Способ получения l-аминокислот, штамм escherichia coli - продуцент l-аминокислоты (варианты) - RU2229513C2
Код документа: RU2229513C2
Описание
Область техники
Настоящее изобретение относится к биотехнологии, в частности к способу получения L-аминокислот методом ферментации, и, более конкретно, к гену, полученному из бактерии Escherichia coli. Указанный ген позволяет улучшить продукцию L-аминокислот, например L-фенилаланина и L-треонина.
Предшествующий уровень техники
Традиционно L-аминокислоты в промышленном масштабе могут быть получены методом ферментации с использованием штаммов микроорганизмов, полученных из природных источников, или их мутантов, специально модифицированных для того, чтобы увеличить продукцию L-аминокислот.
Описано множество методов увеличения продукции L-аминокислот, например, путем трансформации микроорганизма рекомбинантной ДНК (см., например, патент США 4278765). Указанные методы основаны на повышении активности ферментов, вовлеченных в биосинтез аминокислот, и/или уменьшении чувствительности целевого фермента к обратному ингибированию продуцируемой L-аминокислотой (см., например, выложенную патентную заявку Японии № 56-18596 (1981), WO 95/16042 или патенты США 5661012 и 6040160).
С другой стороны, повышенная экскреция L-аминокислот может увеличить продуктивность штамма, продуцирующего L-аминокислоту. Описан штамм бактерии, принадлежащей к роду Corynebacterium, обладающей повышенной экспрессией гена экскреции L-лизина (ген lysE) (WO 9723597 A2). Кроме того, описаны гены, кодирующие белки, способные к секреции L-цистеина, L-цистина, N-ацетилсерина или производных триазолидина (патент США 5972633).
К настоящему времени описаны несколько генов, кодирующих, как предполагается, мембранные белки, которые увеличивают продукцию L-аминокислот. Дополнительная копия гена rhtB делает бактерию более устойчивой к L-гомосерину и увеличивает продукцию L-гомосерина, L-треонина, L-аланина, L-валина и L-изолейцина (Европейская патентная заявка ЕР 994190 А2). Дополнительная копия гена rhtC делает бактерию более устойчивой к L-гомосерину и L-треонину и увеличивает продукцию L-гомосерина, L-треонина и L-лейцина (Европейская патентная заявка ЕР 1013765 А1). Дополнительные копии генов yahN, yeaS, yfiK и yggA увеличивают продукцию L-глутаминовой кислоты, L-лизина, L-треонина, L- аланина, L-гистидина, L-пролина, L-аргинина, L-валина и L-изолейцина (Европейская патентная заявка ЕР 1016710 А2).
Ранее авторы настоящего изобретения получили, исходя из E.coli К12, мутант, содержащий мутацию thrR (упоминаемый здесь и далее как rhtA23), придающую устойчивость к высокой концентрации треонина и гомосерина на минимальной питательной среде (Astaurova, О.В. et al., Appl. Biochem. Microbiol., 21,611-616, 1985). Указанная мутация улучшала продукцию L-треонина (Авторское свидетельство СССР № 974817), гомосерина и глутамата (Astaurova, O.B. et al., Appl. Biochem. Microbiol., 27, 556-561, 1991) соответствующими продуцентами - штаммами E.coli.
Более того, авторы настоящего изобретения выяснили, что ген rhtA расположен в положении 18 min на хромосоме E.coli рядом с опероном glnHPQ, кодирующим компоненты системы транспорта глутамина, а также выяснили, что ген rhtA идентичен открытой рамке считывания ybiF, расположенной между генами рехВ и оmрХ. Указанный ген, экспрессирующий белок, кодируемый открытой рамкой считывания, был обозначен как ген rhtA (rht: resistance to homoserine and threonine - устойчивость к гомосерину и треонину).
Кроме того, авторы настоящего изобретения установили, что амплификация гена rhtA также придает устойчивость к гомосерину и треонину. Мутация rhtA23 является заменой А на G положении-1 относительно старт-кодона ATG (ABSTRACTS of 17thInternational Congress of Biochemistry and Molecular Biology in conjugation with 1997 Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francisco, California, August, 24-29, 1997, abstract No.457). Известно, что состав нуклеотидов между последовательностью SD и старт-кодоном, а в особенности последовательность непосредственно перед старт-кодоном, очень сильно влияет на транслируемость мРНК. Было обнаружено 20-кратное изменение уровня экспрессии в зависимости от природы трех нуклеотидов, предшествующих старт-кодону (Gold et al., Annu. Rev. Microbiol., 35, 365-403, 1981; Hui et al., EMBO J., 3, 623-629, 1984). Поэтому было высказано предположение, что мутация rhtA23 увеличивает экспрессию гена rhtA.
Ген rhtA кодирует состояший из 295 аминокислот сильногидрофобный белок, содержащий 10 предполагаемых трансмембранных сегментов. Поиск с помощью программы PSI-BLAST в нуклеотидной последовательности генома Е.соli К-12, принадлежащего к роду Escherichia, выявил по крайней мере 10 белков, гомологичный белку RhtA. Среди них оказались белки, кодируемые генами ydeD и yedA. Ранее было показано, что ген ydeD вовлечен в процесс выброса из клетки веществ - метаболитов биосинтеза цистеина (Daβler et al., Mol. Microbiol., 36, 1101-1112, 2000; патент США 5972663). Ген yedA известен как ген, кодирующий, как предполагается, трансмембранную субъединицу с неизвестной функцией (нуклеотиды с 8037 по 8957 в последовательности с инвентарным номером АЕ 000287 U 00096 в базе данных GenBank).
Описание изобретения
Целью настоящего изобретения является увеличение продуктивности штаммов, продуцирующих L-аминокислоты, и предоставление способа получения L-аминокислот, например L-фенилаланина и L-треонина, с использованием указанных штаммов.
Данная цель была достигнута путем обнаружения того факта, что yedA, кодирующий мембранный белок, гомологичный RhtA, не вовлеченый в путь биосинтеза целевых L-аминокислот, придает микроорганизму устойчивость к нескольким аминокислотам и их аналогам в случае, когда аллель дикого типа этого гена амплифицировался в микроорганизме на многокопийном векторе. Кроме того, ген yedA увеличивает продукцию аминокислоты в случае, когда дополнительные копии указанного гена введены в клетки соответствующего штамма-продуцента. Таким образом было совершено настоящее изобретение.
Настоящее изобретение включает в себя следующее:
1. Бактерия - продуцент L-аминокислоты, принадлежащая к роду Escherichia, в которой продукция L-аминокислоты упомянутой бактерией увеличена за счет повышения активности белков, описанных в пунктах (А) или (В), в клетке упомянутой бактерии:
(A) белок, который представлен аминокислотной последовательностью, приведенной в списке последовательностей под номером 2;
(B) белок, который представлен аминокислотной последовательностью, включающей делеции, замены, вставки или добавление одной или нескольких аминокислот в аминокислотную последовательность, приведенную в списке последовательностей под номером 2, и который обладает активностью, придающей бактерии повышенную устойчивость к L-аминокислотам, таким как фенилаланин, треонин, гомосерин или цистеин, и/или их аналогам, таким как п-фторфенилаланин, 5-фтор-DL-триптофан, S-(2-аминоэтил)цистеин или 4-аза-DL-лейцин (здесь и далее белки, описанные в вышеупомянутых пунктах (А) или (В), упоминаются как "белки согласно настоящему изобретению").
2. Бактерия в соответствии с вышеупомянутой бактерией, в которой активности белков, описанных в пунктах (А) или (В), повышены путем трансформации бактерии с помощью ДНК, кодирующей белки, описанные в пунктах (А) или (В), или путем изменения регуляции экспрессии указанной ДНК в хромосоме упомянутой бактерии.
3. Бактерия в соответствии с вышеупомянутой бактерией, в которой трансформация осуществляется с использованием многокопийного вектора.
4. Способ получения L-аминокислоты, включающий стадии выращивания бактерии в соответствии с вышеупомянутой бактерией в питательной среде и сбора из культуральной жидкости полученной и накопленной в ней L-аминокислоты.
5. Способ в соответствии с вышеупомянутым способом, в котором L-аминокислотой является L-фенилаланин.
6. Способ в соответствии с вышеупомянутым способом, в котором у указанной бактерии повышена экспрессия генов биосинтеза фенилаланина.
7. Способ в соответствии с вышеупомянутым способом, в котором L-аминокислотой является L-треонин.
8. Способ в соответствии с вышеупомянутым способом, в котором у указанной бактерии повышена экспрессия треонинового оперона.
Способ получения L-аминокислоты включает продукцию L-фенилаланина с использованием бактерии - продуцента L-фенилаланина, в которой повышены активности белков согласно настоящему изобретению, например, представленных аминокислотной последовательностью, приведенной под номером 2. Также способ получения L-аминокислоты включает продукцию L-треонина с использованием бактерии - продуцента L-треонина, в которой повышены активности белков согласно настоящему изобретению, например, представленных аминокислотной последовательностью, приведенной под номером 2.
Настоящее изобретение более детально будет описано ниже.
Вышеуказанной бактерией согласно настоящему изобретению является бактерия - продуцент L-аминокислоты, принадлежащая к роду Escherichia, в которой продукция L-аминокислоты указанной бактерией увеличена за счет повышения активностей белков согласно настоящему изобретению в клетке бактерии.
Вышеуказанной бактерией согласно настоящему изобретению является бактерия - продуцент L-аминокислоты, принадлежащая к роду Escherichia и обладающая повышенной активностью белков, которые увеличивают продукцию целевой L-аминокислоты. Конкретно, бактерией согласно настоящему изобретению является бактерия - продуцент L-аминокислоты, принадлежащая к роду Escherichia и обладающая повышенной активностью белков согласно настоящему изобретению. Более конкретно, бактерия согласно настоящему изобретению содержит ДНК, в которой повышена экспрессия гена yedА на хромосоме или на плазмиде в бактерии, и обладает повышенной способностью к продукции L-аминокислоты, например L-фенилаланина или L-треонина.
К белкам согласно настоящему изобретению относятся белки, описанные в следующих пунктах (А) или (В):
(A) белок, который представлен аминокислотной последовательностью, приведенной в списке последовательностей под номером 2;
(B) белок, который представлен аминокислотной последовательностью, включающей делеции, замены, вставки или добавление одной или нескольких аминокислот в аминокислотную последовательность, приведенную в списке последовательностей под номером 2, и который обладает активностью, придающей бактерии устойчивость к аминокислотам, таким как фенилаланин, треонин, гомосерин или цистеин, и/или их аналогам, таким как п-фторфенилаланин, 5-фтор-DL-триптофан, S-(2-аминоэтил)цистеин или 4-аза-DL-лейцин.
Количество “нескольких” аминокислот различается в зависимости от положения и типа аминокислотного остатка в трехмерной структуре белка. Оно может быть от 2 до 30, предпочтительно от 2 до 15, и более предпочтительно от 2 до 5 для белка (А).
Устойчивость к L-аминокислотам и/или их аналогам означает способность бактерии к росту на минимальной питательной среде, содержащей L-аминокислоту и/или ее аналог в концентрации, при которой штамм дикого типа или родительский штамм не может расти, или способность бактерии расти с большей скоростью на питательной среде, содержащей L-аминокислоту и/или ее аналог, чем штамм дикого типа или родительский штамм. Примерами аналогов L-аминокислот являются п-фторфенилаланин, 5-фтор-DL-триптофан, S-(2-аминоэтил)цистеин или 4-aзa-DL-лейцин. Упомянутая выше концентрация L-аминокислоты составляет обычно от 1 до 50 мг/мл, предпочтительно от 30 до 50 мг/мл в случае L-треонина, предпочтительно от 3 до 5 мг/мл в случае L-гомосерина, предпочтительно от 5 до 7 мг/мл в случае серина и цистеина. Упомянутая выше концентрация аналога L-аминокислоты составляет обычно от 0,1 до 1,0 мкг/мл, предпочтительно от 0,2 до 0,5 мкг/мл в случае 5-фтор-DL-триптофана; обычно от 0,1 до 2,0 мг/мл, предпочтительно от 0,5 до 1,0 мг/мл в случае п-фторфенилаланина; обычно от 0,1 до 2,0 мг/мл, предпочтительно от 0,5 до 10 мг/мл в случае 4-аза-DL-лейцина и S-(2-аминоэтил)цистеина.
К бактерии согласно настоящему изобретению также относятся бактерии, в которых активности белков согласно настоящему изобретению повышены путем трансформации бактерии с помощью ДНК, кодирующей белки, описанные в пунктах (А) или (В), или путем изменения регуляции экспрессии последовательности указанной ДНК в хромосоме упомянутой бактерии.
Упомянутая ДНК, использующаяся для модификации бактерии согласно настоящему изобретению, кодирует, как предполагается, белок, обладающий способностью к экскреции L-аминокислот. Более конкретно, ген yedA является такой ДНК. Ген yedA может быть получен, например, с помощью ПЦР с использованием затравок, полученных на основе нуклеотидной последовательности, приведенной под номером 1.
К ДНК согласно настоящему изобретению относится ДНК, кодирующая белок, включающий делеции, замены, вставки или добавление одной или нескольких аминокислот в одно или несколько положений белка (А) при условии, что они не приводят к утрате активности указанного белка. Хотя количество “нескольких” аминокислот различается в зависимости от положения и типа аминокислотного остатка в трехмерной структуре белка, оно может быть от 2 до 30, предпочтительно от 2 до 15, и более предпочтительно от 2 до 5 для белка (А). ДНК, кодирующая практически такой же белок, как белок, описанный в пункте (А), может быть получена, например, путем модификации нуклеотидной последовательности, кодирующей белок, описанный в пункте (А), с использованием сайт-направленного мутагенеза таким образом, что один или несколько аминокислотных остатков будут удалены, заменены, введены или добавлены. Модифицированная подобным образом ДНК может быть получена традиционными методами, использующими обработку химическими реагентами и содержание в условиях, вызывающих мутации. К подобного рода обработкам относятся обработка ДНК, кодирующей белки согласно настоящему изобретению, с помощью гидроксиламина или обработка бактерии, содержащей ДНК, с помощью УФ излучения или химического реагента, такого как N-метил-N’-нитро-N-нитрозогуанидин или азотистая кислота.
К ДНК согласно настоящему изобретению относятся варианты, которые могут быть найдены в различных штаммах или вариантах бактерий, принадлежащих к роду Escherichia, в виду природного разнообразия. ДНК, кодирующая подобные варианты, может быть получена путем выделения ДНК, которая гибридизуется с геном yedA или частью указанного гена в жестких условиях и которая кодирует белок, увеличивающий продукцию L-аминокислот. Термин “жесткие условия”, упомянутый здесь, означает условия, при которых образуются так называемые специфические гибриды, а неспецифические - не образуются. Например, к жестким условиям относятся условия, при которых гибридизуются ДНК, обладающие высокой степенью гомологии, к примеру ДНК, обладающие гомологией не менее 70% друг относительно друга. В качестве варианта примером жестких условий являются условия, соответствующие условиям отмывки при гибридизации по Саузерну, например 60°С, 1×SSC, 0,1% SDS, предпочтительно 0,1×SSC, 0,1% SDS. В качестве зонда для ДНК, кодирующей варианты и гибридизующейся с геном yedA, также может быть использована часть нуклеотидной последовательности под номером 1. Зонд подобного рода может быть получен в результате ПЦР с использованием в качестве затравок олигонуклеотидов, полученных на основе нуклеотидной последовательности под номером 1, и фрагмента ДНК, содержащего нуклеотидную последовательность под номером 1, в качестве матрицы. В случае, когда в качестве зонда используется фрагмент ДНК длиной около 300 пар оснований, условия отмывки при гибридизации соответствуют, например, 50°С, 2×SSC и 0,1% SDS.
Трансформация бактерии с помощью ДНК, кодирующей белок, означает введение указанной ДНК в клетку бактерии, например, с помощью традиционных методов для того, чтобы усилить экспрессию генов, кодирующих белок согласно настоящему изобретению, и повысить активность белка в клетке бактерии.
К методам увеличения экспрессии генов относятся методы увеличения числа копий гена. Введение гена в вектор, способный к функционированию в бактерии, принадлежащей к роду Escherichia, увеличивает число копий указанного гена. Для подобных целей могут быть предпочтительно использованы многокопийные векторы. Примерами многокопийных векторов являются pBR322, pUC19, pBluescript KS+, pACYC177, pACYC184, pAYC32, pMW119, pET22b и подобные им.
Кроме того, усиление экспрессии гена может быть достигнуто путем введения некоторого числа копий гена в бактериальную хромосому, например, методом гомологичной рекомбинации или подобным.
В случае, когда добиваются усиления экспрессии двух или более генов, указанные гены могут быть расположены вместе на одной и той же плазмиде или раздельно на различных плазмидах. Также допустимо, чтобы одни из генов располагались в хромосоме, а другие гены располагались на плазмиде.
С другой стороны, усиление экспрессии генов может быть достигнуто помещением ДНК согласно настоящему изобретению под контроль сильного промотора. Например, в качестве сильных промоторов известны lac промотор, trp промотор, trc промотор, PL промотор фага лямбда. Использование сильного промотора может быть совмещено с увеличением числа копий гена.
Методы получения хромосомной ДНК, гибридизации, ПЦР, получения ДНК плазмид, расщепления и лигирования ДНК, трансформации, отбора олигонуклеотидов в качестве затравок и другие подобные методы являются обычными методами, хорошо известными для специалиста в указанной области техники. Перечисленные методы описаны в Sambrook, J., and Russell D., "Molecular Cloning A Laboratory Manual, Third Edition", Cold Spring Harbor Laboratory Press (2001) или подобных изданиях.
Бактерия согласно настоящему изобретению может быть получена путем введения вышеуказанных ДНК в бактерию, уже обладающую способностью к продукции L-аминокислоты. С другой стороны, бактерия согласно настоящему изобретению может быть получена путем придания бактерии, уже содержащей указанные ДНК, способности к продукции L-аминокислоты.
Примеры бактерий, принадлежащих в роду Escherichia, - продуцентов.
Бактерии - продуценты фенилаланина.
В качестве родительских штаммов, в которых активности белков согласно настоящему изобретению будут повышены, могут быть использованы бактерии, принадлежащие к роду Escherichia, - продуценты L-фенилаланина, такие как штамм AJ12739 (tyrA:Tn10, tyrR) (ВКМП В-8197); штамм HW1089 (АТСС-55371), содержащий ген pheA34 (патент США 5354672); мутантный штамм MWEC101-b (KR8903681); штаммы NRRL В-12141, NRRL B-12145, NRRL В-12146 и NRRL В-12147 (патент США 4407952) и пободные им. Также в качестве родительских штаммов, в которых активности белков согласно настоящему изобретению будут повышены, могут быть использованы бактерии, принадлежащие к роду Escherichia, - продуценты L-фенилаланина, такие как штамм E.coli K-12[W3110(tyrA)/pPHAB] (FERM ВР-3566), штамм E.coli K-12[W3110(tyrA)/pPHAD] (FERM BP-12659), штамм E.coli K-12[W3110(tyrA)/pPHATerm] (FERM BP-12662) и штамм E.coli K-12[W3110(tyrA)/pBR-aroG4, рАСМАВ], названный как AJ12604 (FERM BP-3579) (Европейский патент ЕР 488424 В1).
Бактерии - продуценты треонина.
В качестве родительских штаммов, в которых активности белков согласно настоящему изобретению будут повышены, могут быть использованы бактерии -продуценты L-треонина, принадлежащие к роду Escherichia, такие как MG442 (ВКПМ В-1628) (Гусятинер М.М. и др. Генетика, 14, 947-956, 1978); ВКПМ В-3996 (патент США 6165756); ВКПМ В-5318 (патент США 6132999); FERM ВР-3756 и ВР-4072 (патент США 5376538) и подобные им.
К способам согласно настоящему изобретению относится способ продукции L-аминокислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления указанной L-аминокислоты в питательной среде и сбора L-аминокислоты из культуральной жидкости. Также к способам согласно настоящему изобретению относится способ продукции L-фенилаланина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-фенилаланина в указанной питательной среде и сбора L-фенилаланина из культуральной жидкости. Также к способам согласно настоящему изобретению относится способ продукции L-треонина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-треонина в указанной питательной среде и сбора L-треонина из культуральной жидкости.
Согласно настоящему изобретению выращивание, сбор и очистка L-аминокислоты из культуральной или подобной ей жидкости могут быть осуществлены способом, подобным традиционным способам ферментации, в которых аминокислота продуцируется с использованием микроорганизма. Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, которые требуются микроорганизму для роста. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, и различные органические кислоты. В зависимости от степени ассимиляции используемого микроорганизма могут использоваться спирты, такие как этанол и глицерин. В качестве источника азота могут использоваться различные соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов и ферментолизат микроорганизмов. В качестве минеральных добавок используются монофосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные соединения. Некоторые питательные добавки могут быть, при необходимости, добавлены в питательную среду. Например, если микроорганизму для роста необходим тирозин (ауксотрофия по тирозину), соответствующее количество тирозина может быть добавлено в питательную среду для выращивания.
Выращивание осуществляется предпочтительно в аэробных условиях, таких как перемешивание, ферментация с аэрацией, при температуре от 20 до 40°С, предпочтительно от 30 до 38°С. рН питательной среды находится в пределах от 5 до 9, предпочтительно от 6.5 до 7.2. рН среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферами. Обычно выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем целевая L-аминокислота может быть собрана и очищена методами ионообменной хроматографии, концентрирования и кристаллизации.
Наилучший способ осуществления изобретения
Настоящее изобретение более детально описано со ссылкой на примеры. В указанных примерах аминокислоты являются аминокислотами L-конфигурации, если не указано иное.
Пример 1. Клонирование гена yedA из E.coli.
Полная нуклеотидная последовательность штамма E.coli К-12 определена (Science, 277, 1453-1474, 1997). Поиск с помощью программы PSI-BLAST выявил, что в геноме E.coli К-12 присутствуют по крайней мере 10 паралогов гена rhtA, включая ген yedА. Ген yedА кодирует, как предполагается, трансмембранную субъединицу, функция которой неизвестна.
На основе приведенной нуклеотидной последовательности были синтезированы затравки, приведенные под номерами 3 (затравка 1) и 4 (затравка 2) в Списке последовательностей (см. в конце описания). Затравка 1 комплементарна последовательности с 179 по 153 нуклеотид перед старт-кодоном и содержит сайт узнавания фермента рестрикции BamHI, введенный на ее 5’-конец. Затравка 2 комплементарна последовательности с 53 по 77 нуклеотид после стоп-кодона и содержит сайт узнавания фермента рестрикции SalI, введенный на ее 5’-конец.
Хромосомная ДНК штамма E.coli TG1 была получена стандартным методом. ПЦР осуществлялась на "Perkin Elmer GeneAmp PCR System 2400" в следующих условиях: 40 секунд при 95°С, 40 секунд при 47°С, 40 секунд при 72°С, 30 циклов с использованием Taq-полимеразы (Fermentas). Полученный фрагмент ДНК, содержащий ген yedA с его собственным промотором, был обработан рестриктазами BamHI и SalI и введен в многокопийные векторы pUC19 и pAYCTER3, предварительно обработанные теми же рестриктазами. Таким образом были получены плазмиды pYEDA1 и pYEDA2 соответственно. Вектор pAYCTER3 является производным вектора pAYC32 - очень стабильного вектора с умеренным числом копийности, сконструированного на основе плазмиды RSF1010 (Christoserdov A.Y., Tsygankov Y.D, Broad-host range vectors derived from a RSF1010 Tnl plasmid, Plasmid, 1986, v.16, pp.161-167). Вектор pAYCTER3 получен путем введения полилинкера из pUC19 и сильного терминатора rrnВ в плазмиду pAYC32 вместо ее промотора следующим образом. Сначала полилинкер из плазмиды pUC19 был получен с помощью ПЦР с использованием затравок, приведенных под номерами 5 и 6 в Списке последовательностей. Полученный продукт ПЦР был обработан рестриктазами EcoRI и BglII. Терминатор rrnВ также был получен с помощью ПЦР с использованием затравок, приведенных под номерами 7 и 8. Полученный продукт ПЦР был обработан рестриктазами BglII и BclII. Затем эти два фрагмента ДНК были лигированы в плазмиду pAYC32, предварительно обработанную рестриктазами EcoRI и BclII. Таким образом была получена плазмида pAYCTER3.
Пример 2. Влияние амплификации гена yedA на устойчивость штамма Е.coli TG1 к аминокислотам и их аналогам.
Плазмиды pYEDAl и pYEDA2, а также векторы pUC19 и pAYCTER3 были введены в штамм E.coli TG1. Таким образом были получены штаммы TG1 (pYEDA1), TG1 (pYEDA2), TG1 (pUC19) и TG1 (pAYCTER3).
Затем была определена способность каждого из указанных штаммов расти в присутствии аминокислот и их аналогов на минимальной агаризованной среде М9 с глюкозой, содержащей ступенчато увеличивающиеся концентрации ингибиторов. Чашки были засеяны от 106 до 107 клеток ночной культуры, выращенной на минимальной среде, содержащей 100 мг/мл ампицилина в случае штаммов с плазмидами. Способность к росту определялась после 44 часов инкубации при 37°С. Результаты экспериментов представлены в таблице 1.
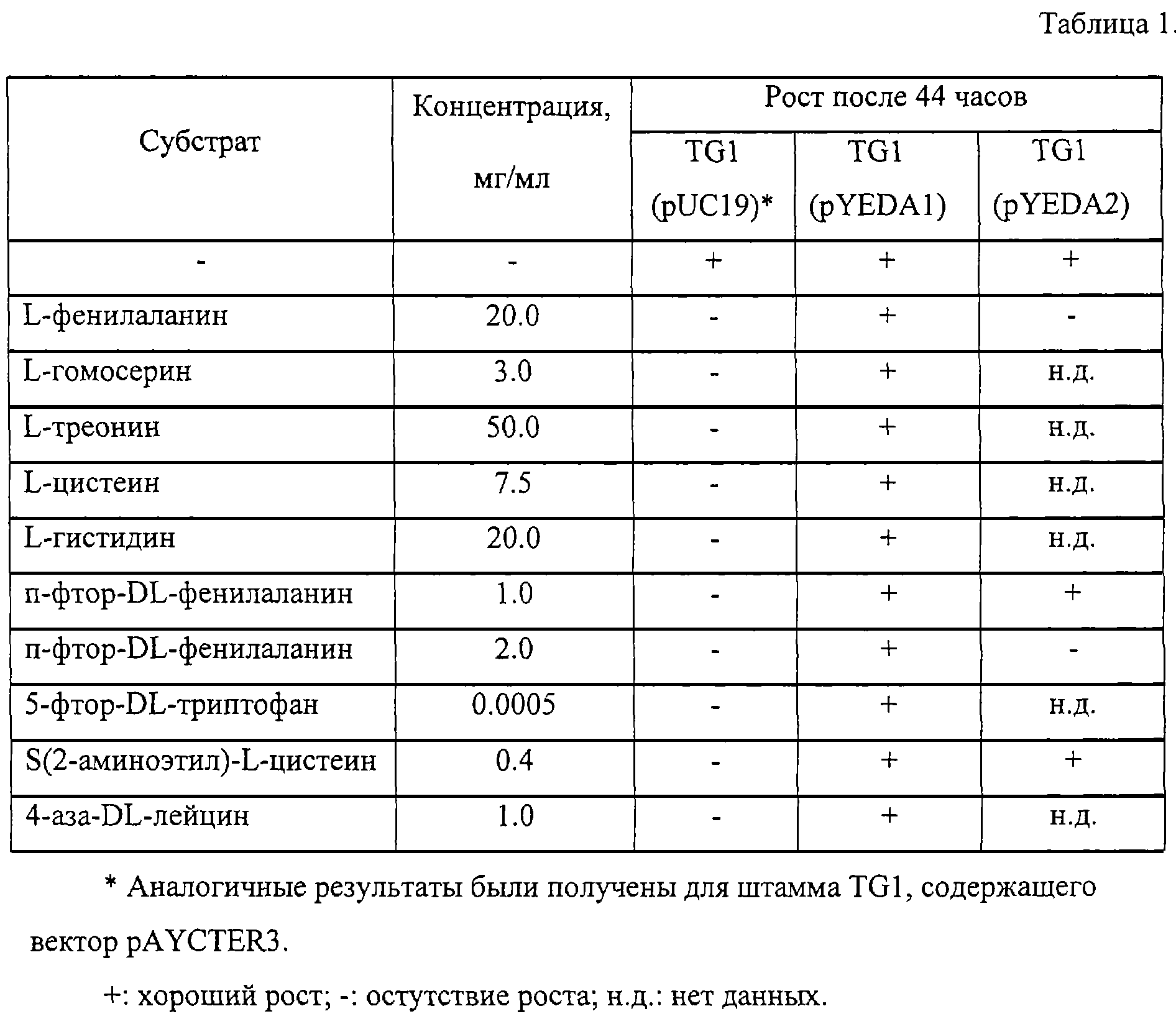
Пример 3. Влияние амплификации гена yedA на продукцию фенилаланина.
Штамм E.coli АJ12739 - продуцент фенилаланина был использован в качестве исходного штамма для трансформации плазмидами, содержащими ген yedA. Штамм АJ12739 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (Россия, 113545, Москва, 1-й Дорожный проезд, 1) 6 ноября 2001 года под инвентарным номером ВКПМ В-8197.
Штамм E.coli АJ12739 - продуцент фенилаланина был трансформирован с помощью плазмиды pYEDA2 или вектора pAYCTER3 с получением штаммов AJ12739/pYEDA2 и AJ 12739/pAYCTER3. Каждый из этих штаммов выращивался при 37°С в течение 18 часов в питательном бульоне, содержащем 100 мг/мл ампицилина; 0,3 мл полученной культуры было перенесено в 3 мл питательной среды для ферментации, содержащей 100 мг/мл ампицилина, в пробирке 20×200 мм и инкубировалось при 37°С в течение 48 часов на роторной качалке. После выращивания количество фенилаланина, накопленное в среде, определялось методом ТСХ. Использовались пластинки для ТСХ размером 10×15 см, покрытые слоем в 0,11 мм силикагелем Sorbfil без флюоресцентного индикатора (Компания “Сорбполимер”, Краснодар, РФ). Пластинки Sorbfil экспонировались с подвижной фазой следующего состава: пропан-2-ол: этилацетат: водный аммиак (25%): вода=40: 40:7:16 (v/v). В качестве реагента для визуализации использовался 2%-ный раствор нингидрина в ацетоне. Результаты представлены в таблице 2.
Состав среды для ферментации (г/л):
Глюкоза 40,0
(NH4)2SO4 16,0
К2НРО4 1,0
MgSО4 7H2О 1,0
FeSО4 7H2O 0,01
MnSО4 5H2O 0,01
Тиамин НСl 0,0002
Дрожжевой экстракт 2,0
Тирозин 0,1
СаСО3 30,0
Глюкоза и сульфат магния стерилизовали раздельно. СаСО3 стерилизовали при 180°С в течение 2 часов. рН поддерживался в районе 7,0. Антибиотик добавляли в питательную среду после стерилизации.

Как видно из таблицы 2, амплификация гена yedA увеличивает продукцию фенилаланина штаммом АJ12739.
Пример 4. Влияние амплификации гена yedA на продукцию треонина.
Известный штамм Е.соli ВНИИГенетика MG442 - продуцент треонина (Гусятинер М.М. и др. Генетика, 14, 947-956, 1978) (депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) в соответствии с Будапештским договором под инвентарным номером ВКПМ В-1628) был трансформирован плазмидой pYEDA1 или вектором pUC19 с получением штаммов MG442/pYEDA1 MG442/pUC19.
Каждый из этих штаммов выращивался при 37°С в течение 18 часов в питательном бульоне, содержащем 100 мг/мл ампицилина; 0,3 мл полученной культуры было перенесено в 3 мл питательной среды для ферментации, содержащей 100 мг/мл ампицилина, в пробирке 20×200 мм и инкубировалось при 37°С в течение 48 часов на роторной качалке. После выращивания количество треонина, накопленное в среде, определялось методом ТСХ. Пластинки Sorbfil экспонировались с подвижной фазой следующего состава: пропан-2-ол-: ацетон: водный аммиак (25%) :вода=25:25:6:7 (v/v). В качестве реагента для визуализации использовался 2%-ный раствор нингидрина в ацетоне. Результаты представлены в таблице 3.
Глюкоза 50,0
(NH4)2SO4 10,0
К2НРО4 1,0
MgSО4 7H2О 0,4
FeSО4 7H2O 0,02
MnSО4 5H2O 0,02
Тиамин НСl 0,0002
Дрожжевой экстракт 1,0
СаСО3 20,0
Глюкоза и сульфат магния стерилизовали раздельно. СаСО3 стерилизовали при 180°С в течение 2 часов. рН поддерживался в районе 7,0. Антибиотик добавляли в питательную среду после стерилизации.

Как видно из таблицы 3, амплификация гена yedA увеличивает продукцию треонина штаммом MG442.



Реферат
Изобретение относится к биотехнологии. L-аминокислоты, такие как L-фенилаланин или L-треонин, получают культивированием бактерии Escherichia coli с увеличенной способностью к продукции L-аминокислот. Затем аминокислоты выделяют из культуральной жидкости. Штаммы бактерии Escherichia coli AJ12739 (pAYCTER-YEDA2) и MG442/pYEDA1 обладают увеличенной способностью к продукции L-фенилаланина и L-треонина соответственно. Изобретение позволяет получать L-аминокислоты с высокой степенью эффективности. 3 с. и 4 з.п.ф-лы, 3 табл.
Комментарии