Гибридное моноклональное антитело, взаимодействующее с cd4-антигеном т-хелперных клеток человека, и способ его получения - RU2112037C1
Код документа: RU2112037C1
Чертежи



























Описание
Изобретение относится к CDR-привитому гибридному моноклональному антителу и способу его получения при использовании технологии рекомбинантных ДНК.
Природные иммуноглобулины (Ig) включают две тяжелые и две легкие цепи, при этом N-терминальные участки каждой пары из тяжелой и легкой цепи соединены между собой и образуют антиген-связывающие сайты.
Обозначения остатков для легкой и тяжелой цепей Ig, приведенные в описании и пунктах формулы изобретения, находятся в соответствии со схемой нумерации, разработанной Кабатом (Kabat et al. in "Seguences of Proteins of Immunological Interest", US Department of Health and Human Services, 1987; Wu and Kabat, J. Exp. Med., 132, 211, 1970). Таким образом, обозначение остатков не всегда прямо соответствует линейной нумерации аминокислотных последовательностей. Действительная линейная аминокислотная последовательность может содержать меньшее число или дополнительные аминокислотные остатки, чем в соответствии с нумерацией Кабата. Таким образом, она содержит вставки или делеции остатков. Эти вставки или делеции могут обнаруживаться в любом участке в пределах цепей. Правильная нумерация остатков для данного Ig может быть определена путем сравнения областей гомологии линеаризованной последовательности Ig со "стандартной" последовательностью, в которой нумерация остатков соответствует системе Кабата. При изучении большого числа Ig было установлено, что вариабельные домены, которые локализованы на N-концевых участках легкой и тяжелой цепей содержат три области, в которых аминокислотные последовательности гипервариабельны. Эти гипервариабельные участки фланкированы с каждой стороны областями, которые изменяются в значительно меньшей степени, как описано Кабатом (см. выше).
Тогда было сделано предложение, что гипервариабельные области участвуют в связывании антигена. Позднее структурными исследованиями с использованием рентгеновской кристаллографии и молекулярного моделирования были определены три области в вариабельных доменах каждой тяжелой и легкой цепи, которые очевидным образом вовлечены в связывание антигена (Chotia and Lesk, J. Mol. Biol., 96, 4, 901). Эти три области обычно называются областями, определяющими комплементарность (CDRS). CDRS объединяются вместе посредством остальных участков вариабельных доменов с образованием, по крайней мере, части антигенсвязывающего центра. Указанные области обычно называются структурными областями. При этом следует отметить, что некоторые специалисты, в частности Кабат (см. выше), рассматривают гипервариабельные области как CDR. Для ясности в данном описании термин "гипервариабельная область" используется только для обозначения антигенсвязывающих участков, выявленных в результате анализа последовательностей, а термин CDR используется для обозначения антигенсвязывающих участков, выделенных в результате структурного анализа.
Сравнение гипервариабельных областей, определенных путем анализа последовательностей, и CDRS, определенных при помощи структурных исследований, показывает, что имеется некоторое, но не полное соответствие между этими областями.
В данном описании термин "гибридное антитело" используется для обозначения белка, включающего по крайней мере антигенсвязывающий участок Ig, соединенный с помощью пептидной связи по крайней мере с частью другого белка. При этом следует отметить, что некоторые специалисты могут также использовать понятие "химерный" для описания таких конструкций, но в описании такие антитела называют гибридными антителами.
Термин "CDR-"привитое" антитело" используется для описания антитела, имеющего по крайней мере один и предпочтительно два или три из CDRS в одном или обоих вариабельных доменах, полученных из антитела из первого вида. Другие участки антитела получают из одного или более других иммуноглобулинов. Вариабельные домены могут быть получены путем использования технологии рекомбинатных ДНК или путем пептидного синтеза.
Так как наиболее доступными моноклональными антителами (МКА) являются антитела, полученные при использовании гибридных клеток грызунов, то такие антитела при введении человеку функционируют как антигены и приводят к развитию нежелательных побочных реакций, поэтому применение МКА грызунов как терапевтических агентов для лечения людей по существу ограничено, поскольку человеческий организм может генерировать иммунный ответ на МКА грызунов, в результате которого данные МКА будут элиминироваться полностью, либо будет снижаться их эффективность. Таким образом, МКА грызунов вообще не рекомендуются для практического применения на людях за исключением одной или небольшого числа инъекций, чтобы избежать развития иммунологического ответа, вызванного их введением.
В связи с этим были предприняты попытки сконструировать антитела человеческого происхождения менее иммуногенными для людей. Такие методики могут быть названы методиками "гуманизации" и они обычно включают применение технологии рекомбинантных ДНК для манипуляций с ДНК-последовательностями, кодирующими полипептидные цепи молекул антитела.
В последние годы достижения молекулярной биологии основаны на получении широкого спектра гетерогенных полипептидов путем трансформации клеток хозяина гетерогенными ДНК-последовательностями, которые кодируют получение желаемых продуктов.
В заявке EP-A-O 088994 (Schering Corporation) предлагается способ конструирования рекомбинантных ДНК, включающих ds-ДНК-последовательность, которая кодирует вариабельный домен легкой или тяжелой цепи Ig, специфического для предварительно определенного лиганда. ds-ДНК-последовательность содержит кодоны инициирования и терминации на 5'- и 3'-концах соответственно, но не включает какие-либо нуклеотиды, кодирующие избыточные аминокислотные остатки в вариабельном домене. ds-ДНК-последовательность используется для трасформации бактериальных клеток.
В заявке не предлагается изменений последовательности вариабельного домена.
В заявке EP-A-O 102634 (Takeda Chemical Industries Limited) описывается клонирование и экспрессия в клетках бактерий-хозяев генов, кодирующих целиком или часть полипептида тяжелой цепи человеческого IgE, но не предполагается изменений в последовательности полипептида.
В заявке EP-A-O 125023 (Genentech Inc.) предлагается использование рекомбинантных ДНК и бактериальных клеток для получения Ig, которые аналогичны Ig позвоночных. Приведенные методики модификации генов используются для конструирования химерных антител или других модифицированных типов антител. Однако полагают, что предложения, описанные в указанной заявке Genentech Inc. , не приводят ни к экспрессии значительных количеств Ig-дополнительных цепей, ни к генерации Ig-активности, ни к секреции и сборке цепей с образованием химерных антител. Острая необходимость в разработке методик, обеспечивающих стабильное введение ДНК Ig генов клетки млекопитающих привела к появлению технологии мутагенеза in vivo и способов трансфекции ДНК для получения рекомбинантных антител, обладающих новыми свойствами.
Однако известно, что функциональная активность молекулы антитела зависит от его трехмерной структуры, которая в свою очередь, опосредована его первичной аминокислотной последовательностью. Таким образом, изменения аминокислотной последовательности антитела могут неблагоприятно воздействовать на его активность. Кроме того, изменения в последовательности ДНК, кодирующей антитело, может оказать влияние на способность клетки, содержащей последовательность ДНК, экспрессировать, секретировать или "собирать" антитело.
Таким образом, возможность получения функционально измененных антител с помощью методики рекомбинантных ДНК является неочевидной.
Однако авторы предлагаемого изобретения разработали способ, посредством которого могут быть получены гибридные антитела, обе части которых функциональны. Этот способ раскрывается в Международной заявке PCT/GB 85/00392. Однако в указанной выше заявке PCT раскрывается только получение гибридных антител, в которых полные вариабельные домены кодируются первым участком ДНК-последовательности и не показано получение гибридных антител с измененными последовательностями вариабельного домена.
В заявке EP-A-O 239400 описывается способ, в котором CDR мышиного МКА были "привиты" на структурные области вариабельных доменов человеческого Ig путем сайт-направленного мутагенеза с использованием длинных олигонуклеотидов. Авторы изобретения указывают также на возможность изменения природной аминокислотной последовательности структурных областей.
Наиболее ранняя работа по изменению МКА путем CDR-"прививки" была проведена на МКА, распознающих синтетические антигены, такие как NP- или NIP-антигены. Однако примеры, в которых мышиное МКА, узнающее лизоцим, и МКА крысы, распознающее антиген на человеческих T-клетках, соответственно были гуманизированы путем CDR-прививки, описаны Verhoeyen et al., Science, 239, 1534, 1988 и Reichmann et al., Nature, 332, 323, 1988.
Однако в работе Reichmann et al. показано, что перенос только CDRS не был достаточным для обеспечения удовлетворительной антигенсвязывающей активности в CDR-привитом продукте. В указанной работе подтверждается необходимость замены серинового остатка в положении 27 человеческой последовательности на соответствующий фенилаланиновый остаток крысы для получения CDR-привитого продукта, имеющего удовлетворительную антигенсвязывающую активность. Этот остаток в положении 27 тяжелой цепи находится в пределах структурной петли смежной с CDR1. Полученная далее конструкция, которая дополнительно содержала замену серина человека на тирозин крысы в положении 30 тяжелой цепи, не имела заметно измененной связывающей активности по сравнению с CDR-привитым антителом с заменой только серина на фенилаланин в положении 27. Эти результаты показывают, что для CDR-привитых антител, которые распознают более сложные антигены, изменения в остатках человеческой последовательности вне CDR-областей, в частности в петле, смежной с CDR1, могут быть необходимы для получения эффективной антигенсвязывающей активности.
Недавно также описаны методики изменения анти-ТАС моноклонального антитела путем CDR-прививки. Человеческие каркасные области были выбраны для максимального увеличения гомологии с последовательностью анти-ТАС антитела, тогда как несколько дополнительных аминокислот вне CDR были сохранены. Анти-ТАС антитело, измененное таким образом, имеет аффинность в отношении p55 цепи человеческого интерлейкина 2, составляющей примерно одну треть таковой анти-ТАС антитела мыши (Queen et al., PNAS-USA, 86, 10029, 1989).
В заявке PCT/US 89/06857 также описываются CDR-привитые антитела, которые являются специфическими в отношении p55 ТАС протеина IL-2-рецептора. В ней показано, что для CDR-привитого антитела может оказаться необходимым, чтобы 3 или более аминокислотных остатков из данного Ig в дополнение к CDRS, обычно по крайней мере один из которых непосредственно примыкает к CDR в донорном Ig, был изменен в соответствии с остатком донорного антитела для того, чтобы получить антигенсвязывающую активность.
Таким образом, совершенно очевидно, что получение CDR-привитого антитела - довольно сложная задача. Часто недостаточно просто привить CDR донорного Ig на каркасные области акцепторного Ig. При этом может оказаться необходимым изменение остатков в каркасных областях акцепторного антитела для того, чтобы обеспечить антигенсвязывающую активность. Однако невозвожно предсказать на основании известных и доступных исследований, какие остатки в каркасных областях, необходимо изменить (если это нужно вообще делать).
В заявке EP-A-O 018794 описывается МКА мыши, которое распознает антиген хелперных T-клеток человека. Данное антитело обозначается OKT4. Антиген, распознаваемый указанным антителом, обозначается как CD4. МКА коммерчески выпускается фирмой "Ortho Diagnostic Systems", Inc. of Raritan, New Jersey, USA. Кроме того, указанная фирма производит МКА мыши, известное как OKT4A; оно распознает другой эпитоп CD4 антигена, отличный от эпитопа, узнаваемого OKT4.
Эксперименты по трансплантации у приматов показали, что оба антитела ОКТ4 и ОКТ4А могут применяться для предотвращения отторжения трансплантантов и могут оказаться полезными для использования в качестве модуляторов человека. Опыт лечения пациентов с пересаженной почкой мышиными МКА ОКТ3 показал, что иногда у больных образуются антитела, нейтрализующие ОКТ3. Этот иммунный ответ мешает повторному введению МКА. Чтобы уменьшить предполагаемый иммунный ответ к анти-CD4 МКА, было бы желательно получить CDR-привитое ОКТ4А, имеющие CDRS мыши и каркасные участки, а также другие области Ig человека.
Однако, как описано выше, простой подход к конструированию CDR-привитого антитела не всегда приводит к получению антитела, которое эффективно связывает заданный антиген. Конкретные аминокислотные остатки, которые входят в состав CDRS , трудно идентифицировать, и они не обязательно соответствуют всем остаткам в гипервариабельных областях. При этом могут существовать также критические каркасные остатки, которые важны для расположения CDRS при взаимодействии с антигеном или которые вовлекаются во взаимодействие между тяжелой и легкой цепями, а также может возникать необходимость изменения определенных остатков в каркасных участках так, чтобы они соответствовали остаткам мыши в этих положениях.
Несмотря на проблемы, которые возникают при получении специфического CDR-привитого антитела, в предпочтительном варианте осуществления изобретения конструируют CDR-привитое антитело, содержащее каркасные области lg человека и распознающее CD4-антиген. В особо предпочтительном варианте осуществления изобретения CDR-привитое антитело обладает аффинностью к CD4-антигену, аналогичной таковой OTK4A MKA мыши. Такие антитела из уровня техники не известны.
Согласно изобретению сконструировано CDR-привитое антитело, имеющее по крайней мере одну цепь, в которой каркасные области преимущественно получены из первого антитела (акцептор) и по крайней мере одна CDR получена из второго антитела (донор), которое способно к связыванию CD4-антигена.
Предпочтительно CDR-привитая цепь имеет две и более предпочтительно все три CDRS, полученные из донорного антитела. Предпочтительно, чтобы в CDR-привитой цепи каждая CDR представляла собой сложную CDR, включающую все остатки CDR и все остатки соответствующей гипервариабельной области донорного антитела.
Предпочтительно также, чтобы по крайней мере один остаток в каркасных областях CDR-привитой цепи был изменен так, чтобы он соответствовал эквивалентному остатку в антителе.
Кроме того, предпочтительно, чтобы каркасные области CDR-привитой цепи были получены из антитела человека.
Предпочтительно, чтобы каркасные области CDR-привитого антитела были получены из тяжелой цепи lg человека. Для таких тяжелых цепей предпочтительно, чтобы остаток 35 в каркасных областях тяжелой цепи изменялся таким образом, чтобы соответствовать эквивалентному остатку в донорном антителе.
Предпочтительно, чтобы в таких тяжелых цепях по крайней мере одна сложная CDR, включающая остатки 26-35, 50-65 или 95-102, соответственно, прививалась на каркасную область человека. В этом случае остаток 35 будет уже соответствовать эквивалентному остатку в донорном антителе.
Предпочтительно, чтобы остатки 23, 24 и 49 в таких тяжелых цепях соответствовали эквивалентным остаткам антитела. Более предпочтительно, чтобы остатки 6, 23, 24, 48 и 49 в таких тяжелых цепях соответствовали остаткам донорного антитела в эквивалентных положениях. При необходимости остатки 71, 73 и 79 могут соответствовать остаткам донорного антитела в эквивалентных положениях.
Чтобы еще более усилить аффинность, любой один или любая комбинация остатков 57, 58, 60, 88 и 91 могут соответствовать остаткам донорного антитела в эквивалентных положениях.
Тяжелая цепь предпочтительно происходит из KOL тяжелой цепи lg человека, однако она может быть также получена из NEWM или EU тяжелой цепи lg человека.
Каркасные области CDR-привитой цепи могут быть получены из каппа или лямбда легкой цепи lg человека. Для такой легкой цепи целесообразно, чтобы по крайней мере одна сложная CDR, содержащая остатки 24-34, 50-65 или 89-97, соответственно, прививалась на каркасную область антитела человека. Предпочтительно, чтобы остаток 49 также соответствовал эквивалентному остатку в донорном антителе.
Для дальнейшей оптимизации аффинности предпочтительно, чтобы остатки 49 и 89 соответствовали эквивалентным остаткам в донорном антителе. При необходимости можно отобрать эквивалентные донорные остатки, которые образуют солевые мостики.
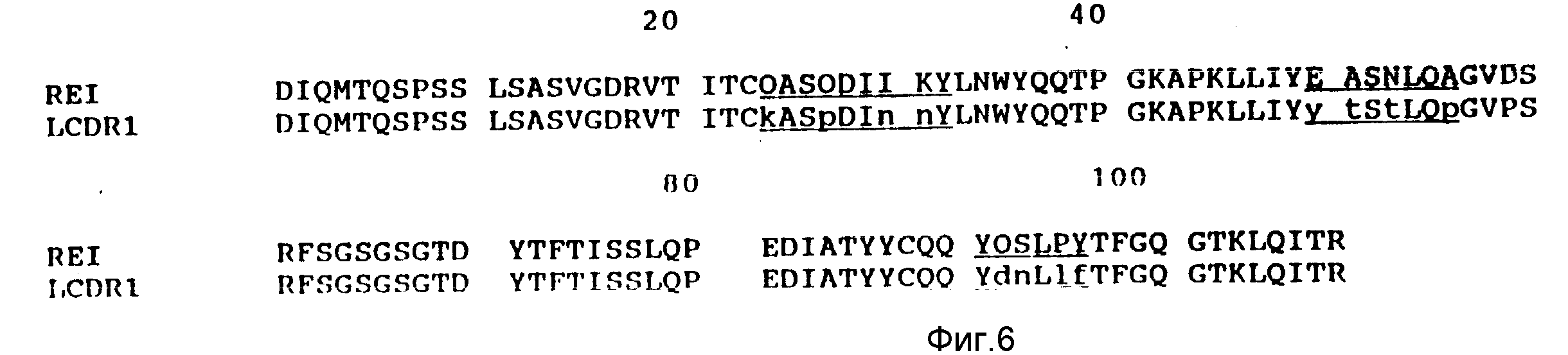
Легкая цепь предпочтительно происходит из REI легкой цепи lg человека, однако она может быть также получена из EU легкой цепи lg человека.
CDR-привитое антитело согласно изобретению предпочтительно содержит легкую цепь и тяжелую цепь, одна или предпочтительно обе из которых включают CDRS, привитые в соответствии с принципами, изложенными выше по отдельности для легких и тяжелых цепей.
В предпочтительном случае целесообразно, чтобы все три CDRS тяжелой цепи были изменены и чтобы минимальные изменения делались в легкой цепи. Возможно, что отсутствие изменений CDRS либо изменения одной или двух CDRS легкой цепи все еще сохраняет связывающую аффинность на приемлемом уровне.
При этом следует отметить, что в некоторых случаях для обеих тяжелой и легкой цепей донорные и акцепторные остатки могут быть идентичны в определенном положении и, таким образом, не потребуется изменений акцепторного остатка каркасной области.
Следует отметить также, что для сохранения, насколько это возможно, человеческой природы CDR-привитого антитела, необходимо осуществлять как можно меньше замен аминокислотных остатков. Предусматривается, что во многих случаях не будет необходимости в больших изменениях, чем затрагивающих CDRS и незначительное число остатков в каркасных областях. Только в исключительных случаях необходимо изменять существенное число остатков в каркасных областях.
Предлагаемое CDR-привитое антитело относится к изотипу IgG, например изотипу IgG1 или IgG4.
При необходимости один или более остатков в константных доменах lg могут быть изменены для того, чтобы изменить эффекторные функции константных доменов.
Предпочтительно CDR-привитое антитело имеет аффинность к антигену, составляющую примерно 105 - 1012 М-1, более предпочтительно по крайней мере 108 М-1 и наиболее предпочтительно аффинность аналогична таковой MKA OKT4 или OKT4A.
Целесообразно, чтобы каждая CDR была получена из антитела млекопитающего, предпочтительно из MKA мыши.
Предлагаемое CDR-привитое антитело получают путем использования технологии рекомбинантных ДНК. Соответствующий способ включает следующие стадии: получение первой последовательности ДНК, кодирующей первую цепь антитела, в которой каркасные области преимущественно получены из первого антитела (акцептора) и по крайней мере одна CDR получена из второго антитела (донор), под контролем подходящих элементов, трансформацию клетки хозяина первой ДНК-последовательностью и культивирование трансформированной клетки хозяина, так что CDR-привитое (гибридное) антитело продуцируется в культуральную среду.
Кроме того, способ включает следующие стадии: получение второй ДНК-последовательности, кодирующей вторую цепь антитела, комплементарную первой цепи, под контролем подходящих элементов и трансформацию клетки хозяина обоеми первой и второй ДНК-последовательностями.
Целесообразно, чтобы вторая ДНК-последовательность кодировала вторую цепь антитела, в которой каркасные области преимущественно получены из первого антитела (акцептор) и по крайней мере одна CDR получена из второго антитела (донор).
Первая и вторая ДНК-последовательности могут содержаться в одном и том же векторе. В этом случае последовательности могут быть под контролем одних и тех же или различных элементов.
Альтернативно первая и вторая ДНК-последовательности могут входить в состав различных векторов.
При этом предусматривается, что гибридные антитела, полученные согласно изобретению, будут использоваться в терапии, особенно для предупреждения отторжения трансплантатов или лечения нарушений, связанных с T-хелперными клетками.
При получении антитела для трансформации соответствующим вектором используют эукариотические клетки, преимущественно клетки CHO.
Для конструирования гибридного моноклонального антитела, во-первых, необходимо установить последовательность вариабельного домена, имеющего желаемые связывающие свойства. В качестве вектора используют любой вектор (плазмиду), содержащий конструктивные элементы, необходимые для экспрессии в данных клетках-хозяевах. Гипервариабельные области могут затем быть определены с помощью метода Кабата (Wu and Kabat, J. Exp. Med., 132, 211, 1970). CDRS могут быть определены методом структурного анализа с использованием рентгеновской кристаллографии или методик молекулярного моделирования. Комплексная CDR может быть затем определена, как содержащая все остатки в одной CDR и все остатки в соответствующей гипервариабельной области. Эти комплексные CDRS наряду с некоторыми выбранными остатками каркасной области преимущественно переносятся как "антигенсвязывающие сайты", тогда как оставшаяся часть антитела, например контактные домены тяжелой и легкой цепи и другие каркасные области, могут принадлежать антителу человека.
В конструировании предлагаемых гибридных антител VH и/или VL сегменты гена изменяют путем мутагенеза.
Типичные методики включают добавление, делецию или неконсервативное замещение органического числа различных нуклеотидов или консервативное замещение многих нуклеотидов при условии, что сохраняется открытая рамка считывания.
Полученные согласно изобретению антитела могут быть очищены в соответствии со стандартными процедурами, включая осаждение сульфатом аммония, аффинную хроматографию и гельэлектрофорез.
Очищенные гибридные моноклональные антитела по крайней мере до 90-95% гомогенности являются предпочтительными, а очищенные до 98-99% или более гомогенности являются наиболее предпочтительными для фармацевтического использования.
В общем полученные антитела могут быть использованы в очищенном виде с фармакологически приемлемыми носителями. Обычно эти носители включают водные или водно-спиртовые растворы, эмульсии или суспензии, включая физиологический раствор и буферные среды.
На фиг. 1 показана нуклеотидная последовательность вариабельного домена тяжелой цепи OKT4A; на фиг. 2 - нуклеотидная последовательность вариабельного домена легкой цепи OKT4A; на фиг. 3 - аминокислотная последовательность вариабельного домена тяжелой цепи OKT4A, в которой CDRS подчеркнуты; на фиг. 4 - аминокислотная последовательность вариабельного домена легкой цепи OKT4A, в которой CDRS подчеркнуты; на фиг. 5 сравнивается последовательность KOL-цепи OKT4A с аминокислотной последовательностью тяжелой цепи CDR-привитого (гибридного) OKT4A, CDRS подчеркнуты, человеческие последовательности обозначены заглавными буквами и последовательности антитела мыши - строчными буквами; на фиг. 6 - последовательность REI-цепи OKT4A с аминокислотной последовательностью легкой цепи CDR-привитого (гибридного) OKT4A, CDRS подчеркнуты, человеческие последовательности обозначены заглавными буквами, последовательности антитела мыши - строчными буквами; на фиг. 7-13 показана ДНК-последовательность и транслированная аминокислотная последовательность тяжелой цепи CDR-привитого антитела; на фиг. 14-16 - ДНК-последовательность и транслированная аминокислотная последовательность легкой цепи CDR-привитого антитела; на фиг. 17 и 18 приведена схема конструирования вектора экспрессии тяжелой цепи CDR-привитого OKT4A; на фиг. 19 - результаты связывания и блокирования конструкций легкой цепи CDR-привитого OKT4A в комбинации с тяжелой цепью химерного OKT4A; на фиг. 20 - результаты анализа связывания и блокирования конструкций тяжелой цепи OKT4A, HCDR1, HCDR2 и HCDR3 в комбинации с OKT4A легкой цепью; на фиг. 21 сравнивается последовательность REI с легкими цепями CDR-привитого OKT4A, LCDR1 и LCDR2, и аминокислотной последовательностью легкой цепи OKT4A мыши, CDRS подчеркнуты, человеческие последовательности обозначены заглавными буквами, мышиные последовательности - строчными буквами; на фиг. 22 и 23 - последовательность KOL с тяжелыми цепями CDR-привитого OKT4A (HCDR1 включительно HCDR10) и аминокислотной последовательностью тяжелой цепи OKT4A мыши, CDRS подчеркнуты, человеческие последовательности обозначены заглавными буквами, и мышиные последовательности - строчными буквами; на фиг. 24 приведены результаты связывания и блокирования конструкций CDR-привитой тяжелой цепи, HCDR1, HCDR2 и HCDR3 в комбинации с CDR-привитой легкой цепью LCDR2; на фиг. 25 - результаты связывания и блокирования конструкций CD2-привитой тяжелой цепи (HCDR4 включительно HCDR10) в комбинации с легкой цепью LCDR2; на фиг. 26 и 27 - результаты блокирования конструкций тяжелой цепи OKT4A, HCDR5, HCDR6, и HCDR10 в комбинации с конструкциями легкой цепи LCDR2, LCDR3, LCDR2Q, LCDR3Q, LCDR4Q и химерной формой OKT4A; на фиг. 28 - результаты относительной аффинности конструкций OKT4A тяжелой цепи HCDR5 и HCDR10 в комбинации с конструкцией легкой цепи LCDR2 и химерной и интактной формой OKT4A, в качестве отрицательных контролей используют химерное и интактное OKT3 мыши; на фиг. 29 - результаты ингибирования MLR различными антителами с использованием T6 в качестве негативного контроля; на фиг. 30 приведены результаты исследований по ингибированию пролиферации различными антителами.
Изобретение иллюстрируется следующим примером.
"Гуманизация" OKT4A.
OKT4A представляет собой моноклональное антитело мыши, которое узнает CD4 антиген, локализованный в основном на хелперных T-лимфоцитах. При этом были получены CDR-привитые антитела (гибридные антитела), в которых CDRS вариабельных доменов обеих тяжелой и легкой цепей были получены из последовательности антитела OKT4A мыши. Каркасные области вариабельных доменов и константные домены были получены из последовательностей антитела человека.
Три CDRS, которые локализованы на обеих тяжелой и легкой цепях, состоят из тех остатков, которые, как показано структурными исследованиями, вовлечены в связывание антигена. Теоретически, если CDRS из OKT4A-антитела мыши прививают на каркасные области антитела человека с образованием CDR-привитого вариабельного домена и этот вариабельный домен присоединяют к константным доменам антитела человека, получаемое в результате CDR-привитое (гибридное) антитело является по существу человеческим антителом со специфичностью OKT4A антитела мыши и способно связывать CD4-антиген на клетках человека. Такое антитело обладает меньшей иммуногенностью при введении человеку по сравнению с OKT4A-антителом мыши.
В результате тестирования связывания антигена CDR-привитым OKT4A-антителом, которое содержит CDRS, соединенные с каркасными участками антитела человека, было показано, что полученное антитело не обладает достаточной аффинностью к CD4 антигену. В связи с этим дополнительные остатки, смежные с некоторыми из CDRS и, критические остатки каркасных областей должны быть замещены в человеческом антителе на соответствующие остатки OKT4A антитела мыши для того, чтобы получить функционально активное антитело.
Выделение кДНК тяжелой и легкой цепи и анализ ДНК-последовательности вариабельного домена.
Для получения CDR-привитого (гибридного моноклонального) OKT4A антитела определяют последовательность вариабельного домена тяжелых и легких цепей OKT4A мыши, а затем последовательность кДНК тяжелой и легкой цепей, которую синтезируют при соответствующей мРНК.
мРНК получают из OKT4A-продуцирующих гибридомных клеток путем гуанидин-тиоцианатной экстракции с последующей очисткой хлоридом цезия.
кДНК синтезируют на основе мРНК. Добавляют EcoR1-линкеры и лигируют в EcoR1-сайт lgt10 клонирующего вектора. Рекомбинантный фаг упаковывают в инфекционные частицы, которые используют для инфицирования E. Coli C600.
Эту библиотеку скринируют для последовательностей тяжелой цепи OKT4A с использованием
олигонуклеотидных проб Cg и FR3. Cg-праймер имеет последовательность
5'GGCCAGTGGATAGAC 3'
и связывается с константным доменом Ig мыши. Проба FR3 имеет последовательность
5'GGCCGTGTCCTCAGACCT 3'
и связывается с третьим каркасным участком вариабельного домена тяжелых цепей Ig мыши. Пять положительных клонов оценивают Саузерн-блоттингом и гибридизацией проб Cg,
FR3 и кДНК IgG2a CH3 мыши. Отбирают единичный клон с 1600 bp EcoRI-вставкой, который гибридизируется со всеми тремя пробами.
Библиотеку скринируют для последовательностей легкой цепи
OKT4A с использованием олигонуклеотидной пробы Ck последовательностью
5' GGCTCCAGGTTGCTGATGCTGAAGG 3',
которая связывается с константным доменом каппа-цепи Ig мыши. Шесть
положительных клонов далее оценивают Саузерн-блоттингом и гибридизацией с олигонуклеотидными пробами T4AK последовательности
5' GGCTCCAGGTTGCTGATGCTGAAGG 3',
которые связываются с
каркасным участком 3 каппа-цепи Ig мыши.
При этом отбирают единичный клон, который содержит 900 bp EcoRl-вставку и гибридизируется с двумя пробами.
кДНК тяжелой цепи размером 1600 bp субклонируют в EcoRl-сайты "pBluescript" плазмидный вектор и вектор секвенирования M13mp8. кДНК легкой цепи размером 900 bp субклонирована в EcoRl-сайты вектора pUC8 и вектора секвенирования M13mp19.
Для определения последовательности одноцепочечной (M13) и двуцепочечной (плазмида) матриц используют метод анализа последовательности ДНК путем дидеоксинуклеотидной терминации цепи. Последовательность 5'-нетраслируемых областей, сигнальных последовательностей, вариабельных доменов и части константных доменов определяют для тяжелой и легкой цепи кДНК. Последовательность ДНК для тяжелой и легкой цепей приведены на фиг. 1 и 2. Аминокислотная последовательность вариабельного домена тяжелой цепи представлена на фиг. 3. Последовательность вариабельного домена легкой цепи представлена на фиг. 4.
При этом следует отметить, что нуклеотидная последовательность легкой цепи имеет A остаток в положении 163 в направлении к началу последовательности, кодирующей CDR1 (см фиг. 2). Трансляция этой последовательности приводит к включению остатка в положении 27 в легкой цепи (фиг. 4). Когда первоначально проводилось определение последовательности, предполагали, что нуклеотидный остаток 163 является C-остатком, обеспечивая наличие пролинового остатка в положении 27 в легкой цепи. Первые CDR-привитые антитела были сконструированы авторами изобретения, исходя из предположения, что остаток 27 в легкой цепи является пролиновым остатком (это может быть видно из фиг. 6, 8 и 12).
Конструирование гибридного моноклонального OKT4A антитела.
Чтобы сконструировать CDR-привитое (гибридное моноклональное) OKT4A антитело, необходимо определить, какие последовательности OKT4A мыши включают CDRS легкой и тяжелой цепей. Рентгеновское исследование кристаллических структур антитела показывают, что антигенсвязывающая поверхность локализуется на трех петлях, охватывающих β-складчатую структуру вариабельного домена. Таким образом, эти петли могут использоваться для определения CDR. Поскольку кристаллическая структура OKT4A мыши не доступна, структура аналогичного антитела мыши известной кристаллической структуры использовалась для определения остатков петель.
На легкой и тяжелой цепях обнаруживаются три гипервариабельных области между менее вариабельными каркасными областями.
В большинстве случаев эти гипервариабельные области соответствуют CDRS, но могут простираться за их пределы. Авторами изобретения было решено, что комбинации остатков CDRS в OKT4A мыши и остатков гипервариабельных областей представляет собой сложные CDRS, которые прививаются на каркасные области человеческого антитела. Аминокислотные последовательности тяжелых и легких цепей OKT4A мыши представлены на фиг. 3 и 4, где подчеркнуты сложные CDRS. Последовательность каркасной области тяжелой цепи человеческого антитела представляет собой последовательность человеческого антитела KOL. Антитело KOL выбирают потому, что его кристаллическая структура определена методом рентгеновской кристаллографии с высокой степенью разрешения. Это позволяет проводить точное молекулярное моделирование антитела. По этой же причине последовательность каркасной области димера легкой цепи антитела человека REI используют конструирование каркасных областей легкой цепи. Аминокислотные последовательности KOL и REI приведены на фиг. 5 и 6 в сравнении с аминокислотными последовательностями вариабельных доменов CDR-привитой тяжелой (HCDR1) и легкой (LCDR1) цепи OKT4A.
CDR-привитую тяжелую цепь конструируют таким образом, чтобы получить константный участок IgG4 человека. IgG4 подкласс выбирают на основании эксперимента с анти-CB3 моноклональным антителом OKT3 мыши, которое используется для предотвращения отторжения почечного трансплантата. OKT3 мыши имеет IgG2a изотип и не фиксирует комплемент человека. Антитело человека IgG4 также не фиксирует комплемент. CDR-привитую OKT4A легкую цепь конструируют таким образом, что она содержит константный домен каппа-цепи Ig человека.
Конструирование генов гибридного моноклонального антитела OKT4A.
CDR-привитые вариабельные домены тяжелой и легкой цепи сконструированы путем соединения синтетических двухцепочечных ДНК-олигомеров аналогично методу, используемому Jones et al. (Nanure, 321, 522. 1986). 5'-конец вариабельных доменов содержит сигнальные последовательности легкой и тяжелой цепей моноклонального антитела мыши B 72.3. Сигнальная последовательность направляет секрецию антитела из клеток млекопитающих. Последовательность Козака непосредственно предшествует AUG стартовому кодону для усиления трансляции. Вариабельные домены лигируют с ДНК, кодирующей константные домены антитела человека для получения гибридных (CDR-привитых) генов тяжелой и легкой цепей.
Конструирование тяжелой цепи CDR-привитого (гибридного) OKT4A.
Восемь комплементарных пар олигомеров с перекрывающимися концами длиной около 30 bp используют для связывания вариабельного домена из XhoI-сайта, локализованного в каркасной области 2, с HindIII-сайтом в начале первого константного домена. Эти восемь олигомерных пар синтезируют, сливают вместе ступенчатым способом и затем лигируют с HindIII 5'-концом ДНК константного домена человеческого IgG4. ДНК IgG4 получают от фирмы Celltech как геномную ДНК. Эта ДНК представляет собой вставку размером 2153 bp в фаговом ДНК-векторе M13 с 5' EcoRI- и 3' BamHI-рестрикционными сайтами. CH1-шарнирный, CH2- и СH3-домены окружают четырьмя интронами. Ген модифицируют таким образом, чтобы заместить C на A в предпоследнем основании CH1-экзона для создания нового HindIII-сайта с целью конструирования CDR-привитого (гибридного) гена.
5'-конец вариабельного домена конструируют путем сшивания двух комплементарных пар синтетических олигомеров, каждый из которых имеет приблизительно 90 bp в длину. Этот фрагмент, который имеет 5' EcoRI-конец и 3' XhoI-конец лигируют с XhoI-концом фрагмента, описанного выше, с получением полного гена CDR-привитой тяжелой цепи. Указанный ген содержит 2364 bp и имеет 5' EcoRI-конец и 3' BamHI-конец. Последовательность ДНК и транслированная аминокислотная последовательность приведены на фиг. 7.
Интересующие исследователей области с соответствующими номерами нуклеотидов:
1-14 - EcoRI сайт и последовательность Козака
15-71 - сигнальная последовательность
72-146 - каркасная область 1
147-176 - CDR1
177-218 - каркасная область 2
219-254 - CDR2
255-362 - каркасная область
3
363-392 - CDR3
393-431 - каркасная область 4
432-727 - CH1 домен
728-1117 - интрон
1118-1153 - шарнирный домен
1154-1271 - интрон
1272-1599 - CH2 домен
1600-1698 - интрон
1699-2016 - CH3 домен
2017-2366 - 3' нетранслируемая область
Конструирование гена CDR-привитой (гибридной) легкой цепи.
Двенадцать комплементарных пар синтетических олигомеров с перекрывающимися концами лигируют одновременно с получением вариабельного домена CDR-привитой (гибридной) легкой цепи. Этот фрагмент имеет 5' EcoRI-конец и 3' NarI-конец. Его лигируют с 5' NarI концом ДНК константного домена каппа-цепи Ig человека. кДНК константной области каппа-цепи Ig человека модифицируют для включения NarI-рестрикционного сайта в третий и четвертый кодоны. Полученный в результате ген CDR-привитой (гибридной) легкой цепи имеет 754 bp в длину и EcoRI-концы. ДНК-последовательность с транслированной аминокислотной последовательностью приведена на фиг. 8.
Интересующие исследователей области с соответствующими номерами нуклеотидов:
1-8 - EcoRI сайт и последовательность
Козака
9-68 - сигнальная последовательность
69-143 - каркасная область 1
144-164 - CDR1
165-215 - каркасная область 2
216-236 - CDR2
237-338
- каркасная область 3
339-356 - CDR3
357-404 - каркасная область 4
405-710 - константный домен каппа-цепи
711-754 - 3' нетранслируемая последовательность
Экспрессия CDR-гибридного антитела OKT4A.
Конструирование вектора экспрессии тяжелой цепи.
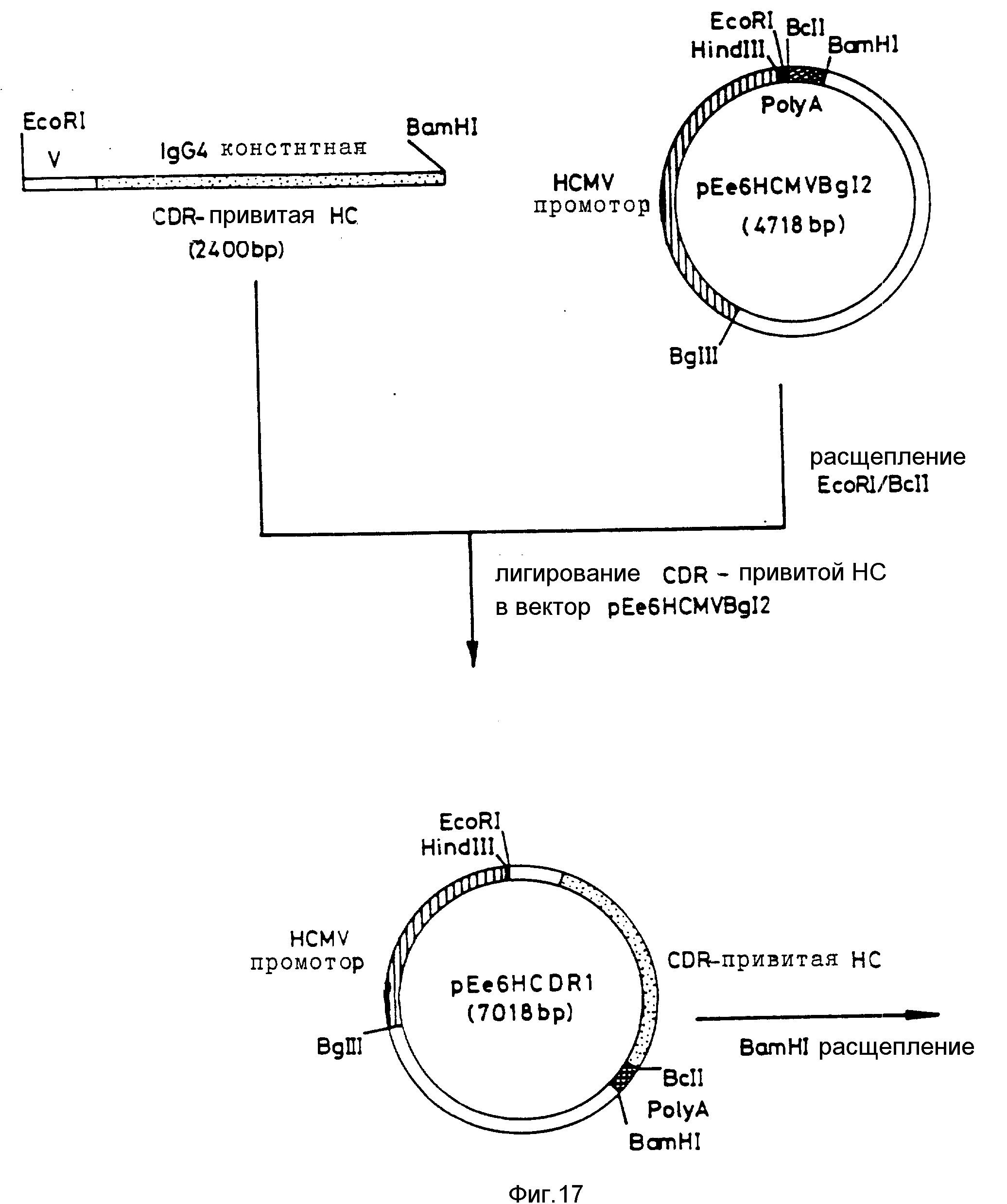
Вектор экспрессии тяжелой цепи гибридного антитела конструируют путем встраивания гена тяжелой цепи в плазмиду экспрессии pEe6HCMVBg12 и добавления GS-фрагмента, который включает ориджин репликации SV40 и миниген глутаминсинтетазы. Указанные стадии представлены в виде диаграммы на фиг. 9. pEe6HCMVBg12 и GS-фрагмент получают от фирмы "Celltech".
pEe6HCMV расщепляют по EcoRI и BclI-сайтам. ДНК pEe6HCMVBg12 деметилируют путем включения ее E. Coli; штамм DAM GM242, который утратил деоксиаденозинметилазу. BclI будет разрезать только ДНК, которая не содержит N6-метилированного деоксиаденозина в сайте, узнаваемом ферментом. Выступающий конец, полученный при BclI рестрикции, совместим с выступающим концом BamHI. EcoRI/BamHI ген CDR-привитой (гибридной) тяжелой цепи (HCDR1) был затем сшит с EcoRI/BclI концами pEe6HCMVBg12 для получения pEe6HCDR1.
BamHI-фрагмент размером 5500 bp, содержащий миниген глутаминсинтетазы, ориджин репликации SV и ранние и поздние промоторы встраивают в BamHI-сайт pEe6HCDR1 с получением pEe6HCDR1gs. Правильную ориентацию GS фрагмента подтверждают путем рестрикционного анализа. pEe6HCDR1gs получают для трансфекции клеток млекопитающих путем щелочного лизиса очисткой в градиенте хлорида цезия.
pEe6HCDR1gs способа экспрессировать тяжелую цепь гибридного OKT4A в COS и CHO клетках. HCMV промотор расположен в направлении 5' по отношению к концу тяжелой цепи и обеспечивает его транскрипцию. Сигнальная последовательность полиаденилирования SV40, расположенная в направлении 3' к указанному гену, действует как терминатор транскрипции. Для транзиторной экспрессии в COS-клетках, ориджин репликации SV40 находится в GS фрагменте размером 5500 bp. GS-миниген является селективным маркером при трансфекции CHO клеток. Экспрессия минигена глутаминсинтетазы обеспечивается поздним промотором SV40. GS-фрагшмент встраивают таким образом, что поздний проматор SV40 обеспечивает транскрипцию в том же направлении, что и HCMV промотор.
При образовании тяжелой цепи гибридного (CDR-привитого) антитела происходят определенные пост-трансляционные модификации. В пределах ядра три последовательности константной области IgG4 удаляются и экзоны соединяются вместе с образованием зрелой мРНК. После трансляции, 19 аминокислотных остатков сигнальной последовательности удаляются в шероховатом одноплазмическом ретикулуме (ER). Один углеводный остаток добавляют к CH2 домену каждой цепи в ER и аппарате Гольджи. Каждая цепь содержит также четыре внутрицепочечных дисульфидных связи. Когда пептид легкой цепи образуется при использовании вектора экспрессии для совместной трансфекции, зрелое антитело собирается путем связывания вместе, через дисульфидные связи двух тяжелых и двух легких цепей.
Конструирование вектора экспрессии цепи CDR-привитого (гибридного) OKT4A.
Вектор экспрессии легкой цепи CDR-привитого (гибридного) OKT4A был сконструирован путем встраивания гена CDR-привитой (гибридной) легкой цепи в вектор экспрессии pEe6HCMVBg12 с последующим введением ориджина репликации SV40 и минигена глутаминсинтетазы, содержащего GS-фрагмент. Вектор экспрессии легкой цепи конструируют, в основном, тем же самым способом, который был использован для получения вектора экспрессии тяжелой цепи, как показано на фиг. 9. Ген легкой цепи встраивают в EcoRI-сайт pEe6HCMVBg12 с получением pEe6LCDRI. Правильную ориентацию гена легкой цепи подтверждают путем рестрикционного анализа. Фрагмент GS размером 5500 bp встраивают в BamHI-сайт с получением pEe6LCDR1gs. Правильную ориентацию GS-фрагмента подтверждают путем рестрикционного анализа. pEe6LCDR1gs используют для трансфекции клеток млекопитающих; данную плазмиду получают методом щелочного лизиса очисткой в градиенте хлорида цезия.
Транскрипция гена легкой цепи в pEe6LCDR1gs обеспечивается HCMV промотором, а терминация сигнальной последовательностью полиаденилирования SV40. Ориджин репликации SV40, содержащийся в GS-фрагменте, обеспечивает автономную репликацию указанной конструкции в COS клетках. Миниген глутаминсинтетазы в GS-фрагмене обеспечивает селекцию и амплификацию в CHO-клетках.
После трансляции лидерная последовательность удаляется в шероховатом ER. Образуются две внутрицепочечные дисульфидные связи. Сборка зрелого антитела происходит, как описано выше.
Транзиторная экспрессия гибридного ОКТ4A в COS-1 клетках.
Транзиторная экспрессия гибридных генов в COS-1 клетках представляет собой быструю и удобную систему для тестирования экспрессии и функциональной способности CDR-привитого антитела ОКТ4A. COS-1 клетки конструктивно экспрессируют большой Т-антиген SV40, который поддерживает транзиторную репликацию эписом, несущих ориджин репликации SV40. Векторы экспрессии гибридных генов pEe6HCDR1gs и pEe6LCDR1gs содержат ориджин репликации SV40 как часть GS-фрагмента. После трансфекции в COS-1 клетки, векторы экспрессии реплицируются в ядро с высоким числом копий, что в результате приводит к относительно высоким уровням экспрессии гибридных генов.
Векторы экспрессии гибридных генов трансфицируют в COS клетки с использованием DEAE-декстрана после шока, который обеспечивается DMSO. Коротко говоря, 0,2 мл 1 мг/мл DEAE-декстрана добавляют в 15 мг векторной ДНК в 0,8 мл DMEM/Tris. Полученную смесь добавляют к 1 - 1,5 • 106 клеток в 60-мл чашке для культуры ткани и инкубируют приблизительно в течение 6 ч. DEAE-декстран/ДНК комплекс удаляют и 10% DMSO в буфере добавляют в чашку в течение 2 мин. Затем раствор удаляют, клетки промывают однократно DMEM и затем инкубируют с DMEM, содержащим 10% эмбриональной сыворотки теленка в течение 3 - 4 дней. В этот момент времени супернатант из лунок собирают и исследуют на уровни антитела и способность связывать CD4 положительные лимфоциты.
Уровни антитела определяют с помощью ELISA. Лунки покрывают козьим антителом, специфичным к Fc-участку антител человека. Различные разведения COS-клеточных супернатантов, содержащих секретируемое антитело, инкубируют в течение одного часа при комнатной температуре во влажной камере и отмывают. Затем добавляют пероксидазу хрена, связанную с антителом козы к каппа-цепи антитела человека, инкубируют в течение 1 ч при комнатной температуре и отмывают, после чего добавляют субстрат для выявления пероксидазы хрена. Уровни CDR-привитого OKT4A после ко-трансфекции pEe6HCDR1gs и pEe6LCDR1gs составляют 200 - 1200 мг/мл супернатанта COS-клеток.
Исследования связывания антигена.
CDR-привитое (гибридное) ОКТ4A, полученное в клетках COS, тестируют на его способность связываться с человеческими лимфоцитами периферической крови человека (ЛПК) и CD4-позитивной клеточной линией лимфоцитов больного, страдающего острым лейкозом периферической крови. Кроме того, гибридное тело тестируют на его способность блокировать связывание ОКТ4A мыши с этими клетками.
Связывание определяют следующим образом. ЛПК выделяют из сыворотки, а клетки острого лейкоза собирают из культурной ткани. Клетки инкубируют при 4oC в течение 1 ч с различными разведениями тестируемого антитела, положительного контрольного антитела и отрицательного контрольного антитела. Клетки однократно отмывают и инкубируют при 4oC в течение 1 ч с меченым ФИТЦ антителом козы к Ig человека (Fc - специфично, абсорбировано Ig мыши). Далее клетки дважды отмывают и анализируют цитофлуорографически. Химерное ОКТ4A (описанное ниже) используют в качестве позитивного контроля. ФИТЦ-меченое ОКТ4A мыши используют в качестве положительного контроля для связывания. Клетки инкубируют с супернатантом мнимо трансформированных клеток COS, образованными ФИТЦ-меченым антителом козы к IgG человека, и используют в качестве отрицательного контроля.
Для тестирования способности гибридного ОКТ4A блокировать связывание ОКТ4A мыши с ЛПК или клетками острого лейкоза, клетки инкубируют при 4oC в течение 1 ч с различными разведениями тестируемого антитела или контрольного антитела, затем добавляют насыщенное количество ФИТЦ ОКТ4A. Образцы инкубируют в течение 1 ч при 4oC, отмывают и анализируют цитофлуорографически. Положительным контролем служит ФИТЦ-меченое ОКТ4A для определения максимального связывания и немеченого ОТК4A мыши в качестве эталонного стандарта для определения блокирования. Отрицательным контролем служат неокрашенные клетки с добавлением или без добавления супернатанта мнимо трансформированных клеток.
Способность легкой цепи CDR-привитого ОКТ4A связывать CD4-положительные клетки и блокировать связывание ОКТ4A мыши первоначально тестируют в комплексе с тяжелой цепью гибридного ОКТ4A. Тяжелая цепь гибридного ОКТ4A состоит из вариабельного домена ОКТ4A и контактной области IgG4 человека. Ген гибридной тяжелой цепи экспрессируется в том же самом векторе, который используется для экспрессии CDR-"привитых" генов. Вектор экспрессии CDR-привитой легкой цепи и вектор экспрессии гибридной тяжелой цепи совместно переносят в COS-клетки. При этом было обнаружено, что полное гибридное ОКТ4A антитело (химерная легкая цепь и химерная тяжелая цепь) способно к полному связыванию CD4-положительных клеток и блокированию связывания ОКТ4A мыши с этими клетками.
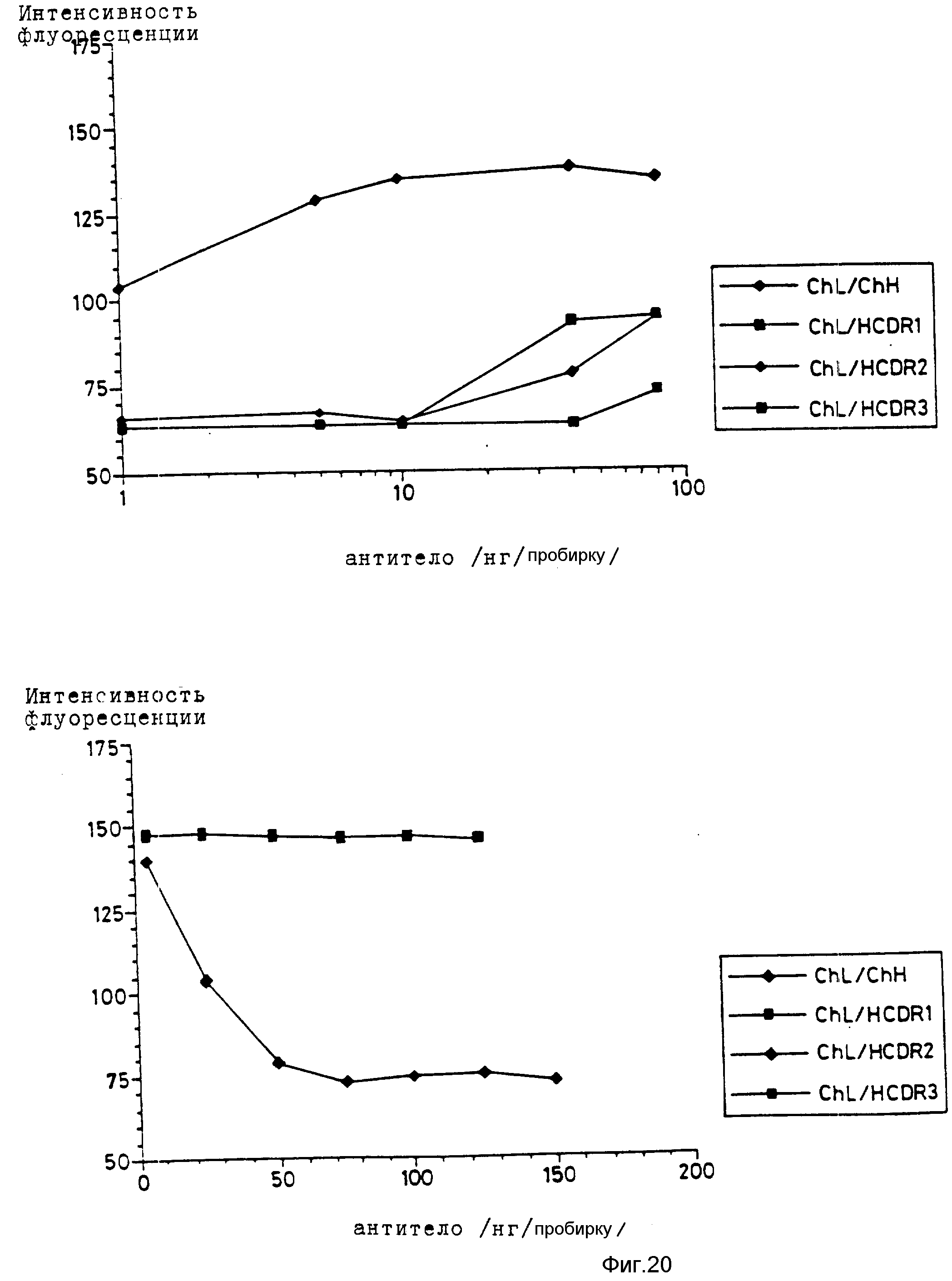
Как показано на фиг. 10, легкая цепь CDR-привитого ОКТ4А, LCDR1, в комбинации с тяжелой цепью химерного ОКТ4A была не способна связывать CD4-положительные клетки или блокировать связывание ОКТ4A мыши с этими клетками.
Фиг. 11 показывает исследования связывания и блокирования, проведенные с тяжелой цепью CDR-привитого ОКТ4A, HCDR1, объединенной с легкой цепью химерного ОКТ4A. Легкая цепь химерного ОКТ4A содержит вариабельный домен ОКТ4A мыши и контактный домен каппа-цепи Ig человека. Оно также экспрессируется в том же самом векторе, который используется для CDR-привитых антител. COS-клетки совместно трансфицируют вектором экспрессии CDR-привитой тяжелой цепи и вектором экспрессии химерной легкой цепи.
Тяжелая цепь CDR-привитого ОКТ4A, HCDR1 в комбинации с легкой цепью химерного ОКТ4A также не способна связывать CD4-положительные клетки или блокировать связывание ОКТ4A мыши с этими клетками.
Модификации CDR-привитого (гибридного) антитела.
Данные по связыванию к блокированию ясно показывают, что первоначальное сконструированное CDR-привитое антитело ОКТ4A не способно распознавать CD4 антиген. Необходима дальнейшая модификация антитела.
Для идентификации остатков, которые являются наиболее критическими для успешного антигенного взаимодействия, было использовано молекулярное моделирование.
Модификация CDR-привитой (гибридной) легкой цепи.
Кристаллическая структура ОКТ4A не была определена, поэтому молекулярная модель ОКТ4A сама по себе не может быть использована в анализе. Для анализа остатков CDR-привитой легкой цепи молекулярную модель легкой цепи антитела человека REI наклыдывают на модель Fab-фрагмента антитела мыши МОРС 603. Последовательность легкой цепи МОРС 603 сходна с аминокислотной последовательностью ОКТ4A.
Затем принимают решение расширить CDR1 путем замены остатков 33 и 34 (leu и asp) в антителе человека на остатки ile и ala ОКТ4A. При этом показано, что остаток glu 38 в REI-цепи антитела человека вовлечен в "упаковку" тяжелой цепи и легкой цепи. Замена этого остатка на his 38 ОКТ4A мыши может быть полезной. Остаток 49 на амино-терминальном участке CDR2 непосредственно "улучшает" CDR2 и также вступает в контакт с CDR3 тяжелой цепи. Остаток 89, близкий к амино-терминальному участку CDR3, взаимодействует с phe 98 в CDR3 легкой цепи и контактирует с CDR3 тяжелой цепи. REI tyr 49 и leu 89 заменяют на остатки his 49 и leu 89 ОКТ4A мыши. Новый ген CDR-привитой легкой цепи, полученный, как указано выше, обозначает LCDR2. Сравнение аминокислотной последовательности вариабельных доментов REI человека, LCDR1, LCDR2 и легкой цепи ОКТ4A мыши показано на фиг. 12. Замены осуществляют путем изменения кодонов при помощи сайт-направленного мутагенеза.
Вектор экспрессии pEe6LCDR2gs конструируют тем же способом, что и LCDR1. COS клетки совместно трансфицируют pEe6GLCDR2gs и вектором экспрессии химерной тяжелой цепи.
Результаты исследования связывания и блокирования приведены на фиг. 10. LCDR2 версия легкой цепи CDR-привитого ОКТ4A в комбинации с тяжелой цепью химерного ОКТ4A способа связывать CD4-положительные клетки и блокировать связывание ОКТ4A мыши. Эти данные показывают, что LCDR2 является функциональной легкой цепью CDR-привитого (гибридного) ОКТ4A.
Модификация CDR-привитой (гибридной) тяжелой цепи.
Для моделирования исследования тяжелой цепи используют молекулярную модель человеческого антитела KOL. Все замены остатков проводят путем сайт-направленного мутагенеза для изменения кодонов, заменяют glu 57 и his 58 KOL на thr 57 и tyr 58 ОКТ4A мыши. Эту CDR-привитую тяжелую цепь обозначают CDR2. В дополнение к заменам остатков 58 и 57 остаток 24 лежит вблизи CDR1 и может быть вовлечен в локализацию CDR1. Остатки 88 и 91 увлекаются в "упаковку" вариабельного домена тяжелой цепи и поверхность раздела между тяжелой и легкой цепью.
Эти три дополнительные замены остатков KOL на остатки ОКТ4A мыши характеризуют HCDR3 тяжелую цепь. Сравнение аминокислотной последовательности вариабельных доменов KOL, HCDR1, HCDR2, HCDR3, тяжелой цепи OKТ4A мыши и полученных выше цепей приведено на фиг. 13.
Векторы экспрессии pEe6HCDR2gs и pEe6HCDR3gs совместно вводят в COS клетки вместе с вектором экспрессии химерной легкой цепи ОКТ4A, pEe6LCDR2gs. Данные по связыванию и блокированию представлены на фиг. 11 и 14. Ни HCDR2, ни HCDR3 не способны эффективно взаимодействовать с антигеном в сочетании с легкой цепью химерного или CDR-привитого ОКТ4А.
Далее заменяют tyr 35 KOL на ser 35 ОКТ4A мыши. Молекулярное моделирование показывает, что остаток 42 вовлечен в локализацию CDR2. Остаток 44 вовлечен в контакты с легкой цепью. Целесообразно заменить gly 42 KOL и gly 44 KOL на glu 42 и arg 44 ОКТ4A мыши. Ala 60 KOL заменяют на pro 60 ОКТ4A мыши. Эти замены осуществляют в различных комбинациях, в то время как сохраняются замены, сделанные в отношении остатков 24, 57, 58, 88 и 91, как указано выше. Последние конструкции обозначают как HCDR4, HCDR5, HCDR6, HCDR7, HCDR8. Замены остатков в каждом случае приведены на фиг. 13. COS-клетки совместно трансфицируют векторами экспрессии новой тяжелой цепи и pEe6LCDR2gs.
Результаты исследований связывания и блокирования в комбинации с LCDR2 (фиг. 15) показывают положительные взаимодействия с CD4 антигеном всех конструкций, за исключением HCDR8. Очевидно, что замена tyr 35 на ser 35 антитела мыши является решающим изменением. Оказывается, что замена остатка в положении 60 антигена мыши устанавливает взаимодействие с антигеном (сравнение HCDR6 и HCDR4), тогда как замена в положении 44 слегка подавляет его (HCDR5 и HCDR4, и HCDR7 и HCDR6).
Определив, достаточно ли замен 35 и 60 для связывания антигена, HCDR9 (остатки антитела мыши в положениях 24, 35, 57, 58, 88 и 91) и HCDR10 (остатки антитела мыши в положениях 24, 35, 57, 58, 60, 88 и 91) получают путем сайт-направленного мутагенеза. Соответствующие векторы экспрессии вводят в клетки COS совместно с pEe6LCDR2gs. Результаты экспериментов по связыванию и блокированию приведены на фиг. 15 наряду с результатами исследований CDR-привитой тяжелой цепи. Тогда становится ясным то, что замены остатков 42 и 44 в предыдущих версиях не были необходимы, в противоположность критериям, изложенным в заявке PCT/US 89/05857. Замена остатка 60, произведенная в HCDR10, но не в LCDR9, является целесообразной.
Общие результаты анализа CDR-привитых тяжелых цепей и их активностей в связывании и блокировании приведены в табл. 1. Наиболее активное CDR-привитое ОКТ4A антитело, которое содержит наименьшее количество остатков антитела мыши представляет собой комбинацию HCDR10 и HCDR2.
Альтернативные конструкции легкой цепи.
Как установлено выше, конструкции легкой цепи получают исходя из предположения, что в положении 27 легкой цепи OKT4A имеется пролиновый остаток. Известно, что в положении 27 должен находиться остаток глутамина. Были получены и экспрессированы три новые конструкции легкой цепи. Они обозначены LCDE2Q, LCDR3Q, LCDR4Q и идентичны LCDR2, LCDR3 и LCDR4 соответственно, за исключением того, что в положении 27 находится глутаминовый (Q) вместо пролинового (P) остатка. Показано, что эти легкие цепи сохраняют полную активность. Данные, подтверждающие указанное, представлены на фиг. 16. При этом следует отметить, что пролин значительно отличается от других аминокислот в том, что имеет планарную структуру. Поэтому вероятно, что структура легкой цепи CDR1, имеющей пролин в положении 27, будет значительно отличаться от структуры легкой цепи CDR1, имеющей глутамин в положении 27. Несмотря на это, показано, что две легкие цепи являются эквивалентными с функциональной точки зрения. Это подтверждает высказанное здесь предположение о необходимости изменения всех 6 CDRS в антителе для того, чтобы получить функциональное CDR-привитое (гибридное) антитело.
Альтернативные модификации CDR-привитых легкой и тяжелой цепей.
Замены остатков, произведенные в более поздних версиях CDR-привитых легкой и тяжелой цепей, основаны на молекулярном моделировании REI, KOL и родственного мышиного антитела МОРС 603, а не CDR-привитых антител самих по себе. Некоторые замены могут быть не обязательными для связывания, особенно при более низких связывающих аффинностях.
Авторы сконструировали несколько генов CDR-привитой легкой и тяжелой цепи, в которых некоторые из каркасных остатков, предварительно замещенные на антитела мыши, были заменены обратно на остатки антитела человека. В целом эти остатки, непосредственно не вовлеченные в удлинение CDRS или локализацию CDRS, заменяются на остатки антитела человека в различных комбинациях.
В табл. 2 приведены указанные гены легкой и тяжелой цепи с некоторыми остатками, которые замещены обратно с мышиных на человеческие. Для конструирования этих генов используют сайт-направленный мутагенез. Они экспрессируются в COS-клетках и тестируются на способность узнавать CD4 в анализах связывания и блокирования. Наиболее желательным CDR-привитым антителом является антитело с наименьшим числом мышиных остатков, которые способны узнавать CD4, с аффинностью, аналогичной аффинности ОКТ4A антитела мыши.
Определение относительной связывающей аффинности.
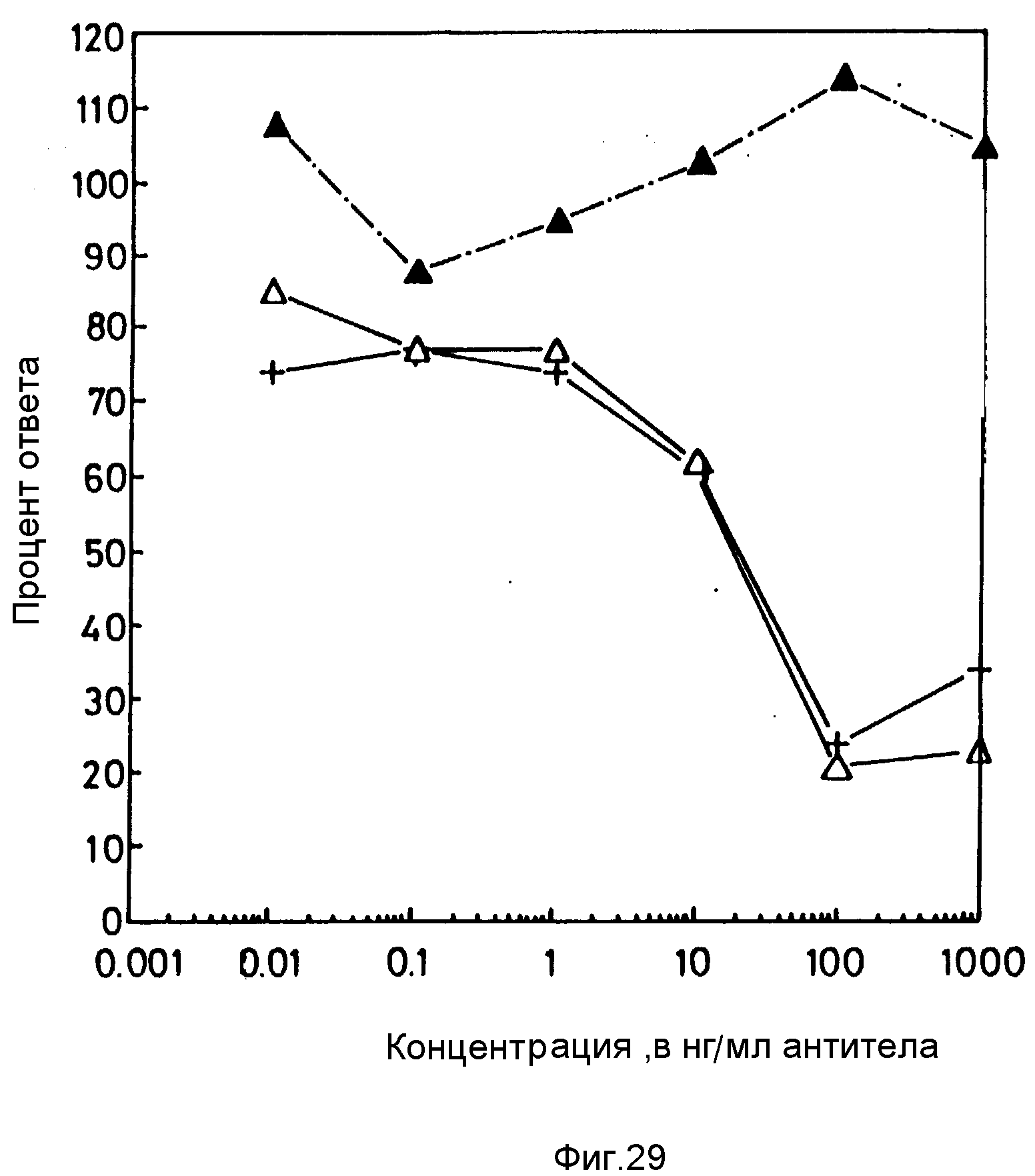
Относительные связывающие аффинности CDR-привитых анти-CD4 моноклональных антител определяют путем конкурентного связывания, используя Т-клеточную линию человека HPB-ALL как источник CD4 антигена и конъюгированное с флуоресцеином ОКТ4A мыши (F1-ОКТ4A) с известной связывающей аффинностью как индикаторное антитело. Связывающую аффинность F1-ОКТ4A индикаторного антитела определяют методом прямого связывания, в котором возрастающее количество F1-ОКТ4A инкубируют с HPB-ALL (5•105) в PBS с 5% эмбриональной сывороткой теленка в течение 60 мин при 4oC. Клетки отмывают и определяют интенсивность флуоресценции на проточном цитофлуориметре. Определяют отношение интенсивности флуоресценции на молекулу антитела (F/P отношение) путем использования микробус, которые несут определенное число антителсвязывающих сайтов IgG мыши. F/P определяют как интенсивности флуоресценции микробус, насыщенных F1-ОКТ4A, деленную на число связывающих сайтов на микробусах. Количество связанного и свободного F1-ОКТ4A вычисляют, исходя из значения интенсивности флуоресценции на клетку. Отношение свободное/связанное антитело в зависимости от числа молей связанного антитела вычисляют графически. Для определения аффинности связывания используют линейную зависимость (абсолютное значение наклона).
Для конкурирующего связывания, возрастающие количества конкурентного антитела добавляют к субнасыщающей дозе F1-ОКТ4A и инкубируют с 5 • 105 HPB-ALL в 200 мкл PBS с 5% эмбриональной сыворотки теленка в течение 60 мин при 4oC. Интенсивность флуоресценции клеток определяют на проточном цитофлуориметре. Вычисляют концентрации связанного и свободного F1-ОКТ4A. Аффинности конкурирующих антител вычисляют из уравнения [x] - [ОКТ4A] = (1/Кх) - (1/Kа), где Ка представляет собой аффинность ОКТ4A мыши; Кх - аффинность конкурирующего антитела; [х] - концентрацию конкурентного антитела, при которой отношение связанное/свободное представляет собой R/2, а R - максимальное отношение связанное/свободное.
Результаты исследований аффинности.
Определяют относительные константы аффинности гуманизированных антител (фиг. 17, табл. 3). LCDR2, комбинированная с HCDR10, сохраняет 68% активности исходного антитела. LCDR2/HCDR5 (табл. 1) сохраняет только 13% аффинности антитела мыши. Эти результаты согласуются с результатами, полученными в исследованиях блокирования (фиг. 16, a и b). Сравнение HCDR6 с HCDR7 (фиг. 15) позволяет предположить, что остаток 60, не являясь необходимым для сохранения активности, желателен для замещения на таковой в последовательности донора. На том же чертеже можно видеть эффект донорного остатка в положении 44 (HCDR4 и HCDR5).
Полученные антитела имеют ту же самую биологическую активность, что и ОКТ4A-антитело мыши.
Реферат
Изобретение относится к гибридному моноклональному антителу, взаимодействующему с CD4 -антигеном человека, содержащему легкие и тяжелые цепи человеческого происхождения, в которых области, определяющие комплементарность (CDRS), имеют принадлежность донорного ОКТ4А-антитела мыши, и способу получения данного антитела. Антитело имеет константу связывания с антигеном 4 • 105 - 1012 М-1 и относится к изотипу IgG. При этом в каркасных областях легких и тяжелых цепей гибридного антитела от двух до тринадцати аминокислот по системе нумерации Кабата дополнительно заменены на аминокислоты донорного антитела мыши. Для получения гибридного антитела конструируют соответствующие легкие и тяжелые цепи, которые встраивают в подходящий(ие) вектор(ы) экспрессии, трансформируют вектором(ами) эукариотические клетки и выделяют гибридное антитело из культуральной среды. 2 с. и 6 з.п.ф-лы, 30 ил., 3 табл.

Формула

Комментарии