Химерный белок боррелии, нуклеиновая кислота, кодирующая такой белок, экспрессирующая кассета, вектор, способ и набор для диагностики лайм-боррелиоза, вакцина для профилактики боррелиоза - RU2549698C2
Код документа: RU2549698C2
Чертежи

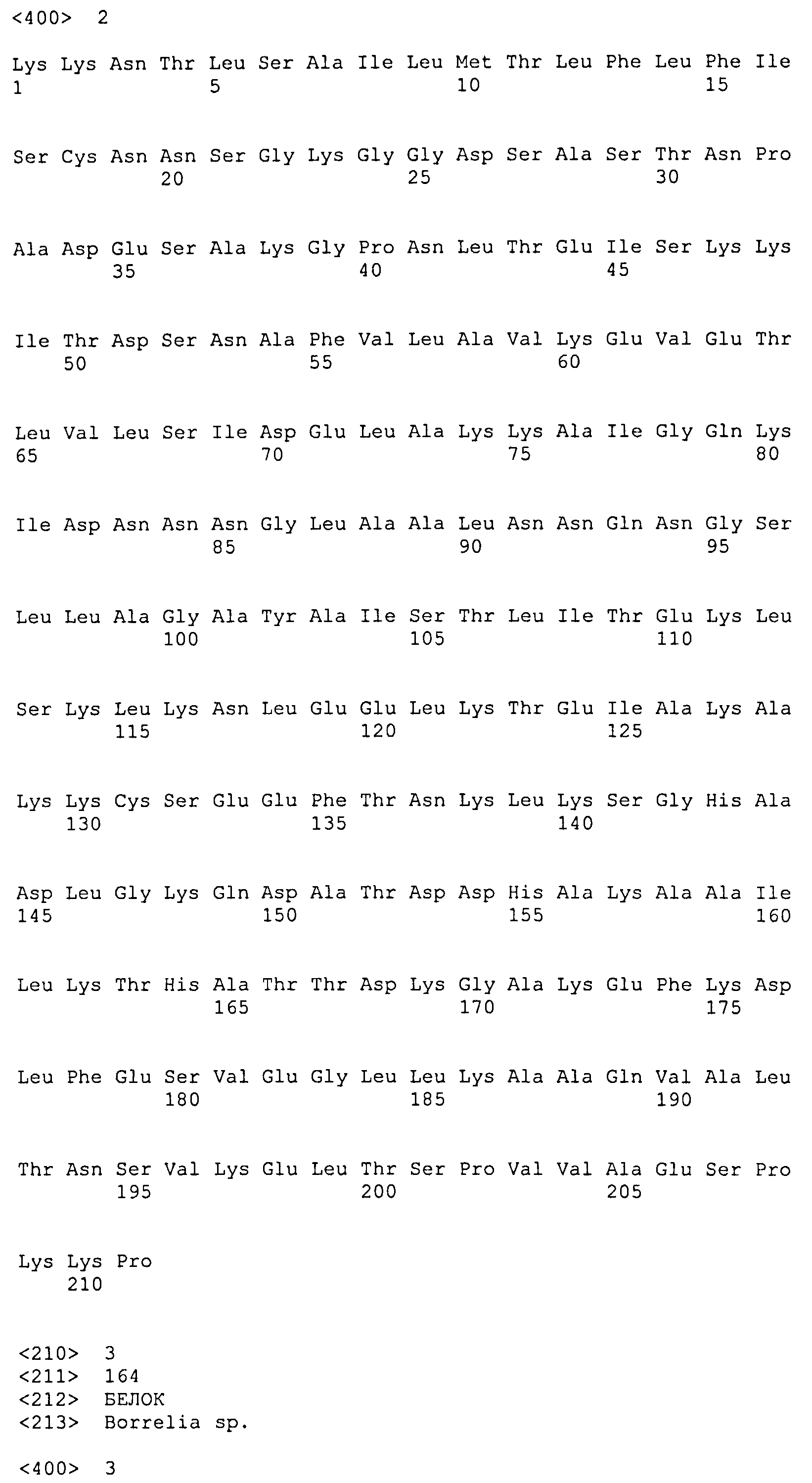



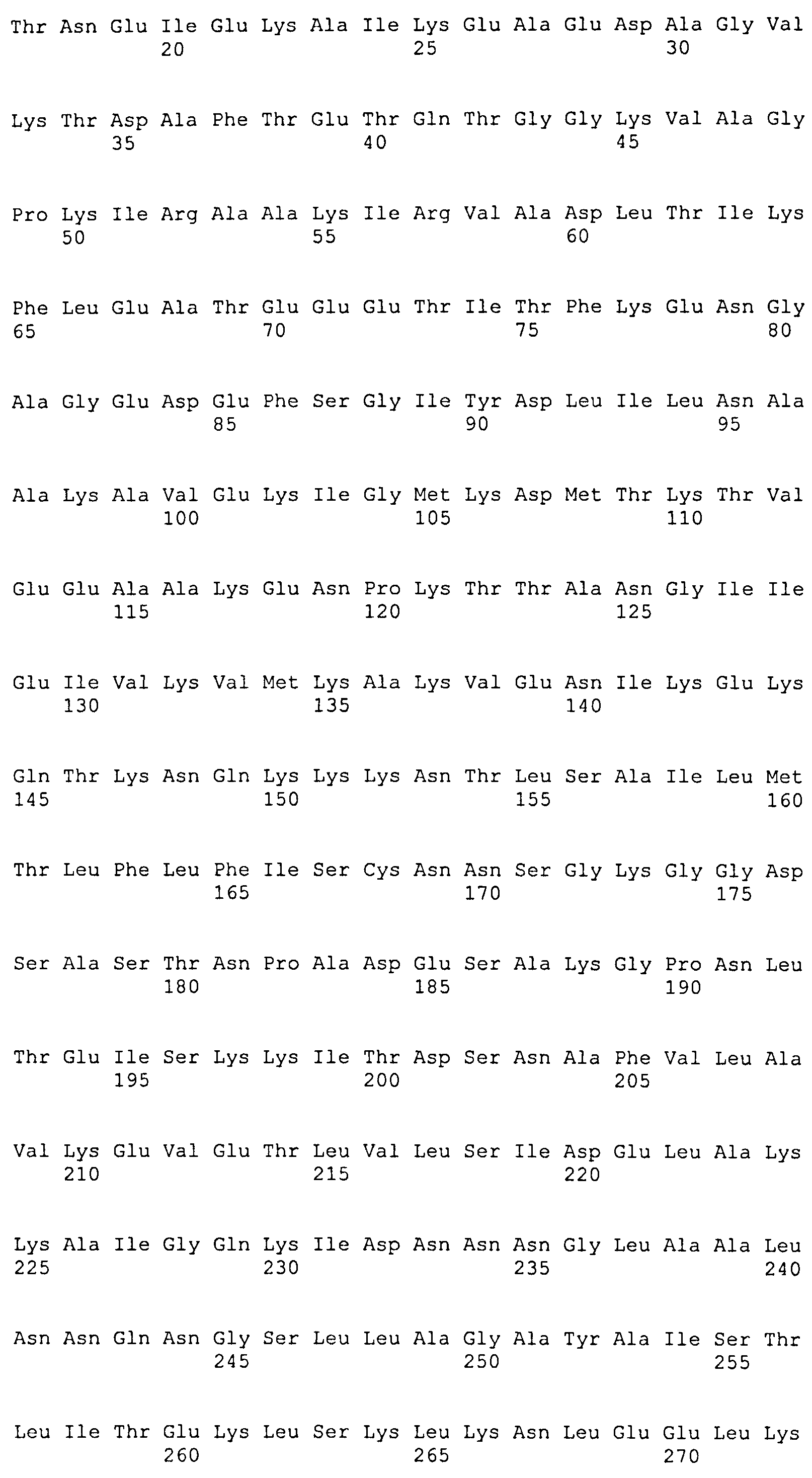

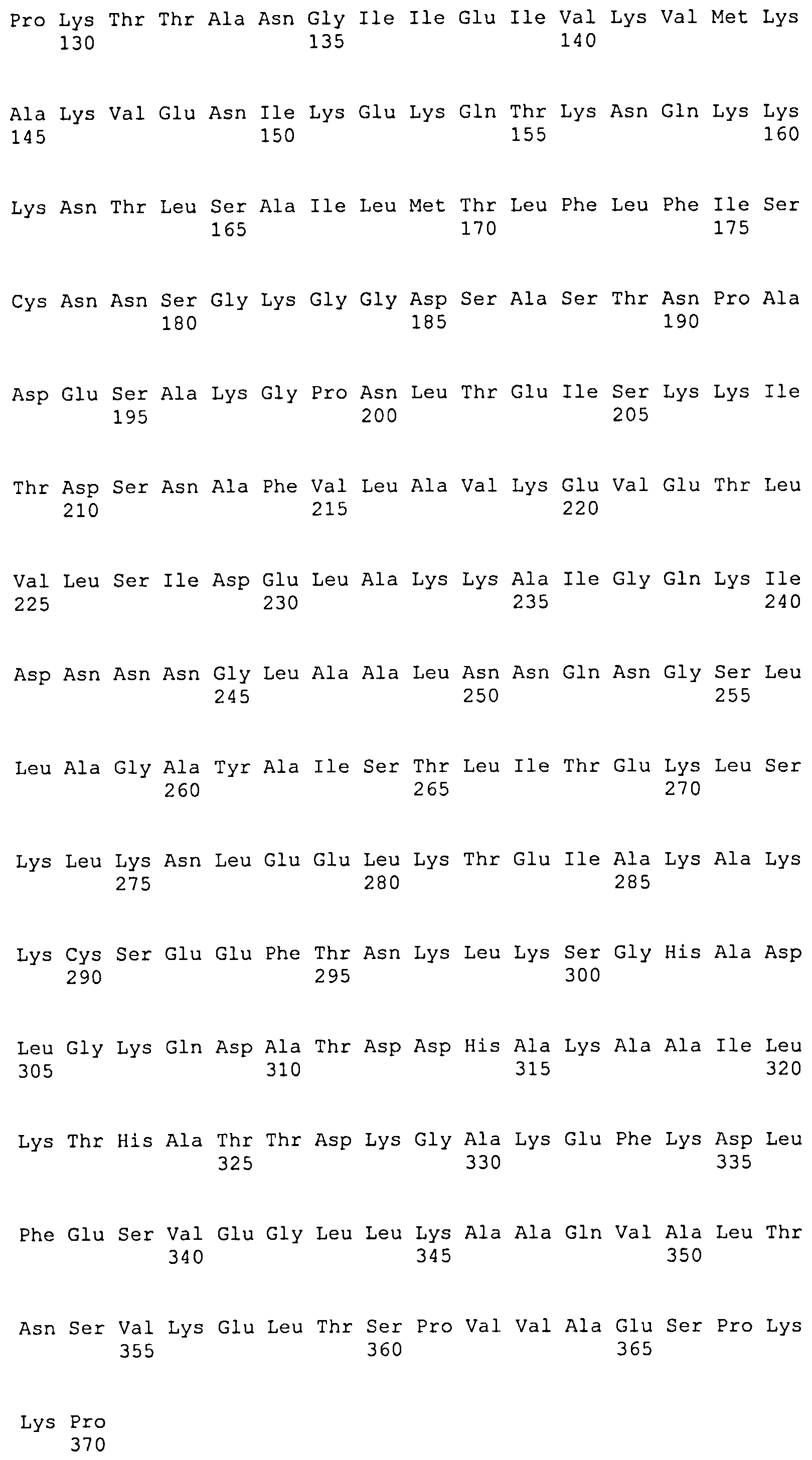
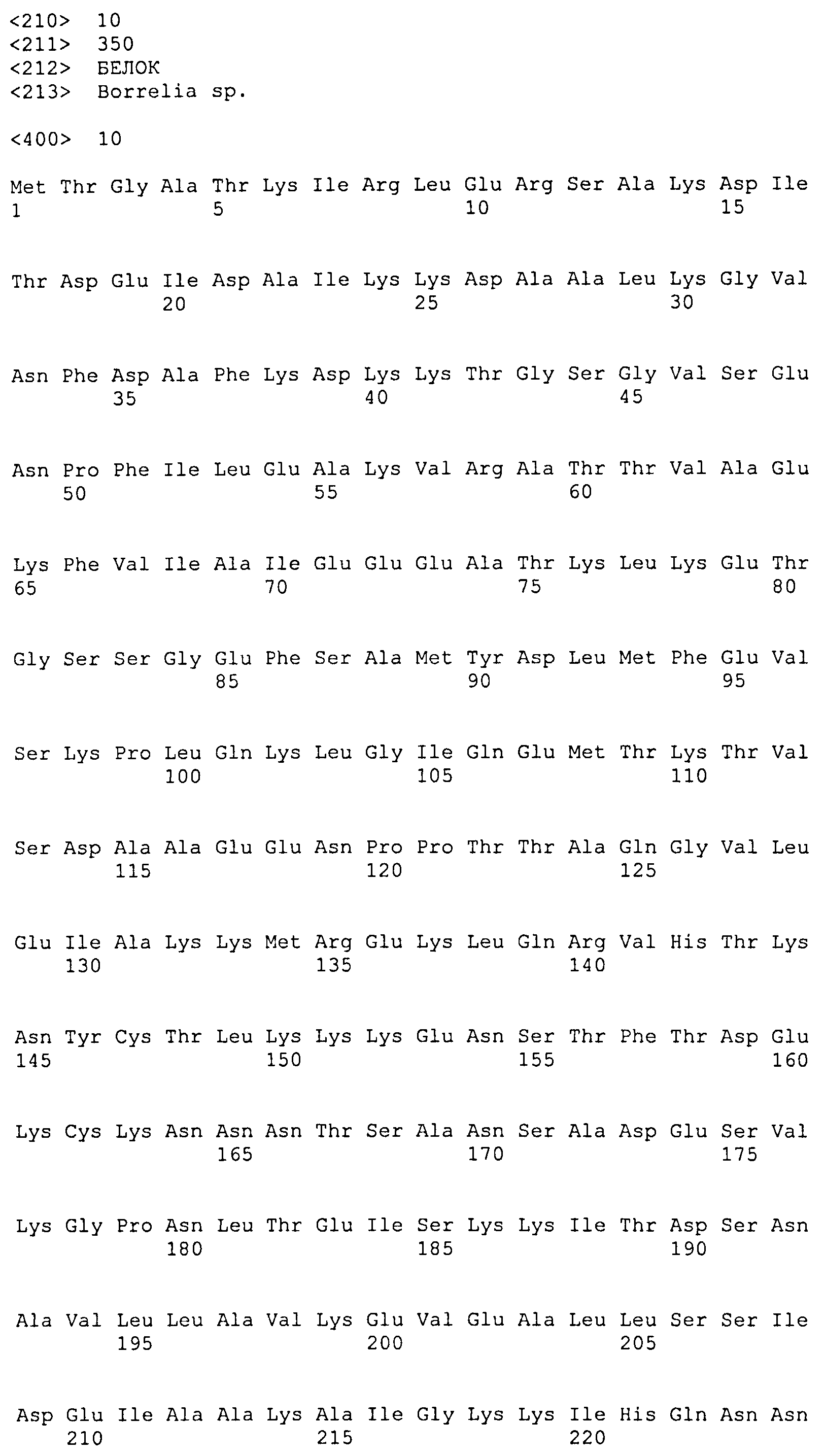

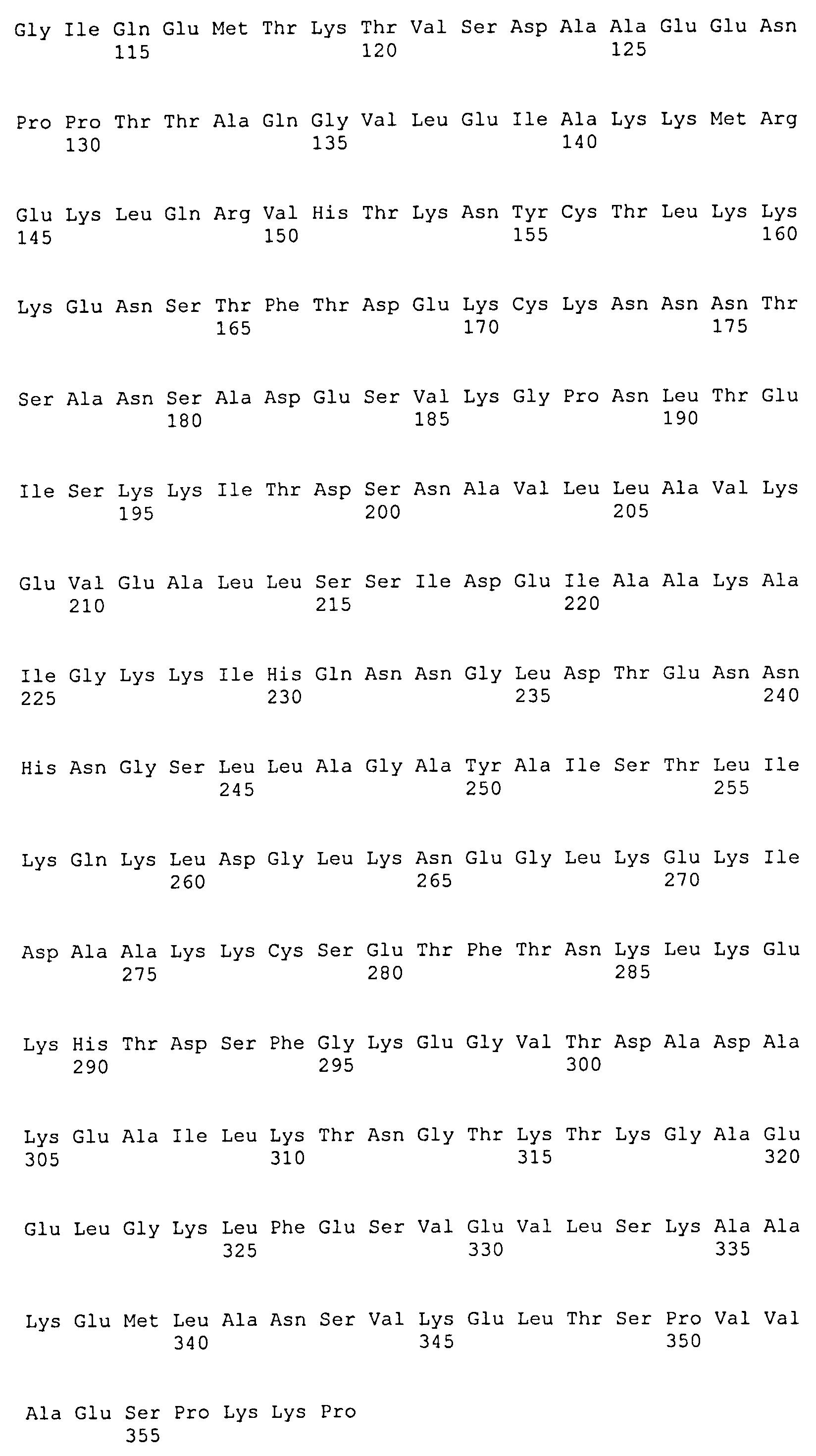

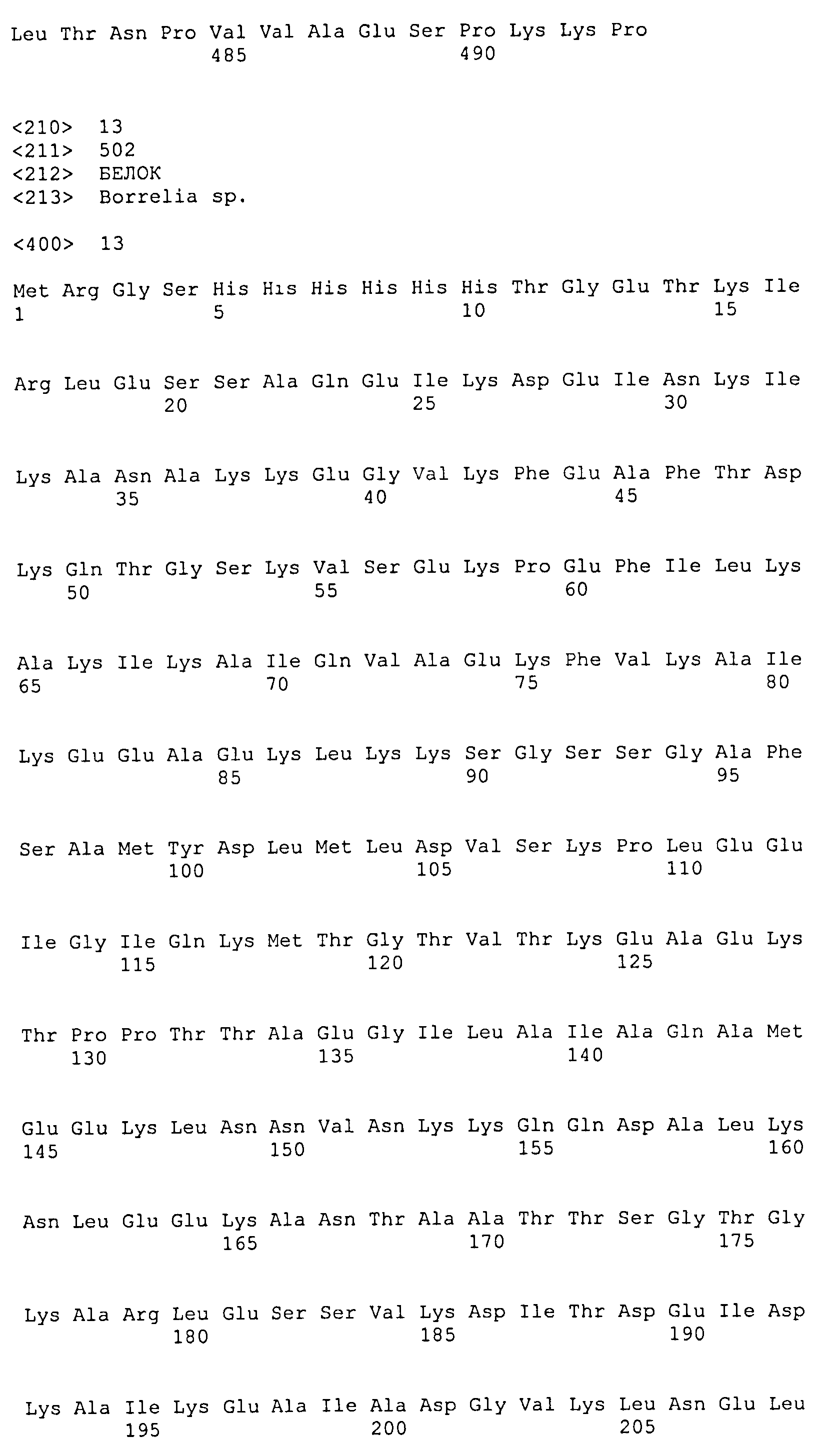











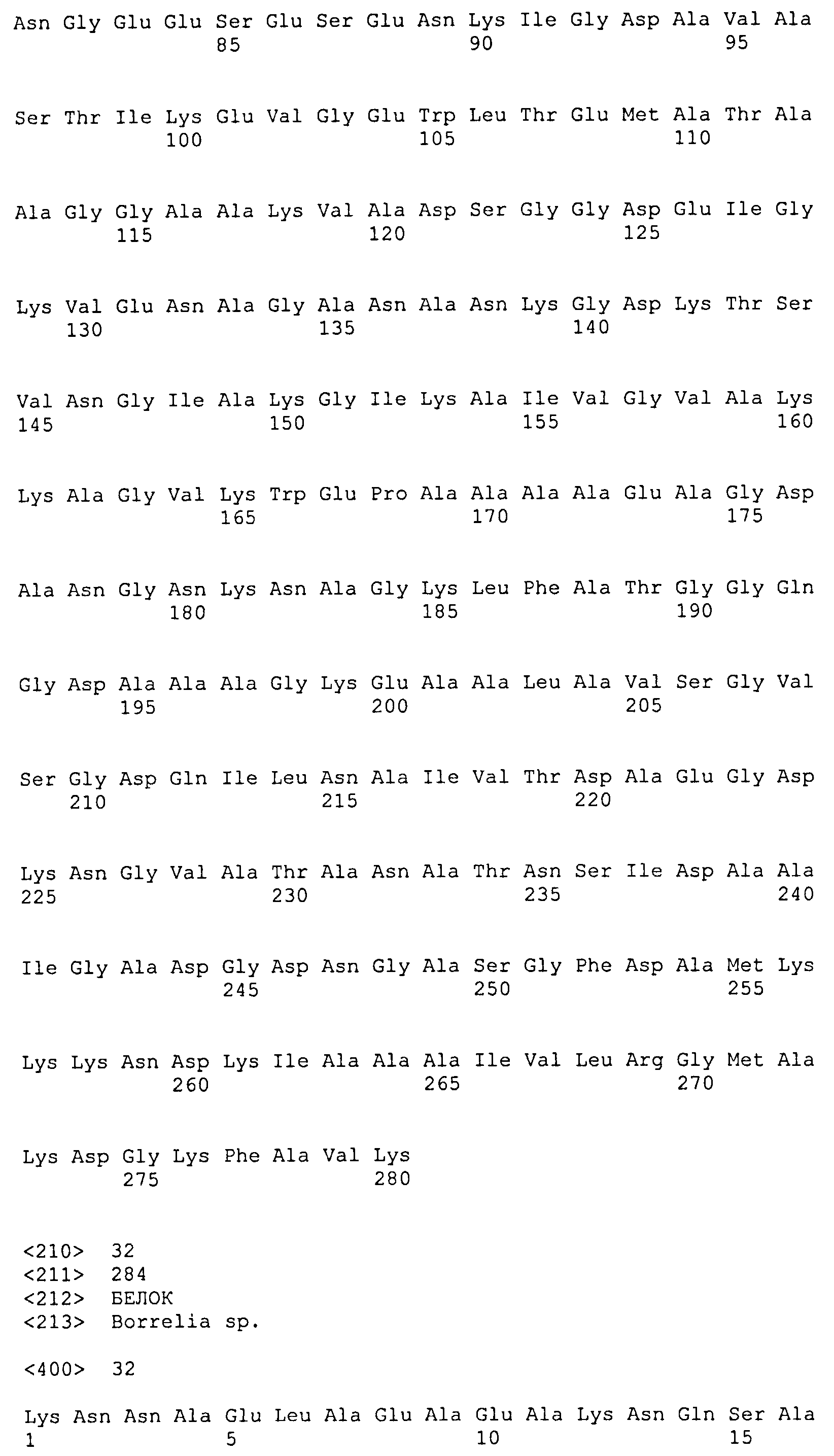
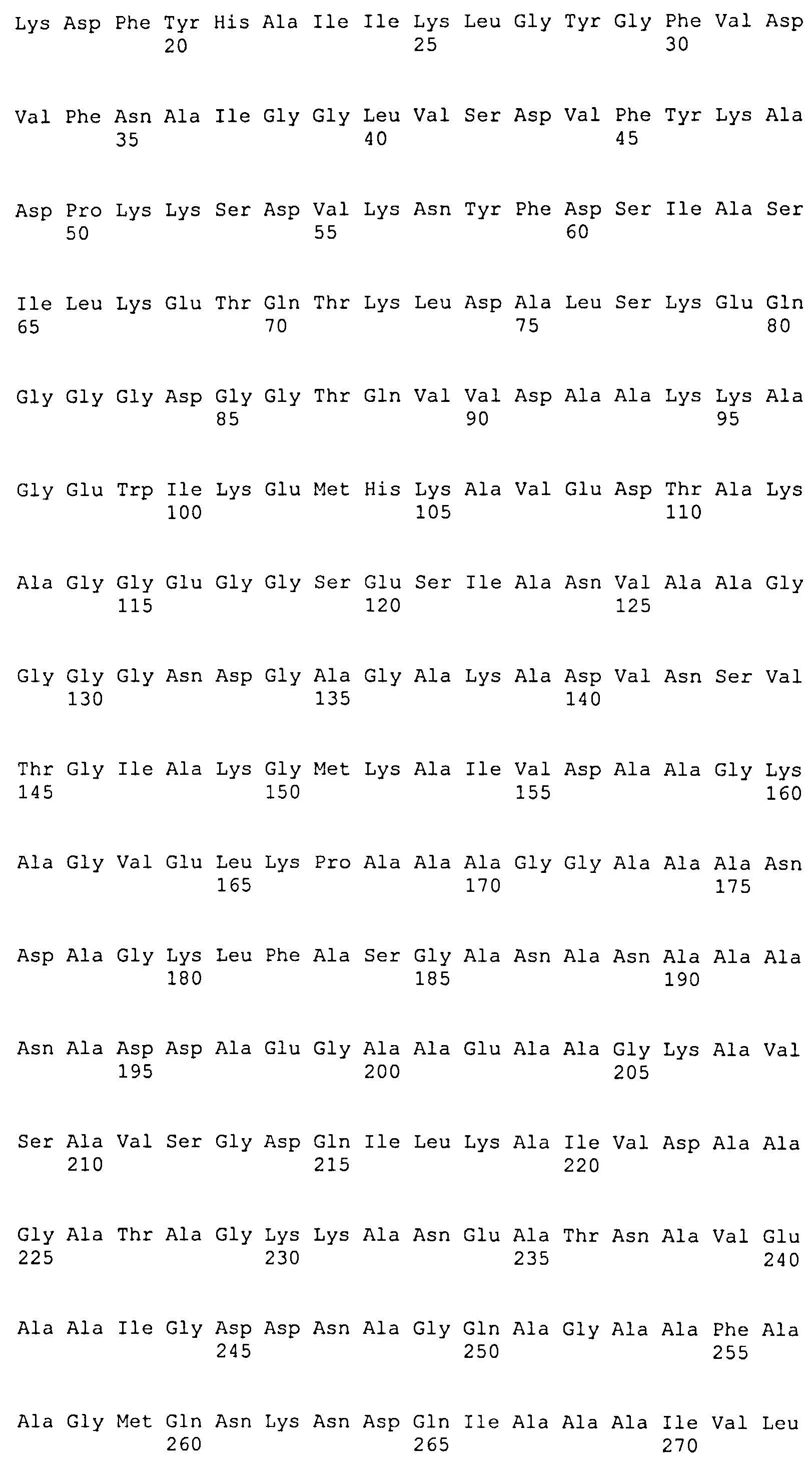
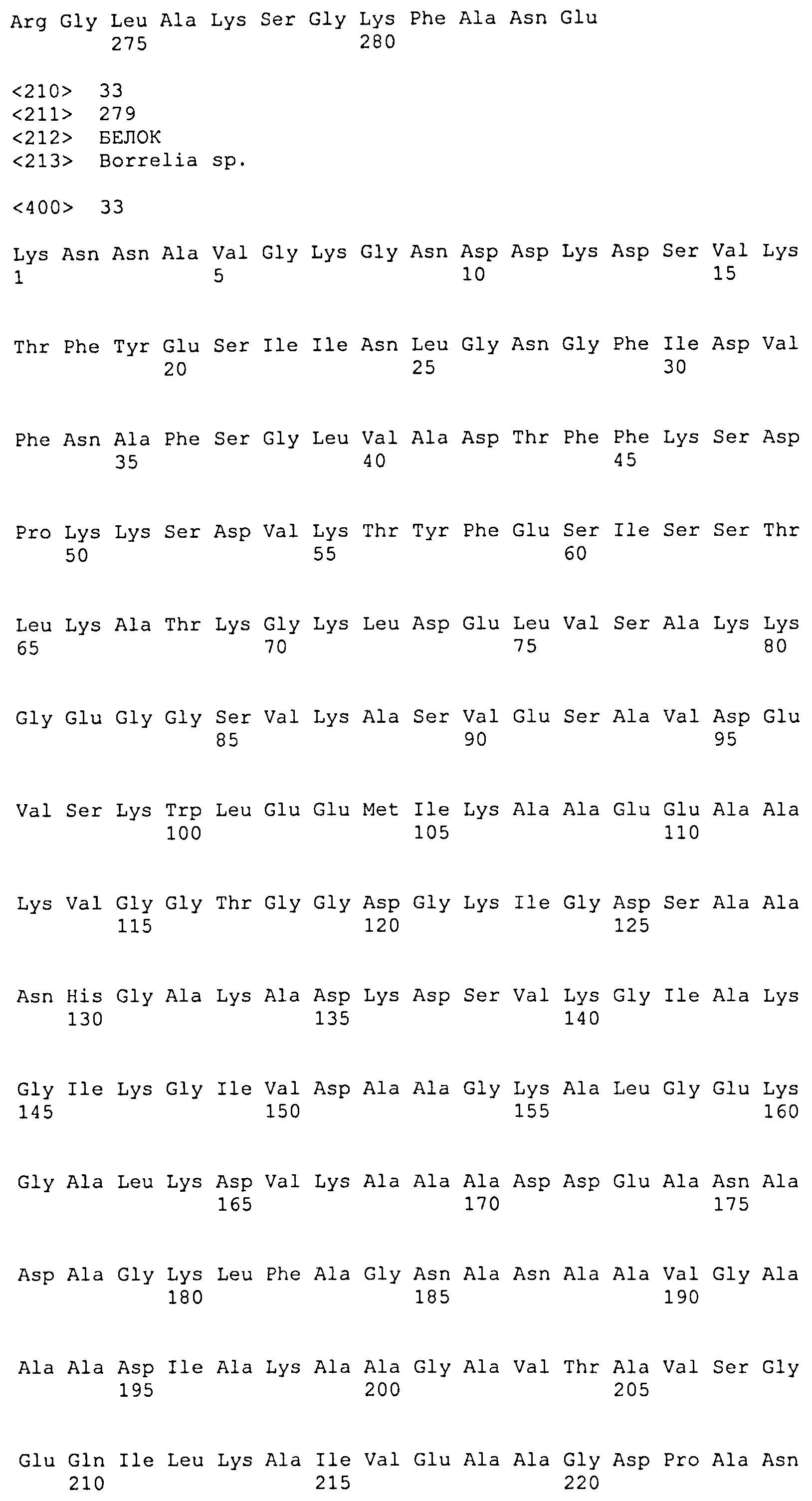



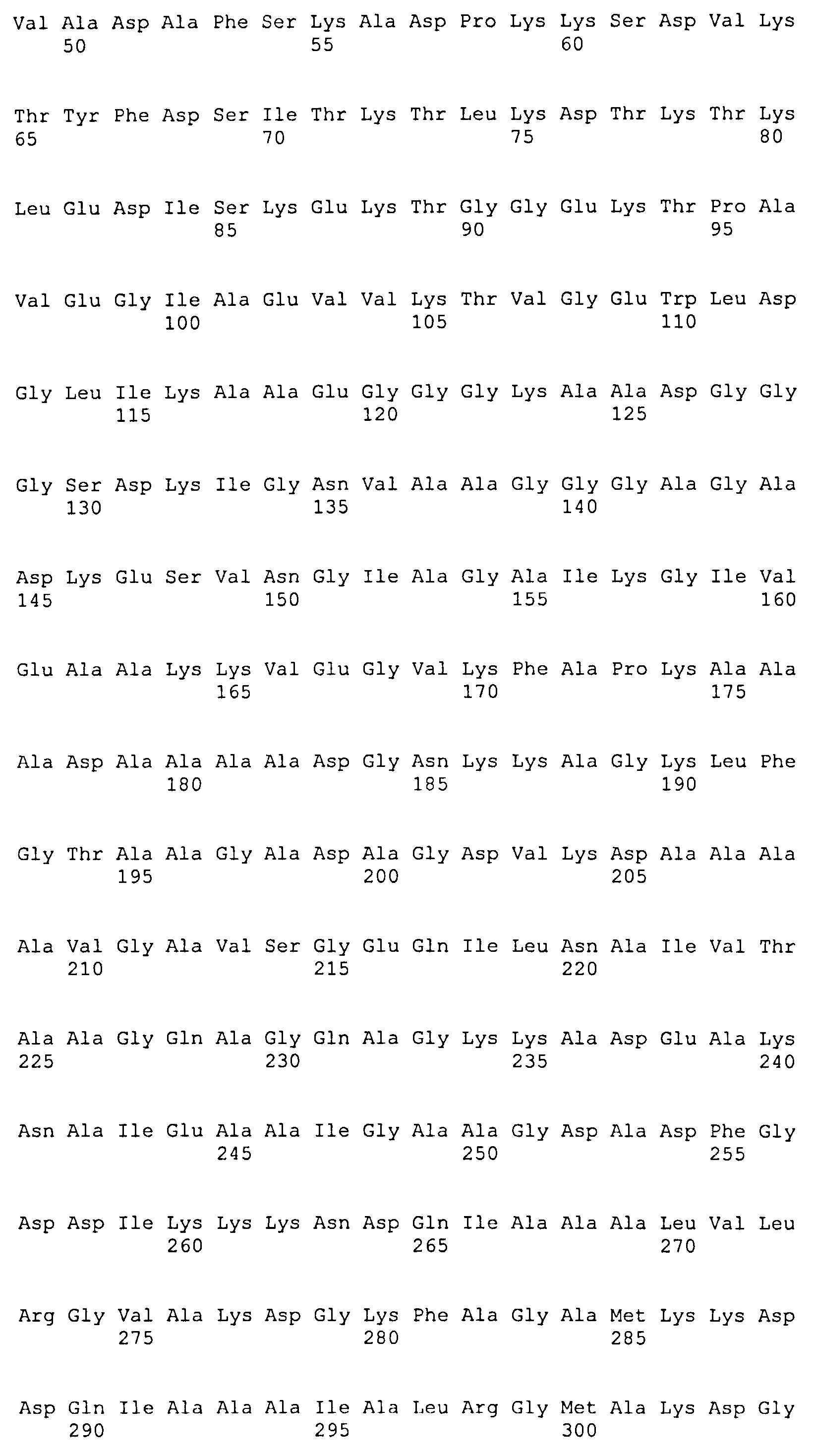




Описание
Лайм-боррелиоз (LB) представляет собой неконтагиозное инфекционное заболевание, вызываемое спирохетой, называемой Borrelia burgdorferi, и передаваемое человеку при укусе клеща семейства иксодовых клещей. LB при отсутствии лечения вызывает различные патологические нарушения (дерматологического, артритического, сердечного, неврологического и иногда офтальмологического характера). Он представляет собой трансмиссионное инфекционное заболевание, наиболее часто встречающееся в США и в некоторых странах северного полушария с умеренным климатом.
В этой инфекции участвуют несколько видов боррелий, в настоящее время обозначаемых как группа burgdorferi или Borrelia burgdorferi sensu lato (включающая в себя Borrelia burgdorferi sensu stricto, B. garinii и B. afzelii). Эти виды являются патогенными для человека.
В США участвующий инфекционный вид представляет собой Borrelia burgdorferi sensu stricto. В Европе к этому виду добавляются B. garinii и B. afzelii. В Азии участвующие виды представляют собой B. garinii и B. afzelii.
В США отмечают приблизительно 10000 случаев в год. В Европе частота появления составляет менее 5 случаев на 100000.
Лайм-боррелиоз эволюционирует, проходя три различные стадии от ранней стадии инфицирования до поздней. Ранняя стадия (стадия I) может быть бессимптомной или выражаться псевдогриппозной картиной. В 50-80% случаев через несколько дней после укуса клеща на коже отмечается появление воспаленной сыпи очень специфического внешнего вида, называемой мигрирующей эритемой (EM). При отсутствии лечения гематогенная диссеминация боррелий проявляется спустя несколько недель во внезапном возникновении воспалительных артритов, неврологических (нейроборрелиоз) и менингеальных поражений, проявлений на коже и в деятельности сердца (стадия II). Через несколько месяцев или лет болезнь переходит в атрофическую хроническую форму, энцефалопатию, энцефаломиелит и хронический артрит (стадия III).
Для каждого вида Borrelia burgdorferi существует особый органический тропизм. Если первая стадия мигрирующей эритемы неясно связана с тремя видами, то переход в неврологическую форму предпочтительно связан с видом B. garinii, артриты чаще связаны с B. burgdorferi sensu stricto, атрофический хронический акродерматит специфичен для B. afzelii.
Сходство клинических симптомов между лайм-боррелиозом и другими не связанными с ним болезнями, а также изменчивость проявлений затрудняет клиническую диагностику. Если доказательства анамнеза отсутствуют (укус клеща или EM), то диагностика боррелиоза на основании клинических наблюдений может быть особенно затрудненной. Ранняя стадия болезни может быть бессимптомной до момента, когда она достигнет очень далеко зашедших клинических стадий.
Именно поэтому диагностика LB основана как на клинических признаках, так и на обнаружении в сыворотке антител, специфических для патогенных Borrelia burgdorferi, чаще всего способом ELISA (Enzyme Linked ImmunoSorbent Assay (ферментный иммуносорбентный тест)) или EIA, IFA. Антитела IgM анти-Borrelia burgdorferi появляются, в общем случае, через несколько дней или недель после начала инфицирования и могут сохраняться в течение развития болезни. Ответ IgG возникает позже. У преобладающей части больных IgG обнаруживаются приблизительно через месяц после начала активного инфицирования, и такие IgG также могут сохраняться в течение многих лет после начального воздействия и исчезновения симптомов.
В Европе оценка серологического ответа осложнена в силу существования трех патогенных видов и межвидовой изменчивости в отношении главных иммунодоминантных антигенов. Антигены, используемые в настоящее время в стандартной программе обнаружения IgG и IgM LB, представляют собой обработанные ультразвуком клеточные образцы Borrelia burgdorferi sensu lato. Результаты серологических исследований с данными антигенами в высокой степени вариативны в отношении специфичности и чувствительности. Таким образом, по причине недостаточной специфичности, вовлекающей перекрестную реактивность с антителами, связанными с различными патогенными бактериями, в частности с Treponema pallidium (этиологический агент сифилиса), спирохетами, риккетсиями, эрлихиями или Helicobacter pylori, диагностика образцов с положительным результатом в испытаниях ELISA должна быть подтверждена иммуноблоттингом. Чувствительность также представляет собой основной фактор. На практике, Borrelia burgdorferi sensu lato экспрессирует различные поверхностные белки вследствие адаптации к различным микросредам, так что генетическое различие и дифференциальная экспрессия генов Borrelia burgdorferi у больных имеют существенное значение для разработки серологических тестов LB. Липопротеин OspC (Outer-surface protein C (поверхностный белок C)) и белок DbpA (Decorin-binding protein A (декорин-связывающий белок A)) составляют часть таких белков. DbpA экспрессируется главным образом у млекопитающего после инфицирования. Данные белки обладают большой вариабельностью последовательностей в видах Borrelia burgdorferi и между видами. Белки DbpA являются особенно вариабельными и подразделяются на четыре группы: группа, соответствующая геновиду Borrelia afzelii, группа, соответствующая геновиду Borrelia sensu stricto, и две группы, соответствующие геновиду Borrelia burgdorferi garinii. Межвидовая идентичность последовательностей аминокислот между белками DbpA составляет только 40-44%. Для белков OspC она равна 54-72%.
Таким образом, требуется разработка набора, который соответствует требуемым критериям специфичности и чувствительности и, в частности, улучшает обнаружение IgM в отношении чувствительности в случае свежего инфицирования.
В настоящем изобретении предлагается устранить комплекс недостатков предшествующего уровня техники благодаря новым химерным рекомбинантным белкам, которые могут быть легко синтезированы и очищены и обладают сильной иммунореактивностью в отношении сыворотки больных, могущих быть инфицированными одним или несколькими патогенными видами Borrelia burgdorferi. Такие слитые химерные белки позволяют смягчить проблемы чувствительности и специфичности, связанные с присутствием нескольких патогенных видов Borrelia burgdorferi, с большой вариабельностью последовательностей поверхностных антигенов Borrelia burgdorferi и необходимостью применять несколько репрезентативных антигенов видов B. garinii, B. burgdorferi sensu stricto и B. afzelii, благодаря разработке диагностического теста на лайм-боррелиоз, основанного по меньшей мере на обнаружении антител анти-OspC и анти-DbpA.
Слитые химерные белки по настоящему изобретению позволяют в то же время устранить затруднения, возникающие вследствие экспрессии некоторых антигенов в рекомбинантной форме в повышенном количестве. На практике, несмотря на значительные работы по конструированию генов для обеспечения оптимизированной экспрессии их в E. coli, авторы изобретения впервые показали, что белки OspC слабо экспрессируются в рекомбинантной форме в E. coli, в то же время совершенно неожиданным образом было обнаружено, что белки DbpA могли экспрессироваться в тех же самых условиях в растворимой форме в условиях, не вызывающих денатурацию, и с более высоким выходом. Легкость экспрессии белков DbpA была использована для создания химерных белков, образующих DbpA и OspC, и авторы смогли показать, что химерные белки экспрессировались лучше, чем изолированные белки OspC, что было полностью неожиданным, поскольку никогда не было описано и даже не сделано предположение, что белки DbpA могли обладать свойствами слитых белков. В силу этого для улучшения уровней экспрессии авторами изобретения были сконструированы химерные белки DbpA-OspC за счет использования неожиданных свойств слияния белков DbpA для улучшения экспрессии и растворимости химерных белков. В молекулярной конструкции для экспрессии химерного белка по настоящему изобретению ген, кодирующий белок DbpA, предпочтительно встраивают в начале одного или нескольких генов, кодирующих один или несколько белков OspC. В зависимости от такой предпочтительной молекулярной конструкции химерный белок по настоящему изобретению на N-концевом участке содержит последовательность, принадлежащую белковой последовательности DbpA, а на C-концевом участке содержит последовательность, принадлежащую белковой последовательности OspC. Такой предпочтительный тип конструкции, помимо того, что он позволяет облегчать и оптимизировать экспрессию и растворимость химерного белка, имеет, кроме того, другое преимущество, состоящее в улучшении узнавания химеры антиборрелиозными антителами в силу лучшей демонстрации им иммунодоминантной области белка OspC. Дополнительное преимущество химерных белков по настоящему изобретению состоит в ограничении числа рекомбинантных белков в комплекте набора для диагностики лайм-боррелиоза. В то же время свойства слияния белка DbpA делают его превосходным кандидатом для экспрессии вакцинных химерных белков DbpA-OspC для профилактики инфицирования боррелиями. Следовательно, химерные белки по настоящему изобретению являются приемлемыми в качестве активного агента в профилактической вакцине против боррелиоза.
Таким образом, настоящее изобретение относится к слитому химерному белку DbpA-OspC боррелии, имеющему не природное происхождение, а являющемуся синтетическим, то есть полученным генной инженерией (рекомбинантный белок) или пептидным синтезом, причем упомянутый белок выбран из группы, в которую входят:
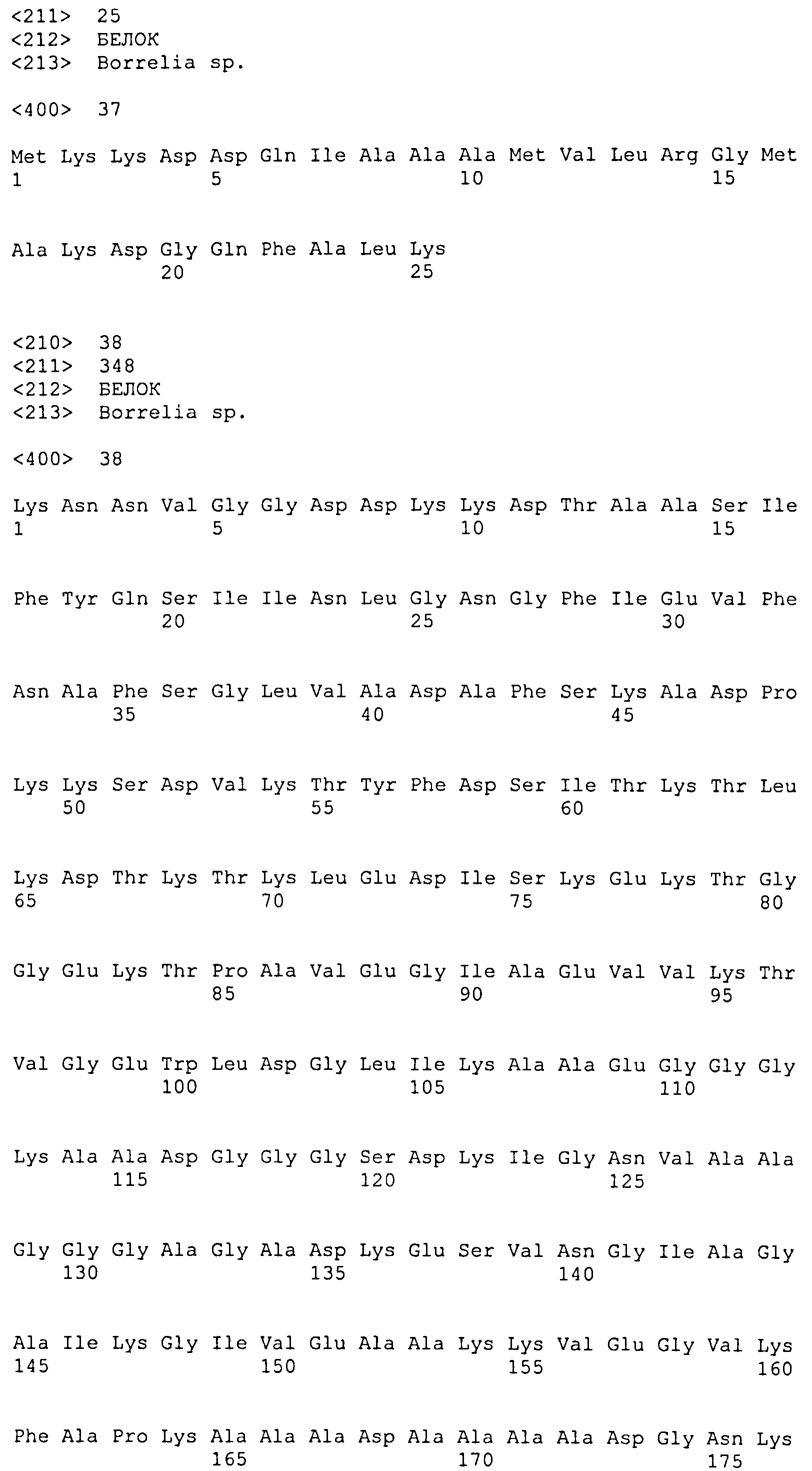
(a) белок, аминокислотная последовательность которого включает (или состоит из нее) последовательность SEQ ID NO: 1 и последовательность SEQ ID NO: 2, или вариант упомянутого белка, аминокислотная последовательность которого включает (или состоит из нее) последовательность, имеющую степень идентичности по меньшей мере 40% относительно SEQ ID NO: 1, и последовательность, имеющую степень идентичности по меньшей мере 50% относительно SEQ ID NO: 2, при условии, что этот вариант способен образовывать иммунологический комплекс с антителами, продуцируемыми вследствие инфицирования боррелиями, или этот вариант способен индуцировать продуцирование антиборрелиозных антител;
(b) белок, аминокислотная последовательность которого включает (или состоит из нее) последовательность SEQ ID NO: 3 и последовательность SEQ ID NO: 4, или вариант упомянутого белка, аминокислотная последовательность которого включает (или состоит из нее) последовательность, имеющую степень идентичности по меньшей мере 40% относительно SEQ ID NO: 3, и последовательность, имеющую степень идентичности по меньшей мере 50% относительно SEQ ID NO: 4, при условии, что этот вариант способен образовывать иммунологический комплекс с антителами, продуцируемыми вследствие инфицирования боррелиями, или этот вариант способен индуцировать продуцирование антиборрелиозных антител;
(c) белок, аминокислотная последовательность которого включает (или состоит из нее) последовательность SEQ ID NO: 5 и последовательность SEQ ID NO: 7, или вариант упомянутого белка, аминокислотная последовательность которого включает (или состоит из нее) последовательность, имеющую степень идентичности по меньшей мере 40% относительно SEQ ID NO: 5, и последовательность, имеющую степень идентичности по меньшей мере 50% относительно SEQ ID NO: 7, при условии, что этот вариант способен образовывать иммунологический комплекс с антителами, продуцируемыми вследствие инфицирования боррелиями, или этот вариант способен индуцировать продуцирование антиборрелиозных антител;
(d) белок, аминокислотная последовательность которого включает (или состоит из нее) последовательность SEQ ID NO: 6 и последовательность SEQ ID NO: 7, или вариант упомянутого белка, аминокислотная последовательность которого включает (или состоит из нее) последовательность, имеющую степень идентичности по меньшей мере 40% относительно SEQ ID NO: 6, и последовательность, имеющую степень идентичности по меньшей мере 50% относительно SEQ ID NO: 7, при условии, что этот вариант способен образовывать иммунологический комплекс с антителами, продуцируемыми вследствие инфицирования боррелиями, или этот вариант способен индуцировать продуцирование антиборрелиозных антител;
(e) белок, аминокислотная последовательность которого включает (или состоит из нее) последовательность SEQ ID NO: 5, последовательность SEQ ID NO: 6 и последовательность SEQ ID NO: 7, или вариант упомянутого белка, аминокислотная последовательность которого включает (или состоит из нее) последовательность, имеющую степень идентичности по меньшей мере 40% относительно SEQ ID NO: 5, последовательность, имеющую степень идентичности по меньшей мере 40% относительно SEQ ID NO: 6, и последовательность, имеющую степень идентичности по меньшей мере 50% относительно SEQ ID NO: 7, при условии, что этот вариант способен образовывать иммунологический комплекс с антителами, продуцируемыми вследствие инфицирования боррелиями, или этот вариант способен индуцировать продуцирование антиборрелиозных антител;
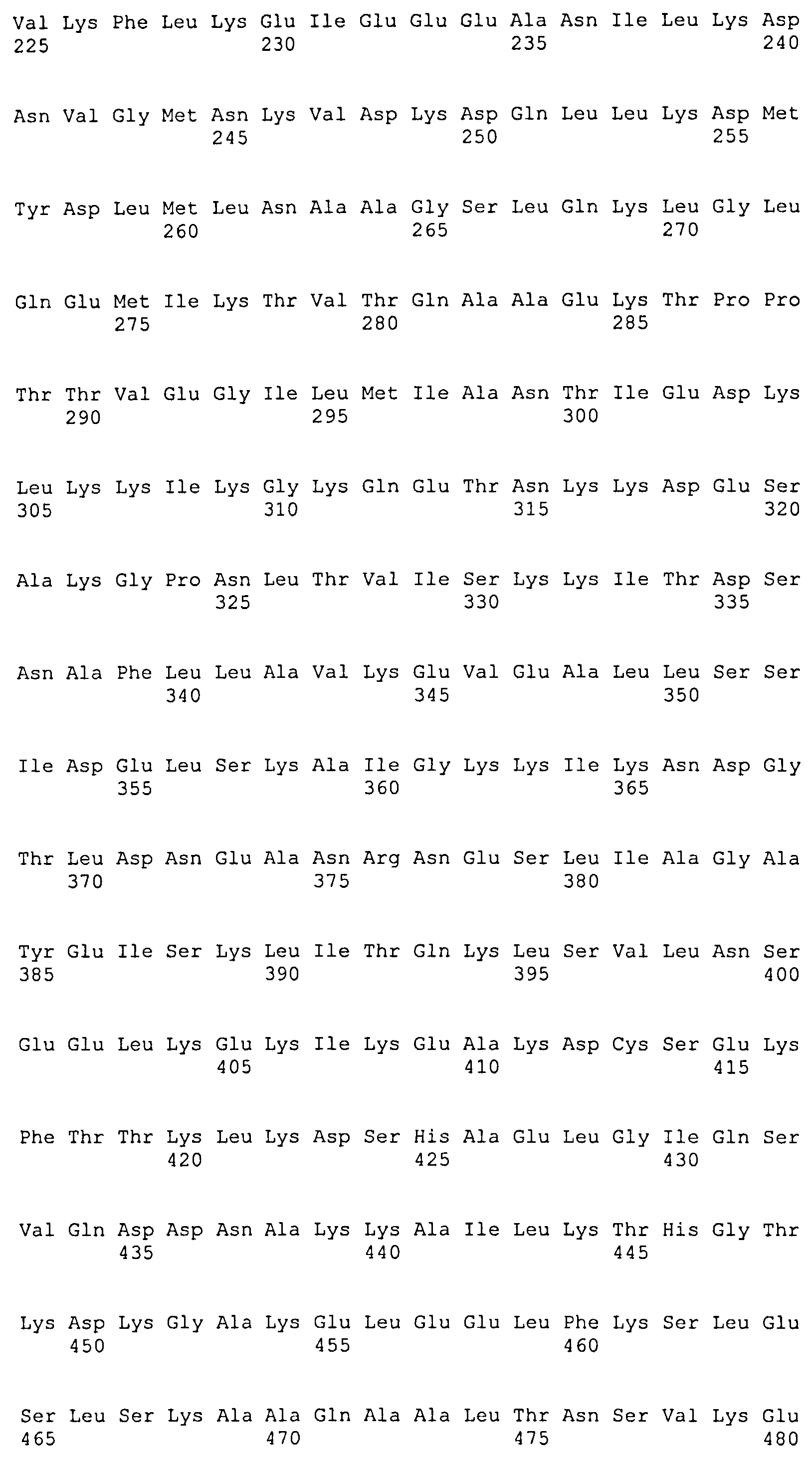
(f) белок, аминокислотная последовательность которого включает (или состоит из нее) последовательность, выбранную из последовательностей SEQ ID NO: 8, 9, 10, 11, 12, 13 и 14.
Каждый из белков, идентифицированных ранее, включает в себя по меньшей мере одну последовательность внеклеточного домена белка DbpA вида боррелии, выбранного из B. afzelii (SEQ ID NO: 1), B. burgdorferi sensu stricto (SEQ ID NO: 3) и B. garinii (группа III: SEQ ID NO: 5) (группа IV: SEQ ID NO: 6), или последовательность, имеющую степень идентичности по меньшей мере 40% относительно упомянутых последовательностей, и по меньшей мере одну последовательность белка OspC B. afzelii (SEQ ID NO: 2), B. burgdorferi sensu stricto (SEQ ID NO: 4) и B. garinii (SEQ ID NO: 7) или последовательность, имеющую степень идентичности по меньшей мере 50% относительно упомянутых последовательностей. Предпочтительно, одна или несколько последовательностей DbpA находятся на N-концевом участке химерного рекомбинантного белка, а последовательность OspC находится на C-концевом участке химерного рекомбинантного белка.
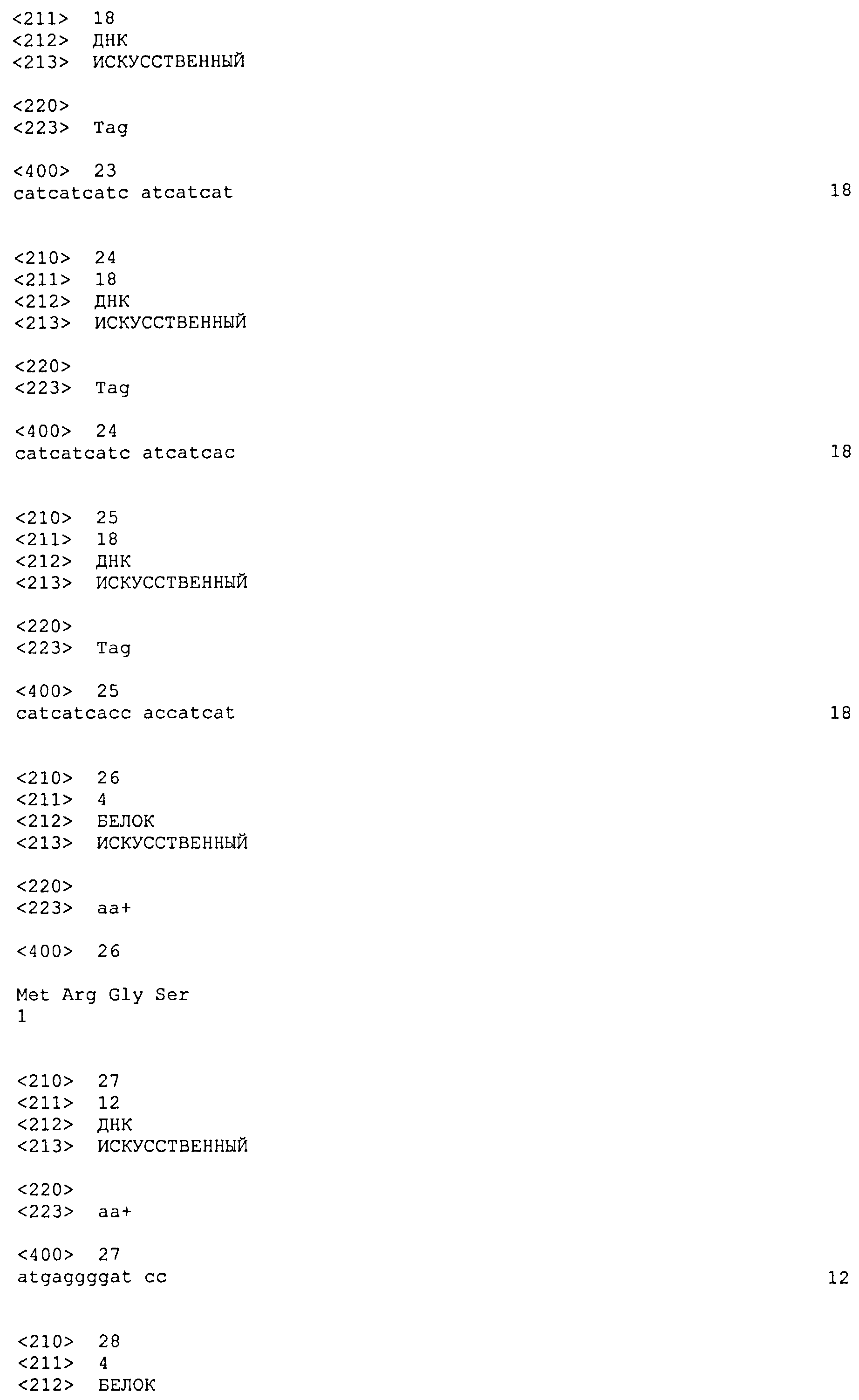
Последовательность по меньшей мере из 6 гистидиновых остатков может быть присоединена на уровне N-концевого или C-концевого участка белка с целью обеспечения его очистки на металлохелатной смоле. Последовательность из 6 гистидиновых остатков, идентифицированная в SEQ ID NO: 22, находится предпочтительно на N-концевом участке конструкции. В качестве примера такое расположение поясняется последовательностями SEQ ID NO: 9, 11, 13 и 14, которые содержат на N-концевом участке цепочку поли-His(6). Цепочка поли-His может кодироваться любой из последовательностей, идентифицированных в SEQ ID NO: 23, 24 и 25.
Добавочные аминокислоты могут находиться в начале цепочки поли-His в результате инсерции в последовательность ДНК, кодирующей малую последовательность, которая позволяет облегчать клонирование требуемой последовательности в экспрессирующей плазмиде. Это относится, в частности, к последовательностям SEQ ID NO: 9, 11, 13 и 14, содержащим звено "MRGS" (SEQ ID NO: 26) в начале цепочки поли-His. Звено "MRGS" кодируется последовательностью ATGAGGGGATCC (SEQ ID NO: 27).
Область связывания может быть введена между каждой из последовательностей DbpA и OspC, образующих химерный рекомбинантный белок. Область такого типа представляет собой область гибкого спейсера, обеспечивающую лучшую доступность потенциальных антител к каждому из доменов. Она содержит много аминокислотных остатков Gly и Ser, представляющих собой аминокислоты, охарактеризованные как аминокислоты, придающие гибкость в третичной структуре белка. Также возможно вводить в требуемую кодирующую последовательность плечо ДНК (или линкер) для облегчения связи между последовательностями, кодирующими два требуемых белка. Это относится, в частности, к последовательности SEQ ID NO: 14, содержащей звено "GSGG" (SEQ ID NO: 28), кодируемое последовательностью GGTTCCGGGGGT (SEQ ID NO: 29) и действующее в качестве плеча связывания между белками DbpA группы IV и OspC B. garinii.
Предпочтительные белки идентифицированы в SEQ ID NO: 8, 9, 10, 11, 12 13 и 14. Они соответственно кодируются соответствующими последовательностями ДНК, идентифицированными в SEQ ID NO: 15, 16, 17, 18, 19, 20 и 21.
Настоящее изобретение относится также к последовательностям ДНК, кодирующим ранее определенные белки, в частности к последовательностям, идентифицированным в SEQ ID NO: 15, 16, 17, 18, 19, 20 и 21.
Настоящее изобретение относится также к экспрессирующей кассете, являющейся функциональной в клетке, производной от прокариотического (пример: Escherichia coli) или эукариотического организма, такого как дрожжи (пример: Pichia, Schizosaccharomyces), и обеспечивающей экспрессию ранее описанной нуклеиновой кислоты (ДНК), когда она находится под контролем элементов, разрешающих ее экспрессию, а также к вектору, включающему в себя такую кассету.
Белки по настоящему изобретению являются предпочтительно приемлемыми для диагностики инфицирования боррелиями. Таким образом, настоящее изобретение относится к способу диагностики in vitro лайм-боррелиоза в биологическом образце (например, в образце сыворотки, крови, плазмы и т.д.), согласно которому биологический образец приводят в контакт по меньшей мере с одним ранее определенным белком и выявляют возможное образование иммунологического комплекса между упомянутым белком и антителами биологического образца (IgG и/или IgM), например, посредством прибавления по меньшей мере одного антииммуноглобулина человека, меченного любым приемлемым маркером. Под маркером понимают индикатор, способный генерировать сигнал. В неограничительный перечень таких индикаторов входят ферменты, производящие сигнал, детектируемый, например, колориметрией, по флуоресценции или люминесценции, такие как пероксидаза хрена, щелочная фосфатаза, β-галактозидаза, глюкозо-6-фосфатдегидрогеназа; хромофоры, такие как флуоресцирующие, люминесцирующие или окрашивающие соединения; группировки, обладающие электронной плотностью, детектируемой электронной микроскопией или по их электрическим свойствам, таким как проводимость, способами амперометрии или вольтамперометрии или по измерению сопротивления; группировки, детектируемые оптическими способами, такими как дифракция, резонанс поверхностного плазмона, изменение краевого угла, или физическими способами, такими как спектроскопия атомных взаимодействий, туннельный эффект, и т.д.; радиоактивные молекулы, содержащие32P,35S или125I. Один или несколько белков предпочтительно иммобилизуют на твердой подложке, которая может представлять собой конус прибора Vidas®, лунки планшета для микротитрования, гель, отдельные частицы и т.д.
В одном из вариантов осуществления настоящего изобретения биологический образец, кроме того, приводят в контакт по меньшей мере с одним химерным белком VlsE, описанным далее.
Белок VlsE (surface expressed lipoprotein with Extensive antigenic Variation (экспрессируемый поверхностью липопротеин с расширенной антигенной вариативностью) главным образом экспрессируется in vivo транзиторным образом и вскоре после инфицирования хозяина. Он является очень иммуногенным у инфицированного хозяина и вызывает продуцирование IgG и IgM. Локус Vls находится в линейной плазмиде длиной 28 т.п.о. (Ip28-1), имеющейся у трех геновидов боррелий, ответственных за лайм-боррелиоз, и состоит из молчащих кассет и экспрессирующего сайта (VlsE). In vivo нерегулярные рекомбинации между экспрессирующими и молчащими кассетами внезапно возникают в ходе инфицирования и лежат в основе антигенной вариабельности VlsE. Белок VlsE состоит из шести вариабельных областей VR1-VR6, расположенных на поверхности белка VlsE и отделенных промежутками от областей IR1-IR6, называемых константными.
Химерный белок VlsE включает (или состоит в основном из):
(i) по меньшей мере одну последовательность, выбранную из последовательностей, идентифицированных в SEQ ID NO: 30, 31, 32, 33 и 34, и последовательностей, имеющих степень идентичности меньшей мере 50%, предпочтительно 60 или 70% и преимущественно по меньшей мере 80 или 85% относительно SEQ ID NO: 30, 31, 32, 33 и 34;
(ii) по меньшей мере одну последовательность, включающую последовательность SEQ ID NO: 35 или последовательность, имеющую степень идентичности по меньшей мере 80%, предпочтительно по меньшей мере 85% и преимущественно по меньшей мере 90% относительно SEQ ID NO: 35, последовательность SEQ ID NO: 36 или последовательность, имеющую степень идентичности по меньшей мере 80%, предпочтительно по меньшей мере 85% и преимущественно по меньшей мере 90% относительно SEQ ID NO: 36, последовательность SEQ ID NO: 37 или последовательность, имеющую степень идентичности по меньшей мере 80%, предпочтительно по меньшей мере 85% и преимущественно по меньшей мере 90% относительно SEQ ID NO: 37, и при необходимости последовательность SEQ ID NO: 43. Химерный белок VlsE предпочтительно включает последовательность SEQ ID NO: 43.
Соответственно описанному ранее полигистидиновая цепочка (×6) может быть присоединена на уровне N-концевого участка химерного белка с целью обеспечения его очистки на металлохелатной смоле, а также могут быть присоединены добавочные аминокислоты в начале цепочки поли-His.
Предпочтительный химерный белок включает (или состоит в основном из, или состоит из):
(i) последовательность SEQ ID NO: 30 или последовательность, имеющую степень идентичности по меньшей мере 50%, предпочтительно по меньшей мере 60 или 70% и преимущественно по меньшей мере 80 или 85% относительно SEQ ID NO: 30;
(ii) последовательность, включающую последовательность SEQ ID NO: 35 или последовательность, имеющую степень идентичности по меньшей мере 80%, предпочтительно по меньшей мере 85% и преимущественно по меньшей мере 90% относительно SEQ ID NO: 35, последовательность SEQ ID NO: 36 или последовательность, имеющую степень идентичности по меньшей мере 80%, предпочтительно по меньшей мере 85% и преимущественно по меньшей мере 90% относительно SEQ ID NO: 36, последовательность SEQ ID NO: 37 или последовательность, имеющую степень идентичности по меньшей мере 80%, предпочтительно по меньшей мере 85% и преимущественно по меньшей мере 90% относительно SEQ ID NO: 37, и последовательность SEQ ID NO: 43.
Предпочтительный химерный белок включает (или состоит в основном из, или состоит из):
(i) последовательность SEQ ID NO: 30;
(ii) последовательность, включающую в себя последовательности SEQ ID NO: 35, 36, 37 и 43.
Белок включает или состоит из последовательности, идентифицированной как SEQ ID NO: 38.
SEQ ID NO: 30 соответствует последовательности внеклеточного домена VlsE B. garinii (штамм pBi), укороченного в его сигнальной последовательности (aa 1-19) и C-концевой области зрелого белка, расположенной после домена IR6.
SEQ ID NO: 31 соответствует последовательности внеклеточного домена VlsE B. garinii (штамм pBr), укороченного в его сигнальной последовательности и C-концевой области зрелого белка, расположенной после домена IR6.
SEQ ID NO: 32 соответствует последовательности внеклеточного домена VlsE B. garinii (штамм pLi), укороченного в его сигнальной последовательности и C-концевой области зрелого белка, расположенной после домена IR6.
SEQ ID NO: 33 соответствует последовательности внеклеточного домена VlsE B. afzelii (штамм pKo), укороченного в его сигнальной последовательности и C-концевой области зрелого белка, расположенной после домена IR6.
SEQ ID NO: 34 соответствует последовательности внеклеточного домена VlsE B. burgdorferi sensu stricto (штамм B31), укороченного в его сигнальной последовательности и C-концевой области зрелого белка, расположенной после домена IR6.
SEQ ID NO: 35 соответствует последовательности домена IR6 B. burgdorferi sensu stricto (штамм B31).
SEQ ID NO: 36 соответствует последовательности домена IR6 B. afzelii (штамм ACA-1).
SEQ ID NO: 37 соответствует последовательности домена IR6 B. garinii (штамм Ip90).
SEQ ID NO: 43 соответствует последовательности вариабельной области VR6 B. burgdorferi sensu stricto (штамм B31).
Настоящее изобретение относится также к набору для диагностики in vitro лайм-боррелиоза, содержащему по меньшей мере один химерный белок DbpA-OspC, описанный ранее, и предпочтительно содержащему по меньшей мере один вид антииммуноглобулина человека, меченного любым приемлемым маркером, соответствующим приведенным ранее определениям. Кроме того, набор может содержать химерный белок VlsE, определенный ранее.
Белки по настоящему изобретению являются также приемлемыми для применения в качестве активного ингредиента для получения вакцинной композиции для профилактики инфицирования боррелиями. Таким образом, настоящее изобретение относится также к вакцинной композиции, содержащей по меньшей мере один белок, определенный ранее, и фармацевтически приемлемый наполнитель.
Приведенные далее примеры даны в качестве пояснения и не имеют какой-либо ограничительный характер. Они позволяют лучше понять настоящее изобретение. Порядок последовательностей, кодирующих различные эпитопные иммунодоминантные области химерных рекомбинантных белков, может быть при необходимости изменен. Эпитопы также могут иметь изменения по сравнению с последовательностями, описанными в примерах, в зависимости от вида Borrelia burgdorferi и одного или нескольких штаммов, представляемых ими. Длина областей связывания также может быть изменена для улучшения гибкости между двумя доменами. В заключение, области фиксации могут быть вставлены внутри областей связывания.
ПРИМЕРЫ
Пример 1. Получение плазмидных конструкций, кодирующих химерные рекомбинантные белки DpbA-OspC
Последовательности ДНК, кодирующие различные описанные последовательности DpbA и OspC, идентифицированы в таблице 1. Последовательности ДНК были оптимизированы для облегчения экспрессии в E. coli с использованием набора GeneOptimizer™ и синтезированы соответственно методике GenScript corporation (Скотч-Плейнс, NJ, США) или GeneArt GmbH (Регенсбург, Германия).
Каждый химерный рекомбинантный белок содержит по меньшей мере одну эпитопную область, соответствующую внеклеточному домену последовательности DbpA Borrelia burgdorferi sensu stricto или B. afzelii, или B. garinii, и по меньшей мере одну эпитопную область, соответствующую внеклеточному домену последовательности OspC Borrelia burgdorferi sensu stricto или B. afzelii, или B. garinii.
Ассоциации различных нуклеотидных последовательностей, кодирующих последовательности DbpA и/или OspC, а также изменения нуклеотидных последовательностей, такие как делеции, добавление последовательности связывания или добавление последовательности-линкера, осуществляли путем генной инженерии с использованием методик ПЦР, хорошо известных специалистам в данной области техники и описанных, например, в Sambrook J. et al., Molecular Cloning: A Laboratory Manual, 1989.
Последовательности ДНК, кодирующие требуемые химерные белки, были введены в экспрессионный вектор pMR [2] между сайтом рестрикции BamHI в положении 5' и сайтом EcoRI или HindIII в положении 3'. Соответствующие плазмидные конструкции и белки, упомянутые в примере (bLYM114, bLYM120 и bLYM121), описаны в таблице 2. MRGS в N-концевом участке рекомбинантных белков и соответствующая нуклеотидная последовательность ATG AGG GGA TCC были введены по методике клонирования, использованной для экспрессионного вектора pMR. В данной последовательности действительно необходим только кодон инициации ATG и, следовательно, аминокислота Met.
Полигистидиновая последовательность (6×His) была введена на N-концевом участке каждого рекомбинантного белка. Данная последовательность обеспечивает очистку рекомбинантных белков на колонке для металлохелатной аффинной хроматографии. Она представляет собой область фиксации на геле Ni-NTA, что позволяет облегчать в дальнейшем стадию очистки химерного рекомбинантного белка. Данный пептид HHHHHH (SEQ ID NO: 22) кодируется нуклеотидными последовательностями CATCATCATCATCATCAT (SEQ ID NO: 23) или CATCATCATCATCATCAC (SEQ ID NO: 24), или CATCATCACCACCATCAT (SEQ ID NO: 25), или любой другой последовательностью, кодирующей последовательность SEQ ID NO: 22. Данная область особой фиксации, включающая в себя гистидиновую последовательность, обеспечивает, в частности, ориентированную фиксацию рекомбинантного белка на подложке, образованной диоксидом кремния или оксидами металлов.
Пример 2. Экспрессия рекомбинантных белков bLYM114, bLYM120 и bLYM121 примера 1 и очистка
Для трансформации бактерии E. coli (штамм BL21) согласно традиционной методике, известной специалистам в данной области техники, была использована плазмидная конструкция, соответствующая последовательности SEQ ID NO: 16, 18 или 21, вставленной в экспрессионный вектор (pMR). Трансформированные бактерии селекционировали по их резистентности к ампициллину, приданной вектором pMR.
Затем осуществляли селекцию клона рекомбинантной бактерии для посева прекультуры в 40 мл среды 2×YT (триптон 16 г/л; экстракт дрожжей 10 г/л; NaCl 5 г/л, pH 7,0), содержавшей 100 мкг/мл ампициллина. После инкубации в течение 15-18 часов при 30°C при перемешивании со скоростью 250 об/мин данную прекультуру использовали для осуществления посева в 1 л среды 2×YT, содержавшей 2% глюкозы и 100 мкг/мл ампициллина. Данную культуру инкубировали при 30°C при перемешивании со скоростью 250 об/мин до достижения оптической плотности DO при 600 нм значения 1,0/1,2. Культуру выдерживали в течение 3 часов 30 мин или 4 часов при 30°C после прибавления 0,4 мМ изопропил-β-D-тиогалактопиранозида (IPTG) и собирали центрифугированием при 6000 g в течение 30 мин. Осадок клеток хранили при -60°C. Для очистки влажную биомассу размораживали и ресуспендировали в лизирующем буферном растворе, содержавшем ингибиторы протеаз без ЭДТА (Roche) и бензоназонуклеазу (Novagen), и осуществляли разрушение клеток при 1,6 кбар в деструкторе клеток (Constant System Ltd, Давентри, Великобритания). Затем лизат центрифугировали при 10000 об/мин в течение 45 мин при 2-8°C. Полученная надосадочная жидкость содержит растворимые белки. Надосадочную жидкость фильтровали через фильтр пористостью 0,45 мкм и очищали аффинной хроматографией на металлохелатной колонке (матрица "никель-нитрилуксусная кислота" (Ni-NTA, Qiagen)). С этой целью надосадочную жидкость вводили (1 мл/мин) при 18-25°C в колонку с 8 мл геля Ni-NTA, кондиционированного буферным раствором A (см. таблицу 3). Затем колонку промывали буферным раствором A до достижения на выходе из колонки DO280 нм=0. Элюирование рекомбинантного белка осуществляли подачей буферного раствора B, соответственно указаниям, приведенным в таблице 3, а очищенный белок подвергали диализу в кассете для диализа с отсечкой 10000 или 20000 а.е.м. (Slide-A-Lyser®, Pierce) против диализного буферного раствора. Условия очистки на геле Ni-NTA описаны в таблице 3.
Образцы анализировали на геле NuPAGE® Novex®, 4-12%, в буферном растворе NuPAGE® MES-SDS согласно руководству изготовителя (Invitrogen). Белки окрашивали бриллиантовым синим Кумасси или электрофоретически переносили на нитроцеллюлозную мембрану. Мембрану блокировали 5% (масс./об.) раствором сухого молока в PBS и инкубировали с анти-пентагистидиновым антителом (Qiagen) в PBS, содержавшем 0,05% Tween 20. Конъюгат антимышиного IgG козы, меченный пероксидазой хрена, (Jackson Immunoresearch laboratories) в PBS/Tween использовали в качестве вторичного антитела.
Определение концентрации белков осуществляли, используя набор Bradford (Pierce Coomassie Plus, Perbio Science) с BSA в качестве стандартного белка.
Пример 3. Обнаружение IgG и IgM человека с химерными рекомбинантными белками способом линейного иммуноблоттинга
Каждый рекомбинантный белок наносили на поливинилидендифторидную мембрану (PVDF, Immobilon, Millipore®, Бедфорд, Массачусетс, США) по приведенной далее методике.
Концентрацию белка в PBS с pH 7,2 устанавливали равной 1 мг/мл и разбавляли PBS с pH 7,2, дополненным 0,03% Tween 20 (разведение 1/200). Мембрану из PVDF смачивали метанолом, промывали в деминерализованной воде и накладывали на бумагу с влажным пятном. Пластмассовую линейку погружали в разбавленный раствор белков и фиксировали на мембране из PVDF. После нанесения белков и высушивания мембрану разрезали в вертикальном направлении на полоски. Перед применением полоски инкубировали с 5%-ным раствором желатина в TBS с pH 7,5 в течение 1 часа при 37°C. Процедуры иммуноблоттинга осуществляли при комнатной температуре соответственно описанному Bretz A.G. и соавт. [3]. Полоски инкубировали в течение 2 часов с пробами сыворотки человека, разбавленными в соотношении 1/200 в TBS с 1% желатина, промывали и инкубировали с антителом анти-IgG или анти-IgM человека, меченным щелочной фосфатазой (Sigma, Сент-Луис, США) и разбавленным в соотношении 1/1000 в TBS с 1% желатина. После промывки полоски инкубировали с субстратом BCIP-NBT щелочной фосфатазы (KPL, Гейтерсбург, MD, США) в течение 30 мин, затем промывали в дистиллированной воде и сушили.
Группа проб испытуемых сывороток
Пробы сыворотки человека отбирали у типичных, клинически хорошо охарактеризованных больных LB, соответствующих различным стадиям LB (22 с мигрирующей эритемой [EM], 5 с кардитом, 20 с нейроборрелиозом [NB], 20 с артритом Лайма [LA], 20 с атрофическим хроническим акродерматитом [ACA] и 10 с доброкачественной лимфоцитомой кожи [LCB]). Антиборрелиозные IgG были найдены описанным ранее иммуноблоттингом при использовании лизатов целых клеток [4] в сыворотке больных LA, ACA и кардитом. EM, NB и LCB были идентифицированы клинически, но все соответствующие пробы сыворотки не были определены как положительные ни посредством штатного иммуноблоттинга [4], ни посредством коммерческих наборов (VIDAS® Lyme (biomérieux), Borrelia IgG (Diasorin®) и Borrelia IgM (r-biopharm®)). Напротив, во всех случаях NB, включенных в исследование, имелись антитела, обнаруживаемые в спинномозговой жидкости [LCR] (индекс находился в интервале от 2 до 27,1 при определении с набором VIDAS® Lyme (biomérieux)). Присутствие IgM было определено только в клинических случаях стадий I и II, но не на хронических стадиях.
Группа отрицательного контроля включала в себя 31 пробу сыворотки, при этом пробы ранее были определены в традиционных испытаниях как отрицательные в отношении присутствия антиборрелиозных антител. Кроме того, с рекомбинантным белком были испытаны 64 пробы сыворотки здоровых доноров крови, проживающих в регионе, эндемичном в отношении болезни Лайма (Монтей, Вале, Швейцария).
Интенсивность реакции оценивали следующим образом: [+], [++], [+++], [-] или неоднозначные результаты. Неоднозначные результаты принимали за отрицательные.
Результаты представлены в таблице 4.
Специфичность равна 100% на основании 31 пробы сыворотки, отобранной у здоровых участников и определенной как лайм-отрицательные посредством стандартных коммерческих тестов.
Обнаружение IgG
Результаты показывают, что слитые химерные рекомбинантные белки представляют собой диагностический инструмент, чувствительный в отношении IgG и IgM на всех стадиях инфицирования. Они позволяют выявить комплементарное действие трех рекомбинантных белков, основанных на последовательностях Borrelia afzelii, B. sensu stricto и B. garinii соответственно, для обнаружения IgG. Комбинированное применение трех химерных рекомбинантных белков позволяет на стадии I инфицирования обнаруживать IgG в 9 случаях больных с EM из 22 (или 40,9% чувствительности).
Обнаружение IgM
Химерные антибелки IgM найдены в 11 случаях из 22 (или 50% чувствительности). Таким образом, в сыворотке больных LB стадии I такие химерные белки позволяют обнаруживать IgM чаще, чем IgG. В испытаниях, осуществленных для контроля посредством штатного иммуноблоттинга [4] и коммерческого набора Borrelia IgM (r-biopharm®), обнаружено не большее число проб сыворотки с положительными результатами на IgM. Кроме того, 3 пробы сыворотки, которые посредством иммуноблоттинга и набора Borrelia IgM (r-biopharm®) определены как отрицательные, определены как положительные посредством трех химерных белков, упомянутых в примере (3/3), или посредством одного из трех белков, упомянутых в примере (1/3). Комбинированное применение трех рекомбинантных белков позволяет улучшить на 13,6% чувствительность обнаружения IgM на стадии I инфицирования.
Пример 4. Получение плазмидных конструкций, кодирующих химерные рекомбинантные белки VlsE
Последовательности ДНК, кодирующие различные последовательности белка, идентифицированы в таблице 5.
Последовательности были оптимизированы в отношении их экспрессии в E. coli с использованием набора GeneOptimizer™ и синтезированы соответственно методике GenScript corporation (Скотч-Плейнс, NJ, США) или GeneArt GmbH (Регенсбург, Германия).
Дополнительные изменения ДНК, делеции или ассоциации различных последовательностей реализовывали способом ПЦР (PCR), применяемым в генной инженерии, с использованием методик ПЦР, хорошо известных специалистам в данной области техники и описанных, например, в Sambrook J. et al., Molecular Cloning: A Laboratory Manual, 1989. Последовательности ДНК были связаны в экспрессионный вектор pMR [2] или pET-3d (Novagen®). Соответствующие плазмидные конструкции и белки, упомянутые в примере (bLYM110, bLYM125), описаны в таблице 6.
Пример 5. Экспрессия рекомбинантных белков примера 4 и очистка
Для трансформации бактерии E. coli (штамм BL21) согласно традиционной методике, известной специалистам в данной области техники, использовали плазмидную конструкцию, описанную в примере 4. Трансформированные бактерии селекционировали по их резистентности к ампициллину, приданной вектором pMR или pET.
Затем осуществляли селекцию клона рекомбинантной бактерии для посева прекультуры в 40 мл среды 2×YT (триптон 16 г/л; экстракт дрожжей 10 г/л; NaCl 5 г/л, pH 7,0), содержавшей 100 мкг/мл ампициллина. После инкубации в течение 15-18 часов при 30°C при перемешивании со скоростью 250 об/мин данную прекультуру использовали для осуществления посева в 1 л среды 2×YT, содержавшей 2% глюкозы и 100 мкг/мл ампициллина. Данную культуру инкубировали при 30°C при перемешивании со скоростью 250 об/мин до достижения оптической плотности DO при 600 нм значения 1,0/1,2. Культуру выдерживали в течение 3 часов 30 мин или 4 часов при 30°C после прибавления 0,4 мМ изопропил-β-D-тиогалактопиранозида (IPTG) и собирали центрифугированием при 6000 g в течение 30 мин. Осадок клеток хранили при -60°C. Для очистки влажную биомассу ресуспендировали в лизирующем буферном растворе, содержавшем ингибиторы протеаз без ЭДТА (Roche) и бензоназонуклеазу (Novagen®), и осуществляли разрушение клеток при 1,6 кбар в деструкторе клеток (Constant System Ltd, Давентри, Великобритания). Затем лизат центрифугировали при 10000 об/мин в течение 45 минут при 2-8°C. После фильтрования через фильтр пористостью 0,22 мкм надосадочную жидкость вводили в колонку Ni-NTA (Quiagen®), кондиционированную лизирующим буферным раствором. Затем смолу промывали тем же самым буферным раствором до достижения A280 нмзначения базовой линии. Элюирование осуществляли элюирующим буферным раствором, а очищенный белок подвергали диализу на кассете Pierce для диализа Slide-A-Lyser® с отсечкой 10000 или 20000 а.е.м. против диализного буферного раствора. Условия очистки на геле Ni-NTA описаны в таблице 7.
Образцы анализировали на геле NuPAGE® Novex®, 4-12%, в циркуляционном буферном растворе NuPAGE® MES-SDS согласно руководству изготовителя (Invitrogen™). Белки окрашивали бриллиантовым синим Кумасси или электрофоретически переносили на нитроцеллюлозную мембрану. Мембрану блокировали 5%-ным (масс./об.) раствором сухого молока в PBS и инкубировали с анти-пентагистидиновым антителом (Qiagen®) в PBS, содержавшем 0,05% Tween 20. Конъюгат антимышиного IgG козы, меченный пероксидазой хрена, (Jackson Immunoresearch laboratories) в PBS/Tween использовали в качестве вторичного антитела.
Определение концентрации белков осуществляли, используя набор Bradford Assay Kit (Pierce Coomassie Plus, Perbio Science) с BSA в качестве стандартного белка.
Пример 6. Обнаружение IgG и IgM человека с химерным рекомбинантным белком bLYM110 примера 5 способом линейного иммуноблоттинга
Рекомбинантный белок наносили на поливинилидендифторидную мембрану (PVDF, Immobilon, Millipore®, Бедфорд, Массачусетс, США) по приведенной далее методике.
Концентрацию белка в PBS с pH 7,2 устанавливали равной 1 мг/мл и разбавляли PBS с pH 7,2, дополненным 0,03% Tween 20 (разведение 1/200). Мембрану из PVDF смачивали метанолом, промывали в деминерализованной воде и накладывали на бумагу с влажным пятном. Пластмассовую линейку погружали в разбавленный раствор белков и фиксировали на мембране из PVDF. После нанесения белков и высушивания мембрану разрезали в вертикальном направлении на полоски. Перед применением полоски инкубировали с 5%-ным раствором желатина в TBS с pH 7,5 в течение 1 часа при 37°C. Процедуры иммуноблоттинга осуществляли при комнатной температуре соответственно описанному Bretz A.G. и соавт. [3]. Полоски инкубировали в течение 2 часов с пробами сыворотки человека, разбавленными в соотношении 1/200 в TBS с 1% желатина, промывали и инкубировали с отличающимися от человеческого IgG или IgM, меченными щелочной фосфатазой (Sigma™, Сент-Луис, США) и разбавленными в соотношении 1/1000 в TBS с 1% желатина. После промывки полоски инкубировали с субстратом BCIP-NBT (KPL, Гейтерсбург, MD, США) в течение 30 минут, промывали в дистиллированной воде и сушили.
Группа проб испытуемых сывороток
Пробы сыворотки человека отбирали у типичных, клинически хорошо охарактеризованных больных LB, соответствующих различным стадиям LB (22 с мигрирующей эритемой [EM], 5 с кардитом, 20 с нейроборрелиозом [NB], 20 с артритом Лайма [LA], 20 с атрофическим хроническим акродерматитом [ACA] и 10 с доброкачественной лимфоцитомой кожи [LCB]). Антиборрелиозные IgG были найдены описанным ранее иммуноблоттингом при использовании лизатов целых клеток [4] в сыворотке больных LA, ACA и кардитом. EM, NB и LCB были идентифицированы клинически, но все соответствующие пробы сыворотки не были определены как положительные ни посредством иммуноблоттинга [4], ни посредством коммерческих наборов (VIDAS® Lyme (biomérieux®), Borrelia IgG (Diasorin®) и Borrelia IgM (r-biopharm®)). Напротив, во всех случаях NB, включенных в исследование, имелись антитела, обнаруживаемые в спинномозговой жидкости [LCR] (индекс находился в интервале от 2 до 27,1).
Группа отрицательного контроля включала в себя 31 пробу сыворотки, при этом пробы ранее были определены в традиционных испытаниях как отрицательные в отношении присутствия антиборрелиозных антител. Кроме того, с рекомбинантным белком были испытаны 64 пробы сыворотки здоровых доноров крови, проживающих в регионе, эндемичном в отношении болезни Лайма (Монтей, Вале, Швейцария). Интенсивность реакции оценивали следующим образом: [+], [++], [+++], [-] или неоднозначные результаты. Неоднозначные результаты принимали за отрицательные.
Результаты представлены в приведенной далее таблице 8.
Обнаружение IgG
Результаты показывают, что рекомбинантный белок bLYM110 представляет собой диагностический антиген, высокочувствительный в отношении IgG на всех стадиях инфицирования. На стадии I инфицирования IgG были обнаружены в 17 случаях больных с EM из 22 (или 77,3% чувствительности). У пяти больных с EM, для которых найдены отрицательные результаты с рекомбинантным белком, такие же результаты определены штатным иммуноблоттингом и с коммерческими наборами. Семь проб сыворотки с EM, найденные положительными с рекомбинантным белком, не были обнаружены иммуноблоттингом, что представляет собой улучшение чувствительности с рекомбинантным белком на 31,8%. На первой стадии инфицирования в отсутствие характерного покраснения диагностика может быть затруднена, так как другие клинические проявления болезни Лайма не являются специфическими. Кроме того, традиционными испытаниями были обнаружены только несколько больных, имевших EM. Таким образом, белок по настоящему изобретению улучшил обнаружение IgG на стадии I инфицирования, обеспечив обнаружение их более чем у 77% больных, имевших EM.
Обнаружение IgM
Химерные антибелки IgM найдены в 23,4% проб сыворотки с LB. В сыворотке больных LB стадий I и II белок позволяет обнаруживать IgG чаще, чем IgM.
Пример 7. Оценка и валидация химерных рекомбинантных белков bLYM114, bLYM120, bLYM121 и bLYM125 посредством теста VIDAS® (bioMérieux)
Данную валидацию осуществляли посредством теста VIDAS®, применяя:
1) химерные рекомбинантные белки bLYM114, bLYM120 и bLYM121, полученные согласно примерам 1 и 2, для обнаружения IgM;
2) химерные рекомбинантные белки bLYM114, bLYM120, полученные согласно примерам 1 и 2, и химерный белок bLYM125, полученный согласно примерам 4 и 5, для обнаружения IgG.
Принцип теста VIDAS® состоит в следующем: конус образует твердую подложку, которая служит также системой подачи реактивов, содержащихся в полости конуса. Один или несколько рекомбинантных белков фиксируют на конусе. После стадии разбавления образец отбирают аспирацией и вводят за несколько приемов внутрь конуса. Данная процедура позволяет антиборрелиозным иммуноглобулинам образца связываться с рекомбинантными белками. Компоненты, оставшиеся несвязанными, удаляют промывкой. Антитело антииммуноглобулинов человека, конъюгированное с щелочной фосфатазой (PAL), инкубируют в конусе, в котором оно прикрепляется к антиборрелиозным иммуноглобулинам. На стадиях промывки удаляют конъюгат, оставшийся неприкрепленным. В ходе последней стадии обнаружения субстрат щелочной фосфатазы (PAL), представляющий собой 4-метилумбеллиферилфосфат, гидролизуют до 4-метилумбеллиферона, флуоресценцию которого измеряют при 450 нм. Интенсивность флуоресценции, измеренная посредством оптической системы Vidas®, пропорциональна содержанию антиборрелиозных иммуноглобулинов, присутствующих в образце. Результаты автоматически анализируются системой VIDAS® и выражаются в RFV (Relative Fluorescent Value (относительные единицы флуоресценции)).
Таким образом, посредством системы Vidas® были проанализированы 255 положительных проб сыворотки (пробы с неоднозначным результатом + пробы с положительным результатом) и 298 отрицательных проб сыворотки (пробы с неоднозначным результатом + пробы с отрицательным результатом).
Конусы Vidas® Lyme IgG сенсибилизировали 300 мкл раствора, содержавшего белки bLYM114, bLYM120 и bLYM125 по настоящему изобретению с концентрацией каждого из них в общем сенсибилизирующем растворе, равной 1 мкг/мл.
На первой стадии пробы сыворотки инкубировали в течение 5,3 мин для образования комплексов "антигены-антитела". На второй стадии анти-IgG человека, меченные PAL, инкубировали в течение 5,3 мин.
Результаты выражали в виде индекса по отношению к порогу положительного значения, составляющего 135 RFV согласно методике.
- Из 255 испытанных положительных проб сыворотки 246 найдены положительными и 9 ложноотрицательными, что соответствует чувствительности 96,5%.
- Из 298 испытанных отрицательных проб сыворотки 284 найдены отрицательными и 14 ложноположительными, что соответствует специфичности 95,3%.
СПИСОК ЛИТЕРАТУРЫ
1. Gottner G. et al., Int. J. Microbiol. 293, Suppl. 37, 172-173 (2004).
2. Arnaud N. et al., Gene 1997; 199:149-156.
3. Bretz A.G., K. Ryffel, P. Hutter, E. Dayer and O. Peter. Specificities and sensitivities of four monoclonal antibodies for typing of Borrelia burgdorferi sensu lato isolates. Clin. Diag. Lab. Immunol. 2001 ; 8: 376-384.
4. Ryffel K., Peter O., Rutti B. and E. Dayer. Scored antibody reactivity by immunoblot suggests organotropism of Borrelia burgdorferi sensu stricto, B. garinii, B. afzelii and B. valaisiana in human.J. Clin. Microbiol. 1999; 37:4086-92.
Реферат
Представленная группа изобретений касается слитых белков, нуклеиновых кислот, кодирующих такие белки, экспрессирующей кассеты, обеспечивающей экспрессию нуклеиновой кислоты, вектора, включающего такую кассету, способа диагностики in vitro-боррелиоза, набора для такой диагностики, в которых используют упомянутые белки, а также вакцинной композиции для профилактики боррелиоза, включающей такие белки. Охарактеризованные слитые белки включают в себя (i) по меньшей мере одну последовательность белка DbpA вида боррелии, выбранного из B. afzelii, B. burgdorferi sensu stricto и B. garinii, и (ii) по меньшей мере одну последовательность белка OspC вида боррелии, выбранного из B. afzelii, B. burgdorferi sensu stricto и B. garinii. Представленная группа изобретений позволяет проводить более чувствительные и специфичные анализы, связанные с присутствием нескольких патогенных видов Borrelia. 7 н. и 4 з.п. ф-лы, 8 табл., 7 пр.
Формула
(a) белок, аминокислотная последовательность которого включает на своем N-конце последовательность SEQ ID NO: 1 и на своем С-конце последовательность SEQ ID NO: 2;
(b) белок, аминокислотная последовательность которого включает на своем N-конце последовательность SEQ ID NO: 3 и на своем С-конце последовательность SEQ ID NO: 4;
(c) белок, аминокислотная последовательность которого включает на своем N-конце последовательность SEQ ID NO: 5 и на своем С-конце последовательность SEQ ID NO: 7;
(d) белок, включающий на своем N-конце последовательность SEQ ID NO: 6 и на своем С-конце последовательность SEQ ID NO: 7;
(e) белок, включающий на своем N-конце последовательность SEQ ID NO: 5, последовательность SEQ ID NO: 6 и на своем С-конце последовательность SEQ ID NO: 7;
(f) белок, аминокислотная последовательность которого включает последовательность, выбранную из последовательностей SEQ ID NO: 8, 9, 10, 11, 12, 13 и 14.
Комментарии