Α -амидирующая монооксигеназа (варианты) и способ получения a -амидированной формы регуляторного полипептида - RU2080385C1
Код документа: RU2080385C1
Чертежи
















Описание
Изобретение является частичным продолжением заявки на патент США Рег. N 655366, поданной 27 сентября 1984, полное описание которой включено в настоящую заявку путем ссылки.
Изобретение относится к альфа-амидирующим ферментам, производству альфа-амидирующих ферментов и их использованию в производстве альфа-амидированных продуктов путем воздействия ферментов на удлиненные глицином субстраты. В определенных предпочтительных вариантах осуществления альфа-амидирующий фермент согласно изобретению может быть использован при получении полезных альфа-амидированных гормонов и продуктов для использования в сельском хозяйстве и медицине, включая кальцитонины, факторов освобождения гормонов роста, пептид, связанный с геном кальцитонина, и другие альфа-амидированные продукты.
Состояние уровня техники
Внутриклеточная обработка (расщепление и/или функциональное изменение групп) предшественников нативных белков после трансляции кодирующей последовательности
нуклеиновых кислот ясно представлено в различных документах.
Обычно клетки млекопитающих или других эукариот могут осуществлять определенные операции посттрансляционной обработки, тогда как прокариоты не могут делать этого. Определенные прокариоты, например, такие как E. Coli, широко используются в качестве хозяев для получения белков млекопитающих с использованием технологии получения рекомбинантной ДНК (r ДНК), поскольку они могут быть легко выращены в процессе ферментации и поскольку они генетически точно охарактеризованы. Однако многие белки млекопитающих, полученные с помощью генной инженерии, требуют определенного типа посттрансляционной обработки, и это часто должно выполняться с использованием сложных химических операций in vitro, которые из-за их высокой стоимости ограничивают широкомасштабное промышленное применение.
Один из видов изменения включает в себя специальное амидирование аминокислот на карбоксильном конце белков. Многие встречающиеся в природе гормоны и пептиды содержат такое изменение, которое часто является существенным, если этот белок должен быть биологически активным. Примером является кальцитонин, где замещение неамидированного пролинового остатка на амидированный пролин нативной формы приводит к 3000-кратному уменьшению биологической активности.
Агент, который влияет на амидацию С-конца (альфа), узнает глициновый остаток, который следует непосредственно за аминокислотой, подвергаемой амидации (R-X-гли, где R основная последовательность белка, Х остаток, который подвергается амидированию и "гли" глициновый остаток). Глицин отщепляется и фактически отдает аминосоставляющую в предпоследнюю аминокислоту, в результате чего происходит ее амидирование.
Первыми авторами, сообщившими приблизительный молекулярный вес для альфа-амидирующего фермента, были Брэдбери и др. (Nature, т. 298, 1982, с. 686-88). Они высказали предположение, что минимальная кажущаяся молекулярная масса, определенная с использованием Сефадекса G-100, составляет приблизительно 60000 Дальтон (1 Дальтон=1,6601 х 10-27 кг).
Последующие исследования показали, что молекулярная масса такого фермента составляет от 60000 до 70000 Дальтон (при измерении с помощью гель-фильтрующей хроматографии). Эти исследования включают в себя работы Хусайни и Тэфта С.С. (FEBS Letters, т. 152, N 2, 1983, с. 277-281); Эйпер и др. (PNAS, т. 80, 1983, с. 5144-5148); Гомес и др. (FEBS Letters, т. 167, N 1, 1984, с. 160-164) и Кайзер Дж. С. и др. (PNAS, т. 81, 1984, с. 3228-3232).
Эйпер и др. (PNAS, т. 80, 1983, с. 5114-48) сообщил, что помимо молекулярного кислорода требуется еще два кофактора для максимальной активности амидирования фермента; ими являются аскорбиновая кислота и ион меди (II).
Химическая реакция, приводящая к амидированию карбоксильного конца пептида, требует источника аминогруппы. Брэдбери А.Ф. и др. (Nature, т. 298, 1982, с. 686-688) показал, что глицин расщепляется и отдает аминосоставляющую предпоследней аминокислоте, приводя к амидированию последней. Требование в отношении глицина как донора аминогруппы было подтверждено другими авторами.
Лэндимор и др. (BBRC, т. 117, N 1, 1983, с. 289-293) показали, что D-аланин также может служить в качестве аминодонора в реакции амидирования. Проведенная Кайзером и др. (PNAS, т. 81, 1984, с. 3228-3232) последующая работа показала две различные ферментные активности в мозгу крысы, которые способны обеспечивать катализ реакции альфа-амидирования. Образцы с большей молекулярной массой (70000 Дальтон) обладают специфичностью, ограниченной глицином у карбоксильного конца субстрата. Фермент с более низкой молекулярной массой превышает субстрат с бета-аланином в качестве последней аминокислоты на карбоксильном конце.
Оптимальное значение pH для альфа-амидирующего фермента, экстрагированного и частично очищенного из свиного гипофиза, по сообщению Брэдбери А. Ф. и Смита Д.Дж. (ВВRC, т. 112, N 2, 1983, с. 372-377), составляло приблизительно 7,0. Эйппер и др. (PNAS, т. 80, 1983, с. 5144-5148) подтвердили эти результаты сообщением об оптимальном pH величиной 7,0 для альфа-амидирующего фермента, который был частично очищен от крысиного гипофиза. Они также отметили, что ферментативная активность быстро спадает при pH ниже 6,5 или выше 7,5.
Во всех вышеуказанных публикациях (включенных здесь путем ссылки на них) экстракты и частично очищенные ферментативные препараты содержат дополнительно протеолитические ферменты, способные вызывать разрушение потенциальных субстратов и продуктов, а также и самих альфа-амидирующих ферментов, таким образом замедляя амидирование пептидов и полипептидов, выделенных очисткой из природных источников или полученных с использованием технологий рекомбинатной ДНК.
В широком смысле все ранее измеренные активности амидирования другими были основаны на преобразовании D-субстратов, таких как трипептиды, D-Тир-Вал-Гли-COOH, в D-Тир-Вал-CO-NH2. Из двух возможных конфигураций (D или L) важнейшие природные аминокислоты представлены в форме L. Однако использование формы D исследователями требовалось, чтобы нейтрализовать присутствие посторонних протеолитических ферментов в загрязненных препаратах амидирующих ферментов, использованных этими экспериментаторами. Эти посторонние ферменты могут иметь ярко выраженное протеолитическое влияние на субстраты с L-аминокислотой, одновременно оказывая слабое влияние на D-субстраты. Исследователи, обремененные проблемами, связанными с наличием протеолитических и других включений в их фермент, использовали ненатуральный D-субстрат, чтобы избежать влияния загрязнений. Никто до настоящего времени не смог показать, что их ферментативные препараты могут эффективно амидировать любые физиологические соответствующие субстраты, т.е. L-субстраты для преобразования в биологически активные альфа-амидированные L-продукты.
Как показано здесь, препараты, полученные согласно настоящему изобретению, могут эффективно амидировать L-сусбтраты и в отношении D-субстратов иметь активность от 60 до 1000 раз более высокую, чем наивысшая активность, указанная в любом ранее известном опубликованном источнике, который известен заявителям.
Ферментативные препараты, которые способны амидировать карбоксильные концы пептидов и белков, известны из различных опубликованных источников. Например, Брэдбери А. Ф. (Nature, т. 298, 1982, с. 686-688) сообщает, что альфа-амидирующая активность присутствует в гипофизе свиней. Препарат из свиного гипофиза, содержащий фермент, был способен преобразовывать пептиды, которые оканчиваются глицином, в соответствующий безглициновый амид пептида. Брэдбери и др. указывают, однако, что препараты не будут амидировать пептиды или полипептиды, выделенные очисткой из природных источников.
Система определения и оценки активности амидирования в гипофизе были основаны на исследовании способности ферментативных препаратов преобразовывать синтетический трипептид D-тирозилвалилглицин в соответствующий амид дипептида D-тирозилвалинамид. D-тирозиновый остаток давал устойчивость к разрушению под действием аминопептидаз, присутствующих в тканевых гомогенатах. Контрольные эксперименты показали, что когда синтетический D-тирозилвалинамид был инкубирован в таких же условиях, он медленно разрушался. Таким образом, образование дипептид амида с помощью гипофизного фермента сопровождалось его разрушением другими ферментами, присутствующими в экстракте гипофиза (с. 686).
Таким образом, Брэдбери и др. показывают, что описанные препараты содержат другие протеолитические ферменты, которые разрушают пептиды или полипептиды, и что неприродный D-тирозиновый остаток был использован, чтобы свести к минимуму такое разрушение.
Кроме того, хотя Блэдбери и др. указывают на использование для очистки амидирующего фермента, гомогенизации или субклеточного фракционирования и последующей гель-фильтрующей хроматографии, он все же остается загрязненным протеолитическими ферментами.
Хусайн И. и Тэйт С. С. (FEBS Letters, т. 152, N 2, 1983, с. 277-281) описывает альфа-амидирующую активность в нейросекреторных железах гипофиза быка.
Эйпер и др. (PNAS, т. 80, 1983, с. 5144-5148) сообщили о наличии альфа-амидирующей активности в передней, и промежуточной и задней долях желез крысиного гипофиза и бычьего промежуточного гипофиза. Однако, в этой ссылке имеется только указание на применение синтетического D-Тир-Вал-Гли в качестве субстрата для исследования альфа-амидирующей активности и признание наличия загрязнений получаемых препаратов.
Гомес и др. (FEBC Letters, т. 167, N 1, 1984, с. 160-164) определили, что гипоталамус крыс также обладает альфа-амидирующей ферментативной активностью.
Брэдбери А. Ф. и Смит Д.Д. в статье Структура пептидов и их функции: Сообщения Восьмого американского симпозиума по пептидам, с. 249-252 (1983), редакторы Хруби В.Дж. и Рич Д.Х. описывают наличие активности альфа-амидирующего фермента в щитовидных железах крыс.
Мэйнс и др. (Endocrinology, т. 114, 1984, с. 1522-1530) сообщили, что относящаяся к мышам или крысам клеточная линия, полученная из передней доли гипофиза (АТТ-20), обладала альфа-амидирующей ферментативной активностью, которая явно снижалась с течением времени в культуре.
Железы или органы, о которых известно, что они содержат амидированные пептиды, могут содержать фермент, способный катализировать реакцию амидирования. Так, низшие формы жизни, например, морская собака (Squalus acanthias), по сообщению Одонохью Т.Л. и др. (Peptides, N 3, 1982, с. 353-395), содержат амидированные пептиды в экстрактах гипофиза. Шеллер Р.Х. и др. (Cell, т. 32, 1983, с. 7-22) сообщили о наличии сигнальных пептидов амидирования у морских улиток. Несмотря на кажущееся всемирное распространение этого вида активности в природе, мало информации опубликовано по вопросу физико-химических характеристик ферментов. Это может быть отнесено к очень низким уровням присутствия ферментов в этих нейроэндокринных органах.
Наличие амидированных пептидов в определенных тканях не обязательно связано с высокими уровнями альфа-амидирующего фермента. Например, ткань передней доли гипофиза крыс обладает высокой альфа-амидирующей активностью, но содержит неизвестные субстраты (Эйпперт и др. PNAS 80, с. 5144-5148, 1983). Ткань задней доли гипофиза крыс содержит амидированные пептиды (окситоцин и вазопрессин), но обладает очень низкой альфа-амидирующей активностью (Эйпперт и др. Endo, 116, с. 2497-2504, 1985). Поэтому, пока отдельные ткани не будут испытаны в отношении альфа-амидирующей активности, существующие или потенциальные уровни фермента не могут быть предсказаны.
Целью изобретения является создание альфа-амидирующих ферментативных препаратов, которые могут обеспечить эффективное производство полезных альфа-амидированных продуктов даже из субстратов, содержащих L-аминокислоты, таких как пептидные или полипептидные субстраты, выделенные очисткой из природных источников или полученные с помощью технологии получения рекомбинантной ДНК.
Дополнительной целью изобретения является создание эффективного и экономичного способа получения ферментативного препарата.
Дополнительной целью является создание моноклональных антител, специфичных в отношении альфа-амидирующего фермента, и иммобилизированных антител, смол для очистки, колонок для проведения хроматографии по иммуносродству и им подобных, которые при использовании указанных антител эффективно очищают альфа-амидирующей фермент.
Дополнительной целью изобретения является получение альфа-амидированных продуктов из субстратов, содержащих глициновый остаток на C-конце, за счет реакции субстрата в присутствии ферментативного препарата.
Поставленная цель достигается прежде всего путем получения новых альфа-амидирующих ферментов (монооксигеназ) такой чистоты, чтобы проявить удельную альфа-амидирующую активность величиной по меньшей мере 25 мЕ на мг белка, и предпочтительно, выше 50 или выше 150 мЕ/мг протеина. Все единицы удельной активности, указанные здесь, приведены в пересчете на преобразование Дансил-Д-Три-Гли-COOH в Дансил-Д-Тир-Вал-CONH2. Одна мЕ определяется как величина активности, необходимая для преобразования одного наномоля (нмоль) Дансил-Д-Тир-Вал-Гли-COOH в один наномоль Дансил-Д-Тир-Вал-Гли-CONH2 за минуту при 37oC и pH 7,0 в присутствии ионов аскорбиновой кислоты при концентрации 3 мМ (в пересчете на суммарную реакционную смесь, включая фермент, субстрат, кофакторы и т.д.), молекулярного кислорода в молярном избытке относительно субстрата и концентрации ионов меди, достаточном, чтобы максимизировать активность (обычно около 2 мМ в зависимости от чистоты фермента).
Альфа-амидирующие ферменты согласно изобретению, такие как пептилглицин-альфа-амидирующая монооксигеназа, способны обеспечить катализ превращения пептидного соединения, имеющего глициновый остаток по крайней мере у C-конца пептидной цепи, в соответствующее пептидил-амидное соединение, имеющее на C-конце аминогруппу вместо глицина. В том смысле, в котором здесь используется термин пептидная цепь, он означает любой полипептид, имеющий по меньшей мере две аминокислоты, соединенные между собой с помощью пептидной связи. Термин пептидное соединение включает в себя любое соединение, имеющее пептидную цепь. Термин соответствующий пептидиламин касается любого продукта реакции, в котором аминогруппа замещает глицин на C-конце пептидной цепи.
Целесообразно, чтобы реакция альфа-амидирования происходила в присутствии кислорода и восстанавливающего вещества. Такие восстановители могут включать в себя, но не ограничиваются, аскорбиновую кислоту, соли аскорбиновой кислоты, дигидроксифумарат, металлические соединения цианида, тетрагидроптерин. Было установлено, что определенные кофакторы способствуют течению реакции и/или замедляют разрушение или снижение активности фермента. Эти кофакторы включают в себя, но не ограничивают, катализу, этанол, калий, иодид и ионы меди. Очищенные ферментативные композиции согласно изобретению предпочтительно свободны от протеолитической активности, способной вызывать разрешение альфа-амидирующего фермента или продуктов или реактантов реакции альфа-амидирования, так что ферментативные композиции могут обеспечивать катализ реакции альфа-амидирования, даже когда субстрат и продукт содержат L-аминокислоты. Протеолитическая активность в смеси реакции амидирования может не прямо отражать концентрацию протеазы. Активность, даже при высоких концентрациях протеазы, может быть подавлена, например, за счет воздействия различных ингибиторов. Недостаток протеолитической активности, которая увеличивает промышленную и практическую привлекательность ферментного препарата при использовании субстратов, содержащих L-аминокислоты, ни коем образом не снижает возможности использования композиции с субстратами, которые не содержат L-аминокислоты.
Как будет подробно описано здесь далее, заявители провели очистку огромного количества образцов специфических белков, которые обладают альфа-амидирующей активностью. "Альфа-амидирующая активность" в том смысле как она здесь используется, означает любую активность, имеющую тенденцию оставлять только одну аминогруппу в положении, ранее занимаемом глицином на C-конце субстрата. Такое замещение может включать в себя отщепление всего, кроме аминогруппы, глицина так, что только аминогруппа остается в положении, ранее занимаемой половиной глицина. "Альфа-амидирующий фермент" в том смысле, как он здесь используется, относится к очищенному соединению, которое проявляет альфа-амидирующую активность.
Ферментные препараты, выделенные в соответствии с настоящим изобретением, могут быть очищены до тех пор, пока не станут однородными. В том смысле, как здесь используется термин "однородный", относится к ферментным препаратам, проявляющим единственную четко определенную полосу после электрофореза на натрий додецил сульфат/полиакриламидном геле и обнаруживающим единственную последовательность аминокислот в ответ на обычные приемы исследования последовательностей.
Конкретные альфа-амидирующие ферменты, очищенные до однородного состояния в соответствии с настоящим изобретением, проявляли удельные ферментативные активности выше, чем 1500 мЕ/мг. белка.
a-амидирующие ферменты (монооксигеназы), приготовленные в соответствии с настоящим изобретением, а в ряде случаев и частично очищенные препараты, могут быть использованы при производстве полезных альфа-амидированных пептидных продуктов путем применения их в качестве для катализатора альфа-амидирования. В качестве субстрата используется пептидное соединение, имеющее глициновый остаток по меньшей мере у C-конца пептидной цепи, где замещение глицином с C-концом аминогруппы приводит к получению требуемого продукта. Субстрат обрабатывается ферментом, полученным в соответствии с настоящим изобретением предпочтительно в присутствии кислорода и восстанавливающего агента в течение времени, достаточного для преобразования пептидильного соединения в соответствующий пептидиламид. Преобразование начинается по существу при первом контакте субстрата и фермента, но скорость реакции в значительной мере изменяется в зависимости от pH, температуры, сродства субстрата, концентрации кофакторов и других параметров, которые могут изменяться известным образом, чтобы оптимизировать реакцию. Реакция обычно должна развиваться в течение времени, выбранного с учетом требуемого процента преобразования (субстрата в продукт). В предпочтительных вариантах осуществления кофакторы, такие, как те, что рассмотрены выше, используются, чтобы способствовать развитию реакции и/или усилению или поддержанию активности фермента.
Огромное количество полезных продуктов, включая природные гормоны и им подобные, для которых альфа-амидирование предпочтительно или необходимо, может быть получено путем проведения реакции удлиненных глицином пептидильных соединений в присутствии альфа-амидирующих ферментативных композиций в соответствии с настоящим изобретением. Эти продукты, которые могут быть использованы, например, в сельском хозяйстве или при медицинском лечении болезней, характеризуемых дефицитом гормонов, включают в себя, но не ограничены, различные кальцитонины, факторы высвобождения гормонов роста, пептиды, связанные с геном кальцитонина и им подобные. Гормоны, на которые здесь приводится ссылка, такие, как вышеуказанные кальцитонины, факторы высвобождения гормонов роста и пептиды, связанные с геном кальцитонина, включают в себя разновидности амидированных на C-конце белков, которые проявляют характеристическую активность указанных гормонов, как это ясно сведущим в этой области специалистам. Например, кальцитонин включает все виды, которые проявляют способность регулировать усвоение кальция в костных тканях, что является характеристикой известных кальцитонинов. И для других белковых видов, рассматриваемых здесь, гомологи включены в определения этих видов. Целесообразно, чтобы по меньшей мере 40% а наиболее целесообразно, чтобы 50% аминокислот пептида соответствовали тем, что заявлены.
Способность полученных по изобретению a-амидирующих ферментов (монооксигеназ) катализировать реакцию альфа-амидирования даже в отношении субстратов, содержащих L-аминокислоты, обеспечивает возможность эффективного и экономически выгодного производства полезных продуктов с использованием субстратов, получаемых с помощью очистки из природных источников или с помощью технологии рекомбинантной ДНК.
В определениях, предпочтительных вариантах осуществления, реакция альфа-амидирования может быть облегчена путем иммобилизации фермента на твердом носителе, который является нерастворимым в водной среде и стойким к разрушению в условиях реакции, и пропускания субстрата через иммобилизованный фермент, предпочтительно в присутствии кофакторов. В том смысле, как здесь используется иммобилизация, подразумевается связь фермента с подложкой или носителем. Носители, которые могут оказаться полезными для этой цели, включают в себя, но вовсе не ограничены этим, пористое стекло или активированный абсорбент, например, сефарозу, активированную бромцианом. Иммобилизированные таким образом ферменты могут быть многократно использованы путем удаления реакционной смеси с твердого носителя, который продолжает удерживать фермент для последующего использования.
Заявители открыли, что ферментные препараты в соответствии с настоящим изобретением могут быть получены и очищены с помощью целого ряда способов. Установлено, что мягкая сердцевидная ткань пораженной раком (карциномой) щитовидной железы, предпочтительно полученная от крыс, клеточные линии ее и/или среда культуры клеток из указанных клеточных линий являются особенно желательными источниками исходного альфа-амидирующего фермента. Загрязненный альфа-амидирующий фермент может быть очищен из исходного сырого препарата как методом хроматографии исключения (вытеснительной хроматографии), так и анионообменной хроматографией, предпочтительно сильной анионообменной хроматографией. В том смысле как здесь используется "сильная" анионообменная хроматография относится к анионообменной хроматографии, проводимой на любой смоле, которая сохраняет постоянный средний положительный заряд в диапазоне pH 2-12. В определенных предпочтительных вариантах осуществления настоящего изобретения хроматография исключения по размерам предшествует сильной анионообменной хроматографии либо предшествует каждому из повторяющихся этапов анионообменной хроматографии. Может применяться и вторая сильная анионообменная хроматография, следующая за первой и в одном предпочтительном варианте осуществления первая сильная анионообменная хроматография или вторая сильная анионообменная хроматография проводятся при основной pH, тогда как другая проводится при кислотном pH.
Используя ферментативные пробы, которые в значительной мере очищены до достижения однородности в соответствии с настоящим изобретением, получали моноклональные и поликлональные антитела, специфичные в отношении фермента, чтобы вызывать иммунную реакцию у мышей и цыплят соответственно. Антитела, полученные таким образом из любых видов, могут быть очищены и иммобилизованы на твердотельном носителе с целью получения колонки для проведения хроматографии по иммуносродству, специфичной для фермента. Эта колонка может быть использована для очистки исходного ферментативного материала, приводя к повышенной эффективности и возможности повторного использования фермента.
Массовое промышленное производство ферментов из природных источников может быть облегчено с помощью методов очистки, включающих в себя как вытеснительную хроматографию, так и анионообменную хроматографию, где ферментативные виды, удерживаемые анионообменной хроматографической колонкой, элюируются с использованием физиологического раствора, имеющего концентрацию выше, чем 250 мМ и предпочтительно выше 350 мМ или 500 мМ. При высокой концентрации физиологического раствора наиболее удерживаемые ферментативные виды будут элюированы. Хроматография исключения по размеру должна быть предусмотрена, чтобы отделять виды с кажущимся молекулярным весом от примерно 58000 до 67000 Дальтон и предпочтительно от примерно 60000 до около 65000 Дальтон. Очищенный препарат способен амидировать пептидильное соединение, которое было получено путем очистки из природных источников или получено с использованием технологии рекомбинантной ДНК, т.е. содержало L-аминокислоты.
Фиг. 1-5 относятся к примеру 1 и поясняют его; фиг.6-12 к примеру 2 и поясняют его; фиг.13-16 к примеру 3 и поясняют его; фиг.17-21 к примеру 4 и поясняют его; фиг.22 к примеру 6 и поясняет его.
Подробное описание определенных предпочтительных вариантов.
Теперь было открыто, что однородный альфа-амидирующий фермент может быть получен с помощью многоэтапного способа, использующего сочетание хроматографии исключения по размеру и ионообменной хроматографии из экстрактов твердых тканей опухолей, клеточных линий опухолей и тканевой культуры из таких клеточных линий.
Фермент экстрагировали из мозговидной карциномы щитовидной железы крыс (МТС), полученной у крыс Вистар, описанных Рузом В.А. и др. (Endocrinology, 1979, т. 150, N 1, с. 27-32). Эта ткань депонирована как IVI-10028. Фермент также экстрагировался из других источников, особенно линий клеток карциномы щитовидной железы крыс. Линия клеток крыс (CA-77) была получена из карциномы щитовидной железы крыс с помощью нескольких пассажей, как описано Мужинским и др. (JBC 1983, т. 258, с. 11678-83). Эта линия клеток депонирована как IVI-10029. Линия клеток человека (НТТ 54/34/) была разработана Рузом в медицинском центре Кливленда, штата Огайо, с использованием мозговидной карциномы щитовидной железы человека для первичной культуры. Линия клеток (НТТ 54/34/) человека была депонирована в качестве экспоната IVI-10031 (см. признание депозита микроорганизмов для целей патентования, свидетельствующее об этих депонентах, и Будапештское свидетельство N 34, 3 ноября 1983, включенные в настоящее описание полностью путем ссылки).
Было показано, что среды культивирования тканевых культур, полученных как от линий клеток человека, так и крыс, обладали значительными уровнями активности альфа-амидирующего фермента, указывая, что часть фермента выделена из клеток. Фермент может быть получен и очищен предварительной обработкой исходного материала (включая расходуемую культурную среду, фермент и включения) анионообменной хроматографией. Проба, например, может быть загружена на препаративный анионообменник, такой как колонка с DEAE типа CUNO 250, поставляемая в продажу фирмой CUNO Корп.
Удаляемый из этой колонки препарат, обладающий альфа-амидирующей активностью, затем подвергают обработке хроматографией исключением на смоле соответствующего разрешения, например, сверхтонкой колонке Сефакрил S-200, которая поставляется фирмой Фармация Файн Кемикал.
Обладающая активностью выделенная фракция затем подвергается ионообменной хроматографии с использованием силовой анионообменной матрицы. Смола, которая может быть в этом случае использована, это сильная анионообменная смола Моно QHR5/5 фирмы Фармация Файн Кемикал, и для однородной очистки фермента может потребоваться одно или более пропусканий через колонку. Колонка Моно QHR 5/5 имеет размер частиц 10 мм1, объемы пор 40% гель, заряженная группа которого CH2-N+(CH3)3, и ионообменную емкость 0,28-0,36 ммоль/мл.
Каждый этап очистки может контролироваться как в отношении содержания белка, так и уровня альфа-амидирующей активности. Эта информация используется для вычисления удельной активности фермента, которая служит индикатором относительной частоты фермента.
Пептидил-глицин альфа-амидирующая монооксигеназа, очищенная в соответствии с настоящим изобретением (депонированный полученный из крыс фермент IVI-10032; депонированный полученный от человека фермент IVI-10033) имеет кажущийся молекулярный вес около 60000-65000 Дальтон, определенный гель-фильтрацией.
Фермент был очищен так, что он проявлял удельную ферментативную активность по меньшей мере приблизительно 25 мЕ/мг белка и предпочтительно по меньшей мере приблизительно 50 мЕ/мг белка. Удельная активность выше около 150 мЕ/мг белка особенно целесообразна. Альфа-амидирующий фермент был также очищен так, что проявлял единственную однородную точно определенную полосу после электрофореза на натрий додецил сульфат-полиакриламидном геле (SDS-PAGE).
Очищенная пептидил-глицин альфа-амидирующая монооксигеназа используется для амидирования альфа-карбоксильной группы полипептида, имеющего концевой глициновый остаток, где глицид работает в качестве донора аминогруппы. Пептид или полипептид субстрата может быть выделен путем очистки из природных источников, синтезирован из его аминокислот или получен с использованием технологии получения рекомбинантной ДНК. Полипептид с глициновым концом соединяется с кислородом в присутствии эффективного количества фермента. Количество фермента, которое требуется при этом, зависит от нескольких переменных, хорошо известных в этой области, включая в себя, в частности, но не ограничиваясь ими, следующие: удельную активность данного ферментативного препарата, количество и химическую природу преобразуемого субстрата, время, в течение которого должно происходить, температуру и pH реакционной смеси. Сведущим в этой области специалистам известны другие переменные, которые могут оказать влияние на точное количество фермента, требуемого в данной ситуации.
Кислород обычно присутствует в реакции в молярном избытке относительно концентрации субстрата. Требуемая концентрация ионов меди может быть обеспечена с помощью любой соли, анион которой не оказывает вредного влияния на реакцию. Для фермента, имеющего удельную ферментативную активность только около 1 мЕ/мг белка, максимальное альфа-амидирование имеет место при относительно высокой концентрации (около 4 мМ) ионов меди. Когда чистота фермента увеличивается, требования к концентрации для экзогенного (внешнего) иона меди снижается. Ферментативная активность может быть также усилена за счет наличия аскорбат ионов, которые могут быть получены с помощью любой соли аскорбиновой кислоты, пока катион соли не оказывает ухудшающего воздействия на реакцию. Для очищенного фермента, имеющего удельную ферментативную активность приблизительно 50 мЕ/мг белка, максимальная активность альфа-амидирования наблюдалась при концентрации около 5 мМ аскорбата. Альфа-амидирующая активность может быть увеличена путем добавления каталазы. Оптимальное значение pH для преобразования биологически соответствующего субстрата в амидированные продукты составляет от 6,5 до 7,5.
Моноклональные и поликлональные антитела, направленные против фермента, были получены с использованием однородного фермента в качестве антигена, чтобы получить иммунную реакцию мышей и цыплят, соответственно. Как моноклональные, так и поликлональные антитела, были получены и очищены заявителями, как это показано в примере 8. Антитела, специфичные к альфа-амидирующему ферменту, хранятся в лабораториях заявителей.
Антитела могут быть иммобилизованы на твердой матрице, которая не растворима в среде, в которой она используется. Целесообразно, чтобы матрица была стойкой к разрушению. Матрица снабжена функциональной группой, способной связывать белки, где функциональные группы затем ковалентно соединены с антителами. Такая иммобилизация антител будет облегчать отделение альфа-амидирующего фермента из природных и/или рекомбинантных источников. Это осуществляется путем смешивания иммобилизированных антител с исходными препаратами фермента. Антитела будут, в частности, связывать молекулы альфа-амидирующего фермента. Загрязняющие белки не будут соединяться с антителами и легко удаляются путем элюирования или легкого центрифугирования. После удаления загрязнений a-амидирующий фермент может быть удален с иммобилизованных антител путем изменения ионной силы или pH или путем добавления хаотропных ионов (Хроматография сродства: Принципы и Способы, Руководство, Фармакия Файн Кемикалс, Уппсала, Швеция) и извлечен в очень хорошо очищенном виде.
Фермент был также достаточно очищен, чтобы определить его аминокислотную последовательность.
Хотя настоящее изобретение описывается в связи с предпочтительными вариантами его осуществления, многие изменения и модификации станут очевидны сведущим в этой области специалистам. Предпочтительные варианты осуществления изобретения далее приведены на примерах.
Пример 1
Процедура очистки препарата альфа-амидирующего фермента из опухолей МТС (Мозговой
карциномы щитовидной железы).
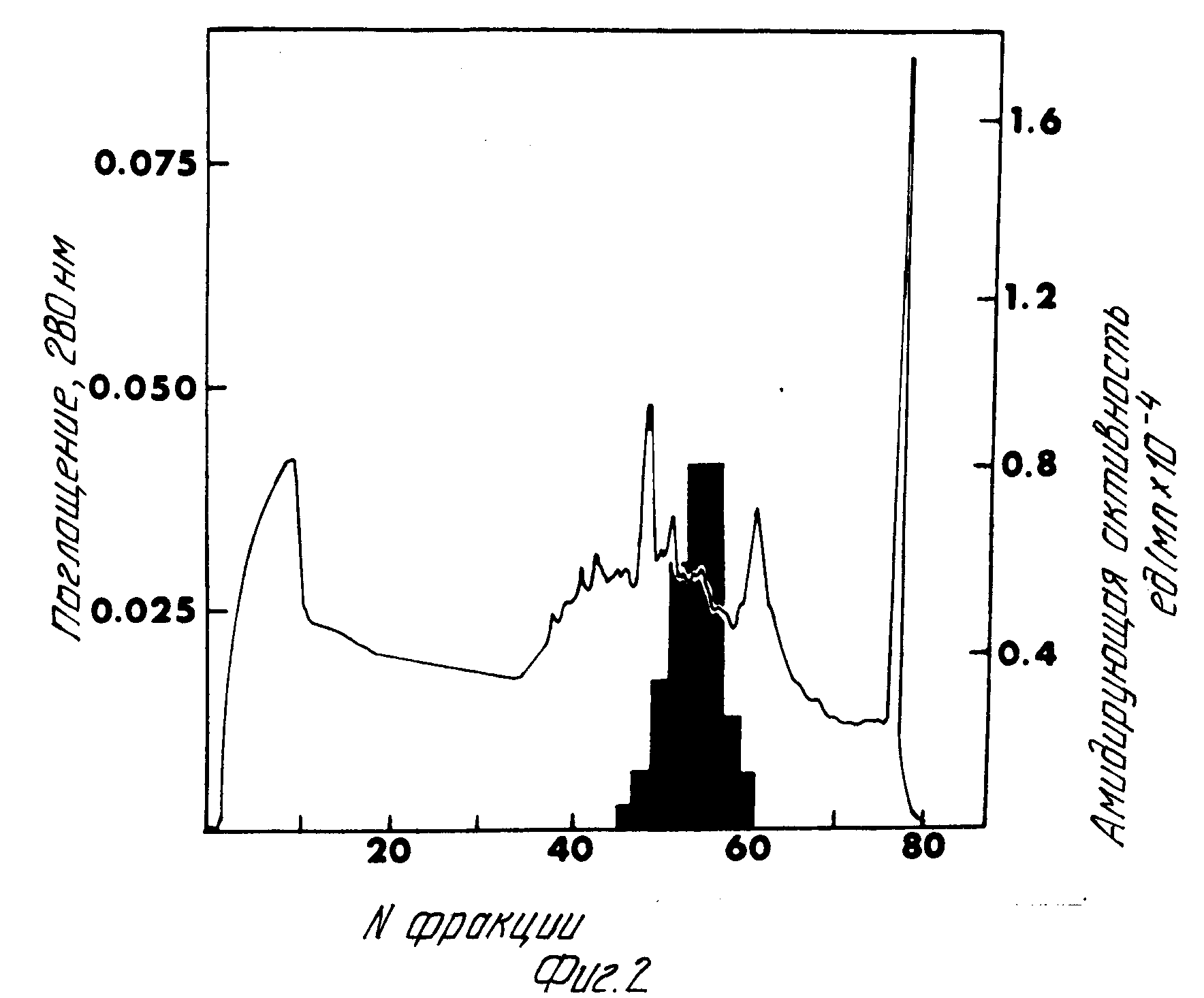
Замороженные опухоли крыс были распылены в виде мелких фрагментов и гомогенизированы в 150 мМ Трис pH 7,5, содержащем 250 мМ сахарозы, 0,25% N-ацетилгликопиранозида, 0,1 мМ PMSF, 20 μг/мл пепстатина и 100 μг/мл ингибитора в виде соевого масла. Обычно 25 г опухолевой ткани гомогенизировались в 200 мл вышеуказанного буфера на льду. Гомогенизация достигалась с помощью четырех импульсов по 20 с каждый с использованием гомогенизатора Политрон (Бринкмана) при насадке 7. Гомогенат центрифугировался в течение 15 мин при 9000 g в роторном устройстве JA-20 (БЕКмана), и супернатант (надосадочная часть) (Суп. 1) декантировался. Гранулы повторно гомогенизировались в 60 мл буфера для гомогенизации в течение 3 импульсов по 20 с каждый. Этот второй гомогенат был аналогично центрифугирован в течение 15 мин при 9000 g. Супернатант этого центрифугирования (Суп. 2) был соединен с супернатантном 1. Соединенные супернатанты были затем центрифугированы в течение 30 мин при 100000 g в роторе SW28 (Бэкман) при 4oC. Для осветления гомогената (Суп. 3) был добавлен кристаллический аммоний-сульфат в количестве достаточном, чтобы получить 25% -ный раствор аммоний-сульфата. Добавление кристаллов осуществлялось постепенно в течение 15 мин при постоянном перемешивании гомогената при температуре 4oC. Через 30 мин перемешивания при 4oC смесь была центрифугирована в течение 15 мин при 27000 g в роторе JA-20 (Бэкман) при 4oC. Супернатант был слит, и дополнительное количество сульфата аммония было добавлено к супернатанту, чтобы довести его до содержания аммоний-сульфата 40% После дополнительных 30 мин перемешивания смесь была вновь центрифугирована в течение 15 мин при 27000 g как было указано выше. Супернатант этого центрифугирования был отброшен, и осадки, которые обладали по меньшей мере 50% суммарной активности фермента гомогената, были повторно взвешены в 50 мМ Трис HCl pH 7,0. Для получения проб. Активность составляла 8,2 мЕ/мг белка. Гель-фильтрационная хроматография исключения по размерам на Сефакрил S-300. Сефакрил S-300 (Фармация) был уравновешен 50 мМ Трис HCl с pH 7,0 и залит в колонку K 50/100 (Фармация) в соответствии с инструкциями изготовителя. Объем слоя колонки составлял приблизительно 1,3 л. Проба была загружена на колонку в объеме, составляющем приблизительно 3% объема слоя колонки. Белки были элюированы из колонки 50 мМ Трис HCl pH 7,0 при расходе потока приблизительно 2 мл/мин. Фракции (10 мл) были собраны и исследованы в отношении альфа-амидирующей активности с использованием субстрата Дансил-Д-Тир-Вал-Гли. Элюированный с колонки фермент в единственном пике активности, показанном на фиг.1, имел молекулярную массу 60000-65000 Дальтон. Активность составляла 50 мЕ/мг белка.
Хроматография Моно Q при pH 6,0 сильная анионообменная хроматография.
Фракция из колонки с Сефакрил S-300, обладающие максимальной ферментативной активностью, были отстоены и подвергнуты диализу против 4 л 20 мМ бис-Трис pH 6,0 буфера. Полученный отстой был затем загружен в колонку Моно Q HR 5/5 (Фармация), которая была предварительно обработана согласно инструкциям изготовления и уравновешена 20 мМ бис-Трис pH 6,0. После того, как белок, не соединяющийся с колонкой, был собран в элюенте колонки (в сквозном потоке), связанные белки были элюированы при линейном солевом градиенте от 0 до 300 мМ в 20 мМ бис-Трис pH 6,0. Колонка работала при расходе 0,5 мл/мин и фракции (2 мл каждая) были собраны. Раствор белков, вытекающих из колонки Моно Q при pH 6,0, был немедленно нейтрализован путем сбора в пробирки, содержащие 1,0 М Трис pH 7,0. Фракции были исследованы в отношении альфа-амидирующей активности фермента, как прежде, и пик активности наблюдался при элюировании из колонки приблизительно 160 мМ NaCl (фиг.11). Активность составляла 161 мЕ/мг белка.
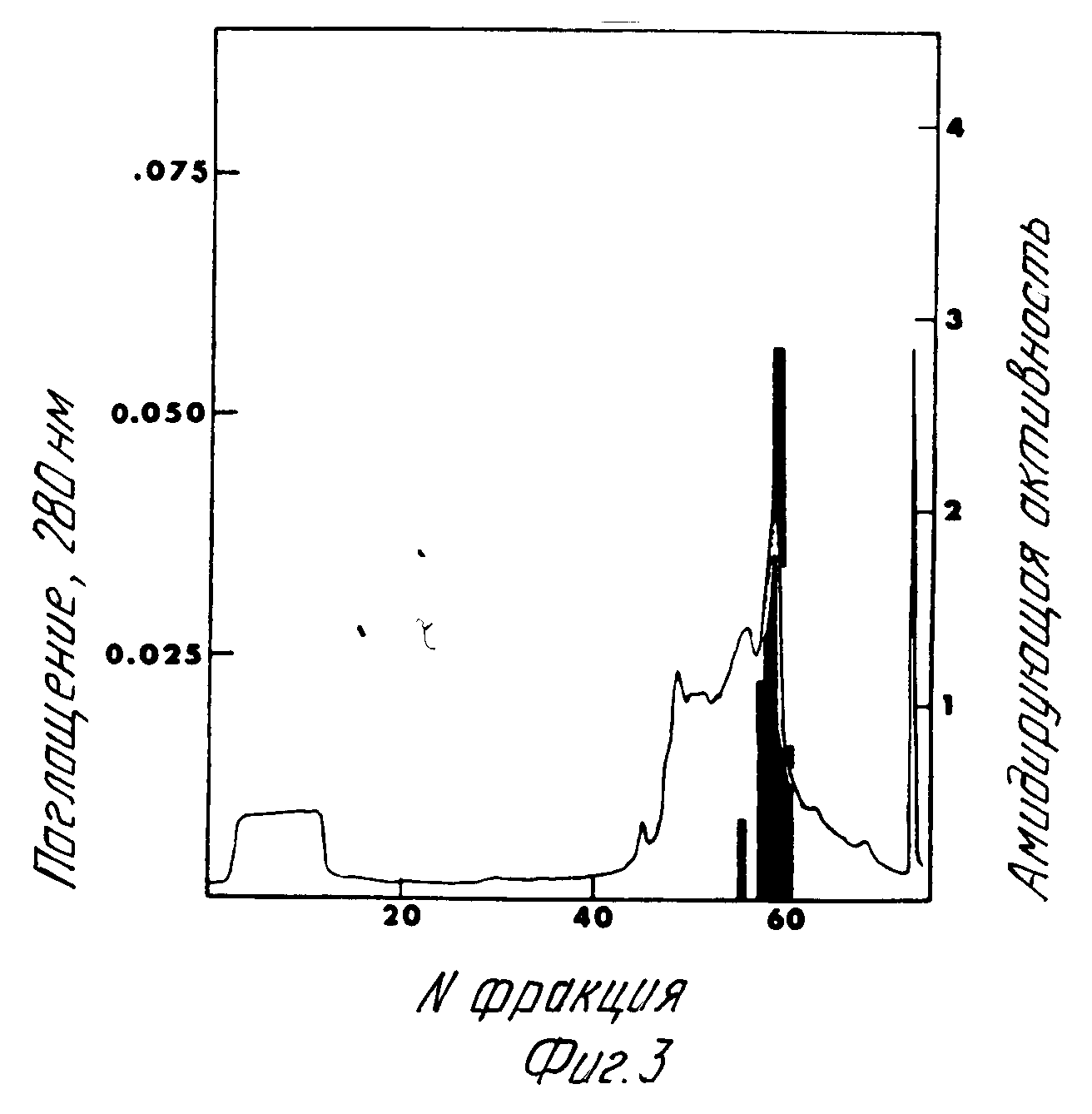
Хроматография Моно Q при pH 8, 0.
Фракции, обладающие пиковой альфа-амидирующей ферментативной активностью при хроматографии Моно Q при pH 6,0, были подвергнуты диализу при двух сменах (по 4 л каждая) 50 мМ Трис HCl pH 8,0. Фермент затем загружен на колонку Моно Q HR 5/5 (Фармация), уравновешенную 50 мМ Трис HCl pH 8,0. Белки были элюированы при солевом градиенте 0 300 мМ NaCl в 50 мМ Трис HCl pH 8,0. Был использован расход 0,5 мл в минуту и собирались 2 мл фракции. Каждая фракция была нейтрализована путем добавления 200 μл 1,0 М Трис pH 7,0 в каждой приборке. Наблюдались два различных пика активности фермента. Элюирующиеся при 190 мМ NaCl и 220 мМ NaCl, соответственно (фиг.3). Активность составляла по меньшей мере 80 и 400 мЕ/мг белка.
Гель-электрофорез главной фракции фермента хроматографии на Моно Q при pH 8,0.
Равные части "пиковой" фракции фермента, вытекающей при 220 мМ NaCl, имеющей активность 408 мЕ/мг белка, были загружены на 10% SDS полиакриламидный гель после тепловой денатурации в буфере, содержащем SDS и β-меркаптоэтанол (фиг.4).
Наблюдалась единственная полоса из фракции при 220 мМ NaCl. Кажущаяся молекулярная масса этой полосы составляет приблизительно 73000-75000 Дальтон.
Подтверждение подлинности полосы 73000-75000 Дальтон в качестве альфа-амидирующего фермента.
Из дополнительного препарата альфа-амидирующего фермента, получаемого путем очистки с использованием критерия, идентичного тем, что описаны выше, одинаковые части были использованы для электрофореза на неденатурирующем 10% акриламидном геле. Равные дозы были загружены на две полоски. После электрофореза белок, находящийся на одной из полосок, был визуализирован с использованием технологии окраски серебром. Другая полоска была нарезана на слои по 3 мм, и каждый срез был помещен в одно гнездо микротитерной пластины. В каждое гнездо было добавлено 100 мл смеси, содержащей субстрат Дансил, каталазу, аскорбиновую кислоту и 150 мМ Трис pH 7,0. Инкубация проводилась в течение 16 ч при 37oC и постоянном встряхивании. Помещенные в каждое гнездо части затем проанализированы в отношении преобразования субстрата в амидированный продукт. Ферментативная активность была обнаружена в двух срезах геля. Активность согласовывалась с полоской сильно окрашивающего белка в соответствующей окрашенной гельной полоске (фиг.5).
Пример 2
Производство рекомбинантного человеческого кальцитонина.
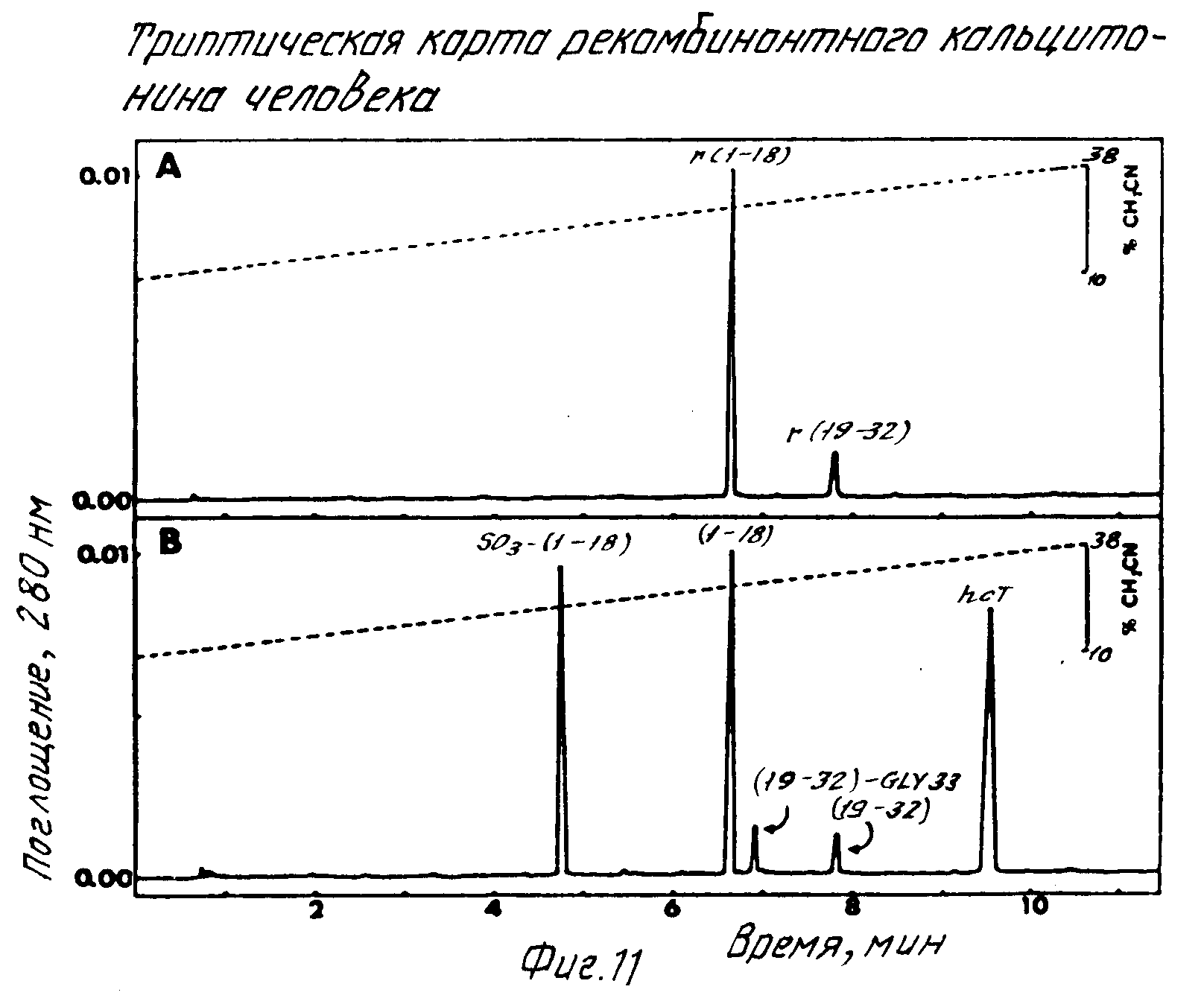
Человеческий кальцитонин это 32-аминокислотный пептидный гормон, имеющий амидированный пролиновый остаток у его карбоксильного конца. Микроорганизм был получен с использованием генной инженерии для получения рекомбинантного искусственного белка, содержащего аминокислотную последовательность, соответствующую человеческому кальцитонину. Ген слитого был подобран так, чтобы последовательность человеческого кальцитонина была фланкирована аргининовым остатком на его аминоконце, а на карбоксильном конце глициновым остатком, который одновременно заканчивал рекомбинатный слитый белок (фиг.7). После связывания гена человеческого кальцитонина в плазмиду для образования соответствующей искусственной плазмиды микроорганизм трансформировался с помощью этой плазмиды и была получена экспрессия (фиг.6). Этот белок, содержащий человеческий кальцитонин, был выделен из лизата рекомбинантных микроорганизмов путем осаждения. Цистеиновые остатки были затем преобразованы в S-сульфонаты, а лизиновые остатки блокировались при этом за счет реакции с цитраконовым ангидридом. Поскольку человеческий кальцитонин не содержит аргинина, триптическое переваривание рекомбинантного искусственного белка создавало пептид, содержащий последовательность человеческого кальцитонина с удлинением глицином карбоксильного конца. Этот пептид был выделен с помощью обращенно-фазовой хроматографии (фиг.8) или ионообменный хроматографии и его структура установлена с помощью аминоанализа и анализа микропоследовательностей.
Глициновый остаток пептида был удален и предпоследний пролиновый остаток преобразован в пролинамид с помощью альфа-амидирующего фермента согласно изобретению. Примером условий, используемых для полупрепаративного альфа-амидирования этого пептида, является следующий. Лиофилизованный пептид (200-300 нмоль) (субстрат) был растворен в 200 μл 150 мМ буфера Трис-HCl pH 7, содержащего приблизительно 750 мЕ альфа-амидирующего фермента. Фермент был получен либо из ткани МТС крыс или среды культуры линии клеток МТС СА-77 крыс. Фермент был очищен в такой степени, что вся протеолитическая активность была устранена. Это осуществлялось сочетанием ионообменной хроматографии и хроматографии исключения (см. вышеприведенные операции). Чистый гомогенный фермент в этом случае не является обязательным. Аскорбиновая кислота и сульфат меди затем добавлены в эту смесь в достаточном количестве, чтобы конечные концентрации составляли приблизительно 3 мМ и 2 UM соответственно. Полученный раствор был перемешан и инкубирован при 37oC в течение 5-6 ч. Обычный процент преобразования субстрата в продукт составлял 70-90% После удаления S-сульфонатных групп рекомбинант человеческого кальцитонина (rhCT) был очищен с помощью обращенно-фазовой хроматографии (фиг.9). Конечный продукт характеризовался его удержанием при обратно-фазовой хроматографии (фиг.10), картированием трипсинового перевара (фиг.11), анализом аминокислот (см. табл. 1) и его биологической активностью (фиг.12). Во всех примерах рекомбинантный человеческий кальцитонин был неотличим от синтетического человеческого кальцитонина.
Вышеописанное показывает, что препараты согласно настоящему изобретению могут осуществлять амидирование физиологически подходящих субстратов, например, человеческого кальцитонина, полученного с использованием технологии получения рекомбинантной ДНК, т.е. содержащей только L-аминокислоты.
Сравнительные примеры
Сравнение активности заявляемого препарата с ранее известным
Сравнение
испытательных систем для определения удельной активности заявляемых препаратов.
Ранее уже использовалось несколько систем испытания активности. Большинство работ, указанных в известном уровне техники, использовали исследования, основанные на преобразовании Д-Тир-Вал-Гли в Д-Тир-Вал-амид. Это исследование является количественным и использует радиоизотопный меченый атом (125 I-Д-Тир-Вал-Гли), который смешан с избыточным намеченным материалом (Д-Тир-Вал-Гли). Измеренное преобразование меченного атома обеспечивает экстраполяцию относительно намеченного материала, и это в свою очередь позволяет произвести расчет активности.
Хотя это исследование было использовано заявителями, определения активности заявляемых препаратов основаны на прямом измерении преобразования Дансил-Тир-вал-Гли в Дансил-Тир-Вал-амид.
Для того, чтобы обеспечить правомерное сравнение удельной активности препаратов согласно известному уровню техники с теми препаратами, которые заявляются согласно настоящему изобретению, был проведен эксперимент по сравнению систем исследования.
Экспериментальные протоколы сведены следующим образом:
1. Монодансил L-Тир-Вал-Гли
Альфа-амидирующий ферментативный препарат, выделенный из опухолей карциномы щитовидной железы крыс и из среды тканевой культуры, выделенной из СА-77, был
использован в качестве источника фермента в этих экспериментах. Концентрация фермента, использованного во всех экспериментах, оставалась постоянной за исключением тех случаев, где это указано.
Реакционная смесь для преобразования заместителя монодансила содержала
5 μ фермента
5 μл 30 мМ аскорбата
5 μл 20 мМ C и SO4
5 μл 100 μг/мл каталаза из поджелудочной железы быка
5μл, содержащих 2 нмоль субстрата
25 μл 150 мМ TeS pH 7,0.
Образцы были подготовлены в двух экземплярах и инкубированы при 37oC в течение 10, 20 и 30 мин. Ферментативная реакция прерывалась при добавлении 10 μл/ 500 мМ EDTA. Субстрат и продукт были разделены с использованием обратно-фазной хроматографии на оборудовании для жидкостной хроматографии высокого давления фирмы Хюллет Паккард 1090 и количественная оценка осуществлялась с использованием интегратора НР-392. Преобразование монодансил L-Тир-Вал-Гли в альфа-амидированный продукт оказалось линейным во времени.
II.125I-Д-Тир-Вал-Гли
Д-Тир-Вал-Гли и Д-Тир-Вал-NH2 были иодированы с использованием шариков иода фирмы Пирси Кемикал Компани. Меченый субстрат и продукт были использованы для калибровки катионно-обменной колонки с сульфул-пропилом.125I-Д-Тир-Вал-Гли добавляли к 650 М Д-Тир-Вал-Гли и использовали в качестве субстрата. Реакционная смесь для преобразования субстрата монодансила содержала.
5 μл
фермента
5 μл аскорбата (30 мМ)
5 μл 100 мг/мл каталазы
5 μл 20 мл CuSO4
5 μл субстрата, 600 μм конечной
концентрации
25 μл 150 мМ TES pH 7,0
Пробы были инкубированы на 10, 20 и 30 мин при 37oC. Реакция останавливалась путем добавления 500 мМ ЕДТА. Вся проба была
разбавлена 10 мМ натрий фосфатного буфера pH 5,2 и помещена в катионообменную колонку на сульфил пропиле. Субстрат не присоединялся к колонке, амидирование меченого субстрата в продукт имело линейный
характер во времени.
III. D- Тир-Вал-Гли
Условия реакции, использованные для амидирования Д-Тир-Вал-Гли, идентичны тем, что описаны для дансилового и иодинированного
субстратов. Концентрация субстратов в реакционной смеси была 650 μМ. Разделение субстрата и продукта достигалось градиентной элюцией при применении жидкостной хроматографии с обращенной фазой,
использующей систему НП-1090. Вытекающая из колонки жидкость контролировалась на длине волны 280 нм. Уровень чувствительности для этого использования значительно ниже, чем чувствительность для
исследования дансиловых или иодированных субстратов. Для того, чтобы соответствовать этому более низкому уровню чувствительности, реакция альфа-амидирования осуществлялась в течение более длительных
отрезков времени и/или с увеличенным количеством альфа-амидирующего фермента.
Анализ данных фракционного преобразования125I-Д-Тир-Вал-Гли в125I-Д-Вал-амид и Дансил-Тир-Вал-Гли в Дансил-Тир-Вал-амид показывает, что в каждый момент времени иодированного субстрата было преобразовано приблизительно в 1,55 раза больше, чем дансилового субстрата. Таким образом, при сравнении уже известной системы исследования с системой исследования, используемой заявителем, коэффициент преобразования приблизительно 1,5 должен быть использован для активности, определяемой по способу с дансиловым сусбтратом (заявителей).
Кроме того, более строгий кинетический анализ был использован для сравнения исследования с дансил-Тир-Вал-Гли (заявителей) с исследованием Д-Тир-Вал-Гли (ранее известный), приведенным в табл. 1.
Как можно видеть, при сравнении максимальной скорости -Vмакс пмоль продукта (мин/μл) для двух субстратов, Д-Тир-Вал-Гли (ранее известный) дает приблизительно 1,48 активности дансил-Тир-Вал-Гли (заявителя). Это подтверждает и согласуется с указанными выше сведениями.
Сравнение активности
Эйперт и др. (PNAS) показывает на странице 5147, фиг.4, Vмакс 39 пмоль/мкг/ч. Это эквивалентно удельной активности 0,65 мЕ/мг белка в минуту. Это наивысшая активность, о которой
сообщалось ранее. При делении этой величины на вышеуказанный коэффициент преобразования, равный 1,5, получим удельную активность 0,4 мЕ/мг белка в минуту. Эта величина может непосредственно
сравниваться с вышеуказанной удельной активностью, достигаемой заявителями. Заявители, как указано ранее, достигли активностей по меньшей мере 25 мЕ/мг белка и более 1500 мЕ/мг белка. Заявители, таким
образом, достигли активности в 60-3750 раз более высокой, чем активность, о которой сообщал Эйперт (PNAS).
Пример 3
Очистка и исследование альфа-амидирующих ферментов,
полученных из опухоли МТС крыс.
Замороженную ткань опухоли МТС крыс распыляли до мелких частиц и гомогенизировали в водном буфере, используя гомогенизатор Политрон. После низкоскоростного центрифугирования супернатант (насадочная жидкость) был слит и осадок был повторно экстрагирован с помощью свежего буфера. Этот второй гомогенат вновь был подвергнут низкоскоростному центрифугированию, и новый супернатант был соединен с первым. Два собранных и соединенных супернатанта были затем осветлены с помощью высокоскоростного центрифугирования, и полученный при этом супернатант был использован в качестве исходного материала для очистки фермента.
Фракционирование аммоний сульфатом было выполнено для этого полученного при высокоскоростном центрифугировании супернатанта. Было установлено, что большая часть активности фермента оседала в 26-40% сульфата аммония, и осадок был очищен дополнительно, как описано ниже.
Хроматография исключения была проведена на колонке Сефакрил S-300. В приведенном здесь примере 1 весь фермент, элюированный с этой колонки, находился в единственном пике активности. В этом примере длина колонки была увеличена, а расход колонки уменьшен. При этих новых условиях элюирования основной пик активности наблюдался, как и в примере 1, и за ним следовал небольшой "хвостовой пик", который мог бы соответствовать ферменту с более низким молекулярным весом. Не ясно в этом вопросе, существует ли фермент с низким молекулярным весом in vivo или образуется за счет частичного протеолитического переваривания во время экстрагирования и очистки.
Основной пик активности из колонки S-300 был получен с помощью хроматографии на колонке Моно-Q при pH 6,0. В этом примере была использована колонка большого размера, чем в примере 1 (Моно Q3 HR 10/10) и элюирование осуществлялось с использованием менее крутого линейного солевого градиента. В результате этих изменений четыре пика альфа-амидирующей активности фермента было обнаружено на этом этапе при элюировании: при NaCl 160, 200, 220 и 240 мМ (Пики I, II, III, и IV соответственно, фиг.13). Это показывает, что существует множество форм фермента и что эти формы обладают определенной неоднородностью. Кроме того, анализ с помощью полиакриламидного геля белков в пиках активности фермента показывают, что пики II, III и IV включают в себя альфа-амидирующий фермент с кажущимся молекулярным весом одного порядка (т. е. 73000-75000 Дальтон), тогда как пик I имел фермент отличного, возможно меньшего молекулярного веса. Активность пика III была очищена до гомогенности следующим образом.
Фермент пика III был собран и подвергнут хроматографической обработке на колонне Моно Q HR 10/10 при pH 8,0 (фиг.14). То, что фермент был элюирован с этой колонки в виде единственного пика при 250 мМ NaCl, а также анализ в геле показали, что фермент очищен до однородного состояния (фиг.15a, полоска 6). Следующие экспериментальные исследования были проведены на очищенном ферменте пика III.
1. Молекулярный вес фермента пика III был определен с помощью анализа на 7% полиакриламидном геле и составлял около 75000 Дальтон (фиг.15b).
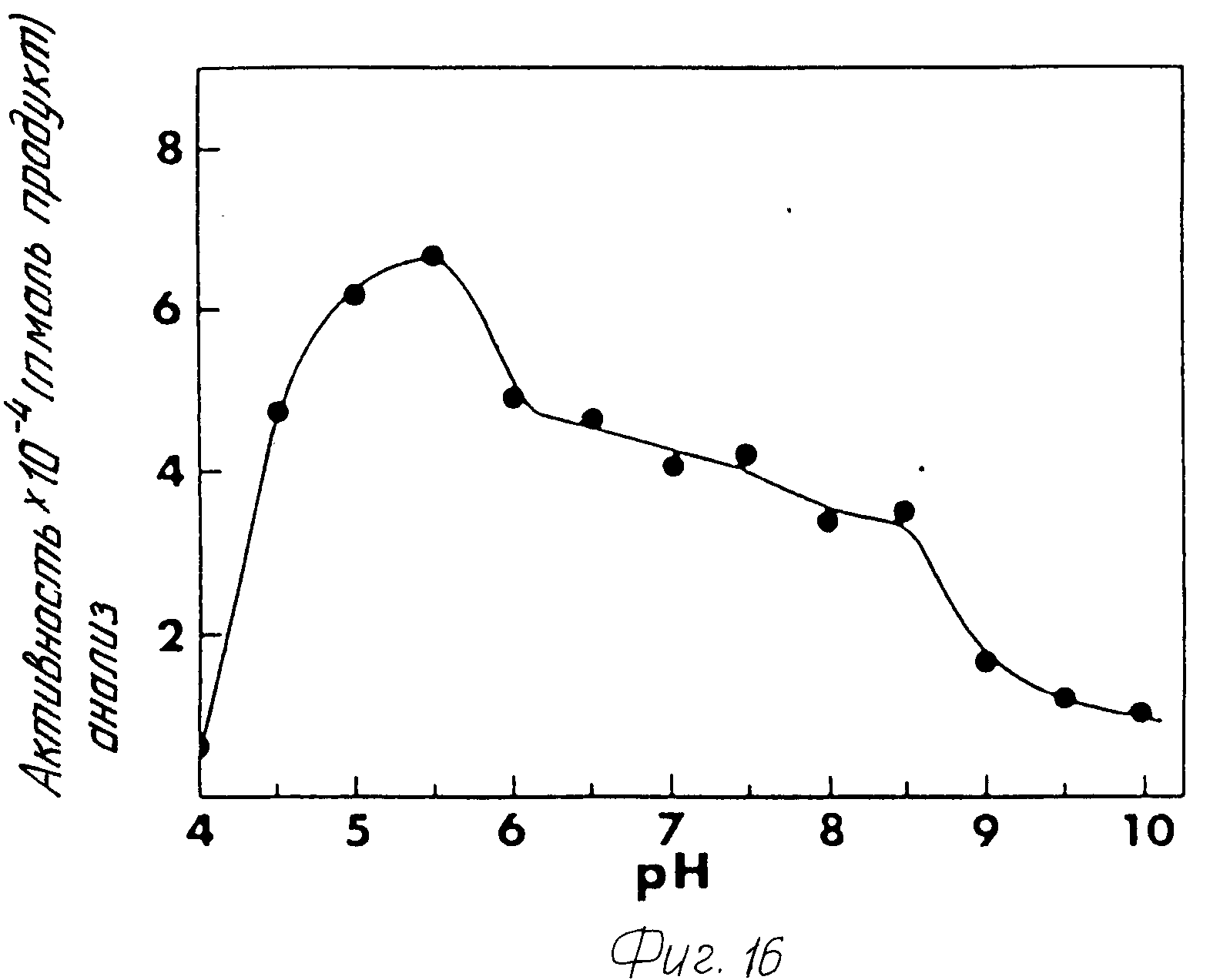
2. Оптимальная pH для активности фермента составила pH 5,0-5,5 при использовании N дансил Тир-Вал-Гли в качестве субстрата (фиг.16). Однако, благодаря повышенной устойчивости фермента при нейтральном pH, может оказаться успешным осуществление реакции амидирования при таком pH.
3. Определено, что количество меди, необходимой в качестве кофактора для активности фермента, было обратно пропорционально чистоте фермента. Очищенный до однородного состояния фермент требовал 0,1 μM или менее Cu++ для максимальной активности, тогда как исходные препараты фермента требовали 2 мМ Cu++.
4. Изоэлектрическая точка (pH) фермента составляла 4,8.
5. Удельная активность гомогенного (однородного) фермента пика III составляла 2,100 мЕ/мг белка.
Фермент пика II был также очищен до однородного состояния. Однако, для этого фермента оказалось, что хроматография Моно Q при pH 8,0 была недостаточной для получения однородного препарата. Поэтому сбор фермента пика II из колонки Моно Q pH 6,0 (фиг.13) был диализирован против 1 M Трис pH 7,0 и загружен на колонку с фенил-сефарозой, уравновешенной таким же буфером. Большая часть загрязняющих белковых проб извлекалась в потоке через колонку, и амидирующий фермент, элюируемый на последней стадии, был по существу чистым. Дополнительная очистка собранного фермента из колонки фенил-сефарозой на колонке Моно QHR 10/10 pH 8,0 привела к получению гомогенного препарата фермента пика II, элюирующегося с колонки при 220 мМ NaCl и выше.
Исследования фермента пика II показало, что
1. Молекулярный вес фермента пика II при электрофорезе с использованием
7% полиакриламидного геля составлял около 73000-75000 Дальтон. Таким образом, ферменты пика II и III были неотличимы по сравнению их молекулярных масс.
2. Оптимальное значение pH для активности фермента пика III составляло 5,0-5,5. И вновь, эта характеристика фермента пика II является такой же, что и для фермента пика III.
3. Изоэлектрическая точка (p1) пика II была приблизительно 5,8.
Пример 4
Очистка и исследование альфа-амидирующего фермента, полученного из среды тканевой культуры клеток CA-77
Клеточные линии CA-77
карциномы щитовидной железы крыс были выращены в виде минослоя при наличии 8% CO2. Культура выдерживалась в определенной среде, состоящей из среды Игла в модификации Дульбекко:F 10(1:1), 3,
7 г/л NaHCO3, 5 μг/мл трансферрина, 10 μг/мл инсулина, 30 нМ селена, 4 μг/мл# пентамицин сульфата. Культуры, выращенные таким образом, могли храниться неограниченное
время, если среда заменялась каждые 48 ч. Для увеличения прироста массы клеток они были дополнительно культивированы и выращивались в среде, содержащей сыворотку крови (5% лошадиной и 2,5% телячьей),
в течение трех дней. Клетки были затем промыты дважды физиологическим раствором с фосфатным буфером.
Среда культуры ткани была собрана с соблюдением асептики по 48-часовой программе и хранилась при -20oC до очистки. Среда культуры ткани (по программе 6 л) была разбавлена 2 л деионизированной воды (3:1) и подана при расходе 50 мл/мин на анионообменную систему (Сипо N 250), которая была предварительно уравновешена 1,0 л 20 мМ бис Трис HCl pH 6,0 при температуре 4oC. Альфа-амидирующий фермент (альфа-АЕ) был элюирован ступенчато с помощью 50 мМ Трис HCl pH 7,0, содержащего 500 мМ NaCl при расходе приблизительно 50 мл/мин. Фракции, обладающие активностью альфа-АЕ (удельная активность 10-15 мЕ/мг) из двух полученных с использованием анионообменных процессов препаратов были собраны и концентрированы 4-5-кратно при пониженном давлении с использованием ротора предварительной обработки Савант RH-100.
Этот материал вводился непосредственно в колонку 5х 50 см, содержащую Сефакрил 300 SF (Фармация). Подвижная фаза представляет собой 100 мМ Трис: HCl pH 7,0 с расходом 1,0 мл/мин. Вся гель-фильтрационная хроматография была осуществлена при 4oC (фиг.XVII).
Препарат амидирующего фермента на этой стадии очистки свободен от неспецифической протеолитической активности и имеет активность по меньшей мере 50 мЕ/мг белка. Препарат амидирующего фермента, полученный на этом этапе, был использован успешно для амидирования рекомбинантного удлиненного глицином человеческого кальцитонина и фактора высвобождения гормона роста. Начиная с плотности клеток 1-1,5•106 клеток/кл (из T-образного флакона) мы постепенно получили выход 200-350 мЕ активности амидирующего фермента/литр среды, проходящей эти два этапа очистки. Фермент устойчив и пригоден для использования в растворе или после иммобилизации его на твердой подложке.
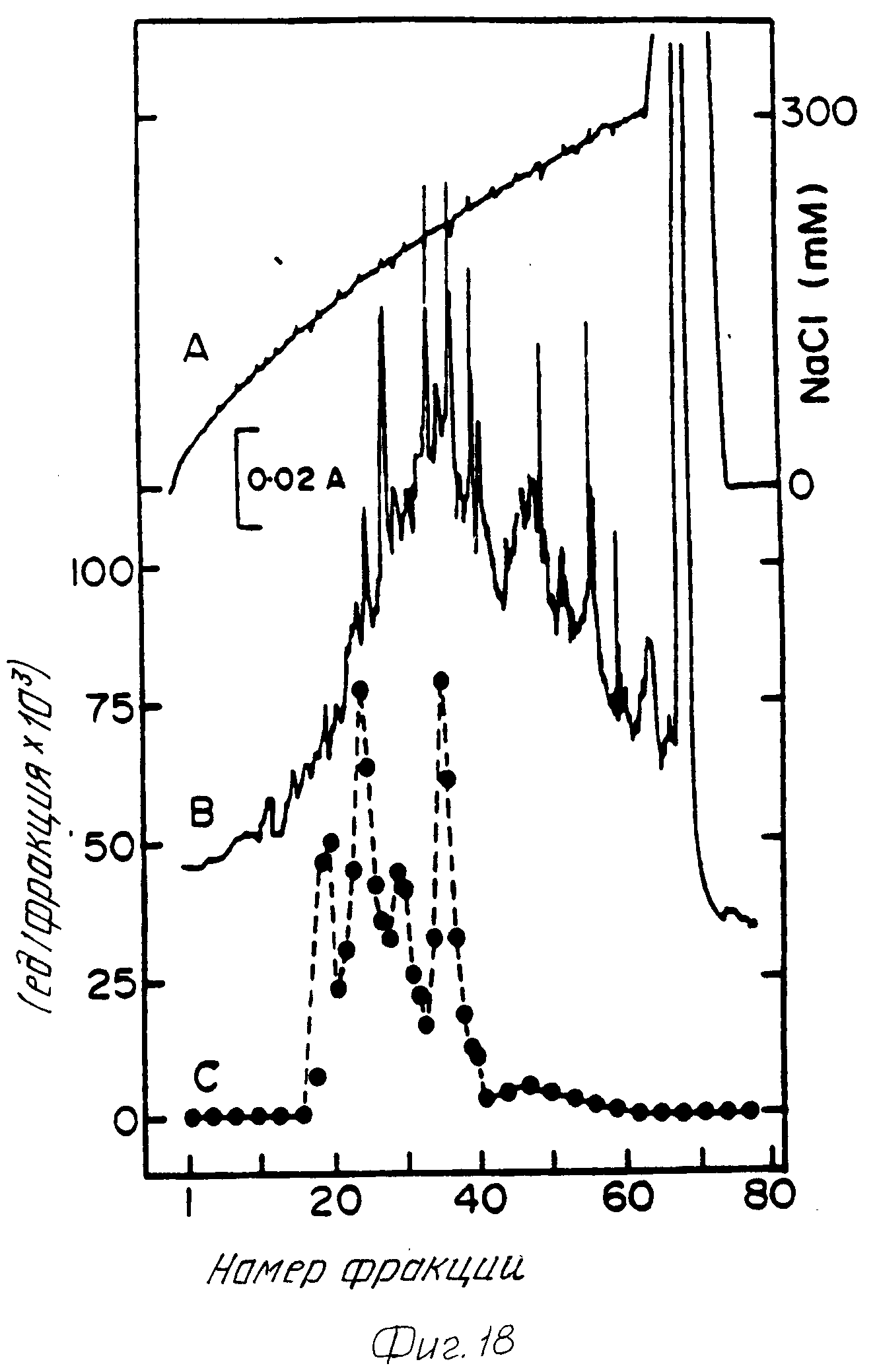
Фракции колонки, содержащей активный альфа-АЕ, были собраны и затем диализированы на 6 л 20 мМ бис Трис HCl pH 6,0. Фермент был введен в колонку с сильным анионообменником pH 6,0. Фермент был элюирован из колонки Моно Q HR 10/10 сильной анионообменной хроматографии с предварительным уравновешиванием 20 мМ бис Трис HCl pH 6,0. Фермент был элюирован с колонки с использованием линейного градиента C-300 мМ NaCl за 3 ч при расходе 2,5 мл/мин. Четыре хроматографически разделяемые пика альфа-амидирующего фермента были получены на этих трех этапах. Пики были пронумерованы в порядке элюирования из колонки (фиг.18). Пики III и IV отображают формы с более высокими молекулярными весами ферментов и соответствуют пикам II и III, полученным из опухолей МТС. Пики I и III отображают формы с более низкими молекулярными весами ферментов, что может отображать протеолитическую активность пиков III и IV.
Четыре формы альфа-амидирующего фермента, идентифицированных в лаборатории, отличаются друг от друга их средним поверхностным зарядом, что доказывается их различными временами удержания в процессе сильной анионообменной хроматографии (фиг. 18). Оптимальное значение pH для этих четырех хроматографически различных форм фермента также различны. Результаты, приведенные на фиг. 19, показывают, что пики III и IV имеют идентичное значение оптимума pH между 5,0 и 5,5. Эти результаты согласуются с pH, определенным для пиков II и III, полученных выделением с очисткой из опухоли МТС. Пики I и II имеют более широкие границы диапазона активности для оптимума pH от 5 до 8,5 (фиг. 19). Эти результаты тесно согласуются с оптимумом pH, о котором сообщали Эйпарт и др. Peptides, т. 4, с. 921-28 (1983) и Марти и др. Biochemistry, т. 261, с. 1815-22 (1986).
Мечение фермента из пиков II и IV изотопами Na125J, последующий SDS-PAGE подтвердили, что активность фермента пика IV характеризуется приблизительной молекулярной массой 73-75 кДа, тогда как активность фермента пика II располагалась ниже 55 кДа. Точный молекулярный вес для фермента пика II не известен, поскольку он не был очищен до однородного состояния (несколько белковых линий проявляются в диапазоне 45-55 кДа).
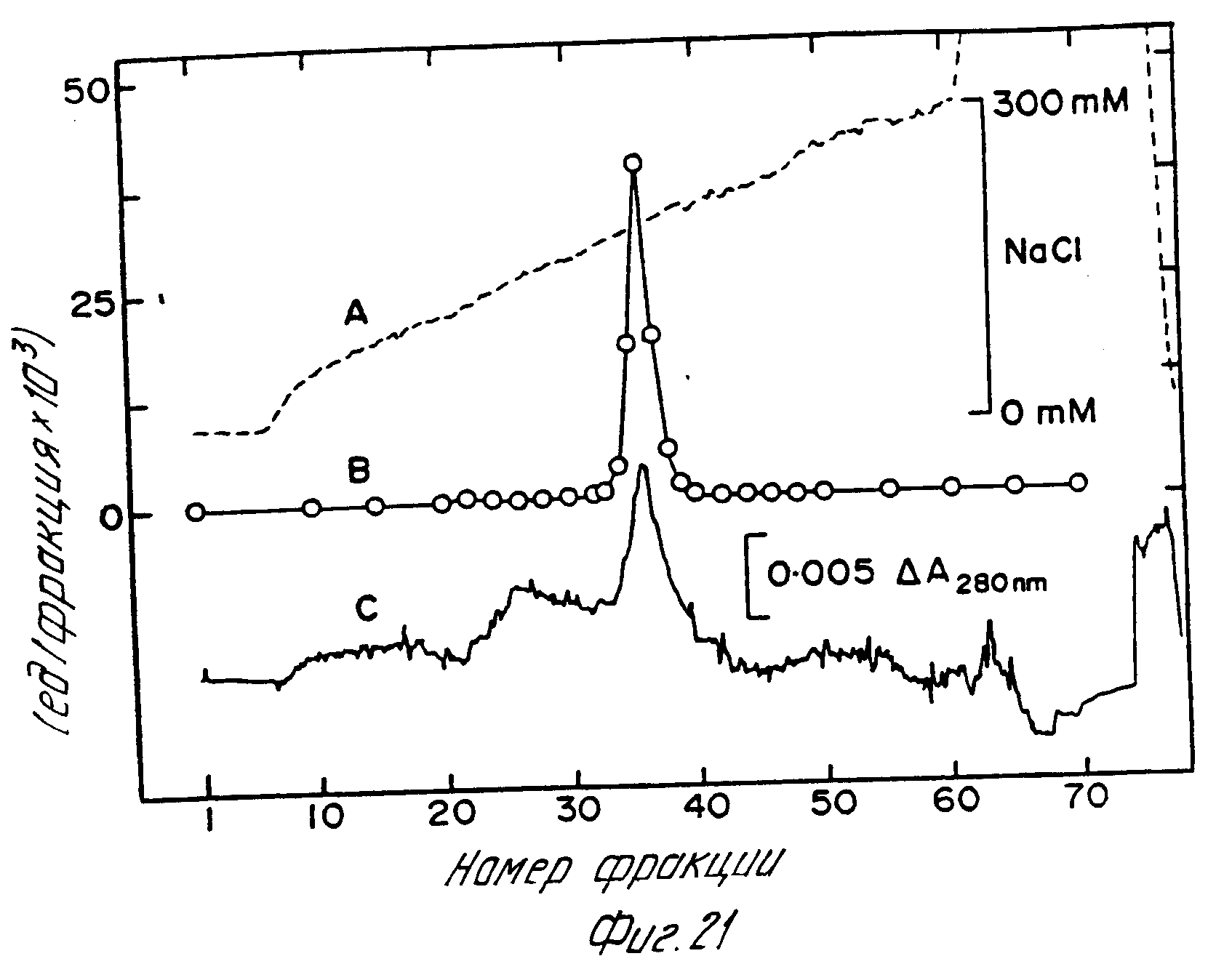
Ферменты пика III и IV могут быть очищены до однородного состояния с использованием сочетания гидрофобной хроматографии и сильной анионообменной хроматографии при pH 8,0. Фермент пика IV (фиг.18) был собран, сконцентрирован до приблизительно 2 мл in vacuo и непосредственно введен в колонку 1,3•8 см с фенилсефарозой (Фармация), уравновешенную 500 мМ Трис HCl pH 7,0. Фракции, обладающие активностью альфа-АЕ, были элюированы уравновешивающим буфером при расходе 0,5 мл/мин (фиг.20). Пиковые фракции, обладающие альфа-амидирующей активностью, были собраны, диализованы на 50 мМ Трис HCl pH 8,0, затем введены в колонку Моно QHR 10/10, уравновешенную 50 мМ Трис HCl pH 8,0. Фермент был элюирован с колонки с использованием линейного градиента 0-300 мМ NaCl в течение 3 ч при расходе 2,0 мл/мин (фиг.21). Фракции, обладающие альфа-АЕ активностью, элюированные при 240 мМ или выше, были собраны, доведены до содержания 0,001% (в объемном отношении) Тритон X-100 и хранились при 4oC. Удельная активность очищенного фермента составляла по результатам измерений приблизительно 1500 мЕ/мг белка при pH 7,0. a-амидирующая активность пика III была получена в гомогенном состоянии с использованием таких же операций, что описаны для пика IV.
Физико-химические исследования "опухолевого" пика (из примера 3) и пика IV тканевой культуры (из примера 4), имеющие молекулярную массу (73000-75000 Дальтон), оптимум pH 5,0-5,5, последовательность аминоконцов и положений элюирования (более чем около 240 натрий хлорида при сильной анионообменной хроматографии, проводимой при pH 8,0) показали, что эти два пика могут отображать один и тот же фермент.
Пример 5
Альфа-амидирование биологически релевантных пептидных гормонов с
использованием альфа-амидирующего фермента
Несколько субстратов рекомбинантных пептидов гормонов, включая те, что предназначены для лососевого и человеческого кальцитонина, фактора
высвобождения гормона роста человека, и пептид, связанный с геном кальцитонина человека, получены и успешно альфа-амидированы препаратом альфа-амидирующего фермента согласно настоящему изобретению.
Для целей иллюстрации процедуры, использованные для производства рекомбинантного лососевого кальцитонина, человеческого пептида, связанного с геном кальцитонина и фактора высвобождения гормона роста
человека приведены ниже. Одинаковые типы подхода к решению задач могут быть использованы для других рекомбинантных пептидов.
Кальцитонин лососевых представляет собой 32-членный пептидный гормон, имеющий альфа-амидированный пролиновый остаток у его карбоксильного конца. Микроорганизмы были созданы с помощью генной инженерии, чтобы получить рекомбинантный слитый белок с аминокислотной последовательностью, соответствующей кальцитонину лосося. Ген искусственного белка был сконструирован так, чтобы последовательность лососевого кальцитонина была заключена между метиониновым остатком на аминоконце и глициновым остатком на карбоксильном конце, который завершал рекомбинантный слитый белок.
После связывания гена кальцитонина лосося в плазмиду микроорганизм был трансформирован с помощью этой плазмиды и была достигнута экспрессия искусственного белка. Этот кальцитонин, содержащий белок, был выведен из лизата рекомбинантного организма путем осаждения, и его цистиновый остаток преобразован в S-сульфонаты. Поскольку лососевый кальцитонин не содержит метионина, цианоген бромидное расщепление рекомбинантного искусственного белка приводило к созданию пептида, содержащего последовательность лососевого кальцитонина с глициновым продолжением карбоксильного конца. Этот пептид был выделен или с помощью жидкостной хроматографии высокого давления с обращенной фазой или ионообменной хроматографии, и его структура была установлена с помощью аминокислотного состава и анализа микропоследовательностей.
Предпоследний пролиновый остаток был преобразован в пролинамид под действием альфа-амидирующего фермента. Пример условий, использованных для альфа-амидирования этого пептида, приведен ниже. Лиофилизированный пептидный субстрат (предшественник кальцитонина лосося, удлиненного глицином, 200-300 наномоль) был разведен в 200 мл буфера 150 мМ Трис HCl pH 7,0, содержащего приблизительно 750 UE альфа-амидирующего фермента. Фермент мог быть извлечен или из опухоли МТС, или из среды культуры ткани клеточной линии крыс МТС СА-77. Фермент должен быть очищен в такой степени, чтобы вся активность внешнего протеолитического фермента была снята. Это обычно осуществляется с помощью сочетания ионообменной и вытеснительной хроматографии (см. пример 4). Аскорбиновая кислота и сульфат меди были затем добавлены в этой смеси в достаточных количествах, чтобы обеспечить конечные концентрации приблизительно 3μM и 2μM соответственно. Каталаза (7,5 мг/мл), этанол (1% по объему) и иодид калия могут быть введены в реакционную смесь, чтобы повысить выход кальцитонина. Полученный раствор был смешан и инкубирован при 37oC в течение 5-6 ч.
После удаления S-сульфонатных групп с помощью бетамеркаптоэтаноловой обработки рекомбинантный лососевый кальцитонин был очищен с помощью жидкостной хроматографии высокого давления с обращенной фазой. Конечный продукт был охарактеризован с помощью показателя удержания в процессе жидкостной хроматографии высокого давления, количественного триптического картирования и аминокислотного анализа. Во всех примерах рекомбинантный кальцитонин лосося не отличается от синтетического кальцитонина лосося.
Человеческий пептид, связанный с геном кальцитонина, представляет собой 37-членный гормон, имеющий альфа-амидированный фенилаланиновый остаток у карбоксильного конца. Ген искусственного белка был сконструирован аналогично гену кальцитонина лосося (см. выше) в том отношении, что последовательность пептида была заключена между метиониновым остатком и глициновым остатком, который оканчивал весь рекомбинантный белок. Освобождение, очистка, альфа-амидирование и исследование рекомбинантного сродственного с геном пептида предшественника было осуществлено аналогично тому, как было это сделано в связи с рекомбинантным кальцитонином лосося.
Фактор высвобождения гормона роста человека (hGHRF) это 44-членный пептидный гормон, имеющий альфа-амидированный лейциновый остаток у его карбоксильного конца. Ген искусственного белка для hGHRF был сконструирован так, что аминокислотная последовательность для пептидного гормона была заключена между триптофановым остатком на его аминоконце и глициновым остатком у его карбоксильного конца, который оканчивал и весь рекомбинантный слитый белок. Содержащий hGHRF слитый белок был выделен из лизата рекомбинантного микроорганизма путем осаждения. Не имеющая hGHRF часть слитого белка была денатурирована путем преобразования цистеиновых остатков в S-сульфонат производные. Поскольку hGHRF не содержит триптофан, химическое переваривание рекомбинантного искусственного белка реактивным BN PS-скатолом вызывало окислительное расщепление искусственного белка, в результате чего создавался неамидированный hGHRF с глициновым удлинением карбоксильного окончания. Одновременно метиониновый остаток в позиции 27 в молекуле hGHRF был окислен до метионин-сульфоксида. Этот удлиненный глицином пептид был выделен с использованием гель-фильтрации и жидкостной хроматографией с обращенной фазой. Его структура была установлена с помощью аминокислотного анализа пептидных фрагментов, полученных посредством трипсинового переваривания.
Предпоследний лейциновый остаток был преобразован в лейцинамид под действием альфа-амидирующего фермента согласно настоящему изобретению. Пример условий, использованных для получения альфа-амидированного hGHRF, приводится ниже. Субстрат лиофилизированного пептида (20-41 наномоль) был разбавлен в 150 μл деионизированной воды и смешан с 90 μл (500 μE) (pH 7,0) препарата альфа-амидирующего фермента из опухоли МТС или среды культивирования культуры ткани клеточной линии Ca-77 крысы. Фермент был очищен для удаления всех видов внешней активности протеолитического фермента путем использования хроматографии исключения и ионообменной хроматографии. Аскорбиновая кислота и сульфат меди были добавлены в эту смесь фермента и субстрата в достаточных количествах, чтобы конечная концентрация составляла 3 μ M и 2 μM, соответственно. Полученный раствор был перемешан и инкубирован при 37oC в течение 4-6 ч. Обычный процент преобразования субстрата в продукт составляет 95% в пересчете на аминокислотный анализ фрагментов, выделенных при переваривании трипсином. Наконец, метионилсульфоксидный остаток был восстановлен до метионина под действием 4 М бетамеркаптоэтанола, забуференного 10 мМ натрий ацетатом до pH 4 при 80oC в течение 1 ч. Конечный продукт был очищен с помощью жидкостной хроматографии высокого давления с обращенной фазой и исследован по времени задержки, анализу переваривания трипсином и аминокислотному анализу. Рекомбинантный альфа-амидированный продукт был также испытан в отношении биологической активности. Во всех примерах рекомбинантный hGHRF был неотличим от синтетического hGHRF.
Помимо вышеприведенных исследований два промышленно важных пептидных гормона, удлиненных глицином, были оценены в отношении их способности действовать в качестве субстрата для α-амидирующей монооксигеназы. Это были предшественники альфа-меланоцитстимулирующего гормона и вещества Р. В обоих случаях результаты показывают, что оба пептида являются пригодными субстратами в отношении альфа-амидирующего фермента.
Пример 6
Анализ последовательностей очищенного альфа-амидирующего фермента
Фракции,
содержащие очищенный альфа-амидирующий фермент, были собраны или из опухолевой ткани МТС крыс или из супернатантов клеточной культуры СА-77, и сульфгидрильные группы фермента были подвернуты
восстановлению, за которым последовало карбоксиметилирование. Полученная реакционная смесь была затем введена в колонку жидкостной хроматографии высокого давления с обращенной фазой (размер частиц
5μ, размер пор 33 нм), которая была уравновешена 0,1%-ным водным раствором трифторуксусной кислоты. Колонка была промыта этим раствором для удаления избыточных буферных солей. Обессоленный
фермент был удален из колонки жидкостной хроматографией высокого давления путем элюирования с помощью 80% ацетонитрила, содержащего 0,08% трифторуксусной кислоты. Вытекающий поток (эффлюент) колонки
контролировался по УФ-излучению на длине волны 220 нм. Полученные белковые фракции были собраны, объединены и лиофилизованы. Этот материал был затем повторно разбавлен в 100 мл 0,1 SDS и затем введен
в устройство исследования последовательностей белка. Апплайд Биосистемс модель 470А. Операции, используемые для анализа микропоследователностей, были такими, как указаны для Апплайд Биосистемс.
Полученные фенилтиогидрантоиновые аминокислоты анализировались с помощью жидкостной хроматографии высокого давления на колонке Гиперсил C18 (размер частиц 5мм, размер пор 10 нм) с контролем поглощения
на длинах волн 269 и 313 нм на системе жидкостной хроматографии Хьюллет Паккард 1090. Амино-концевая последовательность для основных составляющих пика фермента (пик III) из опухолевой ткани была
следующей

Аминоконцевая последовательность для основных компонентов фермента (пик IV) из супернатанта культуры тканей клеток СА-77 показывает, что она идентична той, что имеет место для фермента опухоли (пик III). Однако небольшая компонента была также обнаружена во время анализа микропоследовательностей этого фермента, которая, кажется, содержит продолжение аминоконца при сравнении с основной формой альфа-амидирующего фермента. Наличие этой компоненты, вероятно, обусловлено различием посттрансляционной обработки фермента. Последовательность аминоконца этой компоненты была следующей

Дополнительные данные об аминокислотной последовательности были получены для альфа-амидирующего фермента из опухолевой ткани МТС крыс с помощью следующих операций. Приблизительно 400 г очищенного фермента было подвергнуто восстановлению и карбоксиметилированию. После этой процедуры раствор фермента был перенесен для экструзионного диализа и диализ провели в течение 18 ч против 25 мМ Трис-HCl pH 8,0/0,5 M мочевины. Осадок затем был перенесен в 1,5 мл центрифужную пробирку и концентрирован до объема 600 μл при пониженном давлении. К раствору фермента добавили 2 μл(2 μг.) трипсина и смесь инкубировали один час при температуре 37oC. После этого добавили вторую дозу трипсина 2μг. и продолжали инкубацию еще два часа при 37oC. Переваривание оканчивали путем добавления 200 μл 4M мочевины на 10% уксусной кислоте. Продукт переваривания был затем введен в хроматографическую колонку жидкостной хроматографии высокого давления обращенной фазы (размер частиц 5 μm размер пор 33 нм), которая была уравновешена 0,1% водным раствором трифторуксусной кислоты. Колонка была затем элюирована линейным градиентом ацетонитрилом до концентрации 50% в течение 4 ч, и фракции собирались с двухминутным интервалом. Полученные пики элюента жидкостной хроматографии высокого давления с обращенной фазой для продукта переваривания трипсином графически изображены на фиг.22. Три полученных с использованием триптического расщепления пептида прошли автоматизированный анализ последовательностей, описанный выше, и полученные результаты приводятся ниже.
(Пептиды обозначены их номером фракции).
Триптический пептид N 65.

триптический пептид N 58
-Asn-Gly-Gln-Trp-Thr-Leu-Ile-Gly-Arg-/
триптический пептид N 86
Phe-Val-Thr-Gln-Trp-Gly-Glu
Пример 7
А. Выработка у мышей многоклониальных антител, специфических для альфа-амидирующего фермента
Двенадцать мышей Bal b/сy были иммунизированы и реиммунизированы с очищенным приготовлением амидирующего фермента. У этих мышей была взята кровь и сыворотка была обработана и титрована на очищенный фермент с помощью пробы ELISA. Проба (используемый прием хорошо известен) была осуществлена посредством абсорбирования очищенного фермента на полистироловую пластину, которая затем была промыта и блокирована (для предотвращения постороннего прилипания антител к пластине) бычьим сывороточным альбумином (BSA). Разбавленные мышиные сыворотки (содержащие мнимые антитела против амидирующего фермента) были затем инкубированы на покрытой амидирующим ферментом пластине и промыты. Вторичное антитело (отмеченное маркером, щелочной фосфатазой, которая облегчает закрепление связывания первого антитела к нанесенному на пластину ферменту) было затем инкубировано в ячейках. После промывания и добавления раствора субстрата сигнал был калориметрически измерен с помощью спектрофотометрического записывающего устройства. "Положительные" сыворотки показали отношение сигнала к шуму по меньшей мере 2:1.
Мышь N 7, которая показала более высокий титр в тесте ELISA, была принесена в жертву через четыре дня после окончательной реиммунизации. Селезенка была асептически удалена и измельчена (механически разделена) для получения клеток селезенки в количестве 132,6•106, которые слиты с клетками миеломы NS-I 122,8 (106 mls PEG) полиэтиленгликоль) 4000. Клетки были разделены на пять 24-ячеистых пластин, которые заранее были покрыты тимоцитами и спленоцитами Bal b/cj, которые служили в качестве питательных клеток. Клетки содержались в селективной среде НАТ, которая допускает выживание только гибридных клеток.
Супернатанты из 116 ячеек, которые показали рост клонов, были скринированы с помощью радиоиммунной пробы и метода ELISA для получения антитела. Процедура радиоиммунной пробы была аналогична пробе ELISA, описанной ранее, за исключением того, что второе антитело было помечено с помощью I125, и радиоактивный счет был измерен с помощью гамма-счетчика.
56 из 116 ячеек давали положительный результат на получение антитела и проанализированы на реактивность к альфа-амидирующим ферментам. 25 клонов, которые оказались положительными по отношению к альфа-амидирующему ферменту, были клонированы при помощи процессора последовательного разбавления. Первичные клоны были скринированы против альфа-амидирующих ферментов и BSA (поскольку полистироловые пластины были блокированы с помощью BSA, антитела, которые были связаны с пластиной, могли просто приклеиваться или быть абсорбированы BSA неспецифичным образом) для определения, являлись ли они действительно специфичными для амидирующего фермента. Клоны, которые показали сигнал на альфа-амидирующий фермент и который был по меньшей мере удвоен по сравнению с демонстрируемым для BSA, были клонированы с соотношением распределения I клетка/2 ячейки.
Двадцать одна положительная гибридома (см. табл. III) была перенесена на стадию третичного клонирования, и двадцать из них были охарактеризованы в отношении класса антител при помощи приемов Ouchterlony и ELISA. Семнадцать клонов имело тяжелые цепи IgG2a и три имели IgG1 тяжелые цепи. Все двадцать клонов дали антитела с каппа легкими цепями.
Каждая линия была индивидуально выращена в массовой культуре и аликвоты клеток были заморожены в жидком азоте. Мышам Bal b/cJ была сделана инъекция интраперитониально 5•106 клеток гибридомы. Неделей позднее мышам, у которых не появилась асцитическая припухлость, была сделана усиленная инъекция клеток. Асцитическая жидкость и кровь были удалены 1-2 неделями позднее. После обработки асциты и сыворотки были исследованы и оттитрованы против альфа-амидирующего фермента так же, как и против отрицательных антигенов (например, BSA, углеродной ангидразы и фактора высвобождения гормона роста) для обеспечения специфичности антител.
Результаты в табл. 2.
В. Процедура очистки моноклональных антител, продуцированных мышами
Моноклональные антитела, специфичные для альфа-амидирующего фермента, приготовленные описанными выше методами, очищались следующим образом. Асцитная жидкость, собранная от нескольких мышей, привитых
одним и тем же клоном, была использована в качестве источника антител. Асцитная жидкость была разбавлена (5-кратно) 10 мМ MES с pH 5,6. Разбавленная асцитная жидкость была помещена в 1,5х20 см колонку,
содержащую 40 μm смешанной с АВХ кремниевой смолы (Дж. Т. Бэйкер), предварительно уравновешенной с помощью 10 мМ MES буфером с pH 5,6. Моноклональные антитела были элюированы с колонки с
использованием 0-500 мМ градиента ацетата натрия при pH 7,0. Фракции, содержащие очищенные антитела, были объединены, тестированы на удельную активность и помещены на хранение при 4oC до
последующего использования.
С. Генерация у цыплят поликлональных антител, специфичных для альфа-амидирующего фермента
Двум цыплятам были сделаны внутривенные, внутримышечные
и подкожные инъекции, суммарно содержавшие приблизительно 50 μг очищенного альфа-амидирующего фермента в Ribi-адъюванте на каждого цыпленка. Ribi-адъювант представляет собой полностью
метаболизированную жировую эмульсионную систему, которая состоит из митогена для лимфоцитов цыпленка и адъюванта для усиления реакции антитела на антигены в домашней птице (Ribi Immunochem. Research,
Montana). После начальной иммунизации были сделаны две усиленные инъекции с двухнедельным интервалом приблизительно 50 μг фермента на цыпленка. У животных была взята кровь приблизительно на 21
и 35 день, и сыворотки обработаны и протестированы на присутствие специфических антител с помощью следующей процедуры: сыворотки от обоих цыплят, день 0 (преиммунный/, день 21 и день 35 были
исследованы твердофазным ELISA методом против 100 ng очищенного альфа-амидирующего фермента. Бычий сывороточный альбумин был использован в качестве отрицательного контроля для неспецифического
слипания антитела. Фермент специфичное антитело было детектировано кроличьей сывороткой против 1 gG цыплят, маркированной щелочной фосфатазой.
Результаты описанных выше процедур продемонстрировали, что специфичные антитела могут быть обнаружены в сыворотках цыпленка 257 на 35 день. Разбавленная 1-10000 сыворотка дала отношение сигнала к шуму приблизительно 4:1. Цыпленок 258 показал ферментспецифичные антитела на 21 и 35 день. Для обоих проб крови разбавленная 1:10000 сыворотка дала отношение сигнала к шуму приблизительно 4:1. Сбор яиц от обоих куриц начался на 56 день. Изолированный с помощью осаждения полиэтиленгликолем (PEG) IgY преиммунизированных яиц и постиммунизированных яиц был проанализирован методом Ouchferlony, а ферментспецифичные антитела были скринированы с помощью пробы ELISA.
Д. Очистка IgY- антител цыплят
Поликлональные птичьи антитела, специфичные для альфа-амидирующего фермента, были выработаны в цыплятах,
как описано выше. Яйца от иммунизированных цыплят были собраны и либо погружены в парафин, либо заморожены до использования с целью очистки IgY. Белки яиц были отделены от желтков, которые содержат
специфические антитела альфа-амидирующего фермента. Яичные желтки были разбавлены трехкратно с использованием 10 мМ фосфата натрия pH 7,5, содержащего 0,1 М NaCl и 0,01% азида. Начальная ступень
осаждения PEG была осуществлена с использованием 3,5%-ной конечной концентрации PEG-8000. Осаждение происходило в течение 30 мин при комнатной температуре, затем проводили центрифугирование, и был
оставлен супернатант содержащий IgY. К супернатанту был добавлен дополнительный полиэтиленгликоль для получения конечной концентрации 12,5% PEG. Антитела IgY, осажденные при такой концентрации
полиэтиленгликоля, были отделены при помощи центрифугирования. Антитела IgY на этой стадии очистки очищались дальше с помощью двух методов:
1/ Осажденные антитела IgY были повторно
суспендированы в 10 мМ MES pH 5,6. затем подвергнуты диализу в течение ночи при 4oC против буфера. Образец был затем помещен на 1,5х20 см колонку, содержащую 40 мм смешанной АВХ- кремниевой
смолы (Дж. Т. Бэйкер). Протокол последующей очистки был аналогичен описанному для асцитной жидкости.
2/ Альтернативно, осадок, содержащий IgY, был повторно суспендирован в начальном буфере и затем вновь осажден с использованием насыщенного сульфата аммония (3:1 V/V). Осадок, содержащий IgY, был повторно суспендирован в небольшом объеме дистиллированной H2O и помещен на хранение при 4oC до дальнейшего использования. Процедура иммобилизации для IgY цыплят описана в примере 12.
Пример 8
Кинетическое изучение активности
альфа-амидирующего фермента
Альфа-амидирующий фермент (монооксигеназа), которые мы очистили до гомогенности (как из клеток мозговидной карциномы щитовидной железы СА-77 в культуре, так и из
соответствующей опухолевой ткани, взятой от лабораторных крыс) функционален в превращении неактивных глицин-протяженных пептидных прогормонов в биоактивные C-терминальные амиды. На основании наших
исследований C-терминальная аминокислота прогормона должна быть глициновым остатком для того, чтобы фермент мог распознать его и амидировать его следующим образом:

Проявление in vitro этой активности требует, кроме пептидного субстрата, молекулярного кислорода и восстановителя (L-аскорбиновая кислота). Мы, однако, нашли, что ферментная активность может быть существенно увеличена посредством добавления ионов Cu+2 (сульфат меди) и каталазы. Эта экзогенная медь связывается ферментом и используется как место связывания молекулярного кислорода и активации. С другой стороны, каталаза представляет собой фермент, который служит для удаления перекиси водорода, которая в противном случае будет накапливаться, через побочные реакции вовлекая кислород, абсорбат, медь и разрушая амидирующий фермент.
Мы успешно разработали чувствительные нерадиометрические испытания для обнаружения активности альфа-амидирующего фермента. Эти испытания включают использование синтетических субстратов трипептида. Амидирование этих субстратов отслеживается посредством отделения и определения количества продуктов (амидированный дипептид и трипептид субстрата) с использованием HPLC. Наиболее чувствительное из разработанных исследований используют в качестве субстрата монодансил L-Tyr-Val-Gly.
Это соединение может быть обнаружено при чрезвычайно низких уровнях, поскольку дансильная группа является флуоресцентной. Следовательно, чувствительность этой пробы сравнима с аналогичной радиометрической пробой, разработанной в других лабораториях.
Мы использовали эту пробу для исследования кинетических свойств альфа-амидирующего фермента. В частности, кинетические параметры Км и Vmax были определены посредством исследования концентрации субстрата на ферментативную активность. Км представляет собой меру средства, проявляемого ферментом в отношении определенного субстрата. Чем меньше Км, тем больше средство. Vmax представляет собой максимальную скорость, при которой фермент преобразует субстрат в продукт и излучает при насыщающих концентрациях субстрата (т.е. субстрат присутствует в большом избытке над ферментом).
В типичной пробе для амидирующего фермента реакционная система 50 μl будет включать фермент приблизительно 7 μг, дансил L-Tyr-Val-Gly (lo 40 μм), L-аскорбиновую кислоту (3 мМ), каталазу (jn 0 до 100 μг ml-1) и сульфат меди (jn 0 до 2 μM) в 60 мМ буфера ТЕ при H 7,0. Реакция инициируется при 37oC путем добавления фермента и оканчивается после определенного периода времени путем добавления 0,1 М ЕДТА (конечная концентрация), который связывает медь, делая ее недоступной для фермента.
Альфа-амидирующая моноксигеназа показывает высокое сродство для ферментного амидирования дансила L-Tyr-Val-Gly с Км в диапазоне между 1-2 μM. Эта величина кажется постоянной независимо от степени очистки фермента. Так, ферментный препарат, полученный после хроматографии на Сефакрил S-300, показывает тот же самый Км, что и электрофоретически чистый препарат фермента, полученный на Моно Q pH 8,0 или формы с меньшим молекулярным весом, разделяемые на Моно Q pH 6,0. С другой стороны, как и ожидалось, величины Vmax (выраженные на мг протеина) существенно изменяются в зависимости от степени очистки. Так, если Vmax общей фракции с Сефакрил S-300 составляет приблизительно от 50 до 100 нмоль продукта, образуемого в минуту на мг протеина, Vmax для чистого фермента опухоли составляет около 5000 нмоль продукта, образуемого на мг протеина. Мы нашли, что аскорбат и субстрат могут конкурировать между собой за фермент при некоторых условиях. Например, если концентрация субстрата увеличивается существенно выше уровня насыщения, активность фермента затухает благодаря взаимодействию между ферментом и аскорбиновой кислотой. Таким образом, для каждого конкретного субстрата, по-видимому, имеется тонкий баланс между оптимальными уровнями субстрата/аскорбата, в зависимости от сродства фермента к конкретному субстрату.
Сродство амидирующего фермента к глицин-протяженным
пептидам
Мы использовали пробу амидирования монодансила-L-Tyr-Val-Gly в качестве чувствительной пробы для реакции между ферментом амидирования и несколькими глицин-протяженными прогормонами
пептида. Целью этого исследования было изучение сравнительного сродства фермента при связывании определенных различных типов полипептидных субстратов. Хотя мы показали ранее, что фермент успешно
амидирует глицин-протяженные субстраты человеческого кальцитонина и фактора выделения человеческого гормона роста, это ничего не говорит о способности фермента предпочтительно связывать те или иные
пептиды.
Мы показали, что глицин-протяженный альфа-меланоцитстимулирующий гормон, субстанция Р, аналоги вазопрессина и фактор выделения гормона роста будут взаимодействовать с высоким сродством с амидирующим ферментом, препятствуя превращению дансил L-Tyr-Val-Gly. Далее, С-терминальные 5-членные аминокислотные последовательности глицин-протяженного человеческого кальцитонина, нейропептида Y, холецистокинина, фактора высвобождения кортикотропина, пептида, связанного с кальцитониновым геном, проявляет конкуренцию в этой пробе. Другими словами, фермент имеет способность связывать и, вероятно, амидировать все эти глицин-протяженные пептидные субстраты.
В противоположность этому, когда амидированные пептиды, соответствующие этим субстратам, были изучены, то обнаружилось, что они гораздо менее способны взаимодействовать с ферментом. Способность каталитического сайта фермента распознавать множество различных глицин-протяженных субстратов делает его чрезвычайно полезным общим реагентом для промышленного амидирования прогормона пептида, выработанного с помощью технологии рекомбинантной ДНК.
Пример 9
Иммобилизация
фермента
Очистка α-амидирующего фермента
Перед иммобилизацией альфа-амидирующий фермент был очищен через слабый анионообменник и гель-фильтрацией (пример 4). В некоторых
случаях приготовление фермента может включать дополнительную очистку с использованием либо иммуноафинной хроматографии, либо хроматографии на фенил-сефарозе.
Процедура последовательной иммобилизации не зависит от процедуры очистки; однако специфическая активность должна быть по меньшей мере 25 mU предпочтительно 50 mU или выше.
Иммобилизация a-амидирующего фермента.
Технология иммобилизации для альфа-амидирующего фермента может быть основана на одновременной реакции трех компонентов; фермента, растворимого в воде сополимера акриламида (PAN), и имеющего небольшой молекулярный вес реагента с поперечными связями (ТЕТ). Предварительно созданный полимер (PAN) состоит из акриламида и N-акрилоксисукцинимида, который полимеризован в растворе THF с использованием термальной инициации с азобизом/изобутиронитрил/. Реагент с поперечными связями может быть a, ω-диамином, триэтилентетрамином (ТЕТ), цистамином. Реакция диамина с активными эфирными группами PAN создает поперечные связи цепей полимера через амидную связь и образует нерастворимый гель. Амино-функция фермента (преимущественно ∈ амино группа лизина) в то же самое время реагирует с остаточными активными эфирами на геле и образует стабильные ковалентные амидные связи. Процедура иммобилизации осуществляется в присутствии субстрата и кофакторов. Присутствие субстрата с высокой степенью сродства и кофакторов в концентрации более Km запрещает реакцию между активными эфирами PAN и нуклеофильными группами вблизи или в каталитическом сайте фермента, таким образом защищая фермент от химической инактивации.
Условия иммобилизации:
Частично очищенный α-АЕ растворили в 30 мМ HEPES буфере
с pH 7,0. Отношение PAN и TET устанавливается так, что 15% активных эфиров остаются непрореагировавшими, что служит в качестве связующих сайтов для альфа-амидирующего фермента. Стандартный раствор PAN
представляет собой 20% (W/W). Реактивная смесь альфа-амидирующего фермента состоит из 0,2 μм CuSO4, 40 μм дансил His-Re-Gly и 10 мМ абсорбата и 0,5-2,0 мг α-АЕ (грамм
PAN). Концентрация ТЕТ вычисляется, чтобы равняться 0,85 эквивалентам первичного амина на эквивалент активного эфира. Для определения оптимальных условий реакция запускается без фермента для
установления времени образования геля, то есть времени до добавления ТЕТ, необходимого для достижения образования геля. Оптимум, получаемый для иммобилизации альфа-амидирующего фермента, достигается,
когда добавление фермента смещается ближе к точке образования геля. Общее правило заключается в том, что чем меньше время фермент подвергнут воздействию PAN перед образованием геля, тем выше выход
активного иммобилизированного фермента. Для большинства реакций a-AE (приблизительно 2,0 мг/г PAN) добавляется через 45-60 с после ТЕТ. Фермент-содержащий гель оставляется при комнатной температуре в
течение приблизительно одного часа, чтобы завершить реакцию соединения. После истечения 60 мин гель размельчается с помощью ступки и пестика, приводя к получению фрагментов в среднем с размером 100
μ . Эти частицы вымываются сульфатом аммония для удаления несвязанных реактантов и для преобразования остаточных активных групп эфира в амиды, тем самым покрывая реактивные группы. Выход
активного альфа-амидирующего фермента вслед за иммобилизацией ожидается менее 60% Смыв после иммобилизации может содержать от 30 до 40% начальной активности. Альфа-амидирующий фермент может быть
восстановлен из смыва посредством увеличения концентрации сульфата аммония до 45% что приводит к осаждению активного альфа-амидирующего фермента.
Частицы иммобилизованного геля
проходят для порционной реакции альфа-амидирования, где частицы находятся в суспензии с механической мешалкой. После завершения ферментной реакции частицы осаждаются, и супернатант, содержащий продукт
амидированного пептида, декантируется. Эта процедура не является оптимальной для реакций большего масштаба. Частицы содержащего фермент геля недостаточно прочны, чтобы паковаться в колонну и получить
разумную скорость потока. Для обхода этой проблемы возможны два альтернативных подхода:
1. Гелю дается возможность полимеризации в присутствии стеклянных бус; обычно за минуту до "гелевой
точки" стеклянные бусы добавляются к реакционной смеси и перемешиваются в механической мешалке до покрытия бус слоем раствора PAN. Текучие свойства этого композитного материала значительно лучше, чем
просто гелевых частиц.
2. Альтернативно, частицы PAN смешиваются с фильтрующим Celite 545. Обычно смесь PAN и Celite готовится при PAN, образующем менее 8% W/W (сухой вес) смеси. Для получения колонны этого типа гелевые частицы взвешиваются в 50 мМ Tris HCl pH 7,0 и при постоянном перемешивании добавляется Celite 545 и смешивается в течение двух часов. Колонка упаковывается этим шламмом (3 х 40 см), и температура колонны поддерживается при 37oC для облегчения реакции амидирования. При использовании такого подхода достигается скорость потока 8-10 л в день.
Привлекательной альтернативой колоночной хроматографии является использование системы с тангенциальным потоком. Полимер PAN перед гелевой точкой разливается по листу полисульфоновой пористой подложки 0,45 μм. После образования геля листы могут быть прорезаны для использования в ультрафильтрационных установках с тангенциальным потоком системы Миллипор или Нью Брансвик. В этой конструкции пластины геля с иммобилизованным полисульфоном PAN ферментом укладываются слоями, приводя к значительному увеличению площади поверхности. Этот подход может быть прямо масштабирован до скоростей потока, приближающихся к литрам в минуту. Этот тип системы также обеспечивает удобный способ повторного использования реакционной смеси для максимизации амидирования пептидного субстрата.
Преимущества, достигаемые посредством иммобилизации альфа-амидирующего фермента, включают восстановление и повторное использование фермента. Во-вторых, иммобилизованная матрица увеличивает стабильность фермента и обеспечивает рабочую форму энзима, пригодную для большемасштабных реакций амидирования (от грамм до килограмм субстрата по количеству) в течение протяженных периодов времени.
Пример 10
Приготовление иммуноафинной колонки для очистки альфа-амидирующего фермента
А. Процедура иммобилизации:
Используемая матрица
иммобилизации представляла собой цианоген бромид-активированную Сефарозу 4В (Фармацевтика). Сухой гель был вначале промыт 1 мМ HCl (200 мл/г смолы). Приблизительно 40 мг очищенного поликлонального
антитела, выделенного из желтков куриных яиц, подвергается диализу против 100 мМ NaHCO3 pH 8,3, содержащего 0,5 M NaCl. Соединение осуществляется с использованием 8 мл предварительно
промытой твердой подложки. Реакция осуществлялась в течение 3 ч при
комнатной температуре в Na-бикарбонатном буфере с концентрацией 100 мМ рН 8,3, содержащем 500 мМ NaCl, который был включен для
уменьшения неспецифического связывания протеина с твердым основанием. Остающиеся активные группы геля были блокированы с использованием 0,2 M глицина. После блокирования гель был смыт до пяти раз с
использованием цикла буферов с высоким и низким pH (буфер с высоким pH 100 мМ NaCO pH 8,3 + 500 мМ NaCl, буфер с низким pH 100 мМ ацетат с pH 4,0 + 500 мМ NaCl). Эти ступени промывания удаляют любой
несвязанный белок и блокирующий реагент (глицин) со смолы. Смола с иммунным сродством была помещена при 4oC в буфер с основанием pH, содержащий мертиолат в качестве бактериостатического
агента. Вся последующая хроматография с иммунным сродством осуществляется при 4oC. Аналогичный метод может быть использован для иммобилизации очищенных моноклональных антител.
В. Хроматография с иммунным сродством:
Колонка с иммунным сродством используется как альтернативная стадия высокой эффективности в очистке α-АЕ. Среда культивирования
тканевой культуры клеток Ca-77 (см. пример 4) разбавляется дистиллированной водой и подается под давлением на колонку с DEAE, предварительно уравновешенной 20 мМ Bis-Tris-HCl pH 6,0. Альфа-амидирующий
фермент элюируется с колонки с помощью 50 мМ Трис HCl pH 7,0, содержащего 500 мМ NaCl. Фракции, содержащие a-AE активность, либо подвергаются диализу через буфер Трис HCl pH 7,0, либо очищаются дальше
с использованием гельфильтрации (пример 4) перед иммуноафинной хроматографией. Образцы, содержащие a-АЕ, пропускаются через иммуноадсорбционную колонку с нейтральным pH. Антитела специфично свяжут
альфа-амидирующий фермент, в то время как загрязняющие протеины не будут связаны и будут удалены в элюанте. Активность a-АЕ может быть элюирована с колонки с использованием 100 мМ глицина: HCl буфера
с pH 3,0 (могут быть использованы другие десорбционные агенты, включая мочевину, диоксан, этиленгликоль и NaI). Фракции собираются в 1,0 M Tris HCl pH 7,0 для нейтрализации буферной системы, сохраняя
тем самым активность a-АЕ.
Реферат
Использование: биотехническая, медико-биологическая промышленность. Сущность изобретения: из ткани и клеточных линий мозговидной карциномы щитовидной железы выделены очищенные альфа-амидирующие монооксигеназы, способные обеспечивать катализ преобразования соединений пептидной природы с остатком глицина на C-конце, в том числе и природных L-форм, в соответствующий пептидиламид, для получения активных альфа-амидированных форм регуляторных полипептидов (кальцитонинов, фактора высвобождения гормона роста и др.) полученным ферментом в присутствии кофакторов амидирования (меди, кислорода и восстанавливающего агента) обрабатывают природный или рекомбинантный предшественник с аминокислотной последовательностью амидируемого регуляторного пептида и дополнительным остатком глицина на C-конце. 3 с. и 9 з.п. ф-лы, 22 ил., 2 табл.

Комментарии