Белки шелка - RU2467015C2
Код документа: RU2467015C2
Чертежи



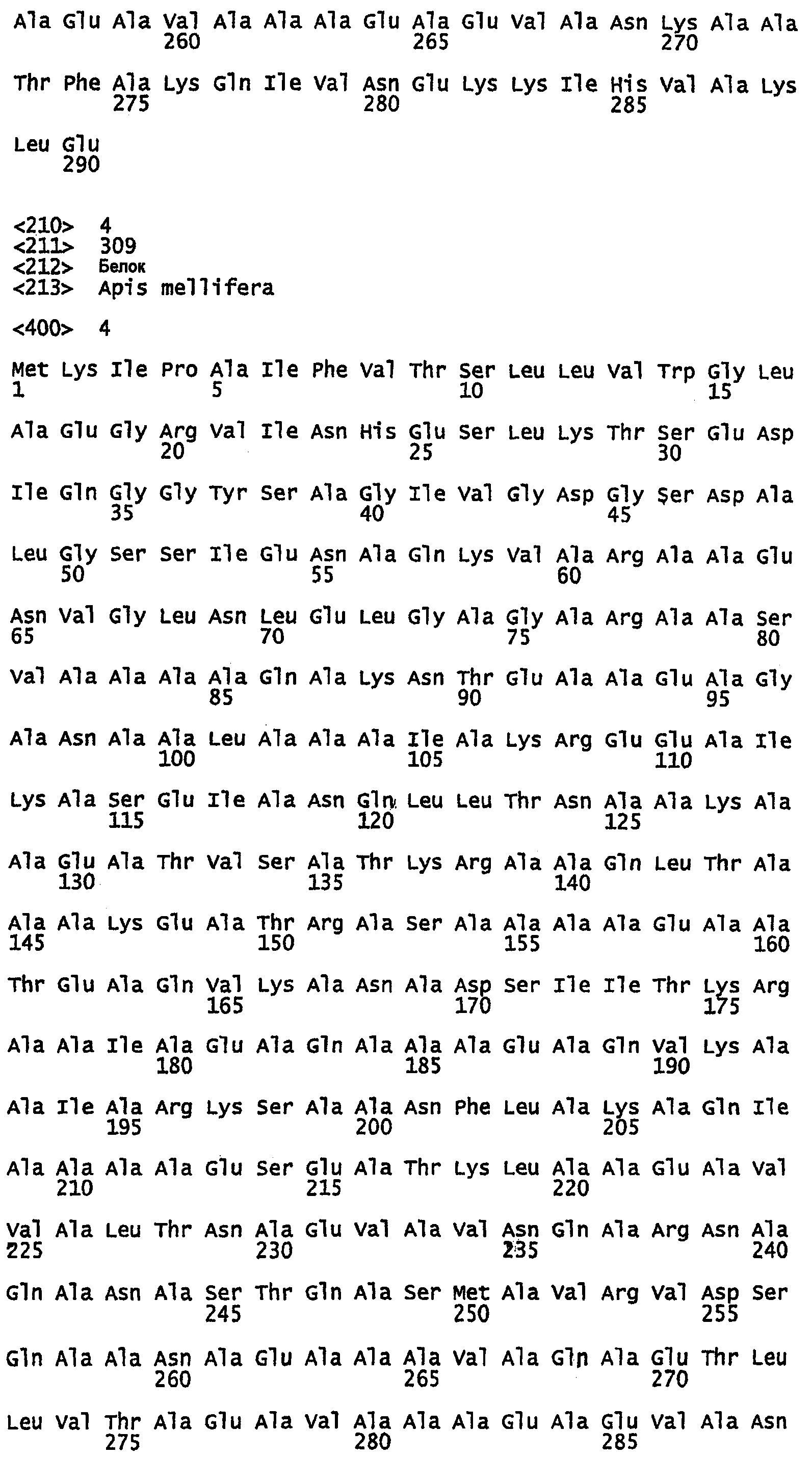
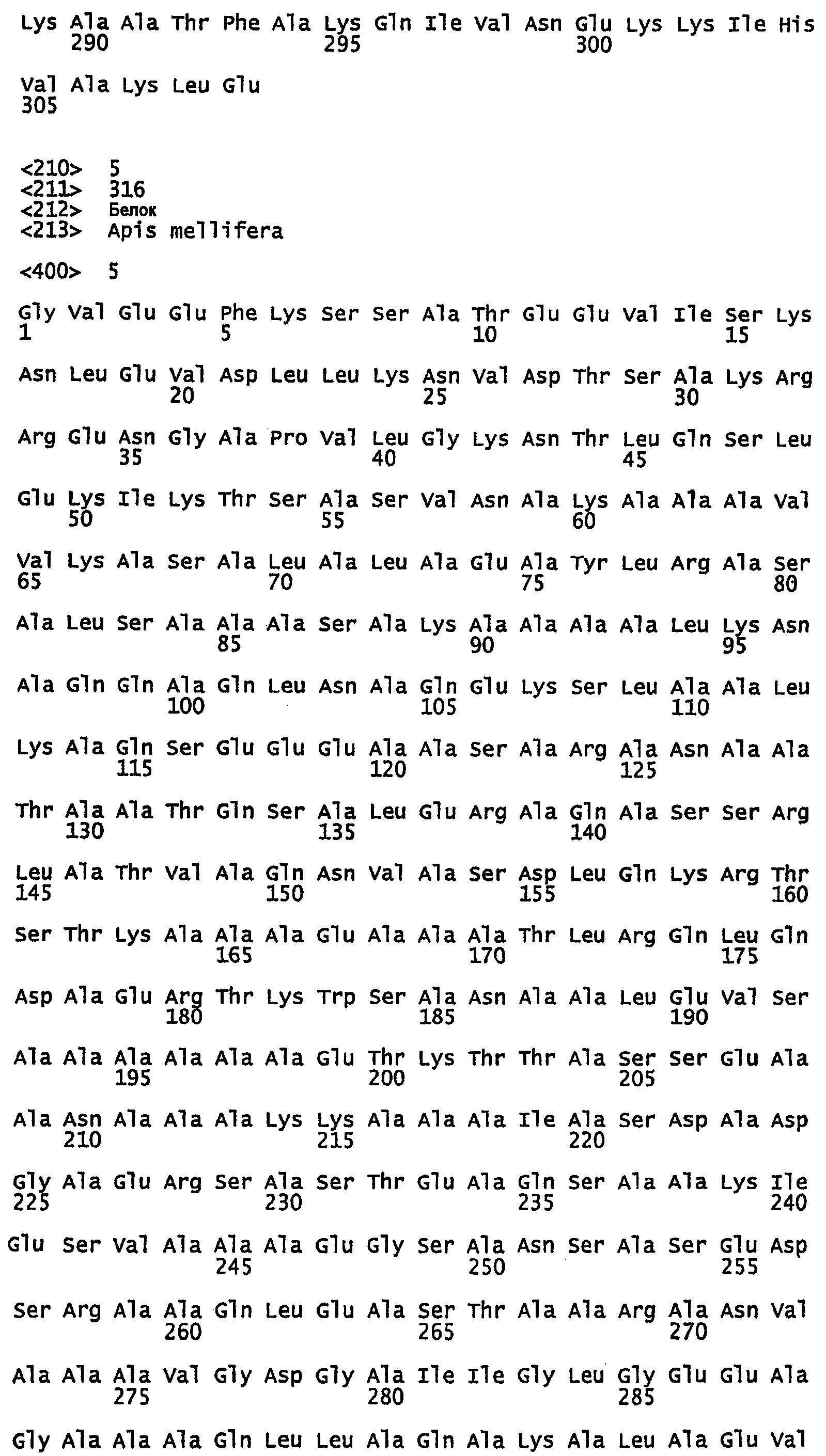
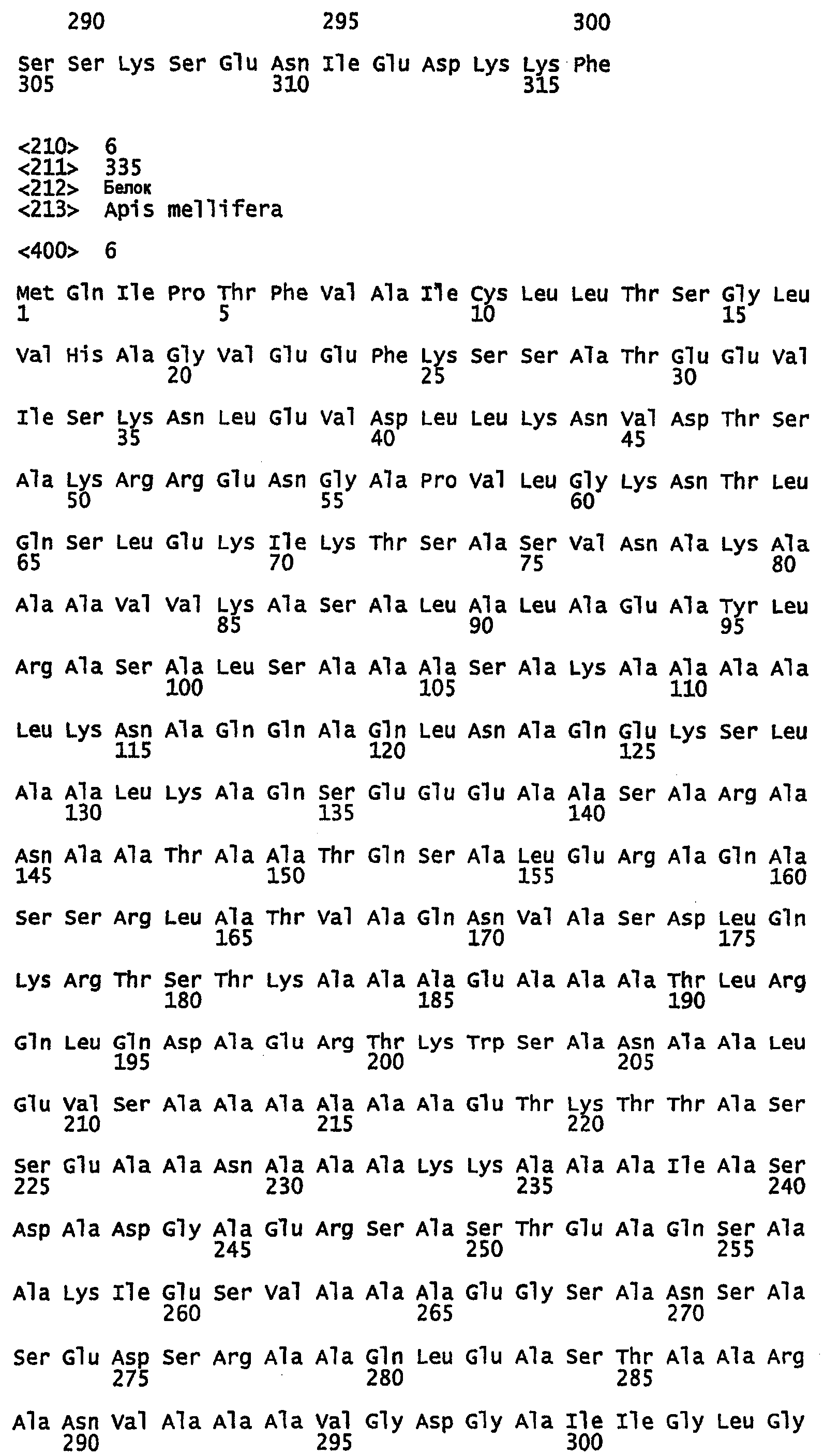
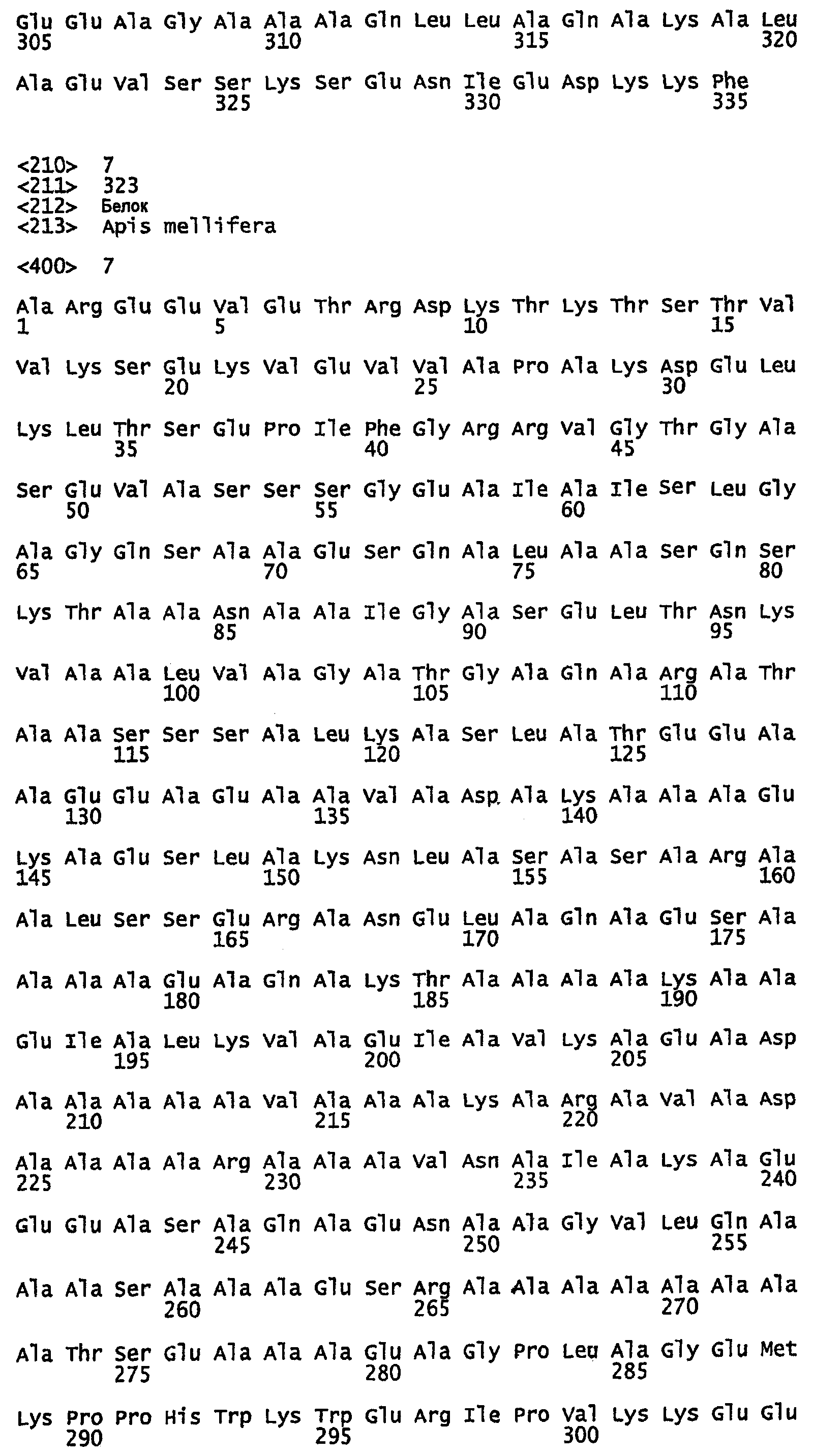








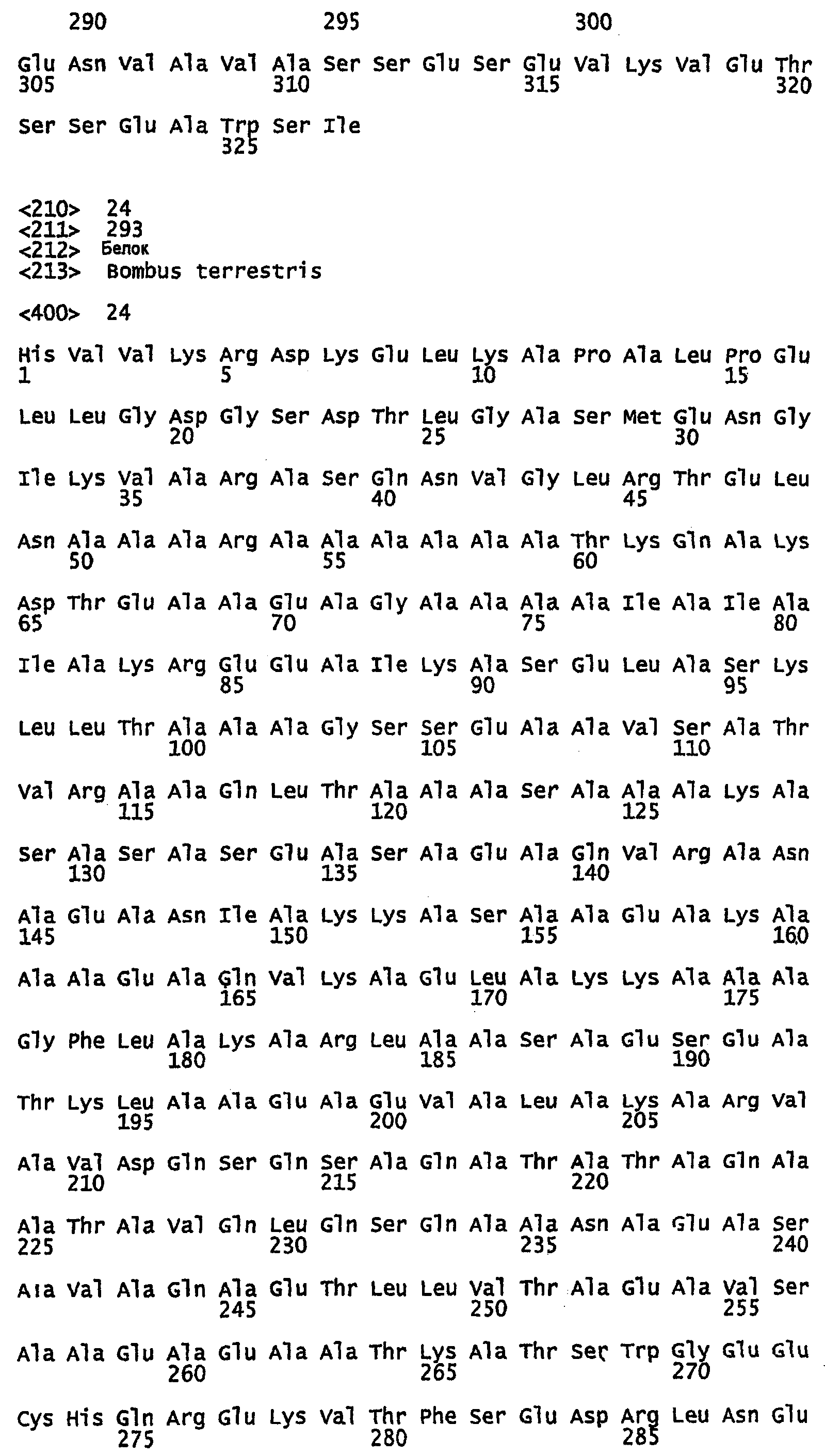
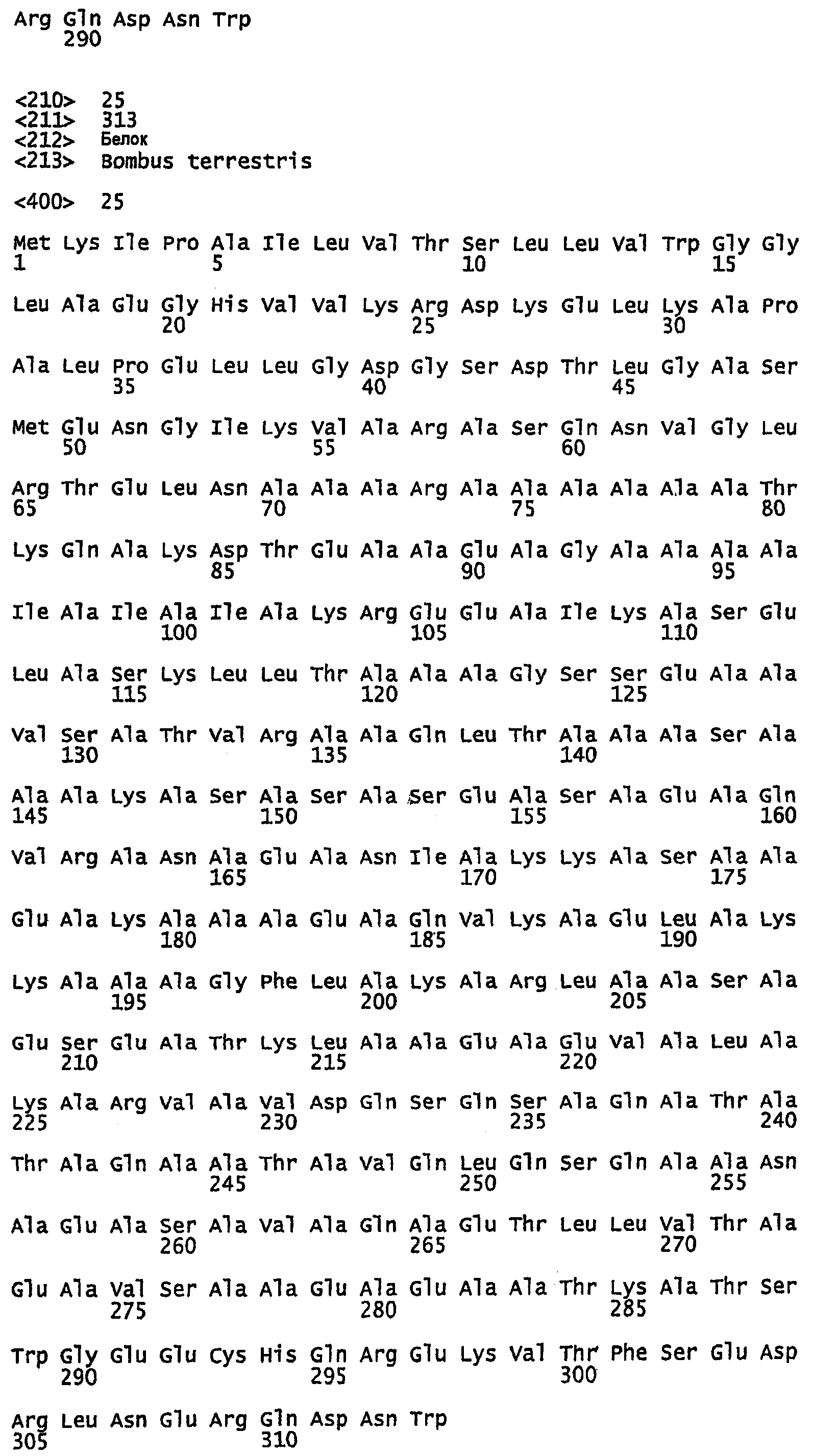
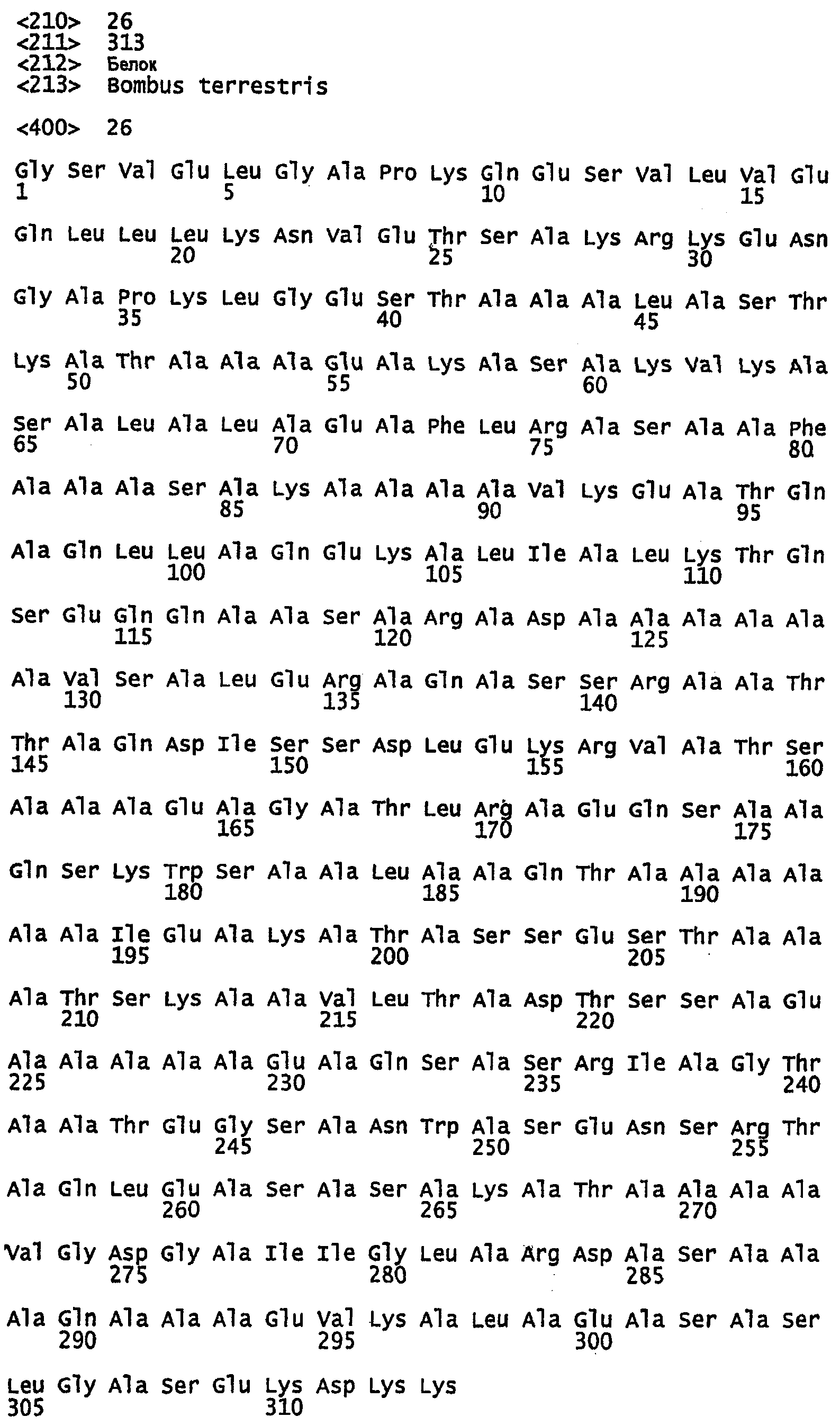
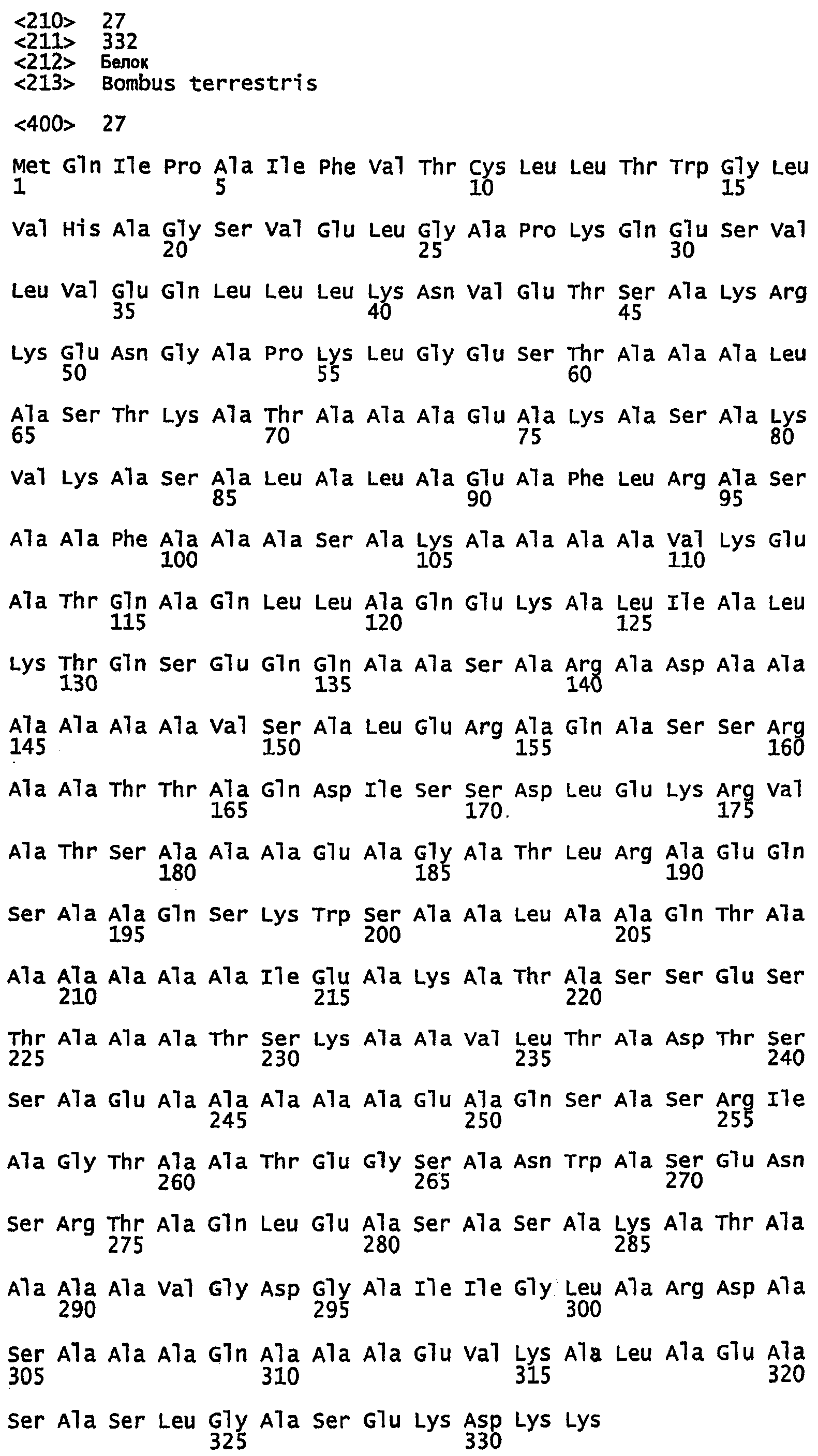
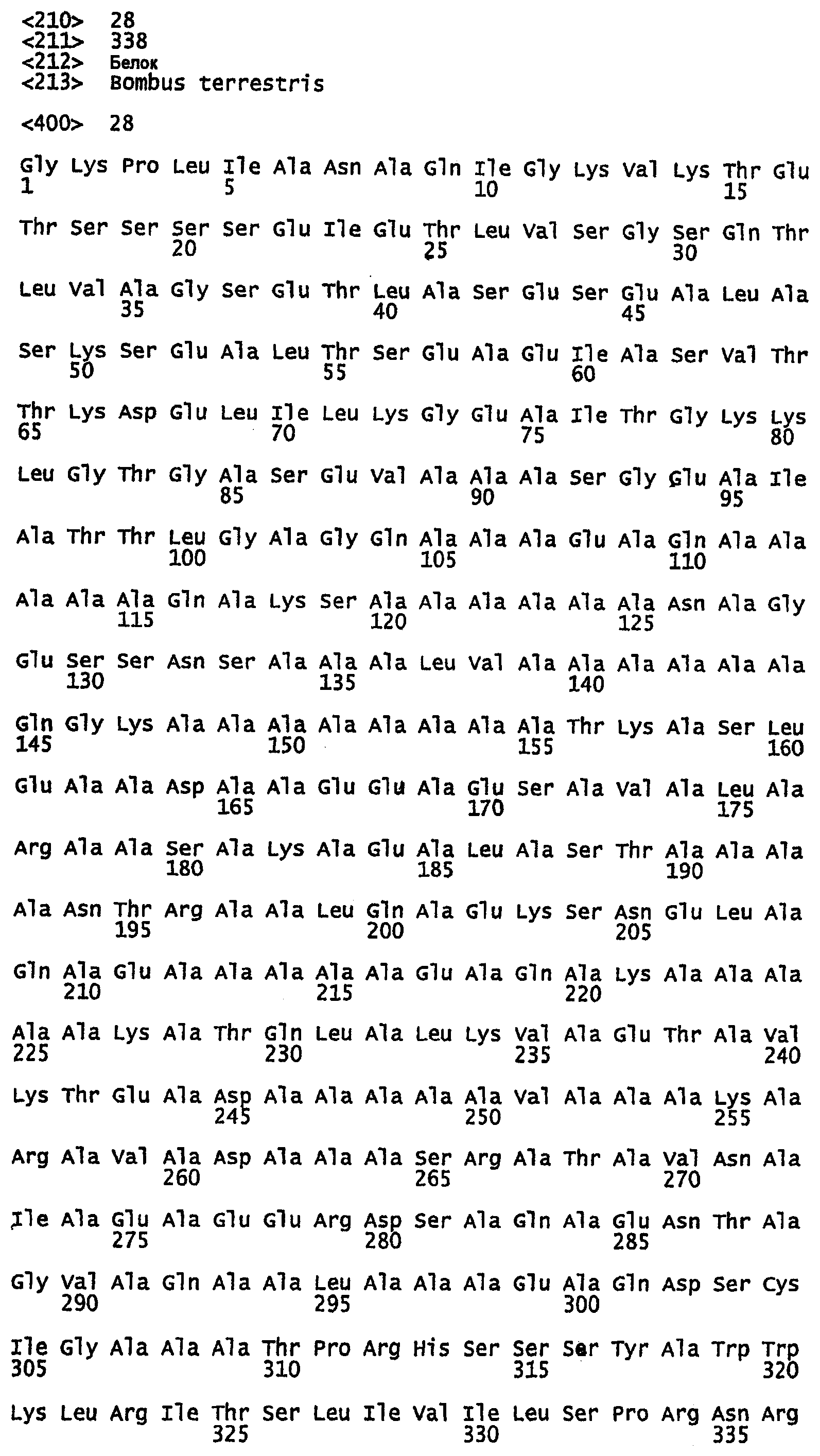
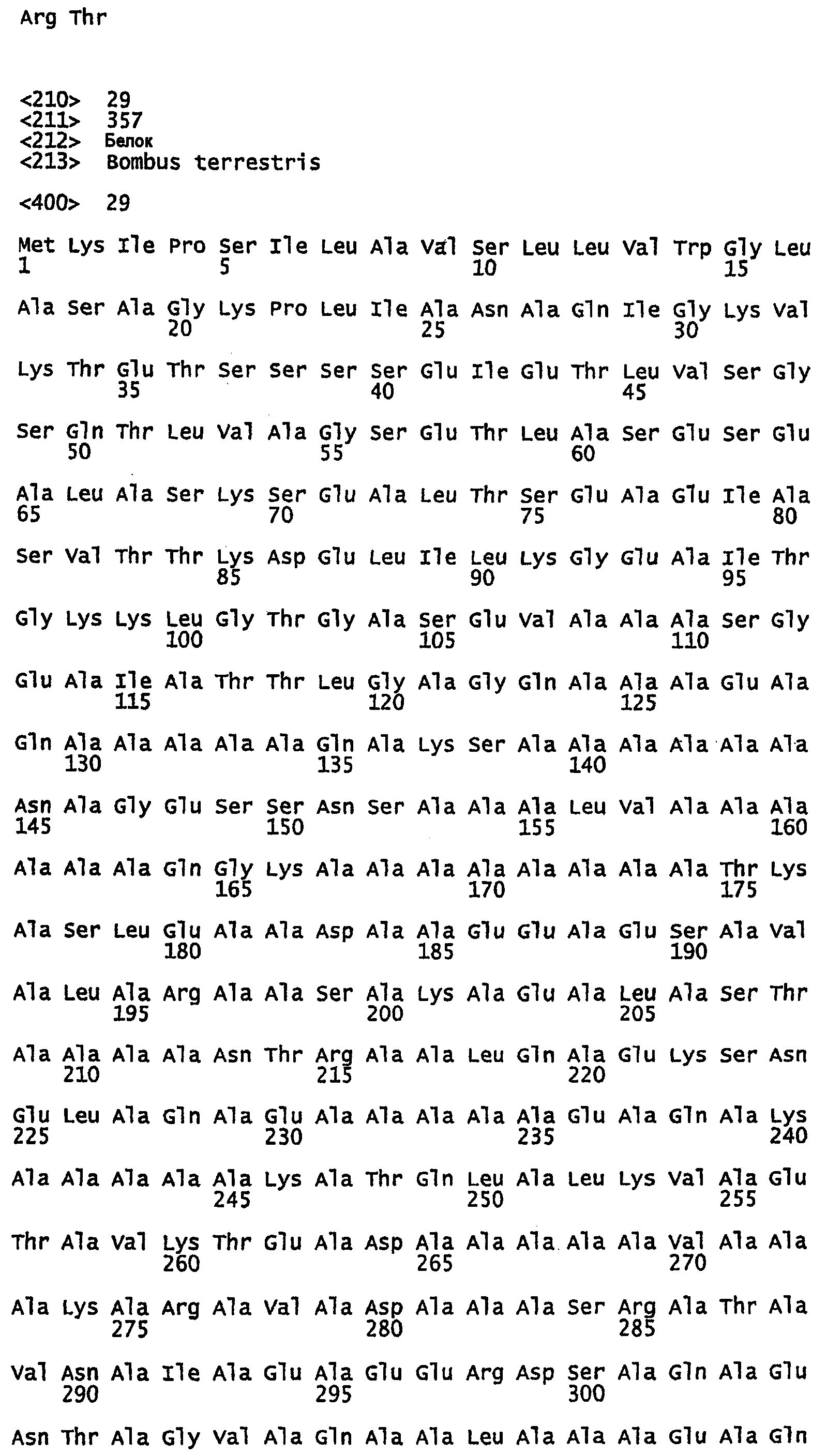
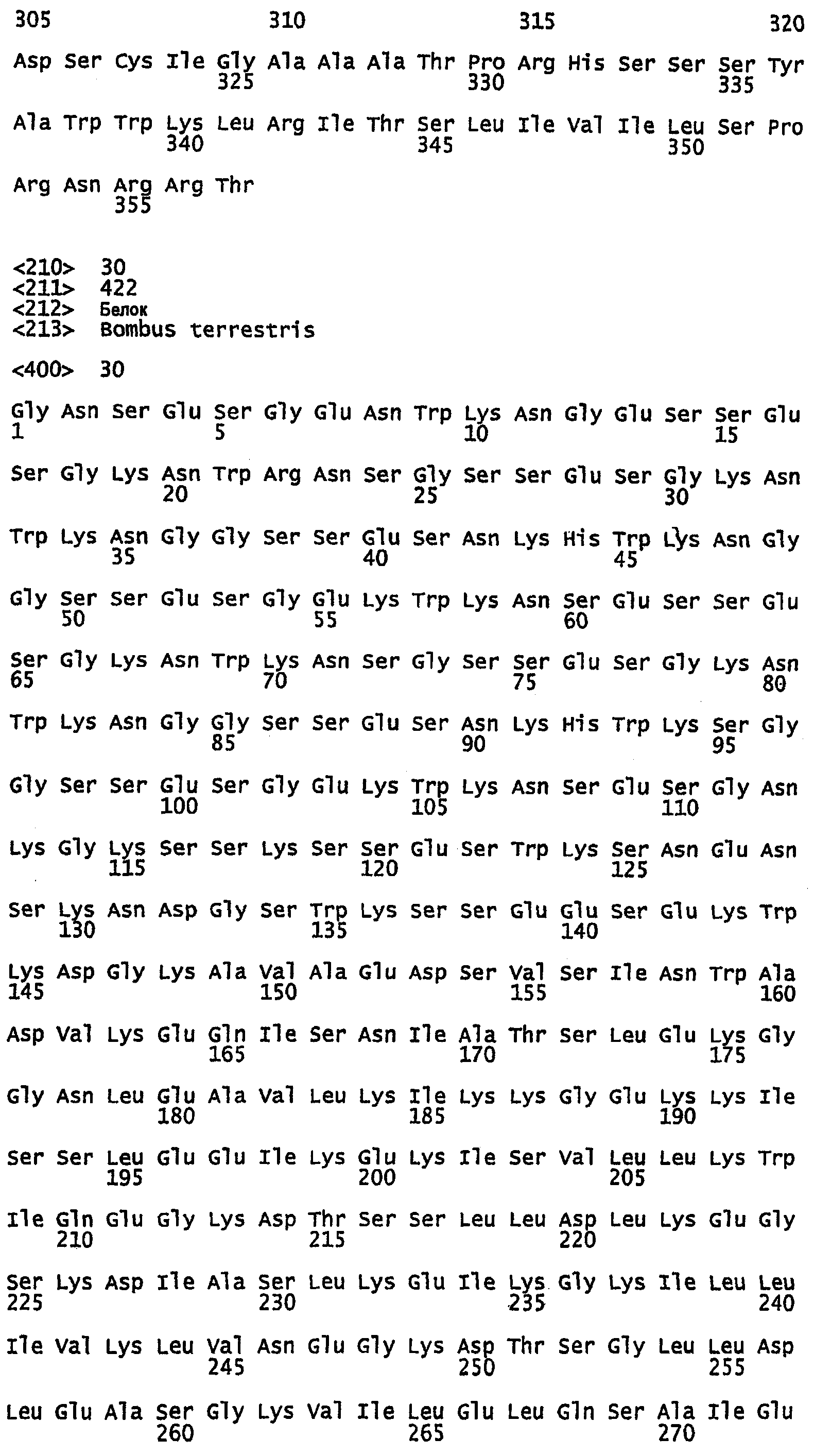





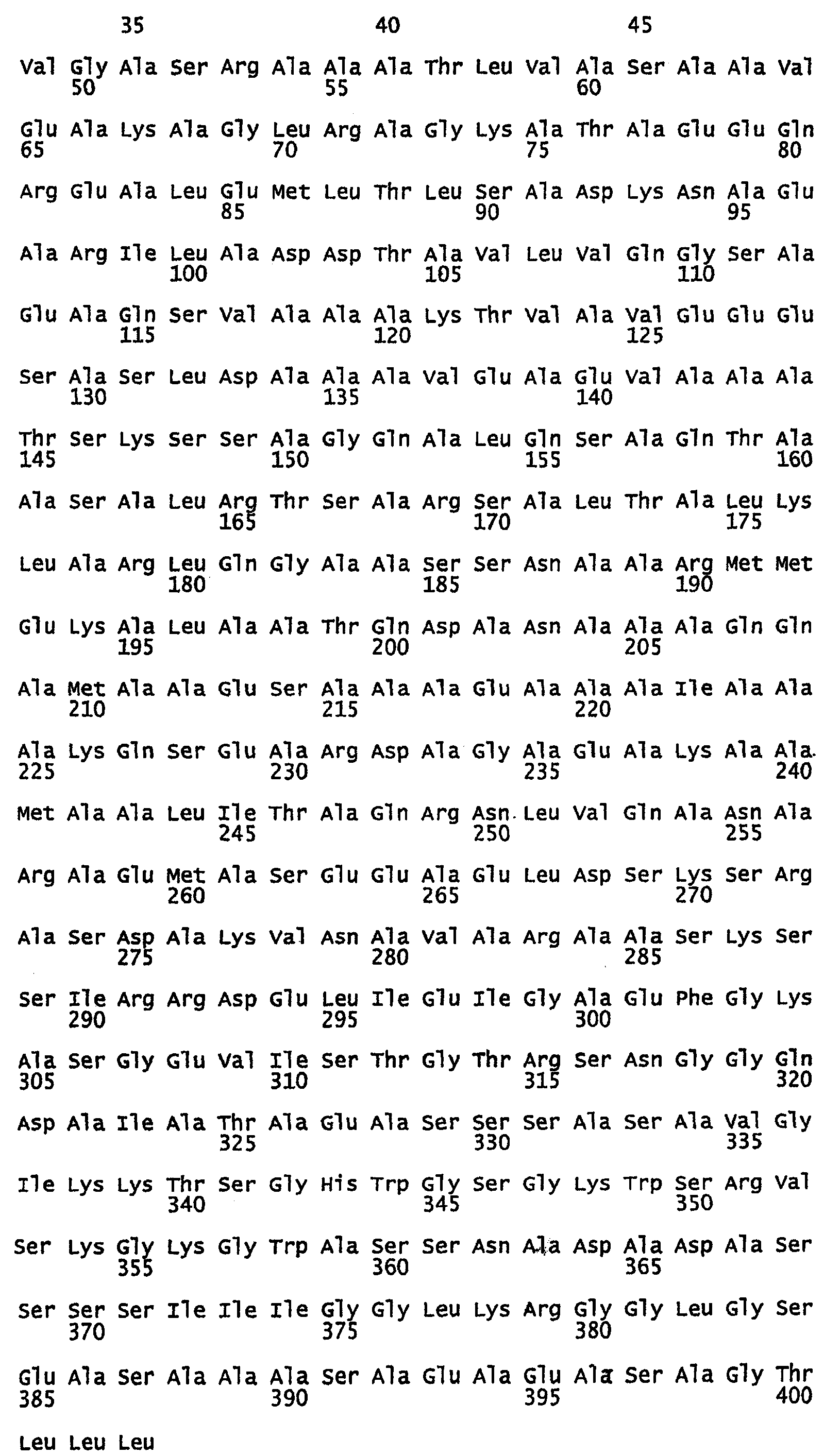
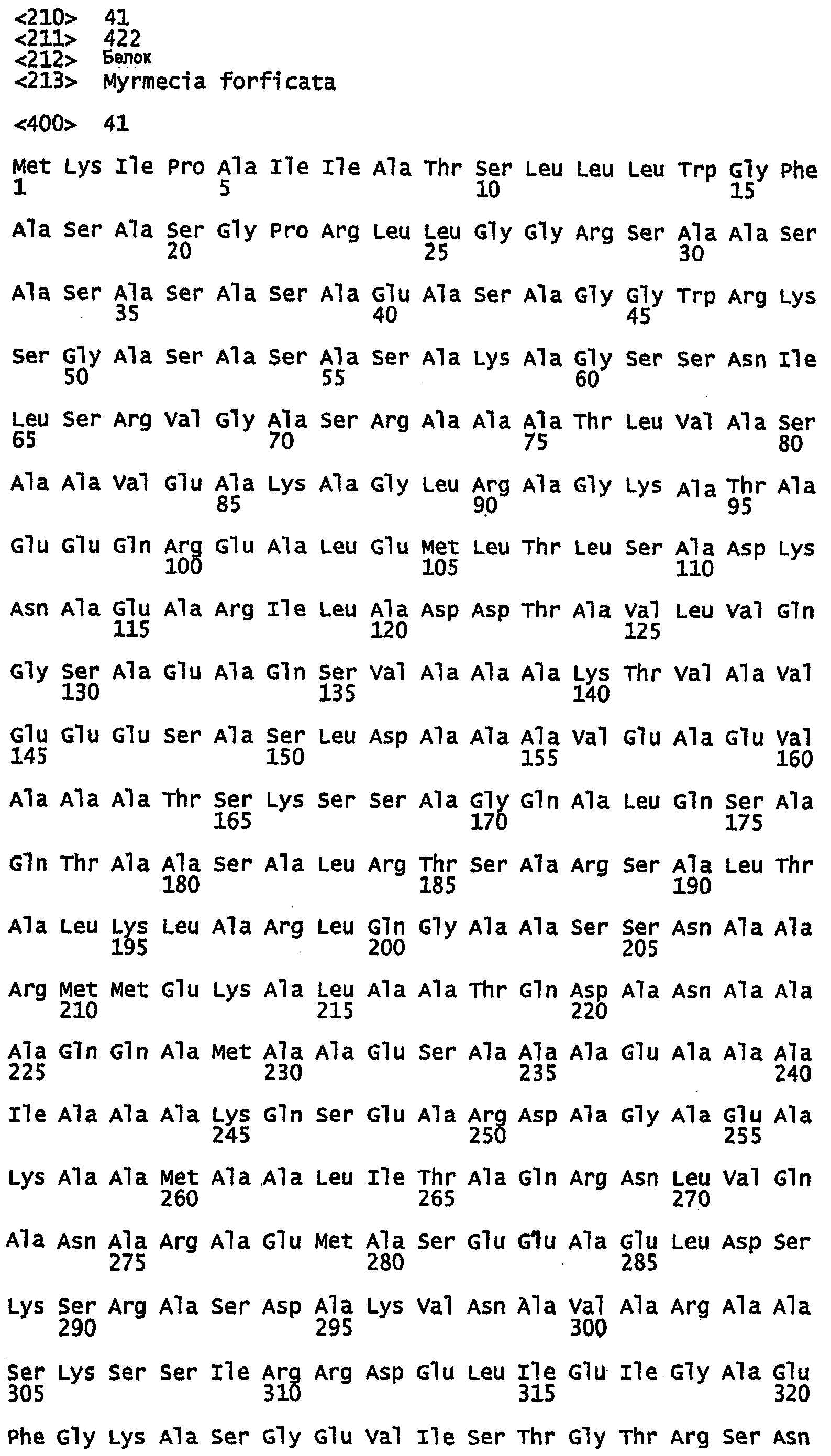



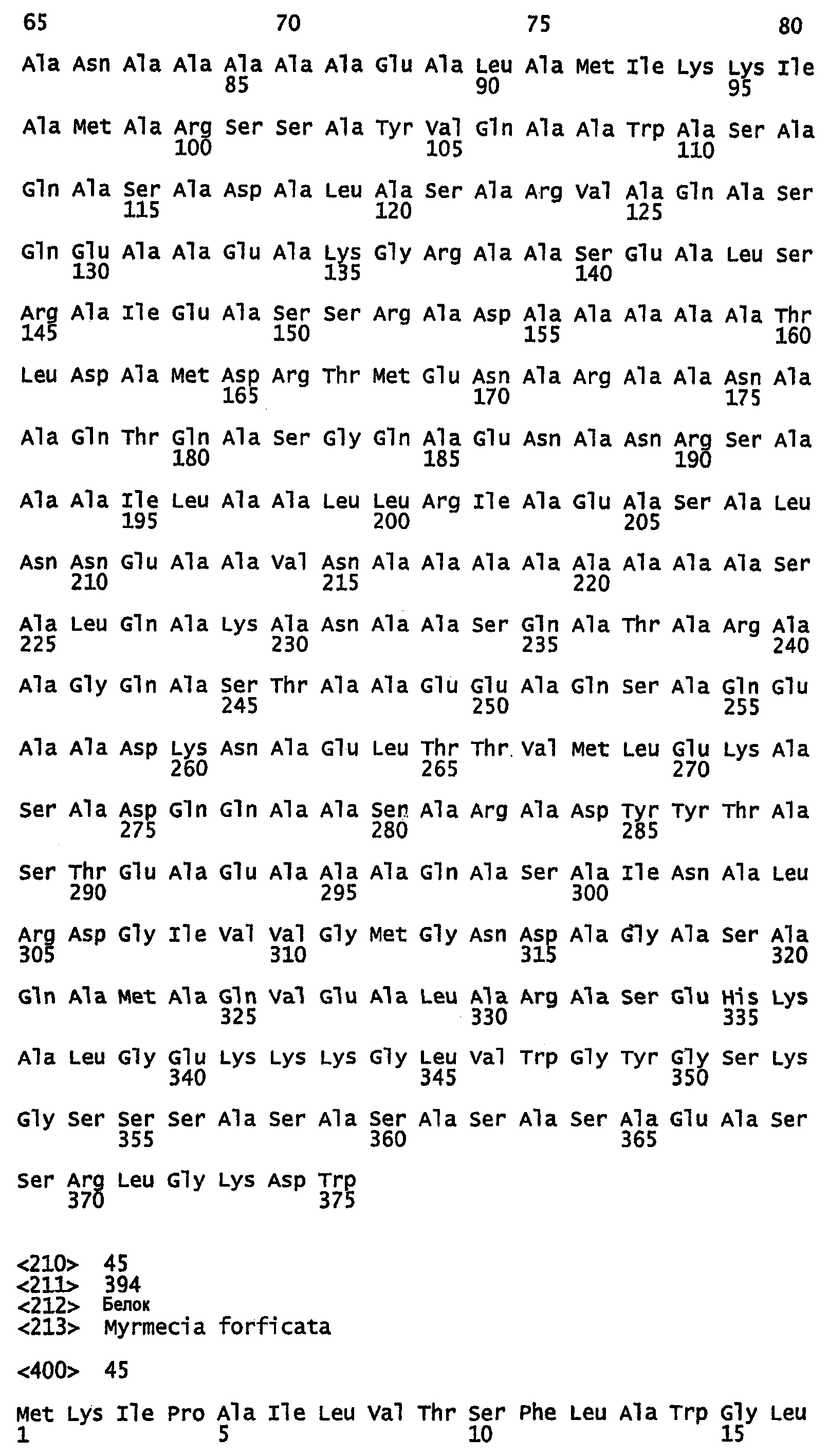
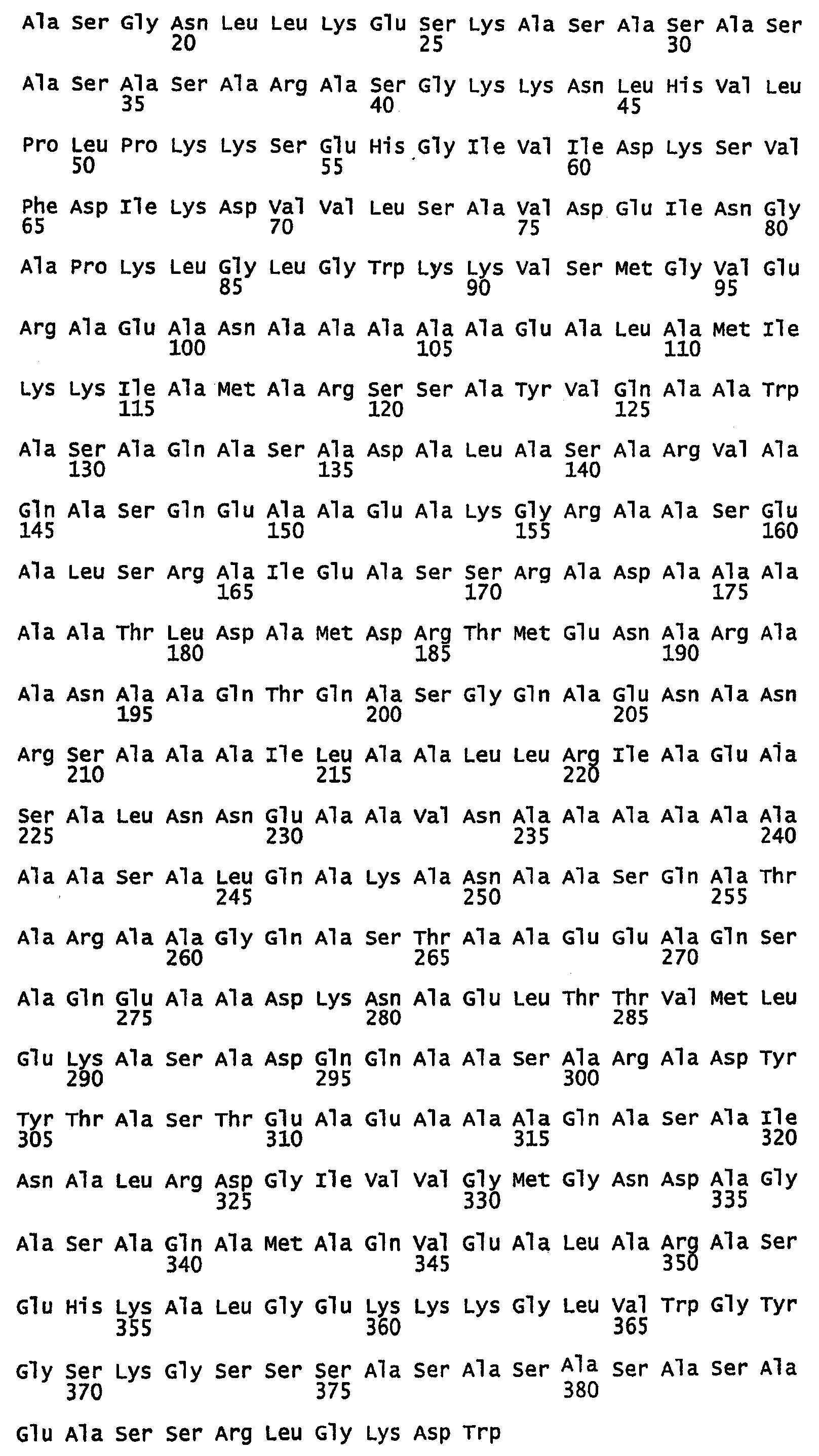
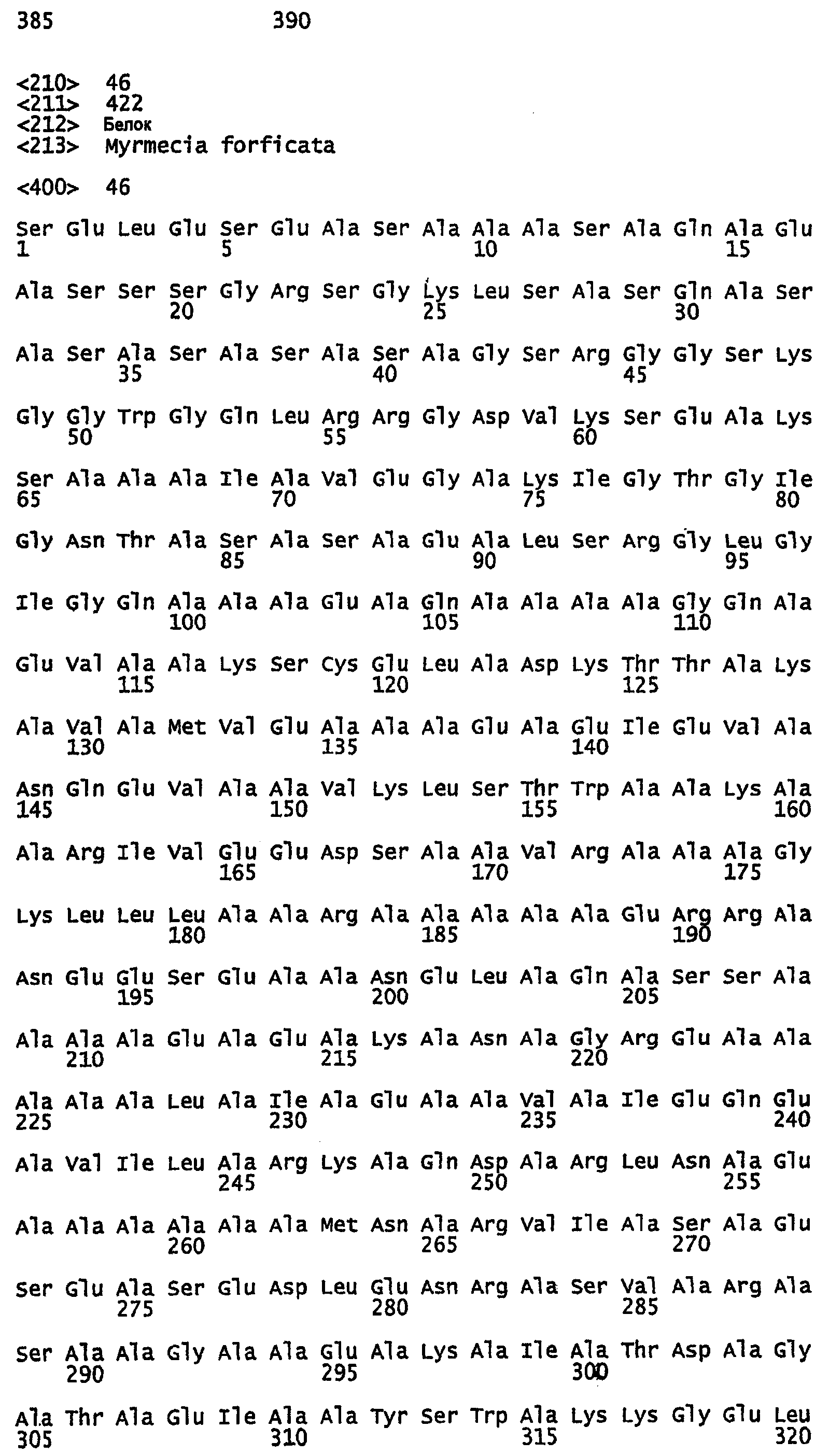
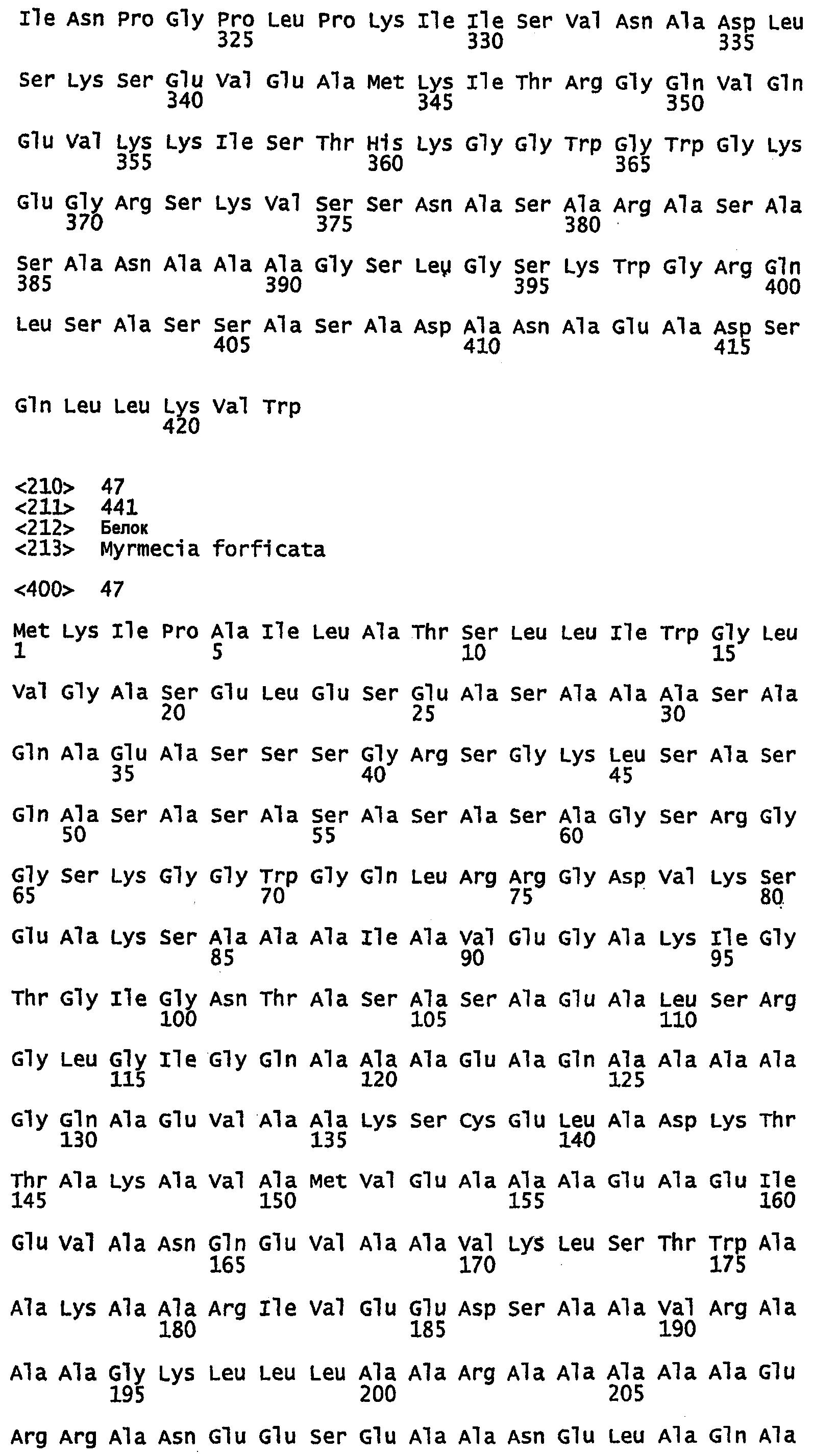










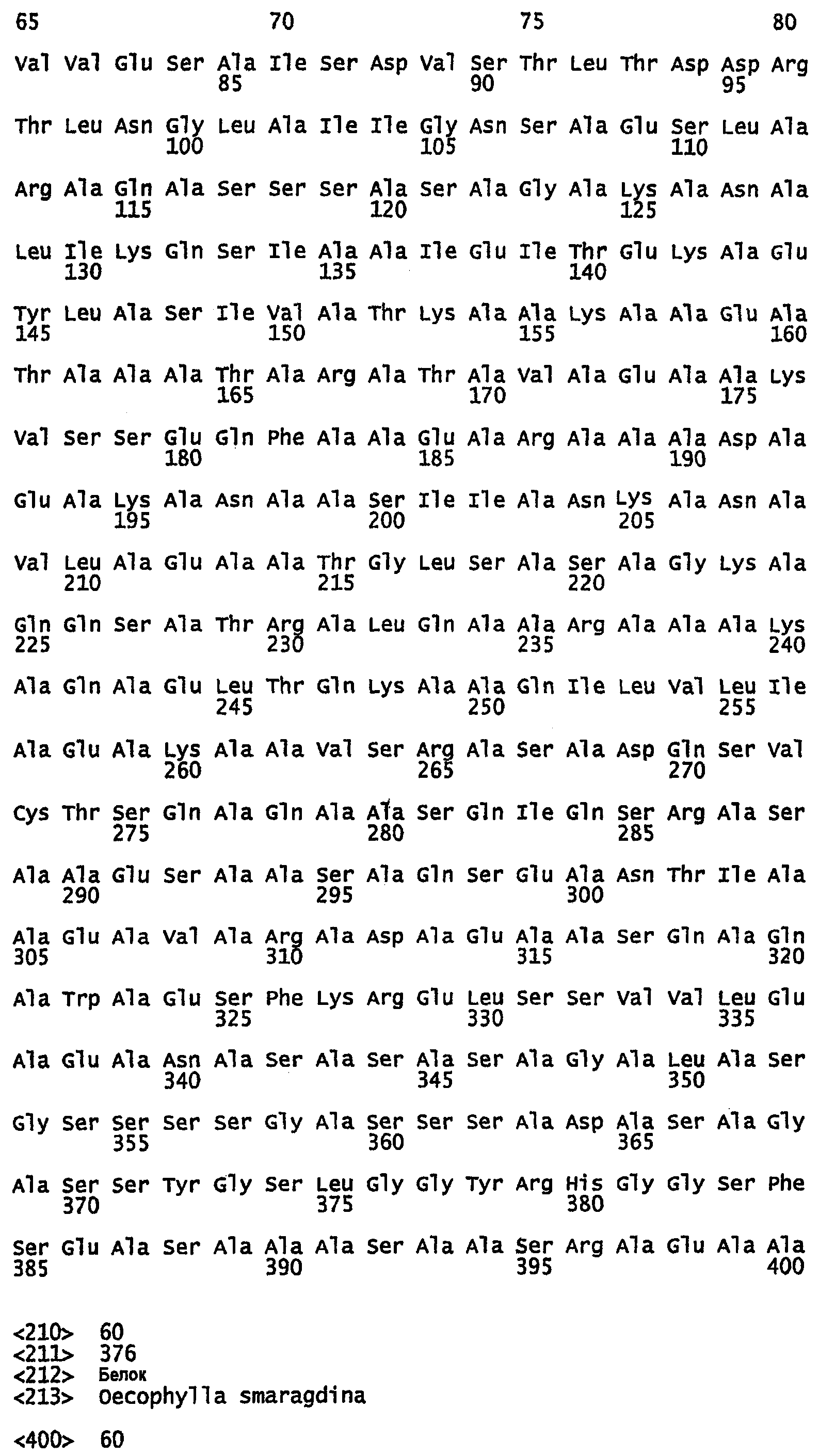
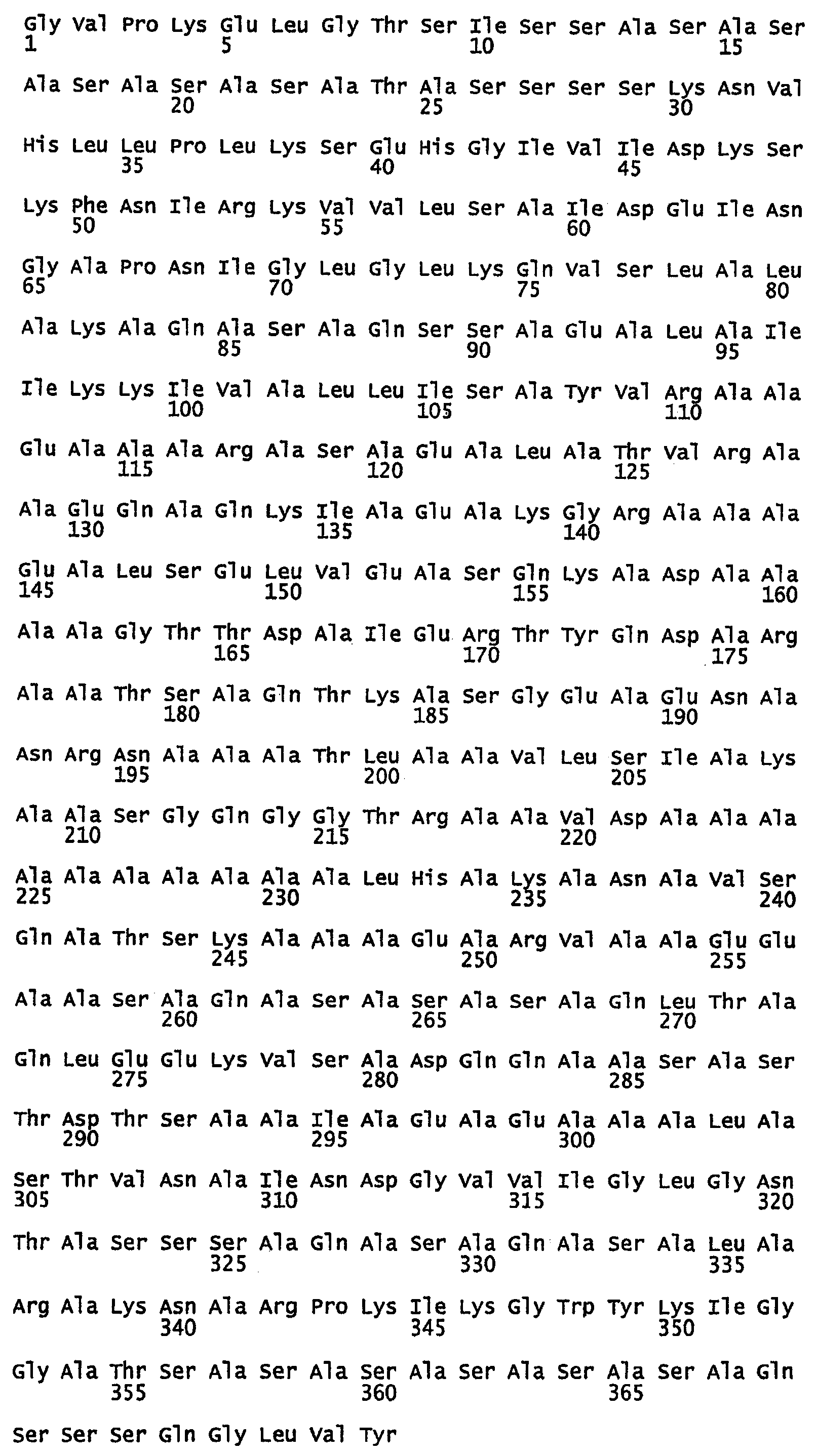


























Описание
Область техники
Изобретение относится к белкам шелка, а также к нуклеиновым кислотам, кодирующим эти белки. Настоящее изобретение также относится к рекомбинантным клеткам и/или организмам, которые синтезируют белки шелка. Белки шелка по изобретению могут применяться для разнообразных целей, таких как производство средств личной гигиены, пластиков, тканей и медико-биологических продуктов.
Предпосылки изобретения
Шелк представляет собой волокнистые белковые секреты, которые проявляют исключительную прочность и сопротивляемость, и поэтому представляют собой мишень для интенсивных исследований. Шелк продуцируют более чем 30000 видов пауков и многие другие насекомые. Из этих типов шелка было охарактеризовано очень мало, причем исследования были в основном сконцентрированы на шелке кокона одомашненного шелкопряда, Bombyx mori и шелке ячеек паука-кругопряда, Nephila clavipes.
У чешуекрылых и пауков гены фиброинов шелка кодируют белки, которые обычно имеют большой размер с выделяющимися гидрофильными концевыми доменами на любом конце, охватывающими обширную область перемежающихся гидрофобных и гидрофильных блоков (Bini et al., 2004). Обычно эти белки содержат различные комбинации кристаллических структур из β-складчатых листов, свободно связанных с β-листами, β-спиралями, α-спиралями и аморфными областями (в качестве обзора, см. Craig and Riekel, 2002).
Поскольку волокна шелка представляют один из наиболее известных природных волокон, они являются предметом интенсивного исследования для того, чтобы попытаться воспроизвести их синтез. Однако существующей проблемой при экспрессии фиброиновых генов чешуекрылых и пауков являются низкие уровни экспрессии в различных рекомбинантных системах экспрессии из-за комбинации повторяющихся нуклеотидных мотивов в гене шелка, которые приводят к разрушающим событиям рекомбинации, большого размера гена и малого числа кодонов, используемых для каждой аминокислоты в гене, что приводит к истощению пулов тРНК в клетках-хозяевах. Рекомбинантная экспрессия приводит к трудностям в ходе трансляции, таким как трансляционные паузы в результате выбора кодонов и потребности в кодонах, и повышенные скорости рекомбинации, приводящие к укорочению генов. Резюмируя, благодаря последовательностям с меньшим количеством повторов, можно было бы избежать множества проблем, связанных с экспрессией гена шелка на сегодняшний день.
В отличие от обширных знаний, накопленных о чешуекрылых (в частности, о шелке кокона Bombyx mori) и пауков (в частности, ячеистом шелке Nephila clavipes), мало известно о химической композиции и молекулярной организации шелков других насекомых.
В начале 1960-х годов с помощью рентгеновских дифракционных изображений из волокон шелка, вытянутых из слюнных желез личинки медоносной пчелы, было показано, что шелк жалящих перепончатокрылых имеет альфа-спиральную структуру (Rudall, 1962). Также показывая, что такой шелк является спиральным, полученные изображения указывали на наличие двойной спиральной системы из альфа-спиральных цепей (Atkins, 1967). Похожие рентгеновские дифракционные изображения были получены для шелка коконов из других видов Aculeata, включая осу, Pseudopompilus humbolti (Rudall, 1962) и шмеля, Bombus lucorum (Lucas and Rudall, 1967).
В отличие от альфа-спиральной структуры, описанной для шелков жалящих перепончатокрылых, шелки филогенетической ветви, родственной жалящим перепончатокрылым, Ichneumonoidea, имеют параллельные β-структуры. Рентгенограммы четырех примеров этой структуры были описаны у Braconidae (Cotesia(=Apenteles) glomerate; Cotesia(=Apenteles) gonopterygis; Apenteles bignelli) и для трех у Ichneumonidae (Dusona sp.; Phytodietris sp.; Branchus femoralis) (Lucas and Rudall, 1967). Кроме того, была описана единственная последовательность шелка Braconidae (Cotesia glomerate) (номер доступа в базе данных Genbank - AB 188680; Yamada et al., 2004). Эта частичная белковая последовательность состоит из высоко консервативного 28-членного Х-аспарагинового повтора (где Х представляет собой аланин или серин) и, предположительно, не содержит двойной спирали, образующей гептадные повторы. Расширенный анализ аминокислотной композиции шелка кокона Braconidae показал, что типы шелка подсемейства Microgastrinae являются уникальными по своему высокому содержанию серина и аспарагина (Lucas et al., 1960; Quicke et al., 2004). Родственные подсемейства продуцируют типы шелка со значительно отличающимися аминокислотными композициями, что позволяет предположить, что шелки Microgastrinae образовались только в этом подсемействе (Yamada et al., 2004). Была выделена частичная кДНК из Cotesia glomerate с использованием ПЦР-праймеров, разработанных на основе последовательности, полученной из внутренних пептидов, полученных из выделенных белков шелка кокона. Предсказанная аминокислотная композиция этой частичной аминокислотной последовательности близко напоминает аминокислотную композицию тщательно промытого шелка из этих видов.
Структура множества шелков из других нежалящих Apocrita и у оставшихся Hymenoptera (Symphata) представляет собой наиболее часто параллельные β-листы с как коллагено-подобными, так и полиглициновыми шелками, продуцируемыми Tenthredinidae (Lucas and Rudall, 1967).
Белки шелка медоносной пчелы синтезируются в середине последней возрастной стадии личинки в виде смеси деполимеризованных белков шелка (Silva-Zacarin et al., 2003). После развития возрастной стадии, вода удаляется из железы и дегидратация приводит к полимеризации белка шелка с образованием высокоорганизованных и нерастворимых шелковых волокон, меченных тактоидами (Silva-Zacarin et al., 2003). Возрастающая дегидратация приводит к дальнейшей реорганизации тактоидов (Silva-Zacarin et al., 2003) и, возможно, к образованию новых связей между филаментами (Rudall, 1962). На изображениях фибрилл из шелковых желез медоносной пчелы, полученных с помощью электронной микроскопии, показаны структуры, приблизительно, 20-25 ангстрем в диаметре (Flower and Kenchington, 1967). Это значение согласуется с трех-, четырех- или пятинитевыми двойными спиралями.
Аминокислотная композиция шелков из различных видов жалящих перепончатокрылых была определена Lucas and Rudall (1967), и было найдено, что они имеют высокое содержание аланина, серина и кислых остатков, аспарагиновой и глутаминовой кислот, и пониженное число остатков глицина по сравнению с классическими фиброинами. Полагают, что спиральное содержание шелка жалящих перепончатокрылых является следствием сниженного содержания глицина и повышенного содержания кислых остатков (Rudall and Kenchington, 1971).
Мало известно о шелке личинок сетчатокрылых (отряд: Neuroptera). Кокон состоит из двух слоев, внутреннего твердого слоя и внешнего волокнистого слоя. Ранее кокон был описан, как содержащий кутикулиновый шелк (Rudall and Kenchington, 1971), описание, относящееся только к внутреннему твердому слою. В 1988 LaMunyon описал вещество, выделяющееся из мальпигиевых сосудов и образующее внешние волокна. После отложения этого слоя создавалась твердая внутренняя стенка из выделений из эпителиальных клеток в сильно ворсинчатом просвете (LaMunyon, 1988).
Также известно, что личинки сетчатокрылых продуцируют клейкое вещество из мальпигиевых сосудов в течение всех возрастных стадий для прикрепления личинки к субстратам, для скрепления предметов маскировки на спине личинки или для ловли добычи (Speilger, 1962). В роде Lomamyia (Bethothidae), личинка продуцирует шелк и адгезивное вещество одновременно, и было высказано предположение, что эти два вещества вполне могут быть одним и тем же продуктом (Speilger, 1962). Адгезивное вещество является высоко растворимым веществом, и полагают, что оно связано с защитой против хищников (LaMunyon & Adams, 1987).
Поскольку шелк, производимый насекомыми, обладает уникальными свойствами, то существует необходимость в идентификации новых нуклеиновых кислот, кодирующих белки шелка из этих организмов.
Краткое изложение сущности изобретения
Авторы изобретения идентифицировали многочисленные белки шелка из насекомых. Эти белки шелка значительно отличаются от известных белков шелка по своей первичной структуре, вторичной структуре и/или аминокислотному составу.
Поэтому, первый аспект настоящего изобретения представляет по существу очищенный и/или рекомбинантный полипептид шелка, где по крайней мере часть полипептида имеет структуру двойной спирали.
Как известно в науке, двуспиральные структуры полипептидов отличаются гептадными повторами, представленными консенсусной последовательностью (abcdefg)n, обычно с гидрофобными остатками в положениях a и d, и, обычно, полярными остатками в других положениях. Удивительно, но при комплексном изучении гептады полипептидов по настоящему изобретению имеют новую композицию с необычно высоким присутствием аланина в «гидрофобном» положении в гептадах, a и d. Дополнительно в этих положениях присутствуют высокие уровни полярных остатков небольшого размера. Кроме того, положение е также имеет высокие уровни аланина и гидрофобных остатков небольшого размера.
Соответственно, в особенно предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 10 копий гептадной последовательности abcdefg, и по крайней мере 25% аминокислот в положениях a и d являются остатками аланина.
В дальнейшем предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 10 копий гептадной последовательности abcdefg, и по крайней мере 25% аминокислот в положениях a, d и е являются остатками аланина.
В дальнейшем предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 10 копий гептадной последовательности abcdefg, и по крайней мере 25% аминокислот в положении a являются остатками аланина.
В дальнейшем предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 10 копий гептадной последовательности abcdefg, и по крайней мере 25% аминокислот в положении d являются остатками аланина.
В дальнейшем предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 10 копий гептадной последовательности abcdefg, и по крайней мере 25% аминокислот в положении е являются остатками аланина.
В особенно предпочтительном варианте осуществления изобретения по крайней мере 10 копий гептадной последовательности являются непрерывными.
В дальнейшем предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 5 копий гептадной последовательности abcdefg, и по крайней мере 15% аминокислот в положениях a и d являются остатками аланина.
В дальнейшем предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 5 копий гептадной последовательности abcdefg, и по крайней мере 15% аминокислот в положениях a, d и е являются остатками аланина.
В дальнейшем предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 5 копий гептадной последовательности abcdefg, и по крайней мере 15% аминокислот в положении a являются остатками аланина.
В дальнейшем предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 5 копий гептадной последовательности abcdefg, и по крайней мере 15% аминокислот в положении d являются остатками аланина.
В дальнейшем предпочтительном варианте осуществления изобретения часть полипептида, которая имеет двуспиральную структуру, содержит по крайней мере 5 копий гептадной последовательности abcdefg, и по крайней мере 15% аминокислот в положении e являются остатками аланина.
В особенно предпочтительном варианте осуществления изобретения по крайней мере 5 копий гептадной последовательности являются непрерывными.
В одном варианте осуществления изобретения полипептид содержит последовательность, выбранную из:
i) аминокислотной последовательности, такой как представленная в любой одной из SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:22, SEQ ID NO:23, SEQ ID NO:40, SEQ ID NO:41, SEQ ID NO:56 и SEQ ID NO:57;
ii) аминокислотной последовательности, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:22, SEQ ID NO:23, SEQ ID NO:40, SEQ ID NO:41, SEQ ID NO:56 и SEQ ID NO:57; и
iii) биологически активного фрагмента из i) или ii).
В другом варианте осуществления изобретения полипептид содержит последовательность, выбранную из:
i) аминокислотной последовательности, такой как представленная в любой одной из SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:42, SEQ ID NO:43, SEQ ID NO:58 и SEQ ID NO:59;
ii) аминокислотной последовательности, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:42, SEQ ID NO:43, SEQ ID NO:58 и SEQ ID NO:59; и
iii) биологически активного фрагмента из i) или ii).
В другом варианте осуществления изобретения полипептид содержит последовательность, выбранную из:
i) аминокислотной последовательности, такой как представленная в любой одной из SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:26, SEQ ID NO:27, SEQ ID NO:44, SEQ ID NO:45, SEQ ID NO:60 и SEQ ID NO:61;
ii) аминокислотной последовательности, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:26, SEQ ID NO:27, SEQ ID NO:44, SEQ ID NO:45, SEQ ID NO:60 и SEQ ID NO:61; и
iii) биологически активного фрагмента из i) или ii).
В другом варианте осуществления изобретения полипептид содержит последовательность, выбранную из:
i) аминокислотной последовательности, такой как представленная в любой одной из SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:28, SEQ ID NO:29, SEQ ID NO:46, SEQ ID NO:47, SEQ ID NO:62 и SEQ ID NO:63;
ii) аминокислотной последовательности, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:28, SEQ ID NO:29, SEQ ID NO:46, SEQ ID NO:47, SEQ ID NO:62 и SEQ ID NO:63; и
iii) биологически активного фрагмента из i) или ii).
В дальнейшем варианте осуществления изобретения полипептид содержит последовательность, выбранную из:
i) аминокислотной последовательности, такой как представленная в SEQ ID NO:72 или SEQ ID NO:73;
ii) аминокислотной последовательности, которая по крайней мере на 30% идентична SEQ ID NO:72 и/или SEQ ID NO:73; и
iii) биологически активного фрагмента из i) или ii).
Далее были идентифицированы белки шелка, которые связаны с белками из первого аспекта. По предсказанию c помощью PROFseс один из этих белков (SEQ ID NO:10) имеет вторичную структуру, состоящую на 41% из альфа-спиралей, на 8% из бета-листов и на 50% из петель и, таким образом, классифицируется как белок со смешанной структурой. MARCOIL анализ этого белка предсказал только короткую область гептадных повторов, характерных для белков с двуспиральной структурой.
Соответственно, второй аспект изобретения представляет по существу очищенный и/или рекомбинантный полипептид шелка, который содержит последовательность, выбранную из:
i) аминокислотной последовательности, такой как представленная в любой одной из SEQ ID NO:9, SEQ ID NO:10 и SEQ ID NO:30;
ii) аминокислотной последовательности, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:9, SEQ ID NO:10 и SEQ ID NO:30; и
iii) биологически активного фрагмента из i) или ii).
Без желания быть ограниченным предположением, представляется, что четыре белка из первого аспекта скручиваются, образуя узел со спиральными осями, практически параллельными друг другу, и этот узел вытягивается по оси в фибриллу. Кроме того, предсказано, что по крайней мере у некоторых видов, таких как медоносная пчела и шмель, белки второго аспекта действуют как «клей», помогая в связывании различных узлов двуспиральных белков первого аспекта вместе, образуя волокнистый белковый комплекс. Однако еще шелковые волокна и сополимеры могут образоваться без полипептида второго аспекта.
В предпочтительном варианте осуществления изобретения полипептид по изобретению может быть очищен из или представлять собой мутант полипептида, очищенного из видов Hymenoptera или Neuroptera. Предпочтительно, видом Hymenoptera являются Apis mellifera, Oecophylla smaragdina, Myrmecia foricata или Bombus terrestris. Предпочтительно, видом Neuroptera является Mallada signata.
В другом аспекте настоящее изобретение представляет полипептид по изобретению, слитый по крайней мере с одним другим полипептидом.
В предпочтительном варианте осуществления изобретения по крайней мере один другой полипептид выбирается из группы, состоящей из: полипептида, который усиливает стабильность полипептида по настоящему изобретению, полипептида, который помогает в очистке слитого белка, и полипептида, который помогает секреции полипептида по изобретению из клетки (например, секреции из растительной клетки).
В другом аспекте, настоящее изобретение представляет выделенный и/или экзогенный полинуклеотид, который кодирует полипептид шелка, где по крайней мере часть полипептида имеет двуспиральную структуру.
В одном варианте осуществления изобретения полинуклеотид содержит последовательность, выбранную из:
i) последовательности нуклеотидов, такой как представленная в любой одной из SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:48, SEQ ID NO:49, SEQ ID NO:64 и SEQ ID NO:65;
ii) последовательности нуклеотидов, кодирующей полипептид по изобретению,
iii) последовательности нуклеотидов, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:11, SEQ ID NO: 12, SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:48, SEQ ID NO:49, SEQ ID NO:64 и SEQ ID NO:65, и
iv) последовательности нуклеотидов, которая гибридизируется с любой одной из i)-iii) при жестких условиях.
В другом варианте осуществления изобретения полинуклеотид содержит последовательность, выбранную из:
i) последовательности нуклеотидов, такой как представленная в любой одной из SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:50, SEQ ID NO:51, SEQ ID NO:66 и SEQ ID NO:67;
ii) последовательности нуклеотидов, кодирующей полипептид по изобретению,
iii) последовательности нуклеотидов, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:50, SEQ ID NO:51, SEQ ID NO:66 и SEQ ID NO:67, и
iv) последовательности нуклеотидов, которая гибридизируется с любой одной из i)-iii) при жестких условиях.
В другом варианте осуществления изобретения полинуклеотид содержит последовательность, выбранную из:
i) последовательности нуклеотидов, такой как представленная в любой одной из SEQ ID NO:15, SEQ ID NO:16, SEQ ID NO:35, SEQ ID NO:36, SEQ ID NO:52, SEQ ID NO:53, SEQ ID NO:68 и SEQ ID NO:69;
ii) последовательности нуклеотидов, кодирующей полипептид по изобретению,
iii) последовательности нуклеотидов, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:15, SEQ ID NO:16, SEQ ID NO:35, SEQ ID NO:36, SEQ ID NO:52, SEQ ID NO:53, SEQ ID NO:68 и SEQ ID NO:69, и
iv) последовательности нуклеотидов, которая гибридизируется с любой одной из i)-iii) при жестких условиях.
В дальнейшем варианте осуществления изобретения полинуклеотид содержит последовательность, выбранную из:
i) последовательности нуклеотидов, такой как представленная в любой одной из SEQ ID NO:17, SEQ ID NO:18, SEQ ID NO:37, SEQ ID NO:38, SEQ ID NO:54, SEQ ID NO:55, SEQ ID NO:70, SEQ ID NO:71 и SEQ ID NO:76;
ii) последовательности нуклеотидов, кодирующей полипептид по изобретению,
iii) последовательности нуклеотидов, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:17, SEQ ID NO:18, SEQ ID NO:37, SEQ ID NO:38, SEQ ID NO:54, SEQ ID NO:55, SEQ ID NO:70, SEQ ID NO:71 и SEQ ID NO:76, и
iv) последовательности нуклеотидов, которая гибридизируется с любой одной из i)=iii) при жестких условиях.
В другом варианте осуществления изобретения полинуклеотид содержит последовательность, выбранную из:
i) последовательности нуклеотидов, такой как представленная в SEQ ID NO:74 или SEQ ID NO:75;
ii) последовательности нуклеотидов, кодирующей полипептид по изобретению,
iii) последовательности нуклеотидов, которая по крайней мере на 30% идентична SEQ ID NO:74 и/или SEQ ID NO:75, и
iv) последовательности нуклеотидов, которая гибридизируется с любой одной из i)-iii) при жестких условиях.
В дальнейшем аспекте изобретение представляет выделенный и/или экзогенный полинуклеотид, полинуклеотид, содержащий последовательность, выбранную из:
i) последовательности нуклеотидов, такой как представленная в любой одной из SEQ ID NO:19, SEQ ID NO:20, SEQ ID NO:21 и SEQ ID NO:39;
ii) последовательности нуклеотидов, кодирующей полипептид по изобретению,
iii) последовательности нуклеотидов, которая по крайней мере на 30% идентична любой одной или нескольким из SEQ ID NO:19, SEQ ID NO:20, SEQ ID NO:21 и SEQ ID NO:39, и
iv) последовательности нуклеотидов, которая гибридизируется с любой одной из i)-iii) при жестких условиях.
В предпочтительном варианте осуществления изобретения полинуклеотид может быть выделен из или представлять собой мутант полинуклеотида, выделенного из видов Hymenoptera или Neuroptera. Предпочтительно, видом Hymenoptera являются Apis mellifera, Oecophylla smaragdina, Myrmecia foricata или Bombus terrestris. Предпочтительно, видом Neuroptera является Mallada signata.
В дальнейшем аспекте настоящее изобретение представляет вектор, содержащий по крайней мере один полинуклеотид по изобретению.
Предпочтительно, вектор представляет собой экспрессионный вектор.
В другом аспекте, настоящее изобретение представляет клетку-хозяина, содержащую по крайней мере один полинуклеотид по изобретению и/или по крайней мере один вектор по изобретению.
Клетка-хозяин может представлять собой любой тип клетки. Примеры включают, но не ограничены этим, бактериальную, дрожжевую или растительную клетку.
Также представлен процесс для получения полипептида по изобретению, процесс, содержащий культивирование клетки-хозяина по изобретению или вектора по изобретению при условиях, которые позволяют экспрессию полинуклеотида, кодирующего полипептид, и выделение экспрессированного полипептида.
Предусмотрено, что трансгенные растения будут особенно пригодными для получения полипептидов по изобретению. Поэтому, в еще одном аспекте настоящее изобретение представляет трансгенное растение, содержащее экзогенный полинуклеотид, полинуклеотид, кодирующий по крайней мере один полипептид по изобретению.
В другом аспекте настоящее изобретение представляет трансгенное животное, кроме человека, содержащее экзогенный полинуклеотид, полинуклеотид, кодирующий по крайней мере один полипептид по изобретению.
В еще одном аспекте настоящее изобретение представляет антитело, которое специфично связывает полипептид по изобретению.
В дальнейшем аспекте настоящее изобретение представляет шелковое волокно, содержащее по крайней мере один полипептид по изобретению.
Предпочтительно, полипептид представляет собой рекомбинантный полипептид.
В варианте осуществления изобретения по крайней мере некоторые из полипептидов являются кросс-сшитыми. В варианте осуществления изобретения по крайней мере некоторые из остатков лизина полипептида являются кросс-сшитыми.
В другом аспекте настоящее изобретение представляет сополимер, содержащий по крайней мере два полипептида по изобретению.
Предпочтительно, полипептиды являются рекомбинантными полипептидами.
В варианте осуществления изобретения сополимер содержит по крайней мере четыре различных полипептида из первого аспекта. В другом варианте осуществления изобретения сополимер далее содержит полипептид второго аспекта.
В варианте осуществления изобретения по крайней мере некоторые из полипептидов являются кросс-сшитыми. В варианте осуществления изобретения по крайней мере некоторые из остатков лизина полипептидов являются кросс-сшитыми.
Специалисту будет ясно, что полипептиды по изобретению имеют широкий спектр приложений, известных в этой области для других типов белков шелка. Поэтому, в дальнейшем аспекте настоящее изобретение представляет продукт, содержащий по крайней мере один полипептид по изобретению, шелковое волокно по изобретению и/или сополимер по изобретению.
Примеры продуктов включают, но не ограничены этим, средства личной гигиены, пластики, ткани и медико-биологические продукты.
В еще одном дальнейшем аспекте настоящее изобретение представляет композицию, содержащую по крайней мере один полипептид по изобретению, шелковое волокно по изобретению и/или сополимер по изобретению и один или более приемлемых носителей.
В одном варианте осуществления изобретения композиция содержит лекарство.
В другом варианте осуществления изобретения композиция используется как лекарственное средство в медицинском устройстве или косметическом средстве.
В другом аспекте настоящее изобретение представляет композицию, содержащую по крайней мере один полипептид по изобретению и один или более приемлемых носителей.
В предпочтительном варианте осуществления изобретения композиция, шелковое волокно, сополимер и/или продукт по изобретению не содержат белок маточного молочка пчел, продуцируемый насекомыми.
В дальнейшем аспекте настоящее изобретение представляет способ лечения или предотвращения болезни, способ, содержащий введение композиции, содержащей лекарство для лечения или предотвращения болезни и фармацевтически приемлемый носитель, где фармацевтически приемлемый носитель выбирается из по крайней мере одного полипептида по изобретению, шелкового волокна по изобретению и/или сополимера по изобретению.
В еще одном аспекте настоящее изобретение представляет применение по крайней мере одного полипептида по изобретению, шелкового волокна по изобретению и/или сополимера по изобретению и лекарства для изготовления лекарственного средства для лечения или предотвращения болезни.
В дальнейшем аспекте настоящее изобретение представляет набор, содержащий по крайней мере один полипептид по изобретению, по крайней мере один полинуклеотид по изобретению, по крайней мере один вектор по изобретению, по крайней мере одно шелковое волокно по изобретению и/или сополимер по изобретению.
Предпочтительно, чтобы набор содержал информацию и/или инструкции для применения набора.
Как будет ясно, предпочтительные свойства и характеристики одного аспекта изобретения являются применимыми для многих других аспектов изобретения.
В этой спецификации слово «содержать» или его вариации, такие как «содержит» или «содержащий» следует понимать, как предполагающее включение указанного элемента, целого или дробного, или группы элементов, целых или дробных, но не как исключение любого другого элемента целого или дробного, или группы элементов, целых или дробных.
Изобретение далее описано путем нижеследующих неограничивающих примеров со ссылками на сопровождающие фигуры.
Краткое описание сопровождающих чертежей
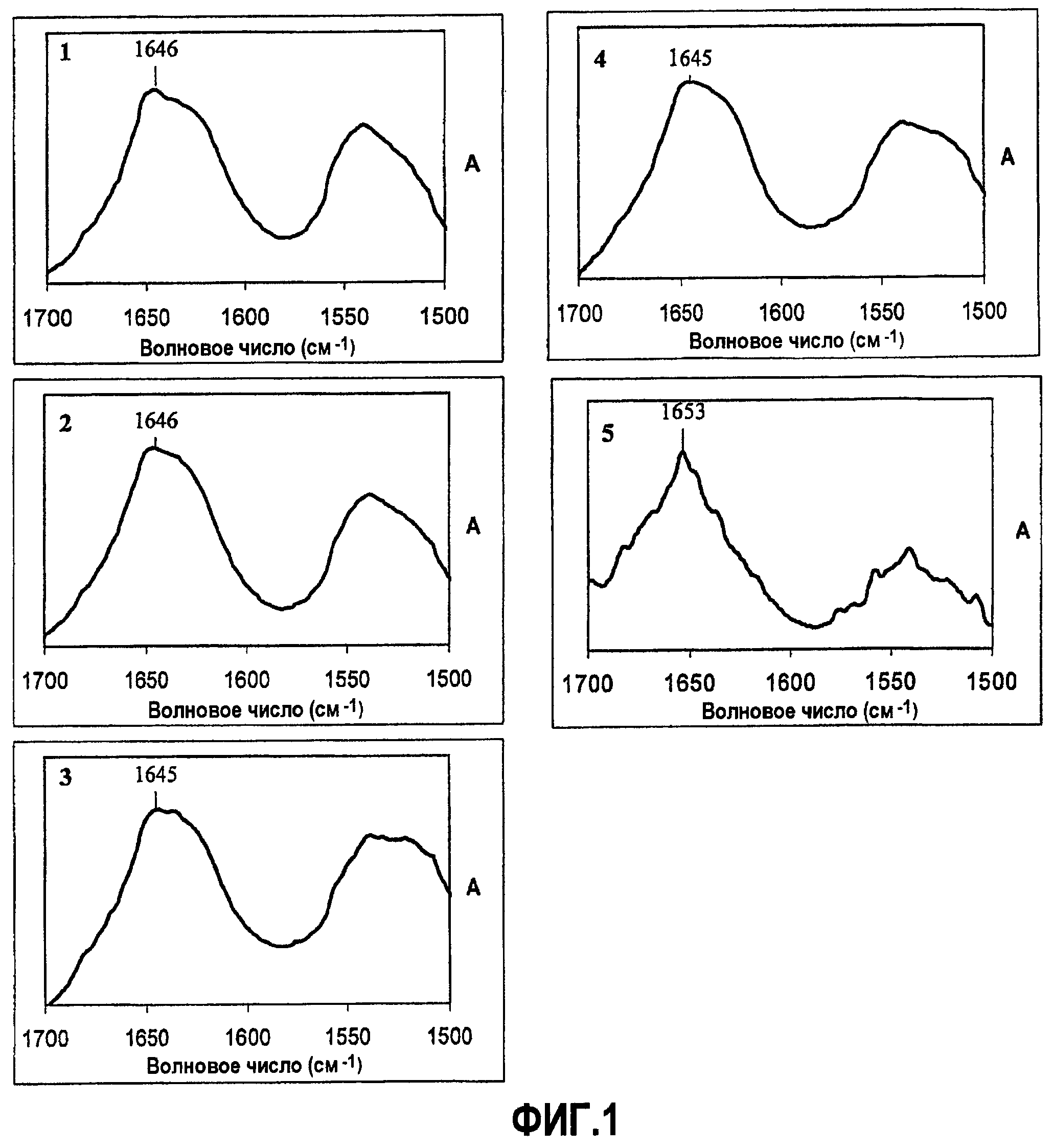
Фиг.1. Фурье инфракрасный спектр амидных областей I и II шелков: 1) шелка медоносной пчелы, 2) шелка шмеля, 3) шелка муравья-бульдога, 4) шелка муравья-портного, 5) шелка личинки златоглазки. Все шелки имеют спектры, ожидаемые для спиральных белков. Шелки перепончатокрылых (муравьев и пчел) имеют спектральный максимум при 1645-1646 см-1 (отмечен), сдвинутый, приблизительно, на 10 см-1 ниже, чем классический альфа-спиральный сигнал и расширенный, что типично для белков с двойной спиралью (Heimburg et al., 1999).
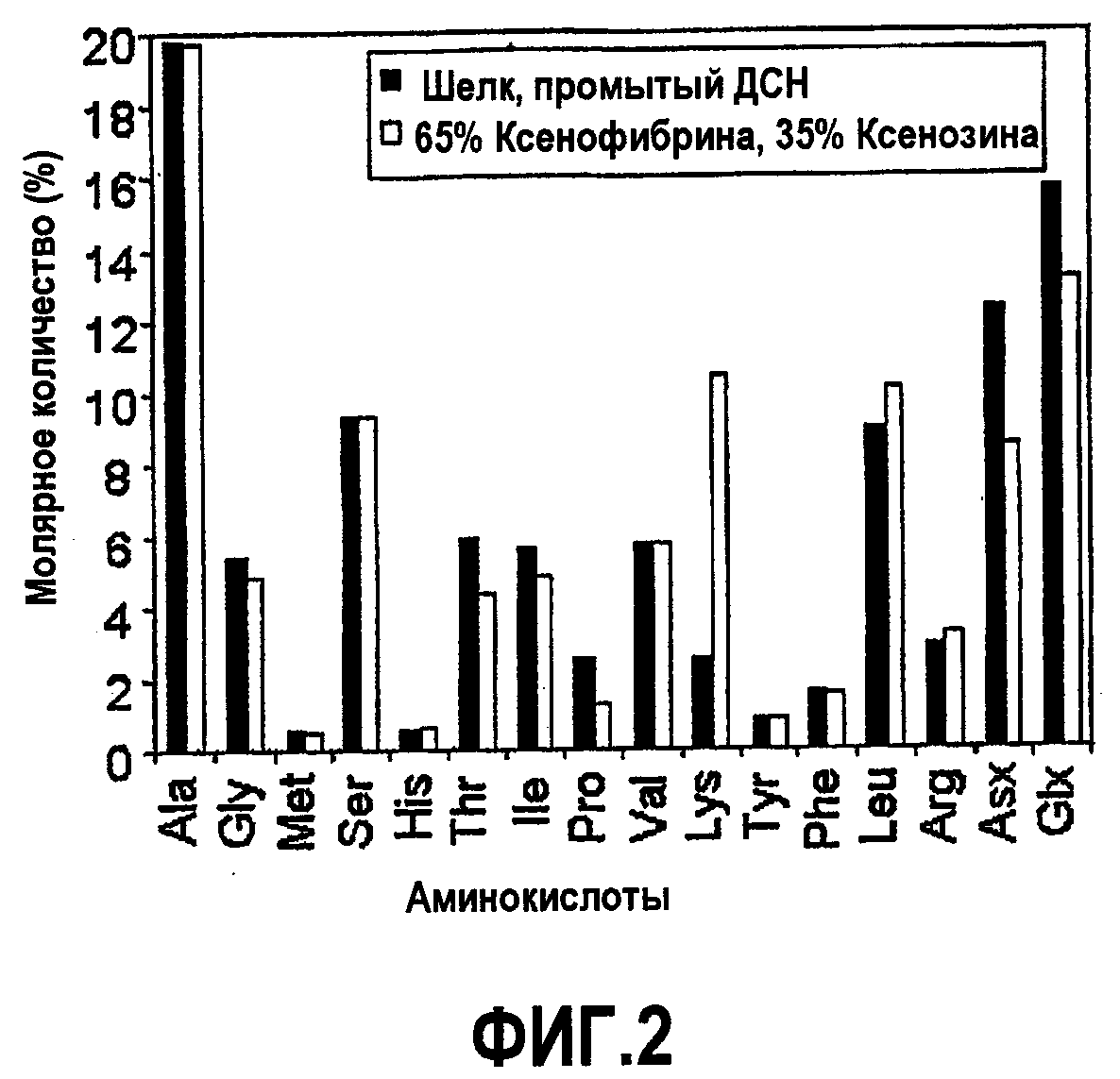
Фиг.2. Сравнение аминокислотной композиции, промытой ДСН шелка сот с расплодом медоносной пчелы с аминокислотной композицией белков Ксеноспира (а именно, Ксеноспира1, Ксеноспира2, Ксеноспира3 и Ксеноспира4) (эквимолярные количества в сумме составляют 65%) и Ксенозина (35%).
Фиг.3. Сравнение аминокислотной композиции шелка с аминокислотной композицией, предсказанных для белков, кодируемых генами шелка.
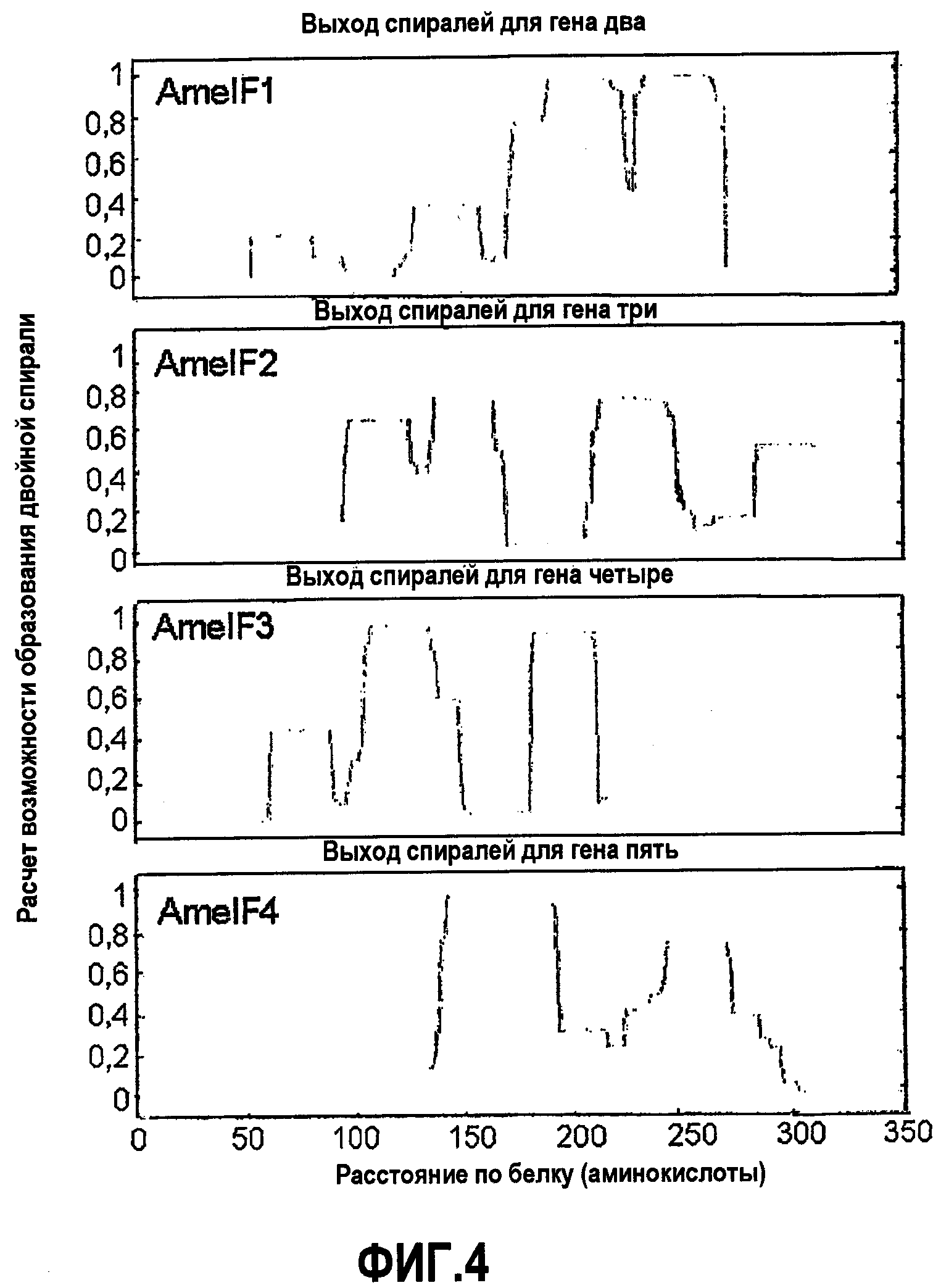
Фиг.4. Предсказание областей двойной спирали белков шелка медоносной пчелы. COILS представляет собой программу, которая сравнивает последовательность с базой данных известных параллельных двухцепочечных двойных спиралей и получает оценку сходства. Путем сравнения этой оценки с распределением оценок в глобулярных и двуспиральных белках, программа затем вычисляет вероятность того, что последовательность примет конформацию двойной спирали, описанную в Lupas et al. (1991). С использованием размера окна 28 эта программа предсказывает следующее число остатков, существующих в каждом белке в двуспиральных доменах: Ксеноспира3: 77; Ксеноспира4: 35; Ксеноспира1: 28; Ксеноспира2: 80.
Фиг.5. Выравнивание белков шелка медоносной пчелы, показывающее предсказание с помощью MARCOIL главных гептад, которые образуют структуру двойной спирали. Последовательности гептад показаны над аминокислотами, остатки аланина в положениях a и d выделены.
Фиг.6. Выравнивание Marciol предсказанных областей двойных спиралей белков шелка перепончатокрылых (пчел и муравьев), показывающее позиционное распределение гептад. Amel, медоносная пчела; ВВ, шмель; ВА, муравей-бульдог; WA, муравей-портной; F1-4, фиброины шелка 1-4. Гептадные последовательности показаны над аминокислотами, и остатки аланина в положениях a, d и е выделены.
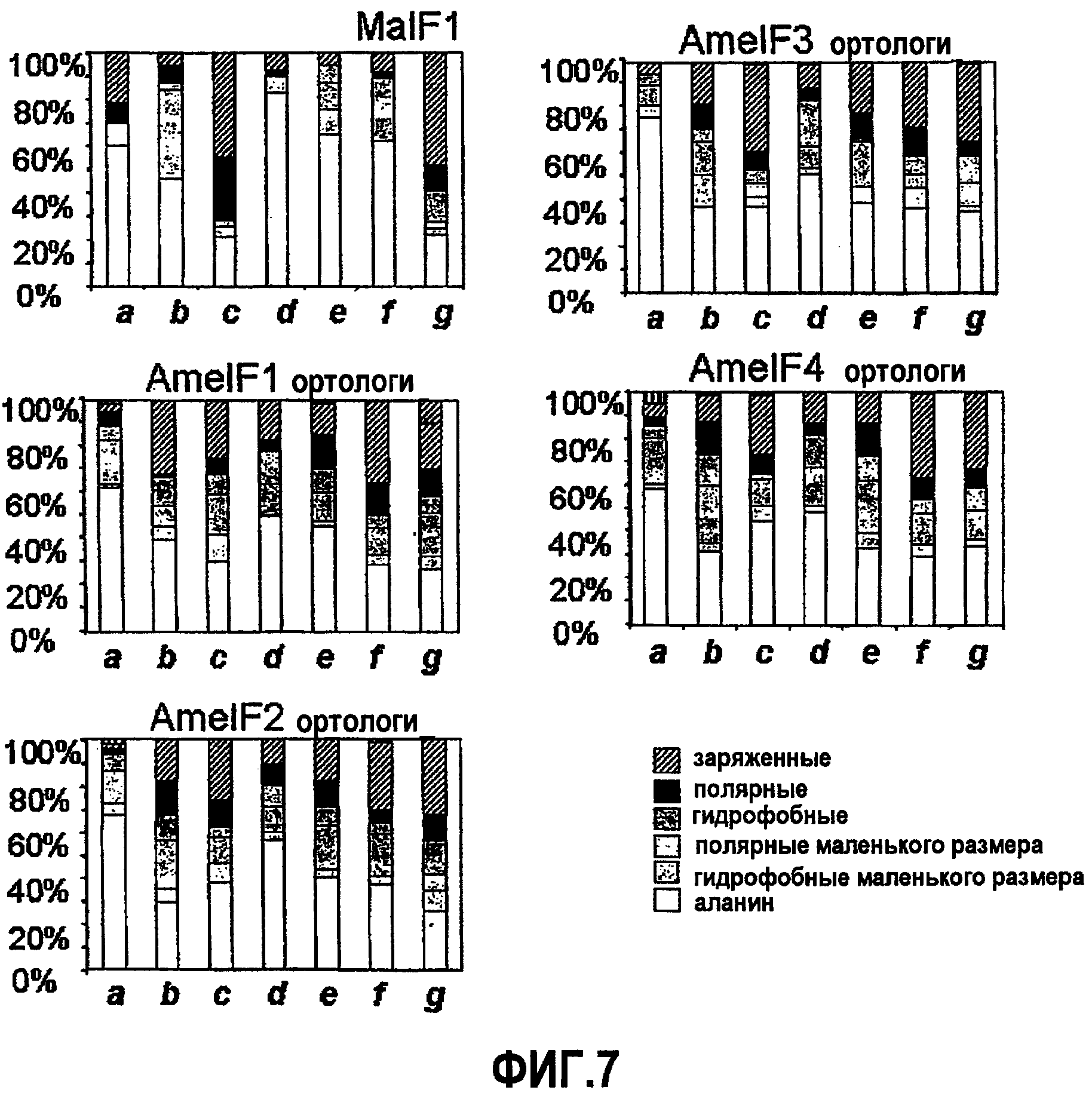
Фиг.7. Аминокислотная характеристика положения гептад в предсказанных областях двойных спиралей белка шелка личинки Mallada signata и ортологичных кластерах белков шелка перепончатокрылых.
Фиг.8. Электрофорез в ДСН-полиакриламидном геле белков слюнных желез поздней последней возрастной стадии личинки. Белки были идентифицированы после расщепления трипсином и анализа совокупности данных масс-спектра с использованием программного обеспечения Spectrum Mill компании Agilent, чтобы совместить данные с предсказанными белковыми последовательностями из белков, идентифицированных из последовательностей кДНК. Программа генерировала счет для оценки качества каждого совпадения между экспериментально наблюдаемыми группами масс фрагментов пептидов и предсказанными фрагментами, которые могут образоваться в соответствии с последовательностями белков в предоставленной базе данных. Все совпадения последовательностей, показанные на фигуре, получили оценку больше 20 при помощи программного обеспечения Spectrum Mill, где оценка 20 была бы достаточной, чтобы автоматически уверенно принять действительное совпадение.
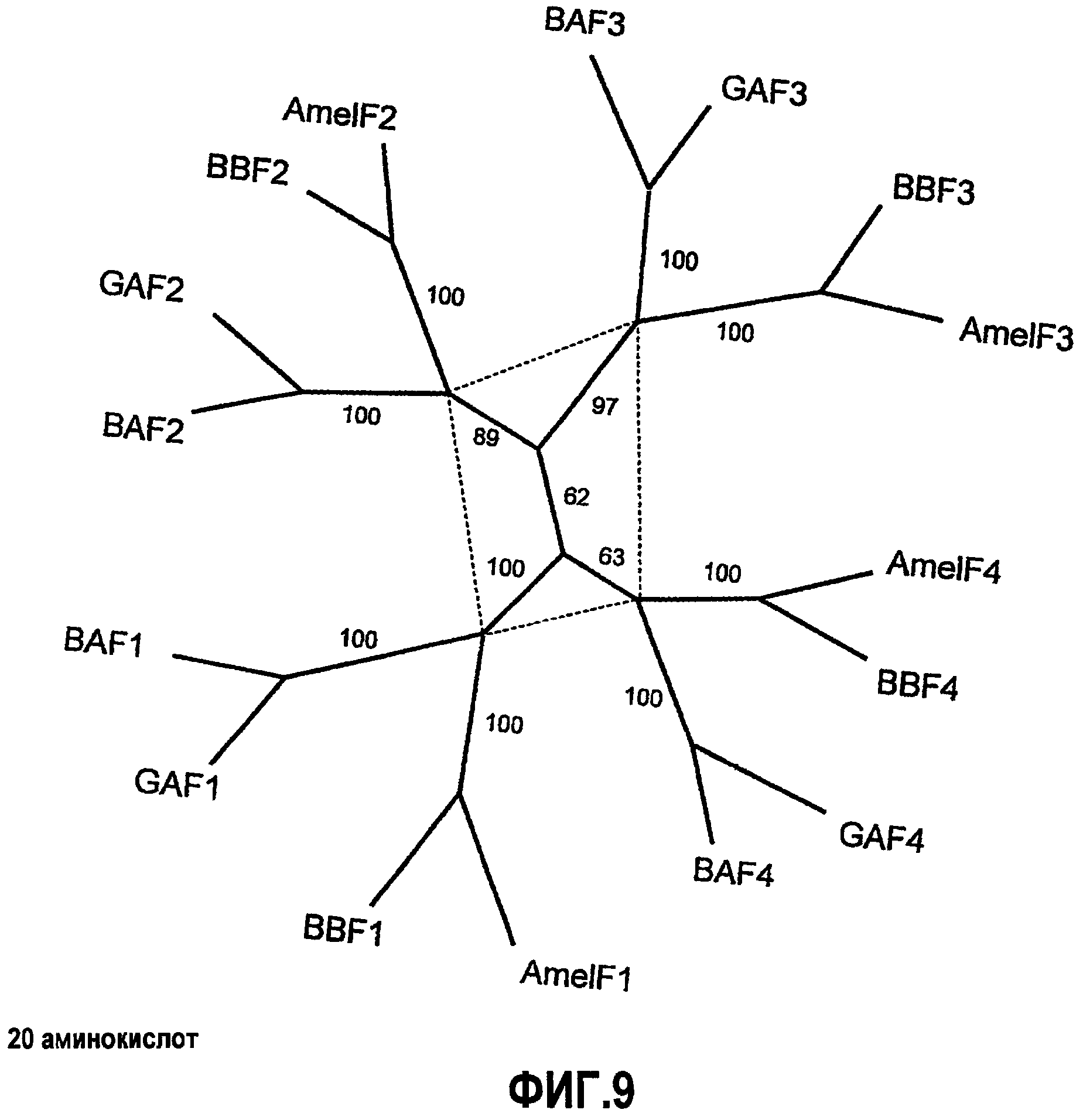
Фиг.9. Экономный анализ двуспиральной области белков шелка. Родство четырех двуспиральных белков позволяет предположить, что гены возникли из общего предшественника, предваряя расхождение Euaculeata. Область, связанная пунктирной линией, указывает на изменение, произошедшее перед расхождением муравьев и ос (Vespoidea) от пчел (Apoidea) в позднем Юрском периоде (155 млн л.; Grimaldi and Engel, 2005). Приведены числа, указывающие начальную загрузку от 1000 итераций.
Фиг.10. А) Белки шелка из Apis mellifera, идентифицированные с помощью масс-спектрометрии пептидов, полученных из шелка пчел после трипсинового гидролиза. Штриховка указывает пептиды, идентифицированные с помощью масс-спектрометрии. Все показанные в этой фигуре совпадения по последовательности получили оценки выше 20 при помощи программного обеспечения Spectrum Mill, где оценка 20 была бы достаточной, чтобы автоматически уверенно принять действительное совпадение.
В) Полноразмерные аминокислотные последовательности белков шелка шмеля, муравья-бульдога, ткача и златоглазки.
Фиг.11. Открытые рамки считывания, кодирующие белки шелка медоносной пчелы, шмеля, муравья-бульдога, муравья-портного и златоглазки.
Фиг.12. Последовательность гена, кодирующего Ксенозин. Приведена полная кодирующая последовательность, которая прерывается единственным интроном (выделен).
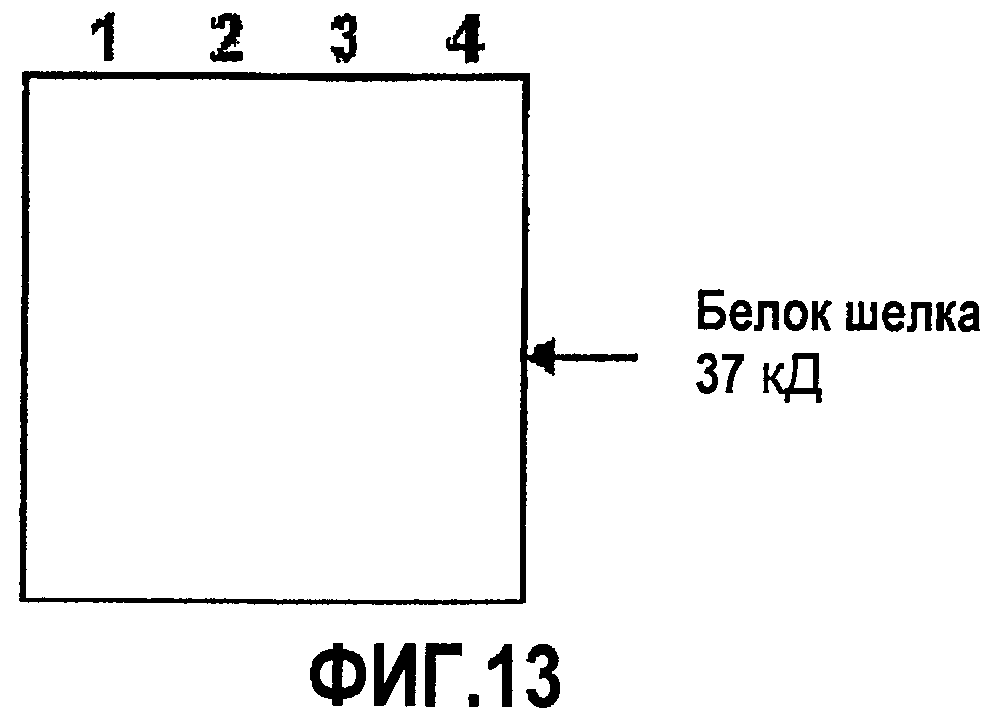
Фиг.13. Экспрессия белка шелка в табаке. Детекция меченных гистидинами белков после анализа методом Вестерн-блота белков из: 1. E. coli, трансформированных пустым экспрессионным вектором, 2. E. coli, трансформированных экспрессионным вектором, содержащим участок, кодирующий AmelF4 (Ксеноспира4), 3. табака, трансформированного пустым экспрессионным вектором, 4. табака, трансформированного экспрессионным вектором, содержащим участок, кодирующий AmelF4 (Ксеноспира4).
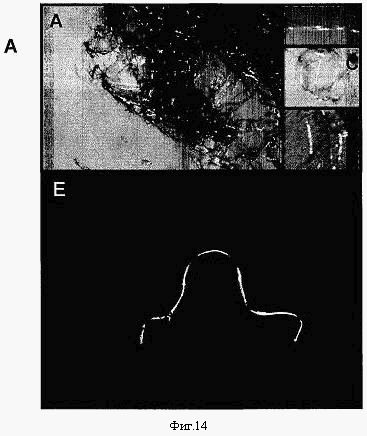
Фиг.14. Волокна, изготовленные из рекомбинантных белков шелка медоносной пчелы, показывающие двойное лучепреломление нитей. Двойное лучепреломление указывает на то, что структура присутствует в нитях. Различные рекомбинантные нити медоносной пчелы показаны на каждой панели А-D, а рекомбинантная нить златоглазки показана на панели Е.
Описание списка последовательностей
SEQ ID NO:1 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксеноспира1 (также именуемый в настоящем описании AmelF1) (без сигнального пептида).
SEQ ID NO:2 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксеноспира1.
SEQ ID NO:3 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксеноспира2 (также именуемый в настоящем описании AmelF2) (без сигнального пептида).
SEQ ID NO:4 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксеноспира2.
SEQ ID NO:5 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксеноспира3 (также именуемый в настоящем описании AmelF3) (без сигнального пептида).
SEQ ID NO:6 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксеноспира3.
SEQ ID NO:7 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксеноспира4 (также именуемый в настоящем описании AmelF4) (без сигнального пептида).
SEQ ID NO:8 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксеноспира4.
SEQ ID NO:9 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксенозин (также именуемый в настоящем описании AmelSA1) (без сигнального пептида).
SEQ ID NO:10 - белок шелка медоносной пчелы, именуемый в настоящем описании Ксенозин.
SEQ ID NO:11 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксеноспира1 (без области, кодирующей сигнальный пептид).
SEQ ID NO:12 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксеноспира1.
SEQ ID NO:13 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксеноспира2 (без области, кодирующей сигнальный пептид).
SEQ ID NO:14 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксеноспира2.
SEQ ID NO:15 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксеноспира3 (без области, кодирующей сигнальный пептид).
SEQ ID NO:16 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксеноспира3.
SEQ ID NO:17 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксеноспира4 (без области, кодирующей сигнальный пептид).
SEQ ID NO:18 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксеноспира4.
SEQ ID NO:19 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксенозин (без области, кодирующей сигнальный пептид).
SEQ ID NO:20 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы Ксенозин.
SEQ ID NO:21 - последовательность гена, кодирующего белок шелка медоносной пчелы Ксенозин.
SEQ ID NO:22 - белок шелка шмеля, именуемый в настоящем описании BBF1 (без сигнального пептида).
SEQ ID NO:23 - белок шелка шмеля, именуемый в настоящем описании BBF1.
SEQ ID NO:24 - белок шелка шмеля, именуемый в настоящем описании BBF2 (без сигнального пептида).
SEQ ID NO:25 - белок шелка шмеля, именуемый в настоящем описании BBF2.
SEQ ID NO:26 - белок шелка шмеля, именуемый в настоящем описании BBF3 (без сигнального пептида).
SEQ ID NO:27 - белок шелка шмеля, именуемый в настоящем описании BBF3.
SEQ ID NO:28 - белок шелка шмеля, именуемый в настоящем описании BBF4 (без сигнального пептида).
SEQ ID NO:29 - белок шелка шмеля, именуемый в настоящем описании BBF4.
SEQ ID NO:30 - частичная аминокислотная последовательность белка шелка шмеля, именуемого в настоящем описании BBSA1.
SEQ ID NO:31 - нуклеотидная последовательность, кодирующая белок шелка шмеля BBF1 (без области, кодирующей сигнальный пептид).
SEQ ID NO:32 - нуклеотидная последовательность, кодирующая белок шелка шмеля BBF1.
SEQ ID NO:33 - нуклеотидная последовательность, кодирующая белок шелка шмеля BBF2 (без области, кодирующей сигнальный пептид).
SEQ ID NO:34 - нуклеотидная последовательность, кодирующая белок шелка шмеля BBF2.
SEQ ID NO:35 - нуклеотидная последовательность, кодирующая белок шелка шмеля BBF3 (без области, кодирующей сигнальный пептид).
SEQ ID NO:36 - нуклеотидная последовательность, кодирующая белок шелка шмеля BBF3.
SEQ ID NO:37 - нуклеотидная последовательность, кодирующая белок шелка шмеля BBF4 (без области, кодирующей сигнальный пептид).
SEQ ID NO:38 - нуклеотидная последовательность, кодирующая белок шелка шмеля BBF4.
SEQ ID NO:39 - частичная нуклеотидная последовательность, кодирующая белок шелка шмеля BBSA1.
SEQ ID NO:40 - белок шелка муравья-бульдога, именуемый в настоящем описании BAF1 (без сигнального пептида).
SEQ ID NO:41 - белок шелка муравья-бульдога, именуемый в настоящем описании BAF1.
SEQ ID NO:42 - белок шелка муравья-бульдога, именуемый в настоящем описании BAF2 (без сигнального пептида).
SEQ ID NO:43 - белок шелка муравья-бульдога, именуемый в настоящем описании BAF2.
SEQ ID NO:44 - белок шелка муравья-бульдога, именуемый в настоящем описании BAF3 (без сигнального пептида).
SEQ ID NO:45 - белок шелка муравья-бульдога, именуемый в настоящем описании BAF3.
SEQ ID NO:46 - белок шелка муравья-бульдога, именуемый в настоящем описании BAF4 (без сигнального пептида).
SEQ ID NO:47 - белок шелка муравья-бульдога, именуемый в настоящем описании BAF4.
SEQ ID NO:48 - нуклеотидная последовательность, кодирующая белок шелка муравья-бульдога BAF1 (без области, кодирующей сигнальный пептид).
SEQ ID NO:49 - нуклеотидная последовательность, кодирующая белок шелка муравья-бульдога BAF1.
SEQ ID NO:50 - нуклеотидная последовательность, кодирующая белок шелка муравья-бульдога BAF2 (без области, кодирующей сигнальный пептид).
SEQ ID NO:51 - нуклеотидная последовательность, кодирующая белок шелка муравья-бульдога BAF2.
SEQ ID NO:52 - нуклеотидная последовательность, кодирующая белок шелка муравья-бульдога BAF3 (без области, кодирующей сигнальный пептид).
SEQ ID NO:53 - нуклеотидная последовательность, кодирующая белок шелка муравья-бульдога BAF3.
SEQ ID NO:54 - нуклеотидная последовательность, кодирующая белок шелка муравья-бульдога BAF4 (без области, кодирующей сигнальный пептид).
SEQ ID NO:55 - нуклеотидная последовательность, кодирующая белок шелка муравья-бульдога BAF4.
SEQ ID NO:56 - белок шелка муравья-портного, именуемый в настоящем описании GAF1 (без сигнального пептида).
SEQ ID NO:57 - белок шелка муравья-портного, именуемый в настоящем описании GAF1.
SEQ ID NO:58 - белок шелка муравья-портного, именуемый в настоящем описании GAF2 (без сигнального пептида).
SEQ ID NO:59 - белок шелка муравья-портного, именуемый в настоящем описании GAF2.
SEQ ID NO:60 - белок шелка муравья-портного, именуемый в настоящем описании GAF3 (без сигнального пептида).
SEQ ID NO:61 - белок шелка муравья-портного, именуемый в настоящем описании GAF3.
SEQ ID NO:62 - белок шелка муравья-портного, именуемый в настоящем описании GAF4 (без сигнального пептида).
SEQ ID NO:63 - белок шелка муравья-портного, именуемый в настоящем описании GAF4.
SEQ ID NO:64 - нуклеотидная последовательность, кодирующая белок шелка муравья-портного GAF1 (без области, кодирующей сигнальный пептид).
SEQ ID NO:65 - нуклеотидная последовательность, кодирующая белок шелка муравья-портного GAF1.
SEQ ID NO:66 - нуклеотидная последовательность, кодирующая белок шелка муравья-портного GAF2 (без области, кодирующей сигнальный пептид).
SEQ ID NO:67 - нуклеотидная последовательность, кодирующая белок шелка муравья-портного GAF2.
SEQ ID NO:68 - нуклеотидная последовательность, кодирующая белок шелка муравья-портного GAF3 (без области, кодирующей сигнальный пептид).
SEQ ID NO:69 - нуклеотидная последовательность, кодирующая белок шелка муравья-портного GAF3.
SEQ ID NO:70 - нуклеотидная последовательность, кодирующая белок шелка муравья-портного GAF4 (без области, кодирующей сигнальный пептид).
SEQ ID NO:71 - нуклеотидная последовательность, кодирующая белок шелка муравья-портного GAF4.
SEQ ID NO:72 - белок шелка златоглазки, именуемый в настоящем описании MalF1 (без сигнального пептида).
SEQ ID NO:73 - белок шелка златоглазки, именуемый в настоящем описании MalF1.
SEQ ID NO:74 - нуклеотидная последовательность, кодирующая белок шелка златоглазки MalF1 (без области, кодирующей сигнальный пептид).
SEQ ID NO:75 - нуклеотидная последовательность, кодирующая белок шелка златоглазки MalF1.
SEQ ID NO:76 - нуклеотидная последовательность, кодирующая белок шелка медоносной пчелы, именуемый в настоящем описании Ксеноспира4, с оптимизированными кодонами для экспрессии в растениях (перед субклонированием в pET14b и pVEC8).
SEQ ID NO:77 - открытая рамка считывания белка шелка медоносной пчелы (Ксеноспира4), оптимизированная для экспрессии в растениях (без трансляционного слияния).
Подробное описание изобретения
Общие методики и определения
Если не указано особо, то все технические и научные термины, используемые в настоящем описании, должны приниматься, как имеющие то же значение, которое обычно понимает под этим термином средний специалист (например, в клеточной культуре, молекулярной генетике, иммунологии, иммуногистохимии, белковой химии и биохимии).
Если не указано особо, методики рекомбинантных белков, клеточной культуры и иммунологические методики, применяемые в настоящем изобретении, представляют собой стандартные процедуры, хорошо известные специалисту. Такие методики описываются и объясняются в литературных источниках, таких как J. Perbal, A Practical Guide to Molecular Cloning, John Wiley and Sons (1984), J. Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbour Laboratory Press (1989), T.A. Brown (editor), Essential Molecular Biology: A Practical Approach, Volumes 1 and 2, IRL Press (1991), D.M. Glover and B.D. Hames (editors), DNA Cloning: A Practical Approach, Volumes 1-4, IRL Press (1995 and 1996) и F.M. Ausubel et al. (editors), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience (1988, включая все обновления на сегодняшний день), Ed Harlow and David Lane (editors) Antibodies: A Laboratory Manual, Cold Spring Harbour Laboratory, (1988) и J.E. Coligan et al. (editors) Current Protocols in Immunology, John Wiley & Sons (включая все обновления на сегодняшний день) и включены в настоящее описание посредством ссылки.
Используемые в настоящем описании термины «белок шелка» и «полипептид шелка» относятся к волокнистому белку/полипептиду, который может применяться для получения шелкового волокна и/или волокнистого белкового комплекса. Природные белки шелка по изобретению образуют часть шелка сот с расплодом насекомых, таких как медоносные пчелы, однако, легко могут быть получены описанные в настоящем документе варианты этих белков, которые будут выполнять те же функции, при экспрессии в соответствующем насекомом.
Используемое в настоящем описании «шелковое волокно» относится к волокну, содержащему белки по изобретению, которое может быть соткано в различные предметы, такие как ткани.
Используемый в настоящем описании «сополимер» представляет собой композицию, содержащую два или более белков шелка по изобретению. Этот термин исключает природные сополимеры, такие как соты с расплодом насекомых.
Термин «растение» включает целые растения, вегетативные структуры (например, листья, стебли), корни, цветочные органы/структуры, семя (включая зародыш, эндосперм и семенную оболочку), растительную ткань (например, проводящую ткань, покровную ткань и т.п.), клетки и потомство всего этого.
«Трансгенное растение» относится к растению, которое содержит генную конструкцию («трансген»), не найденную в растении дикого типа того же вида, штамма или сорта. «Трансген», упоминаемый в настоящем описании, имеет стандартное значение в биотехнологии и включает генетическую последовательность, которая была получена или изменена с помощью методики рекомбинантной ДНК или РНК и которая была встроена в растительную клетку. Трансген может включать генетические последовательности, полученные из растительной клетки. Обычно, трансген встроен в растение посредством манипуляций человека, таких как, например, трансформация, но может быть использован любой способ, известный специалисту.
«Полинуклеотид» относится к олигонуклеотиду, молекуле нуклеиновой кислоты или любому их фрагменту. Это может быть ДНК или РНК геномного или синтетического происхождения, двухцепочечная или одноцепочечная, и соединенная с углеводом, липидами, белком или другими веществами, для выполнения определенного действия, определенного в настоящем описании.
«Функционально связанный», используемый в настоящем описании, относится к функциональному взаимоотношению между двумя или более фрагментами нуклеиновой кислоты (например, ДНК). Обычно, он относится к функциональному взаимоотношению транскрипционного регуляторного элемента к транскрибируемой последовательности. Например, промотор функционально связан с кодирующей последовательностью, такой как определенный в настоящем описании полинуклеотид, если он стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке-хозяине. Обычно, промоторные транскрипционные регуляторные элементы, которые функционально связаны с транскрибируемой последовательностью, физически соединены с транскрибируемой последовательностью, т.е. являются cis-элементами. Однако для некоторых транскрипционных регуляторных элементов, таких как энхансеры, не нужно быть физически присоединенными или находиться в непосредственной близости от кодирующей последовательности, чью экспрессию они усиливают.
Термин «сигнальный пептид» относится к аминоконцевому полипептиду, предшествующему секретируемому зрелому белку. Сигнальный пептид отщепляется и, поэтому, не присутствует в зрелом белке. Сигнальные пептиды имеют функцию направления и trans-расположения секретируемых белков через клеточную мембрану. Сигнальный пептид также именуется сигнальной последовательностью.
Используемая в настоящем описании «трансформация» представляет собой приобретение новых генов в клетке посредством включения полинуклеотида.
Используемый в настоящем описании термин «лекарство» относится к любому соединению, которое может использоваться для лечения или предотвращения определенной болезни, примеры лекарств, которые могут быть введены в рецептуру с белком шелка по изобретению, включают, но не ограничены этим, белки, нуклеиновые кислоты, противоопухолевые агенты, анальгетики, антибиотики, противовоспалительные соединения (стероидные и нестероидные), гормоны, вакцины, меченые вещества и т.п.
Полипептиды
Под «по существу чистым полипептидом» в изобретении понимается полипептид, обычно отделенный от липидов, нуклеиновых кислот, других полипептидов и других примесных молекул, таких как воск, которые ассоциированы с полипептидом в его природной форме. За исключением других белков по изобретению, предпочтительно, чтобы по существу чистый полипептид был по крайней мере на 60% свободен, более предпочтительно по крайней мере на 75% свободен и более предпочтительно по крайней мере на 90% свободен от других компонентов, с которыми он ассоциирован в природе.
Термин «рекомбинантный» в контексте полипептида относится к полипептиду, который продуцируется клеткой, или в бесклеточной системе экспрессии в измененном количестве или с измененной скоростью, по сравнению с ним в природе. В одном варианте осуществления изобретения клетка представляет собой клетку, которая не продуцирует полипептид в природе. Однако клетка может представлять собой клетку, которая содержит не эндогенный ген, вызывающий продукцию измененного, предпочтительно, увеличенного количества полипептида. Рекомбинантный полипептид по изобретению включает полипептиды, которые не были отделены от других компонентов трансгенной (рекомбинантной) клетки или бесклеточной системы экспрессии, в которых он продуцируется, и полипептиды, продуцируемые в таких клетках или бесклеточных системах, которые впоследствии подвергаются очистке по крайней мере от некоторых других компонентов.
Термины «полипептид» и «белок» обычно используются взаимозаменяемо и относятся к одиночной полипептидной цепи, которая может быть модифицирована или может быть не модифицирована посредством добавления групп, не являющихся аминокислотами. Термины «белки» и «полипептиды», используемые в настоящем описании, также включают варианты, мутанты, модификации, аналоги и/или производные полипептидов по изобретению, описанные в настоящем документе.
% идентичности полипептида определяется при помощи GAP (Needleman and Wunsch, 1970) анализа (GCG программа) со штрафом за пропуск = 5 и пропуском за удлинение разрыва = 0,3. Запрашиваемая последовательность представляет собой по крайней мере 15 аминокислот в длину, и GAP анализ выравнивает две последовательности в области из по крайней мере 15 аминокислот. Более предпочтительно, чтобы запрашиваемая последовательность представляла собой по крайней мере 50 аминокислот в длину, и GAP анализ выравнивал две последовательности в области из по крайней мере 50 аминокислот. Более предпочтительно, чтобы запрашиваемая последовательность представляла собой по крайней мере 100 аминокислот в длину, и GAP анализ выравнивал две последовательности в области из по крайней мере 100 аминокислот. Еще более предпочтительно, чтобы запрашиваемая последовательность представляла собой, по крайней мере 250 аминокислот в длину, и GAP анализ выравнивал две последовательности в области из по крайней мере 250 аминокислот. Еще более предпочтительно, чтобы GAP анализ выравнивал две последовательности по всей их длине.
Используемый в настоящем описании «биологически активный фрагмент» является частью полипептида по изобретению, который поддерживает определенную активность полноразмерного полипептида, а именно способность использоваться для получения шелка. Биологически активные фрагменты могут быть любого размера, пока они поддерживают определенную активность.
В отношении определенного полипептида очевидно, что значения % идентичности выше, чем представленные выше, будут охватывать предпочтительные варианты осуществления изобретения. Поэтому, где применимо, в свете минимальных значений % идентичности, предпочтительно, чтобы полипептид содержал аминокислотную последовательность, которая является по крайней мере на 40%, более предпочтительно по крайней мере на 45%, более предпочтительно по крайней мере на 50%, более предпочтительно по крайней мере на 55%, более предпочтительно по крайней мере на 60%, более предпочтительно по крайней мере на 65%, более предпочтительно по крайней мере на 70%, более предпочтительно по крайней мере на 75%, более предпочтительно по крайней мере на 80%, более предпочтительно по крайней мере на 85%, более предпочтительно по крайней мере на 90%, более предпочтительно по крайней мере на 91%, более предпочтительно по крайней мере на 92%, более предпочтительно по крайней мере на 93%, более предпочтительно по крайней мере на 94%, более предпочтительно по крайней мере на 95%, более предпочтительно по крайней мере на 96%, более предпочтительно по крайней мере на 97%, более предпочтительно по крайней мере на 98%, более предпочтительно по крайней мере на 99%, более предпочтительно по крайней мере на 99,1%, более предпочтительно по крайней мере на 99,2%, более предпочтительно по крайней мере на 99,3%, более предпочтительно по крайней мере на 99,4%, более предпочтительно по крайней мере на 99,5%, более предпочтительно по крайней мере на 99,6%, более предпочтительно по крайней мере на 99,7%, более предпочтительно по крайней мере на 99,8% и еще более предпочтительно по крайней мере на 99,9%, идентична соответственной определенной SEQ ID NO.
Мутанты аминокислотных последовательностей полипептидов по настоящему изобретению могут быть получены путем встраивания соответствующих нуклеотидных замен в нуклеиновую кислоту по настоящему изобретению или путем синтеза in vitro желаемого полипептида. Такие мутанты включают, например, делеции, инсерции или замены остатков в аминокислотной последовательности. Может быть создана комбинация делеции, инсерции и замены для того, чтобы получить конечную конструкцию, при условии, что конечный полипептидный продукт обладает желаемыми характеристиками.
Мутантные (измененные) полипептиды могут быть получены с использованием любой известной в данной области методики. Например, полинуклеотид по изобретению может быть подвергнут мутагенезу in vitro. Такие методики мутагенеза in vitro включают субклонирование полинуклеотида в подходящий вектор, трансформацию вектора в «мутаторный» штамм, такой как E. coli XL-1 red (Stratagene) и размножение трансформированных бактерий до нужного числа поколений. В другом примере, полинуклеотиды по изобретению изменяют с использованием методик перетасовки ДНК, в общем описанных Harayama (1998). Эти методики перетасовки ДНК могут включать гены по изобретению возможно в дополнение к генам, родственным генам по настоящему изобретению, таким как гены шелка видов перепончатокрылых или сетчатокрылых, отличающихся от определенных видов, охарактеризованных в настоящем описании. Продукты, полученные из мутированной/измененной ДНК, могут легко быть скринированы с использованием методик, описанных в данном документе, для того, чтобы определить, могут ли они применяться в качестве белков шелка.
В разработке мутантов аминокислотной последовательности местоположение мутируемого участка и природа мутации будет зависеть от характеристики (характеристик), которые должны быть модифицированы. Участки мутации могут быть модифицированы индивидуально или по порядку, например, посредством (1) замещения сначала консервативными аминокислотными вариантами и затем более радикальными вариантами в зависимости от достигнутых результатов, (2) удаления целевого остатка или (3) встраивания других остатков, примыкающих к определенному участку.
Делеции аминокислотной последовательности обычно варьируют от, примерно, 1 до 15 остатков, более предпочтительно, от, примерно, 1 до 10 остатков и обычно, от, примерно, 1 до 5 непрерывных остатков.
Замещенные мутанты имеют по крайней мере один удаленный аминокислотный остаток в полипептидной молекуле и вставленный в это место другой остаток. Участки, представляющие наибольший интерес для заместительного мутагенеза, включают участки, идентифицированные как важные для функционирования. Другими участками, представляющими интерес, являются те, в которых определенные остатки, полученные из различных штаммов или видов, являются идентичными. Эти положения могут быть важны для биологической активности. Эти участки, особенно те, которые попадают внутрь последовательности из по крайней мере трех других идентичных консервативных участков, предпочтительно замещаются относительно консервативным способом. Такие консервативные замены показаны в таблице 1 под заголовком «Примеры замен».
Как изложено выше, часть некоторых полипептидов по изобретению имеет двуспиральную структуру. Двуспиральные структуры полипептидов отличаются гептадными повторами, представленными консенсусной последовательностью (abcdefg)n. В предпочтительном варианте осуществления изобретения часть полипептида, которая имеет структуру двойной спирали, содержит по крайней мере 10 копий гептадной последовательности abcdefg, и по крайней мере 25% аминокислот в положениях a и d представляют собой остатки аланина.

В предпочтительном варианте осуществления изобретения полипептид, который имеет двуспиральную структуру, содержит по крайней мере 12 идущих подряд копий, более предпочтительно по крайней мере 15 идущих подряд копий и еще более предпочтительно по крайней мере 18 идущих подряд копий гептад. В дальнейших вариантах осуществления изобретения полипептид, который имеет двуспиральную структуру, может иметь до, по меньшей мере, 28 копий гептад. Обычно, копии гептад будут повторяться парами. Однако они не обязательно должны быть точными парными повторами, например, как показано на фиг.5 и 6, несколько аминокислот могут быть найдены между двумя гептадами или могут быть найдены несколько укороченных гептад (см., например, белок Ксеноспира1 на фиг.5).
Руководство относительно аминокислотных замен, которые могут быть сделаны в полипептидах по изобретению, которые имеют структуру двойной спирали, представлено на фиг.5 и 6, а также в таблицах 6-10. В случае, где предсказанная пригодная аминокислотная замена на основе экспериментальных данных, приведенных в этом описании, находится так или иначе в противоречии с примерами замен, приведенными в таблице 1, является предпочтительным использование замены, основанной на экспериментальных данных.
Двуспиральные структуры полипептидов по изобретению имеют высокое содержание остатков аланина, особенно в аминокислотных позициях a, d и е гептад. Однако позиции b, c, f и g также имеют высокое содержание остатков аланина. В предпочтительном варианте осуществления изобретения по крайней мере 15% аминокислот в положениях a, d и/или е гептад являются остатками аланина, более предпочтительно по крайней мере 25%, более предпочтительно по крайней мере 30%, более предпочтительно по крайней мере 40% и еще более предпочтительно по крайней мере 50%. В дальнейшем предпочтительном варианте осуществления изобретения по крайней мере 25% аминокислот в обоих положениях a и d гептад представляют собой остатки аланина, более предпочтительно по крайней мере 30%, более предпочтительно по крайней мере 40% и еще более предпочтительно по крайней мере 50%. Кроме того, является предпочтительным, чтобы по крайней мере 15% из аминокислот в положениях b, c, f и g гептад представляли собой остатки аланина, более предпочтительно по крайней мере 20% и еще более предпочтительно по крайней мере 25%.
Обычно, гептады не будут содержать ни пролиновых, ни гистидиновых остатков. Кроме того, гептады будут содержать несколько (1 или 2), если вообще будут содержать, остатки фенилаланина, метионина, тирозина, цистеина, глицина или триптофана. Кроме аланина, общие (например, более чем 5%, более предпочтительно, более чем 10%) аминокислоты в гептадах включают лейцин (особенно в положениях b и d), серин (особенно в положениях b, e и f), глутаминовую кислоту (особенно в положениях c, e и f), лизин (особенно в положениях b, c, d, f и g), а также аргинин в положении g.
Полипептиды (и полинуклеотиды) по изобретению могут быть очищены (выделены) из многих видов перепончатокрылых и сетчатокрылых. Примеры перепончатокрылых включают, но не ограничены этим, любой вид из подотряда Apocrita (пчелы, муравьи и осы), который включает следующие семейства насекомых; Chrysididae (золотых ос), Formicidae (муравьев), Mutillidae (немок), Pompilidae (дорожных ос), Scoliidae, Vespidae (бумажных ос, одиночных ос, шершней), Agaonidae (наездников инжирных), Chalcididae (хальцид), Eucharitidae (эвхаритид), Eupelmidae (эупельмид), Pteromalidae (птеромалид), Evaniidae (таракановых наездников), Braconidae, Ichneumonidae (наездников), Megachilidae, Apidae, Colletidae, Halictidae и Melittidae (пчел-мелиттид). Примеры сетчатокрылых включают насекомых из следующих семейств: Mantispidae, Chrysopidae (златоглазок), Myrmeleontidae (муравьиных львов) и Ascalaphidae (аскалаф). Такие дальнейшие полипептиды (и полинуклеотиды) могут быть охарактеризованы с использованием тех же процедур, описанных в настоящем документе для шелков из Bombus terrestris, Myrmecia forficata, Oecophylla smaragdina и Mallada signata.
Кроме того, если желательно, неприродные аминокислоты или химические аминокислотные аналоги могут быть встроены в качестве замены или дополнения в полипептиды по настоящему изобретению. Такие аминокислоты включают, но не ограничены этим, D-изомеры известных аминокислот, 2,4-диаминобутировую кислоту, α-аминоизобутировую кислоту, 4-аминобутировую кислоту, 2-аминобутировую кислоту, 6-аминогексановую кислоту, 2-аминоизобутировую кислоту, 3-аминопропионовую кислоту, орнитин, норлейцин, норвалин, гидроксипролин, саркозин, цитруллин, гомоцитруллин, цистеиновую кислоту, трет-бутилглицин, трет-бутилаланин, фенилглицин, циклогексилаланин, β-аланин, фтор-аминокислоты, сконструированные аминокислоты, такие как β-метил аминокислоты, Сα-метил аминокислоты, Nα-метил аминокислоты и аналоги аминокислот в целом.
Также включенными в объем изобретения являются полипептиды по настоящему изобретению, которые модифицированы различным образом в ходе или после синтеза, например, посредством биотинилирования, бензилирования, гликозилирования, ацетилирования, фосфорилирования, амидирования, получения производных при помощи известных защитных/блокирующих групп, протеолитического расщепления, присоединения к молекуле антитела или другого клеточного лиганда и т.д. Эти модификации могут служить для увеличения стабильности и/или биологической активности полипептида по изобретению.
Полипептиды по настоящему изобретению могут быть получены различными путями, включая получение и выделение природных полипептидов, получение и выделение рекомбинантных полипептидов и химический синтез полипептидов. В одном варианте осуществления изобретения выделенный полипептид по настоящему изобретению получают при помощи культивирования клетки, способной экспрессировать полипептид при условиях, оптимальных для получения полипептида и выделения полипептида. Предпочтительная клетка для культивирования представляет собой рекомбинантную клетку по настоящему изобретению. Оптимальные условия для культуры включают, но не ограничены этим, оптимальные условия среды, биореактора, температуры, рН и оксигенации, которые позволяют получить полипептид. Оптимальная среда относится к любой среде, в которой культивируемая клетка продуцирует полипептид по настоящему изобретению. Такая среда обычно содержит водную среду, имеющую усваиваемые источники углерода, азота и фосфата, и соответствующие соли, минералы, металлы и другие питательные вещества, такие как витамины. Клетки по настоящему изобретению могут культивироваться в обычных ферментерах, встряхиваемых колбах, пробирках, микротитровочных планшетах и чашках Петри. Культивирование может проводиться при температуре, рН и содержании кислорода, подходящих для рекомбинантной клетки. Такие условия культивирования входят в компетенцию среднего специалиста в данной области.
Полинуклеотиды
Под «выделенным полинуклеотидом», включая ДНК, РНК или их комбинацию, одно- или двухцепочечным, в смысловой или антисмысловой ориентации или их комбинации, дцРНК, или чем-либо еще авторы изобретения понимают полинуклеотид, который по крайней мере частично отделен от полинуклеотидных последовательностей, с которыми он связан или соединен в природе. Предпочтительно, чтобы выделенный полинуклеотид являлся по крайней мере на 60% свободным, предпочтительно по крайней мере на 75% свободным и наиболее предпочтительно по крайней мере на 90% свободным от других компонентов, с которыми он связан в природе. Кроме того, в настоящем описании термин «полинуклеотид» используется взаимозаменяемо с термином «нуклеиновая кислота».
Термин «экзогенный» в контексте полинуклеотида относится к полинуклеотиду, когда тот присутствует в клетке или в бесклеточной системе экспрессии в измененном количестве по сравнению с его естественным состоянием. В одном варианте осуществления изобретения клетка представляет собой клетку, которая в природе не содержит полинуклеотид. Однако клетка может представлять собой клетку, которая содержит неэндогенный полинуклеотид, приводящий к измененному, предпочтительно, увеличенному количеству продукции кодируемого полипептида. Экзогенный полинуклеотид по изобретению включает полинуклеотиды, которые не были отделены от других компонентов трансгенной (рекомбинантной) клетки или бесклеточной системы экспрессии, в которой они присутствуют, и полинуклеотиды, полученные в таких клетках или бесклеточных системах, которые впоследствии очищены от по крайней мере некоторых других компонентов.
% идентичности полипептида определяется при помощи GAP (Needleman and Wunsch, 1970) анализа (GCG программа) со штрафом за создание разрыва = 5 и штрафом за удлинение разрыва = 0,3. Если не указано особо, то запрашиваемая последовательность представляет собой по крайней мере 45 нуклеотидов в длину, и GAP анализ выравнивает две последовательности в области из по крайней мере 45 нуклеотидов. Более предпочтительно, чтобы запрашиваемая последовательность представляла собой по крайней мере 150 нуклеотидов в длину, и GAP анализ выравнивал две последовательности в области из по крайней мере 150 нуклеотидов. Более предпочтительно, чтобы запрашиваемая последовательность представляла собой по крайней мере 300 нуклеотидов в длину, и GAP анализ выравнивал две последовательности в области из по крайней мере 300 нуклеотидов. Еще более предпочтительно, чтобы GAP анализ выравнивал две последовательности по всей их длине.
В отношении определенного полинуклеотида очевидно, что значения % идентичности выше, чем представленные выше, будут охватывать предпочтительные варианты осуществления изобретения. Поэтому, где применимо, в свете минимальных значений % идентичности, предпочтительно, чтобы полинуклеотид по изобретению содержал последовательность, которая является по крайней мере на 40%, более предпочтительно по крайней мере на 45%, более предпочтительно по крайней мере на 50%, более предпочтительно по крайней мере на 55%, более предпочтительно по крайней мере на 60%, более предпочтительно по крайней мере на 65%, более предпочтительно по крайней мере на 70%, более предпочтительно по крайней мере на 75%, более предпочтительно по крайней мере на 80%, более предпочтительно по крайней мере на 85%, более предпочтительно по крайней мере на 90%, более предпочтительно по крайней мере на 91%, более предпочтительно по крайней мере на 92%, более предпочтительно по крайней мере на 93%, более предпочтительно по крайней мере на 94%, более предпочтительно по крайней мере на 95%, более предпочтительно по крайней мере на 96%, более предпочтительно по крайней мере на 97%, более предпочтительно по крайней мере на 98%, более предпочтительно по крайней мере на 99%, более предпочтительно по крайней мере на 99,1%, более предпочтительно по крайней мере на 99,2%, более предпочтительно по крайней мере на 99,3%, более предпочтительно по крайней мере на 99,4%, более предпочтительно по крайней мере на 99,5%, более предпочтительно по крайней мере на 99,6%, более предпочтительно по крайней мере на 99,7%, более предпочтительно по крайней мере на 99,8% и еще более предпочтительно по крайней мере на 99,9%, идентична соответственной определяемой SEQ ID NO.
Полинуклеотиды по настоящему изобретению могут содержать по сравнению с природными молекулами одну или более мутаций, которые представляют собой делеции, инсерции или замены нуклеотидных остатков. Мутанты могут быть или встречающимися в природе (то есть выделенными из природного источника), или синтетическими (например, посредством проведения сайт-направленного мутагенеза на нуклеиновой кислоте).
Олигонуклеотиды и/или полинуклеотиды по изобретению гибридизуются с геном шелка по настоящему изобретению или областью, фланкирующей указанный ген, при жестких условиях. Термин «жесткие условия гибридизации» и т.п., используемый в настоящем описании, относится к параметрам, которые известны в данной области, включая вариации температуры гибридизации в зависимости от длины нуклеотида. Параметры гибридизации нуклеиновой кислоты могут быть найдены в ссылках, которые собирают такие методы, Sambrook, et al. (выше) и Ausubel, et al. (выше). Например, жесткие условия гибридизации, используемые в настоящем описании, могут относиться к гибридизации при 65°С в буфере для гибридизации (3,5×SSC, 0,02% Ficoll, 0,02% поливинилпирролидон, 0,02% бычьего сывороточного альбумина (БСА), 2,5 мМ NaH2PO4 (pH 7), 0,5% ДСН, 2 мМ ЭДТА) с последующими одной или более промывками в 0,2×SSC, 0,01% БСА при 50°С. В качестве альтернативы нуклеиновая кислота и/или олигонуклеотиды (которые также могут именоваться «праймерами» или «пробами») гибридизуются с областью генома насекомого, представляющего интерес, такого как геном медоносной пчелы, при условиях, используемых в методиках амплификации нуклеиновой кислоты, таких как ПЦР.
Олигонуклеотиды по настоящему изобретению могут представлять собой ДНК, РНК или их производные. Хотя термины полинуклеотид и олигонуклеотид имеют перекрывающееся значение, олигонуклеотиды представляют собой обычно относительно короткие одноцепочечные молекулы. Минимальным размером таких олигонуклеотидов является размер, требуемый для образования стабильного гибрида между олигонуклеотидом и комплементарной последовательностью на целевой молекуле нуклеиновой кислоты. Предпочтительно, чтобы олигонуклеотиды представляли собой по крайней мере 15 нуклеотидов, более предпочтительно по крайней мере 18 нуклеотидов, более предпочтительно по крайней мере 19 нуклеотидов, более предпочтительно по крайней мере 20 нуклеотидов, еще более предпочтительно по крайней мере 25 нуклеотидов в длину.
Обычно мономеры полинуклеотида или олигонуклеотида являются связанными фосфодиэфирными связями или их аналогами, образуя олигонуклеотиды, различающиеся по длине от относительно коротких мономерных единиц, например, 12-18, до нескольких сотен мономерных единиц. Аналоги фосфодиэфирных связей включают: фосфоротиоат, фосфородитиоат, фосфороселеноат, фосфородиселеноат, фосфороанилотиоат, фосфоранилидат, фосфорамидат.
Настоящее изобретение включает олигонуклеотиды, которые могут использоваться как, например, пробы для идентификации молекул нуклеиновых кислот, или как праймеры для получения молекул нуклеиновых кислот. Олигонуклеотиды по настоящему изобретению, используемые в качестве проб, обычно соединены с детектируемой меткой, такой как радиоизотоп, фермент, биотин, флуоресцентная молекула или хемилюминесцентная молекула.
Рекомбинантные векторы
Один вариант осуществления настоящего изобретения включает рекомбинантный вектор, который содержит по крайней мере одну выделенную полинуклеотидную молекулу настоящего изобретения, встроенную в любой вектор, способный доставить полинуклеотидную молекулу в клетку-хозяина. Такой вектор содержит гетерологичные полинуклеотидные последовательности, то есть полинуклеотидные последовательности, которые не найдены в природе граничащими с полинуклеотидными молекулами по настоящему изобретению и, предпочтительно, получены из видов, отличных от видов, из которых получены полинуклеотидные молекулы (получена полинуклеотидная молекула). Вектор может быть или РНК, или ДНК, или прокариотическим, или эукариотическим и обычно представляет собой транспозон (такой как описан в US 5792294), вирус или плазмиду.
Один тип рекомбинантного вектора содержит полинуклеотидную молекулу по настоящему изобретению, функционально связанную с экспрессионным вектором. Фраза оперативно связанный относится к встраиванию полинуклеотидной молекулы в экспрессионный вектор таким образом, что молекула способна экспрессироваться, будучи трансформированной в клетку-хозяина. Используемый в настоящем описании экспрессионный вектор является ДНК или РНК-вектором, который способен трансформировать клетку-хозяина и осуществлять экспрессию определенной полинуклеотидной молекулы. Предпочтительно, чтобы экспрессионный вектор имел способность к репликации в клетке-хозяине. Экспрессионные векторы могут быть или прокариотическими, или эукариотическими и представляют собой обычно вирусы или плазмиды. Экспрессионные векторы по настоящему изобретению включают любые векторы, которые функционируют (т.е. направляют экспрессию генов) в рекомбинантных клетках по настоящему изобретению, включая бактериальные, грибные, эндопаразитарные, членистоногих, животные и растительные клетки. Особенно предпочтительные экспрессионные векторы по настоящему изобретению могут направлять экспрессию генов в растительных клетках. Векторы по изобретению могут также использоваться для получения полипептида в бесклеточной системе экспрессии, такие системы хорошо известны в данной области.
В частности, экспрессионные векторы по настоящему изобретению содержат регуляторные последовательности, такие как последовательности для контроля транскрипции, последовательности для контроля трансляции, точки начала репликации и другие регуляторные последовательности, которые являются совместимыми с рекомбинантной клеткой и которые контролируют экспрессию молекул полинуклеотидов по настоящему изобретению. В частности, рекомбинантные молекулы по настоящему изобретению включают последовательности для контроля транскрипции. Последовательности для контроля транскрипции представляют собой последовательности, которые контролируют начало, продолжение и остановку транскрипции. Особенно важными последовательностями для контроля транскрипции являются те, которые контролируют начало транскрипции, такие как промоторные, энхансерные, операторные и репрессорные последовательности. Подходящие последовательности для контроля транскрипции включают любую последовательность для контроля транскрипции, которая может функционировать по крайней мере в одной из рекомбинантных клеток по настоящему изобретению. Ряд таких последовательностей, контролирующих транскрипцию, известен специалистам. Предпочтительные последовательности для контроля транскрипции включают последовательности, которые функционируют в клетках бактерий, дрожжей, членистоногих, растений или млекопитающих, такие как, но не ограниченные этим, tac, lac, trp, trc, oxy-pro, omp/lpp, rrnB, бактериофага лямбда, бактериофага T7, T7lac, бактериофага T3, бактериофага SP6, бактериофага SP01, металлотионеиновую, альфа-фактора дрожжей, алкоголь-оксидазы из Pichia, альфавирусных субгеномных промоторов (таких как субгеномные промоторы вируса Синдбис), гена устойчивости к антибиотикам, бакуловируса, вируса насекомых Heliothis zea, вируса коровьей оспы, вируса герпеса, поксвируса енотов, других поксвирусов, аденовируса, цитомегаловируса (такие как промежуточные ранние промоторы), вируса обезьян 40, ретровируса, актина, ретровирусного длинного концевого повтора, вируса саркомы Рауса, теплового шока, фосфатная и нитратная последовательности для контроля транскрипции, а также другие последовательности, способные контролировать экспрессию генов в прокариотических или эукариотических клетках.
Особенно предпочтительные последовательности для контроля транскрипции представляют собой промоторы, активные в управлении транскрипцией в растениях, или конститутивно или специфически для стадии и/или ткани, в зависимости от использования растения или его частей. Эти промоторы растений включают, но не ограничены этим, промоторы, имеющие конститутивную экспрессию, такие как 35S промотор мозаичного вируса цветной капусты (CaMV), промоторы для экспрессии в листьях, такие как промотор гена малой субъединицы рибулозо-бифосфат карбоксилазы, промоторы для экспрессии в корне, такие как промотор из гена глутамин-синтетазы, промоторы для экспрессии в семенах, такие как промотор круциферина А из Brassica napus, промоторы для экспрессии в клубне, такие как промотор пататина класса I из картофеля или промоторы для экспрессии в плодах, такие как промотор полигалактуроназы (PG) из томата.
Рекомбинантные молекулы по настоящему изобретению могут также (а) содержать сигналы секреции (т.е. последовательности нуклеиновой кислоты сигнального фрагмента) для того, чтобы дать возможность экспрессируемому полипептиду по настоящему изобретению секретироваться из клетки, которая продуцирует полипептид, и/или (б) содержать слитые последовательности, которые приводят к экспрессии молекул нуклеиновой кислоты по настоящему изобретению в виде слитых белков. Примеры подходящих сигнальных фрагментов включают любой сигнальный фрагмент, способный направлять секрецию полипептида по настоящему изобретению. Предпочтительные сигнальные фрагменты включают, но не ограничены этим, тканевый плазминогенный активатор (t-РА), интерферон, интерлейкин, гормон роста, сигнальные фрагменты гликопротеинов вирусного конверта, сигнальный пептид из Nicotiana nectarin (US 5939288), сигнал экстензина табака, сигнал олеозина - связывающего белка жировых телец из сои, сигнальный пептид вакуолярной основной хитиназы из Arabidopsis thaliana, а также природные сигнальные последовательности полипептида по изобретению. В дополнение молекула нуклеиновой кислоты по настоящему изобретению может быть соединена с фрагментом для слияния, направляющим кодируемый полипептид в протеосому, таким как убиквитин. Рекомбинантные молекулы могут также включать промежуточные и/или нетранслируемые последовательности, окружающие и/или находящиеся внутри последовательностей нуклеиновых кислот по настоящему изобретению.
Клетки-хозяева
Другой вариант осуществления настоящего изобретения включает рекомбинантную клетку, содержащую клетку-хозяина, трансформированную одной или более рекомбинантными молекулами по настоящему изобретению, или потомство этих клеток. Трансформация полинуклеотидной молекулы в клетку может быть выполнена любым способом, при помощи которого полинуклеотидная молекула может быть встроена в клетку. Методики трансформации включают, но не ограничены этим, трансфекцию, электропорацию, микроинъекцию, липофекцию, адсорбцию и слияние протопластов. Рекомбинантная клетка может оставаться в виде одиночных клеток или вырасти в ткань, орган или многоклеточный организм. Трансформированные полинуклеотидные молекулы по настоящему изобретению могут оставаться внехромосомными или могут быть встроенными в один или более участков на хромосоме трансформированной (т.е. рекомбинантной) клетки таким образом, чтобы сохранялась их способность к экспрессии.
Подходящие клетки-хозяева для трансформации включают любую клетку, которая может быть трансформирована полинуклеотидом по настоящему изобретению. Клетки-хозяева по настоящему изобретению или могут иметь эндогенную (т.е. природную) способность продуцировать полипептиды по настоящему изобретению, или могут быть способны продуцировать такие полипептиды после того, как они были трансформированы по крайней мере одной полинуклеотидной молекулой по настоящему изобретению. Клетки-хозяева по настоящему изобретению могут представлять собой любую клетку, способную продуцировать по крайней мере один белок по настоящему изобретению, и включают бактериальные, грибные (включая дрожжевые), паразитарные, членистоногих, животные и растительные клетки. Примеры клеток-хозяев включают Salmonella, Escherichia, Bacillus, Listeria, Saccharomyces, Spodoptera, Mycobacteria, Trichoplusia, BHK (почки детеныша хомячка) клетки, MDCK клетки, CRPK клетки, CV-1 клетки, COS (например, COS-7) клетки и Vero клетки. Дальнейшими примерами клеток-хозяев являются E. coli, включая производные K-12 E. coli; Salmonella typhi; Salmonella typhimurium, включая ослабленные штаммы; Spodoptera frugiperda; Trichoplusia ni; и не образующие опухоли клетки миобластов мыши G8 (например, ATCC CRL 1246). Дополнительные подходящие клетки-хозяева из млекопитающих включают другие клеточные линии из почек, другие клеточные линии из фибробластов (например, фибробластоидные клеточные линии из эмбрионов человека, мыши и цыпленка), миеломные клеточные линии, клетки яичников китайского хомячка, мышиные клетки NIH/3T3, LMTK клетки и/или HeLa клетки. Особенно предпочтительными клетками-хозяевами являются растительные клетки, такие как те, что имеются в доступе от Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (германской коллекции микроорганизмов и клеточных культур).
Методики рекомбинантных ДНК могут использоваться для улучшения экспрессии трансформированной полинуклеотидной молекулы посредством манипулирования, например, числом копий полинуклеотидной молекулы в клетке-хозяине, эффективностью, с которой эти молекулы полинуклеотидов транскрибируются, эффективностью, с которой транслируются полученные транскрипты, и эффективностью пост-трансляционных модификаций. Рекомбинантные методики, применимые для увеличения экспрессии молекул полинуклеотидов по настоящему изобретению, включают, но не ограничены этим, функциональное присоединение молекул полинуклеотидов к высококопийным плазмидам, интеграцию полинуклеотидной молекулы в одну или более хромосом клеток-хозяев, добавление к плазмидам последовательностей стабильности вектора, замены и модификации сигналов контроля транскрипции (например, промоторов, операторов, энхансеров), замены и модификации сигналов контроля трансляции (например, участки связывания рибосом, последовательности Шайн-Дальгарно), модификации молекул полинуклеотидов по настоящему изобретению для соответствия частоте встречаемости кодонов в клетке-хозяине и удаление последовательностей, которые дестабилизируют транскрипты.
Трансгенные растения
Термин «растение» относится к целым растениям, органам растения (например, листьям, стеблям, корням и т.д.), семенам, растительным клеткам и т.п. Растения, предполагаемые для применения на практике настоящего изобретения, включат как однодольные, так и двудольные. Растения-мишени включают, но не ограничены этим, следующие: злаки (пшеница, ячмень, рожь, овес, рис, сорго и другие зерновые); свеклу (сахарную свеклу и кормовую свеклу); фрукты, косточковые и ягодные (яблоки, груши, сливы, персики, миндаль, вишню, клубнику, малину и смородину); бобовые растения (бобы, чечевицу, горох, сою); масличные растения (рапс, горчицу, мак, оливки, подсолнечник, кокос, клещевину, какао-бобы, арахис); огуречные растения (кабачки, огурцы, дыни); волокнистые растения (хлопок, лен, пеньку, джут); цитрусовые (апельсины, лимоны, грейпфрут, мандарины); овощи (шпинат, салат, спаржу, капусту, морковь, лук, помидоры, картофель, перец); лавровые (авокадо, корицу, камфору); или растения, такие как маис, табак, орехи, кофе, сахарный тростник, чай, виноград, хмель, дерн, бананы и природные каучуконосные растения, а также декоративные растения (цветы, кустарники, широколиственные деревья и вечнозеленые растения, такие как хвойные).
Трансгенные растения, определенные в контексте настоящего изобретения, включают растения (а также части и клетки указанных растений) и их потомство, которые были генетически модифицированы с использованием рекомбинантных методик, чтобы получить по крайней мере один полипептид по настоящему изобретению в желаемом растении или органе растения. Трансгенные растения могут быть получены с применением методик, известных в данной области, таких как те, что, в общем, описаны в A. Slater et al., Plant Biotechnology - The Genetic Manipulation of Plants, Oxford University Press (2003) и P. Christou and H. Klee, Handbook of Plant Biotechnology, John Wiley and Sons (2004).
Полинуклеотид по настоящему изобретению может экспрессироваться конститутивно в трансгенных растениях в течение всех стадий развития. В зависимости от использования растения или органов растения, полипептиды могут экспрессироваться в зависимости от стадии развития растения. Кроме того, полинуклеотиды могут экспрессироваться тканеспецифическим образом.
В настоящем изобретении могут быть применены регуляторные последовательности, про которые известно или обнаружено, что они вызывают экспрессию гена, кодирующего представляющий интерес полипептид в растениях. Выбор используемых регуляторных последовательностей зависит от представляющих интерес растения-мишени и/или органа-мишени. Такие регуляторные последовательности могут быть получены из растений или вирусов растений или могут быть синтезированы химическим образом. Специалистам хорошо известны такие регуляторные последовательности.
Хорошо известны конститутивные промоторы растений. Дополнительно к ранее упомянутым промоторам некоторые подходящие промоторы включают, но не ограничены этим, промотор нопалин-синтетазы, промотор октопин-синтетазы, промотор CaM 35S, промотор рибулозо-1,5-бисфосфат карбоксилазы, pEmu на основе Adh1, Act1, промотор SAM синтетазы и Ubi-промоторы и промотор белка, связывающего хлорофиллы a/b. В качестве альтернативы может являться желательным иметь трансген(ы) с регулируемой экспрессией. Регулируемая экспрессия полипептидов возможна при помещении кодирующей последовательности белка шелка под контроль промоторов, которые являются тканеспецифическими, зависят от развития или индуцируемыми. Несколько тканеспецифических регулируемых генов и/или промоторов были найдены в растениях. Они включают гены, кодирующие запасные белки семян (такие как напин, круциферин, β-конглицинин, глицинин и фазеолин), зеин или белки жировых телец (такие как олеозин), или гены, вовлеченные в биогенез жирных кислот (включая ацил-переносящие белки, стеароил-АСР десатуразу и десатуразы жирных кислот (fad 2-1)), и другие гены, экспрессирующиеся в ходе развития зародыша (такие как Все4). Особенно применим для экспрессии, специфичной для семян, является промотор вицилина гороха. Другие применимые промоторы для экспрессии в зрелых листьях представляют собой промоторы, которые включаются в начале увядания, такие как SAG промотор из Arabidopsis). Класс промоторов для экспрессии в плодах, экспрессирующихся начиная с периода цветения и в течение развития плода по крайней мере до начала созревания, описан в US 4943574. Другие примеры тканеспецифических промоторов включают промоторы, которые направляют экспрессию в клубнях (например, промотор гена пататина) и в клетках волокон (примером регулируемого в ходе развития белка клетки волокна является белок Е6 волокна).
Другие регуляторные последовательности, такие как терминаторные последовательности и сигналы полиаденилирования, включают любую подобную последовательность, функционирующую как таковая в растениях, выбор которой будет очевиден для опытного специалиста. Область терминации, используемая в экспрессионной кассете, будет выбираться главным образом для удобства, поскольку похоже, что области терминации являются относительно взаимозаменяемыми. Используемая область терминации может быть одного происхождения с областью инициации транскрипции, может иметь одно происхождение с представляющей интерес полинуклеотидной последовательностью или может быть получена из другого источника. Область терминации может иметь природное происхождения или может быть частично или полностью синтетической. Подходящие области терминации доступны из Ti-плазмиды из A. tumefaciens, такие как области терминации октопин-синтетазы и нопалин-синтетазы, или из генов β-фазеолина, химически индуцируемого lant гена, pIN.
Доступны несколько методик для введения в растения-мишени экспрессионной конструкции, содержащей последовательность нуклеиновой кислоты, кодирующей представляющий интерес полипептид. Такие методики включают, но не ограничены этим, трансформацию протопластов с использованием кальциевого/полиэтиленгликолевого метода, электропорацию и микроинъекцию или бомбардировку (покрытыми оболочкой) частицами. В дополнение к так называемым методам прямой трансформации ДНК широко распространены системы трансформации, включающие векторы, такие как вирусные и бактериальные векторы (например, из генов Agrobacterium). После отбора и/или скрининга, протопласты, клетки или части растений, которые были трансформированы, могут быть регенерированы в целые растения с использованием методов, известных в данной области. Выбор методик трансформации и/или регенерации не является критически для этого изобретения.
Чтобы подтвердить присутствие трансгенов в трансгенных клетках или растениях, можно провести амплификацию с помощью полимеразной цепной реакции (ПЦР) или анализ методом Саузерн-блоттинга с применением способов, известных специалисту. Продукты экспрессии трансгенов могут быть детектированы различными путями в зависимости от природы продукта и включают Вестерн-блоттинг и ферментные методы анализа. Одним из особенно применимых путей для качественной оценки экспрессии белка и для детекции репликации в различных растительных тканях является репортерный ген, такой как GUS. После того как получены трансгенные растения, их можно выращивать для получения тканей или частей растения, имеющих желаемый фенотип. Ткань растения или его части могут быть собраны и/или собраны семена. Семена могут служить в качестве источника для выращивания дополнительных растений с тканями или частями, имеющими желаемые характеристики.
Трансгенные животные, кроме человека
Методики для получения трансгенных животных хорошо известны в данной области. Полезным общим учебником по этой тематике является Houdebine, Transgenic animals - Generation and Use (Harwood Academic, 1997).
Гетерологичная ДНК может быть встроена, например, в оплодотворенную яйцеклетку млекопитающего. Например, тотипотентные или плюрипотентные стволовые клетки могут быть трансформированы при помощи микроинъекции, преципитации фосфатом кальция, слияния липосом, ретровирусной инфекции и другими способами, затем трансформированные клетки встраивают в эмбрион, и эмбрион затем развивается в трансгенное животное. В особенно предпочтительном способе развивающиеся эмбрионы заражают ретровирусом, содержащим желаемую ДНК, и получают трансгенных животных из инфицированных эмбрионов. В наиболее предпочтительном способе, однако, соответствующие ДНК совместно вводят с помощью инъекции в пронуклеус или цитоплазму эмбрионов, предпочтительно на стадии одной клетки, и позволяют эмбрионам развиваться до взрослых трансгенных животных.
Другой способ, используемый для получения трансгенного животного, включает микроинъекцию нуклеиновой кислоты в яйцеклетки на пронуклеарной стадии при помощи стандартных способов. Инъецированные яйцеклетки затем культивируют перед переносом в фаллопиевы трубы псевдобеременных реципиентов.
Трансгенные животные могут также быть получены при помощи методики переноса ядер. При применении этого способа, проводят стабильную трансфекцию фибробластов из животных-доноров плазмидой, включающей кодирующие последовательности для интересующих связывающего домена или партнера связывания под контролем регуляторных последовательностей. Затем проводят слияние стабильных трансфектантов с безъядерными ооцитами, культивируют их и переносят в реципиентов женского пола.
Способы выделения и получения шелка
Белки шелка по настоящему изобретению могут быть выделены и очищены из рекомбинантных клеток, таких как растительные, бактериальные или дрожжевые клетки, получая указанный белок различными способами. В одном варианте осуществления изобретения, способ включает удаление нативных клеточных белков из гомогенизированных клеток/тканей/растений и т.д. при понижении рН и нагревании с последующим фракционированием сульфатом аммония. Кратко, все растворимые белки экстрагируются путем гомогенизации клеток/тканей/растений. Нативные белки удаляются осаждением при рН 4,7 и, затем, при 60°С. Полученный супернатант затем фракционируют 40% сульфатом аммония. Полученный в результате белок будет иметь чистоту порядка 95%. Дополнительная степень очистки может быть достигнута с помощью обычной гель- или аффинной хроматографии.
В другом примере клеточные лизаты обрабатывают высококонцентрированной кислотой, например, HCl или пропионовой кислотой, понижая рН до ~1-2 в течение 1 часа или больше, что солюбилизирует белки шелка, но осаждает другие белки.
Из раствора будут образовываться фибриллярные агрегаты при спонтанной самосборке белков шелка по изобретению, когда концентрация белка превышает критическое значение. Агрегаты можно собрать и механически вытянуть в макроволокна по способу O'Brien et al. (I. O'Brien et al., "Design, Synthesis and Fabrication of Novel Self-Assembling Fibrillar Proteins", в Silk Polymers: Materials Science and Biotechnology, pp.104-117, Kaplan, Adams, Farmer and Viney, eds., c.1994 при помощи American Chemical Society, Washington, D.C.).
Из-за своей природной двуспиральной вторичной структуры белки, такие как Ксеноспира1-4, BBF1-4, BAF1-4 и GAF1-4 будут спонтанно образовывать двуспиральную вторичную структуру при дегидратации. Как описано ниже, сила двойной спирали может быть увеличена посредством ферментативных или химических кросс-сшивок остатков лизина, находящихся близко друг от друга.
Волокна шелка и/или сополимеры по изобретению имеют низкие требования к обработке. Белки шелка по изобретению требуют минимальной обработки, например, кручения для образования прочного волокна, поскольку они спонтанно образуют прочные двойные спирали, которые могут быть усилены кросс-сшивкой, такой как лизиновая кросс-сшивка. Это является противоположностью рекомбинантных полипептидов шелка из B. mori и пауков, которые требуют сложных методик прядения для получения вторичной структуры (β-складчатой) и прочности волокна.
Однако волокна могут быть спрядены из растворов, имеющих свойства, характерные для жидкокристаллической фазы. Концентрация волокна, при которой может произойти фазовый переход, зависит от композиции белка и комбинации белков, присутствующих в растворе. Однако можно детектировать фазовый переход посредством наблюдения чистоты и двойного лучепреломления раствора. Начало жидкокристаллической фазы может быть обнаружено, когда раствор приобретает опалесцирующий вид и при просматривании через перекрестные поляризующие фильтры обнаруживается двойное лучепреломление.
По одной методике образования волокон, волокна могут быть экструдированы из белкового раствора через насадку в метанол до получения длины, достаточной для того, чтобы волокно могло быть подхвачено механическими устройствами. Затем волокно может вытягиваться такими механическими устройствами через раствор метанола, собираться и высушиваться. Способы для вытягивания волокна считаются хорошо известными в данной области.
Дальнейшие примеры способов, которые могут использоваться для получения шелковых волокон и/или сополимеров настоящего изобретения, описаны в US 2004/0170827 и US 2005/0054830.
В предпочтительном варианте осуществления изобретения волокна шелка и/или сополимеры по изобретению являются кросс-сшитыми. В одном варианте осуществления изобретения волокна шелка и/или сополимеры являются кросс-сшитыми с представляющими интерес поверхностью/предметом/продуктом и т.д. с использованием методик, известных в данной области. В другом варианте осуществления изобретения (или в сочетании с предыдущим вариантом осуществления изобретения) по крайней мере некоторые белки в волокнах шелка и/или сополимерах являются кросс-сшитыми друг с другом. Предпочтительно, чтобы белки шелка были кросс-сшиты через остатки лизина в белках. Такая кросс-сшивка может быть проведена с применением химических и/или ферментативных методик, известных в данной области. Например, ферментативная кросс-сшивка может быть катализирована лизил-оксидазой, в то время как неферментативные кросс-сшивки могут быть генерированы из гликозилированных остатков лизина (Reiser et al., 1992).
Антитела
Изобретение также представляет моноклональные или поликлональные антитела к полипептидам по изобретению или их фрагментам. Поэтому, настоящее изобретение представляет способ для получения моноклональных или поликлональных антител к полипептидам по изобретению.
Термин «специфически связывается» относится к способности антитела связывать по крайней мере один полипептид по настоящему изобретению, но не другие известные белки шелка.
Используемый в настоящем описании термин «эпитоп» относится к области полипептида по изобретению, которая связывается с антителом. Эпитоп может вводиться животному для получения антител против эпитопа, однако предпочтительно, чтобы антитела по настоящему изобретению специфично связывали область эпитопа в контексте всего полипептида.
Если желательно получить поликлональные антитела, то выбранное млекопитающее (например, мышь, кролик, коза, лошадь и т.д.) иммунизируют иммуногенным полипептидом по изобретению. Сыворотку из иммунизированного животного собирают и обрабатывают по известным методикам. Если сыворотка, содержащая поликлональные антитела, содержит антитела к другим антигенам, то поликлональные антитела могут быть очищены с помощью иммуно-аффинной хроматографии. Методики для получения и обработки поликлональной сыворотки известны в данной области науки. Для того чтобы такие антитела могли быть изготовлены, изобретение также представляет полипептиды по изобретению или их фрагменты, гаптенизированные другим полипептидом для применения в качестве иммуногенов в животных.
Моноклональные антитела, направленные против полипептидов по изобретению, могут также быть легко получены специалистом. Хорошо известна общая методика для создания моноклональных антител при помощи гибридом. Бессмертные клеточные линии, продуцирующие антитела, могут быть созданы посредством слияния клеток, а также с помощью других методик, таких как прямая трансформация В-лимфоцитов онкогенной ДНК или трансфекция вирусом Эпштейна-Барра. Панели полученных моноклональных антител могут быть скринированы для различных целей; т.е. на изотип и аффинность к эпитопу.
Альтернативная методика включает скрининг библиотек фагового дисплея, где, например, на поверхности оболочек фагов экспрессируются scFv фрагменты с большим разнообразием определяющих комплементарность участков (CDR). Эта методика хорошо известна в данной области науки.
Для целей этого изобретения термин «антитело», если не указано противоположное, включает фрагменты целых антител, которые сохраняют их способность связывать антиген-мишень. Такие фрагменты включают Fv, F(ab') и F(ab')2 фрагменты, а также одноцепочечные антитела (scFv). Кроме того, антитела и их фрагменты могут представлять собой гуманизированные антитела, например, описанные в ЕР-А-239400.
Антитела по изобретению могут быть связаны с твердой подложкой и/или упакованы в наборах в подходящий контейнер вместе с соответствующими реагентами, контролями, инструкциями и т.п.
Предпочтительно, чтобы антитела по настоящему изобретению имели детектируемую метку. Примеры детектируемых меток, которые позволяют прямое измерение связывания антитела, включают радиоактивные метки, флюорофоры, красители, магнитные шарики, хемилюминесцентные вещества, коллоидные частицы и т.п. Примеры меток, которые позволяют прямое измерение связывания, включают ферменты, где субстрат обеспечивает окрашенный или флюоресцентный продукт. Дополнительные примеры детектируемых меток включают ковалентно связанные ферменты, способные обеспечить детектируемый сигнал продукта после добавления подходящего субстрата. Примеры подходящих ферментов для использования в конъюгатах включают пероксидазу хрена, щелочную фосфатазу, малат-дегидрогеназу и т.п. Если они не являются доступными в продаже, такие конъюгаты антитела с ферментом легко получить с помощью методик, известных специалистам. Дальнейшие примеры детектируемых меток включают биотин, который с высокой аффинностью связывает авидин или стрептавидин; флюорохромы (например, фикобилипротеины, фикоэритрин и аллофикоцианины; флуоресцеин и Texas red), которые могут быть использованы с проточным цитофлюориметром; гаптены; и т.п. Предпочтительно, чтобы детектируемая метка позволяла прямое измерение в люминометре для микропланшет, например, биотина. Такие меченые антитела могут применяться для детекции полипептидов по изобретению в методиках, известных в данной области науки.
Композиции
Композиции настоящего изобретения могут включать «приемлемый носитель». Примеры таких приемлемых носителей включают воду, физиологический раствор, раствор Рингера, раствор декстрозы, раствор Хэнка и другие водные физиологически сбалансированные солевые растворы. Также могут применяться неводные носители, такие как нелетучие масла, кунжутное масло, этилолеат или триглицериды.
В одном варианте осуществления изобретения «приемлемый носитель» представляет собой «фармацевтически приемлемый носитель». Термин «фармацевтически приемлемый носитель» относится к молекулярным единицам и композициям, которые не вызывают аллергическую, токсическую или какую-либо еще негативную реакцию при введении животному, особенно млекопитающему, и более особенно, человеку. Полезные примеры фармацевтически приемлемых носителей или разбавителей включают, но не ограничены этим, растворители, дисперсионную среду, оболочки, стабилизаторы, защитные коллоиды, адгезивные вещества, загустители, тиксотропные агенты, проникающие агенты, комплексообразующие вещества, изотонические агенты и агенты, откладывающие абсорбцию, которые не влияют на активность полипептида по изобретению. Нужная текучесть может поддерживаться, например, посредством применения оболочки, такой как лецитин, или путем поддержания требуемого размера частиц в случае дисперсии или посредством применения поверхностно-активных веществ. В более общем смысле, полипептиды по изобретению могут быть совмещены с нетоксичными твердыми или жидкими добавками, которые соответствуют обычным рецептурным методикам.
Как указано выше, в некоторых вариантах осуществления полипептид, шелковое волокно и/или сополимер по изобретению используется в качестве фармацевтически приемлемого носителя.
Другие подходящие композиции описаны ниже с определенными ссылками на конкретное применение полипептидов по изобретению.
Применение
Белки шелка применимы для создания новых биоматериалов из-за своей выдающейся прочности и сопротивляемости. Однако на сегодняшний день белки волокон пауков и насекомых представляют собой крупные белки (более 100 кД), и их аминокислотная последовательность содержит множественные повторы. Эти белки кодируются протяженными генами, содержащими редко встречающиеся кодоны, что делает особенно трудным их получение в рекомбинантных системах. По сравнению с ними белки шелка по изобретению являются короткими и не содержат повторов. Эти свойства генов, кодирующих эти белки, делают эти белки особенно привлекательными для рекомбинантного получения новых биоматериалов.
Белки шелка, шелковые волокна и сополимеры по изобретению могут применяться в обширном и разнообразном ряду медицинских, военных, промышленных и коммерческих приложений. Волокна могут использоваться для производства медицинских устройств, таких как шовный материал, кожный трансплантат, матрицы для роста клеток, заместители связок и хирургическая сетка, и в широком спектре промышленных и коммерческих продуктов, таких как, например, кабель, канат, сеть, леска, одежная ткань, подкладка бронезащитных жилетов, контейнерная ткань, рюкзаки, ранцы, ремешки сумок или дамских сумок, адгезивный связывающий материал, неадгезивный связывающий материал, ременный материал, тентовая ткань, непромокаемый брезент, покрытия для бассейнов, покрытия для автомобилей, огораживающий материал, изоляционный материал, строительный материал, материал для защиты от атмосферных воздействий, гибкий разделительный материал, спортивное снаряжение; и, в действительности, в практически любом применении волокна или ткани, для которой прочность при растяжении и эластичность являются желательными характеристиками. Белки шелка, шелковые волокна и/или сополимеры по настоящему изобретению также имеют применение в композициях для предметов личной гигиены, таких как косметические средства, средства для ухода за кожей, средства для ухода за волосами и окрашивания волос; и при покрытии оболочкой частиц, таких как пигменты.
Белки шелка могут применяться в своей нативной форме или могут быть модифицированы, образуя производные, которые обеспечивают более выгодный эффект. Например, белки шелка могут быть модифицированы путем соединения с полимером, чтобы снизить их аллергенность, как описано в US 5981718 и US 5856451. Подходящие модифицирующие полимеры включают, но не ограничены этим, полиалкиленоксиды, поливиниловые спирты, поликарбоксилаты, поливинилпирролидон и декстраны. В другом примере, белки шелка могут быть модифицированы при помощи избирательного расщепления и соединения другими белковыми модификаторами. Например, белки шелка могут быть расщеплены на более мелкие пептидные единицы при помощи обработки кислотой при повышенной температуре, примерно, 60°С. Пригодные кислоты включают, но не ограничены этим, разведенную хлористоводородную, серную или фосфорную кислоты. В качестве альтернативы расщепление белков шелка может быть осуществлено посредством обработки основанием, таким как гидроксид натрия, или может быть применен ферментативный гидролиз с использованием соответствующей протеазы.
Далее белки могут быть модифицированы, чтобы обеспечить рабочие качества, которые являются благоприятными в конкретном применении для средств личной гигиены. Модификация белков для применения в средствах личной гигиены хорошо известна в данной области науки. Например, обычно используемые способы описаны в US 6303752, US 6284246 и US 6358501. Примеры модификаций включают, но не ограничены этим, этоксилирование для улучшения образования эмульсии воды в масле, силоксилирование для обеспечения липофильных свойств и этерификацию для помощи в совместимости в композициях мыла и детергентов. Дополнительно, из белков шелка могут быть получены производные с функциональными группами, включающими, но не ограниченными этим, амины, оксираны, цианаты, эфиры карбоновых кислот, силиконовые сополиолы, силоксановые спирты, четвертичные алифатические амины, уретаны, полиакриламиды, эфиры дикарбоновых кислот и галогениды. Также могут быть получены производные белков шелка посредством реакции с дииминами и путем образования солей металлов.
В соответствии с приведенными выше определениями «полипептида» (и «белка») такие производные молекул или модифицированные молекулы также именуются в настоящем описании в широком смысле как «полипептиды» и «белки».
Белки шелка могут быть спрядены вместе и/или скручены или сплетены с другими типами волокон. Примеры включают, но не ограничены этим, полимерные волокна (например, полипропилен, нейлон, полиэфир), волокна и шелк из других растительных и животных источников (например, хлопка, шерсти, Bombyx mori или шелка пауков), и стекловолокно. Предпочтительный вариант осуществления настоящего изобретения представляет собой шелковое волокно, сплетенное с 10% полипропиленового волокна. Настоящее изобретение предполагает, что получение таких комбинаций волокон может быть легко осуществлено на практике для усиления любых желательных характеристик, например, внешнего вида, мягкости, веса, долговечности, водоотталкивающих свойств, улучшенной себестоимости, что обычно может быть желательным в изготовлении и получении волокон для медицинских, промышленных или коммерческих приложений.
Средства личной гигиены
Косметические композиции и композиции средств личной гигиены могут представлять собой безводные композиции, содержащие эффективное количество белка шелка в косметически приемлемой среде. Применение этих композиций включает, но не ограничено этим, уход за кожей, очищение кожи, декоративную косметику и продукцию против морщин. Эффективное количество белка шелка для косметических композиций и композиций средств личной гигиены в настоящем описании определено как количественное соотношение от, примерно, 10-4 до, примерно, 30% по весу, но, предпочтительно, от, примерно, 10-3 до 15% по весу, относительно общего веса композиции. Это количественное соотношение может варьировать как функция вида косметической композиции или композиции для ухода за кожей. Подходящие композиции для косметически приемлемой среды описаны в US 6280747. Например, косметически приемлемая среда может содержать жир в количественном соотношении обычно от, примерно, 10 до, примерно, 90% по весу относительного общего веса композиции, где жировая фаза содержит по крайней мере одно жидкое, твердое или полутвердое жировое вещество. Жировое вещество включает, но не ограничено этим, масла, воски, смолы и так называемые пастообразные жиры. В качестве альтернативы композиции могут находиться в виде стабильных дисперсий, таких как эмульсии вода-в-масле или масло-в-воде. Дополнительно, композиции могут содержать одну или больше общеупотребительных косметических или дерматологических добавок или адъювантов, включающих, но не ограниченных этим, антиоксиданты, консерванты, наполнители, поверхностно-активные вещества, солнцезащитные средства против спектров А и В ультрафиолета, отдушки, загустители, увлажняющие агенты и анионные, неионные и амфотерные полимеры, и красители или пигменты.
Эмульгированные косметические средства, которые получаются с применением эмульгированных материалов, содержащих по крайней мере один белок шелка по настоящему изобретению, включают, например, очищающую косметику (туалетное мыло, средство для умывания лица, шампунь, ополаскиватель и т.п.), средства для ухода за волосами (краску для волос, косметику для волос и т.п.), основные косметические средства (обычный крем, эмульсию, крем для бритья, кондиционер, одеколон, лосьон для бритья, косметическое масло, маску для лица и т.п.), декоративную косметику (тональный крем, карандаш для бровей, крем для век, тени для век, тушь и т.п.), ароматические косметические средства (духи и т.п.), косметику для загара и солнцезащитную (крем для и против загара, лосьон для и против загара, масло для и против загара и т.п.), косметику для ногтей (крем для ногтей и т.п.), средства для подводки глаз (подводку для глаз и т.п.), косметические средства для губ (помаду, крем для губ и т.п.), продукты для ухода за полостью рта (зубную пасту и т.п.), косметические средства для ванны (продукты для ванны и т.п.) и т.п.
Косметические композиции могут быть также в виде продуктов для ухода за ногтями, такими как лак для ногтей. Лаки для ногтей определены в настоящем описании, как композиции для лечения и окраски ногтей, содержащие эффективное количества белка шелка в косметически приемлемой среде. Эффективное количество белка шелка для использования в лаке для ногтей определено в настоящем описании, как количественное соотношение от, примерно, 10-4 до, примерно, 30% по весу относительно общего веса лака. Компоненты косметически приемлемой среды для лаков для ногтей описаны в US 6280747. Лак для ногтей обычно содержит растворитель и вещество, образующее пленку, такое как производные целлюлозы, производные поливинила, акриловые полимеры или сополимеры, виниловые сополимеры и полиэфирные полимеры. Композиция может также содержать органический или неорганический пигмент.
Композиции по уходу за волосами определены в настоящем описании, как композиции для лечения волос, включая, но не ограничиваясь этим, шампуни, кондиционеры, лосьоны, аэрозоли, гели и муссы, содержащие эффективное количество белка шелка в косметически приемлемой среде. Эффективное количество белка шелка для использования в композиции для ухода за волосами определено в настоящем описании, как количественное соотношение от, примерно, 10-2 до, примерно, 90% по весу относительно общего веса композиции. Компоненты косметически приемлемой среды для композиций для ухода за волосами описаны в US 2004/0170590, US 6280747, US 6139851 и US 6013250. Например, эти продукты для ухода за волосами могут быть водными, спиртовыми или водно-спиртовыми растворами, спирт, предпочтительно, является этанолом или изопропанолом, в количественном соотношении от, примерно, 1 до, примерно, 75% по весу относительно общего веса, для водно-спиртовых растворов. Дополнительно, композиции для ухода за волосами могут содержать одну или более из общеупотребительных косметических или дерматологических добавок или адъювантов, данных выше.
Композиции для окрашивания волос определены в настоящем описании, как композиции для придания цвета, окрашивания или отбеливания волос, содержащие эффективное количества белка шелка в косметически приемлемой среде. Эффективное количество белка шелка для использования в композиции для окрашивания волос определено в настоящем описании, как количественное соотношение от, примерно, 10-4 до, примерно, 60% по весу относительно общего веса композиции. Компоненты косметически приемлемой среды для композиций для окрашивания волос описаны в US 2004/0170590, US 6398821 и US 6129770. Например, композиции для окрашивания волос обычно содержат смесь неорганических пероксид-содержащих агентов для окисления красителя и окисляемый окрашивающий агент. Окисляющий агент для красителя на основе пероксида наиболее часто является пероксидом водорода. Способные к окислению агенты для окрашивания волос образуются посредством окислительного присоединения первичных интермедиатов (например, п-фенилендиаминов, п-аминофенолов, п-диаминопиридинов, гидроксииндолов, аминоиндолов, аминотимидинов или цианофенолов) с вторичными интермедиатами (например, фенолами, резорцинами, м-аминофенолами, м-фенилендиаминами, нафтолами, пиразолонами, гидроксииндолами, катехинами или пиразолами). Дополнительно, композиции для окрашивания волос могут содержать окисляющие кислоты, комплексообразующие вещества, стабилизаторы, загустители, буферные носители, поверхностно-активные вещества, растворители, антиоксиданты, полимеры, неокислительные красители и кондиционеры.
Белки шелка также могут использоваться для покрытия пигментов и косметических частиц в целях улучшения дисперсности частиц для применения в косметике и в покрывающих композициях. Косметические частицы определены в настоящем описании как вещества в виде частиц, такие как пигменты или инертные частицы, которые используются в косметических композициях. Подходящие пигменты и косметические частицы включают, но не ограничены этим, неорганические цветные пигменты, органические пигменты и инертные частицы. Неорганические цветные пигменты включают, но не ограничены этим, оксид титана, оксид цинка и оксиды железа, магния, кобальта и алюминия. Органические пигменты включают, но не ограничены этим, D&C Red № 36, D&C Orange № 17, кальциевые пигменты из D&C Red № 7, 11, 31 и 34, бариевый пигмент из D&C Red № 12, стронциевый пигмент из D&C Red № 13, алюминиевый пигмент из FD&C Yellow № 5 и частицы черного красителя. Инертные частицы включают, но не ограничены этим, карбонат кальция, силикат алюминия, силикат кальция, силикат магния, слюду, тальк, сульфат бария, сульфат кальция, порошковый NylonТМ, перфторированные алканы и другие инертные пластики.
Белки шелка могут также использоваться в зубной нити (см., например, US 2005/0161058). Зубная нить может быть нитью из моноволокна или нитью из мультиволокон и волокна могут быть скручены или нет. Зубная нить может быть упакована в виде индивидуальных отрезков или на бобине с режущей кромкой, чтобы отрезать кусочки любой желаемой длины. Зубная нить может быть представлена в разнообразных формах, отличных от волокон, таких как, но не ограниченных этим, полоски и листки и т.п. Нить может быть покрыта различными материалами, такими как, но не ограниченными этим, воск, политетрафторэтилированная моноволоконная нить для зубной нити.
Белки шелка могут также использоваться в мыле (см., например, US 2005/0130857).
Покрытие пигментов и косметических частиц
Эффективное количество белка шелка для применения в покрытии пигментов и косметических частиц определено в настоящем описании как количественное соотношение от, примерно, 10-4 до, примерно, 50%, предпочтительно, от, примерно, 0,25 до, примерно, 15% по весу относительно сухого веса частицы. Оптимальное количество белка шелка, которое будет использоваться, зависит от пигмента или косметических частиц, которые покрывают оболочкой. Например, количество белка шелка, используемое для покрытия неорганических цветных пигментов, находится, предпочтительно, между, примерно, 0,01% и 20% по весу. В случае органических пигментов, предпочтительное количество белка шелка находится между, примерно, 1% и, примерно, 15% по весу, в то время как для инертных частиц, предпочтительное количество находится между, примерно 0,25% и, примерно, 3% по весу. Способы получения покрытых оболочкой пигментов и частиц описаны в US 5643672. Эти способы включают: добавление водного раствора белка шелка к частицам при барабанной обработке или перемешивании, образование смеси белка шелка и частиц и высушивание, напыление белка шелка на частицы или лиофилизация взвеси белка шелка и частиц. Эти покрытые оболочной пигменты и косметические частицы могут использоваться в косметических рецептурах, красках, чернилах и т.п.
Медико-биологическое применение
Белки шелка могут применяться в качестве покрытия на перевязке для того, чтобы ускорить заживление раны. Для этого применения перевязочный материал покрывается эффективным количеством белка шелка. Для ранозаживляющей перевязки эффективное количество белка шелка в настоящем изобретении определено как количественное соотношение от, примерно, 10-4 до, примерно, 30% по весу относительно веса перевязочного материала. Материал для нанесения покрытия может быть мягкой, биологически инертной, пористой тканью или волокном. Примеры включают, но не ограничены этим, хлопок, шелк, вискозу, ацетат, акрил, полиэтилен, полиэфир и их комбинации. Нанесение покрытия на ткань или волокно может быть выполнено при помощи ряда способов, известных в данной области. Например, материл для нанесения покрытия может быть погружен в водный раствор, содержащий белок шелка. В качестве альтернативы, раствор, содержащий белок шелка, может быть нанесен при распылении на поверхность материала для нанесения покрытия с использованием пистолета-распылителя. Дополнительно, раствор, содержащий белок шелка, может быть нанесен на поверхность с использованием процесса печати на накатных валиках. Перевязочный материал для ран может включать другие добавки, включающие, но не ограниченные эти, дезинфицирующие средства, такие йод, йодид калия, повидон-йод, акринол, пероксид водорода, бензалкония хлорид и хлоргексидин; агенты, ускоряющие заживление, такие как аллантоин, дибукаина гидрохлорид и хлорфениламина малат; сосудосуживающие агенты, такие как нафазолина гидрохлорид; вяжущие агенты, такие как оксид цинка; и образующие струп агенты, такие как борная кислота.
Белки шелка по настоящему изобретению могут также применяться в виде пленки как материал для раневой повязки. Применение белков шелка в виде аморфной пленки в качестве материала для раневой повязки описано в US 6175053. Аморфная пленка содержит плотную и непористую пленку со степенью кристаллизации ниже 10%, которая содержит эффективное количество белка шелка. Для ранозаживляющей пленки эффективное количество белка шелка определено в настоящем описании как находящееся между, примерно, 1 и 99% по весу. Пленка может также содержать другие компоненты, включая, но не ограничиваясь этим, другие белки, такие как серицин и дезинфицирующие средства, агенты, ускоряющие заживление, сосудосуживающие агенты, вяжущие агенты и образующие струп агенты, описанные выше. Другие белки, такие как серицин, могут составлять от 1 до 99% по весу композиции. Количество всех других перечисленных ингредиентов обычно находится ниже 30% по весу, более предпочтительно, между, примерно, 0,5 и 20% по весу композиции. Пленка для раневой повязки может быть получена путем растворения вышеупомянутых материалов в водном растворе, удаления нерастворимых частиц фильтрованием или центрифугированием, и застывания раствора на мягкой твердой поверхности, такой как акриловая пластинка, с последующей сушкой.
Белки шелка по настоящему изобретению могут также использоваться в шовном материале (см., например, US 2005/0055051). Такой шовный материал может быть одним из составляющих оплетки, изготовленной из высокомолекулярных волокон и шелковых волокон. Полиэтилен предоставляет прочность. Полиэфирные волокна могут быть сплетены с высокомолекулярным полиэтиленом, чтобы обеспечить улучшенные затягивающие свойства. Шелк может быть представлен в контрастирующем цвете, чтобы предоставить знак для лучшего распознавания и идентификации шовного материала. Шелк также больше соответствует тканям организма, чем другие волокна, позволяя отрезать концы близко к узлу, не беспокоясь об отрицательном взаимодействии между концами шовного материала и окружающей тканью. Манипуляции с высокопрочным шовным материалом также могут быть улучшены с применением различных материалов для создания оболочки шовного материала. Этот шовный материал имеет преимущество в том, что он обладает прочностью шовного материала Ethibond № 5, хотя имеет диаметр, ощущение и связывающую способность шовного материала № 2. В результате шовный материал является идеальным для большинства ортопедических процедур, таких как восстановление вращательной манжеты, восстановление ахиллесова сухожилия, восстановление коленного сухожилия, воссоздание передних и задних крестовидных связок, процедуры воссоздания бедренного и плечевого суставов и замещение шовного материала, используемого в фиксаторе или с фиксатором. Шовный материал может быть без оболочки или может быть покрытым воском (пчелиным воском, нефтяным парафином, полиэтиленовым воском или другими), силиконом (Dow Corning silicone fluid 202A или другими), силиконовыми резинами, PBA (полибутиловой кислотой), этилцеллюлозой (Filodel) или другими оболочками, для улучшения скользящих свойств плетения, надежности узлов или абразивной устойчивости, например.
Белки шелка по настоящему изобретению могут также применяться в стентах (см., например, US 2004/0199241). Например, представлен стент-трансплантат, который включает эндолюминальный стент и трансплантат, где стент-трансплантат включает шелк. Шелк вызывает ответ в хозяине, который получает стент-трансплантат, где ответ может привести к усиленной адгезии между шелковым стент-трансплантатом и тканью хозяина, которая примыкает к шелку шелкового стент-трансплантата. Шелк может быть прикреплен к трансплантату различными способами, например, путем вплетания шелка в трансплантат или путем налипания шелка на трансплантат (например, с помощью клея или шовного материала). Шелк может иметь форму нити, жгута, полотна, порошка и т.д. Что касается расположения шелка на стент-трансплантате, шелк может быть прикреплен только к внешней поверхности стента, и/или шелк может быть прикреплен к дистальным областям стент-трансплантата, для того, чтобы содействовать закреплению этих дистальных областей в окружающих тканях хозяина. Может применяться широкое разнообразие стент-трансплантатов в контексте настоящего изобретения в зависимости от места и природы желаемого лечения. Стент-трансплантаты могут быть, например, разветвленными или трубчатыми трансплантатами, цилиндрическими или конусовидными, само-расширяющимися или баллон-расширяющимися, цельными или составными и т.д.
В дополнение к шелку стент-трансплантат может содержать оболочку на части или всем шелке, где оболочка разрушается при введении стент-трансплантата в хозяина, таким образом, оболочка задерживает контакт между шелком и хозяином. Подходящие оболочки включают без ограничений желатин, деградируемые полиэфиры (например, PLGA (сополимер молочной и гликолевой кислот), PLA (полимолочную кислоту), MePEG-PLGA (метил-полиэтиленгликоль-сополимер молочной и гликолевой кислот), PLGA-PEG-PLGA (сополимер молочной и гликолевой кислот-полиэтиленгликоль-сополимер молочной и гликолевой кислот) и их сополимеры и смеси), целлюлозу и производные целлюлозы (например, гидроксипропил-целлюлозу), полисахариды (например, гиалуроновую кислоту, декстран, декстран сульфат, хитозан), липиды, жирные кислоты, эфиры сахаров, эфиры нуклеиновых кислот, полиангидриды, полиортоэфиры и поливиниловый спирт (PVA). Содержащий шелк стент-трансплантат может содержать биологически активный агент (лекарственное соединение), где агент высвобождается из стент-трансплантата и затем вызывает усиленный клеточный ответ (например, клеточное или внеклеточное отложение матрикса) и/или фиброзный ответ в хозяине, которому введен стент-трансплант.
Белки шелка по настоящему изобретению могут применяться как матрица для образования связок и сухожилий ex vivo (см., например, US 2005/0089552). Матрица на основе шелковых волокон может быть высеяна с плюрипотентными клетками, такими как клетки стромы костного мозга (BMSC). Созданные с помощью биоинженерных способов связки или сухожилия выгодно отличаются клеточной ориентацией и/или сложной пространственной структурой матрицы в направлении прилагаемых механических усилий, и также продукцией специфических маркеров связок или сухожилий, включая коллаген I типа, коллаген III типа и фибронектиновые белки по оси механической нагрузки, создаваемой механическими усилиями или стимулом, если прилагаются такие усилия. В предпочтительном варианте осуществления изобретения связки или сухожилия отличаются наличием пучков волокон, которые упорядочены в спиральную систему. Некоторые примеры связок и сухожилий, которые могут быть получены, включают передние крестовидные связки, задние крестовидные связки, сухожилия манжетной мышцы, срединные коллатеральные связки локтя и колена, сухожилия сгибающей мышцы руки, латеральные связки щиколотки и сухожилия и связки челюсти или височно-нижнечелюстного сустава. Другие ткани, которые могут быть получены по способам настоящего изобретения, включают хрящ (как суставный, так и менисковый), кость, мышцы, кожу и кровеносные сосуды.
Белки шелка по настоящему изобретению могут также использоваться в гидрогелях (см., например, US 2005/0266992). Гидрогели фиброинов шелка могут отличаться открытой структурой пор, которая позволяет их применение в качестве каркасов в тканевой инженерии, субстрата для клеточной культуры, повязок для ран и ожогов, заменителей мягких тканей, наполнителей костей и также как твердый носитель для фармацевтически или биологически активных соединений.
Белки шелка также могут применяться в дерматологических композициях (см., например, US 2005/0019297). Кроме того, белки шелка по изобретению и их производные могут также применяться в композициях с замедленным высвобождением (см., например, US 2004/0005363).
Текстильные изделия
Белки шелка по настоящему изобретению могут также наноситься на поверхность волокон для последующего применения в текстильных изделиях. Это обеспечивает монослой белковой пленки на волокне, давая в результате гладкую поверхность. US 6416558 и US 5232611 описывают добавление конечного покрытия на волокна. Способы, описанные в этих документах, представляют примеры различных вариантов конечной обработки волокна, для обеспечения хороших тактильных свойств и гладкой поверхности. Для этого применения волокно покрывают эффективным количеством белка шелка. Для нанесения оболочки на волокно для использования в текстильных изделиях эффективное количество белка шелка определено в настоящем описании как количественное соотношение от, примерно, 1 до, примерно 99% по весу относительно веса волокна. Волокно включает, но не ограничено этим, текстильные волокна из хлопка, полиэфиров, таких как вискоза и LycraТМ, нейлона, шерсти и других природных волокон, включая природный шелк. Композиции, подходящие для нанесения белка шелка на волокно, могут включать сорастворители, такие как этанол, изопропанол, гексафторанолы, изотиоцианоуранаты и другие полярные растворители, которые могут быть смешаны с водой с образованием растворов или микроэмульсий. Содержащий белок шелка раствор может быть нанесен на волокно при распылении, или волокно может быть погружено в раствор. Хотя это и не является необходимым, предпочтительной является мгновенная сушка покрытого материала. Альтернативный протокол представляет собой нанесение композиции белка шелка на сплетенные волокна. Идеальным вариантом осуществления этого применения является использованием белков шелка для покрытия оболочкой тянущихся переплетений, таких какие используются для чулок.
Композитные материалы
Волокна шелка могут быть добавлены к полиуретану, другим смолам и термопластическим наполнителям для получения панельных плит и другого конструкционного материала или формованной мебели и покрытия столов, которые заменяют деревянные и древесно-стружечные плиты. Композиты могут также использоваться в строительстве и автомобилестроении, особенно для плоских крыш и дверных панелей. Шелковые волокна усиливают смолу, делая материал более прочным и позволяя получать облегченные конструкции, которые обладают равной или большей прочностью, чем другие древесно-стружечные плиты и композитные материалы. Волокна шелка могут быть выделены и добавлены в синтетическую смолу, образующую композит, или могут использоваться в сочетании с полученными из растений белками, крахмалом и маслами для получения композитных материалов на биологической основе. Процессы для получения таких материалов описаны в JP 2004284246, US 2005175825, US 4515737, JP 47020312 и WO 2005/017004.
Добавки в бумагу
Свойства волокон шелка по изобретению могут добавить прочность и качество текстуры при изготовлении бумаги. Шелковая бумага может быть изготовлена путем добавления шелковых нитей в хлопковую целлюлозу для получения особо мягкой бумаги ручной выделки, используемой для заворачивания подарков, чехлов для ноутбуков, пакетов для покупок. Способы для получения бумажной продукции, которая может включать белки шелка по изобретению, в общем описаны в JP 2000139755.
Материалы с улучшенными свойствами
Шелки по изобретению имеют значительную прочность и выделяются среди других шелков способностью сохранять свойства при намокании (Hepburn et al., 1979).
Интенсивно развивающейся областью в одежно-текстильной промышленности являются технические и «интеллектуальные» ткани. Существует повышающийся спрос на здоровую, высокофункциональную, экологичную и индивидуализированную текстильную продукцию. Волокна, такие как волокна по изобретению, которые не изменяют свойств при намокании и, в особенности, сохраняют прочность и растяжимость, являются пригодными для функциональной одежды для спорта и досуга, а также рабочей одежды и защитной одежды.
Разработки в оружейных технологиях и технических средствах наблюдения представляют собой побудительные нововведения в средствах индивидуальной защиты и боевых системах и структурах. Кроме стандартных требований, таких как устойчивость материала к продолжительным внешним воздействиям, интенсивному износу и защита от внешней среды, шелковые ткани по изобретению могут быть переработаны для сопротивления артиллерийскому снаряду, огню и химикатам. Процессы для получения таких материалов описаны в WO 2005/045122 и US 2005268443.
ПРИМЕРЫ
ПРИМЕР 1
Получение и анализ кДНК слюнных желез поздней последней возрастной стадии личинки
Белки, которые найдены в шелке настоящих жалящих перепончатокрылых и сетчатокрылых (Apis mellifera, Bombus terrestris, Myrmecia forficata, Oecophylla smaragdina, Mallada signata), были идентифицированы при помощи соответствия картины последовательной фрагментации с помощью масс-спектрометра с ионной ловушкой (МС/МС) пептидов, полученных при гидролизе шелка трипсином, с предсказанными масс-спектральными данными белков, кодируемых кДНК, выделенной из слюнных желез поздней конечной возрастной стадии личинки. Для подтверждения того, что ни один белок не был пропущен в результате этого анализа для медоносной пчелы, данные масс-спектров пептидов также сравнивали с виртуальным расщеплением трипсином белков из Apis mellifera, предсказанных с помощью проекта генома пчелы и трансляции геномных последовательностей Amel3 медоносной пчелы во всех шести рамках считывания.
Медоносная пчела
Личинки Apis mellifera были получены от домашних пчел. Ранее было показано, что производство шелка ограничено только слюнными железами в течение последней половины пятой жизненной стадии личинки (Silva-Zacarin et al., 2003). В течение этого периода РНК более представлена в заднем конце железы (Flower and Kenchington, 1967). Области кубических клеток из 50 слюнных желез были вырезаны из личинки поздней пятой жизненной стадии Apis mellifera, погруженной в забуференный физиологический раствор. Задний конец отделенной железы был немедленно помещен в RNAlater® (Ambion, Austin, TX, USA) для стабилизации мРНК и далее хранился при 4°С.
Тотальную РНК (35 мкг) выделяли из слюнных желез личинки поздней конечной жизненной стадии с применением набора RNAqueous for PCR от Ambion (Austin, TX, USA). Матричную РНК выделяли из тотальной РНК с использованием набора для выделения мРНК Micro-FastTrackTM 2.0 mRNA Isolation kit от Invitrogen (Calsbad, CA, USA) по инструкциям производителя с выделенной мРНК, элюируемой в 10 мкл воды, свободной от РНКаз.
Библиотека кДНК была сконструирована из мРНК, выделенной из личинок Apis mellifera, с использованием набора CloneMinerТМ для создания кДНК-библиотек от Invitrogen (Calsbad, CA, USA) со следующими модификациями стандартного протокола: для синтеза первой цепи к 10 мкл РНК добавили 0,5 мкл праймера Biotin-attB2-Oligo(dT) при концентрации 6 пмоль/мкл и 0,5 мкл смеси всех четырех нуклеотидов dNTP при концентрации 2 мМ для каждого нуклеотида. После инкубации 5 мин при 65°С, затем 2 мин при 45°С добавили 2 мкл 5х буфера для синтеза первой цепи, 1 мкл ДТТ с концентрацией 0,1 М и 0,5 мкл обратной транскриптазы SuperScriptтм II с концентрацией 200 ед./мкл. Для синтеза второй цепи общий объем реагентов был уменьшен наполовину, и после осаждения этанолом кДНК ресуспендировали в 5 мкл воды, обработанной DEPC. Адаптер aatB1 (1 мкл) лигировали в общем объеме 10 мкл с 5 мкл кДНК, с 2 мкл 5х буфера для лигирования адаптеров, 1 мкл ДТТ с концентрацией 0,1 М и 1 мкл ДНК-лигазы Т4 (1 ед./мкл) при 16°С в течение 48 ч с дополнительным 0,5 мкл ДНК-лигазы Т4 (1 ед./мкл), добавленным после 16 ч. кДНК фракционировали по размеру согласно инструкциям производителя с элюирующимися между 300-500 мкл образцами, осажденными этанолом, ресуспендированными и трансформированными в предоставленные устойчивые к фагам клетки E. coli DH10BТМ T1, как рекомендовано. Библиотека кДНК содержала приблизительно 1200000 колониеобразующих единиц (кое) с приблизительно 1% исходного вектора. Средний размер вставки был 1,3±1,4 т.п.о.
Восемьдесят два клона были отобраны случайным образом, и их нуклеотидная последовательность была определена с использованием набора GenomeLabТМ DTCS Quick start kit (BeckmanCoulter, Fullerton CA USA), и разделение продуктов реакции было проведено на секвенаторе CEQ8000 компании Biorad. Эти клоны были собраны в 52 группы (таблица 2). Идентификация кДНК, кодирующих белки шелка, описана ниже.
Другие виды
Тотальная РНК была выделена из губных желез поздней личинки 4 шмелей (Bombus terrestris) (2 мкг РНК), 4 муравьев-бульдогов (Myrmecia forficata) (3 мкг РНК), приблизительно 100 муравьев-портных (Oecophylla smaragdina) (0,4 мкг РНК) и приблизительно 50 зеленых златоглазок (Mallada signata) с использованием набора RNAqueous for PCR от компании Ambion (Austin, TX, USA). мРНК выделяли из тотальной РНК с использованием набора для выделения мРНК Micro-FastTrackTM 2.0 mRNA Isolation kit от Invitrogen (Calsbad, CA, USA) в конечный объем 10 мкл воды. кДНК библиотеки были сконструированы из мРНК с использованием набора CloneMinerTM cDNA от компании Invitrogen (Calsbad, CA, USA) со следующими модификациями стандартного протокола: для синтеза первой цепи к 10 мкл мРНК добавили 3 пмоль праймера Biotin-attB2-Oligo(dT) и 1 нмоль каждого из четырех dNTP. После инкубации 5 мин при 65°С, затем 2 мин при 45°С добавили 2 мкл 5х буфера для синтеза первой цепи, 50 нмоль ДТТ и 100 ед. обратной транскриптазы SuperscriptTM II.
Для синтеза второй цепи общий объем всех реагентов был уменьшен наполовину от количеств, рекомендованных изготовителем, и после переосаждения этанолом кДНК ресуспендировали в 5 мкл воды, обработанной DEPC. Адаптер aatB1 (1 мкл) лигировали в общем объеме 10 мкл с 5 мкл кДНК, с 2 мкл 5х буфера для лигирования адаптеров, 50 нмоль ДТТ и 1 ед. ДНК-лигазы Т4 при 16°С в течение 12 ч. Библиотеки кДНК содержали, приблизительно, 2,4×107 (шмель), 5,0×107 (муравей-бульдог) и 6000 (муравей-портной) колониеобразующих единиц (кое) с менее чем 1% исходного вектора для библиотек из шмеля и муравья-бульдога и более чем 80% исходного вектора для библиотеки из зеленого муравья. Средний размер вставки в библиотеках был 1,3 т.п.о.
Данные по последовательностям были получены более чем для 100 случайных клонов из библиотек кДНК из шмеля и муравья-бульдога, 82 клонов из медоносной пчелы и 60 клонов из златоглазки. Технические сложности получения слюнных желез из мелких муравьев-портных (приблизительно 1 мм в длину) снизило эффективность библиотеки из этого вида, и как таковые были исследованы только 40 последовательностей. Обзор найденных белков шелка представлен в таблице 3.
ПРИМЕР 2
Получение и протеомный анализ природного шелка
Соты с расплодом медоносной пчелы после удаления личинок, коконы шмеля после удаления личинок, коконы муравья-бульдога после удаления личинок или шелковые листки муравья-портного интенсивно промывали три раза теплой водой для удаления водорастворимых примесей и затем интенсивно промывали три раза хлороформом, чтобы удалить воск. Хлороформ удаляли промыванием дистиллированной водой, и набор этого шелка оставляли для анализа. Набор образцов шелка перепончатокрылых (муравьев и пчел) далее промывали при помощи кипячения в течение 30 минут в 0,05% растворе карбоната натрия, стандартной процедуры для удаления смолы из шелка шелковичного червя, затем промывали в дистиллированной воде. Шелк златоглазки промывали только в дистиллированной воде. Из набора образцов шелка златоглазки удаляли смолу посредством кипячения в течение 30 минут в 0,05% растворе карбоната натрия.
Набор материала из медоносной пчелы вымачивали в течение ночи в 2% ДСН при 95°С с последующими тремя промывками в дистиллированной воде. Экстракция горячим ДСН солюбилизирует большинство белков, но в этом случае шелковые листки сохраняли свою конформацию.
Чистый шелк анализировали с помощью жидкостной хроматографии, с последующей тандемной масс-спектрометрией (ЖХ/МС), как описано ниже.
Кусочки очищенного шелка помещали в лунку «застегивающегося» планшета компании Millipore, 96-луночного микротитровочного планшета, содержащего пробку из среды С18 для обращеннофазовой хроматографии внизу каждой лунки, в лунку добавляли 20 мкл 25 мМ бикарбоната аммония, содержащего 160 нг трипсина, пригодного для секвенирования (Promega). Затем планшет инкубировали в течение ночи в увлажняемом пластиковом пакете при 30°С.
Материал С18 увлажняли путем нанесения пипеткой ацетонитрила (10 мкл) на стенки каждой лунки и инкубировали планшет при 37°С в течение 15 минут. К каждой лунке добавляли раствор муравьиной кислоты (130 мкл, 1% по весу), и после 30 минут пептиды из гидролизованных белков пчелы захватывались материалом С18 при медленном прохождении растворов из каждой лунки через основание планшета под пониженным давлением. Материал С18 промывали дважды путем его промывки 100 мкл раствора муравьиной кислоты. Пептиды элюировали 6 мкл 1% муравьиной кислоты в 70% метаноле, наносимыми прямо на материал С18 и быстро удаляемыми центрифугированием через пробку С18 на нижележащую микротитровочную плашку. Эту плашку помещали под вакуум до тех пор, пока объем в каждой лунке не уменьшался в 2 раза благодаря испарению. Добавляли раствор муравьиной кислоты (10 мкл) к каждой лунке, и плашку перемещали к устройству забора образцов из лунок планшета из системы Agilent 1100 для жидкостной капиллярной хроматографии.
Проводили связывание пептидов (8 мкл) из экстракта шелка с колонкой Agilent Zorbax SB-C18 5 мкм 150×0,5 мм со скоростью потока 0,1% муравьиной кислоты/5% ацетонитрила 20 мкл/мин в течение одной минуты, затем элюцию градиентом увеличивающейся концентрации ацетонитрила до 0,1% муравьиной кислоты/20% ацетонитрила за одну минуту при 5 мкл/мин, затем до 0,1% муравьиной кислоты/50% ацетонитрила за 28 минут, затем до 0,1% муравьиной кислоты/95% ацетонитрила за одну минуту. Колонку промывали 0,1% муравьиной кислотой/95%-100% ацетонитрилом за 5 минут при 20 мкл/мин и повторно уравновешивали 0,1% муравьиной кислотой/5% ацетонитрилом в течение 7 минут перед нанесением образца пептидов из следующей лунки.
Элюат с колонки вводили в масс-спектрометр с ионной ловушкой Agilent ХСТ посредством электроспрея (источника ионов инструмента, оборудованного микрофорсункой). Кратко, после элюции пептидов с колонки, ионная ловушка собирала полный спектр положительных ионных прогонов (100-2200 m/z) с последующими двумя МС/МС прогонами ионов, наблюдаемых в полном спектре, избегая отбора ионов, которые несут только единичный заряд. Когда ион был выбран для МС/МС анализа, все другие удалялись из ионной ловушки, проводили фрагментацию выбранного иона согласно установкам, рекомендуемым инструментом, «SmartFrag» и «Peptide Scan». Как только два спектра фрагментации были получены для любой конкретной величины m/z, она исключалась из отбора для анализа на дальнейшие 30 секунд для того, чтобы избежать сбора повторяющихся данных.
Наборы данных масс-спектра всего эксперимента были проанализированы с использованием программного обеспечения Spectrum Mill компании Agilent, чтобы совместить данные с предсказанными белковыми последовательностями из библиотек кДНК. Программа генерировала оценки для определения качества каждого совпадения между экспериментально наблюдаемыми наборами масс фрагментов пептидов и предсказанными фрагментами, которые могли бы образоваться из последовательностей белков в представленной базе данных. Все приведенные в этом описании совпадения последовательностей получили оценки выше 20, установленного по умолчанию значения для автоматического, уверенного принятия достоверных совпадений.
С помощью этого анализа было обнаружено, что пять белков, экспрессируемых на высоком уровне в губных железах, совпадали с шелком из каждого родственного вида пчел (показаны в таблицах 2 и 3) и четыре белка, экспрессируемых на высоком уровне в губных железах, совпадали с шелком из каждого родственного вида муравьев (показаны в таблице 3). Распространенность матричной РНК, кодирующей эти белки в губных железах личинки, согласуется с тем, что белки экспрессируются в большом количестве (распространенность мРНК показана в таблице 3).
Чтобы убедиться, что ни один из белков шелка медоносной пчелы не был пропущен в результате этого процесса идентификации, авторы изобретения также сравнили данные масс-спектра пептидов трипсинизированного шелка медоносной пчелы с набором находящихся в общем доступе предсказанных белковых последовательностей из проекта генома медоносной пчелы, созданных компьютерным алгоритмом, который пытается распознать транскрибируемые гены в последовательностях полной геномной ДНК пчелы. Дополнительно, авторы изобретения создали базу данных трансляции в шести возможных рамках считывания каждой непрерывной геномной ДНК-последовательности, предоставленной проектом генома пчелы (релиз Amel3). Эти транслированные ДНК-последовательности были введены в программу Spectrum Mill в виде последовательностей очень больших белков. Совпадение с МС/МС пептидными данными позволило идентифицировать открытые рамки считывания внутри геномных последовательностей, которые имели кодируемые части выделенных белков пчелы, без необходимости сначала предсказывать генную организацию. Этим методом анализа не было обнаружено никаких дополнительных белков в шелке.
ПРИМЕР 3
Структурный анализ природного шелка
Нативные образцы шелка получали, как было описано в примере 2. Образцы шелка исследовали с использованием инфракрасного фурье-спектрометра Bruker Tensor 37 с комплектующей бриллиантовой пластиной ослабленного общего изображения Pike Miracle. Анализ I и II амидных областей спектров шелков медоносной пчелы, шмеля, муравья-портного, муравья-бульдога и шелка личинки златоглазки (фиг.1) показал, что весь этот шелк имеет в основном альфа-спиральную вторичную структуру. Шелк видов настоящих жалящих перепончатокрылых имеет доминантные пики в ФТ-ИК спектре при 1645-1646 см-1, сдвинутые приблизительно на 10 см-1 ниже, чем классический α-спиральный сигнал, и более широкие. Этот сдвиг в α-спиральном сигнале типичен для белков с двойной спиралью (Heimburg et al., 1999). Спектpы из образцов с удаленной смолой были без изменений.
ПРИМЕР 4
Аминокислотная композиция природного шелка близко напоминает состав идентифицированных белков шелка
Аминокислотная композиция нативного шелка была определена после 24 ч газофазного гидролиза при 110°С с использованием химического анализа Waters AccQTag при помощи Australian Proteome Analysis Facility Ltd (Macquarie University, Sydney).
Определенная аминокислотная композиция шелка, промытого ДСН, была похож на состав, предсказанный для последовательностей идентифицированных белков шелка (фиг.2 и 3).
ПРИМЕР 5
Структурный анализ белков шелка
Предсказанные секреторные пептиды
Как и ожидалось для белков шелка, программа предсказания сигнальных пептидов SignalP 3.0 (Bendtsen et al., 2004), которая использует две модели для идентификации сигнальных пептидов, предсказала, что все идентифицированные гены шелка кодируют белки, которые содержат сигнальные пептиды, направляющие секрецию из клетки (данные не показаны). Предсказанные сайты расщепления являются следующими:
Ксеноспира1 (AmelF1) - между позициями 19 и 20 (ASA-GL),
Ксеноспира2 (AmelF2) - между позициями 19 и 20 (AEG-RV),
Ксеноспира3 (AmelF3) - между позициями 19 и 20 (VHA-GV),
Ксеноспира4 (AmelF4) - между позициями 19 и 20 (ASG-AR),
Ксенозин (AmelSA1) - между позициями 19 и 20 (VCA-GV),
BBF1 - между позициями 19 и 20 (ASA-GQ),
BBF2 - между позициями 20 и 21 (AEG-HV),
BBF3 - между позициями 19 и 20 (VHA-GS),
BBF4 - между позициями 19 и 20 (ASA-GK),
BAF1 - между позициями 19 и 20 (ASA-SG),
BAF2 - между позициями 19 и 20 (ASG-RV),
BAF3 - между позициями 19 и 20 (ASG-NL),
BAF4 - между позициями 19 и 20 (VGA-SE),
GAF1 - между позициями 19 и 20 (ADA-SK),
GAF2 - между позициями 19 и 20 (ASG-GV),
GAF3 - между позициями 19 и 20 (ASG-GV),
GAF4 - между позициями 19 и 20 (VGA-SE),
MalF1 - между позициями 26 и 27 (SST-AV).
Все четыре муравьиных и четыре из пяти пчелиных белков шелка являются спиральными и образуют двойные спирали
Моделирование белков и результаты алгоритмов распознавания структуры подтвердили, что большинство из идентифицированных белков шелка были спиральными белками, которые образуют двойную спираль.
Алгоритмы PROFsec (Rost and Sander, 1993) и NNPredict (McClelland and Rumelhart, 1988; Kneller et al., 1990) использовались для исследования вторичной структуры идентифицированных генов шелка. Эти алгоритмы идентифицировали Ксеноспира1 [GB12184-PA] (SEQ ID NO:1), Ксеноспира2 [GB12348-PA] (SEQ ID NO:3), Ксеноспира3 [GB17818-PA] (SEQ ID NO:5) и Ксеноспира4 [GB19585-PA] (SEQ ID NO:7) как преимущественно спиральные белки с количественным содержанием спиральной структуры между 76-85% (таблица 4). Ксенозин [GB15233-PA] (SEQ ID NO:10) имел существенно более низкое содержание спиральной структуры.
Дальнейшее белковое моделирование и результаты алгоритмов распознавания структуры подтвердили, что большинство идентифицированных белков шелка являются спиральными белками, которые образуют двойные спирали. Алгоритмы PredictProtein (Rost et al., 2004) использовались для исследования вторичной структуры идентифицированных генов шелка. Эти алгоритмы идентифицировали Ксеноспира1 (SEQ ID NO:1), Ксеноспира2 (SEQ ID NO:3), Ксеноспира3 (SEQ ID NO:5), Ксеноспира4 (SEQ ID NO:7), BBF1 (SEQ ID NO:22), BBF2 (SEQ ID NO:24), BBF3 (SEQ ID NO:26), BBF4 (SEQ ID NO:28), BAF1 (SEQ ID NO:40), BAF2 (SEQ ID NO:42), BAF3 (SEQ ID NO:44), BAF4 (SEQ ID NO:46), GAF1 (SEQ ID NO:56), GAF2 (SEQ ID NO:58), GAF3 (SEQ ID NO:60), GAF4 (SEQ ID NO:62) и MalF1 (SEQ ID NO:72) как преимущественно спиральные белки с количественным содержанием спиральной структуры между 69-88% (таблица 3). AmelSA1 [GB15233-PA] (Ксенозин) (SEQ ID NO:10) и BBSA1 (SEQ ID NO:30) имели существенно более низкий процент спиральной структуры.
Суперскручивание спиральных белков (двойные спирали) происходит из характерной последовательности гептадных повторов, обычно обозначаемых как (abcdefg)n, с обычно гидрофобными остатками в положении a и d, и обычно заряженными или полярными остатками в остальных положениях. Программы распознавания структуры (MARCOIL (Delorenzi and Speed, 2002), COILS (Lupas et al., 1991)) идентифицировали множественные гептадные повторы, типичные для двойных спиралей в Ксеноспира1 [GB12184-PA] (SEQ ID NO:1), Ксеноспира2 [GB12348-PA] (SEQ ID NO:3), Ксеноспира3 [GB17818-PA] (SEQ ID NO:5) и Ксеноспира4 [GB19585-PA] (SEQ ID NO:7) (MARCOIL: таблица 5; COILS: фиг.4), также как в BBF1 (SEQ ID NO:22), BBF2 (SEQ ID NO:24), BBF3 (SEQ ID NO:26), BBF4 (SEQ ID NO:28), BAF1 (SEQ ID NO:40), BAF2 (SEQ ID NO:42), BAF3 (SEQ ID NO:44), BAF4 (SEQ ID NO:46), GAF1 (SEQ ID NO:56), GAF2 (SEQ ID NO:58), GAF3 (SEQ ID NO:60), GAF4 (SEQ ID NO:62) и MalF1 (SEQ ID NO:72) (MARCOIL: таблица 3).
Идентификация новой двуспиральной последовательности в белках шелка медоносной пчелы
Гептадные повторы аминокислотных остатков, идентифицированные в последовательностях Ксеноспира1 [GB12184-PA], Ксеноспира2 [GB12348-PA], Ксеноспира3 [GB17818-PA], Ксеноспира4 [GB19585-PA], являлись очень характерными для двуспиральной вторичной структуры (фиг.5) (см. таблицу 5 для доверительных уровней). Тот факт, что гептады были найдены последовательно и в большом количестве позволяет предположить, что белки имеют очень упорядоченную структуру. Перекрывающиеся гептады были найдены в двух белках медоносной пчелы: область главной двойной спирали Ксеноспира1 содержала перекрывающие гептады со смещением в 3 остатка с последующим промежутком в 5 остатков и затем четырьмя последовательными гептадами; и вся двуспиральная область Ксеноспира2 имела множественные перекрывающиеся гептады с единичным смещением и смещением из 4 остатков (равным смещению в 3 остатка). Композиция аминокислот в различных положениях главной гептады показана в первой колонке таблицы 6, где положение перекрывающихся гептад указано в соседних колонках.
Неожиданно оказалось, что при совместном изучении главные гептады имеют новую композицию - с необычно высокой встречаемостью аланина в гидрофобных гептадных положениях a и d (см. таблицу 6 и фиг.5). Дополнительно, большая часть гептад имела аланин в обоих a и d положениях внутри одной гептады (33% в Ксеноспира1 [GB12184-PA]; 36% в Ксеноспира2 [GB12348-PA]; 27% в Ксеноспира3 [GB17818-PA]; и 38% в Ксеноспира4 [GB19585-PA]; см. таблицы 6 и 7).
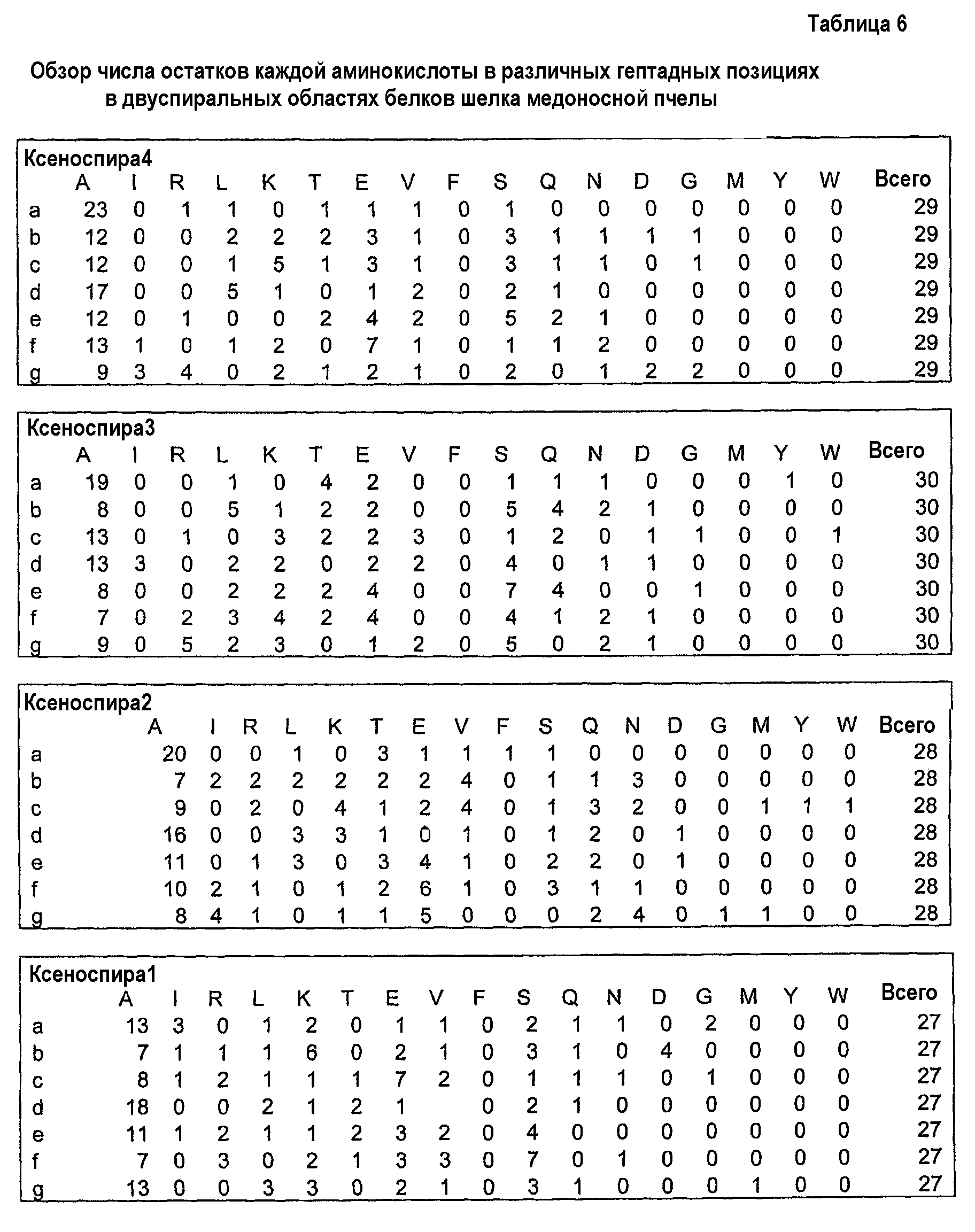
Композиция аминокислот в различных позициях гептады в двуспиральной области шелка перепончатокрылых также кратко изложена на фиг.6 и 7. Как упоминалось выше, положения внутри гептад имели новую композицию - «гидрофобные» гептадные положения a и d из шелка пчелы и муравья содержали очень высокий уровень аланина (средний уровень - 58%) и высокий уровень небольших полярных оснований (средний уровень 21%) по сравнению с другими двойными спиралями. Дополнительно, остатки в положении e были необычно маленьким и гидрофобными (таблица 8, фиг.7). Топографически это положение примыкает к остаткам в положении a внутри спиралей. Его сходство по композиции с остатками в положениях a и d позволяет предположить, что шелк принимает структуру двойной спирали с тремя центральными остатками на α-спираль. Основание из трех остатков предоставляет большую гидрофобную поверхность, чем два остатка в основании (Deng et al., 2006) - свойство, которое помогает образованию и стабильности двойной спирали.
В дополнение, при совместном изучении положения b, c, e, f и g внутри гептады обычно являются более гидрофобными, менее полярными и менее заряженными, чем белковые двуспиральные области, охарактеризованные ранее (см. фиг.7 и таблицы 8 и 9). Поэтому, хотя исторически считалось, что спиральное содержание шелка жалящих перепончатокрылых является следствием пониженного содержания глицина и увеличенного содержания кислых остатков (Rudall and Kenchington, 1971), авторы изобретения обнаружили, что глицин/кислые остатки не отвечают за структуру нового шелка, а скорее положение остатков аланина внутри полипептидных цепей.
ПРИМЕР 6
Белки шелка пчелы, вероятно, имеют высокую степень кросс-сшивки
Белки шелка пчелы все содержат высокое процентное содержание лизина (6,5% - 16,3%). Сравнение между измеренной аминокислотной композицией шелка пчелы и последовательностями идентифицированных белков шелка выявило существенное расхождение в числе остатков лизина, с тем, что было обнаружено больше лизинов в шелке, чем ожидалось (фиг.2 и 3). Это позволяет предположить, что остатки лизина в шелке являются модифицированными таким образом, что они не идентифицируются стандартным аминокислотным анализом. Известно, что лизин образует ряд кросс-сшивок: или ферментативные кросс-сшивки, катализируемые лизил-оксидазой, или неферментативные кросс-сшивки, образуемые гликозилированными остатками лизина (Reiser et al., 1992). Меньшая представительность лизина в аминокислотном анализе шелка медоносной пчелы и шмеля согласуется с наличием лизиновых кросс-сшивок.

Ожидается, что ковалентно кросс-сшитые белки при проведении электрофореза в ДСН-полиакриламидном геле (ПААГ) мигрируют в соответствии с молекулярным весом кросс-сшитого комплекса. Авторы изобретения провели ДСН-ПААГ белков из губных желез поздней последней возрастной стадии личинки медоносной пчелы и измерили миграцию белков шелка по отношению к стандартным белковым маркерам. Наблюдаемые полосы соответствовали мономерам каждого из идентифицированных белков шелка, однако также присутствовали полосы с большим молекулярным весом, содержащие эти белки, как и ожидалось для кросс-сшитых систем (фиг.8).
Как описано выше, губные железы медоносной пчелы содержат смесь организованных и мономерных белков шелка. Наблюдаемые кросс-сшитые белки возможно соответствуют популяции белков передней части железы, где шелк подготавливается для выделения. Логично предположить, что внеклеточный шелк медоносной пчелы содержит существенно большую часть кросс-сшитых белков, чем наблюдается в гетерогенной смеси из всех стадий белков шелка слюнной железы. Маловероятно, что связи являются цистеиновыми кросс-сшивками, так как на шелк не действует восстановительная обработка, и идентифицированные белки шелка не содержат или содержат очень мало цистеиновых остатков.
ПРИМЕР 7
Белки шелка настоящих жалящих перепончатокрылых значительно отличаются от других белков шелка
Шелк настоящих жалящих перепончатокрылых значительно отличается от других описанных генов шелка в отношении аминокислотной композиции (таблица 10), молекулярного веса ее белковых составляющих, вторичной структуры и физических свойств (таблицы 11 и 12). Шелк чешуекрылых в основном состоит из аминокислотных остатков небольшого размера аланина, серина и глицина (например, шелк из Bombyx mori, таблица 10) и в нем превалирует вторичная структура вытянутых бета-листов. Белок шелка из Cotesia glomerata богат аспарагином и серином - распространенность последнего является характерной чертой серицинов шелка чешуекрылых (клеящих веществ) (таблица 10). Моделирование шелка из Cotesia glomerata не выявило спиралей или двойных спиралей во вторичной структуре. В отличие от этого, шелк пчел, муравьев и златоглазок содержит много аланина (таблица 10) и имеет высокий уровень спиральной вторичной структуры, которая образует двойные спирали.
Филогенетический анализ областей двойной спирали белков шелка четырех видов перепончатокрылых (фиг.9) позволяет предположить, что гены возникли из общего предка, который предшествует расхождению Euaculeata от паразитических ос. Последовательности шелка имеют значительные расхождения, и авторы изобретения были в состоянии выровнять только 210 аминокислот, которые содержат область двойной спирали каждого белка. Идентичность по аминокислотам между областями двойных спиралей каждого из белков шелка приведена в этом описании в таблице 13, а идентичность по ДНК соответствующей области показана в таблице 14. Хотя белки и имеют похожий аминокислотный состав (особенно высокое содержание аланина) и третичную структуру, идентичность по первичной аминокислотной последовательности очень низкая. В действительности, ген, кодирующий шелк из Mallada, возник независимо, и как таковая последовательность белка шелка не может быть выровнена с последовательностями перепончатокрылых. Это указывает на то, что значительные вариации могут происходить в составе аминокислот, если они не влияют на биологическую функцию белков.
Из филогенетического анализа можно предсказать, что шелк настоящих жалящих перепончатокрылых ос состоит из белков, родственных шелку муравьев и пчел, и что хотя эти белки будут иметь сходную композицию и архитектуру с белками, описанными в настоящем документе, они будут иметь сильно расходящуюся первичную последовательность.
Проводили сравнение аминокислотных последовательностей белков шелка, представленных в настоящем описании (фиг.10) с белковыми базами данных, однако, не было найдено ни одного прототипного белка с любым значимым уровнем сходства (например, ни одного более чем 30% идентичного по всей длине последовательности белка шелка).
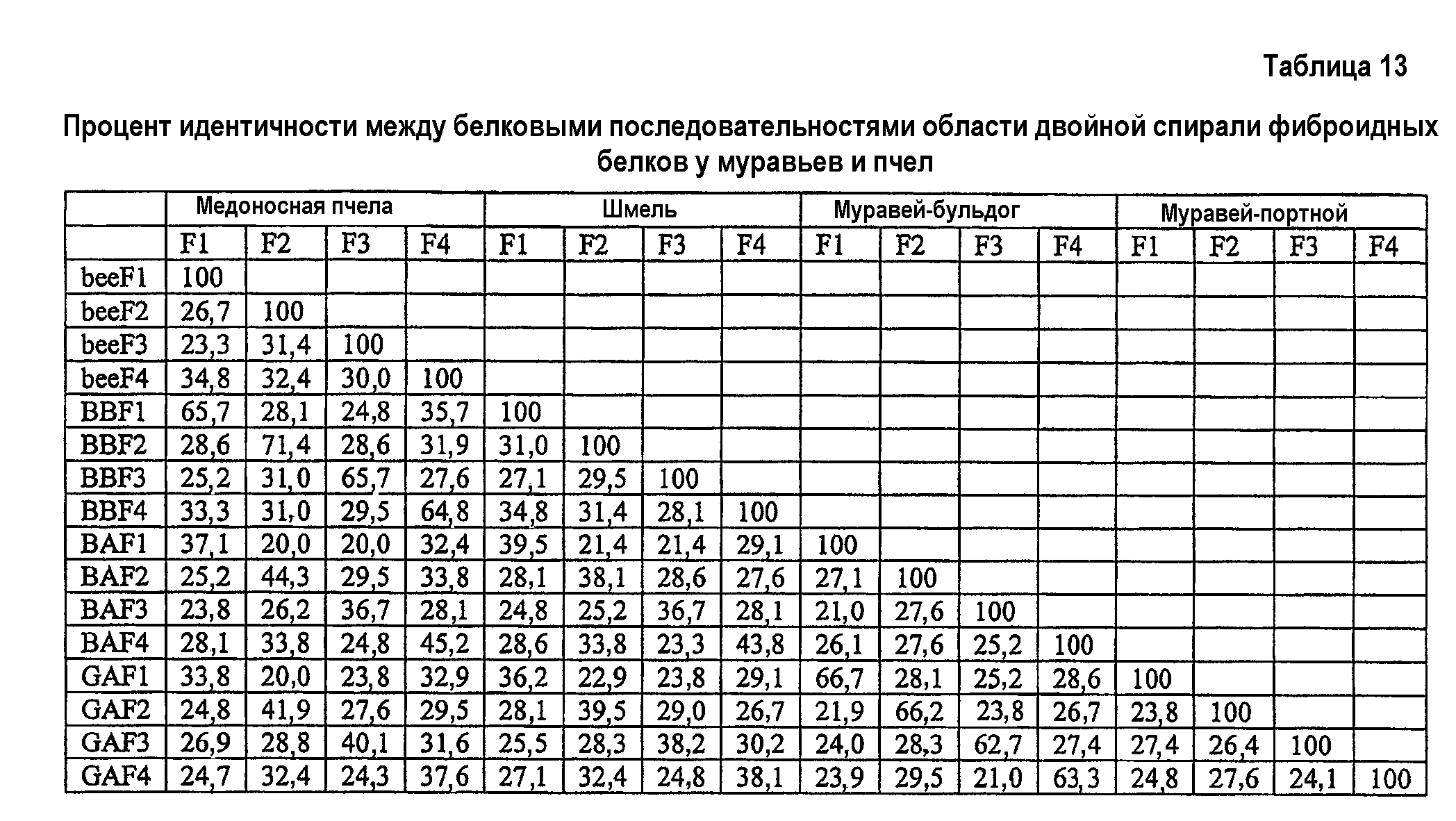
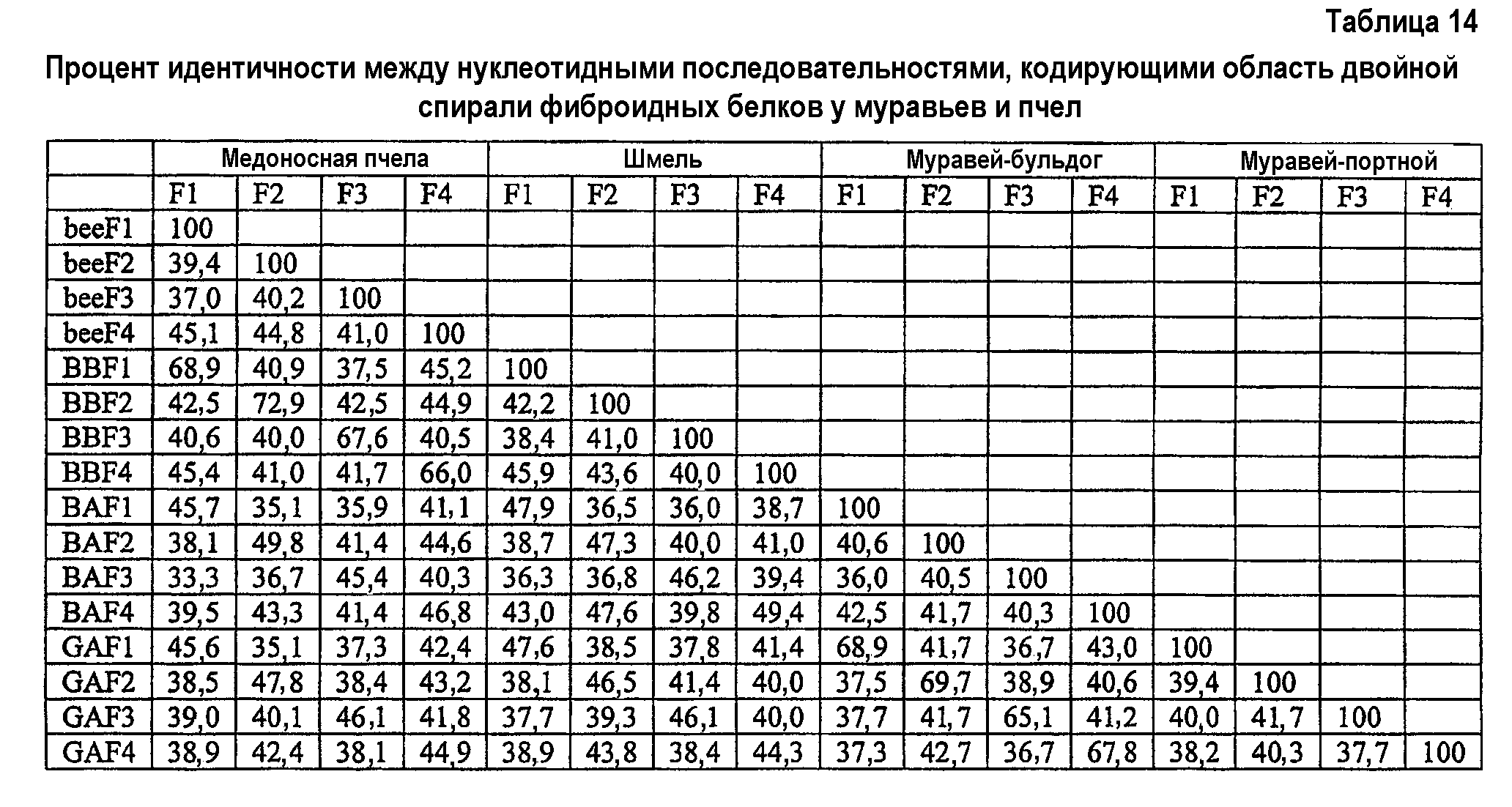
Проводили аналогичный поиск по базам данных для открытых рамок считывания, кодирующих белки шелка (представлены на фиг.11), как было описано выше. Единственные родственные обнаруженные молекулы были опубликованы, как часть геномного проекта медоносной пчелы (www.ncbi.nlm.nih.gov/genome/guide/bee). Открытые рамки считывания были предсказаны в геномном проекте, однако не была предложена функция кодируемых белков. Кроме того, нет свидетельств, что для любой из этих молекул когда-либо был получен полинуклеотид, кодирующий открытую рамку считывания мРНК.
Гены, кодирующие Ксеноспира1, Ксеноспира2, Ксеноспира3 и Ксеноспира4, содержат экзон, покрывающий всю единственную открытую рамку считывания, в то время как ген, кодирующий Ксенозин, содержит по крайней мере один интрон (см. фиг.12).
ПРИМЕР 8
Экспрессия белков шелка в трансгенных растениях
Вектор для экспрессии в растениях, кодирующий белок шелка по изобретению, может состоять из рекомбинантной молекулы нуклеиновой кислоты, кодирующей указанный белок (например, полинуклеотид, приведенный в любой из SEQ ID NO:11-21, 31-39, 48-55, 64-71, 74 или 75), помещенной в 3'-область от промотора CaMV 35S в бинарном векторном каркасе, содержащем ген устойчивости к канамицину (NptII).
Для полинуклеотида, содержащего любую из SEQ ID NO:11, 13, 15, 17, 19, 31, 33, 35, 37, 48, 50, 52, 54, 64, 66, 68, 70 или 74, конструкция далее может содержать область, кодирующую сигнальный пептид, такую как сигнальный пептид вакуолярной основной хитиназы из Arabidopsis thaliana, которую помещают с сохранением рамки считывания в 5'-направлении от последовательности, кодирующей белок шелка.
Отдельно проводят трансформацию конструкции, несущей белок шелка, кодирующий полипептид, в Agrobacterium tumefaciens с помощью электропорации до трансформации в Arabidopsis thaliana. Может применяться гипокотильный способ трансформации, чтобы трансформировать A. thaliana, которая может быть отобрана на выживание в селективной среде, содержащей канамициновую среду. После образования корней на регенератах, их переносят в почву и они образуют первичные трансгенные растения.
Подтверждение процесса трансформации может быть получено посредством ПЦР-скрининга. Включение и экспрессия полинуклеотида могут быть измерены с использованием ПЦР, Саузерн-блот анализа и/или ЖХ/МС расщепленных трипсином экспрессированных белков.
Две или более кодирующих белки шелка конструкций могут быть представлены в одном и том же векторе, или растение можно трансформировать многочисленными различными векторами, каждый из которых кодирует различный белок.
В качестве экспериментального примера экспрессии в растении, кодон-оптимизированную версию AmelF4 (Ксеноспира4) (SEQ ID NO:76) клонировали в pET14b (Novagen), получив pET14b-6xHis:F4op, образующую транслирующийся слитый белок с сохранением рамки считывания с 6хгистидинами на N-конце белка. Последовательность, кодирующая «6xHistidine:F4op», была клонирована в pVEC8 (Wang et al, 1992) под контроль промотора CaMV 35S и регуляторного аппарата полиаденилирования ocs, образуя pVEC8-35S-6xHis:F4op-ocs. Трансформировали pET14b-6xHis:F4op в химически компетентные E. coli, и трансформировали pVEC8-35S-6xHis:F4op в листовые диски табака с помощью опосредуемой Agrobacterium трансформации. Выделяли белки из резистентных к антибиотикам E. coli (индуцируемая экспрессия) и табачных листьев и проводили Вестерн-блот анализ с использованием для детекции антител Tetra-Histidine (Qiagen, Karlsrule, Germany). Пустые векторы pET14b и pVEC8-35S-ocs использовали в качестве отрицательного контроля в соответствующих клетках-хозяевах. Как показано на фиг.13, в результате этих экспериментов было получено растение, экспрессирующее белок Ксеноспира4 (AmelF4).
ПРИМЕР 9
Ферментация и очистка белков шелка
Экспрессионные конструкции были созданы после ПЦР-амплификации областей, кодирующих шелк, из генов медоносной пчелы AmelF1-F4 (Ксеноспира1-4 соответственно) и генов златоглазки MalF1, и клонирования их в экспрессионные векторы pET14b (Novagen, Madison, WI). Затем проводили электропорацию полученных экспрессионных плазмид в клетки E. coli BL21 (DE3) Rosetta и растили в течение ночи на LB-агаре, содержащем ампициллин. Затем использовали одиночную колонию для инокуляции LB-среды, содержащей ампициллин, и растили бактерии в течение ночи при 37°C. Клетки собирали центрифугированием и лизировали детергентом (Bugbuster, Novagen). Тельца включения тщательно промывали и растворяли в 6М гуанидине.
В результате этой процедуры получали белковые смеси с более чем 95% чистотой белков медоносной пчелы и более чем 50% чистотой белка MalF1 златоглазки. Выходы до 50% от сырого веса клеточного осадка E. coli получали регулярно, что указывает на то, что белки легко экспрессируются таким образом.
Солюбилизированные рекомбинантные белки медоносной пчелы наносили на колонку со смолой Talon, приготовленную по инструкциям производителя. Затем элюировали белки с колонки в 100 мМ Tris.HCl, 150 мМ имидазоле, рН 8.
ПРИМЕР 10
Переработка белков шелка в нити
Белки шелка медоносной пчелы и златоглазки легко превратить в нити с использованием ряда способов (см. фиг.14), применяя следующую методику.
Передний сегмент слюнных желез из поздней конечной возрастной стадии личинки Apis mellifera вырезали под фосфатным физиологическим раствором и переносили на плоскую поверхность в капле буфера. Использовали пинцеты для захвата обоих концов сегмента. Один конец поднимали из капли и удаляли от другого с равномерной скоростью. Это давало возможность вытягивать тонкую нить, которая быстро застывала на воздухе.
Рекомбинантные белки шелка личинок медоносной пчелы и златоглазки образовывали нити после дегидратации или концентрирования. Например, при помощи капания растворимого белка в раствор бутанола или путем концентрирования белков на колонке со смолой Talon.
Нити также получали смешиванием рекомбинантных белков шелка медоносной пчелы или златоглазки с органическим растворителем (таким как гексан), чтобы сконцентрировать их в интерфазе с правильной конформацией, и затем добавляя реагент, чтобы исключить их из интерфазы (такой как бутанол). Нити, образуемые в результате этой процедуры, имели сходный ФТ-ИК спектр с нативным шелком, что указывает на то, что они состоят из одинаковой двуспиральной структуры.
Белки шелка из других видов, описанные в настоящем документе, могут также быть получены с помощью этой методики.
Специалистам будет очевидно, что различные вариации и/или модификации могут быть проведены в отношении изобретения, как показано в определенных вариантах осуществления изобретения, не удаляясь от духа или объема изобретения, как приблизительно описано. Настоящие варианты осуществления поэтому должны считаться во всех отношениях как иллюстративные и неограничивающие.
Все обсуждаемые здесь печатные работы целиком включены в это описание.
Все обсуждения документов, актов, материалов, устройств, предметов и т.п., которые были включены в настоящую спецификацию, приведены только для целей предоставления контекста настоящего изобретения. Не должно приниматься предположение, что любой или все из этих предметов образуют часть основы прототипа или известны в области, относящейся к настоящему изобретению, и оно существовало перед приоритетной датой каждого пункта формулы изобретения этого приложения.
Источники информации
Atkins E.D.T. (1967) J. Mol. Biol. 24:139-141.
Bendtsen J.D., Nielsen H., von Heijne G, and Brunak S. (2004) J. Mol. Biol. 340:783-795.
Bini E., Knight D.P. and Kaplan D.L. (2004) J. Mol. Biol. 335:27-40.
Craig C.L. and Riekel C. (2002) Comparative Biochemistry and Physiology Part В 133:493-507.
Delorenzi M. and Speed T. (2002) Bioinformatics 18:617-625.
Deng Y., Liu J., Zheng Q., Eliezer D., Kallenbach N.R. and Lu M. (2006) Structure 14:247-255.
Flower N.E. and Kenchington W.R. (1967) Journal of the Royal Microscopical Society 86:297.
Grimaldi D. and Engel M.S. (2005) Evolution of insects. Cambridge University Press, New York.
Harayama S. (1998) Trends Biotech., 16; 76-82.
Heimburg T, Schunemann J., Weber K., and Geisler N. (1999) Biochemistry 38:12727-12734.
Hepburn H.R., Chandler H. D. and Davidoff M.R. (1979) Insect Biochem. 9:66.
Kneller D.G., Cohen F.E. and Langridge R. (1990) J. Mol. Biol. 214:171-182.
LaMunyon C.W. (1988) Psyche 95:203-209.
LaMunyon C.W. and Adams P.A. (1987) Annals of the Entomological Society of America 80:804-808.
Lucas F. Shaw J.T.B. and Smith S.G. (1960) J. Mol. Biol. 2:339-349.
Lucas F. and Rudall K.M. (1967) In Comprehensive Biochemistry (Ed. Florkin M and Stotz H), Vol.26B pp.475-559 Elsevier Amsterdam.
Lupas A., Van Dyke M. and Stock J. (1991) Science 252:1162-1164.
McClelland J.L. and Rumelhart D.E. (1988) Explorations in Parallel Distributed Processing vol.13, pp.318-362. MITPress, Cambridge MA.
Needleman, S.B. and Wunsch, C.D. (1970) J. Mol. Biol., 48; 443-453.
Quicke D.L.J., Shaw M.R., Takahashi M. and Yanechin B. (2004) Journal of Natural History 38:2167-2181.
Reiser K., McCormick, Rucker R.B. (1992). The FASEB Journal 6:2439-2449.
Rost B. and Sander C. (1993) J. Molecular Biology 232:584-599.
Rost B., Yachdav G. and Liu J. (2004) Nucleic Acids Research 32(Web Server issue):W321-W326.
Rudall K.M. (1962) In Comparative Biochemistry (Ed. By Florkin M and Mason HS) Vol.4, pp.297-435. Academic Press, New York.
Rudall K.M. and Kenchington W. (1971) Annual Reviews in Entomology 16:73-96.
Silva-Zacarin E.C.M., Silva De Moraes R.L.M. and Taboga S.R. (2003) J. Biosci. 6:753-764.
Speilger P.E. (1962) Annals of the Entomological Society of America. 55: 69-77.
Wang M.B., Li Z.Y. et al. (1998). Acta Hort. 461: 401-407.
Yamada H., Shigesada K., Igarashi Y., Takasu Y., Tsubouchi K. and Kato Y. (2004).
Int. J. Wild Silkmoth and Silk 9:61-66.
Реферат
Изобретение относится к области биотехнологии, конкретно к получению белков шелка различных насекомых, и может быть использовано в медицине для создания фармацевтически приемлемого носителя. Получают полипептиды шелка, имеющие двуспиральную структуру и происходящие от медоносной пчелы, шмеля, муравья-бульдога, муравья-портного и златоглазки. Рекомбинантным путем получают клетку-хозяина, трансгенное растение и животное, которые продуцируют полипептид шелка. К полученным полипептидам создают антитела. Изобретение позволяет использовать полученные полипептиды шелка в различных областях промышленности: для создания шелкового волокна, сополимера, продукта - средства личной гигиены, текстиля, пластика и биомедицинского продукта - фармацевтически приемлемого носителя. 14 н. и 7 з.п. ф-лы, 14 ил., 14 табл., 10 пр.
Формула
i) любой аминокислотной последовательности, представленной как SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:22, SEQ ID NO:23, SEQ ID NO:40, SEQ ID NO:41, SEQ ID NO:56 и SEQ ID NO:57;
ii) аминокислотной последовательности, которая по крайней мере на 40% идентична одной или нескольким из SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:22, SEQ ID NO:23, SEQ ID NO:40, SEQ ID NO:41, SEQ ID NO:56 и SEQ ID NO:57;
iii) биологически активного фрагмента i) или ii);
iv) любой аминокислотной последовательности, представленной как SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:42, SEQ.ID NO:43, SEQ ID NO:58 и SEQ ID NO:59;
v) аминокислотной последовательности, которая по крайней мере на 40% идентична одной или нескольким из SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:42, SEQ ID NO:43, SEQ ID NO:58 и SEQ ID NO:59;
vi) биологически активного фрагмента iv) или v);
vii) любой аминокислотной последовательности, представленной как SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:26, SEQ ID NO:27, SEQ ID NO:44, SEQ ID NO:45, SEQ ID NO:60 и SEQ ID NO:61;
viii) аминокислотной последовательности, которая по крайней мере на 40% идентична одной или нескольким из SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:26, SEQ ID NO:27, SEQ ID NO:44, SEQ ID NO:45, SEQ ID NO:60 и SEQ ID NO:61;
ix) биологически активного фрагмента vii) или viii);
x) любой аминокислотной последовательности, представленной как SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:28, SEQ ID NO:29, SEQ ID NO:46, SEQ ID NO:47, SEQ ID NO:62 и SEQ ID NO:63;
xi) аминокислотной последовательности, которая по крайней мере на 40% идентична одной или нескольким из SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:28, SEQ ID NO:29, SEQ ID NO:46, SEQ ID NO:47, SEQ ID NO:62 и SEQ ID NO:63;
xii) биологически активного фрагмента x) или xi);
xiii) аминокислотной последовательности, такой как представленная в SEQ ID NO:72 или в SEQ ID NO:73;
xiv) аминокислотной последовательности, которая по крайней мере на 40% идентична SEQ ID NO:72 и/или SEQ ID NO:73; и xv) биологически активного фрагмента xiii) или xiv).
i) часть полипептида имеет структуру двойной спирали, содержащую по меньшей мере 10 копий гептадной последовательности abcdefq, и где по меньшей мере 25% аминокислот в положении and являются остатками аланина, или
ii) часть полипептида имеет структуру двойной спирали, содержащую по меньшей мере 10 копий гептадной последовательности abcdefq, и где по меньшей мере 25% аминокислот в положении a, d и е являются остатками аланина.
i) любой нуклеотидной последовательности, представленной как SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:48, SEQ ID NO:49, SEQ ID NO:64 и SEQ ID NO:65;
ii) нуклеотидной последовательности, кодирующей полипептид изобретения по любому из пп.1-4;
iii) нуклеотидной последовательности, которая по крайней мере на 40% идентична любой, одной или нескольким, из SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:48, SEQ ID NO:49, SEQ ID NO:64 и SEQ ID NO:65,
iv) нуклеотидной последовательности, которая гибридизируется с любой последовательностью i)-iii) в жестких условиях;
v) любой нуклеотидной последовательности, представленной как SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:50, SEQ ID NO:51, SEQ ID NO:66 и SEQ ID NO:67;
vi) нуклеотидной последовательности, которая по крайней мере на 40% идентична любой, одной или нескольким, из SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:50, SEQ ID NO:51, SEQ ID NO:66 и SEQ ID NO:67;
vii) нуклеотидной последовательности, которая гибридизируется с любой последовательностью v)-vi) в жестких условиях;
viii) любой нуклеотидной последовательности, представленной как SEQ ID NO:15, SEQ ID NO:16, SEQ ID NO:35, SEQ ID NO:36, SEQ ID NO:52, SEQ ID NO:53, SEQ ID NO:68 и SEQ ID NO:69;
ix) нуклеотидной последовательности, которая по крайней мере на 40% идентична любой, одной или нескольким, из SEQ ID NO:15, SEQ ID NO:16, SEQ ID NO:35, SEQ ID NO:36, SEQ ID NO:52, SEQ ID NO:53, SEQ ID NO:68 и SEQ ID NO:69;
x) нуклеотидной последовательности, которая гибридизируется с любой последовательностью viii)-ix) в жестких условиях;
xi) любой нуклеотидной последовательности, представленной как SEQ ID NO:17, SEQ ID NO:18, SEQ ID NO:37, SEQ ID NO:38, SEQ ID NO:54, SEQ ID NO:55, SEQ ID NO:70, SEQ ID NO:71 и SEQ ID NO:76;
xii) нуклеотидной последовательности, которая по крайней мере на 40% идентична любой, одной или нескольким, из SEQ ID NO:17, SEQ ID NO:18, SEQ ID NO:37, SEQ ID NO:38, SEQ ID NO:54, SEQ ID NO:55, SEQ ID NO:70, SEQ ID NO:71 и SEQ ID NO:76;
xiii) нуклеотидной последовательности, которая гибридизируется с любой последовательностью xi)-xii) в жестких условиях;
xiv) нуклеотидной последовательности, представленной как SEQ ID NO:74 или SEQ ID NO:75;
xv) нуклеотидной последовательности, которая по крайней мере на 40% идентична SEQ ID NO:74 и/или SEQ ID NO:75; и
xvi) нуклеотидной последовательности, которая гибридизируется с любой последовательностью xiv)-xv) в жестких условиях.
Комментарии