Применение моноклональных антител, специфичных к о-ацетилированной форме ганглиозида gd2, для лечения некоторых форм рака - RU2462476C2
Код документа: RU2462476C2
Чертежи












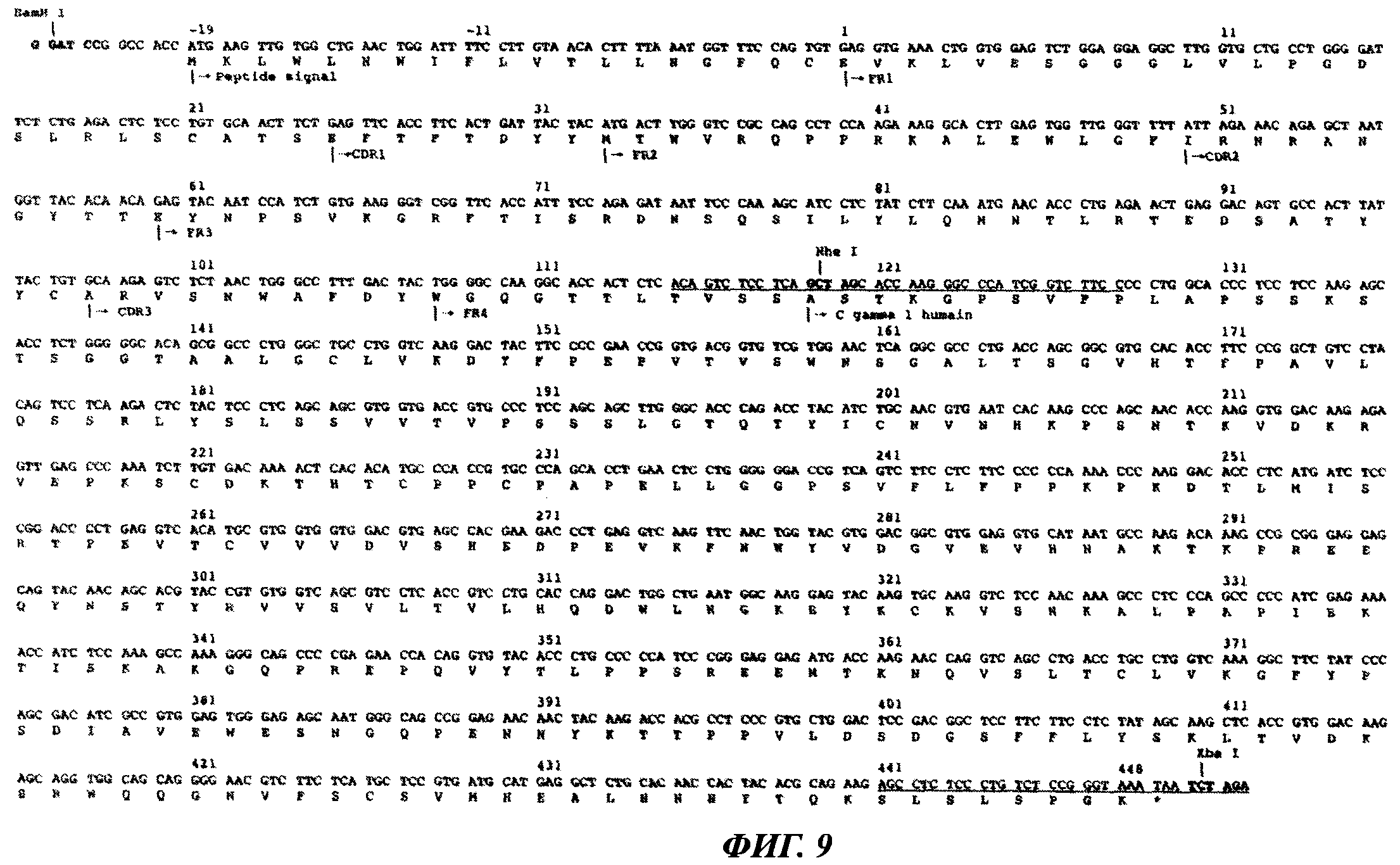







Описание
Область исследовании
Данное изобретение имеет целью создание новых средств для диагностики и терапии форм рака, при котором клетки экспрессируют O-ацетилированную форму ганглиозида GD2. Эти средства в основном включают применение моноклональных рекомбинантных антител, фрагменты которых распознают O-ацетилированный GD2 и не распознают здоровые клетки и нервные волокна периферической нервной системы. Эти новые средства будут более специфичными в отношении раковых клеток и снизят токсичность лечения по сравнению с антителами и производными, применявшимися ранее в лечении таких форм рака.
Предшествующий уровень техники
Опухолевые клетки имеют на своей поверхности определенное количество антигенных детерминант. Среди этих антигенных детерминант некоторые являются специфичными в отношении опухоли. Эти опухолевые антигены экспрессируются преимущественно или исключительно на поверхности раковых клеток.
Эти опухолевые антигены индуцируют выработку иммунной системой различных животных, например мышей, молекул, обозначаемых антителами, отличающихся способностью точно распознавать молекулы антигенов, вызвавших их выработку.
Такое специфическое распознавание между моноклональным антителом и его антигеном человеческой опухоли представляет главный интерес для иммунного фокусирования опухолей. Действительно, оно позволяет сконцентрировать на уровне пораженных органов или тканей детектирующие агенты с целью диагностики или токсические агенты с целью лечения опухолей. Эти детектирующие или токсические агенты могут быть химическими или биологическими соединениями, возможно радиоактивными, искусственным образом присоединенными к антителам до или после их введения. Эти агенты могут также быть биологическими соединениями, естественным образом присутствующими у пациентов (компоненты комплемента, хемокины, цитокины, цитотоксические клетки, например, Т или NK лимфоциты), направляемыми к опухолевым клеткам при помощи антител.
Ганглиозиды представляют собой компоненты клеточной мембраны, и некоторые из них были охарактеризованы как опухолевые антигены. Было продемонстрировано, что ганглиозид GD2 усиленно экспрессируется у человека раковыми клетками нейроэктодермального происхождения, например при меланомах, глиобластомах, мелкоклеточных карциномах легких и нейробластомах. Перечень опухолевых патологий, приведенный здесь, не является исчерпывающим. Ретинобластомы и остеосаркомы также экспрессируют ганглиозид GD2. Определенные формы рака яичников также экспрессируют ганглиозид GD2. Ганглиозид GD2 является кислым гликолипидом, образованным керамидом, связанным с олигосахаридом, последовательность которого представляет собой глюкозу, галактозу и М-ацетил-галактозамин. К молекуле галактозы присоединены две молекулы сиаловых кислот. Концевая сиаловая кислота может быть модифицирована путем присоединения O-ацетильной группировки с образованием O-ацетилированного GD2. С помощью специфических антител к GD2, распознающих также его O-ацетилированную форму, было показано, что определенные формы рака, а именно опухоли нейроэктодермального происхождения, экспрессируют GD2 и его O-ацетилированную форму O-ацетилированный GD2 в различных пропорциях.
Были описаны различные моноклональные антитела к GD2, полученные на мышах. Терапевтическое применение моноклональных антител к GD2, 14.G2a и 3F8, относящихся соответственно к иммуноглобулинам мышей класса lgG2a и lgG3, было протестировано в клинических исследованиях.
Данные исследования показали существование ряда отрицательных побочных эффектов, присущих моноклональным мышиным антителам. Одним из таких отрицательных эффектов является аллергическая или анафилактическая реакция, которая может являться следствием иммунного ответа, развиваемого человеческим организмом против антител мыши.
Для решения данной проблемы иммуногенности моноклональных антител, полученных на мышах, возможно разработать химерные антитела с помощью генной инженерии. «Химерным» антителом называется антитело, модифицированное генетически, в котором более или менее важная часть генетической информации, кодирущей мышиное антитело, заменена на соответствующую часть человеческого антитела. Как правило, вариабельные участки легких и тяжелых цепей антител грызунов (VH и VL) оставляются неизмененными, тогда как остальная часть молекулы берется от человеческого антитела. Такие химерные антитела сохраняют способность специфически распознавать опухолевые антигены, присущую моноклональным антителам мыши, в то же время обладая физическими и эффекторными свойствами, присущими человеческому антителу. Так, клинические исследования показали, что химерные антитела ch.14G2a и ch.14.18 обладают сниженной иммуногеностью и увеличенным периодом полужизни в сыворотке по сравнению с моноклональными антителами мыши, на основе которых они получены. Кроме того, было показано, что моноклональные антитела мыши к GD2 при их химеризации приобретают повышенную способность рекрутировать эффекторные клетки человека, тогда как мышиные антитела обладают только ограниченной способностью рекрутировать эффекторные клетки человека.
Можно продвинуться еще дальше в отождествлении антител грызунов с антителами человека, если от антитела грызунов взять только аминокислоты участков, обозначаемых CDR (от англ. complementary determining regions), формирующих сайт связывания антигена и таким образом определяющих специфичность антител. Генетически модифицированные антитела, в которых сохранены только аминокислоты, расположенные вблизи таких участков CDR исходного антитела мыши, а остальная часть молекулы происходит от антитела человека, обозначаются «гуманизированые».
Недостатки предшествующего уровня техники.
Терапевтическое или диагностическое применение in vivo моноклональных антител требует, чтобы они одновременно обладали определенными показателями специфичности, аффинности и нетоксичности. Однако клинические исследования, проведенные на моноклональных и химерных антителах к GD2, выявили основной отрицательный побочный эффект, не устраняемый при химеризации или гуманизации, а именно нейротоксичность анти-GD2 антител. Действительно, пациенты, имеющие рак нейроэктодермального происхождения, которым вводили анти-GD2 антитела, испытывали боли на фоне введения антител, а у некоторых из этих пациентов развилась нейропатия периферической нервной системы. Данный эффект объясняется присутствием ганглиозида GD2 на поверхности клеток нервных волокон периферической системы.
Такая нейротоксичность, свойственная антителам к GD2, ограничила распространение в клинике иммунотерапии с применением этих антител. В то же время большое число антигенных мишеней GD2 остается фактором, вызывающим большой интерес для иммунного фокусирования опухолей нейроэктодермального происхождения.
Одним из подходов, который бы позволил разрешить проблему нейротоксичности, является использование антител, распознающих опухолевый антиген, ассоциированный с раком нейроэктодермального происхождения, отличный от ганглиозида GD2, и не распознающих нервные волокна периферической системы.
Несколько лет назад изобретатели провели серию иммунизации мышей для получения антител к GD2. Был выделен ряд анти-GD2 антител и их характеризация позволила идентифицировать одно антитело класса lgG3 мыши, обозаченное 8В6, специфически распознающее молекулу ганглиозида GD2, слегка модифицированную благодаря наличию O-ацетильной группы. Данные работы и, в частности, антитело 8В6 описаны в публикации « Variable Region Gene Segments of Nine Monoclonal Antibodies Specific to Disialogangliosides (CG2, GD3) and their O-Acetylated Derivatives », Cerato et al, Hybridoma Volume 16, Number 4, 1997, 307-316, включенной сюда во всей ее полноте путем ссылки.
Была изучена структура этого антитела, но работы над ним не были продолжены. Действительно, это антитело не представляло никакого особенного интереса по сравнению с другими антителами к GD2. В отличие от неацетилированных форм GD2, распределение в тканях человеческого организма O-ацетилированного GD2 остается очень мало документированным по причине отсутствия чувствительных методов определения, таких как иммунологические методы, которые основаны на использовании моноклональных антител, специфичных к ацетилированной форме GD2. Было предположено, что клиническое применение этого нового антитела будет вероятно представлять аналогичные проблемы с нейротоксичностью, как и применение моноклональных антител к GD2. Кроме того, не было выявлено никакой цитотоксической активности антитела 8В6, а антитела lgG3 не являются удобными в обращении из-за их склонности к агглютинации.
Задачи изобретения
Моноклональные антитела, специфичные к O-ацетилированному ганглиозиду GD2, которые не распознают ганглиозид GD2, могут обладать преимуществом нераспознавания нервных волокон периферической системы и, следовательно, не вызывать нейротоксичность. Такие антитела могут представлять большое преимущество по сравнению с моноклональными антителами анти-CD2 при иммунном фокусировании определенных типов злокачественных опухолей человека, таких как злокачественных опухолей нейроэктодермального происхождения, экспрессирующих ганглиозид GD2 в O-ацетилированной форме.
Такие антитела могут быть получены разными способами, известными специалистам. Часть таких методик начинается с иммунизации животных, в частности грызунов, и среди них - мышей, крыс или хомячков, зайцеобразных, и в частности кроликов, верблюдовых, и среди них - лам. Иммуноген, композиция, содержащая антиген, против которого требуется получить антитело, может быть либо собственно гликолипидом, очищенным или содержащимся в неочищенной или частично очищенной смеси, либо гликолипидом или одной из его частей, а именно гидрофильной гликозилированной частью, связанными ковалентной или нековалентной связью с белками или липидами, служащими «носителем», стимулирующим иммунный ответ. При другой процедуре иммунизации иммуноген может быть приготовлен из клеток, экспрессирующих антиген, полученных при культивировании клеток линий опухолевых клеток или первичных клеток, полученных из опухолевой ткани. Среди подходящих клеточных линий можно упомянуть, в частности, линии клеток животных, например, тимомы грызунов EL-4, или человека, например, нейобластомы человека IMR32, глиобластомы человека U87MG, мелкоклеточной карциномы человека HCI-Н82 и меланомы человека М21. Эти клетки могут вводиться в целом виде, фиксированными или живыми, или подвергаться фракционированию для введения, например, только частично очищенной фракции, содержащей компоненты клеточной мембраны или цитоплазмы.
Эти иммуногены могут вводиться животным различными способами, в частности подкожно, интраперитонеально или внутримышечно, самостоятельно или в присутствии адъюванта, в частности адъюванта на основе алюминия или адъюванта Фрейнда. Иммунизации могут повторяться с частотой от нескольких дней до нескольких месяцев.
Для получения антител для использования в промышленном применении существует 3 основных способа. Первый состоит во взятии крови иммунизированных животных и выделении из нее с помощью известных специалистам способов фракций, содержащих более или менее очищенные антитела (сыворотки или плазмы, общих иммуноглобулинов). Антитела могут быть далее очищены с помощью хроматографии или иммуноадсорбции. Второй способ заключается в том, чтобы взять клетки, способные синтезировать антитела, в частности клетки селезенки или лимфоузлов, и их обессмертить, в частности путем вирусной трансформации или соматической гибридизации согласно известным специалистам способам. Среди обессмерченных клеток с помощью клонирования можно селекционировать клетки, продуцирующие антитело, представляющее интерес, а при культивировании клеток выделить из культуральной жидкости антитела и очистить их в больших количествах. Наконец, третий способ состоит в изоляции РНК клеток, способных синтезировать антитела, в частности клеток селезенки, лимфоузлов или периферических лимфоцитов, выделенных у иммунизированных или неиммунизированных животных, или даже человека, и создании библиотек кДНК, из которых путем скрининга согласно методам, известным специалистам, в частности путем экспрессии этих библиотек на поверхности бактериофагов, известной под названием системы экспрессии фаговый дисплей, будут выбраны последовательности антител, представляющих интерес. Создание комбинаторных библиотек участков VH и VL человека, экспрессируемых на поверхности нитевидных фагов («phage display») в форме одноцепочечных фрагментов (scFv, от англ. «single chain Fv»), соединяющих участок VH и участок VL или в форме фрагментов F(ab), состоящих из легкой цепи, связанной с пептидным отрезком VH-CH1, где СН1 соответствует первому домену константного участка.
Антитела согласно данному изобретению могут быть выбраны из группы антител, полученных тем или иным описанным выше способом, отличающихся способностью распознавать в качестве антигена O-ацетилированный GD2, но не GD2, не прошедший O-ацетилирование, это свойство будет обозначено необходимой специфичностью. Такая селекция может быть проведена в ходе выделения антител, например, при размножении исключительно клеток, производящих антитела, обладающие необходимой специфичностью. В альтернативном варианте можно выбирать среди уже выделенных антител те, которые обладают такой специфичностью. С этой целью можно использовать различные способы, в частности непрямую иммунофлуоресценцию клеток для того, чтобы отличить те антитела, которые распознают клетки, известные способностью экспрессировать O-ацетилированный GD2, например IMR32, от антител, которые распознают также клетки, экспрессирующие GD2, но не O-ацетилированный GD2, например Neuro 2A. Можно также использовать иммуноферментный тест ELISA на высушенных клетках, для селекционирования антител, фиксирующихся исключительно на клетках, экспрессирующих O-ацетилированный GD2, но не на клетках, экспрессирующих только GD2. Можно также селекционировать антитела, обладающие необходимой специфичностью, путем тонкослойной хроматографии на силикагеле для разделения компонентов гликолипидов клеток, экспрессирующих как O-ацетилированный GD2, так и GD2, в частности IMR32, и отбора антител, маркирующих исключительно полосу, соответствующую O-ацетилированному GD2. Результаты можно подтвердить путем разрушения O-ацетилированного GD2 в липидном экстракте при алкилировании и исчезновении маркировки. Примеры внедрения таких способов проверки специфичности антител согласно данному изобретению приведены ниже в подробном описании изобретения. Специалистами могут быть разработаны и другие способы, нацеленные на аналогичный результат.
Антитела согласно данному изобретению могут быть модифицированы с помощью ряда методик, известных специалистам, для адаптации к различным способам применения. Известные способы предшествующего уровня техники также могут быть использованы для получения одноцепочечных антител scFv, a также ряда гибридных белков, сохраняющих способность связывать антиген. В частности, известно, что участки антител, обозначаемые константными, могут быть полностью модифицированы, например с целью замены константных участков грызунов на константные участки человеческих антител без потери распознавания антигена. Создание химерного антитела состоит в выделении ДНК, кодирующей участок VH и участок VL моноклонального антитела мыши, и ее слиянии с ДНК, кодирующей константные участки Н и L иммуноглобулина человека. Подобная генетическая конструкция позволяет произвести гибридное антитело, у которого константная часть человеческая и является неиммуногенной или слабоиммуногенной в организме человека (как правило, подразумевается константный участок lgG1 человека и участок Скаппа человека). Возможно также консервировать исключительно участки незаменимые для распознавания антигена, называемые гипервариабельными участками, участки, определяющие комплементарность, или CDR, и заменять все остальные, чтобы произвести так называемую «гуманизацию» антител. Для упрощения описания совокупность таких белков будет обозначаться как «искусственно модифицированные антитела».
Некоторые из таких белков представляют особенный интерес в качестве антител согласно данному изобретению. Например, искусственно модифицированные антитела к O-ацетилированному GD2, сохраняющие способность распознавать O-ацетилированный ганглиозид GD2, но не распознающие ганглиозид GD2, могут представлять большое преимущество по сравнению с моноклональными мышиными антителами к O-ацетилированному ганглиозиду GD2 по своим улучшенным физическим и эффекторным характеристикам.
Искусственно модифицированные антитела, специфичные к O-ацетилированному GD2, должны обладать достаточной для соответствующего антигена аффинностью, чтобы максимально предотвращать диффузию в нормальные ткани этого антитела, возможно несущего токсическое или терапевтическое соединение. В рамках настоящего изобретения, аффинность к ганглиозиду GD2 должна превосходить 10-7 молекул/литр. Аффинность данных антител может быть повышена способами, известными специалистам. Так, экспрессия фрагментов антител на поверхности бактериофагов представляет собой важное средство для поиска высокоаффинных мутантов исходного scFv. Возможно также воспроизводить усиление аффинности, наблюдающееся в ходе развития иммунного ответа. Способы получения случайных мутаций или направленных мутаций, после которых следует селекция путем повторяющихся циклов иммуноадсорбции/элюции, были также успешно воплощены и позволили получить фрагменты антител, обладающие аффинностью, почти в 10 раз большей по сравнению с исходным фрагментом.
Отдельная задача данного изобретения, которая не должна быть лимитирована данным примером, касается искусственно модифицированных антител, произведенных на основе антитела 8В6, о котором известно, что оно обладает необходимой специфичностью, а также рекомбинантных белков, имеющих последовательности CDR, описываемые SEQ ID NO: 3-8.
Также задачей изобретения является предложение модификаций антител, обладающих необходимой специфичностью, для придания им свойств, необходимых для диагностики или лечения некоторых форм рака. Для некоторых воплощений требуется введение антител или их производных больным, страдающим определенными формами рака. В этом случае производные, последовательность которых в большей мере приближается к последовательности человеческого антитела, будут предпочтительными, поскольку они в состоянии ограничить выработку лечащимся пациентом антител против вводимой молекулы, таким образом повышая толерантность и позволяя проведение повторных инъекций. Такие производные также способны облегчать выработку антител к специфическим детерминантам вводимого антитела, так называемых анти-идиотипических антител, представляющих терапевтическое значение.
Для терапевтического применения предпочтительными модификациями являются те, которые позволяют вызвать цитотоксическую активность против опухолевых клеток, экспрессирующих интересующий антиген, в данном случае O-ацетилированный GD2.
Известно, что только некоторые классы иммуноглобулинов обладают способностью активировать комплемент или индуцировать клеточную цитотоксичность. Например, антитело 8В6 распознает O-ацетилированный GD2, но не способно индуцировать достаточно эффективно такую цитотоксичность. Таким образом, еще одна задача данного изобретения заключалась в том, чтобы предложить антитела, распознающие O-ацетилированный GD2 и обладающие цитотоксической активностью. Этого можно добиться при замещении константных участков антител константными участками антител, способными вызывать такую цитотоксичность, а именно константными участками иммуноглобулинов человека класса 1. Цитотоксическая активность далее может быть усилена, в частности, при продукции антител, особым образом гликозилированных и незначительно фукозилированных, например, при их продукции специально отобранными или трансформированными клетками. Для получения аналогичного эффекта в последовательность антител также могут быть введены специфические мутации, описанные в литературе.
Другой способ получения терапевтических антител согласно данному изобретению заключается в связывании производного антитела, обладающего необходимой специфичностью, с цитотоксическим агентом. Указанный цитотоксический агент может являться токсическим химическим агентом, антисмысловыми РНК, и в частности противоопухолевым цитотоксическим препаратом, среди которых можно упомянуть таксаны, алкалоиды барвинка и их производные, антрациклины, алкилирующие агенты, биологическим токсическим агентом, в частности растительными или бактериальными токсинами, среди которых можно назвать рицин или токсин псевдомонад, или даже радиоактивным изотопом, испускающим бета-частицы, например йод-131, иттрий-90, лютеций-177, рений-186 или медь-67, или Оже-электроны, например индий-111, или альфа-частицы, например висмут-213, висмут-212 или астат-211, данные примеры ни в коей мере не должны лимитировать пределы изобретения. Антитела могут также быть получены с помощью методов молекулярной инженерии, известных специалистам, в форме гибридного белка, связанного с цитокином. Используемые цитокины в основном являются представителями семейства интерлейкинов, такими как интерлейкин-2 (IL-2), IL-4, IL-5, IL-6, IL-7, IL-10, IL-12, IL-13, IL-14, IL-15, IL-16 и IL-18, гематопоэтических ростовых факторов, такими как ГМ-КСФ (гранулоцитарно-макрофагальный колониестимулирующий фактор) или Г-КСФ (гранулоцитарный колониестимулирующий фактор), фактор некроза опухоли (ФНО), хемокинов. Полученный при слиянии антитела и цитокина белок обладает биологическими свойствами цитокина и специфичностью антитела, на основе которых он получен.
Антитела согласно данному изобретению могут также иметь преимущества при применении в диагностике, как при использовании in vitro для детекции присутствия антигена O-ацетилированного GD2 в образцах клеток или биологических жидкостей согласно любой методике, известной специалистам, так и при использовании in vivo при введении производного антитела, модифицированного для возможной детекции с помощью одной из медицинских визуализирующих методик, например сцинтиграфии или позитронной эмиссионной томографии. В таком случае производное антитела должно быть связано с радиоактивным изотопом, испускающим гамма-фотоны, таким как йод-131, йод-123, индий-111 или технеций-99 для сцинтиграфической визуализации или томографии одиночных фотонов, или с изотопом, испускающим позитроны, таким как фтор-18, йод-124, иттрий-86, медь-64, скандий-44, при позитронной эмиссионной томографии, причем данные примеры не должны ограничивать область изобретения.
Антитела согласно данному изобретению могут быть связаны с токсичными или радиоактивными компонентами. Токсические соединения связываются химически ковалентным образом с антителами при помощи ряда химических связей, например эфирными, амидными, дисульфидными или тиоэфирными связями. Радиоактивные атомы связываются либо непосредственно путем электрофильного замещения (в случае изотопов йода) или нуклеофильного замещения (в случае фтора-18) или посредством радиоактивно меченого реактивного синтона, в частности реактива Болтона и Хантера для изотопов йода или станнилированных активированных эфиров в случае изотопов йода или астата-211, или также при помощи хелатирующего агента в случае радиоактивного металла. В последнем случае специалисты могут выбрать хелатирующий агент, образующий с металлом стабильный комплекс в биологических жидкостях. Так, DTPA может использоваться преимущественно с индием-111, a DOTA будет предпочтителен для маркировки иттрием-90.
Антитела согласно данному изобретению могут применяться в методах, известных специалистам, в которых токсический или детектируемый агент не связан с антителом непосредственно, а напротив, связан с низкомолекулярной молекулой, вводимой на втором этапе после введения пациенту производного антитела, способного распознавать in vivo эту мелкую молекулу. В этом случае производное антитела представляет собой биспецифичное антитело или иммуноконъюгат или белок, образованный при слиянии производного антитела и авидина. Такой подход может применяться предпочтительно с антителами согласно данному изобретению для диагностики in vivo и лечения опухолей.
Антитела согласно данному изобретению, а также «производные», «производные антител» или «производные соединения» могут быть преимущественно использованы в диагностике и лечении опухолей. Среди них можно отметить опухоли нейроэктодермального происхождения, и в частности меланомы, мелкоклеточный рак легких, глиомы и нейробластомы. Соединения согласно данному изобретению могут предпочтительно использоваться для детекции или лечения таких опухолей, в частности, когда они являются диссеминированными или не поддаются существующим способам лечения.
Краткое описание изобретения.
Данное изобретение описывает применение антител для иммунного фокусирования опухолей человека нейроэктодермального происхождения, например меланом, глиобластом, мелкоклеточных карцином легких и нейробластом.
В ряду различных аспектов данного изобретения, описанных ниже, наиболее удивительным является тот факт, что одно из моноклональных антител, специфичных к O-ацетилированному GD2, не фиксируется на нервных волокнах, экспрессирующих GD2, тогда как оно распознает опухолевые клетки, экспрессирующие ганглиозид GD2 и его O-ацетилированную форму. Такие антитела обладают специфичностью исключительно в отношении опухолевых клеток нейроэктодермального происхождения и не распознают нервные волокна периферической системы. Следствием такой повышенной специфичности является снижение токсичности при терапевтическом применении, что приводит, в частности, к отсутствию фиксации на нормальной периферической нервной ткани, наблюдающейся у антител, распознающих ганглиозид GD2. Данное исследование включает применение таких антител для диагностики и лечения рака с повышенной специфичностью и сниженной токсичностью по сравнению с антителами, распознающими GD2.
Антитела согласно данному изобретению имеют цитотоксическую активность по отношению к опухолевым клеткам, которые они распознают, либо по своей сущности (это одна из причин, по которой антитела химеризуют или гуманизируют), либо поскольку их используют в качестве векторов для токсических агентов, в частности радиоактивных.
Данное изобретение таким образом включает все моноклональные антитела, химерные или гуманизированные, которые распознают исключительно O-ацетилированную форму ганглиозида GD2 или фрагмент этого антитела, указанные антитела или указанный фрагмент распознают молекулы O-ацетилированного GD2, экспрессируемые опухолевыми клетками, и не распознают молекулы GD2, экспрессируемые на поверхности периферических нервов.
Данное изобретение также включает антитела, у которых некоторые аминокислоты были заменены на другие с помощью методов молекулярной генетики, известных специалистам, в частности для модификации свойств исходного антитела, а именно для снижения его иммуногенности или повышения его токсической активности или даже для ускорения или замедления его выведения после инъекции.
Предпочтительно, искусственно модифицированное антитело или его фрагмент согласно изобретению характеризуются тем, что являются IgG каппа с аффинностью выше 10-7 моль/литр по отношению к O-ацетилированному GD2 и аффинностью по меньшей мере в 10 раз ниже по отношению к самому GD2, указанное антитело или указанный фрагмент могут быть моно- или биспецифичными.
В частности, но не исключительно, изобретение включает любое моноклональное искусственно модифицированное антитело или его фрагмент, в котором участки, определяющие комплементарность вариабельного участка цепи Н, имеют аминокислотные последовательности, приведенные в SEQ ID NO:3, SEQ ID NO:4 и SEQ ID NO:5, а участки, определяющие комплементарность вариабельного участка цепи L, имеют аминокислотные последовательности, приведенные в SEQ ID NO:6, SEQ ID NO:7 и SEQ ID NO:8.
В частности, но не исключительно, изобретение включает любое монокпональное искусственно модифицированное антитело или его фрагмент, в котором тяжелая цепь получена путем слияния кДНК, кодирующей вариабельный участок тяжелой цепи антитела, полученного не у человека, и кДНК, кодирующей константный участок иммуноглобулина человека, а легкая цепь получена путем слияния кДНК, кодирующей вариабельный участок легкой цепи того же самого антитела, полученного не у человека, и кДНК, кодирующей константный участок легкой цепи иммуноглобулина человека, характеризующееся тем, что указанное антитело, полученное не у человека, является моноклональным антителом мыши 8В6, а указанное антитело, модифицированное искусственным образом, направлено против O-ацетилированной формы ганглиозида GD2 и не распознает нервные волокна периферической системы.
В частности, но не исключительно, изобретение включает любое моноклональное искусственно модифицированное антитело или его фрагмент, у которого вариабельный участок тяжелой цепи имеет последовательность аминокислот, приведенную в SEQ ID NO:1, а вариабельный участок легкой цепи имеет последовательность аминокислот, приведенную в SEQ ID NO:2.
Предпочтительно, моноклональное искусственно модифицированное антитело или его фрагмент согласно п.5 является антителом КМ8В6, которое можно получить с помощью клеточной линии СНО, или его фрагментом.
Изобретение также включает любую фармацевтическую молекулу, производную искусственно модифицированного антитела или его фрагмента согласно изобретению, в которой антитело или его фрагмент связаны с молекулой X, где Х является токсической молекулой, лекарством, предшественником лекарства или вторым антителом с любой специфичностью.
В одном воплощении указанная токсическая молекула является химической, биологической или радиоактивной токсической молекулой, указанная молекула предназначена для разрушения опухолевых клеток, экспрессирующих O-ацетилированный ганглиозид GD2.
Предпочтительно указанные опухолевые клетки, против которых направлены фармацевтические молекулы согласно изобретению, представляют собой клетки нейробластомы, меланомы, глиобластомы или мелкоклеточного рака легких.
Предпочтительно фармацевтические молекулы согласно данному изобретению изменены на уровне участка Fc путем присоединения Сахаров, для модулирования таким образом активации иммунных клеток и молекул системы комплемента.
Изобретение также включает любую молекулу для диагностики рака, обнаруживающую экспрессию O-ацетилированного ганглиозида GD2 на поверхности опухолевых клеток, указанная молекула является производной искусственно модифицированного антитела или его фрагмента согласно данному изобретению, в которой указанное антитело или указанный фрагмент связан с агентом, обусловливающим детекцию антитела или указанного фрагмента с помощью флуоресценции или радиоактивности.
Изобретение касается любого применения искусственно модифицированного антитела или его фрагмента согласно изобретению и/или молекулы согласно изобретению для производства лекарственных препаратов для лечения рака, при котором клетки экспрессируют O-ацетилированную форму ганглиозида GD2, или для производства соединения для диагностики такой формы рака.
Данное изобретение также включает любое применение моноклонального антитела 8В6 (описанного в публикации « Variable Region Gene Segments of Nine Monoclonal Antibodies Specific to Disialogangliosides (CG2, GD3) and their O-Acetylated Derivatives », Cerato et al, Hybridoma Volume 16, Number 4, 1997, 307-316) для производства фармацевтических молекул, в которых указанное антитело связано с химическим, биологическим или радиоактивным токсическим агентом, указанная молекула предназначена для разрушения опухолевых клеток, экспрессирующих O-ацетилированный ганглиозид GD2.
В частности, но не исключительно, изобретение включает такое применение, при котором указанные клетки являются клетками нейробластомы, меланомы, глиобластомы или мелкоклеточного рака легких.
В частности, но не исключительно, изобретение включает такое применение, при котором указанная терапевтическая молекула изменена на уровне участка Fc путем присоединения сахаров, для модулирования таким образом активации иммунных клеток и молекул системы комплемента.
Изобретение также включает любое применение моноклонального антитела 8В6 для производства молекулы для диагностики рака, демонстрирующей экспрессию O-ацетилированного ганглиозида GD2 на поверхности опухолевых клеток, указанная молекула является производной указанного антитела, в которой указанное антитело связано с агентом, обусловливающим детекцию антитела с помощью флуоресценции или радиоактивности.
Изобретение включает также любую последовательность ДНК, кодирующую искусственно модифицированное антитело согласно данному изобретению, а также любой вектор экспрессии, содержащий такую последовательность ДНК, связанную эффективно с промотором.
Изобретение включает также любую клетку, в частности животную, содержащую такой вектор экспрессии, а также любой трансформант не из организма человека, продуцирующий искусственно модифицированное антитело согласно изобретению.
Наконец, изобретение включает любой способ продукции искусственно модифицировнного антитела против O-ацетилированного ганглиозида GD2, включающий экспрессию последовательности ДНК в клетке или трансформанте не из организма человека в соответствующих условиях и получение антитела.
Предпочтительно, клетку или трансформант культивируют в условиях, при которых происходит накопление антитела.
Краткое описание графического материала.
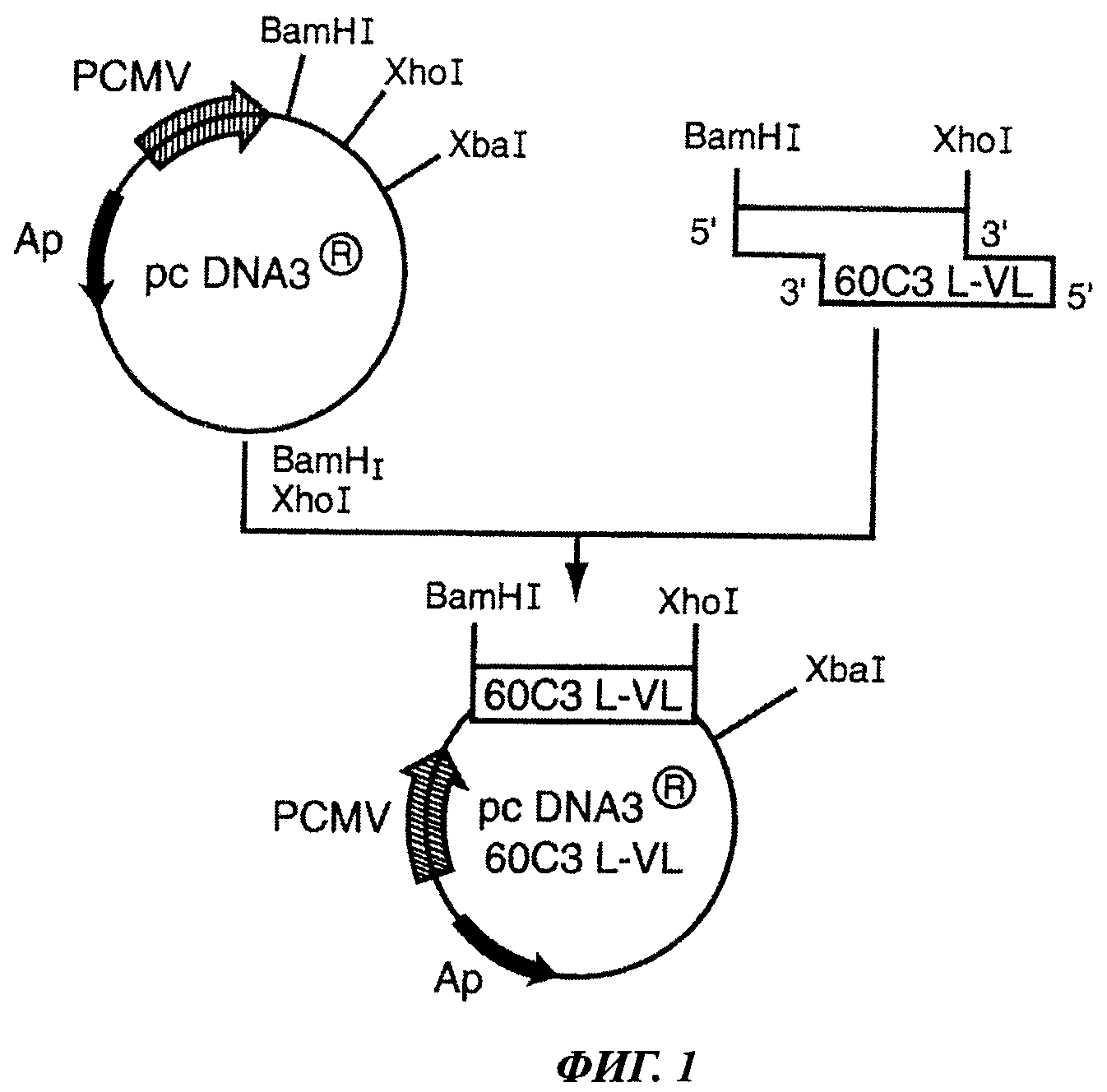
Фиг.1 отображает конструкцию плазмиды pcDNA3®60C3 L-VL.
Фиг.2 отображает конструкцию плазмиды pcDNA3®60C3 L.
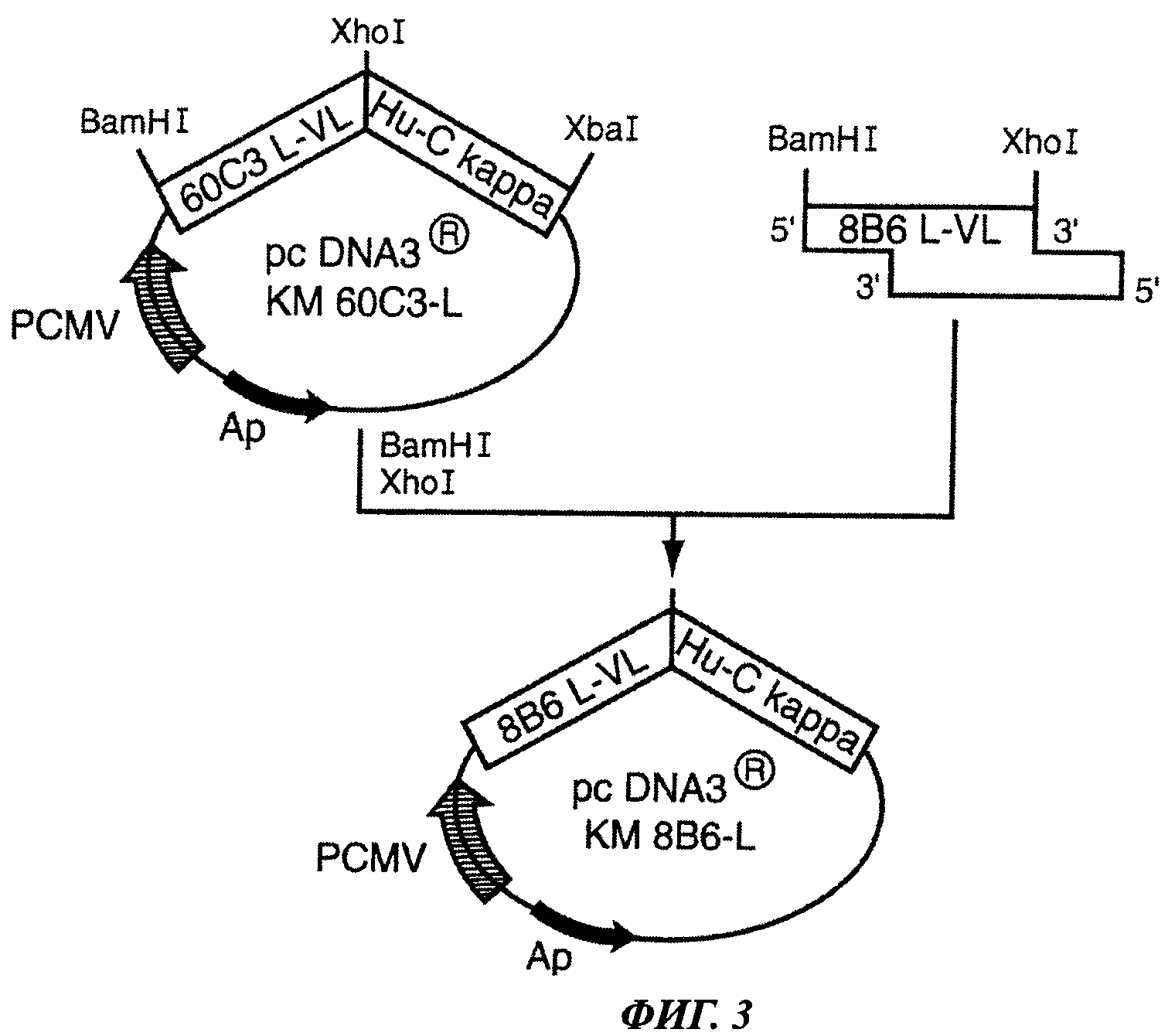
Фиг.3 отображает конструкцию плазмиды pcDNA3® KM8B6L.
Фиг.4 отображает нуклеотидную последовательность и последовательность аминокислот легкой цепи искусственно модифицированного антитела КМ8В6.
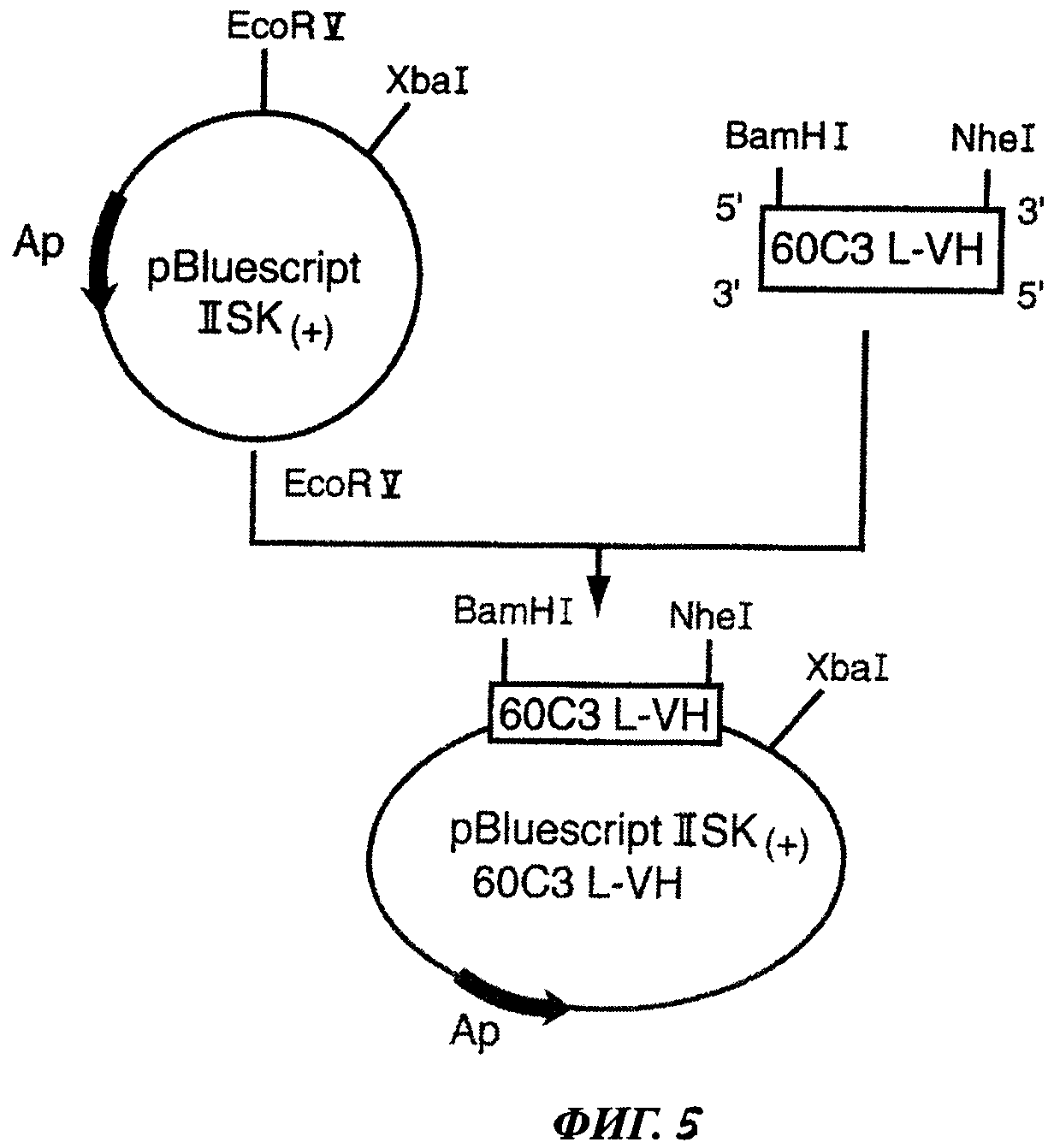
Фиг.5 отображает конструкцию плазмиды pBluescript® II SK (+) 60С3 L-VH.
Фиг.6 отображает конструкцию плазмиды pBluescript® II SK (+) КМ60С3-Н.
Фиг.7 отображает конструкцию плазмиды pBluescript® II SK (+) КМ8В6-Н.
Фиг.8 отображает конструкцию плазмиды pcDNA3.1/Hygro© (+) КМ8В6-Н.
Фиг.9 отображает нуклеотидную последовательность и последовательность аминокислот тяжелой цепи искусственно модифицированного антитела КМ8В6.
Фиг.10 отображает SDS-электрофорез в ПААГ искусственно модифицированного очищенного антитела КМ8В6, направленного против O-ацетилированного GD2. Анализ проведен в редуцирующих условиях (слева) и нередуцирующих условиях (справа). Слева направо, маркер молекулярного веса, КМ8В6, lgG3 8B6 (редуцирующие условия), высокомолекулярный маркер, КМ8В6, и lgG3 8B6 (нередуцирующие условия).
Фиг.11 представляет график, отображающий реактивность антитела 8B6 и антитела КМ8В6 в отношении клеток IMR 32 и NeuroA, соответственно имеющих или не имеющих антигены O-ацетилированного GD2, зарегистрированную при помощи иммунофлуоресценции с отложенным по оси ординат количеством детектированных клеток, а по оси абсцисс - интенсивности флуоресценции. Голубой график соответствует реактивности контроля, а красный график соответствует реактивности продуктов.
Фиг.12 представляет график, отображающий реактивность антител 8B6 и КМ8В6 в отношении клеток IMR 32 и NeuroA, проанализированную с помощью теста ELISA.
Фиг.13 показывает результаты иммуноокрашивания ганглиозидов мозга крысы, разделенных тонкослойной хроматографией на силикагеле, выполненного с применением антитела КМ8В6. Панель А - окраска резорцином миграции различных ганглиозидов мозга крысы.
Фиг.14 отображает результаты иммуногистохимического анализа клеток нейробластомы нервных волокон человека.
Фиг.15 отображает результаты экспериментов по измерению цитотоксичности (ADCC, от англ. antibody-dependent cellular cytotoxicity, антитело-зависимая клеточная цитотоксичность). Процентное выражение активности ADCC искусственно модифицированного антитела КМ8В6 и моноклонального антитела мыши 8 В6, на основе которого оно было получено. *Антитело Rituxan® к CD20 использовано в качестве отрицательного контроля.
Фиг.16 отображает результаты эксперимента in vivo на мышах противоопухолевого эффекта моноклонального антитела 8В6 против клеток линии тимомы мышей EL4, экспрессирующих антиген O-ацетилированного GD2.
Подробное описание изобретения.
Продукция искусственно модифицированного антитела КМ8В6
1. Получение ДНК, кодирующей вариабельный участок L-VL моноклонального антитела мыши 60С3
(а) Экстракция общей РНК гибридомы 60С3, продукция моноклонального антитела 60С3
Общую РНК выделяли из 106 клеток гибридомы 60С3 в экспоненциальной фазе роста при помощи реагента RNAble (EUROBIO, Courtaboeuf, France) согласно инструкциям производителя. Концентрацию общей РНК определяли путем измерения оптической плотности при 260 нм.
(б) Получение нуклеотидной последовательности ДНК 60С3 L-VL
Амплификация ДНК, кодирующей вариабельный участок 60С3 L-VL, была проведена с использованием мессенджерной РНК с помощью RACE-PCR, в ходе которой была получена нуклеотидная последовательность, кодирующая сигнальный пептид (L), связанный со своим вариабельным участком (VL). Такая амплификация была проведена с помощью набора реагентов SMARTTM RACE cDNA Amplification, приобретенного у компании BD Biosciences (San Jose, CA, USA), согласно инструкциям производителя. Количество общей РНК, использованной для обратной транскрипции, составило 1 мкг. Продукт реакции развели в 100 мкл буферного раствора трицин ЭДТА, прилагаемого производителем. Для амплификации было использовано 2,5 мкл раствора продукта. Использовали следующий праймер анти-сенс, специфичный для кДНК 60С3 VL: 3'- 60С3 VL: 5'-ТТТ CAG CTC CAG СТТ GGT CCC AGC -3' (SEQ ID NO:9). Амплификацию проводили при инкубации реакционной смеси в термоциклере Perkin-Elmer (РЕ) DNA thermal Cycler 480 (Perkin Elmer Wellesley, MA, USA) при следующих условиях: 5 циклов (94°С в течение 5 сек, 72°С в течение 3 мин), затем 5 циклов (94°С в течение 5 сек, затем 70°С в течение 10 сек и 3 мин при 72°С) и 25 циклов (94°С в течение 5 сек, затем 69°С в течение 10 сек и 3 мин при 72°С). Продукт реакции RACE-PCR анализировали путем электрофореза в 1% агарозном геле (Q. Biogene, Morgan Irvine, CA, USA) при миграции в буферном растворе Трис ЭДТА с рН 8,0 (40 мМ Tris-base (Sigma Chemicals Co), 25 мМ EDTA (Inerchim, Montlucon, France), 20 мМ уксусная кислота (Carlo Erba Reagenti SpA, Rodano, Ml, Italy). Продукты, имеющие предполагаемый молекулярный вес, затем очищали с использованием набора реагентов QIAquick® Gel Extraction (Qiagen) согласно инструкциям производителя. Определение нуклеотидной последовательности полученных очищенных продуктов проводили в компании GENOME express (Meylan, France) для проверки последовательности ДНК, кодирующей участок L-VL 60С3. Таким образом была определена последовательность, соответствующая сигнальному пептиду, связанному с вариабельным участком легкой цепи антитела 60С3, и были разработаны праймеры для клонирования кДНК 60С3 L-VL в вектор экспрессии.
(в) Амплификация кДНК 60С3 L-VL
Амплификация кДНК 60С3 L-VL была проведена в ходе реакции RT-PCR с использованием экстракта общей РНК клеток гибридомы 60С3. Состав реакционной смеси: Oligo d(T)18 0,5 мкг (New England Biolabs Inc. Beverly, MA, USA), PHK 1 мкг, dNTP 0,5 мМ (Promega, Madison, WI, USA), стерильная вода до 12 мкл. Данную смесь инкубировали в течение 5 мин при 65°С (в твердотельном термостате), затем 2 мин при 4°С (на тающем льду) для денатурации РНК. Затем в реакционную смесь добавляли 4 мкл буферного раствора First-strand 5X (Invitrogen Life Biotechnologies), 10 мМ DTT (Invitrogen Life Biotechnologies), 160 U Rnasine (PROMEGA) и 800 U обратной транскриптазы (Invitrogen Life Biotechnologies). Смесь инкубировали в течение 1 часа при 37°С, затем при 70°С в течение 15 мин для остановки реакции. Копии кДНК 60С3 L-VL получали путем амплификации в ходе реакции PCR с использованием следующих синтетических олигонуклеотидов:
5'-BamHI 60С3 L-VL: 5'-AG GGA TCC AAA GAC AAA ATG GAT-3' (праймер сенс) и 3'-XhoI 60С3 L-VL: 5'-TT CAG CTC GAG СТТ GGT CCC AGC ACC-3' (праймер анти-сенс).
Использованные синтетические праймеры позволили произвести молчащую мутацию в последовательности ДНК, которая не приводила к каким-либо заменам на уровне аминокислотной последовательности и позволяла создать сайты рестрикции BamHI и XhoI, необходимые для клонирования в вектор.
Реакционная смесь имела следующий состав: dNTP (PROMEGa) 500 мкМ, Taq-полимераза (Invitrogen Life Biotechnologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Biotechnologies) 10 мкл, кДНК (матрица) 1 мкл, MgCl2 (Invitrogen Life Biotechnologies) 1,5 мМ, праймер 3'-XhoI 60C3 L-VL и праймер 5'-BamHI 60C3 по 500 мкМ.
Амплификацию проводили в термоцикпере MJ-research PTC 200 (Peltier Thermal Cycler) в следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С.
После верификации продуктов ПЦР с помощью электрофореза в 1% агарозном геле кДНК 60C3 L-VL очищали с использованием набора реагентов QIAquick® Gel Extraction (Qiagen) согласно инструкциям производителя.
2. Конструирование рекомбинантной плазмиды pcDNA3® 60C3 L-VL, имеющей кДНК 60C3 L-VL
(а) Расщепление вектора pcDNA3® и кДНК 60C3 L-VL и дефосфорилирование расщепленного вектора
Расщепление рестрикционными ферментами позволяет получить линейный вектор, имеющий липкие концы, необходимые для встраивания требующейся кДНК. Расщепление проводится при следующих условиях: буфер NEB2 10Х (5 мкл), BSA 100Х (0,5 мкл), вектор pcDNA3® или кДНК 60C3 L-VL (1 мкг), рестриктаза BamHI (10 U), рестриктаза XhoI (10 U). Рестриктазы, реакционный буферный раствор и раствор бычьего сывороточного альбумина (BSA) были приобретены в New England Biolabs Inc. Реакцию проводили в течение 3 ч в водяной бане при 37°С.Расщепленные вектор и вставку очищали с использованием набора реагентов QIAquick® Gel Extraction (Qiagen) согласно инструкциям производителя. Затем вектор дефосфорилировали с использованием CIP (New England Biolabs INC.) в течение 1 ч при 37°С. Реакционная смесь имела следующий состав: буфер NEB3 10Х (5 мкл), вектор (1 мкг), CIP (1 мкл). Данная реакция способствует увеличению производительности процедуры лигации путем снижения уровня самолигирования вектора. Действительно, подавление фосфатных группировок на 5' и 3' концах, высвобожденных в ходе рестрикции, препятствует спонтанному замыканию вектора.
(б) Реакция лигирования
Лигирование позволяет поместить обработанную рестриктазами кДНК 60С3 L-VL в обработанный рестриктазами и дефосфорилированный вектор pcDNAS® с помощью Т4 лигазы (New England Biolabs Inc.). Реакцию проводили на водяной бане при 16°С в течение 16 ч в следующей реакционной среде: лигазный буферный раствор 5Х (4 мкл) (New England Biolabs Inc.), дефосфорилированный обработанный рестриктазами вектор 200 нг, Т4 лигаза (400 U) (New England Biolabs Inc.), очищенная кДНК 170 нг (для вставки размером 1 кб). Молярное соотношение вставка/вектор составляла 3/1.
(в) Трансформация компетентных бактерий Е. coli XL1 blue (Stratagene)
20 мкл продукта лигазной реакции добавляли к 100 мкл суспензии компетентных бактерий. Смесь инкубировали при 4°С в течение 30 мин. Затем бактерии подвергали тепловому шоку в течение 2 мин при 42°С, а затем инкубации в течение 2 при 4°С. Затем прибавляли 1 мл среды LB и бактерии инкубировали в течение 1 ч при 37°С при помешивании (250 об/мин). Бактерии собирали центрифугированием в течение 5 мин при 4000 g при 4°С. Бактериальный осадок растворяли в 100 мкл среды LB для посева в чашках Петри с агаром 2XTY, содержащим 100 мкг/мл ампициллина (Sigma Chemicals Co) и 12,5 мкг/мл тетрациклина (Sigma Chemicals Co). Устойчивые колонии появлялись после инкубации в течение ночи при 37°С.
(г) Верификация наличия в составе вектора pcDNA3® вставки 60С3 L-VL при помощи ПЦР
Изолированные колонии помещали в стерильных условиях в ламинаре в отдельные пробирки, содержащие реакционную смесь для ПЦР: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, кДНК (матрица) 1 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 mM, праймер 3'-XhoI 60С3 L-VL и праймер 5'-BamHI 60С3 по 500 мкМ.
Амплификацию проводили в термоциклере MJ-research PTC 200 при следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С. Размер амплифицированных продуктов ПЦР контролировали с помощью электрофореза в 1% агарозном геле. Клоны, трансформированные должным образом, были единственными, дающими продукты ПЦР ожидаемого размера.
(д) Получение плазмиды pcDNA3® 60С3 L-VL
С помощью стерильной зубочистки колонии переносили в 5 мл жидкой селективной среды LB, содержащей 100 мкг/мл ампициллина (Sigma Chemicals Co) и 12,5 мкг/мл тетрациклина (Sigma Chemicals Co), и инкубировали в течение 16 ч при 37°С. Бактериальную культуру затем использовали для мини-продукции плазмидной ДНК с использованием набора реагентов QIAprep spin miniprep (QIAGEN) согласно инструкциям производителя.
3. Получение кДНК, кодирующей константный участок hu-C kappa моноклонального антитела человека P3Non2
(а) Выделение общей РНК гибридомы Р3Non2, продуцирующей моноклональные антитела P3Non2.
Общую РНК выделяли из 106 клеток гибридомы P3Non2 в экспоненциальной фазе роста при помощи реагента RNAble (Eurobio, Courtaboeuf, France) согласно инструкциям производителя. Концентрацию общей РНК определяли путем измерения оптической плотности при 260 нм.
(б) Амплификация кДНК hu-C kappa
Амплификация кДНК hu-C kappa была проведена в ходе реакции RT-PCR с использованием экстракта общей РНК клеток гибридомы Р3Non2. Реакионная смесь имела следующий состав: Oligo d(T)18 0,5 мкг (New England Biolabs Inc. Beverly, MA, USA), PHK 1 мкг, dNTP 0,5 мМ (Promega, Madison, WI, USA), стерильная вода до 12 мкл. Данную смесь инкубировали в течение 5 мин при 65°С (в твердотельном термостате), а затем 2 мин при 4°С (на тающем льду) для денатурации РНК. Затем в реакционную смесь добавляли 4 мкл буферного раствора First-strand 5X (Invitrogen Life Biotechnologies), 10 мМ DTT (Invitrogen Life Biotechnologies), 160 U Rnasine (Promega) и 800 U обратной транскриптазы (Invitrogen Life Biotechnologies). Смесь инкубировали в течение 1 ч при 37°С, затем при 70°С в течение 15 мин для остановки реакции. Копии кДНК hu-C kappa были получены при амплификации в ходе реакции ПЦР с использованием следующих синтетических олигонуклеотидов: 5'-XhoI hu-C kappa: 5'-AG CTC GAG CTG AAA CGA ACT GTG GCT GCA С-3' (праймер сенс) и 3'-XbaI hu-C kappa: 5'-СТТ СТА GAT ТТА АСА CTC ТСС ССТ GTT GA-3 ' (праймер анти-сенс).
Использованные синтетические праймеры позволили произвести молчащую мутацию в последовательности ДНК, которая не приводила к каким-либо заменам на уровне аминокислотной последовательности и позволяла создать сайты рестрикции XbaI и XhoI, необходимые для клонирования в вектор.
Реакционная смесь имела следующий состав: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, кДНК (матрица) 1 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 мМ, праймер 3'-XbaI hu-C kappa и праймер 5'-XhoI hu-C kappa no 500 мкМ.
Амплификацию проводили в термоциклере MJ-research PTC 200 (Peltier Thermal Cycler) в следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С.
После верификации продуктов ПЦР с помощью электрофореза в 1% агарозном геле кДНК hu-C kappa очищали с использованием набора реагентов QIAquick® Gel Extraction (Qiagen) согласно инструкциям производителя.
4. Конструкция вектора экспрессии легкой цепи искусственно модифицированного антитела КМ60С3: pcDNA3® KM60C3-L
(а) Расщепление вектора pcDNA3® 60С3 L-VL и кДНК hu-C kappa и дефосфорилирование расщепленного вектора
Расщепление проводится при следующих условиях: буфер NEB2 10Х (5 мкл), BSA 100X (0,5 мкл), вектор pcDNA3® 60С3 L-VL или кДНК hu-C kappa (1 мкг), рестриктаза XhoI (10 U), рестриктаза XbaI (10 U). Рестриктазы, реакционный буферный раствор и раствор бычьего сывороточного альбумина (BSA) были приобретены в New England Biolabs Inc. Реакцию проводили в течение 3 ч в водяной бане при 37°С. Расщепленные вектор и вставку очищали с использованием набора реагентов QIAquick® Gel Extraction (Qiagen) согласно инструкциям производителя. Затем вектор дефосфорилировали с использованием CIP (New England Biolabs Inc.) в течение 1 ч при 37°С. Реакционная среда имела следующий состав: буфер NEB3 10Х (5 мкл), вектор (1 мкг), CIP (1 мкл).
(б) Реакция лигирования
Лигирование позволяет поместить обработанную рестриктазами кДНК hu-C kappa в обработанный рестриктазами и дефосфорилированный вектор pcDNA3® 60С3 L-VL с помощью Т4 лигазы (New England Biolabs Inc.). Реакцию проводили на водяной бане при 16°С в течение 16 ч в следующей реакционной среде: лигазный буферный раствор 5Х (4 мкл) (New England Biolabs Inc.), дефосфорилированный обработанный рестриктазами вектор 200 нг, Т4 лигаза (400 U) (New England Biolabs Inc.), очищенная кДНК 170 нг (для вставки размером 1 кб). Молярное соотношение вставка/вектор составляло 3/1. Продукт лигазной реакции использовали для трансформации компетентных бактерий Е. coli XL1 blue (Stratagene).
(в) Верификация наличия в составе вектора pcDNA3® 60C3 L-VL вставки hu-С kappa при помощи ПЦР
Изолированные резистентные колонии помещали в стерильных условиях в ламинаре в отдельные пробирки, содержащие реакционную смесь для ПЦР: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, кДНК (матрица) 1 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 mM, праймер 3'-XbaI hu-C kappa и праймер 5'-XhoI hu-C kappa по 500 мкМ. Амплификацию проводили в термоциклере MJ-research РТС 200 при следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С. Размер амплифицированных продуктов ПЦР контролировали с помощью электрофореза в 1% агарозном геле. Клоны, трансформированные должным образом, были единственными, дающими продукты ПЦР ожидаемого размера.
(г) Получение плазмиды pcDNA3® KM60C3-L
С помощью стерильной зубочистки колонии переносили в 5 мл жидкой селективной среды LB, содержащей 100 мкг/мл ампициллина (Sigma Chemicals Co) и 12,5 мкг/мл тетрациклина (Sigma Chemicals Co), и инкубировали в течение 16 ч при 37°С. Бактериальную культуру затем использовали для мини-продукции плазмидной ДНК с использованием набора реагентов QIAprep spin miniprep (QIAGEN) согласно инструкциям производителя.
5. Конструкция вектора экспрессии легкой цепи искусственно модифицированного антитела KM 8B6: pcDNA3® 8B6-L
(а) Получение вектора pcDNA3® hu-C kappa
Вектор pcDNA3® hu-C kappa был получен на основе вектора pcDNA3® KM60C3-L путем расщепления с помощью рестрикционных ферментов BamHI и XhoI. Реакционная среда имела следующий состав: буфер NEB2 10Х (5 мкл), BSA 100Х (0,5 мкл), вектор (1 мкг), рестриктаза BamHI (10 U), рестриктаза XhoI (10 U). Рестриктазы, реакционный буферный раствор и раствор бычьего сывороточного альбумина (BSA) были приобретены в New England Biolabs Inc. Реакцию проводили в течение 3 ч в водяной бане при 37°С. Расщепленный вектор очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя. Затем вектор дефосфорилировали с использованием CIP (New England Biolabs Inc.) в течение 1 ч при 37°С. Реакционная среда имела следующий состав: буфер NEB3 10Х (5 мкл), вектор (1 мкг), CIP (1 мкл).
(б) Выделение общей РНК гибридомы 8В6, продуцирующей моноклональные антитела 8В6
Общую РНК выделяли из 106 клеток гибридомы 8В6 в экспоненциальной фазе роста при помощи реагента RNAble (Eurobio, Courtaboeuf, France) согласно инструкциям производителя. Концентрацию общей РНК определяли путем измерения оптической плотности при 260 нм.
(в) Получение нуклеотидной последовательности кДНК 8В6 L-VL
Амплификация ДНК, кодирующей вариабельный участок 8В6 L-VL, была проведена с использованием мессенджерной РНК с помощью RACE-PCR, в ходе которой была получена нуклеотидная последовательность, кодирующая сигнальный пептид (L), связанный со своим вариабельным участком (VL). Такая амплификация была проведена с помощью набора реагентов SMART™ RACE cDNA Amplification, приобретенного у компании BD Biosciences (San Jose, CA, USA), согласно инструкциям производителя. Количество общей РНК, использованной для обратной транскрипции, составило 1 мкг. Продукт реакции развели в 100 мкл буферного раствора трицин ЭДТА, прилагаемого производителем. Для амплификации было использовано 2,5 мкл раствора продукта. Использовали следующий праймер анти-сенс, специфичный для кДНК 8В6 VL: 3'-mu С kappa: 5'-GTT CAT ACT CGT CCT TGG TCA ACG TGA GGG-3', который гибридизуется с кДНК, кодирующей константный участок mu С kappa легкой цепи типа каппа антитела мыши. Амплификацию проводили при инкубации реакционной смеси в термоциклере Perkin-Elmer (РЕ) DNA thermal Cycler 480 (Perkin Elmer Wellesley, MA, USA) при следующих условиях: 5 циклов (94°С в течение 5 сек, 72°С в течение 3 мин), затем 5 циклов (94°С в течение 5 сек, затем 70°С в течение 10 сек и 3 мин при 72°С) и 25 циклов (94°С в течение 5 сек, затем 69°С в течение 10 сек и 3 мин при 72°С). Продукт реакции RACE-PCR анализировали путем электрофореза в 1% агарозном геле (Q. Biogene, Morgan Irvine, CA, USA) при миграции в буферном растворе Трис ЭДТА с рН 8,0 (40 мМ Tris-base (Sigma Chemicals Co), 25 мМ EDTA (Inerchim, Montlucon, France), 20 мМ уксусная кислота (Carlo Erba Reagenti SpA, Rodano, Ml, Italy)). Продукты, имеющие предполагаемый молекулярный вес, затем очищали с использованием набора реагентов QIAquick® Gel Extraction (Qiagen) согласно инструкциям производителя. Определение нуклеотидной последовательности полученных очищенных продуктов проводили в компании GENOME express (Meylan, France) для проверки последовательности ДНК, кодирующей участок L-VL 8В6. Таким образом была определена последовательность, соответствующая сигнальному пептиду, связанному с вариабельным участком легкой цепи антитела 8В6, и были разработаны праймеры для клонирования кДНК 8В6 L-VL в вектор экспрессии.
(г) Амплификация кДНК 8В6 L-VL
Амплификация кДНК 8В6 L-VL была проведена в ходе реакции RT-PCR с использованием экстракта общей РНК клеток гибридомы 8В6. Состав реакционной смеси: Oligo d(T)18 0,5 мкг (New England Biolabs Inc. Beverly, MA, USA), PHK 1 мкг, dNTP 0,5 мМ (Promega, Madison, WI, USA), стерильная вода до 12 мкл. Данную смесь инкубировали в течение 5 мин при 65°С (в твердотельном термостате), затем 2 мин при 4°С (на тающем льду) для денатурации РНК. Затем в реакционную смесь добавляли 4 мкл буферного раствора First-strand 5X (Invitrogen Life Biotechnologies), 10 мМ DTT (Invitrogen Life Biotechnologies), 160 U Rnasine (Promega) и 800 U обратной транскриптазы (Invitrogen Life Biotechnologies). Смесь инкубировали в течение 1 часа при 37°С, затем при 70°С в течение 15 мин для остановки реакции. Копии кДНК 8В6 L-VL получали путем амплификации в ходе реакции PCR с использованием следующих синтетических олигонуклеотидов: 5'-BamHI 8В6 L-VL: 5'-AAG GGA TCC GCC ACC ATG AAG TTG CCT GTT-3' (праймер сенс) и 3'- XhoI 8В6 L-VL: 5'-CCG TTT TAT CTC GAG CTT GGT CCC-3' (праймер анти-сенс).
Использованные синтетические праймеры позволили произвести молчащую мутацию в последовательности ДНК, которая не приводила к каким-либо заменам на уровне аминокислотной последовательности и позволяла создать сайты рестрикции BamHI и XhoI, необходимые для клонирования в вектор.
Реакционная смесь имела следующий состав: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, кДНК (матрица) 1 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 мМ, праймер 3'-XhoI 8В6 L-VL и праймер 5'-BamHI 8В6 L-VL no 500 мкМ.
Амплификацию проводили в термоциклере MJ-research PTC 200 (Peltier Thermal Cycler) в следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С.
После верификации продуктов ПЦР с помощью электрофореза в 1% агарозном геле кДНК 8В6 L-VL очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя.
(д) Расщепление кДНК 8В6 L-VL
Расщепление кДНК 8В6 L-VL производится с помощью рестрикционных ферментов BamHI и XhoI. Состав реакционной среды следующий: буфер NEB2 10Х (5 мкл), BSA 100X (0,5 мкл), кДНК 8В6 L-VL (1 мкг), рестриктаза BamHI (10 U), рестриктаза XhoI (10 U). Рестриктазы, реакционный буферный раствор и раствор бычьего сывороточного альбумина (BSA) были приобретены в New England Biolabs Inc. Реакцию проводили в течение 3 ч в водяной бане при 37°С. Расщепленную вставку очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя.
(е) Реакция лигирования
Лигирование позволяет поместить обработанную рестриктазами кДНК 8В6 L-VL в обработанный рестриктазами и дефосфорилированный вектор pcDNA3® hu-C kappa с помощью Т4 лигазы (New England Biolabs Inc.). Реакцию проводили на водяной бане при 16°С в течение 16 ч в следующей реакционной среде: лигазный буферный раствор 5Х (4 мкл) (New England Biolabs Inc.), дефосфорилированный обработанный рестриктазами вектор pcDNA3® hu-C kappa 200 нг, Т4 лигаза (400 U) (New England Biolabs Inc.), очищенная кДНК 8В6 L-VL 170 нг (для вставки размером 1 кб). Молярное соотношение вставка/вектор составляло 3/1. Продукт лигазной реакции использовали для трансформации компетентных бактерий Е. coli XL1 blue (Stratagene).
(ж) Верификация наличия в составе вектора pcDNA3® hu-C kappa вставки 8В6 L-VL при помощи реакции ПЦР
Изолированные резистентные колонии помещали в стерильных условиях в ламинаре в отдельные пробирки, содержащие реакционную смесь для ПЦР: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, кДНК (матрица) 1 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 mM, праймер 3'-XhoI 8В6 L-VL и праймер 5'-BamHI 8В6 L-VL по 500 мкМ. Амплификацию проводили в термоциклере MJ-research PTC 200 при следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С. Размер амплифицированных продуктов ПЦР контролировали с помощью электрофореза в 1% агарозном геле. Клоны, трансформированные должным образом, были единственными, дающими продукты ПЦР ожидаемого размера.
(з) Получение плазмиды pcDNA3® КМ8В6 L
С помощью стерильной зубочистки колонии переносили в 5 мл жидкой селективной среды LB, содержащей 100 мкг/мл ампициллина (Sigma Chemicals Co) и 12,5 мкг/мл тетрациклина (Sigma Chemicals Co), и инкубировали в течение 16 ч при 37°С. Бактериальную культуру затем использовали для мини-продукции плазмидной ДНК с использованием набора реагентов QIAprep spin miniprep (QIAGEN) согласно инструкциям производителя.
6. Получение кДНК, кодирующей вариабельный участок L-VH монокпонального антитела мыши 60С3
(а) Получение нуклеотидной последовательности ДНК 60С3 L-VH
Амплификация ДНК, кодирующей вариабельный участок 60С3 L-VH, была проведена с использованием мессенджерной РНК с помощью RACE-PCR, в ходе которой была получена нуклеотидная последовательность, кодирующая сигнальный пептид (L), связанный со своим вариабельным участком (VH). Такая амплификация была проведена с помощью набора реагентов SMART™ RACE cDNA Amplification, приобретенного у компании BD Biosciences (San Jose, CA, USA), согласно инструкциям производителя. Количество общей РНК, использованной для обратной транскрипции, составило 1 мкг. Продукт реакции развели в 100 мкл буферного раствора трицин ЭДТА, прилагаемого производителем. Для амплификации было использовано 2,5 мкл раствора продукта. Использовали следующий праймер анти-сенс, специфичный для кДНК 60С3 VH: 3'-60С3 L-VH: 5'-TGC AGA GAC AGT GAC CAG CAG AGT AGT CCC-3' (праймер анти-сенс), который гибридизуется с кДНК, кодирующей константный участок mu С kappa легкой цепи типа каппа антитела мыши. Амплификацию проводили при инкубации реакционной смеси в термоциклере Perkin-Elmer (РЕ) DNA thermal Cycler 480 (Perkin Elmer Wellesley, MA, USA) при следующих условиях: 5 циклов (94°С в течение 5 сек, 72°С в течение 3 мин), затем 5 циклов (94°С в течение 5 сек, затем 70°С в течение 10 сек и 3 мин при 72°С) и 25 циклов (94°С в течение 5 сек, затем 69°С в течение 10 сек и 3 мин при 72°С). Продукт реакции RACE-PCR анализировали путем электрофореза в 1% агарозном геле (Q. Biogene, Morgan Irvine, CA, USA) при миграции в буферном растворе Трис ЭДТА с рН 8,0 (40 мМ Tris-base (Sigma Chemicals Co), 25 мМ EDTA (Inerchim, Montlucon, France), 20 мМ уксусная кислота (Carlo Erba Reagenti SpA, Rodano, Ml, Italy)). Продукты, имеющие предполагаемый молекулярный вес, затем очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя. Определение нуклеотидной последовательности полученных очищенных продуктов проводили в компании GENOME express (Meylan, France) для проверки последовательности ДНК, кодирующей участок 60С3 L-VH. Таким образом была определена последовательность, соответствующая сигнальному пептиду, связанному с вариабельным участком тяжелой цепи антитела 60С3, и были разработаны праймеры для клонирования кДНК 60С3 L-VH в вектор экспрессии.
(б) Амплификация кДНК 60С3 L-VH
Амплификация кДНК 60С3 L-VH была проведена в ходе реакции RT-PCR с использованием экстракта общей РНК клеток гибридомы 60С3. Состав реакционной смеси: Oligo d(T)18 0,5 мкг (New England Biolabs Inc. Beverly, MA, USA), PHK 1 мкг, dNTP 0,5 мМ (Promega, Madison, WI, USA), стерильная вода до 12 мкл. Данную смесь инкубировали в течение 5 мин при 65°С (в твердотельном термостате), затем 2 мин при 4°С (на тающем льду) для денатурации РНК. Затем в реакционную смесь добавляли 4 мкл буферного раствора First-strand 5X (Invitrogen Life Biotechnologies), 10 мМ DTT (Invitrogen Life Biotechnologies), 160 U Rnasine (Promega) и 800 U обратной транскриптазы (Invitrogen Life Biotechnologies). Смесь инкубировали в течение 1 часа при 37°С, затем при 70°С в течение 15 мин для остановки реакции. Копии кДНК 8В6 L-VL получали путем амплификации в ходе реакции PCR с использованием следующих синтетических олигонуклеотидов: 5'-BamHI 60С3 L-VH: 5'-CAG GAT CCG AAC АСА CTG ACT СТА ACC ATG G-3' (праймер сенс) и 3'-NheI 60С3 L-VH: 5'-T GCT AGC TGC AGA GAC AGT GAC CAG AGT-3' (праймер анти-сенс).
Использованные синтетические праймеры позволили произвести молчащую мутацию в последовательности ДНК, которая не приводила к каким-либо заменам на уровне аминокислотной последовательности и позволяла создать сайты рестрикции BamHI и NheI, необходимые для клонирования в вектор.
Реакционная смесь имела следующий состав: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, кДНК (матрица) 1 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 мМ, праймер 3'-NheI 60С3 L-VH и праймер 5'-BamHI 60С3 L-VH по 500 мкМ.
Амплификацию проводили в термоциклере MJ-research PTC 200 (Peltier Thermal Cycler) в следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С.
После верификации продуктов ПЦР с помощью электрофореза в 1% агарозном геле кДНК 60С3 L-VH очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя.
7. Создание рекомбинантной плазмиды, содержащей кДНК 60С3 L-VH
(а) Создание клонируемого вектора pBluescript II SK (+) 60С3 L-VH
Для помещения последовательности, кодирующей участок 60С3 L-VH в вектор pBluescript II SK (+), вектор обрабатывали рестрикционным ферментом EcoRV (New England Biolabs Inc.), который производит разрез с тупыми концами. Обработка полученной линейной плазмиды Taq-полимеразой в присутствии dTTP позволяет добавить один тимидин на 3'-конце вектора и предотвратить его самолигирование. Taq-ДНК-полимераза, использовавшаяся для получения кДНК 60С3 L-VH, обладает 5'-3' экзонуклеазной активностью, позволяющей добавить концевой аденин на каждом 3'-конце продуктов ПЦР, которые могут быть непосредственно клонированы в такой вектор. Благодаря комплементарности А/Т эффективность лигирования значительно повышается по сравнению с лигированием тупых концов. Продукт лигазной реакции использовали для трансформации компетентных бактерий Е. coli XL1 blue (Stratagene).
(б) Верификация наличия в составе вектора pBluescript II SK (+) вставки 60С3 L-VH при помощи реакции ПЦР
Изолированные резистентные колонии помещали в стерильных условиях в ламинаре в отдельные пробирки, содержащие реакционную смесь для ПЦР: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 mM, праймер 3'-NheI 60С3 L-VH и праймер 5'-BamHI 60С3 L-VH по 500 мкМ. Амплификацию проводили в термоциклере MJ-research PTC 200 при следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С. Размер амплифицированных продуктов ПЦР контролировали с помощью электрофореза в 1% агарозном геле. Клоны, трансформированные должным образом, были единственными, дающими продукты ПЦР ожидаемого размера.
(в) Получение плазмиды pBluescript II SK (+) 60С3 L-VH
С этой целью колонии переносили помощью стерильной зубочистки в 5 мл жидкой селективной среды LB, содержащей 100 мкг/мл ампициллина (Sigma Chemicals Со) и 12,5 мкг/мл тетрациклина (Sigma Chemicals Co), и инкубировали в течение 16 ч при 37°С. Бактериальную культуру затем использовали для мини-продукции плазмидной ДНК с использованием набора реагентов QIAprep spin miniprep (QIAGEN) согласно инструкциям производителя.
(г) Верификация ориентации вставки 60С3 L-VH в составе вектора pBluescript II SK
(+) 60С3 L-VH путем ферментативного расщепления
Наличие вставки в ориентации "сенс" подтверждается путем ферментативного расщепления вектора pBluescript II SK (+) 60С3 L-VH ферментом BamHI (New England Biolabs Inc.) согласно инструкциям производителя. Фактически технология Т клонирования не предопределяет ориентации вставки, которая таким образом может быть вставлена в ориентации сенс или анти-сенс. Вставка 60С3 L-VH ограничивается двумя сайтами рестрикции BamHI только в случае ориентации сенс. Таким образом, после расщепления ферментом BamHI электрофоретический анализ позволяет отличить и отобрать клоны, имеющие кДНК 60С3 L-VH в ориентации сенс.
8. Получение кДНК, кодирующей константный участок антитела человека hu-С gamma 1
(а) Выделение общей РНК линии миеломы человека LP1, продуцирующей тяжелую цепь изотипа гамма 1
Общую РНК выделяли из 106 клеток гибридомы LP1 в экспоненциальной фазе роста при помощи реагента RNAble (Eurobio, Courtaboeuf, France) согласно инструкциям производителя. Концентрацию общей РНК определяли путем измерения оптической плотности при 260 нм.
(б) Амплификация сегмента кДНК hu-C gamma 1
Амплификация кДНК hu-C gamma 1 была осуществлена с использованием экстракта общей РНК клеток гибридомы LP1. Состав реакционной смеси: Oligo d(T)18 0,5 мкг (New England Biolabs Inc. Beverly, MA, USA), PHK 1 мкг, dNTP 0,5 мМ (Promega, Madison, WI, USA), стерильная вода до 12 мкл. Данную смесь инкубировали в течение 5 мин при 65°С (в твердотельном термостате), затем 2 мин при 4°С (на тающем льду) для денатурации РНК. Затем в реакционную смесь добавляли 4 мкл буферного раствора First-strand 5X (Invitrogen Life Biotechnologies), 10 мМ DTT (Invitrogen Life Biotechnologies), 160 U Rnasine (Promega) и 800 U обратной транскриптазы (Invitrogen Life Biotechnologies). Смесь инкубировали в течение 1 часа при 37°С, затем при 70°С в течение 15 мин для остановки реакции. Копии кДНК hu-C gamma 1 получали путем амплификации в ходе реакции PCR с использованием следующих синтетических олигонуклеотидов: 5'-NheI hu-C gamma 1: 5'-CA GCT AGC ACC AAG GGC CCA TCG GTC TTC С-3'(праймер сенс) и 3'-XbaI hu-C gamma 1: 5'-AGC CTC ТСС CTG TCT CCG GGT ААА ТАА ТСТ AGA CG -3' (праймер анти-сенс).
Использованные синтетические праймеры позволили произвести молчащую мутацию в последовательности ДНК, которая не приводила к каким-либо заменам на уровне аминокислотной последовательности и позволяла создать сайты рестрикции NheI и XbaI, необходимые для клонирования в вектор.
Реакционная смесь имела следующий состав: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, кДНК (матрица) 1 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 мМ, праймер 3'-XbaI hu-C gamma 1 и праймер 5'-NheI hu-C gamma 1 по 500 мкМ.
Амплификацию проводили в термоциклере MJ-research PTC 200 (Peltier Thermal Cycler) в следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С.
После верификации продуктов ПЦР с помощью электрофореза в 1% агарозном геле кДНК hu-C gamma 1 очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя.
9. Создание рекомбинантной плазмиды pBluescript II SK (+) КМ60С3 Н, содержащей кДНК 60С3 L-VH, соединенную с кДНК hu-C gamma 1
(а) Расщепление вектора pBluescript II SK (+) 60С3 L-VH и кДНК hu-C gamma 1 и дефосфорилирование расщепленного вектора
Расщепление проводится при следующих условиях: буфер NEB2 10Х (5 мкл), BSA 100X (0,5 мкл), вектор pBluescript II SK (+) 60С3 L-VH или кДНК hu-C gamma 1 (1 мкг), рестриктаза NheI (10 U), рестриктаза XbaI (10 U). Рестриктазы, реакционный буферный раствор и раствор бычьего сывороточного альбумина (BSA) были приобретены в New England Biolabs Inc. Реакцию проводили в течение 3 ч в водяной бане при 37°С. Расщепленные вектор и вставку очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя. Затем вектор дефосфорилировали с использованием CIP (New England Biolabs Inc.) в течение 1 ч при 37°С. Реакционная среда имела следующий состав: буфер NEB3 10Х (5 мкл), вектор (1 мкг), CIP (1 мкл).
(б) Реакция лигирования
Лигирование позволяет поместить обработанную рестриктазами кДНК hu-C gamma 1 в обработанный рестриктазами и дефосфорилированный вектор pBluescript II SK (+) 60С3 L-VH с помощью Т4 лигазы (New England Biolabs Inc.). Реакцию лигирования проводили на водяной бане при 16°С в течение 16 ч в следующей реакционной среде: лигазный буферный раствор 5Х (4 мкл) (New England Biolabs Inc.), дефосфорилированный обработанный рестриктазами вектор pBluescript II SK (+) 60С3 L-VH 200 нг, Т4 лигаза (400 U) (New England Biolabs Inc.), очищенная кДНК hu-C gamma 1 170 нг (для вставки размером 1 кб). Молярное соотношение вставка/вектор составляло 3/1. Продукт лигазной реакции использовали для трансформации компетентных бактерий Е. coli XL1 blue (Stratagene).
(в) Верификация наличия в составе вектора pBluescript II SK (+) 60С3 L-VH вставки hu-C gamma 1 при помощи ПЦР
Из-за использования сайтов рестрикции NheI и XbaI вставка hu-C gamma 1 может оказаться в ориентации сенс или анти-сенс. Причиной этому является последовательность сайтов рестрикции, которая не предопределяет ориентации вставок. Наличие вставки в ориентации сенс подтверждается при помощи реакции ПЦР. Изолированные резистентные колонии помещали в стерильных условиях в ламинаре соответственно в отдельные пробирки, содержащие реакционную смесь для ПЦР: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 mM, праймеры сенс и анти-сенс по 500 мкМ. Амплификацию проводили в термоциклере MJ-research PTC 200 при следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С. Использовали следующие праймеры: 5'-BamHI 60С3 L-VH: 5'-CAG GAT CCG AAC АСА CTG ACT СТА ACC ATG G-3' (праймер сенс) и 3'-XbaI hu-C gamma 1: 5'-AGC CTC TCC CTG TCT CCG GGT AAA TAA TCT AGA CG-3' (праймер анти-сенс).
Если вставка находится в неверной ориентации, праймеры оказываются на одной цепочке амплифицируемой последовательности и амплификации не происходит. Размер амплифицированных продуктов ПЦР контролировали с помощью электрофореза в 1% агарозном геле. Клоны, трансформированные должным образом, были единственными, дающими продукты ПЦР ожидаемого размера.
(г) Получение плазмиды pBluescript II SK (+) КМ60С3 Н
С помощью стерильной зубочистки колонии переносили в 5 мл жидкой селективной среды LB, содержащей 100 мкг/мл ампициллина (Sigma Chemicals Co) и 12,5 мкг/мл тетрациклина (Sigma Chemicals Co), и инкубировали в течение 16 ч при 37°С. Бактериальную культуру затем использовали для мини-продукции плазмидной ДНК с использованием набора реагентов QIAprep spin miniprep (QIAGEN) согласно инструкциям производителя.
10. Конструкция вектора экспрессии тяжелой цепи искусственно модифицированного антитела KM 8B6: pcDNA3/Hygro 8B6-H
(а) Получение вектора pBluescript II SK (+) hu-C gamma 1
Вектор pBluescript II SK (+) hu-C gamma 1 был получен на основе вектора pBluescript II SK (+) КМ60С3 Н путем расщепления с помощью рестрикционных ферментов BamHI и NheI. Реакционная среда имела следующий состав: буфер NEB2 10Х (5 мкл), BSA 100X (0,5 мкл), вектор (1 мкг), рестриктаза BamHI (10U), рестриктаза NheI (10U). Рестриктазы, реакционный буферный раствор и раствор бычьего сывороточного альбумина (BSA) были приобретены в New England Biolabs Inc. Реакцию проводили в течение 3 ч в водяной бане при 37°С. Затем вектор очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя. Затем вектор дефосфорилировали с использованием CIP (New England Biolabs Inc.) в течение 1 ч при 37°С. Реакционная среда имела следующий состав: буфер NEB3 10Х (5 мкл), вектор (1 мкг), CIP (1 мкл).
(б) Выделение общей РНК гибридомы 8В6, продуцирующей моноклональные антитела 8В6
Общую РНК выделяли из 106 клеток гибридомы 8В6 в экспоненциальной фазе роста при помощи реагента RNAble (Eurobio, Courtaboeuf, France) согласно инструкциям производителя. Концентрацию общей РНК определяли путем измерения оптической плотности при 260 нм.
(в) Получение нуклеотидной последовательности ДНК 8В6 L-VH
Амплификация ДНК, кодирующей вариабельный участок 8В6 L-VH, была проведена с использованием мессенджерной РНК с помощью RACE-PCR, в ходе которой была получена нуклеотидная последовательность, кодирующая сигнальный пептид (L), связанный со своим вариабельным участком (VH). Такая амплификация была проведена с помощью набора реагентов SMART™ RACE cDNA Amplification, приобретенного у компании BD Biosciences (San Jose, CA, USA), согласно инструкциям производителя. Количество общей РНК, использованной для обратной транскрипции, составило 1 мкг. Продукт реакции развели в 100 мкл буферного раствора трицин ЭДТА, прилагаемого производителем. Для амплификации было использовано 2,5 мкл раствора продукта. Использовали следующий праймер анти-сенс, специфичный для кДНК 8В6 L-VH: 3'-mu С gamma 3: 5'-TGA TCA ACT CAG TCT TGC TGG CTG GGT GGG-3', который гибридизуется с кДНК, кодирующей константный участок mu С gamma 3 тяжелой цепи типа гамма 3 антитела мыши. Амплификацию проводили при инкубации реакционной смеси в термоциклере PERKIN-ELMER (РЕ) DNA thermal Cycler 480 (Perkin Elmer Wellesley, MA, USA) при следующих условиях: 5 циклов (94°С в течение 5 сек, 72°С в течение 3 мин), затем 5 циклов (94°С в течение 5 сек, затем 70°С в течение 10 сек и 3 мин при 72°С) и 25 циклов (94°С в течение 5 сек, затем 69°С в течение 10 сек и 3 мин при 72°С). Продукт реакции RACE-PCR анализировали путем электрофореза в 1% агарозном геле (Q. Biogene, Morgan Irvine, CA, USA) при миграции в буферном растворе Трис ЭДТА с рН 8,0 (40 мМ Tris-base (Sigma Chemicals Co), 25 мМ EDTA (Inerchim, Montlucon, France), 20 мМ уксусная кислота (Carlo Erba REAGENTI SpA, Rodano, Ml, Italy)). Продукты, имеющие предполагаемый молекулярный вес, затем очищали с использованием набора реагентов QIAquick® Gel Extraction (Qiagen) согласно инструкциям производителя. Определение нуклеотидной последовательности полученных очищенных продуктов проводили в компании GENOME express (Meylan, France) для проверки последовательности ДНК, кодирующей участок 8В6 L-VH. Таким образом была определена последовательность, соответствующая сигнальному пептиду, связанному с вариабельным участком тяжелой цепи антитела 8В6, и были разработаны праймеры для клонирования кДНК 8В6 L-VH в вектор экспрессии pBluescript II SK (+) hu-C gamma 1.
(г) Амплификация сегментов 8В6 L-VH
Амплификация кДНК 8В6 L-VH была проведена в ходе реакции RT-PCR с использованием экстракта общей РНК клеток гибридомы 8В6. Состав реакционной смеси: Oligo d(T)18 1 мкл (New England Biolabs Inc. Beverly, MA, USA), PHK 1 мкг, dNTP 0,5 мМ (Promega, Madison, Wl, USA), стерильная вода до 12 мкл. Данную смесь инкубировали в течение 5 мин при 65°С (в твердотельном термостате), затем 2 мин при 4°С (на тающем льду) для денатурации РНК. Затем в реакционную смесь добавляли 4 мкл буферного раствора First-strand 5X (Invitrogen Life Biotechnologies), 10 мМ DTT (Invitrogen Life Biotechnologies), 160 U Rnasine (Promega) и 800 U обратной транскриптазы (Invitrogen Life Biotechnologies). Смесь инкубировали в течение 1 часа при 37°С, затем при 70°С в течение 15 мин для остановки реакции. Копии кДНК 8В6 L-VH получали путем амплификации в ходе реакции PCR с использованием следующих синтетических олигонуклеотидов: 5'-BamHI 8В6 L-VH: 5'-CCG TCG GAT CCG GCC ACC ATG AAG TTG TGG-3' (праймер сенс) и 3'-NheI 8В6 L-VH: 5'-CGG GGT GCT AGC TGA GGA GAC TGT-3' (праймер анти-сенс).
Использованные синтетические праймеры позволили произвести молчащую мутацию в последовательности ДНК, которая не приводила к каким-либо заменам на уровне аминокислотной последовательности и позволяла создать сайты рестрикции BamHI и NheI, необходимые для клонирования в вектор.
Реакционная смесь имела следующий состав: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, кДНК (матрица) 1 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 мМ, праймер 3'-NheI 8B6 L-VH и праймер 5'-BamHI 8B6 L-VH по 500 мкМ.
Амплификацию проводили в термоциклере MJ-research PTC 200 (Peltier Thermal Cycler) в следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С.
После верификации продуктов ПЦР с помощью электрофореза в 1% агарозном геле кДНК 8B6 L-VH очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя.
(д) Расщепление кДНК 8B6 L-VH
Расщепление кДНК 8B6 L-VH производится с помощью рестрикционных ферментов BamHI и NheI. Состав реакционной среды следующий: буфер NEB2 10Х (5 мкл), BSA 100Х (0,5 мкл), кДНК 8B6 L-VH (1 мкг), рестриктаза BamHI (10 U), рестриктаза NheI (10 U). Рестриктазы, реакционный буферный раствор и раствор бычьего сывороточного альбумина (BSA) были приобретены в New England Biolabs Inc. Реакцию проводили в течение 3 ч в водяной бане при 37°С. Расщепленную вставку очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя.
(е) Реакция лигирования
Лигирование позволяет поместить обработанную рестриктазами кДНК 8B6 L-VH в обработанный рестриктазами и дефосфорилированный вектор pBluescript II SK (+) hu-C gamma 1. Реакцию проводили на водяной бане при 16°С в течение 16 ч в следующей реакционной среде: лигазный буферный раствор 5Х (4 мкл) (New England Biolabs Inc.), вектор pBluescript II SK (+) hu-C gamma 1, дефосфорилированный и обработанный рестриктазами, 200 нг, Т4 лигаза (400 U) (New England Biolabs Inc.), очищенная кДНК 8B6 L-VH 170 нг (для вставки размером 1 кб). Молярное соотношение вставка/вектор составляло 3/1. Продукт лигазной реакции использовали для трансформации компетентных бактерий Е. coli XL1 blue (Stratagene).
(ж) Получение кДНК, кодирующей тяжелую цепь искусственно модифицированного антитела
Для получения кДНК, кодирующей тяжелую цепь искусственно модифицированного антитела, вектор pBluescript II SK (+) КМ8В6 Н расщепляли рестрикционными ферментами BamHI и XbaI. Реакционная среда имела следующий состав: буфер NEB2 10Х (5 мкл), BSA 100X (0,5 мкл), вектор (1 мкг), рестриктаза BamHI (10 U), рестриктаза XbaI (10 U). Рестриктазы, реакционный буферный раствор и раствор бычьего сывороточного альбумина (BSA) были приобретены в New England Biolabs Inc. Реакцию проводили в течение 3 ч в водяной бане при 37°С. Выделенную вставку очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя.
(з) Расщепление вектора pcDNA3/Hygro и дефосфорилирование расщепленного вектора
Расщепление проводится в следующих условиях: буфер NEB2 10Х (5 мкл), BSA 100X (0,5 мкл), вектор (1 мкг), рестриктаза BamHI (10 U), рестриктаза XbaI (10 U). Рестриктазы, реакционный буферный раствор и раствор бычьего сывороточного альбумина (BSA) были приобретены в New England Biolabs Inc. Реакцию проводили в течение 3 ч в водяной бане при 37°С. Расщепленные вектор и вставку очищали с использованием набора реагентов QIAquick® Gel Extraction (QIAGEN) согласно инструкциям производителя. Затем вектор дефосфорилировали с использованием CIP (New England Biolabs Inc.) в течение 1 ч при 37°С. Реакционная среда имела следующий состав: буфер NEB3 10Х (5 мкл), вектор (1 мкг), CIP (1 мкл).
(и) Реакция лигирования
Реакция лигирования позволяет поместить кДНК, кодирующую тяжелую цепь искусственно модифицировнного антитела, в обработанный рестриктазами и дефосфорилированный вектор pcDNA3.1/Hygro. Реакцию лигирования проводили на водяной бане при 16°С в течение 16 ч в следующей реакционной среде: лигазный буферный раствор 5Х (4 мкл) (New England Biolabs Inc.), дефосфорилированный обработанный рестриктазами вектор pcDNA3/Hygro 200 нг, Т4 лигаза (400 U) (New England Biolabs Inc.), кДНК, кодирующая тяжелую цепь искусственно модифицированного антитела 170 нг (для вставки размером 1 кб). Молярное соотношение вставка/вектор составляло 3/1. Продукт лигазной реакции использовали для трансформации компетентных бактерий Е. coli XL1 blue (Stratagene).
(к) Верификация наличия в составе вектора pcDNA3.1/Hygro кДНК, кодирующей тяжелую цепь искусственно модифицированного антитела, при помощи ПЦР
Изолированные резистентные колонии помещали в стерильных условиях в ламинаре в отдельные пробирки, содержащие реакционную смесь для ПЦР: dNTP (Promega) 500 мкМ, Taq-полимераза (Invitrogen Life Technologies) 1 мкл, буферный раствор для ПЦР (Invitrogen Life Technologies) 10 мкл, MgCl2 (Invitrogen Life Technologies) 1,5 mM, праймер 3' и праймер 5' по 500 мкМ. Амплификацию проводили в термоциклере MJ-research PTC 200 при следующих условиях: 5 мин при 94°С, затем следующие 35 циклов по 30 сек при 94°С, 45 сек при 55°С и 1 мин при 72°С. Использовали следующие праймеры: 5'-BamHI 60C3 L-VH: 5'-CCG TCG GAT CCG GCC ACC ATG AAG TTG TGG-3' (праймер сенс) и 3'-NheI 8B6 L-VH: 5'-CGG GGT GCT AGC TGA GGA GAC TGT-3' (праймер анти-сенс).
Размер амплифицированных продуктов ПЦР контролировали с помощью электрофореза в 1% агарозном геле. Клоны, трансформированные должным образом, были единственными, дающими продукты ПЦР ожидаемого размера.
(л) Получение плазмиды pcDNA3.1/Hygro KM8B6 Н
С помощью стерильной зубочистки колонии переносили в 5 мл жидкой селективной среды LB, содержащей 100 мкг/мл ампициллина (Sigma Chemicals Co) и 12,5 мкг/мл тетрациклина (Sigma Chemicals Co), и инкубировали в течение 16 ч при 37°С. Бактериальную культуру затем использовали для мини-продукции плазмидной ДНК с использованием набора реагентов QIAprep spin miniprep (QIAGEN) согласно инструкциям производителя.
11. Трансфекция клеток СНО для экспрессии искусственно модифицированного антитела KM8B6
Клетки СНО использовали в качестве клеток-хозяев для экспрессии и секреции искусственно модифицированного антитела KM8B6. Клетки ко-трансфецировали плазмидами pcDNA3® KM8B6-L и pDNA3.1/Hygro© KM8B6-H, кодирующими соответственно легкую цепь (Фиг.4) и тяжелую цепь искусственно модифицированного антитела KM8B6 (Фиг.9), при помощи набора реагентов PolyFect® kit (QIAGEN GMBH, Hildn, Germany) согласно инструкциям производителя. Трансформированные клетки селекционировали по их устойчивости к антибиотикам geneticin® и hygromycin В® (Invitrogen Life Technologies, Carlsbad, CA, USA). Устойчивые клоны получали при помощи классического способа клонирования путем лимитирующего разведения. Затем их селекционировали по экспрессии и секреции антитела KM8B6 с помощью иммуноферментного теста ELISA. Был выбран один стабильно трансфецированный клон, секретирующий антитело KM8B6 в концентрации 3,8 мкг/мл.
12. Очистка антител 8В6 и КМ8В6
Моноклональные антитела мыши, специфичные к ганглиозидам, были получены из соответствующей культуральной жидкости гибридомы (Cerato et al, 1997). Их очищали при помощи хроматографии на Protein A (GE Healthcare Amersham Bioscience AB, Uppsala, Sweden) согласно протоколу, разработанному изобретателем, ограничивающим необратимую гомофильную агрегацию lgG3 мыши (Chapman et al, 1990). Вначале колонку уравновешивали буфером 0,1 М Tris-HCl рН 7,6. Образец 2 л культуральной жидкости, содержащей 10% уравновешивающего буфера, наносили на колонку со скоростью 1 мл/мин. Затем колонку промывали 10 объемами уравновешивающего буфера. Связанный с Protein А материал элюировали кислым буфером 0,1 М цитрат, 0,3 М NaCl, рН 3,0, затем собирали фракции при немедленной нейтрализации за счет предварительного внесения буфера 1М Tris-HCl, рН 7,6 в пробирки для сбора. Ход очистки контролировали путем регистрации изменения оптической плотности собираемых фракций. Фракции, представляющие интерес, затем диализовали в растворе 0,3 М NaCl в PBS рН 7,4 и стерилизовали путем фильтрации через 0,22 мкм фильтр. Количество белка определяли путем измерения оптической плотности при 280 нм, и затем концентрацию моноклонального антитела доводили до концентрации, меньшей или равной 0,9 мг антитела в мл для предупреждения феномена гомофильной агрегации lgG3 мышей. Очищенные антитела хранили при 4°С до использования.
Искусственно модифицированное антитело КМ8В6 очищали путем аффинной хроматографии на Protein А из культуральной жидкости клеток СНО, происходящих из одного клона, стабильно ко-трансфецированного плазмидами pcDNA3 KM8B6-L и pcDNA3/Hygro KM8B6-H.
Очищенные антитела анализировали при помощи SDS-электрофореза в ПААГ в денатурирующих восстанавливающих условиях. Образцы растворяли в буферном растворе 0,5 М Tris-HCl рН 6,8, содержащем 10% глицерин (VWR, Fontenay sous Bois, France) и 5% β-меркаптоэтанол (Promega). Их кипятили в течение 5 мин, затем анализировали путем SDS-электрофореза в ПААГ согласно методике Laemmli. Белки наносили в количестве 3 мкг в лунки полиакриламидного геля толщиной 1,5 мм с концентрацией верхнего и нижнего геля соответственно 4,5% и 12% (VWR). После проведения электрофореза при 200 В в течение 45 мин при комнатной температуре гель фиксировали и окрашивали Coomassie blue R250 (Sigma Chemicals Co). Молекулярные массы рассчитывали по миграции маркера молекулярного веса Precision Plus Protein® Standards (BIO-RAD).
SDS-электрофорез в 12% ПААГ в денатурирующих восстанавливающих условиях показал, что молекулярные массы химерных цепей L и Н составляют соответственно 25 и 50 кДа. SDS-электрофорез в 6% ПААГ в денатурирующих невосстанавливающих условиях показал, что цепи L и Н соединены корректно и образуют молекулу антитела, и молекулярная масса искусственно модифицированного антитела составляет около 150 кДа.
13. Исследование специфичности антитела КМ8В6
(а) при помощи непрямой иммунофлуоресценции живых клеток
Специфичность антител изучалась на клетках IMR32, экспрессирующих O-ацетилированный GD2, и на клетках Neuro 2A, которые не экспрессируют O-ацетилированный GD2. Клетки в количестве 106 инкубировали с антителами 8В6 или КМ8В6 в течение 30 мин при 4°С. Затем клетки двукратно отмывали буфером PBS-BSA 1% рН 7,4 и инкубировали снова в течение 30 мин в присутствии F(ab)2 антитела козы против иммуноглобулинов мыши или против иммуноглобулинов человека, маркированного изотиоцианатом флуоресцеина (Jackson ImmunoResearch Europe LTD, Cambrideshire, UK) в разведении 1/100 в PBS-BSA 1%. После трех последующих отмывок в PBS, 10 000 клеток анализировали на проточном цитометре FACScan (Becton-Dickinson, Mountain View, CA) в координатах SSC/FL1-H после исключения мертвых клеток и клеточного дебриса. Позитивными считали клетки, имеющие флуоресценцию выше 1 Log после установления уровня фона при помощи контролей, маркированных неспецифически только с помощью вторичного антитела.
Полученные результаты показали, что антитело КМ8В6, как и антитело 8В6 распознает только клетки IMR32.
(б) при помощи иммуно-ферментного теста ELISA на высушенных клетках в 96-луночных планшетах для микротитрования
Специфичность антител исследовали на клетках IMR32, экспрессирующих O-ацетилированный GD2, и на клетках Neuro 2A, которые не экспрессируют O-ацетилированный GD2. Опухолевые клетки, культивируемые in vitro, отделяли от поверхности для культивирования при помощи трипсина. После трех промывок с помощью PBS клетки вносили в плоскодонные планшеты для микротитрования Maxisorp (Nunc A/S, Roskilde, Denmark) в количестве 105 клеток на лунку в объеме 50 мкл PBS. Затем планшеты помещали в инкубатор с температурой 37°С на ночь для испарения PBS. Планшеты можно было использовать немедленно или хранить в течение нескольких месяцев при комнатной температуре до использования. При проведении анализа планшеты вначале инкубировали при покачивании в течение 1 часа при комнатной температуре с 200 мкл буферного раствора PBS рН 7,4, содержащего 1% BSA, для предупреждения неспецифического связывания. Затем планшет инкубировали в течение 2 ч при покачивании при комнатной температуре со 100 мкл раствора антитела, разведенного в буфере PBS, содержащем 0,1% BSA. После трех отмывок, по 200 мкл буфера PBS на каждую, в каждую лунку вносили по 100 мкл раствора F(ab')2 биотинилированных антител, специфичных либо к общим иммуноглобулинам мыши, либо к общим иммуноглобулинам человека (Jackson ImmunoResearch Europe LTD, Cambrideshire, UK), разведенным 1 к 2500 в PBS-BSA 0.1%. После инкубации в течение 1 часа при комнатной температуре и трех отмывок в PBS проводили реакцию в течение 1 часа со стрептавидин-биотиново-пероксидазным раствором (Jackson ImmunoResearch Europe LTD, Cambrideshire, UK) в PBS-BSA 0,1%, который затем удаляли с помощью отмывки. Обнаружение фиксированного комплекса проводили путем добавления 100 мкл субстрата ABTS (Roche Diagnostics GMBH Mannheim, Germany). Оптическую плотность определяли путем считывания на спектрофотометре (Multisan EX, Thermo Electron Corporation, Waltham, MA) при 405 нм с различными интервалами времени.
Полученные результаты показали, что антитело КМ8В6, как и антитело 8В6, фиксируется в дозозависимым образом исключительно на клетках IMR32.
(в) в тонком слое силикагеля
Эксперимент проводили на экстракте общих ганглиозидов клеток IMR32. Ганглиозиды ткани экстрагировали по методике, описанной Ariga et al. (1991). Образец для экстрагирования измельчали в 10 объемах смеси хлороформ/металон (Х:М) (1:1 в объемном отношении) и оставляли при механическом помешивании при комнатной температуре в течение ночи. После фильтрации осадок смешивали с 2,5 объемами (Х:М) (1:1 в объемном отношении) и внось перемешивали в течение 6 часов. Затем смесь фильтровали и оба фильтрата выпаривали при низком давлении в ротационном выпаривателе. Сухой остаток затем наносили на колонку, содержащую 2 мл DEAE-Sephadex A-25 в форме ацетата (Sigma Chemicals Co). Нейтральные липиды удаляли при помощи 15 мл растворителя А. Ганглиозиды затем элюировали 15 мл метанола, содержащего 0,4 М ацетат натрия (Sigma Chemicals Co). К полученной фракции добавляли 30 мл PBS рН 7,4 для обессоливания на колонке Sep-Pak column (Waters Co., Milford, MA, USA) с гидрофобным гелем С18 по методике McLuer's (1990). Гель С18 вначале обрабатывали метанолом в количестве двух объемов колонки, затем смесью метанол/ PBS 1:2 (в объемном отношении). Экстракт для обессоливания затем наносили на колонку со скоростью 1 мл/мин. Углеводные цепочки ганглиозидов взаимодействовали путем гидрофобных взаимодействий с гелем, а соли и другие негидрофобные молекулы удалялись при помощи дистиллированной воды в количестве двух объемов колонки. Гликосфинголипиды затем элюировали одним объемом метанола и затем одним объемом смеси хлороформ/метанол 2:1 (в объемном отношении). Ганглиозиды, элюированные с этой колонки, концентрировали в подходящем объеме смеси Х/М 2:1 (в объемном отношении) и хранили при -20°С.
Затем ганглиозиды разделяли при помощи тонкослойной хроматографии. Данный метод позволяет оценить общий профиль ганглиозидов. Планшеты HPTLC состоят из силикагеля 60 (MERCK), покрывающего алюминиевую фольгу. Нанесенные ганглиозиды мигрируют в течение 20 мин при комнатной температуре в кювете с раствором для миграции. Этот растворитель (подвижная фаза) состоит из смеси X/M/CaCl2 в концентрации 0,22% в воде (50:45:10 в объемном отношении). Чем более сиалированным является ганглиозид, тем более он полярный и менее подвижный. Детекция ганглиозидов на планшете осуществляется химическим способом при помощи ректива резорцин/НСl (Svennerholm, 1963). Этот реактив реагирует только с сиаловой кислотой, которая характерна для ганглиозидов. После инкубации с резорцином проявляется несколько окрашенных полос, соответствующих миграции ганглиозидов общего экстракта. Идентификация разделенных ганглиозидов проводится либо с помощью моноклональных специфических антител, либо путем сравнения со стандартными ганглиозидами, использующимися в качестве маркеров, мигрировавшими одновременно с экстрактом ганглиозидов, подлежащих анализу.
После миграции ганглиозидов в тонком слое силикагеля планшет погружали в 0,01% раствор поли-(изобутил)-метакрилата в гексане на 1 мин и затем высушивали на воздухе. Это делало планшет пластичным и предотвращало отделение геля от основы во время последующих этапов. Затем выполняли тест ELISA на высушенных клетках за исключением этапа выявления фиксации антител на ганглиозидах, которое проводилось при помощи раствора 4-хлоро-1-нафтола (Sigma Aldrich Chemie GmbH, Stenheim, Germany), приготовленного непосредственно перед анализом из расчета 1 мг вещества, растворенного в 1 мл метанола, разведенного в 20 мл PBS с добавлением 30 мкл 30 объемов перекиси водорода.
Результаты показали, что антитело КМ8В6, как и антитело 8В6, распознает O-ацетилированный GD2 только в отсутствие алкалирующей обработки экстракта ганглиозидов.
14. Изучение распределения ганглиозида GD2 и его O-ацетилированной формы в нервной системе и в опухолевых тканях
Образцы опухолей (глиобластомы, нейробластомы, меланомы, рака легких) были получены при хирургической резекции. Образцы периферических нервов человека получали из дистальной чувствительной ветви мышечно-кожного нерва, латеральной ветви перонеального нерва. Речь идет о взятии образцов для диагностики периферической нейропатии или поражении передних роговидных отростков (нормальные чувствительные нервы), представлявших проблемы в постановке диагноза. Кусочек ткани объемом не более 0,5 см3 выделяли и замораживали в охлажденном изопентане при температуре жидкого азота. После 60 секунд образец извлекали и помещали в пробирку для замораживания, предварительно охлажденную до -70°С. С помощью криостата выполняли срезы замороженной ткани по 10 мкм. Срезы помещали на стеклянные пластинки Suoerfrost Gold+(VWR). Срезы сушили на воздухе в течение 3 мин и затем фиксировали в ацетоне в течение 10 мин и вновь сушили на воздухе. Затем срезы хранили при -20°С до проведения иммуногистохимического анализа. Иммуноокрашивание образцов ткани выполняли при помощи следующих первичных моноклональных антител мыши:
Первичные антитела, специфичные к GD2:
моноклональное антитело 10В8 (lgG3 каппа), специфичное к GD2
моноклональное антитело 8В6 (lgG3 каппа), специфичное к O-ацетилированному GD2
Первичное антитело, служившее в качестве отрицательного контроля -моноклональное антитело МСА2063 (lgG3 каппа), специфичное к DNP (Serotec France, Cergy Saint Christophe, France).
Связывание антител с образцами тестируемой ткани выявляли с помощью набора реагентов DakoCytomation Envision+System, набора Peroxydase HRP (DAKO, Glostrup, Denmark) для использования с первичными антителами мыши согласно инструкциям производителя. Затем образцы заключали между предметным и покровным стеклом с водной гистологической средой Aquadex (VWR). Наличие маркировки устанавливали при помощи световой микроскопии с использованием оптического микроскопа с увеличением 100 и 400. Анализировали цифровые микрофотографии трех полей зрения, выбранных случайным образом, на каждый образец.
Все образцы опухолевой ткани, взятые у разных пациентов, имели положительную маркировку мембраны и цитоплазмы с антителами 10В8 и 8В6, тогда как здоровые клетки не маркировались ни одним из этих антител. Маркировка не была обнаружена при использовании антитела МСА2063 против DNP.
При использовании антитела 10В8 все протестированные образцы нервов имели маркировку на уровне участков миелиновых волокон между соседними перехватами Ранвье, тогда как аксоны и фибробласты не имели маркировки. Такая маркировка не обнаруживалась или едва обнаруживалась при использовании антитела 8В6. При использовании антитела МСА2063 не обнаруживалось никакой маркировки.
15. Исследование цитотоксичности антител 8В6 и КМ8В6 к O-ацетилированному GD2
(а) Приготовление суспензии клеток-мишеней
1×106 клеток IMR32 нейробластомы человека, культивируемых в среде RPMI, инкубировали в присутствии 1,85 MBq Na251CrO4 в течение 1 часа при 37°С. Затем клетки отмывали 3 раза с помощью RPMI и центрифугирования, а затем ресуспендировали в RPMI и инкубировали при 4°С в течение 30 мин для измерения спонтанного высвобождения радиоактивного вещества.
После финального центрифугирования клетки растворяли в 5 мл RPMI и доводили концентрацию до 2×105 клеток/мл.
(б) Приготовление эффекторных клеток
Кровь доноров-добровольцев собирали в пробирки для взятия крови, содержащие гепарин. Лейкоциты периферической крови выделяли из цельной крови путем центрифугирования в градиенте Ficoll® (1800 д в течение 30 мин). Полученные клетки отмывали 3 раза в RPMI с центрифугированием при 1500 g и ресуспендировали в RPMI для получения концентрации клеток 5×106 клеток/мл.
(в) Измерение активности ADCC
В каждую лунку 96-луночного круглодонного планшета для микротитрования, приобретенного в компании Falcon, вносили 50 мкл суспензии клеток-мишеней, полученных, как описано в пункте (а). Затем вносили по 100 мкл суспензии эффекторных клеток, полученных, как описано в пункте (б), т.е. по 50000 клеток на лунку. Соотношение клеток-мишеней к количеству эффекторных клеток составляло 1/50. Затем в каждую лунку вносили антитело 8В6, искусственно модифицированное антитело КМ8В6 и rituxan®, химерное антитело мыши и человека типа lgG1 каппа (анти-CD20, использованное в качестве отрицательного контроля) в концентрации 1 мкг/мл или 10 мкг/мл. Смесь инкубировали в течение 4 ч при 37°С. После инкубации планшет центрифугировали и определяли присутствие в супернатанте51Cr при помощи γ-счетчика. Общее количество высвобожденного51Cr измеряли аналогичным способом, при добавлении среды без антитела и раствора 5 N гидроксида натрия вместо суспензии эффекторных клеток. ADCC рассчитывали по следующей формуле:
% активности ADCC=(51Cr в супернатанте -51Cr высвобожденный спонтанно) / (51Cr общий -51Cr высвобожденный спонтанно)
16. Опухолевая активность антитела 8В6 в сингенной модели лимфомы мышей
Противоопухолевую активность антитела 8В6 определяли на модели подкожной имплантации сингенной культуры клеток Т лимфомы мышей EL-4, экспрессирующих антиген GD2, мышам линии C57BL/6 (Zhang H, Zhang S, Cheung NK, Ragupathi G, Livingston PO. Antibodies against GD2 ganglioside can eradicate syngenic cancer micrometastases. Cancer Res. 1998, 58: 2844-9). Эти клетки EL4 экспрессируют также антиген O-ацетилированного GD2. 24 мышам, выращиваемым в животнике уровня А1, в возрасте 12 недель проводили подкожную инъекцию 20х104 клеток EL-4 в суспензии в PBS. Были сформированы 2 группы по 12 мышей. Мыши группы А получали 70 мкг антитела 8В6 в растворе в 200 мкл буферного раствора PBS путем внутривенного введения, каждые 3 дня начиная с первого дня после инъекции клеток EL-4 и до 21 дня. Мыши группы Б получали таким же образом 200 мкл раствора PBS. Каждые 2 дня замеряли объемы опухолей. Объем опухоли оценивали по следующей формуле: объем (мм3)=длина (мм) × ширина2 (мм) × 0,5 (Zeng G, Li DD, Gao L, Birkle S, Bieberich E, Tokuda A, Yu RK; Alteration of ganglioside composition by stable transfection with antisense vectors against GD3-synthase gene expression. Biochemistry 1999 38: 8762-9). Мышей, имеющих объем опухоли > 3,000 мм3, забивали.
Полученные результаты представлены на Фиг.16. Группа Б, получавшая внутривенно PBS, соответствует группе контроля, не получавшей лечения (Фиг.16, панель Б). Опухоли начинали обнаруживаться через 10 дней после инокуляции и затем характеризовались экспоненциальным ростом. Через 20 дней после инокуляции все мыши, за исключением двух, имели опухоли более 1000 мм3. Все мыши были забиты ранее 30 дней в соответствии с правилам работы с животными, так как их опухоли превосходили 3000 мм3. У мышей группы А, получавших антитело 8В6 (Фиг.16, панель А), наблюдалась задержка появления опухолей. Кроме того, рост опухолей у мышей, получавших это антитело, замедлялся. Через 20 дней после инокуляции ни у одной из мышей, получавших лечение, не обнаруживалось опухоли более 1000 мм3. По окончании 30 дней 58% мышей, получавших антитело 8В6, выжили, тогда как все мыши, не получавшие лечения, погибли. По окончании 40 дней были живы 25% мышей, получавших лечение, а на 50-й день 8% мышей были все еще живы, не имели пальпируемых опухолей и считались излеченными.
Реферат
Настоящее изобретение относится к области иммунологии. Предложено применение моноклональных антител, распознающих О-ацетилированную форму ганглиозида GD2 и не распознающих ганглиозид GD2, для лечения рака, характеризующегося экспрессией О-ацетилированного GD2, в частности, опухолей нейроэктодермального происхождения, а также применение указанного антитела, связанного с токсической молекулой, лекарством или вторым антителом, для лечения рака, характеризующегося экспрессией О-ацетилированного GD2. Использование изобретения может найти дальнейшее применение в лечении рака. 2 н. и 11 з.п. ф-лы, 16 пр., 16 ил.
Комментарии