Человеческие антитела к fel d1 и способы их применения - RU2658491C2
Код документа: RU2658491C2
Чертежи























































































































Описание
Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к человеческим антителам и к антигенсвязывающим фрагментам человеческих антител, которые специфично связываются с кошачьим аллергеном Fel d1, к терапевтическим композициям, содержащим антитела, и способам применения таких антител.
Предшествующий уровень техники настоящего изобретения
Белок Fel d1 представляет собой секретируемый кошачий белок, который принадлежит к семейству секретоглобинов - небольших, связанных дисульфидными мостиками гетеродимерных белков, встречающихся только у млекопитающих (Klug, J. et al. (2000), Ann. N.Y. Acad. Sci. 923:348-354). Он является основной причиной аллергии на кошек у людей (Platts-Mills, Т.А., et al. (1997), J. Allergy Clin. Immunol. 100:S2-S24). Приблизительно у 90-95% больных аллергией на кошек наблюдают реакцию с выработкой IgE на белок Fel d1 (van Ree, et al. (1999), J. Allergy Clin. Immunol. 104:1223-1230). Симптомы у больного, который испытывает аллергическую реакцию на Fel d1, могут варьировать от слабого ринита и конъюнктивита до опасных для жизни астматический реакций. Fel d1 вырабатывается сальными железами, и ороговевающим эпителиальным слоем желез, и клетками плоского эпителия и переносится на шкуру при вылизывании и уходом за шерстью (Bartholome, К. et al. (1985), J. Allergy Clin. Immunol. 76:503-506; Charpin, С. et al. (1991), J. Allergy Clin. Immunol. 88:77-82; Dabrowski, A.J. (1990), et al. J. Allergy Clin. Immunol. 86:462-465). Он также присутствует в слюнных, околоанальных и слезных железах (Andersen, М.С., et al. (1985), J. Allergy Clin. Immunol. 76:563-569; van Milligen, F.J. (1992), et al., Int. Arch. Allergy Appl. Immunol. 92(4):375-378), а основными резервуарами, по видимому, являются шкура и шерсть (Mata, P. et al. (1992), Ann. Allergy 69(4):321-322).
Природный Fel d1 представляет собой гетеродимерный гликопротеин с массой примерно 18 кДа. Каждый гетеродимер содержит две полипептидные цепи, которые ковалентно связаны тремя внутрицепочечными связями и кодируются двумя отдельными генами (Duffort, OA, et al., (1991), Mol. Immunol. 28:301-309; Morgenstern, JP, et al., (1991), PNAS 88:9690-9694; Griffith, I.J., et al. (1992), Gene 113:263-268; Kristensen, A.K. et al. (1997), Biol. Chem. 378:899-908). Цепь 1 содержит 70 аминокислотных остатков, а цепь 2 содержит приблизительно 90-92 аминокислотных остатка. Структурно две цепи похожи, но характеризуются лишь 10-15% идентичности последовательностей (Kaiser, L. et al. (2003) J. Biol. Chem. 278(39):37730-37735). Хотя иногда отдельно каждую цепь называют Fel d1, для полного белка аллергена необходимы обе цепи.
Белок Fel d1 имеет неизвестную функцию у животных, но вызывает реакцию с выработкой IgG или IgE у восприимчивых людей (в виде либо аллергической, либо астматической реакции). Несмотря на то, что известны другие аллергены, в том числе Fel d2 (альбумин) и Fel d3 (цистатин), от 60% до 90% вырабатываемого IgE к кошачьим аллергенам направлено к Fel d1 (Leitermann, K. et al., (1984), J Allergy Clin. Immunol. 74:147-153; Lowenstein, H. et al., (1985), Allergy 40:430-441; van Ree, R. et al., (1999), J. Allergy Clin. Immunol. 104:1223-1230; Ichikawa, K. et al., (2011), Clin. Exp. Allergy, 31:1279-1286).
Иммуноглобулин Ε (IgE) ответственен за гиперчувствительность 1 типа, которая обнаруживается при аллергических ринитах, аллергических конъюнктивитах, сенной лихорадке, аллергической астме, аллергии на пчелиный яд и аллергиях на продукты питания. IgE циркулирует в крови и связывается с высокоаффинными рецепторами FcεR1α к IgE на базофилах и тучных клетках. При большинстве аллергических реакций аллергены попадают в организм посредством вдыхания, приема внутрь или через кожу. Затем аллерген связывается с преформированным IgE, уже связавшимся с высокоаффинным рецептором на поверхностях тучных клеток и базофилов, что в результате приводит к сшиванию нескольких молекул IgE и запуску высвобождения гистамина и других медиаторов воспаления, обуславливая появление различных аллергических симптомов.
Лечение аллергий предусматривает стероиды для подавления иммунной активности и бронхиальные дилататоры для ослабления симптомов астмы. Также для некоторых больных с тяжелой формой аллергии применяют гипосенсибилизирующую терапию. Комбинации пептидных вакцин были протестированы в отношении гипосенсибилизации индивидуумов к конкретным аллергенам, например, Fel d1 (см. US 2010/0239599 A1 и ЕР 2380591 А2). В качестве средств для лечения аллергий были предложены антитела, поскольку они могут блокировать попадание аллергенных молекул в слизистую оболочку ткани или могут связывать аллерген до того, как у него появится возможность связаться с IgE, связавшимся с высокоаффинным рецептором на тучных клетках или базофилах, таким образом предупреждая высвобождение гистамина и других медиаторов высвобождения из этих клеток.
В патенте США №5670626 описано применение моноклональных антител для лечения IgE-опосредованных аллергических заболеваний, таких как аллергический ринит, аллергическая астма и аллергический конъюнктивит, путем блокировки связывания аллергенов со слизистой оболочкой ткани. В патенте США №6849259 описано применение аллерген-специфических антител для подавления аллергического воспаления в in vivo мышиной модели аллергии. Были описаны молочные и яичные системы антител. Например, в US 20030003133 A1 раскрыто применение молока в качестве носителя аллергенов для индукции толерантности при пероральном приеме кошачьей перхоти и других аллергенов. В US 2010/0143266 были описаны композиции и способы снижения аллергической реакции у животных на аллерген в окружающей среде посредством применения молекулы, которая подавляет способность аллергена связываться с тучными клетками. Другие антитела к Fel d1 описаны в работе de Groot et. al. (de Groot et. al., (1988), J. Allergy Clin. Immunol. 82:778-786).
Краткое раскрытие настоящего изобретения
Настоящее изобретение относится к полностью человеческим моноклональным антителам (mAb) и их антигенсвязывающим фрагментам, которые специфично связываются с кошачьим аллергеном Fel d1. Такие антитела могут быть пригодны для связывания аллергена Fel d1 in vivo после того, как сенсибилизированный больной подвергся воздействию кошачьего аллергена, и в связи с этим может содействовать либо ускорению выведения Fel d1, либо блокировке связывания аллергена с преформированным IgE на поверхности тучных клеток или базофилов. Таким образом, антитела согласно настоящему изобретению могут предупреждать высвобождение гистамина или других медиаторов воспаления из тучных клеток или базофилов, таким образом предупреждая или ослабляя нежелательные эффекты, наблюдаемые у больных, сенсибилизированных к кошачьему аллергену. В соответствии с некоторыми вариантами осуществления антитела могут быть способны уменьшать, минимизировать или предупреждать по меньшей мере один симптом у больного, восприимчивого к кошачьему аллергену Fel d1, такой как чихание, гиперемия, заложенность носа, кашель, бронхолегочная обструкция, бронхоконстрикция, ринит или конъюнктивит. В соответствии с некоторыми вариантами осуществления антитела могут быть способны предупреждать еще более серьезные in vivo осложнения, связанные с воздействием на сенсибилизированных индивидуумов кошачьего аллергена, таких как астматические реакции, анафилактическая реакция или даже летальный исход.
Антитела согласно настоящему изобретению могут иметь полную длину (например, антитело IgG1 или IgG4) или могут содержать только антигенсвязывающую часть (например, фрагмент Fab, F(ab')2 или scFv) и могут быть модифицированы с затрагиванием функциональности, например, для устранения остаточных эффекторных функций (Reddy et al., (2000), J. Immunol. 164:1925-1933).
В соответствии с первым аспектом настоящее изобретение относится к выделенному человеческому моноклональному антителу или его антигенсвязывающему фрагменту, которые специфично связываются с Fel d1.
В соответствии с одним вариантом осуществления антитело или его антигенсвязывающий фрагмент относятся к изотипу, отличному от изотипа IgA.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент относятся к изотипу, выбранному из группы, состоящей из IgG1, IgG2 и IgG4.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент специфично связываются с Fel d1 с KD, равной или меньше 10-6 М. В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент специфично связываются с Fel d1 с KD, равной или меньше 1,8 нМ.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат три CDR тяжелой цепи (HCDR1, HCDR2 и HCDR3), содержащиеся в любой из последовательностей вариабельного участка тяжелой цепи (HCVR), выбранной из группы, состоящей из SEQ ID NO: 2, 18, 34, 50, 66, 82, 98, 114, 130, 146, 162, 178, 194, 210, 226, 242, 258, 274, 290, 306, 322, 338, 354, 370 и 460, и три CDR легкой цепи (LCDR1, LCDR2 и LCDR3), содержащиеся в любой из последовательностей вариабельного участка легкой цепи (LCVR), выбранной из группы, состоящей из SEQ ID NO: 10, 26, 42, 58, 74, 90, 106, 122, 138, 154, 170, 186, 202, 218, 234, 250, 266, 282, 298, 314, 330, 346, 362, 378 и 468. Способы и методики определения CDR в аминокислотных последовательностях HCVR и LCVR хорошо известны из уровня техники и могут быть применены для определения CDR в раскрываемых в настоящем описании указанных аминокислотных последовательностях HCVR и/или LCVR. Иллюстративные условные обозначение, которые можно применять для определения границ CDR, включают, например, обозначение по Kabat, обозначение по Chothia и AbM-обозначение. В общих чертах, обозначение по Kabat основано на вариабельности последовательности, обозначение по Chothia основано на расположении структурных петельных участков, и AbM-обозначение является нечто средним между подходами Kabat и Chothia. См., например, Kabat, "Sequences of Proteins of Immunological Interest," National Institutes of Health, Bethesda, Md. (1991); A1-Lazikani et al., (1997), J. Mol Biol 273:927-948; and Martin et al., (1989), Proc. Natl Acad. Sci. USA 56:9268-9272. Находящиеся в общем доступе базы данных также доступны для определения последовательностей CDR в антителе.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат три CDR тяжелой цепи (HCDR1, HCDR2 и HCDR3), содержащиеся в любой из последовательностей вариабельного участка тяжелой цепи (HCVR), выбранной из группы, состоящей из SEQ ID NO: 18, 66, 130, 162, 242, 306, 322, 370 и 460, и три CDR легкой цепи (LCDR1, LCDR2 и LCDR3), содержащиеся в любой из последовательностей вариабельного участка легкой цепи (LCVR), выбранной из группы, состоящей из SEQ ID NO: 26, 74, 138, 170, 250, 314, 330, 378 и 468.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат HCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 2, 18, 34, 50, 66, 82, 98, 114, 130, 146, 162, 178, 194, 210, 226, 242, 258, 274, 290, 306, 322, 338, 354, 370 и 460.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат HCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 18, 66, 130, 162, 242, 306, 322, 370 и 460.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат LCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 10, 26, 42, 58, 74, 90, 106, 122, 138, 154, 170, 186, 202, 218, 234, 250, 266, 282, 298, 314, 330, 346, 362, 378 и 468.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат LCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 26, 74, 138, 170, 250, 314, 330, 378 и 468.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат: (a) HCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 2, 18, 34, 50, 66, 82, 98, 114, 130, 146, 162, 178, 194, 210, 226, 242, 258, 274, 290, 306, 322, 338, 354, 370 и 460, (b) LCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 10, 26, 42, 58, 74, 90, 106, 122, 138, 154, 170, 186, 202, 218, 234, 250, 266, 282, 298, 314, 330, 346, 362, 378 и 468.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат: (a) HCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 18, 66, 130, 162, 242, 306, 322, 370 и 460, и (b) LCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 26, 74, 138, 170, 250, 314, 330, 378 и 468.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат:
(a) домен HCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 4, 20, 36, 52, 68, 84, 100, 116, 132, 148, 164, 180, 196, 212, 228, 244, 260, 276, 292, 308, 324, 340, 356, 372 и 462,
(b) домен HCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 6, 22, 38, 54, 70, 86, 102, 118, 134, 150, 166, 182, 198, 214, 230, 246, 262, 278, 294, 310, 326, 342, 358, 374 и 464,
(c) домен HCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 8, 24, 40, 56, 72, 88, 104, 120, 136, 152, 168, 184, 200, 216, 232,248, 264, 280, 296, 312, 328, 344, 360, 376 и 466,
(d) домен LCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 12, 28, 44, 60, 76, 92, 108, 124, 140, 156, 172, 188, 204, 220, 236, 252, 268, 284, 300, 316, 332, 348, 364, 380 и 470,
(e) домен LCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 14, 30, 46, 62, 78, 94, 110, 126, 142, 158, 174, 190, 206, 222, 238, 254, 270, 286, 302, 318, 334, 350, 366, 382 и 472, и
(f) домен LCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 16, 32, 48, 64, 80, 96, 112, 128, 144, 160, 176, 192, 208, 224, 240, 256, 272, 288, 304, 320, 336, 352, 368, 384 и 474.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат:
(a) домен HCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 20, 68, 132, 164, 244, 308, 324, 372 и 462,
(b) домен HCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 22, 70, 134, 166, 246, 310, 326, 374 и 464,
(c) домен HCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 24, 72, 136, 168, 248, 312, 328, 376 и 466,
(d) домен LCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 28, 76, 140, 172, 252, 316, 332, 380 и 470,
(e) домен LCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 30, 78, 142, 174, 254, 318, 334, 382 и 472, и
(f) домен LCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 32, 80, 144, 176, 256, 320, 336, 384 и 474.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 2/10, 18/26, 34/42, 50/58, 66/74, 82/90, 98/106, 114/122, 130/138, 146/154, 162/170, 178/186, 194/202, 210/218, 226/234, 242/250, 258/266, 274/282, 290/298, 306/314, 322/330, 338/346, 354/362, 370/378 и 460/468.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент содержат пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 18/26, 66/74, 130/138, 162/170, 242/250, 306/314, 322/330, 370/378 и 460/468.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 18/26, 66/74, 130/138 и 162/170.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 18/26 и 322/330.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 18/26 и 306/314.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 18/26 и 370/378.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 242/250 и 306/314.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 242/250 и 322/330.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, взаимодействуют по меньшей мере с одной аминокислотной последовательностью, выбранной из группы, состоящей из аминокислотных остатков, находящихся в диапазоне от приблизительно положения 15 до приблизительно положения 24 в SEQ ID NO: 396, аминокислотных остатков, находящихся в диапазоне от приблизительно положения 85 до приблизительно в положении 103 в SEQ ID NO: 396, аминокислотных остатков, находящихся в диапазоне от приблизительно положения 85 до приблизительно в положении 104 в SEQ ID NO: 396, и аминокислотных остатков, находящихся в диапазоне от приблизительно положения 113 до приблизительно в положении 116 в SEQ ID NO: 396.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют с аминокислотными остатками, находящимися в диапазоне от приблизительно положения 15 до приблизительно в положении 24 в SEQ ID NO: 396.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют с аминокислотными остатками, находящимися в диапазоне от приблизительно положения 85 до приблизительно в положении 103 в SEQ ID NO: 396.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют с аминокислотными остатками, находящимися в диапазоне от приблизительно положения 85 до приблизительно в положении 104 в SEQ ID NO: 396.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют с аминокислотными остатками, находящимися в диапазоне от приблизительно положения 113 до приблизительно в положении 116 в SEQ ID NO: 396.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют по меньшей мере с одной аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 402, 403, 404 и 412.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют с SEQ ID NO: 402.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют с SEQ ID NO: 403.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют с SEQ ID NO: 404.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют с SEQ ID NO: 426.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, взаимодействуют с SEQ ID NO: 412.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые взаимодействуют с SEQ ID NO: 402, 403, 404 и/или 426, содержат три HCDR, содержащиеся в вариабельном участке тяжелой цепи, с SEQ ID NO: 18 и три LCDR, содержащиеся в вариабельном участке легкой цепи, с SEQ ID NO: 26.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые взаимодействуют с SEQ ID NO: 402, 403, 404 и/или 426, содержат HCDR1 с SEQ ID NO: 20, HCDR2 с SEQ ID NO: 22, HCDR3 с SEQ ID NO: 24, LCDR1 с SEQ ID NO: 28, LCDR2 с SEQ ID NO: 30 и LCDR3 с SEQ ID NO: 32.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые взаимодействуют с SEQ ID NO: 412, содержат свободные HCDR, содержащиеся в вариабельном участке тяжелой цепи, с SEQ ID NO: 306 и три LCDR, содержащиеся в вариабельном участке легкой цепи, с SEQ ID NO: 314.
В соответствии с одним вариантом осуществления выделенное человеческое антитело или его антигенсвязывающий фрагмент, которые взаимодействуют с SEQ ID NO: 412, содержат HCDR1 с SEQ ID NO: 308, HCDR2 с SEQ ID NO: 310, HCDR3 с SEQ ID NO: 312, LCDR1 с SEQ ID NO: 316, LCDR2 с SEQ ID NO: 318 и LCDR3 с SEQ ID NO: 320.
В соответствии с одним вариантом осуществления человеческое антитело или его антигенсвязывающий фрагмент, которые связывают Fel d1, содержат аминокислотные последовательности HCDR1, HCDR2 и HCDR3 с SEQ ID NO: 20, 22 и 24, соответственно, и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 с SEQ ID NO: 28, 30 и 32, соответственно.
В соответствии с одним вариантом осуществления человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат аминокислотные последовательности HCDR1, HCDR2 и HCDR3 с SEQ ID NO: 68, 70 и 72, соответственно, и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 с SEQ ID NO: 76, 78 и 80, соответственно.
В соответствии с одним вариантом осуществления человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат аминокислотные последовательности HCDR1, HCDR2 и HCDR3 с SEQ ID NO: 132, 134 и 136, соответственно, и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 с SEQ ID NO: 140, 142 и 144, соответственно.
В соответствии с одним вариантом осуществления человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат аминокислотные последовательности HCDR1, HCDR2 и HCDR3 с SEQ ID NO: 164, 166 и 168, соответственно, и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 с SEQ ID NO: 172, 174 и 176, соответственно.
В соответствии с одним вариантом осуществления человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат аминокислотные последовательности HCDR1, HCDR2 и HCDR3 с SEQ ID NO: 244, 246 и 248, соответственно, и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 с SEQ ID NO: 252, 254 и 256, соответственно.
В соответствии с одним вариантом осуществления человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат аминокислотные последовательности HCDR1, HCDR2 и HCDR3 с SEQ ID NO: 308, 310 и 312, соответственно, и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 с SEQ ID NO: 316, 318 и 320, соответственно.
В соответствии с одним вариантом осуществления человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат аминокислотные последовательности HCDR1, HCDR2 и HCDR3 с SEQ ID NO: 324, 326 и 328, соответственно, и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 с SEQ ID NO: 332, 334 и 336, соответственно.
В соответствии с одним вариантом осуществления человеческое антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат аминокислотные последовательности HCDR1, HCDR2 и HCDR3 с SEQ ID NO: 372, 374 и 376, соответственно, и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 с SEQ ID NO: 380, 382 и 384, соответственно.
Согласно одному варианту осуществления настоящее изобретение относится к полностью человеческому моноклональному антителу или его антигенсвязывающий фрагменту, которые связываются с Fel d1, причем у антитела или его фрагмента наблюдают одну или несколько из следующих характеристик: (i) содержит HCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 18, 66, 130, 162, 242, 306, 322, 370 и 460, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (ii) содержит LCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 26, 74, 138, 170, 250, 314, 330, 378 и 468, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (iii) содержит домен HCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 24, 72, 136, 168, 248, 312, 328, 376 и 466, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен HCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 32, 80, 144, 176, 256, 320, 336, 384 и 474, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (iv) содержит домен HCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 20, 68, 132, 164, 244, 308, 324, 372 и 462, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; домен HCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 22, 70, 134, 166, 246, 310, 326, 374 и 464, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; домен LCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 28, 76, 140, 172, 252, 316, 332, 380 и 470, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен LCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 30, 78, 142, 174, 254, 318, 334, 382 и 472, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (v) связывается с Fel d1 с KD, равной или меньше 10-6 и предпочтительно равной или меньше 10-9; (vi) характеризуется эффективностью по меньшей мере в одной животной модели анафилактической реакции или воспаления; или (vii) конкурирует с эталонным антителом за связывание с Fel d1.
В соответствии с одним вариантом осуществления "эталонное антитело" может включать, например, антитела, имеющие комбинацию пар аминокислотных последовательностей тяжелой цепи и легкой цепи, выбранную из группы, состоящей из 18/26, 66/74, 130/138, 162/170, 242/250, 306/314, 322/330, 370/378 и 460/468.
В соответствии с одним вариантом осуществления полностью человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые связываются с Fel d1, содержат последовательность HCDR1, характеризующуюся формулой X1-X2-X3-X4-X5-X6-X7-X8 (SEQ ID NO: 386), где X1 представляет собой Gly, X2 представляет собой Phe, Tyr или Gly, X3 представляет собой Thr или Ser, X4 представляет собой Phe или Ile, X5 представляет собой Ser, Arg, Thr или Asn, X6 представляет собой Asn, Thr, Asp или Ser, X7 представляет собой Tyr и X8 представляет собой Asn, Tyr или Ala; последовательность HCDR2, характеризующуюся формулой X1-X2-X3-X4-X5-X6-X7-X8 (SEQ ID NO: 387), где X1 представляет собой Ile, X2 представляет собой Tyr, Ser или Asn, X3 представляет собой Tyr, Ser, Gly, Pro или Asp, X4 представляет собой Asp, Arg или Ser, X5 представляет собой Gly, Val или Ser, X6 представляет собой Ser, Gly, Arg или Tyr, X7 представляет собой Tyr, Arg, Thr, Ser или Asn, и X8 представляет собой Ile, Thr, Ala, Ser или отсутствует; последовательность HCDR3, характеризующуюся формулой X1-X2-X3-X4-X5-Χ6-X7-X8-X9-X10-X11-X12-X13-X14-X15-X16 (SEQ ID NO: 388), где X1 представляет собой Ala, X2 представляет собой Lys или Arg, X3 представляет собой Arg, Gly, His, Ser, Asp, Leu или Thr, X4 представляет собой Thr, Pro, Arg, Gly или Glu, X5 представляет собой Leu, Val, Gly, Lys, Tyr или Asn, X6 представляет собой Ser, Arg, Thr, Ala, Tyr, Phe или Trp, X7 представляет собой Tyr, Gly, Arg, Ala, Asn, Asp, His или Asn, X8 представляет собой Tyr, Thr, Ser или His, X9 представляет собой Val, Ser, Ala, Phe, Pro или отсутствует, X10 представляет собой Met, Gly, Asp, Pro, Val, или отсутствует, Χ11 представляет собой Asp, Tyr, Ser, Gly, Phe, или отсутствует, X12 представляет собой Val, Asp, Phe или отсутствует, X13 представляет собой Phe, Asp или отсутствует, X14 представляет собой Phe, Tyr или отсутствует, X15 представляет собой Asp или отсутствует, X16 представляет собой Tyr или отсутствует; последовательность LCDR1, характеризующуюся формулой Х1-X2-X3-X4-X5-X6-X7-X8-X9-X10-X11-X12 (SEQ ID NO: 389), где X1 представляет собой Gln, X2 представляет собой Gly, Ser или Asp, X3 представляет собой Ile или Val, X4 представляет собой Ser, Leu, Asn или Gly, Χ5 представляет собой Asn, Tyr, Gly или Ser, Χ6 представляет собой Tyr, Ser, Phe или Trp, Χ7 представляет собой Ser или отсутствует, X8 представляет собой Asn или отсутствует, X9 представляет собой Asn или отсутствует, X10 представляет собой Lys или отсутствует, X11 представляет собой Gln или отсутствует, X12 представляет собой Tyr или отсутствует; последовательность LCDR2, характеризующуюся формулой X1-X2-X3 (SEQ ID NO: 390), где X1 представляет собой Ala, Trp, Asp, Tyr, Lys, Gly или Ser, X2 представляет собой Ala или Thr, и X3 представляет собой Ser; и последовательность LCDR3, характеризующуюся формулой X1-X2-X3-X4-X5-Χ6-X7-X8-X9 (SEQ ID NO: 391), где X1 представляет собой Gln, Leu или His, X2 представляет собой Lys, Gin или His, Χ3 представляет собой Tyr, Ser или Leu, X4 представляет собой Tyr, Asn, Gly, Asp или Ser, Χ5 представляет собой Ser, Asp или Asn, X6 представляет собой Leu, Ala, Tyr, Thr или Phe, Χ7 представляет собой Pro или Arg, X8 представляет собой Leu, Phe, Tyr или Thr, и Χ9 представляет собой Thr или отсутствует.
В соответствии с одним вариантом осуществления настоящее изобретение относится к человеческому антителу или антигенсвязывающему фрагменту, специфичному к Fel d1, содержащему HCVR, кодируемый сегментами нуклеотидной последовательности, полученными из последовательностей зародышевой линии VH, DH и JH, и LCVR, кодируемый сегментами нуклеотидной последовательности, полученными из последовательностей зародышевой линии VK и JK, с комбинациями, которые описаны в таблице 2.
Настоящее изобретение относится к антителам с модифицированными профилями гликозилирования. В некоторых областях применения может быть пригодна модификация, удаляющая нежелательные сайты гликозилирования, или, например, удаление фукозного фрагмента для повышения функции антителозависимой клеточноопосредованной цитотоксичности (ADCC) (см. Shield et al. (2002) JBC 277:26733). В других областях применения для модификации комплементзависимой цитотоксичности (CDC) можно выполнить модификацию галактозилирования.
Согласно второму аспекту настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, которые конкурируют за специфичное связывание с Fel d1 с антителом или антигенсвязывающим фрагментом, содержащим определяющие комплементарность участки (CDR) вариабельного участка тяжелой цепи (HCVR), причем HCVR имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 18, 34, 50, 66, 82, 98, 114, 130, 146, 162, 178, 194, 210, 226, 242, 258, 274, 290, 306, 322, 338, 354, 370 и 460; и CDR вариабельного участка легкой цепи (LCVR), причем LCVR имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 10, 26, 42, 58, 74, 90, 106, 122, 138, 154, 170, 186, 202, 218, 234, 250, 266, 282, 298, 314, 330, 346, 362, 378 и 468.
Согласно одному варианту осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, которые конкурируют за специфичное связывание с Fel d1 с антителом или антигенсвязывающим фрагментом, содержащим определяющие комплементарность участки (CDR) вариабельного участка тяжелой цепи (HCVR), причем HCVR имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 18, 66, 130, 162, 242, 306, 322, 370 и 460; и CDR вариабельного участка легкой цепи (LCVR), причем LCVR имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID: 26, 74, 138, 170, 250, 314, 330, 378 и 468.
Согласно соответствующему варианту осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающий фрагменту, которые конкурируют за специфичное связывание с Fel d1 с антителом или антигенсвязывающим фрагментом, содержащим CDR тяжелой и легкой цепи, которые содержатся в парах последовательностей тяжелой и легкой цепей, выбранных из группы, состоящей из SEQ ID NO: 18/26, 66/74, 130/138, 162/170, 242/250, 306/314, 322/330, 370/378 и 460/468.
Согласно третьему аспекту настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, которые связывают тот же эпитоп на Fel d1, что и антитело или антигенсвязывающий фрагмент, содержащие определяющие комплементарность участки (CDR) вариабельного участка тяжелой цепи (HCVR), причем HCVR имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 18, 34, 50, 66, 82, 98, 114, 130, 146, 162, 178, 194, 210, 226, 242, 258, 274, 290, 306, 322, 338, 354, 370 и 460; и CDR вариабельного участка легкой цепи (LCVR), причем LCVR имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 10, 26, 42, 58, 74, 90, 106, 122, 138, 154, 170, 186, 202, 218, 234, 250, 266, 282, 298, 314, 330, 346, 362, 378 и 468.
Согласно одному варианту осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, который связывает тот же эпитоп на Fel d1, что и антитело или антигенсвязывающий фрагмент, содержащие определяющие комплементарность участки (CDR) вариабельного участка тяжелой цепи (HCVR), причем HCVR имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 18, 66, 130, 162, 242, 306, 322, 370 и 460; и CDR вариабельного участка легкой цепи (LCVR), причем LCVR имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID: 10, 26, 42, 58, 74, 90, 106, 122, 138, 154, 170, 186, 202, 218, 234, 250, 266, 282, 298, 314, 330, 346, 362, 378 и 468.
Согласно соответствующему варианту осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающий фрагменту, которые связывают тот же эпитоп на Fel d1, что и антитело или антигенсвязывающий фрагмент, содержащие CDR тяжелой и легкой цепи, которые содержатся в парах последовательностей тяжелой и легкой цепей, выбранных из группы, состоящей из SEQ ID NO: 18/26, 66/74, 130/138, 162/170, 242/250, 306/314, 322/330, 370/378 и 460/468.
Согласно четвертому аспекту настоящее изобретение относится к биспецифической антигенсвязывающей молекуле, которая специфично связывает Fel d1 и содержит два антигенсвязывающих домена (два плеча), содержащих аминокислотную последовательность HCVR и аминокислотную последовательность LCVR из любых двух или более описанных в настоящем документе антител.
В соответствии с одним вариантом осуществления биспецифическая антигенсвязывающая молекула содержит первый антигенсвязывающий домен, который содержит аминокислотную последовательность HCVR, приведенную в SEQ ID NO: 370, и аминокислотную последовательность LCVR, приведенную в SEQ ID NO: 378, и второй антигенсвязывающий домен, который содержит аминокислотную последовательность HCVR, приведенную в SEQ ID NO: 18, и аминокислотную последовательность LCVR, приведенную в SEQ ID NO: 378.
В соответствии с одним вариантом осуществления биспецифическая антигенсвязывающая молекула содержит первый антигенсвязывающий домен, который содержит три определяющих комплементарность участка тяжелой цепи (HCDR1, HCDR2 и HCDR3), состоящих из аминокислотных последовательностей, приведенных в SEQ ID NO: 372, 374 и 376, соответственно, и три определяющих комплементарность участка легкой цепи (LCDR1, LCDR2 и LCDR3), состоящих из аминокислотных последовательностей, приведенных в SEQ ID NO: 380, 382 и 384, соответственно; а второй антигенсвязывающий домен содержит три определяющих комплементарность участка тяжелой цепи (HCDR1, HCDR2 и HCDR3), состоящих из аминокислотных последовательностей, приведенных в SEQ ID NO: 20, 22 и 24, соответственно, и три определяющих комплементарность участка легкой цепи (LCDR1, LCDR2 и LCDR3), состоящих из аминокислотных последовательностей, приведенных в SEQ ID NO: 380, 382 и 384, соответственно.
В соответствии с одним вариантом осуществления биспецифическая антигенсвязывающая молекула содержит первый антигенсвязывающий домен, который содержит аминокислотную последовательность HCVR, приведенную в SEQ ID NO: 306, и аминокислотную последовательность LCVR, приведенную в SEQ ID NO: 314, и второй антигенсвязывающий домен, который содержит аминокислотную последовательность HCVR, приведенную в SEQ ID NO: 18, и аминокислотную последовательность LCVR, приведенную в SEQ ID NO: 314.
В соответствии с одним вариантом осуществления биспецифическая антигенсвязывающая молекула содержит три определяющих комплементарность участка тяжелой цепи (HCDR1, HCDR2 и HCDR3), состоящих из аминокислотных последовательностей, приведенных в SEQ ID NO: 308, 310 и 312, соответственно, и три определяющих комплементарность участка легкой цепи (LCDR1, LCDR2 и LCDR3), состоящих из аминокислотных последовательностей, приведенных в SEQ ID NO: 316, 318 и 320, соответственно; а второй антигенсвязывающий домен содержит три определяющих комплементарность участка тяжелой цепи (HCDR1, HCDR2 и HCDR3), состоящих из аминокислотных последовательностей, приведенных в SEQ ID NO: 20, 22 и 24, соответственно, и три определяющих комплементарность участка легкой цепи (LCDR1, LCDR2 и LCDR3), состоящих из аминокислотных последовательностей, приведенных в SEQ ID NO: 316, 318 и 320, соответственно.
В соответствии с одним вариантом осуществления настоящее изобретение относится к выделенному антителу, специфичному к Fel d1, или его антигенсвязывающему фрагменту, которые конкурируют за связывание с Fel d1 с любой из биспецифических антигенсвязывающих молекул по настоящему изобретению.
В соответствии с одним вариантом осуществления настоящее изобретение относится к выделенному антителу, специфичному к Fel d1, или его антигенсвязывающему фрагменту, которые связываются с тем же эпитопом на Fel d1, что и любая из биспецифических антигенсвязывающих молекул по настоящему изобретению.
В соответствии с одним вариантом осуществления биспецифическая антигенсвязывающая молекула представляет собой выделенное человеческое моноклональное антитело, которое специфично связывается с Fel d1.
В соответствии с одним вариантом осуществления биспецифическая антигенсвязывающая молекула представляет собой выделенное человеческое моноклональное антитело, которое специфично связывается с Fel d1, причем человеческое моноклональное антитело является моноспецифическим антителом или биспецифическим антителом.
В соответствии с одним вариантом осуществления настоящее изобретение относится к фармацевтической композиции, содержащей по меньшей мере одну описанную в настоящем документе биспецифическую антигенсвязывающую молекулу и фармацевтически приемлемый носитель или разбавитель.
В соответствии с одним вариантом осуществления настоящее изобретение относится к способу лечения больного, у которого наблюдают чувствительность или аллергическую реакцию на кошку, кошачью перхоть, кошачью шерсть или их экстракт или на белок Fel d1, или лечения по меньшей мере одного симптома или осложнения, ассоциированного с чувствительностью или аллергической реакцией на кошку, кошачью перхоть, кошачью шерсть или их экстракт или на белок Fel d1, предусматривающему введение эффективного количества одной или нескольких биспецифических антигенсвязывающих молекул по настоящему изобретению или фармацевтической композиции, содержащей эффективное количество одной или нескольких биспецифических антигенсвязывающих молекул по настоящему изобретению, нуждающемуся в этом больному, причем у больного наблюдают уменьшенную чувствительность или сниженную аллергическую реакцию на кошку, кошачью перхоть, кошачью шерсть или их экстракт или на белок Fel d1, или он не испытывает какой-либо чувствительности или аллергической реакции на кошку, кошачью перхоть, кошачью шерсть или их экстракт или на белок Fel d1, или у больного наблюдают уменьшение по меньшей мере одного симптома или осложнения, ассоциированного с чувствительностью или аллергической реакцией на кошку, кошачью перхоть, кошачью шерсть или их экстракт или на белок Fel d1, или уменьшение частоты и/или продолжительности по меньшей мере одного симптома или осложнения, ассоциированного с чувствительностью или аллергической реакцией на кошку, кошачью перхоть, кошачью шерсть или их экстракт или на белок Fel d1, после введения биспецифических антигенсвязывающих молекул или композиции, содержащей биспецифические антигенсвязывающие молекулы по настоящему изобретению.
В соответствии с одним вариантом осуществления настоящее изобретение относится к введению эффективного количества второго терапевтического средства наряду по меньшей с мере одной биспецифической антигенсвязывающей молекулой по настоящему изобретению, пригодной для ослабления аллергической реакции на кошку, кошачью перхоть или на белок Fel d1. Второе терапевтическое средство можно выбрать из группы, состоящей из кортикостероида, бронхиального дилататора, антигистамина, эпинефрина, противоотечного средства, кортикостероида, другого отличного антитела к Fel d1 и пептидной вакцины.
В соответствии с одним вариантом осуществления лечение при помощи одной или нескольких биспецифических антигенсвязывающих молекул по настоящему изобретению отдельно или в комбинации со вторым терапевтическим средством может привести в результате к уменьшению аллергического ринита, аллергического конъюнктивита, аллергической астмы или анафилактической реакции после контакта больного с кошкой, кошачьей перхотью или белком Fel d1.
В соответствии с пятым аспектом настоящее изобретение относится к молекулам нуклеиновой кислоты, кодирующим антитела к Fel d1 или их фрагменты. Также настоящее изобретение относится к рекомбинантным векторам экспрессии, несущим нуклеиновые кислоты по настоящему изобретению, и клеткам-хозяевам, в которые были введены такие векторы, в качестве способов получения антител посредством культивирования клеток-хозяев в условиях, позволяющих происходить продукции антител, и выделения полученных антител.
Согласно одному варианту осуществления настоящее изобретение относится к антителу или его фрагменту, содержащему HCVR, кодируемый последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 1, 17, 33, 49, 65, 81, 97, 113, 129, 145, 161, 177, 193, 209, 225, 241, 257, 273, 289, 305, 321, 337, 353, 369 и 459, или практически идентичной последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% гомологию с нею.
В соответствии с одним вариантом осуществления HCVR кодируется последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 17, 65, 129, 161, 241, 305, 321, 369 и 459.
В соответствии с одним вариантом осуществления антитело или его фрагмент дополнительно содержит LCVR, кодируемый последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 9, 25, 41, 57, 73, 89, 105, 121, 137, 153, 169, 185, 201, 217, 233, 249, 265, 281, 297, 313, 329, 345, 361, 377 и 467, или практически идентичной последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% гомологию с нею.
Согласно одному варианту осуществления LCVR кодируется последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO: 25, 73, 137, 169, 249, 313, 329, 377 и 467.
В соответствии с одним вариантом осуществления настоящее изобретение также относится к антителу или антигенсвязывающему фрагменту антитела, содержащему домен HCDR3, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 7, 23, 39, 55, 71, 87, 103, 119, 135, 151, 167, 183, 199, 215, 231, 247, 263, 279, 295, 311, 327, 343, 359, 375 и 465, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен LCDR3, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 15, 31, 47, 63, 79, 95, 111, 127, 143, 159, 175, 191, 207, 223, 239, 255, 271, 287, 303, 319, 335, 351, 367, 383 или 473, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности.
В соответствии с одним вариантом осуществления настоящее изобретение относится к антителу или его фрагменту, дополнительно содержащему домен HCDR1, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 3, 19, 35, 51, 67, 83, 99, 115, 131, 147, 163, 179, 195, 211, 227, 243, 259, 275, 291, 307, 323, 339, 355, 371 и 461, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен HCDR2, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 5, 21, 37, 53, 69, 85, 101, 117, 133, 149, 165, 181, 197, 213, 229, 245, 261, 277, 293, 309, 325, 341, 357, 373 и 463, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен LCDR1, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 11, 27, 43, 59, 75, 91, 107, 123, 139, 155, 171, 187, 203, 219, 235, 251, 267, 283, 299, 315, 331, 347, 363, 379 и 469, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен LCDR2, кодируемый нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 13, 29, 45, 61, 77, 93, 109, 125, 141, 157, 173, 189, 205, 221, 237, 253, 269, 285, 301, 317, 333, 349, 365, 381 или 471, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности.
В соответствии с шестым аспектом настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество одного или нескольких выделенных человеческих антител или их антигенсвязывающих фрагментов, которые специфично связывают Fel d1, вместе с одним или несколькими фармацевтически приемлемыми наполнителями.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит терапевтически эффективное количество двух или более выделенных человеческих антител или их антигенсвязывающих фрагментов, которые специфично связывают Fel d1, вместе с одним или несколькими фармацевтически приемлемыми наполнителями.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
a) выделенное первое полностью человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связывают Fel d1, содержащие HCVR с аминокислотной последовательностью, которая приведена в SEQ ID NO: 18, и LCVR с аминокислотной последовательностью, которая приведена в SEQ ID NO: 26; и
b) выделенное второе полностью человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связывают Fel d1, содержащие HCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 66, 130, 162, 306, 322, 370 и 460, и LCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 74, 138, 170, 314, 330, 378 и 468.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
a) выделенное первое полностью человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связывают Fel d1, содержащие HCVR с аминокислотной последовательностью, которая приведена в SEQ ID NO: 242, и LCVR с аминокислотной последовательностью, которая приведена в SEQ ID NO: 250; и
b) выделенное второе полностью человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связывают Fel d1, содержащие HCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 306, 322 и 460; и LCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 314, 330 и 468.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
a) выделенное первое полностью человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связывают Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 18/26; и
b) выделенное второе полностью человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связывают Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 66/74, 130/138, 162/170, 306/314, 322/330, 370/378 и 460/468.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
a) выделенное первое человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 18/26; и
b) выделенное второе человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 130/138.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
a) выделенное первое человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 18/26; и
b) выделенное второе человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 322/330.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
а) выделенное первое человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 18/26; и
b) выделенное второе человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 306/314.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
a) выделенное первое человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 18/26; и
b) выделенное второе человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 370/378.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
a) выделенное первое полностью человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связывают Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 242/250; и
b) выделенное второе полностью человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связывают Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, выбранную из группы, состоящей из SEQ ID NO: 306/314 и 322/330.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
a) выделенное первое человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 242/250; и
b) выделенное второе человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 306/314.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит:
a) выделенное первое человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 242/250; и
b) выделенное второе человеческое моноклональное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с Fel d1, содержащие пару аминокислотных последовательностей HCVR/LCVR, состоящую из SEQ ID NO: 322/330.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит два или более выделенных человеческих моноклональных антител, которые специфично связываются с Fel d1, или их антигенсвязывающие фрагменты, содержащие пары аминокислотных последовательностей HCVR/LCVR, выбранные из группы, состоящей из SEQ ID NO: 18/26, 66/74, 130/138, 162/170, 242/250, 306/314, 322/330, 370/378 и 460/468.
В соответствии с одним вариантом осуществления фармацевтическая композиция содержит четыре выделенных человеческих моноклональных антитела, которые специфично связываются с Fel d1, или их антигенсвязывающие фрагменты, причем человеческие антитела или их антигенсвязывающие фрагменты содержат пары аминокислотных последовательностей HCVR/LCVR из SEQ ID NO: 18/26, 66/74, 130/138 и 162/170.
В соответствии с одним вариантом осуществления настоящее изобретение относится к композиции, которая представляет собой комбинацию из терапевтически эффективного количества одного или нескольких антител к Fel d1 или их антигенсвязывающих фрагментов по настоящему изобретению, и терапевтически эффективного количества второго терапевтического средства.
Вторым терапевтическим средством может быть низкомолекулярное лекарственное средство, белок/полипептид, антитело, молекула нуклеиновой кислоты, такая как антисмысловая молекула, или siRNA. Второе терапевтическое средство может иметь искусственное или естественное происхождение.
Второе терапевтическое средство может представлять собой любое средство, которое преимущественно скомбинировано с антителом или его фрагментом по настоящему изобретению, например, вторым антителом, отличным от описанных в настоящем документе, которое способно блокировать связывание Fel d1 с IgE, находящимся на тучных клетках или базофилах. Второе терапевтическое средство также может представлять собой любое средство, которое применяют в качестве стандарта лечения при проведении лечения аллергической реакции на любой аллерген. Такое второе терапевтическое средство может представлять собой антигистамин, эпинефрин, противоотечное средство, кортикостероид или пептидную вакцину.
В соответствии с некоторыми вариантами осуществления вторым терапевтическим средством может быть средство, которое способствует препятствованию возникновения или уменьшению любого возможного побочного эффекта(ов), связанного с антителом или антигенсвязывающим фрагментом антитела по настоящему изобретению, при появлении такого побочного эффекта(ов).
Также будет понятно, что антитела и фармацевтически приемлемые композиции по настоящему изобретению можно использовать в разновидностях комбинированной терапии, иначе говоря, антитела и фармацевтически приемлемые композиции можно вводить одновременно, до или после одного или нескольких других необходимых терапевтических средств или медицинских процедур. При конкретной комбинации терапий (терапевтических средств или процедур) для использования в комбинированной схеме будут учтены совместимость необходимых терапевтических средств и/или процедур и подлежащего достижению необходимого терапевтического эффекта. Также будет понятно, что при помощи используемых разновидностей терапии можно достичь необходимый эффект для одного нарушения (например, антитело можно вводить одновременно с другим средством, применяемым для лечения такого же заболевания) или можно достичь различные эффекты (например, контроль каких-либо отрицательных эффектов). Применяемые в настоящем описании дополнительные терапевтические средства, которые в норме вводят для лечения или предупреждения конкретного заболевания или состояния, являются пригодными для лечения такого заболевания или состояния.
При совместном введении множества терапевтических средств дозировки можно соответственным образом скорректировать, как известно из уровня техники.
В соответствии с седьмым аспектом настоящее изобретение относится к способу лечения больного, у которого наблюдают чувствительность или аллергическую реакцию на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1, или лечения по меньшей мере одного симптома или осложнения, ассоциированного с чувствительностью или аллергической реакцией на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1, предусматривающему введение эффективного количества одного или нескольких выделенных человеческих моноклональных антител или их антигенсвязывающих фрагментов, которые специфично связываются с Fel d1, или фармацевтической композиции, содержащей эффективное количество одного или нескольких выделенных человеческих моноклональных антител или их фрагментов, которые специфично связываются с Fel d1, или эффективного количества одной или нескольких биспецифических антигенсвязывающих молекул, которые специфично связывают Fel d1, или фармацевтической композиции, содержащей эффективное количество одной или нескольких из биспецифических антигенсвязывающих молекул, которые специфично связываются с Fel d1, нуждающемуся в этом больному, причем чувствительность или аллергическая реакция на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1 либо предупреждается, либо ослабляется по тяжести и/или продолжительности, или предупреждается или облегчается по меньшей мере один симптом или осложнение, ассоциированные с чувствительностью или аллергической реакцией на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1, или уменьшается такая частота и/или продолжительность или тяжесть чувствительности или аллергической реакции на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1 после введения одного или нескольких из выделенных человеческих моноклональных антител или их фрагментов, которые специфично связываются с Fel d1, или после введения одного или нескольких биспецифических антигенсвязывающих молекул, которые специфично связываются с Fel d1, или после введения композиции, содержащей любое одно или несколько из указанных выше антител или биспецифических антигенсвязывающих молекул.
В соответствии с одним вариантом осуществления настоящее изобретение относится к фармацевтической композиции, содержащей одно или несколько антител по настоящему изобретению или одну или несколько биспецифических антигенсвязывающих молекул, которые специфично связываются с Fel d1, для применения при лечении больного, у которого наблюдается чувствительность или аллергическая реакция на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1, или для лечения по меньшей мере одного симптома или осложнения, ассоциированного с чувствительностью или аллергической реакцией на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1, причем чувствительность или аллергическая реакция на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1 либо предупреждается, либо ослабляется по тяжести и/или продолжительности, или предупреждается или облегчается по меньшей мере один симптом или осложнение, ассоциированные с чувствительностью или аллергической реакцией на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1, или уменьшается такая частота и/или продолжительность или тяжесть чувствительности или аллергической реакции на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1.
В соответствии с одним вариантом осуществления настоящее изобретение относится к применению фармацевтической композиции, содержащей одно или несколько антител по настоящему изобретению или одну или несколько биспецифических антигенсвязывающих молекул, которые специфично связываются с Fel d1, при производстве лекарственного препарата для применения при лечении больного, у которого наблюдается чувствительность или аллергическая реакция на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1, или для лечения по меньшей мере одного симптома или осложнения, ассоциированного с чувствительностью или аллергической реакцией на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1, причем чувствительность или аллергическая реакция на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1 либо предупреждается, либо ослабляется по тяжести и/или продолжительности, или предупреждается или облегчается по меньшей мере один симптом или осложнение, ассоциированные с чувствительностью или аллергической реакцией на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1, или уменьшается такая частота и/или продолжительность или тяжесть чувствительности или аллергической реакции на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1.
В соответствии с одним вариантом осуществления настоящее изобретение относится к применению фармацевтической композиции, которая описана выше, причем композицию вводят в комбинации со вторым терапевтическим средством, пригодным для понижения аллергической реакции на кошку, кошачью перхоть, экстракт кошачьей шерсти или на белок Fel d1. В соответствии с одним вариантом осуществления настоящее изобретение относится к применению описанной выше фармацевтической композиции, причем второе терапевтическое средство выбрано из кортикостероида, бронхиального дилататора, антигистамина, эпинефрина, противоотечного средства, другого отличного антитела к Fel d1 и пептидной вакцины.
В соответствии с некоторыми вариантами осуществления антитела по настоящему изобретению или биспецифические антигенсвязывающие молекулы, которые специфично связываются с Fel d1, могут быть способны уменьшать, минимизировать или предупреждать по меньшей мере один симптом у больного, восприимчивого к кошачьему аллергену Fel d1, такой как чихание, гиперемия, заложенность носа, кашель, бронхолегочная обструкция, бронхоконстрикция, ринит или конъюнктивит.
В соответствии с одним вариантом осуществления антитела по настоящему изобретению или биспецифические антигенсвязывающие молекулы, которые специфично связываются с Fel d1, или композиция, содержащая одно или несколько антител по настоящему изобретению или одну или несколько из антигенсвязывающих молекул, которые специфично связываются с Fel d1, можно применять для предупреждения более серьезных осложнений in vivo, ассоциированных с аллергией на Fel d1, включающей астматические реакции, анафилактический шок или даже летальный исход в результате анафилактической реакции.
В соответствии с одним вариантом осуществления фармацевтическую композицию вводят больному в комбинации со вторым терапевтическим средством.
В соответствии с другим вариантом осуществления второе терапевтическое средство выбирают из группы, состоящей из антигистамина, эпинефрина, противоотечного средства, кортикостероида, другого отличного антитела к Fel d1, пептидной вакцины и любого другого средства для паллиативной терапии, пригодного для уменьшения тяжести аллергической реакции или для ослабления по меньшей мере одного симптома, ассоциированного с аллергической реакцией.
Другие варианты осуществления станут понятны при рассмотрении последующего подробного раскрытия.
Подробное раскрытие настоящего изобретения
Перед описание настоящих способов необходимо понять, что настоящее изобретение не ограничено конкретными способами и описанными экспериментальными условиями, поскольку такие способы и условия могут изменяться. Также следует понять, что применяемая в настоящем описании терминология приведена только с целью описания конкретных вариантов осуществления и не предназначена для ограничения, поскольку объем настоящего изобретения будет ограничен только прилагаемой формулой изобретения.
Если не указано иное, то все применяемые в настоящем документе технические и научные термины имеют такое же значение, которое обычно понимается рядовым специалистом в настоящей области техники, к которой относится настоящее изобретение. Применяемый в настоящем документе термин "приблизительно" при применении в отношении конкретного приведенного числового значения означает, что значение может варьировать от приведенного значения не более чем на 1%. Например, применяемое в настоящем документе выражение "приблизительно 100" включает 99 и 101 и все значения между ними (например, 99,1, 99,2, 99,3, 99,4 и т.п.).
Несмотря на то, что при осуществлении на практике или испытании настоящего изобретения можно применять любые способы и материалы, сходные или эквивалентные описанным в настоящем документе, далее описаны предпочтительные способы и материалы.
Определения
Применяемый в настоящем документе термин "Fel d1" или "FELD1" относится по меньшей мере к одному белку Fel d1, либо в природной/нативной форме, либо полученному рекомбинантно. Белок Fel d1 содержит цепь 1 (также называемую цепью А) Fel d1 (SEQ ID NO: 392) и цепь 2 (также называемую цепью В) Fel d1 (SEQ ID NO: 393). Природный белок Fel d1 представляет собой гетеродимерный гликопротеин с массой примерно 18 кДа, состоящий из двух цепей, полученных от двух независимых генов (См. Duffort, О.А. et al., (1991), Mol. Immunol. 28:301-309; Kristensen, A.K. et al., (1997), Biol. Chem. 378:899-908; Kaiser L. et al. (2003) J. Biol. Chem. 278(39):37730-37735). Рекомбинантно полученный белок Fel d1 также показан как SEQ ID NO: 396, причем эта последовательность содержит аминокислотные остатки 18-109 цепи В Fel d1 под номером доступа GenBank NP_001041619.1 (без сигнальной последовательности), последовательно гибридизированные с аминокислотными остатками 19-88 цепи A Fel d1 под номером доступа GenBank NP_001041618.1 (без сигнальной последовательности и с мутацией D27G, которая соответствует глицину в положении 101 из SEQ ID NO: 396). Другие рекомбинантно полученные конструкции Fel d1 по настоящему изобретению проиллюстрированы в SEQ ID NO: 385, 394, 395 и 397.
"Цепь 1" или "цепь A" Fel d1 представляет собой полипептид, содержащий аминокислотную последовательность из SEQ ID NO: 392 или гомологичную ей последовательность, или в качестве альтернативы состоящий из нее. Применяемый в настоящем документе термин гомологичная последовательность из SEQ ID NO: 392 относится к полипептиду, который имеет идентичность с SEQ ID NO: 392, которая превышает 70%, предпочтительно превышает 80%, более предпочтительно превышает 90% и еще более предпочтительно превышает 95%. В GenBank под номером доступа Р30438 или под номером доступа NP_001041618.1 также представлена аминокислотная последовательность цепи 1 Fel d1, которая также включает сигнальный пептид, удаленный в зрелом белке.
"Цепь 2" или "цепь В" Fel d1 представляет собой полипептид, содержащий аминокислотную последовательность из SEQ ID NO: 393 или гомологичную ей последовательность, или в качестве альтернативы состоящий из нее. Применяемый в настоящем документе термин гомологичная последовательность из SEQ ID NO: 393 относится к полипептиду, который имеет идентичность с SEQ ID NO: 393, которая превышает 70%, предпочтительно превышает 80%, более предпочтительно превышает 90% и еще более предпочтительно превышает 95%. В GenBank под номером доступа Р30440 или под номером доступа NP 001041619.1 также представлена аминокислотная последовательность цепи 2 Fel d1, которая включает сигнальный пептид, удаленный в зрелом белке.
Применяемый в настоящем документе термин "фрагмент Fel d1" относится к полипептиду, содержащему по меньшей мере один иммунодоминантный сайт Fel d1 или в качестве альтернативы состоящему из него. Согласно одному варианту осуществления применяемый в настоящем документе термин "фрагмент Fel d1" относится к полипептиду, содержащему по меньшей мере два иммунодоминантных сайта Fel d1 или в качестве альтернативы состоящему из них. В соответствии с одним вариантом осуществления иммунодоминантные сайты соединены ковалентно. В соответствии с одним вариантом осуществления иммунодоминантные сайты соединены посредством по меньшей мере одной пептидной связи. В соответствии с одним вариантом осуществления иммунодоминантные сайты соединены посредством по меньшей мере одной пептидной связи и спейсера между иммунодоминантными сайтами. В соответствии с одним вариантом осуществления по меньшей мере два иммунодоминантных сайта происходят как из цепи 1 Fel d1, так и из цепи 2 Fel d1. В соответствии с одним вариантом осуществления по меньшей мере два иммунодоминантных сайта включают аминокислотные последовательности 23-92 под номером доступа GenBank Р30438 и аминокислотные последовательности 18-109 под номером доступа GenBank Р30440. В соответствии с одним вариантом осуществления по меньшей мере два иммунодоминантных сайта происходят как из цепи 1 Fel d1, так и из цепи 2 Fel d1. В соответствии с одним вариантом осуществления по меньшей мере два иммунодоминантных сайта включают аминокислотные последовательности 19-88 под номером доступа GenBank NP_001041618.1 и аминокислотные последовательности 18-109 под номером доступа GenBank NP_001041619.1. В соответствии с одним вариантом осуществления по меньшей мере два иммунодоминантных сайта включают аминокислотную последовательность в любой из SEQ ID NO: 385, 394, 395, 396 или 397. В соответствии с одним вариантом осуществления любой из фрагментов Fel d1 способен индуцировать выработку антител in vivo, которые специфично связываются со встречающимся в природе Fel d1 или с рекомбинантно полученным Fel d1.
Применяемый в настоящем документе термин "антитело" означает любую антигенсвязывающую молекулу или молекулярный комплекс, которые содержат по меньшей мере один определяющий комплементарность участок (CDR), который специфично связывается или взаимодействует с конкретным антигеном (например, Fel d1). Применяемый в настоящем документе термин "антитело" предназначен для обозначения молекул иммуноглобулина, в состав которых входят четыре полипептидные цепи, две тяжелые (Н) цепи и две легкие (L) цепи, взаимосвязанные дисульфидными связями (т.е., "молекул целого антитела"), а также его мультимеров (например, IgM) или его антигенсвязывающих фрагментов. В состав каждой тяжелой цепи входят вариабельный участок тяжелой цепи ("HCVR" или "VH") и константный участок тяжелой цепи (в его состав входят домены CH1, CH2 и CH3). В состав каждой легкой цепи входят вариабельный участок легкой цепи ("LCVR или "VL") и константный участок легкой цепи (CL). Участки VH и VL дополнительно можно подразделить на участки гипервариабельности, называемые определяющими комплементарность участками (CDR), чередующиеся с участками, которые являются более консервативными, называемыми каркасными участками (FR). В состав каждого VH и VL входят три CDR и четыре FR, располагающиеся от аминоконца до карбоксиконца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В соответствии с определенными вариантами осуществления по настоящему изобретению FR антитела (или его антигенсвязывающего фрагмента) могут быть идентичными с человеческими последовательностями зародышевой линии или могут быть модифицированы естественным или искусственным путем. Аминокислотную консенсусную последовательность можно определить на основе анализа расположенных параллельно последовательностей двух или более CDR.
Также возможна замена одного или нескольких остатков CDR или исключение одного или нескольких CDR. В научной литературе описаны антитела, для функции связывания у которых не важны один или два CDR. Padlan et al. (1995 FASEB J. 9:133-139) анализировали контактные участки между антителами и их антигенами на основе опубликованных кристаллических структур и пришли к выводу, что фактически приблизительно только от одной пятой до одной трети остатков CDR контактируют с антигеном. Padlan также обнаружил множество антител, у которых один или два CDR не имели аминокислот, контактирующих с антигеном (см. также Vajdos et al. (2002), J Mol Biol 320:415-428).
Остатки CDR, не контактирующие с антигеном, можно определить на основании результатов предыдущих исследований (например, остатки Н60-Н65 в CDRH2 часто не являются необходимыми), исходя из участков CDR по Kabat, лежащих за пределами CDR Chothia, посредством молекулярного моделирования и/или опытным путем. Если CDR или его остаток(остатки) исключены, то он обычно заменен на аминокислоту, занимающую соответствующее положение в другой последовательности человеческого антитела или консенсусной последовательности таких последовательностей. Положения для замены в CDR и аминокислотах для замены можно также подобрать опытным путем. Эмпирические замены могут быть консервативными или неконсервативными заменами.
Раскрываемые в настоящем документе полностью человеческие моноклональные антитела, которые специфично связываются с Fel d1, могут содержать одну или несколько аминокислотных замен, вставок и/или делеций в каркасных и/или CDR участках вариабельных доменов тяжелых и легких цепей по сравнению с соответствующими последовательностями зародышевой линии. Такие мутации можно легко выявить путем сравнения раскрытых в настоящем документе аминокислотных последовательностей с последовательностями зародышевой линии, доступными, например, из находящихся в общем доступе баз данных последовательностей антител. Настоящее изобретение относится к антителам и к их антигенсвязывающим фрагментам, которые получены из любой из раскрытых в настоящем документе аминокислотных последовательностей, причем одна или несколько аминокислот в одном или нескольких каркасных и/или CDR участках являются мутировавшими по соответствующему остатку(остаткам) последовательности зародышевой линии, из которой было получено антитело, или по соответствующему остатку(остаткам) другой последовательности человеческой зародышевой линии, или по консервативной аминокислотной замене соответствующего остатка(остатков) зародышевой линии (такие изменения последовательности собирательно называются в настоящем документе как "мутации зародышевой линии"). Рядовой специалист в настоящей области, исходя из раскрываемых в настоящем документе последовательностей вариабельных участков тяжелых и легких цепей, легко может получить многочисленные антитела и антигенсвязывающие фрагменты, которые содержат одну или несколько отдельных мутаций зародышевой линии или их комбинации. В соответствии с определенными вариантами осуществления все каркасные и/или CDR остатки в доменах VH и/или VL являются мутировавшими обратно до остатков, обнаруживаемых в исходной последовательности зародышевой линии, из которой было получено антитело. Согласно другим вариантам осуществления только определенные остатки являются мутировавшими обратно до исходной последовательности зародышевой линии, например, только мутировавшие остатки, обнаруживаемые в первых 8 аминокислотах FR1 или в последних 8 аминокислотах FR4, или мутировавшие остатки, обнаруживаемые только в CDR1, CDR2 или CDR3. Согласно другим вариантам осуществления один или несколько каркасных и/или CDR остатков являются мутировавшими до соответствующих остатков отличной последовательности зародышевой линии (т.е. последовательности зародышевой линии, которая отличается от последовательности зародышевой линии, из которой исходно было получено антитело). Вместе с тем, антитела по настоящему изобретению могут содержать любую комбинацию из двух или более мутаций зародышевой линии в каркасных и/или CDR участках, в которой, например, некоторые отдельные остатки являются мутировавшими до соответствующего остатка конкретной последовательности зародышевой линии, в то время как некоторые другие остатки, которые отличаются от исходной последовательности зародышевой линии, сохранены или являются мутировавшими до соответствующего остатка отличной последовательности зародышевой линии. После получения антитела и антигенсвязывающие фрагменты, которые содержат одну или несколько мутаций зародышевой линии, можно легко протестировать на предмет одного или нескольких необходимых свойств, таких как улучшенная специфичность связывания, повышенная аффинность связывания, улучшенные или увеличенные антагонистические или агонистические биологические свойства (при соответствующих обстоятельствах), уменьшенная иммуногенность и т.д. Настоящее изобретение относится к антителам и антигенсвязывающим фрагментам, которые, в целом, получены таким способом.
Настоящее изобретение также относится к полностью человеческим моноклональным антителам, содержащим варианты любой из раскрываемых в настоящем документе аминокислотных последовательностей HCVR, LCVR и/или CDR, имеющей одну или несколько консервативных замен. Например, настоящее изобретение относится к антителам, имеющим аминокислотные последовательности HCVR, LCVR и/или CDR, например, с 10 или менее, 8 или менее, 6 или менее, 4 или менее и т.д. консервативными аминокислотными заменами относительно любой из раскрываемых в настоящем документе аминокислотных последовательностей HCVR, LCVR и/или CDR.
Применяемый в настоящем документе термин "человеческое антитело" подразумевают как включающий антитела, имеющие вариабельные и константные участки, полученные из иммуноглобулиновых последовательностей человеческой зародышевой линии. Человеческие mAb по настоящему изобретению могут включать аминокислотные остатки, не кодируемые иммуноглобулиновыми последовательностями человеческой зародышевой линии (например, мутации, введенные в результате неспецифического или сайт-направленного мутагенеза in vitro или соматической мутации in vivo), например, в CDR и, в частности, в CDR3. Тем не менее, применяемый в настоящем документе термин "человеческое антитело" не подразумевают как включающий mAb, у которых последовательности CDR, полученные из зародышевой линии других видов млекопитающих (например, мыши), были пришиты на человеческие последовательности FR.
Применяемое в настоящем документе выражение "антигенсвязывающая молекула" означает белок, полипептид или молекулярный комплекс, включающий по меньшей мере один гипервариабельный участок (CDR) или состоящий из них него, который отдельно, или в комбинации с одним или несколькими дополнительными CDR и/или каркасными участками (FR), специфично связывает конкретный антиген. В соответствии с некоторыми вариантами осуществления антигенсвязывающая молекула представляет собой антитело или фрагмент антитела, согласно определений этим терминам, приведенным в других частях настоящего документа.
Применяемое в настоящем документе выражение "биспецифическая антигенсвязывающая молекула" означает белок, полипептид или молекулярный комплекс, содержащий по меньшей мере первый антигенсвязывающий домен и второй антигенсвязывающий домен (т.е. два плеча). Каждый антигенсвязывающий домен в биспецифической антигенсвязывающей молекуле содержит по меньшей мере один CDR, который отдельно, или в комбинации с одним или несколькими дополнительными CDR и/или FR, специфично связывается с конкретным антигеном. В контексте настоящего изобретения первый антигенсвязывающий домен специфично связывает первый антиген на Fel d1, а второй антигенсвязывающий домен специфично связывает второй, отличный антиген на Fel d1.
Термин "специфично связывает", или "специфично связывается с", или т.п. означает, что антитело или его антигенсвязывающий фрагмент формирует с антигеном комплекс, который является относительно стабильным в физиологических условиях. Специфичное связывание можно охарактеризовать равновесной константой диссоциации, составляющей по меньшей мере приблизительно 1×10-6 M или менее (например, KD с меньшим значением указывает на более прочное связывание). Способы определения, специфично ли связываются две молекулы, хорошо известны в настоящей области техники и включают, например, равновесный диализ, поверхностный плазмонный резонанс и т.п. Описанные в настоящем документе антитела, которые специфично связываются с Fel d1, были определены с помощью поверхностного плазмонного резонанса, например, BIACORE™. Кроме того, применяемые в настоящем документе мультиспецифические антитела, которые связываются с Fel d1 и одним или несколько дополнительных антигенов, или биспецифическое антитело, которое связывается с двумя различными участками Fel d1 (например, цепь 1 и/или цепь 2 Fel d1), тем не менее считают антителами, которые "специфично связывают".
Термин антитело с "высокой аффинностью" относится к mAb, обладающим аффинностью связывания Fel d1, выраженной как KD, которая составляет по меньшей мере 10-8 М, предпочтительно 10-9 М, более предпочтительно 10-10 М, еще более предпочтительно 10-11 М, еще более предпочтительно 10-12 М, по результатам измерения с помощью поверхностного плазмонного резонанса, например, BIACORE™, или с помощью ELISA на аффинность в растворе.
Под термином "медленная скорость диссоциации", "Koff" или "kd" подразумевают антитело, которое диссоциирует от Fel d1 с константой скорости реакции 1×10-3 с-1 или менее, предпочтительно 1×10-4 с-1 или менее, по результатам измерений с помощью поверхностного плазмонного резонанса, например, BIACORE™.
Применяемые в настоящем документе термины "антигенсвязывающая часть" антитела, "антигенсвязывающий фрагмент" антитела и т.п. включают любой встречающийся в природе, получаемый ферментативно, синтетический или генетически сконструированный полипептид или гликопротеин, который специфично связывается с антигеном с формированием комплекса. Применяемые в настоящем документе термины "антигенсвязывающая часть" антитела или "фрагмент антитела" относятся к одному или нескольким фрагментам антитела, которые сохраняют способность связываться с Fel d1.
В соответствии с конкретными вариантами осуществления антитело или фрагменты антитела по настоящему изобретению могут быть конъюгированы с терапевтическим фрагментом ("иммуноконъюгат"), таким как кортикостероид, второе антитело к Fel d1 или эпинефрин, вакцина или любой другой терапевтический фрагмент, пригодный для лечения аллергической реакции на Fel d1.
Применяемый в настоящем документе термин "выделенное антитело" предназначен для обозначения антитела, которое практически не содержит других антител (Ab), имеющих отличные антигенные специфичности (например, выделенное антитело, которое специфично связывает Fel d1, или его фрагмент, практически не содержит Ab, которые специфично связывают антигены, отличные от Fel d1).
Применяемый в настоящем документе термин "блокирующее антитело" или "нейтрализующее антитело" (или "антитело, которое нейтрализует активность Fel d1") предназначен для обозначения антитела, или его антиген связывающей части, связывание которого с Fel d1 в результате приводит к ингибированию по меньшей мере одной биологической активности Fel d1. Например, антитело по настоящему изобретению может способствовать предупреждению первичной аллергической реакции на Fel d1. Альтернативно у антитела по настоящему изобретению можно наблюдать способность предупреждать вторичную аллергическую реакцию на Fel d1 или по меньшей мере один симптом аллергической реакции на Fel d1, включая чихание, кашель, астматическое состояние или анафилактическую реакцию, спровоцированную Fel d1. Такое ингибирование биологической активности Fel d1 можно оценить путем измерения одного или нескольких показателей биологической активности Fel d1 при помощи одного или более из нескольких стандартных анализов in vitro или in vivo (таких как описанный в настоящем документе анализ пассивной кожной анафилаксии) или других in vivo анализов, известных из уровня техники (например, других животных моделей для проверки защиты от провокационной дозы Fel d1 после введения одного или нескольких описанных в настоящем документе антител).
Применяемый в настоящем документе термин "поверхностный плазмонный резонанс" относится к оптическому явления, которое позволяет анализировать в реальном времени биомолекулярные взаимодействия путем определения изменений концентраций белка в матрице биосенсора, например, при помощи системы BIACORE™ (Pharmacia Biosensor АВ, Уппсала, Швеция и Пискатауэй, Нью-Джерси).
Применяемый в настоящем документе термин "KD" предназначен для обозначения равновесной константы диссоциации конкретного взаимодействия антитело-антиген.
Термин "эпитоп" относится к антигенной детерминанте, которая взаимодействует со специфичным антигенсвязывающим сайтом в вариабельном участке молекулы антитела, известном как паратоп. Один антиген может иметь более одного эпитопа. Таким образом, различные антитела могут связываться с различными областями на антигене и могут оказывать различные биологические эффекты. Термин "эпитоп" также относится к сайту на антигене, на который реагируют В- и/или Т-клетки. Он также относится к участку антигена, который связывается антителом. Эпитопы могут быть либо линейными, либо конформационными. Линейный эпитоп является эпитопом, получаемым посредством смежных аминокислотных остатков в полипептидной цепи. Конформационный эпитоп получают посредством пространственно соединенных аминокислот из различных сегментов линейной полипептидной цепи. В соответствии с определенными вариантами осуществления эпитопы могут включать детерминанты, которые являются химически активными поверхностными группировками таких молекул, как аминокислоты, сахарными боковыми цепями, фосфорильными группами или сульфонильными группами и, в соответствии с определенными вариантами осуществления, могут иметь характеристики, касающиеся трехмерной структуры, и/или характеристики удельного заряда. Эпитопы также можно определить как структурные или функциональные. Функциональные эпитопы в целом представляют собой подгруппу структурных эпитопов и имеют такие остатки, которые вносят непосредственный вклад в аффинность взаимодействия. Эпитопы, сформированные из смежных аминокислот, обычно остаются после воздействия денатурирующими растворителями, при этом эпитопы, сформированные в результате образования третичной структуры, обычно оказываются утраченными после обработки денатурирующими растворителями. Эпитоп, как правило, включает по меньшей мере 3 или, более характерно, по меньшей мере 5 или 8-10 аминокислот в уникальной пространственной конформации.
Термин "существенная идентичность" или "практически идентичный" в отношении нуклеиновой кислоты или ее фрагмента указывает на то, что при оптимальном выравнивании при помощи соответствующих нуклеотидных вставок или делеций с другой нуклеиновой кислотой (или комплементарной ей нитью) имеет место идентичность нуклеотидной последовательности по меньшей мере у приблизительно 90% и более предпочтительно по меньшей мере приблизительно 95%, 96%, 97%, 98% или 99% нуклеотидных оснований согласно результатам измерения при помощи любого хорошо известного алгоритма определения идентичности последовательностей, такого как FASTA, BLAST или GAP, который обсуждают ниже. Молекула нуклеиновой кислоты, характеризующаяся существенной идентичностью с эталонной молекулой нуклеиновой кислоты, может в определенных случаях кодировать полипептид, имеющий такую же или практически сходную аминокислотную последовательность, что и полипептид, кодируемый эталонной молекулой нуклеиновой кислоты.
Применительно к полипептидам термин "существенное сходство" или "практически сходный" означает, что две пептидные последовательности при оптимальном выравнивании, таком как посредством программ GAP или BESTFIT, с применением штрафов за открытие гэпов по умолчанию, имеют по меньшей мере 90% идентичность последовательности, еще более предпочтительно по меньшей мере 95%, 98% или 99% идентичность последовательности. Предпочтительно, положения остатков, которые не являются идентичными, отличаются по консервативным аминокислотным заменам. "Консервативная аминокислотная замена" является заменой, при которой аминокислотный остаток заменен на другой аминокислотный остаток, имеющий боковую цепь (группа R) со сходными химическими свойствами (например, зарядом или гидрофобностью). В целом, консервативная аминокислотная замена практически не будет изменять функциональные свойства белка. В случаях, когда две или более аминокислотные последовательности отличаются друг от друга по консервативным заменам, процент или степень сходства можно скорректировать до надлежащей по консервативной природы замены. Способы осуществления такой корректировки хорошо известны специалистам в настоящей области техники. (См., например, Pearson (1994) Methods Mol. Biol. 24: 307-331). Примеры групп аминокислот, которые имеют боковые цепи со сходными химическими свойствами, включают: 1) алифатические боковые цепи: глицин, аланин, валин, лейцин и изолейцин; 2) алифатически-гидроксильные боковые цепи: серии и треонин; 3) амидсодержащие боковые цепи: аспарагин и глутамин; 4) ароматические боковые цепи: фенилаланин, тирозин и триптофан; 5) основные боковые цепи: лизин, аргинин и гистидин; 6) кислотные боковые цепи: аспартат и глутамат; и 7) серосодержащие боковые цепи: цистеин и метионин. Предпочтительными группами консервативных аминокислотных замен являются: валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин, глутамат-аспартат и аспарагин-глутамин. Альтернативно, консервативным замещением является любое изменение, имеющее положительное значение в матрице логарифмического правдоподобия РАМ250, раскрытой в работе Gonnet et al. ((1992) Science 256: 1443 45). "Умеренно консервативное" замещение представляет собой любое изменение, имеющее неотрицательное значение в матрице логарифмического правдоподобия РАМ250.
Сходство последовательностей для полипептидов обычно измеряют при помощи программного обеспечения для анализа последовательностей. Программное обеспечение для анализа белка сопоставляет сходные последовательности при помощи измерений сходства, присвоенного различным заменам, делециям и другим модификациям, включая консервативные аминокислотные замены. Например, программное обеспечение GCG содержит программы, такие как GAP и BESTFIT, которые можно применять с параметрами по умолчанию для определения гомологии последовательностей или идентичности последовательностей между близкородственными полипептидами, такими как гомологичные полипептиды от различных видов организмов, или между белком дикого типа и его мутеином. См., например, GCG Version 6.1. Полипептидные последовательности также можно сравнивать при помощи FASTA с параметрами по умолчанию или рекомендованными параметрами, программы в GCG Version 6.1. FASTA (например, FASTA2 и FASTA3) выводит результаты выравнивания и процентную идентичность последовательности для участков с наилучшим совпадением между запрашиваемой и поисковой последовательностями (Pearson (2000) supra). Другим предпочтительным алгоритмом при сравнении последовательности по настоящему изобретению с базой данных, содержащей большое количество последовательностей различных организмов, является компьютерная программа BLAST, в особенности BLASTP или TBLASTN, с применением параметров по молчанию. (См., например, Altschul et al. (1990) J. Mol. Biol. 215: 403 410 и (1997) Nucleic Acids Res. 25:3389 402).
Согласно конкретным вариантам осуществления антитело или фрагмент антитела для применения согласно способу по настоящему изобретению может быть моноспецифическим, биспецифическим или мультиспецифическим.
Мультиспецифические антитела могут быть специфичными к различным эпитопам одного целевого полипептида или могут содержать антигенсвязывающие домены, специфичные к эпитопам более одного целевого полипептида. Иллюстративный формат биспецифического антитела, который можно применять в контексте настоящего изобретения, включает применение первого домена CH3 иммуноглобулина (Ig) и второго домена CH3 Ig, причем первый и второй домены CH3 Ig отличаются друг от друга по меньшей мере по одной аминокислоте, и причем по меньшей мере одно аминокислотное отличие уменьшает связывание биспецифического антитела с белком А по сравнению с биспецифическим антителом без аминокислотного отличия. В соответствии с одним вариантом осуществления первый домен CH3 Ig связывается с белком А, а второй домен Ig CH3 содержит мутацию, которая уменьшает или ликвидирует связывание белка А, такую как модификация H95R (согласно нумерации экзонов IMGT; H435R согласно нумерации EU). Второй CH3 может дополнительно содержать модификацию Y96F (согласно IMGT; Y436F согласно EU). Дополнительные модификации, которые можно обнаружить во втором CH3, включают: D16E, L18M, N44S, K52N, V57M и V82I (согласно IMGT; D356E, L358M, N384S, K392N, V397M и V422I согласно EU) в случае mAb IgG1; N44S, K52N и V82I (IMGT; N384S, K392N и V422I согласно EU) в случае mAb IgG2; и Q15R, N44S, K52N, V57M, R69K, E79Q и V82I (согласно IMGT; Q355R, N384S, K392N, V397M, R409K, E419Q и V422I согласно EU) в случае mAb IgG4. Описанные выше варианты формата биспецифических антител предусмотрены объемом настоящего изобретения.
Под фразой "терапевтически эффективное количество" подразумевают количество, которое производит необходимый эффект у того, кому его вводят. Точное количество будет зависеть от цели лечения и может быть установлено специалистом в настоящей области техники при помощи известных методик (см., например, Lloyd (1999) The Art, Science and Technology of Pharmaceutical Compounding).
Антитела по настоящему изобретению можно применять для "гипосенсибилизирования" чувствительного к кошачьим аллергенам индивидуума. В настоящем документе термин "гипосенсибилизировать" обозначает уменьшать аллергическую реактивность чувствительного к кошачьим аллергенам индивидуума при контакте с кошками, кошачьей перхотью или их продуктами, например, Fel d1 (до уровня, меньшего, чем уровень, который чувствительный к кошачьим аллергенам индивидуум мог бы испытывать в обратном случае).
Общее описание
Домашняя кошка является источником многих комнатных аллергенов, а степени тяжести симптомов у индивидуумов, у которых наблюдают чувствительность к кошачьим аллергенам, варьируют от относительно слабого ринита и конъюнктивита до потенциально опасного для жизни астматического состояния (Lau, S. et al. (2000), Lancet 356:1392-1397). Не смотря на то, что у больных, у которых наблюдают такую чувствительность к кошкам, судя по всему, восприимчивы к различным молекулам, встречаемым в кошачьей перхоти и коже, основным аллергеном, по-видимому, является Fel d1 (аллерген 1 Felis domesticus). Было показано, что более 80% больных, которые являются аллергиками на кошек, имеют антитела IgE к этому антигену (van Ree, R. et al. (1999), J. Allergy Clin. Immunol 104:1223-1230).
Белок Fel d1 представляет собой гетеродимерный кислый гликопротеин с массой примерно 18 кДа, который содержит приблизительно 10-20% N-связанных углеводов. Каждый гетеродимер содержит две полипептидные цепи, которые кодируются двумя отдельными генами (Duffort, OA, et al., (1991), Mol. Immunol. 28:301-309; Morgenstern, JP, et al., (1991), PNAS 88:9690-9694; Griffith, I.J., et al. (1992), Gene 113:263-268). Цепь 1 содержит приблизительно 70 аминокислотных остатков, а цепь 2 содержит приблизительно 90-92 аминокислотных остатка. Было выдвинуто предположение о трех межцепочечных дисульфидных связях, связывающих две цепи в природном Fel d1 (Kristensen, А.K. et al. (1997), Biol. Chem. 378:899-908), и они были подтверждены для рекомбинантного Fel d1 в кристаллической структуре (Kaiser, L. et al. (2003) J. Biol. Chem. 278:37730-37735; Kaiser, L. et al., (2007), J. Mol. Biol. 370:714-727). Хотя иногда отдельно каждую цепь называют Fel d1, для полного белка аллергена необходимы обе цепи.
Fel d1 вырабатывается сальными железами, ороговевающим эпителиальным слоем желез и клетками плоского эпителия и переносится на шкуру при вылизывании и уходом за шерстью (Bartholome, K. et al. (1985), J. Allergy Clin. Immunol. 76:503-506; Charpin, С. et al. (1991), J. Allergy Clin. Immunol. 88:77-82; Dabrowski, A.J. (1990), et al. J. Allergy Clin. Immunol. 86:462-465). Он также присутствует в слюнных, околоанальных и слезных железах (Andersen, М.С., et al. (1985), J. Allergy Clin. Immunol. 76:563-569; van Milligen, F.J. et al., (1992), Int. Arch. Allergy Appl. Immunol. 92:375-378) и основными источниками оказываются кожа и мех (Mata, P. et al. (1992), Ann. Allergy 69(4):321-322).
Белок Fel d1 имеет неизвестную функцию у животных, но вызывают реакцию с выработкой IgG или IgE у восприимчивых людей (в виде либо аллергической, либо астматической реакции). Несмотря на то, что известны другие аллергены, в том числе Fel d2 (альбумин) и Fel d3 (цистатин), от 60% до 90% вырабатываемого IgE к аллергенам кошки направлено к Fel d1 (Leitermann, K. et al., (1984), J Allergy Clin. Immunol. 74:147-153; Lowenstein, H. et al., (1985), Allergy 40:430-441; van Ree, R. et al., (1999), J. Allergy Clin. Immunol. 104:1223-1230; Ichikawa, K. et al., (2011), Clin. Exp. Allergy, 31:1279-1286).
Иммуноглобулин Ε (IgE) ответственен за гиперчувствительность 1 типа, которая обнаруживается при аллергических ринитах, аллергических конъюнктивитах, сенной лихорадке, аллергической астме, аллергии на пчелиный яд и аллергиях на продукты питания. IgE циркулирует в крови и связывается с высокоаффинными рецепторами Fe к IgE на базофилах и тучных клетках. При большинстве аллергических реакциях аллергены попадают в организм посредством вдыхания, приема внутрь или через кожу. Затем аллерген связывается с преформированным IgE, уже связавшимся с высокоаффинным рецептором на поверхностях тучных клеток и базофилов, что в результате приводит к сшиванию нескольких молекул IgE и запуску высвобождения гистамина и других медиаторов воспаления, обуславливая появление различных аллергических симптомов.
Лечение аллергий на кошек включает гипосенсибилизирующую терапию, которая предусматривает повторные введения инъекцией с повышающимися дозировками либо неочищенного экстракта кошачьей перхоти, либо коротких пептидов, полученных из Fel d1. Применение неочищенного экстракта кошачьей перхоти, Lilja et al., продемонстрировало, что после трех лет такого лечения у больных с аллергией на кошек все еще наблюдали системные симптомы (Lilja, Q. et al. (1989), J. Allergy Clin. Immunol. 83:37-44; и Hedlin, et al. (1991), J. Allergy Clin. Immunol. 87:955-964). Применение коротких пептидов, полученных из Fel d1, для гипосенсибилизации приводили в результате к незначимому отличию между группой пептида и контрольной группой плацебо (Oldfield, W.L. et al., (2002), Lancet, 360:47-53). Эффективность наблюдали только при введении больным больших количеств (750 мкг) короткого пептида (Norman, P.S. et al. (1996), Am. J. Respir. Crit. Care Med. 154:1623-1628). Более того, астматические реакции отмечали у больных, которые получали как неочищенные экстракты из кошачьей перхоти, так и у больных, которые получали лечение коротким пептидом Fel d1. Соответственно, в области лечения аллергии на кошек существует потребность в альтернативных стратегиях лечения больных, чувствительных к кошачьим аллергенам, в частности Fel d1.
В качестве общей стратегии лечения аллергий были предложены антитела, поскольку они могут блокировать попадание аллергенных молекул в слизистую оболочку ткани или могут связывать аллерген до того, как у него появится возможность связаться с IgE, связавшимся с высокоаффинным рецептором на тучных клетках или базофилах, таким образом предупреждая высвобождение гистамина и других медиаторов высвобождения из этих клеток. В патенте США №5670626 описано применение моноклональных антител для лечения IgE-опосредованных аллергических заболеваний, таких как аллергический ринит, аллергическая астма и аллергический конъюнктивит, путем блокировки связывания аллергенов со слизистой оболочкой ткани. В патенте США №6849259 описано применение аллерген-специфических антител для подавления аллергического воспаления в in vivo мышиной модели аллергии. Были описаны молочные и яичные системы антител. Например, в US 20030003133 A1 раскрыто применение молока в качестве носителя аллергенов для индукции толерантности при пероральном приеме кошачьей перхоти и других аллергенов. В US 2010/0143266 были описаны композиции и способы снижения аллергической реакции у животных на аллерген в окружающей среде посредством применения молекулы, которая подавляет способность аллергена связываться с тучными клетками. Другие антитела к Fel d1 описаны в работе de Groot et. al. (de Groot et. al., (1988), J. Allergy Clin. Immunol. 82:778-786).
Описанные в настоящем документе полностью человеческие антитела демонстрируют специфичное связывание с Fel d1 и могут быть пригодными для лечения больных, страдающих от кошачьих аллергенов, в частности, больных, у которых наблюдают чувствительность к аллергену Fel d1. Применение таких антител может представлять собой эффективное средство для лечения больных, страдающих от аллергий на кошачью перхоть, или их можно применять для предупреждения повышенной реакции на Fel d1 при вторичном контакте с аллергеном или сопутствующих симптомов, ассоциированных с аллергией, или их можно применять для ослабления тяжести и/или продолжительности аллергической реакции, ассоциированной с первичным контактом с контактом с кошкой, являющейся носителем аллергена Fel d1, или с рецидивом симптомов при вторичном контакте с аллергеном. Их можно применять отдельно или в качестве вспомогательной терапии с другими терапевтическими веществами или средствами, известными из уровня техники, для лечения таких аллергий, такими как без ограничения лечение при помощи кортикостероидов или эпинефрина. Их можно применять в сочетании со вторым или третьим отличным антителом, специфичным к Fel d1. Их можно применять с аллерген-специфической иммунотерапией (SIT).
В соответствии с некоторыми вариантами осуществления антитела по настоящему изобретению получают из мышей, иммунизированных первичным иммуногеном, таким как природный Fel d1, который можно приобрести коммерчески (см., например, Indoor Biotech, № NA-FD1-2) или можно получить рекомбинантно. В соответствии с некоторыми вариантами осуществления иммуноген может представлять собой либо цепь 1 Fel d1, либо цепь 2 Fel d1 или может представлять собой комбинацию из цепи 1 и цепи 2, вводимых последовательно или одновременно. Аминокислотная последовательность полной длины цепи 1 (также называемая как FELD1 А) показана как SEQ ID NO: 392. Аминокислотные последовательности полной длины для цепи 1 также можно найти в GenBank под номерами доступа Р30438 и NP_001041618.1. Аминокислотная последовательность полной длины цепи 2 (также называемая как FELD1 В) показана как SEQ ID NO: 393. Аминокислотные последовательности полной длины для цепи 2 также можно найти в GenBank под номерами доступа РР30440 и NP_001041619.1.
В соответствии с некоторыми вариантами осуществления рекомбинантно полученный иммуноген Fel d1 можно получить путем прямой гибридизации двух цепей Fel d1, как описано в работе Kaiser et. al., с получением гибридного продукта, который имеет паттерн рефолдинга, сходный с паттерном природного Fel d1 (Kaiser, L. et al., (2003), J. Biol. Chem. 278(39):37730-37735). В соответствии с некоторыми вариантами осуществления иммуноген может представлять собой гибридный белок, такой как показанный в конструкциях SEQ ID NO: 385, 394, 395, 396 или 397, с последующей иммунизацией вторичным иммуногеном или иммуногенно активным фрагментом природного или рекомбинантно полученного Fel d1.
Иммуноген может быть биологически активным и/или иммуногенным фрагментом природного или рекомбинантно полученного Fel d1 или ДНК, кодирующей его активный фрагмент. Фрагмент можно получить или с N-конца, или с С-конца либо цепи 1, либо цепи 2, или с N-конца, или С-конца как цепи 1, так и цепи 2. Фрагменты можно получить из любого сайта в цепи 1 или цепи 2 для применения в качестве иммуногена для получения антитела к Fel d1.
В соответствии с некоторыми вариантами осуществления иммуноген может представлять собой гибридный белок, содержащий любое одно или несколько из следующих: i) аминокислотные остатки 18-109 цепи 2 в Fel d1 (см. номер доступа в GenBank Р30440, а также SEQ ID NO: 393), гибридизированные через С-конец непосредственно с N-концом аминокислотных остатков 23-92 цепи 1 в Fel d1 (см. номер доступа в GenBank Р30438, а также SEQ ID NO: 392); ii) аминокислотные остатки 23-92 цепи 1 в Fel d1 (см. номер доступа в GenBank Р30438, а также SEQ ID NO: 392), гибридизированные через С-конец с N-концом аминокислотных остатков 18-109 цепи 2 в Fel d1 (см. номер доступа в GenBank Р30440, а также SEQ ID NO: 393); iii) аминокислотные остатки 18-109 цепи 2 в Fel d1 (см. номер доступа в GenBank NP_001041619.1), гибридизированные через С-конец непосредственно с N-концом аминокислотных остатков 19-88 цепи 1 в Fel d1 (см. номер доступа в GenBank NP_001041618.1), например, конструкция, показанная в SEQ ID NO: 394 или 396; iv) аминокислотные остатки 19-88 цепи 1 в Fel d1 (см. номер доступа в GenBank NP_001041618.1), гибридизированные через С-конец с N-концом аминокислотных остатков 18-109 цепи 2 в Fel d1 (см. номер доступа в GenBank NP_001041619.1). См. также SEQ ID NO: 395. В соответствии с некоторыми вариантами осуществления гибридный белок мог иметь метку на C-терминальном конце конструкции, такую как myc-myc-гексагистидиновая метка (см. SEQ ID NO: 385, 396 или 397 для таких конструкций). Согласно соответствующим вариантам осуществления гибридный белок может иметь мышиный участок Fc антитела, присоединенный на C-терминальном конце конструкции (см. SEQ ID NO: 394 или 395 для таких конструкций). В соответствии с некоторыми вариантами осуществления цепи 1 и 2 соединены с помощью линкера, который известен специалистам в настоящей области техники, например (G4S)3 (см. SEQ ID NO: 395 и 397 для такой конструкции).
В соответствии с некоторыми вариантами осуществления антитела, которые специфично связываются с Fel d1, можно получить с помощью фрагментов указанных выше участков, или пептидов, которые выходят за рамки обозначенных участков на от приблизительно 5 до приблизительно 20 аминокислотных остатков с любого, или с обоих, N- или C-терминальных концов описанных в настоящем документе участков. В соответствии с некоторыми вариантами осуществления при получении специфичных к Fel d1 антител можно применять любую комбинацию указанных выше участков или их фрагментов. В соответствии с некоторыми вариантами осуществления для получения моноспецифических, биспецифических или мультиспецифических антител можно применять любой один или несколько из указанных выше участков Fel d1 или их фрагментов.
Антигенсвязывающие фрагменты антител
Если специально не указано иное, применяемый в настоящем документе термин "антитело" необходимо понимать как охватывающий молекулы антител, содержащих две иммуноглобулиновые тяжелые цепи и две иммуноглобулиновые легкие цепи (т.е. "целые молекулы антител"), а также их антигенсвязывающие фрагменты. Применяемые в настоящем документе термины "антигенсвязывающая часть" антитела, "антигенсвязывающий фрагмент" антитела и т.п. включают любой встречающийся в природе, получаемый ферментативно, синтетический или генетически сконструированный полипептид или гликопротеин, который специфично связывается с антигеном с формированием комплекса. Применяемые в настоящем документе термины "антигенсвязывающая часть" антитела или "фрагмент антитела" относятся к одному или нескольким фрагментам антитела, которые сохраняют способность специфично связываться либо с цепью 1 и/либо с цепью 2 Fel d1. Фрагмент антитела может включать фрагмент Fab, фрагмент F(ab')2, фрагмент Fv, фрагмент dAb, фрагмент, содержащий CDR, или выделенный CDR. Антигенсвязывающие фрагменты антитела могут быть получены, например, от целых молекул антител с применением любых подходящих стандартных методик, таких как протеолитическое расщепление или рекомбинантные методики генетической инженерии, предусматривающие манипуляцию и экспрессию ДНК, кодирующей вариабельные и (необязательно) константные домены антитела. Такая ДНК известна и/или легко доступна из, например, коммерческих источников, библиотек ДНК (включая, например, библиотеки фаг-антитело), или ее можно синтезировать. ДНК можно секвенировать и с ней можно проводить манипуляции химическими способами или при помощи методик молекулярной биологии, например, для организации одного или нескольких вариабельных и/или константных доменов в подходящую конфигурацию или введения кодонов, создания цистеиновых остатков, модификации, добавления или удаления аминокислот и т.д.
Неограничивающие примеры антигенсвязывающих фрагментов включают: (i) фрагменты Fab; (ii) фрагменты F(ab')2; (iii) фрагменты Fd; (iv) фрагменты Fv; (v) одноцепочечные молекулы Fv (scFv); (vi) фрагменты dAb и (vii) минимальные единицы распознания, состоящие из аминокислотных остатков, которые имитируют гипервариабельный участок антитела (например, выделенный определяющий комплементарность участок (CDR), такой как пептид CDR3), или ограниченный пептид FR3-CDR3-FR4. Также применяемым в настоящем документе выражением "антигенсвязывающий фрагмент" охвачены другие сконструированные молекулы, такие как домен-специфичные антитела, антитело с одним доменом, антитела с удаленным доменом, химерные антитела, антитела с пришитым CDR, диатела, триотела, тетратела, миниантитела, нанотела (например, моновалентные нанотела, бивалентные нанотела и т.д.), иммунопрепараты на основе модульного белка с малым размером молекул (SMIP) и вариабельные домены IgNAR акулы.
Антигенсвязывающий фрагмент антитела, как правило, будет содержать по меньшей мере один вариабельный домен. Вариабельный домен может иметь любой размер или аминокислотный состав и, как правило, будет содержать по меньшей мере один CDR, который является смежным с одной или несколькими каркасными последовательностями или находится в их пределах. В антигенсвязывающих фрагментах, имеющих домен VH, связанный с доменом VL, домены VH и VL могут быть расположены относительно друг друга в любом подходящем порядке. Например, вариабельный участок может быть димерным и содержать димеры VH-VH, VH-VL или VL-VL. Альтернативно, антигенсвязывающий фрагмент антитела может содержать мономерный домен VH или VL.
В соответствии с определенными вариантами осуществления антигенсвязывающий фрагмент антитела может содержать по меньшей мере один вариабельный домен, ковалентно связанный по меньшей мере с одним константным доменом. Неограничивающие иллюстративные конфигурации вариабельных или константных доменов, которые можно обнаружить в антигенсвязывающем фрагменте антитела по настоящему изобретению, включают: (i) VH-CH1; (ii) VH-CH2; (iii) VH-CH3; (iv) VH-CH1-CH2; (v) VH-CH1-CH2-CH3; (vi) VH-CH2-CH3; (vii) VH-CL; (viii) VL-CH1; (ix) VL-CH2; (x) VL-CH3; (xi) VL-CH1-CH2; (xii) VL-CH1-CH2-CH3; (xiii) VL-CH2-CH3; и (xiv) VL-CL. В любой конфигурации вариабельных и константных доменов, включая любые из перечисленных выше иллюстративных конфигураций, вариабельные и константные домены могут быть либо непосредственно соединены друг с другом, либо могут быть соединены при помощи полного или частичного шарнирного или линкерного участка. Шарнирный участок может состоять по меньшей мере из 2 (например, 5, 10, 15, 20, 40, 60 или более) аминокислот, что в результате дает гибкую или полугибкую связь между смежными вариабельными и/или константными доменами в отдельной полипептидной молекуле. Кроме того, антигенсвязывающий фрагмент антитела по настоящему изобретению может содержать гомодимер или гетеродимер (или другой мультимер) в любой из перечисленных выше конфигураций вариабельных и константных доменов в нековалентной связи друг с другом и/или с одним или несколькими мономерными доменами VH или VL (например, посредством дисульфидной связи(связей)).
Как и в случае с целыми молекулами антител, антигенсвязывающие фрагменты могут быть моноспецифическими или мультиспецифическими (например, биспецифическими). Мультиспецифический антигенсвязывающий фрагмент антитела, как правило, будет содержат по меньшей мере два различных вариабельных домена, причем каждый вариабельный домен способен специфично связываться с отдельным антигеном или с различным эпитопом на одном антигене. Любой мультиспецифический формат антитела, включая раскрываемые в настоящем документе иллюстративные биспецифические форматы антител, можно адаптировать для применения в контексте антигенсвязывающего фрагмента антитела по настоящему изобретения с помощью доступных в настоящей области техники стандартных методик.
Получение человеческих антител
Способы создания человеческих антител в трансгенных мышах известны из уровня техники. Для создания человеческих антител, которые специфично связываются с Fel d1, в контексте настоящего изобретения можно применять любые такие известные способы.
С помощью технологии VELOCIMMUNE™ (см., например, патентный документ US 6596541, Regeneran Pharmaceuticals, VELOCIMMUNE®) или любого другого известного способа создания моноклональных антител, изначально выделяют с химерные антитела с высокой аффинностью к Fel d1 человеческим вариабельным участком и мышиным константным участком. Технология VELOCIMMUNE® предусматривает создание трансгенной мыши, имеющей геном, содержащий локусы человеческих вариабельных участков тяжелых и легких цепей, функционально связанные с эндогенными локусами мышиных константных участков так, чтобы мышь в ответ на стимуляцию антигеном продуцировала антитело, содержащее человеческий вариабельный участок и мышиный константный участок. ДНК, кодирующую вариабельные участки тяжелых и легких цепей антитела, выделяют и функционально связывают с ДНК, кодирующей человеческие константные участки тяжелых и легких цепей. Затем ДНК экспрессируют в клетке, способной экспрессировать полностью человеческое антитело.
Как правило, мыше VELOCIMMUNE® вводят представляющий интерес антиген и выделяют лимфатические клетки (такие как В-клетки) из мышей, которые экспрессируют антитела. Лимфатические клетки можно гибридизировать с линией миеломных клеток с получением бессмертных линий гибридомных клеток, и такие линии гибридомных клеток подвергают скринингу и отбирают для выявления линий гибридомных клеток, которые продуцируют антитела, специфичные к представляющему интерес антигену. ДНК, кодирующая вариабельные участки тяжелой цепи и легкой цепи, можно выделить и соединить с необходимыми изотипическими константными участками тяжелой цепи и легкой цепи. Такой белок антитела может продуцироваться в такой клетке, как клетка СНО. Альтернативно, ДНК, кодирующую антиген-специфические химерные антитела или вариабельные домены легких и тяжелых цепей, можно выделить непосредственно из антиген-специфических лимфоцитов.
Изначально, химерные антитела с высокой аффинностью выделяют с человеческим вариабельным участком и мышиным константным участком. Так же, как и в приведенном ниже экспериментальном разделе, у антител определяют характеристики и их отбирают по необходимым характеристикам, в том числе аффинности, избирательности, эпитопу и т.д. Мышиные константные участки замещают необходимыми человеческими константными участками для создания полностью человеческого антитела по настоящему изобретению, например, дикого типа или модифицированного IgG1 или IgG4. Несмотря на то, что выбранный константный участок может варьировать в соответствии с конкретным применением, характеристики связывания антигена с высокой аффинностью и специфичности к мишени присущи вариабельному участку.
В целом, антитела по настоящему изобретению обладают очень высокоаффинными характеристиками, как правило обладая KD, составляющей от приблизительно 10-12 до приблизительно 10-9 М, при измерении по связыванию с антигеном, либо иммобилизированным на твердой фазе, либо в жидкой фазе. Мышиные константные участки замещают необходимыми человеческими константными участками для создания полностью человеческих антител по настоящему изобретению. Несмотря на то, что выбранный константный участок может варьировать в соответствии с конкретным применением, характеристики связывания антигена с высокой аффинностью и специфичности к мишени присущи вариабельному участку.
Биоэквиваленты
Антитела к Fel d1 и фрагменты антител по настоящему изобретению охватывают белки с аминокислотными последовательностями, которые отличаются от последовательностей описанных антител, но которые сохраняют способность связывать Fel d1. Такие варианты антител и фрагментов антител содержат одно или несколько добавлений, делеций или замен аминокислот по сравнению с исходной последовательностью, но характеризуются биологической активностью, которая фактически равна активности описанных антител. Аналогичным образом, кодирующие антитела последовательности ДНК по настоящему изобретению охватывают последовательности, которые содержат одно или несколько добавлений, делеций или замен нуклеотидов по сравнению с раскрываемой последовательностью, но которые кодируют антитело или фрагмент антитела, которые фактически биоэквивалентны антителу или фрагменту антитела по настоящему изобретению.
Два антигенсвязывающих белка, или антитела, считают биоэквивалентом, если, например, они являются фармацевтическими эквивалентами или фармацевтическими альтернативами, в скорости и степени абсорбции которых не наблюдают значительного различия при введении в одинаковой молярной дозе при сходных экспериментальных условиях, либо в единоразовой дозе, либо в многократной дозе. Некоторые антитела будут считать эквивалентами или фармацевтическими альтернативами, если они являются эквивалентными по степени их абсорбции, но не по скорости их абсорбции, и их все еще можно считать биоэквивалентными, так как такие различия в скорости абсорбции являются преднамеренными и учтены при введении меток, не являются существенным для достижения эффективных концентраций лекарственного средства в организме, например, при затяжном приеме, и их считают незначительными в медицинском плане для конкретной исследуемой готовой лекарственной формы.
В соответствии с одним вариантом осуществления два антигенсвязывающих белка являются биоэквивалентными, если они не имеют клинически существенных отличий по их безопасности, чистоте и активности.
В соответствии с одним вариантом осуществления два антигенсвязывающих белка являются биоэквивалентами, если больной может менять один или несколько раз эталонный продукт и биологический продукт без предположительного повышения риска отрицательных эффектов, включая клинически значимое изменение иммуногенности или сниженную эффективность, по сравнению с продолжительной терапией без такой замены.
В соответствии с одним вариантом осуществления два антигенсвязывающих белка являются биоэквивалентными, если они оба действуют по общему механизму или механизмам действия по отношению к условию или условиям применения, при условии, что такие механизмы известны.
Биоэквивалентность можно продемонстрировать посредством способов in vivo и/или in vitro. Измерения биоэквивалентности включают, например, (а) тестирование in vivo на людях или других млекопитающих, при котором концентрацию антитела или его метаболитов измеряют в крови, плазме крови, сыворотке крови или другой биологической жидкости в зависимости от времени; (b) тестирование in vitro, для которого установлена корреляция, и оно является достаточно прогнозируемым по отношению к данным биологической доступности in vivo у человека; (с) тестирование in vivo на людях или других млекопитающих, при котором соответствующий быстрый фармакологический эффект антитела (или его мишени) измеряют в зависимости от времени; и (d) строго контролируемое клиническое испытание, при котором устанавливают безопасность, эффективность, или биологическую доступность, или биоэквивалентность антитела.
Биоэквивалентные варианты антител по настоящему изобретению можно сконструировать, например, посредством создания различных замещений остатков или последовательностей или удаления концевых или внутренних остатков или последовательностей, которые не являются необходимыми для биологической активности. Например, цистеиновые остатки, несущественные для биологической активности, можно удалить или заместить на другие аминокислоты для предупреждения формирования нежелательных или неправильных внутримолекулярных дисульфидных мостиков при ренатурации. Согласно другому контексту, биоэквивалентные антитела могут включать варианты антител, содержащие аминокислотные изменения, которые модифицируют характеристики гликозилирования у антител, например, мутации, которые исключают или удаляют участки гликозилирования.
Биологические характеристики антител
Как правило, антитела по настоящему изобретению могут функционировать, связываясь либо с цепью 1, либо с цепью 2 Fel d1, либо как с цепью 1, так и с цепью 2 Fel d1, или с фрагментом либо цепи 1, либо цепи 2.
В соответствии с определенными вариантами осуществления антитела по настоящему изобретению могут связываться с эпитопом, расположенным по меньшей мере в C-концевом участке либо цепи 1, либо цепи 2 Fel d1. В соответствии с одним вариантом осуществления антитела по настоящему изобретению могут связываться с эпитопом в N-терминальном участке либо цепи 1, либо цепи 2 Fel d1.
В соответствии с некоторыми вариантами осуществления антитела по настоящему изобретению могут функционировать путем блокировки или ингибирования связывания IgE с тучными клетками или базофилами у больных, чувствительных к аллергену Fel d1.
В соответствии с некоторыми вариантами осуществления антитела по настоящему изобретению могут функционировать путем связывания с любым другим участком или фрагментом цепи 1 или цепи 2 полной длины природного белка Fel d1, аминокислотная последовательность которого показана в SEQ ID NO: 392 (цепь 1) и SEQ ID NO: 393 (цепь 2).
В соответствии с определенными вариантами осуществления антитела по настоящему изобретению могут представлять собой биспецифические антитела. Биспецифические антитела по настоящему изобретению могут связывать один эпитоп в цепи 1 и также могут связывать один эпитоп в цепи 2. В соответствии с некоторыми вариантами осуществления биспецифические антитела по настоящему изобретению могут связывать два различных эпитопа в цепи 1. В соответствии с некоторыми вариантами осуществления биспецифические антитела по настоящему изобретению могут связывать два различных эпитопа в цепи 2. В соответствии с некоторыми вариантами осуществления биспецифические антитела по настоящему изобретению могут связываться с двумя различными сайтами в одной спирали на одной из либо цепи 1, либо цепи 2, или могут связываться с одной спиралью как на цепи 1, так и на цепи 2. Структура Fel d1 более подробно описана в работе Kaiser et. al. (Kaiser, L. et. al. (2003) J. Biol. Chem. 278 (39):37730-37735), где авторы отмечают, что Fel d1 состоит из восьми спиралей, Н1-Н4 и Н5-Н8, которые в природном естественном Fel d1 соответствуют цепям 2 и 1, соответственно.
В соответствии с одним вариантом осуществления настоящее изобретение относится к полностью человеческому моноклональному антителу или его антигенсвязывающий фрагменту, которые связываются с цепью 1 и/или цепью 2 Fel d1, причем у антитела или его фрагмента наблюдают одну или несколько из следующих характеристик: (i) содержит HCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 2, 18, 34, 50, 66, 82, 98, 114, 130, 146, 162, 178, 194, 210, 226, 242, 258, 274, 290, 306, 322, 347, 338, 354 и 370, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (ii) содержит LCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 10, 26, 42, 58, 74, 90, 106, 122, 138, 154, 170, 186, 202, 218, 234, 250, 266, 282, 298, 314, 330, 347, 346, 362 и 378, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (ii) содержит домен HCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 8, 24, 40, 56, 72, 88, 104, 120, 136, 152, 168, 184, 200, 216, 232 248, 264, 280, 296, 312, 328, 344, 347, 346, 360 и 376, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен LCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 16, 32, 48, 64, 80, 96, 112, 128, 144, 160, 176, 192, 208, 224, 240, 256, 272, 288, 304, 320, 336, 347, 352, 368 и 384, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (iv) содержит домен HCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 4, 20, 36, 52, 68, 84, 100, 116, 132, 148, 164, 180, 196, 212, 228, 244, 260, 276, 292, 308, 324, 347, 340, 356 и 372, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; домен HCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 6, 22, 38, 54, 70, 86, 102, 118, 134, 150, 166, 182, 198, 214, 230, 246, 262, 278, 294, 310, 326, 347, 342, 358 и 374, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; домен LCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 12, 28, 44, 60, 76, 92, 108, 124, 140, 156, 172, 188, 204, 220, 236, 252, 268, 284, 300, 316, 332, 347, 348, 364 и 380, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен LCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 14, 30, 46, 62, 78, 94, 110, 126, 142, 158, 174, 190, 206, 222, 238, 254, 270, 286, 302, 318, 334, 350, 366 и 382, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (v) связывается с цепью 1 и/или цепью 2 Fel d1 с KD, равной или меньше 10-9; (vi) не вступает в перекрестную реакцию с утероглобин или не связывается с ним; или (vii) блокирует проникновение красителя из сосудов in vivo в мышиной модели пассивной кожной анафилаксии (PCA) при помощи мышиного IgE, специфичного к Fel d1.
В соответствии с одним вариантом осуществления настоящее изобретение относится к применению комбинации из двух или более полностью человеческих антител по настоящему изобретению, или их фрагментов, для получения композиции, причем антитела связываются с цепью 1 и/или цепью 2 Fel d1, и причем у каждого содержащегося в композиции антитела или его фрагмента наблюдают одну или несколько из следующих характеристик: (i) содержит HCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 2, 18, 34, 50, 66, 82, 98, 114, 130, 146, 162, 178, 194, 210, 226, 242, 258, 274, 290, 306, 322, 347, 338, 354 и 370, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (ii) содержит LCVR с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 10, 26, 42, 58, 74, 90, 106, 122, 138, 154, 170, 186, 202, 218, 234, 250, 266, 282, 298, 314, 330, 347, 346, 362 и 378, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (ii) содержит домен HCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 8, 24, 40, 56, 72, 88, 104, 120, 136, 152, 168, 184, 200, 216, 232 248, 264, 280, 296, 312, 328, 344, 347, 346, 360 и 376, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен LCDR3 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 16, 32, 48, 64, 80, 96, 112, 128, 144, 160, 176, 192, 208, 224, 240, 256, 272, 288, 304, 320, 336, 347, 352, 368 и 384, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (iv) содержит домен HCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 4, 20, 36, 52, 68, 84, 100, 116, 132, 148, 164, 180, 196, 212, 228, 244, 260, 276, 292, 308, 324, 347, 340, 356 и 372, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; домен HCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 6, 22, 38, 54, 70, 86, 102, 118, 134, 150, 166, 182, 198, 214, 230, 246, 262, 278, 294, 310, 326, 347, 342, 358 и 374, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; домен LCDR1 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 12, 28, 44, 60, 76, 92, 108, 124, 140, 156, 172, 188, 204, 220, 236, 252, 268, 284, 300, 316, 332, 347, 348, 364 и 380, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; и домен LCDR2 с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 14, 30, 46, 62, 78, 94, 110, 126, 142, 158, 174, 190, 206, 222, 238, 254, 270, 286, 302, 318, 334, 350, 366 и 382, или практически сходной с ней последовательностью, имеющей по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности; (v) связывается с цепью 1 и/или цепью 2 Fel d1 с KD, равной или меньше 10-9; (vi) не вступает в перекрестную реакцию с утероглобин или не связывается с ним; или (vii) блокирует проникновение красителя из сосудов in vivo в мышиной модели пассивной кожной анафилаксии (РСА) при помощи мышиного IgE, специфичного к Fel d1; или (viii) при комбинировании со вторым антителом или его антигенсвязывающим фрагментом по настоящему изобретению понижает частоту секретирующих слизь клеток в легких животных, которым была введена провокационная доза Fel d1.
Некоторые антитела к Fel d1 по настоящему изобретению при применении отдельно или в комбинации способны связывать и нейтрализовать по меньшей мере один биологический эффект Fel d1, согласно результатам анализов in vitro или in vivo. Описанную в настоящем документе способность антител по настоящему изобретению связывать и нейтрализовать активность Fel d1 можно измерить при помощи любого стандартного способа, известного специалистам в настоящей области техники, включая анализы связывания или анализы нейтрализации активности (например, защита от анафилаксии).
Неограничивающие иллюстративные анализы измерения in vitro связывающей активности проиллюстрированы в настоящем документе в примере 4. В примере 4 связывающие аффинности и константы скорости реакции человеческих антител к Fel d1 определяли с помощью поверхностного плазмонного резонанса, и измерения осуществляли на приборе Т200 Biacore.
Белки или пептиды Fel d1 можно модифицировать с включением добавлений или замен определенных остатков для введения метки или с целью конъюгации с молекулами-носителями, такими как KLH. Например, цистеин можно добавить либо в N-, либо в С-конец пептида, или можно добавить линкерную последовательность с получением пептида для конъюгации, например, с KLH для иммунизации. Антитела, специфичные к Fel d1, могут не содержать дополнительные метки или фрагменты, или они могут содержать N-концевую или C-концевую метку или фрагмент. В соответствии с одним вариантом осуществления меткой или фрагментом является биотин. При анализе связывания по расположению метки (при наличии) можно определить ориентацию пептида относительно поверхности, с которой связан пептид. Например, если поверхность покрыта авидином, то содержащий N-концевой биотин пептид будет ориентирован так, что C-концевая часть будет удаленной по отношению к поверхности.
Картирование эпитопов и родственные методики
Применяемый в настоящем описании термин "эпитоп" относится к антигенной детерминанте, которая взаимодействует со специфичным антигенсвязывающим сайтом в вариабельном участке молекулы антитела, известном как паратоп. Один антиген может иметь более одного эпитопа. Таким образом, различные антитела могут связываться с различными областями на антигене и могут оказывать различные биологические эффекты. Эпитопы могут быть либо конформационными, либо линейными. Конформационный эпитоп получают посредством пространственно соединенных аминокислот из различных сегментов линейной полипептидной цепи. Линейный эпитоп является эпитопом, получаемым посредством смежных аминокислотных остатков в полипептидной цепи. В определенных условиях эпитоп может включать фрагменты сахаридов, фосфорильных групп или сульфонильных групп на антигене.
Настоящее изобретение относится к антителам к Fel d1, которые взаимодействуют с одной или несколькими аминокислотами, находящимися в одном или нескольких участках цепи 1 или цепи 2 молекулы Fel d1, включая, например, цепь 1 (цепь А), которая показана в SEQ ID NO: 392, или цепь 2 (цепь В), которая показана в SEQ ID NO: 393, или в сравниваемых участках рекомбинантно полученного белка Fel d1, которые показаны в любой из SEQ ID NO: 385, 394, 395, 396 или 397. Эпитоп, с которым связываются антитела, может состоять из отдельной непрерывной последовательности из 3 или более (например, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более) аминокислот, расположенных в любых вышеупомянутых участках или сегментах молекулы Fel d1 (например, линейный эпитоп либо в цепи 1 или в цепи 2, либо в участке, который охватывает как цепь 1, так и цепь 2). Альтернативно, эпитоп может состоять из множества несмежных аминокислот (или аминокислотных последовательностей), расположенных в любом или обоих из вышеупомянутых участках или сегментах молекулы Fel d1 (например, конформационный эпитоп).
Для определения того, "взаимодействует ли антитело с одной или несколькими аминокислотами" в полипептиде или белке, можно применять различные методики, известные рядовым специалистам в настоящей области. Иллюстративные методики включают, например, стандартные эпитоп-перекрестные конкурентные анализы, такие как описанные в работе Antibodies, Harlow and Lane (Cold Spring Harbor Press, Cold Spring Harb., NY). Другие способы включают анализ при помощи сканирующего аланином мутагенеза, пептидный блот-анализ (Reineke (2004) Methods Mol Biol 248:443-63), анализ расщепления пептидов, кристаллографические исследования и NMR-анализ. Кроме того, можно использовать такие способы, как вырезание эпитопов, экстракция эпитопов и химическая модификация антигенов (Tomer (2000) Protein Science 9: 487-496). Другим способом, который можно применять для определения аминокислот в полипептиде, с которым взаимодействует антитело, является водородно/дейтериевый обмен, определяемый посредством масс-спектрометрии. В общих чертах, способ водород/дейтериевого обмена предусматривает мечение дейтерием представляющего интерес белка с последующим связыванием антитела с помеченным дейтерием белком. Затем, комплекс белок/антитело переносят в вводу и способные к обмену протоны в аминокислотах, которые защищены комплексом с антителом, подвергаются обратному обмену дейтерия-на-водород с более медленной скоростью, чем способные к обмену протоны в аминокислотах, которые не являются частью границы раздела. В результате, аминокислоты, которые формируют часть границы раздела белок/антитело, могут удерживать дейтерий и, таким образом, характеризуются относительно более высокой массой по сравнению с аминокислотами, не включенными в границу раздела. После диссоциации антитела белок-мишень подвергают протеазному расщеплению и масс-спектрометрическому анализу, таким образом выявляя меченные дейтерием остатки, которые соответствуют конкретным аминокислотам, с которыми взаимодействует антитело. См., например, Ehring (1999) Analytical Biochemistry 267(2):252-259; Engen and Smith (2001) Anal. Chem. 73:256A-265А. Для картирования эпитопа также можно применять рентгенокристаллографию комплекса антиген/антитело.
Анализ антител на основе модификаций (MAP), также известный как анализ антител на основе структуры антигена (ASAP), является способом, с помощью которого классифицируют большие количества моноклональных антител (mAb) к одному антигену в соответствии со схожестью профиля связывания у каждого антитела с поверхностями химически или ферментативно модифицированных антигенов (US 2004/0101920). Каждая категория может отражать уникальный эпитоп, либо явно отличающийся от эпитопа, представленного в другой категории, либо практически совпадающий с ним. Такая методика позволяет быстро отфильтровывать генетически идентичные антитела так, чтобы характеристику можно было сфокусировать на генетически отличных антителах. Применительно к скринингу гибридом MAP может облегчить определение редких гибридомных клонов, которые продуцируют mAb с необходимыми характеристиками. MAP можно применять для разделения антител по настоящему изобретению на группы антител, связывающих различные эпитопы.
В соответствии с некоторыми вариантами осуществления антитела к Fel d1 или их антигенсвязывающие фрагменты связывают эпитоп в любом одном или нескольких участках, представленных в цепи 1 или цепи 2 Fel d1, либо в природной форме, которая проиллюстрирована в SEQ ID NO: 392 (цепь 1) и SEQ ID NO: 393 (цепь 2), либо полученных рекомбинантно, которые проиллюстрированы в любой из SEQ ID NO: 385, 394, 395, 396 и 397, или их фрагмент. В соответствии с некоторыми вариантами осуществления антитела по настоящему изобретению, которые показаны в таблице 1, взаимодействуют по меньшей мере с одной аминокислотной последовательностью, выбранной из группы, состоящей из аминокислотных остатков, находящихся в диапазоне от приблизительно положения 15 до приблизительно положения 24 в SEQ ID NO: 396, аминокислотных остатков, находящихся в диапазоне от приблизительно положения 85 до приблизительно в положении 103 в SEQ ID NO: 396, аминокислотных остатков, находящихся в диапазоне от приблизительно положения 85 до приблизительно в положении 104 в SEQ ID NO: 396, аминокислотных остатков, находящихся в диапазоне от приблизительно положения 113 до приблизительно в положении 116 в SEQ ID NO: 396. Такие участки дополнительно проиллюстрированы в SEQ ID NO: 402, 403, 404, 412 и 426.
Настоящее изобретение также относится к антителам к Fel d1, которые связываются с тем же эпитопом, или частью эпитопа, что и любое из конкретных иллюстрированных антител, описанных в настоящем документе в таблице 1, или антитело с последовательностями CDR из любых иллюстративных антител, описанных в таблице 1. Аналогичным образом, настоящее изобретение также относится к антителам к Fel d1, которые конкурируют за связывание с Fel d1 или с фрагментом Fel d1 с любым из конкретных иллюстрированных антител, описанных в настоящем документе в таблице 1, или антителом с последовательностями CDR из любых иллюстративных антител, описанных в таблице 1.
С помощью стандартных известных из уровня техники способов можно легко определить, связывается ли антитело с тем же эпитопом или конкурирует за связывание с эталонным антителом к Fel d1. Например, для определения того, связывается ли тестовое антитело с тем же эпитопом, что и эталонное антитело к Fel d1 по настоящему изобретению, эталонному антителу позволяют связаться с белком или пептидом Fel d1 в насыщающих условиях. Затем оценивают способность тестового антитела связываться с молекулой Fel d1. Если тестовое антитело способно связываются с Fel d1 после насыщающего связывания с эталонным антителом к Fel d1, то можно прийти к заключению, что тестовое антитело связывается с эпитопом, отличным от эталонного антитела к Fel d1. С другой стороны, если тестовое антитело не способно связываться с молекулой Fel d1 после насыщающего связывания с эталонным антителом к Fel d1, то тестовое антитело может связываться с тем же эпитопом, что и эпитоп, связываемый посредством эталонного антитела к Fel d1 по настоящему изобретению.
Для определения того, конкурирует ли антитело за связывание с эталонным антителом к Fel d1, осуществляют описанные выше методики связывания в двух адаптациях. Согласно первой адаптации эталонному антителу позволяют связаться с молекулой Fel d1 в насыщающих условиях с последующей оценкой связывания тестового антитела с молекулой Fel d1. Согласно второй адаптации тестовому антителу позволяют связаться с молекулой Fel d1 в насыщающих условиях с последующей оценкой связывания эталонного антитела с молекулой Fel d1. Если в обоих адаптациях только первое (насыщающее) антитело способно связываться с молекулой Fel d1, то делают вывод, что тестовое антитело и эталонное антитело конкурируют за связывание Fel d1. Рядовому специалисту в настоящей области техники будет понятно, что антитело, которое конкурирует за связывание с эталонным антителом, не обязательно может связываться с идентичным эпитопом, что и эталонное антитело, а может стерически блокировать связывание эталонного антитела посредством связывания совпадающего или смежного эпитопа.
Два антитела связываются с одинаковым или перекрывающимся эпитопом, если каждое конкурентно ингибирует (блокирует) связывание другого с антигеном. А именно, 1-, 5-, 10-, 20- или 100-кратный избыток одного антитела ингибирует связывание другого по меньшей мере на 50%, но предпочтительно на 75%, 90% или даже 99%, согласно результатам измерения в анализе конкурентного связывания (см., например, Junghans et al., Cancer Res. 1990 50:1495-1502). Альтернативно, два антитела характеризуются одинаковыми эпитопами, если фактически все аминокислотные мутации в антигене, которые уменьшают или исключают связывание одного антитела, уменьшают или исключают связывание другого. Два антитела характеризуются перекрывающимися эпитопами, если несколько аминокислотных мутаций, которые уменьшают или исключают связывание одного антитела, уменьшают или исключают связывание другого.
Затем, можно осуществить дополнительный стандартный эксперимент (например, анализы на мутации и связывание пептида) для подтверждения того, является ли наблюдаемое отсутствие связывания тестового антитела фактическим следствием связывания с тем же эпитопом, что и эталонное антитело, или является ли стерическое блокирование (или другое явление) ответственным за отсутствие наблюдаемого связывания. Эксперименты такого рода можно осуществить при помощи ELISA, RIA, поверхностного плазмонного резонанса, проточной цитометрии или любого другого количественного или качественного анализа связывания антитела, который доступен в настоящей области техники.
Иммуноконъюгаты
Настоящее изобретение относится к человеческому моноклональному антителу к Fel d1, конъюгированному с терапевтическим фрагментом ("иммуноконъюгат"), таким как антиген, который способен уменьшить тяжесть аллергической реакции на антиген Fel d1, присутствующий в кошачьей перхоти или на кошках, или в зоне окружающей среды, где могут проживать кошки, или ослабить по меньшей мере один симптом, ассоциированный с контактом с кошками, кошачьей перхотью или с аллергеном Fel d1, включая ринит, конъюнктивит или затруднения дыхания, или их тяжесть. Такое средство может представлять собой кортикостероид, второе отличное антитело к Fel d1 или вакцину. Тип терапевтического фрагмента, который может быть конъюгирован с антителом к Fel d1, будет принимать в учет подлежащее лечению состояние и подлежащий достижению необходимый терапевтический эффект. Альтернативно, если необходимый терапевтический эффект предназначен для лечения последствий или симптомов, ассоциированных с контактом с аллергеном Fel d1 или любым другим состоянием, которое является результатом такого контакта, таких как без ограничения риниты или конъюнктивит, то может быть предпочтительно конъюгирование средства, подходящего для лечения последствий или симптомов состояния или для ослабления любых побочных эффектов антител по настоящему изобретению. Примеры подходящих средств для формирования иммуноконъюгатов известны в настоящей области техники, см., например, WO 05/103081.
Мультиспецифические антитела
Антитела по настоящему изобретению могут быть моноспецифическими, биспецифическими или мультиспецифическими. Мультиспецифические антитела могут быть специфичными к различным эпитопам одного целевого полипептида или могут содержать антигенсвязывающие домены, специфичные к нескольким целевым полипептидам. См., например, Tutt et al., 1991, J. Immunol. 147:60-69; Kufer et al., 2004, Trends Biotechnol. 22:238-244. Антитела по настоящему изобретению могут быть связаны или совместно экспрессироваться с другой функциональной молекулой, например, другим пептидом или белком. Например, антитело или его фрагмент могут быть функционально связаны (например, посредством образования химической связи, генетической гибридизацией, нековалентным связыванием или иначе) с одной или несколькими другими молекулярными структурами, такими как другое антитело или фрагмент антитела, с получением биспецифического или мультиспецифического антитела со второй специфичностью связывания. Например, настоящее изобретение относится к биспецифическим антителам, причем одно плечо иммуноглобулина может быть специфичным к цепи 1 Fel d1 или ее фрагменту, а другое плечо иммуноглобулина может быть специфичным к цепи 2 Fel d1 или второй терапевтической мишени, или может быть конъюгировано с терапевтическим фрагментом.
Согласно определенным иллюстративным вариантам осуществления настоящее изобретение относится к биспецифической антигенсвязывающей молекуле, которая представляет собой биспецифическое антитело. Каждый антигенсвязывающий домен биспецифического антитела содержит вариабельный домен тяжелой цепи (HCVR) и вариабельный домен легкой цепи (LCVR). HCVR также может называться участком VH, а LCVR может называться участком VL. Обычно, каждый HCVR и LCVR содержит три CDR, чередующиеся с четырьмя FR, располагающиеся от амино-конца до карбокси-конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Три CDR в HCVR в настоящем описании могут называться HCDR1, HCDR2 и HCDR3; в то время как три CDR в LCVR в настоящем описании могут называться LCDR1, LCDR2 и LCDR3.
В биспецифических антигенсвязывающих молекулах по настоящему изобретению каждый антигенсвязывающий домен может включать целую молекулу антитела или антигенсвязывающий фрагмент антитела или может состоять из них. Применяемые в настоящем документе термины "антигенсвязывающая часть" антитела, "антигенсвязывающий фрагмент" антитела и т.п. включают любой встречающийся в природе, получаемый ферментативно, синтетический или генетически сконструированный полипептид или гликопротеин, который специфично связывается с антигеном с формированием комплекса. Антигенсвязывающие фрагменты антитела могут быть получены, например, от целых молекул антител с применением любых подходящих стандартных методик, таких как протеолитическое расщепление или рекомбинантные методики генетической инженерии, предусматривающие манипуляцию и экспрессию ДНК, кодирующей вариабельные и необязательно константные домены антитела. Такая ДНК известна и/или легко доступна из, например, коммерческих источников, библиотек ДНК (включая, например, библиотеки фаг-антитело), или ее можно синтезировать. ДНК можно секвенировать и с ней можно проводить манипуляции химическими способами или при помощи методик молекулярной биологии, например, для организации одного или нескольких вариабельных и/или константных доменов в подходящую конфигурацию или введения кодонов, создания цистеиновых остатков, модификации, добавления или удаления аминокислот и т.д.
Неограничивающие примеры антигенсвязывающих фрагментов, которые могут быть включены в биспецифические антигенсвязывающие молекулы по настоящему изобретению, включают: (i) фрагменты Fab; (ii) фрагменты F(ab')2; (iii) фрагменты Fd; (iv) фрагменты Fv; (v) одноцепочечные молекулы Fv (scFv); (vi) фрагменты dAb и (vii) минимальные единицы распознания, состоящие из аминокислотных остатков, которые имитируют гипервариабельный участок антитела. Также применяемым в настоящем документе выражением "антигенсвязывающий фрагмент" охвачены другие сконструированные молекулы, такие как домен-специфичные антитела, антитело с одним доменом, антитела с удаленным доменом, химерные антитела, антитела с пришитым CDR, диатела, триотела, тетратела, миниантитела, нанотела (например, моновалентные нанотела, бивалентные нанотела и т.д.), иммунопрепараты на основе модульного белка с малым размером молекул (SMIP) и вариабельные домены IgNAR акулы.
Антигенсвязывающий фрагмент антитела, как правило, будет содержать по меньшей мере один вариабельный домен. Вариабельный домен может иметь любой размер или аминокислотный состав и, как правило, будет содержать по меньшей мере один CDR, который является смежным с одной или несколькими каркасными последовательностями или находится в их пределах. В антигенсвязывающих фрагментах, имеющих домен VH, связанный с доменом VL, домены VH и VL могут быть расположены относительно друг друга в любом подходящем порядке. Например, вариабельный участок может быть димерным и содержать димеры VH-VH, VH-VL или VL-VL. Альтернативно, антигенсвязывающий фрагмент антитела может содержать мономерный домен VH или VL.
В соответствии с определенными вариантами осуществления антигенсвязывающий фрагмент биспецифической антигенсвязывающей молекулы может содержать по меньшей мере один вариабельный домен, ковалентно связанный по меньшей мере с одним константным доменом. Неограничивающие иллюстративные конфигурации вариабельных и константных доменов, которые можно обнаружить в антигенсвязывающем домене биспецифической антигенсвязывающей молекулы, могут включать: (i) VH-CH1; (ii) VH-CH2; (iii) VH-CH3; (iv) VH-CH1-CH2; (v) VH-CH1-CH2-CH3; (vi) VH-CH2-CH3; (vii) VH-CL; (viii) VL-CH1; (ix) VL-CH2; (x) VL-CH3; (xi) VL-CH1-CH2; (xii) VL-CH1-CH2-CH3; (xiii) VL-CH2-CH3; и (xiv) VL-CL. В любой конфигурации вариабельных и константных доменов, включая любые из перечисленных выше иллюстративных конфигураций, вариабельные и константные домены могут быть либо непосредственно соединены друг с другом, либо могут быть соединены при помощи полного или частичного шарнирного или линкерного участка. Шарнирный участок может состоять по меньшей мере из 2 (например, 5, 10, 15, 20, 40, 60 или более) аминокислот, что в результате дает гибкую или полугибкую связь между смежными вариабельными и/или константными доменами в отдельной полипептидной молекуле. Кроме того, антигенсвязывающий домен биспецифической антигенсвязывающей молекулы может содержать гомодимер или гетеродимер (или другой мультимер) в любой из перечисленных выше конфигураций вариабельных и константных доменов в нековалентной связи друг с другом и/или с одним или несколькими мономерными доменами VH или VL (например, посредством дисульфидной связи(связей)).
Первый антигенсвязывающий домен и второй антигенсвязывающий домен могут непосредственно или опосредованно быть соединены друг с другом с формированием биспецифической антигенсвязывающей молекулы. Альтернативно, каждый из первого антигенсвязывающего домена и второго антигенсвязывающего домена может быть соединен с отдельным мультимеризирующим доменом. Связь одного мультимеризирующего домена с другим мультимеризирующим доменом способствует образованию связи между двумя антигенсвязывающими доменами, таким образом формируя биспецифическую антигенсвязывающую молекулу. Применяемый в настоящем документе термин "мультимеризирующий домен" относится к любой молекуле, белку, полипептиду, пептиду или аминокислоте, которые могут связываться со вторым мультимеризирующим доменом такой же или сходной структуры или строения. Например, мультимеризирующий домен может представлять собой полипептид, содержащий домен CH3 иммуноглобулина. Неограничивающим примером мультимеризирующего компонента является Fc часть иммуноглобулина, например, Fc домен IgG, выбранного из изотипов IgG1, IgG2, IgG3 и IgG4, а также любого аллотипа в каждой группе изотипов. В соответствии с некоторыми вариантами осуществления мультимеризирующий домен может представлять собой Fc фрагмент или аминокислотную последовательность от 1 до приблизительно 200 аминокислот в длину, содержащую по меньшей мере один цистеиновый остаток. Согласно другим вариантам осуществления мультимеризирующий домен может представлять собой цистеиновый остаток или короткий, содержащий цистеин пептид. Другие мультимелизирующие домены включают пептиды или полипептиды, содержащие лейциновую застежку, мотив спираль-петля или заспирализованный мотив или состоящие из них.
Любой формат или методику биспецифического антитела можно применять для получения биспецифических антигенсвязывающих молекул по настоящему изобретению.
Например, антитело или его фрагмент, характеризующийся первой антигенсвязывающей специфичностью, можно функционально связать (например, посредством образования химической связи, генетической гибридизации, нековалентным связыванием или иначе) с одной или несколькими другими молекулярными структурами, такими как другое антитело или фрагмент антитела, характеризующиеся второй антигенсвязывающей специфичностью, с получением биспецифической антигенсвязывающей молекулы.
Иллюстративный формат биспецифического антитела, который можно применять в контексте настоящего изобретения, включает применение первого домена CH3 иммуноглобулина (Ig) и второго домена CH3 Ig, причем первый и второй домены CH3 Ig отличаются друг от друга по меньшей мере по одной аминокислоте, и причем по меньшей мере одно аминокислотное отличие уменьшает связывание биспецифического антитела с белком А по сравнению с биспецифическим антителом без аминокислотного отличия. В соответствии с одним вариантом осуществления первый домен CH3 Ig связывается с белком А, а второй домен CH3 Ig содержит мутацию, которая уменьшает или делает невозможным связывание белка А, такую как модификация H95R (согласно нумерации экзонов IMGT; H435R согласно нумерации EU). Второй CH3 может дополнительно содержать модификацию Y96F (согласно IMGT; Y436F согласно EU). Дополнительные модификации, которые можно обнаружить во втором CH3, включают: D16E, L18M, N44S, K52N, V57M и V82I (согласно IMGT; D356E, L358M, N384S, K392N, V397M и V422I согласно EU) в случае антител IgG1; N44S, K52N и V82I (IMGT; N384S, K392N и V422I согласно EU) в случае антител IgG2; и Q15R, N44S, K52N, V57M, R69K, E79Q и V82I (согласно IMGT; Q355R, N384S, K392N, V397M, R409K, E419Q и V422I согласно EU) в случае антител IgG4. Описанные выше варианты формата биспецифических антител предусмотрены объемом настоящего изобретения.
Другие иллюстративные биспецифические форматы, которые можно применять в контексте настоящего изобретения, включают без ограничения, например, биспецифические форматы на основе scFv или диатела, гибриды IgG-scFv, Ig с двойным вариабельным доменом (DVD), квадрогибридому, выступы-во-впадины, общую легкую цепь (например, общая легкая цепь с выступами-во-впадины и т.д.), CrossMab, CrossFab, (SEED)тело, лейциновую застежку, Duobody, IgG1/IgG2, IgG с Fab двойного действия (DAF) и биспецифические форматы Mab2 (см., например, Klein et al. 2012, mAbs 4:6, 1-11, и материалы, упомянутые в ней, для обзора упомянутых выше форматов). Биспецифические антитела также можно построить при помощи, например, конъюгации пептида/нуклеиновой кислоты, причем не встречающиеся в природе аминокислоты с ортогональной химической реакционной способностью применяют для получения конъюгатов сайт-специфического антитела и олигонуклеотида, которые затем самостоятельно собираются в мультимерные комплексы с определенным составом, валентностью и геометрией. (См., например, Kazane et al., J. Am. Chem. Soc. [Epub: Dec. 4, 2012]).
Терапевтическое введение и композиции
Настоящее изобретение относится к терапевтическим композициям, содержащим антитела к Fel d1 или их антигенсвязывающие фрагменты по настоящему изобретению. Введение терапевтических композиций в соответствии с настоящим изобретением будут осуществлять через подходящий путь, включая без ограничения внутривенно, подкожно, внутримышечно, с подходящими носителями, наполнителями и другими средствами, которые включены в лекарственные формы для обеспечения улучшенного переноса, доставки, стойкости и т.п. Большое количество соответствующих лекарственных форм можно найти в известном всем фармакологам фармакологическом справочнике: Remington's Pharmaceutical Sciences, Mack Publishing Company, Easton, PA. Такие лекарственные формы включают, например, порошки, пасты, мази, желе, воски, масла, липиды, содержащие липиды (катионные или анионные) везикулы (такие как LIPOFECTIN™), ДНК-конъюгаты, безводные абсорбционные пасты, эмульсии по типу масло-в-воде и вода-в-масле, эмульсии карбовакса (полиэтиленгликоли с различными молекулярными массами), полужидкие гели и полужидкие смеси, содержащие карбовакс. См. также Powell et al. "Compendium of excipients for parenteral formulations" PDA (1998) J Pharm Sci Technol 52:238-311.
Доза антитела может варьировать в зависимости от возраста и размера субъекта, которому будут ее вводить, целевого заболевания, условий, пути введения и т.п. При применении антитела по настоящему изобретению для лечения ринита или конъюнктивита, ассоциированного с контактом с кошкой или с кошачьей перхотью, у индивидуума, имеющего чувствительность к Fel d1, или для предупреждения анафилактической реакции на кошачий аллерген, или для ослабления тяжести аллергической реакции, предпочтительно внутривенное введение антитела по настоящему изобретению в норме за одну дозу, составляющую от приблизительно 0,01 до приблизительно 30 мг/кг массы тела, более предпочтительно от приблизительно 0,02 до приблизительно 7, от приблизительно 0,03 до приблизительно 5 или от приблизительно 0,05 до приблизительно 3 мг/кг массы тела. В зависимости от тяжести состояния частоту введения и длительность лечения можно откорректировать. В соответствии с определенными вариантами осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению можно вводить в качестве изначальной дозы, составляющей по меньшей мере от приблизительно 0,1 мг до приблизительно 800 мг, от приблизительно 1 до приблизительно 500 мг, от приблизительно 5 до приблизительно 300 мг, или от приблизительно 10 до приблизительно 200 мг, до приблизительно 100 мг или до приблизительно 50 мг. В соответствии с определенными вариантами осуществления после изначальной дозы может следовать введение второй или нескольких последующих доз антитела или его антигенсвязывающего фрагмента в количестве, которое может быть примерно таким же или меньше, чем в изначальной дозе, причем последующие дозы разделены по меньшей мере 1-3 днями, по меньшей мере одной неделей, по меньшей мере 2 неделями, по меньшей мере 3 неделями, по меньшей мере 4 неделями, по меньшей мере 5 неделями, по меньшей мере 6 неделями, по меньшей мере 7 неделями, по меньшей мере 8 неделями, по меньшей мере 9 неделями, по меньшей мере 10 неделями, по меньшей мере 12 неделями или по меньшей мере 14 неделями.
Известны различные системы доставки, и их можно применять для введения фармацевтической композиции по настоящему изобретению, например, инкапсуляция в липосомах, микрочастицы, микрокапсулы, рекомбинантные клетки, способные экспрессировать мутантные вирусы, опосредованный рецепторами эндоцитоз (см., например, Wu et al. (1987) J. Biol. Chem. 262:4429-4432). Способы введения включают без ограничения внутрикожный, чрезкожный, внутримышечный, внутрибрюшинный, внутривенный, подкожный, интраназальный, эпидуральный и пероральный пути. Композицию можно вводить любым подходящим путем, например, при помощи инфузии или инъекции ударной дозы вещества, посредством всасывания через эпителиальные или слизистые оболочки (например, слизистую оболочку полости рта, слизистую оболочку прямой кишки или тонкого кишечника и т.д.), и можно вводить вместе с другими биологически активными средствами. Введение может быть системным или локальным.
Фармацевтическую композицию также можно доставлять в везикуле, в частности, липосоме (см., например, Langer (1990) Science 249:1527-1533).
В определенных ситуациях фармацевтическую композицию можно доставлять в системе с контролируемым высвобождением. В соответствии с одним вариантом осуществления можно применять помпу. Согласно другому варианту осуществления можно применять полимерные материалы. В соответствии с еще одним вариантом осуществления систему с контролируемым высвобождением можно поместить вблизи мишени композиции, таким образом, будет необходима лишь часть системной дозы.
Инъекционные формы могут включать готовые лекарственные формы для внутривенных, подкожных, интракутанных и внутримышечных инъекций, капельных внутривенных вливаний и т.д. Такие инъекционные формы можно получить посредством общеизвестных способов. Например, инъекционные формы можно получить, например, растворением, суспендированием или эмульгированием описанных выше антитела или его соли в стерильной водной среде или масляной среде, обычно применяемой для инъекций. Водной средой для инъекций являются, например, физиологический раствор, изотонический раствор, содержащий глюкозу и другие вспомогательные средства и т.д., которые можно применять в комбинации с соответствующим солюбилизирующим средством, таким как спирт (например, этанол), многоатомный спирт (например, пропиленгликоль, полиэтиленгликоль), неионное поверхностно-активное вещество [например, полисорбат 80, НСО-50 (аддукт полиоксиэтилена (50 моль) и гидрогенизованного касторового масла)] и т.д. Масляными средами являются, например, сезамовое масло, соевое масло и т.д., которые можно применять в комбинации с солюбилизирующим средством, таким как бензилбензоат, бензиловый спирт и т.п.Полученный таким образом инъекционный препарат предпочтительно заполняют в подходящую ампулу.
Фармацевтическую композицию по настоящему изобретению можно доставлять подкожно или внутривенно при помощи стандартной иглы и шпица. Кроме того, что касается подкожной доставки, то при доставке фармацевтической композиции по настоящему изобретению с легкостью можно применять устройство для доставки по типу шприц-ручка. Такое устройство для доставки по типу шприц-ручка может быть многоразовым или одноразовым. В многоразовом устройстве для доставки по типу шприц-ручка обычно используют сменный картридж, который содержит фармацевтическую композицию. После того, как вся фармацевтическая композиция из картриджа была введена и картридж опустел, пустой картридж можно легко выбросить и поместить новый картридж, который содержит фармацевтическую композицию. Устройство для доставки по типу шприц-ручка затем можно повторно использовать. В случае одноразового устройства для доставки сменный картридж отсутствует. Вернее, одноразовое устройство для доставки по типу шприц-ручка заполнено фармацевтической композицией, удерживаемой в резервуаре устройства. После удаления из резервуара фармацевтической композиции выбрасывают полностью все устройство.
Для подкожной доставки фармацевтической композиции по настоящему изобретению применяют разнообразные многоразовые шприц-ручки и автоинжекторные устройства для доставки. Примеры включают без ограничения AUTOPEN™ (Owen Mumford, Inc., Вудсток, Великобритания), шприц-ручку DISETRONIC™ (Disetronic Medical Systems, Бургдорф, Швейцария), шприц-ручку HUMALOG MIX 75/25™, шприц-ручку HUMALOG™, шприц-ручку HUMALIN 70/30™ (Eli Lilly and Co., Индианаполис, Индиана), NOVOPEN™ I, II и III (Novo Nordisk, Копенгаген, Дания), NOVOPEN JUNIOR™ (Novo Nordisk, Копенгаген, Дания), шприц-ручку BD™ (Becton Dickinson, Франклин-Лейке, Нью-Джерси), OPTIPEN™, OPTIPEN PRO™, OPTIPEN STARLET™ и OPTICLIK™ (sanofï-aventis, Франкфурт, Германия), не говоря уже о других. Примеры одноразовых устройств для доставки по типу шприц-ручка, применяемых для подкожной доставки фармацевтической композиции по настоящему изобретению, включают без ограничения шприц-ручку SOLOSTAR™ (sanofi-aventis), FLEXPEN™ (Novo Nordisk) и KWIKPEN™ (Eli Lilly), SURECLICK™ Autoinjector (Amgen, Таузенд-Оукс, Калифорния), PENLET™ (Haselmeier, Штутгарт, Германия), EPIPEN (Dey, L.P.) и шприц-ручку HUMIRA™ (Abbott Labs, Abbott Park, Иллинойс), не говоря уже о других.
Предпочтительно, описанные выше фармацевтические композиции для перорального или парентерального применения получают в готовых лекарственных формах в стандартной дозе, подобранной в соответствии с дозой активных ингредиентов. Такие готовые лекарственные формы в стандартной дозе включают, например, таблетки, пилюли, капсулы, инъекции (ампулы), суппозитории и т.п. Количество вышеизложенного включенного антитела обычно составляет от приблизительно 5 до приблизительно 500 мг на готовую лекарственную форму в стандартной дозе; особенно в форме инъекции, предпочтительно, чтобы вышеизложенное антитело содержалось в количестве от приблизительно 5 до приблизительно 100 мг и от приблизительно 10 до приблизительно 250 мг на каждую готовую лекарственную форму.
Терапевтические применения антител
В связи с их взаимодействием с Fel d1 настоящие антитела пригодны для лечения первичной реакции после контакта индивидуума с кошкой, кошачьей перхотью или со средой, содержащей белок Fel d1, или по меньшей мере одного симптома, ассоциированного с аллергической реакцией, такой как зуд глаз, конъюнктивит, ринит, бронхолегочная обструкция, затруднение дыхания, или для предупреждения вторичной реакции на аллерген Fel d1, включающей более серьезную анафилактическую реакцию, или для снижения тяжести, продолжительности и/или частоты появления симптомов после повторного контакта с кошачьим аллергеном. Соответственно, предусматривают, что антитела по настоящему изобретению можно применять профилактически или терапевтически.
Согласно следующему варианту осуществления по настоящему изобретению настоящие антитела применяют для получения фармацевтической композиции для лечения больных, страдающих от чувствительности к кошкам, кошачьей перхоти, кошачьей шерсти или их экстракта и/или белка Fel d1. Согласно еще одному варианту осуществления по настоящему изобретению настоящие антитела применяют для получения фармацевтической композиции для уменьшения тяжести первичного контакта с Fel d1 или для уменьшения тяжести, продолжительности и/или числа аллергических реакций на Fel d1. Согласно следующему варианту осуществления по настоящему изобретению настоящие антитела применяют в качестве средства вспомогательной терапии с любым другим средством, пригодным для лечения аллергий на кошек, включая кортикостероиды, вакцины, антигенспецифическую иммунотерапию (SIT) или любое другое паллиативное средство терапии, известное специалистам в настоящей области техники.
Средства комбинированной терапии
Средства комбинированной терапии могут включать антитело к Fel d1 по настоящему изобретению и любое дополнительное терапевтическое средство, которое можно успешно комбинировать с антителом по настоящему изобретению или с биологически активным фрагментом антитела по настоящему изобретению.
Например, для содействия уменьшению аллергических симптомов после контакта с кошкой, кошачьей перхотью, кошачьей шерстью или их экстрактом или Fel d1 или контакта со средой, в которой проживает кошка, можно использовать второе терапевтическое средство, такое как кортикостероид. Антитела также можно применять в сочетании с другими средствами терапии, такими как вакцина, специфичная к аллергену Fel d1. Дополнительный терапевтически активный компонент(ы) можно вводить до, одновременно или после введения антитела к Fel dl по настоящему изобретению. В целях настоящего раскрытия такие схемы введения рассматривают как введение антитела к Fel d1 "в комбинации со" вторым терапевтически активным компонентом.
Схемы введения
В соответствии с определенными вариантами осуществления по настоящему изобретению многократные дозы одного или нескольких антител к Fel d1 (комбинации антител) или биспецифической антигенсвязывающей молекулы можно вводить субъекту через определенный период времени. Способы в соответствии с этим аспектом по настоящему изобретению предусматривают последовательное введение субъекту многократных доз антитела, комбинации антител или биспецифической антигенсвязывающей молекулы по настоящему изобретению. Применяемое в настоящем описании "последовательное введение" означает, что каждую дозу антитела, комбинации антител или биспецифической антигенсвязывающей молекулы вводят субъекту в различные моменты времени, например, в различные дни, разделенные предварительно определенным интервалом (например, часами, днями, неделями или месяцами). Настоящее изобретение относится к способам, которые предусматривают последовательное введение больному однократной изначальной дозы антитела, комбинации антител или биспецифической антигенсвязывающей молекулы с последующим введением одной или нескольких вторичных доз антитела и необязательно с последующей одной или несколькими третичными дозами антитела.
Термины "изначальная доза," "вторичные дозы" и "третичные дозы" относятся к временной последовательности введения антитела, комбинации антител или биспецифической антигенсвязывающей молекулы по настоящему изобретению. Таким образом, "изначальной дозой" является доза, которую вводят в начале схемы лечения (также называемая как "исходная доза"), "вторичные дозы" являются дозами, которые вводят после изначальной дозы, и "третичные дозы" являются дозами, которые вводят после вторичных доз. Все из изначальной, вторичных и третичных доз могут содержать одинаковое количество антитела, комбинации антител или биспецифической антигенсвязывающей молекулы, но, в целом, могут отличаться друг от друга касательно частоты введения. Тем не менее, в соответствии с определенными вариантами осуществления количество антитела, комбинации антител или биспецифической антигенсвязывающей молекулы, содержащееся в изначальной, вторичной и/или третичной дозах, отличается друг от друга (например, повышено или понижено, по необходимости) в течение курса лечения. В соответствии с определенными вариантами осуществления две или более (например, 2, 3, 4 или 5) доз вводят в начале схемы лечения в качестве "ударных доз", за которыми следуют последующие дозы, которые вводят не так часто (например, "поддерживающие дозы").
Согласно одному иллюстративному варианту осуществления по настоящему изобретению каждую вторичную и/или третичную дозу вводят через 1-26 (например, 1, 11/2, 2, 21/2, 3, 31/2, 4, 41/2, 5, 51/2, 6, 61/2, 7, 71/2, 8, 81/2, 9, 91/2, 10, 101/2, 11, 111/2, 12, 121/2, 13, 131/2, 14, 141/2, 15, 151/2, 16, 161/2, 17, 171/2, 18, 181/2, 19, 191/2, 20, 201/2, 21, 211/2, 22, 221/2, 23, 231/2, 24, 241/2, 25, 251/2, 26, 261/2 или более) недель после непосредственно предшествующей дозы. Применяемая в настоящем документе фраза "непосредственно предшествующая доза" означает в последовательности многократных введений дозу антитела, комбинации антител или биспецифической антигенсвязывающей молекулы, которую вводят больному перед введением непосредственно следующей дозы в последовательности при отсутствии промежуточных доз.
Способы в соответствии с этим аспектом по настоящему изобретению могут предусматривать введение больному любого числа вторичных или третичных доз антитела, комбинации антител или биспецифической антигенсвязывающей молекулы, которые специфично связывают Fel d1. Например, в соответствии с определенными вариантами осуществления больному вводят только одну вторичную дозу. Согласно другим вариантам осуществления больному вводят две или более (например, 2, 3, 4, 5, 6, 7, 8 или более) вторичных доз. Аналогичным образом, в соответствии с определенными вариантами осуществления больному вводят только одну третичную дозу. Согласно другим вариантам осуществления больному вводят две или более (например, 2, 3, 4, 5, 6, 7, 8 или более) третичных доз.
Согласно вариантам осуществления, предусматривающим множественные вторичные дозы, каждую вторичную дозу можно вводить с такой же частотой, как и другие вторичные дозы. Например, каждую вторичную дозу можно вводить больному спустя 1-2 недели после непосредственно предшествующей дозы. Аналогичным образом, согласно вариантам осуществления, предусматривающим множественные третичные дозы, каждую третичную дозу можно вводить с такой же частотой, как и другие третичные дозы. Например, каждую третичную дозу можно вводить больному спустя 2-4 недели после непосредственно предшествующей дозы. Альтернативно, частота, с которой вводят больному вторичные и/или третичные дозы, может изменяться в ходе схемы лечения. Частота введения также может корректироваться в течение курса лечения лечащим врачом в зависимости от индивидуальных потребностей больного после клинического обследования.
Диагностические применения антител
Антитела к Fel d1 по настоящему изобретению также можно применять для обнаружения и/или измерения Fel d1 в образце, например, в диагностических целях. Предусмотрено, что подтверждение аллергической реакции, которая, как полагают, обусловлена Fel d1, можно осуществить посредством измерения наличия либо Fel d1 посредством применения любого одного или нескольких из антител по настоящему изобретению. Иллюстративные диагностические анализы на Fel d1 могут предусматривать, например, приведение в контакт образца, полученного от больного, с антителом к Fel d1 по настоящему изобретению, причем антитело к Fel d1 помечено поддающейся обнаружению меткой или репортерной группой, или его применяют в качестве захватывающего лиганда для избирательного выделения белка Fel d1 из образцов больного. Альтернативно, непомеченное антитело к Fel d1 можно применять для диагностических целей в комбинации со вторичным антителом, которое само по себе помечено для возможности обнаружения. Поддающаяся обнаружению метка или репортерная группа могут представлять собой радиоизотоп, такой как3Н,14С,32Р,35S или125I; флуоресцентный или хемилюминесцентный фрагмент, такой как флуоресцеинизотиоцианат или родамин, или фермент, такой как щелочная фосфатаза, β-галактозидаза, пероксидаза хрена или люцифераза. Конкретные иллюстративные анализы, которые можно применять для обнаружения или измерения Fel d1 в образце, включают твердофазный иммуноферментный анализ (ELISA), радиоиммунный анализ (RIA) и сортировку флуоресцентно-активированных клеток (FACS).
Образцы, которые можно применять в диагностических анализах на Fel d1, в соответствии с настоящим изобретением включают любой образец ткани или жидкости, получаемый от больного, который содержит поддающиеся обнаружению количества белка Fel d1 или его фрагментов в нормальных или патологических условиях. Обычно уровни Fel d1 в конкретном образце, полученном от здорового/не аллергика (например, больного, не страдающего от чувствительности, ассоциированной с наличием Fel d1), будут измерять, изначально устанавливая исходный или стандартный уровень Fel d1. Такой базовый уровень Fel d1 затем можно сравнить с уровнями Fel d1, измеренными в образцах, полученных от людей, у которых подозревают наличие чувствительности к Fel d1 в кошачьей перхоти или симптомов, ассоциированных с таким заболеванием или состоянием.
Примеры
Следующие примеры приведены с целью предоставления специалистам в настоящей области техники полного раскрытия и описания того, как осуществлять и применять способы и композиции по настоящему изобретению, и не предназначены для ограничения рассматриваемого авторами объема настоящего изобретения. Были предприняты попытки обеспечить точность в отношении используемых чисел (например, количеств, температуры и т.д.), но следует учитывать некоторые экспериментальные ошибки и отклонения. Если не указано иное, части являются частями по массе, молекулярная масса является средней молекулярной массой, температура приведена в градусах шкалы Цельсия, а давление равно атмосферному или почти атмосферное.
Пример 1: Создание человеческих антител к Fel d1
Для создания антител к Fel d1 можно использовать иммуноген, включающих любой из приведенных далее. В соответствии с некоторыми вариантами осуществления антитела по настоящему изобретению получали от мышей, иммунизированных первичным иммуногеном, таким как полноразмерный природный Fel d1 (nFel d1), который доступен на рынке для приобретения (например, от компании Indoor Biotechnologies, № LTN-FD1-1), или выделить из кошачьей шерсти или перхоти путем многоэтапной колоночной хроматографии (см., например, работу Chapman MD, et al. (1988), J. Immunol. 140:812-818), или который можно получить рекомбинантно (см. номера доступа в GenBank Р30438 или NP_001041618.1 для полноразмерной аминокислотной последовательности цепи 1 для Fel d1 (также называемой цепью А или FELD1 А; см. также SEQ ID NO: 392) и номер доступа в GenBank Р30440 или NP_001041619.1 для полноразмерной аминокислотной последовательности цепи 2 для Fel d1 (также называемой цепью В или FELD В; см. также SEQ ID NO: 393), или фрагменты либо цепи 1, либо цепи 2, или фрагменты как от цепи 1, так и от цепи 2 белка Fel d1, с последующей иммунизацией вторичным иммуногеном или иммуногенно активным фрагментом природного белка. Животных можно иммунизировать либо только белком цепи 1, либо только белком цепи 2, либо белками как цепи 1, так и цепи 2, вводимыми последовательно или одновременно. С помощью частей цепи 1 и цепи 2 наряду с различными приемами образования связей или разделения спейсерами, которые известны специалистам в настоящей области, можно получить различные конструкции. Такие конструкции можно применять отдельно или в различных комбинациях для вызова гуморальных иммунных реакций in vivo. Например, в качестве иммуногенов можно применять рекомбинантные конструкции Fel d1, такие как проиллюстрированные в SEQ ID NO: 385, 394, 395, 396 или 397, или их фрагменты.
В соответствии с некоторыми вариантами осуществления антитела по настоящему изобретению получали от мышей, иммунизированных первичным иммуногеном, таким как биологически активный и/или иммуногенный фрагмент природного Fel d1, или ДНК, кодирующей его активный фрагмент. Фрагмент можно получить из N-концевого или С концевого домена либо цепи 1 и/либо цепи 2 у Fel d1.
В соответствии с некоторыми вариантами осуществления рекомбинантно полученный иммуноген Fel d1 можно получить путем прямой гибридизации двух цепей Fel d1, как описано в работе Kaiser et. al., с получением гибридного продукта, который имеет паттерн рефолдинга, сходный с паттерном природного Fel d1 (Kaiser, L. et al., (2003), J. Biol. Chem. 278(39):37730-37735). В соответствии с некоторыми вариантами осуществления иммуноген может представлять собой гибридный белок, такой как показанный в конструкциях SEQ ID NO: 385, 394, 395, 396 или 397, с последующей иммунизацией вторичным иммуногеном или иммуногенно активным фрагментом природного или рекомбинантно полученного Fel d1.
В соответствии с некоторыми вариантами осуществления рекомбинантные конструкции белка Fel d1, применяемые в описанных в настоящем документе исследованиях, состояли либо из i) цепи В Fel d1 (цепь 2) и цепи A Fel d1 (цепь 1), связанных в виде непрерывного, последовательного гибрида (с цепью В Fel d1 на N-конце), либо из ii) непрерывного, последовательного гибрида с цепью A Fel d1 на N-конце, за которой шел гибкий линкер [(Gly4Ser)3], за которым шла цепь В Fel d1. Такие конструкции также могли включать C-концевую метку (myc-myc-His6 или Fc участок мышиного IgG2a), как указано ниже. Белки экспрессировали в клетках яичника китайского хомяка (СНО). Экзогенную сигнальную последовательность, используемую для облегчения экспрессии в клетках СНО, не включали в перечни последовательностей.
В соответствии с некоторыми вариантами осуществления иммуноген мог представлять собой гибридный белок, содержащий любое одно или несколько из следующих: i) аминокислотные остатки 18-109 цепи 2 в Fel d1 (см. номер доступа в GenBank Р30440, а также SEQ ID NO: 393), гибридизированные через С-конец непосредственно с N-концом аминокислотных остатков 23-92 цепи 1 в Fel d1 (см. номер доступа в GenBank Р30438, а также SEQ ID NO: 392); ii) аминокислотные остатки 23-92 цепи 1 в Fel d1 (см. номер доступа в GenBank Р30438, а также SEQ ID NO: 392), гибридизированные через С-конец с N-концом аминокислотных остатков 18-109 цепи 2 в Fel d1 (см. номер доступа в GenBank Р30440, а также SEQ ID NO: 393); iii) аминокислотные остатки 18-109 цепи 2 в Fel d1 (см. номер доступа в GenBank NP_001041619.1), гибридизированные через С-конец непосредственно с N-концом аминокислотных остатков 19-88 цепи 1 в Fel d1 (см. номер доступа в GenBank NP_001041618.), например, конструкция, показанная в SEQ ID NO: 394 или 396; iv) аминокислотные остатки 19-88 цепи 1 в Fel d1 (см. номер доступа в GenBank NP_001041618.1), гибридизированные через С-конец с N-концом аминокислотных остатков 18-109 цепи 2 в Fel d1 (см. номер доступа в GenBank NP_001041619.1). См. также SEQ ID NO: 395). В соответствии с некоторыми вариантами осуществления гибридный белок мог иметь метку на C-терминальном конце конструкции, такую как myc-myc-гексагистидиновая метка (см. SEQ ID NO: 385, 396 или 397 для таких конструкций). Согласно соответствующим вариантам осуществления гибридный белок мог иметь мышиный Fc, присоединенный на C-терминальном конце конструкции (см. SEQ ID NO: 394 или 395 для таких конструкций). В соответствии с некоторыми вариантами осуществления цепи 1 и 2 соединены с помощью линкера, который известен специалистам в настоящей области техники, например (G4S)3 (см. SEQ ID NO: 395 и 397 для такой конструкции).
В соответствии с некоторыми вариантами осуществления антитела, которые специфично связываются с Fel d1, можно получить с помощью фрагментов указанных выше участков, или пептидов, которые выходят за рамки обозначенных участков на от приблизительно 5 до приблизительно 20 аминокислотных остатков с любого, или с обоих, N- или C-терминальных концов описанных в настоящем документе участков. В соответствии с некоторыми вариантами осуществления при получении специфичных к Fel d1 антител можно применять любую комбинацию указанных выше участков или их фрагментов. В соответствии с некоторыми вариантами осуществления для получения моноспецифических, биспецифических или мультиспецифических антител можно применять любой один или несколько из указанных выше участков Fel d1 или их фрагментов.
Полноразмерные белки или их фрагменты, которые применяли в качестве иммуногенов, как указано выше, вводили непосредственно, с адъювантом для стимуляции иммунной реакции, мыше VELOCIMMUNE®, содержавшей ДНК, кодирующую вариабельные участки тяжелой и каппа легкой цепей человеческого иммуноглобулина. Иммунную реакцию с продуцированием антител отслеживали с помощью иммунологического анализа, специфичного по отношению к Fel d1. При достижении необходимой иммунной реакции спленоциты собирали и гибридизировали с мышиными миеломными клетками для сохранения их жизнеспособности и создания линий гибридомных клеток. Линии гибридомных клеток подвергали скринингу и отбору для выявления линий клеток, которые продуцировали специфичные к Fel d1 антитела. С помощью такой методики и различных описанных выше иммуногенов получали несколько антител к Fel d1, химерных антител (т.е. антител, обладающих человеческими вариабельными доменами и мышиными константными доменами); некоторые полученные таким образом иллюстративные антитела обозначали H1M1230N, H1M1234N, H1M1241N, H2M1233N, H2M1236N, H2M1237N и H2M1242N.
Антитела к Fel d1 также непосредственно выделяли из антиген-положительных В-клеток без гибридизации с миеломными клетками как описано в патентном документе U.S. 2007/0280945 А1. С помощью такого способа получали несколько полностью человеческих антител к Fel d1 (т.е. антител, обладающих человеческими вариабельными доменами и человеческими константными доменами); полученные таким образом иллюстративные антитела обозначали следующим образом: Н4Н2574Р, H4H2590S, Н4Н2592В, H4H2594S, Н4Н2597Р, Н4Н2606В, Н4Н2607В, Н4Н2608В, Н4Н2636Р, Н4Н2645Р, Н4Н2793Р, Н4Н2797Р и Н4Н2864Р.
Биологические свойства иллюстративных антител, полученных в соответствии со способами согласно настоящему примеру, подробно описаны в приведенных ниже примерах.
Пример 2: Аминокислотные последовательности вариабельных участков тяжелой и легкой цепей
В таблице 1 изложены пары аминокислотных последовательностей вариабельных участков тяжелой и легкой цепей выбранных антител, специфичных к Fel d1, и их соответствующие идентификаторы антител. В настоящем документе антитела, как правило, обозначены согласно следующей номенклатуре: приставка Fc (например, "Н4Н", "H1M, "Н2М"), с последующим числовым идентификатором (например, "1232", как показано в таблице 1), с последующим суффиксом "Р" или "N". Таким образом, согласно настоящей номенклатуре антитело может быть обозначено, например, "H1M1232N". Приставки Н4Н, H1M и Н2М в используемых в настоящем документе обозначениях антител указывают на конкретный Fc участок антитела. Например, антитело "Н2М" имеет Fc мышиного IgG2, в то время как антитело "Н4Н" имеет Fc человеческого IgG4. Рядовому специалисту в настоящей области будет понятно, что антитело H1M или Н2М можно преобразовать в антитело Н4Н, и наоборот, но в любом случае вариабельные домены (включающие CDR), которые указаны с помощью показанных в таблице 1 числовых идентификаторов, остаются теми же. Антитела с одинаковыми числовыми обозначениями антител, но различающиеся по буквенному суффиксу N, В, S или Р, относятся к антителам, имеющим тяжелые и легкие цепи с идентичными последовательностями CDR, но с вариациями последовательностей в участках, которые выходят за пределы последовательностей CDR (т.е. находятся в каркасных участках). Таким образом, варианты N, В, S и Р конкретного антитела имеют идентичные последовательности CDR в своих вариабельных участках тяжелых и легких цепей, но отличаются друг от друга по своим каркасным участкам.
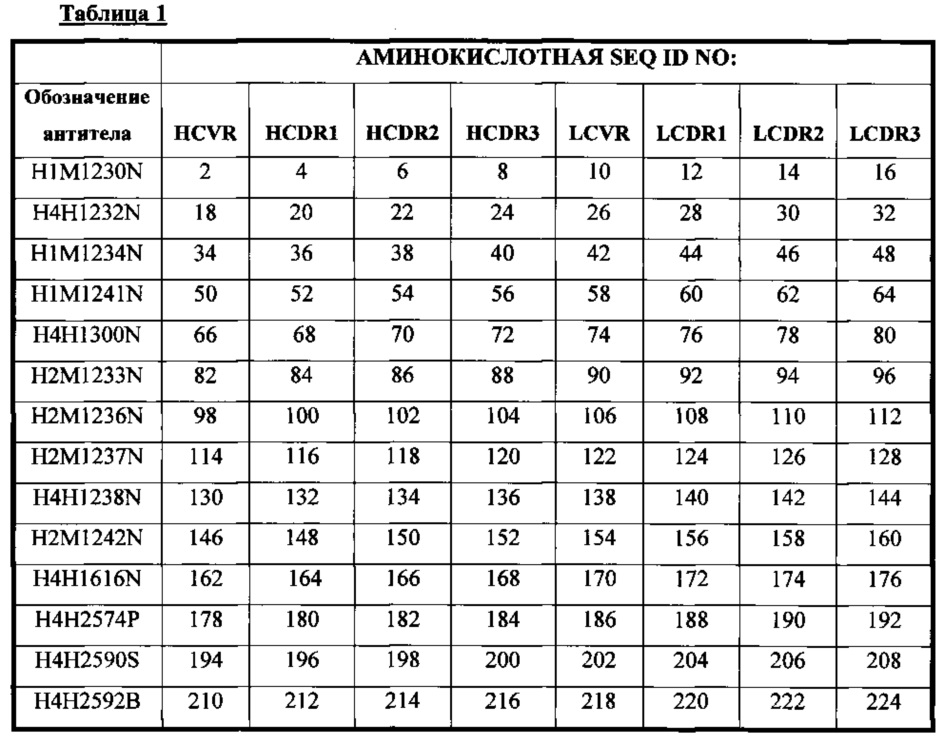
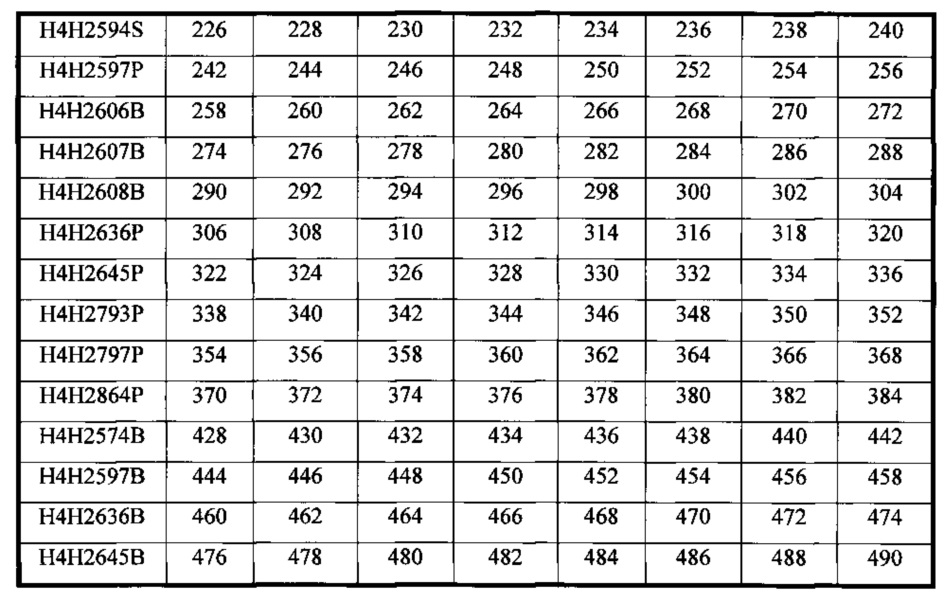
Пример 3: Анализ использования гена вариабельных участков
Для анализа структуры полученных антител клонировали и секвенировали нуклеиновые кислоты, кодирующие вариабельные участки антител. Из последовательности нуклеиновой кислоты и прогнозируемой аминокислотной последовательности антитела выявляли используемость гена (VH, D, JH, VK или JK) для каждого вариабельного участка тяжелой цепи (HCVR) и вариабельного участка легкой цепи (LCVR). В таблице 2 изложена используемость гена для выбранных антител в соответствии с настоящим изобретением.
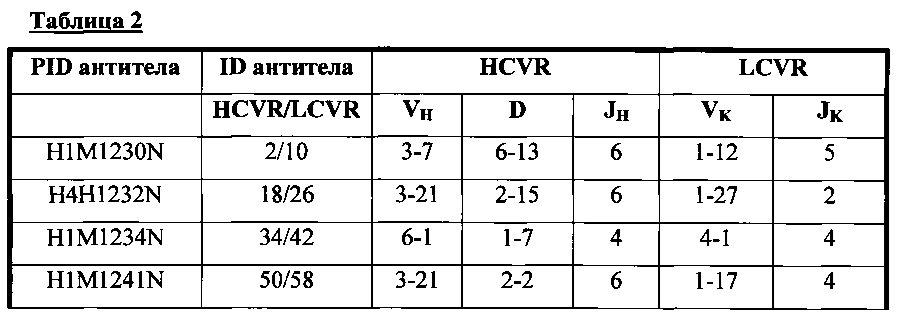
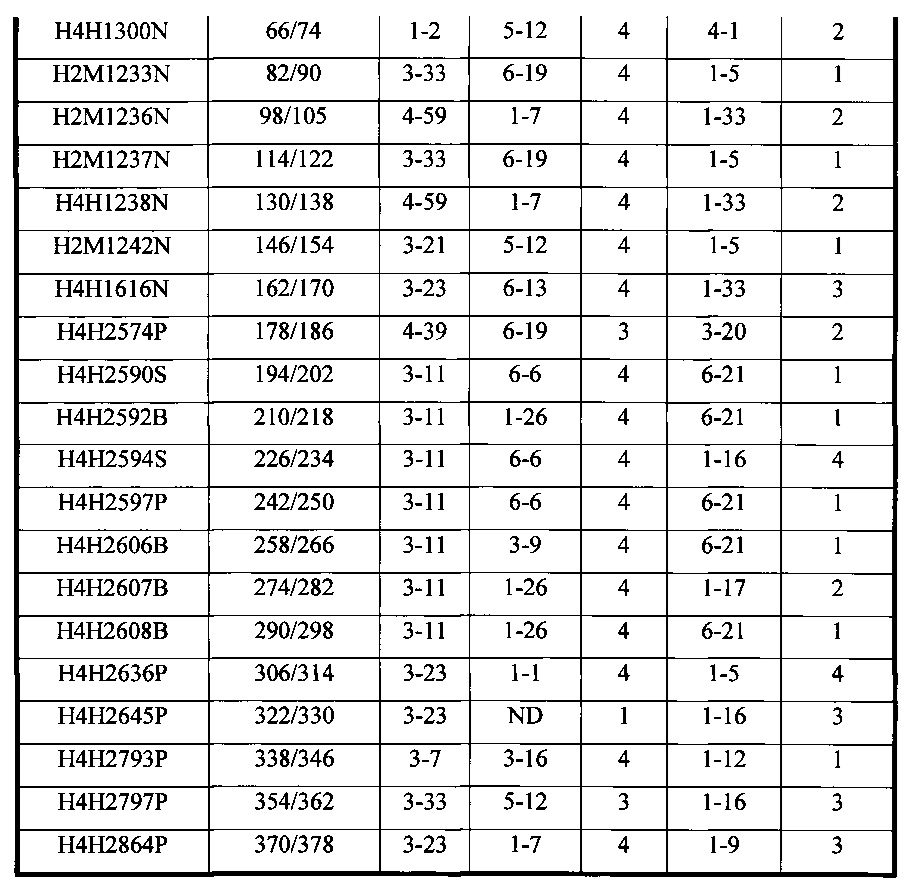
Пример 4: Связывание антител с Fel d1 по результатам определения с помощью поверхностного плазмонного резонанса
Константы скорости ассоциации и диссоциации связывания (ka и kd, соответственно) и расчетные равновесные константы диссоциации и периоды полудиссоциации (KD и t1/2, соответственно) для связывания антигена с моноклональными антителами к Fel d1 определяли с помощью анализа с применением биосенсора на основе технологии поверхностного плазмонного резонанса в режиме реального времени (Biacore Т200 или Biacore 2000). Поверхность биосенсора Biacore дериватизировали либо поликлональным антителом кролика к антителу мыши (GE Healthcare, № BR-1008-38), либо моноклональным антителом мыши к человеческому Fc (GE Healthcare, № BR-1008-39) для захвата антител к Fel d1, которые экспрессировались с мышиным Fc (приставка ID антитела H1M, Н2М, Н2аМ, H2bM) или человеческим Fc IgG4 (приставка ID антитела Н4Н), соответственно. Для построения графиков кинетики по меньшей мере две различные концентрации (в диапазоне от 390 пМ до 67 нМ) природного Fel d1 (Indoor Biotech, № NA-FD1-2) или рекомбинантного варианта белка Fel d1 (B-A)-mmH (SEQ ID NO: 396) вводили на поверхность с захваченным моноклональным антителом к Fel d1 при 25°С со скоростью потока, составлявшей 50 мкл/мин. в подвижном буфере (10 мМ HEPES, 150 мМ NaCl, 0,05% Р20, 3 мМ MgCl2, 3 мМ CaCl2). Fel d1 (B-A)-mmH экспрессировался в клетках яичника китайского хомяка (CHO) и состоял из аминокислот 18-109 из Fel d1 В (№ доступа Р30440), последовательно гибридизованных с аминокислотами 23-92 из Fel d1 А (№ доступа Р30438) с С-концевой myc-myc-гексагистидиновой меткой. Ассоциацию антитело-антиген отслеживали в течение 3-5 минут, а диссоциацию антигена от захваченного моноклонального антитела (только в подвижном буфере при 25°С) отслеживали в течение 10 или 15 минут. Константы скорости реакции ассоциации (ka) и диссоциации (kd) определяли путем обработки и приближения данных к 1:1 модели связывания с помощью программного обеспечения для приближенного изображения функций Scrubber 2.0. Равновесные константы диссоциации связывания (KD) и периоды полудиссоциации (t1/2) рассчитывали из констант скорости реакции следующим образом: KD=kd/ka и t1/2=ln(2)/kd. Параметры связывания для различных моноклональных антител к Fel d1 сведены в таблице 3 и таблице 4. В таблице 3 показаны аффинности согласно Biacore при 25°С для связывания природного Fel d1 с захваченными моноклональными антителами к Fel d1, а в таблице 4 показаны аффинности согласно Biacore при 25°С для связывания рекомбинантного Fel d1 с захваченными моноклональными антителами к Fel d1.
Как показано в таблице 3, 10 из 25 протестированных антител характеризовались значениями Ко ниже 1 нМ для связывания с природным Fel d1, варьировавшим в диапазоне от 207 пМ до 982 пМ. Как показано в таблице 4, 17 из 25 протестированных антител характеризовались значениями KD ниже 1 нМ для связывания с рекомбинантным Fel d1, варьировавшим в диапазоне от 144 пМ до 924 пМ. Два антитела, Н4Н2574В и Н4Н2793Р, связывались в таких экспериментальных условиях с рекомбинантным, но не с природным Fel d1.
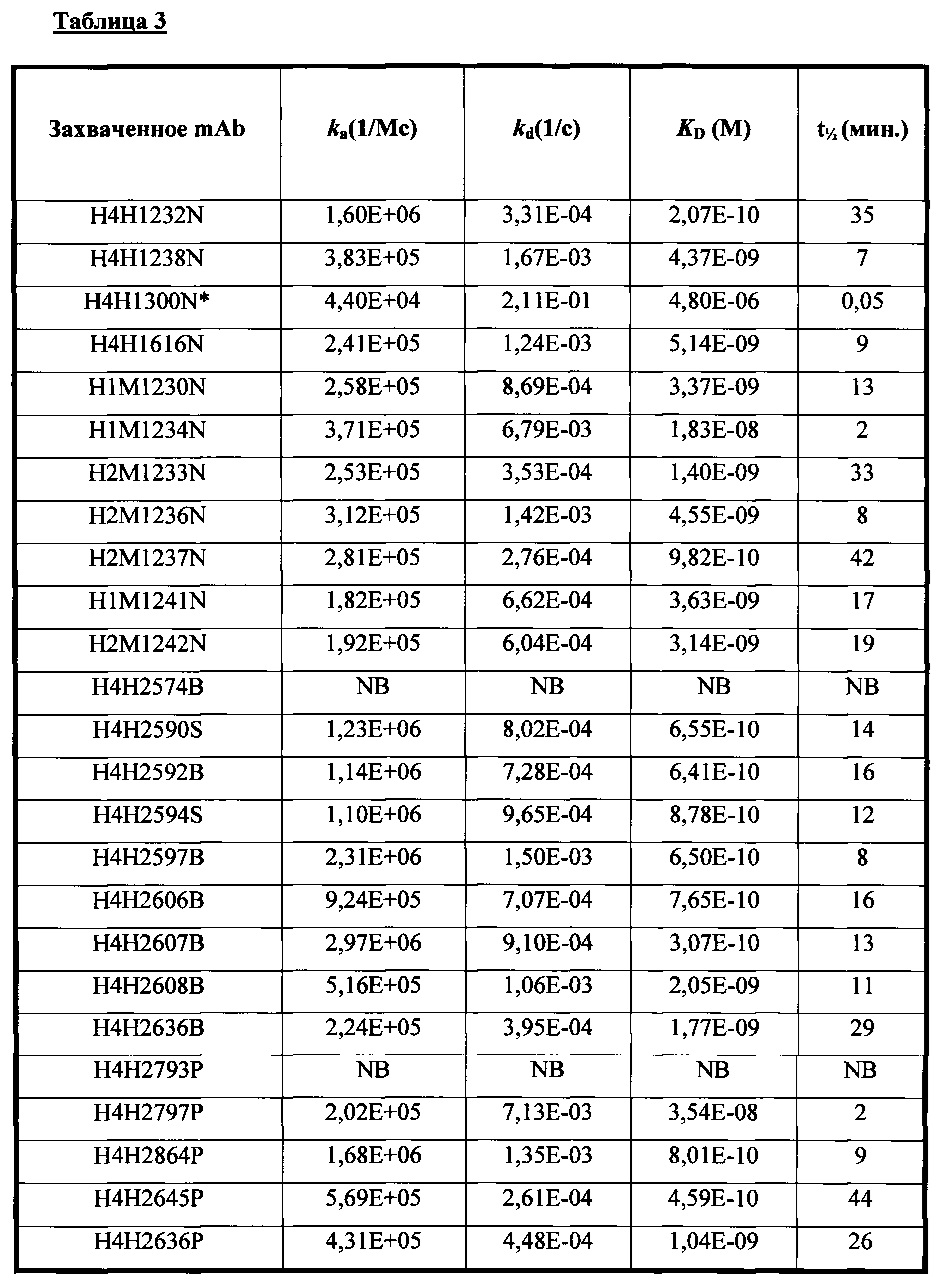
*По причине более низкой наблюдаемой аффинности связывания в этом образце использовали более высокие вводимые концентрации природного Fel d1 (67 нМ, 200 нМ и 600 нМ).
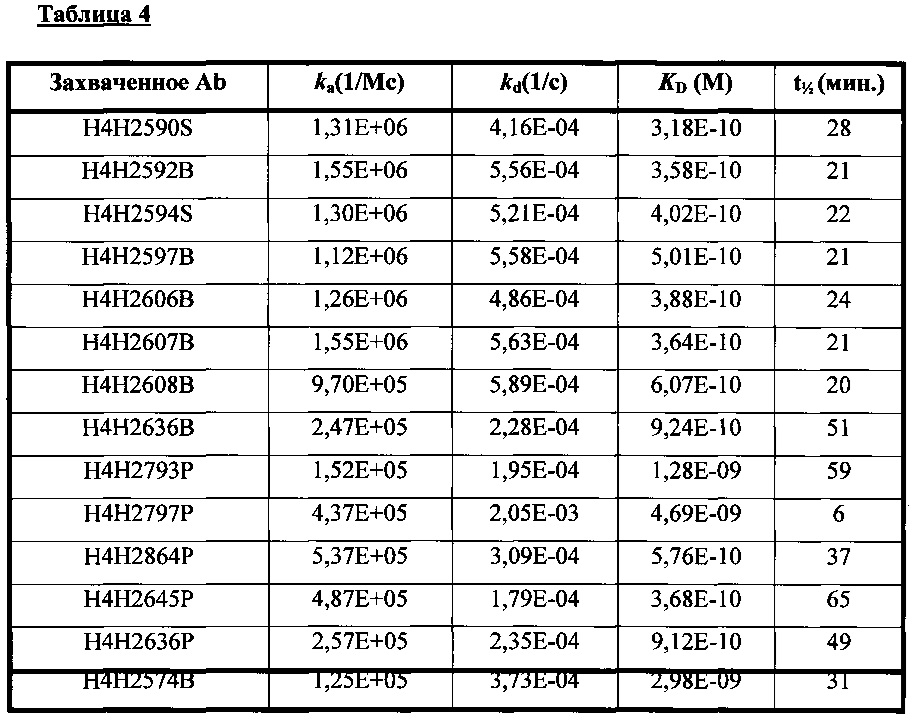
*По причине более низкой наблюдаемой аффинности связывания в этом образце использовали более высокие вводимые концентрации рекомбинантного Fel d1 (67 нМ, 200 нМ и 600 нМ).
Пример 5: Перекрестная конкуренция антител к Fel d1 за связывание с природным (n) Fel d1
Эксперимент по связыванию проводили с применением системы биосенсора Octet Red (Fortebio Inc.) для определения перекрестной конкуренции для панели из 8 антител к Fel d1, связывающихся с природным Fel d1 (nFel d1; Indoor Biotechnologies, № NA-FD1-2). Эксперимент проводили при 25°С в HBST буфере (0,01 М HEPES, рН 7,4, 0,15 М NaCl, 3 мМ EDTA, 0,05% об./об. Surfactant Р20), содержавшем 0,1 мг/мл BSA. Этап промывки HBST буфером осуществляли между каждым этапом связывания, а планшеты встряхивали во время этапов связывания и промывки с помощью орбитального встряхивателя для планшетов со скоростью 1000 об/мин. Первое антитело к Fel d1 (mAb-1) захватывали в течение 2 минут на поверхности биосенсора к hFc из маточных растворов антитела в количестве 10 мкг/мл (конечные уровни захвата ~1,5 нм единиц ответа). Покрытые наконечники биосенсора затем блокировали в течение 5 минут 100 мкг/мл раствором нерелевантного антитела. Наконечники биосенсора затем погружали в лунки, содержавшие 500 нМ nFel d1, на 5 минут, а затем в лунки, содержавшие 50 мкг/мл растворов второго антитела к Fel d1 (mAb-2). Растворы mAb-2 дополняли 100 мкг/мл нерелевантного антитела для минимизации неспецифичного связывания. Реакции связывания для связывания mAb-2 с nFel d1, который предварительно образовал комплекс с mAb-1, измеряли для 8×8 матрицы антител (таблица 5). Из каждого значения связывания для связывания mAb-2 с другой поверхностью захвата mAb-1/Fel d1 (внизу столбца таблицы 5) вычитали значение самоконкуренции mAb-1/Fel d1/mAb-2 (где mAb-1=mAb-2; по диагонали в таблице 5). Значения ниже 0,10 нм указывали на перекрестную конкуренцию mAb-1 и mAb-2 за общий сайт связывания на Fel d1.
Четыре антитела, Н4Н2636Р, H4H1616N, Н4Н2645Р и Н4Н2864Р, двунаправленно конкурировали с каждым из остальных за связывание с nFel d1, но не конкурировали с никаким из антител к Fel d1. Два антитела, H4H1232N и Н4Н2597Р, двунаправленно конкурировали с каждым из остальных за связывание с nFel d1. Как H4H1232N, так и Н4Н2597Р однонаправленно конкурировали с H4H1300N. Двунаправленную конкуренцию с H4H1300N нельзя было определить, поскольку H4H1300N предварительно не образовывал комплекс с nFel d1. H4H1238N не конкурировал с никаким из антител к Fel d1 за связывание с nFel d1.

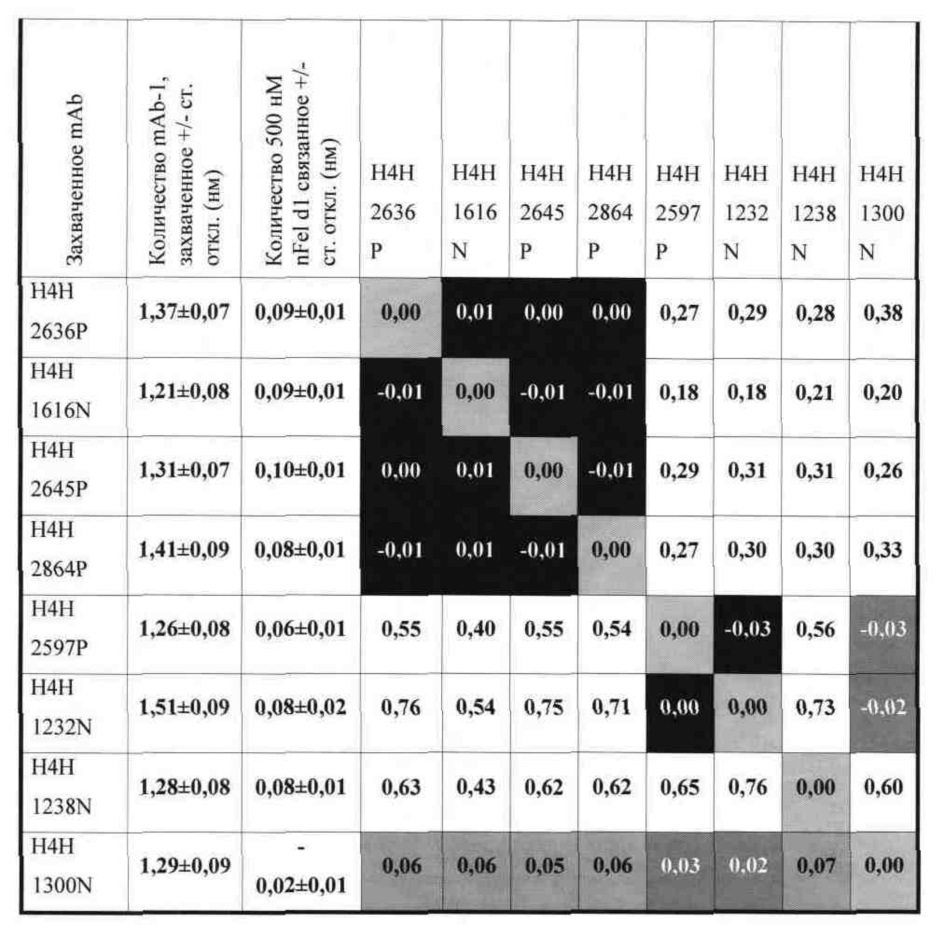
Пример 6; Влияние антител к Fel d1 в in vivo модели пассивной кожной анафилаксии (РСА)
Для оценки in vivo дегрануляции тучных клеток использовали in vivo модель пассивной кожной анафилаксии (РСА). Модель включала внутрикожную инъекцию аллерген-специфичной антисыворотки в локальный участок на коже с последующей внутрикожной инъекцией только антигена с красителем.
Аллергическая реакция вызывала расширение капилляром и повышала проницаемость сосудов в месте сенсибилизации, что приводило к избирательному накоплению красителя в данном месте. Краситель можно было экстрагировать из ткани и количественно оценить с помощью спектрофотометрии. Выход красителя из сосудов в ткань, сенсибилизированную тестируемой антисывороткой, сравнивали с выходом из сосудов в ткань, сенсибилизированную нерелевантной антисывороткой.
Антисыворотку получали путем иммунизации Balb/c мышей посредством 5 мкг природного белка Fel d1, очищенного из экстракта кошачьей шерсти (Indoor Biotechnologies, № LTN-FD1-1), 5 мкг неочищенного экстракта аллергена арахиса (Greer Laboratories, № XPF171D3A25) или 1250 биоэквивалентные аллергенные единицы (BAU) стандартизированного экстракта кошачьей шерсти (Greer Laboratories, № GTE3A01) в растворе 1 мг/мл квасцовой муки (Pierce, №77161) в 1X фосфатно-солевом буферном растворе. Спустя две недели (день 14) сенсибилизированных мышей стимулировали дозами аллергена, идентичного аллергену, использованного для начальной иммунизации. Спустя две недели после стимуляции (день 28) мышей умерщвляли и собирали сыворотку. Суммарную концентрацию IgE в выделенной антисыворотке определяли с помощью ELISA. Конечную концентрацию антисыворотки разводили до 2400 нг/мл IgE в 1X фосфатно-солевом буферном растворе.
Для определения влияния антител к Fel d1 на дегрануляцию тучных клеток в модели РСА перед сенсибилизацией уха антисывороткой, полученной как описано выше, группам Balb/c мышей сначала подкожно вводили инъекцией либо контрольное человеческое антитело с изотипом IgG4, либо антитело к Fel d1, либо комбинацию антител к Fel d1 с дозами, составлявшими 5 мг/кг (общая доза антитела, 2,5 мг/кг каждого антитела), для экспериментов с одной дозой, если не было указано иное, или с концентрациями, варьировавшими в диапазоне от 0,06 мг/кг до 2 мг/кг, для экспериментов с диапазонами доз. Спустя три дня предварительного лечения при помощи антитела одну группу мышей ("группу природного Fel d1") сенсибилизировали внутрикожной инъекцией 10 мкл полученной из природного Fel d1 антисыворотки или 10 мкл полученной из арахиса антисыворотки (отрицательного контроля) в правое и левое ухо, соответственно, каждой мыши. Вторую группу мышей ("группу кошачьего экстракта") сенсибилизировали 20 мкл полученной из экстракта кошачьей шерсти антисыворотки или 20 мкл полученной из арахиса антисыворотки (отрицательного контроля) в правое и левое ухо, соответственно, каждой мыши. Спустя двадцать четыре часа после сенсибилизации мышам в группе природного Fel d1 вводили провокационную дозу путем внутривенной инъекции (100 мкл на мышь) раствора 0,25 мкг/мл природного Fel d1 (Indoor Biotechnologies, № LTN-FD1-1), растворенного в 1X фосфатно-солевом буферном растворе, содержавшем 0,5% (объем/масса) красителя Evan's blue (Sigma, № Е2129). Аналогично, спустя 24 часа после сенсибилизации мышам в группе кошачьего экстракта вводили провокационную дозу 250BAU стандартизированного экстракта кошачьей шерсти [стандартизированный экстракт кошачьей шерсти (Greer Laboratories, № GTE3A01)], растворенного в 1X фосфатно-солевом буферном растворе, содержащем 0,5% (объем/масса) красителя Evan's blue (Sigma, № Е2129). Спустя один час после введения провокационной дозы антигена мышей умерщвляли, уши отрезали и помещали в 1 мл формамида и инкубировали в течение 3 дней при температуре 56°С для экстракции красителя Evan's blue из ткани. Затем ткань уха удаляли из формамида, промакивали для удаления избытка жидкости и взвешивали. Аликвоты размером двести микролитров каждого экстракта формамидом переносили в 96-луночные планшеты в двух повторах. Поглощение полученных надосадочных жидкостей измеряли при 620 нм. OD преобразовывали в концентрацию красителя Evan's blue с помощью стандартной кривой. Средняя концентрация красителя Evan's blue, проникшего из сосуда в ткань сенсибилизированного антисывороткой уха (нормализованная по массе ткани уха), рассчитывали для группы, обработанной антителом изотипического контроля и обозначаемой F(изотип, средн.). Уменьшение проникновения из сосуда красителя Evan's blue, произошедшее в результате предварительной обработки антителом, рассчитывали на мышь путем вычитания количества красителя Evan's blue для Fel d1 обработанной антителом группы или экстракта сенсибилизированного уха, обозначенного F(mAb,i), из F(изотип, средн.). Это число затем делили на разницу между F(изотип, средн.) и количеством красителя уха, сенсибилизированного арахисом, из обработанной антителом группы [P(mAb,i)] и умножали на 100 с получением общего процента уменьшения проникновения из сосуда красителя для каждой мыши (% уменьшения).
% уменьшения (на мышь) = 100*[F(изотип, средн.) - F(mAb,i)]/[F(изотип, средн.) - P(mAb,i)]
Затем для каждой группы антитела рассчитывали средний процент уменьшения вытекания красителя. Результаты, выраженные как (среднее ± SD) процента уменьшения Evan's blue, показаны в таблице 6 и таблице 7 для группы природного Fel d1 и в таблице 8 для группы экстракта кошачьей шерсти.
Как показано в таблице 6, у семи групп мышей из группы природного Fel d1, при обработке конкретными комбинациями антител к Fel d1 с фиксированными концентрациями, наблюдали уменьшения проникновений из сосуда красителя, варьировавшие от 79% до 103%, по сравнению с мышами, получавшими контрольное антитело. У мышей, обработанных парными комбинациями антител H4H2590S/H4H1238N, H4H2590S/H4H2574P или H4H1232N/H4H1616N, наблюдали менее 3% уменьшения проникновения из сосуда красителя по сравнению с мышами, получавшими контрольное антитело, что демонстрировало, что не все тестируемые в этой модели антитела к Fel d1 были эффективны.
Кроме того, с мышами из группы природного Fel d1 проводили варьирующие по дозе эксперименты, результаты которых показаны в таблице 7. Отдельные антитела не были настолько же эффективны при уменьшении проникновения из сосудов красителя, как комбинации антител к Fel d1 в тестируемых дозах.
Отдельную пару антител к Fel d1 (Н4Н2636Р и H4H1232N) с множеством уровней доз, а также каждое из этих антител к Fel d1 отдельно с одним (наиболее высоким) уровнем дозы, дополнительно тестировали в модели РСА с применением мышей, которых сенсибилизировали и вводили провокационную дозу экстракта кошачьей шерсти, результаты чего показаны в таблице 8. На уровне 2 мг/кг такие единичные отдельные антитела к Fel d1 не были настолько эффективны в уменьшении проникновения красителя из сосудов, насколько комбинация двух антител. Комбинация Н4Н2636Р и H4H1232N на уровне как 2 мг/кг, так и 1 мг/кг уменьшала проникновение красителя из сосудов на более чем 90% по сравнению с изотипическим контролем в модели РСА с применением экстракта кошачьей шерсти в качестве антигена.
Все уменьшения, которые были статистически значимы (р<0,05) по сравнению с изотипическим контролем по результатам двухфакторного дисперсионного анализа с апостериорным критерием Бонферронни отмечены звездочкой (*). Количество использованных на группу мышей (n) в таблицах приведено в скобках.
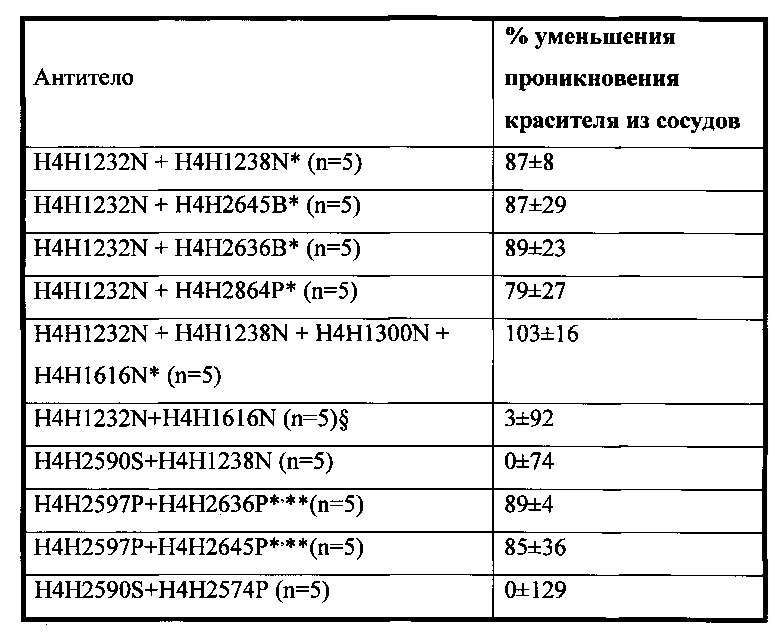
§10 мг/кг суммарная концентрация антител; **0,5 мг/кг суммарная концентрация антител
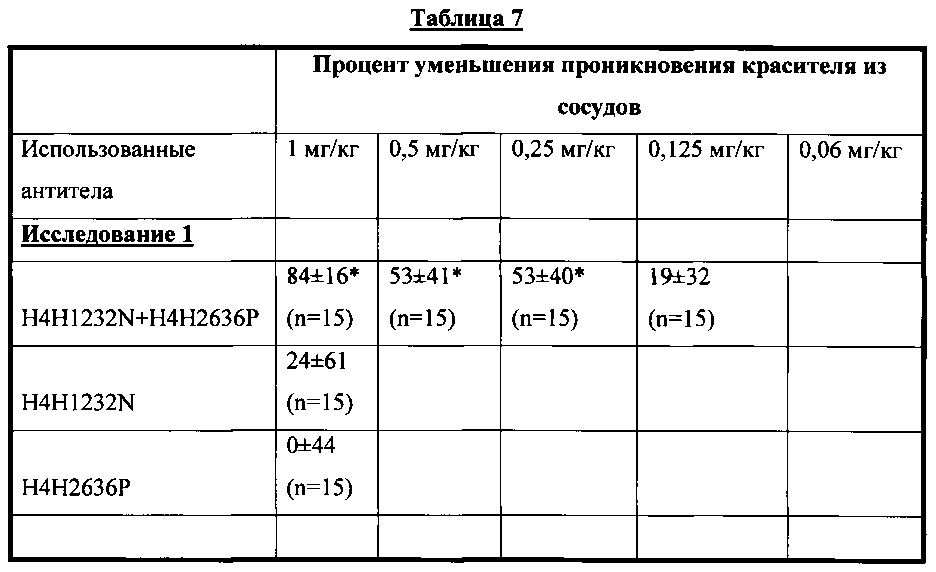
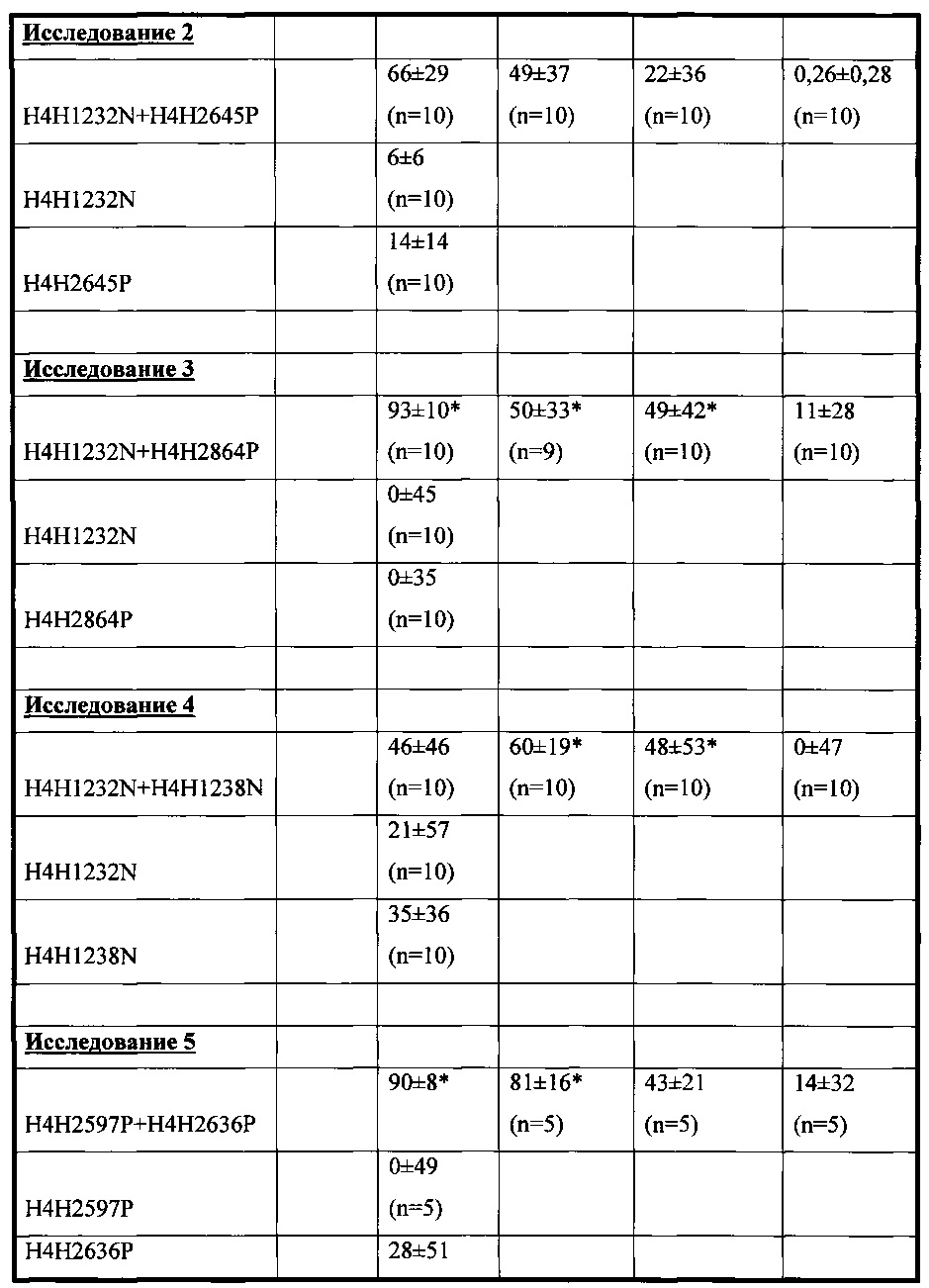
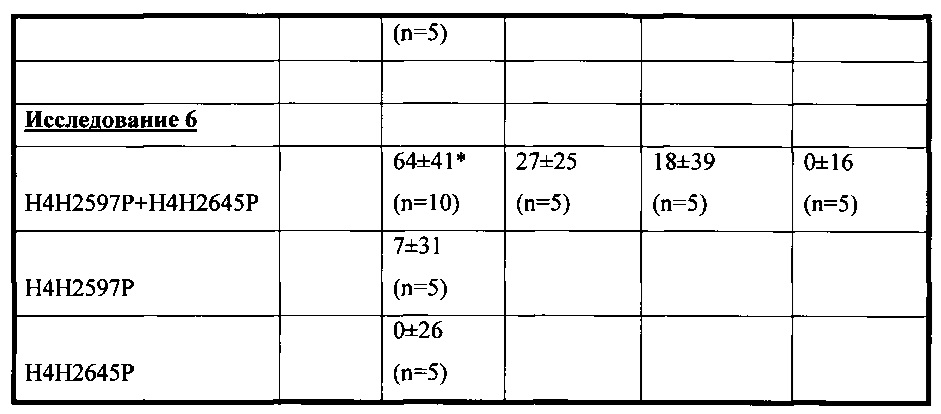
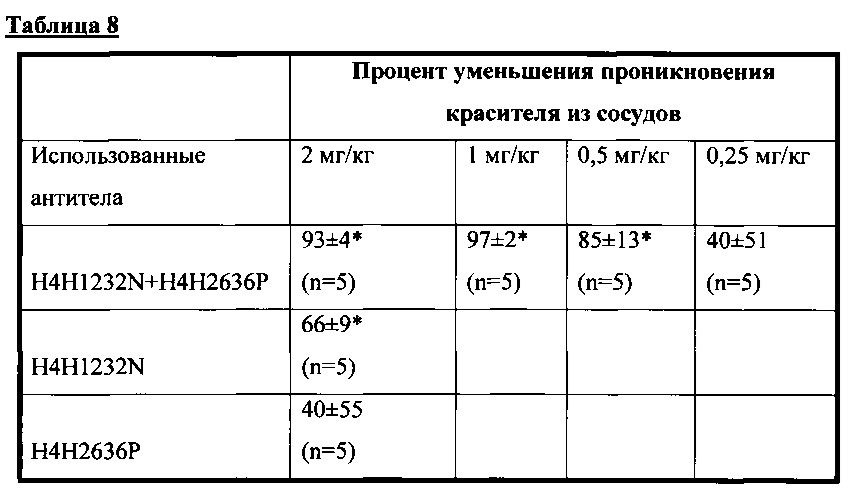
Пример 7: Влияние антител к Fel d1 в in vivo модели воспаления легких
In vivo мышиную модель воспаления легких использовали для оценки индуцированного аллергеном воспаления легких и накопления слизи, которые могли быть связаны с астмой или риноконъюнктивитом. Модель включает повторное интраназальное введение аллергена предварительно сенсибилизированным аллергеном мышам. Связанное с аллергеном воспаление может вызывать повышения накопления слизи в легких, миграции эозинофилов в легкое, общего IgE сыворотки крови и уровней аллерген-специфичного IgG1.
Мышей Balb/c внутрибрюшинно иммунизировали 1 мкг природного белка Fel d1, очищенного из экстракта кошачьей шерсти (Indoor Biotechnologies, № LTN-FD1-1), в растворе 1 мг/мл квасцовой муки (Pierce, №77161) в 1X фосфатно-солевом буферном растворе. Спустя семь дней сенсибилизированных мышей внутрибрюшинно стимулировали 1 мкг природного Fel d1 в растворе 1 мг/мл квасцовой муки в 1X фосфатно-солевом буферном растворе. На дни 17, 21 и 25 группам мышей (n=5) подкожно вводили инъекцией человеческий IgG4 в качестве антитела изотипического контроля или 1:1 комбинацию антител к Fel d1 H4H1232N и Н4Н2636Р в количестве 20 мг/кг (суммарная доза антител). На дни 20, 24 и 28 мышам интраназально вводили провокационную дозу 0,05 мкг природного Fel d1, разведенного в 20 мкл 1X фосфатно-солевого буферного раствора. В те же дни контрольным мышам вводили провокационную дозу 20 мкл 1X фосфатно-солевого буферного раствора. На день 32 всех мышей умерщвляли и собирали их легкие. Экспериментальное введение дозы и протокол лечения для групп мышей показаны в таблице 9.
Для определения находящегося в системном кровообращении общего IgE и специфичного к Fel d1 IgG1 в сыворотке крови мышей собирали образцы сыворотки у каждой мыши посредством концевой пункции сердца с помощью 1 мл ТВ шприца 27G1/2 (Becton Dickinson, №309306) с прикрепленной иглой. Образцы крови помещали в пробирки для отделения сыворотки BD microtainer® (Becton Dickinson, №365956), центрифугировали, а затем сыворотку переносили в свежую пробирку для хранения до анализа.
Для определения суммарной концентрации IgE в образцах сыворотки крови использовали набор ELISA OPTELA (BD Biosciences, №555248) согласно инструкциям производителя. Образцы сыворотки разводили и инкубировали с антителом захвата к IgE, нанесенным на 96-луночные планшеты. Общий IgE определяли с помощью биотинилированного вторичного антитела к IgE мыши. В качестве стандарта использовали очищенный мышиный IgE, меченный пероксидазой хрена (HRP). Для детектирования активности HRP использовали хромаген 3,3',5,5'-тетраметилбензидин (ТМВ) (набор субстратных реагентов BD OPTEIA, BD, №555214). Затем добавляли стоп-раствор, представлявший собой 1М серную кислоту, и измеряли поглощение на 450 нм на планшет-ридере Molecular Devices SpectraMax М5. Анализ данных проводили с помощью программного обеспечения Prism™. Средние величины уровней циркулирующего IgE для каждой экспериментальной группы выражали как нг/мл (± SEM), как показано в таблице 10. У мышей, которым была интраназально введена провокационная доза Fel d1, при лечении комбинацией антител к Fel d1 наблюдали значимое снижение величины IgE [6683 (±1394) нг/мл] по сравнению с мышами, получавшими антитело изотипического контроля [14080 (±1505) нг/мл].
Для определения уровней специфичного к Fel d1 IgG1 в образцах сыворотки крови каждой мыши использовали ELISA. Планшеты с нанесенным Fel d1 инкубировали с серийно разведенными образцами сыворотки крови мышей с последующей инкубацией с конъюгированным с HRP антителом к IgG1 мыши (BD Biosciences, #559626). Все образцы проявляли с помощью раствора ТМВ и анализировали как описано выше. Относительные уровни циркулирующего IgG1 в сыворотке крови представляли в виде титрационных единиц (титрационные единицы рассчитывали путем умножения измеренной OD на фактор разведения, необходимый для достижения OD450, которая превышала фон более чем в два раза). Средние уровни циркулирующего Fel d1-специфичного IgG1 для каждой экспериментальной группы выражали в виде титр × 103 (± SEM), как показано в таблице 11. У мышей, которым была интраназально введена провокационная доза Fel d1, при лечении комбинацией антител к Fel d1 наблюдали значимое снижение величины уровней циркулирующего Fel d1-специфичного IgG1 [титр 105,3 (±31,33)×103] по сравнению с мышами, получавшими антитело изотипического контроля [526,1(±144,0) нг/мл].
Сбор легких для анализа клеточного инфильтрата
После обескровливания у каждой мыши удаляли правое легко и помещали его в небольшую чашку Петри, содержавшую среду Игла, модифицированную по Дульбекко (DMEM) (Irvine Scientific, №9033), и разрубывали на кубики, которые имели размер примерно 2-3 мм. Кубики затем переносили в пробирку, содержавшую раствор 20 мкг/мл ДНКазы (Roche, №10104159001) и 0,7 ед./мл либеразы ТН (Roche, №05401151001), разведенные в сбалансированном солевом растворе Хенкса (HBSS) (Gibco, №14025), и помещали в водяную баню с температурой 37°С на 20 минут со встряхиванием на вортексе каждые 5 минут. Затем реакцию останавливали путем добавления этилендиаминтетрауксусной кислоты (EDTA) (Gibco, №15575) с конечной концентрацией 10 мМ. Каждое легкое разминали, фильтровали через 70 мкм фильтр, центрифугировали, а затем остаток легкого ресуспендировали в 4 мл лизирующего буфера АСК (Gibco, №10492) для удаления эритроцитов. После 3-минутной инкубации при комнатной температуре добавляли DMEM для дезактивации буфера АСК. Суспензии клеток центрифугировали, а затем клеточные осадки ресуспендировали в 10 мл буферного раствора MACS [смеси раствора для промывания Miltenyi auto MACS Rinsing Solution (Militenyi Biotec, №130-091-222) и BSA MACS (Militenyi Biotec, №130-091-376)]. Ресуспендированные образцы фильтровали через 70 мкм фильтр и 1×106 клеток высеивали в 96-луночном планшете с V-образным дном. Затем клетки центрифугировали, а осадки ресуспендировали в очищенном Fc Block крысы к мышиному CD16/CD32, (BD Biosciences Clone: 2.4G2, №553142), разведенному в MACS буфере в течение 15 минут при 40°С. Клетки два раза промывали, а затем инкубировали в смеси соответствующих антител (описанной в таблице 12), разведенной в буфере MACS в течение 30 минут при 4°С в защищенном от света месте. После инкубации с антителами клетки два раза промывали в буфере MACS и ресуспендировали в cytofix от BD (BD Biosciences, №554655) в течение 15 минут при 4°С в защищенном от света месте. Клетки промывали, ресуспендировали в буфере MACS, а затем переносили в пробирки для FACS от BD (BD Biosciences, №352235) для анализа эозинофилов с помощью проточной цитометрии. Эозинофилы определяли как клетки, которые были CD45+, GR1-, CD11clo, SiglecFhi. Данные выражали как частоту эозинофилов среди CD45+ клеток (± SEM) в таблице 13.
У мышей, которым была интраназально введена провокационная доза Fel d1, при лечении комбинацией антител к Fel d1 наблюдали значимое снижение частоты эозинофилов в популяции CD45+ клеток по сравнению с мышами, не получавшими антитело (67% снижение) или получавшими антитело изотипического контроля (46% снижение), как показано в таблице 13.
Сбор легких для гистологического анализа
После обескровливания удаляли левые легкие и помещали в пробирки, содержавшие 5 мл раствора 4% (объем/масса) параформальдегида (Boston Bioproducts, № ВМ-155) в 1X фосфатно-солевом буферном растворе и хранили при комнатной температуре в течение 3 дней. Образцы легких потом промакивали досуха и переносили в пробирки, содержавшие 70% этанол, для гистологического анализа. Образцы отсылали в Histoserv, Inc (Джермантаун, Мэриленд) для получения срезов и окрашивания реактивом Шиффа (PAS).
Получали примерно 35 цифровых изображений по всей области каждого окрашенного PAS среза легкого с помощью светового микроскопа для получения изображений Zeiss Axioplan 2 с камерой Zeiss AxioCam MRc. Затем воссоздавали изображение целого легкого из меньших изображений и анализировали с применением программного обеспечения ImageJ с помощью плагина для цветового порога. Участки скопления слизи в бронхиальном просвете выявляли и количественно оценивали посредством выбранного пользователем цветового порога и нормализовали к общей области просвета, которую выявляли и количественно оценивали с помощью отдельных настроек цветового порога. Процент бронхиального просвета, занятого скоплением слизи, для каждого легкого выражали как [(зона слизи/площадь просвета) × 100] и рассчитывали для каждой группы лечения. Результаты, выраженные как средний процент обструкции легкого (± SEM), показаны в таблице 14.
У мышей, которых лечили комбинацией антител к Fel d1, наблюдали тенденцию к уменьшенному скоплению слизи в бронхах легкого (5,21+/-0,81% скопления слизи) по сравнению с мышами, получавшими контрольное антитело (10,81+/-1,13% скопления слизи) в модели воспаления легкого, как показано в таблице 14. Среди групп мышей не наблюдали различий в размере бронхиального просвета или общего размера легкого.
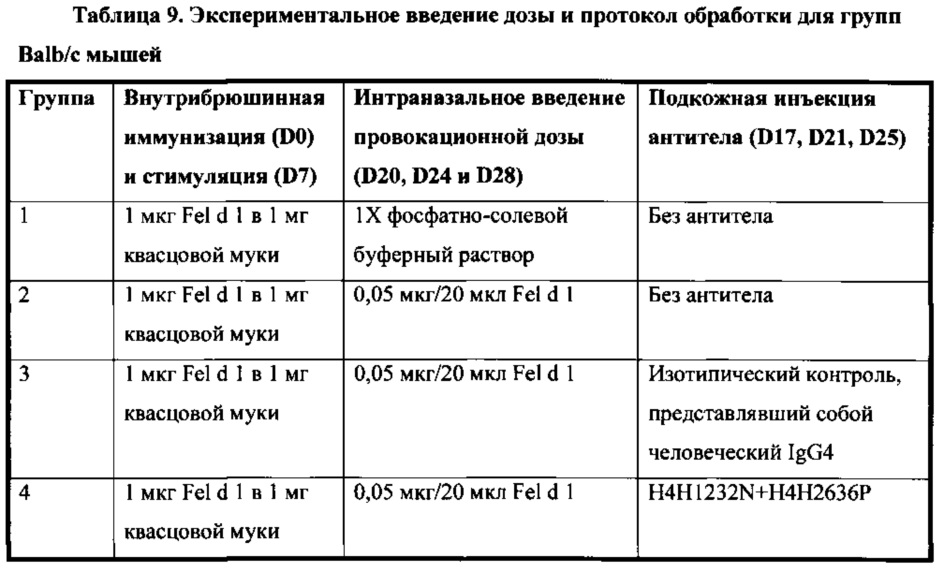



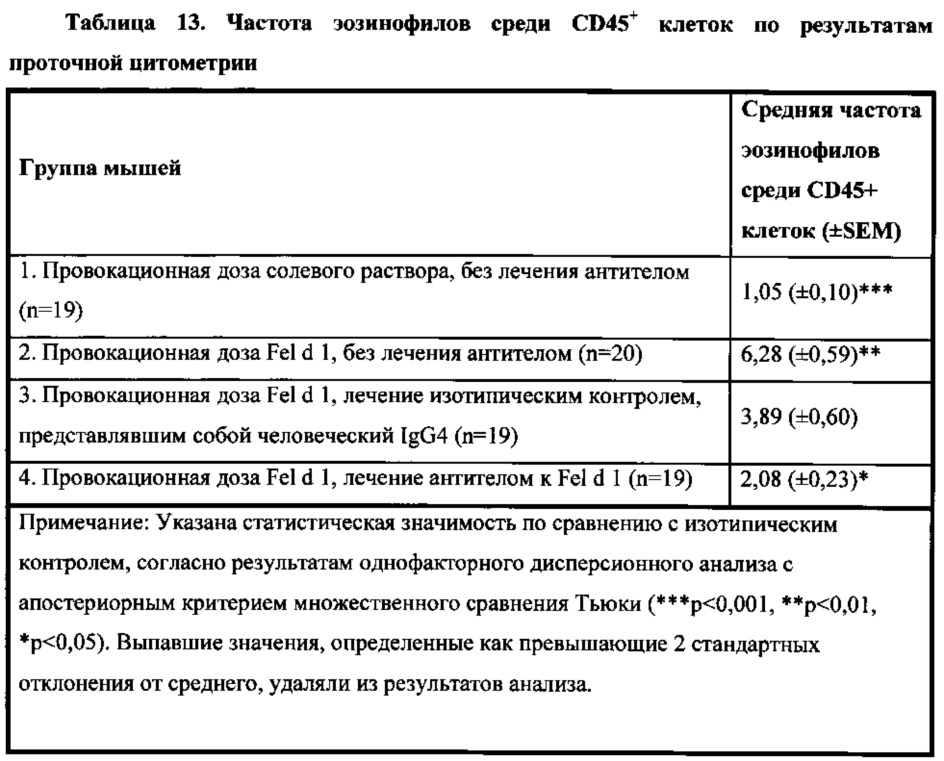

Пример 8: Картирование эпитопов на основе обмена водород-дейтерий
Для определения эпитопов Fel d1 (гетеродимерного белка, состоящего из цепи A Fel d1 и цепи В FELD1), распознаваемого двумя антителами к Fel d1, проводили исследования на основе обмена водород-дейтерий (H/D) для каждого антитела, образующего совместный комплекс с Fel d1. Перед проведением экспериментов по обмену H/D экспрессируемый клетками СНО рекомбинантный Fel d1, содержащий в своем составе аминокислоты 18-109 цепи В Feld 1 (номер доступа GenBank NP_001041619.1), последовательно гибридизированные с аминокислотами 19-88 FELD1 А (№ доступа GenBank NP_001041618.1), экспрессированными с С-концевой myc-myc-гексагистидиновой меткой и мутацией D27G (Fel d1B-A-mmH; SEQ ID: 396), дегликозилировали при 37°С в течение 4 часов в естественных условиях с помощью ПНГазы F (New England BioLabs, №0704). Для настоящего исследования два антитела к FELD1 (H4H1232N и Н4Н2636Р) ковалентно связывали с N-гидроксисукцинимид (NHS) агарозными гранулами (GE Lifescience, №17-0906-01) согласно протоколу производителя.
Для картирования эпитопа связывания у Fel d1B-A-mmH, распознаваемого H4H1232N, проводили два набора экспериментов по обмену H/D (все реакции по связыванию и обмену проводили при комнатной температуре). В первом эксперименте использовали формат 'обмен в растворе/обратный обмен на гранулах' (осуществление обмена в растворе с последующим обратным обменом на гранулах). Для осуществления обмена дегликозилированный белок Fel d1B-A-mmH дейтеризовали в течение 5 и 10 минут (в двух отдельных подэкспериментах) в PBS буфере с рН 7,4, приготовленном с D2O (PBS-D), а затем связывали с гранулами с H4H1232N в течение 2-минутной инкубации в PBS-D. Совместный комплекс Fel d1B-A-mmH, связанного с гранулами с H4H1232N, затем промывали PBS буфером с pH 7,4, приготовленном с H2O (PBS-H), и инкубировали в PBS-Н в течение половины времени осуществления обмена (обратный обмен), что позволяло только эпитопам на Fel d1B-A-mmH, защищенном в результате связывания антитела H4H1232N, оставаться дейтеризованным. После обратного обмена связанный Fel d1B-A-mmH элюировали с гранул с помощью водного раствора ледяной 0,1% трифторуксусной кислоты (TFA). Элюированный Fel d1B-A-mmH затем расщепляли с помощью иммобилизированного пепсина (Thermo Scientific, №20343) в течение 5 минут при 4°С. Полученные в результате пептиды обессоливали при 4°С с помощью хроматографических микродозаторов ZipTip (Millipore, № ZTC18S096) согласно протоколу производителя, а затем сразу анализировали на масс-спектрометре (MS) для анализа методом времени пролета при ионизации лазерной десорбцией при содействии матрицы (MALDI-TOF) UltrafleXtreme.
Второй эксперимент назывался 'обмен на гранулах/обратный обмен на гранулах' (осуществление обмена на гранулах с последующим обратным обменом на гранулах). Для такого эксперимента дегликозилированный Fel d1B-A-mmH сначала связывали с гранулами с H4H1232N, а затем инкубировали в течение 5 или 10 минут (в отдельных подэкспериментах) в PBS-D для осуществления обмена. Последующие этапы (обратный обмен, расщепление пепсином и MS-анализ) проводили как описано выше для процедуры 'обмен в растворе/обратный обмен на гранулах'. Центроидные значения или средние отношения массы к заряду (m/z) у всех детектируемых пептидов рассчитывали и сравнивали между результатами экспериментов обмен в растворе/обратный обмен на гранулах и обмен на гранулах/обратный обмен на гранулах. Пептиды, характеризовавшиеся повышенной массой после процедуры обмен в растворе/обратный обмен на гранулах по сравнению с процедурой обмен на гранулах/обратный обмен на гранулах, включали аминокислоты в белке Fel d1, защищенные от обмена в результате связывания антитела, и, следовательно, у них обнаруживали связывающие участки-эпитопы.
Эксперимент по обмену H/D в отношении связывания Fel d1B-A-mmH с антителом к Fel d1 Н4Н2636Р проводили с помощью той же процедуры, что описана выше для H4H1232N, но с заменой гранул с H4H1232N на гранулы с Н4Н2636Р.
Результаты сравнения центроидных значений m/z для всех пептидов, детектированных в экспериментах по обмену H/D у Fel d1B-A-mmH с H4H1232N, показаны в таблице 15. Такие пептиды выявляли с помощью MS на основе жидкостной хроматографии - ионизации лазерной десорбцией при содействии матрицы (LC-MALDI). Большая часть полученных в результате расщепления пепсином пептидов давали похожие центроидные значения (различия <0,3 единиц m/z) как для протокола обмен в растворе/обратный обмен на гранулах, так и для протокола обмен на гранулах/обратный обмен на гранулах, для каждого из двух различных периодов обмена и обратного обмена. Тем не менее, три пептида с аминокислотами, занимающими положения 85-103, 85-104 и 113-127 в Fel d1B-A-mmH (SEQ ID NO: 396), имели отличия по центроидным значениям m/z>0,3 как в 5-минутных, так и в 10-минутных экспериментах. Отличия между центроидными значениями у протокола обмен в растворе/обратный обмен на гранулах от протокола обмен на гранулах/обратный обмен на гранулах в таблице 15 обозначены жирным шрифтом. Поскольку у другого пептида, с аминокислотами 117-127 в SEQ ID NO: 396, не наблюдали сохранения дейтеризации после обратного обмена, участок с защитой от обмена в пептиде 113-127 можно было уменьшить до остатков 113-116 в SEQ ID NO: 396. Два участка, с остатками 85-104 (SEQ ID NO: 403) и 113-116 (SEQ ID NO: 426), были защищены от полного обратного обмена в результате связывания H4H1232N с Fel d1B-A-mmH после обмена. Таким образом, эти два сегмента определяли с помощью способа обмена H/D как непрерывный эпитоп для связывания антитела H4H1232N с белком Fel d1B-A-mmH.
Результаты сравнений центроидных значений m/z для пептидов, детектированных в экспериментах по обмену H/D у Fel d1B-A-mmH, образующего комплекс с Н4Н2636Р, показаны в таблице 16. Лишь один пептид, с аминокислотами 15-24 в FELD1B-A-mmH, характеризовался повышением центроидных значений m/z>0,3 m/z для состояния обмен в растворе/обратный обмен на гранулах по сравнению с состоянием обмен на гранулах/обратный обмен на гранулах, что указывало на то, что этот сегмент был защищен от полного обратного обмена в результате связывания Н4Н2636Р. Отличия центроидных значений более чем на 0,3 m/z в таблице 16 выделены жирным шрифтом. Таким образом, аминокислоты в таком участке 15-24 (SEQ ID NO: 412), исходя из результатов, полученных по способу обмена H/D, включали эпитоп для связывания антитела Н4Н2636Р с белком Fel d1B-A-mmH.
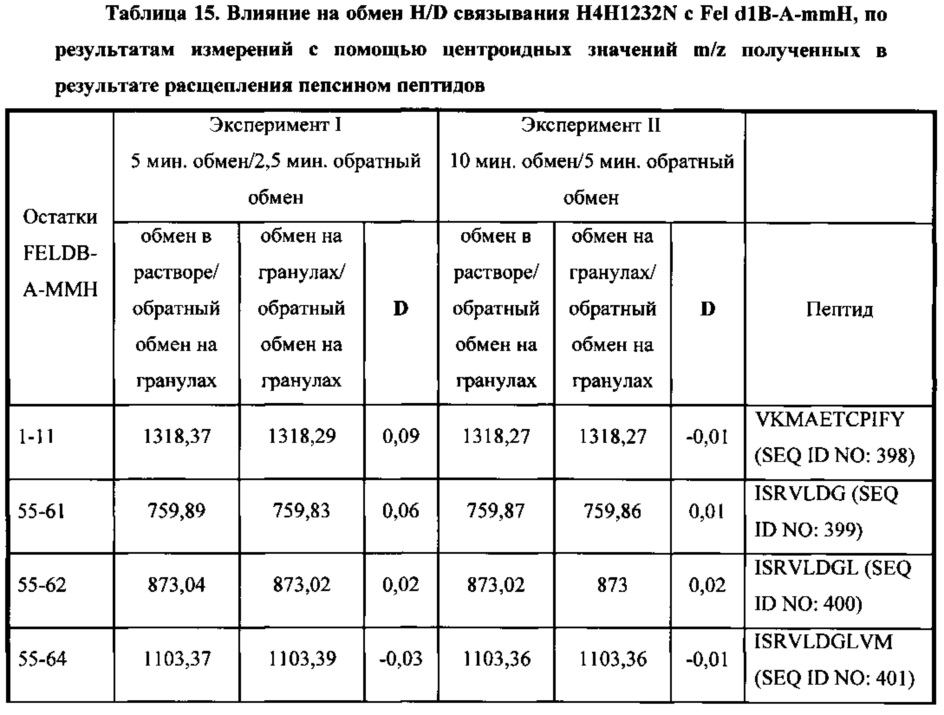
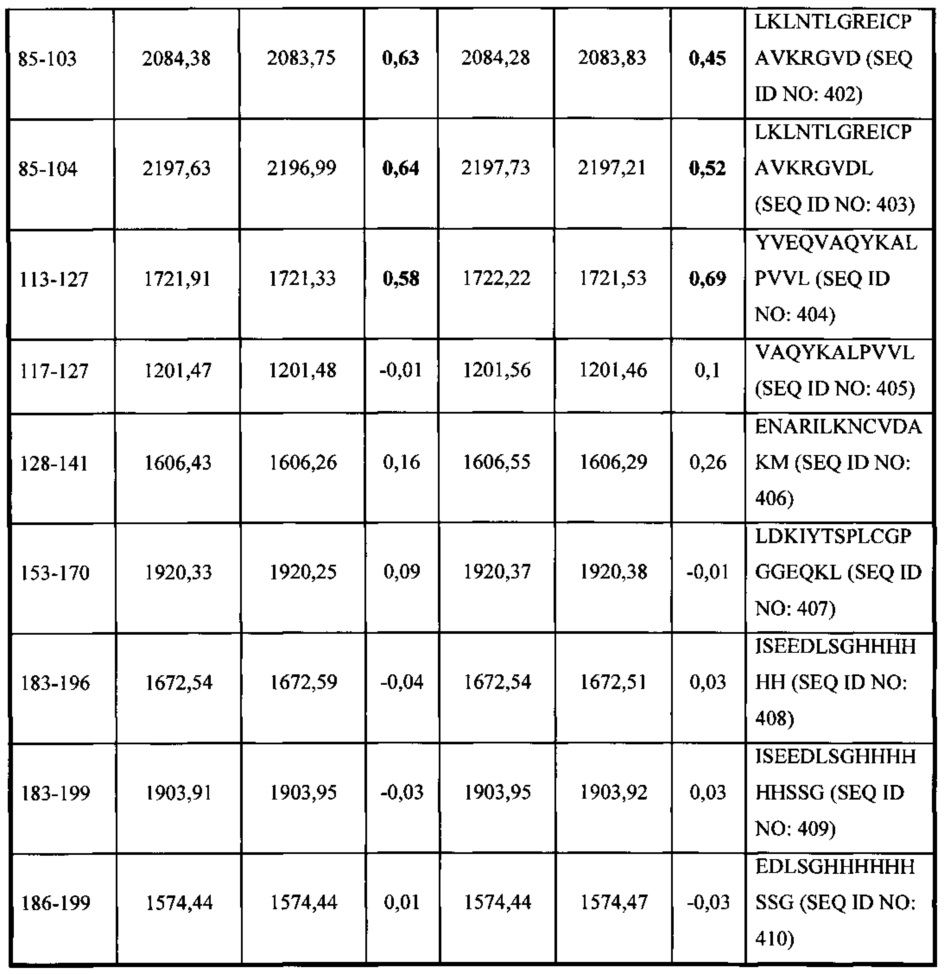
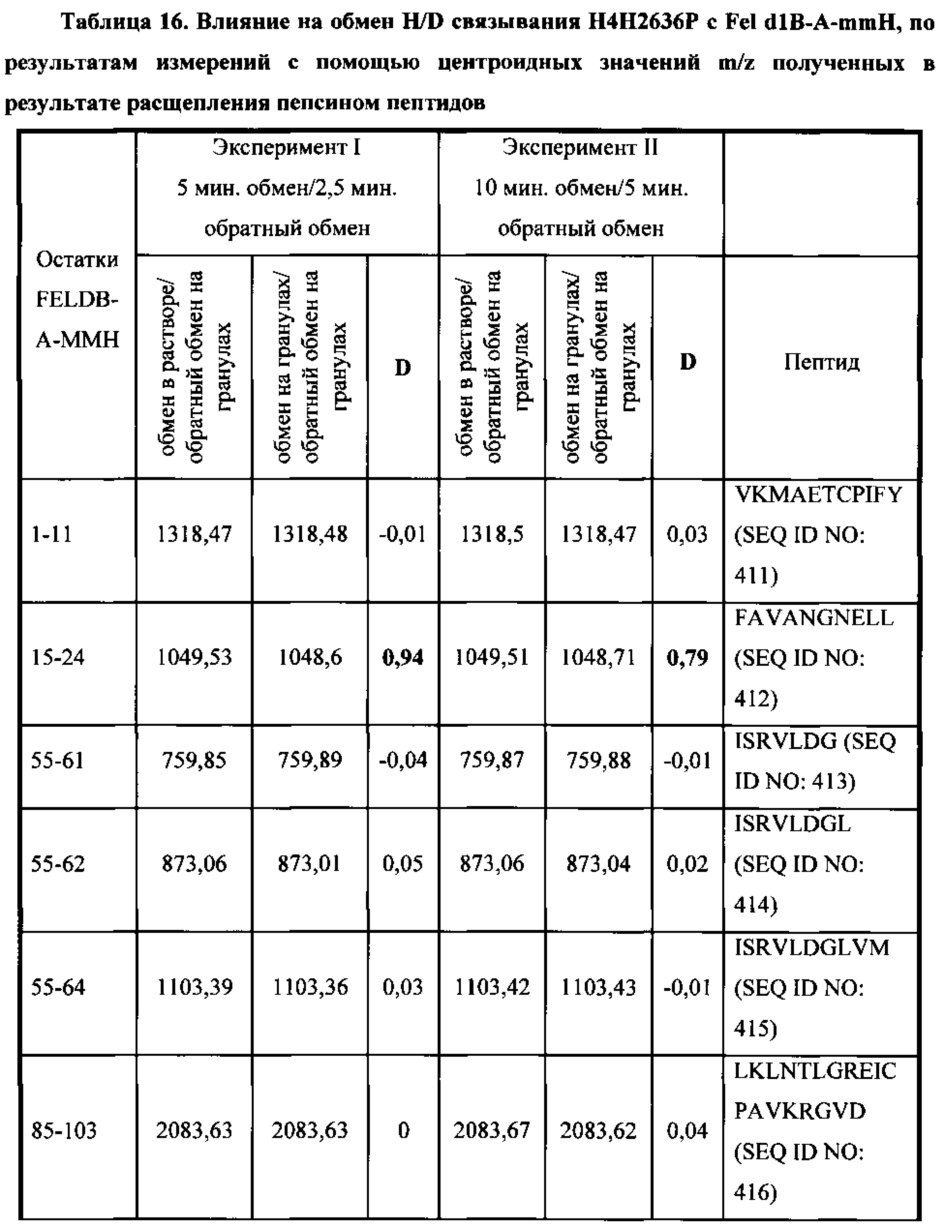
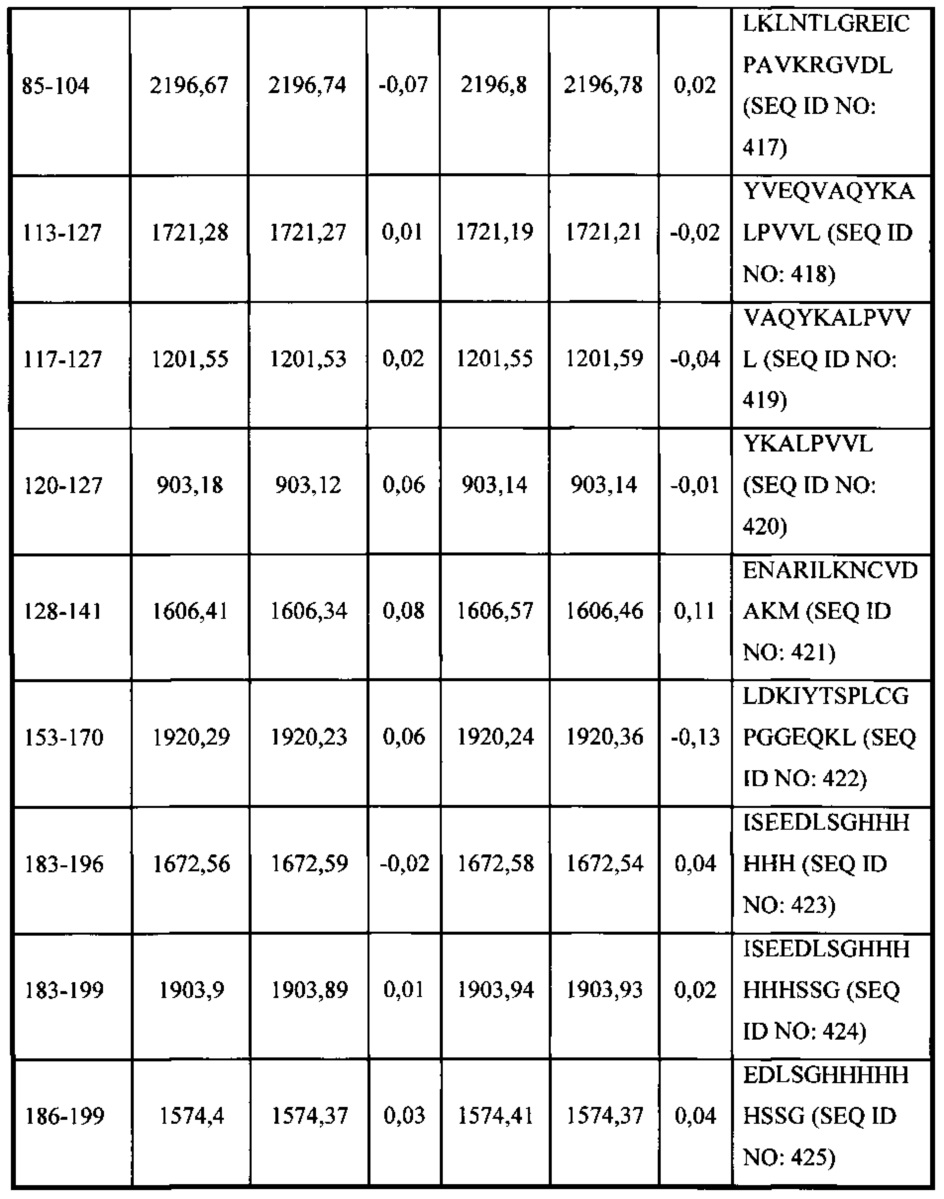
Пример 9: Создание биспецифических антител
Описание полученных биспецифических антител к Fel d1
Биспецифические антитела, содержащие домены связывания тяжелой и легкой цепи из пары некоторых описанных в настоящем изобретении антител к Fel d1, строили с помощью стандартных методик. Антитела к Fel d1, использованные для построения биспецифических антител настоящего примера, получали в результате иммунизации мыши VelocImmune® первичным иммуногеном, таким как полноразмерный природный Fel d1, который доступен на рынке для приобретения (например, от компании Indoor Biotechnologies, № LTN-FD1-1), или выделяли из кошачьей шерсти или перхоти путем многоэтапной колоночной хроматографии (см., например, работу Chapman MD, et al. (1988), J. Immunol. 140:812-818), или который можно получить рекомбинантно (см. номера доступа в GenBank Р30438 или NP_001041618.1 для полноразмерной аминокислотной последовательности цепи 1 для Fel d1 (также называемой цепью А или FELD1 А; см. также SEQ ID NO: 392) и номер доступа в GenBank Р30440 или NP_001041619.1 для полноразмерной аминокислотной последовательности цепи 2 для Fel d1 (также называемой цепью В или FELD В; см. также SEQ ID NO: 393), или фрагменты либо цепи 1, либо цепи 2, или фрагменты как от цепи 1, так и от цепи 2 белка Fel d1, с последующей иммунизацией вторичным иммуногеном или иммуногенно активным фрагментом природного белка. В соответствии с одним вариантом осуществления использованный иммуноген проиллюстрирован в SEQ ID NO: 394 (последовательная гибридизация цепь 2 Fel d1 - цепь 1-mFc) или SEQ ID NO: 395 (гибридизация цепи 1 Fel d1 с помощью линкера и цепи 2 -mFc).
Биспецифические антитела, полученные в соответствии с настоящим примером, содержали два антигенсвязывающих домена (т.е. "плечи связывания 1 и 2").
Одно из биспецифических антител, обозначенное H4H3467D, содержало общую легкую цепь каппа на обоих Fab-плечах, полученных из антитела Н4Н2864Р (SEQ ID NO: 378). В одном Fab-плече H4H3467D задействован вариабельный участок тяжелой цепи (VH) из антитела Н4Н2864Р (SEQ ID NO: 370), в то время как в другом Fab-плече задействован VH участок из H4H1232N (SEQ ID NO: 18).
Второе биспецифическое антитело по настоящему изобретению, обозначаемое H4H8751D, содержит общую легкую цепь каппа на обоих Fab-плечах, полученных из антитела Н4Н2636Р (SEQ ID NO: 314). В одном Fab-плече H4H8751D задействован VH участок из Н4Н2636Р (SEQ ID NO: 306), в то время как в другом Fab-плече задействован VH участок из H4H1232N (SEQ ID NO: 18).
В приведенной ниже таблице 17 изложены составные части антигенсвязывающих доменов двух биспецифических антител, полученных в соответствии с примером 9. Также в таблице 17 приведены идентификаторы аминокислотных последовательностей для различных вариабельных участков тяжелой цепи и легкой цепи, которые были получены из исходных антител (использованных для получения биспецифических антител).
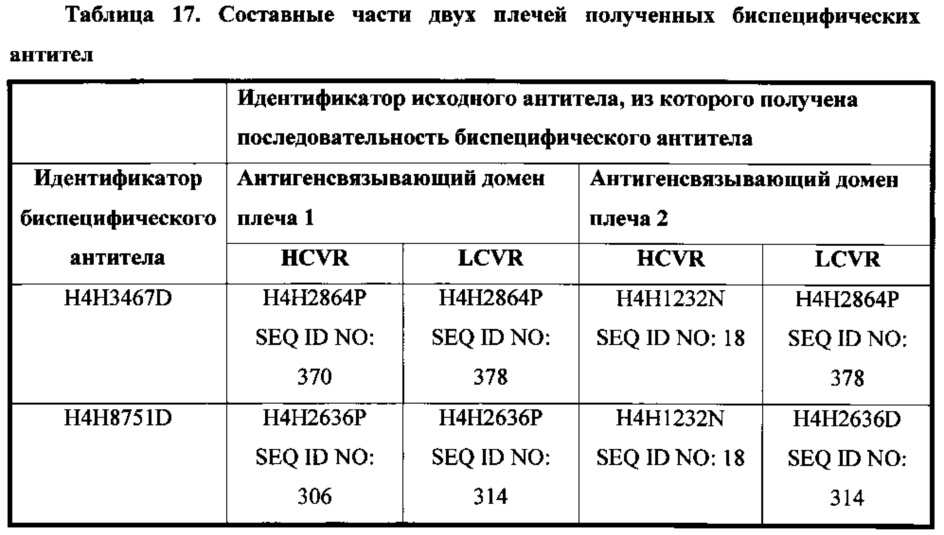
В приведенных ниже таблицах 18А и 18В изложены идентификаторы аминокислотных последовательностей для различных вариабельных участков тяжелой цепи (таблица 18А) и вариабельных участков легкой цепи (таблица 18В) и последовательности их соответствующих гипервариабельных участков (CDR) для двух описанных в настоящем документе биспецифических антител.
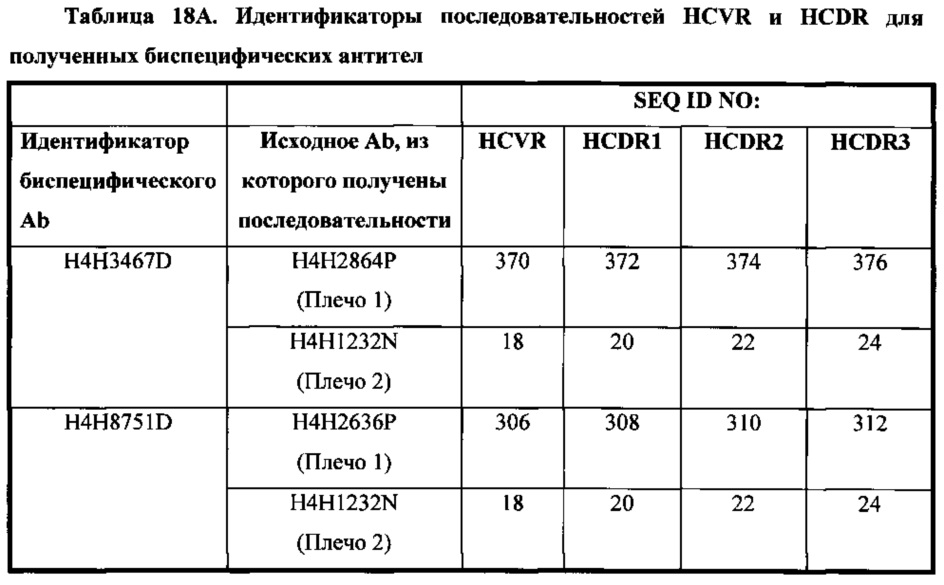
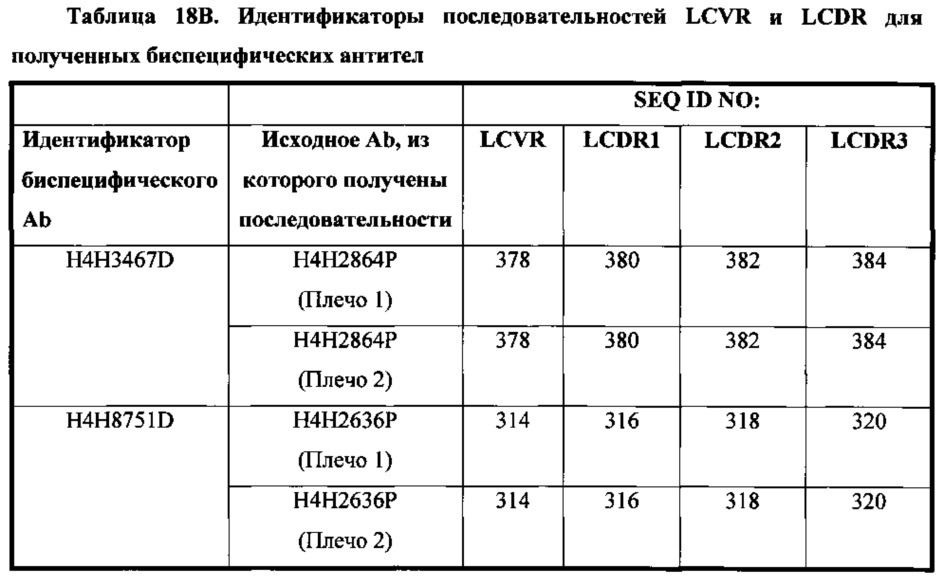
Анализ Biacore биспецифических антител для определения величин ассоциации и диссоциации
Константы скорости ассоциации и диссоциации связывания (ka и kd, соответственно), равновесные константы диссоциации и периоды полудиссоциации (KD и t1/2, соответственно) для связывания природного Fel d1 (далее обозначаемого nFel d1) с очищенными моноспецифическими и биспецифическими антителами к Fel d1 определяли с помощью анализа с применением биосенсора на основе технологии поверхностного плазмонного резонанса в режиме реального времени на приборе Biacore 2000. На чипе СМ5 с помощью химической реакции EDC-NHS поверхность биосенсора Biacore дериватизировали моноклональным мышиным антителом к человеческому Fc (GE, № BR-1008-39) для захвата моноспецифических и биспецифических антител к Fel d1. Все исследования по связыванию Biacore проводили при 25°С в HBSP+ подвижном буфере (0,01 M HEPES, pH 7,4, 0,15 M NaCl, 3 мМ CaCl2, 3 мМ MgCl2, 0,05% об./об. Surfactant Р20). На поверхность с захваченным антителом к Fel d1 со скоростью потока, составлявшей 50 мкл/мин. вводили различные концентрации nFel d1 (Indoor Biotech, № NA-FD1-2) (варьировавшие в диапазоне от 600 нМ до 2,34 нМ, с 6-кратным разведением), приготовленные в HBSP+ подвижном буфере. Ассоциацию nFel d1 с захваченными моноклональными антителами отслеживали в течение 4 минут, а диссоциацию nFel d1 в HBSP+ подвижном буфере отслеживали в течение 7 минут. Константы скорости реакции ассоциации (ka) и диссоциации (kd) определяли путем приближения сенсограмм в реальном времени к 1:1 модели связывания с ограничением массопереноса с помощью программного обеспечения для приближенного изображения функций Scrubber 2.0 с. Затем равновесные константы диссоциации связывания (KD) и периоды полудиссоциации (t1/2) рассчитывали из констант скорости реакции следующим образом: KD(M)=kd/ka и t1/2 (мин.)=[ln2/(60*kd)].
Константы скорости реакции связывания nFel d1 с различными моноспецифическими и биспецифическими антителами к Fel d1 при 25°С приведены в таблице 19. Три моноспецифических антитела к Fel d1 связывались с nFel d1 со значениями KD, варьирующими от 155 пМ до 1,6 нМ. Два биспецифических антитела к Fel d1, H4H3467D и H4H8751D, связывались с nFel d1 со значениями KD, составлявшими 250 пМ и 347 пМ, соответственно.
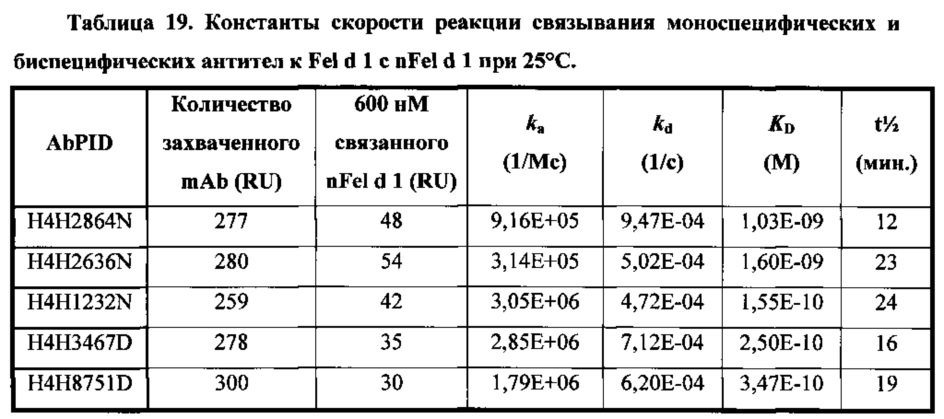
Для определения in vivo эффективности биспецифических антител к Fel d1 по сравнению с их моноспецифическими исходными антителами такие антитела одновременно с антителом изотипического контроля тестировали в in vivo модели РСА с применением природного Fel d1 как для сенсибилизации, так и для введения провокационной дозы, что было описано ранее (см. пример 6). Антитела в настоящем исследовании вводили в концентрации 1 мг/кг суммарного антитела (использовали 0,5 мг/кг каждого антитела при одновременном введении двух антител) с использованием 8 мышей на экспериментальную группу. Данные для каждой экспериментальной группы, выраженные как процент уменьшения проникновения красителя из сосудов ±SD, показаны в таблице 20.
Моноспецифические антитела H4H1232N и Н4Н2864Р соответственно вызывали 67 (±26)% и 81 (±26)% уменьшение проникновения красителя из сосудов. Комбинация моноспецифических антител H4H1232N и Н4Н2864Р вызывала 98 (±3,5)% уменьшение проникновения красителя из сосудов, в то время как биспецифическое антитело H4H3467D, состоящее из моноспецифических антител, H4H1232N и Н4Н2864Р, вызывало 93 (±11)% уменьшение проникновения красителя из сосудов.
В другом эксперименте моноспецифические антитела H4H1232N и Н4Н2636Р соответственно вызывали 64 (±33)% и 8,7 (±79)% уменьшение проникновения красителя из сосудов. Комбинация моноспецифических антител H4H1232N и Н4Н2636Р вызывала 90 (±15)% уменьшение проникновения красителя из сосудов, в то время как биспецифическое антитело H4H8751D, состоящее из моноспецифических антител, H4H1232N и Н4Н2636Р, вызывало 77 (±20)% уменьшение проникновения красителя из сосудов.
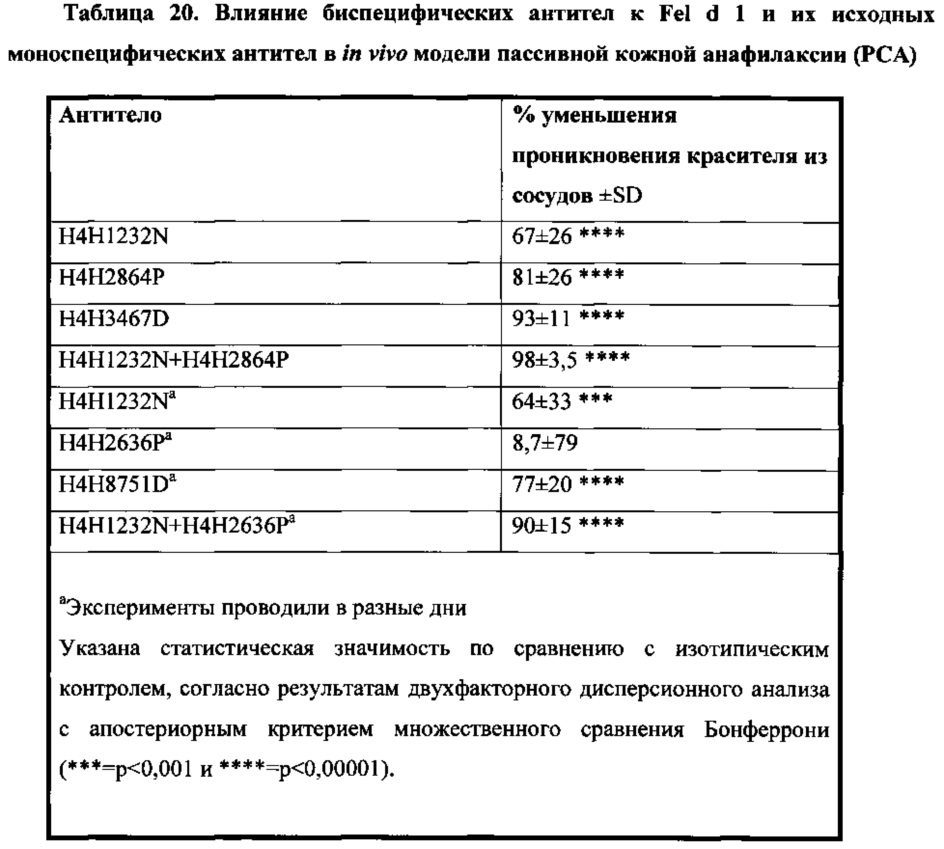
Реферат
Настоящее изобретение относится к области биохимии, а именно к антителу, которое связывается с кошачьим аллергеном, Fel d1, композициям, содержащим указанное антитело, нуклеиновой кислоте, его кодирующей, а также к способу применения указанного антитела. Антитела по настоящему изобретению также могут быть пригодны для диагностических целей, для определения, имеет ли больной аллергию на кошачий аллерген Fel d1. Изобретение позволяет предупреждать высвобождение гистамина или других медиаторов воспаления из тучных клеток или базофилов, таким образом предупреждая или ослабляя нежелательные эффекты, наблюдаемые у больных, сенсибилизированных к кошачьему аллергену. 18 н. и 36 з.п. ф-лы, 20 табл., 9 пр.
Комментарии