локализации (NLS) pVII (pVII-2; SEQ ID NO:28, нуклеиновая кислота; SEQ ID NO:57, аминокислоты). Встроенная последовательность EYLNKIQNSLSTEWSPCSVT подчеркнута.
[0054] На Фиг.37 приведены последовательности нуклеиновой кислоты и аминокислот модифицированного белка pVII, содержащего последовательность эпитопа PfCSP, распознаваемого CD4+-лимфоцитами, EYLNKIQNSLSTEWSPCSVT, (SEQ ID NO:62), между двумя NLS белка pVII (pVII-3; SEQ ID NO:29, нуклеиновая кислота; SEQ ID NO:58, аминокислоты). Встроенная последовательность EYLNKIQNSLSTEWSPCSVT подчеркнута.
[0055] На Фиг.38 показана экспрессия белка PyCS в клетках AD293 после временной трансфекции вектором PyCS-GFP/pShuttle-CMV. Белок PyCS определяли методом вестерн-блоттинга с применением моноклональных анти-PyCS антител мыши (9D3).
[0056] На Фиг.39 показаны результаты окрашивания серебром и вестерн-блоттинга (А) и твердофазного иммуноферментного анализа (ELISA) (В) очищенных рекомбинантных аденовирусов PyCS-GFP с модифицированным капсидом, проводимых для подтверждения встраивания эпитопа (QGPGAP)3 (SEQ ID NO:59; n=3) в белки капсида аденовируса. При проведении ELISA планшеты для ELISA покрывали непосредственно очищенными аденовирусами и определяли встроенный эпитоп в частицах аденовируса при помощи анти-PyCS антитела.
[0057] На Фиг.40 показаны результаты окрашивания серебром и вестерн-блоттинга (А) и ELISA (В) очищенных рекомбинантных аденовирусов PyCS с модифицированным капсидом, проводимых для подтверждения встраивания эпитопа (QGPGAP)n (SEQ ID NO:59) в белки капсида аденовируса. При проведении ELISA планшеты для ELISA покрывали непосредственно очищенными аденовирусами, и определяли встроенный эпитоп в частицах аденовируса при помощи анти-PyCS антитела.
[0058] Фиг.41 иллюстрирует режим однократной вакцинации аденовирусами PyCS с модифицированным капсидом, содержащими повторы (QGPGAP)n (SEQ ID NO:59, n=3, 4, 5, 6, 9, 12) (А), и CD8+-ответ, развивающийся через две недели после вакцинации (В).
[0059] Фиг.42 иллюстрирует режим первичной и поддерживающей вакцинации аденовирусами PyCS с модифицированным капсидом, содержащими повторы (QGPGAP)n (SEQ ID NO:59, n=3) (A), PyCS-специфичные гуморальные ответы к неделе 10 (В), и концентрацию малярийного плазмодия в печени через 42 часа после инфицирования спорозоитом (С).
[0060] На Фиг.43 показан титр антител против спорозоита, определяемый методом непрямого иммунофлуоресцентного анализа (ИФА) (А) и по нейтрализующей активности в отношении спорозоита in vitro (В) в объединенных пробах сыворотки, полученных от мышей, подвергшихся вакцинации в соответствии с режимом на Фиг.42.
[0061] Фиг.44 иллюстрирует режим первичной и поддерживающей вакцинации аденовирусами PyCS с модифицированным капсидом, содержащими повторы (QGPGAP)n (SEQ ID NO:59, n=4,6) в HVR1(A), PyCS-специфичные гуморальные ответы к неделе 9 (В), концентрацию малярийного плазмодия в печени через 42 часа после инфицирования спорозоитом (С) и нейтрализующую активность в отношении спорозоита in vitro в объединенных пробах сыворотки (D). Мышей вакцинировали с адъювантом или без него.
[0062] Фиг.45 иллюстрирует режим первичной и поддерживающей вакцинации аденовирусами PyCS с модифицированным капсидом, содержащими повторы (QGPGAP)n (SEQ ID NO:59, n=6, 9, 12) в HVR1(A), PyCS-специфичные гуморальные ответы к неделе 9 (В), PyCS-специфичные клеточные ответы CD8+Т-лимфоцитовк неделе 9 (С) и концентрацию малярийного плазмодия в печени через 42 часа после инфицирования спорозоитом (D). Мышей вакцинировали с адъювантом или без него.
[0063] На Фиг.46 показана экспрессия белка PfCSP в клетках AD293 после временной трансфекции вектором PfCSP/pShuttle-CMV. Белок PfCSP определяли методом вестерн-блоттинга с применением моноклональных анти-NANP антител мыши (2А10).
[0064] На Фиг.47 показаны результаты окрашивания серебром и вестерн-блоттинга (А) и ELISA (В) очищенных рекомбинантных аденовирусов PfCSP с модифицированным капсидом, проводимых для подтверждения встраивания эпитопа (NANP)4 (SEQ ID NO:60; n=4) в белки капсида аденовируса. Встроенный эпитоп (NANP)4 (SEQ ID NO:60; n=4) определяли с использованием моноклонального анти-NANP антитела мыши (2А10). При проведении ELISA планшеты для ELISA покрывали непосредственно очищенными аденовирусами.
[0065] На Фиг.48 показаны результаты окрашивания серебром и вестерн-блоттинга (А) и ELISA (В) очищенных рекомбинантных аденовирусов PfCSP с модифицированным капсидом, проводимых для подтверждения встраивания эпитопа (NANP)n (SEQ ID NO:60; n=4, 6, 8, 10, 12, 14, 16, 18, 20, 22) в белки капсида аденовируса. При проведении ELISA планшеты для ELISA покрывали непосредственно очищенными аденовирусами, и определяли встроенный эпитоп в частицах аденовируса при помощи анти-PfCSP антитела (2А10).
[0066] Фиг.49 иллюстрирует режим первичной и поддерживающей вакцинации рекомбинантным аденовирусами PfCSP с модифицированным капсидом, содержащими (NANP)4 (SEQ ID NO:60; n=4) (А), и PfCSP-специфичные гуморальные ответы к неделе 9 (В).
[0067] Фиг.50 иллюстрирует режим первичной и поддерживающей вакцинации рекомбинантным аденовирусами PfCSP с модифицированным капсидом, содержащими (NANP)n (SEQ ID NO:60; n=10, 16, 22) в HVR1 (А) и PfCSP-специфичные гуморальные ответы к неделе 9 (В).
[0068] Фиг.51 иллюстрирует режим первичной и поддерживающей вакцинации рекомбинантным аденовирусами PfCSP с модифицированным капсидом, содержащими (NANP)n (SEQ ID NO:60; n=4, 6, 8, 10) в HVR1 (А) и PfCSP-специфичные гуморальные ответы к неделе 9 (В). Мышей вакцинировали с адъювантом или без него.
[0069] На Фиг.52 показаны результаты окрашивания серебром очищенного аденовируса с белком нити и белком pVII-, модифицированным повторами (QGPGAP)3 ((QGPGAP)3-Fib/CD4-pVII-1/PyCS-GFP) (А) и титр анти-QGPGAP антител к неделе 10 у мышей, вакцинированных (QGPGAP)3-Fib/PyCS-GFP и (QGPGAP)3-Fib/CD4-pVII-1/PyCS-GFP, как описано на Фиг.49 (В). Результаты двух независимых экспериментов отложены на графике после нормировки с указанием медиан титров антител в B-Fib/PyCS-GFP-вакцинированной группе.
[0070] На Фиг.53 приведены схематические изображения структуры белков pVII аденовируса с последовательностью эпитопа PfCSP, распознаваемого CD4+-лимфоцитами, EYLNKIQNSLSTEWSPCSVT (SEQ ID NO:62), встроенными перед первым клеточным сигналом внутриядерной локализации (NLS) pVII (PfCD4-pVII-2; SEQ ID NO:29, нуклеиновая кислота; SEQ ID NO:58, аминокислоты) (А) и результат окрашивания серебром проводимых для подтверждения встраивания указанного эпитопа в pVII (В).
[0071] Фиг.54 иллюстрирует режим первичной и поддерживающей вакцинации рекомбинантными аденовирусами PfCSP, модифицированными по белкам HVR1 и pVII (А), PfCSP-специфичные гуморальные ответы к неделе 6 (В), PfCSP-специфичные клеточные ответы CD4+Т-лимфоцитов(EYLNKIQNSLSTEWSPCSVT; SEQ ID NO:62) к неделе 9 (С).
[0072] Фиг.55 иллюстрирует нейтрализацию рекомбинантного аденовируса in vitro пробами сыворотки человека. Клетки AD293 инфицировали рекомбинантными аденовирусами в присутствии разведенной сыворотки человека в течение ночи и измеряли экспрессию GFP (зеленого флуоресцентного белка) в качестве маркера инфицирования.
[0073] Фиг.56 иллюстрирует действие иммунитета против аденовируса на индукцию PyCS-специфичного Ответа Т-лимфоцитов на аденовирусы PyCS-GFP с модифицированным капсидом in vivo. На панели (А) приведено краткое описание дизайна исследования. На панели (В) показан PyCS-специфичный CD8+-Ответ Т-лимфоцитов у мышей, вакцинированных диким типом (wt)/«пустым» аденовирусом дважды, и в дальнейшем примированных аденовирусами PyCS-GFP с модифицированным капсидом.
[0074] Фиг.57 иллюстрирует действие иммунитета против аденовируса на индукцию PyCS-специфичного гуморального иммунного ответа на аденовирусы PyCS-GFP с модифицированным капсидом in vivo. На панели (А) приведено краткое описание дизайна исследования. На панели (В) показан PyCS-специфичный гуморальный иммунный ответ у мышей, вакцинированных диким типом (wt)/«пустым» аденовирусом дважды, и далее - двумя дозами аденовирусов PyCS-GFP с модифицированным капсидом.
ПУТИ РЕШЕНИЯ ЗАДАЧ
[0075] Авторы настоящего изобретения обнаружили новый рекомбинантный аденовирус, обладающий новой структурой с модифицированным капсидом, который получают из плазмидного вектора на основе рекомбинантного аденовируса. Указанный рекомбинантный аденовирус способен инфицировать клетки млекопитающих, что вызывает экспрессию белка circumsporozoite Plasmodium указанными клетками. Также указанный рекомбинантный аденовирус содержит один или более белков капсида, которые были модифицированы путем встраивания целевого иммуногенного антигена, такого как эпитоп белка circumsporozoite Plasmodium, распознаваемый В-лимфоцитами или Т-лимфоцитами. Указанный рекомбинантный аденовирус получают путем трансфекции клеток линеаризованным рекомбинантным плазмидным вектором на основе аденовируса. Авторы настоящего изобретения применили полученный рекомбинантный аденовирус для проведения расширенного исследования фармацевтических средств, содержащих в качестве активного ингредиента рекомбинантный аденовирус, обладающий профилактическим и лечебным действием в отношении малярии. В результате авторы настоящего изобретения показали, что полученный рекомбинантный аденовирус обладает желаемым фармацевтическим действием.
ПОДРОБНОЕ ОПИСАНИЕ
[0076] В следующем описании приведены конкретные детали для всестороннего объяснения и описания вариантов реализации настоящего изобретения. Однако специалисты в данной области техники понимают, что настоящее изобретение может быть осуществлено без приведенных деталей. В других случаях хорошо известные структуры и функции не были подробно показаны или описаны, чтобы излишне не усложнять описание вариантов реализации настоящего изобретения.
[0077] Сокращения, применяемые для аминокислот, пептидов, последовательностей оснований и нуклеиновых кислот в настоящем описании, основаны на сокращениях, указанных в Договоренности по биохимической номенклатуре IUPAC-IUB (Международный союз теоретической и прикладной химии - Международный биохимический союз), Eur. J. Biochem., 138: 9 (1984), "Guideline for Preparing Specifications Including Base Sequences and Amino Acid Sequences" (Бюро по патентам и товарным знакам США), и соответствуют сокращениям, широко применяемым в данной области техники.
[0078] «Последовательность нуклеотидов», «полинуклеотид» или «молекула ДНК» в настоящем описании могут включать двухцепочечную ДНК или одноцепочечную ДНК (например, смысловую цепь и антисмысловую цепь, составляющие двухцепочечную ДНК) и не ограничены по своей полной длине. Последовательности нуклеотидов, кодирующие иммуногенный чужеродный ген, такой как ген, раскрываемый ниже в настоящей заявке, включают двухцепочечную ДНК, включая геномную ДНК, одноцепочечную ДНК (смысловую цепь), включая кДНК, одноцепочечную ДНК (антисмысловую цепь), обладающую последовательностью, комплементарной к смысловой цепи, и их фрагменты, если не указано другое.
[0079] В настоящем описании термины «последовательности нуклеотидов», «полинуклеотиды» или «молекулы ДНК» не ограничиваются функциональной областью, и могут включать по меньше мере одну область подавления экспрессии, кодирующую область, лидерную последовательность, экзон и интрон. Также примеры последовательностей нуклеотидов в полинуклеотидах могут включать РНК или ДНК. Полипептид, содержащий специфическую последовательность аминокислот, и полинуклеотид, содержащий специфическую последовательность ДНК, может включать фрагменты, гомологи, производные и мутанты полинуклеотида. Примеры мутантов последовательности нуклеотидов или полинуклеотида (такие как мутантная ДНК) включают существующие в природе аллельные мутанты; искусственные мутанты и мутанты, содержащие делеции, замены, присоединения и/или вставки. Очевидно, что такие мутанты кодируют полипептиды, обладающие по существу такой же функцией, что и полипептиды, кодируемые исходным немутированным полинуклеотидом.
[0080] Настоящее изобретение относится к рекомбинантному аденовирусу, который может экспрессировать антигенную детерминанту плазмодия Plasmodium и содержит один или более модифицированных белков капсида и/или сердцевины. Указанный рекомбинантный аденовирус получают из плазмидного вектора на основе рекомбинантного аденовируса. Применение аденовируса в качестве вектора подробно обсуждается далее. Плазмидные векторы на основе рекомбинантного аденовируса, описанные в настоящей заявке, можно применять в качестве вакцины против малярии или фармацевтической композиции, что обеспечивает индукцию как гуморального, так и клеточный иммунного ответа.
[0081] Из известных видов Plasmodium (P.) можно выбрать любой плазмодий вида Plasmodium, например, Р. falciparum, P. malariae, P. ovale, P. vivax, Р. knowlesi, P. berghei, P. chabaudi и Р. yoelii. Согласно некоторым вариантам реализации антигенную детерминанту получают из специфичного для грызунов Plasmodium yoelii или из специфичного для человека Plasmodium falciparum.
[0082] Согласно одному варианту реализации вектор на основе рекомбинантного аденовируса с модифицированным капсидом (также называемый в настоящей заявке вектором на основе рекомбинантного аденовируса) представляет собой плазмиду, которая кодирует и воспроизводит рекомбинантный вирус с модифицированным капсидом и/или сердцевиной (также называемый в настоящей заявке рекомбинантным аденовирусом), обладающий структурой с одним или более модифицированных белков капсида и/или сердцевины. Согласно указанным вариантам реализации настоящего описания модификация белков капсида и/или сердцевины может сопровождаться встраиванием по меньшей мере одного иммуногенного эпитопа белка circumsporozoite Plasmodium. В качестве альтернативы, по меньшей мере часть белка капсида и/или сердцевины может быть делегирована и заменена по меньшей мере одним иммуногенным эпитопом белка circumsporozoite Plasmodium. Согласно некоторым вариантам реализации иммуногенным эпитопом является эпитоп белка circumsporozoite Plasmodium, распознаваемый В-лимфоцитами или Т-лимфоцитами. Добавление эпитопа, распознаваемого В-лимфоцитами или Т-лимфоцитами, может служить для повышения эффективности вектора аденовируса, применяемого в качества вакцины против малярии, за счет возникновения или усиления гуморального иммунного ответа на белок CS. Модифицированные белки капсида и сердцевины и их значение для применения в рекомбинантном аденовирусе, описанном в настоящей заявке, также обсуждаются далее.
[0083] Один или более модифицированных белков капсида и/или сердцевины могут быть модифицированным белком гексона, модифицированным белком нити, модифицированным белком pVII или их сочетанием. Согласно одному варианту реализации часть гипервариабельного участка гексона (HVR) и/или часть белка нити заменяют по меньшей мере на один эпитоп белка circumsporozoite Plasmodium, распознаваемый В-лимфоцитами и/или Т-лимфоцитами. В качестве альтернативы, один или более эпитопов белка circumsporozoite Plasmodium, распознаваемых В-лимфоцитами и/или Т-лимфоцитами можно вводить в HVR белка нити или гексона. Согласно некоторым аспектам модифицированным HVR может быть HVR1, HVR2, HVR3, HVR4, HVR5, HVR6 или HVR7. Согласно другим аспектам модифицированным HVR может быть HVR1 или HVR5. Согласно некоторым вариантам реализации HVR-модифицированный гексон может обладать последовательностью нуклеиновой кислоты SEQ ID NO:3 (Фиг.11), SEQ ID NO:4 (Фиг.12), SEQ ID NO:5 (Фиг.13), SEQ ID NO:6 (Фиг.14), SEQ ID NO:7 (Фиг.15), SEQ ID NO:8 (Фиг.16), SEQ ID NO:9 (Фиг.17), SEQ ID NO:10 (Фиг.18), SEQ ID NO:11 (Фиг.19), SEQ ID NO:12 (Фиг.20), SEQ ID NO:13 (Фиг.21), SEQ ID NO:14 (Фиг.22), SEQ ID NO:15 (Фиг.23), SEQ ID NO:16 (Фиг 24), SEQ ID NO:17 (Фиг.25), SEQ ID NO:18 (Фиг.26), SEQ ID NO:19 (Фиг.27), SEQ ID NO:20 (Фиг.28), SEQ ID NO:21 (Фиг.29), SEQ ID NO:22 (Фиг.30) или SEQ ID NO:23 (Фиг.31). Согласно другим вариантам реализации модифицированный белок нити может обладать последовательностью нуклеиновой кислоты SEQ ID NO:24 (Фиг.32) или SEQ ID NO:25 (Фиг.33).
[0084] Согласно еще одному варианту реализации эпитоп белка circumsporozoite Plasmodium, распознаваемый Т-лимфоцитами, может быть встроен в белок сердцевины pVII в любом из следующих сайтов: С-конец, перед первым клеточным сигналом ядерной локализации (NLS) или между двумя NLS. В качестве альтернативы, эпитоп белка circumsporozoite Plasmodium, распознаваемый Т-лимфоцитами, может заменять часть белка pVII. Согласно некоторым вариантам реализации модифицированный белок pVII может характеризоваться последовательностью нуклеиновой кислоты SEQ ID NO:26 (Фиг.34), SEQ ID NO:27 (Фиг.35), SEQ ID NO:28 (Фиг.36) или SEQ ID NO:29 (Фиг.37).
[0085] Рекомбинантный аденовирус может обеспечивать экспрессию трансгенного белка или рекомбинантного трансгенного белка. Согласно некоторым вариантам реализации трансгенный белок или рекомбинантный трансгенный белок представляет собой белок circumsporozoite Plasmodium или антигенную детерминанту, кодируемые плазмидным вектором на основе рекомбинантного аденовируса, как описано в настоящей заявке, и экспрессируются рекомбинантным аденовирусом, полученным из указанного плазмидного вектора на основе рекомбинантного аденовируса после инфицирования одной или более клеток-хозяев.
[0086] Таким образом, согласно некоторым вариантам реализации, плазмидные векторы на основе рекомбинантного аденовируса содержат последовательность нуклеотидов рекомбинантного трансгенного белка. Согласно одному варианту реализации рекомбинантный трансгенный белок может содержать антигенную детерминанту Р. yoelii, плазмодия, специфичного для грызунов, причем антигенная детерминанта содержит ген белка circumsporozoite (CS) P. Yoelii или его антигенной части. Согласно еще одному варианту реализации рекомбинантный трансгенный белок может содержать антигенную детерминанту Р. falciparum, плазмодия, специфичного для человека, причем антигенная детерминанта содержит ген белка circumsporozoite (CS) P. falciparum или его антигенной части. Было показано, что белок CS P. falciparum обеспечивает профилактику малярии при применении его в качества основы для активной иммунизации людей против инфекции, распространяемой комарами. Также антигенная детерминанта может содержать иммуногенный эпитоп, такой как эпитоп, распознаваемый В-лимфоцитами и/или Т-лимфоцитами.
[0087] Согласно некоторым вариантам реализации белок CS является кодон-оптимизированным в целях повышения уровня экспрессии в организме субъекта. Оптимизация кодонов основана на необходимом аминокислотном составе, на общей оптимальной частоте использования кодонов у рассматриваемого субъекта, а также факторах, которых следует избегать, чтобы экспрессия была адекватной. К таким факторам можно отнести донорные или акцепторные сайты сплайсированного фрагмента, терминирующие кодоны, сигналы полиаденилирования (рА), GC- и АТ-богатые последовательности, внутренние ТАТА-боксы или любые другие аспекты, известные в данной области техники. Согласно некоторым вариантам реализации последовательность ДНК кодон-оптимизированного трансгена показана на Фиг.9 (SEQ ID NO:1, Р. yoelii) и Фиг.10 (SEQ ID NO:2, P. falciparum).
[0088] Согласно некоторым вариантам реализации плазмидным вектором на основе рекомбинантного аденовируса может быть один из следующих плазмидных векторов на основе рекомбинантного аденовируса, модифицированного последовательностью Р. falciparum: вектор на основе HVR1-модифицированного аденовируса (NANP-HVR1/PfCSP), сконструированный как показано на Фиг.2, с применением последовательности (NANP)n, кодирующей эпитоп, распознаваемый В-лимфоцитами (SEQ ID NO:60); вектор на основе аденовируса с модифицированным белком нити (NANP-Fib/PfCSP), сконструированный как показано на Фиг.4, с применением последовательности (NANP)n, кодирующей эпитоп, распознаваемый В-лимфоцитами (SEQ ID NO:60); вектор на основе аденовируса с модифицированными HVR1 и белком нити (NANP-HVRI/B-Fib/PfCSP), сконструированный как показано на Фиг.5, с применением последовательности (NANP)n, кодирующей эпитоп, распознаваемый В-лимфоцитами (SEQ ID NO:60); вектор на основе аденовируса с модифицированными HVR1 и pVII (NANP-HVR1/CD4-pVII/PfCSP), сконструированный как показано на Фиг.7, с применением последовательности (NANP)n, кодирующей эпитоп, распознаваемый В-лимфоцитами (SEQ ID NO:60) и последовательности EYLNKIQNSLSTEWSPCSVT, кодирующей эпитоп, распознаваемый CD4-лимфоцитами (SEQ ID NO:62); вектор на основе аденовируса с модифицированным белком нити и pVII (NANP-Fib/CD4-pVII/PfCSP), сконструированный как показано на Фиг.6, с применением эпитопа (NANP)n, распознаваемый В-лимфоцитами (SEQ ID NO:60) и последовательности EYLNKIQNSLSTEWSPCSVT, кодирующей эпитоп, распознаваемый CD4-лимфоцитами (SEQ ID NO:62); и вектор на основе аденовируса с модифицированными HVR1, белком нити и pVII (NANP-HVR1/Fib/CD4-pVII/PfCSP), сконструированный как показано на Фиг.8, с применением эпитопа (NANP)n, распознаваемый В-лимфоцитами (SEQ ID NO:60) и последовательности EYLNKIQNSLSTEWSPCSVT, кодирующей эпитоп, распознаваемый CD4-лимфоцитами (SEQ ID NO:62).
[0089] Согласно другим вариантам реализации плазмидным вектором на основе рекомбинантного аденовируса может быть один из следующих плазмидных векторов на основе рекомбинантного аденовируса, модифицированного последовательностью Р. yoelii: вектор на основе HVR1-модифицированного аденовируса (QGPGAP-HVR1/PyCS), сконструированный как показано на Фиг.2, с применением последовательности (QGPGAP)n, кодирующей эпитоп, распознаваемый В-лимфоцитами (SEQ ID NO:59); вектор на основе аденовируса с модифицированным белком нити (QGPGAP-Fib/PyCS), сконструированный как показано на Фиг.4, с применением последовательности(QGPGAP)n, кодирующей эпитоп, распознаваемый В-лимфоцитами (SEQ ID NO:59); вектор на основе аденовируса с модифицированными HVR1 и белком нити(QGPGAP-HVR1/B-Fib/PyCS), сконструированный как показано на Фиг.5, с применением последовательности (QGPGAP)n, кодирующей эпитоп, распознаваемый В-лимфоцитами (SEQ ID NO:59); вектор на основе аденовируса с модифицированными HVR1 и pVII (QGPGAP-HVR1/CD4-pVII/PyCS), сконструированный как показано на Фиг.7, с применением последовательности(QGPGAP)n, кодирующей эпитоп, распознаваемый В-лимфоцитами (SEQ ID NO:59) и последовательности YNRNIVNRLLGDALNGKPEEK, кодирующей эпитоп, распознаваемый CD4-лимфоцитами (SEQ ID NO:61); вектор на основе аденовируса с модифицированным белком нити и pVII (QGPGAP-Fib/CD4-pVII/PyCS), сконструированный как показано на Фиг.6, с применением эпитопа (QGPGAP)n, распознаваемый В-лимфоцитами (SEQ ID NO:59) и последовательности YNRNIVNRLLGDALNGKPEEK, кодирующей эпитоп, распознаваемый CD4-лимфоцитами (SEQ ID NO:61); и вектор а на основе аденовируса с модифицированными HVR1, белком нити и pVII (QGPGAP-HVR1/Fib/CD4-pVII/PyCS), сконструированный как показано на Фиг.8, с применением эпитопа (QGPGAP)n, распознаваемый В-лимфоцитами (SEQ ID NO:59) и последовательности YNRNIVNRLLGDALNGKPEEK, кодирующей эпитоп, распознаваемый CD4-лимфоцитами (SEQ ID NO:61).
[0090] Согласно другим вариантам реализации рекомбинантный аденовирус может быть получен с использованием одного из следующих плазмидных векторов на основе рекомбинантного аденовируса, модифицированного последовательностями Р. falciparum или Р. yoelii: NANP-HVRI/PfCSP или QGPGAP-HVR1/PyCS (Фиг.2), NANP-Fib/PfCSP или QGPGAP-Fib/PyCS (Фиг.4), NANP-HVR1/B-Fib/PfCSP или QGPGAP-HVR1/B-Fib/PyCS (Фиг.5), NANP-HVR1/CD4-pVII/PfCSP или QGPGAP-HVR1/CD4-pVII/PyCS (Фиг.7), NANP-Fib/CD4-pVII/PfCSP или QGPGAP-Fib/CD4-pVII/PyCS (Фиг.6), NANP-HVR1/Fib/CD4-pVII/PfCSP или QGPGAP-HVR1/Fib/CD4-pVII/PyCS (Фиг.8). Рекомбинантный аденовирус может быть получен описанными в настоящей заявке способами получения плазмидного вектора на основе рекомбинантного аденовируса, обладающего способностью экспрессировать рекомбинантный трансгенный белок (например, белок CS Plasmodium) в клетках млекопитающих.
[0091] Очистка рекомбинантного аденовируса может быть осуществлена при помощи известных способов очистки вирусов. Например, очистка 0,5-1,0 мл культуры вируса, полученной согласно способу получения белков рекомбинантного аденовируса путем заражения клеток насекомых (1×107клеток/чашку диаметром 10 см), таких как клетки AD293. Надосадочную жидкость культуры отбирают через несколько дней после инфицирования, и осадок, содержащий вирус, отделенный в результате центрифугирования, разводят буфером, таким как фосфатно-солевой буферный раствор (ФСБ). Полученную суспензию подвергают действию градиента сахарозы от 10 до 60%, затем центрифугируют (25000 об/мин в течение 60 минут при 4°С), в результате отбирают полосу с вирусом. Полученный вирус дополнительно разбавляют в ФСБ, после чего центрифугируют при тех же условиях, что и предыдущее центрифугирование, и полученные очищенный осадок, содержащий вирус хранят при 4°С в буфере, таком как ФСБ.
[0092] Еще один вариант реализации относится к фармацевтической композиции, по существу содержащей по меньшей мере один активный ингредиент. Согласно одному варианту реализации активный ингредиент указанной фармацевтической композиции может включать рекомбинантный аденовирус, который может быть получен методами генной инженерии, описанными в настоящей заявке. Более конкретно, активный ингредиент может быть рекомбинантным аденовирусом, содержащим модифицированные белки капсида и/или сердцевины, в которых часть гипервариабельного участка гексона (HVR), часть белка нити, часть белка pVII или их комбинации заменены по меньшей мере одним иммуногенным эпитопом белка circumsporozoite Plasmodium. В качестве альтернативы, в белок нити, HVR гексона или белок pVII можно встраивать один или более эпитопов белка circumsporozoite Plasmodium, распознаваемых В-лимфоцитами и/или Т-лимфоцитами. Указанный вектор на основе рекомбинантного аденовируса также содержит трансгенный белок или рекомбинантный трансгенный белок, который экспрессируется рекомбинантным аденовирусом после инфицирования одной или более клеток-хозяев. Указанным трансгенным белком или рекомбинантным трансгенным белком может быть белок circumsporozoite Plasmodium или малярийный антиген белка circumsporozoite Plasmodium, причем указанный малярийный антиген содержит по меньшей мере один иммуногенный эпитоп (например, распознаваемый В-лимфоцитами или Т-лимфоцитами) белка circumsporozoite Plasmodium.
[0093] Согласно некоторым вариантам реализации активным ингредиентом указанной фармацевтической композиции является рекомбинантный аденовирус, полученный из плазмидного вектора на основе рекомбинантного аденовируса, причем указанный вектор на основе рекомбинантного аденовируса является одним из следующих плазмидных вектором на основе рекомбинантного аденовируса, модифицированного последовательностями Р. falciparum или Р. yoelii: NANP-HVR1/PfCSP или QGPGAP-HVR1/PyCS (Фиг.2), NANP-Fib/PfCSP или QGPGAP-Fib/PyCS (Фиг.4), NANP-HVR1/B-Fib/PfCSP или QGPGAP-HVR1/B-Fib/PyCS (Фиг.5), NANP-HVR1/CD4-pVII/PfCSP или QGPGAP-HVR1/CD4-pVII/PyCS (Фиг.7), NANP-Fib/CD4-pVII/PfCSP или QGPGAP-Fib/CD4-pVII/PyCS (Фиг.6), NANP-HVR1/Fib/CD4-pVII/PfCSP или QGPGAP-HVR1/Fib/CD4-pVII/PyCS (Фиг.8). Указанные плазмидные векторы на основе рекомбинантного аденовируса способны к выработке рекомбинантных аденовирусов при их трансфекции в клетки (например, клетки AD293), причем указанный рекомбинантный трансгенный белок может экспрессироваться в клетках млекопитающих, включая клетки человека.
[0094] Фармацевтическая композиция, содержащая в качестве активного ингредиента описанный в настоящей заявке рекомбинантный аденовирус, при введении субъекту усиливает противомалярийное профилактическое действие, направленное против малярийного антигена, и снижает титр инфекционности, как описано далее в разделе «Примеры». Таким образом, рекомбинантный аденовирус можно применять для лечения малярии, ассоциированной с инфекционным поражением клеток и тканей-мишеней. Примеры клеток-мишеней, поражаемых при малярии, включают клетки крови, клетки печени, клетки почек, клетки легких, эпителиальные и мышечные клетки. Примеры тканей, содержащих указанные клетки, включают легкие, печень, почки, головной мозг, артерии и вены, желудок, кишечник, мочеиспускательный канал, кожу и мышцы.
[0095] Согласно некоторым аспектам указанная фармацевтическая композиция может усиливать противомалярийное профилактическое действие, направленное против антигенов возбудителя, например, антигенов малярийного плазмодия, таких как поверхностные антигены спорозоита (белок circumsporozoite (CSP) и родственный тромбоспондину адгезивный белок (TRAP)) малярийных плазмодиев, поверхностный мембранный белок мерозоита (MSPI), малярийный S-антиген, секретируемый эритроцитами, инфицированными малярией, и белок-1 Р. falciparum на мембране эритроцитов (PfEMPI)), присутствующие на поверхности эритроцитов, инфицированных малярией. Указанная фармацевтическая композиция может усиливать профилактическое действие в отношении заражения любым плазмодием, выбранным из известных видов Plasmodium (P), например, Р. falciparum, P. malariae, P. ovale, P. vivax, P. knowlesi, P. berghei, P. chabaudi и Р. yoelii, путем снижения титра инфекционности. При введении указанной фармацевтической композиции субъекту снижение титра инфекционности может приводить к увеличению периода выживаемости, стадии ремиссии или периода выживаемости без инфекции, увеличению уровня выживаемости или выживаемости без инфекции по сравнению с субъектами, которым не вводили указанную фармацевтическую композицию. Таким образом, согласно некоторым аспектам указанную фармацевтическую композицию можно применять в качества профилактического или лечебного средства при малярии, вызванной такими патогенами, как Plasmodium. Согласно дополнительным аспектам указанная фармацевтическая композиция применима в качества профилактического или лечебного средства при осложнениях малярии, вызванной такими патогенами, как Plasmodium.
[0096] Профилактическое действие рекомбинантного аденовируса согласно настоящему изобретению у субъекта можно обеспечить, например, путем введения фармацевтической композиции, содержащей рекомбинантный аденовирус с модифицированным капсидом согласно настоящему изобретению и вспомогательные вещества для введения препарата, позвоночным, в частности млекопитающим, включая человека, внутримышечным (в/м), подкожным (п/к), внутрикожным (в/к), внутрикожным (в/д), внутрибрюшинным (в/б), назальным или ингаляционным путем, и последующей многократной вакцинации позвоночных фармацевтической композицией, содержащей в качестве активного ингредиента рекомбинантный аденовирус, описанный в настоящей заявке. Чтобы оценить профилактическое действие, можно сравнивать уровень выживаемости, стадии ремиссии или выживаемости без инфекции у субъектов, многократно вакцинированных указанной фармацевтической композицией и впоследствии инфицированных целевым патогенном (например, выбранным из видов Plasmodium), с уровнем выживаемости, стадии ремиссии или выживаемости без инфекции у субъектов, не получавших указанную фармацевтическую композицию.
[0097] Согласно некоторым вариантам реализации указанная фармацевтическая композиция может содержать фармацевтически эффективное количество рекомбинантного аденовируса с модифицированным капсидом и/или сердцевиной, как описано в настоящей заявке, и фармацевтически приемлемую основу, которая дополнительно описана далее.
[0098] Еще один вариант реализации относится к вакцине, по существу содержащей по меньшей мере один активный ингредиент. Согласно одному варианту реализации активный ингредиент указанной вакцины может включать рекомбинантный аденовирус, полученный из плазмидного вектора на основе рекомбинантного аденовируса, описанного в настоящей заявке. Более конкретно, активный ингредиент может быть рекомбинантным аденовирусом, содержащим модифицированные белки капсида или сердцевины, в которых часть гипервариабельного участка гексона (HVR), часть белка нити, часть белка pVII или их комбинации заменены по меньшей мере одним иммуногенным эпитопом белка circumsporozoite Plasmodium. В качестве альтернативы, в белок pVII, белок нити или HVR гексона, или в их сочетание, можно встраивать по меньшей мере один иммуногенный эпитоп белка circumsporozoite Plasmodium. Согласно некоторым вариантам реализации активный ингредиент указанной вакцины можно получить из плазмидного вектора на основе рекомбинантного аденовируса, проиллюстрированного на Фиг.2-8, например, NANP-HVR1/PfCSP или QGPGAP-HVR1/PyCS (Фиг.2), NANP-Fib/PfCSP или QGPGAP-Fib/PyCS (Фиг.4), NANP-HVR1/B-Fib/PfCSP или QGPGAP-HVR1/B-Fib/PyCS (Фиг.5), NANP-HVR1/CD4-pVII/PfCSP или QGPGAP-HVR1/CD4-pVII/PyCS (Фиг.7), NANP-Fib/CD4-pVII/PfCSP или QGPGAP-Fib/CD4-pVII/PyCS (Фиг.6), NANP-HVR1/Fib/CD4-pVII/PfCSP или QGPGAP-HVR1/Fib/CD4-pVII/PyCS (Фиг.8).
[0099] Согласно некоторым аспектам указанная вакцина при ее введении субъекту первоначально содержит рекомбинантный аденовирус, обладающий одной или более антигенных частей белка CS Plasmodium (например, эпитоп, распознаваемый В-лимфоцитами, эпитоп, распознаваемый Т-лимфоцитами или оба), встроенными по меньшей мере в одну часть белка капсида или сердцевины, или заменяющими указанную часть. Затем указанная вакцина может экспрессировать рекомбинантный трансгенный белок, причем указанным рекомбинантным трансгенным белком является белок CS Plasmodium, содержащий эпитоп, распознаваемый В-лимфоцитами, эпитоп, распознаваемый Т-лимфоцитами или оба. Антигенные части белка CS Plasmodium обнаруживаются в рекомбинантном трансгенном белке, и модифицированные белки капсида или сердцевины способствуют развитию или усилению приобретенного гуморального иммунитета, клеточного иммунитета или обоих, как описано далее в разделе «Примеры». Таким образом, согласно некоторым аспектам рекомбинантный аденовирус, описанный в настоящей заявке, применим в качества вакцины для обеспечения возникновения или усиления гуморального иммунитета, клеточного иммунитета или обоих типов.
[00100] Согласно дополнительным вариантам реализации указанная вакцина может усиливать профилактическое действие, направленное против инфекционного антигена, например малярийных антигенов, таких как поверхностные белки спорозоита малярийного плазмодия (CSP и TRAP), поверхностный мембранный белок мерозоита MSPI, малярийный S-антиген, секретируемый эритроцитами, инфицированными малярией, белок PfEMPI, присутствующий на поверхности эритроцитов, инфицированных малярией, богатый серином антиген (SERA), богатый тирозином кислый матричный белок (TRAMP) и антиген-1 апикальной мембраны (AMAI). Кроме того, снижение титра инфекционности (например, снижение титра инфекционности вируса), возникающее вследствие введения вакцины, описанной в настоящей заявке, может приводить к увеличению периода выживаемости, стадии ремиссии или периода выживаемости без инфекции, увеличению уровня выживаемости, ремиссии или выживаемости без инфекции по сравнению с субъектами, которым не вводили указанную фармацевтическую вакцину. Таким образом, согласно некоторым аспектам указанная вакцина также применима в качества профилактического или лечебного средства против малярии, вызванной такими патогенами, как Plasmodium. Согласно дополнительным аспектам указанная вакцина также применима в качества профилактического или лечебного средства при борьбе с осложнениями малярии, вызванной такими патогенами как Plasmodium.
[00101] Вакцина, описанная в настоящей заявке, может содержать терапевтически эффективное количество рекомбинантного аденовируса, описанного в настоящей заявке, и дополнительно содержать фармацевтически приемлемую основу в соответствии со стандартным способом. Примеры приемлемых основ включают физиологически приемлемые растворы, такие как стерильный физиологический раствор и стерильный буферный раствор.
[00102] Согласно некоторым вариантам реализации указанную вакцину или фармацевтическую композицию можно применять в сочетании с фармацевтически приемлемым количеством адъюванта для усиления противомалярийного действия. В качестве адъюванта можно применять любой иммунологический адъювант, который может стимулировать иммунную систему и увеличивать ответ на вакцину и сам по себе не обладает каким-либо специфическим антигенным действием. Многие иммунологические адъюванты имитируют эволюционно консервативные молекулы, называемые патоген-ассоциированными молекулярными структурами (РАМ3) и распознаются комплексом иммунных рецепторов, таких как Toll-подобные рецепторы (TLR). Примеры адъювантов, которые можно применять согласно вариантам реализации настоящего изобретения, включают полный адъювант Фрейнда, неполный адъювант Фрейнда, двухцепочечную РНК (лиганд TLR3), липополисахариды (LPS), аналоги LPS, такие как монофосфорил-липид A (MPL) (лиганд TLR4), флагеллин (лиганд TLR5), липопротеины, липопептиды, одноцепочечные ДНК, одноцепочечные ДНК, аналоги имидазохинолина (лиганды TLR7 и TLR8), CpG ДНК (лиганд TLR9), адъювант Риби (монофосфорил-липид А/ трегалоза-дикориноиголат), гликолипиды (аналоги α-GalCer), неметилированные CpG-островки, масляные эмульсии, липосомы, виросомы, сапонины (активные фракции сапонина, такого как QS21), мурамил-дипептид, квасцы, гидроксид алюминия, сквален, БЦЖ, цитокины, такие как GM-CSF (гранулоцито-макрофаго-колониестимулирующий фактор) и ИЛ-12, хемокины, такие как MIP 1-α и RANTES (хемокин, выделяемый Т-лимфоцитами при активации), N-ацетилмурамин-L-аланил-D-изоглутамин (MDP), тимозин αl и MF59. Применяемое количество адъюванта может быть выбрано должным образом в зависимости от выраженности симптомов, таких как размягчение кожи, боль, эритема, лихорадка, головная боль и боль в мышцах, которые могут проявляться как часть иммунного ответа у людей или животных после введения вакцины данного типа.
[00103] Согласно некоторым вариантам реализации, вакцину или фармацевтическую композицию, описанные в настоящей заявке, можно применять в сочетании с другими известными фармацевтическими препаратами, такими как пептиды, способствующие развитию иммунного ответа, и противомикробные препараты (синтетические противомикробные препараты). Указанная вакцина или фармацевтическая композиция также может содержать другие лекарственные или вспомогательные вещества. Примеры лекарственных или вспомогательных веществ, которые можно применять в сочетании с вакциной или фармацевтической композицией, описанными в настоящей заявке, включают лекарственные вещества, которые способствуют захвату рекомбинантного аденовируса или рекомбинантного трансгенного белка согласно настоящему изобретению клетками, липосомы и другие лекарственные и/или вспомогательные вещества, облегчающие трансфекцию (например, фторуглеродные эмульгаторы, со-хелаторы, трубочки, частицы золота, биодеградируемые микросферы и катионные полимеры).
[00104] Согласно некоторым вариантам реализации, количество активного ингредиента, содержащееся в вакцине или фармацевтической композиции, описанных в настоящей заявке, может быть выбрано из широкого диапазона концентраций, единиц вирусных частиц (VPU), бляшкообразующих единиц (БОЕ), процентов по массе на объем (м/о %) или других количественных мер для оценки количества активного ингредиента, при условии, что это терапевтически или фармацевтически эффективное количество. Дозу вакцины или фармацевтической композиции можно надлежащим образом выбирать из широкого диапазона в зависимости от желаемого терапевтического действия, способа введения (пути введения), периода введения, возраста и пола пациента, других его состояний, и др.
[00105] Согласно некоторым аспектам при введении человеку рекомбинантного аденовируса в качестве активного ингредиента указанной вакцины или фармацевтической композиции доза указанного рекомбинантного аденовируса может составлять приблизительно от 102 до 1014 PFU (бляшкообразующих единиц), предпочтительно от 105 до 1012 PFU, и более предпочтительно от 106 до 1010 PFU на пациента, рассчитываемая как PFU рекомбинантного вируса.
[00106] Согласно некоторым аспектам при введении субъекту рекомбинантного аденовируса в качестве активного ингредиента указанной вакцины или фармацевтической композиции доза может быть выбрана выбирать из широкого диапазона в пересчете на количество экспрессируемой ДНК, вводимой хозяину с вакциной, или на количество транскрибируемой РНК. Также доза зависит от эффективности промоторов транскрипции и трансляции, применяемых в любом из используемых векторов переноса.
[00107] Согласно некоторым вариантам реализации, вакцину или фармацевтическую композицию, описанные в настоящей заявке, можно вводить путем прямых инъекций суспензии рекомбинантного аденовируса, приготовленной путем разведения указанного рекомбинантного аденовируса в ФСБ (фосфатно-солевом буферном растворе) или физиологическом растворе, в локальный участок организма (например, в ткань легкого, печени, мышцы или головного мозга), путем назального введения или ингаляции, или путем внутрисосудистого (в/с) (например, внутриартериального, внутривенного или в воротную вену), подкожного (п/к), внутрикожного (в/к), внутрикожного (в/к) или внутрибрюшинного (в/б) введения. Вакцину или фармацевтическую композицию согласно настоящему изобретению можно вводить более одного раза. Более конкретно, после первичного введения можно проводить одну или более дополнительных вакцинаций в качестве поддерживающей вакцинации. Одна или более вторичных вакцинаций могут усиливать желаемое действие. После введения указанной вакцины или фармацевтической композиции можно проводить вторичную вакцинацию фармацевтической композицией, содержащей рекомбинантный аденовирус, описанный в настоящей заявке.
[00108] Согласно дополнительным вариантам реализации, применение различных других адъювантов, лекарственных или вспомогательных веществ в сочетании с вакциной согласно настоящему изобретению, как описано выше, может усиливать терапевтическое действие, достигаемое при введении указанной вакцины или терапевтической композиции. Фармацевтически приемлемая основа может содержать следовые количества вспомогательных веществ, таких как вещества, повышающие изотоничность и химическую стабильность. Такие вспомогательные вещества должны быть нетоксичными для человека или других млекопитающих в применяемых дозах и концентрациях, и примеры таких вспомогательных веществ включают буферы, например фосфорную кислоту, лимонную кислоту, янтарную кислоту, уксусную кислоту и другие органические кислоты, а также их соли; антиоксиданты, например аскорбиновую кислоту; низкомолекулярные пептиды (например, менее 10 остатком аминокислот) (например, полиаргинин и трипептид), белки (например, сывороточный альбумин, желатин и иммуноглобулин); аминокислоты (например, глицин, глутаминовая кислота, аспарагиновая кислота и аргинин): моносахариды, дисахариды и другие углеводы (например, целлюлоза и ее производные, глюкоза, манноза и декстрин), хелатирующие агенты (например, ЭДТА/этилендиаминтетрауксусная кислота/); сахароспирты (например, маннитол и сорбитол); противоионы (например, натрий); неионные поверхностно-активные вещества (например, полисорбат и полоксамер); и полиэтиленгликоль (ПЭГ).
[00109] Вакцину или фармацевтическую композицию, содержащие рекомбинантный аденовирус, описанный в настоящей заявке, можно хранить в форме водного раствора или лиофилизированного препарата в контейнере с однократной дозой или несколькими дозами, таком как запаянная ампула или флакон.
[00110] Согласно еще одному варианту реализации предложен способ профилактики малярии или способ лечения малярии, включающий введение эффективного количества вакцины, рецептуры или фармацевтической композиции на основе рекомбинантного аденовируса. Согласно настоящему изобретению также предложен способ стимуляции иммунного ответа, включающий введение субъекту эффективного количества вакцины, рецептуры или фармацевтической композиции на основе рекомбинантного аденовируса, или их сочетания. Субъекты могут включать людей, животных (таких как млекопитающие, птицы, рептилии, рыбы и амфибии) или любых других субъектов, которые могут быть инфицированы малярийным плазмодием. Малярийный плазмодий может включать плазмодия Plasmodium, любого выбранного из видов Plasmodium (Р), например Р. falciparum, P. malariae, P. ovate, Р. vivax, P. knowlesi, P. berghei, P. chabaudi и Р. yoelii.
[00111] Согласно некоторым вариантам реализации рекомбинантный аденовирус, описанный в настоящей заявке, может один или вместе с фармацевтически приемлемой основой входить в состав вакцины, рецептуры или фармацевтической композиции, и его можно вводить субъекту. Путем введения может быть, например, любой из путей введения, перечисленных выше. Фармацевтически приемлемую основу для применения в рамках настоящего изобретения можно надлежащим образом выбрать из основ, обычно применяемых в данной области техники в зависимости от формы фармацевтической композиции, которую требуется получить. Например, если фармакологическую композицию готовят в форме водного раствора, в качестве основы можно применять очищенную воду (стерильную воду) или физиологический буферный раствор. Когда фармацевтическую композицию создают в форме других пригодных растворов, в качестве основы можно применять органические эфиры, которые можно вводить в организм, такие как гликоль, глицерин и оливковое масло. Указанная композиция может содержать стабилизаторы, вспомогательные и другие вещества, обычно применяемые в данной области техники, и в частности в области производства вакцин.
[00112] Согласно дополнительным вариантам реализации количество рекомбинантного аденовируса, применяемое в вакцине, лекарственной форме или фармацевтической композиции, может быть соответственно выбрано из широкого диапазона концентраций, VPU, PFU, процентов по массе на объем (м/о %) или других количественных мер для оценки количества активного ингредиента. Согласно некоторым аспектам подходящий диапазон количества рекомбинантного вируса в указанной композиции предпочтительно составляет приблизительно от 0,0002 до приблизительно 0,2 (м/о %), а более предпочтительно - от 0,001 до 0,1 (м/о %). Способ введения вакцины, рецептуры или фармацевтической композиции на основе рекомбинантного аденовируса согласно некоторым вариантам реализации можно соответствующим образом выбрать в зависимости от лекарственной формы, возраста и пола пациента, других условий, таких как тяжесть заболевания. Подходящей лекарственной формой является форма для парентерального введения, такая как растворы для инъекций, капли, назальные капли и средства для ингаляции. Когда указанную композицию создают в форме растворов для инъекций или капель, композицию вводят внутривенно и смешивают со средствами для восполнения объема жидкости, такими как раствор глюкозы или аминокислот, как предписано, или могут вводить внутримышечно (в/м), внутрикожно (в/к), подкожно (п/к), внутрикожно (в/д) или внутрибрюшинно (в/б).
[00113] Согласно другим вариантам реализации суточная доза вакцины, рецептуры или фармацевтической композиции на основе рекомбинантного аденовируса может варьировать в зависимости от состояния субъекта, массы тела, возраста, пола и др. Согласно некоторым аспектам доза рекомбинантного аденовируса составляет приблизительно от 0,001 до 100 мг на кг массы тела в день. Вакцину, рецептуру или фармацевтическую композицию согласно настоящему изобретению можно вводить один или более раз в день.
[00114] Согласно дополнительным вариантам реализации при введении человеку рекомбинантного аденовируса в качестве активного ингредиента вакцины, рецептуры или фармацевтической композиции вводимая доза рекомбинантного аденовируса приблизительно соответствует от 102 до 1014 PFU, предпочтительно от 105 до 1012 PFU, и более предпочтительно от 106 до 1010 PFU на одного пациента, рассчитываемая как PFU частиц рекомбинантного вируса. Вакцину согласно настоящему изобретению следует вводить в соответствии с принципами Надлежащей медицинской практики, с учетом клинического состояния (например, состояние, которое требуется предупреждать или лечить) каждого пациента, участка введения вакцины, содержащей аденовирус, ткани-мишени, способа введения, режима дозирования и других факторов, известных специалистам в данной области техники. Следовательно, надлежащую дозу вакцины, описанной в настоящей заявке, определяют с учетом всего вышеперечисленного.
[00115] Еще один вариант реализации настоящего изобретения относится к способу лечения или профилактики малярии у субъекта, включающему введение иммунологического или терапевтического количества вакцины против малярии, содержащей рекомбинантный аденовирус. Рекомбинантный аденовирус в составе указанной вакцины против малярии может содержать антигенную детерминанту Plasmodium, а также может содержать один или более модифицированных белков капсида или сердцевины. Иммунологически, фармакологически или терапевтически приемлемым количеством может быть любое подходящее количество, при введении которого генерируется эффективный иммунный ответ на одну или более антигенных частей белка (CS) (т.е. трансген, эпитоп, распознаваемый В-лимфоцитами, или эпитоп, распознаваемый CD4+Т-лимфоцитами), причем предотвращается развитие малярии, или уменьшается ее тяжесть.
[00116] Когда конкретный субъект впервые подвергается воздействию или «примируется» вектором на основе аденовируса, его иммунная система вырабатывает нейтрализующие антитела против указанного конкретного вектора. Иммунный ответ на аденовирус главным образом направлен против белков капсида. Следовательно, последующее введение того же вектора на основе аденовируса, или «поддерживающая вакцинация», может снижать эффективность экспрессии трансгена. Следовательно, согласно некоторым вариантам реализации способ лечения или профилактики малярии, описанный выше, может включать этап примирования с применением первого вектора на основе рекомбинантного аденовируса, а затем один или более этапов поддерживающей вакцинации с применением одного или более других векторов на основе рекомбинантного аденовируса. Данный способ можно применять у субъектов, которые еще не подвергались воздействию аденовируса дикого типа, или у субъекта, который ранее подвергался действию вектора на основе аденовируса дикого типа, причем вектор на основе рекомбинантного аденовируса на этапе примирования применяют для того, чтобы преодолеть существующий иммунитет на аденовирус. Дополнительные варианты реализации и примеры приведены ниже.
Аденовирус в качестве вектора
[00117] Аденовирусы представляют собой ДНК-содержащие вирусы, лишенные липопротеиновой оболочки, которые содержат комплекс белков капсида (описаны ниже) и вирусный геном, который получил широкое применение в качестве средств доставки одного или более терапевтических или антигенных трансгенов в клетки разных типов in vitro и in vivo. Существует много серотипов аденовирусов. Среди известных серотипов аденовирусов в качестве вектора для трансдукции чужеродных генов преимущественно применяют серотип 5 (Ad5) благодаря его сильной инфекционности в условиях in vivo (Abbink et al. 2007). Экспрессию антигенного трансгена можно контролировать при помощи любого промотора или энхансера, известного в данной области. Промоторы, которые можно применять для контроля экспрессии генов, включают промотор цитомегаловируса (CMV) предранней стадии, ранний промотор вируса обезьян 40 (SV40), промотор клеточного фактора элонгации полипептидных цепей-1 альфа (EF1), промотор вируса саркомы Рауса (RSV) и тетрациклин-регулируемый промотор (TR), но не ограничиваются ими. Сигнал полиаденилирования (рА) после кодирующей последовательности также можно применять для эффективной транскрипции и трансляции. Вектор на основе рекомбинантного аденовируса, описанный в настоящей заявке, может быть с дефективной репликацией, может содержать делецию по меньшей мере в участке Е1 генома аденовируса, поскольку участок Е1 необходим для процессов репликации, трансляции и упаковки. Согласно некоторым аспектам также можно удалить участки Е2, Е3 и/или Е4. Согласно дополнительным аспектам для более эффективной трансляции можно применять консенсусную последовательность Козака (Kozak 1987).
[00118] Система на основе аденовируса являет собой перспективный вектор для разработки рекомбинантных вакцин по нескольким причинам. Одна из причин заключается в том, что векторы на основе рекомбинантных аденовирусов инфицируют большинство типов клеток млекопитающих (как репликативных, так и нерепликативных), включая типы клеток человека и мыши, но не ограничиваясь ими. Так, один и тот же вектор можно с успехом применять и в моделях на мышах, и в клинических исследованиях с участием человека. Вторая причина заключается в том, что любая переданная генетическая информация остается вне хромосом, и удается избежать инсерционного мутагенеза и изменения генотипа клеток (Crystal 1995). Еще одна причина заключается в том, что трансген остается неизменным после успешных циклов репликации вирусов. Другие преимущества применения аденовируса включают то, что рекомбинантный аденовирус: 1) обладает высоко стабильным вирионом, 2) хорошо переносится, 3) его титр может возрастать до высоких уровней, 4) может вмещать крупные трансгены, 5) обладает геномом, который детально изучался в течение многих лет, и известна полная последовательность ДНК нескольких серотипов, что облегчает манипуляции с геномом Ad путем технологии рекомбинантных ДНК (Graham and Prevec 1992).
[00119] Согласно одному варианту реализации платформу для создания вакцин на основе аденовируса применяют в качестве вирусного вектора для разработки вакцины, которая направлена против преэритроцитарной формы малярийного плазмодия и обеспечивает защиту от малярии. Было показано, что среди векторов на основе известных рекомбинантных аденовирусов (Rodrigues et al. 1997, Bruna-Romero et al. 2001, Anderson et al. 2004, Tao et al. 2005) для получения вакцины против малярии подходит вектор на основе аденовируса, поскольку он может вызывать эффективный защитный клеточный иммунный ответ на преэритроцитарные формы малярийного плазмодия (Rodrigues et al. 1997). Малярийный плазмодий может принадлежать к любому семейству Plasmodium. Согласно некоторым вариантам реализации целевым плазмодием может быть Р. yoelii или Р. falciparum.
Векторы на основе аденовируса, экспрессирующие в качестве трансгена PyCS, вызывают ответ CD8+Т-лимфоцитов
[00120] Аденовирус представляет собой перспективный вектор для индукции выраженного иммунитета против малярии, опосредуемого CD8+Т-лимфоцитами (Rodrigues et al. 1997, Rodrigues et al. 1998). Иммуногенность рекомбинантного аденовируса, экспрессирующего белок CS P. yoelii (малярийного плазмодия, специфичного для грызунов), AdPyCS, определяли при помощи модели малярии у грызунов. Иммунизация мышей AdPyCS вызывает развитие полного иммунитета у значительного процента мышей, что препятствует возникновению паразитемии (Rodrigues et al. 1997). Указанный защитный эффект главным образом опосредуется CD8+Т-лимфоцитами, о чем свидетельствует истощение популяции Т-лимфоцитов, и что подтверждается тем фактом, что AdPyCS не мог вызвать высокие титры антител против малярийного плазмодия.
[00121] Чтобы количественно оценить инфекционность аденовируса с модифицированным капсидом, в челночный вектор можно включить кассету экспрессии зеленого флуоресцентного белка (GFP) и сайты клонирования для трансгена. Полученный в результате челночный вектор (GFP/pShuttle-CMV) содержит двойные промоторы pCMV и SV40pAs для трансгена, а также GFP из pmaxGFP («Amaxa», Германия). Оптимизированный фрагмент PyCS встраивали в сайты KpnI и HindIII вектора GFP/pShuttle-CMV.
[00122] Иммуногенность Ad(PyCS+GFP) определяли путем измерения величины CS-специфичного ответа CD8+-Т-лимфоцитов и уровня защитного иммунитета против плазмоида в печеночной стадии. Введение Ad(PyCS+GFP) разными путями при оптимальной дозе 1010 вирусных частиц (в.ч.), вызывало такой же характер противомалярийных защитных ответов, которые, как было показано, вызывал AdPyCS, причем при п/к и в/м пути введения развивался наиболее сильный ответ, приводящий к наибольшей степени угнетения печеночной стадии плазмодия у мышей, инфицированных живыми спорозоитами Р. yoelii. Данный факт показывает, что в качестве вакцины Ad(PyCS+GFP) ведет себя аналогично AdPyCS (Rodrigues et al. 1997), и возможно применим при определении тропизма AdPyCS in vivo.
Белки капсида и сердцевины аденовируса
[00123] Описанные выше исследования подтверждают, что векторы на основе рекомбинантного аденовируса, экспрессирующие белок CS, обеспечивают сильный клеточный иммунный ответ, опосредуемый CD8+ Т-лимфоцитами, но не вызывают заметного гуморального ответа. Соответственно, поскольку гуморальный ответ на аденовирус дикого типа часто может быть связан с белками капсида, были сконструированы векторы на основе рекомбинантного аденовируса с модифицированными белками капсида и сердцевины, чтобы 1) усилить гуморальный иммунитет за счет активации В-лимфоцитов, 2) усилить гуморальный иммунитет за счет активации Т-лимфоцитов и 3) преодолеть существующий иммунитет на аденовирусы.
[00124] Аденовирус представляет собой ДНК-содержащий вирус, лишенный липопротеиновой оболочки, с «оголенной» двухцепочечной ДНК, икосаэдрической формы с 20 гранями в виде равносторонних треугольников. Капсид аденовируса состоит из 252 капсомеров, из которых 240 - это тримеры гексона, а 12 - пентамеры пентонов. Белок нити, который отходит от каждого основания пентона, обусловливает присоединение к клетке-хозяину за счет взаимодействия с рецепторами клетки. Вторичное взаимодействие возникает между мотивом RGD (Асп-Арг-Гли) в основании пентона с αvβ3, αvβ5 и аналогичными интегринами, облегчающими последующую интернализацию аденовируса в клетку (Mathias et al. 1994, Wickham et al. 1993). Большинство аденовирусов используют рецепторы к Коксэки-аденовирусу (CAR), в качестве рецептора клеток (Bergelson et al. 1997). Кроме того, было показано, что молекулы МСН (Главный комплекс гистосовместимости) I класса, VCAM (молекула адгезии сосудистого эндотелия 1 типа) и гепаран сульфат опосредуют присоединение и вход Ad5 (Chu et al. 2001, Hong et al. 1997). После входа путем эндоцитоза Ad5 быстро выходит из внутриклеточных компартментов в цитозоль (Meier and Greber 2003, Leopold and Crystal 2007). Затем вирион перемещается в ядро при помощи микротрубочек. Белок нити «сбрасывается», как ранний белок капсида, в ходе данного процесса (Nakano et al. 2000, Hong et al. 2003). Аденовирусы разных серотипов демонстрируют разные паттерны миграции (Miyazawa et al. 1999, Miyazawa et al. 2001). Изменение или модификация белка нити может влиять на миграцию, что может быть особенно важно для переработки и представления антигена после инфицирования антиген-презентирующей клетки (АПК).
[00125] Белок нити аденовируса представляет собой тример, который подразделяют на хвостовой, стволовой и головной домен белка нити (Henry et al. 1994, Rux and Burnett 2004, Chroboczek et al. 1995). Известна третичная структура головного домена, и в совокупности с исследованиями мутагенеза указанные исследования позволяют визуализировать области, участвующие в CAR-взаимодействии и тримеризации (Kirby et al. 1999, Xia et al. 1995). Стволовые отростки белка нити, отходящие от вириона, и головка белка нити содержат домен взаимодействия с рецепторами к Коксэки-вирусу и аденовирусу (CAR) (Roelvink et al. 1999, Bewley et al. 1999). Сайт связывания CAR в головке белка нити состоит первично из остатков из петли АВ и петли CD, а вторично распространяется к FG и петле HI, а также к β-складчатостям В, Е и F (Roelvink et al. 1999, Bewley et al. 1999). Петля HI является наиболее хорошо изученным сайтом встраивания в головке белка нити (Worgall et al. 2004, Mizuguchi and Hayakawa 2004, Koizumi et al. 2003, Belousova et al. 2002, Noureddini and Curiel 2005, Nicklin et al. 2001), и встраивание эпитопа в петлю HI (остатки 543 и 544) привело к образованию белка с иммунитетом против такого эпитопа (Krause et al. 2006). Следовательно, иммунодоминантный полученный из CS эпитоп, распознаваемый В-лимфоцитами, сначала встраивали в петлю HI белка нити.
[00126] Гексон представляет собой наиболее многочисленный белок в капсиде аденовируса; его количество составляет 720 копий на вирион. В зрелом вирусе гексон существует в форме гомотримерных капсомеров, которые образуют собой фасетки икосаидрического вириона (Rux and Burnett 2004). Была получена кристаллическая структура гексонов аденовирусов 2 и 5 серотипов (Ad2 и Ad5), что выявило сложную молекулярную архитектуру (Athapilly et al. 1994, Roberts et al. 1986, Rux and Burnett 2000). Основание каждой мономерной субъединицы состоит из двух мотивов бета-складчатостей, которые присутствуют в белках капсида многих икосаидрических вирусов. Три длинных петли (DE1, FG1 и FG2) проходят от структуры основания и образуют опорную область каждой молекулы (Rux and Burnett 2004). Последовательности в указанных петлевых доменах выдаются над поверхностью капсида, образуя внешнюю часть вириона. Сопоставление аденовирусов разных серотипов показывает, что последовательности, расположенные на внешней стороне капсида, мало консервативны, как по длине, так и по последовательности аминокислот (Crawford-Miksza and Schnurr 1996). Кроме того, было показано, что последовательности, расположенные в указанных мало консервативных доменах, именуемых гипервариабельными участками (HVR), содержат детерминанты, к которым образуются серотип-специфичные антитела (Тор 1975, Rux and Burnett 2000, Top et al. 1971).
[00127] На основании сопоставления ранних последовательностей HVR были идентифицированы во всей молекуле гексона (Crawford-Miksza and Schnurr 1996, Roberts et al. 2006). Поскольку HVR мало консервативны при сравнении разных серотипов и, по-видимому, не участвуют в поддержании структурной целостности гексона, можно вносить в указанные домены небольшие изменения, не оказывая влияния на жизнеспособность вируса (Rux and Burnett 2000). Например, в HVR2, HVR3, HVR5, HVR6 и HVR7 можно встраивать гексагистидиновый маркер, не снижая жизнеспособности вируса (Wu et al. 2005). Так HVR гексонов часто применяют в качестве мишеней для эффективной индукции ответа антител на пептиды, расположенные в HVR гексона (Worgall et al. 2005, Crompton et al. 1994). В связи с малой консервативностью HVR5 гексона между серотипами в отношении длины и его расположением ближе всего к наружной поверхности капсида аденовируса (Rux and Burnett 2000, Crawford-Miksza and Schnurr 1996) он первоначально был выбран в качестве сайта для встраивания эпитопа. Также кристаллическая структура гексона показывает, что HVR5 представляет собой гибкую петлю на поверхности капсида, что указывает на способность HRV5 вместить относительно крупные пептиды без нарушения структурной целостности капсида (Roberts et al. 1986). Недавно были идентифицированы гексон-специфичные эпитопы, распознаваемые CD4+ и CD8+-лимфоцитами (Leen et al. 2008), и ответ CD4+Т-клеток сосредоточен у человека против консервативных остатков в белке гексоне (Onion et al. 2007, Heemskerk et al. 2006).
[00128] Сердцевина аденовируса состоит из вирусного генома и четырех белков сердцевины. Терминальный белок (ТР) ковалентно связан с 5'-концом каждой линейной цепи ДНК вируса в двух копиях вириона. Три других белка сердцевины - mu (µ), V (pV) и VII (pVII) - связаны с ДНК вируса нековалентно и неспецифически. Основным белком сердцевины является pVII, его количество составляет приблизительно 700-800 копий на вирион, и он служит гистоноподобным центром, вокруг которого накручивается ДНК вируса, образуя нуклеосомную структуру.
Модификация белков капсида аденовируса для повышения гуморального иммунитета
[00129] Согласно некоторым вариантам реализации описаны векторы на основе аденовируса со спорозоитами (CS), содержащие иммунодоминантный белок CS в качестве эпитопа, распознаваемого В-лимфоцитами, в белке капсида аденовируса (встроенный в гексон или белок нити). Трансген может находиться под контролем такого промотора как CMV в целях повышения клеточного и гуморального иммунного ответа на белок CS.
[00130] Центральная область повторов представляет собой консервативную структуру белка CS, характерную для всех видов Plasmodium, и было показано, что антитело на такую повторяющуюся последовательность обладает нейтрализующей активностью в отношении спорозоитов. Примеры повторяющихся последовательностей в белке CS Plasmodium включают повтор (NANP)n (P. falciparum; SEQ ID NO:60), повтор ANGAGNQPG (P. vivax; SEQ ID NO:63) и повтор NAAG (P. malariae; SEQ ID NO:64), которые можно встраивать в белки капсида аденовируса. Согласно некоторым вариантам реализации в HVR1 гексона аденовируса 5 серотипа можно встраивать четыре или более повторов PfCSP - (NANP)n (SEQ ID NO:60). Согласно некоторым вариантам реализации в HVR1 гексона аденовируса 5 серотипа можно встраивать два, четыре, шесть, восемь, десять, четырнадцать, шестнадцать, восемнадцать, двадцать, двадцать два, двадцать четыре, двадцать шесть или двадцать восемь повторов PfCSP - (NANP)n (SEQ ID NO:60; n=2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28). Повторяющуюся последовательность (NANP)n можно дополнительно встраивать в петлю HI белка нити.
[00131] Согласно некоторым вариантам реализации были картированы иммунодоминантные эпитопы, распознаваемые нейтрализующими В-лимфоцитами, встраиваемые в CS, в целях разработки улучшенных вакцин на основе аденовируса с белком CS. У мышей, вакцинированных рекомбинантным белком CS P. yoelii CS (PyCS), генерировались высокие титры антител против двух основных иммунодоминантных эпитопов, распознаваемых В-лимфоцитами, QGPGAP (SEQ ID NO:59) и QQPP (SEQ ID NO:65), но проба на нейтрализацию in vitro показывает, что нейтрализующая активность может быть обусловлена гуморальным иммунным ответом на эпитоп QGPGAP (SEQ ID NO:59) как таковой, поскольку нейтрализацию удается обратить путем добавления к среде пептида (QGPGAP)3 (SEQ ID NO:59; n=3). Согласно некоторым вариантам реализации в HVR1 гексона аденовируса 5 серотипа встраивают три или более повторов PyCS - (QGPGAP)n (SEQ ID NO:59). Согласно некоторым вариантам реализации в HVR1 гексона аденовируса 5 серотипа можно встраивать три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать или двенадцать повторов PyCS - (QGPGAP)n (SEQ ID NO:59; n=3, 4, 5, 6, 7, 8, 9, 10, 11, 12). Повторяющуюся последовательность (QGPGAP)n можно дополнительно встраивать в петлю HI белка нити.
[00132] Пептид - эпитоп, распознаваемый В-лимфоцитами, должен присутствовать на поверхности вирионов аденовируса, чтобы иммунная система могла эффективно распознавать указанный эпитоп. Такими сайтами встраивания могут быть HVR гексона и структуры петли в волоконном белке, и можно сочетать разные сайты встраивания.
Модификация белков капсида и сердцевины аденовируса для повышения активации Т-хелперов
[00133] Согласно еще одному варианту реализации эпитоп, распознаваемый CD4+-лимфоцитами, специфичный для трансгена, можно встраивать в такие белки аденовируса как pVII, pV и гексон в целях повышения иммуногенности вакцины на основе аденовируса. Профессиональные антиген-презентирующие клетки (АПК), такие как дендритные клетки (ДК) и В-лимфоциты, могут захватывать разделенные на части патогены, такие как вирусы, путем эндоцитоза и представлять CD4+-эпитопы в патогене CD4+ Т-лимфоцитам, которые действуют как клетки-хелперы при развитии гуморального и/или клеточного иммунного ответа. pVII и гексон можно легко использовать в качестве целевых белков аденовируса для встраивания антигенных CD4+-пептидов, поскольку в одном вирионе велико число их копий (700-800 копий pVII и 720 копий гексона).
Модификация белков капсида аденовируса для преодоления существующего иммунитета к аденовирусам
[00134] Согласно некоторым вариантам реализации белки капсида аденовируса - белок нити и гексон. можно модифицировать путем встраивания эпитопов, распознаваемых В- и Т-лимфоцитами, в целях преодоления проблемы существующего иммунитета к аденовирусу и/или для усиления гуморального ответа на вакцину на основе аденовируса. По оценкам у 80% молодых людей в популяции в крови циркулируют нейтрализующие антитела на аденовирус (Douglas 2007), особенно на 5 серотип (Ad5). В исследованиях, где аденовирус применяли в качестве генотерапевтического вектора, было показано, что наличие нейтрализующих антител у животных ограничивает экспрессию трансгенов, доставляемых аденовирусами. Помимо нейтрализующих антител ограничению экспрессии рекомбинантных генов также способствуют ответы CD8+T-лимфоцитов (Yang et al. 1995, Yang et al. 1996). Ранее сообщалось, что такой предсуществующий иммунитет к аденовирусу снижает эффективность вакцин на основе рекомбинантных аденовирусов (Рарр et al. 1999), а также снижает иммуногенность вакцины на основе аденовирусов в клинических исследованиях (Priddy et al. 2008).
[00135] Гексон является основной мишенью иммунных ответов, направленных на капсид аденовируса (Roy et al. 2005, Wohlfart 1988), и, вероятно, обусловливает эффективное адъювантное действие аденовируса, включая индукцию ответов CD4+ и CD8+ Т-лимфоцитов. Следовательно, одна из стратегий, которые были применены для преодоления иммунитета к аденовирусу, состоит в том, чтобы заменить весь гексон или его часть другим белком, например, белком редких серотипов, таких как аденовирус 11, 24, 26 и 35. Поскольку гексон является основной мишенью нейтрализующих антител против аденовируса (Youil et al. 2002, Sumida et al. 2005), можно заменить гексон или HVR гексона на аналогичные участки редких серотипов (Wu et al. 2002, Roberts et al. 2006).
[00136] Согласно еще одной стратегии, описанной в одном из вариантов реализации настоящей заявки, гексон аденовируса можно модифицировать путем замены HVR1 или HVR5 на антигенный пептид в целях преодоления предсуществующего иммунитета к аденовирусу или наличия нейтрализующих антител на аденовирус, образовавшихся вследствие ранее проводимой вакцинации вектором на основе аденовируса. Согласно некоторым вариантам реализации антигенным пептидом может быть иммуногенный эпитоп белка CS Plasmodium, и согласно определенным аспектам указанный эпитоп может содержать центральную повторяющуюся последовательность, последовательность эпитопа, распознаваемого CD4+-лимфоцитами, или последовательность эпитопа, распознаваемого CD8+-лимфоцитами.
[00137] Повторное введение вектора на основе Ad того же серотипа недопустимо из-за развития анти-Ad иммунитета после вакцинации. Следовательно, для многих вакцин на основе Ad поддерживающая вакцинация затруднена из-за нарушения экспрессии и представления антигена, кодируемого трансгеном (Yang 1995, Hackett et al. 2000, Harvey et al. 1999, Mastrangeli et al. 1996). Добавление специфического эпитопа в капсид Ad, такого как эпитопы, описанные в следующих примерах, может сократить или устранить данное препятствие согласно некоторым вариантам реализации.
[00138] Следующие примеры приведены с целью проиллюстрировать варианты реализации, и их не следует интерпретировать как ограничивающие объем каких-либо завяленных вариантов реализации. В случае если упоминаются конкретные материалы, это сделано исключительно в целях иллюстрации, но не в целях ограничения изобретения. Специалист в данной области может разработать эквивалентные средства или реактивы без дополнительной изобретательской активности, и не выходя за пределы области настоящего изобретения. Очевидно, процедуры, описанные в настоящей заявке, допускают разнообразные вариации, выходящие за рамки настоящего изобретения. По замыслу авторов изобретения такие варианты включены в объем настоящего изобретения.
Пример 1: Конструирование плазмидных векторов на основе аденовируса с модифицированным капсидом, содержащим белок circumsporozoite плазмодия, и частиц рекомбинантного аденовируса
Картирование эпитопов
[00139] Сначала выбирали иммунодоминантный эпитоп, распознаваемый нейтрализующими В-лимфоцитами. Не вакцинированных ранее мышей Balb/c трижды иммунизировали рекомбинантным белком PyCS с неполным адъювантом Фрейнда, и объединенную сыворотку применяли для определения критического эпитопа нейтрализующего антитела.
[00140] Вкратце, при проведении теста на нейтрализацию в качестве клеток-мишеней применяли клетки HepG2 человека, экспрессирующие CD81. CD81 представляет собой молекулу, необходимую малярийному плазмодию для образования паразитофорных вакуолей в гепатоцитах, где они размножаются и переходят в стадию шизонтов (Silvie et al. 2006), что приводит к значительному увеличению инфекционности спорозоитов in vitro.
[00141] В указанном тесте синтетические пептиды PyCS добавляли в лунки, где они блокировали пептид-специфичное антитело. Результаты картирования эпитопов показали, что центральная повторяющаяся последовательность PyCS, QGPGAP (SEQ ID NO:59), является более эффективным нейтрализующим эпитопом PyCS, чем последовательность QQPP (SEQ ID NO:65). Соответственно, для модификации белков капсида, гексона и/или белка нити применяли встраивание эпитопа (QGPGAP)n (SEQ ID NO:59).
Конструирование плазмидных векторов с модифицированным капсидом
[00142] «Челночный» вектор на основе аденовируса pShuttle-CMV (STRATAGENE) модифицировали путем встраивания кассеты экспрессии GFP ниже сайта клонирования. Сначала у фрагмента BsmBI-SacI (pCMV+GFP) и фрагмента SacI-BsmBI (сигнал полиА SV40) pmaxGFP («Lonza», Кельн, Германия) «затупляли концы» и встраивали их в «затупленные» сайты Sall и KpnI в pUC19, соответственно. Фрагмент BamHI-EcoRI полученного в результате pCMV-GFP/pUC19 встраивали в те же сайты SV40pA/pUC19 с получением фрагмента SV40pA-pCMV-GFP. У указанного фрагмента «затупляли концы» и встраивали его в сайт EcoRV вектора pShuttle-CMV. Полученный в результате челночный вектор (GFP/pShuttle-CMV) содержал двойные промоторы pCMV и SV40pAs для трансгена и GFP.
[00143] Еще одну модификацию челночного вектора на основе аденовируса получали путем замены области промотора CMV на промотор CMV5 из pQBI-AdCMV5 (QBIOgene). Чтобы сконструировать вектор pShuttle-CMV5, фрагмент SgrAI-KpnI вектора заменяли фрагментом, содержащим последовательность промотора CMV5 и расположенную выше последовательности из промотора CMV в векторе pShuttle-CMV.
[00144] Ген CS (PyCS) P. yoelii подвергали оптимизации кодонов за исключением повторов (QGPGAP)n (SEQ ID NO:59) путем перекрывающейся ПЦР на основе алгоритма оптимизации кодонов JCat (http://www.jcat.de/).
[00145] В качестве шаблонной последовательности для оптимизации кодонов применяли последовательность аминокислот PfCSP штамма 3D7 Р. falciparum. Оптимизацию кодонов для экспрессии белка у человека проводили при помощи оптимизирующей компьютерной программы Integrated DNA Technologies (Технологии интегрированных ДНК, Coralville, IA США). Фрагменты ДНК, которые кодируют весь PfCSP за исключением GPI-заякоренного мотива на С-конце (Фиг.10; SEQ ID NO:2), синтезировали при помощи Технологии интегрированных ДНК.
[00146] Ген с оптимизацией кодонов PyCS (Фиг.9; SEQ ID NO:1) или ген PfCSP (Фиг.10; SEQ ID NO:2) встраивали в сайты KpnI и HindIII вектора pShuttle-CMV, pShuttle-CMV5 или GFP/pShuttle-CMV. Полученные челночные векторы на основе аденовируса, кодирующие белок circumsporozoite плазмодия, применяли для гомологичной рекомбинации с использованием AdEasy-1 для конструирования генома аденовируса, который содержал ген антигена circumsporozoite плазмодия и последовательность, кодирующую интактный белок аденовируса. Вкратце, челночные векторы на основе аденовируса, кодирующие белок circumsporozoite плазмодия, линеаризовали путем расщепления ферментом PmeI, и совместно трансформировали клетки Е.coli BJ5183 линеаризованным челночным вектором и вектором pAdEasy-1 (Bruna-Romero et al. 2003) в целях гомологичной рекомбинации.
[00147] На Фиг.1 в обобщенном виде представлена и проиллюстрирована модификация белков капсида аденовируса. На Фиг.2 проиллюстрирована модификация последовательности HVR1 в ДНК генома аденовируса. Вкратце, AdEasy-1 расщепляли при помощи фермента Sfil, и фрагмент 6,4 т.п.н. субклонировали в сайты EcoRI и PstI pUC19 при помощи линкерных олигомеров EcoRI-Sfil и PstI-Sfil. Для замены HVR1 на эпитоп circumsporozoite плазмодия, распознаваемый В-лимфоцитами, участок, содержащий сайты Agel и Ndel, амплифицировали путем двухэтапной ПЦР с применением праймеров, которые содержали последовательность эпитопа вместо последовательности HVR1. Продукт ПЦР расщепляли при помощи Agel и Ndel, и затем применяли для замены нативного участка Agel-Ndel фрагмента Sfil в векторе Sfil/pUC19. После подтверждения последовательности фрагмент Sfil в ДНК генома аденовируса заменяли фрагментом Sfil, содержащим последовательность эпитопа спорозоита и получали гексон, модифицированный по HVR1. Согласно некоторым вариантам реализации гексон, модифицированный по HVR, может содержать последовательность нуклеиновой кислоты в соответствии с SEQ ID NO:3 (Фиг.11), SEQ ID NO:4 (Фиг.12), SEQ ID NO:5 (Фиг.13), SEQ ID NO:6 (Фиг.14), SEQ ID NO:7 (Фиг.15), SEQ ID NO:8 (Фиг.16), SEQ ID NO:9 (Фиг.17), SEQ ID NO:10 (Фиг.18), SEQ ID NO:11 (Фиг.19), SEQ ID NO:12 (Фиг.20), SEQ ID NO:13 (Фиг.21), SEQ ID NO:14 (Фиг.22), SEQ ID NO:15 (Фиг.23), SEQ ID NO:16 (Фиг 24), SEQ ID NO:17 (Фиг.25), SEQ ID NO:18 (Фиг.26), SEQ ID NO:19 (Фиг.27), SEQ ID NO:20 (Фиг.28), SEQ ID NO:21 (Фиг.29), SEQ ID NO:22 (Фиг.30) или SEQ ID NO:23 (Фиг.31).
[00148] Чтобы встроить (NANP)28 (SEQ ID NO:60; n=28) в HVR1, часть центрального повторяющегося участка PfCSP с оптимизацией кодонов амплифицировали путем ПЦР с использованием праймеров, содержащих гексон-специфичную последовательность на 5'-конце и NANP- специфичную последовательность на 3'-конце, и полученный в результате фрагмент ДНК встраивали в участок Agel-Ndel путем второй ПЦР.
[00149] Для модификации по HVR5, как показано на Фиг.3, в петлю L1 HVR5 гексона в Ad Easy-1 вводили сайт XbaI, а затем синтезированный фосфорилированный двухцепочечный олигомер, кодирующий эпитоп circumsporozoite плазмодия, встраивали в сайт XbaI. Встраивание подтверждали путем секвенирования (Фиг 31; SEQ ID NO:23).
[00150] Для модификации белка нити, как показано на Фиг.4, фрагмент Spel-PacI вектора AdEasy-1 субклонировали в сайты EcoRI и PstI pUC19 при помощи линкерных олигомеров EcoRI-PacI и PstI-Spel. Чтобы встроить последовательность эпитопа circumsporozoite плазмодия, распознаваемого В-лимфоцитами, в петлю HI головки белка нити, участок, содержащий сайты EcoNI (или Nhel) и Mfel амплифицировали путем двухэтапной ПЦР с применением праймеров, которые содержали последовательность эпитопа. Продукт ПЦР расщепляли при помощи EcoNI (или Nhel) и Mfel и затем применяли для замены нативного участка EcoNI (или Nhel)-Mfel белка нити в векторе Spel-PacI/pUC19. После подтверждения последовательности (Фиг.32, SEQ ID NO:24; Фиг.33, SEQ ID NO:25) фрагмент Spel-PacI вектора Ad Easy-1 заменяли фрагментом Spel-PacI, содержащим последовательность эпитопа. Полученную ДНК аденовируса с модифицированным белком нити применяли для гомологичной рекомбинации челночным вектором на основе аденовируса, кодирующим белок circumsporozoite плазмодия, и получали ДНК аденовируса с белком нити, модифицированным путем встраивания circumsporozoite плазмодия.
[00151] Чтобы сконструировать ДНК аденовируса с модифицированным HVR1 и белком нити, которая содержит две вставки эпитопов, фрагмент Sfil-Sfil ДНК аденовируса с модифицированным белком нити заменяли фрагментом Sfil-Sfil, содержащим эпитоп белка circumsporozoite в HVR1, как показано на Фиг.5.
[00152] Чтобы модифицировать С-конец pVII, участок, содержащий сайты Sfi I и Sal I, амплифицировали путем двухэтапной ПЦР с применением праймеров, которые содержали последовательность эпитопа белка спорозоита. Продукт ПЦР расщепляли при помощи Sfil и Sall, и затем применяли для замены нативного участка Sfil-Sall вектора Sfil/pUC19 (Фиг.6, 7 и 8). После подтверждения последовательности (Фиг.34, SEQ ID NO:26; Фиг.35, SEQ ID NO:27) фрагмент Sfil-Sfil в ДНК аденовируса с HVR1 и/или белком нити, модифицированным путем встраивания circumsporozoite плазмодия, заменяли фрагментом Sfil-Sfil, содержащим эпитоп белка circumsporozoite плазмодия в pVII.
[00153] Чтобы встроить последовательность эпитопа белка спорозоита, распознаваемого CD4+-лимфоцитами, EYLNKIQNSLSTEWSPCSVT (SEQ ID NO:62), в pVII, фрагмент длиной приблизительно 7,7 т.н. вектора pAdEasy-1 подготавливали путем расщепления RsrII и клонировали между сайтами EcoRI и HindIII плазмиды pUC19 при помощи линкера RsrII (RsrII/pUC19). Участок, содержащий сайты AscI и BgIII в RsrII/pUC19, амплифицировали путем двухэтапной ПЦР с применением праймеров, которые содержали последовательность эпитопа. Продукт ПЦР расщепляли при помощи AscI и BgIII, и затем применяли для замены нативного участка AscI и BgIII в плазмиде RsrII/pUC19. После подтверждения последовательности (Фиг.36, SEQ ID NO:28; Фиг.37, SEQ ID NO:29), фрагмент RsrII в ДНК аденовируса, модифицированного по HVR1, заменяли фрагментом RsrII, содержащим последовательность эпитопа.
[00154] Рекомбинантные аденовирусы, перечисленные в Таблице 1 (Р. yoelii) и Таблице 2 (Р. falciparum) ниже, были сконструированы для оценки влияния встраивания эпитопа на инфекционность, иммуногенность и чувствительность к предсуществующему иммунитету к аденовирусу. Применяемые векторы на основе рекомбинантных аденовирусов были векторами с дефектной репликацией на основе аденовируса 5 серотипа с делегированными Е1 и Е3 (STRATAGENE). На Фиг.1 приведена структурная схема рекомбинантного аденовируса с модифицированным капсидом с встроенным белком спорозоита плазмодия.



[00155] Плазмиду на основе геномной ДНК аденовируса с модифицированным капсидом, очищали, переводили в линейную форму путем расщепления при помощи PacI и применяли для трансфекции клеток AD293.
[00156] Частицы аденовируса готовили из трансфецированных клеток AD293 путем проведения четырех циклов замораживания/размораживания и применяли для последующей амплификации вируса. После последней амплификации частицы аденовируса очищали путем центрифугирования в градиенте плотности CsCI. Затем отбирали дорожку и подвергали ее диализу против буфера для диализа, чтобы удалить CsCI. Количество частиц вируса (v.p.) определяли на основании OD (оптической плотности) на длине волны 260 нм (1 OD260=1,25×1012v.p./мл) (Bruna-Romero et al. 2003).
[00157] Во время процедуры амплификации среди аденовирусов с модифицированным капсидом наблюдались лишь небольшие различия в росте аденовируса, что указывает на то, что модификация не оказывает никакого неблагоприятного действия на инфекционность и продуктивность аденовирусов.
Пример 2: Иммунный ответ, специфичный в отношении белка circumsporozoite Plasmodium yoelii
Проверка рекомбинантных аденовирусов с белком Plasmodium yoelii
[00158] Челночные векторы на основе аденовируса, несущие белок circumsporozoite плазмодия, применяли для временной трансфекции, чтобы подтвердить экспрессию белка circumsporozoite плазмодия с применением клеток AD293 (Фиг.38). Через 24 часа после трансфекции клетки подвергали лизису в буфере для разведения проб с SDS (додецил-сульфатом натрия), после чего проводили электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия (SDS PAGE) и вестерн-блоттинг с моноклональным анти-PyCS антителом (9D3).
[00159] Чтобы подтвердить встраивание эпитопа в белки капсида аденовируса, очищенные рекомбинантные аденовирусы анализировали методом электрофореза в SDS-ПААГ (2×109 v.p./дорожку), а также проводили вестерн-блоттинг (1×109 v.p./дорожку) с антителом к спорозоиту, которое распознавало повторы (QGPGAP)n (SEQ ID NO:59), как показано на Фиг.39А и 40А. Интенсивность полос на Фиг.39А коррелировала с числом копий белка капсида в вирионе аденовируса: число копий белка нити (36 копий на вирион) в двадцать раз меньше, чем число копий гексона (720 копий на вирион). Нижняя полоса на дорожке 4 на Фиг.39А, вероятно, соответствует расщепленному гексону. Интенсивность полос на Фиг.40А коррелировала с числом повторов (QGPGAP)n (SEQ ID NO:59), встроенных в HVR1.
[00160] Чтобы оценить, присутствует ли эпитоп PyCS, распознаваемый В-лимфоцитами, на поверхности вириона, серийные разведения частиц очищенного рекомбинантного аденовируса наносили на планшет для твердофазного иммуноферментного анализа (ELISA) и проводили детекцию при помощи анти-PyCS антитела, которое распознает повторы (QGPGAP)n (SEQ ID NO:59). Антитело распознало все аденовирусы с модифицированным капсидом (Фиг.39В и 40В). Результаты ELISA указывают на то, что эпитоп PyCS, распознаваемый В-лимфоцитами, встроенный в белки капсида, стабильно появляется на поверхности вирионов аденовируса.
Иммунный ответ, специфичный для белка circumsporozoite Plasmodium yoelii, после вакцинации аденовирусом с модифицированным капсидом, содержащим PyCS
[00161] Самок мышей BALB/c в возрасте шести-восьми недель приобретали у «Taconic» (Гудзон, Нью Йорк, США) и содержали в стандартных условиях в Центре исследований лабораторных животных Университета Рокфеллера. Для вакцинации аденовирусы разводили в ФСБ и вводили внутримышечно в указанных дозах.
[00162] Для оценки иммуногенности рекомбинантных аденовирусов после однократной вакцинации группы ранее не вакцинированных мышей BALB/c (пять животных на группу) вакцинировали путем внутримышечного введения разных рекомбинантных PyCS аденовирусов в количестве 1×109 v.p. и оценивали клеточные иммунные ответы (КИО) через 2 недели после вакцинации методом иммуноферментных пятен (Фиг.41А).
[00163] Количество PyCS-специфичных секретирующих IFN-γ CD8+ Т-лимфоцитов в селезенке вакцинированных мышей определяли методом иммуноферментных пятен (ELISPOT) с применением синтетического пептида, соответствующего эпитопу, распознаваемому CD8+ Т-лимфоцитами (SYVPSAEQI; SEQ ID NO:66), в белке PyCS. Вкратце, планшеты на 96 лунок из нитроцеллюлозы (Milititer НА, «Millipore») на одну ночь покрывали антимышиным интерфероном у mAb, R4. после инкубации в течение ночи при комнатной температуре лунки многократно промывали культуральной средой и блокировали культуральной средой в течение 4 часов. В лунки для теста ELISPOT добавляли 5×105спленоцитов, взятых у вакцинированных мышей, в присутствии или в отсутствие 10 мкг/мл пептида эпитопа, распознаваемого CD8+ Т-лимфоцитами, и инкубировали в течение 24 часов при 37°С и 5% CO2. После обильной промывки планшетов ФСБ, содержащим 0,05% Tween 20 (PBST), добавляли биотинилированные моноклональные антитела к интерферон γ мыши, XMG1.2, в PBST, и инкубировали в течение ночи при 40°С. После промывания буфером PBST планшеты инкубировали с меченным пероксидазой авидином («eBiosciences»). Пятна появлялись при добавлении субстрата 3-амино-9-этилкарбазола (АЭК) («BD Biosciences»).
[00164] Все аденовирусы с модифицированным капсидом вызывали сопоставимый уровень клеточного иммунитета против аденовирусов с интактным белком капсида в данной дозе (Фиг.41В).
[00165] Далее не подвергавшимся ранее вакцинации мышам BALB/c вводили несколько доз рекомбинантных аденовирусов с увеличением дозы, т.е. 1×108, 1×109 и 1×1010 v.p., с 3-недельными интервалами, как показано на Фиг.42А. PyCS-специфичный гуморальный ответ определяли методом ELISA. Из хвостовой вены вакцинированных мышей брали по пять микролитров крови и разводили в 495 мкл ФСБ, затем указанные пробы центрифугировали при 5000 об/мин в течение 5 мин для приготовления проб разведенной плазмы (×100). Планшеты для ELISA Maxisorp покрывали CS-специфичным пептидом ((QGPGAP)3; SEQ ID NO:59, n=3) в концентрации 5 мкг/мл в 0,1М натрий-карбонатном буфере (рН 9,5) и оставляли на ночь при 4°С. Планшеты промывали и блокировали 1х разбавителем в течение 2 часов при комнатной температуре. Затем планшеты промывали повторно и добавляли в них по 100 мкл плазмы или сыворотки, подвергнутой последовательному двукратному разведению в 1х разбавителе; далее планшеты инкубировали в течение одного часа при комнатной температуре. Планшеты промывали и инкубировали с 100 мкл меченного пероксидазой хрена (HRP-меченного) антитела козы к IgG мыши. Все пептиды были синтезированы компанией «Biosyntheis» (Луисвилл, Техас, США).
[00166] При таком же режиме вакцинации все аденовирусы с модифицированным капсидом вызывали гуморальный ответ с образованием анти-(QGPGAP)3, который по уровню значительно превышал ответ на wt/PyCS-GFP к неделе 10 (Фиг.42В).
[00167] Для определения эффективности вакцины на основе аденовируса с модифицированным капсидом вакцинированных мышей инфицировали, вводя им 2×104 контагиозных спорозоитов Р. yoelii путем инъекции в хвостовую вену на неделе 10. Через 42 часа после инфицирования спорозоитами концентрацию паразитов определяли, подсчитывая число плазмодий-специфичных рибосомальных РНК в печени мышей и выражая его как отношение абсолютного числа копий рибосомальной РНК плазмодия к абсолютному числу мРНК GADPH (глицеральдегид-3-фосфатдегидрогеназы) мыши. Для статистического анализа значения подвергли логарифмическому преобразованию, и далее для определения различий был применен односторонний ANOVA (дисперсионный анализ) с последующим применением критерия Дуннетта.
[00168] Вакцинации с применением (QGPGAP)3-HVR1/PyCS-GFP, (QGPGAP)3-Fib/PyCS-GFP или (QGPGAP)3-HVR1/Fib/PyCS-GFP вызывали более высокий уровень защиты, чем вакцинация с применением wt/PyCS-GFP, и приводили к значительно более низким концентрациям плазмодия у мышей, инфицированных малярией (Фиг.42С).
[00169] Далее оценивали функциональные свойства PyCS-специфичного антитела, индуцируемого в ответ на аденовирусы с модифицированным капсидом. Сначала, чтобы проверить, способна ли сыворотка мышей, вакцинированных аденовирусом на неделе 10 (Фиг.42А), распознавать интактные спорозоиты, проводили непрямой иммунофлюоресцентный анализ (ИФА). В тесте ИФА высушенные на воздухе спорозоиты на предметных стеклах с множественными мазками инкубировали с 3% бычьим сывороточным альбумином (БСА) в ФСБ в течение одного часа, а затем инкубировали с разведенной сывороткой в течение одного часа. После промывки предметные стекла инкубировали с вторичными антителами с флуоресцентной меткой в течение одно часа. Далее предметные стекла промывали, и на них определяли ИФА титры как максимальное разведение, дающее флуоресценцию под флуоресцентным микроскопом. И (QGPGAP)3-HVR1/PyCS-GFP, и (QGPGAP)3-HVR1/Fib/PyCS-GFP давали максимальные ИФА-титры против спорозоитов (Фиг.43А), что указывает на то, что встраивание эпитопа (QGPGAP)3 в HVR1 гексона аденовируса придает аденовирусу способность к индукции полноценного ответа антител не только против синтетического пептида, но также и против нативного эпитопа, присутствующего в малярийном плазмодии.
[00170] На втором этапе, чтобы определить, вырабатываются ли у мышей, вакцинированных аденовирусом с модифицированным капсидом (Фиг.42А), «функциональные» антитела, способные нейтрализовать инфекционность спорозоитов, проводили тест на нейтрализацию спорозоитов in vitro.
[00171] В тесте на нейтрализацию in vitro спорозоиты Р. yoelii добавляли к CD81/HepG2 в планшете на 96 лунок в присутствии 30-кратно разведенной объединенной сыворотки, полученной от вакцинированных аденовирусом мышей. После двухчасовой инкубации неинфицированные спорозоиты отмывали средой и затем указанные клетки культивировали в течение 42 часов. Отношение количества рибосомальной РНК плазмодия к мРНК GADPH человека измеряли при помощи ПЦР в режиме реального времени (Ophorst et al. 2006).
[00172] Пробы объединенной сыворотки, взятой у мышей, вакцинированных аденовирусом с модифицированным капсидом, в частности (QGPGAP)3-HVR1/PyCS-GFP и (QGPGAP)3-HVR1/Fib/PyCS-GFP, почти полностью (99%) подавляли инфекционность спорозоитов in vitro (Фиг.43В). Примечательно, что степень подавления в данном тесте была обратно пропорциональна титрам ИФА, показанным на Фиг.43А.
Защита от эритроцитарной стадии малярии
[00173] Далее определяли, защищает ли вакцинация rAd с модифицированным капсидом от развития эритроцитарной стадии малярии после заражения спорозоитом. Эксперименты проводили дважды, и в каждом эксперименте 20 мышей BALB/c в каждой группе вакцинировали три раза с применением wt/PyCS-GFP или (QGPGAP)3-HVR1/PyCS-GFP, как показано на Фигуре 42А, и через 4 недели после последней вакцинации мышей инфицировали, вводя внутривенно 50 спорозоитов Р. yoelii. Мазки крови, окрашенные по Гимзе, исследовали через 3-12 дней после заражения в целях выявления эритроцитарной стадии малярии. В группе, вакцинированной wt/CS-GFP, были инфицированы 30 из 40 мышей (75%), а в группе без вакцинации были инфицированы 35 из 40 мышей (87,5%) (Таблица 3, ниже). Мыши, вакцинированные (QGPGAP)3-HVR1/CS-GFP, были лучше защищены, чем мыши, вакцинированные wt/CS-GFP; в данной группе были инфицированы только 15 из 40 мышей (37,5%), что согласуется с результатом оценки защиты, в которой измеряли концентрацию паразитов в печени (Фигура 42С).
Первичная-поддерживающая вакцинация 1
[00174] Затем ранее не вакцинированные мыши BALB/c получали «бустер-дозы» HVR1-модифицированных аденовирусов с PyCS, которые содержали четыре или шесть повторов (QGPGAP)n (SEQ ID NO:59; n=4, 6), с адъювантом или без него, в нескольких возрастающих дозах (т.е. 1×108, 1×109 и 1×1010 v.p.) с3-недельными интервалами, как показано на Фиг.44А. В данном эксперименте в качестве адъюванта применяли Sigma Adjuvant System («Sigma-Aldrich»), содержащую 200 мкг/мл сапонина («Sigma-Aldrich»). Один флакон Sigma Adjuvant System (1 мл) содержит 0,5 мг монофосфорил-липида А (обезвреженного эндотоксина) из Salmonella minnesota и 0,5 мг синтетической трегалозы-дикориноиголата в 2% масле (сквален) - Tween 80 в воде. Перед вакцинацией раствор аденовируса смешивали с равным количеством адъюванта. 100 микролитров смеси аденовирус-адъювант вводили внутримышечно. PyCS-специфичный гуморальный и клеточный иммунный ответ оценивали, как было описано выше. Была выявлена тенденция, заключающаяся в том, что HVR1-модифицированный аденовирус, содержащий шесть повторов, приводил к более высокому титру антител, чем аденовирус, содержащий четыре повтора, а применение адъюванта увеличивало титр антител (Фиг.44В). Напротив, адъювант не оказывал никакого действия на клеточный иммунитет (данные не показаны).
[00175] Чтобы определить эффективность вакцины на основе HVR1-модифицированных аденовирусов с PyCS, по пять мышей в каждой группе инфицировали, вводя им 2×104 контагиозных спорозоитов Р. yoelii путем инъекции в хвостовую вену на неделе 9. Через 42 часа после инфицирования спорозоитами концентрацию паразитов определяли, как описывалось выше. Для статистического анализа значения подвергли логарифмическому преобразованию, и далее для определения различий был применен односторонний ANOVA с последующим применением критерия Дуннетта. Была выявлена такая тенденция, что HVR1-модифицированный аденовирус, содержащий шесть повторов, более эффективно сокращал концентрацию паразитов, чем аденовирус, содержащий четыре повтора, а применение адъюванта усиливало защиту (Фиг.44С).
[00176] Для оценки функциональных свойств антитела, индуцируемого в ответ на аденовирусы с модифицированным капсидом, содержащим PyCS, мы проводили пробу нейтрализации in vitro, как описывалось выше. Пробы объединенной сыворотки, отобранной у мышей, вакцинированных аденовирусом с модифицированным капсидом, содержащим PyCS, на неделе 9, нейтрализовали инвазию спорозоитов при 50-кратном разведении (Фиг.44D).
Первичная-поддерживающая вакцинация 2
[00177] Ранее не вакцинированные мыши BALB/c получали «бустер-дозы» HVR1-модифицированных аденовирусов с PyCS, которые содержали шесть, девять или двенадцать повторов (QGPGAP)n (SEQ ID NO:59; n=6, 9, 12), с адъювантом или без него, трехкратно в дозировке 1×1010 v.p. с 3-недельными интервалами, как показано на Фиг.45А. В данном эксперименте в качестве адъюванта применяли Sigma Adjuvant System («Sigma-Aldrich»), содержащую 200 мкг/мл сапонина («Sigma-Aldrich»). Раствор аденовируса перед вакцинацией смешивали с равным количеством адъюванта. PyCS-специфичный гуморальный и клеточный иммунный ответ оценивали, как было описано выше. HVR1-модифицированный аденовирус с PyCS, который содержал двенадцать повторов (QGPGAP)n (SEQ ID NO:59; n=12) с адъювантом приводил к самому высокому титру антител среди указанных групп к неделе 9 (Фиг.45В). Что касается PyCS-специфичного клеточного иммунитета, среди групп не было различий, что указывает на то, что встраивание более длинного эпитопа (до двенадцати повторов) не снижает инфекционности аденовируса in vivo (Фиг.45С). Кроме того, адъювант не влияет на способность аденовируса вызывать клеточный иммунный ответ (Фиг.45С).
[00178] Чтобы определить эффективность вакцины на основе HVR1-модифицированных аденовирусов с PyCS, по пять мышей в каждой группе инфицировали, вводя им 2×104 контагиозных спорозоитов Р. yoelii путем инъекции в хвостовую вену на неделе 9. Через 42 часа после инфицирования спорозоитами концентрацию паразитов определяли, как описывалось выше. Все HVR1-модифицированные аденовирусы с PyCS, содержащие повторы (QGPGAP)n(SEQ ID NO:59, n=6, 9, 12), с адъювантом или без него, демонстрировали повышенную защиту. Однако HVRI-модифицированный аденовирус с PyCS, содержащий двенадцать повторов (QGPGAP)n (SEQ ID NO:59, n=12) с адъювантом демонстрировал максимальную защиту (Фиг.45D), и обладал значительно большими защитными свойствами, чем любое другое средство.
Пример 3: Иммунный ответ, специфичный в отношении белка circumsporozoite Plasmodium falciparum
[00179] Валидация рекомбинантных аденовирусов с белками Plasmodium falciparum
Челночные векторы на основе аденовируса, кодирующие белок circumsporozoite плазмодия, применяли для временной трансфекции, чтобы подтвердить экспрессию белка circumsporozoite плазмодия с использованием клеток AD293 (Фиг.46А). Через 24 часа после трансфекции клетки подвергали лизису в буфере для разведения проб с SDS, после чего проводили электрофорез в SDS-ПААГ и вестерн-блоттинг с моноклональным анти-NANP антителом (2А10).
[00180] Чтобы подтвердить встраивание эпитопа в белки капсида, очищенные рекомбинантные аденовирусы исследовали методом электрофореза в SDS-ПААГ (2×109 v.p./дорожку), а также проводили вестерн-блоттинг (1×109v.p./дорожку) с антителом к спорозоиту, которое распознает повторы (NANP)n (SEQ ID NO:60), как показано на Фиг.47А и 48А. Интенсивность полос на Фиг.47А коррелировала с числом копий белка капсида в вирионе аденовируса: число копий белка нити (36 копий на вирион) в двадцать раз меньше, чем число копий гексона (720 копий на вирион). Интенсивность полос на Фиг.48А коррелирует с числом повторов NANP в HVR1.
[00181] Чтобы оценить, присутствует ли эпитоп PfCSP, распознаваемый В-лимфоцитами, на поверхности вириона аденовируса, последовательные разведения частиц очищенного рекомбинантного аденовируса наносили на планшет для твердофазного иммуноферментного анализа (ELISA) и проводили детекцию при помощи анти-PyCS антитела, которое распознает повторы (NANP)n (SEQ ID NO:60). Указанное антитело распознало все аденовирусы с модифицированным капсидом (Фиг.47В и 48В). Результаты ELISA указывают на то, что эпитоп PfCSP, распознаваемый В-лимфоцитами, встроенный в белки капсида, стабильно присутствует на поверхности вирионов аденовируса.
Первичная-поддерживающая вакцинация 3
[00182] Ранее не вакцинированные мыши BALB/c получали несколько возрастающих доз аденовирусов с PfCSP (т.е. 1×108, 1×109 и 1×1010 v.p.) с 3-недельными интервалами, как показано на Фиг.49А. PfCSP-специфичный гуморальный и клеточный иммунный ответ оценивали методом ELISA, как было описано выше, с применением планшетов для ELISA, покрытых 1 мкг/мл (Т1В)4 - пептидом с повторами CS, который содержал повторяющиеся последовательности (NANP)n (SEQ ID NO:60) (Calvo-Calle et al. 2006). Для статистического анализа значения подвергли логарифмическому преобразованию, и далее для определения различий между wt/PfCSP и аденовирусами с модифицированным капсидом был применен односторонний ANOVA с последующим применением критерия Дуннетта. Все аденовирусы с модифицированным капсидом приводили к титру анти-NANP антител, который статистически значимо превышал титр, вызываемый wt/PfCSP.
Первичная-поддерживающая вакцинация 4
[00183] Затем PfCSP-специфический гуморальный ответ измеряли, как описано выше. Все HVRI-модифицированные аденовирусы приводили к образованию более высоких титров анти-NANP антител, чем wt/PfCSP к неделе 9 (Фиг.50В). Для статистического анализа значения подвергли логарифмическому преобразованию, и далее для определения различий был применен односторонний ANOVA с последующим применением критерия Дуннетта.
Первичная-поддерживающая вакцинация 5
[00184] Ранее не вакцинированные мыши BALB/c получали «бустер-дозы» HVR1-модифицированных аденовирусов, которые содержали десять, шестнадцать или двадцать два повтора (NANP)n (SEQ ID NO:60; n=10, 16, 22), с адъювантом или без него, трехкратно в дозировке 1×1010 v.p. с3-недельными интервалами, как показано на Фиг.51А. В данном эксперименте в качестве адъюванта применяли Sigma Adjuvant System («Sigma-Aldrich»), содержащую 200 мкг/мл сапонина («Sigma-Aldrich»). Перед вакцинацией раствор аденовируса смешивали с равным количеством адъюванта. PfCSP-специфичный гуморальный иммунный ответ оценивали, как описано выше, и выявили, что HVR1-модифицированный аденовирус с более длинным эпитопом, распознаваемым В-лимфоцитами, приводил к более высокому титру антител (Фиг.51В).
Пример 4: Встраивание эпитопа PyCS, распознаваемого CD4-лимфоцитами. в белок сердцевины аденовируса pVII
[00185] Выработка и пролиферация антиген-специфичных CD4 Т-лимфоцитов. Соответственно, чтобы определить, возможно ли усилить PyCS-специфичный гуморальный иммунный ответ, вызываемый аденовирусом с модифицированным капсидом, путем встраивания эпитопа PyCS, распознаваемого CD4-лимфоцитами, в белок аденовируса, был сконструирован (QGPGAP)3-Fib/PyCS-GFP, который содержит эпитоп PyCS, распознаваемый CD4-лимфоцитами, в pVII ((QGPGAP)3-Fib/CD4-pVII-1/PyCS-GFP). pVII представляет собой один из белков сердцевины аденовируса, и число его копий на вирион составляет 700-800, что создает идеальные условия для эффективной презентации CD4 эпитопа на молекуле МНС II класса. Как показано на Фиг.52А, на SDS-PAGE геле полоса pVII сдвигается при встраивании эпитопа PyCS, распознаваемого CD4-лимфоцитами, в белок pVII.
[00186] Для проверки влияния встраивания эпитопа PyCS, распознаваемого CD4-лимфоцитами, в pVII, ранее не вакцинированных мышей BALB/c вакцинировали с применением (QGPGAP)3-Fib/PyCS-GFP или (QGPGAP)3-Fib/CD4-pVII-1/PyCS-GFP, как показано на Фиг.42А, и определяли титр анти-QGPGAP антител методом ELISA на неделе 10. (QGPGAP)3-Fib/CD4-pVII-1/PyCS-GFP приводил к более высокому титру анти-QGPGAP антител, чем (QGPGAP)3-Fib/PyCS-GFP (Фиг.52В), и было показано, что встраивание эпитопа PyCS, распознаваемого CD4-лимфоцитами, в pVII усиливало гуморальный иммунный ответ, вызываемый аденовирусом с модифицированным капсидом.
Встраивание эпитопа PfCSP, распознаваемого CD4-лимфоцитами, в белок сердцевины аденовируса pVII
[00187] Для проверки влияния встраивания эпитопа PfCSP, распознаваемого CD4-лимфоцитами, в разные участки белка сердцевины аденовируса pVII на вызываемый аденовирусом иммунный ответ, конструировали HVR1-модифицированные аденовирусы с PfCSP, содержащие эпитоп PfCSP, распознаваемый CD4+-лимфоцитами, непосредственно перед первым клеточным сигналом внутриядерной локализации (NLS) или между двумя NLS (Фиг.53А).
[00188] Чтобы подтвердить встраивание эпитопа в pVII, очищенные рекомбинантные аденовирусы анализировали методом электрофореза в SDS-ПААГ, как описано выше. Как показано на Фиг.53В, полосы, соответствующие (NANP)4-HVR1/CD4-pVII-2/PfCSP и (NANP)4-HVR1/CD4-pVII-3/PfCSP, сдвигались вверх из-за встраивания эпитопа.
PfCSP-специфичный иммунный ответ, вызываемый аденовирусом с модифицированными HVR1 и pVII, содержащими PfCSP
[00189] Затем ранее не вакцинированные мыши BALB/c получали «бустер-дозы» аденовирусов с модифицированными HVR1 и pVII и с PfCSP, которые содержали четыре повтора NANP в HVR1, и эпитоп PfCD4+, в нескольких возрастающих дозах (т.е. 1×108, 1×109 и 1×1010 v.p.) с 3-недельными интервалами, как показано на Фиг.54А. PfCSP-специфичный гуморальный иммунный ответ оценивали, как описано выше. (NANP)4-HVR1/CD4-pVII-2/PfCSP и (NANP)4-HVR1/CD4-pVII-3/PfCSP приводили к более высокому титру анти-NANP антител, чем (NANP)4-HVR1/PfCSP на неделе 6 (Фиг.54В). Что касается CMI, (NANP)4-HVR1/CD4-pVII-3/PfCSP приводил к более высокому уровню IFNγ и IL-4-секретирующих PfCSP-специфичных CD4+ Т-лимфоцитов, чем (NANP)4-HVR1/PfCSP (Фиг.54С).
Пример 5: Влияние модификации капсида на иммунитет к аденовирусу
[00190] Для проведения экспериментов по нейтрализации аденовирусов in vitro сыворотку добавляли к клеткам AD293 в указанных разведениях, а затем инфицировали аденовирусами. Пробы сыворотки людей европеоидной расы получали из компании Innovative Research (Novi, Мичиган, США). Все данные, полученные методом проточной цитометрии, анализировали при помощи компьютерной программы FlowJo v8.8 («Tree Star, Inc», Ashland, Орегон, США). Клетки AD293 инфицировали каждым из аденовирусов с модифицированным капсидом в присутствии проб нейтрализующей аденовирус сыворотки человека в указанных разведениях, после чего измеряли экспрессию GFP методом проточной цитометрии. Замена HVR1 на эпитоп PyCS-B явно делала аденовирус устойчивым к сыворотке, направленной против аденовируса 5 серотипа, а модификация HVR5 или белка нити не оказывала никакого действия (Фиг.55).
[00191] Затем определяли, является ли HVR1 критичной молекулой для нейтрализации in vivo. Для этой цели мышей инфицировали, вводя им 1×1010 v.p. wt/«пустого» аденовируса дважды для создания достаточного предсуществующего иммунитета к аденовирусу (Фиг.56А) и рандомизировали их в зависимости от титра антител, определенного у них методом ELISA. Затем мышам вводили однократную дозу вакцины на основе аденовируса с модифицированным капсидом или на основе немодифицированного аденовируса и оценивали уровень PyCS-специфичного опосредуемого CD8+ Т-лимфоцитами ответа, как описано выше. Только вакцинация (QGPGAP)3-HVR1/PyCS-GFP или (QGPGAP)3-HVR1/Fib/PyCS-GFP могла вызвать значительно более эффективный CS-специфичный ответ CD8+ Т-лимфоцитов по сравнению с ответами, вызываемыми другими аденовирусами с модифицированным капсидом или немодифицированными аденовирусами (Фиг.56В).
[00192] Также оценивали уровень ответа антител на эпитоп (QGPGAP)3, который присутствовал на белках капсида rAd у мышей, инфицированных wt/«пустым» Ad и впоследствии вакцинированных rAd с модифицированным капсидом (Фигура 57А). Только мыши, вакцинированные (QGPGAP)3-HVR1/PyCS-GFP и (QGPGAP)3-HVR1/Fib/PyCS-GFP, обладали способностью поддерживать значительно более высокий титр анти-QGPGAP антител, чем титр у мышей, вакцинированных wt/PyCS-GFP (Фигура 57В).
Приведенные выше примеры предназначены для того, чтобы более детально проиллюстрировать варианты реализации, и их не следует интерпретировать как ограничение объема каких-либо заявленных вариантов реализации. Кроме того, литература, цитируемая в настоящем описании, и все источники литературы, перечисленные ниже, полностью встроены в настоящую заявку путем, как если бы они были изложены в настоящем описании.
Ссылки
Abbink, P., Lemckert, A.A., Ewald, B.A., Lynch, D.M., Denholtz, M., et al. 2007. Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J Virol, 81:4654-4663.
Alonso, P.L., Sacarlal, J., Aponte, J.J., Leach, A., Macete, E., et al. 2004. Efficacy of the RTS,S/AS02A vaccine against Plasmodium falciparum infection and disease in young African children: randomised controlled trial. Lancet. 364:1411-1420.
Alonso, P.L., Sacarlal, J., Aponte, J.J., Leach, A., Macete, E., et al. 2005. Duration of protection with RTS,S/AS02A malaria vaccine in prevention of Plasmodium falciparum disease in Mozambican children: single-blind extended follow-up of a randomised controlled trial. Lancet. 366:2012-2018.
Anderson, R.J., Hannan, C.M., Gilbert, S.C., Laidlaw, S.M., Sheu, E.G., et al. 2004. Enhanced CD8+ T cell immune responses and protection elicited against Plasmodium berghei malaria by prime boost immunization regimens using a novel attenuated fowlpox virus. J Immunol. 172:3094-3100.
Athappilly, F.K., Murali, R., Rux, J.J., Cai, Z. & Burnett, R.M. The refined crystal structure of Hexon, the major coat protein of adenovirus type 2, at 2.9 A resolution. Journal of molecular biology 242, 430-455 (1994).
Barrat, F.J., Meeker, T., Gregorio, J., Chan, J.H., Uematsu, S., et al. Nucleic acids of mammalian origin can act as endogenous ligands for Toll-like receptors and may promote systemic lupus erythematosus. J Exp Med. 202, 1131-1139 (2005)
Bejon, P., Lusingu, J., Olotu, A., Leach, A., Lievens, M., et al. 2008. Efficacy of RTS,S/AS01 E vaccine against malaria in children 5 to 17 months of age. N Engl J Med. 359:2521-2532.
Belousova, N., Krendelchtchikova, V., Curiel, D.T. & Krasnykh, V. Modulation of adenovirus vector tropism via incorporation of polypeptide ligands into the Fiber protein. Journal of virology 76, 8621-8631 (2002).
Bergelson, J.M. et al. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science (New York, N.Y 276, 1320-1323 (1997).
Bewley, M.C., Springer, K., Zhang, Y.B., Freimuth, P. & Flanagan, J.M. Structural analysis of the mechanism of adenovirus binding to its human cellular receptor, CAR. Science (New York, N.Y 286, 1579-1583 (1999).
Bruña-Romero, O., Schmieg, J., Del Val, M., Buschle, M. & Tsuji, M. The dendritic cell-specific chemokine, dendritic cell-derived CC chemokine 1, enhances protective cell-mediated immunity to murine malaria. J Immunol 170, 3195-3203 (2003). Hong, S.S., Karayan, L, Tournier, J., Curiel, D.T. & Boulanger, P.A. Adenovirus type 5 Fiber knob binds to MHC class I alpha2 domain at the surface of human epithelial and B lymphoblastoid cells. The EMBO journal 16, 2294-2306 (1997).
Bruña-Romero, O., Gonzalez-Aseguinolaza, G., Hafalla, J.C., Tsuji, M., and Nussenzweig, R.S. 2001. Complete, long-lasting protection against malaria of mice primed and boosted with two distinct viral vectors expressing the same plasmodial antigen. Proc Natl Acad Sci USA. 98:11491-11496.
Bruña-Romero, O., Rocha, C.D., Tsuji, M. & Gazzinelli, R.T. Enhanced protective immunity against malaria by vaccination with a recombinant adenovirus encoding the circumsporozoite protein of Plasmodium lacking the GPI-anchoring motif. Vaccine 22, 3575-3584 (2004).
Calvo-Calle, J.M., Oliveira, G.A., Watta, C.O., Soverow, J., Parra-Lopez, C., et al. 2006. A linear peptide containing minimal T- and B-cell epitopes of Plasmodium falciparum circumsporozoite protein elicits protection against transgenic sporozoite challenge. Infect Immun. 74:6929-6939.
Clyde, D.F., Most, H., McCarthy, V.C., and Vanderberg, J.P. 1973. Immunization of man against sporozite-induced falciparum malaria. Am J Med Sci. 266:169-177.
Chroboczek, J., Ruigrok, R.W. & Cusack, S. Adenovirus Fiber. Current topics in microbiology and immunology 199 (Pt 1), 163-200 (1995).
Chu, Y., Heistad, D., Cybulsky, M.I. & Davidson, B.L. Vascular cell adhesion molecule-1 augments adenovirus-mediated gene transfer. Arterioscler Thromb Vase Biol 21, 238-242(2001).
Crawford-Miksza, L. & Schnurr, D.P. Analysis of 15 adenovirus Hexon proteins reveals the location and structure of seven hypervariable regions containing serotype-specific residues. Journal of virology 70, 1836-1844 (1996).
Crompton, J., Toogood, C.I., Wallis, N. & Hay, R.T. Expression of a foreign epitope on the surface of the adenovirus Hexon. The Journal of general virology 75 (Pt 1), 133-139(1994).
Crystal, R.G. Transfer of genes to humans: early lessons and obstacles to success. Science (New York, N.Y 270, 404-410 (1995).
Douglas, J.T. Adenoviral vectors for gene therapy. Molecular biotechnology 36, 71-80 (2007).
Edelman, R., Hoffman, S.L, Davis, J.R., Beier, M., Sztein, M.B., et al. 1993. Long-term persistence of sterile immunity in a volunteer immunized with X-irradiated Plasmodium falciparum sporozoites. J Infect Dis. 168:1066-1070.
Grillot, D., Valmori, D., Lambert, P.M., Corradin, G., and Del Giudice, G. 1993. Presentation of T cell epitopes assembled as multiple-antigen peptides to murine and human T lymphocytes. Infect Immun. 61:3064-3067.
Graham, F.L & Prevec, L. Adenovirus-based expression vectors and recombinant vaccines. Biotechnology (Reading, Mass 20, 363-390 (1992).
Gwadz, R.W., Cochrane, A.H., Nussenzweig, V., and Nussenzweig, R.S. 1979. Preliminary studies on vaccination of rhesus monkeys with irradiated sporozoites of Plasmodium knowlesi and characterization of surface antigens of these parasites. Bull World Health Organ.57 Suppl 1:165-173.
Hackett, N.R. et al. Use of quantitative TaqMan real-time PCR to track the time-dependent distribution of gene transfer vectors in vivo. Mol Ther2, 649-656 (2000).
Harvey, B.G. et al. Airway epithelial CPTR mRNA expression in cystic fibrosis patients after repetitive administration of a recombinant adenovirus. The Journal of clinical investigation 104, 1245-1255 (1999).
Heemskerk, B. et al. Adenovirus-specific CD4+ T cell clones recognizing endogenous antigen inhibit viral replication in vitro through cognate interaction. J Immunol 177, 8851-8859 (2006).
Henry, L.J., Xia, D., Wilke, M.E., Deisenhofer, J. & Gerard, R.D. Characterization of the knob domain of the adenovirus type 5 Fiber protein expressed in Escherichia coli. Journal of virology 68, 5239-5246 (1994).
Hong, S.S., Habib, N.A., Franqueville, L, Jensen, S. & Boulanger, P.A. Identification of adenovirus (ad) penton base neutralizing epitopes by use of sera from patients who had received conditionally replicative ad (addl1520) for treatment of liver tumors. Journal of virology 77, 10366-10375(2003).
Kester, K.E., Cummings, J.F., Ockenhouse, C.F., Nielsen, R., Hall, B.T., et al. 2008. Phase 2a trial of 0, 1, and 3 month and 0, 7, and 28 day immunization schedules of malaria vaccine RTS,S/AS02 in malaria-naive adults at the Walter Reed Army Institute of Research. Vaccine. 26:2191-2202.
Kirby, I. et al. Mutations in the DG loop of adenovirus type 5 Fiber knob protein abolish highaffinity binding to its cellular receptor CAR. Journal of virology 73, 9508-9514 (1999).
Koizumi, N., Mizuguchi, H., Utoguchi, N., Watanabe, Y. & Hayakawa, T. Generation of Fiber-modified adenovirus vectors containing heterologous peptides in both the HI loop and C terminus of the Fiber knob. The journal of gene medicine 5, 267-276 (2003).
Kozak M. 1987. An analysis of 5'-noncoding sequences from 699 vertebrate messenger RNAs. Nucleic Acids Res. 15:8125-148.
Krause, A., Joh, J.H., Hackett, N.R., Roelvink, P.W., Bruder, J.T., et al. 2006. Epitopes expressed in different adenovirus capsid proteins induce different levels of epitope-specific immunity. J Virol. 80:5523-5530.
Labow, D., Lee, S., Ginsberg, R.J., Crystal, R.G. & Korst, R.J. Adenovirus vector-mediated gene transfer to regional lymph nodes. Human gene therapy 11, 759-769 (2000).
Leen, A.M. et al. Identification of Hexon-specific CD4 and CD8 T cell epitopes for vaccine and immunotherapy. Journal of virology 82, 546-554 (2008).
Leopold, P.L. & Crystal, R.G. Intracellular trafficking of adenovirus: many means to many ends. Advanced drug delivery reviews 59, 810-821 (2007).
Mastrangeli, A. et al. "Sero-switch" adenovirus-mediated in vivo gene transfer: circumvention of anti-adenovirus humoral immune defenses against repeat adenovirus vector administration by changing the adenovirus serotype. Human gene therapy 7, 79-87 (1996).
Mathias, P., Wickham, T., Moore, M. & Nemerow, G. Multiple adenovirus serotypes use alpha v integrins for infection. Journal of virology 68, 6811-6814 (1994).
McConnell, M.J., Danthinne, X., and Imperiale, M.J. 2006. Characterization of a permissive epitope insertion site in adenovirus Hexon. J Virol. 80:5361-5370.
Meier, O. & Greber, U.F. Adenovirus endocytosis. The journal of gene medicine 5, 451-462 (2003).
Miyazawa, N. et al. Fiber swap between adenovirus subgroups B and C alters intracellular trafficking of adenovirus gene transfer vectors. Journal of virology 73, 6056-6065(1999).
Miyazawa, N., Crystal, R.G. & Leopold, P.L. Adenovirus serotype 7 retention in a late endosomal compartment prior to cytosol escape is modulated by Fiber protein. Journal of virology 75, 1387-1400 (2001).
Mizuguchi, H. & Hayakawa, T. Targeted adenovirus vectors. Human gene therapy 15, 1034-1044 (2004).
Nakano, M.Y., Boucke, K., Suomalainen, M., Stidwill, R.P. & Greber, U.F. The first step of adenovirus type 2 disassembly occurs at the cell surface, independently of endocytosis and escape to the cytosol. Journal of virology 74, 7085-7095 (2000).
Nicklin, S.A. et al. Ablating adenovirus type 5 Fiber-CAR binding and HI loop insertion of the SIGYPLP peptide generate an endothelial cell-selective adenovirus. Mol Ther 4, 534-542 (2001).
Noureddini, S.C. & Curiel, D.T. Genetic targeting strategies for adenovirus. Molecular pharmaceutics 2, 341-347 (2005).
Nussenzweig, R.S., Vanderberg, J., Most, H., and Orton, C. 1967. Protective immunity produced by the injection of x-irradiated sporozoites of plasmodium berghei. Nature. 216:160-162.
Nussenzweig, R.S. & Long, C.A. Malaria vaccines: multiple targets. Science (New York, N.Y 265, 1381-1383(1994).
Onion, D. et al. The CD4+ T cell response to adenovirus is focused against conserved residues within the Hexon protein. The Journal of general virology 88, 2417-2425 (2007).
Ophorst, O.J., Radosevic, K., Havenga, M.J., Pau, M.G., Holterman, L., et al. 2006. Immunogenicity and protection of a recombinant human adenovirus serotype 35-based malaria vaccine against Plasmodium yoelii in mice. Infect Immun. 74:313-320.
Oualikene, W., Gonin, P. & Eloit, M. Short and long term dissemination of deletion mutants of adenovirus in permissive (cotton rat) and non-permissive (mouse) species. The Journal of general virology 75 (Pt 10), 2765-2768 (1994).
Priddy, F.H., Brown, D., Kublin, J., Monahan, K., Wright, D.P., et al. 2008. Safety and immunogenicity of a replication-incompetent adenovirus type 5 HIV-1 clade B gag/pol/nef vaccine in healthy adults. Clin Infect Dis. 46:1769-1781.
Roberts, M.M., White, J.L, Grutter, M.G. & Burnett, R.M. Three-dimensional structure of the adenovirus major coat protein Hexon. Science (New York, N.Y 232, 1148-1151 (1986).
Roberts, D.M., Nanda, A., Havenga, M.J., Abbink, P., Lynch, D.M., et al. 2006. Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immunity. Nature. 441:239-243.
Rodrigues, E.G., Zavala, F., Eichinger, D., Wilson, J.M., and Tsuji, M. 1997. Single immunizing dose of recombinant adenovirus efficiently induces CD8+ T cell-mediated protective immunity against malaria. J Immunol. 158:1268-1274.
Rodrigues, E.G., Zavala, F., Nussenzweig, R.S., Wilson, J.M. & Tsuji, M. Efficient induction of protective anti-malaria immunity by recombinant adenovirus. Vaccine 16, 1812-1817(1998).
Roelvink, P.W., Mi Lee, G., Einfeld, D.A., Kovesdi, I. & Wickham, T.J. Identification of a conserved receptor-binding site on the Fiber proteins of CAR-recognizing adenoviridae. Science (New York, N.Y 286, 1568-1571 (1999).
Rux, J.J. & Burnett, R.M. Type-specific epitope locations revealed by X-ray crystallographic study of adenovirus type 5 Hexon. Mol Ther 1, 18-30 (2000).
Rux, J.J. & Burnett, R.M. Adenovirus structure. Human gene therapy 15, 1167-1176 (2004).
Roy, S. et al. Use of chimeric adenoviral vectors to assess capsid neutralization determinants. Virology 333, 207-214 (2005).
Silvie, O., Greco, C., Franetich, J.F., Dubart-Kupperschmitt, A., Hannoun, L., et al. 2006. Expression of human CD81 differently affects host cell susceptibility to malaria sporozoites depending on the Plasmodium species. Cell Microbiol. 8:1134-1146.
Sumida, S.M., Truitt, D.M., Lemckert, A.A., Vogels, R., Ousters, J.H., et al. 2005. Neutralizing antibodies to adenovirus serotype 5 vaccine vectors are directed primarily against the adenovirus Hexon protein. J Immunol. 174:7179-7185.
Sun, P., Schwenk, R., White, K., Stoute, J.A., Cohen, J., et al. 2003. Protective immunity induced with malaria vaccine, RTS,S, is linked to Plasmodium falciparum circumsporozoite protein-specific CD4+ and CD8+ T cells producing IFN-gamma. J Immunol. 171:6961-6967.
Tao, D., Barba-Spaeth, G., Rai, U., Nussenzweig, V., Rice, C.M., and Nussenzweig, R.S. 2005. Yellow fever 17D as a vaccine vector for microbial CTL epitopes: protection in a rodent malaria model. J Exp Med. 201:201-209.
Teramoto, S. et al. Investigation of effects of anesthesia and age on aspiration in mice through LacZ gene transfer by recombinant El-deleted adenovirus vectors. American journal of respiratory and critical care medicine 158, 1914-1919 (1998).
Top, F.H., Jr., Dudding, B.A., Russell, P.K. & Buescher, E.L. Control of respiratory disease in recruits with types 4 and 7 adenovirus vaccines. American journal of epidemiology 94, 142-146 (1971).
Top, F.H., Jr. Control of adenovirus acute respiratory disease in U.S. Army trainees. Yate J Biol Med 48, 185-195 (1975).
Tsuji, M., Romero, P., Nussenzweig, R.S., and Zavala, F. 1990. CD4+ cytolytic T cell clone confers protection against murine malaria. J Exp Med. 172:1353-1357.
Tsuji, M., Rodrigues, E.G. & Nussenzweig, S. Progress toward a malaria vaccine: efficient induction of protective anti-malaria immunity. Biol Chem 382, 553-570 (2001).
Wickham, T.J., Mathias, P., Cheresh, D.A. & Nemerow, G.R. Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment. Cell 73, 309-319(1993).
Wohlfart, C. Neutralization of adenoviruses: kinetics, stoichiometry, and mechanisms. Journal of virology 62, 2321-2328 (1988).
Worgall, S. et al. Modification to the capsid of the adenovirus vector that enhances dendritic cell infection and transgene-specific cellular immune responses. Journal of virology 78, 2572-2580 (2004).
Worgall, S., Krause, A., Rivara, M., Hee, K.K., Vintayen, E.V., et al. 2005. Protection against P. aeruginosa with an adenovirus vector containing an OprF epitope in the capsid. J Clin Invest. 115:1281-1289.
Worgall, S., Krause, A., Qiu, J., Joh, J., Hackett, N.R., and Crystal, R.G. 2007. Protective immunity to pseudomonas aeruginosa induced with a capsid-modified adenovirus expressing P. aeruginosa OprF. J Virol. 81:13801-13808.
Wu, H., Dmitriev, I., Kashentseva, E., Seki, T., Wang, M., et al. 2002. Construction and characterization of adenovirus serotype 5 packaged by serotype 3 Hexon. J Virol, 76:12775-12782.
Wu, H. et al. Identification of sites in adenovirus Hexon for foreign peptide incorporation. Journal of virology 79, 3382-3390 (2005).
Xia, D., Henry, L, Gerard, R.D. & Deisenhofer, J. Structure of the receptor binding domain of adenovirus type 5 Fiber protein. Current topics in microbiology and immunology 199 (Pt 1), 39-46 (1995).
Yang, Y., Li, Q., Ertl, H.C. & Wilson, J.M. Cellular and humoral immune responses to viral antigens create barriers to lung-directed gene therapy with recombinant adenoviruses. Journal of virology 69, 2004-2015 (1995).
Youil, R., Toner, T.J., Su, Q., Chen, M., Tang, A., et al. 2002. Hexon gene switch strategy for the generation of chimeric recombinant adenovirus. Hum Gene Ther. 13:311-320.
Реферат
Настоящее изобретение относится к модификациям белков аденовирусов с целью усиления иммунного ответа на трансген рекомбинантного аденовируса и преодоления предсуществующего иммунитета к аденовирусам. Некоторые варианты реализации относятся к рекомбинантному аденовирусу, полученному из плазмидного вектора на основе рекомбинантного аденовируса. Указанный плазмидный вектор на основе рекомбинантного аденовируса содержит последовательность нуклеотида, кодирующую белок circumsporozoite плазмодия, функционально связанную с гетерологичным промотором, и модифицированный белок капсида или сердцевины. При этом иммуногенный эпитоп спорозоита плазмодия встроен по меньшей мере в часть белка капсида или сердцевины или заменяет ее. Другие варианты реализации относятся к фармацевтической композиции или вакцине против малярии, содержащей рекомбинантный аденовирус. Дополнительные варианты реализации включают способ лечения, профилактики или диагностики малярии, включающий введение терапевтического количества указанной фармацевтической композиции или вакцины против малярии. Предложенная группа изобретений может быть использована в медицине. 6 н. и 20 з.п. ф-лы, 57 ил., 3 табл., 5 пр.
Формула
ген белка circumsporozoite плазмодия (Plasmodium) или его антигенной части, функционально связанный с последовательностью гетерологичного промотора, и
один или более генов модифицированных белков капсида и/или сердцевины, причем последовательность иммуногенного эпитопа белка circumsporozoite плазмодия была встроена по меньшей мере в часть одного или нескольких генов белков капсида или сердцевины или замещает указанную часть,
где последовательность иммуногенного эпитопа представляет собой последовательность эпитопа, распознаваемого В-лимфоцитами, выбранную из (NANP)12, (NANP)14, (NANP)16, (NANP)18, (NANP)20 или (NANP)22.
a) встроена в последовательность HVR1 или HVR5; или
b) замещает часть последовательности HVR1 или HVR5.
ген белка circumsporozoite плазмодия (Plasmodium) или его антигенной части, функционально связанный с последовательностью гетерологичного промотора, и
один или более генов модифицированных белков капсида и/или сердцевины, причем последовательность иммуногенного эпитопа белка circumsporozoite плазмодия была встроена по меньшей мере в часть одного или нескольких генов белков капсида или сердцевины или замещает указанную часть,
где ген белка сердцевины содержит ген белка pVII, и последовательность эпитопа, распознаваемого CD4+Т-лимфоцитами, встроена в ген белка pVII.
белок cirmumsporozoite плазмодия или его антигенную часть, функционально связанные с последовательностью гетерологичного промотора, и
модифицированный белок капсида или сердцевины, в котором иммуногенный эпитоп белка circumsporozoite плазмодия встроен по меньшей мере в часть белка капсида и/или сердцевины или замещает по меньшей мере часть белка капсида и/или сердцевины,
где последовательность иммуногенного эпитопа представляет собой последовательность эпитопа, распознаваемого В-лимфоцитами, выбранную из (NANP)12, (NANP)14, (NANP)16, (NANP)18, (NANP)20 или (NANP)22.
первой примирующей дозы первого рекомбинантного вируса, и
последующей бустер-дозы второго рекомбинантного вируса,
где указанный первый рекомбинантный аденовирус получен из плазмидного вектора на основе рекомбинантного аденовируса, и где указанный плазмидный вектор на основе рекомбинантного аденовируса включает последовательность нуклеотидов, кодирующую белок circumsporozoite плазмодия или его антигенную часть, функционально связанные с последовательностью гетерологичного промотора, и
где указанный второй рекомбинантный аденовирус получен из плазмидного вектора на основе рекомбинантного аденовируса, и где указанный плазмидный вектор на основе рекомбинантного аденовируса включает последовательность нуклеотидов, кодирующую белок circumsporozoite плазмодия или его антигенную часть, функционально связанные с последовательностью гетерологичного промотора, и модифицированный белок капсида или сердцевины, где иммуногенный эпитоп белка circumsporozoite плазмодия встроен по меньшей мере в часть белка капсида и/или сердцевины или замещает указанную часть, и где последовательность иммуногенного эпитопа представляет собой последовательность эпитопа, распознаваемого В-лимфоцитами, выбранную из (NANP)12, (NANP)14, (NANP)16, (NANP)18, (NANP)20 или (NANP)22.
Документы, цитированные в отчёте о поиске
Способы вакцинации против малярии
Комментарии