Новые антитела к тканевому фактору в качестве антикоагулянтов - RU2345789C2
Код документа: RU2345789C2
Чертежи







Описание
Для нормального гемостаза важную роль играет поддержание требуемого баланса между прокоагулирующей и антикоагулирующей активностью в кровеносных сосудах (Davie E.W., Biochemistry, 30(43), 1991, сс.10363-10370). Сдвиг баланса в сторону коагуляции приводит к тромбозу, что может приводить к сердечному приступу, "удару", легочной эмболии и венозному тромбозу. В связи с вышеуказанным существует потребность в более эффективных и безопасных антикоагулянтах, предназначенных для лечения специфических связанных с тромбозом заболеваний.
Тканевый фактор ("ТФ") представляет собой трансмембранный гликопротеин, который является основным инициатором каскада коагуляции (Nemerson Y., Thromb. Haemost, 74(1), 1995, сс.180-184). В нормальных физиологических условиях ТФ не имеет контакта с кровью. При повреждении сосуда попадание в кровь субэндотелиального ТФ и коллагена приводит к активации факторов свертывания и тромбоцитов и, как следствие, к образованию гемостатической пробки. Попавший в кровь ТФ действует в качестве кофактора для фактора VIIa ("FVIIa"), катализирующего активацию фактора IX ("FIX") и фактора Х ("FX"), которые представляют собой имеющие решающие значение компоненты присущих организму комплексов теназы и протромбиназы соответственно. Это приводит к быстрому формированию FXa и тромбина. Затем тромбин расщепляет фибриноген с образованием фибрина, который, в свою очередь, полимеризуется с образованием фибринового сгустка. Несоответствующая индукция экспрессии ТФ при различных клинических симптомах может приводить к угрожающему жизни тромбозу и/или вызывать патологические осложнения. Попадание в кровь ТФ после разрушения бляшки, вероятно, приводит к тромботической окклюзии, которая является причиной острого инфаркта миокарда и "удара". В этих случаях активация факторами свертывания путей сигналов провоспаления также приводит к образованию отека и увеличению размера инфаркта. Повреждение сосуда, связанное с пластической операцией на сосуде, приводит к повышающей регуляции ТФ на поверхности клеток гладкой мускулатуры (SMC), что, вероятно, индуцирует пути клеточных сигналов, связанных с рестенозом. Сверхэкспрессия ТФ в опухоли и при связанном с грамотрицательными бактериями сепсисе приводит к угрожающему жизни тромбозу и активации воспалительных путей.
Комплекс FVIIa/ТФ участвует в механизме патогенеза при различных тромботических заболеваниях, и наличие в кровотоке определенной концентрации ТФ является фактором риска для некоторых типов пациентов. FVIIa и ТФ играют уникальную роль в нарушении функции сосудов в поддержании гемостаза и инициации тромбоза при повреждении сосудов.
ТФ в норме экспрессируется в адвентиции, но при заболевании сосудов происходит повышающая регуляция и не присущая организму в норме экспрессия ТФ в медии и неоинтиме. Экспрессия ТФ в атеросклеротических бляшках повышается, и при контакте с ТФ защита крови, осуществляемая с помощью тонкой фиброзной оболочки ("шапочки"), может нарушаться. Хирургические вмешательства, такие как пластическая операция на сосудах с использованием баллонов, введение стентов или эндартерэктомия, повреждают стенки сосудов и приводят к контакту с находящимся в более глубоких слоях ТФ. Спонтанное разрушение или эндотелиальная эрозия атеросклеротической богатой липидами тонкостенной бляшки приводит к контакту с ТФ и тромбозу, что вызывает нестабильную стенокардию и инфаркт миокарда. ТФ может находиться в кровотоке в микрочастицах клеточного происхождения, и уровни ТФ в кровотоке повышаются при нестабильной стенокардии, что позволяет предположить, что находящийся в кровотоке ТФ может принимать участие в образовании тромба (Soejima Н. и др. Circulation, 99(22), 1999, сс.2908-2913). Рак часто связан с состоянием гиперкоагуляции, обусловленной сверхэкспрессией ТФ на поверхности опухолевых клеток. Это приводит к предрасположенности пациента к тромбозу глубоких вен, эмболии легких и к ДВС-синдрому (диссеминированное внутрисосудистое свертывание крови) невысокой степени. ДВС приводит к отложению фибрина в микрососудах, приводящему к нарушению многих органов.
Антикоагулянты на основе белков, мишенью которых является ТФ, включают нейтрализующие ТФ антитела, ингибирующие активный сайт фактора VIIa ("FVIIai"), ингибитор пути метаболизма тканевого фактора ("TFPI") и антикоагулирующий белок нематод ("NAPC2"). Результаты, полученные с использованием моделей тромбоза на основе острого артериального повреждения, свидетельствуют о том, что белковые ингибиторы FVIIa/ТФ являются эффективными антитромботическими агентами, которые характеризуются меньшей способностью по сравнению с гепарином вызывать кровотечение, непосредственными ингибиторами тромбина, ингибиторами тромбоцитов и ингибиторами FXa (Himber J. и др., Thromb. Haemost. 85, 2001, сс.475-481; Harker L.A. и др., Thromb. Haemost. 74(1), 1995, сс.464-472). Кроме того, ингибиторы FVIIa/ТФ являются более эффективными, чем другие антикоагулянты (например, гепарин, ингибитор FXa), в отношении способности предупреждать утолщение неоинтимы и сосудистый стеноз после повреждения баллоном (Jang Y. и др., Circulation, 92(10), 1995, сс.3041-3050).
С помощью экспериментальных моделей сепсиса установлено, что ингибирование ТФ, FVIIa или комплекса FVIIa/ТФ представляет собой эффективный антитромботический путь предупреждения ДВС и снижения смертности. Аналоги TFPI предупреждают индуцированное как тромбопластином, так и эндотоксином ДВС у кроликов (Day K..C. и др., Blood, 76, 1990, сс.1538-1545); Bregengard С. и др., Blood Coagul. Fibrinolysis, 4, 1993, сс.699-706). Моноклональные антитела к FVIIa (Biemond B.J. и др., Thromb. Haemost., 73, 1993, сс.223-230 или Levi M. и др., J.Clin. Invest., 93, 1994, сс.114-120) предупреждают индуцированное эндотоксином ДВС у обезьян. Нейтрализующие ТФ антитела, FVIlai и TFPI ингибируют ДВС и снижают смертность павианов при использовании модели индуцированного Е.coli сепсиса (Creasey А.А. и др., J. Clin. Invest. 91, 1993, сс.2850-2860; Taylor F.В. и др., Blood, 78, 1991а, сс.364-368; Taylor F. В. и др., Circ. Shock, 33, 1991b, сс.127-134; Taylor F.В. Haemostasis Suppl. 126, 1996, сс.83-91). Известно, что как FXa, так и комплекс FVIIa/ТФ/РХа индуцируют производство провоспалительных цитокинов, которые связаны с повышенном риском смерти пациентов, страдающих сепсисом (Riewald M. и др., Proc. Natl. Acad. Sci. USA, 98, 2001, сс.7742-7747). Интересно отметить, что, как установлено с использованием в качестве модельных животных павианов, FVIIai снижает уровни в плазме IL-6 и IL-8 (Taylor F.B. и др., Blood, 91, 1998, сс.1609-1615), что позволяет предположить, что ингибирование FVIIa/ТФ может оказывать дополнительное противовоспалительное действие, не связанное с другими антикоагулирующими механизмами.
Из литературы известно несколько антител, которые являются эффективными антикоагулянтами и которые связываются или нейтрализуют либо ТФ, либо комплекс FVIIa/ТФ, либо и их обоих (см., например, Carson S.D. и др., Blood, 66(1), 1985, сс.152-156; Tanaka H. и др., Thromb. Res., 40(6), 1985, сс.745-756; Kirchhofer D. и др., Throomb. Haemost., 84(6), 2000, сс.1072-1081; Kirchhofer D. и др., Biochemistry, 40(3), 2001, сс.675-682; Faelber К. и др., J. Mol. Biol.,313, 2001, сс.83-87 и патенты США 5506134, 5986065 и 6274142). Антитела к ТФ, предлагаемые в настоящем изобретении, являются эффективными антикоагулянтами, которые обладают улучшенными характеристиками по сравнению с описанными ранее антителами к ТФ. В частности, антитела, предлагаемые в изобретении, связываются с большей аффинностью с комплексом FVIIa/ТФ, чем со свободным ТФ, и они не конкурируют с FVII или с FX за связывание с ТФ.
Краткое изложение сущности изобретения
Настоящее изобретение относится к антителам, действующим в качестве антикоагулянтов, которые связываются с большей аффинностью с комплексом фактор VIIa/тканевый фактор ("FVIIa/ТФ"), чем со свободным тканевым фактором ("ТФ"). Согласно одному из вариантов осуществления изобретения антитела, предлагаемые в изобретении, обладают по меньшей мере в 2 раза более высокой аффинностью к связыванию с комплексом FVIIa/ТФ, чем со свободным ТФ, при оценке с помощью микрокалориметрического анализа. Согласно предпочтительному варианту осуществления изобретения антитела, предлагаемые в изобретении, обладают по меньшей мере в 5 раз более высокой аффинностью к связыванию с комплексом FVIIa/ТФ, чем со свободным ТФ. Согласно еще более предпочтительному варианту осуществления изобретения, антитела, предлагаемые в изобретении, обладают по меньшей мере в 10 раз более высокой аффинностью к связыванию с комплексом FVIIa/ТФ, чем со свободным ТФ. Согласно другому варианту осуществления изобретения антитела, предлагаемые в изобретении, не конкурируют за связывание с TF с одним или несколькими факторами свертывания, выбранными из группы, включающей факторы VII ("FVII"), IX ("FIX") и Х ("FX"). Согласно предпочтительному варианту осуществления изобретения антитела, предлагаемые в изобретении, не конкурируют за связывание с TF с факторами FVII и FX. Согласно более предпочтительному варианту осуществления изобретения антитела, предлагаемые в изобретении, связываются с большей аффинностью с комплексом FVIIa/ТФ, чем со свободным ТФ, и не конкурируют за связывание с ТФ с факторами FVII и FX.
Антикоагулирующее антитело, предлагаемое в изобретении, осуществляет направленный перенос и связывается с комплексом FVIIa/ТФ в области повреждения и ингибирует активацию фактора Х ("FX"), предупреждая тем самым образование тромба, и вследствие этого обладает эффективностью в качестве антикоагулянта при лечении определенных заболеваний, включая (но, не ограничиваясь ими) сепсис, диссеминированное внутрисосудистое свертывание крови, ишемический "удар", тромбоз глубоких вен, острый коронарный синдром, связанные с тромбозом осложнения после пластической операции на сосудах, и коагулопатия при прогрессирующем раке. Кроме того, антитело можно применять при микрососудистой хирургии, при трансплантации кожи и вен и трансплантации органов.
Еще одним объектом изобретения являются фармацевтические композиции, содержащие предлагаемые в изобретении антитела.
Следующим объектом изобретения является способ защиты пациента от образования тромба, заключающийся в том, что пациенту вводят терапевтически эффективное количество антитела, предлагаемое в изобретении, и тем самым ингибируют образование тромбина, не оказывая при этом непосредственного воздействия на другие параметры свертывания, такие как активация и агрегация тромбоцитов.
Следующим объектом изобретения является способ уменьшения и лечения тромбоза глубоких вен ("ТГВ"), или диссеминированного внутрисосудистого свертывания ("ДВС"), или острого коронарного синдрома, или рака с признаками коагулопатии у пациента, заключающийся в том, что пациенту вводят терапевтически эффективное количество антитела, предлагаемого в изобретении.
Следующим объектом изобретения является способ регуляции воспалительного ответа у пациента, заключающийся в том, что пациенту вводят терапевтически эффективное количество антитела, предлагаемого в изобретении.
Еще одним объектом изобретения является антитело, предлагаемое в изобретении, которое можно применять для нанесения нетромбогенного покрытия на поверхность медицинских приспособлений, которые контактируют с кровью.
Следующим объектом изобретения является набор, который содержит антитело, предлагаемое в изобретении, которое связывается с комплексом FVIIa/ТФ. В другом варианте набор может содержать последовательности ДНК, кодирующие компоненты антитела.
В изобретении предложены также способы получения антител, предлагаемых в изобретении, как путем рекомбинации, так и путем синтеза.
Описание чертежей
На чертежах показано:
на фиг.1 - данные об активности ТФ-связывающих одноцепочечных антител, полученные на основе анализа способности рТФ/FVIIa осуществлять гидролиз пептида (пептидный гидролиз). Анализ способности рТФ/FVIIa осуществлять пептидный гидролиз осуществляли согласно методу, описанному в примере 4, с применением уравновешенной смеси FVIIa (5 нМ) и рТФ (10 нМ), который основан на аффинности FVIIa к рТФ (КD(арр) составляет ˜10 нМ). Гидролиз хромогенного пептидного субстрата S2266 оценивали согласно описанному методу. Представлены конечные концентрации экспрессируемых в бактериях одноцепочечных антител и контрольных белков FVIIai (FVIIa, инактивируемый хлорметилкетонным пептидом, РРАСК) и МАт №4504 (фирма American Diagnostica);
на фиг.2 - данные, свидетельствующие о том, что связывание scFv(TF)3e10 с рТФ повышает кажущуюся аффинность рТФ к FVIIa. Анализ способности рТФ/FVIIa осуществлять пептидный гидролиз осуществляли согласно методу, описанному в примере 4, с применением 2 нМ FVIIa в присутствии 800 нМ экспрессируемого в бактериях антитела scFv(TF)3e10 или без него. При осуществлении этого анализа рТФ титровали и определяли скорость расщепления хромогенного пептидного субстрата S2266. Кажущееся значение kD для рТФ рассчитывали с помощью стандартной аппроксимации 4-параметрической зависимости;
на фиг.3 - данные об активности ТФ-связывающих антител, полученные путем оценки протромбинового времени (ПВ). Анализ ПВ осуществляли согласно методу, описанному в примере 4, с применением рекомбинантного человеческого тромбопластина (фирма Dade, Inc.), содержащего полноразмерный человеческий ТФ в фосфолипидных пузырьках. Определяли конечные концентрации экспрессируемых в бактериях одноцепочечных антител и контрольного белка FVIIai;
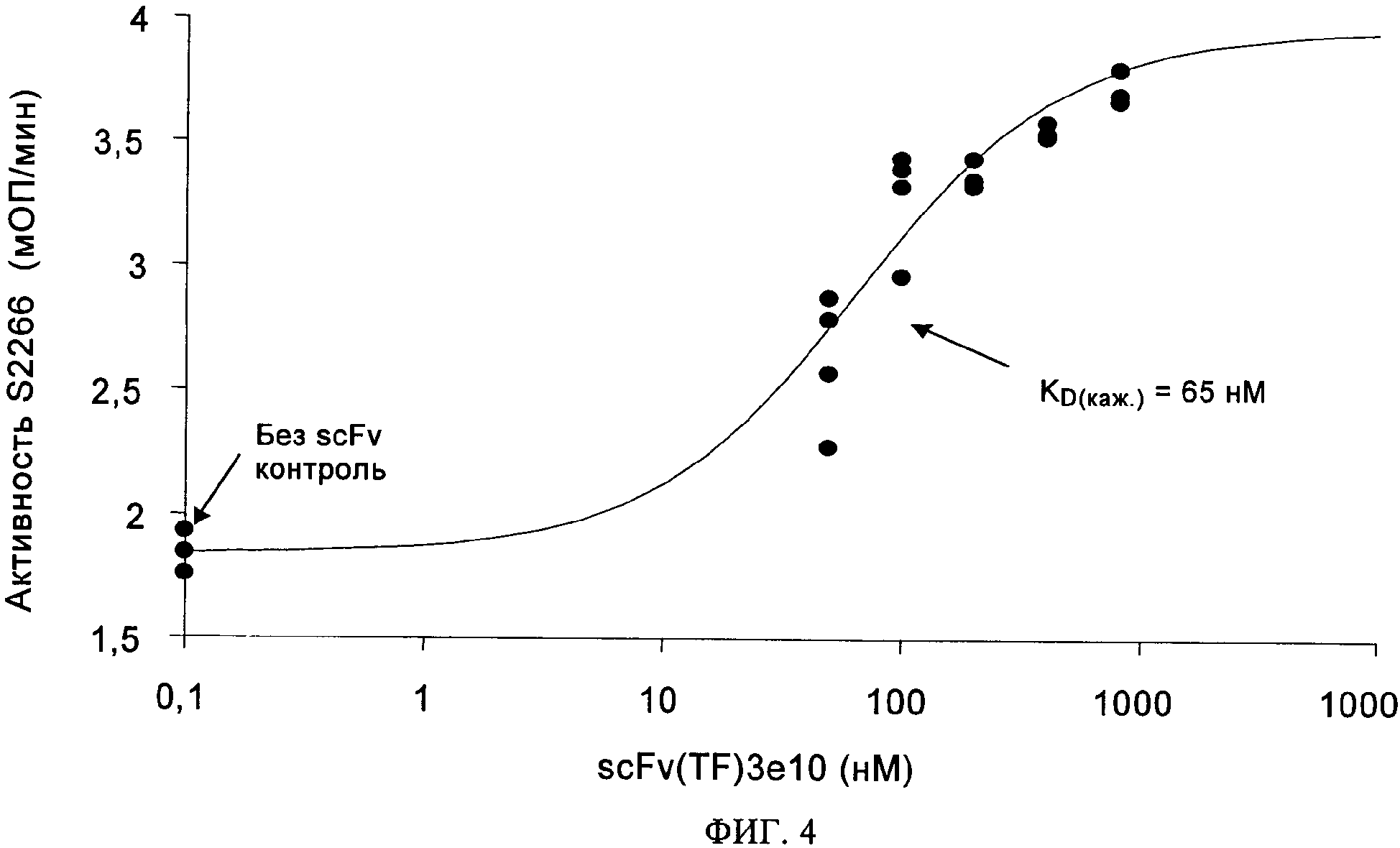
на фиг.4 - данные о кажущейся аффинности к связыванию scFv(TF)3e10 с рТФ. Анализ способности рТФ/FVIIa осуществлять пептидный гидролиз проводили согласно методу, описанному в примере 4, с применением 3 нМ рТФ и 2 нМ FVIIa. Применяемая концентрация рТФ была ниже значения kD, характеризующей связывание с FVIIa. Вносили возрастающие концентрации экспрессируемого в бактериях scFv(TF)3e10 и возрастающую скорость реакции использовали для определения кажущихся значений kD, характеризующих связывание этого антитела с рТФ, с помощью стандартной аппроксимации 4-параметрической зависимости. Аффинность scFv(TF)3e10 к комплексу рТФ / FVIIa (КD(арр)=65 нМ) оказалась выше, чем аффинность scFv(TF)3e10 к рТФ при измерении с помощью BIAcore (КD(арр)=470 нМ);
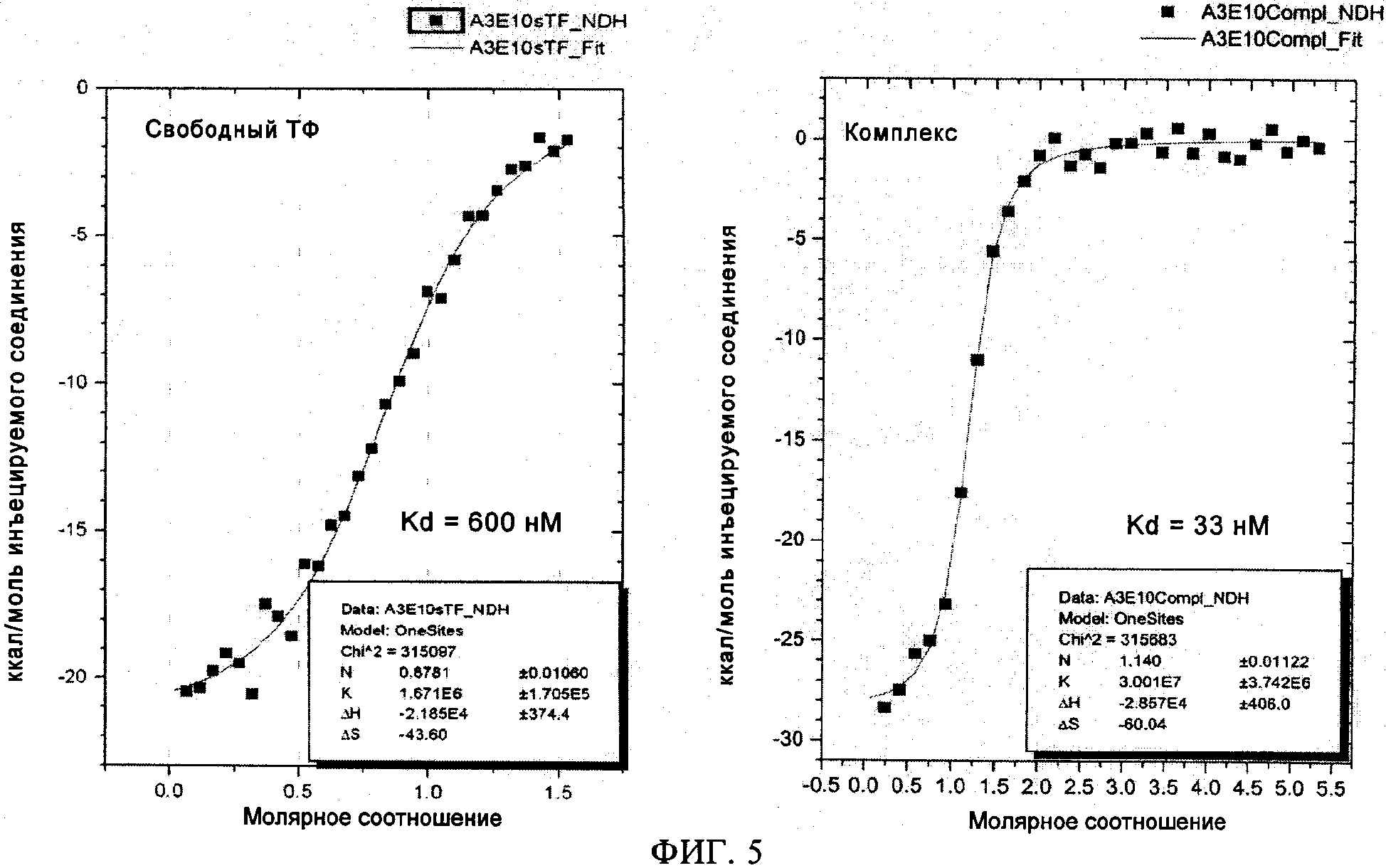
на фиг.5 - данные об аффинности к связыванию scFv(TF)3e10 с ТФ и комплексом FVIIa/ТФ. Микрокалориметрический анализ осуществляли согласно методу, описанному в примере 4, используя антитело scFv(TF)3e10, экспрессируемое в СНО-клетках. Этот анализ показал, что scFv(TF)3e10 обладает ˜ в 20 раз более высокой аффинностью к связыванию с комплексом рТФ/FVIIa ("комплекс"), чем к рТФ ("свободный ТФ");
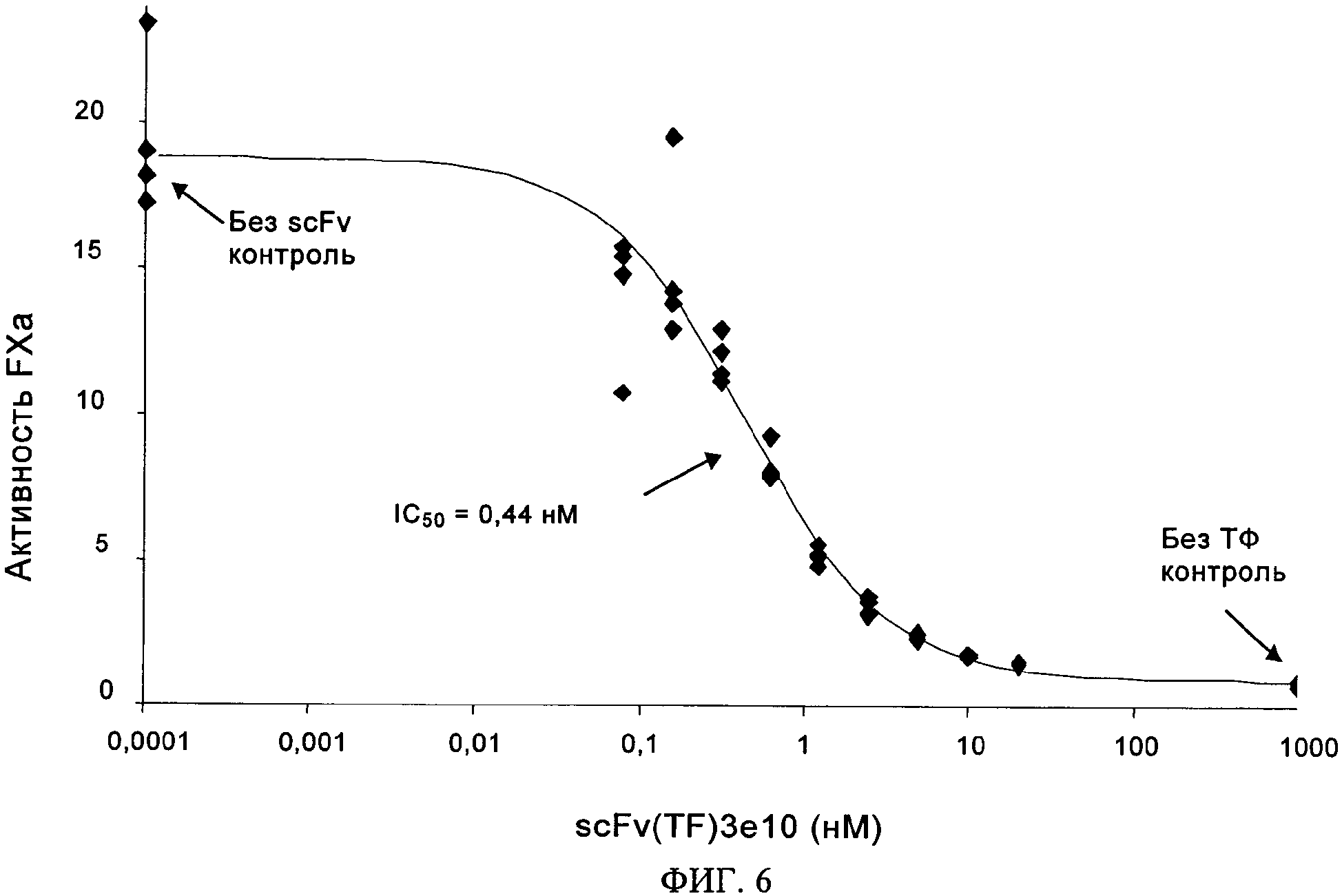
на фиг.6 - данные о зависящей от дозы способности scFv(TF)3e10 ингибировать активацию FX. Анализ активации FX осуществляли с помощью метода, описанного в примере 4, используя возрастающие концентрации экспрессируемого в бактериях scFv(TF)3e10, 250 нМ FX и комплекс FVIIa/ТФ на поверхности фосфолипидов (10 пМ FVIIa). Значение IC50 представляет собой дозу, необходимую для достижения 50%-ного ингибирования;
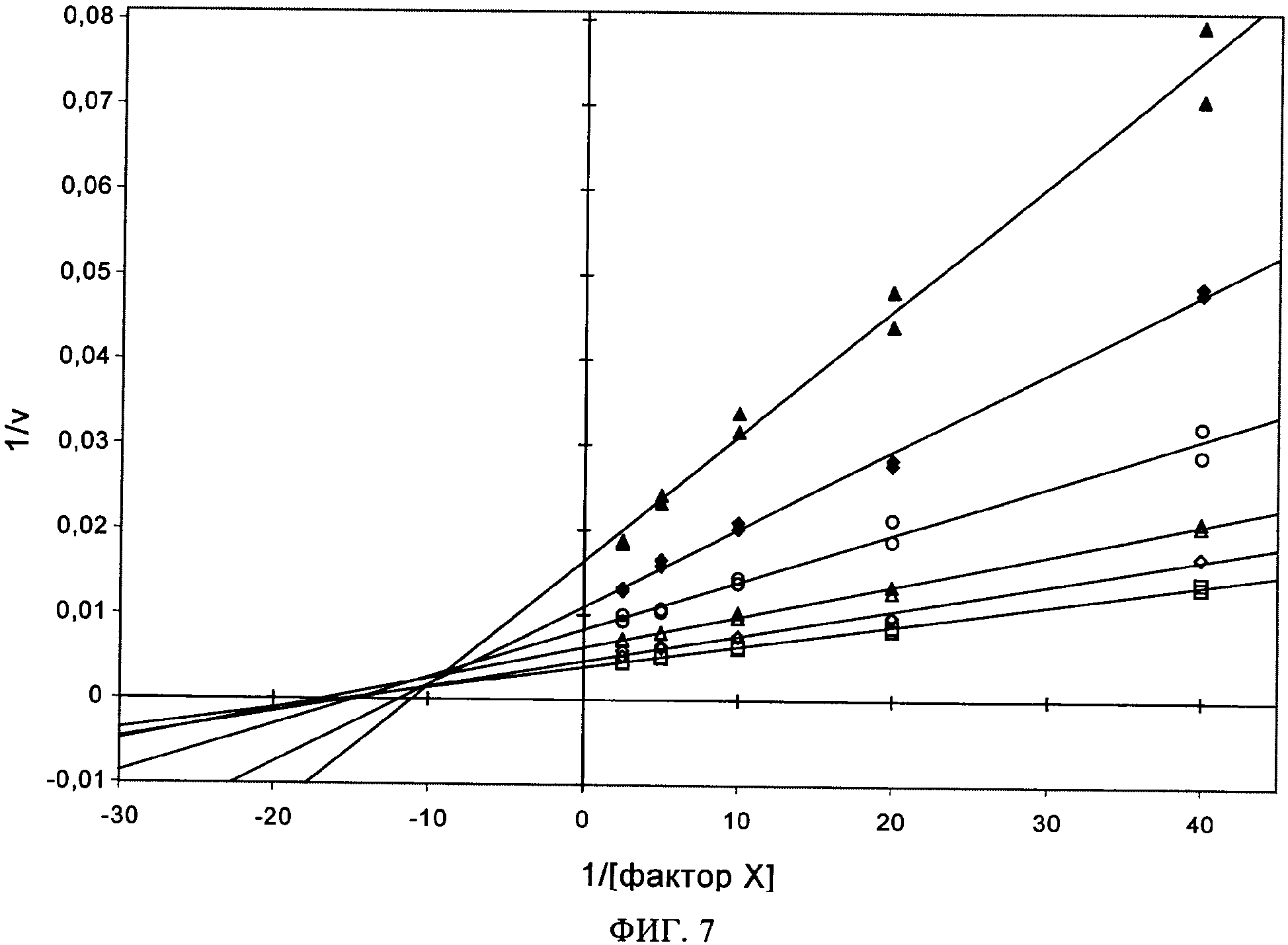
на фиг.7 - данные, свидетельствующие о том, что scFv(TF)3e10 неконкурентно ингибирует активацию FX с помощью комплекса FVIIa/ТФ. Анализ активации FX осуществляли согласно методу, описанному в примере 4, используя экспрессируемое в бактерии scFv(TF)3e10, 25-400 нМ FX и комплекс FVIIa/ТФ на поверхности фофолипида (10 пМ FVIIa). В этом анализе осуществляли титрование с использованием возрастающих концентраций scFv(TF)3e10 (0 нМ, незакрашенный квадрат; 0,25 нМ, незакрашенный ромб; 0,74 нМ, незакрашенный треугольник, 2,2 нМ, незакрашенный круг; 6,7 нМ, закрашенный ромб; 20 нМ, закрашенный треугольник). Представленный график зависимости Lineweaver-Burk (1/[S], где S (субстрат) обозначает фактор Х (мкМ); относительно 1/v, где v (скорость) выражают в виде мОП/мин, характеризующей гидролиз S2222 с помощью FXa, происходящий в течение 5 мин) свидетельствует о том, что scFv(TF)3e10 является неконкурентным ингибитором в отношении субстрата, т.е. FX. Все кривые пересекаются на (или вблизи) оси абсцисс, что характерно для неконкурентного ингибирования (при конкурентном ингибировании все кривые должны пересекаться на оси ординат);
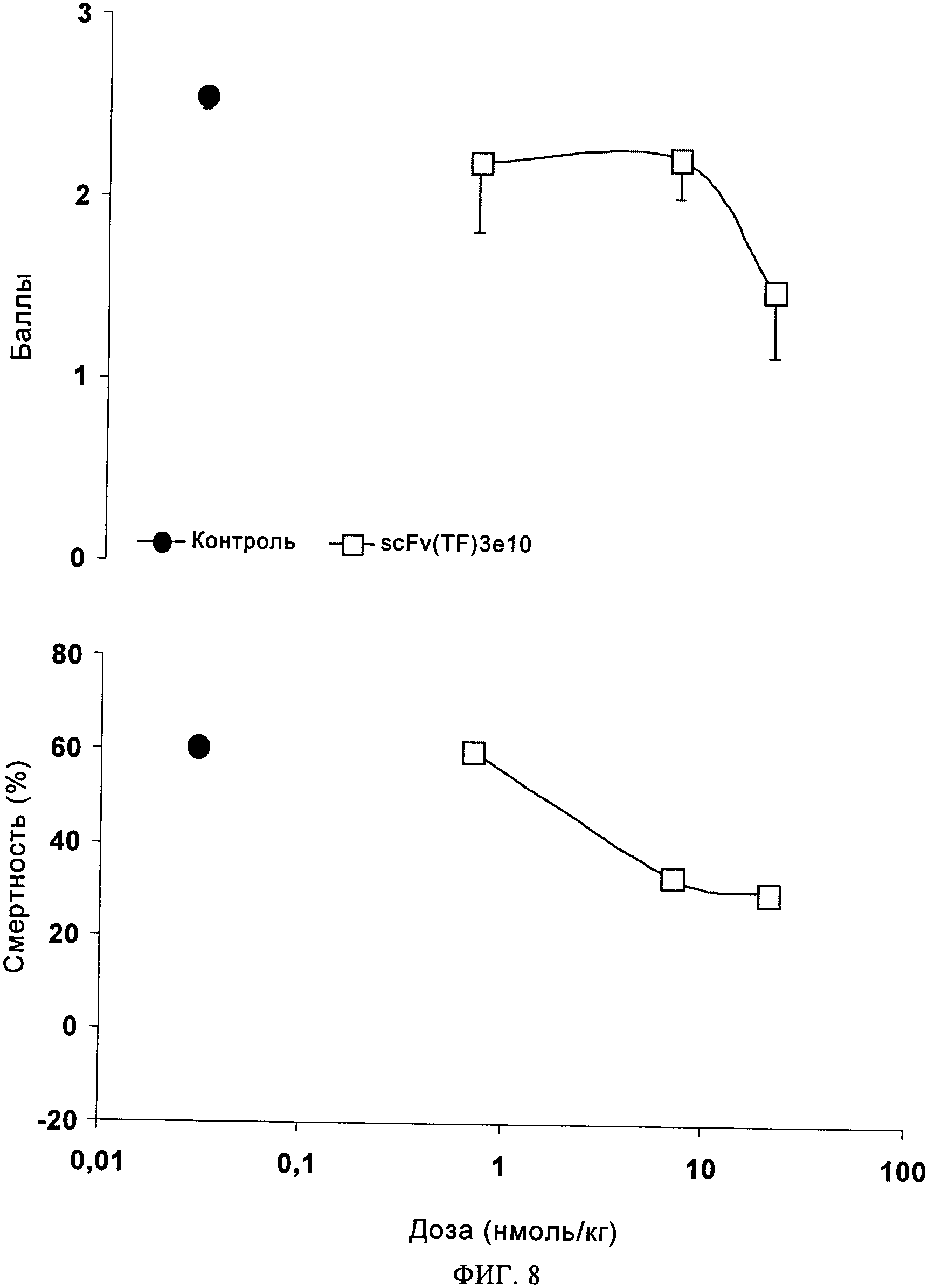
на фиг.8 - данные, подтверждающие эффективность scFv(TF)3e10 на модели in vivo диссеминированного внутрисосудистого свертывания крови ("ДВС"). Антитело к ТФ scFV(TF)3e10, экспрессируемое в СНО-клетках, оценивали с использованием модели тромбоэмболии, индуцированной у крыс, которая описана в примере 6, на основе (А) процента смертности и (Б) показателя заболеваемости-смертности. (А) Для обработанной носителем группы применяемая доза ТФ вызывала 60%-ную смертность (LD60). Антитело scFv(TF)3e10 в дозе 0,7 нмоль/кг не оказывало воздействия на уровень смертности, но в дозе 7,0 нмоль/кг снижало смертность до уровня <40%. (Б) В обработанной носителем группе in vivo применяемая доза ТФ приводила к получению среднего показателя заболеваемости-смертности 2,6. Антитело scFv(TF)3e10 в дозе 0,7 нмоль/кг не оказывало воздействия на смертность и оказывало небольшое воздействие на респираторный дистресс-синдром или не оказывало его вообще, но при использовании в дозе 14 нмоль/кг снижало средний показатель заболеваемости-смертности до ˜1,5.
Подробное описание изобретения
Антикоагулирующее антитело, предлагаемое в настоящем изобретении, представляет собой антитело, которое связывается с большей аффинностью с комплексом фактор VIIa/тканевый фактор ("FVIIa/ТФ"), чем со свободным тканевым фактором ("ТФ"). Антитело, предлагаемое в изобретении, обладает по меньшей мере в 2 раза большей аффинностью, предпочтительно по меньшей мере в 5 раз большей аффинностью и более предпочтительно по меньшей мере в 10 раз большей аффинностью к связыванию с комплексом FVIIa/ТФ, чем со свободным ТФ. Кроме того, антитело, предлагаемое в изобретении, не конкурирует за связывание с ТФ с одним или несколькими факторами свертывания, выбранными из ряда, включающего фактор VII ("FVII"), фактор IX ("FIX") и фактор Х ("FX"). Предпочтительно антитело, предлагаемое в изобретении, не конкурирует за связывание с ТФ с FVII и с FX.
Определения
В настоящем описании следующие понятия имеют указанные ниже значения.
Понятие "рекомбинантные белки или полипептиды" относится к белкам или полипептидам, полученным с помощью рекомбинантной ДНК, т.е. полученным из клеток, микроорганизмов или млекопитающих, трансформированных экзогенной конструкцией ДНК, которая кодирует требуемый полипептид. Белки или полипептиды, экспрессируемые в большинстве бактериальных культур, должны быть свободны от гликана. Белки или полипептиды, экспрессируемые в дрожжах, могут иметь схему гликозилирования, отличную от схемы гликозилирования экспрессируемых в клетках млекопитающих белков или полипептидов.
Понятие "нативные" белки или полипептиды относится к белкам или полипептидам, выделенным из встречающегося в естественных условиях источника. Понятие "нативное антитело" включает встречающиеся в естественных источниках антитела и их фрагменты. Понятие "кодирующая последовательность" ДНК обозначает последовательность ДНК, в результате транскрипции которой синтезируется мРНК и которая транслируется в полипептид в клетке-хозяине, если она помещена так, что находится под контролем соответствующих регуляторных последовательностей. Границы кодирующей последовательности определены стартовым кодоном на 5'N-конце и стоп-кодоном трансляции на 3'С-конце. Кодирующая последовательность может включать прокариотические последовательности, кДНК, полученную на основе эукариотической мРНК, геномные последовательности ДНК из эукариотической ДНК и синтетические последовательности ДНК. Последовательность терминатора транскрипции, как правило, должна быть локализована на 3'-конце относительно кодирующей последовательности.
Понятие "нуклеотидная последовательность" обозначает полимер дезооксирибонуклеотидов (основания аденин, гуанин, тимин или цитозин). Последовательности ДНК, которые кодируют антитела, предлагаемые в настоящем изобретении, могут быть собраны из полученных из синтетической кДНК ДНК-фрагментов и коротких олигонуклеотидных линкеров для создания синтетического гена, который обладает способностью экспрессироваться в рекомбинантном экспрессионном векторе. При обсуждении структуры конкретных двухцепочечных молекул ДНК в настоящем описании подразумевается общепринятое обозначение последовательностей только в направлении 5'→3' нетранскрибируемой цепи кДНК.
Понятие "рекомбинантный экспрессионный вектор" обозначает способную к репликации конструкцию ДНК, применяемую либо для амплификации, либо для экспрессии ДНК, которая кодирует антитела, предлагаемые в настоящем изобретении. Экспрессионный вектор содержит контролирующие последовательности и кодирующую последовательность ДНК. Контролирующие последовательности ДНК включают промоторные последовательности, сайты связывания рибосом, сигналы полиаденилирования, последовательности терминаторов транскрипции, расположенные против хода транскрипции регуляторные домены и энхансеры. Представленные в описании рекомбинантные экспрессионные системы должны экспрессировать антитела после индукции регуляторных элементов.
Понятие "трансформированные клетки-хозяева" относится к клеткам, трансформированным и трансфектированным экзогенной ДНК. Экзогенная ДНК может не быть интегрирована или может быть интегрирована (ковалентно связана) в хромосомную ДНК, дополняя геном клетки. У прокариот и дрожжей, например, экзогенная ДНК может поддерживаться на эписомальном элементе, таком как плазмида, или быть стабильно интегрирована в хромосомную ДНК. Что касается эукариотических клеток, то стабильно трансформированная клетка представляет собой клетку, в которой экзогенная ДНК интегрирована в репликативную хромосому. Стабильность можно продемонстрировать по способности линий или клонов эукариотических клеток продуцировать популяцию дочерних клеток, содержащих экзогенную ДНК.
Понятие "аналог", "фрагмент", "производное" и "вариант" применительно к антителам, предлагаемым в изобретении, обозначает аналоги, фрагменты, производные и варианты антител, которые сохраняют практически такую же биологическую функцию или активность и которые будут более подробно описаны ниже.
Понятие "аналог" включает прополипептид, в состав которого входит аминокислотная последовательность антитела, предлагаемого в изобретении. Активное антитело, предлагаемое в изобретении, можно отщеплять от дополнительных аминокислот, которые входят в состав молекулы проантитела, с помощью естественных процессов in vivo или с помощью методов, хорошо известных в данной области, таких как ферментативное или химическое расщепление. Например, рекомбинантный полипептид scFV(TF)3e10 (SEQ ID NO:1) экспрессируется в виде состоящего из 282 аминокислот прополипептида, который процессируется in vivo с высвобождением активного зрелого полипептида, состоящего из 264 аминокислот.
Понятие "фрагмент" обозначает часть антитела, предлагаемого в изобретении, которая сохраняет практически такую же функциональную активность, которая выявлена в анализах in vitro, более подробно описанных ниже.
Понятие "производное" включает все модификации антител, предлагаемых в изобретении, которые в основном сохраняют требуемые функции и несут дополнительную структуру и обладают сопутствующей функцией, например ПЭГилированные антитела, которые имеют более продолжительное время полужизни, и биотинилированные антитела, которые дополнительно будут описаны ниже. Производное включает также N- или O-связанные гликозилированные антитела, которые можно получать путем встраивания сайтов N- или O-гликозилирования в последовательности антител с помощью стандартных методов рекомбинантной ДНК. Понятия "практически такая же функциональная активность" и "практические такая же биологическая функция или активность" каждое обозначает, что уровень биологической активности составляет примерно 30-100% или более от биологической активности, свойственной полипептиду, с которым проводят сравнение, если биологическую активность каждого полипептида определяют с помощью одинаковых метода или анализа. Например, можно говорить, что антитело имеет практически такую же функциональную активность, что и антитело из примера 1 (SEQ ID NO:1), если оно при оценке пептидного гидролиза с помощью рТФ/FVIIa и активации FX с использованием анализов, которые описаны в примере 4, демонстрирует способность связываться и нейтрализовать комплекс FVIIa/ТФ.
"Сходство" между двумя полипептидами определяют путем сравнения аминокислотной последовательности и консервативных аминокислотных замен одного полипептида с последовательностью второго полипептида. Такие консервативные замены включают замены, описанные выше в The Atlas of Protein Sequence and Structure 5 by Dayhoff (1978) и у Argos EMBO J. 8, 1989, сс.779-785. Например, аминокислоты, принадлежащие к одной из следующих групп, представляют собой консервативные замены:
- Ala, Pro, Gly, Gln, Asn, Ser, Thr;
- Cys, Ser, Tyr, Thr;
- Val, Ile, Leu, Met, Ala, Phe;
- Lys, Arg, His;
- Phe, Tyr, Trp, His; и
- Asp, Glu.
Понятие "антитело" в контексте настоящего описания включает интактные молекулы иммуноглобулинов ("Ig"), а также их фрагменты, такие как Fab-, F(ab')2- и Fv-фрагменты, которые обладают способностью связываться с эпитопом выбранного белка-мишени, например растворимого ТФ ("рТФ"). Как правило, для образования эпитопа требуется по меньшей мере 6, 8, 10 или 12 смежных аминокислот. Однако для образования эпитопов, которые включают не смежные аминокислоты, может требоваться большее количество, например по меньшей мере 15, 25 или 50 аминокислот.
Все другие технические понятия, применяемые в настоящем описании, имеют значения, которые обычно применяют специалисты в данной области.
Антитела, предлагаемые в изобретении, и их получение
Антикоагулирующие антитела, предлагаемые в настоящем изобретении, связываются с большей аффинностью с комплексом фактор VIIa/тканевый фактор ("FVIIa/ТФ"), чем со свободным тканевым фактором ("ТФ"). Согласно предпочтительному варианту осуществления изобретения антитела, предлагаемые в изобретении, обладают по меньшей мере в 2 раза большей аффинностью, более предпочтительно по меньшей мере в 5 раз большей аффинностью и еще более предпочтительно по меньшей мере в 10 раз большей аффинностью к связыванию с комплексом FVIIa/ТФ, чем со свободным ТФ при оценке с помощью микрокалориметрического анализа. Согласно другому предпочтительному варианту осуществления изобретения антитела также не конкурируют за связывание с ТФ с одним или несколькими факторами свертывания, выбранными из ряда, включающего фактор VII ("FVII"), фактор IX ("FIX") и фактор Х ("FX"). Согласно наиболее предпочтительному варианту осуществления изобретения антитела, предлагаемые в изобретении, не конкурируют за связывание с ТФ с FVII и с FX. Согласно наиболее предпочтительному варианту осуществления изобретения антитела, предлагаемые в изобретении, связываются с большей аффинностью с комплексом FVIIa/ТФ, чем со свободным ТФ, и не конкурируют за связывание с ТФ с FVII и с FX.
В целом, антитело, которое специфично связывается с выбранным белком-мишенью (например, комплексом FVIIa/ТФ или ТФ), при оценке с помощью иммунохимического анализа дает сигнал обнаружения по меньшей мере в 5, 10 или 20 раз более высокий, чем сигнал обнаружения других белков. Предпочтительно антитела, которые специфично связываются с выбранным белком-мишенью, не обнаруживают другие белки при оценке с помощью иммунохимических анализов и могут осуществлять иммунопреципитацию белка-мишени из раствора.
Выбранный белок-мишень можно использовать для иммунизации млекопитающего, такого как мышь, кролик, морская свинка, обезьяна или человек, для продуцирования поликлональных антител. При необходимости белок-мишень можно конъюгировать с белком-носителем, таким как бычий сывороточный альбумин, тироглобулин и гемоцианин лимфы улитки. В зависимости от видов хозяев для повышения иммунологического ответа можно использовать различные адъюванты. Такие адъюванты включают (но не ограничиваясь ими) адъювант Фрейнда, минеральные гели (например, гидроксид алюминия) и поверхностно-активные вещества (например, лизолецитин, полиолы плюроника, полианионы, пептиды, масляные эмульсии, гемоцианин лимфы улитки и динитрофенол). Из применяемых для человека адъювантов особенно предпочтительными являются БЦЖ (бацилла Кальмета-Герена) и Cornybacterium parvum.
Моноклональные антитела, которые специфично связываются с выбранным белком-мишенью, можно получать с помощью любого метода, который обеспечивает получение молекул антител с помощью стабильных клеточных линий в культуре. Эти методы включают (но не ограничиваясь ими) метод на основе гибридом, метод на основе человеческой В-клеточной гибридомы и метод на основе EBV-гибридомы (Kohler и др. Nature 256, 1985, сс.495-497; Kozbor и др., J. Immunol. Methods 81, 1985, сс.31-42; Cote и др. Proc. Natl. Acad. Sci. USA 80, 1983, сс.2026-2030; и Cote и др., Mol. Cell Biol. 62, 1984, сс.109-120).
Кроме того, можно применять методы, разработанные для получения "химерных антител", сплайсинга мышиных генов антител и человеческих генов антител для получения молекулы с соответствующей антигенной специфичностью и биологической активностью (Morrison и др., Proc. Natl. Acad. Sci. USA 81, 1984, сс.6851-6855; Neuberger и др. Nature 312, 1984, сс.604-608; Takeda и др. Nature 314, 1985, сс.452-454). Моноклональные и другие антитела могут также представлять собой "гуманизированные" антитела, предназначенные для предупреждения у пациента иммунного ответа на антитело при его применении в качестве терапевтического агента. Для непосредственного применения последовательность таких антител может обладать достаточным сходством с последовательностью человеческих антител или может нуждаться в изменении нескольких имеющих решающее значение остатков. Различия в последовательности антител грызунов и человеческих последовательностей можно минимизировать, заменяя остатки, которые отличаются от остатков в человеческих последовательностях, путем сайтнаправленного мутагенеза или путем трансплантации полных гибервариабельных участков. В другом варианте гуманизированные антитела можно получать с помощью методов рекомбинации, описанных в GB 2188638B. Антитела, которые специфически связываются с выбранным белком-мишенью, могут содержать антигенсвязывающие центры, которые являются либо частично, либо полностью гуманизированными, как описано в U.S. 5565332.
В другом варианте методики, предназначенные для получения одноцепочечных антител, можно адаптировать с помощью известных в данной области методов для получения одноцепочечных антител ("scFv"), которые специфично связываются с выбранным белком-мишенью. Антитела с родственной специфичностью, но имеющие другой идиотипический состав, можно получать путем перестановки цепей из случайных комбинаторных библиотек Ig (Burton, Proc. Natl. Acad. Sci. USA, 88, 1991, сс.11120-11123).
Одноцепочечные антитела можно также конструировать с помощью метода амплификации ДНК, такого как ПЦР, используя кДНК гибридомы в качестве матрицы (Thirion и др., Eur. J. Cancer Prev. 5, 1996, сс.507-511). Одноцепочечные антитела могут быть моно- или биспецифическими и могут быть бивалентными или тривалентными. Конструирование тетравалентных биспецифических одноцепочечных антител описано, например, у Coloma и Morrison, Natl. Biotechnol. 15, 1997, сс.159-163. Конструирование бивалентных биспецифических одноцепочечных антител описано у Mallendar и Voss, J. Biol. Chem., 269, 1994, сс.199-216.
Нуклеотидную последовательность, кодирующую одноцепочечное антитело, можно конструировать с помощью ручного или автоматического синтеза нуклеотидов, клонирования в предназначенной для экспрессии конструкции с помощью стандартных методов рекомбинантной ДНК и интродукции в клетку для экспрессии кодирующей последовательности. В другом варианте одноцепочечные антитела можно получать непосредственно, применяя, например, технологию на основе дисплейной библиотеки нитчатого фага (Verhaar и др. Int. J. Cancer 61, 1995, сс.497-501; и Nicholls и др., J. Immunol. Meth. 165, 1993, сс.81-91).
Антитела, которые специфично связываются с выбранным белком-мишенью, можно получать также путем индукции in vivo производства популяции лимфоцитов или путем скрининга библиотек Ig или панелей связывающихся с высокой специфичностью реагентов согласно описанным в литературе методам (Orlandi и др., Proc. Natl. Acad. Sci. USA 86, 1989, сс.3833-3837; Winter и др., Nature 349, 1991, сс.293-299).
ДНК, которая клонирует антитело, предлагаемое в изобретении, можно клонировать в форме кДНК или геномной ДНК с помощью любой процедуры клонирования, известной специалисту в данной области (см., например, Sambrook, J.F. и др. Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (1989), публикация включена в настоящее описание в качестве ссылки).
В случае, когда антитело представляет собой моноклональное антитело, то если идентифицирована последовательность ДНК, кодирующая Fv-фрагмент, который при экспрессии обладает способностью к специфическому связыванию, то антитела, содержащие этот Fv-фрагмент, можно получать с помощью методов, хорошо известных специалисту в данной области. Так, например, у Chaudhary V.K. и др. Nature, 339(6223), 1989, сс.394-397; Batra J.K. и др., J. Biol. Chem. 265(25), 1990, сс.15198-15202; Batra J.K. и др., Proc. Natl. Acad. Sci. USA 86(21), 1989, сс.8545-8549; Chaudhary V.K. и др., Proc. Natl. Acad. Sci. USA 87(3), 1990, сс.1066-1070 (все публикации включены в настоящее описание в качестве ссылки), описано получение белков различных одноцепочечных антител.
Согласно предпочтительному варианту ТФ-связывающие антитела, предлагаемые в изобретении, представляют собой одноцепочечные антитела, которые получают с помощью фаговой дисплейной библиотеки. Конструируют эпитопсвязывающий центр одноцепочечного антитела, состоящий из двух доменов вариабельной области: один из тяжелой цепи, а второй из легкой цепи. На первой стадии конструирования фаговой дисплейной библиотеки гены вариабельных цепей (VH (из IgM), Vκ и VL) клонируют с помощью ПЦР из пулированной мРНК из человеческого костного мозга, лимфатического узла и селезенки с использованием набора специфических для семейства праймеров. Полученные библиотеки pCITE-VН (3,8×109 членов), pZ604-Vκ (1,6×107) и pZ604-VL (3,2×107) представляют собой библиотеки постоянных и имеющих широкое разнообразие V-генов. Затем VH-гены амплифицируют из библиотеки pCITE-VH. С помощью ПЦР из библиотек pZ604-Vκ и pZ604-VL амплифицируют Vκ- и VL-гены, несущие обратную JН-область и линкерную последовательность на 5'-конце. Затем очищенные на геле содержащие VH, Vκ и VL ПЦР-продукты лигируют друг с другом, получая набор генов scFv. Набор генов scFv клонируют в фагмидном векторе pZ603 и полученный в результате лигирования продукт вводят путем электропорации в компетентные клетки Е.coli линии TG1, создавая фаговую дисплейную библиотеку scFv, обозначенную HuPhabL3, которая несет 5,2×109 индивидуальных трансформантов (Кау В.К. и др. (1996) Phage Display of Peptides and Proteins: A Laboratory Manual, изд-во Academic Press, San Diego CA; Marks J.D. и др. J. Mol. Biol. 222(3), 1991, сс.581-597; Sheets M.D. и др., Proc. Natl. Acad. Sci. USA 95(11), 1998, сс.6157-6162).
ТФ-связывающий фаг из фаговой дисплейной библиотеки scFv отбирают, амплифицируют и затем идентифицируют с помощью методов пэннинга, хорошо известных в данной области. Растворимый ТФ иммобилизуют на пластиковых пробирках, при этом для уменьшения неспецифического связывания с пластиком можно использовать обезжиренное молоко. Популяцию фага, экспрессирующего scFv (scFv-фага), наносят на иммобилизованный рТФ в пластиковые пробирки и несвязанный фаг удаляют путем интенсивной отмывки. ТФ-связывающий scFv-фаг элюируют из пробирок и затем амплифицируют, заражая клетки Е.coli TG1 в растворе. Указанную процедуру пэннинга повторяют трижды и полученный ТФ-связывающий scFv-фаг выделяют путем трансформации клеток линии TG1. Трансформанты, которые экспрессируют ТФ-связывающие антитела, идентифицируют с помощью стандартного метода ELISA, для осуществления которого используют рТФ, иммобилизованный на пластиковых 96-луночных планшетах. Секвенируют ДНК-вставки одноцепочечного антитела позитивных по данным ELISA трансформантов. На основе секвенирования выявлено 6 уникальных одноцепочечных антител, т.е. scFv(TF)2c1, scFv(TF)2c11, scFv(TF)2d3, scFv(TF)2h6, scFv(TF)3e10 и scFv(TF)3h2, которые экспрессируют и выделяют из E.coli и характеристики которых описаны ниже в примере 5.
Экспрессия и очистки антител, предлагаемых в изобретении
Существует несколько путей экспрессии in vitro рекомбинантных антител, предлагаемых в изобретении, включая Е.coli, бакуловирусы, клетки дрожжей и млекопитающих или другие системы экспрессии. Методы экспрессии клонированных генов в бактериях хорошо известны. Для получения высокого уровня экспрессии клонированного гена в прокариотической системе важную роль играет конструирование экспрессионных векторов, которые содержат, как минимум, сильный промотор для обеспечения терминации транскрипции мРНК. Примерами регуляторных областей, которые можно применять для этой цели, являются промоторная и операторная область гена бета-глюкозидазы Е.coli, гены пути биосинтеза триптофана Е.coli или "обратный" промотор фага лямбда. Можно включать селектируемые маркеры в векторные ДНК, которыми трансформируют Е.coli. Примерами таких маркеров являются гены, обеспечивающие устойчивость к ампициллину, тетрациклину или хлорамфениколу.
Для экспрессии антител, предлагаемых в изобретении, и их аналогов, фрагментов, производных или вариантов можно применять системы на основе клеток высших эукариотических организмов, которые можно выбирать из многочисленных клеточных систем. Приведенными с целью иллюстрации примерами клеточных линий млекопитающих являются (но не ограничиваясь ими) клетки RPMI 7932, VERO и HeLa, линии клеток яичника китайского хомячка (СНО), WI38, ВНК, COS-7, С127 или MDCK. Предпочтительной линией клеток млекопитающих является CHL-1. Когда применяют линию CHL-1, то в качестве маркера селекции для эукариотических клеток используют гигромицин. CHL-1-клетки получают из клеток меланомы линии RPMI 7032, которая представляет собой легко доступную человеческую линию клеток. Линия клеток CHL-1 в соответствии с Будапештским договором депонирована в АТСС 18 июня 1987 г. под регистрационным номером CRL 9446. Клетки, которые можно применять согласно настоящему изобретению, можно получать из АТСС. В качестве иллюстративных примеров клеток можно привести клетки насекомых Spodoptera frugiperda и Bombyx mori.
В прокариотической системе, такой как Е.coli, нельзя осуществлять посттрансляционную модификацию, такую как гликозилирование. Кроме того, белки со сложной системой дисульфидных мостиков часто имеют ошибки в укладке при экспрессии в Е.coli. При применении прокариотической системы экспрессируемый белок либо присутствует в клеточной цитоплазме в нерастворенной форме в виде так называемых внутриклеточных телец, которые находятся в растворимой фракции после лизиса клеток, либо его переносят в периплазму, добавляя соответствующие секреторные сигнальные последовательности. Если экспрессируемый белок находится в нерастворимых внутриклеточных тельцах, то, как правило, требуется солюбилизация и последующая повторная укладка внутриклеточных телец.
Специалистам в данной области известен целый ряд поступающих в продажу прокариотических экспрессионных векторов, таких как рКК223-3 (фирма Pharmacia Fine Chemicals, Уппсала, Швеция), рКК233-2 (фирма Clontech, Пало-Альто, шт. Калифорния, США) и pGEMl (фирма Promega Biotech, Мэдисон, шт. Висконсин, США).
Промоторы, которые обычно применяют в рекомбинантных экспрессионных системах на основе микроорганизмов, включают промоторные системы бета-лактамазы (пенициллиназы) и лактозы (Chang A.C. и др.. Nature, 275(5681), 1978, сс.617-624; Goeddel D.V. и др. Nature 281(5732), 1979, сс.544-548), промоторную систему триптофана (trp) (Goeddel D.V. и др., Nucl. Acids Res. 8(18), 1980, сс.4057-4074) и промотор tac (Maniatis T. и др. Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (1982)). В других пригодных бактериальных системах экспрессии применяют промотор фага лямбда pL и термоиндуцируемый репрессор cIts857 (Bernard H.U. и др.. Gene, 5(1), 1979, сс.59-76; Love C.A. и др. Gene 176(1-2), 1996, сс.49-53). Рекомбинантные антитела можно экспрессировать также в хозяевах-дрожжах, таких как Saccharomyces cerevisiae. Это, как правило, позволяет осуществлять различные посттрансляционные модификации. Экспрессируемое антитело можно секретировать в супернатант культуры, в котором не присутствуют остатки многих других белков, что упрощает очистку. Векторы на основе дрожжей, предназначенные для экспрессии антител, предлагаемых в изобретении, несут определенные требуемые структуры. Элементы вектора, как правило, получают из дрожжей и бактерий для осуществления размножения плазмиды в обоих типах клеток. Бактериальные элементы включают сайт инициации репликации и селектируемый маркер. Элементы дрожжей включают последовательность сайта инициации репликации (ARS), селектируемый маркер, промотор и терминатор транскриции.
Пригодными промоторами в экспрессионных векторах на основе дрожжей являются промоторы гена TRP1, гена ADH1 или ADHI, гена кислой фосфатазы (РН03 или РН05), гена изоцитохрома или промоторы, участвующие в гликолизе, такие как промотор энолазы, глицеральдегид-3-фосфат-дегидрогеназы (GADPH), 3-фосфоглицераткиназы (PGK), гексокиназы, пируваткиназы, триозофосфатизомеразы и глюкозофосфатизомеразы (Hitzeman R.A. и др., J. Biol. Chem. 255(24), 1980, сс.12073-12080; Hess В. и др., J. Adv. Enzyme Reg. 7, 1968, сс.149-167; и Holland M.J. и Holland, J.P., Biochemistry 17(23), 1978, сс.4900-4907).
Поступающие в продажу векторы на основе дрожжей включают pYES2, рРIС9 (фирма Invitrogen, Сан-Диего, шт. Калифорния), Yepc-pADH2a, pYcDE-1 (Washington Research, Сиэтл, шт. Вашингтон), рВС102-К22 (АТСС №67255) и YpGX265GAL4 (АТСС №67233). Линии клеток млекопитающих, которые можно применять для экспрессии рекомбинантных антител, предлагаемых в изобретении, включают (но не ограничиваясь ими) COS-7, L-клетки, С127, 3Т3, клетки яичника китайского хомячка (СНО), HeLa, BHK, CHL-1, NSO и НЕК293. Рекомбинантные белки, продуцируемые в клетках млекопитающих, как правило, являются растворимыми и гликозилированными и имеют аутентичные N-концы. Экспрессионные векторы на основе клеток млекопитающих могут содержать нетранскрибируемые элементы, такие как сайт инициации репликации, промотор и энхансер и 5'- или 3'-нетранслируемые последовательности, такие сайты связывания рибосом, сайт полиаденилирования, акцепторный сайт и донор для сплайсинга, и последовательности терминаторов транскрипции. Примерами промоторов, которые обычно применяют в экспрессионных векторах на основе клеток млекопитающих, являются вирусные промоторы, такие как промоторы вируса полиомы, аденовируса, вируса Т-клеточного лейкоза (HTLV), обезьяньего вируса 40 (OB 40) и человеческого цитомегаловируса (CMV).
В зависимости от выбранных системы экспрессии и хозяина гомогенное рекомбинантное антитело можно получать с помощью различных сочетаний общепринятых методов хроматографии, применяемых для очистки белков. Они включают иммуноаффинную хроматографию, хроматографию с обращенной фазой, катионообменную хроматографию, анионообменную хроматографию, хроматографию на основе гидрофобных взаимодействий, гель-фильтрацию и ЖХВР. Если система экспрессии секретирует антитело в питательные среды, то белок можно очищать непосредственно из сред. Если не происходит секреция антитела, то его выделяют из клеточных лизатов. Разрушение клеток можно осуществлять с помощью любого общепринятого метода, включая циклы замораживания-оттаивания, облучение ультразвуком, механическое разрушение или применение лизирующих клетки агентов.
Для экспрессии в бактериях одноцепочечного антитела, предлагаемого в изобретении, применяют плазмидную конструкцию на основе pCANTAB5 (фирма Pharmacia). Например, плазмида, содержащая одноцепочечное антитело scFv(TF)3e10, представляет собой плазмиду pZ612/3e10, и она кодирует последовательность одноцепочечного антитела, за которой расположена последовательность метки e-tag, применяемой для очистки белка. Аминокислотная последовательность антитела scFV(TF)3e10 соответствует SEQ ID NO:1 (пример 1), а последовательность ДНК, которая кодирует scFv(TF)3e10 соответствует SEQ ID NO:2.
Плазмидную конструкцию, основой которой является pTHR525 (см. US 5827824), используют для экспрессии одноцепочечных антител, предлагаемых в настоящем изобретении, в клетках млекопитающих. Например, конструируют праймеры для создания ДНК-фрагмента, перекрывающего аминокислотную последовательность про-scFv(TF)3e10, включая N-концевую сигнальную последовательность и С-концевую e-tag-последовательность, в бактериальной экспрессионной плазмиде. Осуществляют ПЦР для создания ДНК-фрагмента, который встраивают в экспрессионную плазмиду млекопитающих, содержащую ген, который обусловливает устойчивость к ампициллину, и гены маркеров селекции гигромицина и DHFR (дигидрофолатредуктазы). Экспрессия одноцепочечных антител, предлагаемых в изобретении, находилась под контролем промотора LTR MPSV.
Согласно предпочтительному варианту осуществления изобретения экспрессионными конструкциями, предназначенными для клеток млекопитающих, трансфектируют СНО-клетки линии DXB11. Отбирают стабильные популяции, используя 400 мкг/мл гигромицина В в среде HAMS/F12. Уровни экспрессии составляют примерно 500 мкг/л. Для повышения уровней экспрессии для отбора популяции используют 100 нМ метотрексат в среде альфа-МЕМ. Уровень экспрессии этой популяции составляет примерно 5 мг/л.
Антитела, предлагаемые в настоящем изобретении, можно очищать с помощью методов, хорошо известных в данной области. Например, антитела можно очищать с помощью аффинной хроматографии на колонке, с которой связан ТФ. Затем комплекс ТФ-антитела можно элюировать из колонки с помощью буфера с высокой концентрацией соли.
Одноцепочечные антитела, предлагаемые в изобретении, содержат на С-конце белка последовательность e-tag. Колонки для аффинной хроматографии, содержащие антитела к e-tag, получают от фирмы American/Pharmacia Biotech. Среды для культуры клеток фильтруют через фильтр с размером отверстий 0,22 мкм и вносят в 5-миллилитровую колонку, загруженную e-tag, со скоростью 2 мл/мин. Колонку отмывают 0,2М фосфатным буфером, дополненным 0,05% NaN3, рН 7,0, и затем продукт собирают в пробирки, содержащие 0,1 объема 1М Трис-буфера, рН 8,2, для нейтрализации буфера для элюирования. В другом варианте профильтрованную культуральную среду вносят в колонку, загруженную протеином А. В этом случае колонку отмывают 50 мМ лимонной кислотой, 300 мМ NaCl, рН 6,5 и элюируют этим же буфером, рН 3.0. В обоих случаях очищенные образцы затем вносят в колонку, заполненную Sephadex 200, для отделения мономерных форм антитела от димерных форм.
Отбор антител, предлагаемых в изобретении
После того как антитела получены, экспрессированы и очищены, их можно дополнительно характеризовать с помощью BIAcore, анализа оценки рТФ-зависимого фактора VIIa (анализ способности рТФ/FVIIa осуществлять пептидный гидролиз), анализа активации FX и оценки ПВ, которые описаны в примере 4, для идентификации антител, предлагаемых в настоящем изобретении.
Согласно предпочтительному варианту осуществления изобретения ТФ-связывающие scFv-антитела, которые связываются с большей аффинностью с комплексом FVIIa/ТФ, чем со свободным ТФ, отбирают с помощью анализа способности рТФ/FVIIa осуществлять пептидный гидролиз. При оценке этим анализом для антител к ТФ, которые связываются с большей аффинностью с комплексом FVIIa/ТФ, чем со свободным ТФ, должны быть характерны более высокие значения КD(арр). На фиг.2 показано, что для одноцепочечного антитела scFv(TF)3e10 значение КD(арр) повышается ˜ в 5 раз. Для оценки аффинности антител к ТФ к комплексу FVIIa/ТФ по сравнению с одним ТФ применяют микрокалориметрический анализ. На фиг.5 показано, что одноцепочечное антитело scFv(TF)3e10 характеризуется значениями KD связывания с комплексом FVIIa/ТФ и со свободным ТФ, составляющими 600 нМ и 33 нМ соответственно, что свидетельствует о ˜20-кратном повышении аффинности к комплексу FVIIa/ТФ по сравнению со свободным ТФ. Антитела, предлагаемые в настоящем изобретении, имеют по меньшей мере в 2 раза, предпочтительно по меньшей мере в 5 раз и наиболее предпочтительно по меньшей мере в 10 раз более высокую аффинность к комплексу FVIIa/ТФ, чем к ТФ.
ТФ-связывающие scFv-антитела, которые не конкурируют с FVII за связывание с ТФ, отбирают с помощью анализа способности рТФ/FVIIa осуществлять пептидный гидролиз. В этом анализе антитела к ТФ, которые конкурируют с FVIIa за связывания с ТФ, должны ингибировать гидролиз хромогенного субстрата S2266. На фиг.1 показано, что одноцепочечное антитело scFv(TF)3e10 не ингибирует, а фактически повышает активность FVIIa.
ТФ-связывающие scFv-антитела, которые ингибируют активацию FX, отбирают с помощью оценки ПВ и анализа активации FX. При осуществлении анализа ПВ следует ожидать, что антитела к ТФ, которые пролонгируют ПВ, должны ингибировать активацию FX. На фиг.3 показано, что одноцепочечное антитело scFv(TF)3e10 пролонгирует ПВ, что позволяет предположить, что это антитело ингибирует активацию FX. При анализе активации FX антитела к ТФ, которые ингибируют активность FXa, должны ингибировать гидролиз хромогенного субстрата S2222. На фиг.6 показано, что одноцепочечное антитело scFV(TF)3e10 ингибирует активность FXa.
ТФ-связывающие scFv-антитела, которые не конкурируют с FX за связывание с ТФ, отбирают с помощью анализа активации FX. В этом анализе антитела к ТФ, которые не конкурируют с FX, должны ингибировать активацию FX независимо от применяемой концентрации FX. На фиг.7 показано, что одноцепочечное антитело scFv(TF)3e10 ингибирует активность FXa независимо от концентрации FX.
Согласно предпочтительному варианту осуществления настоящего изобретения выявлено одноцепочечное антитело (scFv(TF)3e10), которое несет один VH/VL-связывающий центр для ТФ. Аминокислотная последовательность scFv(TF)3e10 представлена в примере 1 и соответствует SEQ ID NO:1. Последовательность ДНК, кодирующая scFv(TF)3e10, соответствует SEQ ID NO:2.
Аналоги, фрагменты, производные и варианты антител, предлагаемых в изобретении
Под аналогом, фрагментом, производным или вариантом антител, предлагаемых в настоящем изобретении, подразумевают аналог, фрагмент, производное или вариант, в котором: (I) один или несколько аминокислотных остатков заменены консервативным или неконсервативным аминокислотным остатком (предпочтительно консервативным аминокислотным остатком), и такой замененный аминокислотный остаток может кодироваться таким же или отличным генетическим кодом; или (II) в котором один или несколько аминокислотных остатков включают группу заместителя, или (III) в котором зрелое антитело слито с другим соединением, например с соединением, которое увеличивает время полужизни антитела (например, с полиэтиленгликолем), или (IV) в котором дополнительные аминокислоты слиты со зрелым антителом, такие как лидерная или секреторная последовательность или последовательность, которую применяют для очистки зрелого антитела. Из приведенного описания специалистам в данной области должны быть очевидны понятия "аналоги", фрагменты", "производные" и "варианты".
Предпочтительно производные, предлагаемые в настоящем изобретении, должны содержать консервативные аминокислотные замены (дополнительно описанные ниже) одного или нескольких предсказанных, предпочтительно заменимых аминокислотных остатков. "Заменимый" аминокислотный остаток представляет собой остаток, который может быть заменен в последовательности белка дикого типа без изменения биологической активности, а "незаменимый" аминокислотный остаток необходим для проявления биологической активности. Понятие "консервативная аминокислотная замена" обозначает замену, при которой аминокислотный остаток заменяют аминокислотным остатком, который имеет аналогичную боковую цепь. Семейства аминокислотных остатков, которые имеют аналогичные боковые цепи, известны в данной области. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серии, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Неконсервативные замены не могут затрагивать консервативные аминокислотные остатки или аминокислотные остатки, расположенные в консервативном домене белка, если замены не делают с целью отбора варианта антител, что будет дополнительно описано ниже. Фрагменты или биологически активные белки включают полипептидные фрагменты, пригодные для применения в качестве лекарственного средства, реагента для исследования и т.п. Фрагменты включают пептиды, которые содержат аминокислотные последовательности, аналогичные или выведенные из аминокислотных последовательностей антитела, предлагаемого в настоящем изобретении, и которые обладают по меньшей мере одной активностью указанного полипептида, но которые включают меньше аминокислот по сравнению с описанными полноразмерными полипептидами.
Кроме того, предпочтительные производные, предлагаемые в настоящем изобретении, включают зрелые антитела, слитые с другим соединением, например с соединением, которое удлиняет время полужизни полипептида и/или снижает потенциальную иммуногенность полипептида (например, с полиэтиленгликолем, "ПЭГ"), ПЭГ можно применять для воздействия на водорастворимость, размер, медленную скорость почечного клиренса и для снижения иммуногенности антитела (см., например, US 6214966). При ПЭГилировании слияние антитела с ПЭГ можно осуществлять с помощью любых методов, известных специалистам в данной области. Например, ПЭГилирование можно осуществлять сначала путем интродукции цистеина посредством мутации в антитело с последующей сайтспецифической дериватизацией с использованием ПЭГ-малеимида. Цистеин можно добавлять к С-концу пептидов (см., например, Tsutsumi и др., Proc. Natl. Acad. Sci. USA 97(15), 2000, сс.8548-8553). Другая модификация, которой можно подвергать антитело, включает биотинилирование. В определенных случаях может потребоваться, чтобы антитело было биотинилировано так, чтобы легко вступать в реакцию со стрептавидином. Методы биотинилирования белков хорошо известны в данной области. Кроме того, можно интродуцировать сайты N- или О-гликозилирования в последовательности антител с тем, чтобы посттрансляционное связанное с N или О гликозилирование антител могло происходить in vivo.
Варианты антител, предлагаемые в настоящем изобретении, включают полипептиды, аминокислотная последовательность которых аналогична аминокислотной последовательности исходных антител. Понятие "аналогична" означает, что первая аминокислотная последовательность содержит достаточное или минимальное количество идентичных или эквивалентных аминокислотных остатков относительно второй аминокислотной последовательности, так что первая и вторая аминокислотные последовательности имеют общий структурный домен и/или общую функциональную активность. Например, аминокислотные последовательности, которые содержат общий структурный домен, идентичный по меньшей мере примерно на 45%, предпочтительно примерно на 75-98%, рассматриваются в контексте настоящего описания как аналогичные. Предпочтительно варианты должны быть аналогичны аминокислотной последовательности предпочтительных антител, предлагаемых в настоящем изобретении. Варианты включают варианты антител, кодируемых полинуклеотидом, который гибридизуется с полинуклеотидом, предлагаемым в настоящем изобретении, или его комплементом в строгих условиях. Такие варианты, как правило, сохраняют функциональную активность антител, предлагаемых в настоящем изобретении. Для создания разнородной популяции фрагментов, предназначенной для скрининга и последующего отбора, можно применять библиотеки фрагментов полинуклеотидов. Например, библиотеку фрагментов можно создавать путем обработки двухцепочечного ПЦР-фрагмента полинуклеотида нуклеазой в условиях, при которых происходит примерно только один одноцепочечный разрыв ("ник") на молекулу, денатурации двухцепочечной ДНК, ренатурации ДНК с образованием двухцепочечной ДНК, которая может включать смысловые/антисмысловые пары из различных, полученных в результате "ников" продуктов, удаления одноцепочечных участков из преобразованных дуплексов путем обработки нуклеазой S1 и встраивания путем лигирования полученной библиотеки фрагментов в экспрессионный вектор. С помощью этого метода можно создавать экспрессионную библиотеку, которая кодирует N-концевые и внутренние фрагменты различных размеров антител, предлагаемых в настоящем изобретении.
Варианты включают антитела, которые имеют различия в аминокислотной последовательности в результате мутагенеза. Например, мутагенез можно осуществлять с помощью методов рекомбинантной ДНК, которые хорошо известны в данной области, модифицируя VL - и VН-области легких и тяжелых цепей Ig, создавая варианты, которые обладают более высокой аффинностью к связыванию с комплексом FVIIa/ТФ, чем со свободным ТФ. Особенно предпочтительные варианты включают антитела, которые имеют модификации в гипервариабельных участках VL- и VН-областей.
Варианты включают также антитела, которые имеют различия в аминокислотной последовательности из-за вставок или делеций аминокислотных остатков, полученных в результате мутагенеза. Например, мутагенез можно осуществлять с помощью методов рекомбинантной ДНК, которые хорошо известны в данной области, для встраивания или делеций аминокислот в N-концевых или С-концевых участках Vl- и VН-областей легких и тяжелых цепей Ig, получая варианты, которые сохраняют практически такую же функциональную активность. Особенно предпочтительные варианты включают одноцепочечные антитела с вставками или делециями аминокислотных остатков в VН-VL-линкере между VH- и VL-областями. VH-VL-линкерная последовательность в одноцепочечном антителе, описанном в примере 1, состоит из 5 аминокислотных остатков. Очевидно, что можно использовать другие короткие линкерные последовательности, состоящие из 0-20 аминокислот, если антитело сохраняет практически аналогичную функциональную активность. Модификации существующего VH-VL-линкера можно использовать для повышения стабильности димерной формы одноцепочечного антитела.
Согласно одному из вариантов осуществления изобретения библиотеку с широким разнообразием вариантов получают путем комбинаторного мутагенеза на нуклеотидном уровне, и она кодируется библиотекой с широким разнообразием генов. Библиотеку с широким разнообразием вариантов можно получать, например, путем ферментативного лигирования смеси синтетических олигонуклеотидов в генных последовательностях, таким образом, чтобы вырожденный набор потенциальных вариантов аминокислотных последовательностей мог экспрессироваться в виде индивидуальных полипептидов, или в другом варианте в виде более крупных слитых белков (например, для фаговой дисплейной библиотеки), содержащих набор последовательностей. Известен целый ряд методов, которые можно применять для создания библиотек потенциальных вариантов из вырожденной олигонуклеотидной последовательности. Химический синтез вырожденной генной последовательности можно осуществлять в автоматическом синтезаторе ДНК и затем встраивать путем лигирования синтетический ген в соответствующий экспрессионный вектор. Применение вырожденного набора генов позволяет объединять в одной смеси все последовательности, кодирующие требуемый набор последовательностей потенциальных вариантов. Методы синтеза вырожденных олигонуклеотидов известны в данной области (см., например, Narang, Tetrahedron 39, 1983, c.3; Itakura и др,. Annu. Rev. Biochem. 53, 1984а, с.323; Itakura и др., Science 198, 1984б, c.1056; Ike и др., Nucleic Acid Res. 11, 1983, c.477).
В данной области известно несколько методов скрининга генных продуктов комбинаторных библиотек, созданных с помощью точечных мутаций или укорачивания, и для скрининга библиотек кДНК с целью введения генных продуктов, несущих отобранный признак. Такие методы можно адаптировать для быстрого скрининга генных библиотек, созданных путем комбинаторного мутагенеза антител, на способность связываться с ТФ- или с комплексом FVIIa/ТФ или на способность ингибировать активацию FX. Наиболее широко используемые методы, которые можно применять для крупномасштабного скрининга больших генных библиотек, как правило, включают клонирование генной библиотеки в способных к репликации экспрессионных векторах, трансформацию соответствующих клеток с получением библиотеки векторов и экспрессию комбинаторных генов в условиях, при которых выявление требуемой активности облегчает выделение вектора, кодирующего ген, продукт которого был выявлен. Для идентификации требуемых вариантов можно применять в сочетании с методами скрининга рекурсивный групповой мутагенез (REM), метод, который позволяет увеличивать частоту функциональных мутантов в библиотеках.
В примере 1 представлена аминокислотная последовательность одного из ТФ-связывающих одноцепочечных антител, а именно scFv(TF)3e10 (SEQ ID NO:1), и делинеаризированные VH-VL-линкер и VH- и VL-области. Варианты, которые обладают способностью связывать ТФ или комплекс FVIIa/ТФ или ингибировать активацию FX, можно выявлять путем скрининга комбинаторных библиотек мутантов, например инсерционных, укороченных или точечных мутантов антител, предлагаемых в настоящем изобретении, с помощью анализа способности рТФ/FVIIa осуществлять пептидный гидролиз или анализа активации FX, которые описаны в примере 4. Антитела, предлагаемые в настоящем изобретении, включают антитела, описанные в примерах 1 и 3 (SEQ ID NO:1 и 3), а также антитела, которые имеют несущественные вариации последовательности. Понятие "несущественная вариация" включает любой вариант последовательности, связанный с заменой или делецией, который сохраняет в значительной степени по меньшей мере одну биологическую функцию антител, предлагаемых в настоящем изобретении, например способность ингибировать активацию FX. Такие функциональные эквиваленты предпочтительно могут включать антитела, в которых домены VL- или VH-области идентичны по меньшей мере на 90% указанным доменам одноцепочечного антитела, которое имеет последовательность SEQ ID NO:1 или SEQ ID NO:3, и более предпочтительно по меньшей мере на 95% идентичны доменам VL- или VH-области одноцепочечного антитела, которое имеет последовательность SEQ ID NO:1 или SEQ ID NO:3, и еще более предпочтительно по меньшей мере на 97% идентичны доменам VL- или VH-области одноцепочечного антитела, которое имеет последовательность SEQ ID NO:1 или SEQ ID NO:3, а также включают участки такого антитела, которые обладают практически одинаковой биологической активностью. Однако под объем настоящего изобретения подпадает любое антитело, аминокислотная последовательность которого имеет незначительную вариацию относительно антитела, имеющего SEQ ID NO:1 или SEQ ID NO:3, которое является функциональным эквивалентом, как это будет описано ниже.
Фармацевтические композиции
Изобретение относится также к фармацевтическим композициям, которые можно вводить пациенту для достижения терапевтического действия. Фармацевтические композиции, предлагаемые в настоящем изобретении, можно приготавливать для введения, объединяя антитело, предлагаемое в настоящем изобретении, которое имеет требуемую степень чистоты и взято в фармацевтически эффективном количестве, с физиологически приемлемыми носителями.
Антитела, предлагаемые в настоящем изобретении, можно применять в виде фармацевтических композиций, предназначенных для внутривенного введения, или подкожного введения, или внутритрахеального введения. Таким образом, описанные выше антитела предпочтительно объединяют с приемлемым стерильным фармацевтическим носителем, таким как 5%-ная декстроза, лактированный раствор Рингера, обычный физиологический раствор, стерильная вода или любой другой поступающий в продажу физиологический буферный раствор, предназначенный для внутривенной инфузии. Должно быть очевидно, что выбор раствора-носителя и доза, а также путь введения композиции должны варьироваться в зависимости от пациента и конкретного клинического требования и должны регулироваться стандартными медицинскими процедурами.
Согласно способам, предлагаемым в настоящем изобретении, указанные фармацевтические композиции можно вводить в количествах, эффективных для ингибирования патологических состояний, которые связаны с избыточным образованием тромбина у индивидуума.
Введение антитела можно осуществлять путем болюсной внутривенной инъекции, путем постоянной внутривенной инфузии или путем объединения обоих путей. В другом или дополнительном варианте антитело, смешанное с соответствующими эксципиентами, может попадать в кровоток из внутримышечной области. Системное лечение с помощью антитела можно оценивать, определяя активированное парциальное тромбопластиновое время (АПТВ) на серийных образцах крови, взятой у пациента. Определяемое в этом опыте время свертывания удлиняется, когда в кровотоке достигается достаточный титр антитела.
Рекомбинантные антитела и фармацевтические композиции, предлагаемые в настоящем изобретении, можно применять для парентерального, местного, внутривенного или локального введения. Фармацевтические композиции можно вводить в виде различных стандартных доз лекарственных средств в зависимости от метода введения. Например, стандартные дозы лекарственных средств можно вводить в форме, которая включает (но не ограничиваясь ими) таблетки, капсулы, порошок, растворы и эмульсии.
Рекомбинантные антитела и фармацевтические композиции, предлагаемые в настоящем изобретении, наиболее пригодны для внутривенного введения. Предназначенные для введения композиции, как правило, содержат раствор одноцепочечного антитела в фармацевтически приемлемом носителе, предпочтительно водном носителе. Можно применять различные водные носители, например забуференный физиологический раствор и т.п. Эти растворы являются стерильными и, как привило, не содержат нежелательные продукты. Композиции можно стерилизовать с помощью общепринятых хорошо известных методов стерилизации.
Типичную фармацевтическую композицию для внутривенного введения может легко выбрать специалист в данной области. Вводимые количества, безусловно, определяются специфичностью белка и зависят от его эффективности и фармакокинетического профиля. Современные методы приготовления вводимых парентерально композиций хорошо известны специалистам в данной области и более подробно описаны в Remington's Pharmaceutical Science, 15oe изд., изд-во Mack Publishing Company, Easton, Pa (1980).
Композиции, которые содержат антитела, предлагаемые в настоящем изобретении, или их смесь (т.е. смесь с другими белками) можно вводить в качестве терапевтического лекарственного средства. Для терапевтического применения композиции вводят пациенту, страдающему связанным с кровотечением нарушением или заболеванием, в количестве, достаточном для лечения или по меньшей мере частичного прекращения кровотечения. Необходимое для этого количество называют "терапевтически эффективным количеством". Эффективное для указанной цели количество должно зависеть от серьезности заболевания и общего состояния здоровья пациента.
Композиции можно применять в виде однократной дозы или нескольких доз в зависимости от уровня дозы и необходимой частоты введения и переносимости пациентом. В любом случае композиция должна обеспечивать достаточное количество белков, предлагаемых в настоящем изобретении, для эффективного лечения пациента.
Антитела, предлагаемые в изобретении, или их фармацевтически приемлемые композиции вводят в терапевтически эффективном количестве, которое должно варьироваться в зависимости от различных факторов, таких как активность специфического применяемого антитела; метаболическая стабильность и продолжительность действия антитела; возраст, вес тела, общее состояние здоровья, пол и рацион пациента; путь и время введения; скорость выведения; комбинация лекарственных средств; серьезность конкретного болезненного состояния; и лечение, которому подвергается хозяин. Как правило, суточное терапевтически эффективное количество составляет от примерно 0,14 до примерно 14,3 мг/кг веса тела в день антитела, предлагаемого в изобретении, или его фармацевтически приемлемой композиции; предпочтительно от примерно 0,7 до примерно 10 мг/кг веса тела в день и наиболее предпочтительно от примерно 1,4 до примерно 7,2 мг/кг веса тела в день. Например, для введения человеку весом 70 кг дозы должны составлять от примерно 10 мг до примерно 1,0 г в день антитела, предлагаемого в изобретении, или его фармацевтически приемлемой композиции, предпочтительно от примерно 50 до примерно 700 мг в день и наиболее предпочтительно от примерно 100 до примерно 500 мг в день.
Клеточная и генная терапия
Антитело, предлагаемое в изобретении, можно применять, согласно настоящему изобретению, путем экспрессии такого антитела in vivo с помощью метода, который называют "клеточная терапия". Так, например, можно создавать клетки, несущие полинуклеотид (ДНК или РНК), который кодирует антитело ех vivo, и затем вводить созданные клетки пациенту для лечения с помощью антитела. Такие методы хорошо известны в данной области. Например, клетки можно создавать с помощью процедур, хорошо известных в данной области, путем применения ретровирусной частицы, содержащей РНК, которая кодирует антитело, предлагаемое в настоящем изобретении.
Антитело, предлагаемое в изобретении, можно применять также согласно настоящему изобретению путем экспрессии такого антитела in vivo с помощью метода, который называют "генная терапия". Так, например, можно создавать вирус, несущий полинуклеотид (ДНК или РНК), который кодирует антитело, и затем вводить созданный вирус пациенту для лечения с помощью антитела. Такие методы хорошо известны в данной области. Например, с помощью процедур, хорошо известных в данной области, можно создавать рекомбинантные аденовирусы, которые содержат ДНК, кодирующую антитело, предлагаемое в настоящем изобретении.
Локальное введение антикоагулирующих антител, предлагаемых в настоящем изобретении, с использованием клеточной или генной терапии может обеспечивать введение терапевтического агента в область-мишень, т.е. выстилку кровеносных сосудов, состоящую из эпителиальных клеток.
Под объем изобретения подпадают методы in vitro и in vivo как клеточной, так и генной терапии. Известно несколько методов переноса потенциально терапевтических генов в определенные клеточные популяции (см., например, Mulligan, Science 260, 1993, сс.926-931). Эти методы включают:
1) непосредственный перенос генов (см., например, Wolff и др, Science, 247, 1990, сс.1465-1468;
2) опосредуемый липосомами перенос ДНК (см., например, Caplen и др., Nature Med. 3, 1995, сс.39-46; Crystal, Nature Med. 1, 1995, cc.15-17; Gao и Huang, Biochem. Biophys. Res. Comm. 179, 1991, сс.280-285;
3) опосредуемый ретровирусом перенос ДНК (см., например, Кау и др., Science 262, 1993, сс.117-119; Anderson, Science 256, 1992, сс.808-813;
4) опосредуемый ДНКовыми вирусом перенос ДНК. Такие ДНКовые вирусы включают аденовирсусы (предпочтительно векторы на основе Ad2 или Ad5), вирусы герпеса (предпочтительно векторы на основе вируса герпеса простого) и парвовирусы (предпочтительно векторы на основе "дефектных" или не автономных парвовирусов, более предпочтительно векторы на основе аденоассоциированного вируса, более предпочтительно векторы на основе AAV-2) (см., например, Ali и др. Gene Therapy I, 1994, сс.367-384; US 4797368, включенный в настоящее описание в качестве ссылки, и US 5139941, включенный в настоящее описание в качестве ссылки).
Выбор конкретной векторной системы для переноса представляющего интерес гена должен зависеть от ряда факторов. Одним из важных факторов является природа клеточной популяции-мишени. Хотя ретровирусные векторы широко изучены и нашли применение в различных аспектах генной терапии, эти векторы, как правило, нельзя применять для заражения неделящихся клеток. Кроме того, ретровирусы обладают потенциальной канцерогенностью. Однако современные разработки в области лентивирусных векторов позволяют преодолеть некоторые из этих ограничений (см. Naldini и др., Science 272, 1996, сс.263-267).
Ретровирусы, на основе которых можно создавать указанные выше ретровирусные плазмидные векторы, включают (но не ограничиваясь ими) вирус мышиного лейкоза Молони, вирус некроза селезенки, ретровирусы, такие как вирус саркомы Рауса, вирус саркомы Гарвея, вирус лейкоза птиц, вирус лейкоза гиббонов, вирус иммунодефицита человека, аденовирус, вирус миелопролиферативной саркомы и вирус опухоли молочной железы. В одном из вариантов осуществления изобретения ретровирусный плазмидный вектор получают из вируса мышиного лейкоза Молони.
Преимуществом аденовирусов является широкий спектр их хозяев, их способность заражать покоящиеся или полностью дифференцированные клетки, такие как нейроны или гепатоциты, и то, что, вероятно, они в основном не являются канцерогенными (см., например, Ali и др. (1994), выше, с.367). Аденовирусы, вероятно, не интегрируются в геном хозяина. Поскольку они существует вне хромосом, риск инсерционного мутагенеза в значительной степени снижен (Ali и др. (1994), выше, с.373).
Аденоассоциированные вирусы обладают такими же преимуществами, что и векторы на основе аденовирусов. Однако для AAV характерна сайтспецифическая интеграция в человеческую хромосому 19 (Ali и др. (1994), выше, с.377).
Согласно предпочтительному варианту осуществления изобретения ДНК, которая кодирует антитела, предлагаемые в настоящем изобретении, применяют в клеточной или генной терапии для нарушений, которые включают (но не ограничиваясь ими) тромбоз глубоких вен, диссеминированное внутрисосудистое свертывание крови, острый коронарный синдром или рак с признаком коагулопатии.
Согласно этому варианту осуществления изобретения клеточную или генную терапию с использованием ДНК, которая кодирует антитела, предлагаемые в настоящем изобретении, применяют для лечения пациента, который нуждается в этом, при установлении диагноза или сразу после этого.
Специалистам в данной области должно быть очевидно, что согласно этому варианту осуществления изобретения можно применять любой пригодный для генной терапии вектор, содержащий ДНК, которая кодирует антитело, предлагаемое в изобретении, или ДНК, которая кодирует аналоги, фрагменты, производные или варианты антитела, предлагаемого в изобретении. Методы конструирования такого вектора хорошо известны (см., например, Anderson W.F. Nature 392, 1998, сс.25-30; Verma I.M. и Somia N. Nature 389, 1998, сс.239-242). Интродукцию вектора, содержащего ДНК антитела, в область-мишень можно осуществлять известными методами.
Векторы, применяемые для клеточной или генной терапии, включают один или несколько промоторов. Приемлемые промоторы, которые можно применять, включают (но не ограничиваясь ими) LTR ретровируса; промотор ОВ-40 и промотор человеческого цитомегаловируса (CMV), описанные у Miller и др., Biotechniques 7(9), 1989, сс.980-990, или любой другой промотор (например, клеточные промоторы, такие как промоторы эукариотических клеток, которые включают (но не ограничиваясь ими), промоторы гистона, ро1 III и (β-актина). Другие вирусные промоторы, которые можно применять, включают (но не ограничиваясь ими) промоторы аденовирусов, промоторы тимидинкиназы (ТК) и промоторы парвовируса В19. Выбор пригодного промотора должен быть очевиден специалистам в данной области после ознакомления с настоящим описанием.
Нуклеотидная последовательность, которая кодирует антитело, предлагаемое в настоящем изобретении, находится под контролем приемлемого промотора. Приемлемые промоторы, которые можно применять, включают (но не ограничиваясь ими) промоторы аденовирусов, например основной поздний промотор аденовируса; или гетерологичные промоторы, например промотор цитомегаловируса (CMV); промотор респираторно-синцитиального вируса (РСВ); индуцибельные промоторы, например промотор ММТ, промотор металлотионеина; промоторы теплового шока; промотор альбумина; промотор ApoAl; промоторы человеческого глобина; промоторы вирусной тимидинкиназы, например промотор тимидинкиназы вируса герпеса простого; ретровирусные LTR (включая модифицированные ретровирусные LTR, которые описаны выше); промотор β-актина и промотор человеческого гормона роста.
Ретровирусный плазмидный вектор применяют для трансдукции упаковывающих клеточных линий для получения клеточных линий-продуцентов. Примеры упаковывающих клеток, которые можно трансфектировать, включают (но не ограничиваясь ими) РЕ501, РА317, ψ-2, ψ-АМ, РА12, Т19-14Х; VT-19-17-Н2, ψCRE, ψCRIP, GP+#-86, GP+envAml2 и клеточных линий DAN, которые описаны у Miller, Human Gene Therapy 1, 1990, сс.5-14, публикация полностью включена в настоящее описание в качестве ссылки. Вектором можно трансдуцировать упаковывающие клетки с помощью любых методов, известных в данной области. Такие методы включают (но не ограничиваясь ими) электропорацию, применение липосом и осаждение с помощью CaPO4. В качестве одной из альтернатив ретровирусный плазмидный вектор можно капсулировать в липосому или сшивать с липидом, а затем вводить хозяину. Клеточная линия-продуцент образует инфекционные ретровирусные частицы, которые включают нуклеотидную(ые) последовательность(и), которая(ые) кодирует(ют) полипептиды. Такие частицы ретровирусных векторов можно затем применять для трансдукции эукариотичесикх клеток либо in vitro, либо in vivo. Трансдуцированные эукариотические клетки должны экспрессировать нуклеотидную(ые) последовательность(и), которая(ые) кодирует(ют) полипептиды. Эукариотические клетки, которые можно трансдуцировать, включают (но не ограничиваясь ими) эмбриональные стволовые клетки, эмбриональные раковые клетки, а также гематопоэтические стволовые клетки, гепатоциты, фибробласты, миобласты, кератиноциты, эндотелиальные клетки и бронхиальные эпителиальные клетки.
Альтернативным генной терапии подходом является "транскариотическая терапия", при осуществлении которой клетки пациента обрабатывают ех vivo для того, чтобы индуцировать у молчащих хромосомных генов способность продуцировать представляющий интерес белок после обратной интродукции в организм пациента. Транскариотическая терапия подразумевает, что пациент имеет нормальный комплемент генов, необходимый для активации. Транскариотическая терапия предусматривает интродукцию промотора или другой экзогенной регуляторной последовательности, которая обладает способностью активировать находящиеся на стадии возникновения гены в хромосомную ДНК клеток пациента ех vivo, культивирование и отбор клеток, продуцирующих активный белок, и последующую обратную интродукцию активированных клеток в организм пациента с тем, чтобы они затем полностью восстановились. После этого "активированная геном" клетка продуцирует представляющий интерес белок в течение некоторого значительного промежутка времени, возможно, в течение всей жизни пациента. В US 5641670 и 5733761 подробно изложена эта концепция, и они полностью включены в настоящее описание в качестве ссылки.
Наборы
Настоящее изобретение относится также к наборам для исследования или диагностических целей. Наборы, как правило, включают один или несколько контейнеров, которые содержат антитела, предлагаемые в настоящем изобретении. Согласно предпочтительному варианту осуществления изобретения наборы включают контейнеры, которые содержат одноцепочечные антитела в форме, пригодной для дериватизации с использованием второй молекулы. Согласно наиболее предпочтительному варианту осуществления изобретения наборы включают контейнеры, содержащие антитело, которое имеет последовательность SEQ ID NO:1 или SEQ ID NO:3.
Согласно другому варианту осуществления изобретения наборы могут содержать последовательности ДНК, которые кодируют антитела, предлагаемые в изобретении. Предпочтительно последовательности ДНК, кодирующие эти антитела, присутствуют в плазмиде, которую можно применять для трансфекции клетки-хозяина и экспрессии в ней. Плазмида может содержать промотор (часто индуцибельный промотор) для регуляции экспрессии ДНК в клетке-хозяине. Плазмида может содержать также соответствующие сайты рестрикции для облегчения встраивания других последовательностей ДНК в плазмиду с целью получения различных антител. Плазмиды могут содержать также многочисленные другие элементы, предназначенные для облегчения клонирования и экспрессии кодируемых белков. Такие элементы хорошо известны специалистам в данной области и включают, например, селектируемые маркеры, инициирующие кодоны, терминирующие кодоны и т.п. Согласно более предпочтительному варианту осуществления изобретения наборы включают контейнеры, содержащие последовательности ДНК SEQ ID NO:2 или SEQ ID NO:4.
Терапевтические показания
Болезни, в этиологии которых образование тромба играет очень важную роль, включают инфаркт миокарда, диссеминированное внутрисосудистое свертывание крови, тромбоз глубоких вен, эмболию легких, ишемический "удар", септический шок, острый респираторный дистресс-синдром, нестабильную стенокардию и другие артериальные и венозные связанные с окклюзией состояния. Антитела, предлагаемые в настоящем изобретении, можно применять во всех указанных случаях, а также при других заболеваниях, при которых образование тромба является патологическим. Другие патологические состояния, при которых можно применять антитело, предлагаемое в настоящем изобретении, включают рак, сопровождающийся коагулопатией и воспалением. Антитела могут найти также применение при трансплантации кожи и вен и трансплантации органов. Под понятием "можно применять" ("полезны") понимают, что антитела приносят пользу при лечении, либо предупреждая заболевание, либо предупреждая его переход в более серьезную стадию. Антитела, предлагаемые в настоящем изобретении, являются также безопасными и эффективными антикоагулянтами, например, их можно использовать для пациентов, которые имеют биопротез, например, клапанов сердца. Эти антитела могут заменять гепарин и варфарин при лечении, например, эмболии легких или острого инфаркта миокарда. Антитела, предлагаемые в настоящем изобретении, могут найти также применение в качестве покрытия медицинских устройств, в которых может иметь место свертывание.
Анализы
Известен ряд лабораторных анализов, которые можно применять для оценки in vitro антикоагулирующей активности антитела, предлагаемого в настоящем изобретении. Антикоагулирующее действие антитела можно оценивать на основе анализов времени свертывания в плазме, таких как активированное парциальное тромбопластиновое время ("АПТВ"), время образования тромбина (время свертывания под воздействием тромбина) ("ТСВ") и/или протромбиновое время ("ПВ"). Эти анализы позволяют различать различные механизмы ингибирования свертывания и включают активацию белка С. Пролонгирование времени свертывания, установленное в одном из этих анализов, свидетельствует о том, что молекула может ингибировать свертывание в плазме.
Вышеперечисленные анализы применяют для выявления антител с антикоагулирующей активностью, которые могут связывать ТФ как в очищенных системах, так и в плазматических средах. Кроме того, для оценки активности антител, предлагаемых в изобретении, применяют дополнительные анализы, такие как анализ катализируемого тромбином образования фибрина из фибриногена (Jakubowski H.V. и др., J. Biol. Chem. 261(8), 1986, сс.3876-3882), анализ ингибирования активации тромбином фактора V (Esmon С.Т. и др., J. Biol. Chem. 257(14), 1982, cc.7944-7947), анализ усиления ингибирования тромбина антитромбином III гепариновым кофактором II (Esmon N.L. и др., J. Biol. Chem. 258(20), 1983, с.с.12238-12242), анализ ингибирования активации тромбином фактора XIII (Polgar J. и др., Thromb. Haemost. 58(1), 1987, с.140), анализ ингибирования опосредуемой тромбином инактивации белка S (Thompson Е.А. и Salem H.H., J. Clin. Inv. 78(1), 1986, сс.13-17) и анализ ингибирования опосредуемых тромбином активации и агрегации тромбоцитов (Esmon N.L. и др. (1983), выше).
Следующие, подробно описанные в примере 4, анализы, применяют для оценки in vitro эффективности или оценки in vitro аффинности к связыванию антител, предлагаемых в изобретении: 1) анализ способности рТФ/FVIIa осуществлять пептидный гидролиз; 2) анализ активации фактора X; 3) оценка ПВ; и 4) микрокалориметрический анализ.
При осуществлении методов, применяемых в настоящем изобретении, естественно должно быть очевидно, что ссылку на конкретные буферы, среды, реагенты, клетки, условия культивирования и т.п., не следует рассматривать как ограничивающую объем изобретения, а как включающую все родственные продукты, которые, как очевидно специалистам в данной области, могут представлять интерес или иметь значение в конкретном контексте настоящего описания. Например, часто может заменять одну буферную систему или культуральную среду на другую, достигая при этом аналогичного или идентичного результата. Специалистам в данной области известны системы и методики, с помощью которых без излишних экспериментов можно осуществлять такие замены и которые могут служить для оптимизации методов и процедур, представленных в настоящем описании.
Ниже настоящее изобретение будет более подробно описано на примерах, не ограничивающих его объем. Должно быть очевидно, что вместо методов, применяемых в конкретных примерах, специалист в данной области, безусловно, может сам предложить другие варианты их осуществления.
Выше и в приведенных ниже примерах, если не указано иное, все температуры даны в градусах Цельсия, все части и проценты даны в мас.ч. или в мас.%.
Полное описание всех процитированных выше заявок на патенты, патентов и публикаций включено в настоящее описание в качестве ссылок.
Пример 1
Конструкция одноцепочечного антитела к ТФ scFv(TF)3e10

Одноцепочечное антитело к ТФ scFv(TF)3e10 (SEQ ID NO:1) состоит из сигнального пептида (от положения -18 до положения -1), VH-области (1-126), VH-VL-линкера (127-131), VL-области (132-246) и последовательности е-tag (247-264). Последовательность ДНК scFv(TF)3e10 (SEQ ID NO:2) кодирует 25 аминокислотную последовательность SEQ ID NO:1.
Пример 2
Конструкция 1 слитого белка scFv(TF)3e10-TMi456

Слитый белок scFv(TF)3e10-TMi456 (SEQ ID NO:2) состоит из сигнального пептида (от положения -18 до положения -1), VH-области (1-126), VH-VL-линкера (127-131), VL-области (132-246), VL-ТМ-линкера (247-264), TMi-домена (265-382) и последовательности е-tag (383-400). Мутации H381G, M388L, R456G и H457Q в TMi456 подчеркнуты.
Пример 3
Конструкция 2 слитого белка scFv(TF)3e10-TMi456

Слитый белок scFv(TF)3e10-TMi456 (SEQ ID NO:3) состоит из сигнального пептида (от положения -18 до положения -1), VH-области (1-126), VH-VL-линкера (127-131), VL-области (132-246), VL-ТМ-линкера (244-261), TMi-домена (262-379). Мутации H381G, M388L, R456G и H457Q в TMi456 подчеркнуты.
В изменениях, внесенных в письменную и компьютерную версию описания и перечня последовательностей, исключены ошибки. К этим изменениям никаких дополнений не сделано.
Пример 4
Анализы in vitro эффективности и аффинности к связыванию
1. Анализ способности рТФ/FVIIa осуществлять пептидный гидролиз
Ниже описан принцип этого анализа. Трипептидную пара-нитроанилидамидную связь субстрата гидролизуют с помощью комплекса рТФ/FVIIa. Высвобождение хромофора, т.е. пара-нитроанилида (p-NA), оценивают при 405 нм и концентрацию продукта, образовавшегося в единицу времени, рассчитывают с использованием коэффициента молярной экстинкции 9920 М-1 см-1. Значения IC50 (С) определяют путем аппроксимации начальных скоростей с помощью 4-параметрического уравнения Y=(A-D)/(1+(x/C)^B)+D

Реагенты и растворы:
1. Буфер для анализа (БА): 50 мМ Трис-HCl, 150 мМ NaCl, 5 мМ CaCl2, 0,1% БСА, рН 7,5.
2. Человеческий FVIIa (HCVIIA-0060, фирма Haematologic Technologies Inc.): 10-кратный (×) рабочий раствор - приготавливают 20 нМ раствор в буфере для анализа перед применением.
3. Растворимый ТФ (фирма Berlex): 10-кратный рабочий раствор - приготавливают 30 нМ раствор в буфере для анализа перед применением.
4. Хромогенный субстрат S2266 (фирма Kabi Pharmacia Hepar Inc.): Маточный раствор: 10 мМ в Н2О, хранившийся при 4°С. 2,5-кратный рабочий раствор - приготавливают 2,5 нМ раствор в буфере для анализа перед применением.
5. Антитело: приготавливают 2,5-кратные разведения в буфере для анализа перед применением.
Условия проведения анализа
Анализы осуществляют в 96-луночном титрационном микропланшете при комнатной температуре. Конечные концентрации компонентов составляют:
Процедура анализа
1. Вносят пипеткой 0,1 мл 2,5 × БА (или контрольного буфера) в каждую лунку.
2. Добавляют 0,025 мл 10 × рТФ и инкубируют 10 мин при комнатной температуре, осторожно встряхивая.
3. Добавляют 0,025 мл 10 × FVIIa, инкубируют 10 мин при комнатной температуре, осторожно встряхивая.
4. Добавляют 0,1 мл 2,5 × субстрата S2266, немедленно переносят планшет в планшет-ридер и измеряют кинетические характеристики фермента при 405 нм в течение 15 мин с 10 с интервалами.
Этот анализ можно использовать для оценки кажущегося значения KD, которое характеризует связывание антител, предлагаемых в изобретении, с рТФ или с комплексом FVIIa/ТФ и для определения, конкурируют ли антитела, предлагаемые в изобретении, с FVII за связывание с ТФ.
2. Анализ активации фактора Х
Ниже описан принцип этого анализа. FVIIa инкубируют с пузырьками, содержащими рекомбинантный человеческий ТФ, получая протеазный комплекс, который обладает способностью активировать субстрат, т.е. FX. Этот комплекс формируют в присутствии (или в отсутствие) различных концентраций антитела, затем вводят субстрат FX и реакции дают протекать до образования продукта, т.е. активной протеазы FXa, которая обладает способностью гидролизовать пара-нитроанилидамидную связь хромогенного субстрата S2222. Высвобождение хромофора, т.е. пара-нитроанилида, оценивают при 405 нм и концентрацию продукта, образовавшегося в единицу времени, рассчитывают с использованием коэффициента молярной экстинкции 9920 М-1 см-1. Значения IC50 (С) определяют путем аппроксимации начальных скоростей с помощью 4-параметрического уравнения: Y=(A-D)/(1+(х/С^В)+D

Реагенты и растворы:
1. Буфер для анализа: 50 мМ Трис-HCl, 150 мМ NaCl, 5 мМ CaCl2, 0,1% БСА,
рН 7,5.
2. Человеческий FVIIa (HCVIIA-0031, фирма Haematologic Technologies Inc.): 4-кратный рабочий раствор - приготавливают 100 пМ раствор в буфере для анализа перед применением.
3. Рекомбинантный человеческий ТФ (восстановленный в лаборатории заявителей из препарата фирмы Innovin, Dade): рабочий раствор -приготавливают разбавление 1:480 в буфере для анализа перед применением.
4. Человеческий фактор Х (НСХ-0060, фирма Haematologic Technologies Inc.): 4-кратный рабочий раствор - приготавливают 1000 нМ раствор в буфере для анализа перед применением.
5. Хромогенный субстрат S2222 (фирма Kabi Pharmacia Hepar Inc.):
Маточный раствор: 6 мМ в Н2O, хранившийся при 4°С.
Рабочий раствор - приготавливают 0,78 мМ раствор в 3,57 мМ ЭДТК (для прекращения реакции), 150 мМ NaCl, 50 мМ Трис-HCl, рН 7,5 перед применением.
6. Антитело:
Приготавливают 4-кратные разведения в буфере для анализа перед применением.
Условия проведения анализа:
Анализы осуществляют в 96-луночном титрационном микропланшете при комнатной температуре. Конечные концентрации компонентов составляют:
Пузырьки рТФ: 1/4 от разведения 1:480
Процедура анализа:
1. Вносят пипеткой 0,015 мл 4×БА (или контрольного буфера) в каждую лунку.
2. Добавляют 0,015 мл 4 × пузырьков рТФ.
3. Добавляют 0,015 мл 4 × FVIIa, инкубируют в течение 10 мин при комнатной температуре, осторожно встряхивая.
4. Добавляют 0,015 мл 4 × FX, инкубируют в течение 5 мин при комнатной температуре, осторожно встряхивая.
5. Добавляют 0,14 мл S2266, немедленно переносят планшет в планшет-ридер и измеряют кинетические характеристики фермента при 405 нм в течение 15 мин с 10 с интервалами.
Этот анализ можно использовать для определения того, могут ли антитела, предлагаемые в изобретении, ингибировать активацию FX комплексом FVIIa/ТФ и могут ли антитела, предлагаемые в изобретении, конкурировать с FX за связывание с комплексом FVIIa/ТФ.
3. Определение протромбинового времени
При осуществлении стандартной реакции для определения ПВ 90 мкл соответствующей концентрации антитела или ЗФР добавляют в кювету к смеси, содержащей 20 мкл тромбопластина IS (фирма Dade) и 90 мкл 25 мМ CaCl2. Смесь инкубируют в течение 1 мин при 37°С, затем добавляют 100 мкл цитрированной плазмы (фирма Helena Laboratories). В другом варианте соответствующий объем концентрированного антитела добавляют к 100 мкл рекомбинантного человеческого тромбопластина (фирма Ortho Recombiplastin) и примерно через 2 мин добавляют 100 мкл восстановленной человеческой плазмы. Время свертывания в каждом индивидуальном опыте по оценке коагуляции оценивают, определяя среднее значение для двух измерений, полученных с помощью устройства для измерения свертывания (коагулометра) типа Electra 900C (фирма Hemoliance), и определяют средние значения для проведенных с дублированием анализов (n=3 или 4). Получают кривые зависимости реакции от дозы для каждого ингибитора и затем применяют регрессионный анализ для расчета концентрации (в нМ), необходимой для двукратного увеличения времени свертывания.
Этот анализ можно использовать для оценки воздействия антител, предлагаемых в изобретении, на внешний (нехарактерный) путь свертывания крови. Определяют количество антитела, необходимое для удвоения ПВ, которое можно сравнивать с другими антикоагулянтами, определяя тем самым относительную эффективность антител, предлагаемых в изобретении.
4. Микрокалориметрический анализ
Микрокалориметрический (изотермическая титрационная калориметрия) анализ применяют для определения аффинности к связыванию (КD) антител, предлагаемых в изобретении, со свободным ТФ или с комплексом FVIIa/ТФ. Этот анализ осуществляют с помощью устройства MicroCal VP-ITC. Предварительно получают комплекс FVIIa/ТФ, добавляя 2,3-кратный молярный избыток FVIIai к рТФ. Для подтверждения того, что рТФ в этом анализе полностью входит в комплекс, применяют гель-фильтрацию. Для определения аффинности антитела к комплексу 1,2 мкМ комплекс VIIa/ТФ добавляют в ячейку микрокалориметра и 65 мкМ раствор антитела вносят в шприц. Для определения аффинности антитела к свободному рТФ 10 мкМ рТФ добавляют в ячейку и 141 мкМ раствор антитела вносят в шприц. Для анализа данных применяют программное обеспечение MicroCal Origin. Данные соответствуют одному сайту связывания.
Пример 5
Характеристики in vitro антител к ТФ, предлагаемых в настоящем изобретении
Шесть различных ТФ-связывающих антител выделяют из фаговой дисплейной библиотеки полных человеческих одноцепочечных антител: scFv(TF)2c1, scFv(TF)2c11, scFv(TF)2d3, scFv(TF)2h6, scFv(TF)3e10 и scFv(TF)3h2. Аффинность этих рТФ-связывающих антител, экспрессируемых в E.coli, при оценке с помощью BIAcore, составляет 35-470 нМ. Анализ способности рТФ/VIIa осуществлять пептидный гидролиз, который описан в примере 4, используют для определения того, могут ли эти антитела блокировать образование активного комплекса VIIa/ТФ. При таком анализе связывание VIIa с рТФ увеличивает скорость расщепления хромогенного пептидного субстрата S2266 более чем в 20 раз. Антитела, которые ингибируют связывание FVIIa с ТФ, блокируют это увеличение скорости. Для 5 одноцепочечных антител, т.е. scFv(TF)2c1, scFv(TF)2c11, scFv(TF)2d3, scFv(TF)2h6 и scFv(TF)3h2, выявлена способность ингибировать гидролиз S2266, что позволяет предположить, что они ингибируют связывание FVIIa с рТФ (фиг.1). В противоположность этому одноцепочечное антитело scFv(TF)3e10 не ингибирует пептидный гидролиз, осуществляемый рТФ/VIIa, а фактически это антитело повышает скорость гидролиза S2266, что позволяет предположить, что scFv(TF)3e10 повышает аффинность рТФ к FVIIa. С помощью анализа способности рТФ/VIIa осуществлять пептидный гидролиз установлено, что антитело scFv(TF)3e10 повышает аффинность FVIIa к рТФ, снижая тем самым кажущееся значение KD в 5 раз (фиг.2). Антитело scFv(TF)3e10 не оказывает воздействия на скорость гидролиза, осуществляемого FVIIa, в отсутствие рТФ, что свидетельствует о том, что это антитело не взаимодействует со свободным FVIIa. Значение KD, характеризующее связывание антитела scFv(TF)3e10 с рТФ, которое определяют с помощью анализа способности рТФ/FVIIa осуществлять пептидный гидролиз, составляет 65,4 нМ (фиг.4). Микрокалориметрический анализ применяют для сравнения аффинности scFv(TF)3e10 к ТФ и к комплексу FVIIa/ТФ. Эти эксперименты позволяют установить, что экспрессируемое в клетках млекопитающих scFv(TF)3e10 имеет ˜ в 20 раз более высокую аффинность к комплексу ТФ/FVIIa, чем к свободному рТФ (33 нМ против 600 нМ, фиг.5).
Шесть ТФ-связывающих одноцепочечных антител, экспрессируемых в бактериях, сравнивают с использованием анализа активации FX и анализа ПВ. Ни одно из пяти антител, которые ингибируют скорость гидролиза S2266 с помощью комплекса рТФ/FVIIa, а именно scFv(TF)2c1, scFv(TF)2c11, scFv(TF)2d3, scFv(TF)2h6 и scFv(TF)3h2, не удлиняют ПВ по сравнению с контролем (ЗФР-буфер) (фиг.3). В противоположность этому, хотя антитело scFv(TF)3e10 не обладает самой высокой аффинностью по данным, полученным с помощью BIAcore, и повышает аффинность FVIIa к рТФ, scFv(TF)3e10 представляет собой единственное антитело в группе, которое ингибирует активацию FX (фиг.6 и не представленные данные) и пролонгирует время свертывания при анализе ПВ (фиг.3). Значение IC50, которое характеризует способность антитела scFv(TF)3e10 (димер) ингибировать активацию FX, составляет 0,44 нМ (фиг.6). И, наконец, анализ активации FX используют для определения того, может ли антитело scFv(TF)3e10 конкурировать с FX за связывание с комплексом FVIIa/ТФ. Антитело scFv(TF)3e10 в зависимости от дозы ингибирует активацию FX, при этом значение КD(арр) остается одинаковым при всех концентрациях субстрата FX, что свидетельствует о том, что scFv(TF)3e10 не конкурирует с FX и не связывается с ТФ или с комплексом FVIIa/ТФ в том же сайте, что и FX (фиг.7).
Антитело scFv(TF)3e10 выявлено на основе связывания с рекомбинантным человеческим растворимым ТФ. Гомология последовательности ТФ у человека и мыши или человека и кролика составляет 58 и 71% соответственно. Вероятно, антитело связывается с уникальным эпитопом на человеческом ТФ, который оказывает воздействие на активацию FX комплексом FVIIa/ТФ. Физиологически это антитело имеет преимущество по сравнению с антителами, которые конкурируют с FVII или FVIIa за связывание с ТФ. Значение KD, характеризующее связывание FVIIa с растворимым ТФ, составляет ˜10 нМ (фиг.2), это значение согласуется с опубликованными значениями для связывания FVIIa с рТФ (4,8 нМ, Neuenschwander P.F. и Morrissey J. H., J. Biol. Chem. 269(11), 1994, сс.8007-8013) или для связывания FVII, FVIIa и инактивированного с помощью DIP FVIIai с полноразмерным ТФ, восстановленным в нейтральных фосфатидилхолиновых пузырьках (Bach R. и др. Biochemistry 25, 1986, сс.4007-4020). Аффинность связывания FVII или FVIIa с полноразмерным ТФ повышается в значительной степени, когда заряженный фосфотидилом серин включают в фосфолипидные пузырьки (Bach R. и др. (1986), выше), из-за взаимодействия GLA-домена FVII или FVIIa с заряженной мембранной поверхностью (Neuenschwander P. F. и Morrissey J. H. (1994), выше). В этих оптимальных условиях FVIIa связывается с полноразмерным ТФ с очень высокой аффинностью (41 пМ). У антитела к ТФ, которое конкурирует за связывание с FVII или FVIIa, должна быть затруднена способность конкурировать за связывание с этим обладающим высокой аффинностью комплексом FVIIa/ТФ. В противоположность этому антитело scFv(TF)3e10 не только обладает более высокой аффинностью к комплексу FVIIa/ТФ, чем к ТФ, но и не конкурирует с FVIIa за связывание с ТФ. Антитело, такое как scFv(TF)3e10, которое не конкурирует с FVIIa, должно ингибировать активацию FX не зависимо от концентрации FVII в плазме, которая составляет ˜10 нМ.
Антитело scFv(TF)3e10 также имеет преимущество по сравнению с антителами, которые конкурируют с FX за связывание с ТФ. Значение Кm, характеризующее связывание FX с комплексом VIIa/ТФ, составляет 0,061-0,099 мкМ, что видно из данных, представленных на фиг.7, это согласуется с опубликованными значениями (0,1 мкМ, Baugh R. J. и др., J. Biol. Chem. 275(37), 2000, сс.28826-28833). Концентрация FX в человеческой плазме составляет 140 нМ (значение Кm выше в 1,4-2 раза). Антитело, такое как scFv(TF)3e10, которое не конкурирует с FX, что видно из фиг.7, должно ингибировать активацию FX не зависимо от концентрации FX в плазме.
Пример 6
Модель тромбоэмболии in vivo, созданная на крысах
ТФ-связывающее одноцепочечное антитело scFv(TF)3e10 является специфичным в отношении ТФ приматов. Модель тромбоэмболии, вызванной человеческим ТФ (с применением в качестве реагента тромбопластина, содержащего человеческий рекомбинантный ТФ, фирма Ortho), создают у находящихся в сознании самцов крыс Sprague-Dawley (350-400 г, n>7/группу). На этой модели диссеминированного внутрисосудистого свертывания крови (ДВС) ТФ в результате инъекции тромбопластина индуцирует отложение фибрина в легких, вызывая тем самым нарушение дыхания и смерть. Определенные дозы клеток млекопитающих, экспрессирующих scFv(TF)3e10, или носителя инъецируют в хвостовую вену, а затем через 15 мин осуществляют болюсную инъекцию тромбопластина (0,5 мл/кг). В группе, обработанной носителем, эта доза ТФ вызывает гибель 60% особей (LD60), как правило, через 5 мин инъекции тромбопластина. Состояние крыс оценивают с помощью следующей балльной системы заболеваемости-смертности: 0 - отсутствие действия; 1 - слабый респираторный дистресс-синдром (восстановление через 30 мин); 2 - серьезный респираторный дистресс-синдром (состояние агонии, для восстановления требуется более 60 мин; и 3 - смерть. Для сравнения эффективности лечения различных групп используют средний балл. Результаты, полученные в этом анализе in vivo, представлены на фиг.8. В этом опыте установлено, что антитело, предлагаемое в изобретении, обладает способностью снижать смертность и уменьшать респираторный дистресс-синдром.
Описанные выше примеры можно воспроизводить, получая аналогичный результат, заменяя в целом или некоторые описанные реактанты и/или условия проведения испытаний, применяемые при осуществлении вышеприведенных примеров.
Несмотря на то, что изобретение проиллюстрировано на примере получения определенных конструкций антител, очевидно, что можно осуществлять вариации и модификации без отклонения от сущности и объема изобретения.
Реферат
Настоящее изобретение относится к биотехнологии. Описано антикоагулирующее антитело, которое связывается с большей аффинностью с комплексом FVIIa/ТФ, чем со свободным ТФ. Раскрыта фармацевтическая композиция для предупреждения и лечения тромбоза, содержащая описанное антитело. Предложен способ предупреждения образования тромба, заключающийся в том, что вводят терапевтически эффективное количество описанного антитела. Также предложен способ снижения и лечения тромбоза глубоких вен (ТГВ), диссеминированного внутрисосудистого свертывания крови (ДВС), острого коронарного тромбоза или рака с признаками коагулопатии у пациента, заключающийся в том, что пациенту вводят терапевтически эффективное количество описанного антитела. Раскрыт способ регуляции воспалительного ответа у пациента, заключающийся в том, что пациенту вводят терапевтически эффективное количество описанного антитела. Представлена полинуклеотидная последовательность, кодирующая описанное антитело. Настоящее изобретение позволяет получить антитело, которое обладает большей аффинностью при связывании с комплексом фактор FVIIa/ТФ, чем при связывании со свободным тканевым фактором. 10 н. и 14 з.п. ф-лы, 8 ил.
Формула
Документы, цитированные в отчёте о поиске
Способ получения белка, обладающего опосредованной фактором vii активностью в свертывании крови
Комментарии