Гибридный белок, гибридный ген, способ получения гибридного белка, вакцина против вирусных заболеваний верхних дыхательных путей, бакуловирусный вектор экспрессии (варианты) - RU2123047C1
Код документа: RU2123047C1
Чертежи

































Описание
Настоящее изобретение относится к конструированию и экспрессии мультимерных гибридных генов, содержащих генные последовательности, кодирующие иммуногенные белки или фрагменты белков от многочисленных патогенов.
Предшествующий уровень техники.
Преимуществом технологии, разработанной авторами настоящего изобретения, является получение единых иммуногенов, содержащих протективные антигены от ряда патогенов. Указанные химеры в значительной степени упрощают разработку комбинированных вакцин, в частности продуцирование поливалентных вакцин одноразового применения. В настоящее время поливалентные вакцины изготавливают путем независимого продуцирования патогенов и/или соответствующих антигенов с последующим их объединением в различных препаратах. Такой метод изготовления вакцин является трудоемким, дорогостоящим и требует применения сложных технологий. В противоположность этому возможность получения одного единственного иммуногена, способного обеспечить защиту от целого ряда заболеваний, позволила бы решить множество проблем, связанных с изготовлением поливалентной вакцины. Например, несколько химерных иммуногенов, полученных в соответствии с настоящим изобретением, могут быть объединены друг с другом, а это позволило бы снизить число отдельных антигенов, необходимых для продуцирования поливалентной вакцины.
Вирус парагриппа человека типов 1, 2, 3 и респираторно-синцитиальный вирус типов A и B являются основными вирусными патогенами, ответственными за возникновения тяжелых инфекций дыхательных путей у детей грудного и младшего возраста. По оценкам специалистов только в США RSV-инфекциями с тяжелым клиническим течением ежегодно заболевают около 1,6 миллионов детей в возрасте до 1 года, и кроме того, около 1,4 миллионов детей в этом возрасте ежегодно заболевают парагриппом-3. В США от осложнений, возникающих в результате тяжелых поражений дыхательных путей, вызываемых вирусами RSV и PIV-3, ежегодно умирает около 4000 детей в возрасте до 1 года. По данным консультативных комиссий при BOC (WHO-Всемирная организация здравоохранения) и NIALD (Национальный институт артрита и инфекционных заболеваний, США) исследования по разработке вакцин против RSV занимают второе место после работы по ВИЧ-вакцинации, а исследования по получению эффективных вакцин против PIV-3 находятся в числе десяти приоритетных областей деятельности по вакцинации.
В настоящее время не существует надежных и эффективных вакцин против вирусных инфекций, которые можно было бы использовать для детей младшего возраста, а поэтому совершенно очевидно, что разработка таких вакцин является крайне необходимой. Клинические испытания показали, что инактивированные формальдегидом и живые, но ослабленные вирусные вакцины не дают адекватной защиты против указанных инфекций. Фактически, у детей, которым была введена формалин-инактивированная RSV-вакцина, наблюдались более серьезные заболевания нижних дыхательных путей при последующем естественном RSV-инфицировании, чем у детей контрольной группы (Am. J. Epidemiology 89, 1969, стр. 405-421; J.Inf.Dis. 145, 1982, стр. 311-319). Кроме того, RSV-гликопротеины, очищенные посредством иммуноаффинной хроматографии с элюированием при кислотном значении pH, вызывали потенцирование иммунного ответа у хлопковых хомяков. (Vaccine, 10(7), 1992, стр. 475 - 484). Разработка эффективных вакцин против PIV-3 и RSV, которые не вызывали бы обострения легочных заболеваний после инфицирования вирусом дикого типа, связана со значительными терапевтическими осложнениями. Есть основания ожидать, что получение единого рекомбинантного иммуногена, обладающего протективным действием одновременно против PIV- и RSV-инфекций у детей младшего возраста, позволит значительно снизить процент заболеваемости и смертности в результате поражения указанными вирусами.
Известно, что протективный иммунный ответ против вируса парагриппа (PIV) и респираторно-синцитиального вируса (RSV) зависит от индуцирования нейтрализующих антител против мажорных гликопротеинов вирусной поверхности. В случае PIV такими протективными иммуногенами являются HN- белок, который имеет молекулярную массу 72 кДа и обладает как гемагглютинирующей, так и нейраминидазной активностью, и белок F (белок слияния), который имеет молекулярную массу 65 кДа и является ответственным за слияние вирусной и клеточной мембраны и за распространение этого вируса от клетки к клетке. В случае RSV двумя мажорными иммуногенными белками являются гликопротеин G (80-90 кДа) и белок F (70 кДа). Указанные белки G и F являются, очевидно, функциональными аналогами HN - беков и F - белков PIV соответственно. Гликопротеины F PIV и RSV были синтезированы в виде неактивных предшественников (FO), которые протеолитически расщепляются на N-концевой F2- и C-концевой F1- фрагменты, связанные между собой дисульфидными связями.
Рекомбинантные поверхностные гликопротеины PIV и RSV были по отдельности экспрессированы в клетках насекомых с использованием бакуловирусных систем (Ray и др. , (1989), Virus Research, 12: 169-180, Coelingh и др., (1987), Virology, 160: 465-472, Wathen и др., (1989), J. of Lnf. Dis., 159: 253-263), а также в клетках млекопитающих, инфицированных рекомбинантными поксвирусами (Spriggs и др., (1987), J.Virol. 61: 3416-3423; Stott и др., (1987), J. Virol. 61: 3855-3861). Было установлено, что рекомбинантные антигены, продуцируемые в этих системах, обеспечивают защиту хлопковых хомяков от контрольного заражения их живым вирусом. Позднее были сконструированы и продуцированы в клетках млекопитающих и насекомых гибридные рекомбинантные антигены F-G RSV (Wathan и др. , (1989), J.Gen. Virol. 70: 2625 - 2635; Wathen, опубл., международная пат. заявка WO 89/05823) и F-HN PIV-3 (Wathen, опубл. международная пат. заявка WO 89/10405). Было установлено, что гибридный антиген F-G RSV является протективным для хлопковых хомяков (Wathan и др., (1989) J.Gen.Virol. 70: 2637-2644), хотя он индуцирует слабый гуморальный ответ против белка G (Connors и др., (1992)., Vaccine 10: 475-484). Однако в этой опубликованной патентной заявке ничего не сообщается о протективной способности F- HN - белка PIV-3. Указанные антигены были сконструированы лишь в целях защиты против гомологического корпуса, то есть либо RSV, либо PIV-3. При этом совершенно очевидно, что было бы целесообразней и экономичней сконструировать и продуцировать один рекомбинантный иммуноген, содержащий по крайней мере один протективный антиген от каждого вируса, так, чтобы он мог служить защитой одновременно против PIV- или RSV - инфекций у детей младенческого и младшего возраста. Химерные белки, полученные в этих целях в соответствии с настоящим изобретением, могут быть введены также беременным женщинам и женщинам детородного возраста для стимуляции образования материнских антител как против PIV, так и против RSV. Кроме того, указанная вакцина может быть введена и другим индивидуумом с повышенной восприимчивостью, например лицам пожилого возраста.
Краткое описание изобретения.
В наиболее широком варианте своего осуществления настоящее изобретение относится к мультимерному гибридному гену, содержащему генную последовательность, кодирующую антигенную область белка от первого патогена и связанную с генной последовательностью, кодирующей антигенную область белка от второго патогена, и кроме того, настоящее изобретение относится к химерному белку, кодированному указанным мультимерным гибридным геном. Этот химерный белок включает в себя антигенную область белка от первого патогена, связанную с антигенной областью белка от второго патогена.
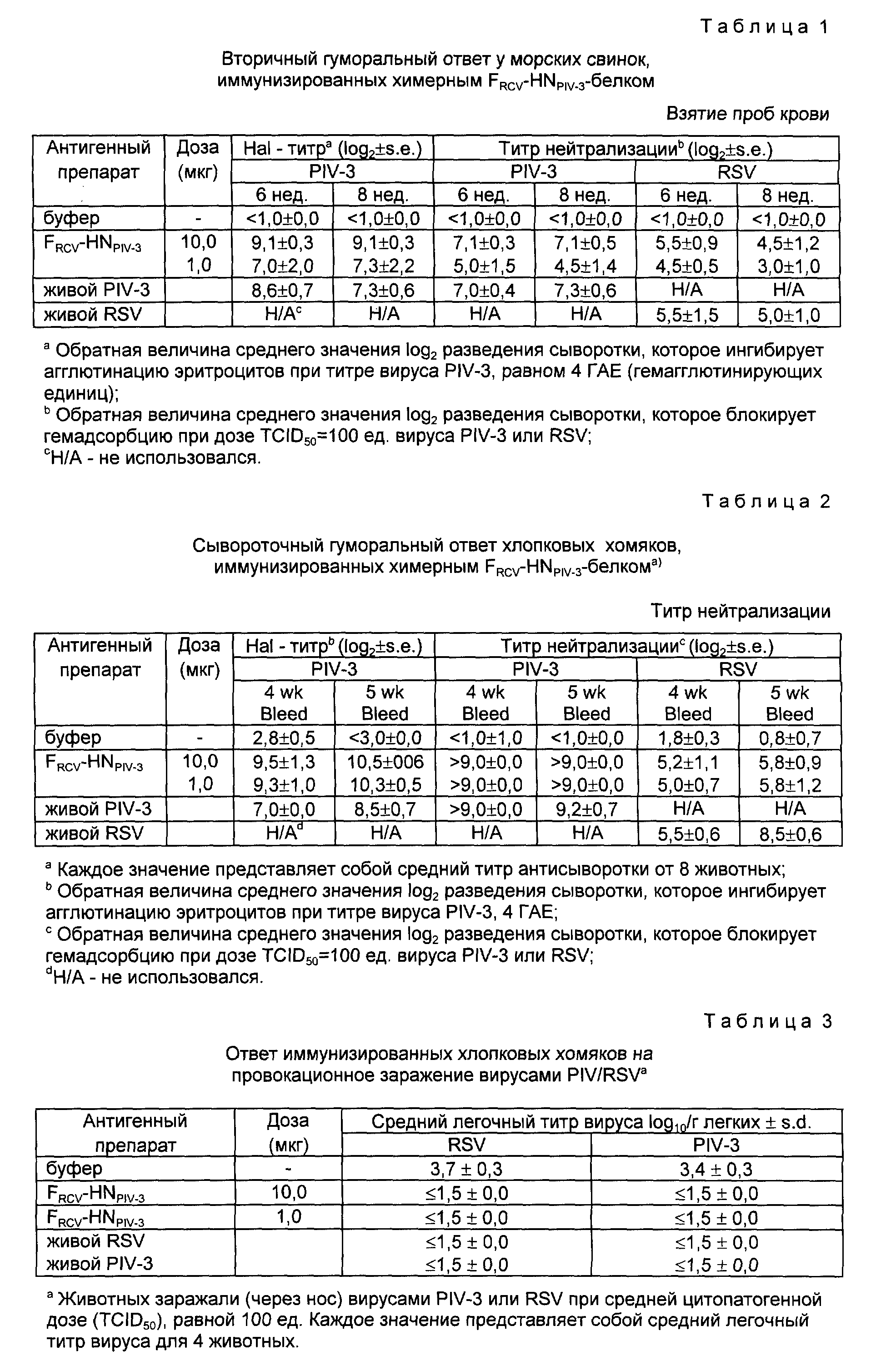
Указанные первый и второй патогены выбирают в основном из бактериальных и вирусных патогенов, а в одном из вариантов осуществления настоящего изобретения оба указанных патогена могут быть вирусными патогенами. Предпочтительно, если первый и второй патогены выбирают из таких патогенов, которые вызывают различные заболевания дыхательных путей, например заболевания верхних и нижних дыхательных путей. В предпочтительном варианте осуществления настоящего изобретения первым патогеном является парагриппозный вирус, а вторым патогеном является респираторно-синцитиальный вирус. В качестве белка парагриппозного вируса (PIV) использовали предпочтительно F- и HN- белки PIV-3, а в качестве белка респираторно-синцитиального вируса (RSV) использовали предпочтительно G - F белки RSV.
В другом своем варианте настоящее изобретение относится к клеткам, содержащим мультимерный гибридный ген и обеспечивающим экспрессию химерного белка, кодируемого указанным геном. Эти клетки могут быть бактериальными клетками, клетками млекопитающих, клетками насекомых, дрожжевыми клетками, или грибковыми клетками. Кроме того, настоящее изобретение относится к "живому" вектору, предназначенному для доставки антигена и содержащему мультимерный гибридный ген (причем таким вектором может быть вирусный или бактериальный вектор), а также его фармацевтически приемлемый носитель. Указанный живой вектор может образовывать активный компонент вакцины против заболеваний, вызываемых множеством патогенных инфекций. Эта вакцина может быть изготовлена в виде препарата, предназначенного для инъекций, для введения через нос, или для перорального введения.
В другом варианте своего осуществления настоящее изобретение относится к способу получения химерного белка, заключающемуся в том, что выделяют генную последовательность, кодирующую антигенную область белка первого патогена; выделяют генную последовательность, кодирующую антигенную область белка второго патогена; указанные генные последовательности сшивают, в результате чего получают мультимерный гибридный ген; и указанный мультимерный гибридный ген экспрессируют в клеточной экспрессирующей системе. Такой экспрессирующей клеточной системой может быть бактериальная клетка, клетка млекопитающего, клетка насекомого, дрожжевая клетка или грибковая клетка. Химерный белковый продукт, образующийся в результате экспрессии гена, может быть выделен из культуры клеточной экспрессирующей системы и очищен.
Настоящее изобретение также относится к вакцине против заболеваний, вызываемых множественными патогенными инфекциями, причем указанная вакцина включает в себя химерный белок, кодируемый мультимерным гибридным геном, и физиологически приемлемый носитель. Такая вакцина может быть изготовлена в соответствующей форме, предназначенной для инъекций, для введения через нос или для перорального введения.
Вакцины, полученные в соответствии с настоящим изобретением, могут быть использованы для иммунизации хозяина против заболеваний, вызываемых множественными патогенными инфекциями, а в частности против заболеваний, вызываемых парагриппозным вирусом и респираторно-синцитиальным вирусом; причем указанную иммунизацию осуществляют путем введения хозяину эффективного количества указанной вакцины. Как указывалось выше, для PIV и RSV человека таким хозяином могут быть дети младенческого и младшего возраста, беременные женщины, женщины детородного возраста и другие индивидуумы, восприимчивые к указанным вирусам, например люди пожилого возраста.
Химерный белок настоящего изобретения может быть также использован в соответствующих анализах в качестве диагностического реагента для выявления инфекции, вызываемой у хозяина множеством различных патогенов.
При этом следует отметить, что хотя описание настоящего изобретения сосредоточенно главным образом на химерной молекуле, которая является эффективной для иммунизации против заболеваний, вызываемых PIV- и RSV - инфекцией; но тем не менее в своем более широком смысле настоящее изобретение относится к любому химерному белку, который является эффективным для иммунизации против заболеваний, вызываемых множеством патогенов, и который содержит антиген от каждого из указанных патогенов; причем эти антигены связаны между собой в одной молекуле; и кроме того, настоящее изобретение относится к генам, кодирующим указанные химерные молекулы.
Термин "мультимерные гибридные гены", используемый в настоящем описании, означает гены, кодирующие антигенные области белков от различных патогенов; а термин "химерные белки" означает иммуногены, содержащие антигенные области белков от различных патогенов.
Краткое описание чертежей.
На фиг. 1 представлена нуклеотидная (SEQ ID N 1) и аминокислотная (SEQ ID N 2) последовательности PCR- амплифицированого F- гена и F- белка PIV соответственно.
На фиг. 2 представлена рестрикционная карта гена, кодирующего белок F PIV-3.
На фиг. 3 представлены нуклеотидная (SEQ ID N 3) и аминокислотная (SED ID N 4) последовательности гена HN PIV и белка HN PIV соответственно.
На фиг. 4 представлена рестрикционная карта гена, кодирующего белок HN PIV-3.
На фиг. 5 представлена нуклеотидная (SEQ ID N 5) и аминокислотная (SED ID N 6) последовательности гена F RSV и белка F RSV соответственно.
На фиг. 6 представлена рестрикционная карта гена, кодирующего F - белок RSV.
На фиг. 7 представлены нуклеотидная (SEQ ID N 7) и аминокислотная (SEQ ID N 8) последовательности G RSV и белка G RSV соответственно.
На фиг. 8 представлена рестрикционная карта гена, кодирующего белок G RSV.
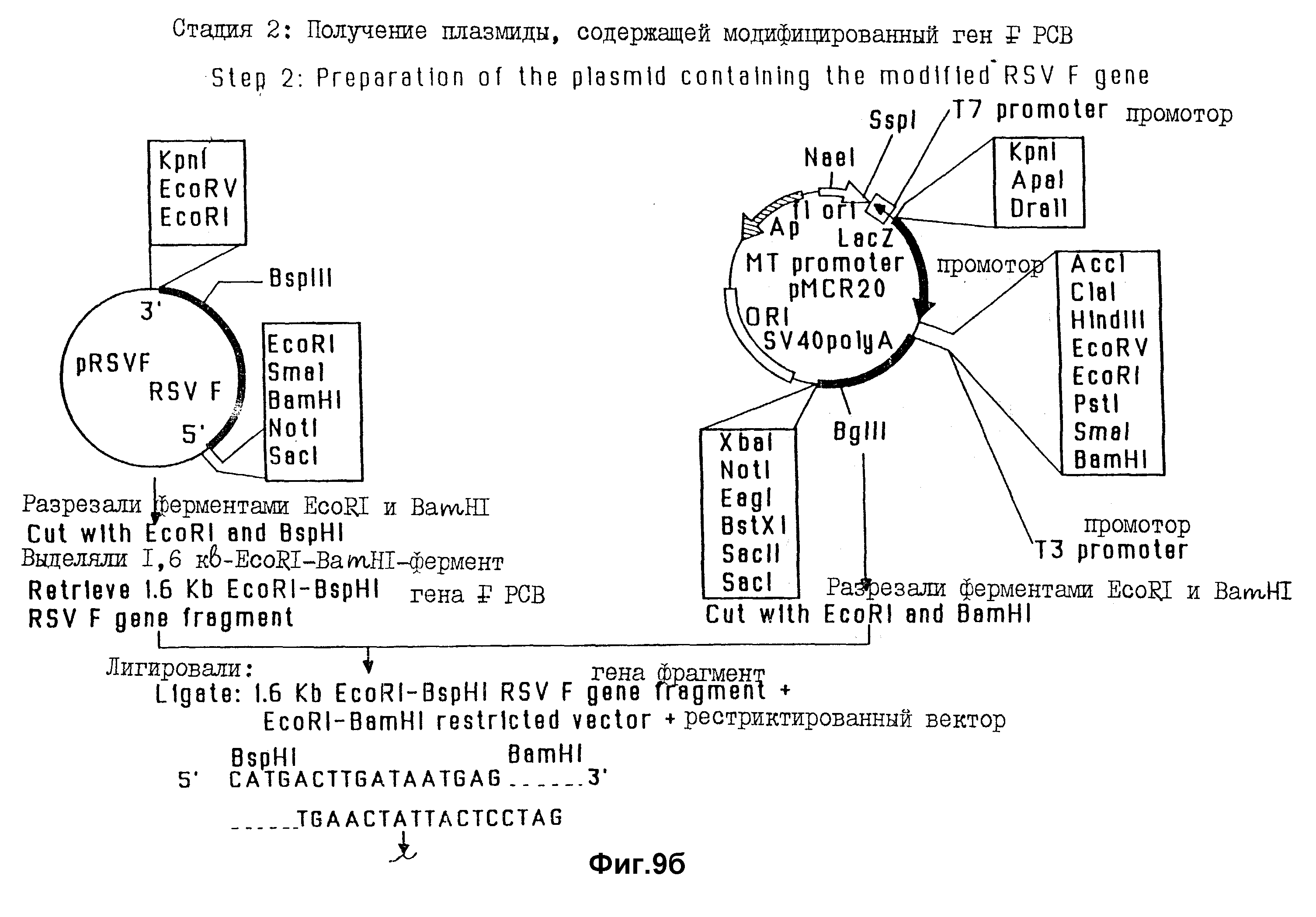
На фиг. 9. проиллюстрированы стадии конструирования экспрессирующего вектора, содержащего химерный ген FPIV-3 - FRSV.
На фиг. 10 проиллюстрированы стадии конструирования экспрессирующего вектора, содержащего FPIV-3 - ген, в котором отсутствует 5'-нетранслируемая последовательность и области, кодирующие трансмембранный "якорь" - домен и цитоплазматический концевой сегмент.
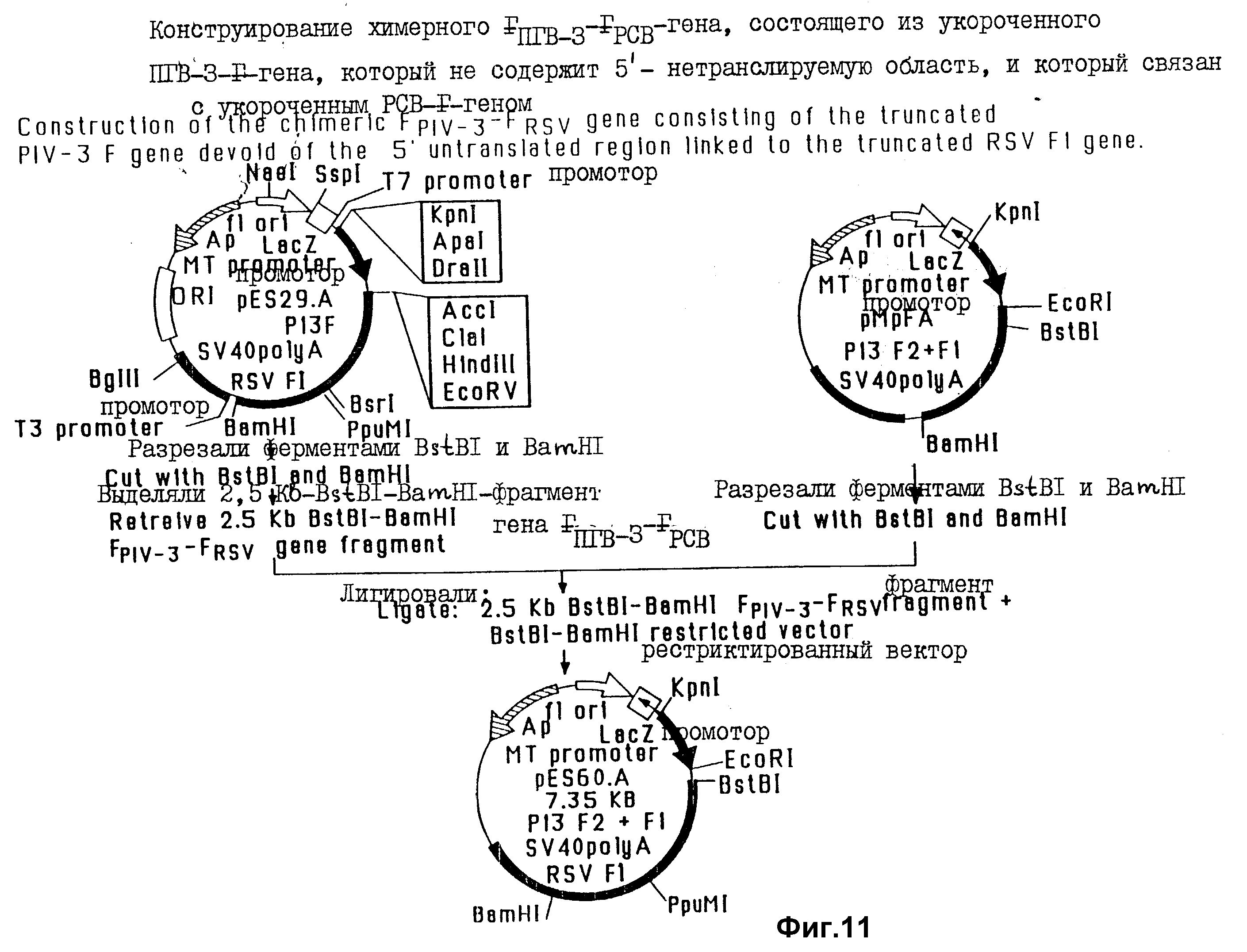
На фиг. 11 проиллюстрированы стадии конструирования экспрессирующего вектора, содержащего химерный ген FPIV-3 - FRSV, который включает в себя укороченный ген F PIV-3, лишенный 5'-нетранслируемой области и связанный с укороченным геном F1 RCV.
На фиг. 12 проиллюстрированы стадии конструирования экспрессирующего вектора, построенного на основе модифицированного бакуловируса 610 pAC и содержащего химерный ген FPIV-3 - FRSV, который включает в себя ген F PIV-3, лишенный 5'-нетранслируемой последовательности, а также области, колирующей трансмембранный домен и цитоплазматический концевой сегмент, и связанную с укороченным геном F1 RSV.
На фиг. 13 представлены иммуноблоты клеточных лизатов от клеток Sf9, инфицированных рекомбинантными бакуловирусами.
На фиг. 14 проиллюстрированы стадии конструирования вектора переноса бакуловируса (pD2).
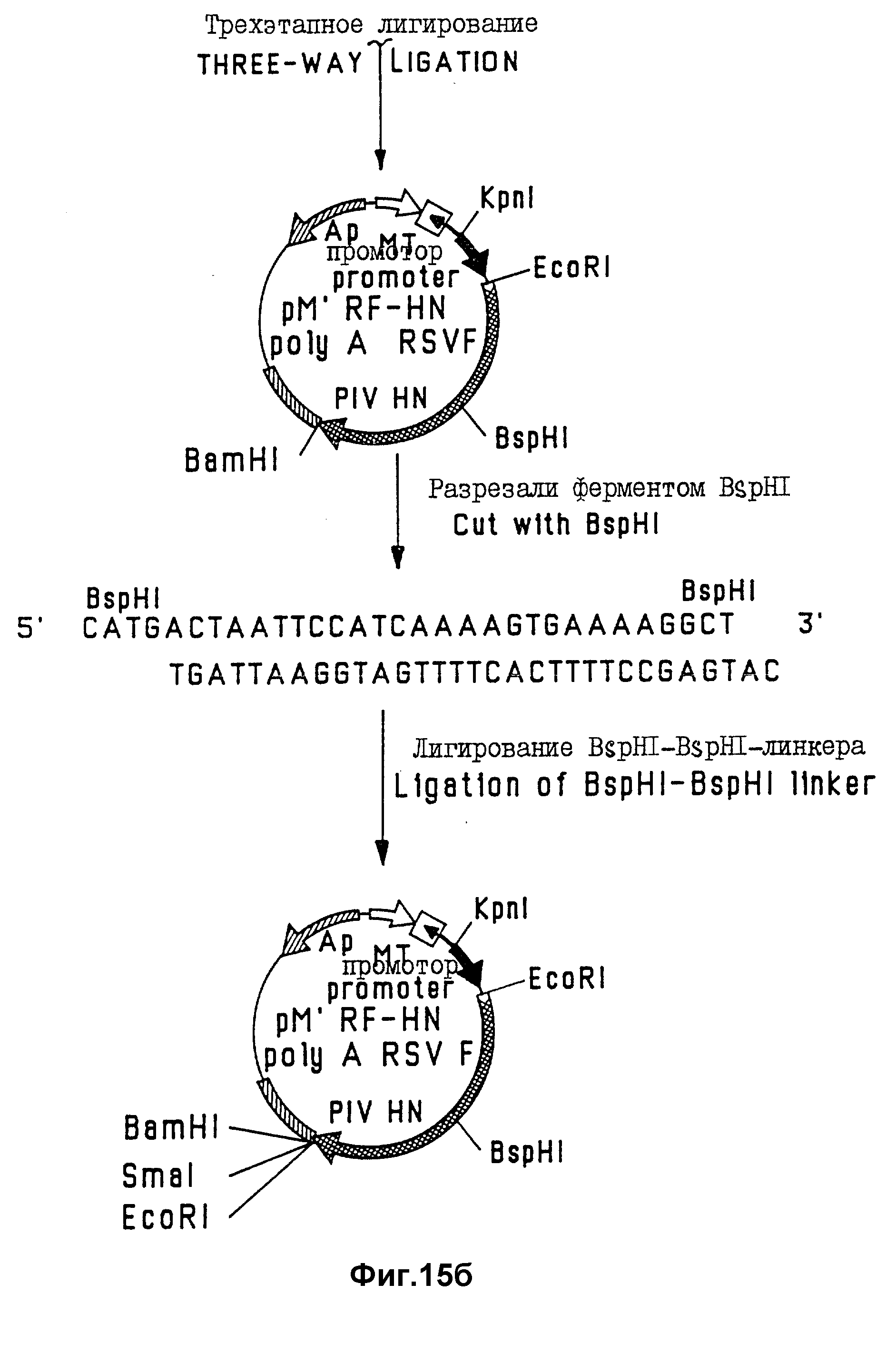
На фиг. 15 проиллюстрированы стадии конструирования химерного гена FRSV-HNPIV-3.
На фиг. 16 показаны ДСH-ПАА-гель и иммуноблоты очищенного химерного FRSV-HNPIV-3- белка.
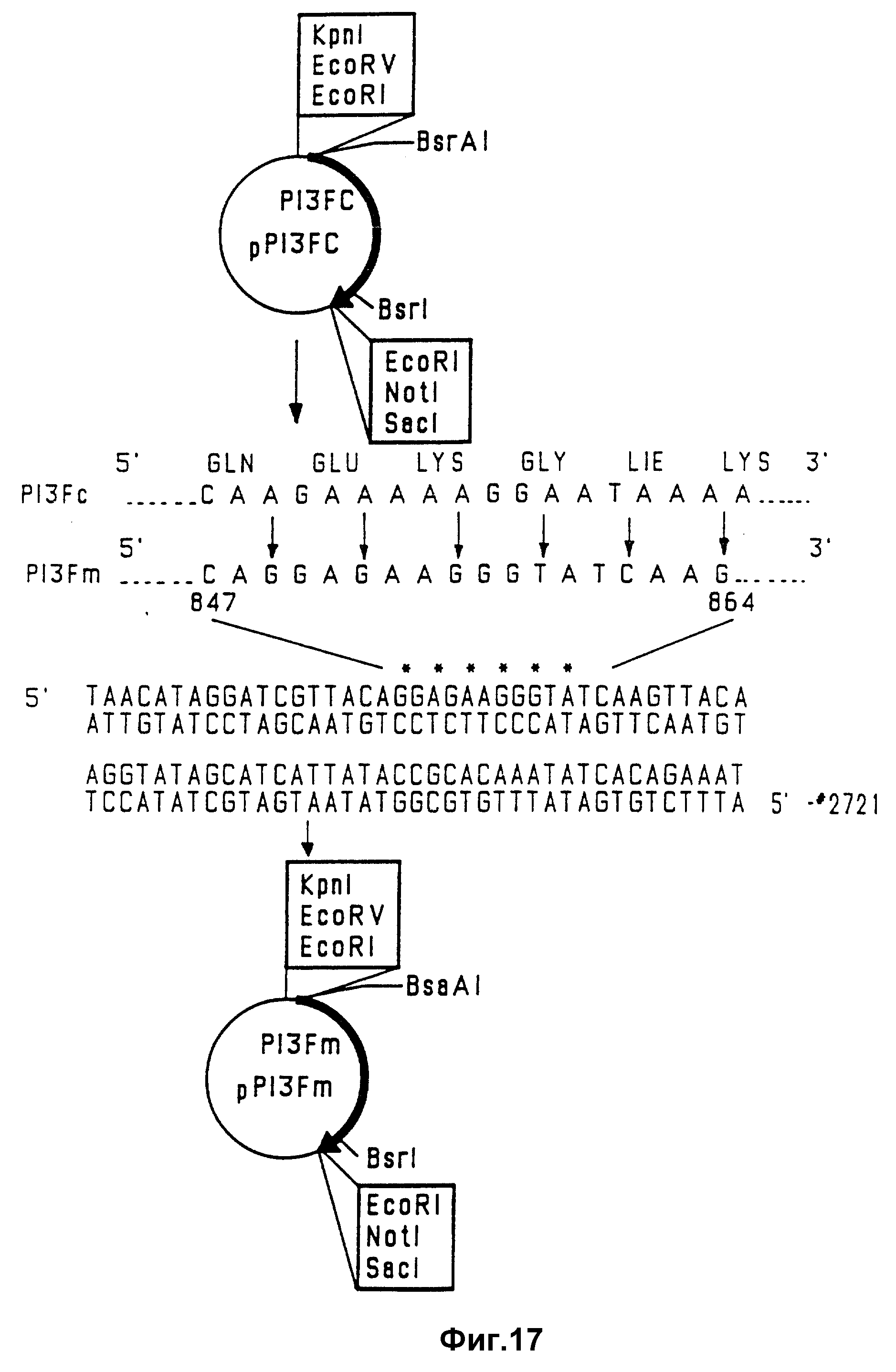
На фиг. 17 проиллюстрирован мутагенез гена PIV-3.
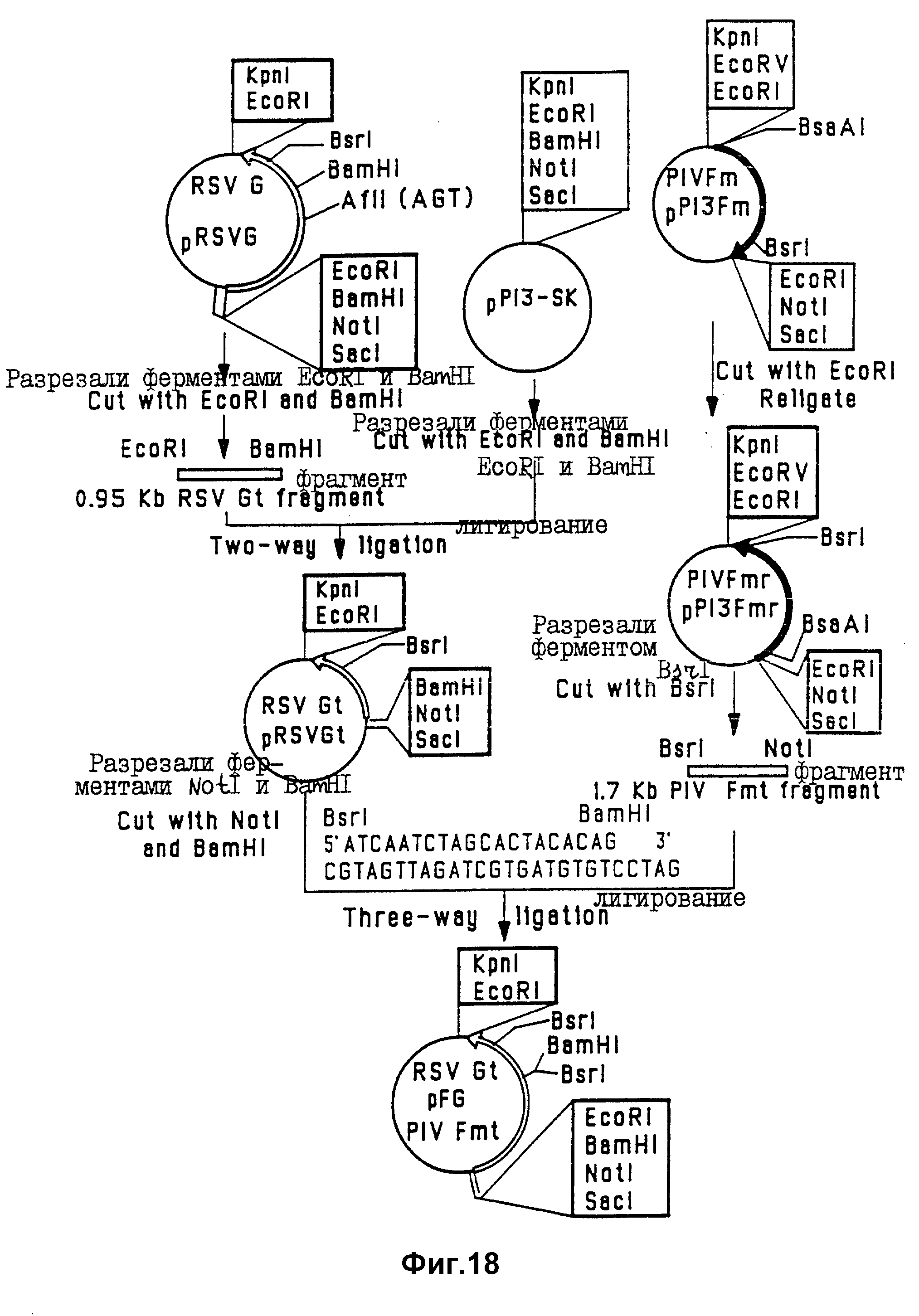
На фиг. 18 проиллюстрированы стадии конструирования химерного гена FPIV-3-GRSV.
Общее описание изобретения
В настоящем изобретении была
получена химерная молекула, обеспечивающая защиту от двух основных заболеваний детей младшего возраста. Более конкретно настоящее изобретение относится к комбинации различных рекомбинантных
иммуногенов, а именно рекомбинантного парагриппозного вируса ( PIV) и респираторно-синцитиального вируса (RSV), предназначенной для получения безопасных и эффективных вакцин, способных обеспечить
защиту детей младенческого и младшего возраста, а также других индивидуумов, восприимчивых к этим вирусам, от заболеваний, вызываемых обеими PIV - и RSV - инфекциями. Однако, как указывалось выше,
настоящее изобретение распространяется и на конструирование мультимерных гибридных генов, включающих в себя гены, кодирующие протективные антигены от многих патогенов. Такие вакцины могут быть введены
любым способом, например путем инъекции, через нос или перорально.
В частности, авторами настоящей заявки были сконструированы несколько моделей химерных PIV/RSV - генов, содержащих соответствующие последовательности от выбранных генов, кодирующих PIV-3 и RSV - поверхностные гликопротеины, сцепленные друг с другом тандемом. Все гены в химерных конструкциях, описанных в настоящей заявке, были получены из поздних клинических изолятов вирусов PIV-3 и RSV. Указанные химерные генные конструкции могут содержать последовательности либо от генов F - PIV-3, либо от генов HN PIV-3, сцепленных тандемом либо с генами F RSV, либо с генами G RSV во всех возможных ориентациях относительно друг друга, и во всех возможных комбинациях.
Химерные генные конструкции настоящего изобретения могут содержать либо последовательности полного гена, либо его сегментов, кодирующих иммуногенные и протективные эпитопы. Природная нуклеотидная последовательность указанных генов может быть модифицирована путем мутации при сохранении антигенности; причем указанные модификации могут включать в себя удаление соответствующих предтранскрипционных терминаторов для оптимизации экспрессии этих генов в эукариотических клетках. Сконструированные гены кодируют PIV - RSV - гибридные поверхностные гликопротеины, сцепленные тандемом в единой конструкции, в результате чего продуцируются генные продукты, способствующие образованию протективных антител против парагриппозного и респираторно-синцитиального вирусов. Указанные мультимерные гибридные гены состоят из генной последовательности, кодирующей F- или HN- белок человеческого PIV-3 или его иммуногенного эпитоп-содержащего фрагмента, связанного с генной последовательностью, кодирующей F- или G-белок человеческого RSV, или его иммуногенного эпитоп-содержащего фрагмента. Примерами конкретных генных конструкций могут служить гибридные гены FPIV-3-FRSV, FRSV-HNPIV-3 и FPIV-3 - GRSV.
Кроме того, настоящее изобретение также относится к конструированию других мультимерных генов, например тримерных генов, содержащих PIV - и RSV- гены или сегменты этих генов, связанные во всех возможных ориентациях по отношению друг к другу.
Например: FPIV - HNPIV-F или GRSV
FPIV - FRSV - GRSV
HNPIV - FRSV - GRSV
Мультимерные гены настоящего изобретения могут также содержать по крайней мере один
ген, кодирующий по крайней мере одну иммуногенную и/или иммуностимулирующую молекулу.
Мультимерные гибридные гены настоящего изобретения могут быть субклонированы в соответствующие векторы для экспрессии в клеточных экспрессирующих системах. Такими клеточными экспрессирующими системами могут быть бактериальные клетки, клетки млекопитающих, насекомых и грибков, например дрожжей.
Химерные белки, полученные в соответствии с настоящим изобретением, могут быть также представлены иммунной системе посредством использования "живого" вектора, включая "живые" вирусные векторы, такие как рекомбинантные посквирусы, аденовирусы, ретровирусы, вирусы лесов Семлики; и "живые" бактериальные векторы, такие как векторы на основе сальмонеллы и микобактерий (например, бациллы Кальмета-Герена, BCG).
Затем химерные белки, такие как PIV/RSV - химеры, присутствующие либо в супернатантах, либо в клеточных лизатах трансфецированных, трансформированных или инфицированных клеток, могут быть очищены любым традиционным способом.
Для оценки иммуногенности и протективной способности химерных белков подходящие экспериментальные животные были иммунизированы различными дозами очищенных химерных белков, таких как PIV/RSV - химеры, и/или живыми рекомбинантными векторами, описанными выше. Указанные химерные белки могут быть представлены иммунной системе либо путем использования физиологически приемлемых носителей, таких как фосфат алюминия, либо путем использования систем доставки, таких как ISCOMS и липосомы. Кроме того, указанные химеры могут быть изготовлены таким путем, что они будут обладать способностью индуцировать мукозальный ответ, например, путем конъюгирования или объединения с носителями, обеспечивающими направленную иммунодоставку данных химер, например, такими как субъединица холерного токсина B; либо путем введения этих химер в микрочастицы. Кроме того, такие вакцины могут служить средством для доставки мультимерного белка непосредственно в клети иммунной системы, например, для доставки молекул токсина или антител. Для усиления иммунозащитной функции химерных белков они могут быть использованы в сочетании с другими иммуногенными и/или иммуностимулирующими молекулами. Например, описанные в настоящей заявке химерные PIV/RSV - белки в сочетании с адъювантом, таким как фосфат алюминия, могут быть использованы для получения готовой к употреблению инъецируемой вакцины против заболеваний, вызываемых обоими вирусами PIV-3 и RSV. Указанные химерные белки могут быть также введены через нос или перорально. Кроме того, эти химерные белки могут быть использованы в тест-наборах для диагностики PIV-3 и RSV - инфекций.
Настоящее изобретение не ограничивается получением лишь PIV-3 и RSV - химерных белков, оно может быть также использовано для получения химерных иммуногенов, состоящих либо из полных последовательностей ли из областей иммуногенных белков, происходящих по крайней мере от двух патогенов, и последовательно соединенных друг с другом в одной единой молекуле. Кроме того, химерные антигены могут быть синтезированы таким образом, что они будут содержать иммунодоминантные эпитопы несколько белков от различных патогенов. Эти химерные антигены могут быть использованы в качестве вакцин или в качестве диагностических реагентов.
Идентификация последовательностей
В настоящей заявке рассматривается несколько нуклеотидных и аминокислотных последовательностей.
Идентификация и локализация этих последовательностей представлены в таблице 4.
Данные о депонировании
Некоторые плазмидные ДНК, которые описаны, и на которые имеются ссылки в настоящем описании, были
депонированы Американской коллекцией типовых культур (АТСС) (находящейся в Роквиле, шт. Мэриленд, США) в соответствии с Будапештским договором до подачи настоящей заявки. Эти депонированные очищенные
плазмиды будут доступны специалистам после выдачи патента на основании данной заявки на патент США или после публикации соответствующей заявки на Европатент независимо от того, будет ли выдан
упомянутый патент. При этом следует отметить, что объем изобретения, описанного и заявленного в настоящей заявке, не ограничивается плазмидными ДНК депонированных конструкций, поскольку указанные
депонированные плазмиды используются лишь в иллюстративных целях. Нижеприведенные очищенные плазмиды были депонированы 17 декабря 1992 г. в АТСС под следующими входящими номерами (таблица 5).
Любые эквивалентные плазмиды, которые могут быть использованы для продуцирования эквивалентных антигенов, описанных в настоящей заявке, входят в объем настоящего изобретения.
Примеры.
Представленное выше описание раскрывает настоящее изобретение в общих чертах. Для более ясного понимания сущности изобретения ниже приводятся более конкретные примеры. Эти примеры описаны лишь с иллюстративной целью и не должны рассматриваться как некое ограничение изобретения. Все модификации и эквивалентные замещения представляют собой в основном предположительные или целесообразные варианты осуществления изобретения. Кроме того, используемые в настоящем описании конкретные термины имеют главным образом описательный смысл и не должны рассматриваться как некое ограничение изобретения.
В настоящем описании не приводится подробного раскрытия методом клонирования и секвенирования генов PIV-3 и RSV, а также техники субклонирования генов в соответствующих векторах и экспрессии генных конструкций в клетках млекопитающих и насекомых, поскольку указанные методы являются хорошо известными специалистам.
Пример 1
В
этом примере описывается стратегия, используемая для клонирования и секвенирования генов F PIV-3 и HN PIV-3, и генов G и F RSV (из изолята типа A). Эти гены были использованы для конструирования
химерных генов FPIV-3 - FRSV, FRSV-HNPIV-3 и FPIV-3-GRSV, которые подробно описаны в примерах 2-4, 9 и 15 соответственно.
Сначала путем PCR - амплицикации кДНК, происходящей от вирусной РНК, экстрагированной из позднего клинического изолята PIV-3, получали клоны гена, кодирующего F-белок вируса PIV-3 (сокращенно ген F PIV-3. Прим. пер.). Два других клона гена F PIV-3, а также гены HN PIV-3, F RSV и G RSV клонировали из кДНК- библиотеки, полученной от мРНК, выделенной из клеток MRC-5, которые были инфицированы клиническими изолятами либо вируса PIV-3, либо вируса RSV (изолят типа A). Клоны генов F PIV-3 (оба PCR -амплифицированный и неамплифицированный клона), HN PIV-3, F RSV и G RSV секвенировали дидезокси-методом обрыва цепи. Секвенирование обеих цепей генов осуществляли как вручную, так и автоматически.
Нуклеотидная (SEQ ID N 1) и аминокислотная (SEQ ID N 2) последовательности PCR-амплифицированного гена F PIV-3 и F- белка PIV-3 соответственно представлены на фиг. 1, а рестрикционная карта этого гена показана на фиг. 2. Анализ последовательности из 1844 нуклеотидов двух PCR-амплифицированных клонов гена F PIV-3 подтвердили идентичность этих клонов. Сравнение кодирующей последовательности PCR-амплифицированного клона гена F PIV-3 с известной последовательностью гена F PIV-3 выявлено 2,6%-ное расхождение между этими двумя последовательностями, которое привело к замещениям в 14 аминокислотах.
Нуклеотидная последовательность не амплифицированного посредством PCR клона гена F PIV-3 отличалась от PCR-амплифицированного клона этого гена следующим образом: неамплифицированный клон имел 10 дополнительных нуклеотидов (AGGACAAAAG) в 5'-нетранслируемой области гена, и имел расхождения в 4 положениях, а именно : 8 (T PCR - амплифицированном гене, и C в не-PCR-амплифицированном гене), 512 (C в PCR - амплифицированном гене, и T в не-PCR-амплифицированном гене), 518 (G в PCR - амплифицированном гене, и A в не - PCR - амплифицированном гене), 1376 (A в PCR - амплифицированном гене, и G в не - PCR - амплифицированном гене). Эти изменения привели к трем заменам в аминокислотной последовательности F - белка, кодированного не PCR - амплифицированным геном F PIV-3. Серии (положение 110), глицин (положение 112) и аспарагиновая кислота (положение 398) в первичной аминокислотной последовательности F - белка, кодированного PCR - амплифицированным геном F PIV-3, были заменены фенилаланином (положение 110), глутаминовой кислотой (положение 112) и глицином (положение 398) соответственно в первичной аминокислотной последовательности F - белка, кодированного PCR - амплифицированным клоном.
На фиг. 3 показаны нуклеотидная (SEQ ID N 3) и аминокислотная (SEQ ID N 4) последовательности гена и белка HN PIV-3 соответственно, а рестрикционная карта указанного гена представлена на фиг. 4. Анализ 1833 нуклеотидов последовательности двух HN-клонов подтвердили идентичность этих последовательностей. При сравнении этой последовательности с известной кодирующей последовательностью HN PIV-3 было выявлено 4,4%-ное расхождение в этих последовательностях. Это расхождение привело к 17 замещениям в аминокислотной последовательности белка, кодированного геном HN PIV-3.
Нуклеотидная (SEQ ID N 5) и аминокислотная (SEQ ID N 6) последовательности гена F RSV и F-белка RSV соответственно показаны на фиг. 5, а рестрикционная карта этого гена показана на фиг. 6. Анализ 1887 - нуклеотидной последовательности двух F- клонов RSV подтвердил полную гомологию между последовательностями этих двух клонов. Сравнение этой нуклеотидной последовательности с известной последовательностью гена F RSV выявлено приблизительно 1,8%-ное расхождение в кодирующей последовательности, которое привело к замещениям в 11 аминокислотах.
Нуклеотидная (SEQ ID N 7) и аминокислотная (SEQ ID N 8) последовательности гена G RSV и G белка RSV соответственно представлены на фиг. 7, а рестрикционная карта этого гена показана на фиг. 8. Сравнение 920-нуклеотидной последовательности клона гена G с известной G - последовательностью (изолят типа A) выявило 4,2%-ное расхождение в нуклеотидной последовательности и 6,7%-ное расхождение в аминокислотной последовательности генного продукта. Это расхождение привело к замещениям в 20 аминокислотах.
Полные гены F PIV-3 (не - PCR - амплифицированный), HN PIV-3, F PSV и G RSV клонировали в λgt 11 и субклонировали в сайт множественного клонирования вектора Bluescript M13-SK, либо путем лигирования по "тупым" концам или путем использования соответствующих линкеров. PCR-амплифицированный ген F PIV-3 клонировали непосредственно в вектор Biuescript. Клонирующие векторы, содержащие PCR - амплифицированный ген F PIV-3, не - PCR - амплифицированный ген F PIV-3, ген HN PIV-3, ген F RSV и ген G RSV, обозначали соответственно pP13F, pP13Fc, pPIVHN, pRSVF и pRSVG.
Пример 2.
В этом примере проиллюстрировано конструирование экспрессирующего вектора, получаемого на основе Bluescript (pMCP20), и содержащего FPIV-3-FRSV-химерный ген. Эта химерная генная конструкция содержит 5'-нетранслируемую область гена F PIV-3, но не содержит области, кодирующие гидрофобный домен и цитоплазматический концевой домен от обоих F-генов PIV-3 и RSV. Все стадии конструирования указанной плазмиды систематизированы на фиг. 9.
Для получения PIV-3 - части химерного гена (фиг. 9, стадия 1) полный ген PIV-3, в котором отсутствует область, кодирующая трансмембранный домен и цитоплазматический концевой домен, выделяли из плазмиды pP13F путем отрезания полилинкера ферментом Bam H1, затупления концов линеаризированной плазмиды с использованием полимеразы Кленова; и вырезания гена ферментом BsrI. Кассету BsrI-BamHI олигонуклеотидов (SEQ ID N 9), содержащую Ppu MI - сайт и три последовательно расположенные кодона терминации трансляции, лигировали с укороченным 1,6 kb Bam HI-BsrI-фрагментом гена F PIV-3 и клонировали в EcoRV-BamHI-сайты экспрессирующего вектора Bluescript M 13-SK, содержащего промотор человеческого металлотионена и poly A- и IVS-последовательности генома SV40 (обознач. pMCR20), в результате чего получали pME1.
Для получения RSV - F - генного компонента химерной конструкции (фиг. 9, стадия 2). Ген F RSV, в котором отсутствовали области, кодирующие трансмембранный домен и цитоплазматический концевой домен, восстанавливали и плазмиды pPSVF путем отрезания полилинкера ферментом Eco RI и вырезания гена ферментом BspHI. Синтетическую кассету BspHI- Bam HI-олигонуклеотидов (SEQ ID N 10), содержащую три последовательно расположенных кодона терминации трансляции, лигировали с укороченным 1,6 kb - геном F RSV и клонировали в EcoRI-BamHI-сайты вектора, полученного на основе Biuescript (pM CR20), в результате чего получали плазмиду pES 13A. Затем плазмиду pES 13A разрезали ферментами EcoRI и PpuMI для удаления лидерной и F2- кодирующей последовательности из укороченного гена F RSV. Лидерную последовательность восстанавливали с использованием EcoRI-PpuMI - олигонуклеотидную кассету (SEQ ID N 11) и лигировали с RSV - F1-генным сегментом, в результате чего получали плазмиду pES23A.
Для получения химерного FPIV-3-FRSV - гена (фиг. 9, стадия 3), содержащего 5'-нетранслируемую область PIV-3 - F-гена, сцепленного с укороченным фрагментом гена F1 RSV, плазмиду pMEI (содержащую укороченный 1,6 kb - ген F PIV-3) сначала разрезали ферментами PpuMI и BamHI. PpuMI - BamHI - рестриктированный вектор pME1 дефосфорилировали кишечной щелочной фосфатазой. 1,1-kb - фрагмент гена F1 RSV выделяли из плазмиды pES23A путем разрезания плазмиды ферментами PpuMI и BamHI. Этот 1,1 kb- PpuMI - BamHI-фрагмент гена F1 RSV клонировали в PpuMI-BamHI-сайты дефосфорилированного вектора pMEI, в результате чего получали плазмиду pES29A. Эта химерная генная конструкция содержала 5'-нетранслируемую область гена PIV-3, но не содержала нуклеотидные последовательности, кодирующие гидрофобные "якорные" домены, и цитоплазматические концы обоих F- белков PIV-3 и RSV.
Пример 3
В этом примере проиллюстрировано конструирование экспрессирующего вектора, полученного на основе Bluescript и содержащего ген F PIV-3, в котором отсутствуют 5'-нетранслируемая область и области,
кодирующие трансмембранный домен - "якорь" и цитоплазматический концевой домен. Стадии конструирования этой плазмиды показаны на фиг. 10.
Плазмиду pP113F, содержащую полный ген F PIV-3, разрезали ферментом BamHI, затупляли по концам с использованием полимеразы Кленова, а затем разрезали ферментом BsrI для удаления трансмембранного домена и цитоплазматических концевых кодирующих областей. Вектор, полученный на основе Bluescript (pMCR20) разрезали ферментами SmaI и BamHI. Синтетическую кассету BsrI-BamHI - олигонуклеотидов (SEQ ID N 12), содержащую кодон терминации трансляции, лигировали с затупленным по концу BsrI 1,6 Kb - фрагментом гена F PIV-3 и с SmaI-BamHI-рестриктированным вектором pMCR20, в результате чего получали плазмиду pMp FB. Ген F PIV-3 этой конструкции не содержал ДНК-фрагмент, кодирующий трансмембранный и цитоплазматические якорные домены, но содержал 5'-нетранслируемую область. Для конструирования плазмиды, содержащей PIV-3 - F-ген, лишенный 5'-нетранслируемой области и ДНК-фрагмента, кодирующего гидрофобный домен, плазмиду pMpFB разрезали ферментами EcoRI и BstBI. Кассету EcoRI-Bst BI - олигонуклеотидов (SEQ ID N 13), содержащую последовательность для восстановления сигнального пептида и кодирующую последовательности, удаленные путем Eco RI-BstBI-рестрикции, лигировали с EcoRI-BstBI-рестриктированным вектором pMp FB, в результате чего получали плазмиду pMpFA.
Пример 4
В этом примере иллюстрируется конструирование химерного FPIV-3-FRSV-гена, состоящего из укороченного
PIV-3 - F - гена, лишенного 5'-нетранслируемой области и лигированного с укороченным геном FI RSV. Все стадии конструирования этой плазмиды показаны на фиг. 11.
Для получения указанной химерной генной конструкции плазмиду pES29A (пример 2) разрезали ферментами BstBI и BamHI для высвобождения химерного генного 2,5 kb - Bst BI-BamHI-фрагмента FI PIV-3 - F - RSV. Этот Bst BI - BamHI-фрагмент выделяли из легкоплавкого агарозного геля и клонировали в BstBI-BamHI-сайты дефосфорилированного вектора pMpFA, в результате чего получали плазмиду p ES60A. Эта конструкция содержала ген F PIV-3, в котором отсутствовали обе 5'-нетранслируемая область и последовательности, кодирующие гидрофобный домен и цитоплазматический концевой домен, и который связан с FI-кодирующей областью укороченного гена F RSV. Этот химерный ген затем субклонировали в бакуловирусный вектор переноса (см. пример 5).
Пример 5
В этом примере проиллюстрировано конструирование
модифицированного бакуловирусного вектора pAC 610, содержащего нативный полиэдриновый промотор и химерный FPIV-3 - FRSV - ген, включающий в себя ген F PIV, в котором отсутствует
5'-нетранслируемая последовательность и нуклеотидная последовательность, кодирующая гидрофобный якорный домен и цитоплазматический конец, и который связан с укороченным геном FI RSV. Конструирование
этой плазмиды проиллюстрировано на фиг. 12.
Бакуловирусный экспрессирующий вектор pAC 610 модифицировали нижеописанным способом так, чтобы он содержал нативный полиэдриновый промотор. Бакуловирусный 9,4 kb-вектор, в котором отсутствовала EcoRV-BamHI-последовательность, ДНК, выделяли из низкоплавкого агарозного геля и обрабатывали кишечной щелочной фосфатазой. В трехэтапном лигировании кассету Eco RV-Eco RI-олигонуклеотидов (SEQ ID N 14), содержащую нуклеотиды, необходимые для выделения нативного поли эдринового промотора, лигировали с 1,6 kb - EcoRI-BamHI-фрагментом укороченного гена F RSV, выделенным из конструкции p ES13A (пример 2, стадия 2), и с EcoRV-BamHI-рестриктиворанным и обработанным фосфатазой вектором, в результате чего получали плазмиду pES47A. Для получения экспрессирующего вектора на основе pAC 610, содержащего химерный FPIV-3 - FRSV - ген, плазмиду pES47A разрезали ферментами ECoRI и BamHI для удаления 1,6 kb - вставки укороченного гена F RSV. Химерный FPIV-3-FRSV-ген (2,8 kb) выделяли путем разрезания плазмиды pES 60A (пример 4) ферментами EcoRI и BamHI. Этот химерный EcoRI-BamHI-ген (2,8 kb) лигировали с EcoRI-BamHI-рестриктированным вектором pES47A, в результате чего получали плазмиду pAC DR7 (ATCC 75387).
Пример 6
В этом примере описывается получение очищенных
методом бляшек рекомбинантных бакуловирусов, содержащих химерный ген FPIV-3-FRSV.
Клетки Spodoptera frugiperda (Sf9) трансфецировали 1,0 микрограммами AcM NPV-ДНК дикого типа и 2,5 микрограммами FPIV-3-FRSV-плазмидной ДНК (плазмида pAC DR7- пример 5). Предлагаемые рекомбинантные бакуловирусы (очищенные один раз путем серийного разведения), содержащие FPIV-3-FRSV- химерный ген, идентифицировали с помощью дот-блотгибридизации. Лизаты клеток насекомого, инфицированных предполагаемыми рекомбинантными бакуловирусами, зондировали с использованием32F-меченной вставки химерного FPIV-3-FRSV-гена. Перед использованием в экспериментах по экспрессии рекомбинантные бакуловирусы дважды очищали методом бляшек. Все процедуры проводили по методике, описанной M.D. Summers and G.E. Smith in "A Manual of Methods for Baculovirus Vectors and Insect Cell Culture Procedures", Texas Agricultural Experiment Station, Bulletin 1555, 1987.
Пример 7
Этот пример иллюстрирует анализы на присутствие химерного FPIV-3-FRSV-белка в
супернатантах и клеточных лизатах -Sf9 - инфицированных клеток.
Клетки насекомого инфицировали рекомбинантными бакуловирусами, очищенными, как описано в примере 6, при множественности
заражения = 8. Концентрированные супернатанты, полученные от клеток, инфицированных рекомбинантными вирусами, показали положительную реакцию при специфическом ELISA - анализе на белок F PIV-3. Кроме
того, после ДСН-ПААГ-электрофореза лизатов35S-метионин-меченных инфицированных клеток и анализа гелей с помощью авторадиографии в лизатах клеток, инфицированных рекомбинантными вирусами,
присутствовала яркая полоса с кажущейся молекулярнойй массой приблизительно 90 кДа, однако эта полоса отсутствовала с лизатах клеток, инфицированных вирусом дикого типа. Присутствие химерного FPIV-3-FRSV-белка в лизатах клеток, инфицированных рекомбинантными бакуловирусами, было также подтверждено Вестерн-блотнализом, проведенным с использованием моноспецифических
антисывороток против PIV-3-F и против RSV-F и/или моноклональных антител (Mads). При иммуноблоттинге лизаты клеток, инфицированных рекомбинантными бакуловирусами, вступают в реакцию с антисыворотками
против PIV-3 и против RSV. Как показали иммунолот-анализ (фиг. 13), лизаты клеток, инфицированных либо RSV-F- либо FPIV-3-FRSV - рекомбинантными бакуловирусами, обнаруживали
положительную реакцию с моноклональными антителами против F RSV. Как и ожидалось, лизаты клеток, инфицированных вирусом дикого типа, не реагировали с этими моноклональными антителами. Кроме того,
только лизаты клеток, инфицированных химерными FPIV-3-FRSV-рекомбинантными вирусами, реагировали с антисывороткой против F1PIV-3
Пример 8
Этот пример
иллюстрирует модификацию бакуловирусного вектора переноса pVL1392 (полученного от Invitrogen), в котором полиэдриновый инициирующий кодон ATG был заменен на кодон ATT, а последовательность CCG
находится за геном полиэдрина (5'->3') в положениях +4, 5, 6. Известно, что инсерцию нескольких пар оснований структурного гена ниже (5'->3') ATT - кодона осуществляют для усиления
трансляции. Стадии конструирования такого модифицированного бакуловирусного вектора переноса показаны на фиг. 14.
Бакуловирусный экспрессирующий вектор pVL 1392 разрезали ферментами Eco RV и Bam HI. Рестриктированный 9,5 kb - вектор pVL 1392 лигировали с кассетой Eco RV - Bam HI - олигонуклеотидов (SEQ ID N 15) и получали в результате вектор pD2.
Пример 9
Этот пример иллюстрирует конструирование бакуловирусного экспрессирующего вектора, содержащего химерный FRSV-HNPIV-3-ген, состоящий из укороченных RSV-F- PIV-3-HN-генов,
сцепленных тандемом. Стадии конструирования этой плазмиды схематически показаны на фиг. 15.
Для конструирования FRSV-HNPIV-3-гена ген F RSV, в котором отсутствует нуклеотидная последовательность, кодирующая трансмембранный домен и цитоплазматический концевой домен гликопротеина F RSV, выделяли из плазмиды pRSVE (пример 1) путем отрезания полилинкера ферментом Eco RI, а гена ферментом BspHI. Ген HN PIV-3, в котором отсутствует ДНК-фрагмент, кодирующий гидрофобный якорный домен, выделяли из плазмиды pPIVHN (пример 1) путем отрезания гена ферментом BspHI, а полилинкера ферментом BamHI. 1,6 kb - Eco RI-BspHI-фрагмент гена F RSV и 1,7 Kb- BspHI-Bam HI-фрагмент гена HN PIV-3 удаляли из легкоплавкого агарозного геля. В целях клонирования два BspHI - сайта в векторе экспрессии для клеток млекопитающих, полученном на основе Bruescript были мутированы. Эти мутации были введены в BspHI - сайты плазмиды pMC R20 путем разрезания экспрессирующего вектора ферментом BapHI, обработки BspHI-рестрикционного вектора и полученного в результате этой рестрикции 1,1 kb-фрагмента полимеразой Кленова; и лигирования затупленного по концам 1,1 kb-фрагмента с затупленным по концам экспрессирующего вектора на основе Bruescript, в результате чего получали плазмиду pM'. Поскольку инсерция затупленного по концам 1,1 kb - фрагмента в вектор экспрессии для клетки млекопитающего была проведена в неправильной ориентации и должна поэтому изменить Ampr-ген экспрессирующего Bruescript - вектора, то в присутствии ампициллина будут выживать только колонии HB101 - клеток, трансформированных плазмидной ДНК pM, содержащей затупленный по концам 1,1 kb - фрагмент в правильной ориентации. Плазмидную ДНК очищали от ампициллин-резистивных колоний HB 101-клеток, трансформированных плазмидой pMN, путем равновесного центрифугирования в градиентах хлорида цезия-бромида этидия. 1,6 kb-Eco RI-Bsp HI-фрагмент гена F RSV и 1,7 kb - Bsp HI- Bam HI - фрагмент гена HN PIN-3 были клонированы непосредственно в Eco RI-Bam HI- сайты вектора pM' при 3-этапном лигировании, в результате чего получали плазмиду pM RF-HN.
Для восстановления специфических кодирующих последовательностей генов F RSV и HN PIV-3, удаленных при BspHI - разрезании, кассету BspHI - BspHI - олигонуклеотидов (SEQ ID N 16), содержащую соответствующие RSV-F и PIV-3-HN - генные последовательности, лигировали посредством BspHI - сайта с BspHI - рестриктированной плазмидой pM RF-HN, в результате чего получали плазмиду pM RF-HN. Клоны, содержащие кассету BspHI - BspHI - олигонуклеотидов в правильной ориентации, идентифицировали посредством секвенирования олигонуклеотидного линкера и фланкирующих его областей.
Для клонирования химерного гена FRSV - HNRIV-3 в бакуловирусный экспрессирующий вектор pD2 (пример 8), сначала укороченный ген FRSV -HNPIV-3 выделяли из плазмиды pM RF-HN путем разрезания этой плазмиды ферментом EcoRI. Затем 3,3 kb - ген FRSV -HNPIV-3 клонировали в EcoRI - сайт бакуловирусного плазмидного вектора переноса pD2, в результате чего получали плазмиду pD2 RF-HN (АТСС 75388). Правильная ориентация 3,3 kb-EcoRI - вставки химерного гена FRSV -HNPIV-3 в плазмиде pD2 RF-HN была подтверждена анализом последовательности.
Пример 10
В этом примере описывается получение очищенного методом бляшек рекомбинантного бакуловируса, содержащего химерный ген FRSV -HNPIV-3.
Клетки Spodoptera fruqiperda (Sf9) совместно трасфицировали 1 микрограммами ДНК Ac NPV дикого типа и 2 микрограммами плазмидной FRSV -HNPIV-3 - ДНК (плазмида pD2 RF-HN, пример 9). Предлагаемые рекомбинантные бакуловирусы (очищенные один раз путем серийного разведения), содержащие химерный ген FRSV -HNPIV-3, идентифицировали с помощью дотблот-гибридизации. Лизаты клеток насекомых, инфицированных предлагаемыми рекомбинантными бакуловирусами, зондировали32P-меченными RSV-F- или PIV-3 -HN -генными олигонуклеотидными зондами. Перед использованием в экспериментах по экспрессии рекомбинантные бакуловирусы очищали три раза методом бляшек. Все процедуры проводили в соответствии со схемой, описанной Summers и Smith (пример 6).
Пример 11
В этом примере проиллюстрирован анализ на присутствие химерного FRSV -HNPIV-3 - белка в супернатантах Sf9-High-5 инфицированных
клеток.
Клетки насекомого (Sf9 и High 5), выдерживаемые в бессывороточной среде EX 401, инфицировали очищенными рекомбинантными бакуловирусами (пример 10) при множественности заражения от 5 до 10 БОЕ/кл. Супернатанты от клеток, инфицированных испытуемыми рекомбинантными бакуловирусами, показали положительные результаты в отношении экспрессии белка в обоих RSV-F- и PIV-3-HN-специфических EIISA-анализах. Кроме того, супернатанты от инфицированных клеток обнаруживали положительную реакцию с моноклональным антителом против F RSV и с антисывороткой против пептида HN при иммуноблотировании. В иммуноблотах присутствовала заметная полоса приблизительно 105 кДа. Полученные результаты подтверждают секрецию химерного FRSV-HNPIV-3-белка в супернатанты Sf-9 и High5-клеток, инфицированных рекомбинантными бакуловирусами.
Пример 12
В этом примере описывается очистка химерного FRSV-HNPIV-3
-белка из супернатантов от High 5-инфицированных клеток.
Клетки High 5, поддерживаемые в бессывороточной среде, инфицировали очищенными рекомбинантными бакуловирусами (пример 10) при множественности заражения 5 БОЕ/кл. Супернатанты от инфицированных вирусом клеток собирали через 2 дня после инфицирования. Растворимый FRSV-HNPIV-3 -химерный белок очищали от супернатантов инфицированных клеток посредством иммуноаффинной хроматографии с использованием моноклонального антитела против HN PIV-3. Моноклональное антитело против HN связывали с CN Br-активированной Сефарозой 4B с использованием стандартной техники. Иммуноаффинную колонку промывали 10 объемами слоя промывочного буфера (10 мм Трис-HCl, pH 7,5, 150 мМ NaCl, 0,02% (объем/объем) Тритон-Х 100) перед ее использованием. После загрузки образца колонку промывали 10 объемами слоя промывочного буфера, а затем 3 объемами слоя высококонцентрированного солевого буфера (10 мм Трис-HCl, pH 7,5, 500 мМ NaCl, 0,02% (об/об.) Тритон-Х 100). Химерный FRSV-HNPIV-3-белок элюировали из иммуноаффинной колонки глицином (100 мМ, pH 2,5) в присутствии 0,02% Тритона Х-100. Элюированный белок сразу нейтрализовали 1М Трис-HCl, pH 10,7.
Анализ с помощью электрофореза в полиакриламидном геле (фиг. 16, дорожка A) FRSV-HNPIV-3-белка, очищенного посредством иммуноаффинной хроматографии, выявил присутствие одной полосы мажорного белка с кажущейся молекулярной массой 105 кДа. При иммуноблотировании (фиг. 16, панель B, дорожки 1 и 2 соответственно).
Пример 13
В этом примере проиллюстрирован анализ на иммуногенность FRSV-HNPIV-3-белка в морских свинках.
Группам из четырех морских свинок инъецировали внутримышечно либо 1,0 либо 10,0 мкг химерного FRSV-HNPIV-3-белка, очищенного как описано в примере 12, и дополненного адъювантом, а именно фосфатом алюминия. Группы контрольных животных иммунизировали либо плацебо, либо "живым" вирусом PIV-3 или RSV (через нос). У животных брали кровь через 2 и 4 недели после первичной инъекции и через 4 недели вводили вторичную инъекцию антигенного препарата в эквивалентной дозе. Образцы сыворотки также брали через 2 и 4 недели после введения вторичной дозы. Для оценки способности химерного белка продуцировать ответное образование PIV-3 и RSV-специфических антител образцы сыворотки анализировали на присутствие PIV-3-специфических антител, ингибирующих и нейтрализующих гемагглютинацию, а также RSV-нейтрализующих антител. Как показано в таблице 1 (таблицы приводятся в конце описания), сыворотки животных, иммунизированных двумя 10 мкг - дозами химерного белка, имели титры PIV-3-специфического ингибирования гемаглютинации (HAI) и титры PIV-4/RSV-нейтрализующих антител (через 6 и 8 недель), которые были эквивалентны уровням, полученным после интраназальной инокуляции либо живым PIV-3, либо живым RSV. Кроме того, у животных, иммунизированных лишь двумя 1-мкг-дозами химерного белка, наблюдалось сильное продуцирование PIV-3 и RSV-специфических нейтрализующих антител. Полученные результаты подтвердили иммуногенность обоих компонентов (RSV и PIV-3) химерного белка и явились неоспоримым свидетельством того, что единый рекомбинантный иммуноген может продуцировать нейтрализующие антитела против обоих вирусов RSV и PIV-3.
Пример 14
В этом примере проиллюстрирован анализ на иммуногенность FRSV-HNPIV-3-белка в хлопковых хомяках.
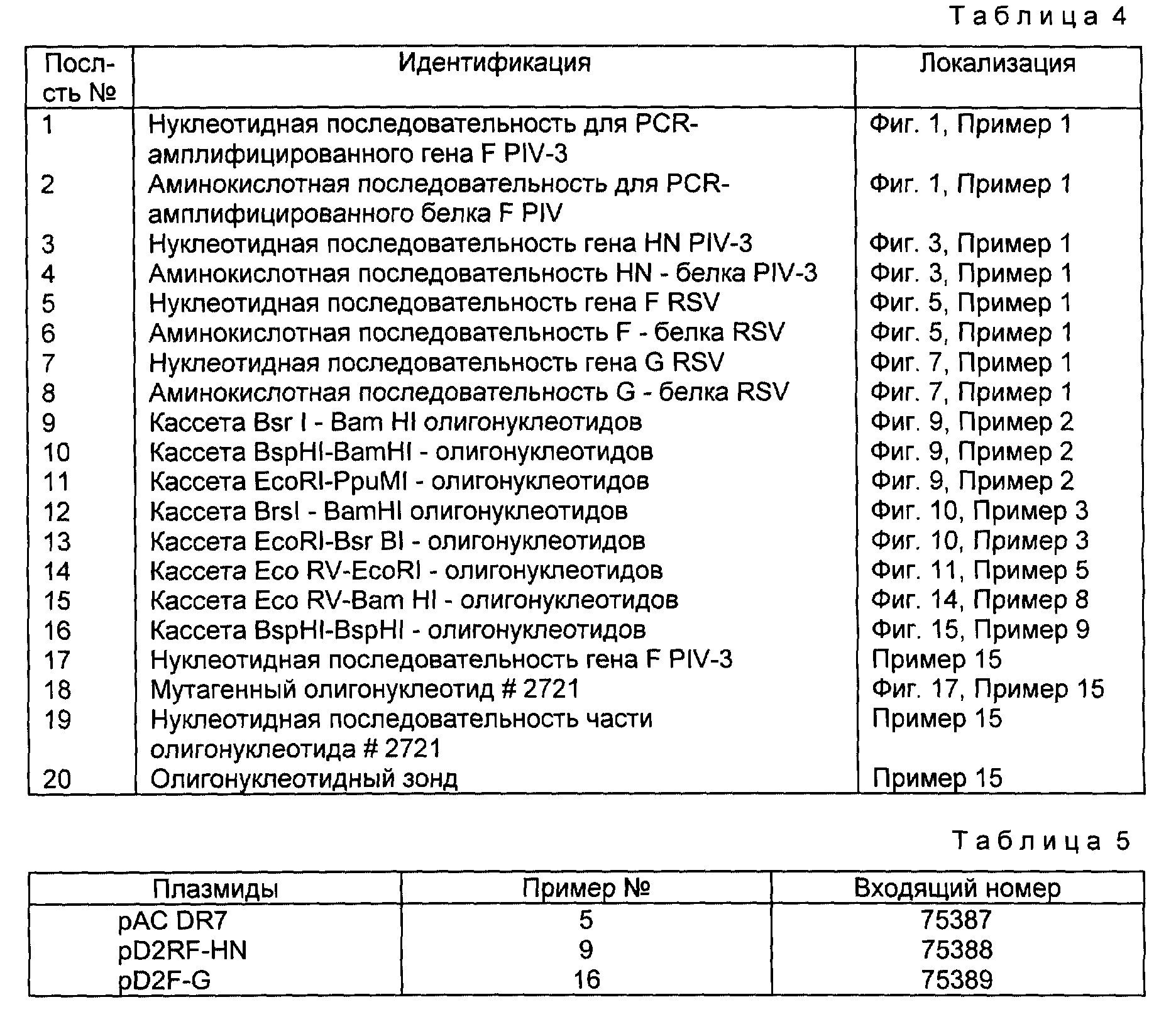
Группам из восьми хлопковых хомяков инъецировали внутримышечно либо 1,0, либо 10,0 мкг FRSV-HNPIV-3-химерного белка, очищенного как описано в примере 12, и дополненного фосфатом алюминия. Группы контрольных животных иммунизировали либо плацебо (SPB+фосфат алюминия), либо живым вирусом PIV-3 или PSV (вводимого через нос). У животных брали кровь через 4 недели после первичной инъекции и через 4 недели вводили вторичную эквивалентную дозу антигенного препарат. Образцы сыворотки брали также через 1 неделю после введения вторичной дозы. Как показано ниже в таблице 2, данные, взятые через 4 недели после инъекции, свидетельствуют о том, что обе 1 и 10 мкг - дозы химерного белка способны индуцировать сильный первичный иммунный ответ. Обратная величина log2 титров PIV-3-специфического ингибирования гемагглютинации (HAI) и PIV-3/RSV-нейтрализующих титров была эквивалентна титрам, полученным с использованием живого PIV-3 и RSV. Таким образом, одной инокуляции химерного белка было достаточно для продуцирования нейтрализующих антител против PIV-3 и RSV. Высокие PIV-3 и RSV-нейтрализующие титры также наблюдались после введения вторичной дозы (взятие пробы через 5 недель). Эти результаты служат еще одним свидетельством того, чтоб RSV- и PIV-3-компоненты химерного белка являются в высокой степени иммуногенными.
Для оценки способности химерного иммуногена к одновременной защите животных от заражения вирусами RSV и PIV-3 четырех хлопковых хомяков от каждой группы подвергали контрольному заражению (через нос) либо вирусом PIV-3, либо вирусом RSV при средней цитопатогенной дозе (TCID50) 100 ед. Через 4 дня после заражения животных забивали. Вирусные титры определяли в гомогенатах легких. Как показано ниже, в таблице 3, животные, иммунизированные либо 1, либо 10 мкг химерного FRSV-HNPIV-3-белка, были абсолютно невосприимчивы к заражению либо PIV-3PIV-3, либо RSV. Полученные результаты свидетельствуют о том, что химерный белок является не только в высокой степени иммуногенным, но и также способен обеспечить защиту хлопковых хомяков одновременно от заболеваний, вызываемых вирусами PIV-3 и RSV.
Пример 15
В этом примере проиллюстрировано конструирование Bluescript M13-SK-вектора, содержащего химерный FPIV-3-GRSV-ген. Эта генная
конструкция содержит 5'-нетранслируемую область мутированного PIV-3-гена, но не содержит нуклеотидную последовательность, кодирующую гидрофобный домен и цитоплазматический концевой домен мутированного
PIV-3-F-гена и нативного RSV-G-гена. Стадии конструирования этой плазмиды показана на фиг. 17 и 18.
В первой стадии (фиг. 17) для получения PIV-3-F-компонента химерного гена FPIV-3-GRSV удаляют предполагаемые предтерминационные сайты внутри 18-нуклеотидной последовательности 5'CAAGAAAAAGGAATAAAA 3' (SEQ ID N 17), расположенной между положениями 857 и 874 не-PCR-амплифицированного PIV-3-F-гена и положениями 847 и 864 PCR-амплифицированного PIV-3-F-гена (см. фиг. 1). А затем PIV F-кДНК не PCR-амплифицированного PIV-3-F-гена разрезали в BsaAI - и EcoRI - сайтах. BsaAI- EcoRI-фрагмент PIV-3-F-гена клонировали в EcoRI-сайт - в Bluescript M13-SK-вектор с использованием EcoRI-BsaAI-линкера. Затем 857-874 - область - мишень PIV-3-F-гена (не-PCR-амплифицированного) подвергали мутации с помощью сайтнаправленного мутагенеза с использованием олигонуклеотидов и в соответствии с методом, описанным Morinaga и др. (1984, Biotechnology, 2: 636-639). Плазмиду pP13Fc (пример 1) разрезали ферментом Scal в Ampr-гене, и дефосфорилировали щелочной фосфатазой (плазмида #1). Второй образец плазмиды pP13Fc разрезали ферментами Bst EII и Nsil, в результате чего получали рестриктированную 3,9-ко-плазмиду, в которой отсутствовал 0,9 kb-BstEII-Nsil-фрагмент гена PIV-3-F (плазмида #2). Для специфического мутирования 857-874-сегмента ДНК без изменения последовательности F-белка синтезировали мутагенный 78-мерный синтетический олигонуклеотид (# 2721; показан на фиг. 17, SEQ ID N 18), содержащий последовательность 5'CAGGAGAAGGGTATCAAG 3' (SEQ ID N 19).
Этот олигонуклеотид добавляли к плазмидной ДНК #1 и #2, денатурировали при 100oC в течение 3 минут и ренатурировали путем постепенного охлаждения. После этого смесь инкубировали в присутствии ДНК-полимеразы, dNTP и T4-лигазы и трансформировали в клетки HB101. Бактерии, содержащие мутированный 1,8 kb-ген F PIV-3, выделяли на УТ-агаровых чашках, содержащих 100 мкг/мл ампициллина. Для подтверждения присутствия мутированного PIV-3-F-гена осуществляли гибридизацию с олигонуклеотидным зондом 5'AGGAGAAGGGTATCAAG 3' (SEQ ID N 20). Присутствие мутированной последовательности гена было подтверждено с помощью ДНК-секвенирования. Плазмиду, содержащую мутированный PIV-3-ген, обозначали pP13Fm.
Вторая стадия (фиг. 18) конструирования химерного гена заключалась в получении вектора на основе Bluescript так, чтобы он содержал укороченный PIV-3-F-ген, в котором отсутствует нуклеотидная последовательность, кодирующая трансмембранный якорный домен и цитоплазматическую концевую область PIV-3-F-белка, и который связан тандемом с RSV-G-геном, не содержащим 5'-лидерную последовательность и нуклеотидную последовательность, кодирующую трансмембранный якорный домен и цитоплазматический концевой домен гликопротеина G.
Для получения этого химерного гена ориентацию мутированного PIV-F-гена в плазмиде pP13Fm сначала "перевертывали" посредством EcoRI-переваривания, а затем снова лигировали, в результате чего получали плазмиду pP13Fmr. Для получения PIV-3-F-компонента химерного гена плазмиду pP13 Fmr разрезали ферментами Not1 и Bsr1 в целях выделения укороченного 1,7 kb-гена PIV-3-F. Для получения RSV-G-компонента, 0,95 kb-ген с RSV, в котором отсутствовали 5'-лидерная последовательность и ДНК-сегмент, кодирующий "якорный" домен белка G и цитоплазматический концевой домен, выделяли из плазмиды pRSVG (пример 1) путем отрезания полилинкера ферментом EcoRI и гена ферментом BamHI. EcoRI-BamHI-фрагмент (0,95 kb) гена G RSV субклонировали в EcoRI-BamHI-сайты рестриктированного Bluescript вектора pM13-SK, в результате чего получали плазмиду pRSVG. EcoRI-BamHI-фрагмент (0,95 kb) гена G и NotI-BstI-фрагмент (1,5 kb) укороченного гена F PIV-3 сшивали с помощью кассеты BstI-BamHI-олигонуклеотидов (SEQ ID N 9). Восстанавливая при этом F и G-генные кодирующие последовательности, а затем клонировали в вектор pRSVG, рестриктированный ферментами BamHI и NotI при 3-этапном лигировании. Полученную таким образом плазмиду обозначали pFG.
Пример 16
В этом примере описывается конструирование бакуловирусного вектора переноса pD2 (описанного в Примере 8), содержащего химерный FPIV-3-GRSV-ген, состоящий из
мутированного гена F PIV-3, в котором отсутствуют области, кодирующие гидрофобный домен и цитоплазматический домен, и который сцеплен с геном G RSV, не содержащим 5'-лидерную последовательность и
нуклеотидные последовательности, кодирующие трансмембранный якорный домен и цитоплазматический концевой домен белка G.
Для получения этой конструкции плазмиду pFG (пример 15) разрезали ферментом EcoRI и получали химерный ген (1,6 kb) FPIV-3-GRSV. Затем 2,6 kb-фрагмент EcoRI-рестриктированного химерного гена субклонировали в EcoRI-сайт дефосфорилированного вектора pD2, в результате чего получали 12,1 kb плазмиду pD2 F-G (ATCC 75389).
Пример 17
В этом примере описывается получение очищенного методом бляшек
рекомбинантного бакуловируса, содержащего химерный FPIV-3-GRSV-ген.
Клетки Spodoptera frugiperda (Sf9) совместно трансфицировали 1 микрограммами pD2F-G-плазмидной ДНК (пример 16) и 1 микрограммами линейной ДНК AcNPV дикого типа (полученной от фирмы Invitrogen). Рекомбинантные бакуловирусы, содержащие FPIV-3-GRSV-ген, дважды очищали методом бляшек в соответствии с процедурой, описанной в примере 10.
Пример 18
В этом примере проиллюстрирован анализ на присутствие химерного FPIV-3
-GRSV-белка в супернатантах клеток Sf9 и High5, инфицированных рекомбинантными бакуловирусами.
Клетки Sf9 и High5 были инфицированы рекомбинантными бакуловирусами, содержащими FPIV-3-GRSV-ген (пример 16), при множественности инфекции от 5 до 10 БОЕ/кл. Супернатанты, полученные от клеток, инфицированных рекомбинантными вирусами, показали положительную реакцию при специфическом ELISA-анализе на экспрессированный белок F PIV-3. Супернатанты от инфицированных клеток реагировали с моноклональными антителами против F PIV-3 и против G RSV при иммуноблоттинге. Полученные результаты подтвердили присутствие химерного FPIV-3-GRSV белка в супернатантах инфицированных клеток Sf9 и High5.
Пример 19
В этом примере описывается получение рекомбинантных вирусов осповакцины, содержащих гены FPIV-3 - FRSV и FRSV - HNPIV-3.
Рекомбинантные вирусы осповакцины, экспрессирующие гены FPIV-3 - FRSV (обозначенный vP1192) и ген FRSV - HNPIV-3 (обозначенный vP1195) продуцировали в Корпорации по вирусной генетике (Virogenetics Corporation, Troy, NY) (самостоятельное юридич. лицо, относящееся к его правоприемнику) с использованием селекционной системы COPAK с несколькими хозяевами. В указанной системе COPAK, содержащей ген коровьей оспы KIL с несколькими хозяевами (Perkus и др. (1990), Virology 179: 276 - 286) и модифицированный промотор вируса коровьей оспы H6 (Perkus и др. (1989), J. Virology 63: 3829 - 3836) использовали инсерционные плазмиды. В этих инсерционных плазмидах ген KIL, промотор H 6 и полилинкерная область расположены между франкирующими плечами штамма Copenhagen коровьей оспы вместо ATI-области (открытые рамки считывания (OPF) A25L, A26L, Goebel и др., (1990), Virology 179: 247 - 266; 517 - 563). Инсерционные плазмиды COPAK конструировали для использования в in vivo-рекомбинации с помощью вируса "спасения" NYVAC (P866) (Tartaglia и др. (1992), Virology 188: 217 - 232). Селекцию рекомбинантных вирусов осуществляли на клетках почки кролика.
Рекомбинантные вирусы vP1192 и vP1195 генерировали с использованием инсерционных плазмид pES 229A-6 и PSD.RN соответственно. Для получения плазмиды pES 229A-6, содержащей FPIV-3 - FRSV-ген, инсерционную плазмиду (COPAK-H6) pSD555 разрезали ферментом Smal и дефосфорилировали кишечной щелочной фосфатазой. FPIV-3 - FRSV-ген (2,6 кb) выделяли из плазмиды pES60A (пример 4) путем разрезания плазмиды ферментами EcoRI и BamHI. Eco RI-Bam HI-ген (2.6 кb) FPIV-3 - FRSV затупляли по концам с помощью полимеразы Кленова, выделяли из легкоплавкого агарозного геля и клонировали в Smal-сайт инсерционной плазмиды (COPAK-H6) pSD555, в результате чего получали плазмиду pES 229A-6. При этом положение OPC FPIV-3 - FRSV таково, что 5'-конец находится ближе всего к промотору H6.
Для получения плазмиды PSD.RN.pSD555-вектор сначала разрезают ферментами Smal и BamHI. Плазмиду pM RF-HN (пример 9), содержащую укороченный FRSV - HNPIV-3-ген, разрезали ферментом Clal, затупляли по концам с помощью полимеразы Кленова, а затем разрезали ферментом BamHI. FRSV - HNPIV-3-ген (3,3 кb) клонировали в Smal-BamHI-сайты вектора pSD555, в результате чего получали плазмиду PSD.RN.OPC. FRSV - HNPIV-3 располагалась так, что 5'-конец H6 находился ближе всего к промотору H6.
Плазмиды pES229A-6 и PSD.RN использовали в экспериментах по in vitro рекомбинации в клетках с использованием NYVAC (vP866) в качестве вируса "спасения". Вирус рекомбинантного потомства отбирали на клетках почки (PK) - 13 кролика (АТСС # CCL 37). Несколько бляшек два раза пассировали на клетки PK-13. Вирусы, содержащие химерные гены, идентифицировали стандартным методом in situ гибридизации бляшек (Piccini и др. (1987), Methods in Enzymology 153: 545 - 563) с использованием радиоактивно меченных зондов, специфичных к PIV - и RSV - инсертированных ДНК-последовательностей. Очищенные методом бляшек вирусы, содержащие химерные гены FPIV-3 - FRSV и FRSV - HNPIV-3, обозначали vP1192 и vP1195 соответственно.
Для подтверждения экспрессии химерных генов в vP1192 и vP1195 - инфицированных клетках проводили радиоиммунопреципитацию. Эти анализы осуществляли с использованием лизатов, полученных от инфицированных клеток Vero (в соответствии с процедурой, описанной Tay lor и др., (1990), J. Virology 64, 1441 - 1450), и с использованием моноспецифической антисыворотки морских свинок против HN PIV-3 и F PIV-3, и кроличьей антисыворотки против F RSV. Обе антисыворотки (против F PIV-3 и против F RSV) осаждали белок кажущейся молекулярной массой приблизительно 90 кДа из vP1192-инфицированных клеток Vero. Обе антисыворотки (морской свинки против PIV-HV и кроличьей против RSV-F) осаждали белок с кажущейся молекулярной массой приблизительно 100 кДа из vP1195-инфицированных клеток. Полученные результаты подтвердили продуцирование химерных FPIV-3 - FRSV - и FRSV - HNPIV-3-белков в клетках Vero, инфицированных рекомбинантными поксвирусами.
Краткое резюме
Резюмируя, можно сказать, что настоящее изобретение относится к мультимерным
гибридным генам, кодирующим химерные белки, способные обеспечивать иммунную защиту против заражения, вызываемого множеством патогенов, в частности вирусами PIV и RSV. При этом возможны модификации, не
выходящие за рамки объема настоящего изобретения.
Реферат
Гибридный белок предназначен для приготовления вакцины против вирусных заболеваний верхних дыхательных путей. Гибридный белок содержит иммуногенный белок или его фрагмент вируса парагриппа типа 3(ПГВ-3), связанный с иммуногенным белком или его фрагментом респираторно-синцитиального вируса типа А(РСВ). Предложены также гибридный ген, кодирующий гибридный белок, способ получения гибридного белка и вакцина против заболеваний верхних дыхательных путей, содержащая гибридный белок. Предположены бакуловирусные векторы экспрессии рАC DR7, pD2RF-HN, pD2F-G, обеспечивающие экспрессию гибридного белка. Преимуществом настоящего изобретения является получение единых иммуногенов, содержащих претективные антигены, что упрощает разработку комбинированных вакцин. 7 с. и 5 з.п. ф-лы , 18 ил., 5 табл.

Комментарии