Семейство пептидов - ингибиторов активности белка reca, блокирующих sos-ответ у бактерий - RU2621862C1
Код документа: RU2621862C1
Чертежи

























































































Описание
Изобретение относится к области биотехнологии, а именно к новым пептидным структурам, обладающим антибактериальными свойствами. В настоящее время одной из главных проблем фармакологии и медицины является способность бактерий адаптироваться к действию применяемых антибиотиков. В результате широкого применения антибиотиков для борьбы с различными инфекциями они постепенно теряют свою эффективность. При этом постоянно увеличивается количество бактериальных штаммов, устойчивых к применяемым антибиотикам (Cassell G.H. Emergent antibiotic resistance: health risks and economic impact // FEMS Immunol Med. Microbiology. 1997, Vol. 18, P. 271-274.). Известно, что такая адаптация бактерий к новым антибиотикам связана с активацией бактериального SOS-ответа (Wu N., et al. Ranking of persister genes in the same Escherichia coli genetic background demonstrates varying importance of individual persister genes in tolerance to different antibiotics // Frontiers in Microbiology. - 2015. - Vol. 6 - No 01003).
Основные классы современных антибиотиков воздействуют на весьма ограниченное количество белков, занятых в таких ключевых клеточных процессах, как биосинтез белков и компонентов клеточных мембран, репликации и репарации ДНК (Cirz R.T., et al Inhibition of mutation and combating the evolution of antibiotic resistance // PLoS Biol. - 2005. - Vol. 3. - P. e176). Известно, что бактериальные клетки адаптируются к воздействию этих антибиотиков с помощью активации клеточного механизма SOS-ответа и связанного с этим повышенного мутагенеза и перестройки бактериального генома (Smith Р.А., Romesberg F.E. Combating bacteria and drag resistance by inhibiting mechanisms of persistence and adaptation // Nat. Chem. Biol. 2007. Vol. 3. P. 549-556). В свою очередь, стратегии преодоления этого сопротивления обычно включают химические модификации существующих антибиотиков.
Одной из потенциальных возможностей выхода из этого тупика является разработка новых способов блокирования SOS-ответа у бактерий с помощью ингибиторов основного белка - инициатора бактериального SOS-ответа - белка RecA. Белок RecA является центральным ферментом гомологической рекомбинации и рекомбинационной репарации ДНК. Структура и функции RecA сохраняют высочайшую консервативность среди патогенных бактерий. Белок RecA необходим для осуществления рекомбинационной репарации ДНК бактерий, а также для восстановления синтеза ДНК в местах вынужденной остановки репликативной вилки. Поэтому белок RecA является хорошей мишенью для конструирования соединений, блокирующих бактериальный SOS-ответ.
В последние годы предпринято несколько попыток поиска ингибиторов белка RecA (Peterson E.J., Janzen W.P., Kireev D., Singleton S.F. High-throughput screening for RecA inhibitors using a transcreener adenosine 5'-O-diphosphate assay // Assay Drug Dev Technol. - 2012. - Vol. 10. - P. 260-268). В частности, были обнаружены несколько химических соединений, которые ингибируют АТФазную активность RecA in vitro, например, природный фенол (куркумин) ингибирует SOS-ответ, индуцируемый антибиотиком левофлоксацином (Bellio P., et al. Curcumin inhibits the SOS response induced by levofloxacin in Escherichia coli // Phytomedicine. - 2014. - Vol. 21. - P. 430-434). Кроме того, в числе таких соединений оказались некоторые аналоги нуклеотидов (Lee A.M., Ross С.Т., Zeng В.В., Singleton S.F. A molecular target for suppression of the evolution of antibiotic resistance: inhibition of the Escherichia coli RecA protein by N(6)-(1-naphthyl)-ADP // J Med Chem. - 2005. - Vol. 48. - P. 5408-5411), малые органические молекулы на основе 2-амино-4,6-диарилпиридина, 1,2,4-оксадиазол, хиназолинон, бензимидазола и диазепинона (Sexton J.Z., et al Novel Inhibitors of E. coli RecA ATPase Activity // Cur. Chem. Genomics 2010, Vol. 4, P. 34-42), полисульфатированные и полисульфанированные нафтил соединения (Nautiyal A., Patil K.N., Muniyappa K. Suramin is a potent and selective inhibitor of Mycobacterium tuberculosis RecA protein and the SOS response: RecA as a potential target for antibacterial drug discovery // J Antimicrob Chemother. - 2014. - Vol. 69. - P. 1834-1843). Однако, все эти соединения недостаточно специфичны, и имеют широкую область побочного действия, поскольку могут связываться не только с бактериальными белками RecA, но и со многими человеческими АТФазами, имеющими сходное с RecA строение центра связывания и гидролиза АТФ и поэтому потенциально токсичны.
В отличие от этих соединений, небольшие пептидные фрагменты, представляющие функциональные участки природных ингибиторов, являются одними из лучших кандидатов для достижения блокирования SOS-ответа у бактерий. Такие соединения содержат только природные аминокислоты, весьма специфичны для их белков мишеней и поэтому не обладают широкой областью побочного действия и нетоксичны для человека.
В качестве наиболее близкого аналога рассмотрена заявка на патент № US 20100292135 А1 от 18.11.2010 «Inhibitors of RecA activities for control of antibiotic-resistant bacterial pathogens» («Ингибиторы активности RecA для контроля анибиотикоустойчивых бактериальных патогенов»; МПК: A61K 31/095; A61K 31/12; A61K 31/538).
Кроме большого набора органических ингибиторов АТФазной активности белков RecA на основе производных АДФ, приведены две пептидные последовательности (INPEP-1 и INPEP-2), построенных по аналогии с аминокислотной последовательностью N-концевого домена RecA, участвующего в образовании межмолекулярного интерфейса белков RecA при образовании ими спирального филамента на онДНК.
К недостаткам этого аналога можно отнести следующее. В заявке US 20100292135 А1 не приведено никаких данных о степени блокирования SOS-ответа у бактерий in vivo, а пептид рассматривается как ингибитор активности белка RecA только in vitro. Используя данные, приведенные в описании к заявке, можно прийти к выводу, что степень блокирования SOS-ответа у бактерий крайне низка. Например, представленные сведения демонстрируют, что пептиды INPEP препятствуют сборке спирального филамента RecA на онДНК, но она все равно будет происходить, но с замедленной скоростью, и на ДНК будет какое-то количество RecA, что служит сигналом для других белков бактерий, участвующих в SOS-ответе, и поэтому он не будет полностью заблокирован. Из фиг. 11а заявки US 20100292135 А1 следует, что величина концентрации полумаксимального ингибирования (IC50) заявленного пептида даже in vitro весьма высока и составляют 35-500 мкМ, а эффективное блокирования бактериального SOS-ответа с помощью этих пептидов является практически не реализуемой. А вариант пептида, где удалось понизить IC50 до ~3 мкМ, бесполезен с точки зрения практического использования, поскольку требуют образования дисульфидных связей, которое в бактериальных клетках, как известно, блокируется высокими концентрациями глутатиона. Это соединение обладает антиоксидантными свойствами, и не позволит окисляться тиоловым группам аминокислотных остатков цистеина в пептиде, образующих дисульфидную связь, и поэтому пептид потеряет необходимую конформационную стабильность и эффект блокирования бактериального SOS-ответа достигнут не будет. Кроме того, необходимо отметить, что для того, чтобы получить достаточно низкие значения IC50 ~3 мкМ, авторы примерно в три раза понизили концентрации белка RecA, по сравнению с их обычными значениями для такого рода измерений IC50. Поэтому реальные значения IC50 для лучшего их пептида INPEP-2, скорее всего ~9 мкМ.
Как и все природные аминокислотные последовательности глобулярных белков в случае их использования в виде мономерных пептидов, они не обладают достаточной конформационной стабильностью, что и объясняет их низкую эффективность в качестве ингибиторов тех или иных биохимических реакций. Это объясняется тем, что ингибиторы связываются и действует на свои мишени только в определенной биологически активной конформации. В данном случае в конформации при которой боковые группы пептида, взаимодействующих с RecA аминокислот, имеют правильное взаимное расположение в пространстве для связывания с RecA и препятствованию формирования его филамента. Чем выше стабильность биологически активной конформации пептида, тем выше концентрация готового к взаимодействию с RecA пептида и соответственно выше удельная эффективность блокирования SOS-ответа.
Таким образом, в заявке US 20100292135 А1 не удалось получить достаточно хорошие величины IC50 с использованием нескольких традиционных способов конформационной стабилизации пептидов, таких как: дополнительных солевых мостиков, взаимодействий заряженных групп с макродиполем альфа-спирали, использованием аминокислот с высокой склонностью к образованию альфа-спиралей и бета-структур, а также так называемых "кэпинг" взаимодействий. Именно поэтому для конформационной стабилизации пептидов INPEP авторам пришлось использовать искусственные дисульфидные мостики, которые, однако, совершенно бесполезны в условиях бактериальных клеток.
Задачей данного изобретения является создание семейства пептидов для практического применения, обладающих ингибирующей активностью против бактериальных белков RecA, обеспечивающих высокую эффективность блокировки SOS-ответа у бактерий и нетоксичных для организма человека.
Техническим результатом является повышение эффективности блокирования SOS-ответа у бактерий, за счет повышенной конформационной стабильности.
Сущность изобретения заключается в том, что предлагается семейство пептидов (см. таблицу 1), общей формулы Ac-Glu-Glu-Glu-Glu-Lys-AAA-Lys-Ile-HAA-Arg-Tyr-AAA-AAA-Tyr-Arg-HAA-HAA-HAA-NH2, где Ас - N-концевая ацетильная группа, AAA- одна из алифатических аминокислот из набора Leu, Ile, Val, НАА - одна из гидрофобных аминокислот из набора Leu, Ile, Val, Phe и Tyr, a NH2 - C-концевая амидная группа, обладающих ингибирующей активностью против бактериальных белков RecA, а также свойством блокирования SOS-ответа у бактерий.
В отличие от аналога, обладающего недостаточной эффективностью для практического применения, заявляемый нами пептиды имеют совершенно другую длину, аминокислотную последовательность, структуру и механизм действия. Пептиды заявляемого семейства обладают такой же способностью, как и природный белок RecX, быстро разбирать уже собранные фрагменты филаментов белков RecA на онДНК, что доказано в заявляемом изобретении экспериментально, и, таким образом, полностью исключать их появление в клетках в достаточном количестве для активации бактериального SOS-ответа и, соответственно, начала процесса адаптации бактерий к действию антибиотиков.
Изобретение относится к направлению создания препаратов на основе конформационно стабильных пептидов, состоящих только из 20 природных аминокислот, построенных на основе модифицированного фрагмента белка RecX, природного ингибитора белков RecA. Поскольку регуляторный белок RecX довольно нестабилен in vitro и в чистом состоянии он быстро теряет свою активность, его использование в качестве антибиотического лекарственного средства, к сожалению, практически невозможно. Кроме того его аминокислотная последовательность имеет длину 166 остатков и поэтому его твердофазный синтез в чистом виде, в отличие от коротких пептидов, слишком трудоемок и дорог. В заявляемом изобретении используется только короткий фрагмент на основе модифицированной последовательности белка RecX, который непосредственно вовлечен во взаимодействия с комплексом белка RecA и онДНК.
Короткие аминокислотные последовательности природных альфа-спиралей обычно не обладают достаточной конформационной стабильностью. Поэтому для конструирования пептидов с высокой конформационной стабильностью была использована программа SEQOPT (свидетельство о государственной регистрации программы для ЭВМ №2013614525 от 14.05.2013) (Yakimov A., Rychkov G., Petukhov М. De novo design of stable alpha-helices // Methods. Mol. Biol. 2014. Vol. 1216. P. 1-14).
Для решения поставленной задачи за основу был взят участок белка RecX и его аминокислотная последовательность была оптимизирована. В результате получено семейство пептидов, которое не встречается в природе, и с достаточно низким значением IC50 ингибирует активность белка RecA, что в свою очередь приводит к блокированию SOS-ответа у бактерий.
На фиг. 1 приведены результаты глобальной оптимизации аминокислотной последовательности альфа-спирали из белка RecX. № aa/RecX - номера и имена остатков белка RecX из E. coli. 4Е1 - аминокислотная последовательность пептида 4Е1 (SeqNo 1), выбранного для экспериментальной проверки заявляемых свойств.
На фиг. 2 представлена сравнительная динамика гидролиза АТФ белком RecA в присутствии и отсутствии белка RecX и пептида 4Е1.
На фиг. 3 показана динамика изменения длины ДНК при формировании филаментов RecA и последующей разборке филаментов в результате добавления пептида 4Е1.
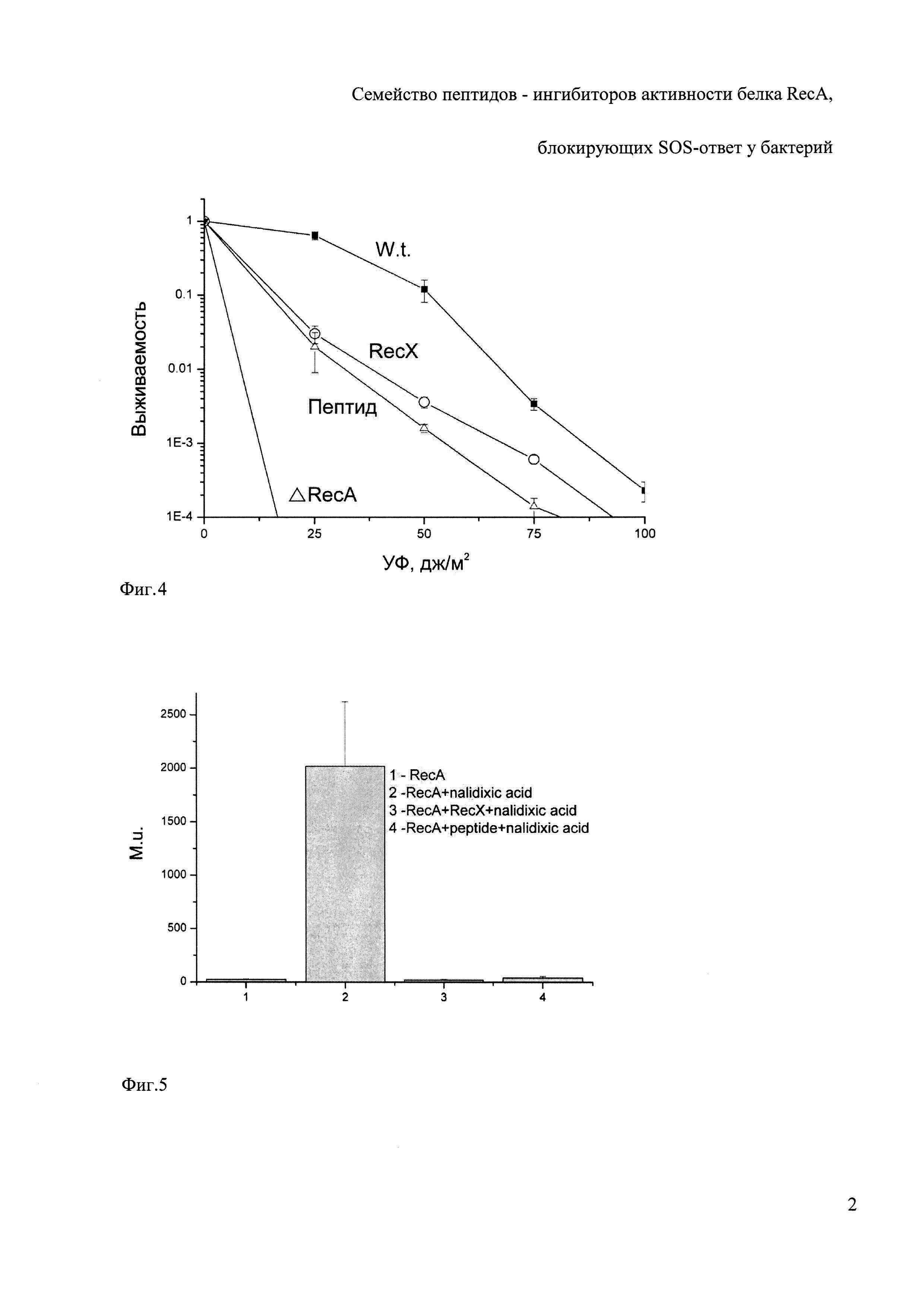
На фиг. 4 представлены данные по изучению зависимости выживаемости клеток E. coli K-12 от дозы УФ-излучения.
На фиг. 5 показана величина SOS-ответа при воздействии на бактериальные клетки E. coli GY6871 налидиксовой кислотой.
По результатам оптимизации аминокислотных последовательностей программой SEQOPT, для экспериментальной проверки был выбран пептид 4Е1 (Seq No 1), обладающий не менее чем 90% конформационной стабильностью и достаточно хорошей растворимостью в воде. Также для проведения экспериментов в реакции ингибирования АТФазной активности использовался пептид с аминокислотной последовательностью, соответствующий фрагменту белка RecX, без модификаций и оптимизации последовательности, с расчетной конформационной стабильностью составляющей 16%. Для немодифицированного фрагмента белка RecX не было обнаружено никакого влияния на белок RecA.
Экспериментальные результаты, представленные на фиг. 2-5, получены для пептида 4Е1. Общая структура этого пептида приведена на фиг. 1. Однако основываясь на хорошо экспериментально проверенной теоретической модели AGADIR (Munoz V., Serrano L. Elucidating the folding problem of helical peptides using empirical parameters // Nat. Struct. Biol. 1994, V. 1 (6), P. 399-409), описывающей конформационные переходы альфа-спираль - случайный клубок в мономерных пептидах, любой пептид из семейства высоко гомологичных пептидов, имеющих общую формулу Ac-Glu-Glu-Glu-Glu-Lys-AAA-Lys-Ile-HAA-Arg-Tyr-AAA-AAA-Tyr-Arg-HAA-HAA-HAA-NH2, где Ас - N-концевая ацетильная группа, AAA- - одна из алифатических аминокислот из набора Leu, Ile, Val, НАА - одна из гидрофобных аминокислот из набора Leu, Ile, Val, Phe и Tyr, a NH2 - C-концевая амидная группа, будет обладать похожей биологической активностью по ингибированию бактериальных белков RecA и блокированию бактериального SOS-ответа.
Пептиды синтезированы стандартными методами с помощью твердофазного синтезатора пептидов с использованием 5-кратного избытка L-аминокислот, имеющих защитные группы Fmoc ((9-фторенил)-метоксикарбонил) (фирма Novabiochem), и реагентов FastMoc (гексафторфосфат/гидроксибензотриазол 2-(1Н-бензотриазол-1-ил)-1,1,3,3-тетраметилурония (ГБТУ/ГОБТ) (фирма Advanced Chemical Europe) в качестве агента для осуществления реакции образования пептидной связи. После отщепления от смолы и удаления защитных групп осуществляли обессоливание путем гель-фильтрации, с помощью системы для жидкостной хроматографии FPLC типа AktaPurifier 10 (фирма GE Healthcare, Великобритания), с использованием в качестве элюента 0,1%-ного водного раствора ТФК (трифторуксусная кислота). Окончательную очистку с помощью препаративной хроматографии с обращенной фазой осуществляли на колонке типа С18, присоединенной к системе для UFPLC типа Shimadzu LC-20. Пептиды элюировали с использованием линейного градиента ацетонитрила в 0,1%-ном водном растворе ТФК и наблюдение за процессом элюирования осуществляли при длине волны 220-230 нм. Массы молекул были подтверждены масс-спектрометрией ионно-циклотронного резонанса с Фурье преобразованием (Varian FT-ICR MS 9.4Т MALDI) с матрично-активированной лазерной десорбцией/ионизацией. Содержание основного вещества, по данным оптической плотности, не менее 95%. Аминокислотный состав пептидов соответствовал приведенным на фиг. 1.
Для in vivo экспериментов использованы следующие штаммы E. coli K-12: AB1157 (F-recA+ara14 galK2 his_4 proA2 argE3 thil tsx33 thr1 leu6lacY1 mtl1 xyl5 rpsl31 supE37), штамм GY6871 sfiA::lacZ из лаборатории P. Деворе. В экспериментах использованы плазмиды precXEc и pPeptid, несущие нормальные аллели генов recX Е. Coli, пептид 4Е1 соответственно. Плазмида precXEc (pEAW224) любезно предоставлена проф. М. Кокс (Университет Висконсин-Мэдисон, США). Для индукции экспрессии гена recX с промотора гена lac проводили инкубацию с изопропил-β-D-тиогалактопиранозидом (IPTG) при концентрации 1 мМ в течение 2 ч перед УФ облучением. Для индукции использовали двуплазмидную систему, включающую плазмиду рТ7, которая несет ген Т7 РНК полимеразы. Ген, несущий аминокислотную последовательность пептида 4Е1, был синтезирован и клонирован в плазмиду pET21b.
Экспериментальная проверка и доказательство работоспособности изобретения проводилась при помощи двух in vitro (в пробирке) и двух in vitro (на живых бактериях) экспериментах, полностью подтвердивших заявляемые свойства. Эксперименты 1 и 2 показали влияние на биохимические реакции, проводимые белком RecA и ингибирование его активности. Эксперименты 3 и 4 показали что заявляемое изобретение оказывает влияние на бактерии E. coli и приводит к наблюдаемому снижению выживаемости при воздействии ультрафиолетового (УФ) излучения, и отсутствию активации SOS-ответа.
Эксперимент 1. Влияние пептида 4Е1 на реакцию АТФ-гидролиза белка RecA in vitro; (фиг. 2)
Известно, что белок RecA является ДНК зависимой АТФазой. Поэтому блокирование его АТФазной активности пептидом 4Е1 в присутствии онДНК было доказано экспериментально. Степень гидролиза АТФ, катализируемого белком RecA, измеряли с помощью спектрофотометрического теста, в котором уровень образования АДФ сопряжен с окислением NADH в NAD+ (Shan Q., Bork J.M., Webb B.L., Inman R.B., Cox M.M. RecA protein filaments: end-dependent dissociation from ssDNA and stabilization by RecO and RecR proteins // J Mol Biol. - 1997. - Vol. 265. - P. 519-540). Концентрацию NADH определяли по оптической плотности реакционной смеси при λ=380 нм с использованием коэффициента экстинкции 1.21 мМ-1 см-1. Реакцию проводили при 37°C в стандартном буфере TMD (25 мМ Трис-HCl (pH 7.5), 10 мМ MgCl2, 1 мМ дитиотреитол), содержащем 2 мМ АТФ, АТФ-регенерирующую систему (4 мМ фосфоенолпирувата и 30 ед/мл пируваткиназы), а также NADH и 30 ед/мл лактатдегидрогеназы. Реакционная смесь содержала 5 мкМ поли(dT) и 3 мкМ RecA. Уровень гидролиза определяли с использованием спектрофотометра Cary 5000 и кюветы с длиной оптического пути 1 см. Концентрацию белков RecA дикого типа определяли по поглощению при 280 нм с использованием коэффициента экстинкции 2.23×104 М-1 см-1. Концентрацию белка RecX и пептида 4Е1 определяли при λ=280 нм с использованием коэффициентов молярной экстинкции 2.57×104 М-1 см-1.
Количество белка RecA, находящегося в состоянии филамента на онДНК, обычно оценивают по скорости гидролиза АТФ. Соответственно, изменение скорости гидролиза АТФ отражает не только количество RecA, которое перешло в свободное или, наоборот, связанное состояние, но и количество онДНК в филаменте. Использование поли(dT), лишенной сложной вторичной структуры, позволило нам вовлечь в реакцию RecA и RecX в отсутствии SSB.
Добавление RecX либо пептида 4Е1 в реакционную смесь, содержащую поли(dT), приводит к одномоментному резкому снижению скорости гидролиза до некоторого постоянного значения. Добавление RecX указано на фиг. 2 стрелкой, в точке 10 мин по оси времени. Эффективность ингибирования активности RecA пептидом 4Е1 примерно в 10-20 раз меньше, чем в случае с белком RecX, если считать по молекулярной концентрации. Вместе с тем, в отличие от белка RecX, который связывает сразу три мономера RecA, пептид 4Е1 взаимодействует лишь с одним мономером. Таким образом, различие в эффективности ингибирования активности белка RecA пептидом 4Е1 по сравнению с полнофункциональным белком RecX различается менее чем в 10 раз.
Таким образом, пептид 4Е1 ингибирует АТФазную активность белка RecA, что является необходимым для блокирования бактериального SOS-ответа.
Эксперимент 2. Влияние пептида 4Е1 на стабильность филамента белка RecA на днДНК in vitro; (фиг. 3)
Известно, что белок RecA инициирует бактериальный SOS-ответ только после образования им спирального филамента на онДНК. Поэтому блокирование пептидом 4Е1 этой реакции было доказано экспериментально. Исследование влияния пептида 4Е1 на стабильность филаментов RecA проводилось на уровне индивидуальных молекул ДНК при помощи метода оптического захвата. В ходе эксперимента молекула ДНК, биотинилированная по обоим концам, фиксировалась между двух полистироловых микросфер диаметром 2,13 мкм, покрытых стрептавидином. Манипуляция ДНК осуществлялась за счет оптического захвата микросфер при помощи двух оптических ловушек. Эксперименты проводились на уникальной установке «Лазерный пинцет» (СПбПУ), которая позволяет исследовать динамику изменения длины ДНК в реальном времени при поддержании постоянной силы натяжения ДНК (Pobegalov G., et al Deinococcus radiodurans RecA nucleoprotein filaments characterized at the single-molecule level with optical tweezers // Biochemical and Biophysical Research Communications. 2015, V. 466(3), P. 426-430.).
На фиг. 3 представлена полученная экспериментальная зависимость изменения длины молекулы ДНК в присутствии RecA, до и после добавления пептида 4Е1. Так как связывание RecA приводит к увеличению длины ДНК на величину порядка 50% по сравнению с В-формой спирали, то в ходе построения филаментов RecA на ДНК происходит постепенное увеличение ее длины. Соответственно, диссоциация RecA приводит к сокращению длины ДНК до исходной. Таким образом, наблюдение за длиной молекулы ДНК при постоянной растягивающей силе позволяет отслеживать динамику построения и разборки филаментов RecA, которая свидетельствует о скорости ингибирования RecA пептидом 4Е1.
В эксперименте была использована ДНК бактериофага λ, длина которой составляет порядка 16 мкм (48502 пары оснований). ДНК помещали в раствор, содержащий Трис-HCl 10 мМ, MgCl2 1 мМ, RecA 1 мкМ и АТФ 1 мМ, в результате чего, при приложении силы растяжения в 20 пН, наблюдалось увеличение длины ДНК, что соответствует связыванию RecAc ДНК. На 490-й секунде в реакцию был добавлен пептид 4Е1 в концентрации 10 мкМ. В присутствии данного пептида рост филаментов RecA прекращался. При этом наблюдалось постепенное уменьшение длины ДНК до исходной, что свидетельствует о том, что исследуемый пептид 4Е1 препятствует связыванию RecA с ДНК и стимулирует разборку филамента RecA на ДНК. Данное наблюдение указывает на то, что пептид 4Е1 способен ингибировать активность RecA на стадии формирования пресинаптического комплекса.
Таким образом, пептид 4Е1 приводит к разборке филамента белка RecA на ДНК, что является необходимым для блокирования бактериального SOS-ответа.
Эксперимент 3. Влияние пептида 4Е1 на выживаемость клеток E. coli при УФ облучении in vivo; (Фиг. 4).
Известно, что блокирование активности белков RecA в клетках Е. coli резко снижает их способность выживать при действии высоких доз ультрафиолетового облучения. Способность пептида 4Е1 блокировать действие белков RecA in vivo доказано экспериментально. Клетки Е. coli, несущие плазмиды с геном recX и аминокислотной последовательности пептида 4Е1, выращивали в среде LB до OD600, равной 0.4-0.6, добавляли IPTG до конечной концентрации 0.3 мМ и растили в течение еще 1.5 ч для индукции синтеза белка RecX либо пептида 4Е1. Контрольные клетки Е. coli также выращивали в среде LB, но без индукции. Клетки, после достижения OD600, равной 0.7-0.9, осаждали центрифугированием (5000 об/мин, 10 мин) и ресуспендировали в исходном объеме солевой среды (NaCl 0.9%). УФ-облучение (λ=254) проводили в чашках Петри на приборе "Хроматоскоп" при мощности дозы 100 мДж/м /с, отбирая аликвоты культуры объемом 0.1 мл через равные временные интервалы (2 с). Облученные клетки разбавляли солевой средой, рассевали на LB-агар и через 24 ч подсчитывали число колоний. УФ облучение клеток, их разведение, рассев и выращивание проводили при освещении красным светом для избегания фотореактивации. С каждой культурой проводили по три независимых опыта. Для оценки погрешностей экспериментальных измерений средняя квадратичное отклонение результата серии измерений умножали на коэффициент Стьюдента для заданной надежности (0.9) и находили границы соответствующего доверительного интервала, т.е. погрешность измерений (Кассандрова О.Н., Лебедев В.В. Обработка результатов наблюдений: «Наука», Москва, 1979). Погрешность измерения во всех случаях не превышала 10%.
Экспрессия RecX приводит к падению выживаемости клеток E. coli. Данный результат был продемонстрирован ранее и свидетельствует о том, что белок RecX является ингибитором in vivo (Stohl Е.А., et al Escherichia coli RecX inhibits RecA recombinase and coprotease activities in vitro and in vivo // J Biol Chem. - 2003. - Vol. 278. - P. 2278-2285). Вместе с тем ингибирование не является максимальным в сравнении со штаммом, где отсутствует ген RecA (дельта RecA). Кривая падения выживаемости клеток, несущих ген пептида, подобна кривой падения выживаемости клеток с RecX. Представленные на фиг. 4 экспериментальные данные доказывают, что экспрессия пептида 4Е1 приводит к уменьшению выживаемости при УФ облучении столь же эффективно как при экспрессии RecX.
Таким образом, пептид 4Е1 приводит к снижению выживаемости бактерий, что показывает отсутствие активности белка RecA на ДНК, что является необходимым условием для блокирования SOS-ответа у бактерий.
Эксперимент 4. Влияние пептида 4Е1 на бактериальный SOS-ответ в клетках E. coli in vivo, (фиг. 5)
Поскольку в живых клетках в принципе могут быть и альтернативные RecA пути активации SOS-ответа, способность пептида 4Е1 блокировать бактериальный SOS-ответ in vivo доказана экспериментально с помощью стандартного метода Миллера. Известно, что единицы Миллера, в которых стандартно измеряется величина SOS-ответа, пропорциональны количеству увеличения О-нитрофенола в минуту на бактерию при воздействии на бактериальные клетки налидиксовой кислоты.
Ген бета-галактозидазы встроен под промотор sfiA таким образом, что о силе SOS-ответа можно судить по количеству выработанной бета-галактозидазы, при SOS-ответе вызванным налидиксовой кислотой. Промотор sfiA (sulA) гена является наиболее чувствительным промотором в ходе индуцируемого SOS-ответа. Экспрессия белков с помощью этого промотора в зависимости от дозы и времени воздействия может увеличиваться до 100 раз (McCool J.D., et al Measurement of SOS expression in individual Escherichia coli K-12 cells using fluorescence microscopy // Mol Microbiol. - 2004. - Vol. 53.
- P. 1343-1357; Cole S. Characterisation of the promoter for the LexA regulated sulA gene of Escherichia coli // Mol. Gen. Genet. 1983, Vol. 189, P. 400-404). В норме при выбранных нами условиях RecA вызывает SOS-ответ примерно до 2000 единиц Миллера, то есть увеличивает в 80-100 раз относительно его базовых значений (измерение №2, фиг. 5). Однако экспрессия белка RecX либо пептида 4Е1 подавляет активность RecA и приводит к ингибированию SOS-ответа почти до базовых значений (измерение №3, 4) Из этого следует, что пептид 4Е1 подавляет SOS-ответ, а вместе с ним и экспрессию всех генов SOS-ответа, включая полимеразу 5. Полимераза 5 является причиной множественного мутагенеза, то есть образует мутации, которые в свою очередь являются материалом для селективного отбора новых генов и последующей адаптации бактерии к антибиотикам.
Активность белка рассчитывалась в условных единицах в соответствии со стандартными рекомендациями (Миллер Д. Эксперименты в молекулярной генетике: «Мир», Москва, 1976): β-gal=TOOO(OD420/OD600tν), где OD420 - оптическая плотность реакционной смеси при 420 нм, OD600 - оптическая плотность клеточной суспензии в Z-буфере при длине волны 600 нм, t - время инкубации, a ν - объем пробы в миллилитрах. Из ночной культуры, выращенной в LB с ампициллином 100 мкГ/мл и канамицином 40 мкГ/мл или без них (для контрольного штамма), 100 мкл вносили в 4 мл такой же среды. Выращивали 50-60 минут и добавляли IPTG до конечной концентрации 1 мМ и проводили индукцию в течение 90 минут. Затем пробы делили пополам и к одной из них добавляли налидиксовую кислоту до конечной концентрации 40 мкГ7/мл. Индуцировали 60 минут, 1 мл клеток смешивали с 1 мл Z-буфера и измеряли OD600. Z-буфер (pH 7.0) На 1 л: Na2HPO4 × 7H2O 16.1 г NaH2PO4 × H2O 5.5 г KCl 0.75 г MgSO4 × 7H2O 0.246 г
Таким образом, пептид 4Е1 приводит блокированию SOS-ответа у бактерий.
Таким образом, приведенные выше доказательства свидетельствуют о том, что подмножество заявляемого семейства пептидов - пептид 4Е1 отличается от известных пептидов новыми свойствами: ингибирующей активностью против бактериальных белков RecA, а также свойством блокирования SOS-ответа у бактерий. При этом обладает соизмеримыми с белком RecX активностями: 1) ингибировать АТФазную активность RecA; 2) инициировать разборку филамента белка RecA на онДНК; 3) понижать уровень выживаемости бактериальных клеток при облучении УФ излучением 4) блокировать систему SOS-ответа бактериальных клеток. Все это позволяет считать пептид 4Е1 и заявляемое семейство пептидов заявляемой структуры хорошими кандидатами для разработки лекарственных средств, предотвращающих адаптацию бактерий к новым антибиотикам.
Реферат
Изобретение относится к области биотехнологии, конкретно к новым пептидным структурам, обладающим антибактериальными свойствами, и может быть использовано в медицине. Заявляется семейство пептидов, обладающих ингибирующей активностью против бактериальных белков RecA, а также свойством блокирования SOS-ответа у бактерий. В отличие от известных аналогов предложенные пептиды обладают большей конформационной стабильностью и способны ингибировать активность белка RecA с достаточно низким IC. 4 пр., 1 табл., 5 ил.
Комментарии