Микроорганизм с улучшенной способностью продуцировать l-лизин и способ получения l-лизина с использованием этого микроорганизма - RU2668176C1
Код документа: RU2668176C1
Чертежи






















Описание
Область изобретения
Настоящее изобретение относится к новому модифицированному полипептиду сигма-фактора А РНК-полимеразы (SigA), кодирующему его полинуклеотиду, микроорганизму, содержащему этот полипептид, и способу получения L-лизина с использованием этого микроорганизма.
Предшествующий уровень техники
Штаммы микроорганизмов с повышенной продуктивностью могут быть созданы путем открытия и соответствующего использования генетических факторов, которые непосредственно/опосредованно вовлечены на ранних стадиях гликолиза, для получения штаммов, продуцирующих полезные продукты, такие как аминокислоты, в большом масштабе. Репрезентативные примеры технологий, которые можно применять для этого, могут включать глобальное репрограммирование транскриптома (gTME), которое представляет собой подход к регуляции экспрессии всех генов в клетке путем индуцирования случайного мутагенеза рекрутирующих белков РНК-полимеразы. Например, исследовательская группа из Массачусетского технологического института добилась успеха в значительном повышении способности Е.coli продуцировать тирозин с использованием технологии gTME (Патент США No. 8735132).
РНК-полимераза, применяемая на стадии транскрипции у микроорганизмов, представляет собой макромолекулу, состоящую из 5 небольших субъединиц, а именно двух факторов α, одного фактора β, одного фактора β' и одного фактора ω, и ее холофермент обозначают как α2ββ'ω. Сигма(σ)-факторы вместе с холоферментом представляют собой факторы, необходимые для стадии инициации транскрипции, которые обеспечивают специфичность связывания РНК-полимеразы с промотором. Штамм Corynebacterium имеет 7 типов сигма-факторов (SigA, SigB, SigC, SigD, SigE, SigH и SigM), которые регулируют транскрипцию определенных групп генов в соответствии с изменениями в окружающей среде (Journal of Biotechnology 154, 2011, 101-113). В частности, SigA представляет собой главный регулятор среди сигма-факторов, вовлеченных в регуляцию большинства генов "домашнего хозяйства" и "коровых" генов. Согласно сообщениям по результатам предыдущих исследований, были предприняты попытки улучшить способность продуцировать целевые вещества путем случайного мутагенеза SigA (Metabolic Engineering 9, 2007, 258-267), и также было сообщение, касающееся исследования повышения способности продуцировать L-лизин с использованием штамма Corynebacterium (Международная публикация No. WO 2003-054179).
Описание
Техническая задача
В этих обстоятельствах авторы настоящего изобретения приложили усилия для создания микроорганизма, обладающего способностью продуцировать L-лизин в повышенной концентрации без ингибирования роста клетки-хозяина. В результате они подтвердили, что микроорганизм рода Corynebacterium с повышенной способностью продуцировать L-лизин можно создать путем введения нового модифицированного типа полипептида SigA РНК-полимеразы в микроорганизм после разработки микроорганизма, в чем и заключается настоящее изобретение.
Техническое решение
Задача настоящего изобретения заключается в том, чтобы предложить модифицированный полипептид, состоящий из аминокислотной последовательности SEQ ID NO: 2, обладающей активностью сигма-фактора А РНК-полимеразы, в котором часть аминокислот полипептида заменена.
Другая задача настоящего изобретения заключается в том, чтобы предложить полинуклеотид, кодирующий этот модифицированный полипептид.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить трансформированный микроорганизм, кодирующий этот полипептид.
Еще одна задача настоящего изобретения заключается в том, чтобы предложить способ получения L-лизина, включающий получение культуральной среды путем культивирования микроорганизма и выделения L-лизина из культуральной среды или из культивируемого микроорганизма.
Полезные эффекты изобретения
Настоящее изобретение позволяет подтвердить, может ли какой-либо модифицированный полипептид, имеющий модифицированный сигма-фактор А РНК-полимеразы, вызывать повышение способности продуцировать L-лизин. Кроме того, на основании вышеизложенного, микроорганизм, способный экспрессировать соответствующий модифицированный полипептид, обладает в высшей степени превосходной способностью продуцировать L-лизин. Соответственно, ожидается, что микроорганизм обеспечит эффекты по снижению производственных затрат, удобству в получении и так далее в промышленном аспекте.
Наилучшее воплощение изобретения
Для решения вышеизложенных задач в одном аспекте настоящего изобретения предложен модифицированный полипептид, обладающий активностью нового сигма-фактора А РНК-полимеразы.
Как его используют здесь, термин "сигма-фактор A (SigA) РНК-полимеразы" относится к фактору инициации транскрипции, который действует вместе с РНК-полимеразой, и он представляет собой белок (SigA), соответствующий одному из сигма-факторов. Сигма-факторы вовлечены в регуляцию транскрипции путем взаимодействия с ДНК выше по ходу транскрипции (UP-элемент), которая присутствует выше по ходу транскрипции от определенного промотора, и различными транскрипционными факторами. В частности, SigA известен как главный регулятор, контролирующий большинство "коровых" генов. Информацию по белку SigA можно получить из известных баз данных, таких как NCBI GenBank, например по белку с № доступа NP_601117. В частности, белок SigA может включать аминокислотную последовательность SEQ ID NO: 2, но последовательность не ограничена этим, при условии что белок обладает такой же активностью как SigA по настоящему изобретению.
Как его используют здесь, термин "модифицированный полипептид" относится к полипептиду, в котором аминокислотная последовательность полипептида дикого типа частично или полностью заменена. В настоящем описании модифицированный полипептид относится к полипептиду, обладающему активностью сигма-фактора A (SigA) РНК-полимеразы, в котором аминокислотная последовательность полипептида, имеющего сигма-фактор А (SigA) РНК-полимеразы, частично заменена, таким образом имеющему аминокислотную последовательность, отличную от таковой аминокислотной последовательности дикого типа. То есть, в настоящем изобретении предложен модифицированный полипептид SigA, который вносит вклад в способность продуцировать L-лизин, вместо полипептида SigA дикого типа.
В частности, модифицированный полипептид может представлять собой полипептид, обладающий активностью сигма-фактора А РНК-полимеразы, в котором по меньшей мере одна аминокислота, выбранная из группы, состоящей из аминокислот в следующих положениях полипептида, состоящего из аминокислотной последовательности SEQ ID NO: 2, заменена на другую аминокислоту:
136-ая аминокислота, 254-ая аминокислота, 268-ая аминокислота, 281-ая аминокислота, 381-ая аминокислота, 429-ая аминокислота и с 445-ой по 495-ую аминокислоты относительно инициирующего метионина, обозначенного как первая аминокислота.
То есть, модифицированный полипептид может представлять собой полипептид, в котором аминокислота по меньшей мере в одном положении из 57 аминокислотных положений для модификации (136-ая, 254-ая, 268-ая, 281-ая, 381-ая, 429-ая и от 445-ой до 495-ой) заменена на другую аминокислоту.
Более конкретно, среди аминокислот от 445-ой до 495-ой, по меньшей мере одна аминокислота, выбранная из группы, состоящей из 447-ой аминокислоты, 451-ой аминокислоты, 455-ой аминокислоты, 479-ой аминокислоты, 483-ей аминокислоты, 488-ой аминокислоты и 491-ой аминокислоты, может быть заменена на другую аминокислоту, но без ограничения этим.
В частности, аминокислотная замена может представлять собой комбинацию по меньшей мере одной из следующих аминокислотных замен:
замена 136-ой аминокислоты на глицин (D136G); замена 254-ой аминокислоты на аспарагин (I254N); замена 268-ой аминокислоты на серии (A268S); замена 281-ой аминокислоты на серии (T281S); замена 381-ой аминокислоты на аргинин (L381R); замена 429-ой аминокислоты на аргинин (Q429R); замена 447-ой аминокислоты на гистидин (L447H); замена 451-ой аминокислоты на изолейцин (L451I); замена 455-ой аминокислоты на валин (M455V); замена 479-ой аминокислоты на аргинин (K479R); замена 483-ей аминокислоты на аргинин (K483R); замена 488-ой аминокислоты на треонин (S488T) и замена 491-ой аминокислоты на аргинин (Q491R) относительно инициирующего метионина.
Более конкретно, полипептид может представлять собой модифицированный полипептид, в котором 136-ая аминокислота и 281-ая аминокислота заменены на глицин и серии, соответственно (D136G, T281S); 254-ая аминокислота на аспарагин (I254N); 268-ая аминокислота на серии (A268S); 381-ая аминокислота на аргинин (L381R); 429-ая аминокислота на аргинин (Q429R); 447-ая аминокислота на гистидин (L447H); 451-ая и 491-ая аминокислоты на изолейцин и аргинин, соответственно (L451I, Q491R); 455-ая аминокислота на валин (M455V); 479-ая аминокислота на аргинин (K479R); 483-ая аминокислота на аргинин (K483R) и 488-ая аминокислота на треонин (S488T) относительно инициирующего метионина полипептида, состоящего из аминокислотной последовательности SEQ ID NO: 2, или имеет место комбинация по меньшей мере из одной из представленных выше 11 аминокислотных замен.
Согласно одному примеру воплощения настоящего изобретения, модифицированный полипептид может представлять собой полипептид, имеющий по меньшей мере одну аминокислотную последовательность из аминокислотных последовательностей SEQ ID NO: 12-22.
Модифицированный полипептид по настоящему изобретению включает, без ограничения, не только аминокислотные последовательности, представленные SEQ ID NO: 12-22, но также аминокислотные последовательности, имеющие гомологию с вышеизложенными аминокислотными последовательностями 70% или больше, более конкретно 80% или больше, еще более конкретно 90% или больше или еще более конкретно 99%, где эти белки вносят вклад в улучшение способности продуцировать L-лизин по сравнению с таковой белка SigA дикого типа. Очевидно, что любая аминокислотная последовательность, обладающая биологической активностью по существу такой же или соответствующей таковой у белка, имеющего аминокислотную последовательность модифицированного белка SigA, дополнительно к вышеуказанной гомологии последовательности, также будет попадать в объем настоящего изобретения, хотя аминокислотная последовательность может иметь делецию, модификацию, замену или вставку в части последовательности.
Как его используют здесь, термин "гомология" относится к степени идентичности между двумя различными последовательностями нуклеотидов или аминокислотных остатков в определенном участке сравнения нуклеотидных или аминокислотных последовательностей гена, кодирующего белок, после выравнивания обеих последовательностей до максимального соответствия. Когда гомология является достаточно высокой, продукты экспрессии соответствующего гена могут иметь такую же или похожую активность. Гомологию можно определить с использованием программы для сравнения последовательностей, известной в области техники, например BLAST (NCBI), CLC Main Workbench (CLC bio), MegAlign™ (DNASTAR Inc.) и им подобных.
Другой аспект настоящего изобретения включает полинуклеотид, кодирующий модифицированный полипептид, или вектор, включающий этот полинуклеотид.
Как его используют здесь, термин "полинуклеотид" относится к полимеру из нуклеотидов, простирающемуся в длину в виде цепи посредством ковалентной связи между нуклеотидными единицами, то есть цепи ДНК или РНК, имеющей по меньшей мере определенную длину, и более конкретно он относится к полинуклеотидному фрагменту, кодирующему модифицированный полипептид.
В настоящем изобретении ген, кодирующий аминокислотную последовательность сигма-фактора A (SigA) РНК-полимеразы, представляет собой ген rpoD, и более конкретно, ген, имеющий происхождение из Corynebacterium glutamicum. На основании вырожденности генетического кода нуклеотидные последовательности, кодирующие одну и ту же аминокислоту и ее варианты, попадают в объем настоящего изобретения и, в частности, они могут быть представлены SEQ ID NO: 1, но не ограничиваться этим.
Дополнительно, в отношении модифицированного полипептида, нуклеотидные последовательности, кодирующие одну и ту же аминокислоту и ее варианты, попадают в объем настоящего изобретения на основании вырожденности генетического кода. Более конкретно, любая нуклеотидная последовательность, кодирующая любую из аминокислотных последовательностей 12-22 и их варианты, может быть включена, но без ограничения ими.
Еще в одном аспекте настоящего изобретения предложена клетка-хозяин, включающая полинуклеотид, кодирующий модифицированный полипептид, и микроорганизм, трансформированный вектором, включающим полинуклеотид, кодирующий модифицированный полипептид. Более конкретно, введение может быть выполнено путем трансформации, но без ограничения этим.
Более конкретно, микроорганизм, включающий модифицированный полипептид SigA может улучшать способность продуцировать L-лизин без ингибирования роста клетки-хозяина и, таким образом, может получать L-лизин с высоким выходом по сравнению с микроорганизмом, включающим полипептид SigA дикого типа.
Как его используют здесь, термин "вектор" относится к любому носителю для клонирования и/или переноса нуклеотидов в клетку-хозяина. Вектор может представлять собой репликон для обеспечения возможности репликации фрагмента, комбинированного с другим фрагментом ДНК. Термин "репликон" относится к любой генетической единице, действующей как самореплицирующаяся единица для репликации ДНК in vivo, то есть произвольные генетические единицы, способные к репликации путем саморегуляции (например плазмиды, фаги, космиды, хромосомы и вирусы). Как его используют здесь, термин "вектор" может включать вирусные и невирусные носители для введения нуклеотидов в клетку-хозяина in vitro, ex vivo или in vivo, и также может включать мини-сферическую ДНК. Например, вектор может представлять собой плазмиду без последовательности бактериальной ДНК. Дополнительно вектор может включать транспозоны (Annu Rev Genet. 2003; 37: 3-29) или искусственные хромосомы. Более конкретно, вектор может включать векторы pACYC177, pACYC184, pCL1920, pECCG117, pUC19, pBR322, pDZ и pMW118, но без ограничения ими.
Как его используют здесь, термин "трансформация" относится к процессу введения гена в клетку-хозяина, тем самым давая возможность экспрессии гена в клетке-хозяине. Трансформированный ген может включать, без ограничения, как ген, вставленный в хромосому клетки-хозяина, так и ген, локализованный за пределами хромосомы, при условии, что они могут экспрессироваться в клетке-хозяине.
Дополнительно ген может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой полинуклеотидную конструкцию, включающую все основные элементы, необходимые для самостоятельной экспрессии. Экспрессионная кассета обычно может включать промотор, функционально связанный с геном, сигнал терминации транскрипции, домен связывания с рибосомой и сигнал терминации трансляции. Экспрессионная кассета может быть в форме вектора экспрессии, способного к саморепликации. Дополнительно полинуклеотид может быть введен в клетку-хозяина как есть или в форме полинуклеотидной конструкции, и может быть функционально связан с последовательностью, необходимой для его экспрессии в клетке-хозяине, но без ограничения этим.
Клетка-хозяин или микроорганизм могут представлять собой любую клетку-хозяин или микроорганизм, которые включают полинуклеотид, кодирующий модифицированный полипептид, или которые трансформированы вектором, включающим полинуклеотид, кодирующий модифицированный полипептид, таким образом обладая способностью экспрессировать модифицированный полипептид, и для задач настоящего изобретения клетка-хозяин или микроорганизм могут представлять собой любую клетку-хозяин или микроорганизм, которые могут продуцировать L-лизин посредством включения SigA-модифицированного полипептида. Конкретные примеры микроорганизма могут включать микробные штаммы, принадлежащие роду Escherichia, роду Serratia, роду Erwinia, роду Enterobacteria, роду Salmonella, роду Streptomyces, роду Pseudomonas, роду Brevibacterium, роду Corynebacterium и так далее, и конкретно микроорганизм, принадлежащий роду Corynebacterium, и более конкретно Corynebacterium glutamicum, но без ограничения ими.
В примере воплощения по настоящему изобретению в различные штаммы Corynebacterium glutamicum, обладающие способностью продуцировать L-лизин (KCCM11016P, KFCC10750, KCCM10770P и CJ3P), вводили модифицированный полипептид, который включает одну любую аминокислотную последовательность, выбранную из аминокислотных последовательностей SEQ ID NO: 12-22, и сравнивали их продуктивности по L-лизину. В результате было подтверждено, что все штаммы, в которые вводили модифицированный полипептид, демонстрировали повышенный уровень продукции L-лизина по сравнению со штаммом, включающим полипептид SigA дикого типа (Примеры 7-10 и Таблицы 8-11). Эти результаты позволяют предположить, что микроорганизм, включающий модифицированный полипептид по настоящему изобретению, представляет собой штамм с усиленной способностью продуцировать L-лизин, и этот микроорганизм обеспечивает экономическое преимущество при получении L-лизина с высоким выходом при культивировании этого микроорганизма.
Авторы настоящего изобретения обозначили штамм с усиленной способностью продуцировать L-лизин как таковой "KCCM11016P::SigA(L447H)", как "Corynebacterium glutamicum СА01-2277" и депонировали его в Корейском центре культур микроорганизмов, признанном международным органом депонирования согласно Будапештскому договору, 22 ноября 2013 года с номером доступа KCCM11479P.
Еще в одном аспекте настоящего изобретения предложен способ получения L-лизина, включающий культивирование микроорганизма рода Corynebacterium, описанного выше, и выделение L-лизина из культивируемого микроорганизма или культуральной среды.
Как его используют здесь, термин "культивирование" относится к выращиванию микроорганизма в подходящих и искусственно контролируемых условиях окружающей среды. Процесс культивирования по настоящему изобретению может быть выполнен на основе подходящих культуральной среды и условий культивирования, известных в области техники. Конкретные условия, такие как температура культивирования, время культивирования, рН культуральной среды и так далее, обычный специалист в данной области техники может определить на основании общих знаний или путем традиционного способа, известного в данной области техники, и соответственно отрегулировать подходящим образом. Более конкретно, эти известные способы культивирования подробно описаны в источниках [Chmiel; Bioprozesstechnik 1. Einfuhrung indie Bioverfahrenstechnik (Gustav Fischer Verlag, Stuttgart, 1991) и Storhas; Bioreaktoren und periphere Einrichtungen (Vieweg Verlag, Braunschweig / Wiesbaden, 1994)]. Дополнительно, способы культивирования могут включать периодическое культивирование, непрерывное культивирование и культивирование с подпиткой, и в особенности, культивирование можно осуществлять непрерывно в процессе культивирования с подпиткой или периодического культивирования с подпиткой, но без ограничения этим.
Среда, используемая при культивировании, должна соответствующим образом удовлетворять требованиям конкретных штаммов. Примеры источников углерода для использования в этой среде могут включать сахара и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол, и органические кислоты, такие как уксусная кислота, но без ограничения ими. Эти источники углерода можно использовать по отдельности или в комбинации, но без ограничения этим. Примеры источников азота для использования в среде могут включать пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, жидкий кукурузный экстракт, соевую муку и мочевину или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота также можно использовать по отдельности или в комбинации, но без ограничения этим. Примеры источников фосфора для использования в среде могут включать дигидрофосфат калия, гидроортофосфат калия, соответствующие натрий-содержащие соли и так далее, но без ограничения ими. Дополнительно среда может содержать соли металлов, такие как сульфат магния или сульфат железа, необходимые для роста. Наконец, вещества, необходимые для роста, такие как аминокислоты и витамины, также могут содержаться в дополнение к веществам, описанным выше. Дополнительно можно использовать предшественники, подходящие для культуральной среды. Эти источники можно добавлять в культуру подходящим способом во время культивирования путем периодического культивирования или непрерывного культивирования, но способы не ограничены этим.
Дополнительно, значение рН культуры может быть скорректировано во время культивирования путем добавления в культуру такого соединения, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота, подходящим образом. Во время культивирования можно добавлять пеногаситель, такой как полигликолевый сложный эфир жирной кислоты, для предупреждения образования пены. Дополнительно для поддержания аэробного состояния культуры в культуру может быть введен кислород или газ, содержащий кислород; или азот, водород или углекислый газ могут быть введены без введения газа для поддержания анаэробного или микроаэробного состояния культуры. Температура в культуре обычно может находиться в диапазоне от 27°С до 37°С, и более конкретно, от 30°С до 35°С. Культивирование можно продолжать до тех пор, пока не будет получено желаемое количество полезных веществ, и в особенности от 10 часов до 100 часов. L-лизин может выделяться в культуральную среду, в которой проводится культивирование, или может содержаться в клетках микроорганизма.
Дополнительно, в отношении способа получения L-лизина по настоящему изобретению, способ выделения L-лизина из культивируемого микроорганизма или из культуры широко известен в данной области техники. Способы выделения L-лизина могут включать фильтрацию, анионообменную хроматографию, кристаллизацию, HPLC (высокоэффективную жидкостную хроматографию) и так далее, но не ограничиваются ими.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Здесь и далее настоящее изобретение будет описано более подробно со ссылкой на следующие примеры. Однако эти примеры приведены исключительно для иллюстративных целей и не предназначены ограничивать объем настоящего изобретения.
Пример 1: Получение модифицированного полипептида SigA путем искусственного мутагенеза
В этом примере библиотеку векторов для первичной вставки в хромосому путем кроссинговера для получения модифицированного SigA получали следующим способом. Выполняли ПЦР (полимеразная цепная реакция) пониженной точности (error-prone PCR) для гена rpoD (SEQ ID NO: 1), кодирующего SigA Corynebacterium (SEQ ID NO: 2), и таким образом были получены фрагменты (1497 п.н.) вариантов гена rpoD с введенной случайным образом модификацией - нуклеотидной заменой. ПЦР пониженной точности проводили с помощью набора Genemorphll Random Mutagenesis Kit (Stratagene) с использованием геномной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы наряду с праймером 1 (SEQ ID NO: 3) и праймером 2 (SEQ ID NO: 4). ПЦР пониженной точности проводили таким образом, что модификации могли быть введены в амплифицированный фрагмент гена в соотношении от 0 до 4,5 мутаций на 1 т.п.н. амплифицированного фрагмента. ПЦР выполняли в течение 30 циклов в следующих условиях: денатурация при 96°С в течение 30 сек, отжиг при 53°С в течение 30 сек и полимеризация при 72°С в течение 2 мин.

Амплифицированные фрагменты гена соединяли с вектором pCR2.1-ТОРО (здесь и далее 'pCR2.1') с использованием набора для клонирования pCR2.1-TOPO ТА Cloning Kit (Invitrogen), трансформировали в Е.coli DH5a и высевали на твердую среду LB. Из трансформированных таким образом колоний выбрали двадцать, и их нуклеотидные последовательности анализировали после выделения из них плазмид. В результате было подтверждено, что мутации были введены в различные положения с частотой 1,5 мутаций/т.п.н. Плазмиды экстрагировали примерно из 20000 трансформированных колоний Е. Coli, и они получили название "библиотека pCR2.1-rpoD(mt)". Затем была получена плазмида, включающая ген rpoD дикого типа, для применения в качестве контроля. ПЦР проводили с использованием геномной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы наряду с праймером 1 (SEQ ID NO: 3) и праймером 2 (SEQ ID NO: 4) в таких же условиях как описано выше. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene), и плазмиды, полученные таким образом получили название "pCR2.1-rpoD (WT)".
Пример 2: Получение библиотеки штаммов с модифицированным SigA с повышенной способностью продуцировать L-лизин
Библиотеку pCR2.1-rpoD(mt), полученную с использованием штамма KCCM11016P (патент Кореи No. 10-0159812) в качестве родительского штамма, трансформировали путем гомологичной хромосомной рекомбинации, высевали на комплексную среду, содержащую канамицин (25 мг/л) и компоненты, описанные ниже, и из нее было получено примерно 25000 колоний. Колонии получили названия от "KCCM11016P/pCR2.1-rpoD(mt)-1" до "KCCM11016P/pCR2.1-rpoD(mt)-25000". Дополнительно, полученным таким образом вектором pCR2.1-rpoD(WT) трансформировали штамм KCCM11016P для получения контрольного штамма, и он получил название "KCCM11016P/pCR2.1-rpoD(WT)".
Полученные таким образом трансформанты будут иметь две копии гена rpoD в хромосоме. Однако, в библиотеку pCR2.1-rpoD(mt) и вектор pCR2.1-rpoD(WT) вставлены фрагменты гена rpoD без какого-либо промотора, и следовательно, когда они вставлены в хромосому штамма путем гомологичной рекомбинации, только одна копия из двух копий гена rpoD экспрессируется, таким образом давая возможность экспрессии модифицированного белка SigA или белка SigA дикого типа.
Комплексная среда (рН 7,0)
Глюкоза (10 г), пептон (10 г), экстракт говядины (5 г), дрожжевой экстракт (5 г), сердечно-мозговая вытяжка (18,5 г), NaCl (2,5 г), мочевина (2 г), сорбит (91 г), агар (20 г) (на основе 1 л дистиллированной воды)
Пример 3: Скрининг библиотеки штаммов с модифицированным SigA с улучшенной способностью продуцировать L-лизин
Примерно 25000 колоний, полученных в Примере 2, были соответственно инокулированы в селективную среду (300 мкл), содержащую компоненты, описанные ниже, и их культивировали в 96-луночном планшете с глубокими лунками при 32°С на скорости 1000 об/мин в течение 24 часов. Количество L-лизина, продуцируемого во время культивирования, анализировали нингидриновым методом (J. Biol. Слет. 1948. 176: 367-388). После завершения культивирования 10 мкл культурального супернатанта и 190 мкл раствора для нингидриновой реакции ((63% глицерин, 27% нингидриновый реактив (7,1 г/л в 0,5 М цитратном буфере, рН 5,5)) взаимодействовали при 65°С в течение 30 минут. Поглощение при длине волны 570 нм измеряли с помощью спектрофотометра и сравнивали с таковым контроля, то есть KCCM11016P/pCR2.1-rpoD(WT), и было выбрано примерно 935 модифицированных штаммов, демонстрирующих поглощение, увеличенное по меньшей мере на 10%. Другие колонии демонстрировали похожее или уменьшенное поглощение по сравнению с контролем.
Селективная среда (рН 8,0)
Глюкоза (10 г), сульфат аммония (5,5 г), MgSO4⋅7H2O (1,2 г), KH2PO4 (0,8 г), K2HPO4 (16,4 г), биотин (100 мкг), тиамин HCl (1000 мкг), кальций-пантотеновая кислота (2000 мкг) и никотинамид (2000 мкг) (на основе 1 л дистиллированной воды).
Вышеописанный способ неоднократно повторяли для выбранных 935 штаммов, и были выбраны лучшие 231 тип штаммов со способностью продуцировать L-лизин, улучшенной на 15% или больше по сравнению с таковой штамма KCCM11016P /pCR2.1-rpoD(WT).
Пример 4: Анализ способности продуцировать L-лизин KCCM11016P/dCR2.1-moD(mt)
231 тип штаммов, выбранных в Примере 3, анализировали в отношении их способности продуцировать L-лизин после их культивирования следующим способом.
Каждый из штаммов инокулировали в колбу на 250 мл с угловыми перегородками, содержащую 25 мл среды для посевной культуры, соответственно, и культивировали во встряхиваемом инкубаторе (200 об/мин) при 30°С в течение 20 часов. Затем в каждую из колб на 250 мл с угловыми перегородками, содержащих 24 мл культуры, которая содержала компоненты, описанные ниже, инокулировали 1 мл посевной культуральной жидкости и культивировали во встряхиваемом инкубаторе (200 об/мин) при 30°С в течение 72 часов. Концентрацию L-лизина в каждой культуре анализировали путем высокоэффективной жидкостной хроматографии (HPLC).
Посевная культуральная среда (рН 7,0)
Глюкоза (20 г), пептон (10 г), дрожжевой экстракт (5 г), мочевина (1,5 г), KH2PO4 (4 г), K2HPO4 (8 г), MgSO4⋅7H2O (0,5 г), биотин (100 мкг), тиамин HCl (1000 мкг), кальций-пантотеновая кислота (2000 мкг) и никотинамид (2000 мкг) (на основе 1 л дистиллированной воды).
Среда для продуцирования (рН 7,0)
Глюкоза (100 г), (NH4)2SO4 (40 г), соевый белок (2,5 г), твердая фракция кукурузного экстракта (5 г), мочевина (3 г), KH2PO4 (1 г), MgSO4⋅7H2O (0,5 г), биотин (100 мкг), тиамин HCl (1000 мкг), кальций-пантотеновая кислота (2000 мкг), никотинамид (3000 мкг) и CaCO3 (30 г) (на основе 1 л дистиллированной воды).
Из выбранных 231 типа штаммов было выбрано 17 типов штаммов, воспроизводимо демонстрирующих повышенную концентрацию L-лизина по сравнению с контролем, и культивирование и анализ были выполнены в повторах. Результаты анализа концентрации L-лизина показаны в Таблице 2 ниже.
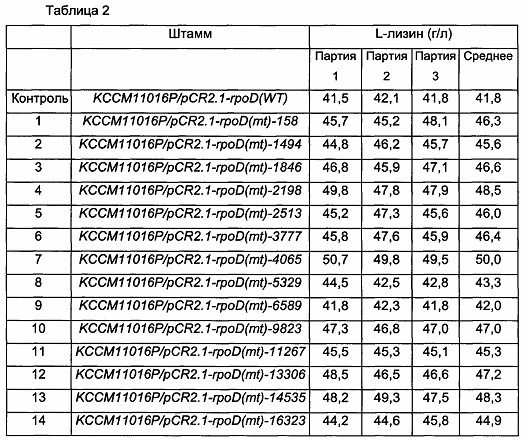

В результате анализа концентрации L-лизина 17 выбранных штаммов было подтверждено, что способность продуцировать L-лизин 15 штаммов, исключая два штамма, то есть KCCM11016P/pCR2.1-rpoD(mt)-6589 и KCCM11016P/pCR2.1-rpoD(mt)-18904, была повышена максимум на 20% по сравнению с таковой в контроле, штамме KCCM11016P/pCR2.1-rpoD(WT).
Пример 5: Подтверждение мутаций в гене rpoD у штаммов, выбранных из библиотеки с искусственным мутагенезом SigA
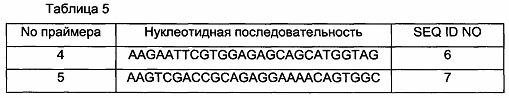
Для подтверждения мутаций, введенных в SigA в 15 штаммах, демонстрирующих улучшенную способность продуцировать L-лизин из 17 выбранных штаммов из Примера 4, были проанализированы нуклеотидные последовательности мутантов rpoD. Для определения нуклеотидных последовательностей проводили ПЦР с использованием праймера 1 (SEQ ID NO: 3) и праймера 3 (SEQ ID NO: 5).

Нуклеотидная последовательность модифицированного гена rpoD была подтверждена на основании Генного банка Национального института здоровья (NIH Genbank) путем анализа нуклеотидных последовательностей фрагментов модифицированного гена rpoD для каждого из полученных таким образом 15 штаммов, и нуклеотидная последовательность модифицированного SigA была подтверждена. Результаты анализа модифицированной аминокислотной последовательности SigA полученных таким образом 15 штаммов показаны в Таблице 4 ниже.

В результате было подтверждено, что были заменены аминокислоты в диапазоне минимум от одной максимум до четырех. В Таблице 4 выше цифры представляют номер аминокислоты SigA, буква напротив цифры представляет аминокислоту до замены, и буква после цифры представляет аминокислоту, на которую произведена замена.
Пример 6: Получение вектора для введения в хромосому модифицированного rpoD для получения штамма, способного продуцировать L-лизин в высокой концентрации
Для подтверждения эффекта применения модификации SigA, которая была подтверждена в Примере 4, был получен вектор, подходящий для введения модификации SigA в хромосому.
Праймер 4 (SEQ ID NO: 6), в который был вставлен сайт рестрикции EcoRI на 5'-конце, и праймер 5 (SEQ ID NO: 7), в который был вставлен сайт рестрикции Sa/I на 3'-конце, синтезировали на основании описанных нуклеотидных последовательностей. Затем проводили ПЦР с использованием каждой из хромосом 15 выбранных типов в качестве матрицы с использованием пары праймеров, и таким образом амплифицировали 15 типов модифицированных фрагментов гена rpoD(mt). ПЦР проводили в следующих условиях: денатурация при 94°С в течение 5 мин; 30 циклов денатурации при 94°С в течение 30 сек, отжига при 56°С в течение 30 сек и полимеризации при 72°С в течение 2 мин; и полимеризация при 72°С в течение 7 мин.
15 типов фрагментов гена, амплифицированных путем ПЦР, обрабатывали EcoRI и Sa/I для получения соответствующих фрагментов ДНК и соединяли фрагменты с вектором pDZ (патент Кореи No. 10-0924065) для введения в хромосому, который включает в себя сайты рестрикции EcoRI и Sa/I, трансформировали им Е.coli DH5a и высевали на твердую среду LB, содержащую канамицин (25 мг/л). Колонии, трансформированные вектором с вставленным методом ПЦР геном-мишенью, отбирали и плазмиды получали путем традиционного способа выделения плазмид. Плазмиды получили названия pDZ-SigA(D136G, T281S), pDZ-SigA(L381R), pDZ-SigA(M230T), pDZ-SigA(M455V), pDZ-SigA(S488T), pDZ-SigA(R279L), pDZ-SigA(I254N), pDZ-SigA(A268S), pDZ-SigA(L451I, Q491R), pDZ-SigA(Q429R), pDZ-SigA(K90E, K105Y, D250G, I254L), pDZ-SigA(L447H), pDZ-SigA(K483R), pDZ-SigA(K479R) и pDZ-SigA(D238V, N263S, E358D), соответственно, в соответствии с модификацией вставки в SigA каждой плазмиды. Дополнительно в качестве другого контроля для вышеизложенных вариантов праймеры 6-9 (SEQ ID NO: 8-11) получали для конструирования векторов для вставки SigA, модифицированного A414V, о котором сообщалось, что он обладает действием по повышению способности продуцировать L-лизин (Международная публикация WO 2003-054179), в хромосому. Затем проводили ПЦР с использованием геномной ДНК Corynebacterium glutamicum АТСС13032 наряду с парой праймера 6 и праймера 9 и парой праймеров из праймера 7 и праймера 8, соответственно. ПЦР проводили в следующих условиях: денатурация при 94°С в течение 5 мин; 30 циклов денатурации при 94°С в течение 30 сек, отжига при 56°С в течение 30 сек и полимеризации при 72°С в течение 1 мин; и полимеризация при 72°С в течение 7 мин.

В результате были получены фрагменты гена размером примерно 500 п.н. и продукты амплификации были связаны с вектором pDZ с помощью набора для клонирования Infusion (Invitrogen). Полученный таким образом вектор получил название pDZ-SigA(A414V).
Пример 7: Получение штамма KCCM11016P с введенным модифицированным полипептидом SigA для штаммов, способных продуцировать L-лизин в высокой концентрации, и сравнение их способностей продуцировать L-лизин
15 типами векторов с введенными новыми мутациями, полученными в Примере 6, и одним типом вектора с введенной мутацией, о которой сообщалось ранее, трансфицировали Corynebacterium glutamicum KCCM11016P, который представляет собой штамм, продуцирующий L-лизин, путем двухэтапной рекомбинации гомологичных хромосом. Затем штаммы с введенными модификациями SigA отбирали путем анализа нуклеотидных последовательностей. Штаммы с введенными модификациями SigA получили названия KCCM11016P::SigA(D136G, T281S), KCCM11016P::SigA(L381R), KCCM11016P::SigA(M230T), KCCM11016P::SigA(M455V), KCCM11016P::SigA(S488T), KCCM11016P::SigA(R279L), KCCM11016P::SigA(I254N), KCCM11016P::SigA(A268S), KCCM11016P::SigA(L451I, Q491R), KCCM11016P::SigA(Q429R), KCCM11016P::SigA(K90E, K105Y, D250G, I254L), KCCM11016P::SigA(L447H), KCCM11016P::SigA(K483R), KCCM11016P::SigA(K479R), KCCM11016P::SigA(D238V, N263S, E358D) и KCCM11016P::SigA(A414V), соответственно.
Штаммы культивировали таким же способом, как в Примере 4, и анализировали их концентрации L-лизина. Результаты показаны в Таблице 7 ниже.

В результате два новых штамма с введенными модификациями (KCCM11016P::SigA(M230T) и KCCM11016P::SigA(R279L)) показали способность продуцировать L-лизин, эквивалентную таковой у родительского штамма, в то время как два других новых штамма с введенными мутациями (KCCM11016P::SigA(K90E, K105Y, D250G, I254L) и KCCM11016P::SigA(D238V, N263S, E358D)) продемонстрировали значительно более низкую скорость роста и значительно сниженный уровень продукции L-лизина. Однако остальные 11 новых мутантных штаммов продемонстрировали максимальное увеличение продукции L-лизина на 19% по сравнению с таковым родительского штамма, и также продемонстрировали повышение продукции L-лизина на 16% по сравнению с таковой KCCM11016P::SigA(A414V), который представляет собой мутантный штамм с введенной мутацией SigA(A414V), о котором сообщалось ранее. Авторы настоящего изобретения обозначили штамм с улучшенной способностью продуцировать L-лизин как таковой "KCCM11016P::SigA(L447H)" "Corynebacterium glutamicum СА01-2277" и депонировали его в Корейском центре культур микроорганизмов, признанном как международный орган депонирования в соответствии с Будапештским соглашением, 22 ноября 2013 года с номером доступа KCCM11479P.
Эти результаты позволяют предположить, что 11 новых модифицированных полипептидов SigA обладают превосходной способностью продуцировать L-лизин.
Пример 8: Получение штамма KFCC10750 с введенным модифицированным полипептидом SigA для штаммов, способных продуцировать L-лизин в высоких концентрациях, и сравнение их способностей продуцировать L-лизин
Для подтверждения эффекта введения 11 типов вариантов SigA, выбранных в Примере 7, в другие штаммы рода Corynebacterium glutamicum, штаммы с введенной, соответственно, каждой из 11 типов мутаций SigA Corynebacterium glutamicum KFCC10750 (KCCM11347P, патент Кореи No. 10-0073610), обладающие способностью продуцировать L-лизин, были получены таким же способом как в Примере 7. Эти штаммы получили названия KFCC10750::SigA(D136G, T281S), KFCC10750::SigA(L381R), KFCC10750::SigA(M455V), KFCC10750::SigA(S488T), KFCC10750::SigA(I254N), KFCC10750::SigA(A268S), KFCC10750::SigA(L451I, Q491R), KFCC10750::SigA(Q429R), KFCC10750::SigA(L447H), KFCC10750::SigA(K483R) и KFCC10750::SigA(K479R), соответственно. Дополнительно, штамм с введенной мутацией SigA(A414V), о котором сообщалось ранее, был получен и получил название "KFCC10750::SigA(A414V)".
Штаммы культивировали таким же образом как в Примере 4 и анализировали их концентрации L-лизина. Результаты показаны в Таблице 8 ниже.

В результате 11 типов штаммов с введенными новыми мутациями демонстрировали максимальное увеличение продукции L-лизина на 17% по сравнению с таковой родительского штамма и также демонстрировали увеличение продукции L-лизина примерно на 14% по сравнению с таковой KFCC10750::SigA(A414V), который представляет собой штамм с введенной мутацией SigAA414V, о котором сообщалось ранее.
Пример 9: Получение штамма KCCM10770P с введенным модифицированным полипептидом SigA для штаммов, способных продуцировать L-лизин в высокой концентрации, и сравнение их способностей продуцировать L-лизин
Для подтверждения эффекта введения 11 типов вариантов SigA, выбранных в Примере 7, в другие штаммы рода Corynebacterium glutamicum, штаммы с введенными, соответственно, модификациями SigA Corynebacterium glutamicum KFCC10770 (KCCM11347P, патент Кореи No. 10-0924065), обладающие способностью продуцировать L-лизин, были получены таким же способом как в Примере 7. Эти штаммы получили названия KCCM10770P::SigA(D136G, T281S), KCCM10770P::SigA(L381R), KCCM10770P::SigA(M455V), KCCM10770P::SigA(S488T), KCCM10770P::SigA(I254N), KCCM10770P::SigA(A268S), KCCM10770P::SigA(L451I, Q491R), KCCM10770P::SigA(Q429R), KCCM10770P::SigA(L447H) и KCCM10770P::SigA(K483R), KCCM10770P::SigA(K479R), соответственно. Дополнительно штамм с введенной мутацией SigA(A414V), о котором сообщалось ранее, был получен и получил название "KFCC10770::SigA(A414V)".
Эти штаммы культивировали таким же образом, как в Примере 4, и анализировали их концентрации L-лизина. Результаты показаны в Таблице 9 ниже.


В результате 11 типов штаммов с введенными новыми мутациями демонстрировали максимальное увеличение продукции L-лизина на 20% по сравнению с таковой родительского штамма, и также демонстрировали увеличение продукции L-лизина примерно на 16% по сравнению с таковой KFCC10770::SigA(A414V), который представляет собой штамм с введенной мутацией SigAA414V, о котором сообщалось ранее.
Пример 10: Получение штамма CJ3P с введенным модифицированным полипептидом SigA для штаммов, способных продуцировать L-лизин в высокой концентрации, и сравнение их способностей продуцировать L-лизин
Для подтверждения эффекта на других штаммах рода Corynebacterium glutamicum, штаммы, в которые были введены модификации SigA, были соответственно введены в Corynebacterium glutamicum CJ3P (Binder et al. Genome Biology 2012, 13: R40), обладающие способностью продуцировать L-лизин, были получены таким же образом как в Примере 7. Эти штаммы получили названия CJ3P::SigA(D136G, T281S), CJ3P::SigA(L381R), CJ3P::SigA(M455V), CJ3P::SigA(S488T), CJ3P::SigA(I254N), CJ3P::SigA(A268S), CJ3P::SigA(L451I, Q491R), CJ3P::SigA(Q429R), CJ3P::SigA(L447H), CJ3P::SigA(K483R) и CJ3P::SigA(K479R), соответственно. Дополнительно, штамм с введенной мутацией SigA(A414V), о котором сообщалось ранее, был получен и получил название "CJ3P::SigA(A414V)". CJ3P представляет собой штамм Corynebacterium glutamicum, который обладает способностью продуцировать L-лизин благодаря введению 3 типов мутаций в микроорганизм дикого типа на основе известной технологии.
Штаммы культивировали таким же образом как в Примере 4 и анализировали их концентрации L-лизина. Результаты показаны в Таблице 10 ниже.

В результате 11 типов штаммов с введенными новыми мутациями демонстрировали максимальное увеличение продукции L-лизина на 20% по сравнению в таковой родительского штамма, и также демонстрировали увеличение продукции L-лизина примерно на 15% по сравнению с таковой CJ3P::SigA(A414V), который представляет собой штамм с введенной мутацией SigAA414V, о котором сообщалось ранее.
Суммируя вышеизложенные результаты, было показано, что 11 новых модифицированных полипептидов, обладающих активностью SigA, полученных в настоящем изобретении (то есть, SigA(D136G, T281S), SigA(L381R), SigA(M455V), SigA(S488T), SigA(I254N), SigA(A268S), SigA(L451I, Q491R), SigA(Q429R), SigA(L447H), SigA(K483R) и SigA(K479R)), обладают превосходным действием по улучшению продукции L-лизина, соответственно, у различных микроорганизмов рода Corynebacterium, и эти результаты позволяют предположить, что модифицированные полипептиды обладают превосходным действием по улучшению продукции L-лизина по сравнению с модифицированным SigA(A414V), о котором сообщалось ранее.
На основании вышеизложенного специалист в области техники, к которой принадлежит данное изобретение, понимает, что настоящее изобретение может быть воплощено в других конкретных формах без изменения технической идеи или существенных признаков настоящего изобретения. В этом отношении примеры воплощений, раскрытые здесь, служат исключительно для иллюстративных целей и не предназначены ограничивать объем настоящего изобретения. Наоборот, настоящее изобретение предназначено охватывать не только воплощения, представленные в примерах, но также различные альтернативные, модифицированные, эквивалентные и другие воплощения, которые могут быть включены в пределах сущности и объема настоящего изобретения, как определено в прилагаемой формуле изобретения.
Реферат
Группа изобретений относится к области биотехнологии. Предложен модифицированный полипептид, обладающий активностью сигма-фактора А РНК-полимеразы, где по меньшей мере одна аминокислота, выбранная из группы, состоящей из аминокислот в следующих положениях полипептида, имеющего аминокислотную последовательность, изложенную в SEQ ID NO: 2, заменена, и аминокислотная замена представляет собой по меньшей мере одну из следующих аминокислотных замен: 136G, 254N, 268S, 281S, 381R, 429R, 447H, 451I, 455V, 479R, 483R, 488T и 491R. Предложен полинуклеотид, кодирующий указанный модифицированный полипептид. Предложен микроорганизм, обладающий повышенной способностью продуцировать L-лизин, содержащий указанный модифицированный полипептид. Предложен способ получения L-лизина с использованием указанного микроорганизма. Группа изобретений позволяет повысить продукцию L-лизина. 4 н. и 3 з.п. ф-лы, 10 табл., 10 пр.
Комментарии