Постоянные линии клеток человека для получения вирусов гриппа - RU2565546C2
Код документа: RU2565546C2
Чертежи












Описание
Заявленное изобретение относится к способу получения вакцины на основе вируса гриппа с использованием постоянных клеток-амниоцитов человека, а также к применению постоянной клетки-амниоцита человека для получения вакцины на основе вируса гриппа.
Вакцинация в здравоохранении является самой важной мерой предотвращения заболевания, вызываемого ежегодной эпидемией гриппа. Успешное применение вакцин зависит от быстрейшего производства достаточно больших объемов вакцин, например, инактивированных вирусов, из стабильных и простых в применении исходных материалов. Ускоренная разработка вакцин и их адекватная доступность крайне важны в эффективной борьбе со многими болезными человека и животных. Как результат задержек в производстве вакцин и количественных потерь, могут возникать проблемы с контролем над вспышками заболеваний. Данное обстоятельство послужило в последнее время причиной целенаправленности исследований на культивирование вирусов в клеточных культурах для использования в качестве вакцин.
В настоящее время, доступные вакцины против гриппа производят из эмбрионов куриных яиц. Данные куриные яйца должны пройти контроль на отсутствие определенных вирусных и бактериальных загрязнений. Эти так называемые «свободные от специфической патогенной флоры» (SPF) куриные яйца доступны в коммерческих объемах. Даже несмотря на то, что куриные яйца весьма успешно применяют для размножения вирусов животных и человека, они обладают рядом недостатков в производстве вакцин. Например, в случае пандемии, резко увеличивается спрос на куриные яйца производстве вакцин, учитывая то, что для получения одной дозы обычной вакцины требуется одно яйцо. Исходя из того, что объем куриных яиц ограничен, требуется примерно год для накопления необходимого количества куриных яиц. Помимо этого, существуют подтипы гриппа, крайне патогенные для куриц, которые могут послужить причиной недостатка куриных яиц в случае пандемии. Дополнительно, сам процесс производства высоко затратный и долговременный. Еще одним недостатком производства вакцин в куриных яйцах является невозможность полного исключения куриного белка из вакцин, и, соответственно, проявления аллергической реакции у некоторых пациентов. И, наконец, возможная селекция субпопуляции, отличающейся от встречающегося в природе вируса, требует применения систем альтернативных клеток-хозяев.
В отличие от куриных яиц, клетки для производства вакцин против гриппа на основе клеточных культур всегда доступны. Они хранятся в глубокой заморозке и могут быть быстро разморожены и репродуцированы в требуемых объемах в любой момент времени при необходимости. Таким образом, производство вакцин можно запустить в любой требуемый момент времени. В случае резко возросшего спроса, или более частой циркуляции новых непрогнозируемых штаммов вирусов, необходимую вакцину могут изготовить в кратчайшее время.
Процесс производства с использованием клеточной культуры позволяет осуществлять получение вирусов в форме вакцин в закрытой, стандартизированной системе при определенных, контролируемых параметрах. Благодаря контролируемости процесса производства, окончательная вакцина против гриппа не требует добавления антибиотиков. Поскольку получение вакцин против гриппа на основе клеточных культур полностью независимо от наличия куриных яиц, вакцина не содержит яичного белка и, таким образом, не вызывает аллергических реакций у пациентов с непереносимостью белка куриных яиц.
В настоящее время, для производства вакцин против гриппа используют преимущественно три линии клеток, а именно, клетки PER.C6 человека, клетки почек собак производства Madin Darby Canine Kidney (MDCK), и клетки почек мартышек (Vero). Дополнительно, в настоящее время в разработке находятся линии клеток сетчатки уток (AGE1.CR) и линии птичьих эмбриональных стволовых клеток. Производство вакцин в клетках млекопитающих является альтернативой производству вакцин на основе куриных яиц, однако данные клетки требуют наличия сыворотки и/или прикрепления для надежной поддержки их роста. Данное обстоятельство затрудняет и повышает стоимость производства вакцин в данных клетках, поскольку, из соображений мер предосторожности, сыворотку необходимо полностью отделить, а рост на твердых прикреплениях ограничен, что приводит к более низкой производительности.
Преимуществом производства вакцин в клетках млекопитающих является тот факт, что изоляцию и репликацию вируса в клеточной культуре не генерируют какой-либо пассажир-зависимой селекции фенотипа, отличающегося от клинического дикого типа. Таким образом, вирусный гликопротеин гемагглютинин, посредством которого осуществляют инфицирование прикрепления к клетке и интеграцию вируса в клетку, экспрессируют аналогично природной форме, и он обладает улучшенной специфичностью и авидностью, соответственно, способствует появлению клеточно-опосредованного иммунитета у людей.
Таким образом, целью заявленного изобретения является предоставление постоянных линий клеток человека с улучшенными характеристиками для производства вакцин на основе вирусов гриппа.
Цель достигается практическим воплощением сути заявленного изобретения, определенной в формуле изобретения.
Фиг. иллюстрируют заявленное изобретение.
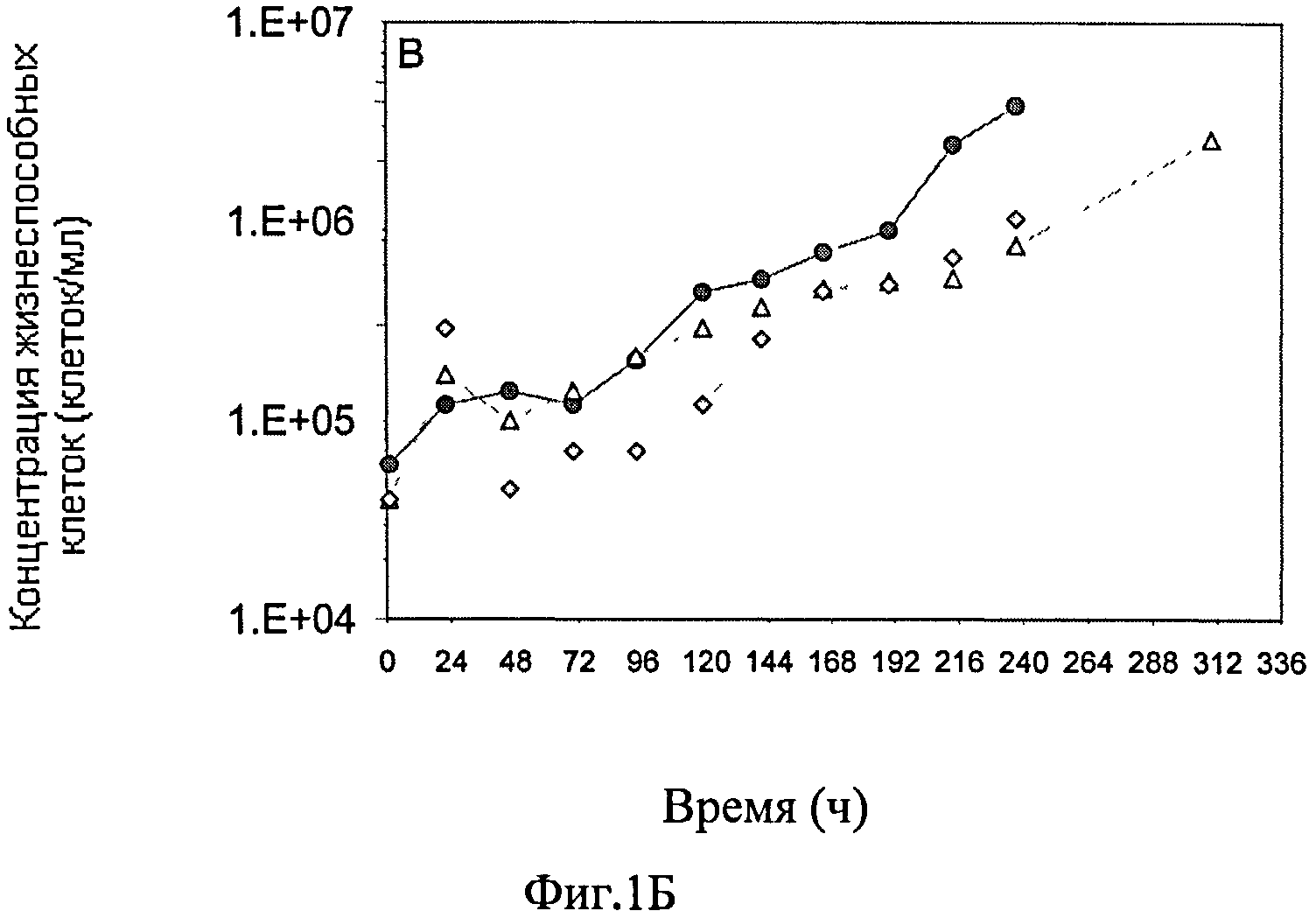
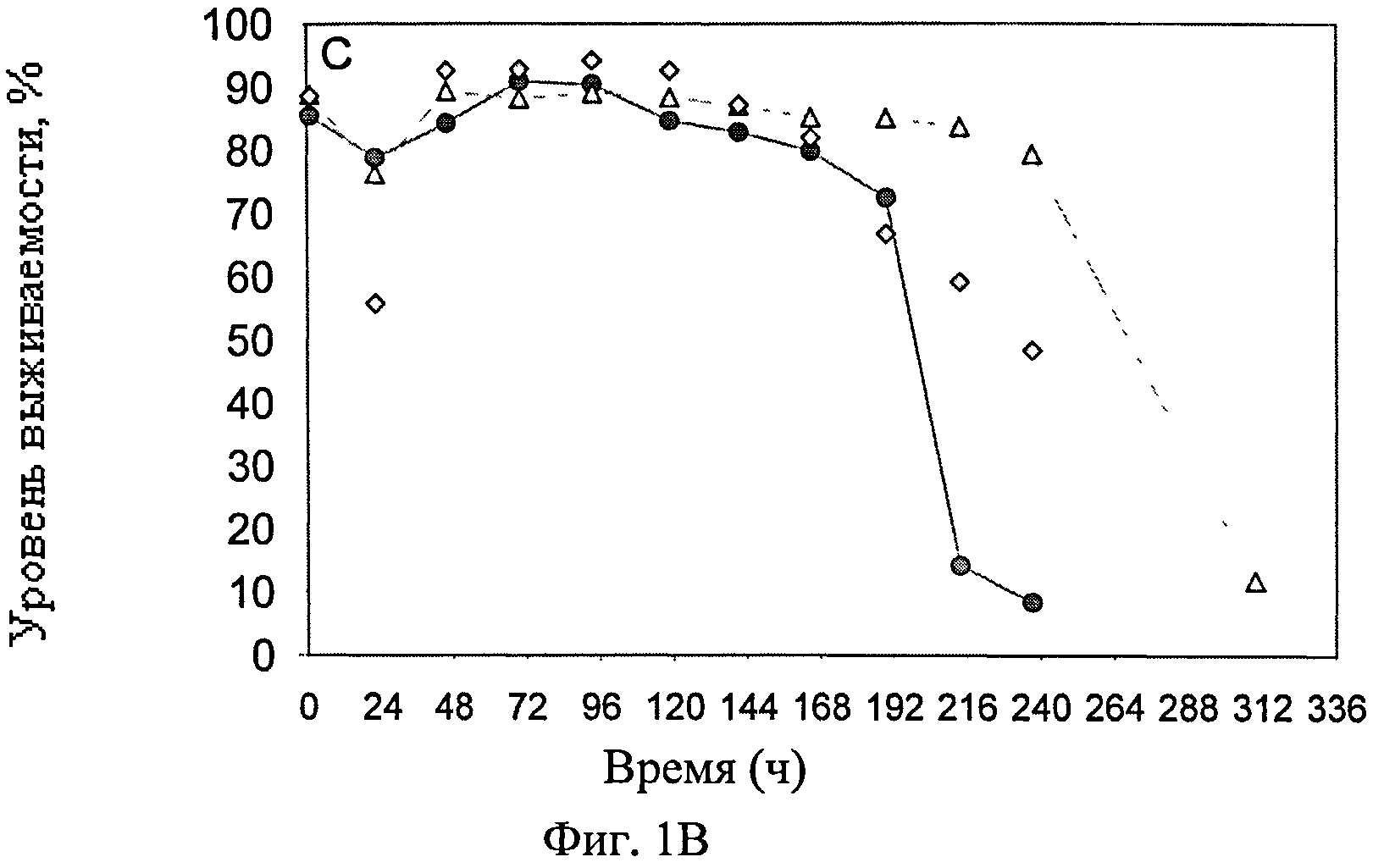
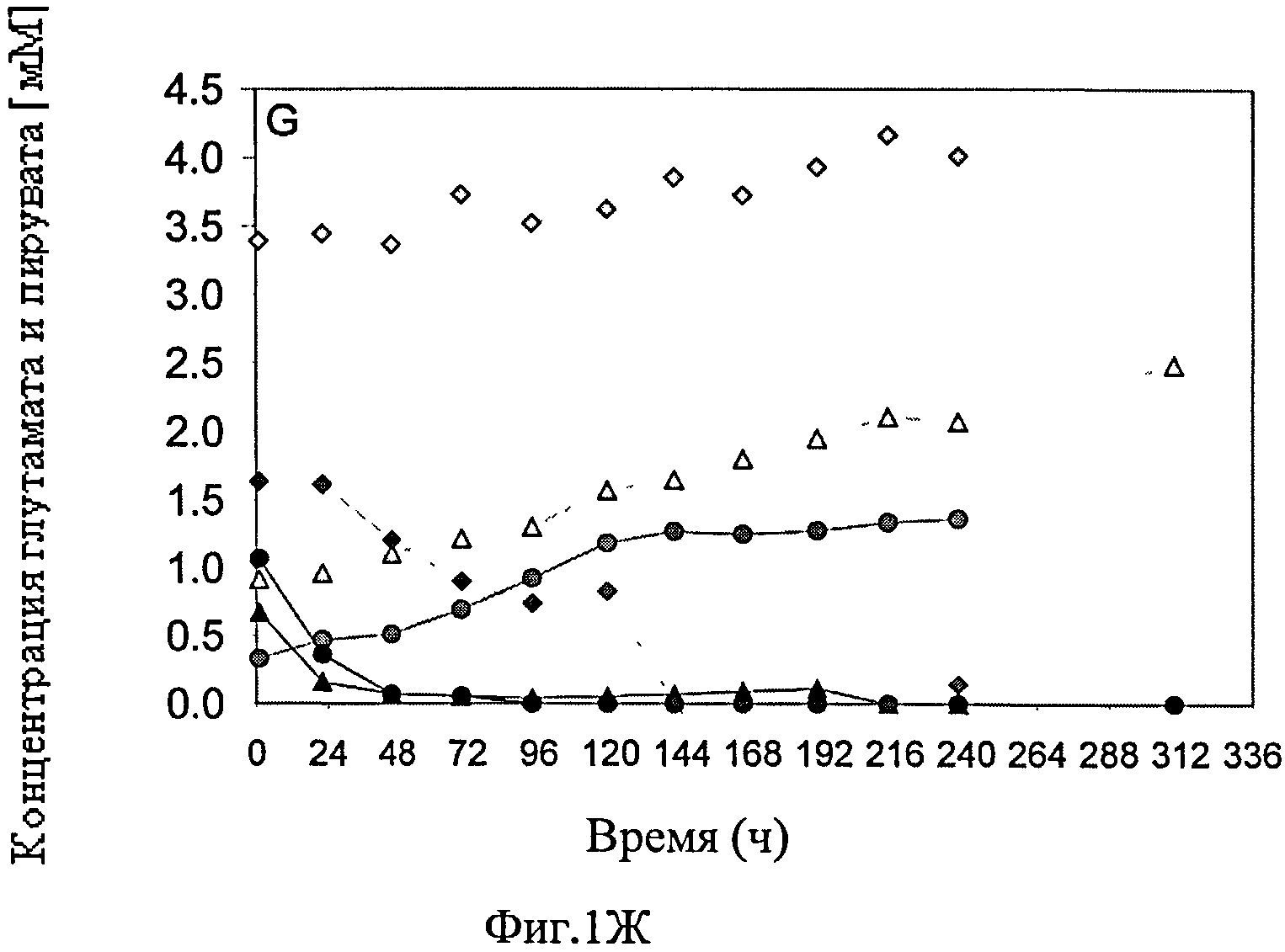
Фиг.1А-Ж схематически представляют динамику изменения различных параметров во время культивирования постоянной линии клеток-амниоцитов CAP 1D5 в 293SFMII среде (●), постоянной линии клеток-амниоцитов CAP 1D5 в PEM среде (▲) и постоянной линии клеток почек собак MDCK.SUS2 (Madin Darby Canine Kidney) в SMIF8 среде (♦) во встряхиваемых колбах емкостью 100 мл. Фиг.1А графически представляет динамику изменения концентрации жизнеспособных клеток трех линий клеток, в сравнении; фиг.1Б представляет динамику изменения концентрации мертвых клеток линий клеток; а фиг.1В представляет динамику колебания уровня выживаемости линий клеток. Фиг.1Г-Ж схематически представляют динамику изменения значения pH (Г), концентрации (Д) глюкозы (светлые значки) и лактозы (темные значки), темные значки (Е) глутамина (Gln) (светлые значки) и аммония (темные значки), и концентрации (Ж) глутаминовой кислоты (Glu) (светлые значки) и пирувата (темные значки).
Фиг.2 представляет гистограмму, изображающую замеры титров вирусов как значение TCID50 в 4 проходах штаммов гриппа A/PR78/34 (H1N1) и A/Uruguay/716/2007 (H3N2) в CAP-1D5 клетках 293SFMII и PEM среды. Сокращения: A/PR 293: штамм гриппа A/PR/8/34 (H1N1) в CAP-1D5 клетках в 293SFMII среде, A/PR PEM: штамм гриппа A/PR/8/34 (H1N1) в CAP-1D5 клетках в PEM среде; A/Urug 293: A/Uruguay/716/2007 штамм гриппа (H3N2) в CAP-1D5 клетках в 293SFMII среде; A/Urug PEM: штамм гриппа A/Uruguay/716/2007 (H3N2) в CAP-1D5 клетках в PEM среде; значение TCID50 - вирусный титр в числе вирусов/мл, которое требуется для инфицирования 50% клеток-хозяев.
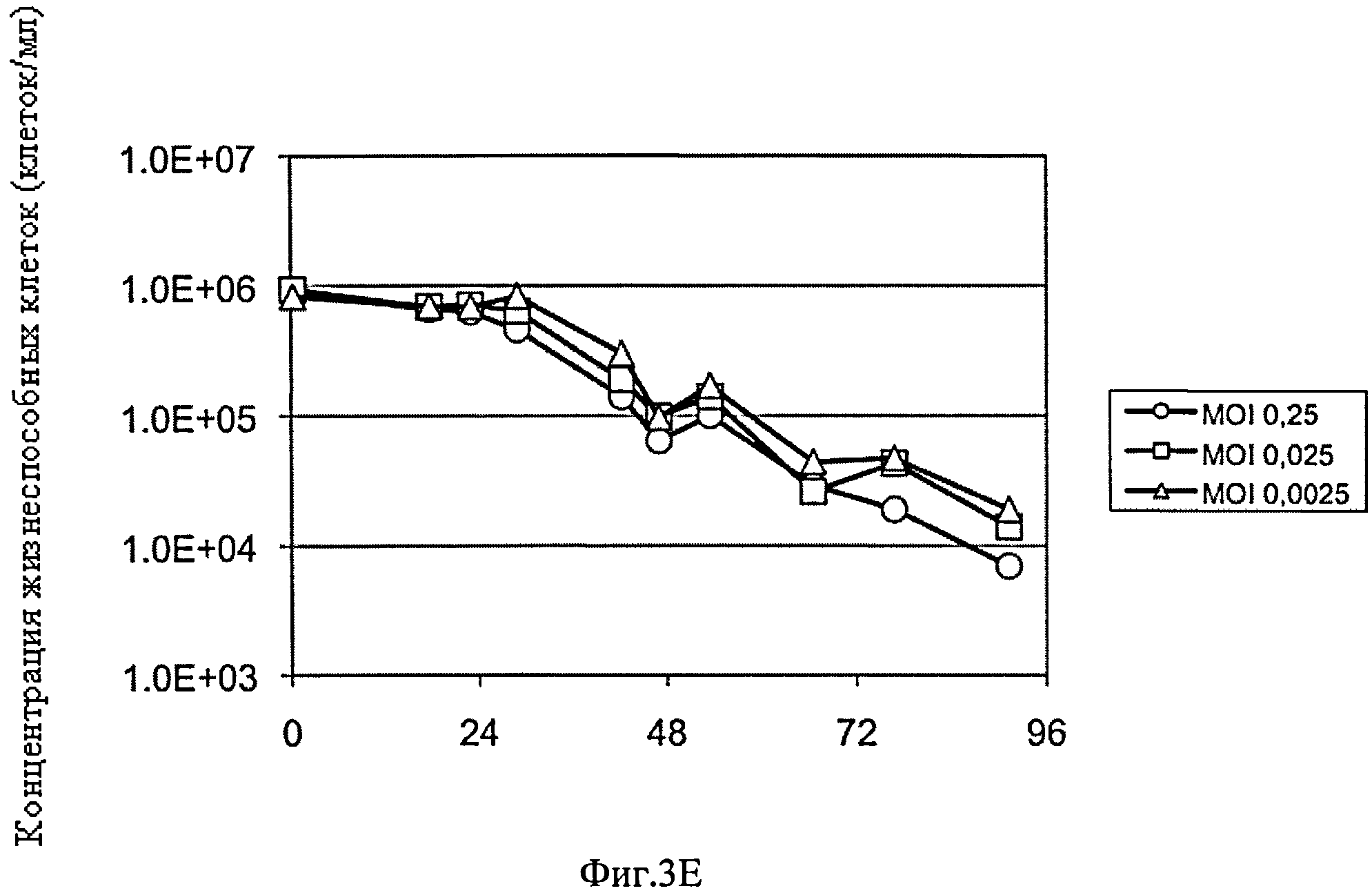
Фиг.3А-Е схематически представляют динамику изменения количества вирусных частиц, обозначенных как log HA (гемагглютинин) единицы/100 мкл и концентрации жизнеспособных клеток в культуре постоянных клеток-амниоцитов CAP-1D5 в 293SFMII-(A, Б) и в PEM среде (В, Г), постоянных клеток почек собак MDCK.SUS2 в SMIF8 среде (Д, Е) после инфицирования клеток штаммом вируса гриппа A/PR/8/34 при использовании различных объемов вируса, обозначенных как величины MOI (множественность заражения): MOI: 0.0025 (Δ) MOI: 0.025 (□), MOI: 0.25 (○). MOI (множественность заражения) представляет соотношение числа инфицированных частиц к клеткам-мишеням.
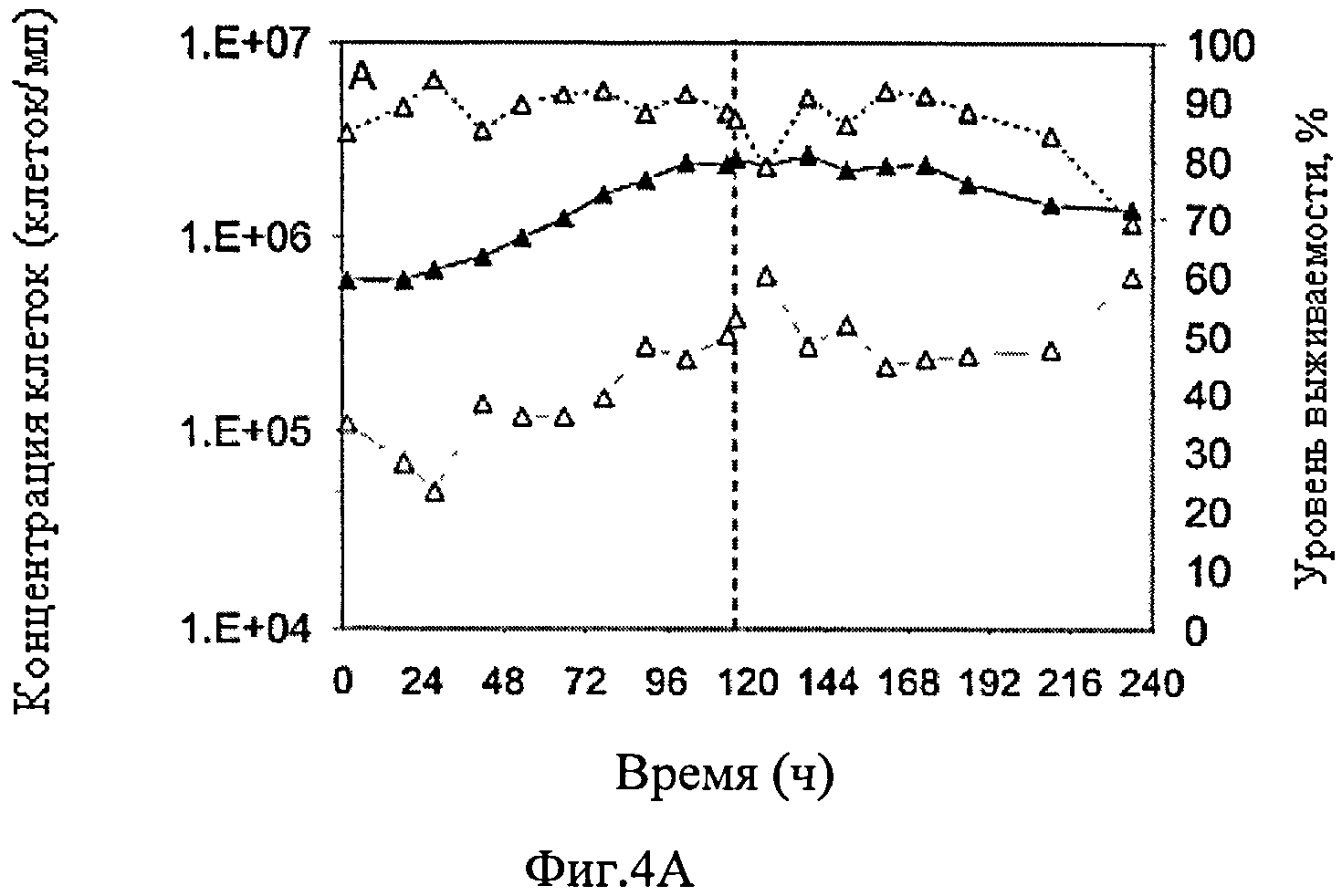
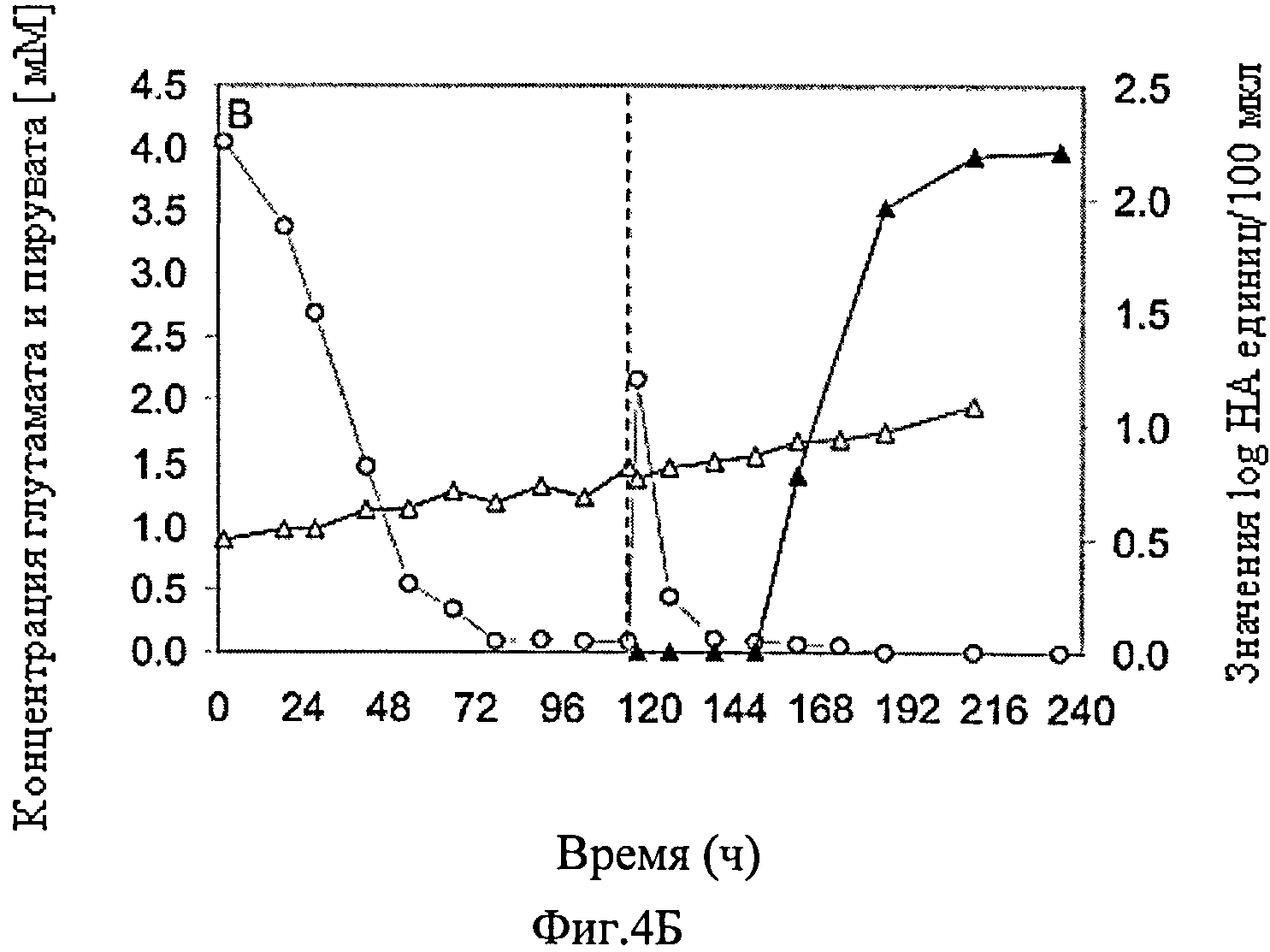
Фиг.4А-Г схематически представляют динамику изменения различных параметров культивирования постоянной линии клеток-амниоцитов CAP-1D5 в PEM среде в биореакторе объемом 1 л, в котором инфекция происходит после 114 ч с различным объемом вируса, обозначенным как MOI от 0,025 с вирусом гриппа A/PR/8/34 (адаптированный). Фиг.4А схематически представляет динамику изменения концентрации жизнеспособных клеток (▲), концентрации мертвых клеток (Δ) и уровня выживаемости клеток (


Фиг.5А и Б показывают гистограммы, представляющие титры вирусов, измеренные как log HA единиц/100 мкл (А) или значение TCID50 (Б) вдоль 4 проходов штаммов гриппа A/Brisbane/59/2007, B/Florida/4/2006, свиного гриппа (A/Swine (H1N2) Bakum/1832/00) и лошадиного гриппа (A/Equine, A/Newmarket/1/93 (H3N8)) на клетках CAP-1D5 в 293SFMII и PEM среде. Сокращения: A/Bris 293: штамм гриппа A/Brisbane/59/2007 на CAP-1D5 клетках в 293SFMII среде; A/Bris PEM: штамм гриппа A/Brisbane/59/2007 на CAP-1D5 клетках в PEM среде; B/Flor 293: штамм гриппа B/Florida/4/2006 на CAP-1D5 клетках в 293SFMII среде; B/Flor PEM: штамм гриппа B/Florida/4/2006 на CAP-1D5 клетках в PEM среде; Schw 293: штамм гриппа A/Swine (H1N2) Bakum/1832/00 на CAP-1D5 клетках в 293SFMII среде; Schw PEM: штамм гриппа A/Swine (H1N2) Bakum/1832 00 на CAP-1D5 клетках в PEM среде; horse 293: штамм гриппа A/Equine, A/Newmarket/1/93 (H3N8) на CAP-1D5 клетках в 293SFMII среде; horse PEM: штамм гриппа A/Equine, A/Newmarket/1/93 (H3N8) на CAP-1D5 клетках в PEM среде; значение TCID50 показывает титр вируса как число вирусов/мл, которое требуется для инфицирования 50% клеток-хозяев.
Фиг.6А и Б схематически представляют динамику изменения концентрации жизнеспособных клеток и значения pH в культивировании постоянной линий клеток-амниоцитов CAP-1D5 в 100 мл PEM среды во встряхиваемых колбах, причем первоначальная концентрация клеток составляет 5×105 клеток/мл, а среда дополнительно содержит 4 мМ пирувата (♦), или первоначальная концентрация клеток составляет 8×105 клеток/мл, а среда дополнительно содержит 4 мМ пирувата (▲), или первоначальная концентрация клеток составляет 8×105 клеток/мл, а среда дополнительно содержит 10 мМ пирувата плюс дополнительные аминокислоты (●).
Фиг 7А-В схематически представляют динамику изменения титров вирусов, измеренных в log HA единиц/100 мкл культуры, причем CAP-1D5 клетки инфицировали адаптированным штаммом гриппа A/PR/8/34. До инфицирования, не проводили никакой смены среды (А), либо осуществили разбавление 1:2 с PEM средой (Б), либо полную смену среды. Фиг.7А схематически представляет динамику изменения титра вируса CAP-1D5 клеточных культур без смены среды, причем для инфицирования использовали различные концентрации трипсина 1×10-4 Е/клетку (♦), 3×10-5 E/клетку (▲) и 5×10-5 E/клетку (■). Фиг.7Б схематически представляет динамику изменения титра вируса CAP-1D5 клеточных культур с разбавлением 1:2 с PEM средой, причем для инфицирования использовали различные концентрации трипсина 1×10-4 Е/клетку (♦), 3×10-5 E/клетку (▲), и 5×10-5 E/клетку (■). Фиг.7 В схематически представляет динамику изменения титра вируса CAP-1D5 клеточных культур с полной сменой среды, причем для инфицирования не использовали трипсин () или использовали различные концентрации трипсина 1×10-4 Е/клетку (▲), 1×10-5 Е/клетку (♦), 5×10-5 Е/клетку (■) и 1×10-6 E/клетку (x).
Фиг.8A-E схематически представляют динамику изменения титра вирусов в CAP-1D5 клеточных культурах, которые инфицировали вирусами гриппа A/PR/8/34, A/Brisbane/59/2007 or B/Florida/4/200, причем до инфицирования либо осуществили смену среды (А-В), либо не проводили смену среды (Г-Е). Инфицирование вирусом гриппа A/PR/8/34 и B/Florida/4/2006 осуществили соответственно с объемами вируса, обозначенными как MOI 0.25, 0.025 и 0.0025. Инфицирование вирусом гриппа A/Brisbane/59/2007 происходило при значениях MOI 0.1, 0.025 и 0.0025. MOI (множественность заражения) представляет соотношение числа инфицированных частиц к клеткам-мишеням.
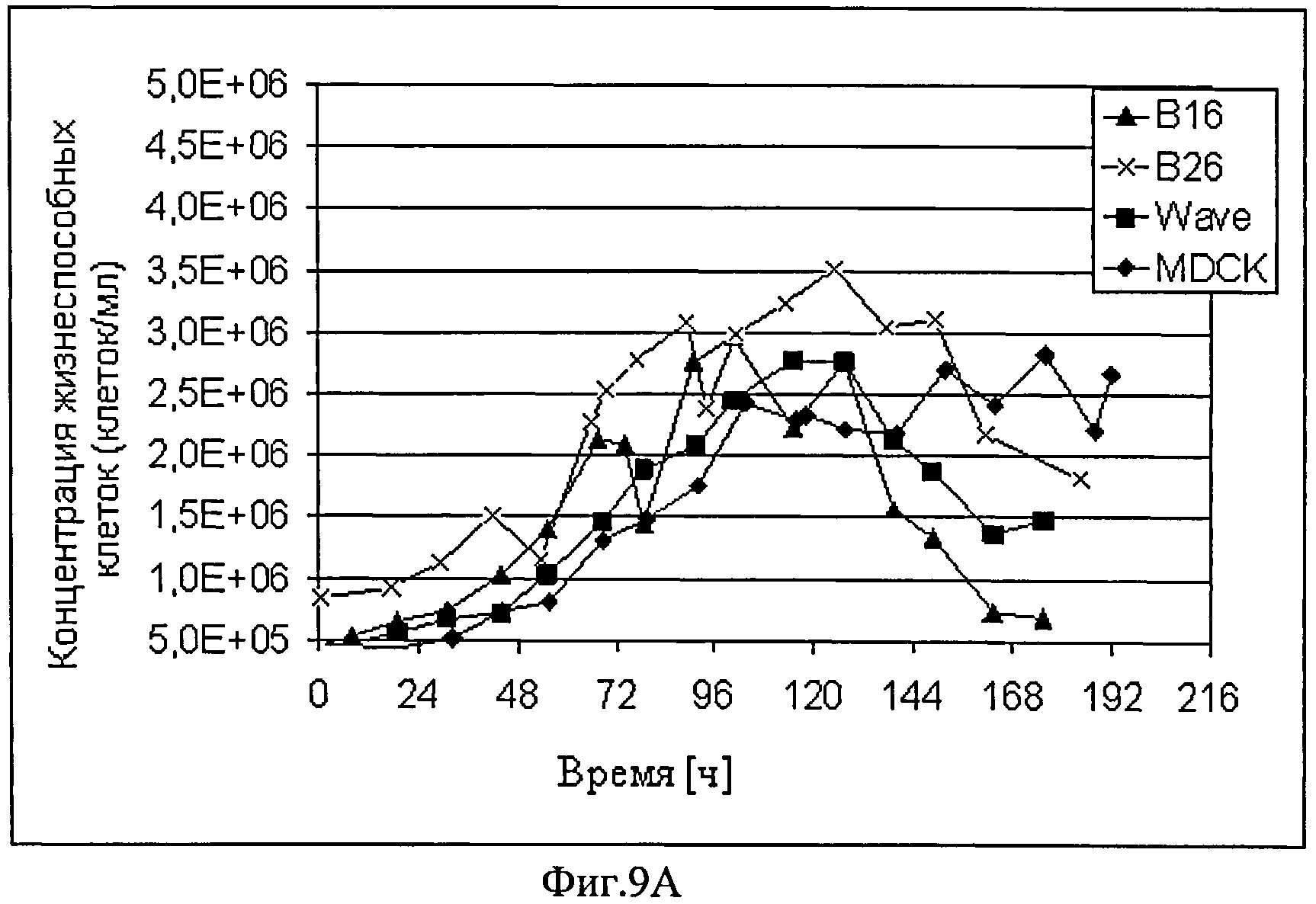
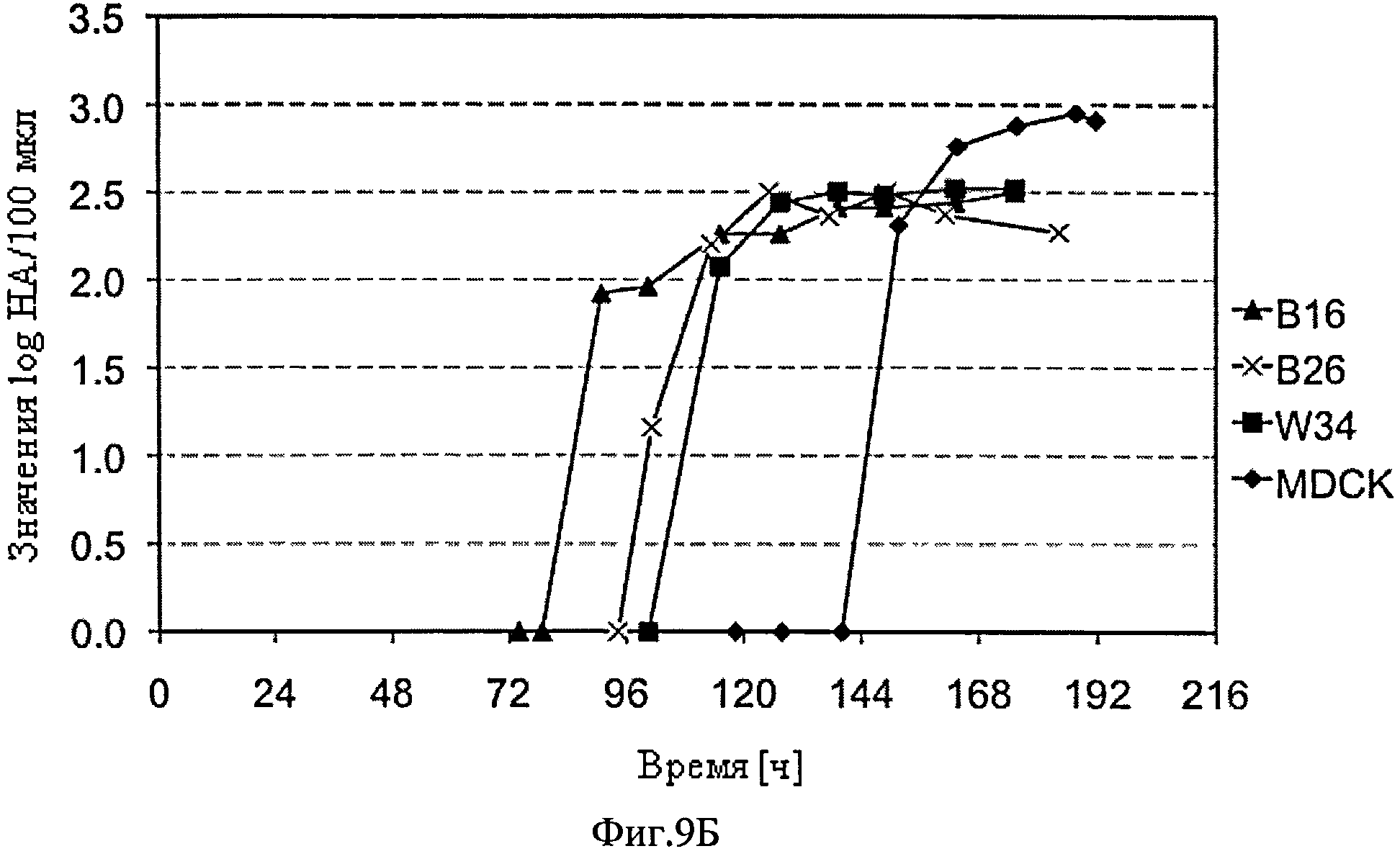
Фиг.9А и Б схематически представляют динамику изменения концентрации жизнеспособных клеток и титра вируса CAP-1D5 клеточных культур (B16, B26, и Wave) и почек собак-MDCK. SUS2 культура (MDCK), которую инфицировали с адаптированным A/PR/8/34 вирусом гриппа и культивировали в объеме 1 л в STR (Sartorius) (В16, В26, и MDCK) или Wave Bioreactors (Wave Biotech AG) (Wave). До инфицирования осуществили смену среды, в случае использования В26 культур и Wave.
Фиг.10А-В схематически представляют динамику изменения титра вируса, измеренного в log HA единиц/100 мкл, концентрации жизнеспособных клеток и значения pH CAP-1D5 клеточных культур, которые были инфицированы адаптированным вирусом гриппа A/PR/8/34 и культивированы в PEM 100 мл среды во встряхиваемых колбах. До инфицирования, осуществили либо 1:1 смену среды (светлые значки) со 293SFMII средой (□) или PEM средой (◊), либо полную смену среды (темные значки) со 293SFMII средой (■) или PEM средой (♦).
Термин «вирус гриппа», в том значении, в котором его используют в описании заявленного изобретения, относится к членам ортомиксовирусов, которые могут инфицировать человека и животных. Их классифицируют на вирус гриппа типов A, B и C. Вирусы гриппа A и B объединены в род (таксон). Вирусы гриппа C различают благодаря их семи геномным сегментам. Вирусы гриппа A и B имеют восемь геномных сегментов. Дополнительно, вирусы гриппа A и B каждый кодирует гемагглютинин (HA) и нейроминидазу (NA); в отличие от них, вирус гриппа С кодирует поверхностный белок, который сочетает два свойства гемагглютинин-эстераза-слитый белок (HEF). Вирусы гриппа A дополнительно делят на подтипы, на основе последовательности молекул гемагглютинина (H1-H15) и нейраминидазы (N1-N9).
Термин «белок вируса гриппа», в том значении, в котором его используют в описании заявленного изобретения, относится к белкам или производным вируса гриппа. Производное вируса гриппа обычно представляет собой белок или его часть вируса гриппа, который могут использовать в целях иммунизации. Белки вируса гриппа или их производные образуют белки вирусной оболочки или ее частей. В частности, белки вируса гриппа образуют белки гриппа A, белки гриппа В или белки гриппа C, например, гемагглютинин (HA), нейраминидазу (NA), нуклеопротеин (NP), матричные белки (M1) и (M2), белки полимеразы (PB1), (PB2) и (PA) и неструктурные белки (NS1) и (NS2) и их части. Части (фрагменты) белков вирусов гриппа содержат один или более эпитопов белков гриппа A, белков гриппа B или белков гриппа C. Эпитопы могут являться CD4+T-клеточными эпитопами, которые представляют пептиды, содержащие связывающий мотив класса ГКГ класса II и репрезентированы на поверхности антигенпрезентирующих клеток, при помощи молекул ГКГ класса II, или CD8+T-клеточными эпитопами, которые представляют пептиды, содержащие связывающий мотив класса ГКГ класса I и репрезентированы на поверхности антигенпрезентирующих клеток, при помощи молекул ГКГ класса I. Например, алгоритмическая модель, анализы связывания ГКГ, методы идентификации антигена in silico, и методы рентгенографической кристаллографии позволяют осуществлять идентификацию антигенов, способных связывать различные молекулы ГКГ.
Термин «вакцина», в том значении, в котором его используют в описании заявленного изобретения, относится к биологически или генетически модифицированному (инжинирингованному) антигену, содержащему белки, белковые субъединицы, пептиды, углеводороды, липиды, нуклеиновые кислоты, убитые или аттенуированные вирусы, причем оные могут быть целыми вирусными частицами или частями вирусных частиц, или их комбинацией. Антиген может являться как минимум эпитопом, например, T-клеточным и/или B-клеточным эпитопом. Упомянутый антиген детектируется иммунологическими рецепторами, например, T-клеточным рецептором или B-клеточным рецептором. Вакцину используют после применения в специфической активации иммунной системы в отношении конкретного вируса. Таким образом, реакцию иммунной системы используют как инициации иммунного ответа в присутствии вирусов и их специфических антигенов, соответственно. Это приводит к образованию антител и специфических T-хелперных клеток, способных предоставлять долговременную защиту от конкретного заболевания, которое, в зависимости от вируса, длится от нескольких лет до всей продолжительности жизни. Вакцины содержат живые или инактивированные вакцины. Живая вакцина, например, содержит аттенуированные вирусы, сохраняющие способность к репродуцированию вирусов, которые не могут вызвать заболевание. В случае использования инактивированной вакцины, данные вирусы убивают или содержат только фрагменты вируса (антигены). Инактивирование (убийство) вируса происходит, например, путем воздействия химических веществ, таких, как формальдегид, бета-пропиолактон и псоралин. Вирусная оболочка остается неизменной. Существуют также токсоидные вакцины, содержащие только биологически инактивированную часть (токсоид) токсина вируса (например, столбнячный токсин), которые также включают в убитые вакцины. В частности, инактивированная вакцина может представлять собой расщепленную вакцину, содержащую фрагменты белков вирусной оболочки. Разрушение или расщепление вирусной оболочки может иметь место, например, при использовании детергентов или сильных органических растворителей. Вирусы могут инактивировать и убивать добавлением химических агентов, соответственно. Помимо этого, субъединичные вакцины также являются частью убитых вакцин; они содержат специфические компоненты вируса, например, белки гемагглютинина и нейраминидазы.
Термин «вакцина на основе вируса гриппа», в том значении, в котором его используют в описании заявленного изобретения, относится ко всем белкам, пептидам или их частям, а также нуклеиновым кислотам, кодирующим данные белки, пептиды или части вируса гриппа, а также сами частицы вируса гриппа, рекомбинантные белки вируса гриппа, включая белки оболочки вируса гриппа, субвирусные частицы, вирусоподобные частицы (VLP), комплексы с вирусоподобными частицами и/или их части, которые могут использовать для иммунизации против гриппа.
Термин «адъювант», в том значении, в котором его используют в описании заявленного изобретения, относится к веществам, способным модулировать иммуногенность антигена. Адъювантами являются, например, минеральные соли, скваленовые смеси, мурамилпептиды, производные сапонина, препараты клеточной стенки микобактерий, определенные эмульсии, монофосфорил липид A, производные миколевой кислоты, сурфактанты неионных блок-полимеров, Quil A, B-субъединица холерного токсина, полифасфазены и их производные, иммуностимулирующие комплексы, цитокиновые адъюванты, MF59 адъювант, липидные адъюванты, мукозные адъюванты, определенные бактериальные экзотоксины, специфические олигонуклеотиды и PLG.
Термин «амниоциты», в том значении, в котором его используют в описании заявленного изобретения, относится ко всем клеткам, которые присутствуют в амниотической жидкости, и могут быть изъяты путем амниоцентеза. Эти клетки происходят либо из амниона, либо тканей плода, соприкасающихся с амниотической жидкостью. По морфологическому признаку, амниоциты подразделяют на три основных класса: фибробластоподобные клетки (Ф-клетки), эпителиоидные клетки (Э-клетки) и клетки амниотической жидкости (АЖ-клетки) (Hohn et al., Pediat. Res. 8:746-754, 1974). АЖ-клетки являются доминирующим классом.
Термин «постоянные линии клеток», в том значении, в котором его используют в описании заявленного изобретения, относится к клеткам, подвергнутым генетической модификации таким образом, что они могут продолжать расти перманентно в клеточной культуре при стабильных условиях культуры. Такие клетки также получили название «иммортализованных».
Термин «первичные клетки» в том значении, в котором его используют в описании заявленного изобретения, относится к клеткам, которые были получены путем прямого изъятия из организма или ткани, и помещены в культуру. Первичные клетки имеют очень ограниченную продолжительность жизни.
Термин «трансфекция», в том значении, в котором его используют в описании заявленного изобретения, относится к любому способу, подходящему для введения упомянутой нуклеиновой кислоты (кислот) в клетки. В качестве примеров (в т.ч., в сочетании) можно привести способ кальциево-фосфатный, электропорацию, липосомальные системы различных типов.
Термин «метилированный кэп» («CAP»), в том значении, в котором его используют в описании заявленного изобретения, относится к постоянной линии амниоцитов человека, полученной путем иммортализации первичных амниоцитов человека с генными факторами аденовируса E1A и E1B.
Термин «метилированный кэп-Т» («CAP-Т»), в том значении, в котором его используют в описании заявленного изобретения, относится к CAP-клеткам, которые в дополнение были подвергнуты стабильной трансфекции с молекулой нуклеиновой кислоты, содержащей последовательность большого SV40 Т-антигена.
Объект заявленного изобретения относится к способу получения вакцины на основе вируса гриппа, включающему следующие этапы:
(i) контакт вируса гриппа с постоянной клеткой человека,
(ii) культивирование постоянной клетки человека,
(iii) обеспечение экспрессии вируса гриппа, и
(iv) изолирование вируса гриппа из среды.
Согласно заявленному способу, постоянные клетки человека культивируют при параметрах (например, температура, среда, pH), которые благоприятны для роста клеток. Условия касательно температуры, среды, значения pH и прочие параметры роста, известны специалистам в данной области техники, или могут быть определены обычными методами. Как только культура достигла требуемой плотности роста, для инфицирования клеток добавляют вирусы гриппа. Для размножения вируса внутри клеток может потребоваться несколько дней. Во время этого процесса размножения, большая часть клеток погибает, а вирусы высвобождаются в среду. Содержащий вирус раствор отделяют от клеточных остатков, например, путем центрифугирования. Вирус затем могут отделить от раствора среды при помощи, например, хроматографической колонки, а объем могут уменьшить. Затем, вирусы могут инактивировать, например, химическим процессом, с последующим расщеплением вируса, при необходимости. После дополнительной очистки и концентрирования, получают концентрат антигена штамма вируса.
В предпочтительном варианте практического воплощения заявленного изобретения, штаммы вирусов гриппа A/PR/8/34, A/Uruguay/716/2007, A/Brisbane/59/2007, B/Florida/4/2006, свиного гриппа (A/Swine (H1N2) Bakum/1832/00) или лошадиного гриппа (A/Equine, A/Newmarket/1/93 (H3N8)) используют для инфицирования постоянных клеток человека.
В еще одном предпочтительном варианте практического воплощения заявленного изобретения, вирусы гриппа, используемые для инфицирования постоянных линий клеток человека, предварительно адаптируют к клеткам; предпочтительно, они представляют собой вышеперечисленные вирусы гриппа. Предпочтительно, данная адаптация проходит в 4 прохода. Предпочтительно, адаптацию вирусов гриппа осуществляют в 293SFMII среде или PEM среде.
Дополнительно, объект заявленного изобретения относится к способу получения вакцины на основе вируса гриппа, включающему следующие этапы:
(i) контакт молекулы нуклеиновой кислоты, кодирующей белок вируса гриппа, с постоянной клеткой человека,
(ii) культивирование постоянной клетки человека,
(iii) обеспечение репликации молекулы нуклеиновой кислоты, кодирующей белок вируса гриппа и/или экспрессии белка гриппа, и
(iv) изолирование молекулы нуклеиновой кислоты, кодирующей белок гриппа и/или белок вируса гриппа из среды.
В предпочтительном варианте практического воплощения заявленного изобретения, постоянные клетки человека, применяемые в способе согласно заявленному изобретению, представляют собой постоянные клетки-амниоциты человека.
В предпочтительном варианте практического воплощения заявленного изобретения, постоянные клетки человека культивируют во встряхиваемых колбах или биореакторах, предпочтительно STR или Wave Bioreactors. Постоянные клетки человека могут культивировать в различных средах, но предпочтительно в 293SFMII или PEM среде. Дополнительно, к среде могут добавить пируват, глутамин, глюкозу и другие аминокислоты. Предпочтительно, среда содержит 4 мМ или 10 мМ пирувата или других аминокислот.
В еще одном предпочтительном варианте практического воплощения заявленного изобретения, первоначальная концентрация постоянных клеток человека при культивировании во встряхиваемых колбах составляет 5×105 клеток/мл, более предпочтительно 8×105 клеток/мл.
В еще одном предпочтительном варианте практического воплощения заявленного изобретения, значение pH клеточной культуры находится в пределах от 7.1 до 7.8, более предпочтительно в пределах от 7.3 до 7.5, наиболее предпочтительно в пределах от 7.3 до 7.5.
В еще одном предпочтительном варианте практического воплощения заявленного изобретения, осуществляют полную смену среды или 1:2 разбавление со средой до инфицирования постоянных клеток человека вирусом гриппа.
В предпочтительном варианте практического воплощения заявленного изобретения, используют трипсин в концентрации 1×10-4 Е/клетку, 1×10-5 Е/клетку, 3×10-5 Е/клетку, 5×10-5 Е/клетку или 1×10-6 Е/клетку для инфицирования постоянных клеток человека вирусом гриппа. При отсутствии смены среды до инфицирования постоянных клеток человека, предпочтительно используют трипсин в концентрации 1×10-4 Е/клетку для инфицирования упомянутых клеток вирусом гриппа. При 1:2 разбавлении со средой до инфицирования постоянных клеток человека, предпочтительно используют трипсин в концентрации 5×10-5 E/клетку для инфицирования упомянутых клеток вирусом гриппа. При полной смене среды до инфицирования постоянных клеток человека, предпочтительно используют трипсин в концентрации 5×10-6 E/клетку для инфицирования упомянутых клеток вирусом гриппа.
В предпочтительном варианте практического воплощения заявленного изобретения, объем вируса, определенный как значение MOI (множественность заражения) в пределах от 0.001 до 0.3 используют для инфицирования постоянных клеток человека. В предпочтительном варианте практического воплощения заявленного изобретения, объем вируса, определенный как значение MOI в пределах от 0.25, 0.1, 0.06, 0.025 или 0.0025 используют для инфицирования постоянных клеток человека. Предпочтительно, когда постоянные клетки человека инфицируют вирусом гриппа A/PR/8/34 без смены среды до инфицирования, объем вируса, определенный как значение MOI 0.25 используют для инфицирования постоянных клеток человека вирусом гриппа; когда постоянные клетки человека инфицируют вирусом гриппа A/Brisbane/59/2007 без смены среды до инфицирования, объем вируса, определенный как значение MOI 0.1 используют для инфицирования постоянных клеток человека вирусом гриппа. Предпочтительно, когда постоянные клетки человека инфицируют вирусом гриппа A/PR/8/34 со сменой среды до инфицирования, объем вируса, определенный как значение MOI 0.1 или 0.25 используют для инфицирования постоянных клеток человека вирусом гриппа; когда постоянные клетки человека инфицируют вирусом гриппа A/Brisbane/59/2007 со сменой среды до инфицирования, объем вируса, определенный как значение MOI 0.06 или 0.25 используют для инфицирования постоянных клеток человека вирусом гриппа; и когда постоянные клетки человека инфицируют вирусом гриппа B/Florida/4/2006 со сменой среды до инфицирования, объем вируса, определенный как значение MOI 0.01, 0.025 или 0.0025 используют для инфицирования постоянных клеток человека вирусом гриппа.
В предпочтительном варианте практического воплощения заявленного изобретения, концентрация клеток в момент инфицирования, в случае культивирования во встряхиваемых колбах, находится в пределах от 1×106 до 6×106 клеток/мл. Предпочтительно, концентрация клеток в момент инфицирования составляет 2.3×106 клеток/мл, 4.5×106 клеток/мл или 5×106 клеток/мл. В предпочтительном варианте практического воплощения заявленного изобретения, концентрация клеток в момент инфицирования составляет 4.5×106 клеток/мл, в случае отсутствия смены среды до инфицирования. В еще одном предпочтительном варианте практического воплощения заявленного изобретения, концентрация клеток в момент инфицирования составляет 2.3×106 клеток/мл, и до инфицирования проводят 1:2 разбавление со свежей PEM средой. В еще одном предпочтительном варианте практического воплощения заявленного изобретения, концентрация клеток в момент инфицирования составляет 5×106 клеток/мл, и до инфицирования проводят полную смену среды.
В особенно предпочтительном варианте практического воплощения заявленного изобретения, постоянные клетки человека культивируют в биореакторе STR (Sartorius) емкостью 1 литр в PEM среде с 4 мМ глутамина и 4 мМ пирувата, первоначальная концентрация клеток составляет 5×105 клеток/мл, причем, при концентрации клеток 2.1×106 клеток/мл с вирусом гриппа в объеме, определенном как значение MOI 0.025, проводят инфицирование без предварительной смены среды. Предпочтительно, инфицирование осуществляют в присутствие трипсина в финальной концентрации 3×10-5 Е/мл.
В особенно предпочтительном варианте практического воплощения заявленного изобретения, постоянные клетки человека культивируют в биореакторе STR (Sartorius) емкостью 1 литр в PEM среде, при этом первоначальная концентрация клеток составляет 8×105 клеток/мл, и при этом инфицирование проводят с вирусом гриппа, используя объем вируса, определенный как значение MOI 0.025, со сменой среды до инфицирования. Предпочтительно, инфицирование проводят в присутствие трипсина в финальной концентрации 3×10-5 Е/мл.
В особенно предпочтительном варианте практического воплощения заявленного изобретения, постоянные клетки человека культивируют в биореакторе Wave (Wave Biotech AG) емкостью 1 литр в PEM среде с 4 мМ глутамина, 4 мМ пирувата и 20 мМ глюкозы в PEM среде, первоначальная концентрация клеток составляет 5×105 клеток/мл, а концентрация клеток до инфицирования составляет 2.1×106 клеток/мл, при этом инфицирование проводят с объемом вируса, определенным как значение MOI 0.025, без смены среды до инфицирования. Предпочтительно, инфицирование проводят в присутствие трипсина в финальной концентрации 3×10-5 Е/мл.
В предпочтительном варианте практического воплощения заявленного изобретения, постоянные клетки человека культивируют в PEM среде с 4 мМ глутамина и 4 мМ пирувата во встряхиваемых колбах, причем проводят смену среды до инфицирования клеток вирусом гриппа, с объемом вируса, определенным как значение MOI 0.025, в присутствие концентрации трипсина 1×10-6 E/клетку.
В предпочтительном варианте практического воплощения заявленного изобретения, постоянные клетки человека культивируют в PEM среде с 4 мМ глутамина и 4 мМ пирувата во встряхиваемых колбах, причем проводят 1:1 смену среды до инфицирования клеток вирусом гриппа, с объемом вируса, определенным как значение MOI 0.025, в присутствие концентрации трипсина 1×10-5 Е/клетку.
В производстве белков гриппа и молекул нуклеиновых кислот, кодирующих белок гриппа, культивированные клетки человека с молекулами нуклеиновых кислот, кодирующими белок гриппа, трансфектируют, и затем белок вируса гриппа или молекулы нуклеиновых кислот, кодирующие белок гриппа, изолируют и очищают, используя известные способы.
В еще одном предпочтительном варианте практического воплощения заявленного изобретения, клетки человека находятся в или между средне-экспоненциальной фазой роста и стационарной фазой роста, согласно заявленному способу, в момент инфицирования вирусной частицей, или в момент трансфекции с молекулой нуклеиновой кислоты, кодирующей белок вируса гриппа или его часть. Типичная кривая роста, в которой изображена зависимость концентрации клеток от времени, представляет собой сигмоидную кривую. Она начинается с так называемой лаг-фазы, с последующими лог-фазой или экспоненциальной фазой роста и стационарной фазой роста. Средняя экспоненциальная фаза роста в данном случае соответствует первой точке перегиба типичной кривой роста, причем точка перегиба является точкой на кривой роста в том месте, где форма кривой меняется от вогнутой к выпуклой, или наоборот. Стационарная фаза начинается, когда кривая роста достигает плоской части, т.е. число клеток остается постоянным.
Нуклеиновые кислоты, получаемые с применением способа согласно заявленному изобретению, кодирующие белок гриппа, предоставляемый изобретательским способом, могут быть использованы для иммунизации нуклеиновыми кислотами, или так называемых ДНК вакцин. При иммунизации нуклеиновыми кислотами, иммуногенные антигены, т.е. антигены, запускающие иммунный ответ у человека, инокулируются. Такие иммуногенные антигены кодируются ДНК или РНК, присутствуют как кассеты экспрессии или векторы, или интегрированы в вирусные векторы с целью индукции иммунного ответа на генный продукт. ДНК вакцины могут предоставлять в различных системах доставки, например, как ДНК или РНК, в форме линеаризованных или кольцевых плазмид или кассет экспрессии, причем они снабжены необходимыми элементами для экспрессии, как, например, промотором, сайтами полиаденилирования, репликатором (точкой начала репликации) и т.п. В случае введения ДНК, аналогичный комплект присутствует в буфере с или без адъюванта, или связанными с наночастицами, или в адъювант-содержащем компаунде, или интегрированными в вирусный или бактериальный вектор. DNA вакцины запускают как гуморальный, так и клеточный иммунный ответ. Преимуществом использования ДНК вакцин является то, что антиген эксперссируют в его нативной форме, что приводит к улучшенной иммунизации. Другим преимуществом использования ДНК вакцин является то, что, в отличие от ослабленных живых вакцин, они неинфекционные и не могут быть снова вирулентными.
Введение ДНК вакцин в форме ДНК или РНК, плазмидных или линейных фрагментов ДНК, которые связаны с частицами, могут осуществлять путем инъекций или генетической пушки. Например, ДНК вакцины для инъекций могут присутствовать в физиологическом или забуференном растворах.
Нуклеиновые кислоты, получаемые с применением способа согласно заявленному изобретению, кодирующие белок гриппа, белок гриппа и вирус гриппа, могут использовать в качестве вакцины против вируса гриппа типов A и/или B и/или C.
Вакцина на основе вируса гриппа, полученная с применением способа в соответствии с заявленным изобретением, содержит все белки, пептиды или их части, а также нуклеиновые кислоты, кодирующие данные белки, пептиды или их части, -- вируса гриппа, а также сами частицы вируса гриппа, рекомбинантные белки вирусов гриппа, включая белки оболочки вируса гриппа, субвирусные частицы, вирусоподобные частицы (VLP), комплексы с вирусоподобными частицами и/или их части, которые могут использовать для иммунизации против гриппа.
Предпочтительно, белки гриппа, полученные с применением способа в соответствии с заявленным изобретением, представляют собой белки или производные вируса гриппа, предпочтительно штаммов вирусов гриппа A/PR/8/34, A/Uruguay/716/2007, A/Brisbane/59/2007, B/Florida/4/2006, вируса свиного гриппа (A/Swine (H1N2) Bakum/1832/00) или вируса лошадиного гриппа (A/Equine, A/Newmarket/1/93 (H3N8)).
Изолирование и очистку нуклеиновых кислот, кодирующих белок вируса гриппа или его части, полученных с применением способа в соответствии с заявленным изобретением, осуществляют с применением традиционных методов, известных специалистам в данной области техники.
Изолирование и очистку белков вирусов гриппа, полученных с применением способа в соответствии с заявленным изобретением, осуществляют с применением традиционных методов, известных специалистам в данной области техники. Очистка белков первоначально зависит от их происхождения. Различие делают между внутри- и внеклеточными белками. Если белки расположены внутри клеток, вначале необходимо разрушить клеточные стенки, например, используя силу трения или осмолизис. После этого, осуществляют сепарирование нерастворимого материала, например, клеточных мембран и клеточных стенок, к примеру, путем центрифугирования (способ, применяемый по умолчанию для сепарирования клеток, клеточных органелл (органоидов) и белков). Более эффективным способом, с точки зрения емкости сепарирования, является импульсный электрофорез. Дополнительно, после сепарирования прочих клеточных компонентов, все еще присутствует необходимость сепарировать белки разных размеров, пептиды и аминокислоты. Сепарирование белков могут проводить одно- или двумерным гель-электрофорезом или капиллярным электрофорезом. Для аминокислот и пептидов применяют, например, хроматографические методы, а именно, аффинную хроматографию, ионообменную хроматографию (IEC), или обращенно-фазовую хроматографию (RPC). Присутствие липидов и необходимость удаления или деактивирования протеаз является неблагоприятным моментом процесса очистки. Белки, присутствующие во внеклеточной матрице, не нуждаются в экстрагировании из клеток, однако, после сепарирования всех нерастворимых компонентов, они сильно разбавлены и наличествуют в гораздо меньших количествах, чем как внутриклеточные белки.
Изолирование и очистку частиц вирусов гриппа, полученных с применением способа в соответствии с заявленным изобретением, осуществляют с применением традиционных методов, известных специалистам в данной области техники, например, путем дифференциального центрифугирования в градиенте плотности или зонального центрифугирования.
Постоянные клетки человека, применяемые в способе в соответствии с заявленным изобретением, генерируют иммортализацией первичных клеток человека. Первичные клетки человека получают путем прямого изъятия из организма или ткани, взятой из организма, и помещают в культуру. Предпочтительны такие первичные клетки человека, которые хорошо конвертируются в линии постоянных клеток человека путем экспрессии с клеточными трансформирующими факторами, в частности, амниоцитами, зародышевыми клетками сетчатки и зародышевыми клетками нейронного происхождения.
Клеточными трансформирующими факторами являются T-антиген SV40 (Genbank Асе. No. J02400), E6 и E7 генные продукты вируса папилломы человека ВПЧ (HPV) (например, HPV 16, Genbank Асе. No. K02718), E1A и E1B генные продукты аденовирусов человека (например, аденовирус человека серотип-5, Genbank Асе. No. Х02996). Первичные клетки трансфектируют для иммортализации путем экспрессии аденовируса человека E1 с двумя последовательностями нуклеиновых кислот для генных продуктов E1A и E1B. В случае экспрессии природного ВПЧ, E6 и E7 могут экспрессировать из транскрипта РНК. Аналогичные параметры применимы для экспрессии E1A и E1B природного аденовируса. Клеточные трансформирующие факторы, например, аденовирусный E1 генный фактор, вызывают иммортализацию или трансформацию, и, соответственно, способность клеток к долговременному культивированию.
Экспрессию клеточных трансформирующих факторов могут осуществлять под контролем гомологичного промотора, и элементов транскрипционных стоп-кодонов, например, природного E1A промотора и природного E1A сайта полиаденилирования для экспрессии аденовирусного E1A генного фактора. Этого могут достичь, используя молекулы нуклеиновых кислот для трансфекции соотвествующих вирусных геномных фрагментов, например, аденовирусного генома, который содержит упомянутые генные факторы, как E1A, E1B. Экспрессию клеточных трансформирующих факторов могут также осуществлять под контролем гетерологичных промоторов, промоторов, обычно отсутствующих с использованными кодирующими областями, или элементов транскрипционных стоп-кодонов. В качестве гетерологичных промоторов могут, к примеру, выступать промотор ЦМВ (CMV, цитомегаловирус) (Makrides, 9-26 in: Makrides (Eds.), Gene Transfer and Expression in Mammalian Cells, Elsevier, Amsterdam, 2003), промотор EF-1α (Kim et al, Gene 91st:217-223, 1990), промотор ЦАГ (CAG, гибридный промотор предраннего энхансера человеческого цитомегаловируса и модифицированного куриного β-активного промотора с первым интроном) (Niwa et al., Gene 108:193-199, 1991), промотор человеческой или мышиной pgk (фосфоглицераткиназа) (Adra et al, Gene 60:65-74, 1987), промотор BCP (RSV, вирус саркомы Рауса) (Makrides, 9-26 in: Makrides (ed.), Gene Transfer and Expression in Mammalian Cells, Elsevier, Amsterdam, 2003), или промотор SV40 (вирус обезьяны 40) (Makrides, 9-26 in: Makrides (ed.), Gene Transfer and Expression in Mammalian Cells, Elsevier, Amsterdam, 2003). Например, последовательности полиаденилирования большого SV40 T-антигена (GenBank асе. no. J02400) или человеческого ГКСФ/G-CSF (гранулоцитарный колониестимулирующий фактор) гена (Mizushima und Nagata, Nucl. Acids Res. 18:5322, 1990) могут выбирать в качестве сайтов полиаденилирования.
Клетки иммортализуют путем трансфекции первичных клеток человека с молекулой нуклеиновой кислоты, содержащей последовательности нуклеиновых кислот, кодирующие E1A и E1B. Молекула нуклеиновой кислоты, используемая для иммортализации упомянутых первичных клеток человека, содержит последовательности нуклеиновых кислот для E1A и E1B, более предпочтительно получаемые из аденовирусов человека, в частности, серотипа 5 человеческого аденовируса. В предпочтительном варианте практического воплощения заявленного изобретения, молекула нуклеиновой кислоты, используемая для иммортализации, содержит последовательность нуклеиновой кислоты, кодирующую фактор (функцию) pIX гена аденовируса, в дополнение к последовательностям нуклеиновых кислот, кодирующих E1A и E1B. Полипептид pIX, вирусный структурный белок, выполняет роль транскрипционного активатора на различных вирусных и клеточных промоторах, как, например, тимидинкиназа и промотор бета-глобина. Транскрипционно-активирующее влияние полипептида pIX дополнительно экспрессируется на клетках и может вызвать повышение уровня экспрессии рекомбинантного полипептида в процессе получения линий клеток согласно заявленному изобретению, в случае, если кодирующая последовательность рекомбинантного полипептида находится под контролем одного из вышеупомянутых промоторов. Типовую последовательность можно найти в GenBank acc. no. X02996. В частности, молекулы нуклеиновых кислот содержат нуклеотиды с 1 по 4344, с 505 по 3522 или нуклеотиды с 505 по 4079 аденовируса человека серотип-5.
В предпочтительном варианте практического воплощения заявленного изобретения, молекула нуклеиновой кислоты содержит для иммортализации первичных клеток, в частности, амниоцитов, нуклеотидную последовательность аденовируса серотип-5 от нуклеотида 505 до нуклеотида 4079. В более предпочтительном варианте практического воплощения заявленного изобретения, молекула нуклеиновой кислоты содержит для иммортализации первичных клеток, в частности, амниоцитов, нуклеотидную последовательность аденовируса серотип-5 от нуклеотида 505 до нуклеотида 3522. В еще одном более предпочтительном варианте практического воплощения заявленного изобретения, молекула нуклеиновой кислоты содержит для иммортализации первичных клеток, в частности, амниоцитов, нуклеотидную последовательность аденовируса серотип-5 от нуклеотида 1 до нуклеотида 4344, соответствуя аденовирусной ДНК в клетках HEK 293 (Louis et al, Virology 233rd: 423-429, 1997). Помимо этого, иммортализованные клетки человека могут экспрессировать фактор репликации вируса. Данный фактор репликации может связываться с точкой начала репликации (ori, «начало репликации») молекулы нуклеиновой кислоты, введенной путем трансфекции и, таким образом, инициируя эписомальную репликацию молекулы нуклеиновой кислоты. Эписомальная репликация молекул нуклеиновых кислот, в частности, плазмидной ДНК, в клетки приводит к значительному увеличению числа копий трансфектированных молекул нуклеиновых кислот, и, таким образом, повышению экспрессии рекомбинантного полипептида, кодируемого данной молекулой, а также ее поддержанию во время множества клеточных делений. Таким фактором вирусной репликации является, например, T-антиген вируса обезьяны 40 (SV40), который после связывания с последовательностью, обозначенной как точка начала репликации SV40 (SV40 ori, начало репликации) на молекуле нуклеиновой кислоты, например, плазмидной ДНК, инициирует ее репликацию. Белок вируса Эпштейн-Барр EBNA-1 (ядерный антиген-1 вируса Эпштейн-Барр) распознает точку начала репликации, определенную как ori-P и катализирует экстрахромосомную репликацию ori-P, несущую молекулу нуклеиновой кислоты. T-антиген вируса обезьяны 40 (SV40) активирует репликацию не только как фактор репликации, но также оказывает активирующее действие на транскрипцию некоторых вирусных и клеточных генов (Brady, John and Khoury, George, 1985, Molecular and Cellular Biology, vol 5, no.6, p.1391 to 1399).
Иммортализованная клетка человека, используемая в способе согласно заявленному изобретению, представляет собой в частности иммортализованную клетку-амниоцит человека. В предпочтительном варианте практического воплощения заявленного изобретения, иммортализованная клетка человека, используемая в способе согласно заявленному изобретению, экспрессирует большой T-антиген SV40 или ядерный антиген-1 (EBNA-1) вируса Эпштейна-Барр (ЭБВ). В особенно предпочтительном варианте практического воплощения заявленного изобретения, иммортализованная клетка-амниоцит человека, используемая в способе согласно заявленному изобретению, экспрессирует большой T-антиген SV40 или ядерный антиген-1 (EBNA-1) вируса Эпштейна-Барр (ЭБВ). В еще одном особенно предпочтительном варианте практического воплощения заявленного изобретения, иммортализованная клетка человека, а именно, клетка-амниоцит, используемая в способе согласно заявленному изобретению, экспрессирует большой T-антиген SV40 под контролем промоторов ЦАГ SV40, ВСР или ЦМВ.
Постоянные амниоциты человека, используемые в способе согласно заявленному изобретению, подробно описаны в патентах EP 1230354 и EP 1948789. В особенно предпочтительном варианте практического воплощения заявленного изобретения, постоянная клетка-амниоцит человека, используемая в способе согласно заявленному изобретению, представляет собой CAP или CAP-T.
В случае использования CAP-клеток, первичные амниоциты трансфектировали с плазмидой, содержащей промотор мышиной pgk, Ad5 последовательности nt. 505-3522, содержащие всю область E1, 3' сигнал полиаденилирования и сплайса SV40 и pIX область Ad5 (nt. 3485-4079). Данная плазмида подробно описана в EP 1948789.
Для получения CAP-T-клеток, CAP-клетки трансфектировали с плазмидой, содержащей кассету экспрессии для Т-антигена SV 40, фланкированную интроном из SV40 и сайтом полиаденилирования. Дополнительно, плазмида может содержать промотор ЦАГ (гибридный промотор, состоящий из ЦМВ энхансера и промотора куриного β-актина) (Niwa et al., Gene 108:193-199, 1991), промотор ВСР (промотор вируса саркомы Рауса) (GenBank асе. no. DQ075935) или промотор ЦМВ (ранний промотор человеческого цитомегаловируса) (SEQ ID NO:5). Для генерации стабильных линий клеток, плазмида содержит кассету экспрессии бластицидина с промотором убиквитина (pUB/Bsd, Invitrogen # V512-20).
Кроме этого, заявленное изобретение предоставляет способ согласно заявленному изобретению, при котором клетка человека, в особенности, клетка-амниоцит, может расти в суспензии. Помимо этого, клетку человека, в особенности, клетку-амниоцит, используемую в способе согласно заявленному изобретению, могут культивировать в бессывороточной среде.
Еще одной целью заявленного изобретения является использование постоянной клетки человека, в особенности, клетки-амниоцита, для получения вакцины на основе вируса гриппа.
В предпочтительном варианте практического воплощения заявленного изобретения, постоянная клетка-амниоцит человека, используемая для получения вакцины на основе вируса гриппа, представляет собой CAP- или CAP-T клетку.
Вакцина на основе вируса гриппа, полученная с применением способа согласно заявленному изобретению, может представлять собой вирус гриппа и/или белок вируса гриппа или молекулу нуклеиновой кислоты, кодирующую белок гриппа. Вакцину могут вводить парентерально, при помощи шприца. Различают внутрикожные, подкожные или внутримышечные инъекции. Внутрикожную инъекцию можно выполнять с использованием прививочного медицинского пистолета или ланцета. Внутримышечную инъекцию можно проводить в верхнее предплечье, бедро или ягодицу. Помимо этого, вакцину можно вводить перорально или назально, человеку и животным.
Вакцина на основе вируса гриппа, полученная с применением способа согласно заявленному изобретению, может предоставлять как активную, так и пассивную иммунизацию, сопротивляемость в отношении одного или нескольких вирусов гриппа. Для активной иммунизации, вакцину используют после применения для специфической активации иммунной системы человека и животных, в отношении конкретного вируса. В данном случае, реакцию иммунной системы используют для инициации иммунного ответа в присутствие вирусов или их специфических антигенов. Это приводит к образованию антител и специализированных T-хелперных клеток, которые могут обеспечить долговременную защиту от конкретного заболевания, длительность которой, в зависимости от вируса, может составлять от нескольких лет или пожизненно.
Вакцина на основе вируса гриппа, полученная с применением способа согласно заявленному изобретению, может представлять собой, например, живую или инактивированную вакцину. Живая вакцина, например, содержит аттенуированные вирусы, сохраняющие способность к репродуцированию вирусов, которые не могут вызвать заболевание. В инактивированной вакцине, данные вирусы убивают или включают только фрагменты вируса (антигены). Инактивирование (убийство) вируса происходит, например, путем воздействия химических веществ, таких, как формальдегид, бета-пропиолактон и псоралин. Вирусная оболочка остается неизменной. Существуют также токсоидные вакцины, содержащие только биологически инактивированную часть (токсоид) токсина вируса (например, столбнячный токсин), которые также включают в убитые вакцины. В частности, инактивированная вакцина может представлять собой расщепленную вакцину, содержащую фрагменты белков вирусной оболочки. Разрушение (расщепление) вирусной оболочки может иметь место, например, при использовании детергентов или сильных органических растворителей. Вирусы могут инактивировать (убивать) добавлением химических агентов. Помимо этого, субъединичные вакцины также являются частью убитых вакцин; они содержат специфические компоненты вируса, например, белки гемагглютинина и нейраминидазы.
В случае пассивной иммунизации, вакцину на основе вируса гриппа, полученную с применением способа согласно заявленному изобретению, вводят в хозяина (например, млекопитающее), индуцированную антисыворотку экстрагируют и затем вводят реципиенту, который инфицирован как минимум одним вирусом гриппа.
Помимо этого, для введения вакцину смешивают с одним или несколькими добавками, например, стабилизаторы, нейтрализаторы, носители и консерванты. Данные вещества включают формальдегид, тимеросал, фосфат алюминия, ацетон и фенол. Дополнительно, вакцину могут смешивать со вспомогательными веществами для усиления действия вакцины. Эти так называемые адъюванты сами по себе не должны оказывать какого-либо фармакологического действия, лишь выполняя функцию солюбилизатора, эмульгатора или их смесей. Адъювантами являются, например, минеральные соли, скваленовые смеси, мурамилпептиды, производные сапонина, препараты клеточной стенки микобактерий, определенные эмульсии, монофосфорил липид A, производные миколевой кислоты, сурфактанты неионных блок-полимеров, Quil A, B-субъединица холерного токсина, полифасфазены и их производные, иммуностимулирующие комплексы, цитокиновые адъюванты, MF59 адъювант, липидные адъюванты, мукозные адъюванты, определенные бактериальные экзотоксины, специфические олигонуклеотиды и PLG.
Следующие примеры иллюстрируют заявленное изобретение и не носят ограничительного характера относительно сути заявленного изобретения. Если не указано иначе, используют стандартные методы молекулярного биологического исследования, как, например, описано Sambrook et al, 1989, Molecular Cloning. A Laboratory Manual 2nd Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Пример 1: Опыты по культивированию постоянной линии амниоцитов CAP-1D5 в PEM или 293SFMII средах
Постоянную линию клеток-амниоцитов CAP-1D5 культивировали в 100 мл бессывороточной 293SFMII среды (Invitrogen) или PEM среды (Invitrogen) при 37°C, 8% CO2 и 100 оборотах в минуту. В качестве контроля, постоянную линию клеток почек собак MDCK.SUS2 (Madin Darby Canine Kidney, адаптированную к росту в суспензии) (Lohr et al., Vaccine, 2010, 28(38):6256-64) использовали в 100 мл бессывороточной SMIF8 среды во встряхиваемых колбах емкостью 100 мл.
Во временной точке 0 (= старт культуры), и в каждом случае, промежутками через 24 ч, определяли концентрацию жизнеспособных клеток, концентрацию мертвых клеток, уровень выживаемости линий клеток, а также значение pH, концентрацию глюкозы, лактозы, глутамина, аммония, глутаминовой кислоты и пирувата в среде (система анализа биохимических мультипараметров) (Lohr et al, Vaccine, 2009, 27 (37), 4975-4982; Genzel et al, Appl Microbiol Biotechnol, 2010, 88 (2):461-75).
Результаты представлены на фиг.1. Концентрация жизнеспособных клеток постоянной линии клеток-амниоцитов CAP 1D5 в 293SFMII, а также в PEM среде, а также клеток MDCK.SUS2, начиная от примерно 2×105 клеток/мл культуры в начале культивирования, показали одинаковую динамику, и после 168 часов, концентрация жизнеспособных клеток составляла примерно 2×106 клеток/мл. От 192 ч и далее, концентрация жизнеспособных клеток MDCK.SUS2 клеток упала до 9×105 клеток/мл и оставалась постоянной вплоть до 240 ч после начала кривой роста. Концентрация жизнеспособных клеток постоянной линии клеток-амниоцитов CAP-1D5 в 293SFMII среде упала только в отметке 216 ч, до первоначального значения 2×105 клеток/мл культуры, однако, концентрация жизнеспособных клеток CAP-1D5 оставалась стабильной в PEM среде до отметки 240 ч до примерно 2.5×106 клеток/мл культуры, и, после 312 ч, достигла первоначального значения концентрации жизнеспособных клеток 2×105 клеток.
В начале кривой роста и до отметки 168 ч, уровень выживаемости CAP-1D5 клеток в 293SFMII среде и в PEM среде, а также MDCK.SUS2 клеток, находился в пределах от 80 до 90%. До 240 ч, уровень выживаемости CAP-1D5 клеток в PEM среде оставался на 80%, затем падал, и после 312 ч оставался на 10%. Уровень выживаемости CAP-1D5 клеток в 293SFMII среде уже понижался после отметки 168 ч и установился на примерно 10% после 216 ч. Уровень выживаемости MDCK.SUS2 клеток постоянно снижался после 168 ч, оставался стабильным и достигал 45% на отметке 240 ч.
Значение pH MDCK.SUS2 клеток оставалось относительно стабильным в течение 240 ч, в пределах от 7.7 до 7.6. Тем не менее, значение pH (первоначально 7.4) в культуре CAP-1D5 клеток в 293SFMII среде, а также в PEM среде, неуклонно снижалось до pH 6.4 после 240 ч (293SFMII среда) и 312 ч (PEM среда), соответственно.
Концентрация лактозы увеличилась с первоначальной на уровне ниже 5 мМ в 293SFMII среде, в PEM среде соответствующей культуры CAP-1D5 клеток до 30-35 мМ вплоть до 240 ч. В культуре MDCK.SUS2 клеток, концентрация лактозы увеличивалась меньшими темпами, но достигла аналогичного значения в CAP-1D5 клеточной культуре в отметке 240 ч. Концентрация глюкозы уменьшалась на 20-25 мМ от начала культуры CAP-1D5 клеток в 293SFMII среде и PEM среде, а также в культуре MDCK.SUS2 клеток до уровня ниже 10 мМ, причем самый резкий спад зафиксировали в культуре CAP-1D5 клеток в 293SFMII среде.
Повышение концентрации аммония в культуре CAP-1D5 клеток в 293SFMII среде, а также в PEM среде, проходило аналогично (от <0.5 до 4-5 мМ). В отличие от этого, концентрация аммония в культуре MDCK.SUS2 клеток повышалось значительными темпами (до 7 мМ). Снижение концентрации глутамина также проходило аналогичным образом в культуре CAP-1D5 клеток в 293SFMII среде, а также в PEM среде, причем самый резкий спад зафиксировали в PEM среде.
Самую высокую концентрацию глутаминовой кислоты зафиксировали в культуре MDCK.SUS2 клеток, с очень небольшими колебаниями. Концентрация глутаминовой кислоты была выше в начале культивирования в культуре CAP-1D5 клеток в PEM среде, в отличие от 293SFMII среды, и повышалась устойчивыми темпами. Концентрация пирувата в культуре MDCK.SUS2 клеток понижалась до нулевой отметки от 144 ч и далее. Тем не менее, весь объем пирувата был использован после 48 ч в культурах CAP-1D5 клеток в PEM среде и в 293SFMII среде, соответственно.
Таким образом, CAP-1D5 клетки показали в PEM среде более благоприятный рост, чем в 293SFMII среде. CAP-1D5 клетки в PEM среде показали самое высокое потребление глюкозы и самый высокий уровень образования лактозы. Ограничение образования лактозы от отметки 192 ч и далее может объяснить снижение числа клеток CAP-1D5 в PEM среде. Путем оптимизации параметров культивирования клеточной культуры CAP-1D5 в PEM среде и 293SFMII среде, соответственно, стабилизация значения pH, добавление пирувата и глутамина, а также глюкозы, могут быть релевантными.
Пример 2: Вирусное инфицирование с неадаптированными вирусами
Для опытов по инфицированию с неадаптированными вирусами, линию клеток-амниоцитов CAP-1D5 культивировали в 55 мл бессывороточной 293SFMII среды (Invitrogen) и PEM среды (Invitrogen) при 37°C, 8% CO2, 100 оборотах в минуту во встряхиваемых колбах. В качестве контроля, использовали постоянную линию клеток почек собак MDCK.SUS2 (Madin Darby Canine Kidney, адаптированную к суспензии).
В табл.1 представлены показатели плотности клеток, которые инфицировали вирусом B/Florida/4/2006 (NIBSC, Национальный институт биологических стандартов и контроля) и A/PR/8/34 (H1N1) (RKI, Институт им. Роберта Коха) без смены среды, и с добавлением 5×10-6 Е/мл трипсина, соответственно. Объем полученных вирусных частиц определили титрованием гемагглютинина (НА, гемагглютинация стандартными методами) (в log HA единицах/100 мкл; Таблица 1) (Kalbfuss et al, 2008;. Biologicals 36 (3):145-61).
Также представлены в табл.1 соответствующие значения pH среды в соответствующих временных отметках.
Для постоянной линии клеток-амниоцитов CAP-1D5 при культивировании в PEM среде, получение вирусных частиц невозможно осуществить как с инфицированием вирусом B/Florida/4/2006 (B/Florida), так и вирусом A/PR/8/34. Максимальное значение НА для A/PR/8/34 (H1N1) в 293SFM среде было ниже максимального значения НА в MDCK.SUS2. Значение pH при максимальном HA сравнимо со всеми примерами инфицирования в CAP-1D5 (pH 6.6), но значительно ниже, чем в инфицированных MDCK.SUS2 клетках (pH 7.7).
Таким образом, инфицирование постоянной линии клеток-амниоцитов CAP 1D5 при параметрах опыта происходило только при наличии вируса B/Florida. Однако, в случае использования постоянной линии клеток почек собак MDCK.SUS2 (Madin Darby Canine Kidney), инфицирование детектировали только в случае вируса A/PR/8/34 (H1N1). В последующих опытах, устанавливали, можно ли достичь улучшенного инфицирования и, таким образом, более высокого выхода вируса путем предварительной адаптации вирусов к CAP-1D5 клеткам.
Пример 3: Вирусная адаптация в CAP-1D5 клетках в PEM и 293SFMII средах во встряхиваемых колбах
Вирусную адаптацию вирусов гриппа A/PR/8/34 (H1N1) (RKI, Институт им. Роберта Коха), A/Uruguay/716/2007 (H3N2) (NYMC Х-175С, NIBSC, Национальный институт биологических стандартов и контроля) и B/Florida/4/2006 (NIBSC, Национальный институт биологических стандартов и контроля), соответственно, осуществили путем инфицирования CAP-1D5 в 4 прохода во встряхиваемых колбах в PEM и 293SFMII средах. До каждого инфицирования осуществили смену среды. Урожай вируса во время каждого прохода определили титрованием гемагглютинина (log НА единиц/100 мкл) и квантифицировали с использованием Tissue Culture Infectious Doseso Assay (TCID50 вирусов/мл) (Genzel and Reichl, Vaccine production - state of the art and future needs in upstream processing in Methods of Biotechnology: Animal Cell Biotechnology - Methods and Protocols, Ed R. Portner, Humana Press Inc., Totowa, NJ, 2007, 457-473; Kalbfuss et al, 2008; Biological 36 (3):145-61).
Результаты представлены на фиг.2. Инфицирование CAP-1D5 клеток вирусом гриппа A/PR/8/34 (H1N1) и A/Uruguay/716/2007 (H3N2) вызвало значительное повышение, как в PEM, так и в 293SFMII средах, титров вируса после 4 проходов. Инфицирование CAP-1D5 клеток вирусом гриппа B/Florida/4/2006 в культивировании в 293SFMII и в PEM средах, привело к значительному росту титра вируса во втором проходе. Аналогичный рост зафиксирован в титровании значения НА во время инфицирования CAP-1D5 клеток вирусом гриппа A/PR/8/34 (H1N1) или A/Uruguay/716/2007 (H3N2) как в PEM, так и в 293SFMII средах.
Наряду с повышением титра вируса во время адаптации, с каждым проходом репликация вируса проходила быстрее и, в конечном итоге, могла быть значительно увеличена. В общем, представляется, что можно достичь небольшого роста титра вируса в 293SFMII среде, в сравнении с PEM средой.
Пример 4: Зависимость MOI (множественность заражения) от инфицирования CAP-1D5 и MDCK.SUS2 клеток адаптированным гриппом A/PR/8/34
С адаптированным вирусом гриппа A/PR/8/34 (H1N1, RKI, Институт им. Роберта Коха), испытали зависимость MOI при 3 различных значениях MOI (множественность заражения), измерили число вирусных частиц на клетку-хозяина, 0.0025, 0.025 и 0.25. Постоянные клетки-амниоциты в CAP-1D5 инфицировали в 293SFMII и PEM средах, соответственно, и также постоянные клетки почек собак MDCK.SUS2 в SMIF8 среде, во встряхиваемых колбах, с различными MOI адаптированного вируса гриппа A/PR/8/34. При инфицировании, также осуществили смену среды, что привело к почти постоянному значению pH примерно pH=7.5. В течение 96 ч, определяли концентрацию жизнеспособных клеток, а число вирусных частиц (log HA единиц/100 мкл) - в течение 144 ч титрованием гемагглютинина в опыте по гемагглютинации с использованием стандартных методов.
Результаты представлены на фиг.3. Динамика изменения концентрации жизнеспособных клеток, а также динамика изменения числа образовавшихся вирусных частиц в культуре CAP-1D5 клеток в 293 SFM среде и в культуре MDCK.SUS2 клеток в SMIF8 среде показали отсутствие зависимости от значений MOI. Титр вируса повысился до примерно 2.5 log HA - единиц/100 мкл в двух культурах. Культура CAP-1D5 клеток в PEM среде показала для всех значений MOI более низкие объемы (приблизительно 2.0 log HA единиц/100 мкл) вирусных частиц. Соответственно, концентрация жизнеспособных клеток культуры CAP-1D5 клеток в PEM среде в течение 48 ч оставалась постоянной 1×106 клеток/мл, а затем упала до уровня менее 1×104 клеток/мл. В отличие от этого, концентрация жизнеспособных клеток постоянно снижалась в культурах CAP-1D5 клеток в 293SFM среде и MDCK.SUS2 клеток с 1×106 клеток/мл до менее чем 1×104 клеток/мл после 96 ч. Во всех культурах наблюдали небольшую задержку репликации вируса при MOI 0.0025, подтвержденную замедленным по времени повышением log HA единиц, по сравнению с более высокими значениями MOI.
Пример 5: Культивирование в объеме 1 л с инфицированием (адаптированный A/PR/8/34)
Клетки CAP-1D5 культивировали в биореакторе емкостью 1 л в PEM среде, с 4 мМ глутамина и 4 мМ пирувата при 85 оборотах в минуту, pH=7.2, и с парциальным давлением кислорода pO2 40% чистого кислорода. Первоначальная концентрация клеток составила 5×105 клеток/мл.
После 114 ч роста, и при концентрации клеток 2.4×106 клеток/мл, CAP-IDS клетки инфицировали вирусом гриппа A/PR/8/34 (адаптированный: в PEM, 4-й проход, 1.78×107 вирусов/мл). Без смены среды, но с добавлением 80 мл PEM среды, а также глутамина и пирувата в финальной концентрации 2 мМ. Значение MOI на уровне 0.025, добавили трипсин в финальной концентрации 1×10-5 Е/мл. В течение 240 часов и в промежутки по 24 часа, определяли концентрацию жизнеспособных клеток, концентрацию мертвых клеток, уровень выживаемости линий клеток, а также значение pH, концентрацию глюкозы, лактозы, глутамина, аммония, глутаминовой кислоты и пирувата в среде. Далее, от времени инфицирования (114 ч), детектировали log HA-единиц/100 мкл и значение TCID50 (Genzel and Reichl, Vaccine production determined - state of the art and future needs in upstream processing in Biotechnology: Animal Cell Biotechnology - Methods and Protocols, Eds., R. Portner, Humana Press Inc., Totowa, NJ, 2007, 457-473; Kalbfuss et al, 2008; Biologicals 36 (3): 145-61).
Результаты представлены на фиг.4. Концентрация жизнеспособных клеток CAP-1D5 клеток первоначально выросла, вплоть до инфицирования вирусом гриппа A/PR/8/34, с 6×105 клеток/мл до 2.4×106 клеток/мл и незначительно снизилась после инфицирования. Уровень выживаемости CAP-1D5 клеток в течение всего периода времени был на уровне от 80 до 90%, а после 240 ч опустилась до 70%. Концентрация пирувата в культуре снижалась в течение 72 ч до нуля, объем пирувата, добавляемого в инфицировании вирусом гриппа, также расходовался в течение 10 ч. Концентрация глутамата повышалась устойчивыми темпами в течение всего времени от примерно 1 мМ до примерно 1.8 мМ. С небольшими вариациями, значение pH культуры в течение наблюдаемого времени от 7.1 до 7.4. Максимальный TCID50 титр достиг 2.4×107 вирусов/мл, максимальный HA титр 2.2 log HA-единиц/100 мкл.
Результаты данного опыта по росту показали, что благодаря введению пирувата не наблюдалось дефицита глюкозы, и, таким образом, поддерживалась концентрация жизнеспособных клеток.
Пример 6: Адаптация вируса к CAP-1D5 клеткам в PEM и 293SFMII средах в пробирках производства фирмы Falcon емкостью 50 мл
Вирусную адаптацию вирусов гриппа A/Brisbane/59/2007 (H1N1-like HGR; IVR-148, NIBSC, Национальный институт биологических стандартов и контроля), B/Florida/4/2006 (NIBSC, Национальный институт биологических стандартов и контроля), свиного гриппа (A/Swine (H1N2) Bakum/1832/00; IDT biologies) и лошадиного гриппа (A/equine 2 (H3N8); A/Newmarket/1/93; NIBSC, Национальный институт биологических стандартов и контроля) осуществили путем инфицирования CAP-1D5 в 4 прохода в пробирке Falcon 50 мл в PEM и 293SFMII среде. До каждого инфицирования, осуществили смену среды. Урожай вируса во время каждого прохода квантифицировали титрованием гемагглютинина (log HA единиц/100 мкл) и Tissue Culture Infectious Dose50 Assay (TCID50 вирусов/мл), (Genzel and Reichl, Vaccine production - state of the art and future needs in upstream processing in Methods in Biotechnology. Animal Cell Biotechnology - Methods and Protocols, Ed R. Portner, Humana Press Inc., Totowa, NJ, 2007, 457-473; Kalbfuss et al, 2008; Biologicals 36 (3): 145-61).
Результаты представлены на фиг.5. Инфицирование CAP-1D5 клеток вирусом гриппа A/Brisbane/59/2007 и B/Florida/4/2006 в PEM и 293SFMII средах вызвало значительное повышение титров вируса после 4 проходов, причем повышение титра вируса в 293SFMII среде было более значительным. Аналогичный рост зафиксирован в титровании значения НА во время инфицирования CAP-1D5 клеток вирусом гриппа A/Brisbane/59/2007 и B/Florida/4/2006, соответственно, в PEM и также в 293SFMII среде. Инфицирование CAP-1D5 клеток вирусом свиного гриппа приводит к повышению титрования значения HA, как в PEM, так и в 293SFMII средах.
Наряду с повышением титра вируса вследствие адаптации, с каждым проходом репликация вируса проходила быстрее и, в конечном итоге, могла быть значительно увеличена. В общем, представляется, что можно достичь небольшого роста титров вирусов в 293SFMII среде, в сравнении с PEM средой.
Пример 7: Опыты по культивированию постоянной линии клеток-амниоцитов при повышенной первоначальной концентрации клеток
Постоянную линию клеток-амниоцитов CAP-1D5 культивировали в 100 мл PEM среды (Invitrogen) при 37°C, 8% CO2 и 185 оборотах в минуту. Первоначальная концентрация клеток составляла 5×105 клеток/мл и 8×105 клеток/мл, соответственно. Дополнительно, добавили пируват в сериях с повышенной первоначальной концентрацией клеток при финальной концентрации 4 мМ и 10 мМ, соответственно, и также прочие аминокислоты.
Во временной точке 0 (= старт культуры), и в каждом случае, промежутками через 24 ч, определяли концентрацию жизнеспособных клеток, концентрацию мертвых клеток, уровень выживаемости линий клеток, а также значение pH, концентрацию глюкозы, лактозы, глутамина, аммония, глутаминовой кислоты и пирувата в среде (система анализа биохимических мультипараметров) (Lohr et al, Vaccine, 2009, 27(36), 4975-4982; Genzel et al, Appl Microbiol Biotechnol, 2010, 88(2):461-75).
Результаты представлены на фиг.6. Повышением первоначальной концентрации клеток от примерно 5×105 клеток/мл культуры в начале культуры до 8×105 клеток/мл культуры постоянной линии клеток-амниоцитов CAP-1D5 в PEM среде, наблюдали дополнительный урожай 1×106 клеток/мл культуры после 90 ч. В обычное время инфицирования (примерно от 96 ч до 120 ч после старта культуры), получили концентрацию клеток 5-6×106 клеток/мл культуры.
Значение pH культуры в типичное время инфицирования (примерно от 96 ч до 120 ч после старта культуры), находилось в достаточно критическом диапазоне от 6.6 до 6.8. Предпочтительно, значение pH при инфицировании должно быть примерно на уровне 7.2-7.4.
Пример 8: Влияние изменений активности трипсина и параметров смены среды и 1:2 разбавления со средой на титр вируса
Данный опыт направлен на исследование влияния различных концентраций трипсина в вирусном инфицировании и смене среды, или 1:2 разбавления со средой на титр вируса.
Постоянную линию клеток-амниоцитов CAP-1D5 культивировали в 100 мл PEM среды (Invitrogen) с 4 мМ пирувата и 4 мМ глутамина при 37°C, 8% CO2 и 185 оборотах в минуту во встряхиваемых колбах. Линию клеток инфицировали во время старта культуры CAP-1D5 клеток адаптированным вирусом гриппа A/PR/8/34 (H1N1, RKI, Институт им. Роберта Коха). Число клеток в культуре составляло во время инфицирования 4.5×106 клеток/мл культуральной среды, без смены среды до инфицирования, и 2.3×106 клеток/мл культуры, с 1:2 разбавлением с PEM средой до инфицирования, и 5×106 клеток/мл, с полной сменой среды до инфицирования. Помимо этого, в культурах без смены среды и культурах с 1:2 разбавлением с PEM средой, использовали трипсин в концентрации 1×10-4 Е/клетку, 3×10-5 Е/клетку, и 5×10-5 Е/клетку. В культурах с полной сменой среды использовали трипсин в концентрации 1×10-4 Е/клетку, 1×10-5 Е/клетку, 5×10-5 Е/клетку и 1×10-6 Е/клетку или не использовали трипсин.
Результаты представлены на фиг.7. Разбавление 1:2 со свежей PEM средой приводит к раннему повышению HA (примерно 12 ч, вместо 24 ч) и более высоким максимальным уровням HA - 2.70 log HA в сравнении с 2.30 log HA культур без смены среды. Полная смена среды привела к повышенным значениям HA, превышавшим 3.0 log HA.
В культурах без смены среды и с 1:2 разбавлением с PEM средой, зафиксировали практически идентичные значения log HA, вне зависимости от концентрации трипсина. В культурах, в которых осуществили полную смену среды, динамика изменения значений log HA была аналогичной при концентрациях трипсина 1×10-5 E/клетку, 5×10-5 E/клетку и 1×10-6 E/клетку. В культуре без трипсина, значение log HA достигло значения только примерно 2 log HA - единиц/100 мкл и в культуре, в которой для инфицирования использовали трипсин 1×104 E/клетку, значение log HA находилось ниже 1 log HA - единиц/100 мкл.
Пример 9: Зависимость MOI-(множественность заражения) от инфицирования CAP-1D5 клеток в PEM среде с различными адаптированными штаммами вируса гриппа, с или без смены среды до инфицирования
Данный опыт направлен на исследование зависимости между значением MOI, т.е. численного соотношения числа вирусных частиц, используемых для инфицирования, и числа CAP-1D5 клеток, подлежащих инфицированию, и титра вируса (в log HA единицах на 100 мкл культуры).
Для этого, постоянную линию клеток-амниоцитов CAP-1D5 культивировали в 50 мл PEM среды (Invitrogen), с 4 мМ пирувата и 4 мМ глутамина при 37°C, 8% CO2 и 185 оборотах в минуту, во встряхиваемых колбах. Зависимость MOI исследовали как со сменой, так и без смены среды культуры до инфицирования. Использовали три адаптированных штамма гриппа: A/PR/8/34 (RKI, Институт им. Роберта Коха), A/Brisbane/59/2007 (IVR-148, NIBSC, Национальный институт биологических стандартов и контроля) и B/Florida/4/2006 (NIBSC, Национальный институт биологических стандартов и контроля). Инфицирование проводили во встряхиваемых колбах емкостью 50 мл с концентрацией клеток при инокуляции 4.9×106 клеток/мл (без смены среды) и 5.0×106 клеток/мл (со сменой среды). Инфицирование осуществили при значениях MOI 0.25 и 0.10 (для вируса гриппа A/Brisbane), 0.025 и 0.0025, без смены среды, и при значениях MOI 0.10 и 0.06 (для вируса гриппа A/Brisbane), 0.025 и 0.0025, со сменой среды. Без смены среды, активность трипсина составляла 1×104 E/клетку, а со сменой среды - 1×106 E/клетку. После этого, определили число вирусных частиц в течение 144 часов (log HA - Единиц/100 мкл) (Kalbfuss et al, 2008; Biologicals 36 (3):145-61).
Результаты представлены на фиг.8. Осуществление смены среды приводит к более стабильным результатам в репликации вируса. Низкую зависимость MOI можно отметить только в клетках, инфицированных вирусом гриппа A/PR/8/34 без смены среды, и в клетках, инфицированных вирусом гриппа A/Brisbane со сменой среды. Со сменой среды, можно достичь значительно более высоких значений log HA. Значения pH без смены среды находились частично в критическом диапазоне от 6.6 до 6.8, а при смене среды в диапазоне от 7.3 до 7.5.
Пример 10: Дополнительное культивирование в объеме 1 л в биореакторах STR и Wave Bioreactors с инфицированием (адаптированный A/PR/8/34)
В первом опыте (B16) CAP-1D5 клетки культивировали в биореакторе STR (Sartorius) емкостью 1 л в PEM среде с 4 мМ глутамина и 4 мМ пирувата при 120 оборотах в минуту, pH=7.2, и с парциальным давлением кислорода pO2 40% чистого кислорода. Первоначальная концентрация клеток составила 5×105 клеток/мл. После 72.75 ч роста, и при концентрации клеток 2.1×106 клеток/мл, CAP-1D5 клетки инфицировали вирусом гриппа A/PR/8/34 (адаптированный: в PEM, 4-й проход, 2.01×106 вирусов/мл). Без смены среды. Значение MOI на уровне 0.025, добавили трипсин в финальной концентрации 3×10-5 E/мл.
Во втором опыте (B26) CAP-1D5 клетки культивировали в биореакторе STR (Sartorius) емкостью 1 л в PEM среде при 120 оборотах в минуту, pH=7.4 до 7.2, и с парциальным давлением кислорода pO2 40% чистого кислорода. Первоначальная концентрация клеток составила 8×105 клеток/мл. После 92 ч роста CAP-1D5 клетки (адаптация: в PEM, 4-й проход, 3.75×106 вирусов/мл) инфицировали вирусом гриппа A/PR/8/34. До этого, осуществили полную смену среды, а значение pH достигло 7.6. Значение MOI на уровне 0.025, добавили трипсин в финальной концентрации 3×10-5 E/мл.
В третьем опыте (Wave) CAP-1D5 клетки культивировали в биореакторе Wave (Wave Biotech) емкостью 1 л в PEM среде с добавлением 4 мМ глутамина, 4 мМ пирувата и 20 мМ глюкозы при частоте вибрации 13 оборотов в минуту, угол 7°, pH=7.3 до 6.9, с парциальным давлением кислорода pO2 40% чистого кислорода и с парциальным давлением CO2 7.5%. Первоначальная концентрация клеток составила 5×105 клеток/мл. После 72 ч роста CAP-1D5 клетки (адаптация: в PEM, 4-й проход, 1.87×106 вирусов/мл) инфицировали вирусом гриппа A/PR/8/34. Концентрация клеток до инфицирования составила 2.1×106 клеток/мл. Без смены среды. Значение MOI на уровне 0.025, добавили трипсин в финальной концентрации 3×10-5 E/мл.
В четвертом опыте MDCK.SUS2 клетки культивировали в биореакторе STR (Sartorius) емкостью 1 л в АЕМ среде. Первоначальная концентрация клеток составила 5×105 клеток/мл. После 118.25 часов роста, MDCK.SUS2 клетки инфицировали вирусом гриппа A/PR/8/34 (Lohr et al., Vaccine, 2010, 28 (38):6256-64).
Результаты представлены на фиг.9. В течение 192 ч, определили концентрацию жизнеспособных клеток и концентрацию мертвых клеток, а также значение pH, концентрацию глюкозы, лактозы, глутамина, аммония, глутаминовой кислоты и пирувата в среде. Помимо этого, начиная с момента инфицирования, определили значения log HA-единиц/100 мкл и TCID50 (Genzel and Reichl, Vaccine production determined - state of the art and future needs in upstream processing in Methods in Biotechnology: Animal Cell Biotechnology - Methods and Protocols, Eds R. Portner; Humana Press Inc., Totowa, NJ, 2007, 457-473; Kalbfuss et al, 2008; Biologicals 36 (3): 145-61).
CAP-1D5 клетки растут быстрее во всех трех опытах и в более высокой плотности, по сравнению с MDCK.SUS2 клетками. Титры вирусов в культурах клеток CAP-1D5 достигают максимального значения примерно 2.5 log HA-единиц/100 мкл. Титр вируса культуры клеток MDCK.SUS2 достигает максимального значения примерно 3 log HA - единиц/100 мкл. Титры вирусов в культурах клеток CAP-1D5 повышаются намного раньше, по сравнению с титром вируса в культуре клеток MDCK.SUS2.
Пример 11: Опыт по культивированию для повышения урожая вируса во встряхиваемых колбах в PEM или 293SFMII средах с полной сменой среды или 1:1 сменой среды до инфицирования
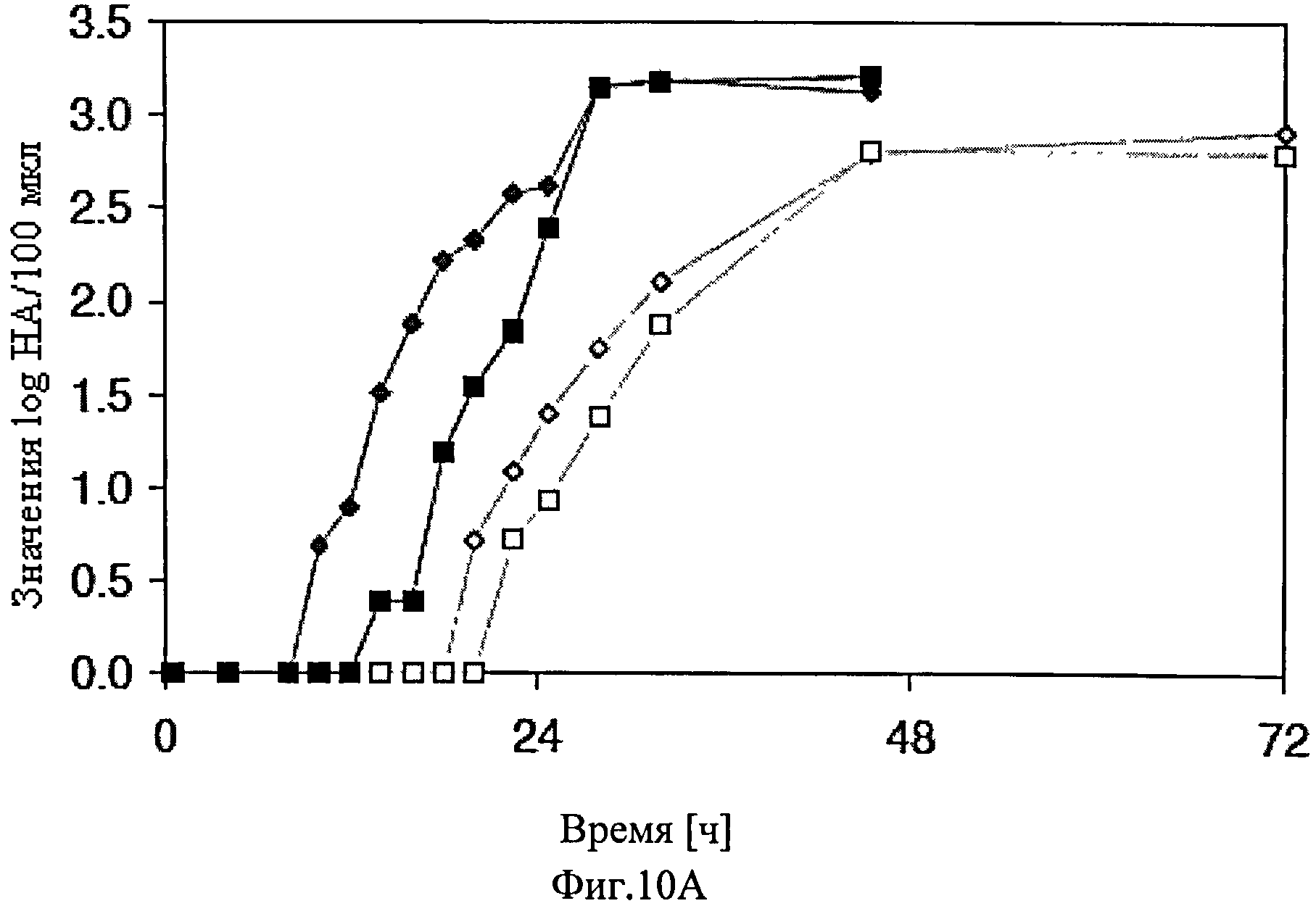
Для оптимизации урожая вируса в культурах клеток CAP-1D5, которые культивируют во встряхиваемых колбах, CAP-1D5 клетки культивировали в средах 293SFMII (Invitrogen) и PEM (Invitrogen) и осуществили смену среды 1:1 или полную смену среды.
Постоянную линию клеток-амниоцитов CAP-1D5 культивировали в 50 мл в PEM среде, с 4 мМ глутамина и 4 мМ пирувата при 37°C, 8% CO2 и 100 оборотах в минуту во встряхиваемых колбах емкостью 100 мл. До инфицирования клеток адаптированным вирусом гриппа A/PR/8/34 при MOI 0.025, осуществили смену среды. При смене среды 1:1, для инфицирования клеток использовали трипсин в концентрации 1×10-5 Е/клетку. При полной смене среды, для инфицирования клеток использовали трипсин в концентрации 1×10-6 E/клетку. Смену среды осуществили в случае использования как PEM среды, так и 293SFMII среды. Концентрация клеток в момент инфицирования составила 5×106 клеток/мл.
Результаты представлены на фиг.10. В течение 72 ч, концентрацию жизнеспособных клеток, уровень выживаемости, значение pH, а также log HA-единиц/100 мкл определили путем титрования гемагглютинина с использованием стандартного метода (Lohr et al., Vaccine, 2009, 27 (36), 4975-4982; Genzel et al, Appl Microbiol Biotechnol, 2010, 88 (2):461-75).
Титр вируса повышался быстрее в тех клеточных культурах, в которых проводили полную смену среды до инфицирования вирусом гриппа A/PR/8/34, по сравнению с клеточными культурами, в которых проводили смену среды 1:1. Помимо этого, титр вируса в клеточных культурах, в которых, до инфицирования вирусом гриппа A/PR/8/34, осуществили полную смену среды, достиг наивысшего максимального значения, по сравнению с теми клеточными культурами, в которых осуществили смену среды 1:1.
Концентрация жизнеспособных клеток в культурах с полной сменой среды до инфицирования упала с 5×106 клеток/мл после 24 ч, и после 48 ч составила примерно 2×104 клеток/мл. В клеточных культурах с PEM средой и сменой среды 1:1 до инфицирования концентрация жизнеспособных клеток снизилась менее резко, и составила после 72 ч примерно 7×105 клеток/мл.
Значение pH во всех клеточных культурах в течение всего периода времени 72 ч составило от 7.6 до 7.2.
Реферат
Настоящее изобретение относится к биотехнологии. Предложен способ получения вакцины на основе вируса гриппа и применение постоянной клетки-амниоцита человека для получения вакцины на основе вируса гриппа. Предложенный способ включает инфицирование постоянной клетки-амниоцита человека вирусом гриппа, последующее культивирование клетки-амниоцита, экспрессию вируса гриппа, отделение полученного вируса гриппа из среды. При этом клетка-амниоцит человека экспрессирует аденовирусные генные продукты E1A и Е1В. Использование постоянных клеток-амниоцитов человека для получения вакцины на основе вируса гриппа позволяет использовать для получения вакцины вирусы гриппа, фенотип которых не отличается от вирусов гриппа дикого типа, что способствует появлению клеточно-опосредованного иммунитета у людей. Предложенный способ получения вакцины и применение постоянной клетки-амниоцита человека для получения вакцины на основе вируса гриппа может быть использовано в медицине для получения эффективных вакцин на основе вируса гриппа. 2 н. и 9 з.п. ф-лы, 36 ил., 1 табл., 11 пр.
Формула
а) инфицирование постоянной клетки-амниоцита человека вирусом гриппа,
б) культивирование постоянной клетки-амниоцита человека,
в) экспрессию вируса гриппа, и
г) изолирование вируса гриппа из среды,
причем постоянная клетка-амниоцит человека экспрессирует аденовирусные генные продукты E1A и E1B.
Комментарии