Новые слитые белки тромбомодулина, обеспечивающиенаправленный перенос к тканевому фактору, в качестве антикоагулянтов - RU2320366C2
Код документа: RU2320366C2
Чертежи









Описание
Сохранение надлежащего баланса между прокоагулирующей и антикоагулирующей активностью в кровеносных сосудах существенно для нормального гемостаза (Е.W.Davie и др., Biochemistry, 30 (43), 1991, с. 10363-10370). Нарушение баланса в направлении коагуляции приводит к тромбозам, которые могут вызвать сердечный приступ, инсульт, легочную эмболию и венозный тромбоз. Существует необходимость в создании более эффективных и безопасных антикоагулянтов для лечения специфических тромбозных нарушений.
Тканевой фактор (TF) - это трансмембранный гликопротеин, который является основным инициатором каскада коагуляции (Y.Nemerson, Thromb. Haemost. 74 (1), 1995, с. 180-184). При нормальных физиологических условиях TF не находится в контакте с кровью. При сосудистом повреждении воздействие на кровь субэндотелиального TF и коллагена приводит к активации факторов коагуляции и тромбоцитов и впоследствии к образованию гемостатической пробки. Неуместная индукция экспрессии TF в различных клинических ситуациях может привести к угрожающему жизни тромбозу и/или содействовать патологическим осложнениям. Воздействие TF на последующее разрушение тромбоцита считается ответственным за тромбозную окклюзию, приводящую к острому инфаркту миокарда или инсульту. В этих ситуациях противоспалительные сигнальные пути, активированные факторами коагуляции, содействуют также образованию отека и увеличению объема инфаркта. Сосудистое повреждение, ассоциированное с ангиопластикой, приводит к активации TF на клетках гладкой мышцы (SMC), которая, как считается, индуцирует пути передачи клеточных сигналов, связанные с рестенозом. Сверхэкспрессия TF при раке и грамотрицательном сепсисе приводит к угрожающему жизни тромбозу и активации воспалительных путей.
Комплекс факторов VIIa (FVIIa)/TF включается в патогенетический механизм при различных тромбозных заболеваниях, и циркулирующий уровень TF является фактором риска для некоторых пациентов. Факторы VIIa и TF играют уникальные роли при сосудистом повреждении в сохранении гемостаза и инициации тромбоза. Экспрессируется TF обычно в адвентициальной оболочке, но при сосудистом заболевании несоответственно активируется и экспрессируется в середине и неоинтиме сосуда. Экспрессия TF в атеросклеротических бляшках повышена и экранирована от крови толстым волокнистым слоем, который может разрушаться, высвобождая TF. Хирургические вмешательства, такие как баллонная ангиопластика, установка стента или эндартерэктомия, повреждают сосудистую стенку и высвобождают находящийся под ней TF. В атеросклеротической толстостенной бляшке с высоким содержанием липидов спонтанное разрушение или эрозия эндотелия приводит к воздействию TF и тромбозу, заканчивающимся нестабильной стенокардией и инфарктом миокарда. TF может циркулировать в происходящих от клеток микрочастицах, и уровни циркулирующего TF при нестабильной стенокардии являются повышенными, допуская предположение о том, что этот циркулирующий TF может содействовать образованию тромба (Н.Soejima и др., Circulation, 99 (22), 1999, с. 2908-2913). Часто злокачественная опухоль ассоциируется с состоянием гиперкоагуляции, объясняемым сверхэкспрессией TF на опухолевых клетках. Это предрасполагает пациента к тромбозу глубоких вен, легочной эмболии и низкой степени коагулопатии потребления (DIC). Коагулопатия потребления приводит к отложениям фибрина в капиллярных сосудах, способствуя повреждению многих органов. Результаты, полученные на моделях тромбоза от острого артериального нарушения, указывают, что основанные на белках ингибиторы FVIIa/TF, такие как ингибитор активного центра фактора VIIa (FVIIai) и ингибитор метаболических путей тканевого фактора (TFPI), являются эффективными антитромбозными средствами с меньшей кровопотерей, по сравнению с ингибиторами тромбина и фактора Ха (FXa). Кроме того, ингибиторы FVIIa/TF превосходят другие антикоагулянты (например, гепарин, ингибиторы FXa) в предотвращении образования утолщения неоинтимы и сосудистого стеноза после нарушения, вызванного баллонной ангиопластикой (Y. Jang и др., Circulation, 92 (10), 1995, с. 3041-3050).
Тромбомодулин (ТМ) - это трансмембранный гликопротеин, который обладает антикоагулирующими свойствами и преимущественно экспрессируется на полостной поверхности эндотелиально-клеточной выстилки кровеносных сосудов (N.L.Esmon и др., J.Biol. Chem. 257 (2), 1982, с. 859-864; H.H. Salem и др., J. Biol. Chem. 259 (19), 1983, с. 12246-12251). Зрелый с полной последовательностью ТМ является модульным белком из 557 аминокислотных остатков, составленным из 5 структурных доменов: N-концевого гидрофобного участка (остатки 1-226); богатого цистеином участка (остатки 226-462); O-гликозилированного Ser/Thr-богатого участка (остатки 463-497); гидрофобного трансмембранного участка (остатки 498-521); и С-концевого цитоплазматического хвоста (остатки 522-557).
Богатый цистеином участок включает шесть повторяющихся структур, гомологичных предшественнику эпидермального фактора роста (EGF), названного EGF-подобный, EGF-гомологичный или EGF-домены. Богатый цистеином участок далее может быть разделен на 3 домена: EGF-подобные повторы 1, 2 и 3 (EGF 123, остатки 226-344), междоменная петля между EGF3 и EGF4 (остатки 345-349) и EGF-подобные домены 4, 5 и 6 (EGF456, остатки 350-462). Функция EGF456 состоит в том, чтобы опосредовать связывание и активацию белка С. Одно исследование предполагает, что пятый и шестой EGF-подобные повторы (EGF5, остатки 390-407 и EGF6, остатки 427-462 соответственно) обладают способностью связывать тромбин (S. Kwosawa и др., J. Biol. Chem. 263 (13), 1988, с. 5993-5996); другое предполагает, что домена EG456 достаточно, чтобы действовать в качестве кофактора опосредованной тромбином активирующей белок С активности (М. Zushi и др., J. Biol. Chem. 264 (18), 1989, с. 10351-10353). Ser/Thr-богатый домен усиливает опосредованное EGF456 связывание тромбина. Третий EGF-подобный повтор (EGF3, остатки 311-344) требуется для активации ингибитора фибринолиза, активируемого тромбином (TAFI). Описано несколько точечных мутантов, которые препятствуют активации TAFI (W. Wang и др., J. Biol. Chem. 275 (30), 2000, с. 22942-22947). Комплекс тромбин/ТМ превращает белок С в активированный белок С (АРС), который, в свою очередь, разрушает факторы Va и Villa, препятствуя тем самым генерации тромбина. Следовательно, ТМ функционирует как молекулярный переключатель, превращая тромбин из прокоагулянта в антикоагулянт.
Значение Кm белка C для комплекса тромбин/ТМ снижается 10-кратно, когда ТМ локализован на поверхности мембраны (C.N.Esmon, ESEB J. 9 (10), 1995, с. 946-955). Концентрация белка С в крови (0,065 мкМ) значительно ниже сообщенного значения Кm (5 мкМ) для растворимого комплекса ТМ/тромбин, отсюда утверждение, что ТМ на прокоагулирующей мембранной поверхности приведет к заметному локальному повышению скорости генерации белка С.
ТМ ингибирует тромбоз по механизму, отличному от гепарина или его производных. Гепарин является кофактором антитромбина III и ингибирует как FXa, так и тромбин по антитромбин III-зависимому механизму. Связанный с тромбом тромбин защищен от действия антитромбина III, который ограничивает антитромбозную эффективность гепарина или гепарина с низкой молекулярной массой (LMWH) на существующих ранее сгустках. Это объясняет отсутствие у гепарина или LMWH способности ингибировать рост тромба, запускаемый связанным со сгустком тромбином или протромбиназой в исследованиях на приматах, кроме человека. В противоположность этому рекомбинантный ТМ ослабляет индуцированную сгустком генерацию тромбина и образование фибрина дозозависимым способом (М. Mohri и др., Thromb. Haemost. 80 (6), 1998, с. 925-929). Ингибирующий эффект ТМ аннулируется антителом к белку С. Подавление связанной со сгустком прокоагулянтной активности является клинически актуальным, т.к. связанная со сгустком прокоагулирующая активность приводит к более быстрому росту тромба и в конечном итоге к сосудистой окклюзии или тромбоэмболическим осложнениям. Ингибирование роста тромба позволяет эндогенной фибринолитической системе удалять сгустки быстрее и полностью. Кроме того, ожидается также, что ТМ более эффективен, чем гепарин, при патологических состояниях, при которых антитромбин в плазме снижен, таких как коагулопатия потребления (DIC). Несмотря на то что как ТМ, так и гепарин ингибируют поглощение тромбоцитов и фибриногена при экспериментальной DIC, только ТМ был эффективен, когда уровни антитромбина III снижались.
Краткое содержание изобретения
Настоящее изобретение обеспечивает новые слитые белки, которые действуют как антикоагулянты и включают белок, который взаимодействует или с тканевым фактором (TF), или с комплексом фактора VIIa/тканевой фактор (FVIIa/TF), и функционально связан с тромбомодулиновым (ТМ) доменом EGF456, одним или в комбинации с другим доменом ТМ, выбранным из группы, состоящей из домена N-концевого гидрофобного участка, домена EGF123, междоменной петли между EGF3 и EGF4 и O-гликозилированного Ser/Thr-богатого домена или их аналогов, фрагментов, производных или вариантов.
Антикоагулирующий слитый белок по данному изобретению направлен на и связывает TF или комплекс FVIIa/TF в участке повреждения, локализуя ТМ в участке повреждения и таким образом предотвращая образование тромба и в связи с этим действуя более эффективно в качестве антикоагулянта, по сравнению или с растворимым антителом к TF, или растворимым ТМ, или фрагментами ТМ. Слитый белок более эффективен, чем гепарин с низкой молекулярной массой (LMWH), при лечении некоторых заболеваний, включая, но без ограничения, сепсис, коагулопатию потребления, ишемический удар, тромбоз глубоких вен, острые коронарные синдромы, тромбозные осложнения после ангиопластики и коагулопатию при прогрессирующей злокачественной опухоли. Кроме того, слитый белок применяется в хирургических операциях на капиллярных сосудах, в трансплантатах кожи и вен и при пересадке органов.
В другом аспекте изобретение обеспечивает фармацевтические композиции, включающие в качестве объекта слитые белки.
В другом аспекте изобретение обеспечивает способ предохранения пациента от образования тромба, включающий введение упомянутому пациенту терапевтически эффективного количества слитого белка, и в связи с этим подавление генерации тромбина без непосредственного воздействия на другие параметры коагуляции, такие как активация и агрегация тромбоцитов.
В другом аспекте изобретение относится к способу предупреждения и лечения тромбоза глубоких вен (DVT) или коагулопатии потребления (DIC), или острого коронарного синдрома, или злокачественной опухоли с проявлением коагулопатии у пациента, включающему введение упомянутому пациенту терапевтически эффективного количества слитого белка.
В другом аспекте изобретение относится к способу регулирования воспалительного ответа у пациента, включающему введение упомянутому пациенту терапевтически эффективного количества слитого белка.
Еще в одном аспекте слитый белок по изобретению может быть использован для образования не являющегося тромбогенным покрытия медицинских инструментов, контактирующих с кровью.
В другом аспекте изобретение относится к набору, содержащему слитый белок, включающий белок, который обеспечивает направленный перенос и связывает TF или комплекс FVIIa/TF и домены ТМ. Альтернативно набор может включать последовательности ДНК, кодирующие компоненты слитого белка.
Раскрываются также способы получения слитых белков по изобретению как рекомбинантные, так и синтетические.
Описание графиков
Фиг.1. Связывание антитела scFv(TF)3e10 с растворимым TF (sTF) увеличивает кажущееся сродство sTF к FVIIa. Активационный анализ sTF/FVIIa осуществляли, как описано в примере 5, озаглавленном «Анализ активации sTF/FVIIa», используя 2 нМ FVIIa в присутствии и в отсутствие 800 нМ scFv(TF)3e10. Титровали sTF в пробе и определяли скорость расщепления хромогенного субстрата (S-2266). Значение кажущейся KD для sTF рассчитывали, используя стандартную 4-параметрическую аппроксимацию.
Фиг.2. Измерение сродства связывания scFv(TF)3e10 к sTF. Анализ sTF/FVIIa осуществляли, как описано в примере 5, озаглавленном «Анализ активации sTF/FVIIa», используя 3 нМ sTF и 2 нМ FVIIa. Использованная концентрация sTF была ниже значения KD для связывания с FVIIa. Связывание антитела scFv (TF)3e10 снижало значение KD sTF для связывания с FVIIa, приводя к повышению образования комплекса sTF/FVIIa и, следовательно, скорости расщепления хромогенного субстрата S2266. Прибавляли scFv(TF)3e10 при увеличивающихся концентрациях и повышенную скорость реакции использовали, чтобы определить кажущуюся KD антитела для sTF, используя стандартную 4-параметрическую аппроксимацию.
Фиг.3. Микрокалориметрический анализ показывает, что scFv(TF)3e10 имеет 20-кратно более высокое сродство к комплексу sTF/FVIIa, чем к одному sTF. Комплекс sTF/FVIIa был предварительно получен прибавлением 2,3-кратного молярного избытка FVIIai по отношению к sTF. Гранулометрическую эксклюзионную хроматографию использовали, чтобы подтвердить, что sTF полностью комплексирован. Для определения сродства антитела к комплексу прибавляли 1,2 мкМ комплекса sTF/FVIIa в ячейку микрокалориметра и 65 мкМ антитела scFv(TF)3e10 прибавляли в шприц. Для определения сродства антитела к одному sTF прибавляли 10 мкМ sTF в ячейку и 141 мкМ scFv(TF)3e10 прибавляли в шприц. Анализ данных проводили, используя программу MicroCal Origin. Данные были аппроксимированы к одному сайту связывания.
Фиг.4. Антитело scFv(TF)3e10 дозозависимо ингибирует анализ активации фактора Х (FX). Подробности анализа описаны в примере 5, озаглавленном «Анализ активации фактора X». IC50 означает дозу, требуемую для достижения 50% ингибирования.
Фиг.5. Слитый белок сильнее подавляет коагуляцию, чем антитело TF или один ТМi456. Анализ протромбинового времени (ПВ) проводили, чтобы сравнить слитый белок с антителом TF или одним TMi456. Соответствующий объем концентрированного ингибитора или антитела TF (scFv(TF)3e10), TMi456 или слитого белка (scFv(TF)3e10-TMi456) прибавляли к 100 мкл рекомбинантного человеческого тромбопластина (Ortho Recombiplastin). Приблизительно 2 мин позже прибавляли 100 мкл восстановленной человеческой плазмы. Время коагуляции определяли на коагулометре Haemoliance. Кривые ответа на дозу получали для каждого ингибитора и затем использовали регрессионный анализ, чтобы рассчитать концентрацию (в нМ), необходимую для двукратного увеличения времени свертывания крови.
Фиг.6. Слитый белок сохраняет полную кофакторную активность для активации белка С. Анализ, описанный в примере 5, озаглавленный «Анализ (хромогенный) активации белка С», включал 20 мкл образца ТМ или ТМ1456, который содержал домены EGF4-6 и междоменную петлю между EGF3 и EGF4, или слитый белок (scFv(TF)3e10-TMi456), 20 мкл 1,5 мкМ белок С и 20 мкл 3 нМ α-тромбин. Активации давали возможность проходить в течение 1 ч. Фазу активации останавливали путем добавления 20 мкл 0,16 ед/мл гирудина. Затем прибавляли 100 мкл 1 мМ хромогена S2266 и А405 определяли каждые 10 сек в течение 30 мин. Скорость реакции зависит от количества генерированного активированного белка С. Данные выражены в мОП/мин.
Фиг.7. Скорость активации белка С слитым белком повышается на содержащих TF фосфолипидных поверхностях. На скорость активации белка С не оказывает влияния прибавление везикул TF. Образец для анализа, описанного в примере 5, озаглавленном «Анализ активации белка С (на TF-богатой поверхности)», содержал 20 мкл образца ТМ, или ТМ1456, или слитый белок (scFv (TF)3e10-TMi456), 20 мкл 1,5 мкМ белка С, 20 мкл 3 нМ α-тромбина и 20 мкл буфера или везикул TF (инновин, человеческий рекомбинантный TF, 4х нормальная концентрация для ПВ). Активации давали проходить в течение 1 ч. Фазу активации останавливали добавлением 20 мкл 0,16 ед/мл гирудина. Затем прибавляли 100 мкл 1 мМ хромогена S2266 и А405 определяли каждые 10 сек в течение 30 мин. Скорость реакции зависела от количества генерированного активированного белка С. Данные выражены в мОП/мин.
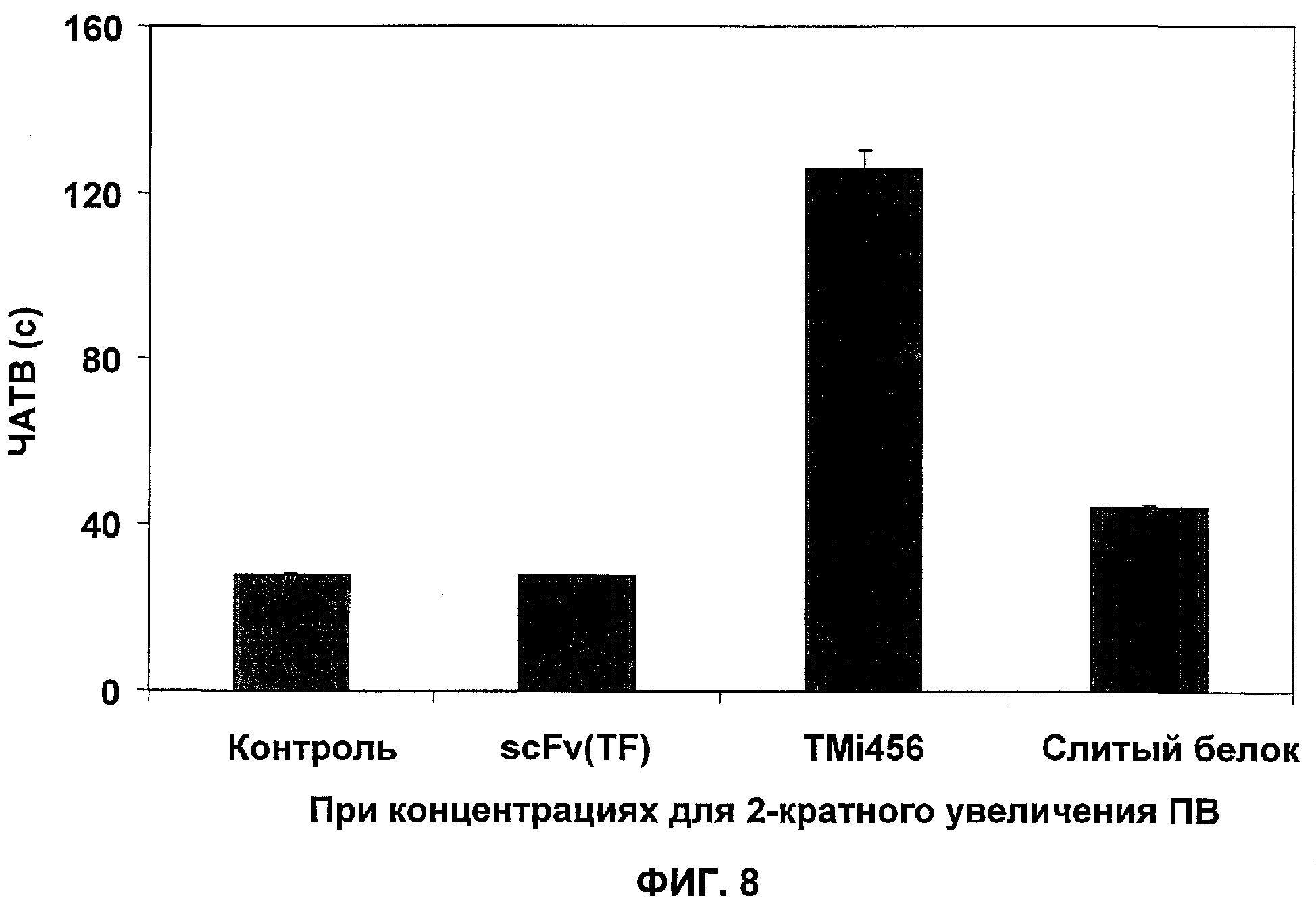
Фиг.8. Слитый белок показывает более высокую специфичность к индуцированной TF коагуляции, чем TMi456. Анализ частичного активированного тромбопластинового времени (ЧАТВ) чувствителен к ингибиторам внутреннего и центрального метаболических путей коагуляции. Коагуляция, которая происходит в данном анализе, независима от TF. Ингибиторы - или антитело TF (scFv(TF)3e10), TMi456, или слитый белок (scFv(TF)3e10-TMi456) - разбавляли в 50 мкл восстановленной человеческой плазмы до конечной концентрации, которая давала двукратное увеличение в анализе протромбинового времени (ПВ). В коагулометр затем прибавляли 50 мкл реагента АРТТ (кефалин из кроличьего мозга в 0,1 мМ эллаговой кислоте, с буфером, стабилизаторами и консервантами; Alexin HS) и 50 мкл раствора CaCl2 (0,02 моль/л) и определяли время свертывания крови в сек.
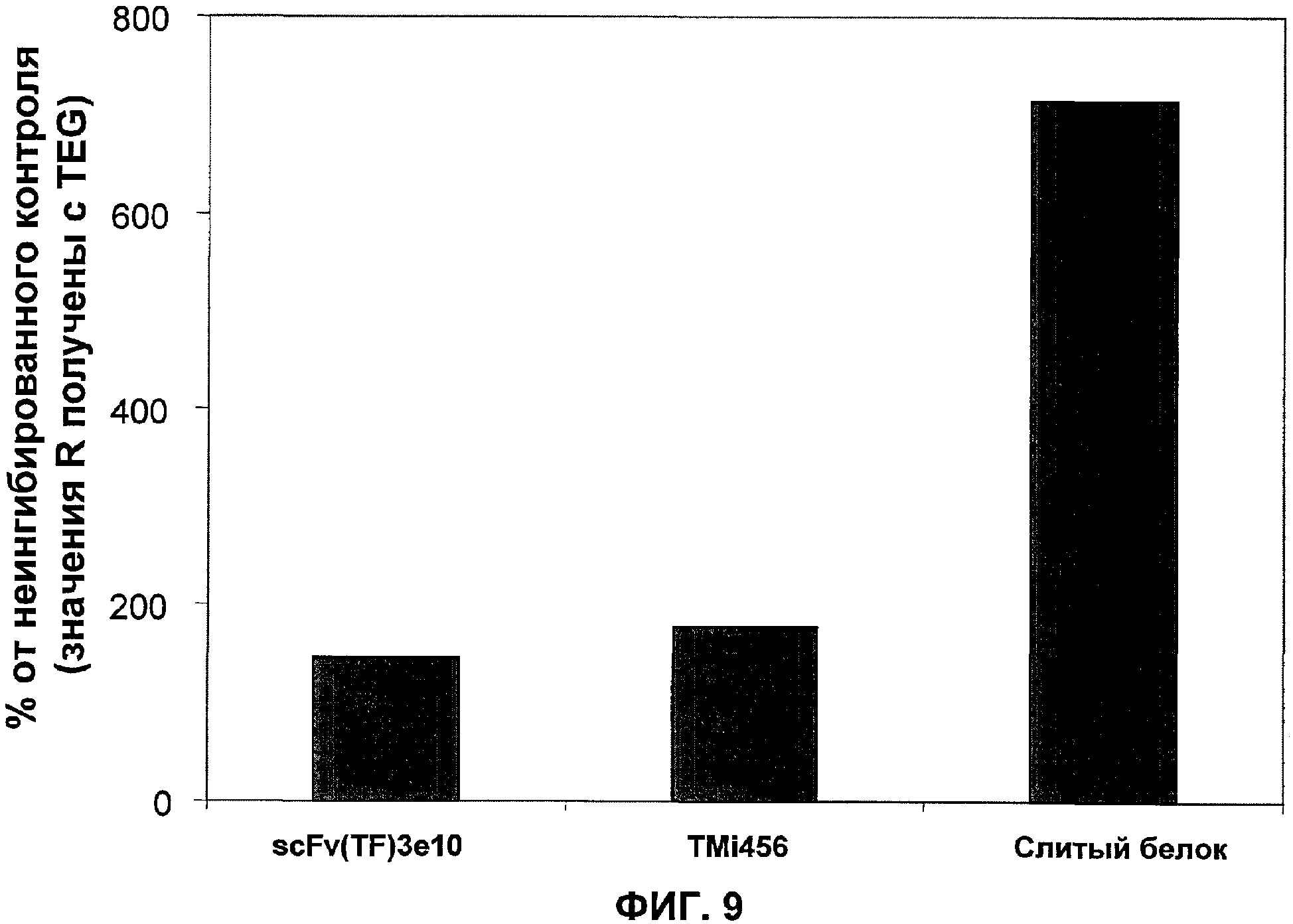
Фиг.9. Слитый белок сильнее подавляет индуцированную TF коагуляцию цельной крови, чем любой из его компонентов в отдельности. Коагуляцию цельной крови анализировали, используя анализатор тромбоэластограф (TEG) Haemoscope. К цельной крови в цитратном буфере прибавляли 120 нМ антитела TF (scFv(TF)3e10), TMi456 или слитый белок (scFv(TF)3e10-TMi456) вместе с 10 мкл тромбопластинового реагента (разведение 1:64) и 20 мкл 0,2 М CaCl2. Значение R (время до начального образования фибрина) получали для каждого образца. Данное значение затем превращали в % не ингибированного контрольного значения R.
Фиг.10. Слитый белок проявляет более предсказуемый дозовый ответ, чем LMWH, в анализе коагуляции цельной крови (TEG). К цельной крови в цитратном буфере прибавляли увеличивающиеся концентрации (15 нМ начальная и увеличенные с помощью инкрементов 2х) слитого белка (scFv(TF)3e10-TMi456) или увеличивающиеся концентрации (0,15 ед/мл начальная и увеличенные 2х) эноксапарина (LMWH) наряду с 10 мкл тромбопластинового реагента (разведение 1:64) и 20 мкл 0,2 М CaCl2. Значение R (время до начала образования фибрина) получали для каждого образца и строили график зависимости от относительной концентрации (принимали самую низкую концентрацию за 1 для каждого (подобного значения R), затем последующие концентрации увеличивали 2х).
Фиг.11. Слитый белок scFv(TF)3e10-TMi456 эффективен in vivo на модели коагулопатии потребления (DIC). Антитело TF (scFv(TF)3e10) и слитый белок (scFv(TF)3e10-TMi456) оценивали на крысиной модели тромбоэмболии, описанной в примере 8, по (А) проценту смертности и (Б) количественному показателю распространенность заболевания-смертности. (А) В обработанной наполнителем группе использованная доза TF привела к 60% летальности (LD60). Слитый белок scFv(TF)3e10-TMi456 при 0,7 нмоль/кг полностью предотвращал гибель. В противоположность этому, scFv(TF)3e10 при 0,7 нмоль/кг не влияло на гибель. Слитый белок scFv(TF)3e10-TMi456 был более эффективным, чем 10-кратно более высокая доза scFv(TF)3e10. (Б) В обработанной наполнителем группе применяемая in vivo доза TF приводила к средней количественной оценке распространенности заболевания-смертности в 2,6 балла, основанной на следующей оценке в баллах: 0 - нет воздействия; 1 - слабая дыхательная недостаточность (восстановление в течение 30 мин); 2 - острая дыхательная недостаточность (агонирующее состояние, восстановление требует более чем 60 мин); и 3 - гибель. Слитый белок scFv(TF)3e10-TMi456 дозозависимым образом предотвращал индуцированную TF гибель и дыхательную недостаточность при значении 50% эффективной дозы ED50 0,46 нмоль/кг (0,019 мг/кг). В дозе 7,0 нмоль/кг scFv(TF)3e10-TMi456 полностью предотвращал как гибель, так и дыхательную недостаточность, а в дозе 0,7 нмоль/кг полностью предотвращал гибель и значительно уменьшал дыхательную недостаточность. В противоположность этому, scFv(TF)3e10 при 0,7 нмоль/кг не оказывало влияния на гибель и слабо либо совсем не действовало на дыхательную недостаточность. Слитый белок scFv(TF)3e10-TMi456 был более эффективен, чем 10-кратно более высокая доза scFv(TF)3e10.
Подробное описание изобретения
Антикоагулирующий слитый белок по настоящему изобретению включает белок, который обеспечивает направленный перенос и взаимодействует или с тканевым фактором (TF), или с комплексом фактор VIIa/тканевой фактор (FVIIa/TF), который функционально связан с доменом EGF456 тромбомодулина (ТМ), одним или в комбинации по меньшей мере с другим доменом ТМ, выбранным из группы, состоящей из N-концевого домена гидрофобного участка, домена EGF123, междоменной петли между EGF3 и EGF4, O-гликозилированного Ser/Thr-богатого домена или их аналогов, фрагментов, производных или вариантов.
Определения
В описании по настоящему изобретению определены следующие термины, как указано ниже.
Термин «рекомбинантные белки или пептиды» относится к белкам или пептидам, полученным с использованием методик рекомбинантной ДНК, т.е. полученным из микробных клеток и клеток млекопитающих, трансформированных с помощью конструкции экзогенной ДНК, кодирующей требуемый полипептид. Белки или полипептиды, экспрессированные в большинстве бактериальных культур, будут свободны от гликана. Белки и полипептиды, экспрессированные в дрожжевых клетках, имеют гликозилированную структуру, отличную от таковой, экспрессированной в клетках млекопитающих.
Термин «нативные» белки или полипептиды относится к белкам или полипептидам, выделенным из природных источников. Термин «нативный ТМ» будет включать существующий в природе ТМ и его фрагменты.
Термин «кодирующая последовательность» ДНК означает последовательность ДНК, которая транскрибируется в мРНК и транслируется в полипетид в клетке-хозяине при помещении под контроль соответствующих регуляторных последовательностей. Границы кодирующей последовательности определяют с помощью инициирующего кодона на 5'-N-конце и трансляционного терминирующего кодона на 3'-С-конце. Кодирующая последовательность может включать прокариотические последовательности, кДНК из эукариотической мРНК, геномные последовательности ДНК и синтетические последовательности ДНК. Последовательность терминации транскрипции обычно будет располагаться в положении 3' по отношению к кодирующей последовательности.
Термин «слитый белок» означает белок, возникающий в результате экспрессии по меньшей мере двух функционально связанных гетерологичных кодирующих последовательностей. Слитый белок по изобретению состоит из белка, который обеспечивает направленный перенос и взаимодействует или с TF, или с комплексом FVIIa/TF, который функционально связан только с доменом EGF456 тромбомодулина (ТМ) или находится в комбинации по меньшей мере с другим доменом ТМ, выбранным из группы, состоящей из N-концевого домена гидрофобного участка, домена EGF123, междоменной петли между EGF3 и EGF4, O-гликозилированного Ser/Thr-богатого домена или их аналогов, фрагментов, производных или вариантов.
Термин «белок, который обеспечивает направленный перенос» означает белок, который связывается или взаимодействует с другим белком или белковым комплексом. Белок, обеспечивающий направленный перенос, по данному изобретению означает белок, который связывается или взаимодействует с TF или комплексом FVIIa/TF. Например, антитело к TF или к комплексу FVIIa/TF является белком, обеспечивающим направленный перенос, по данному изобретению. Двумя другими примерами белков, обеспечивающих направленный перенос, являются ингибированный по активному центру фактор VIIa (FVIIai), который может связывать TF, образуя неактивный комплекс FVIIai/TF, и ингибитор метаболизма тканевого фактора (TFPI), который может связывать и инактивировать комплекс FVIIa/TF.
Термин «нуклеотидная последовательность» означает гетерополимер из дезоксирибонуклеотидов (основания - аденин, гуанин, тимин или цитозин). Последовательности ДНК, кодирующие слитые белки по данному изобретению, могут быть составлены из синтетических фрагментов ДНК, происходящих от кДНК, и коротких олигонуклеотидных линкеров, чтобы обеспечить получение синтетического гена, который способен экспрессироваться с помощью рекомбинантного вектор экспрессии. При обсуждении структуры отдельных двухцепочечных молекул ДНК последовательности могут быть описаны в контексте согласно обычному правилу приведения последовательности только в направлении от 5'-конца к 3'-концу вдоль не транскрибированной цепи кДНК.
Термин «рекомбинантный вектор экспрессии» означает конструкцию реплицируемой ДНК, использованную для того, чтобы или амплифицировать, или экспрессировать ДНК, кодирующую слитые белки по настоящему изобретению. Вектор экспрессии содержит контролирующие последовательности ДНК и кодирующую последовательность. Контролирующие последовательности ДНК включают промоторные последовательности, сайты связывания рибосом, сигналы полиаденилирования, последовательности терминации транскрипции, расположенные против хода транскрипции домены и энхансеры. Рекомбинантные системы экспрессии, как определено в контексте, будут экспрессировать слитые белки при индукции регуляторных элементов.
Термин «трансформированные клетки-хозяева» относится к клеткам, которые были трансформированы и трансфицированы экзогенной ДНК. Экзогенная ДНК может быть или может не быть интегрирована (ковалентно связана) с хромосомной ДНК, составляющей геном клетки. Например, в прокариотах и дрожжах экзогенная ДНК может сохраняться в эписомальном элементе, таком как плазмида, или устойчиво интегрировать в хромосомную ДНК. Что касается эукариотических клеток, то устойчиво трансформированная клетка означает клетку, в которой экзогенная ДНК стала интегрированной в репликацию хромосом. Эта стабильность демонстрируется способностью линий или клонов эукариотических клеток продуцировать популяцию дочерних клеток, содержащих экзогенную ДНК.
Термин «тромбомодулин (ТМ)» относится к гликопротеину поверхности клеток эндотелия, который образует с высоким сродством к комплексу с тромбином. Гены, кодирующие нативный ТМ (как его геномную форму, так и в форме кДНК), были выделены из бычьего и человеческого источников и секвенированы (R.W.Jackman и др., Proc. Natl. Acad. Sci. USA, 83 (23), 1986, с. 8834-8838 и R.W. Jackman и др., Proc. Natl. Acad. Sci. USA, 84 (18), 1987, с. 6425-6429, обе ссылки включены в контекст путем цитирования). Последовательности ТМ для быка, человека и мыши обладают высокой степенью гомологии друг с другом. В случае человеческого ТМ кДНК кодирует белок 60,3 кДа из 575 аминокислот, включающий сигнальную последовательность примерно из 18 аминокислот, см., например, патент US 5827824.
Когда тромбин связывается с ТМ, может быть тысячекратное или большее увеличение скорости активации белка С, которая образует активированный антикоагулирующим ферментом белок С. Кроме того, когда тромбин связывается с ТМ, тромбин больше не работает в качестве прокоагулирующего фермента. А именно, катализируемые тромбином образование фибрина, активация фактора V и активация тромбоцитов, - все ингибируются в присутствии ТМ. Таким образом, ТМ превращает тромбин в физиологический антикоагулянт.
Термин «тромбомодулиновый (ТМ) домен» относится к дискретной аминокислотной последовательности, которая может быть ассоциирована с особой функцией или особенностью ТМ, такой как структура, характерная для третичной структуры. Ген ТМ полной протяженности кодирует предшественник или прополипептид, содержащий следующие домены: аминокислоты 18-1 - сигнальная последовательность; аминокислоты 1-226 - N-концевой гидрофобный участок; аминокислоты 227-462 - богатый цистеином участок; состоящий из 6 тандемных EGF-подобных повторов, соединенных небольшими междоменными пептидами или петлями; аминокислоты 463-497 - O-гликозилированный Ser/Thr-богатый участок; аминокислоты 498-521 - гидрофобный трансмембранный участок и аминокислоты 522-557 - С-концевой цитоплазматический хвост. Богатый цистеином участок может быть далее разделен на 3 домена: аминокислоты 226-344 - EGF123, состоящий из EGF-подобных повторов 1, 2 и 3 (остатки 226-344); аминокислоты 345-349 - междоменная петля между EGF3 и EGF4 и аминокислоты 350-462 - EGF456, состоящий из EGF-подобных доменов 4, 5 и 6 (см., например, C.S. Yost и др., Cell, 34 (3): 1983, с. 759-766; D.Z. Wen и др., Biochemistry, 26 (14), 1987, с. 4350-4357, и W. Wang и др., 2000, см. выше, при этом все публикации включены в настоящее описание в качестве ссылки).
Термины «аналог», «фрагмент», «производное» и «вариант», когда относятся к слитым белкам по данному изобретению, а также к белкам, обеспечивающим направленный перенос и доменам ТМ, означают аналоги, фрагменты, производные и варианты слитых белков, белков, обеспечивающих направленный перенос, и доменов ТМ, которые фактически сохраняют такую же биологическую функцию или активность, как описанные ниже.
Термин «аналог» включает прополипептид, который содержит внутри себя аминокислотную последовательность слитого белка по данному изобретению. Активный слитый белок по данному изобретению может быть отщеплен от дополнительных аминокислот, которые замыкают молекулу предшественника слитого белка, с помощью природных превращений in vivo или с помощью методик, хорошо известных специалистам, таких как энзиматическое или химическое расщепление. Например, нативный ТМ естественно экспрессируется как полипептид из 575 аминокислот, который затем подвергается процессингу in vivo, высвобождая активный зрелый полипептид из 557 аминокислот.
Термин «фрагмент» означает часть слитого белка, белка, обеспечивающего направленный перенос, или доменов ТМ, которые фактически сохраняют одинаковую функциональную активность, как показано в анализах in vivo, раскрытых в контексте, как описано далее.
Термин «производное» включает все модификации слитого белка, которые фактически сохраняют функции, раскрытые в контексте, и включают дополнительную структуру с сопутствующей функцией, например, слитые белки, модифицированные полиэтиленгликолем (PEG), которые имеют большее время полувыведения, O-гликозилированные слитые белки, модифицированные добавлением сульфата хондроитина, и биотинилированные слитые белки, как описано далее.
Термины «по существу подобная функциональная активность» и «по существу та же самая биологическая функция или активность» означают каждый, что интенсивность биологической активности находится в интервале 30-100% или более от биологической активности, продемонстрированной полипептидом, с которым происходит сравнение, когда биологическая активность каждого полипептида определена с использованием той же методики или того же анализа. Например, слитый белок или домен ТМ, который имеет по существу подобную функциональную активность, что и слитый белок примера 2 (последовательность с идентификационным номером 2 (SEQ ID No:2)), означает, что белок при тестировании в анализе (хромогенном) активации белка С, описанном в примере 5, демонстрирует накопление активированного белка С. Белок, который обеспечивает направленный перенос и имеет по существу подобную функциональную активность, что и антитело к TF примера 1 (SEQ ID No:1), означает, что белок при тестировании в анализе sTF/FVIIa или в анализах активации FX, описанных в примере 5, демонстрирует способность связывать или нейтрализовать TF или комплекс FVIIa/TF.
Термин «подобие» между двумя полипептидами определяется путем сравнения аминокислотной последовательности и ее консервативных аминокислотных заместителей одного полипептида с последовательностью второго полипептида. Такие консервативные заместители включают таковые, описанные выше в The Atlas of Protein Sequence and Structure, 5, Dayhoff и Argos (1989) EMBO J. 8, 1987, с. 779-785. Например, аминокислоты, принадлежащие к одной из следующих групп, представляют консервативные изменения:
-Ala, Pro, Gly, GLN, Asn, Ser, Thr;
-Cys, Ser, Tyr, Thr;
-Val, ILe, Leu, Met, Ala, Phe;
-Lys, Arg, His;
-Phe, Tyr, Trp, His и
-Asp, Glu.
Все другие технические термины, использованные в контексте, имеют такое же значение, которое используют специалисты в области, к которой принадлежит настоящее изобретение.
Белок, обеспечивающий направленный перенос
Белок, обеспечивающий направленный перенос, по изобретению означает белок, который обладает способностью специфически связываться с конкретной предварительно выбранной молекулой-мишенью, например TF или комплексом FVIIa/TF, и затем служит для направления слитого белка в клетку или ткань, содержащую предварительно выбранную молекулу-мишень.
В одном варианте воплощения по данному изобретению белок, обеспечивающий направленный перенос, означает антитело, которое может связываться и нейтрализовать TF или комплекс FVIIa/TF. Термин «антитело», как он использован в контексте, включает интактные молекулы иммуноглобулина (Ig), а также их фрагменты, такие как Fab, F(ab')2 и Fv, которые способны связывать эпитоп TF или комплекса FVIIa/TF. Обычно требуется по меньшей мере 6, 8, 10 или 12 смежных аминокислот, чтобы образовать эпитоп. Однако для эпитопов, которые включают не являющиеся смежными аминокислоты, может потребоваться больше аминокислот, например, по крайней мере 15, 25 или 50.
Обычно антитело, которое специфически связывается с TF или комплексом FVIIa/TF, обуславливает детектирующий сигнал по меньшей мере 5-, 10- или 20-кратно более сильный, чем детектирующий сигнал, обусловленный другими белками при использовании в иммунохимическом анализе. Предпочтительно антитела, которые связываются специфически с TF или комплексом FVIIa/TF, не детектируют другие белки в иммунохимических анализах и могут образовывать иммунопреципитат TF или комплекса FVIIa/TF из раствора.
TF или комплекс FVIIa/TF могут быть использованы для иммунизации млекопитающего, такого как мышь, крыса, кролик, морская свинка, обезьяна или человек, чтобы продуцировать поликлональные антитела. Если требуется, TF или комплекс FVIIa/TF могут быть конъюгированы с белком-носителем, таким как бычий сывороточный альбумин, тироглобулин и гемоцианин из Megathura crenulata. В зависимости от вида хозяина могут применяться различные адъюванты, чтобы усилить иммунологический ответ. Такие адъюванты включают, но без ограничения, адъювант Фрейнда, минеральные гели (например, гидроксид алюминия) и поверхностно-активные вещества (например, лизолецитин, плюроновые полиолы, полианионы, пептиды, масляные эмульсии, гемоцианин из Megathura crenulata и динитро фенол). Среди адъювантов, применяемых у человека, особенно полезными являются BCG (bacilli Calmette-Guerin) и Cornybacteriuparvum.
Моноклональные антитела, которые специфически связываются с TF или комплексом FVIIa/TF, могут быть получены с использованием любой методики, которая обеспечивает продукцию молекул антител стабильными клеточными линиями в культуре. Эти методики включают, но без ограничения, гибридомную технику, технику человеческой В-клеточной гибридомы и технику гибридомы вируса Эпштейна-Барра (EBV) (Kohler и др., Nature 256, 1987, с. 495-497; Kozbor и др., J. Immunol. Methods 81, 1985, с. 31-42; Cote и др., Proc. Natl. Acad. Sci. USA 80, 1983, с. 2026-2030; и Cote и др., Mol. Cell Biol. 62, 1984, с. 109-120).
Кроме того, могут быть использованы методики, созданные для продукции «химерных антител», сплайсинг генов мышиного и человеческого антител, чтобы получить молекулу с соответствующей антигенной специфичностью и биологической активностью (Morrison и др., Proc.Natl. Acad. Sci. USA 81, 1984, с. 6851-6855; Neuberger и др., Nature 312, 1984, с. 604-608; Takeda и др., Nature 314, 1985, с. 452-454). Моноклональные и другие антитела могут быть также «гуманизированы», чтобы предотвратить у пациента формирование иммунного ответа к антителам, когда оно применяется в терапевтических целях. Такие антитела обладают достаточным сходством с последовательностью человеческого антитела, чтобы непосредственно использоваться в слитом белке, или могут потребовать изменения нескольких ключевых остатков. Различия в последовательностях между антителами грызунов и человеческими последовательностями может быть сведено к минимуму путем замены остатков, которые отличаются от таковых в человеческих последовательностях, с помощью сайт-направленного мутагенеза отдельных остатков или с помощью трансплантации полностью комплементарных детерминирующих участков. Альтернативно гуманизированные антитела могут быть продуцированы с использованием рекомбинантных способов, как описано в патенте GB 2188638В. Антитела, которые связываются специфично с TF или комплексом FVIIa/TF, могут содержать антиген-связывающие сайты, которые или частично, или полностью гуманизированы, как раскрывается в патенте US 5565332.
Альтернативно методики, описанные для продуцирования одноцепочечных антител, могут быть адаптированы, используя способы, известные специалистам, для продуцирования одноцепочечных антител, которые специфично связываются с TF или комплексом FVIIa/TF. Антитела с родственной специфичностью, но отличающегося идиотипического состава, могут быть получены путем перестановки цепи из случайных комбинаторных библиотек Ig (Burton, Proc. Natl. Acad. Sci. USA 88, 1991, с. 11120-11123).
Одноцепочечные антитела могут быть также сконструированы с применением способа амплификации ДНК, такого как полимеразная цепная реакция, используя гибридомную кДНК в качестве матрицы (Thirion и др., Eur. J. Cancer Prev.5, 1996, с. 507-511). Одноцепочечные антитела могут быть моно- или биспецифичными и могут быть бивалентными или тетравалентными. Конструкция тетравалентных биспецифичных одноцепочечных антител сообщена, например, Coloma Morrison Natl. Biotechnol. 15, 1997, с. 159-163. Конструкция бивалентных биспецифичных одноцепочечных антител сообщена Mallendar и Voss J. Biol. Chem. 269, 1994, с. 199-216.
Нуклеотидная последовательность, кодирующая одноцепочечное антитело, может быть построена с применением ручного или автоматизированного синтеза нуклеотидов, клонирована в конструкцию экспрессии с использованием стандартных способов рекомбинантной ДНК и введена в клетку, чтобы экспрессировать кодирующую последовательность. Альтернативно одноцепочечные антитела могут быть продуцированы непосредственно с использованием, например, техники отображения нитевидного фага (Verhaar и др., Int. J. Cancer, 61, 1996, с. 497-501; и Nicholls и др., J. Immunol. Meth. 165, 1993, c. 81-91).
Антитела, которые связываются специфично с TF или комплексом FVIIa/TF, могут быть также получены путем индуцирования in vivo продукции в популяции лимфоцитов или путем скрининга библиотек Ig или панелей реагентов с высокой специфичностью связывания, как раскрывается в литературе (Orlandi и др., Proc. Natl. Acad. Sci. USA 86:3, 1989, с. 833-3837; Winter и др, Nature 349,1991, с. 293-299).
В другом варианте воплощения по данному изобретению белок, обеспечивающий направленный перенос, является частью, обеспечивающей направленный перенос, иного, чем антитело, белка, который может связывать и нейтрализовать TF. Двумя такими примерами являются ингибированный по активному центру фактор FVIIa (FVIIai) и ингибитор метаболизма тканевого фактора (TFPI).
Как FVIIa, так и FVIlai с высоким сродством образуют комплекс с TF (В.В. Sorenson, В.В и L.V. Rao, Blood Coagul. Fibrinolysis 9 (доп.1), 1998, с. 67-71). Фактор FVIIai является нейтрализующим TF антикоагулянтом, который действует, конкурируя с эндогенным FVIIa за связывание с экспонированным TF. Фактор FVIIai подавляет способность протеолитически активного FVIIa образовывать компетентный комплекс FVIIa-TF и таким способом подавляет инициацию коагуляции. Путем генетического слияния доменов ТМ в FVIIai тромбомодулин мог быть нацелен на богатые TF протромбозные поверхности.
кДНК, кодирующая человеческий FVII, была выделена и секвенирована (H.S. Hagen и др., Proc. Natl. Acad. Sci. USA 83 (8), 1986, с. 2412-2416, ссылка включена в контекст путем цитирования). Человеческий FVIIa, кодируемый кДНК, может быть получен по стандартным методикам рекомбинантной ДНК, исходя из мРНК, выделенной из печени человека. Фактор FVIIai может быть получен с помощью изменения серина в активном центре по стандартным методикам рекомбинантной ДНК или химической обработкой каталитически активного FVIIa пептидилхлорметилкетоном, который необратимо модифицирует и ингибирует активный центр.
TFPI нацелен на комплекс FVIIa/TF и ингибирует его FXa-зависимым образом (I. Salemink и др., J. Biol. Chem. 274 (40), 1999, с. 28225-28232). TFPI сначала связывается с FXa, и затем комплекс TFPI-FXa связывается с комплексом FVIIa/TF и ингибирует его. При генетическом слиянии доменов ТМ в TFPI тромбомодулин (ТМ) мог быть нацелен на TF-богатые протромбозные поверхности.
кДНК, кодирующая человеческий TFPI, была выделена и секвенирована (Т.С. Wun и др., J. Biol. Chem. 263 (13), 1988, с. 6001-6004, ссылка включена в контекст цитированием). Человеческий TFPI, кодируемый кДНК, может быть получен с помощью стандартных методик рекомбинантной ДНК, исходя из мРНК, выделенной из печени человека.
Белок, обеспечивающий направленный перенос, по данному изобретению (т.е. антитело или другие относящиеся к делу белки) может быть экспрессирован и очищен способами, хорошо известными специалистам. Например, антитела и белки могут быть очищены аффинным способом путем пропускания через колонку, в которой TF находится в связанном состоянии. Связанные антитела или белки могут затем элюироваться из колонки с использованием буфера с высокой концентрацией соли.
В одном предпочтительном варианте воплощения по данному изобретению белком, обеспечивающим направленный перенос, является TF-связывающее антитело scFv, которое подавляет активацию FX с помощью комплекса FVIIa/TF и не конкурирует со связыванием FVIIa. Для того чтобы получить TF-связывающее антитело scFv, библиотека человеческих антител HuPhaBL3 на нитевидном фаге была отобрана с помощью иммобилизованного растворимого TF. Антитела TF-презентирующего фага были сверхэкспрессированы в Е.coli и аффинно очищены с использованием колонки с последовательностью e-tag. Очищенные антитела были далее охарактеризованы, используя BIAcore, анализ sTF-зависимого фактора Vila (анализ sTF/FVIIa), анализ активации FX и анализ протромбинового времени (ПВ). Последовательность TF-связывающего антитела scFv, обозначенного scFv(TF)3e10, показана в примере 1 и соответствует SEQ ID No:1. Выделение, продукция и характеристика TF-связывающего антитела scFv описаны более подробно ниже.
Тромбомодулин
Тромбомодулиновая (ТМ) доменная часть слитого белка действует как кофактор катализируемой тромбином активации белка С, который, в свою очередь, подвергает деградации факторы Va и VIIa, предотвращая тем самым дальнейшее тромбообразование. Домены ТМ включают, например, домен N-концевого гидрофобного участка, домен EGF123, междоменную петлю между EGF3 и EGF4, домен EGF456 и домен O-гликозилированного Ser/Thr-богатого участка. Домен EGF456, в частности, опосредует связывание тромбина и активацию белка С (S. Kurosawa и др. (1988), выше; и М. Zushi и др. (1989), выше). В предпочтительных воплощениях по данному изобретению тромбомодулиновая доменная часть слитого белка включает домен EGF456, один или в комбинации с одним доменом или несколькими другими доменами ТМ. В еще более предпочтительных вариантах воплощения по данному изобретению домен EGF456 содержит точечные мутации, которые делают белок более устойчивым к окислительному повреждению и действию протеаз и/или увеличивают каталитическую эффективность.
Полная протяженность последовательности ДНК, кодирующей человеческий ТМ, способствует получению генов и используется как исходная точка для создания последовательностей ДНК, кодирующих пептиды ТМ и слитые белки, содержащие ТМ и фрагменты/пептиды ТМ.
Ген полной длины ТМ может быть получен несколькими способами. Человеческие геномные библиотеки коммерчески доступны. Олигонуклеотидные зонды, специфичные для этих генов, могут быть синтезированы с использованием опубликованной генной последовательности. Известны способы скрининга геномных библиотек с олигонуклеотидными зондами. Опубликованные генные последовательности ТМ демонстрируют, что внутри кодирующего участка нет интронов. Таким образом, геномный клон обеспечивает необходимый исходный материал, чтобы сконструировать плазмиду экспрессии для ТМ, используя известные способы.
Кодирующий ТМ фрагмент ДНК может быть возвращен в прежнее состояние путем использования сайтов эндонуклеаз рестрикции, которые были идентифицированы в участках, расположенных сбоку или являющихся внутренними по отношению к гену (R.W.Jackman и др. (1987), выше). Альтернативно гены полной длины могут также быть получены из банка кДНК. Например, матричная РНК, полученная из клеток эндотелия, обеспечивает соответствующий исходный материал для получения кДНК. Способы создания банков кДНК хорошо известны (смотри, например, J.F.Sambrook и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (1989), ссылка включена в контекст путем цитирования).
Слитый белок
Антикоагулирующий слитый белок по данному изобретению включает белок, который обеспечивает направленный перенос и связывается или с TF, или с комплексом FVIIa/TF и функционально связан с доменом EGF456 тромбомодулина (ТМ), одним или в комбинации по меньшей мере с другим доменом ТМ, выбранным из группы, состоящей из домена N-концевого гидрофобного участка, домена EGF123, междоменной петли между EGF3 и EGF4 и O-гликозилированного Ser/Thr-богатого домена или их аналогов, фрагментов, производных или вариантов. Слитый белок может включать белок, обеспечивающий направленный перенос, связанный с доменами ТМ в любой комбинации.
В одном особенно предпочтительном варианте воплощения слитый белок включает антитело, которое связывает TF, функционально связанный с тромбомодулиновым доменом EGF456 и междоменной петлей между EGF3 и EGF4 (ТМ1456) или их аналогами, фрагментами, производными или вариантами.
Слитый белок по настоящему изобретению включает, но без ограничения, конструкции, в которых С-концевая часть одноцепочечного антитела соединяется с N-концевой частью аналога, фрагмента, производного или варианта домена ТМ, С-концевая часть антитела IgG соединяется с N-концевой частью аналога, фрагмента, производного или варианта домена ТМ, С-концевая часть антитела Fab соединяется с N-концевой частью аналога, фрагмента, производного или варианта домена ТМ, N-концевая часть одноцепочечного антитела соединяется с С-концевой частью аналога, фрагмента, производного или варианта домена ТМ, N-концевая часть антитела IgG соединяется с С-концевой частью аналога, фрагмента, производного или варианта домена ТМ, N-концевая часть антитела Fab соединяется с С-концевой частью аналога, фрагмента, производного или варианта домена ТМ, более чем одно одноцепочечное антитело соединяется как с N-концевой, так и с С-концевой частями аналога, фрагмента, производного или варианта домена ТМ, более чем одно антитело IgG соединяется как с N-концевой, так и с С-концевой частями аналога, фрагмента, производного или варианта домена ТМ, более чем одно антитело Fab соединяется как с N-концевой, так и с С-концевой частями аналога, фрагмента, производного или варианта домена ТМ, более чем один аналог, фрагмент, производное или вариант домена ТМ соединяется как с N-концевой, так и с С-концевой частями одноцепочечного антитела, более чем один аналог, фрагмент, производное или вариант домена ТМ соединяется как с N-концевой, так и с С-концевой частями антитела IgG, более чем один аналог, фрагмент, производное или вариант домена ТМ соединяется как с N-концевой, так и с С-концевой частями антитела Fab, один или более чем один аналог, фрагмент, производное или вариант домена ТМ соединяется как с N-концевой, так и с С-концевой частями димерного одноцепочечного антитела.
Слитые белки по настоящему изобретению включают слитые белки примера 2 (SEQ ID No:2) и 3 (SEQ ID No:3), а также те слитые белки, которые имеют несущественные отклонения в последовательности от них. Термин «несущественное отклонение» включает любую последовательность, замещение или делеционный вариант, которые сохраняют в значительной степени по меньшей мере одну биологическую функцию полипептидов по данному изобретению, предпочтительно кофакторную активность для опосредованной тромбином активации белка С. Эти функциональные эквиваленты могут предпочтительно включать слитые белки, которые имеют по меньшей мере примерно 90% идентичность с слитыми белками SEQ ID Nos:2 или 3 и более предпочтительно - по меньшей мере 95% идентичность со слитыми белками SEQ ID Nos:2 или 3 и еще более предпочтительно - по меньшей мере 97% идентичность со слитыми белками SEQ ID Nos:2 или 3, и также включают части таких слитых белков, имеющих в значительной мере такую же биологическую активность. Однако любой слитый белок, имеющий несущественное отклонение в аминокислотной последовательности от слитых белков SEQ ID Nos:2 или 3, который проявляет функциональную эквивалентность, как описано далее в контексте, включен в описание по настоящему изобретению.
В другом варианте воплощения изобретения слитый белок включает антитело, которое связывает TF, функционально связанный с тромбомодулиновым (ТМ) доменом EGF3, который требуется для активации ингибитора активируемого тромбином фибринолиза (TAFI).
Аналоги, фрагменты, производные и варианты
Аналог, фрагмент, производное или вариант слитых белков, а также белки, обеспечивающие направленный перенос, и домены ТМ по настоящему изобретению могут быть: (i) одним, в котором один или несколько аминокислотных остатков замещены консервативным или неконсервативным аминокислотным остатком (предпочтительно консервативным аминокислотным остатком), и такой замещенный аминокислотный остаток может или не может быть остатком, кодированным с помощью генетического кода; или (ii) одним, в котором один или несколько аминокислотных остатков включают заместитель; или (iii) одним, в котором зрелый слитый белок соединяется с другим соединением, таким как соединения для увеличения времени полувыведения слитого белка (например, полиэтиленгликоль); или (iv) один, в котором дополнительные аминокислоты соединены со зрелым слитым белком, таким как лидерная или секреторная последовательность, или последовательность, которая используется для очистки зрелого слитого белка; или (v) одним, в котором полипептидная последовательность соединена с более длинным полипептидом, т.е. человеческим альбумином, антителом или белком Fc, для увеличения продолжительности эффекта. Считается, что такие аналоги, фрагменты, производные и варианты доступны специалисту в данной области из сообщений в контексте.
Предпочтительно производные по настоящему изобретению будут содержать консервативные аминокислотные замещения (определенные ниже), сделанные в одном или нескольких прогнозируемых остатках, предпочтительно в остатках заменимых аминокислот. Термин «заменимый» аминокислотный остаток означает остаток, который может меняться в белковой последовательности дикого типа без изменения биологической активности, тогда как «незаменимый» аминокислотный остаток необходим для биологической активности. Термин «консервативное аминокислотное замещение» означает замещение, при котором аминокислотный остаток заменяется аминокислотным остатком, имеющим подобную боковую цепь. Семейства аминокислотных остатков, имеющих подобные боковые цепи, определены в данной области. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), β-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Неконсервативные замещения не проводятся для консервативных аминокислотных остатков или аминокислотных остатков, располагающихся внутри консервативного белкового домена, если неконсервативные замещения не проводятся, чтобы сделать образующийся слитый белок более устойчивым к окислительному повреждению и действию протеаз и/или повысить его каталитическую эффективность. Фрагменты биологически активных частей включают полипептидные фрагменты, подходящие для применения в качестве лекарственного средства, в качестве реагента для исследований и тому подобного. Фрагменты включают пептиды, содержащие аминокислотные последовательности, подобные аминокислотным последовательностям или происходящие от аминокислотных последовательностей слитого белка по данному изобретению и по меньшей мере обладающие активностью того полипептида, но которые включают меньше аминокислот, чем полипептид полной протяженности, раскрытый в контексте. Обычно биологически активные части включают домен или мотив по меньшей мере с активностью полипептида. Биологически активная часть полипептида может быть пептидом, т.е., например, длиной в 5 или более аминокислот. Такие биологически активные части могут быть получены синтетически или с помощью рекомбинантных методик и могут быть оценены на одну или несколько функциональных активностей полипептида по данному изобретению способами, раскрытыми в контексте и/или хорошо известными специалистам.
Более того, предпочтительные производные по настоящему изобретению включают зрелые слитые белки, которые соединены с другим соединением, таким как соединение для увеличения времени полувыведения полипептида и/или для снижения потенциала иммуногенности полипептида (например, полиэтиленгликоль, PEG). Полиэтиленгликоль может применяться, чтобы придать слитому белку растворимость в воде, габариты, малую скорость почечного клиренса и сниженную иммуногенность. Смотри, например, патент US 6214966. В случае модификации полиэтиленгликолем соединение слитого белка с PEG может быть выполнено любыми способами, известными специалистам в этой области. Например, модификация полиэтиленгликолем может быть осуществлена сначала введением цистеиновой мутации в слитый белок с последующим получением производного с PEG-малеимидом. Цистеин может быть добавлен к С-концу пептидов. Смотри, например, Tsutsumi и др., Proc. Natl. Acad. Sci. USA, 97 (15), 2000, с. 8548-8553. Другая модификация, которая может быть проведена со слитым белком, включает биотинилирование. В некоторых случаях может быть полезным иметь биотинилированный слитый белок, чтобы он мог легко реагировать со стрептавидином. Способы биотинилирования белков хорошо известны специалистам. Кроме того, со слитым белком может быть связан сульфат хондроэтина.
Варианты слитых белков, белков, обеспечивающих направленный перенос, и доменов ТМ по изобретению включают полипептиды, имеющие аминокислотную последовательность, в значительной мере подобную аминокислотной последовательности исходных слитых белков, белков, обеспечивающих направленный перенос, и доменов ТМ. Термин «подобные» означает первую аминокислотную последовательность, которая содержит достаточное или минимальное число идентичных или эквивалентных аминокислотных остатков относительно второй аминокислотной последовательности, так что первая и вторая аминокислотные последовательности имеют общий структурный домен и/или общую функциональную активность. Например, аминокислотные последовательности, которые содержат общий структурный домен, который идентичен по меньшей мере примерно на 45%, предпочтительно примерно на 75-98%, определяются в контексте как идентичные. Предпочтительно варианты будут подобны аминокислотной последовательности преимущественных слитых белков по данному изобретению. Варианты включают варианты слитых белков, кодированных полинуклеотидом, который гибридизуется с полинуклеотидом по данному изобретению или комплементарен ему в жестких условиях. Такие варианты, как правило, сохраняют функциональную активность слитых белков по данному изобретению. Библиотеки фрагментов полинуклеотидов могут быть использованы, чтобы получать разнообразную популяцию фрагментов для скрининга и последующей селекции. Например, библиотека фрагментов может быть получена обработкой с нуклеазой полученного ПЦР двухцепочечного фрагмента полинуклеотида в условиях, где одноцепочечный разрыв происходит только примерно один раз на молекулу, денатурацией двухцепочечной ДНК, ренатурацией ДНК, чтобы образовать двухцепочечную ДНК, которая может включать смысловые/антисмысловые пары от различных продуктов с одноцепочечным разрывом, удалением одноцепочечных частей от реформированных дуплексов путем обработки с нуклеазой S1 и лигированием образующейся библиотеки фрагментов в вектор экспрессии. Этим способом можно получить библиотеку экспрессии, которая кодирует N-концевой и внутренний фрагменты слитых белков различной величины по данному изобретению.
Варианты включают слитые белки, а также белки, обеспечивающие направленный перенос, и домен ТМ, которые отличаются по аминокислотной последовательности из-за мутагенеза. Варианты, которые обладают кофакторной активностью для опосредованной тромбином активации белка С, могут быть идентифицированы с помощью скрининговых комбинаторных библиотек мутантов, например отсечение или точечные мутанты слитых белков или доменов ТМ по данному изобретению, и использованием анализа активации белка С, описанного в примере 5. Варианты, которые обладают активностью связывания TF или комплекса FVIIa/TF, могут быть идентифицированы с помощью скрининговых комбинаторных библиотек мутантов, например расщепление или точечные мутанты слитых белков или белков, обеспечивающих направленный перенос, по данному изобретению с использованием анализа sTF/FVIIa или анализы по активации FX примера 5, описанные в примере 5. Кроме того, биоэквивалентные аналоги слитых белков могут также быть образованы путем проведения различных замещений по остаткам или последовательностям в доменной части ТМ слитого белка, которые могут сделать слитый белок более устойчивым к окислительному повреждению или действию протеазы, смотри, например, патент US 5827824, или увеличить каталитическую эффективность слитого белка, смотри, например, М. Adier и др., J. Biol. Chem. 270 (40), 1995, с. 23366-23372 и заявку WO 01/98352, опубликованную 27.12.2001, все полностью включены в контекст путем цитирования.
В одном варианте воплощения изобретения разнообразная библиотека вариантов генерируется путем комбинаторного мутагенеза на уровне нуклеиновых кислот и кодируется разнообразной генной библиотекой. Разнообразная библиотека вариантов может быть получена, например, энзиматическим лигированием смеси синтетических олигонуклеотидов в генные последовательности таким образом, что вырожденный набор потенциальных вариантных аминокислотных последовательностей экспрессируется в виде индивидуальных полипептидов, или альтернативно в виде набора слитых белков большей протяженности (например, для развертки фага), содержащих набор их последовательностей. Существует множество способов, которые могут применяться для получения библиотек потенциальных вариантов их вырожденной олигонуклеотидной последовательности. Химический синтез вырожденной генной последовательности может быть осуществлен в автоматическом синтезаторе ДНК, и затем синтетический ген встраивают в соответствующий вектор экспрессии. Применение вырожденного набора генов допускает обеспечение в одной смеси всех последовательностей, кодирующих требуемый набор потенциальных вариантных последовательностей. Способы синтеза вырожденных олигонуклеотидов известны специалистам (смотри, например, Narang, Tetrahedron 39, 1983, с.3; Itakura и др., Annu. Rev.Biochem. 53, 1984а, с.323; Itakura и др.. Science 198, 1984b, с.1056; Ike и др., Nucleic Acid Res. 11, 1983 с.47).
Специалистам известны несколько методик скрининга генных продуктов комбинаторных библиотек, полученных с помощью точечных мутаций или расщепления, и скрининга библиотек кДНК для генных продуктов, имеющих выбранное свойство. Такие методики адаптируются для быстрого скрининга генных библиотек, полученных путем комбинаторного мутагенеза слитых белков, а также белков, обеспечивающих направленный перенос, и доменов ТМ, на кофакторную активность опосредованной тромбином активации белка С или активности связывания TF или комплекса FVIIa/TF. Наиболее широко применяемые методики, которые подлежат анализу с высокой пропускной способностью для скрининга больших генных библиотек, обычно включают клонирование генной библиотеки в реплицируемые векторы экспрессии, трансформацию соответствующих клеток с образующейся библиотекой векторов и экспрессию комбинаторных генов в условиях, при которых детектирование требуемой активности благоприятствует выделению вектора, кодирующего ген, чей продукт был детектирован. Рекуррентный групповой мутагенез (REM), методика, которая повышает частоту функциональных мутантов в библиотеках, может быть применен в комбинации со скрининговыми анализами, чтобы идентифицировать требуемые варианты.
Получение слитых белков
Слитый белок по изобретению получают соединением белка, обеспечивающего направленный перенос, или иначе - связыванием его с доменами ТМ или их аналогами, фрагментами, производными или вариантами любым способом, известным специалистам в этой области. Два компонента могут быть химически связаны вместе с помощью множества хорошо известных химических методик. Например, связь может быть образована с помощью гетеробифункциональных мостиковых линкеров, например, N-сукцинимидил-3-(2-пиридилдитио)пропионата (SPDP), карбодиимида, глутарового альдегида и тому подобного.
В более предпочтительном варианте воплощения белок, обеспечивающий направленный перенос, по данному изобретению может быть соединен с доменами ТМ рекомбинантными способами, такими как использование методик рекомбинантной ДНК для получения нуклеиновой кислоты, которая кодирует как белок, обеспечивающий направленный перенос, так и полипептид, кодирующий домены ТМ, и экспрессия последовательности ДНК в клетке-хозяине, такой как Е.coli или клетке млекопитающего. ДНК, кодирующая слитый белок, может быть клонирована в кДНК или в геномной форме с помощью любой методики клонирования, известной специалистам в этой области. Смотри, например, J.F. Sambrook и др. (1989), выше.
В случае, когда белок, обеспечивающий направленный перенос, является антителом, если была идентифицирована последовательность ДНК, которая кодирует участок Fv, который при экспрессии проявляет специфическую связывающую активность, слитые белки, включающие тот участок Fv, могут быть получены способами, известными специалисту. Так, например, V.K. Chaudhary и др., Nature 339 (6223), 1989, с. 394-397; J.K. Batra и др., J. Biol. Chem. 265 (25), 1990, с. 15198-15202; J.K. Batra и др., Proc. Natl. Acad. Sci. USA 86 (21), 1989, с. 8545-8549; V.K. Chaudhary и др., Proc. Natl. Acad. Sci. USA 87 (3) 1990, с. 1066-1070, все публикации, включенные путем цитирования, описывают получение слитых белков на основе различных одноцепочечных антител. Участок Fv может быть соединен непосредственно с доменами ТМ или может быть связан посредством линкерной последовательности. Линкерная последовательность может присутствовать просто для того, чтобы обеспечить пространство между частью, обеспечивающей направленный перенос, и доменами ТМ или содействовать мобильности между этими участками, чтобы дать возможность каждому из них достигнуть своей оптимальной конформации. Последовательность ДНК, включающая соединяющий фрагмент, может также обеспечить последовательности (такие как праймерные или рестрикционные сайты) для содействия клонированию или может сохранять рамку считывания между последовательностью, кодирующей часть, обеспечивающую направленный перенос, и последовательностью, кодирующей домены ТМ. Создание таких связующих пептидов хорошо известно специалистам в данной области.
По настоящему изобретению линкерные последовательности могут применяться для связывания белка, обеспечивающего направленный перенос, с доменами ТМ. В одном предпочтительном варианте воплощения по настоящему изобретению две линкерные последовательности применяются в создании конструкции слитого белка, состоящего из одноцепочечного антитела и тромбомодулинового (ТМ) домена EGF456 и междоменной петли между EGF3 и EGF4 (TMi456). Первая связывает тяжелый и легкий домены одноцепочечного антитела. Первая линкерная последовательность имеет протяженность из 5 аминокислот. Будет очевидным, что могут применяться и другие короткие линкерные последовательности, от 0 до 10 аминокислот. Второй линкер по настоящему изобретению является линкером из 15 аминокислот, который связывает антитело с доменами ТМ. Будет очевидным для специалистов в данной области, что много различных линкерных последовательностей может использоваться и по-прежнему приводить к слитому белку, который сохраняет антикоагулирующую активность и способность к активации белка С. Модификации существующего линкера будут ставить своей целью самое большое усиление активации белка С на содержащих TF фосфолипидных поверхностях.
В предпочтительном подходе одноцепочечное антитело получали, используя фаговую дисплейную библиотеку. На первом этапе конструирования фаговой дисплейной библиотеки гены (VH (из IgM), VK и VL) были с помощью ПЦР клонированы от объединенной мРНК из человеческого костного мозга, лимфатического узла и селезенки, используя набор семейства специфических праймеров. Образовавшиеся библиотеки pCITE-VH (3.8×109 членов), pZ604-VK(1.6×107) и pZ604-VL (3.2×107) представляют перманентное и большое разнообразие генов V. Гены VH были амплифицированы с помощью библиотеки pCITE-VH. Гены VK и VL были амплифицированы с помощью ПЦР от библиотеки pZ604-VK и pZ604-VL с обратной JH и линкерной последовательностью на 5'-конце. Очищенные в геле VH, VK и VL, содержащие продукты ПЦР, были затем подвергнуты совместному сплайсингу, чтобы получить набор генов scFv. Набор генов scFv клонировали в фагмидный вектор pZ603 и продукт лигирования электропорировали в компетентные клетки Е.coli TG1, чтобы получать фаговую дисплейную библиотеку scFV, HuPabL3, с 5,2×109 индивидуальных трансформантов (В.К.Кау и др., Phage Display of Peptides and Proteins: A Laboratory Manual, Academic Press, San Diego, CA, 1996; J.D. Marks и др., J. Mol. Biol. 222 (3), 1991, с. 581-597; M.D. Sheets и др., Proc. Natl. Acad. Sci. USA 95 (11), 1998, с. 6157-6162).
В предпочтительном варианте воплощения по настоящему изобретению получали одноцепочечное антитело (scFv(TF)3e10), которое имело один связывающий сайт VH/VL для TF. Аминокислотная последовательность scFv(TF)3e10 (SEQ ID No:1) изображена в примере 1.
В предпочтительном варианте воплощения по настоящему изобретению фрагмент ПЦР, содержащий последовательность TMi456 (с мутациями M388L и H381G), фланкированную с помощью сайтов Notl, субклонировали в сайт Notl вектора pZ612/3e10 (бактериальный вектор экспрессии для scFv(TF)3e10, основанный на pCANTAB5 от Pharmacia). В контексте точечные мутации в частях ТМ слитых белков по изобретению обозначаются однобуквенным символом аминокислотного остатка нативного ТМ, сопровождающимся номером положения аминокислотного остатка в зрелом ТМ и однобуквенным символом аминокислотной мутации. Например, M388L указывает на то, что метионин в положении 3888 аминокислоты зрелого ТМ заменен лейцином. Сайт Notl расположен между последовательностью антитела и последовательностью e-tag. Это позволяет получить бактериальную конструкцию экспрессии (pKM101) для слитого белка, состоящую из scFv(TF)3e10 - линкера из 15 аминокислот - TMi456 и последовательность e-tag. Чтобы получить вектор экспрессии для млекопитающего, фрагмент ПЦР сначала получали на матрице pKM101. Этот фрагмент был создан для встраивания в сайты Stul/Msd тромбомодулинового вектора экспрессии pTHR525. Это позволило получить вектор (рКМ113), который имел сигнальную последовательность Solulin, последовательность зрелого слитого белка, последовательность e-tag. Вектор содержит ген устойчивости к ампициллину и селектируемые маркеры гигромицина и дигидрофолатредуктазы (DHFR). Экспрессия запускается промотором MPSV LTR. Сайт-специфический мутагенез осуществляли на данном векторе, чтобы включить мутации R456G и H457Q, которые придают части ТМ устойчивость к протеазе. Образующийся вектор обозначали как рКМ115. Вектор рКМ115 содержал линкер из 15 аминокислот, разделяющий домены VH и VL, и другой линкер из 15 аминокислот, отделяющий домен VL от TMi456. Линкер, разделяющий VH и VL, был уменьшен до 5 аминокислот, чтобы провести образование димера более высокой авидности, обозначенного как рНМ115,5. Слитый белок, кодированный рНМ115,5, scFv(TF)3e10-TMi456 (SEQ ID No:2), изображен в примере 2. Дополнительный вектор рКМ125 генерировали, используя стандартные методики рекомбинантной ДНК, путем удаления 3 аминокислот (GAP) между линкером из 5 аминокислот, разделяющим домены VHи VL, и удаления e-tag на С-конце слитого белка. Образующийся слитый белок scFv(TF)3e10-TMi456A (SEQ ID No:3) изображен в примере 3.
Экспрессия и очистка слитых белков
Существует несколько путей, чтобы экспрессировать рекомбинантные слитые белки in vitro, включая Е.coli, бакуловирусы, клетки дрожжей и млекопитающих или другие системы экспрессии. Способы экспрессии клонированных генов в бактериях хорошо известны. Чтобы получить высокий уровень экспрессии клонированного гена в прокариотической системе, необходимо создать векторы экспрессии, которые содержат как минимум сильный промотор, чтобы направлять терминацию транскрипции мРНК. Примерами регуляторных участков, подходящих для этой цели, являются промоторный и операторный участок гена β-глюкозидазы Е.coli, биосинтетический путь триптофана Е.coli или левый промотор фага К. Полезным является включение селектируемых маркеров в векторы ДНК, трансформирующие в Е.coli. Примеры таких маркеров включают гены, определяющие устойчивость к ампициллину, тетрациклину или хлорамфениколу.
Существуют многочисленные клеточные системы, которые могут быть использованы для обора эукариотических клеточных систем, наиболее пригодных для экспрессии слитых белков и их аналогов. Иллюстративные примеры линий клеток млекопитающих включают, но без ограничения, клетки RPMI 7932, VERO и HeLa, линии клеток яичника китайского хомяка (СНО), линии клеток W138, ВНК, COS-7, С 127 или MDCK. Предпочтительной линией клеток млекопитающих является CHL-1. При использовании клеток CHL-1 гигромицин включают в качестве эукариотического селектируемого маркера. Клетки CHL-1 происходят от клеток меланомы RPMI 7032, легко доступной линии человеческих клеток. Линия клеток CHL-1 была заложена на хранение Американской коллекцией тканевых культур (АТСС) в соответствии с будапештским договором и была обозначена #CRL 9446, депонирована 18.06.1987. Клетки, подходящие для применения по данному изобретению, коммерчески доступны от АТСС. Иллюстративные линии клеток включают Spodoptera frugiperda и Bombix mori.
Прокариотическая система Е.coli не способна осуществлять посттрансляционную модификацию, такую как гликозилирование. Кроме того, часто нарушается укладка белков со сложными дисульфидными структурами при экспрессии в Е.coli. Для слитого белка, описанного в контексте, наблюдалось заметное снижение тромбомодулиновой кофакторной активности при экспрессии в Е.coli, хотя обе активности еще присутствовали. В случае прокариотической системы экспрессированный белок или присутствует в цитоплазме клетки в нерастворимой форме, в виде так называемых телец включения, найденных в растворимой фракции после лизиса клетки, или направляется в периплазму при добавлении соответствующей секреторной сигнальной последовательности. Если экспрессированный белок находится в виде телец включения, обычно требуются растворение и последующая повторная укладка телец включения.
Специалистам известно много прокариотических векторов экспрессии, таких как рКК.223-3 (Pharmacia Fine Chemicals, Uppsala, Sweden), pKK.233-2 (Clontech, Palo Alto, CA, USA) и pGEMl (Promega Biotech, Madison, WI, USA), которые коммерчески доступны.
Промоторы, обычно применяемые в рекомбинантных микробных системах экспрессии, включают р-лактамазу (пенициллиназу) и систему лактозного промотора (F.C. Chang и др., Nature 275 (5681), 1978, с. 617-624; D.V. Goeddel и др., Nature 281 (5732), 1979, с. 544-548), промоторную систему триптофана (trp) (D.V. Goeddel и др., Nucl. Acids Res. 8 (18), 1980, с. 4057-4074) и промотор tac (Т. Maniatis и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (1982)). Другая подходящая бактериальная система экспрессии использует промотор pL фага К и термоиндуцируемый репрессор clts857 (H.U. Bernard и др., Gene 5 (1), 1979, с. 59-76; С.А. Love и др., Gene 176 (1-2), 1996, с. 49-53). Рекомбинантные слитые белки могут быть также экспрессированы в дрожжах-хозяевах, таких как Saccharomyces cerevisiae. Обычно это дает возможность проводить различные посттрансляционные модификации. Экспрессированный слитый белок может быть секретирован в культуральный супернатант, где остается немного других белков, что облегчает очистку. Дрожжевые векторы для экспрессии слитых белков по данному изобретению сохраняют некоторые необходимые свойства. Элементы вектора обычно получают из дрожжей и бактерий, чтобы сделать возможным воспроизведение плазмиды в обеих системах. Бактериальные элементы включают источник репликации и селектируемый маркер. Дрожжевые элементы включают источник репликационной последовательности (ARS), селектируемый маркер, промотор и терминатор транскрипции.
Соответствующие промоторы в дрожжевых векторах для экспрессии включают промоторы гена TRP1, ADH1 или гена ADHII, ген кислой фосфатазы (РН03 или РН05), ген изоцитохрома или промоторы, включенные в гликолитический путь обмена, такие как промотор енолазы, глицеральдегид-3-фосфатдегидрогеназы (GADPH), 3-фосфоглицераткиназы (PGK), гексокиназы, пируваткиназы, триозафосфатизомеразы и фосфоглюкозаизомеразы (R.A. Hitzeman и др., J. Biol. Chem. 255 (24), 1980, 12073-12080; В. Hess и др., J. Adv.Enzyme Reg. 7,1968, с. 149-167; и M.J. Holland и J. P. Holland., Biochemistry, 17 (23), 1978, с. 4900-4907).
Коммерчески доступные дрожжевые векторы включают pYES2, pPIC9 (Invitrogen, San Diego, CA), Yepc-pADH2a, pYcDE-1 (Washington Research, Seattle, WA), pBC102-K22 (ATCC # 67255) и YpGX265GAL4 (ATCC # 67233). Линии клеток млекопитающих, включающие, но без ограничения, COS-7, клетки L, C127, ЗТЗ, клетки яичника китайского хомяка (СНО), HeLa, BHK, CHL-1, NSO и НЕК293, могут быть использованы для экспрессии рекомбинантных слитых белков по данному изобретению. Рекомбинантные белки, продуцированные в клетках млекопитающих, обычно растворимы, гликозилированы и имеют аутентичные N-концы. Векторы экспрессии млекопитающих могут содержать нетранскрибируемые элементы, такие как источник репликации, промотор и энхансер, и нетранслируемые 5'- или 3'-последовательности, такие как сайты связывания рибосом, сайты полиаденилирования, акцепторный сайт и сплайсинг-донор, и последовательности терминации транскрипции. Промоторы, пригодные для использования в векторах экспрессии млекопитающих, обычно являются, например, промоторами вирусов, таких как вирус полиомы, аденовирус, вирус человеческого Т-клеточного лейкоза (HTLV), вирус обезьяны 40 (SV 40) и человеческий цитомегаловирус (CMV).
В зависимости от системы экспрессии и выбранного хозяина гомогенный рекомбинантный слитый белок может быть получен с использованием различных комбинаций общеизвестной хроматографии, применяемой для очистки белка. Это включает иммуноаффинную хроматографию, обращеннофазовую хроматографию, катионообменную хроматографию, анионообменную хроматографию, хроматографию гидрофобного взаимодействия, гель-фильтрационную хроматографию и ВЭЖХ. Если система экспрессии секретирует слитый белок в ростовую среду, белок может быть очищен непосредственно из среды. Если слитый белок не секретируется, он выделяется из клеточных лизатов. Разрушение клеток может быть осуществлено любым общеизвестным способом, включая чередование замораживания-оттаивания, облучение ультразвуком, механическое разрушение или применение агентов, лизирующих клетки.
В предпочтительном варианте воплощения по данному изобретению конструкции экспрессии млекопитающих были трансфицированы в клетки СНО DXB11. Устойчивые популяции были отобраны с использованием 400 мкг/мл гигромицина В среде HAMS/F12. Уровни экспрессии составили приблизительно 500 мкг/л. Чтобы повысить уровни экспрессии, была отобрана популяция с использованием 100 нМ метотрексата в среде α-МЕМ. Примерный уровень экспрессии данной популяции составил 5 мкг/л.
Эта конструкция содержала последовательность e-tag на С-конце белка. Колонки со сродством к e-tag были приобретены у American/Pharmacia Biotech. Среду клеточной культуры фильтровали через фильтр 0,22 мкм и загружали в колонку на 5 мл с последовательностью e-tag со скоростью 2 мл/мин. Колонку промывали 0,2 М фосфатным буфером с 0,05% NaNs, pH 7,0 и затем элюат собирали в пробирки, содержащие 0,1 объема 1 М трис-буфера, pH 8,2, чтобы нейтрализовать буфер элюции. Альтернативно фильтрованную культуральную среду загружали в колонку с белком А. В данном случае колонку промывали 50 мМ лимонной кислотой, 300 мМ NaCl, pH 6,5 и элюировалй тем же самым буфером при pH 3,0. В обоих случаях очищенные образцы затем загружали в колонку с сефадексом 200, чтобы отделить мономер от димерной формы слитого белка.
Фармацевтические композиции
Изобретение обеспечивает также фармацевтические композиции, которые могут быть введены пациенту для достижения терапевтического эффекта. Фармацевтические композиции по данному изобретению могут быть получены для введения путем комбинирования слитого белка, обладающего требуемой степенью чистоты и фармацевтически эффективным количеством, с физиологически приемлемыми носителями.
Слитые белки по настоящему изобретению могут применяться в фармацевтических композициях для внутривенного введения, или подкожного введения, или подоболочечного введения. Так, описанные выше слитые белки предпочтительно будут комбинироваться с приемлемым стерильным фармацевтическим носителем, таким как 5% декстроза, модифицированный лактатом раствор Рингера, нормальный физиологический раствор, стерильная вода или любой другой приготовленный для продажи физиологический буферный раствор, созданный для внутривенной инфузии. Будет понятно, что выбор раствора-носителя и дозы и способа введения композиции будут меняться в зависимости от субъекта и конкретной клинической ситуации и будут обусловлены стандартными медицинскими процедурами.
В соответствии со способами по настоящему изобретению эти фармацевтические композиции могут быть введены в количествах, эффективных для подавления патологических последствий, ассоциированных с избыточной генерацией тромбина у субъекта.
Введение слитого белка может быть болюсной внутривенной инъекцией, непрерывной внутривенной инфузией или комбинацией обоих путей. Альтернативно или дополнительно слитый белок, смешанный с соответствующими наполнителями, может попасть в систему кровообращения из внутримышечного участка. Систематическую обработку слитым белком можно контролировать определением активированного неполного тромбопластинового времени (ПВ) на серийных образцах крови, взятых у пациента. Время коагуляции, наблюдаемое в этом анализе, является длительным, когда в системе кровообращения достигается достаточный уровень слитого белка.
Рекомбинантные слитые белки и фармацевтические композиции по данному изобретению применимы для парентерального, местного, внутривенного, перорального или локального введения. Фармацевтические композиции могут быть введены в разнообразных стандартных дозированных формах в зависимости от способа введения. Например, стандартные дозированные формы могут быть введены в лекарственной форме, включающей, но без ограничения, таблетки, капсулы, порошки, растворы и эмульсии.
Рекомбинантные слитые белки и фармацевтические композиции по данному изобретению применимы для внутривенного введения. Композиции для введения обычно будут содержать раствор одноцепочечного антитела или слитого белка, включающего одноцепочечное антитело, растворенное в фармацевтически приемлемом носителе, предпочтительно в водном носителе. Могут применяться разнообразные водные носители, например забуференный физиологический раствор и тому подобное. Эти растворы стерильны и, как правило, свободны от нежелательного материала. Композиции могут стерилизоваться обычными, хорошо известными способами.
Типичная фармацевтическая композиция для внутривенного вливания может быть легко определена специалистом в данной области. Вводимые количества совершенно очевидно являются специфическим белком и зависят от его эффективности и фармакокинетического профиля. Современные способы получения парентерально вводимых композиций будут известны или очевидны специалистам и описаны более подробно в таких публикациях, как Remington's Pharmaceutical Science, 15оеизд., Mack Publishing Company, Easton, Pa (1980).
Композиции, содержащие настоящие слитые белки или коктейль на их основе (т.е. с другими белками), могут вводиться в виде терапевтических курсов лечения. При терапевтическом применении композиции вводят пациенту, страдающему кровотечением в результате нарушения или заболевания, в количестве, достаточном, чтобы лечить или по меньшей мере остановить кровотечение. Количество, достаточное для того, чтобы это осуществить, определено как «терапевтически эффективное количество». Количества, эффективные для данного применения, будут зависеть от тяжести заболевания и общего состояния здоровья пациента.
Единственное или множественное введение композиций может проводиться в зависимости от дозы и частоты, которые требуются и переносимы пациентом. В любом случае композиция должна обеспечить достаточное количество белков по данному изобретению для эффективного лечения пациента.
Слитые белки по изобретению или их фармацевтически приемлемые композиции вводят в терапевтически эффективном количестве, которое будет меняться в зависимости от множества факторов, включая активность применяемого специфического слитого белка; метаболическую стабильность и длительность действия слитого белка; возраст, массу тела, общее состояние здоровья, пол и диету пациента; способ действия и время введения, скорость экскреции; комбинацию препаратов; тяжесть конкретного заболевания-состояния и реципиента, подвергающегося терапии. Как правило, ежедневное терапевтически эффективное количество составляет от примерно 0,14 мг до примерно 14, 3 мг/кг массы тела/день слитого белка по изобретению или его фармацевтически приемлемой композиции; предпочтительно от примерно 0,7 мг до 10 мг/кг массы тела/день и наиболее предпочтительно от примерно 1,4 мг до примерно 7,2 мг/кг массы тела/день. Например, для введения человеку с массой 70 кг дозовый диапазон был бы от примерно 10 мг до примерно 1,0 г/день слитого белка по изобретению или его фармацевтически приемлемой композиции, предпочтительно от примерно 50 мг до примерно 700 мг/день и наиболее предпочтительно от примерно 100 мг до примерно 500 мг/день.
Генная терапия
Слитый белок по изобретению может быть также использован согласно настоящему изобретению путем экспрессии такого слитого белка in vivo, которую часто относят к «генной терапии». Так, например, клетки могут быть сконструированы с полинуклеотидом (ДНК или РНК), кодирующим слитый белок ех vivo, затем созданные клетки вводят пациенту, который должен быть подвергнут терапии слитым белком. Такие способы хорошо известны специалистам. Например, клетки могут быть сконструированы с помощью методик, известных специалистам, с использованием ретровирусных частиц, содержащих РНК, кодирующую слитый белок по настоящему изобретению.
Локальная доставка антикоагулирующих слитых белков по настоящему изобретению, использующая генную терапию, может обеспечить терапевтический агент в области-мишени, эндотелиальной клеточной выстилке кровеносных сосудов.
Рассматриваются методологии генной терапии как in vitro, так и in vivo. Известно несколько способов переноса потенциально терапевтических генов, чтобы определять клеточную популяцию. Смотри, например, Mulligan, Science 260, 1993, с. 936-931. Эти способы включают:
(1) прямой перенос гена. Смотри, например, Wolff и др. (1990) Science 247: 1465-1468;
(2) опосредованный липосомой перенос ДНК. Смотри, например, Caplen и др., Nature Med. 3, 1995, с. 39-46; Crystal, Nature Med. 1, 1995, c.15-17; Gao и Huang, Biochem. Biophys. Res. Comm. 179, 1991, с. 280-285;
(3) опосредованный ретровирусом перенос ДНК. Смотри, например, Кау и др., Science 262, 1993, с. 117-119; Anderson, Science 256, 1992, с. 808-813;
(4) опосредованный ДНК-содержащим вирусом перенос ДНК. Такие ДНК-содержащие вирусы включают аденовирусы (предпочтительно векторы на основе Ad2 или Ad5), вирусы герпеса (предпочтительно векторы на основе вируса простого герпеса) и парвовирусы (предпочтительно векторы на основе «дефектных» или неавтономных парвовирусов, более предпочтительно векторы на основе адено-ассоциированных вирусов, наиболее предпочтительно векторы на основе AAV-2). Смотри, например, Ali и др., Gene Therapy, I, 1994, с. 367-384; патент US 4797368, включенный в контекст путем цитирования, и патент US 5139941, включенный в контекст путем цитирования.
Выбор конкретной векторной системы для переноса гена, представляющего интерес, будет зависеть от множества факторов. Одним важным фактором является клеточная популяция-мишень. Хотя ретровирусные векторы широко изучены и использовались во многих случаях применения генной терапии, эти векторы, как правило, не подходят для заражения неделящихся клеток. Кроме того, ретровирусы обладают потенциалом онкогенности. Однако недавние разработки в области лентивирусных векторов могут обойти некоторые из этих ограничений. Смотри Naldini и др., Science 272, 1996, с. 263-267.
Ретровирусы, на основе которых могут быть получены ретровирусные плазмидные векторы, упомянутые выше, включают, но без ограничения, вирус мышиного лейкоза Молони, вирус некроза селезенки, ретровирусы, такие как вирус саркомы Рауса, вирус саркомы Харви, вирус лейкоза птиц, вирус лейкоза гиббонов, вирус иммунодефицита человека, аденовирус, вирус миелопролиферативной саркомы и вирус опухоли молочной железы. В одном варианте воплощения изобретения ретровирусный плазмидный вектор создается на основе вируса мышиного лейкоза Молони.
Аденовирусы обладают тем преимуществом, что они имеют широкий диапазон хозяев, могут заражать покоящиеся или полностью дифференцированные клетки, такие как нейроны или гепатоциты, и, по-видимому, в существенной мере не являются онкогенными. Смотри, например, Ali и др. (1994), выше, стр.367. Аденовирусы, по-видимому, не интегрируют в геном хозяина. Из-за того, что они существуют экстрахромосомально, опасность инсерционного мутагенеза сильно снижена. Ali и др. (1994), выше, стр.373.
Адено-ассоциированные вирусы обладают подобными преимуществами, что и векторы на основе аденовирусов. Однако AAVs обладают свойством сайт-специфичной интеграции в человеческую хромосому 19 (Ali и др., 1994, с.377).
В предпочтительном варианте воплощения изобретения ДНК, кодирующая слитые белки по данному изобретению, применяется в генной терапии нарушений, включая, но без ограничения, тромбоз глубоких вен, коагулопатию потребления, острый коронарный синдром или злокачественную опухоль с признаками коагулопатии.
Согласно данному варианту воплощения генная терапия с ДНК, кодирующей слитые белки по данному изобретению, проводится нуждающемуся в этом пациенту параллельно или незамедлительно после постановки диагноза.
Квалифицированный специалист признает, что любой соответствующий вектор для генной терапии, содержащий ДНК, кодирующую слитый белок по изобретению, или ДНК, кодирующую аналоги, фрагменты, производные или варианты слитого белка по изобретению, может быть применен в соответствии с данным вариантом воплощения. Методики создания такого вектора известны. Смотри, например, W.F. Anderson, Nature 392,1998, с. 25-30; I.M. Verma и N. Somia, Nature 389, 1998, c. 239-242. Введение ДНК-содержащего вектора слитого белка в сайт-мишень может быть выполнено с использованием известных методик.
Вектор для генной терапии включает один или несколько промоторов. Соответствующие промоторы, которые могут использоваться, включают, но без ограничения, ретровирусный длинный концевой повтор (LTR); промотор вируса SV40 и промотор человеческого цитомегаловируса (CMV), описанный Miller и др., Biotechniques 7 (9), 1989, c. 980-990, или любой другой промотор (например, клеточные промоторы, такие как эукариотические клеточные промоторы, включающие, но без ограничения, гистоновый, pol III и β-актиновый промоторы). Другие вирусные промоторы, которые могут использоваться, включают, но без ограничения, аденовирусные промоторы, тимидинкиназные (ТК) промоторы и промоторы парвовируса В 19. Выбор соответствующего промотора из содержащихся в контексте указаний будет очевидным для специалиста в данной области.
Последовательность нуклеиновой кислоты, кодирующая слитый белок по настоящему изобретению, находится под контролем соответствующего промотора. Соответствующие промоторы, которые могут использоваться, включают, но без ограничения, аденовирусные промоторы, такие как аденовирусный основной поздний промотор; или гетерологичные промоторы, такие как промотор цитомегаловируса (CMV); промотор респираторного синтициального вируса (RSV); индуцируемые промоторы, такие как промотор мышиной опухоли молочной железы (ММТ), промотор металлотионеина; промоторы теплового шока; альбуминовый промотор; промотор аполипопротеина Al (ApoAl); промоторы человеческого глобина; промоторы вирусной тимидинкиназы, такие как промотор тимидинкиназы вируса простого герпеса; ретровирусные LTRs (включая модифицированные ретровирусные LTRs, описанные выше); промотор β-актина и промотор человеческого гормона роста.
Ретровирусный плазмидный вектор используется, чтобы трансдуцировать упакованные клеточных линий с целью образования продуцирующих линий клеток. Примерами упакованных клеток, которые могут быть трансфицированы, включают, но без ограничения, линии клеток РЕ501, РА317, ψ-2, ψ-AM, PA12, Т19-14Х; VT-19-17-H2, ψCRE, ψCRIP, GP+#-86, GP+envAm12 и DAN, как описано в публикации Miller, Human Gene Therapy, 1990, c. 15-14, которая включена в контекст полностью путем цитирования. Вектор может трансдуцировать упакованные клетки посредством любых способов, известных специалистам. Такие способы включают, но без ограничения, электропорацию, применение липосом и осаждение фосфатом кальция. По одной из альтернативных возможностей ретровирусный плазмидный вектор может быть инкапсулирован в липосому или соединен с липидом и затем введен хозяину. Продуцирующая линия клеток генерирует инфекционные ретровирусного частицы вектора, которые включают нуклеиновые последовательности, кодирующие полипептиды. Такие частицы ретровирусного вектора затем могут использоваться для трансдукции эукариотических клеток или in vitro, или in vivo. Трансдуцированные эукариотические клетки будут экспрессировать нуклеиновые последовательности, кодирующие полипептид. Эукариотические клетки, которые могут быть трансдуцированы, включают, но без ограничения, эмбриональные стволовые клетки, эмбриональные клетки карциномы, а также гемопоэтические стволовые клетки, гепатоциты, фибробласты, миобласты, кератиноциты, эндотелиальные клетки и бронхиальные эпителиальные клетки.
Другим подходом к генной терапии является «транскариотическая терапия», где клетки пациентов обрабатывают ex vivo для индукции у покоящихся хромосомных генов способности продуцировать представляющий интерес белок после обратного введения пациенту. Транскариотическая терапия предполагает, что реципиент имеет нормальный комплект генов, необходимых для активации. Транскариотическая терапия включает введение промотора или другой экзогенной регуляторной последовательности, способной активировать находящиеся на стадии возникновения гены в хромосомную ДНК клеток пациента ех vivo, культивирование и отбор клеток, продуцирующих активный белок, и затем повторное введение активированных клеток пациенту с намерением, что они там полностью восстанавливаются. Клетки, «активированные геном», затем производят представляющий интерес белок в течение некоторого значительного периода времени, возможно, такой продолжительности, как жизнь пациента. Патенты US 5641670 и 5733761 раскрывают детали этой концепции и тем самым включены во всей полноте путем цитирования.
Наборы
Кроме того, данное изобретение относится к наборам для исследовательских или диагностических целей. Обычно набор включает один или несколько контейнеров, содержащих одноцепочечные антитела по настоящему изобретению. В предпочтительном варианте воплощения изобретения наборы включают контейнеры, содержащие одноцепочечные антитела в форме, подходящей для получения производного со второй молекулой, например доменом ТМ или его фрагментами. В более предпочтительном варианте воплощения наборы включают контейнеры, содержащие слитые белки с последовательностями SEQ ID No:2 или SEQ ID No:3.
В другом варианте воплощения изобретения наборы могут содержать последовательности ДНК, кодирующие слитые белки. Предпочтительно последовательности ДНК, кодирующие эти слитые белки, обеспечиваются в плазмиде, подходящей для трансфекции в клетку-хозяин и экспрессии в хозяйской клетке. Плазмида может содержать промотор (часто индуцируемый промотор) для регуляции экспрессии ДНК в клетке-хоязяине. Плазмида может также содержать соответствующие сайты рестрикции для содействия вставке в плазмиду другой последовательности ДНК, чтобы продуцировать различные слитые белки. Плазмиды могут также содержать многочисленные другие элементы, чтобы содействовать клонированию и экспрессии кодированных белков. Такие элементы хорошо известны специалистам и включают, например, селектируемые маркеры, кодоны инициации, кодоны терминации и тому подобное.
Терапевтические показания
Заболевания, при которых образование тромба играет важную этиологическую роль, включают инфаркт миокарда, коагулопатию потребления, тромбоз глубоких вен, легочную эмболию, ишемический удар, септический шок, острый респираторный дистресс-синдром, нестабильную стенокардию и другие артериальные и венозные обтурирующие состояния. Слитые белки по настоящему изобретению полезны при всех этих, а также других заболеваниях, при которых тромбообразование является патологическим. Другие патологические состояния, при которых слитый белок по данному изобретению может быть полезен, включают злокачественную опухоль с коагулопатией и воспаление. Соединения могут также найти применение при пересадках кожи и вен и трансплантации органов. Термин «полезный» означает, что соединения полезны для терапии, или для предупреждения заболевания, или для его прогрессирования в более тяжелое состояние. Соединения по данному изобретению обеспечивают также безопасный и эффективный антикоагулянт, например, у пациентов, получающих биопротезы, такие как сердечные клапаны. Эти соединения могут заменить гепарин и варфарин в лечении, например, легочной эмболии или острого инфаркта миокарда. Слитые белки по данному изобретению могут также найти применение в нанесении покрытия на медицинские приспособления, где коагуляция является предметом озабоченности.
Анализы
Много лабораторных анализов для измерения активности ТМ в слитом белке по изобретению являются доступными. Активность белка С может быть измерена в анализе, описанном Н.Н. Salem и др., 1984 выше, и J.B. Galvin и др., J. Biol. Chem. 262 (5), 1987, с. 2199-2205. Кратко анализ состоит из двух этапов. Первым этапом является инкубация тестируемого слитого белка с тромбином и белком С при определенных условиях. На втором этапе тромбин инактивируют гирудином или антитромбином III и гепарином, и активность вновь активированного белка С определяют с применением хромогенного субстрата, посредством чего хромофор высвобождается с помощью протеолитической активности активированного белка С. Этот анализ проводят с очищенными реагентами.
Альтернативно эффект слитого белка может быть измерен с использованием анализов времени свертывания плазмы, такого как время частично активированного тромбопластина (ЧАТВ), время свертывания тромбина (ВСТ) и/или протромбиновое время (ПВ). Эти анализы проводят различие между разными механизмами подавления коагуляции и включают активацию белка С. Удлинение времени свертывания в любом из этих анализов показывает, что молекула может подавить коагуляцию в плазме.
Приведенные выше анализы применяются для идентификации слитых белков с активностью ТМ, которые способны связывать тромбин как в очищенных системах, так и в окружающей среде плазмы. Дополнительные анализы затем используют, чтобы оценить другие виды активности нативного ТМ, такие как подавление катализируемого тромбином образования фибрина из фибриногена (H.V. Jakubowski и др., J. Biol. Chem. 261 (8), 1986, с. 3876-3882), подавление активации тромбином фактора V (С.Т. Esmon и др., J. Biol. Chem. 257 (14), 1982, 7944-7947), ускоренное подавление тромбина антитромбином III и гепариновым кофактором II (N.L. Esmon и др., J. Biol. Chem. 258 (20), 1983, с. 12238-12242), подавление тромбином фактора XIII (J. Polgar и др., Thromb. Haemost. 58 (1), 1987, с.140), подавление опосредованной тромбином инактивации белка S (Е.А. Thompson и Н.Н. Salem., J. Clin. Inv.78 (1), 1986, с. 13-17) и подавление опосредованной тромбином активации и агрегации тромбоцитов (N.L. Esmon, и др., 1983, см. выше).
Следующие анализы, описанные в деталях ниже в примере 5, использовали для измерения эффективности in vitro слитых белков по изобретению: (1) анализ (хромогенный) активации белка С; (2) анализ активации sTF/FVIIa; (3) анализ активации фактора X; и (4) анализ активации белка С (на богатой TF поверхности).
Подразумевается, что при осуществлении методик по настоящему изобретению, безусловно, следует понимать, что ссылки на конкретные буферы, среды, реагенты, условия культивирования и тому подобное не являются ограничивающими, но должны восприниматься так, будто включают все родственные материалы, которые специалист в данной области признал бы представляющими интерес или ценность в особом контексте, в котором это обсуждение представлено. Например, часто можно заменить одну буферную систему или культуральную среду другой и все же достичь подобных, если не идентичных, результатов. У таких специалистов будет достаточно информации о таких системах и методологиях, чтобы суметь без ненадлежащего экспериментирования сделать такие замены, которые будут оптимально служить своим целям при применении способов и методик, раскрытых в контексте.
Настоящее изобретение теперь будет, кроме того, описываться с помощью следующих не ограничивающих его примеров. При использовании раскрытия примера следует четко помнить, что другие и разные варианты воплощения способов, раскрытых по настоящему изобретению, будут, без сомнения, предлагаться специалистам в имеющей отношение к делу области.
Без дальнейшего уточнения считается, что любой специалист в этой области, используя предшествующее описание, применит настоящее изобретение в его самом полном объеме. В связи с этим следующие предпочтительные специфические варианты воплощения изобретения должны рассматриваться как просто иллюстративные, а не ограничивающие никоим образом оставшееся раскрытие.
В упомянутых и в следующих примерах все температуры не исправлены и даны в градусах Цельсия, все части и проценты являются массовыми, если не указано иначе.
Полные раскрытия заявок, патентов и публикаций, цитированных выше, тем самым включены путем цитирования.
Следующие примеры предусмотрены в качестве руководства, чтобы содействовать применению изобретения на практике.
Пример 1
Конструкция однопепочечного анти-TF антитела scFv(TF)3e10
(-18) M L G V L V L G A L A L A G L V F P E M A Q V N L R E
S G G T L V Q P G G S L R L S C A A S G F S F T D A W M S
W V R Q A P G K E L E W V S S I S G S G G S T Y Y A G S V K
G R F T I S R D N S K N T L Y L Q M N S L R A E D Т A V Y Y
C A R V L S L T D Y Y W Y G M D V W G Q G T L V T V S A G
G G G S G A P N F M L T Q P H S V S A S P G K T V T I S C T
R S S G S V A S Y Y V Q W Y Q Q R P G S S P T T V I Y E D N
H R P S G V P D R F S G S I D T S S N S A S L T I S G L K T E
D E A D Y Y C Q S Y D S N N L V V F G G G T K L T V L G A
A A G A P V P Y P D P L E P R A A (264)
Одноцепочечное анти-TF антитело scFv(TF)3e10 (SEQ ID No:1) состоит из сигнального пептида (-18 до -1), домена VH (1-126), линкера VH-VL (127-131), домена VL (132-246) и последовательности e-tag (247-264).
Пример 2
Конструкция 1 слитого белка scFv(TF)3e10-TMi456


Слитый белок scFv(TF)3e10-TMi456 (SEQ ID No:2) состоит из сигнального пептида (-18 до -1), домена VH (1-126), линкера VH-VL (127-131), домена VL(132-246), линкера VL-TM(247-264), домена TMi456 (265-382) и последовательности е-tag (383-400). В TMi456 подчеркнуты мутации H381G, M388L, R456G и H457Q.
Пример 3
Конструкция 2 слитого белка scFv(TF)3e10-TMi456A

Слитый белок scFv(TF)3e10-TMi456A (SEQ ID No:3) состоит из сигнального пептида (-18 до -1), домена VH (1-126), линкера VH-VL (127-131), домена VL (132-243), линкера VL-тм (244-261) и домена TMi456 (262-379). В TMi456 подчеркнуты мутации H381G, M388L, R456G и H457Q.
Пример 4
Экспрессия слитого белка в бактериальных клетках/клетках млекопитающих
Экспрессия в бактериальных клетках возможна, но она дает белок, который обладает сильно сниженной кофакторной активностью ТМ. Слитый белок экспрессировали в клетках СНО. Плазмида экспрессии содержала как гигромицин В, так и селектируемые маркеры DHFR. Первоначальная селекция была проведена при 400 мкг/мл гигромицина, чтобы отобрать популяцию. Образующуюся популяцию затем подвергали селекции с 100 нМ метотрексатом. В ходе данной селекции клетки, которые имеют амплифицированные копии участка ДНК, содержащего селектируемый маркер, и ген-мишень, отбирают среди популяции. В результате данной селекции уровни экспрессии были повышенными от приблизительно 0,3 мг/л до примерно 6 мг/л.
Пример 5
Анализы in vitro
1. Анализ (хромогенный) активации белка С
Данный анализ проводили смешиванием 20 мкл каждого из следующих белков в планшете для микротитрования: образец ТМ (неизвестный или стандартный), тромбин (3 нМ) и белок С (1,5 мкМ). Разбавителем в анализе для каждого белка был 20 мМ трис-HCl, 0,1 М NaCl, 2,5 мМ CaCl2, 2,5 мг/мл бычьего сывороточного альбумина (BSA), рН 7,4. Лунки инкубировали в течение 2 ч при 37°С, после чего активацию белка С прерывали добавлением 20 мкл гирудина (0,16 ед/мкл, 370 нМ) в разбавителе для анализа и инкубировали в течение дополнительных 10 мин.
Количество образовавшегося активированного белка С определяли добавлением 100 мкл 1 мМ субстрата S2266 (в разбавителе для анализа) и продолжали инкубировать планшет при 37°С. Поглощение при 405 нм в каждой лунке считывали каждые 10 сек в течение 30 мин, используя считывающее устройство для планшетов Molecular Devices. Данные поглощения сохраняли, и изменение поглощения в секунду (угловой коэффициент) рассчитывали для каждой лунки. Изменение поглощения в секунду было пропорционально пмолям/мл активированного белка С.
Соотношение определяли эмпирически, используя изменяющиеся концентрации активированного белка С. Образцы, содержащие 100% активированного белка С, генерировали смешиванием белка С при 0-1,5 мкМ с 60 нМ кроличьим ТМ и 30 нМ тромбином, инкубацией в течение 0-4 ч, добавлением гирудина и измерением активности S2266, как описано выше. Условия, при которых активировалось 100% белка С, определяли, как те, при которых активность S2266 (поглощение А405/сек) достигало плато. Единицу активности определяли как 1 пмоль активированного белка С, генерированного в мл/мин с реагентами в условиях, определенных выше. Альтернативно значения активности приведены в сравнении с нативным кроличьим ТМ, растворенным с помощью детергента.
2. Анализ активации sTF/FVIIa
Принцип данного анализа описан ниже. Амидную связь субстрата n-нитроанилида трипептида гидролизовали с помощью комплекса sTF/FVIIa. Выделенный хромофорный продукт n-нитроанилид контролировали при 405 нм и концентрацию продукта, образованного в единицу времени, рассчитывали, используя коэффициент молярной экстинкции 9920 М-1 см-1. Значения IC50 (С) определяли путем аппроксимации начальных скоростей в 4-параметрическом уравнении: Y=(A-D)/(1+(х/С)^B)+D

Реагенты и растворы
1. Буфер для анализа: 50 мМ трис-HCl, 150 мМ NaCl, 5 мМ CaCl2, 0,1% BSA, рН 7,5
2. Человеческий FVIIa (HCVIIA-0060, Haematologic Technologies Inc.): 10x рабочий раствор - приготовить 20 нМ раствор в буфере для анализа перед использованием.
3. Растворимый TF (Berlex): 10х рабочий раствор - приготовить 30 нМ раствор в буфере для анализа перед использованием.
4. Хромогенный субстрат S2266 (Kabi Pharmacia Hepar Inc.): исходный раствор: 10 мМ в Н2О, сохраняемый при 4°С; 2,5х рабочий раствор - приготовить 2,5 мМ раствор в буфере для анализа перед использованием.
5. Антитело: приготовить 2,5х разбавления в буфере для анализа перед использованием.
Условия анализа
Анализы проводили в 96-луночном планшете для титрования при комнатной температуре. Конечные концентрации компонентов были следующими:
Методика анализа
1. Пипетку на 0,1 мл с 2,5х буфера для анализа (или буферного контроля) в каждую лунку.
2. Прибавить 0,025 мл 10х sTF и инкубировать 10 мин при комнатной температуре при легком встряхивании.
3. Прибавить 0,025 мл 10х FVIIa, инкубировать 10 мин при комнатной температуре при легком встряхивании.
4. Прибавить 0,1 мл 2,5х субстрата S2266, незамедлительно перенести планшет в приспособление для считывания планшетов и измерять ферментную кинетику при 405 нм с интервалом в 10 сек в течение 15 мин.
5. Анализ активации фактора Х
Принцип данного анализа описан ниже. Инкубировали FVIIa с везикулами рекомбинантного человеческого TF, чтобы образовать протеазный комплекс, способный активировать субстрат FX. Данный комплекс образуется в присутствии (или в отсутствие) различных концентраций антитела, затем вводили субстрат FX и давали проходить реакции с образованием продукта - активной протеазы Fxa, которая способна гидролизовать амидную связь n-нитроанилида хромогенного субстрата S2222. Выделенный хромофорный продукт, n-нитроанилид, контролировали при 405 нм и концентрацию продукта, образованного в единицу времени, рассчитывали, используя коэффициент молярной экстинкции 9920 М-1 см-1. Величины IC50 (С) определяли путем аппроксимации начальных скоростей в 4-параметрическом уравнении: Y=(А-D)/ (l+ (х/С^В)+D
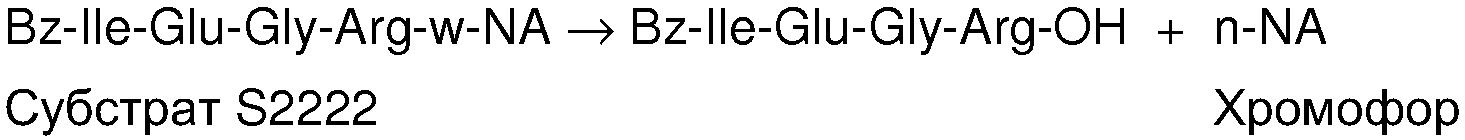
Реагенты и растворы
1. Буфер для анализа: 50 мМ трис-HCl, 150 мМ NaCl, 5 мМ CaCl, 0,1% BSA, рН7,5
2. Человеческий FVIIa (HCVIIA-0031, Haematologic Technologies Inc.): 4x рабочий раствор - приготовить 100 пМ раствор в буфере для анализа перед использованием.
3. Рекомбинантный человеческий TF (восстановленный в лаборатории заявителя из Innovin, Dade): рабочий раствор - приготовить разведение 1:480 в буфере для анализа перед использованием.
4. Фактор Х (НСХ-0060, Haematologic Technologies Inc.): 4х рабочий раствор - приготовить 1000 нМ раствор в буфере для анализа перед использованием.
5. Хромогенный субстрат S2222 (Kabi Pharmacia Hepar Inc.): исходный раствор: 6 мМ в Н2О, сохраняемый при 4°С. Рабочий раствор - приготовить 0,78 мМ раствор в 3,57 мМ этилендиаминтетрауксусной кислоте (EDTA) (чтобы остановить реакцию), 150 мМ NaCl, 50 мМ трис-HCl, рН 7,5 перед использованием.
6. Антитело: приготовить 4х разведения в буфере для анализа перед использованием.
Условия анализа
Анализы проводили в 96-луночном планшете для микротитрования при комнатной температуре. Конечные концентрации компонентов были следующими.
Методика анализа
1. Пипетку на 0,015 мл с 4х буфера для анализа (или буферного контроля) в каждую лунку.
2. Прибавить 0,015 мл 4х пузырьков TF.
3. Прибавить 0,015 мл 4х FVIIa, инкубировать 10 мин при комнатной температуре при легком встряхивании.
4. Прибавить 0,015 мл 4х FX, инкубировать 5 мин при комнатной температуре при легком встряхивании.
5. Прибавить 0,14 мл субстрата S2222, незамедлительно перенести планшет в приспособление для считывания планшетов и измерять ферментную кинетику при 405 нм с интервалом в 10 сек в течение 15 мин.
4. Анализ активации белка С (на TF-богатой поверхности)
Данный анализ проводили, как и хромогенный анализ активации белка С, приведенный выше, за исключением того, что в данном анализе человеческие TF-содержащие везикулы пептидного хромофора/пептидного субстрата (PC/PS) прибавляли к слитому белку или контрольному ТМ до прибавления тромбина и белка С.
Пример 6
Характеристика антител против TF
Семь разных связывающих TF антител выделяли из фаговой дисплейной библиотеки человеческих одноцепочечных антител. Значения сродства связывающих sTF антител, измеренные с использованием BIAcore, были между 35 и 470 нм. Анализ sTF/VIIa использовали, чтобы определить, блокируют ли антитела образование активного комплекса VIIa/TF. В анализе sTF/VIIa связывание Vila с sTF более чем 20-кратно увеличивает скорость расщепления по сравнению с хромогенным пептидным субстратом S2266. Антитела, которые подавляют связывание FVIIa с TF, блокируют данное ускорение. Среди выделенных семи разных антител только одно из них, scFv(TF)3e10, не вызывало подавления в анализе sTF/VIIa. Данное антитело увеличивало сродство FVIIa к sTF, понижая константу диссоциации kd пятикратно (фиг.1). Значение k.d антитела scFv(TF)3e10 для sTF, измеренное с использованием анализа sTF/FVIIa, составило 65,4 нМ (фиг.2). Микрокалориметрию использовали, чтобы сравнить сродство scFv(TF)3e10 к TF при сопоставлении с комплексом FVIIa/TF. Эти эксперименты показали, что антитело имело 20-кратно более высокое сродство к комплексу TF/FVIIa по сравнению со свободным sTF 9 (33 нМ против 600 нМ, фиг.3). Антитела сравнивали, используя анализ активации FX, который состоял из TF полной протяженности в фосфолипидных везикулах, FVIIa и FX. Количество генерированного FXa определяли, используя хромогенный субстрат S2765. Хотя антитело scFv(TF)3e10 не обладало самым высоким сродством, как измерено с помощью BIAcore, и это повышало сродство FVIIa к sTF, оно было единственным антителом в группе, которое подавляло активацию FX и удлиняло время свертывания в анализе ПВ. Значение IC50 (димерного) антитела scFv(TF)3e10 для ингибирования в анализе активации FX составило 0,44 нМ (фиг.4), и двукратное увеличение ПВ происходило при 417 нМ (фиг.5).
Антитело scFv(TF)3e10 идентифицировали на основании связывания с рекомбинантным человеческим растворимым TF. Гомология последовательностей TF между человеческой и мышиной или человеческой и кроличьей составила 58% и 71% соответственно. Антитело связывается с уникальным эпитопом на человеческом TF, который препятствует активации FX комплексом FVIIa/TF. Физиологически антитело имеет преимущество над антителами, которые конкурируют со связыванием FVII или FVIIa с TF. Значение KD как FVII, так и FVIIa в человеческой плазме составило 10 нМ или между 100- и 1000-кратным значением, более высоким, чем KD. Диссоциация для комплекса с высоким сродством будет медленной (70-700 сек, предполагая значение константы скорости ассоциации kon=108 М-1 сек-1). В противоположность этому, константа Михаэлиса Кm фактора Х для комплекса VIIa/TF была между 0,200 и 4 мкМ, и концентрация FX в человеческой плазме составила 130 нМ (между 0,03- и 0, 65-кратным КD). Первичная функция комплекса FVIIa/TF при коагуляции состоит в превращении FX в FXa.
Пример 7
Свойства in vitro слитого белка scFv(TF)3e10-TMi456
Свойства слитого белка по изобретению scFv(TF)3e10-TMi456 оценивали в различных анализах in vitro. Слитый белок scFv(TF)3e10-TMi456 сохранял способность подавлять активацию FX с помощью комплекса FVIIa/TF (IC50=0,5 нМ, данные не приведены) и действовать в качестве кофактора катализируемой тромбином активации белка С (хромогенный анализ, фиг.6). Не наблюдалось никакого значительного различия в кофакторной активности ТМ между слитым белком и одним TMi456 в отсутствие содержащих TF фосфолипидных пузырьков. В противоположность этому, кофакторная активность ТМ слитого белка, но не TMi456, повышалась более чем 5-кратно в присутствии содержащих TF фосфолипидных везикул (фиг.7). Активность in vitro слитого белка scFv(TF)3e10-TMi456 против индуцируемой TF коагуляции (анализ ПВ, несвойственный путь коагуляции) была в 6 раз выше, чем антитела scFv(TF)3e10, и в 17 раз выше, чем одного TMi456 (фиг.5). В противоположность этому, эффективность слитого белка in vitro против присущих и общих путей коагуляции не была затронута значительно (анализы АРТТ и ТСТ, данные не приведены). Следовательно, доза слитого белка, которая вызывала двукратное увеличение ПВ, оказывала только умеренное действие на АРТТ, тогда как TMi456 в эквивалентной дозе в анализе ПВ вызывал 4-кратное увеличение АРТТ (фиг.8). Профиль in vitro был совместим с таковым, ожидаемым для антикоагулянтов, направленных на TF/FVIIa, которые, как известно, обладают исключительным воздействием на соотношения кровотечения на животных моделях тромбоза. В соответствии с анализами основанной на плазме коагуляции слитый белок scFv(TF)3e10-TMi456 был более мощным в анализе индуцированной TF коагуляции цельной крови (тромбоэластограф, TEG), чем или антитело scFv(TF)3e10, или один TMi456 (фиг.9). Кроме того, слитый белок scFv(TF)3e10-TMi456 давал более предсказуемый дозовый отклик при коагуляции цельной крови, индуцированной TF, чем низкомолекулярный гепарин (LMWH, фиг.10). В итоге приведенные выше данные демонстрируют, что слитые белки по изобретению являются мощными и селективными антикоагулянтами in vitro.
Пример 8
Модель крысиной тромбоэмболии in vivo
TF-часть антитела слитого белка scFv(TF)3e10-TMi456 является специфичной для TF приматов. Модель тромбоэмболии, инициированная с помощью человеческого TF (тромбопластиновый реагент, содержащий человеческий рекомбинантный TF, Ortho), была создана у находящихся в сознании самцов крыс линии Sprague-Dawley (350-400 г, n>7/группу). На этой модели коагулопатии потребления (DIC) фактор ткани через посредство инъекции тромбопластина индуцирует отложение легочного фибрина, нарушение дыхания и смерть. Эквимолярные дозы scFv(TF)3e10-TMi456, или scFv(TF)3e10, или наполнителя вводили с помощью инъекции в хвостовую вену, затем через 15 мин с помощью болюсной инъекции вводили тромбопластин (0,5 мг/кг). В группе, обработанной наполнителем, данная доза TF приводила к 60% летальности (LD60) обычно в течение 5 мин после инъекции тромбопластина.
Крыс оценивали в соответствии со следующей системой показателей в баллах, характеризующих распространенность заболевания-смертность: 0 - без воздействия; 1 - слабое расстройство дыхания (восстановление к норме через 30 мин); 2 - острое расстройство дыхания (агонирующее состояние, восстановление к норме требовало более чем 60 мин); и 3 - гибель. Средняя оценка была выбрана для сравнения эффективности 4 различных обработанных групп. Результаты использования данного анализа in vivo приведены на фиг.11. В этом анализе слитый белок по изобретению был способен подавить гибель и расстройство дыхания.
Предшествующие примеры могут быть повторены с одинаковым успехом с помощью замещающих в родовом отношении или конкретно описанных реагентов и/или рабочих условий по данному изобретению для тех, которые использованы в предшествующих примерах.
Несмотря на то что изобретение иллюстрировано в отношении получения некоторых конструкций слитых белков, очевидно, что могут быть сделаны изменения и модификации по изобретению без отхода от сущности и объема изобретения.





Реферат
Изобретение относится к области медицины и касается новых слитых белков тромбомодулина, обеспечивающих направленный перенос к тканевому фактору, которые могут применяться в качестве антикоагулянтов. Сущность изобретения включает антикоагулирующий слитый белок, содержащий белок, обеспечивающий направленный перенос, который функционально связан с тромбомодулиновым (ТМ) доменом EGF456 и с доменом ТМ, включающим междоменную петлю между EGF3 и EGF4 или их производных или вариантов, и взаимодействующий с тканевым фактором (TF) или комплексом фактора VIIa/TF. Слитый белок связывается в участке повреждения и предотвращает инициацию тромбоза, а также может применяться для терапии различных тромбозных состояний, включая, но без ограничения, тромбоз глубоких вен, коагулопатию потребления и острый коронарный синдром. Преимущество изобретения заключается в расширении области применения. 7 н. и 17 з.п. ф-лы, 11 ил.
Комментарии