Cd19xcd3-специфические полипептиды и их применение - RU2228202C2
Код документа: RU2228202C2
Чертежи


















Описание
Настоящее изобретение относится к новым одноцепочечным многофункциональным полипептидам, включающим по меньшей мере два антигенсвязывающих участка, специфических для антигенов CD19 и CD3 соответственно. Кроме того, настоящее изобретение относится к полипептиду, где указанный выше полипептид включает по меньшей мере один дополнительный домен, предпочтительно с заранее определенной функцией. Кроме того, настоящее изобретение относится к полинуклеотидам, кодирующим эти полипептиды, а также к векторам, включающим эти полинуклеотиды, и к трансформированным ими клеткам-хозяевам, и к их применению для получения указанных полипептидов. Изобретение также относится к композициям, предпочтительно фармацевтическим и диагностическим композициям, включающим любые из вышеуказанных полипептидов, полинуклеотидов или векторов. Еще одним объектом настоящего изобретения является применение указанных выше полипептидов, полинуклеотидов и векторов для приготовления фармацевтических композиций для иммунотерапии, предпочтительно для лечения злокачественных заболеваний В-клеток, таких как не-ходжкинская лимфома.
В настоящем описании процитировано несколько документов. Каждый из процитированных документов (включая любые спецификации производителей, инструкции и т.д.) включен в настоящее описание в виде ссылки; однако не подразумевается, что все процитированные документы действительно являются прототипами настоящего изобретения.
Несмотря на важное медицинское значение, при исследовании опосредованных В-клетками болезней, таких как не-ходжкинская лимфома, получено лишь небольшое количество приемлемых для клинического применения данных, и обычные подходы к лечению таких болезней остаются тривиальными и болезненными и/или имеют высокий риск рецидива. Например, хотя химиотерапия высокими дозами в качестве первичного лечения высокозлокачественной не-ходжкинской лимфомы может повысить общий уровень выживаемости, примерно 50% пациентов до сих пор умирают от этой болезни (2-4). Кроме того, низкозлокачественный напоминающий не-ходжкинскую лимфому хронический лимфатический лейкоз и лимфома клеток покрывающего слоя все еще являются неизлечимыми болезнями. Эти причины стимулировали разработку альтернативных стратегий типа иммунотерапии. Антитела, которые вырабатываются к молекулам клеточной поверхности, для которых характерны антигены CD, дают уникальную ценную возможность разработки терапевтических реагентов.
Экспрессия определенных антигенов CD в значительно степени ограничена специфическими линиями дифференцировки лимфогематопоэтических клеток и в последние несколько лет антитела, полученные к специфическим для лимфоидной ткани антигенам, применялись для разработки методов лечения, которые проявили эффективность либо in virto, либо на животных-моделях (5-13). В этой связи CD19 представляет собой очень ценную мишень. CD19 экспрессируется во всей В-клеточной линии дифференцировки от про-В-клетки до зрелой В-клетки, он не исчезает, единообразно экспрессируется на поверхности всех лимфатических клеток и отсутствует в стволовых клетках (8, 14). Представляет интерес применение биспецифического антитела специфического как по отношению к антигену CD19, так и к антигену CD3 на Т-клетках. Однако недостатком доступных в настоящее время биспецифических антител является их низкая Т-клеточная цитотоксичность, и для проявления достаточно высокой биологической активности необходимы дополнительные стимулирующие агенты.
Таким образом, техническая проблема, лежащая в основе настоящего изобретения, - это разработка способов и методик, пригодных для лечения опосредованных В-клетками болезней, таких как различные формы не-ходжкинской лимфомы. Решение этой технической проблемы достигается с помощью вариантов осуществления, которые представлены в формуле изобретения.
Таким образом, настоящее изобретение относится к одноцепочечному многофункциональному полипептиду, включающему
(а) первый домен, содержащий антигенсвязывающий участок цепи иммуноглобулина или антитела, который специфически распознает антиген CD19, и
(б) второй домен, содержащий антигенсвязывающий участок цепи иммуноглобулина или антитела, который специфически распознает антиген CD3.
Понятие "первый домен" и "второй домен" согласно настоящему изобретению обозначает, что один из антигенсвязывающих участков распознает pan-В-клеточный маркер CD19, который единообразно экспрессируется на поверхности практически всех злокачественных В-клеток, а другой антигенсвязывающий участок распознает антиген CD3 человеческих Т-клеток.
Понятие "антигенсвязывающий участок" в контексте настоящего изобретения обозначает домен, включающий трехмерную структуру, обладающую способностью специфически связываться с эпитопом, аналогичную таковой нативных антител, свободных scFv-фрагментов, или одной из их соответствующих цепей иммуноглобулина, предпочтительно VH -цепи. Таким образом, этот домен может включать VH- и/или VL-область антитела или цепи иммуноглобулина, предпочтительно по меньшей мере VH-область. С другой стороны, эти антигенсвязывающие участки, входящие в полипептид по изобретению, могут включать по меньшей мере один гипервариабельный участок (CDR) антитела или цепи иммуноглобулина, распознающий антигены CD19 и CD3 соответственно. В этой связи следует отметить, что домены антигенсвязывающих участков, которые присутствуют в полипептиде по изобретению, могут не только происходить из антител, но также из других CD19- или CD3-связывающих протеинов, таких как встречающиеся в естественных условиях поверхностные рецепторы или лиганды. Согласно изобретению, этот антигенсвязывающий участок включен в домен.
Понятие "многофункциональный пептид" в контексте настоящего описания обозначает полипетид, включающий по меньшей мере две аминокислотные последовательности, происходящие из разных источников, т.е. из двух различных молекул, необязательно происходящие из различных видов, причем по меньшей мере два из этих источников определяют специфичность антигенсвязывающих участков. Таким образом, эти антигенсвязывающие участки определяют специфичность функций или по меньшей мере некоторых функций этого многофункционального пептида. Такие полипептиды включают, например, биспецифические одноцепочечные (bsc) антитела.
Понятие "одноцепочечный" в контексте настоящего описания обозначает, что указанные первый и второй домен полипептида ковалентно связаны, предпочтительно в форме колинейной аминокислотной последовательности, кодируемой молекулой нуклеиновой кислоты. CD19 обозначает антиген, который экспрессируется В-клеточной линией дифференцировки, такой как про-В-клетка и зрелая В-клетка, он не исчезает, единообразно экспрессируется на поверхности всех лимфатических клеток и отсутствует в стволовых клетках (8, 14).
CD3 обозначает антиген, который экспрессируется на поверхности Т-клеток как часть многомолекулярного комплекса Т-клеточного рецептора, и он состоит из трех различных цепей CD3


Антитела, которые специфично распознают антиген CD19 или CD3, известны из существующего уровня техники, например описаны в (24), (25) и (43) соответственно, и могут быть получены с помощью известных в данной области методов.
Ранее было установлено, что биспецифические антитела CD19×CD3, которые не имеют одноцепочечный формат, повторно направляют Т-клеточную цитотоксичность на клетки лимфомы независимым от МНС образом, обладают эффективностью in vitro (5, 6, 9-11, 13, 43), на животных-моделях (7, 28), а также в некоторых пилотных клинических испытаниях (12, 29, 30). Для этой цели антитела конструировали с помощью методов на основе межвидовых гибридом путем ковалентного связывания моноклональных антител (31) или использования биспецифических антител (43). Проведение более обширных клинических исследований затруднено тем фактом, что эти антитела обладают низкой биологической активностью, что приводит к необходимости использовать высокие дозы, кроме того, применение одних антител не оказывало требуемого терапевтического действия. Кроме того, доступность клинически чистого материала была ограниченной.
Не вдаваясь в теорию, можно предположить, что полученные с помощью описанного выше биспецифического напоминающего антитела формата полипептиды, такие как биспецифические антитела CD19×CD3, как правило, могут разрушить CD19-позитивные клетки-мишени путем рекрутмента цитотоксичных Т-лимфоцитов, не нуждаясь при этом в пре- и/или костимуляции Т-клеток. Это находится в прямой противоположности со всеми известными биспецифическими антителами CD19×CD3, которые продуцируются согласно другим молекулярным форматам и, как правило, не зависят от конкретной CD19- или CD3-специфичности антитела, примененного в конструкции, например, биспецифического одноцепочечного антитела. Независимость от пре- и/или костимуляции Т-клеток может в значительной степени определять чрезвычайно высокую цитотоксичность, опосредуемую полипептидом по изобретению, что видно на примере использования конкретных биспецифических антител CD19×CD3, описанных в примерах.
Еще одним ценным качеством полипептида по изобретению является то, что в из-за своей небольшой, относительно компактной структуры он может быть легко получен и очищен, что решает проблемы, связанные с низким уровнем выхода, получением вредных побочных продуктов, или с процедурами лабораторной очистки (15-19), которые известны для CD19× CD3-специфических антител, получаемые с помощью межвидовых гибридом, путем химического связывания или путем ренатурации из бактериальных внутриклеточных телец. Далее преимущества и неожиданные свойства полипептида по изобретению будут обсуждены в приведенных примерах, которые не направлены на ограничение объема изобретения, включая некоторые приведенные ниже предпочтительные варианты осуществления, которые иллюстрируют широкую концепцию настоящего изобретения.
Согласно настоящему изобретению, применяли эукариотическую систему экспрессии, которая была разработана для получения рекомбинанатных биспецифических одноцепочечных антител (1) с целью производства рекомбинанатного биспецифического одноцепочечного антитела CD19×CD3 путем экспрессии в СНО-клетках. Полностью функционально активное антитело легко может быть очищено из супернатанта культуры с помощью его С-концевой гистидиновой метки хроматографией на Ni-NTA-колонке. Специфическое связывание с CD19 и CD3 было продемонстрировано анализом с использованием FASC (клеточный сортер с возбуждением флуоресценции). У полученной в результате молекулы bscCD19×CD3 (биспецифическое одноцепочечное антитело CD19×CD3) обнаружены некоторые неожиданные свойства:
- она индуцирует высокую, направленную к лимфоме, Т-клеточную цитотоксичность in vitro и in vivo. Даже в очень низких концентрациях на уровне 10-100 пг/мл и низких соотношения Е (эффектор): Т (мишень), таких как 5:1 и 2,5:1, наблюдали значительный специфичный лизис линий клеток лимфомы. Кроме того, при щадящем применении 3-10 мкг молекулы bscCD19×CD3 по изобретению обнаружено четкое и существенное улучшение медицинского состояния. По сравнению с известными к настоящему времени антителами CD19× CD3, полученными с помощью межвидовых гибридом и методов с использованием двойных антител (которые также представляют собой различные форматы), которые проявляют цитотоксическую активность в диапазоне от нескольких нанограмм/мл или даже мкг/мл, антитело bscCD19×CD3 по изобретению, вероятно, является намного более эффективным (5-7, 27, 43), например, как показано в приведенных далее примерах 4, 5 и 7.
Даже низкие концентрации bscCD19×CD3 по изобретению обладали способностью индуцировать быструю направленную к лимфоме цитотоксичность (через 4 ч) при низких соотношениях Е:Т и без необходимости в какой-либо предварительной стимуляции Т-клеток. В противоположность этому обычное биспецифическое антитело CD19×CD3 (5-7, 27) в этих условиях (а именно без предварительной стимуляции Т-клеток и при низком соотношении Е:Т) не проявило никакой заметной цитотоксической активности даже при высоких концентрациях, достигающих 3000 нг/мл. Хотя индукция цитотоксической активности без предварительной стимуляции ранее была установлена для другого обычного антитела CD19×CD3, этот эффект достигался только при более высоких концентрациях и высоких соотношениях Е:Т (100 нг/мл, 27:1) (9) по сравнению с bscCD19×CD3 по изобретению (100 пг/мл; 2,5:1). Кроме того, цитотоксическое действие этого обычно антитела наблюдали только через 1 день после предварительной стимуляции самим биспецифическим антителом, в то время как bscCD19×CD3 по изобретению индуцирует направленную к лимфоме цитотоксичность уже через 4 ч. По данным заявителей, такая быстрая и специфическая цитотоксическая активность нестимулированных Т-клеток при таких низких концентрациях и соотношениях Е:Т не описана для других применяемых ранее биспецифических антител. Хотя недавно было установлено, что F(ab)2-фрагмент биспецифического антитела к p185HER2/CD3 индуцирует цитотоксическую активность в таких же концентрациях, что и bscCD19×CD3 по изобретению, для этого антитела требуется предварительная стимуляция в течение 24 ч IL-2 (32). Таким образом, bscCD19×CD3-антитело по изобретению обладает уникальными свойствами цитотоксичности, что отличает эту молекулу от других известных биспецифических антител.
BscCD19×CD3 по изобретению опосредует цитотоксические действия, которые являются антигенспецифическими, о чем свидетельствуют такие факты:
- это антитело не обладает способностью осуществлять лизис линий клеток плазмацитомы NCI и L363, которые относятся к линиях клеток В-клеточной дифференцировки, не экспрессирующим антиген CD19; и
- цитотоксичность в отношении клеток лимфомы может блокироваться родительским антителом к CD19 HD37 (антитело HD37 происходит из гибридомы HD37 (22)).
Блокада пути биосинтеза перфорина путем истощения кальция с помощью ЭГТК (этиленгликольтетрауксусная кислота) полностью подавляла опосредуемую bscCD19×CD3 цитотоксичность, что позволяет предположить, что специфический лизис скорее опосредован Т-клетками, а не связан с непосредственным воздействием самого антитела.
Обобщая вышесказанное, можно заключить, что сконструированное согласно общим концепциям изобретения антитело bscCD19×CD3 превосходит ранее описанные биспецифические антитела CD19×CD3 как с точки зрения его значительно более высокой биологической активности, так и с точки зрения возможности его быстрого и простого способа получения, дающего большие количества высококачественного клинически чистого материала.
Таким образом, молекулы bscCD19×CD3 по изобретению могут рассматриваться в качестве приемлемых кандидатов для доказательства того, что терапевтически ценные биспецифические антитела могут использоваться для лечения опосредуемых В-клетками болезней, таких как не-ходжкинская лимфома, в клинических условиях.
В предпочтительном полипептиде по изобретению указанные домены соединены полипептидным линкером. Этот линкер расположен между первым и вторым доменом, причем этот полипептидный линкер предпочтительно включает многочисленные, гидрофильные, связанные пептидом аминокислоты, и он соединяет N-конец первого домена и С-конец второго домена.
Согласно еще одному предпочтительному варианту осуществления первый и/или второй домен описанного выше полипептида имитирует или соответствует VH- и VL-области природного антитела. Антитело, несущее антигенсвязывающий участок полипептида по изобретению, может представлять собой, например, моноклональное антитело, поликлональное антитело, химерное антитело, "гуманизированное" антитело, биспецифическое антитело, синтетическое антитело, фрагмент антитела, такой как Fab-, Fv- или scFv-фрагмент и т.д., или химические модифицированное производное любого из них. Моноклональные антитела могут быть получены, например, методами, первоначально описанными Кohler и Milstein, Nature 256 (1975), 495 и Galfre, Meth. Enzymol. 73, 3 (1981), которые включают слияние клеток мышиной миеломы с клетками селезенки, полученными из иммунизированных млекопитающих, и с помощью модификаций, принятых в данной области.
Кроме того, антитела к вышеуказанным антигенам или их фрагменты могут быть получены с использованием методов, описанных, например, у Harlow и Lane в "Antibodies, A Laboratory Manual", CSH Press, Cold Spring Harbor, 1988. Антитела могут быть получены из нескольких видов, включая человека. Когда производные указанных антител получают с помощью доступной методики с использованием фагов, то для повышения эффективности антител фагов, которые связываются с эпитопом антигена CD19 или CD3, может применяться поверхностный плазменный резонанс (Schier, Human Antibidies Hybridomas 7, 97-105 (1996); Malmbord, J. Immunol. Methods 183, 7-13 (1995)). Получение химерных антител описано, например, в WO 89/09622. Методы получения гуманизированных антител описаны, например, в ЕР-А 10239400 и WO 90/07861. Дополнительным источником антител, которые могут применяться согласно настоящему изобретению, являются так называемые ксеногенные антитела. Общие принципы получения ксеногенных антител, таких как человеческие антитела в мышах, описаны, например, в WO 91/10741, WO 94/02602, WO 96/34096 и WO 96/33735.
Антитела, которые применяются согласно изобретению, и их соответствующая(ие) иммуноглобулиновая(ые) цепь(и), могут быть дополнительно модифицированы с помощью обычных методов, известных в данной области, например, с использованием аминокислотной(ых) делеции(ий), инсерции(ий), замены(н), добавки(ок) и/или рекомбинации(ий) и/или другой (их) модификации(ий), известных в данной области, отдельно или в сочетании. Методы введения таких модификаций в последовательность ДНК, кодирующую аминокислотную последовательность цепи иммуноглобулина, хорошо известны специалистам в данной области и описаны, например, у Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (1989), N.Y. Описанные в этом руководстве модификации предпочтительно осуществляют на нуклеотидном уровне.
Согласно еще одному предпочтительному варианту осуществления изобретения, по меньшей мере один из указанных доменов в описанном выше полипептиде представляет собой одноцепочечный фрагмент вариабельной области антитела.
Как хорошо известно, Fv1, т.е. минимальный фрагмент антитела, который содержит полный сайт узнавания антигена и антигенсвязывающий участок, состоит из димера, включающего одну вариабельную область тяжелой цепи и одну вариабельную область легкой цепи (VH- и VL-области), которые не являются ковалентно связанными. В этой конфигурации, которая обнаружена в природных антителах, три гипервариабельных участка (CDR) каждой вариабельной области взаимодействуют для того, чтобы определить антигенсвязывающий участок на поверхности VH-VL-димера. В целом шесть CDR обусловливают специфичность связывания антигена антителом. Каркасные участки (FR), фланкирующие CDR, имеют третичную структуру, которая в основном является консервативной в природных иммуноглобулинах столь различных видов, как человек и мышь. Эти FR служат для поддержания CRD в соответствующей ориентации. Постоянные области не требуются для функции связывания, но они могут принимать участие в стабилизации VH-VL-взаимодействия. Даже отдельная вариабельная область (или половина Fv-фрагмента, включающего только три специфичных для антигена CDR) обладает способностью узнавать и связывать антиген, хотя, как правило, с более низкой аффинностью, чем полный антигенсвязывающий участок (Painter, Biochem., 11, 1327-1337 (1972)). Следовательно, этот домен антигенсвязывающего участка полипептида по изобретению может представлять собой пару VH-VL-, VH-VH- или VL-VL-областей либо одного и того же, либо различных иммуноглобулинов. Порядок расположения VH- и VL-областей в полипептидной цепи не имеет решающего значения для настоящего изобретения, приведенный выше порядок доменов может быть изменен, как правило, без какой-либо потери функции.
Однако важно, чтобы VH- и VL-области были расположены так, чтобы антигенсвязывающий участок мог быть соответствующим образом уложен (изогнут).
В предпочтительных полипептидах по изобретению эти домены расположены в следующем порядке: VLCD19-VHCD19-VHCD3-VLCD3, где VL и VH обозначают вариабельную область легкой и тяжелой цепи специфических антител к CD19 и CD3.
Как обсуждалось ранее, указанные антигенсвязывающие участки предпочтительно соединены гибким линкером, предпочтительно полипептидным линкером, который расположен между указанными доменами, причем этот полипептидный линкер предпочтительно включает многочисленные, гидрофильные, связанные пептидом аминокислотные последовательности, которые имеют длину, достаточную для того, перекрывать расстояние между С-концом одного из указанных доменов, включающего указанные антигенсвязывающие участки, и N-концом второго домена, включающего указанные антигенсвязывающие участки, когда полипептид по изобретению принимает пригодную для связывания конформацию при помещении его в водный раствор. Предпочтительно полипептидный линкер включает многочисленные остатки глицина, аланина и/или серина. Также предпочтительно, чтобы этот полипептидный линкер включал множество последовательных копий аминокислотных последовательностей. Как правило, полипептидный линкер включает 1-15 аминокислот, хотя полипептидные линкеры, состоящие более чем из 15 аминокислот, могут также хорошо выполнять свою функцию. Согласно предпочтительному варианту осуществления изобретения, полипептидный линкер включает 1-5 аминокислотных остатков.
Согласно особенно предпочтительному варианту осуществления изобретения, полипептидный линкер в полипептиде по изобретению состоит из 5 аминокислот. Как продемонстрировано в приведенных далее примерах, предпочтительно, чтобы полипептидный линкер включал аминокислотную последовательность Gly Gly Gly Gly Ser.
Согласно еще одному предпочтительному варианту осуществления изобретения, первый домен полипептида по изобретению включает по меньшей мере один CDR VH- и VL-области, который имеет аминокислотную последовательность, кодируемую последовательностью ДНК, представленной на фиг.8 от нуклеотида 82 до 414 (VL) и от нуклеотида 460 до 831 (VH), и/или второй домен включает по меньшей мере один CDR, более предпочтительно два, еще более три CDR VH- и VL-области, который(е) имеет(ют) аминокислотную последовательность, кодируемую последовательностью ДНК, представленной на фиг.8 от нуклеотида 847 до 1203 (VH) и от нуклеотида 1258 до 1575 (VL), необязательно в комбинации с каркасными участками, которые встречаются вместе в этих CDR в родительских антителах. CDR, содержащие указанные на фиг.8 вариабельные области, могут быть выявлены, например, согласно Кэбату, Kabat, "Sequences of Proteins of Immunological Interest" (U.S. Department of Health and Human Services, 3-е изд. 1983 г; 4-е изд. 1987 г.; 5-е изд. 1990 г.). Специалисту в данной области должно быть очевидно, что антигенсвязывающий участок или по меньший мере один полученный из него CDR может применяться для конструирования полипептида по изобретению. Предпочтительно этот полипептид включает аминокислотную последовательность, кодируемую последовательностью ДНК, представленной на фиг.8 от нуклеотида 82 по 1575. Специалисту в данной области должно быть очевидно, что антигенсвязывающие участки полипептида по изобретению могут быть сконструированы с помощью методов, известных в данной области, например описанных в ЕР-А10451261 и ЕР-А10549581.
Домены антигенсвязывающих участков полипептида по изобретению предпочтительно обладают специфичностью, по меньшей мере практически идентичны по специфичности связывания, например, антителу или цепи иммуноглобулина, из которых они получены. Такие домены антигенсвязывающих участков могут обладать аффинностью к связыванию по меньшей мере 105M-1, предпочтительно не выше чем 107M-1 в отношении антигена CD3 и предпочтительно до 1010М-1 или выше в отношении антигена CD19.
В предпочтительном полипептиде по изобретению
(а) антигенсвязывающий участок первого домена обладает аффинностью по меньшей мере примерно 10-7М, предпочтительно по меньшей мере примерно 10-9М и наиболее предпочтительно по меньшей мере примерно 10-11М и/или
(б) антигенсвязывающий участок второго домена обладает аффинностью ниже, чем примерно 10-7М, предпочтительно ниже, чем примерно 10-6 и наиболее предпочтительно порядка 10-5М.
Согласно указанным выше предпочтительным вариантам осуществления, целесообразно, чтобы антигенсвязывающий участок, который узнает антиген CD19, имел высокую аффинность для того, чтобы осуществлять захват клеток-мишеней, которые должны быть разрушены с высокой эффективностью. С другой стороны, аффинность связывания антигенсвязывающего участка, узнающего антиген CD3, должна иметь такой же порядок, что и природный CD3-рецептор, или соответствовать аффинности, которая наблюдается при взаимодействии Т-клеточного рецептора с лигандом, представляющим собой МНС-пептидный комплекс на поверхности клетки-мишени.
Согласно еще одному предпочтительному варианту осуществления изобретения, описанный выше полипептид представляет собой биспецифическое одноцепочечное антитело.
Настоящее изобретение также относится к полипептиду, включающему по меньшей мере один дополнительный домен, эти домены связаны ковалентными или нековалентными связями.
Связь может быть основана на генетическом слиянии, которое осуществляют согласно описанным выше известным в данной области методам, или может быть получена, например, путем химического перекрестного сшивания, как описано, например, в WO 94/04686. Дополнительный домен, присутствующий в полипептиде по изобретению, может предпочтительно быть связан гибким линкером, предпочтительно полипептидным линкером, с одним из доменов антигенсвязывающего участка, причем этот полипептидный линкер предпочтительно включает многочисленные, гидрофильные, связанные пептидом аминокислотные последовательности, которые имеют длину, достаточную для того, перекрывать расстояние между С-концом одного из указанных доменов и N-концом второго домена, когда полипептид по изобретению принимает пригодную для связывания конформацию при помещении его в водный раствор. Предпочтительно этот полипептидный линкер представляет собой полипептидный линкер, описанный ранее в предпочтительных вариантах осуществления. Полипептид по изобретению может также включать расщепляемый линкер или сайт расщепления протеиназами, такими как энтерокиназа (см. приведенные далее примеры).
Кроме того, этот дополнительный домен может иметь предварительно определенную специфичность или функцию. Например, в научной литературе имеется большое количество сведений о направленном переносе биологически активных веществ, таких как лекарства, токсины и ферменты, к определенным точкам в организме с целью разрушения или локализации злокачественных клеток или с целью индукции локализованного действия лекарства или фермента. Было предложено достигать этого действия путем конъюгации биологически активного вещества с моноклональными антителами (см., например, N.Y. Oxford University Press, и Ghose, J. Natl. Cancer Inst. 61, 657-676 (1978)).
В этом контексте также понятно, что полипептиды по изобретению могут быть дополнительно модифицированы обычными известными в данной области методами. Это позволяет конструировать химерные протеины, включающие полипептид по изобретению, и другие функционально активные аминокислотные последовательности, например сигналы ядерной локализации, трансактивирующие домены, ДНК-связывающие домены, гормонсвязывающие домены, протеины-метки (GST, GFP, пептид h-myc, FLAG, НА-пептид), которые могут быть получены из гетерологичных протеинов. Как описано ниже в примерах, полипептид по изобретению предпочтительно содержит метку FLAG, имеющую в длину 8 аминокислот (см. фиг.8).
Полипептиды по изобретению могут применяться для лечения пациентов, страдающих опосредованными В-клетками нарушениями, такими как В-клеточная лимфома, обусловленный В-клетками хронический лимфатический лейкоз (B-CLL), и/или имеющих связанное с В-клетками аутоимунное заболевание, такое как миостения тяжелая псевдопаралитическая, базедова болезнь, тиреоидит Хашимото или синдром Гудпасчера. Такая терапия может быть осуществлена, например, путем введения полипептидов по изобретению. Для такого введения могут использоваться как немеченые, так и меченые полипептиды.
Например, полипептиды по изобретению могут вводиться в виде полипептидов, меченных терапевтическим агентом. Эти агенты могут быть связаны либо непосредственно, либо опосредованно с антителами или антигенами по изобретению. Одним из примеров опосредованного связывания является связывание с помощью спейсерных фрагментов. Эти спейсерные фрагменты, в свою очередь, могут быть либо нерастворимыми, либо растворимыми (Diener, Science 231, 148 (1986)), и могут быть выбраны таким образом, чтобы позволять лекарству высвобождаться из антигена в месте-мишени. Примерами применяемых для иммунотерапии терапевтических агентов, которые могут быть связаны с полипептидами по изобретению, являются лекарственные средства, радиоактивные изотопы, лектины и токсины. Лекарственные средства, которые могут быть конъюгированы с полипептидами по изобретению, включают соединения, являющиеся классическими представителями лекарственных средств, такие как митомицин С, даунорубицин и винбластин.
При использовании полипептидов по изобретению, конъюгированных с радиоактивными изотопами, например для иммунотерапии, некоторые изотопы могут быть более предпочтительны, чем другие, в зависимости от таких факторов как распределение в лейкоцитах, а также стабильность и излучение. В зависимости от аутоиммуного ответа некоторые излучатели являются более предпочтительными, чем другие. В целом для иммунотерапии предпочтительными являются радиоактивные изотопы, испускающие α- и β-частицы. Предпочтительными являются короткоживущие, обладающие большой энергией α-излучатели, такие как212Bi. Примерами радиоактивных изотопов, которые могут быть связаны с полипептидами по изобретению для терапевтических целей, являются125 I,131I,90Y,67Cu,212Bi,212At,211Pb,47Sc,109Pd и188Re.
Лектины представляют собой протеины, обычно выделяемые из растительного материла, которые связываются с определенными фрагментами сахаров. Многие лектины также способны склеивать клетки и стимулировать лимфоциты. Причем рицин представляет собой токсичный лектин, который уже ранее применялся для иммунотерапии. Иммунотерапию проводили путем связывания α-пептидной цепи рицина, которая ответственна за токсичность, с полипептидом, способным обеспечивать сайтспецифичную направленность токсического действия.
Токсины - это ядовитые соединения, которые продуцируются растениями, животными или микроорганизмами и которые в соответствующей дозе часто являются летальными. Токсин дифтерии представляет собой вещество, продуцируемое Corynebacterium diphtheria, которое может применяться в терапевтических целях. Этот токсин состоит из α- и β-субъединиц, которые в соответствующих условиях могут быть разделены. Токсичный компонент А может быть связан с полипептидом по изобретению и может применяться для сайтспецифичного введения к взаимодействующим В-клеткам и Т-клеткам, которые находятся вблизи друг от друга в результате связывания с полипептидом по изобретению.
Другие терапевтические агенты из числа описанных выше, которые могут быть связаны с полипептидом по изобретению, а также соответствующие протоколы опытов ex vivi и in vitro являются известными или могут быть легко разработаны специалистами в данной области. При необходимости специалист в данной области может применять вместо самого белкового материала описанный ниже полинуклеотид по изобретению, кодирующий любой из полипептидов по изобретению, или соответствующие векторы.
Таким образом, специалисту в данной области должно быть очевидно, что полипептид по изобретению может применяться для конструирования других полипептидов с требуемой специфичностью и биологической функцией. Предполагается, что полипептиды по изобретению могут играть важную роль в области медицины как с терапевтической, так и с научной точки зрения, например, при создании новых способов лечения, связанных с В-клетками болезней, таких как определенные формы рака и аутоиммунных болезней, или в качестве перспективных инструментов анализа и модуляции путей трансдукции соответствующих клеточных сигналов.
Согласно еще одному варианту осуществления, указанный по меньшей мере один дополнительный домен содержит молекулу, выбранную из группы, включающей эффекторные молекулы, имеющие конформацию, пригодную для проявления биологической активности, аминокислотные последовательности, которые обладают способностью секвестровать ион, и аминокислотные последовательности, которые обладают способностью избирательно связываться с твердой подложкой или с предварительно отобранным антигеном.
Предпочтительно этот дополнительный домен включает фермент, токсин, рецептор, антигенсвязывающий участок, антигенсвязывающий участок синтезированного биологическим путем антитела, фактор роста, фактор клеточной дифференцировки, лимфокин, цитокин, гормон, обнаруживаемый на расстоянии фрагмент, антиметаболит, радиоактивный атом или антиген. Этот антиген может представлять собой, например, опухолевый антиген, вирусный антиген, микробный антиген, аллерген, аутоантиген, вирус, микроорганизм, полипептид, пептид или разнообразные опухолевые клетки.
Кроме того, последовательность, которая обладает способностью секвестровать ион, предпочтительно выбирают из последовательности калмодулина, металлотионеина, их функционально активного фрагмента, или из аминокислотной последовательности, богатой по меньшей мере одной из аминокислот, выбранных их глутаминовой кислоты, аспарагиновой кислоты, лизина и аргинина.
Кроме того, последовательность, которая обладает способностью избирательно связываться с твердой подложкой, может представлять собой положительно или отрицательно заряженную аминокислотную последовательность, содержащую цистеин-аминокислотную последовательность, авидин, стрептавидин, функционально активный фрагмент протеина A Staphylococcus, GST, His-метку, FLAG-метку или Lex А. Как описано в приведенных ниже примерах, полипептид по изобретению, примером которого является одноцепочечное антитело, также экспрессировали с FLAG-меткой на N-конце и/или с His-меткой на С-конце, что облегчает его очистку и определение. Используемая в примере FLAG-метка (см. фиг.8), состоит из 8 аминокислот, и ее предпочтительно применяют согласно изобретению. Однако также предпочтительными являются используемые в примерах FLAG-метки, представляющие собой укороченные версии FLAG, такие как аминокислотная последовательность Asp-Tyr-Lys-Asp.
Описанные выше эффекторные молекулы и аминокислотные последовательности могут присутствовать в виде предварительной формы (проформы), которая сама либо является, либо не является активной, и может быть удалена, например, при попадании в определенное окружение клетки.
Согласно наиболее предпочтительному варианту осуществления изобретения, этот рецептор представляет собой дополнительно стимулируемую поверхностную молекулу, важную для активации Т-клеток, или включает эпитопсвязывающий участок или гормонсвязывающий сайт.
Согласно еще одному из наиболее предпочтительных вариантов осуществления изобретения, эта дополнительно стимулируемая поверхностная молекула представляет собой CD80 (В7-1) или CD86 (В7-2).
Согласно еще одному предпочтительному варианту осуществления, настоящее изобретение относится к полинуклеотидам, которые при экспрессии кодируют вышеописанные полипептиды. Эти полинуклеотиды могут быть слиты в приемлемыми контролирующими экспрессию последовательностями, известными в данной области, для того, чтобы гарантировать правильную транскрипцию и трансляцию полипептида.
Такие полипептиды могут представлять собой, например, ДНК, кДНК, РНК или полученную синтетическим путем ДНК или РНК или полученную рекомбинантным путем химерную молекулу нуклеиновой кислоты, которая включает любой из этих полинуклеотидов по отдельности или в сочетании. Предпочтительно такой полинуклеотид является частью вектора. Такие векторы могут включать дополнительные гены, такие как маркерные гены, которые позволяют осуществлять отбор вектора в приемлемой клетке-хозяине и в пригодных средах. Предпочтительно полинуклеотиды по изобретению функционально связаны с контролирующими экспрессию последовательностями, которые позволяют осуществлять экспрессию в прокариотических и эукариотических клетках. Экспрессия полинуклеотида включает транскрипцию полинуклотида с получением транслируемой РНК. Специалистам в данной области хорошо известны регуляторные элементы, обеспечивающие экспрессию в эукариотических клетках, предпочтительно в клетках млекопитающих. Они, как правило, включают регуляторные последовательности, обеспечивающие инициацию транскрипции, или необязательно поли-А-сигналы, обеспечивающие терминацию транскрипции и стабилизацию транскрипта. Дополнительные регуляторные элементы могут включать энхансеры транскрипции, а также трансляции и/или встречающиеся в естественных условиях или гетерологичные промоторные области. Возможные регуляторные элементы, обеспечивающие экспрессию в прокариотических клетках-хозяевах, включают, например, промоторы PL, lac, trp или tac E. в coli, а примерами регуляторных элементов, обеспечивающих экспрессию в эукариотических клетках-хозяевах, являются промотор АОХ1 или GAL1 в дрожжах или промотор CMV, SSV40, RSV (вирус саркомы Рауса), энхансер CMV, энхансер SV40 или интрон глобина в клетках млекопитающих и в других клетках животных. Помимо элементов, ответственных за инициацию транскрипции, такие регуляторные элементы могут также представлять собой сигналы терминации транскрипции, такие как сайт SV40-поли-А или сайт tk-поли-А, расположенные по ходу транскрипции относительно полинуклеотида.
Кроме того, в зависимости от используемой системы экспрессии, лидерные последовательности, которые обладают способностью осуществлять направленный перенос полипептида в клеточный компартмент или обеспечивать направленную секрецию в среду, могут быть добавлены к кодирующей последовательности полинуклеотида по изобретению, и они хорошо известны в данной области; см., например, приведенные ниже примеры. Лидерную(ые) последовательность(и) объединяют на соответствующей фазе с последовательностями трансляции, инициации и терминации, и предпочтительная лидерная последовательность обладает способностью обеспечивать направленную секрецию транслируемого протеина или его части в периплазматическое пространство или внеклеточную среду. Необязательно гетерологичная последовательность может кодировать слитый протеин, который включает N-концевой идентифицирующий пептид, придающий требуемые характеристики, например, осуществляющий стабилизирующую функцию или упрощающий очистку экспрессируемого рекомбинантного продукта; см. выше. В этом контексте приемлемыми экспрессионными векторами являются известные в данной области векторы, такие как экспрессионный вектор кДНК Okayama-Berg (Окаяма-Берга) pcDV1 (фирма Pharmacia), pCDM8, pRc/CMV, pcDNA1, pcDNA3 (фирма In-vitrogen) или pSPORT1 (фирма GIBCO BRL).
Предпочтительно в векторах, которые обладают способностью трансформировать трансфектируемые эукариотические клетки-хозяева, контролирующие экспрессию последовательности, должны представлять собой системы эукариотических промоторов, но также могут использоваться и контролирующие последовательности прокариотических хозяев. После того, как вектор встроили в соответствующего хозяина, хозяина содержат в условиях, пригодных для достижения высокого уровня экспрессии нуклеотидных последовательностей, и при необходимости после этого может быть осуществлен сбор и очистка полипептида по изобретению (см. например, приведенные ниже примеры).
Как описано выше, полинуклеотид по изобретению может использоваться по отдельности или в качестве части вектора для экспрессии полипептида по изобретению в клетках, например, для генной терапии или диагностики болезней, связанных с В-клеточными нарушениями. Полинуклеотиды или векторы, несущие последовательность(и) ДНК, которая(ые) кодирует(ют) любой из описанных выше полипептидов, встраивают в клетки, которые, в свою очередь, продуцируют представляющий интерес полипептид. Генная терапия, основой которой является введение терапевтических генов в клетки с помощью методов ex-vivo или in-vitro, представляет собой наиболее важное применение переноса генов. Приемлемые векторы, методы или системы введения генов для генной терапии in-vitro или in-vivo описаны в литературе и известны специалистам в данной области, см., например, Giordano, Nature Medicine 2, 534-539 (1996); Schaper, Circ. Res. 79, 911-919 (1996); Anderson, Science, 256, 808-813 (1992); Verma, Nature, 389, 239 (1994); Inser, Lancet 348, 370-374 (1996); Muhlhauser, Circ. Res. 77, 1077-1086 (1995); Onodera, Blood, 91, 30-36 (1998); Verma, Gene Ther. 5, 692-699 (1998); Nabel, Ann. N.Y. Acad. ScL, 811, 289-292 (1997); Vewrzeletti, Hum. Gene Ther., 9, 2243-2251 (1998); Wang, Nature Medicine, 2, 714-716 (1996); WO 94/29469; WO 97/00957, US 5580859; US 5589466 или Schaper, Current Opinion in Biotechnology, 7, 635-640 (1996), и приведенные в этих публикациях ссылки. Полинуклеотиды и векторы по изобретению могут быть сконструированы для прямого введения или для введения с помощью липосом или вирусных векторов (например, аденовирусных, ретровирусных) в клетки. Предпочтительно эти клетки представляют собой зародышевую линию клеток, эмбриональную клетку или яйцеклетку или их производные, более предпочтительно эта клетка представляет собой стволовую клетку. Примером эмбриональной стволовой клетки среди прочего может быть стволовая клетка, описанная у Nagy, Proc. Natl. Acad. Sci. USA 90, 8424-8428 (1993).
Согласно вышесказанному, настоящее изобретение относится к векторам, прежде всего плазмидам, космидам, вирусам и бактериофагам, которые обычно применяют в генной инженерии, включающим полинуклеотид, кодирующий полипептид по изобретению. Предпочтительно этот вектор представляет собой экспрессионный вектор и/или вектор для переноса или для направленного переноса гена. Экспрессионные векторы, выведенные из вирусов, таких как ретровирусы, вирус коровьей оспы, аденоассоциированные вирусы, вирус герпеса простого или вирус бычьей папилломы, могут использоваться для введения полинуклеотидов или вектора по изобретению в популяцию клеток-мишеней. Для конструирования рекомбинантных векторов могут использоваться методы, хорошо известные специалистам в данной области; см., например, методики, описанные у Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory (1989), N.Y. и у Ausubel, Curren protocols in Molecular Biology, Green Publishinh Associates and Wiley Interscience, N.Y. (1989). В альтернативном варианте полинуклеотиды и векторы по изобретению могут быть восстановлены в липосомах для ведения в клетки-мишени. Векторы, содержащие полинуклеотиды по изобретению, могут быть перенесены в клетку-хозяина хорошо известными способами, которые могут варьироваться в зависимости от типа клетки-хозяина. Например, трансфекцию хлоридом калия обычно применяют для прокариотических клеток, а обработку фосфатом кальция или электропорацию можно использовать для других клеток-хозяев; см. Sambrook, выше. После экспрессии полипептиды по настоящему изобретению могут быть очищены согласно стандартным методикам в данной области, включая осаждение сульфатом аммония, хроматографию на аффинных колонках, хроматографию на колонках, гель-электрофорез и т.д.; см. у Scopes, "Protein Purification", Springer-Verlag, N.Y. (1982). Для фармацевтического применения предпочтительны практически чистые полипептиды, гомогенные по меньшей мере на 90-95%, более предпочтительно гомогенные на 98-99%. После очистки, частичной или при необходимости до гомогенного состояния, полипептиды могут использоваться в терапевтических целях (в том числе экстракорпорально) или для разработки или осуществления процедур анализов.
Согласно еще одному варианту осуществления, настоящее изобретение относится к клетке, содержащей полинуклеотид или вектор, которые описаны выше. Предпочтительно клетка представляет собой эукариотическую клетку, наиболее предпочтительно клетку млекопитающего, если предусматривается терапевтическое применение полипептида. Естественно, что также особенно, если полипептид применяют в диагностических целях, могут применяться клетки дрожжей и менее предпочтительно прокариотические клетки, например бактериальные клетки.
Полинуклеотид или вектор по изобретению, который находится в клетке-хозяине, может либо быть интегрирован в геном клетки-хозяина, либо может существовать вне хромосом.
Подразумевается, что понятие "прокариотический" включает все бактерии, которые могут быть трансформированы или трансфектированы молекулами ДНК или РНК для экспрессии полипептида по изобретению. Прокариотические хозяева могут включать грамотрицательные, а также грамположительные бактерии, например, такие как Е. coli, S. typhimurium, Serratia marcescens и Bacillus subtilis. Подразумевается, что понятие "эукариотический" включает клетки дрожжей, высших растений, насекомых и предпочтительно млекопитающих. В зависимости от хозяина, примененного для процедуры получения рекомбинантной молекулы, полипептиды по настоящему изобретению могут быть гликозилированы или могут не быть гликозилированы. Полипептиды по изобретению могут также включать аминокислотный остаток сайта инициации метионина. Полинуклеотид, кодирующий полипептид по изобретению, может применяться для трансформации или трансфекции хозяина с помощью любого из способов, обычно известных специалистам в данной области. Особенно предпочтительным является использование плазмид или вируссодержащей кодирующей последовательности полипептида по изобретению и генетически слитой с ней N-концевой FLAG-метки и/или С-концевой His-метки. Предпочтительно FLAG-метка имеет в длину 4-8 аминокислот, более предпочтительно 8 аминокислот. Методы получения слитых, функционально связанных генов и их экспрессии, например, в клетках млекопитающих и бактерий хорошо известны в данной области (Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor N.Y. (1989)). Описанные в этом руководстве генетические конструкции и методы могут применяться для экспрессии полипептида по изобретению в эукариотических или прокариотических хозяевах. В целом экспрессионные векторы, содержащие промоторные последовательности, которые облегчают эффективную транскрипцию встроенного полинуклеотида, применяют в зависимости от выбранного хозяина. Экспрессионные векторы, как правило, содержат сайт инициации репликации, промотор и терминатор, а также определенные гены, которые позволяют осуществлять отбор трансформированных клеток по фенотипическому признаку. Кроме того, для крупномасштабного производства полипептида по изобретению могут применяться трансгенные животные, предпочтительно млекопитающие, включающие клетки по изобретению.
Таким образом, согласно еще одному варианту осуществления, настоящее изобретение относится к способу получения описанного выше полипептида, предусматривающему культивирование клетки по изобретению в условиях, пригодных для экспрессии полипептида и выделения полипептида из клетки или из культуральной среды.
Трансформированные хозяева могут выращиваться в ферментерах и культивироваться согласно методам, известным в данной области, для достижения оптимального роста клеток. Полипептид по изобретению затем может быть выделен из среды для выращивания клеточных лизатов или фракций клеточных мембран. Выделение и очистка, например, экспрессируемых в микроорганизмах полипептидов по изобретению могут быть осуществлены любыми общепринятыми способами, такими как, например, препаративное хроматографическое разделение и иммунологическое разделение, в том числе включающие применение моноклинальных или поликлональных антител, например, к метке полипептида по изобретению, или согласно описанным ниже в примерах способам.
Таким образом, изобретение позволяет получить рекомбинантным путем полипептиды, включающие антигенсвязывающие участки, обладающие аффинностью и специфичностью в отношении эпитопа антигена CD19 и CD3 соответственно, и необязательно дополнительный функционально активный домен. Как ясно из вышеизложенного, изобретение относится к большому семейству полипептидов, включающих такие антигенсвязывающие участки, которые могут применяться для любых терапевтических и диагностических целей. Специалистам в данной области должно быть очевидно, что полипептиды по изобретению могут быть дополнительно слиты с другими описанными выше фрагментами, например, для направленного переноса лекарственного средства и с целью визуализации. Такое слияние может быть осуществлено химическим путем после экспрессии полипептидов с сайтом присоединения, или слитый продукт может быть создан в полипептиде по изобретению на уровне ДНК. ДНК затем экспрессируют в пригодной системе хозяина и полученные в результате экспрессии протеины собирают и при необходимости ренатурируют. Как описано выше, антигенсвязывающие участки предпочтительно получают из вариабельной области антител. В этой связи метод с использованием гибридом позволяет получить линии клеток, секретирующих антитело к практически любому требуемому веществу, которое вызывает иммунный ответ. Затем из цитоплазмы гибридомы может быть получена РНК, кодирующая легкие и тяжелые цепи иммуноглобулина. Согласно способу по изобретению, 5'-конец мРНК может использоваться для получения кДНК. Затем в клетках, предпочтительно в клетках млекопитающих может быть экспрессирована ДНК, кодирующая полипептиды.
В зависимости от выбора клетки-хозяина, для достижения соответствующей конформации могут требоваться методы ренатурации. При необходимости в ДНК могут быть сделаны точковые замены, способствующие оптимизации связывания, с использованием общепринятого мутагенеза и использованием кассет или другой методологии конструирования протеинов, в частности приведенной в данном описании. Получение полипептидов по изобретению также может зависеть от данных об аминокислотной последовательности (или соответствующей последовательности ДНК или РНК) биологически активных протеинов, таких как ферменты, токсины, факторы роста, факторы клеточной дифференцировки, рецепторы, антиметаболиты, гормоны или различные цитокины или лимфокины. Такие последовательности описаны в литературе и доступны из компьютеризированных банков данных. Например, может быть сконструирован полипептид по изобретению, который, например, состоит из одноцепочечного Fv-фрагмента и внеклеточной области человеческого дополнительно стимулируемого протеина CD80 (В7-1), соединенного с помощью линкера (Gly4Ser1)1. Дополнительно стимулируемый протеин CD80 принадлежит надсемейству Ig. Он представляет собой высокогликозилированный протеин, состоящий из 262 аминокислотных остатков. Наиболее подробное его описание опубликовано у Freeman, J. Immunol. 143, 2714-2722 (1989). Стабильная экспрессия может быть осуществлена, например, в СНО-клетках с дефицитом дигидрофолатредуктазы (DHFR), как описано у Kaufmann, Methods EnzymoL, 185, 537-566 (1990). Протеин затем может быть выделен с помощью His-метки, присоединенной к С-концу, с помощью Ni-NTA-колонки (Mack, Proc. Natl. Acad. Sci. USA, 92, 7021-7025 (1995)).
Кроме того, настоящее изобретение относится к композициям, включающим вышеуказанные полипептид, полинуклеотид или вектор по изобретению.
Предпочтительно настоящее изобретение относится к композициям, которые представляют собой фармацевтические композиции, включающие эти вышеуказанные полипептид(ы), полинуклеотид(ы) или вектор(ы) по изобретению.
Фармацевтическая композиция по настоящему изобретению может дополнительно включать фармацевтически приемлемый носитель. Примеры фармацевтически приемлемых носителей хорошо известны в данной области и включают забуференные фосфатом физиологические растворы, воду, эмульсии, такие как эмульсии масло/вода, различные типы смачивающих агентов, стерильные растворы и т.д. Композиции, включающие такие носители, могут быть приготовлены хорошо известными общепринятыми методами. Эти фармацевтические композиции могут вводиться пациенту в пригодной дозе. Введение приемлемых композиций может быть осуществлено различными путями, например путем внутривенного, внутрибрюшинного, подкожного, внутримышечного, местного или внутрикожного введения. Схема приема лекарственного средства может быть определена штатным врачом и она зависит от клинических факторов. Как хорошо известно из области медицины, для каждого пациента дозы зависят от многих факторов, включая рост пациента, площадь поверхности тела, возраст, конкретное соединение, подлежащее введению, пол, временя и путь введения, общее состояние здоровья, и от других лекарственных средств, которые применяют одновременно. В целом в случае регулярного применения фармацевтической композиции схема приема лекарственного средства должна включать введение от 1 мкг до 10 мг стандартных доз в день. Если схема приема лекарственного средства представляет собой продолжительную инфузию, то она также должна вводиться в диапазоне от 1 мкг до 10 мг стандартных доз на 1 кг веса тела в минуту соответственно. Однако более предпочтительная доза для продолжительной инфузии может составлять от 0,01 мкг до 10 мг стандартных доз на 1 кг веса тела в час. Особенно предпочтительные дозы приведены ниже. Их поступление можно определять с помощью периодической оценки. Дозы могут варьироваться, но предпочтительная доза для внутривенного введения ДНК составляет примерно от 106 до 1012 копий молекулы ДНК. Композиции по изобретению могут применяться локально или системно. Введение обычно является парентеральным, например внутривенным; ДНК также может вводиться непосредственно к месту действия, например, с помощью биобаллистического введения к внутреннему или внешнему месту действия или с помощью катетера к определенной области в артерии. Препараты для парентерального введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, включая физиологический раствор и забуференные среды. Носители для парентерального введения включают растворы хлористого натрия, декстрозу Рингера, декстрозу и хлорид натрия, лактированный раствор Рингера или жидкие жиры. Носители для внутривенного введения включают жидкие и питательные наполнители, электролитические наполнители (например, на основе декстрозы Рингера) и т.п. Могут присутствовать консерванты и другие добавки, такие как, например, антимикробные агенты, антиоксиданты, хелатирующие агенты и инертные газы и т.п. Кроме того, фармацевтическая композиция по настоящему изобретению может включать белковые носители типа, например, сывороточного альбумина или иммуноглобулина, предпочтительно полученные из организма человека. Кроме того, предусматривается, что фармацевтическая композиция по изобретению может включать дополнительные биологически активные агенты в зависимости от предполагаемого применения фармацевтической композиции. Такие агенты могут представлять собой лекарственные средства, действующие на желудочно-кишечную систему, лекарственные средства, действующие в качестве цитостатических агентов, лекарственные средства, предупреждающие гиперурикемию, и/или такие агенты, как костимулируемые Т-клеточные молекулы или цитокины, известные в данной области.
Согласно настоящему изобретению, подразумевается, что различные полинуклеотиды и векторы по изобретению вводят либо по отдельности, либо в любой комбинации с использованием стандартных векторов и/или систем введения генов и необязательно вместе с фармацевтически приемлемым носителем или эксципиентом. После введения эти полинуклеотиды или векторы могут быть стабильно интегрированы в геном пациента.
С другой стороны, могут применяться вирусные векторы, которые являются специфичными для определенных клеток или тканей и сохраняются в этих клетках. Фармацевтически приемлемые носители и эксципиенты хорошо известны в данной области. Фармацевтические композиции, полученные согласно изобретению, могут применяться для профилактики или лечения или замедления развития различных типов болезней, которые связаны с В-клеточными иммунодефицитами и злокачественными заболеваниями.
Кроме того, можно применять фармацевтическую композицию по изобретению, которая включает полинуклеотид или вектор по изобретения, для генной терапии. Приемлемые системы введения генов могут включать липосомы, опосредованные рецептором системы переноса, депротеинизированную ДНК и вирусные векторы, среди прочего такие как вирус герпеса, ретровирус, аденовирус и аденоассоциированный вирус. Перенос нуклеиновых кислот к определенному месту в организме с целью генной терапии также может быть осуществлен с использованием биобаллистической системы введения, такой как описана у Williams (Proc. Natl. Acad. Sci. USA, 88, 2726-2729 (1991)). Дополнительные способы введения нуклеиновых кислот включают опосредованный частицами перенос генов, например такой, как описан у Verma, Gen. Ther. 15, 692-699 (1998). Должно быть очевидно, что интродуцированные полинуклеотиды и векторы экспрессируют генный продукт после интродукции в клетку и предпочтительно сохраняются в этом состоянии на протяжении времени жизни клетки. Например, клеточные линии, которые стабильно экспрессируют полинуклеотид под контролем соответствующих регуляторных последовательностей, могут быть сконстурированы с помощью методов, хорошо известных специалистам в данной области. Клетки-хозяева могут быть трансформированы полинуклеотидом по изобретению с селектируемым маркером, находящимися на одной или на разных плазмидах, а не только экспрессионными векторами, которые содержат вирусные сайты инициации репликации. После интродукции чужеродной ДНК сконструированным клеткам можно позволить расти в течение 1-2 дней в обогащенной среде, а затем их переносят в среду для селекции. Селектируемый маркер в рекомбинантной плазмиде обусловливает устойчивость к агенту селекции и позволяет отбирать клетки, в хромосомы которые стабильно интегрирована плазмида, и выращивать их с получением локусов, которые, в свою очередь, могут быть клонированы и размножены с получением линий клеток. Такие сконструированные линии клеток также особенно пригодны для использования в системах скрининга для выявления соединений, участвующих, например, во взаимодействии В-клеток/Т-клеток.
Может применяться большое число систем селекции, включая тимидинкиназу вируса герпеса простого (Wigler, Cell, 11, 223 (1977), гипоксантин-гуанинфосфорибозилтрансферазу (Szybalska, Proc. Natl. Acad. Sci. USA, 48, 2026 (1962)) и аденинфосфорибозилтрансферазу (Lowy, Cell, 22, 817 (1980)) в tk-, hgpt- или aptt--клетках соответственно. Кроме того, может использоваться устойчивость к антиметаболитам, основанная на отборе гена dhfr, который придает устойчивость к метотрексату (Wigler, Proc. Natl. Acad. Sci. USA, 77, 3567 (1980); O'Hare, Proc. Natl. Acad. Sci. USA, 78, 1527 (1981)), гена gpt, который придает устойчивость к микофенольной кислоте (Mulligan, Proc. Nail. Acad. Sci. USA, 78, 2072 (1981), гена gpt, который придает устойчивость к аминогликозиду G-418 (Colberre-Garapin, J. Mol. BioL, 150, 1 (1981)), гена hygro, который придает устойчивость к гигромицину (Santerre, Gene, 30, 147 (1984) или пуромицину (pat, пуромицин-N-ацетаттрансфераза). Известны также другие селектируемые гены, например trpB, который позволяет клеткам утилизировать индол вместо триптофана, hisD, который позволяет клеткам утилизировать гистинол вместо гистидина (Hartman, Proc. Natl. Acad. Sci. USA, 85, 8047 (1982) и ODC (орнитиндекарбоксилаза), который придает устойчивость к ингибитору орнитиндекарбоксилазы, 2-(дифторметил)-DL-орнитину, DFMO (McCologue, (ред.) 1987 в: Current Communications in Molecular Biology, Cold Spring Harbor Laboratory).
Согласно еще одному варианту осуществления настоящее изобретение относится к диагностической композиции, включающей любые из вышеописанных полипептидов, полинуклеотидов или векторов по изобретению, и необязательные приемлемые способы обнаружения.
Полипептиды по изобретению также могут применяться для иммуноанализов, в которых они могут использоваться в жидкой фазе или в форме, связанной с твердофазным носителем. Примерами иммуноанализов, в которых может использоваться полипептид по изобретению, являются конкурентные и бесконкурентные иммуноанализы, проведенные либо в прямом или в непрямом формате. Примерами таких иммуноанализов является радиоиммноанализ (РИА), сэндвич-анализ (иммунометрический анализ) и Вестерн-блоттинг.
Полипептиды по изобретению также могут быть связаны с многочисленными различными носителями и применяться для выделения клеток, специфично связанных с этими полипептидами. Примеры хорошо известных носителей включают стекло, полистирол, поливинилхлорид, полипропилен, полиэтилен, поликарбонат, декстран, найлон, амилозы, природные и модифицированные целлюлозы, коллоидные металлы, полиактириламиды, агарозы и магнетит. Для целей изобретения носитель по своей природе может быть либо растворимым, либо нерастворимым.
Специалистам в данной области известны многочисленные метки и методы введения метки. Примеры типов меток, которые могут применяться согласно настоящему изобретению, включают ферменты, радиоактивные изотопы, коллоидные металлы, флуоресцентные соединения, хемилюминисцентные соединения и биолюминисцентные соединения, см. также описанные выше варианты осуществления.
Настоящее изобретение также относится к применению вышеописанного полипептида, полинуклеотида и вектора по изобретению для приготовления фармацевтической композиции для лечения В-клеточных злокачественных заболеваний, опосредуемых В-клетками аутоиммуных болезней или истощения В клеток.
Современные клинические испытания с использованием переориентированной цитотоксической активности человеческих Т-клеток с помощью биспецифических антител дали обещающие результаты в отношении лечения трудноизлечимой болезни Ходжкина (33), рака молочной железы и яичника (34-37) и злокачественной глиоамы (38). С учетом того, что
- bsc-антитела вследствие их низкой молекулярной массы облегчают проникновение к опухоли (что было доказано для Fab- и Fv-фрагментов) (39), и
- bsc-антитела, вероятно, уменьшают зависящую от дозы и ограниченную дозой токсичность, вызванную высвобождением системного цитокина, что связано с Fc-фрагментами обычных биспецифических антител (40), и
- даже интактное моноклональное антитело (синтезированное к CD20) приводит к регрессии опухолей на продвинутых стадиях не-ходжкинской лимфомы (NHL) (41, 42), можно ожидать, и это было продемонстрировано, что полипептиды по изобретению являются представляющими интерес молекулами, которые могут внести вклад в усовершенствование терапевтических методов лечения.
Таким образом, согласно предпочтительному варианту осуществления фармацевтическую композицию по изобретению используют для лечения не-ходжкинской лимфомы.
Диапазоны доз при введении полипептидов, полинуклеотидов и векторов по изобретению являются достаточно большими для того, чтобы вызвать требуемое действие, при котором симптомы опосредованных В-клетками болезней будут облегчены. Доза не должна быть столь высокой, чтобы вызывать значительные вредные побочные действия, такие как нежелательные перекрестные реакции, анафилактические реакции и т.п. В целом доза может варьироваться в зависимости от возраста, состояния, пола и состояния болезни у пациента и может быть определена специалистом в данной области. Доза может регулироваться каждым врачом в случае любых противопоказаний. Предусматривается, что диапазон доз составляет, например, от 0,01 мкг до 10 мг полипептида по изобретению. Особенно предпочтительными являются дозы, находящиеся в диапазоне от 0,1 мкг до 1 мг, еще более предпочтительно от 1 мкг до 100 мкг и наиболее предпочтительными от 3 мкг до 10 мкг, что, например, проиллюстрировано в приведенном далее примере 7.
Кроме того, изобретение относится к способу идентификации активирующих или дополнительно стимулирующих Т-клетки соединений или идентификации ингибиторов Т-клеточной активации или стимуляции, который предусматривает
(а) культивирование СD19-позитивных клеток (предпочтительно В-клеток) и Т-клеток в присутствии полипептида по изобретению и необязательно в присутствии соединения, обладающего способностью обеспечивать обнаруживаемый сигнал в ответ на Т-клеточную активацию соединением, подлежащим скринингу, в условиях, обеспечивающих взаимодействие соединения с клетками, и
(б) обнаружение присутствия или отсутствия сигнала, полученного в результате взаимодействия соединения с клетками.
Этот вариант осуществления особенно важен для тестирования способности соединений являться дополнительно стимулирующими молекулами. Согласно этому способу, CD19-позитивная клетка/В-клетка обеспечивает первичную активацию сигнала для Т-клетки, не используя таким образом клонотипичный Т-клеточный рецептор. Затем согласно изобретению может быть установлено, какое из тестируемых соединений также требуется для действительной активации Т-клетки. Согласно способу по изобретению, CD19-позитивная клетка/В-клетка функционируют как стимулирующая клетка, которая связывает биспецифические молекулы, соединенные с CD3-комплексами на поверхности этой же Т-клетки. Биологические методы культивирования, обнаружения и необязательно тестирования очевидны специалисту в данной области.
Понятие "соединение" в способе по изобретению включает отдельное вещество или несколько веществ, которые могут быть одинаковыми или неодинаковыми.
Понятие "соединение" может включать, например, такие образцы, как клеточные экстракты, полученные, например, из растений, животных или микроорганизмов. Кроме того, эти соединения могут быть известными в данной области, но для них может быть не известна способность ингибировать Т-клеточную активацию или не известно, что они могут применяться для дополнительной стимуляции Т-клетки соответственно. Многочисленные соединения могут быть, например, добавлены в культуральную среду или введены в клетки путем инъекции.
Если образец, содержащий соединение(я), идентифицируют согласно способу по изобретению, то далее либо можно выделить соединение из исходного образца, для которого установлено, что он содержит представляющий интерес соединение, либо можно дополнительно разделить исходный образец, если он состоит из нескольких различных соединений, для того, чтобы уменьшить количество различных веществ на образец и повторить процедуру с использованием подгрупп исходного образца. Затем с помощью методов, известных в данной области, таких, как приведены в настоящем описании и описаны ниже в примерах, можно определить, обладает ли образец или соединение требуемыми свойствами. В зависимости от сложности образцов описанные выше стадии могут быть проведены несколько раз, предпочтительно до тех пор, пока выявленный с помощью способа по изобретению образец не будет состоять только из ограниченного количества веществ или из одного вещества. Предпочтительно этот образец содержит вещества со сходными химическими и/или физическим свойствами и наиболее предпочтительно эти вещества являются идентичными. Способы по настоящему изобретению могут быть легко осуществлены и воспроизведены специалистами в данной области, например в опытах на других клетках, известных из уровня техники, или с использованием модификации способов, как описано в приведенных ниже примерах. Кроме того, специалист в данной области может легко установить, какие дополнительные соединения и/или клетки могут применяться для осуществления способов по изобретению, например интерлейкины или при необходимости ферменты, которые превращают определенное соединение в предшественник, который, в свою очередь, стимулирует или подавляет Т-клеточную активацию. Такая адаптация способа по изобретению хорошо известна специалистам в данной области и может быть осуществлена без проведения масштабных экспериментов.
Соединения, которые могут применяться согласно способу по настоящему изобретению, включают пептиды, протеины, нуклеиновые кислоты, антитела, небольшие органические соединения, лиганды, пептидомиметики, PNA (агглютинины из земляных орехов) и т.п. Эти соединения также могут представлять собой функционально активные производные или аналоги известных Т-клеточных активаторов или ингибиторов. Способы получения химических производных и аналогов хорошо известны специалистам в данной области и описаны, например, у Beilstein, Handbook of Organic Chemistry, Springer, ред. New York Inc., 175 Fifth Avenue, New York, N.Y. 10010 USA и Organic Synthesis, Wiley, New York, USA. Кроме того, эти производные и аналоги могут тестироваться в отношении их воздействий согласно известным методам или описанным, например, в приведенных ниже примерах. Также могут применяться пептидомиметики и/или созданные с помощью компьютерной технологии соответствующие активаторы или ингибиторы Т-клеточной активации, например, с помощью описанных ниже способов. Соответствующие компьютерные программы могут применяться для идентификации интерактивных сайтов предполагаемого ингибитора и антигена по изобретению на основе компьютерных анализов комплементарных структурных мотивов (Fassina, Immunomethods 5, 114-120 (1994)). Дополнительные соответствующие компьютерные системы, предназначенные для создания с помощью компьютерной технологии, протеина и пептидов, известны из существующего уровня техники, например описаны у Berry, Biochem. Soc. Trans. 22, 1033-1036 (1994); Wodak, Ann. N.Y. Acad. Sci. 501, 1-13 (1987); Pabo, Biochemistry 25, 5987-5991 (1986). Результаты, полученные с помощью описанного выше компьютерного анализа, могут использоваться в сочетании со способом по изобретению, например для оптимизации известных Т-клеточных активаторов или ингибиторов. Пригодные пептидомиметики также могут быть выявлены путем синтеза комбинаторных библиотек пептидомиметиков посредством последовательной химической модификации и тестирования полученных соединений, например согласно представленному в настоящем описании способу и нижеприведенных примеров. Методы создания и применения комбинаторных библиотек пептидомиметиков известны из существующего уровня техники, например описаны у Ostresh, Methods in Enzymology, 267, 220-234 (1996) и у Dorner, Bioorg. Med. Chem. 4, 709-715 (1996). Кроме того, для создания ингибиторов или активаторов Т-клеточной активации, которые могут тестироваться согласно способу по изобретению, может использоваться трехмерная и/или кристаллографическая структура ингибиторов или активаторов В-клеточного/Т-клеточного взаимодействия (Rose, Biochemistry 35, 12933-12944 (1996); Rutenber, Bioorg. Med. Chem. 4, 1545-1558 (1996)).
В целом настоящее изобретение относится к способам выявления соединений, которые обладают способностью модулировать опосредованные В-клеткой/Т-клеткой иммунные ответы.
Соединения, для которых обнаружена способность активировать опосредованные В-клеткой/Т-клеткой иммунные ответы, могут применяться для лечения рака и аналогичных болезней. Кроме того, также представляется возможным специфично ингибировать вирусные заболевания, тем самым предотвращая вирусную инфекцию или распространение вируса. Соединения, выявленные в качестве супрессоров Т-клеточной активации или стимуляции, могут применяться при трансформации органов с целью предотвращения отторжения трансплантата; см. также выше.
Также предполагается, что соединения, выявленные или полученные согласно способу по настоящему изобретению, могут применяться для диагностики и в конкретных терапевтических целях. Таким образом, еще один вариант осуществления изобретения относится к способу получения фармацевтической композиции, которая включает приготовление фармацевтически приемлемой формы соединения, выявленного на стадии (б) описанных выше способов по изобретению. Кроме того, подразумевается, что этот компонент может быть модифицирован с помощью пептидомиметиков. Способы получения и применения комбинаторных библиотек пептидомиметиков известны из существующего уровня техники и описаны, например, у Ostresh, Methods in Enzymology, 267, 220-234 (1996), Dorner, Bioorg. Med. Chem. 4, 709-715 (1996), Beeley, Trends Biotechnol. 12, 213-216 (1994) или у al-Obeidi, Mol. Biotech. 9, 205-223 (1998).
Терапевтически приемлемые соединения, выявленные согласно способу по изобретению, могут вводиться пациенту с помощью любого пути, приемлемого для этого соединения, например орально, внутривенно, парентерально, чрескожно или через слизистую оболочку, или с помощью хирургического вмешательства или имплантации (например, если соединение находится в форме твердого или полутвердого биологически совместимого и способного растворяться матрикса) в место или вблизи того места, где требуется действие соединения. Терапевтические дозы могут быть легко определены специалистом в данной области, см. выше.
Кроме того, настоящее изобретение относится к способу лечения злокачественных В-клеточных заболеваний, опосредованных В-клетками аутоиммунных болезней или истощения В-клеток, и/или к способу замедления развития патологического состояния, которое вызывается В-клеточными нарушениями, предусматривающему введение полипептида, полинуклеотида или вектора по изобретению млекопитающему, страдающему этими злокачественными болезнями и/или патологическим состоянием. Также предпочтительно, если это млекопитающее представляет собой человека.
Эти и другие варианты осуществления представлены и включены путем описания и примеров в настоящее изобретение. Дополнительные литературные данные, касающиеся любых антител, способов, особенностей применения и соединений, которые могут применяться согласно настоящему изобретению, могут быть получены из опубликованных библиотек и баз данных, с помощью, например, электронных систем. Например, опубликованная база данных "Medline" может быть получена через Интернет, например, по следующему адресу электронной почты: http://www.ncbi.nIm.nih.gov./PubMed/medline.html. Другие базы данных и адреса, такие как http://www.ncbi.nlm.nih.gov./, http://www.infobiogen.fr/, http://www.fmi.ch/biology/research_tools.html, http://www.tigr.org/, известны специалистам в данной области или также могут быть получены, например, по адресу http://www.lycos.com. Обзор патентной информации в области биотехнологии и относящихся к делу источников патентной информации, которые могут использоваться для ретроспективного поиска и получения современных данных, приведен у Berks, TIBTECH 12, 352-364 (1994).
Описание фиг.1-22:
Фиг.1:
ДСН-ПААГ: окрашивание с помощью кумасси очищенного bscCD19xCD3-фрагмента с различными количествами протеинов. Слева показана молекулярная масса (кДа) маркера
Фиг.2:
FACS-анализ bscCD19xCD3 (200 мкг/мл) из различных CD19-позитивных линий В-клеток (BJAB, SKW6.4, Blin-1, Daudi, Raji) CD19-негативной линии В-клеток BL60 и CD3-позитивных клетках линии Jurkat и первичных человеческих РВМС (мононуклеары периферической крови). Штриховыми линиями показаны отрицательные контроли.
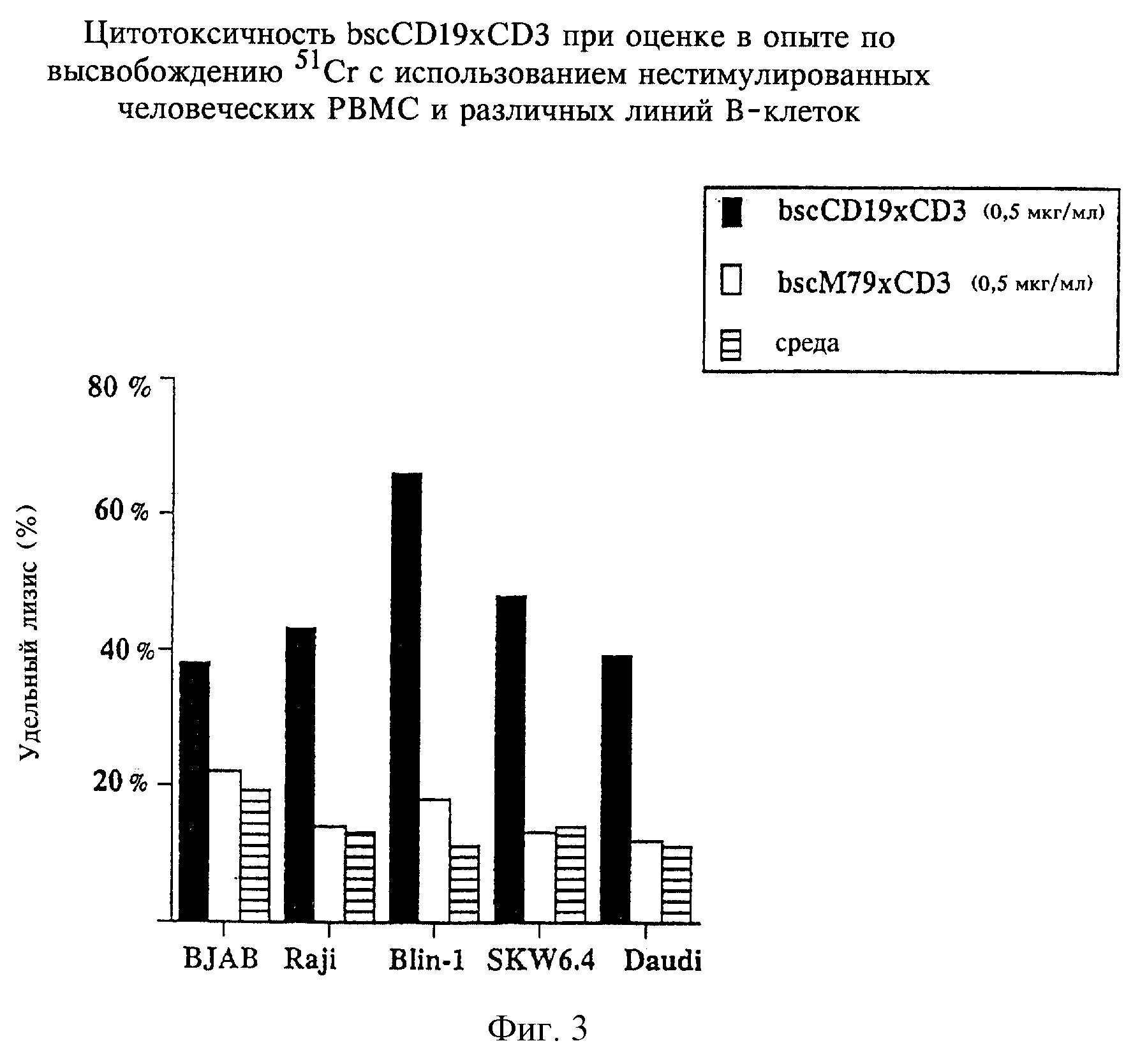
Фиг.3:
Цитотоксичность bscCD19×CD3 при оценке в опыте по высвобождению5lCr с использованием нестимулированных человеческих РВМС и различных линий В-клеток.
Соотношение клеток-эффекторов: клеток-мишеней составляет 10:1; время инкубации 4 ч. Стандартные отклонения по всем 3 повторностям ниже 7%.
Фиг.4:
Цитотоксичность при оценке в опыте по высвобождению хрома с использованием нестимулированных первичных человеческих PBL (лимфоциты периферической крови) к плазмацитомным линиями клеток L363 и NCI и линии клеток лимфомы Daudi. Соотношение Е:Т 20:1, время инкубации 8 ч.
Фиг.5:
Ингибирование цитотоксичности bscCDl9×CD3 родительским антителом к CD19 HD37 в опыте по высвобождению хрома; время инкубации 8 ч; соотношение Е:Т 20:1, концентрация bscCDl9×CD3 1 нг/мл.
Фиг.6:
Оценка цитотоксичности с использованием нестимулированных РВМС в отношении клеток линии Daudi после добавления возрастающих количеств ЭГТК; соотношение Е:Т 10:1, время инкубации 4 ч.
Фиг.7:
Цитотоксичность bscCDl9×CD3 при оценке в опыте по высвобождению15Сr с использованием нестимулированных человеческих РВМС и линии Blin-1 в качестве клеток-мишеней при различных соотношениях Е:Т; время инкубации 4 ч.; концентрация обычного биспецифического антитела 3 мкг/мл; концентрация bsc17-1A×CD3 100 нг/мл; соотношения Е:Т, как указано выше.
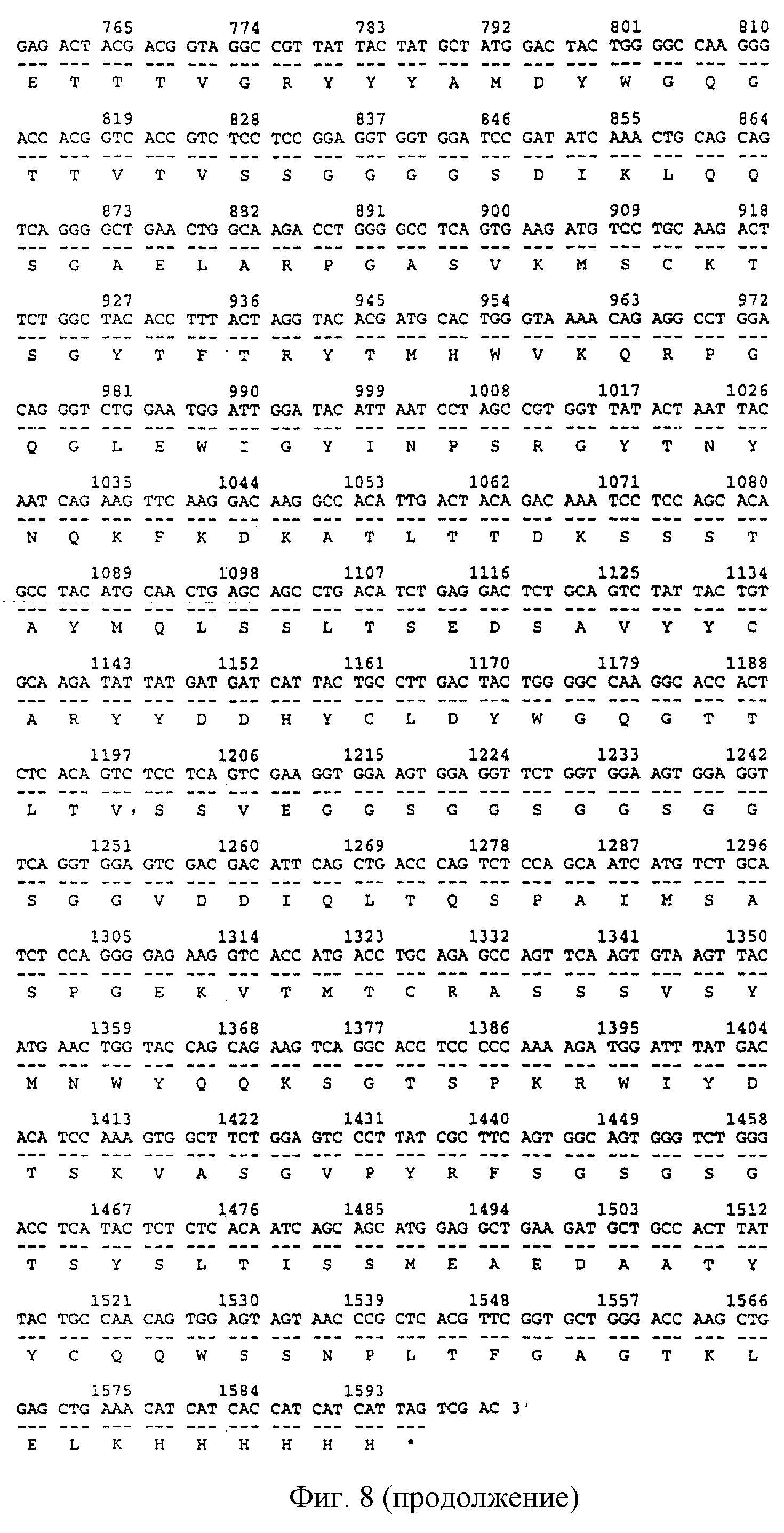
Фиг.8:
Последовательность ДНК и протеина антитела bscCDl9×CD3 (вариант с FLAG-меткой). Цифрами обозначены положения нуклеотидов (нт), соответствующая аминокислотная последовательность приведена под нуклеотидной последовательностью. Кодирующая последовательность ДНК биспецифического антитела начинается с положения 1 и заканчивается положением 1593. Первые 6 нт (положения от -10 до -5) и последние 6 нт (положения 1596-1601) содержат сайты расщепления рестриктазами EcoRI и Sail соответственно. Нуклеотиды 1-57 специфичны для лидерной последовательности; нуклеотиды 82-414 и 460-831 кодируют VL-область CD19 и VH-область CD19 соответственно; нуклеотиды 847-1203 и 1258-1575 кодируют VH-область CD3 и VL-область CD3 соответственно и нуклеотиды 1576-1593 кодируют His-метку.
Фиг.9:
Уменьшение первичных (злокачественных) СD19+-B- клеток путем рекрутмента аутологичных первичных Т-лимфоцитов с помощью bscCD19×CD3
А. Исходный момент времени (t=0): n=3·106 PBL/лунку высевают в 24-луночные планшеты для культуры ткани в каждую лунку в 1 мл RPMI 1640, дополненной 10%-ФТС. Определяют начальное процентное содержание CD19+-В-клеток, а также CD4+- и CD8+-Т-клеток.
Б-Ж. Относительное содержание В- и CD4+ - и CD8+-Т-клеток после инкубации в течение 5 дней (t=5 дней) при 37°С/5% СО2 в отсутствии (Б-В) или в присутствии (Д-Ж) bscCD19×CD3 (концентрации указаны) с добавлением 60 ед./мл IL2 или без IL2. Негативные контроли включают либо биспецифическое одноцепочечное антитело (17-1A×CD3) с несоответствующей специфичностью в отношении клеток-мишеней или вообще не включают биспецифическое антитело (В).
Фиг.10:
Стадии очистки bscCD19×CD3
Фиг.11:
ДСН-ПААГ анализ чистоты bscCD19×CD3
Приведены данные, полученные с помощью окрашенного коллоидным кумасси голубым ДСН с использованием 4-12%-ного градиента полиакриламидного геля. Полосы 1 и 6 - маркеры молекулярной массы; полоса 2 - супернатант клеточной культуры; полоса 3 - активная фракция, полученная с помощью катионообменной хроматографии; полоса 5 - активная фракция, полученная с помощью гель-фильтрации. Анализировали одинаковые количества протеина (2 мкг) из супернатанта клеточной культуры и различных колоночных фракций. Справа показаны стандарты молекулярной массы в кДа. Стрелкой показано положение bscCD19×CD3.
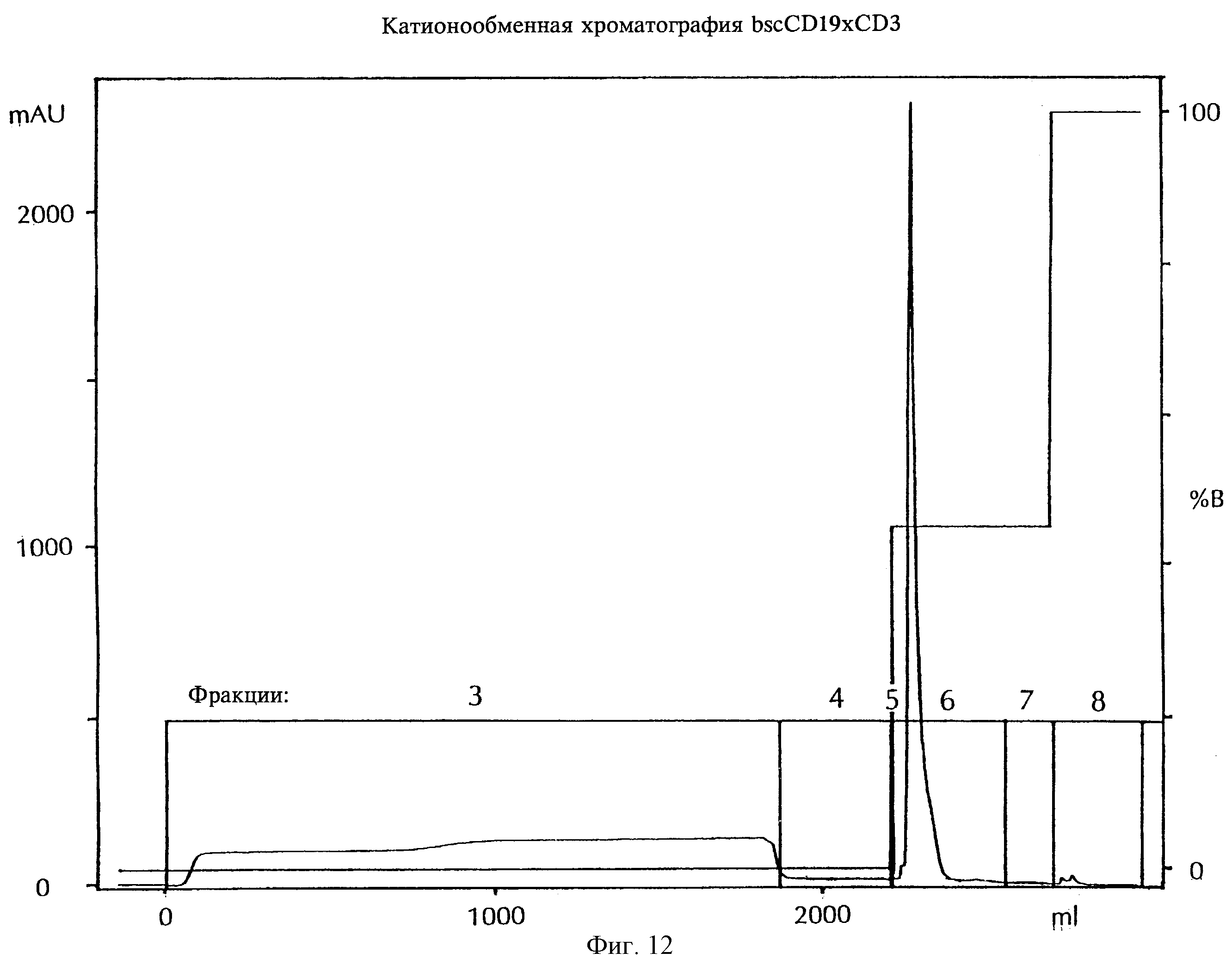
Фиг.12:
Катионообменная хроматография bscCD19×CD3
Концентрацию протеина измеряли по абсорбции при 280 нм (mAU, слева). Жирной линией показан профиль элюции протеина. Профиль, полученный с использованием ступенчатого градиента NaCl, показан прямой жирной линией (% В, справа), и собранные фракции показаны штриховыми линиями, bscCD19×CD3 обнаружено во фракции F6.
Фиг.13:
Кобальтовая хелатная аффинная хроматография bscCD19×CD3
Концентрацию протеина измеряли по абсорбции при 280 нм (mAU, слева). Жирной линией показан профиль элюции протеина. Профиль, полученный с использованием градиента имидазола, показан прямой жирной линией (% В, справа), и собранные фракции показаны штриховыми линиями, bscCD19×CD3 обнаружено во фракции F7.
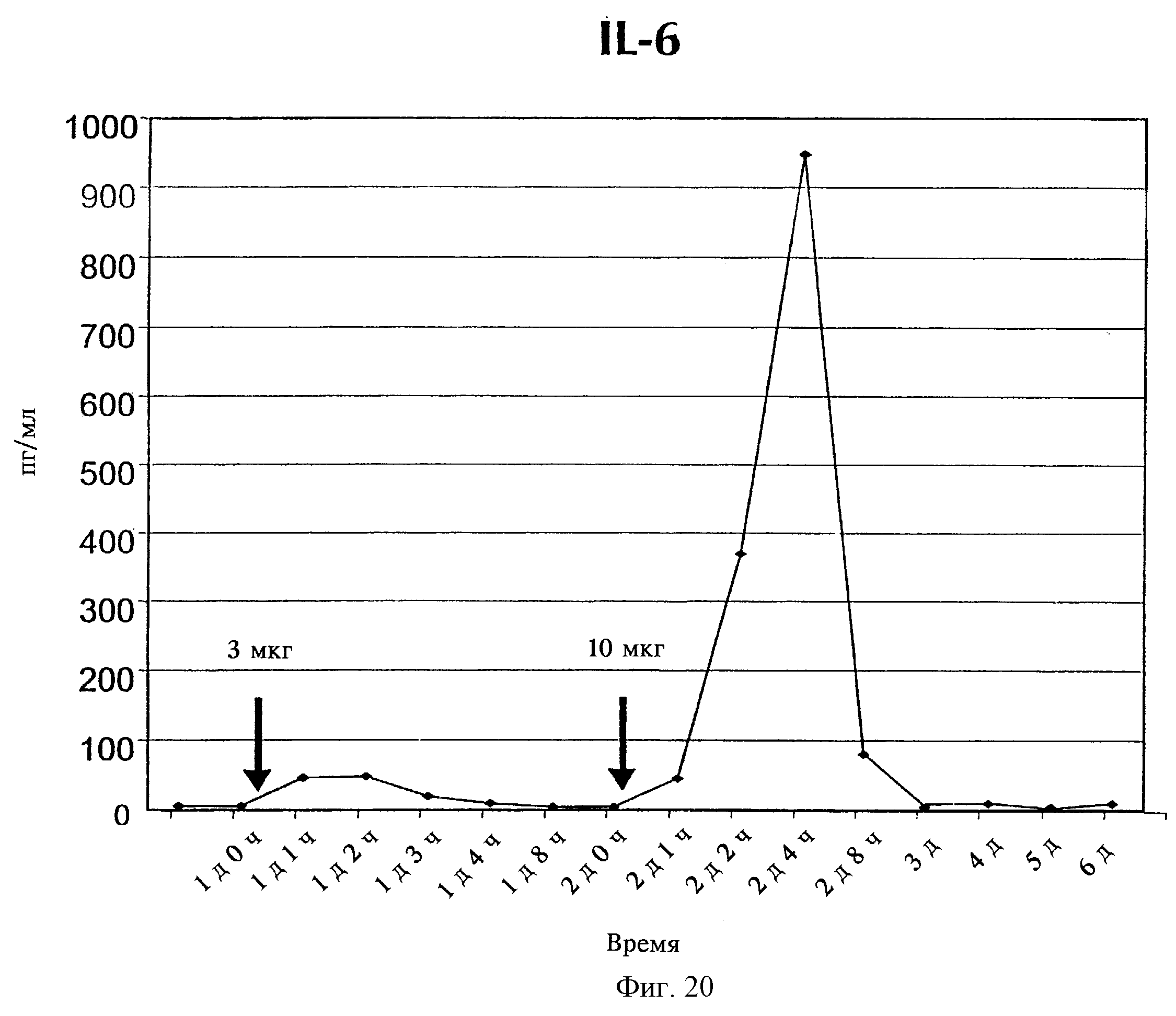
Фиг.14:
Гель-фильтрация bscCD19×CD3
Концентрацию протеина измеряли по абсорбции при 280 нм (mAU, слева). Жирной линией показан профиль элюции протеина. Штриховыми линиями показаны собранные фракции, bscCD19×CD3 обнаружено во фракции F7, соответствующей молекулярной массе примерно 60 кДа.
Фиг.15:
Уровни в крови гамма-глутамилтрансферазы (GGT) в ответ на обработки bscCD19× CD3
Уровни GGT определяли стандартным методом клинической биохимии и выражали в виде ед./л. На оси абсцисс показано время в днях (д) после начала первой обработки лекарственным средством и, начиная с 0 часов (ч) после каждого введения лекарственного средства. Стрелками показаны моменты времени введения лекарственного средства.
Фиг.16:
Измерения с помощью ультразвука селезенки пациентки А-Б
А. Определение размера селезенки, датированное 12 апреля 1999 г., до терапии с использованием bscCD19×CD3. На чертеже видна увеличенная по размеру селезенка (размер 146 мм × 69,2 мм), что является результатом инфильтрации злокачественных В-клеток.
Б. Определение размера селезенки, датированное 16 апреля 1999 г., после введения 3 мкг 14 апреля и 10 мкг 15 апреля. На чертеже видно уменьшение размера селезенки до 132 мм × 58,9 мм, вызванное системной обработкой bscCD19×CD3. Различия между отдельными данными измерений, которые видны из таблицы 1, объясняются определением с помощью ультразвука размеров в разных пространственных плоскостях. Два положения обозначены с помощью (+) и (×).
Фиг.17:
Содержание лейкоцитов в крови в ответ на обработку bscCD19×CD3.
Количество лейкоцитов дано в Гч./л. На оси абсцисс показано время в днях (д) после начала первой обработки лекарственным средством, и начиная с 0 часов (ч), после каждого введения лекарственного средства. Стрелками показаны моменты времени введения лекарственного средства.
Фиг.18:
Содержание в крови С-реактивного протеина (CRP) в ответ на обработку bscCD19×CD3
Уровни CRP определяли стандартным методом клинической биохимии и выражали в виде мг/дл. На оси абсцисс показано время в днях (д) после начала первой обработки лекарственным средством и, начиная с 0 часов (ч), после отдельных добавлений лекарственного средства. Стрелками показаны моменты времени введения лекарственного средства.
Фиг.19:
Содержание в крови фактора некроза опухоли альфа (TNF) в ответ на обработку bscCD19×CD3
Уровни TNF определяли с помощью ELISA и выражали в нг/мл. На оси абсцисс показано время в днях (д) после начала первой обработки лекарственным средством и, начиная с 0 часов (ч), после каждого введения лекарственного средства. Стрелками показаны моменты времени введения лекарственного средства.
Фиг.20:
Содержание в крови интерлейкина-6 (IL-6) в ответ на обработку bscCD19×CD3
Уровни IL-6 определяли с помощью ELISA и выражали в пг/мл. На оси абсцисс показано время в днях (д) после начала первой обработки лекарственным средством и, начиная с 0 часов (ч), после каждого введения лекарственного средства. Стрелками показаны моменты времени введения лекарственного средства.
Фиг.21:
Содержание в крови интерлейкина-8 (IL-8) в ответ на обработку bscCD19×CD3
Уровни IL-8 определяли с помощью ELISA и выражали в пг/мл. На оси абсцисс показано время в днях (д) после начала первой обработки лекарственным средством и, начиная с 0 часов (ч), после каждого введения лекарственного средства. Стрелками показаны моменты времени введения лекарственного средства.
Фиг.22:
Содержание в крови альфа-цепи растворимого рецептора интерлейкина-2 (IL-2R) в ответ на обработку bscCD19×CD3
Уровни IL-2R определяли с помощью ELISA и выражали в пг/мл. На оси абсцисс показано время в днях (д) после начала первой обработки лекарственным средством и, начиная с 0 часов (ч), после каждого введения лекарственного средства. Стрелками показаны моменты времени введения лекарственного средства.
Ниже изобретение описано со ссылкой на следующие биологические примеры, которое даны только с целью иллюстрации и не направлены на ограничение объема настоящего изобретения.
Пример 1: Клонирование вариабельных (V) областей иммуноглобулина
V-область легкой цепи (VL) и V-область тяжелой цепи (VH) из гибридомы HD37 (22) клонировали согласно стандартным способам ПЦР (23). Синтез кДНК осуществляли с использованием олиго-дТ-праймеров и полимеразы Taq.
Перечень праймеров
5'L1:GAAGCACGCGTAGATATCKTGMTSACCCAAWCTCCA [SEQ ID:1] 3'K:GAAGATGGATCCAGCGGCCGCAGCATCAGC [SEQ ID:2] 5'H1:CAGCCGGCCATGGCGCAGGTSCAGCTGCAGSAG [SEQ ID:3] 3'G:ACCAGGGGCCAGTGGATAGACAAGCTTGGGTGTCGTTTT [SEQ ID:4] 5'VLB5RRV:AGGTGTACACTCCATATCCAGCTGACCCAGTCTCCA [SEQ ID:5] 3'VLGS15: GGAGCCGCCGCCGCCAGAACCACCACCTTTGATCTCGAGCTTGGTCCC [SEQ ID:6] 5'VHGS15:
GGCGGCGGCGGCTCCGGTGGTGGTGGTTCTCAGGTSNARCTGCAGSAGTCWGG [SEQ ID:7]
3'VHBspE1:AATCCGGAGGAGACGGTGACCGTGGTCCCTTGGCCCCAG [SEQ ID:8]
Для амплификации V-областей с помощью ПЦР использовали праймеры 5'L1 и 3'К, фланкирующие VL-область, и 5'Н1 и 3'G для тяжелой цепи, основой которых являются праймеры, описанные у Dubel и др. (24).
кДНК scFv-Фрагмента антитела к CD3 любезно предоставлена А. Traunecker (25).
Пример 2: Конструирование биспецифических одноцепочечных фрагментов и экспрессия в эукариотических организмах
Для получения scFv-фрагмента к CD19 соответствующие VL- и VH-области, клонированные в разных плазмидных векторах, служили в качестве матриц для VL- и VH-специфичной ПЦР с использованием пар олигонуклеотидных праймеров 5'VLB5RRV/3'VLGS15 и 5'VHGS15/3'VHBspEI соответственно. Таким образом, в ПЦР-продукты интродуцировали перекрывающиеся комплементарные последовательности, которые объединяли для получения кодирующей последовательности состоящего из 15 аминокислот (Gly4 Ser1)3-линкера в процессе последующих ПЦР-слияний. Эту стадию амплификации осуществляли с использованием пары праймеров 5'VLB5RRV/3'VHBsEI и полученный слитый продукт (или обычно scFv-фрагмент к CD19) расщепляли с помощью рестриктаз EcoRV и BspEI и после этого клонировали в векторе bluescript KS (фирмы Stratagene), который содержал либо (EcoRI/SalI-клонированную) кодирующую последовательность биспецифического одноцепочечного антитела к 17-1А/К CD3 с N-концевой FLAG-меткой (1) или его модифицированную версию без FLAG/эпитопа (21), тем самым заменяя анти-17-1A- на анти-CD-19-специфичность и сохраняя состоящий из 5 аминокислот (Gly4Ser1)1- линкер, соединенный с С-концом scFv-фрагмента к CD3 соответственно. Затем фрагменты ДНК, кодирующие обе версии биспецифического одноцепочечного антитела к CD19/K CD3 со следующим расположением областей: VLCD19-VHCD19-VHCD3-VLCD3, субклонировали с использованием EcoRI/SalI в описанном векторе экспрессии pEF-DHRF (1) соответственно. Полученными плазмидными ДНК трансфектировали путем электропорации СНО-клетки с дефицитом DHFR, отбор, амплификацию гена и получение протеина проводили, как описано ранее (1). В приведенных ниже примерах проиллюстрированы результаты, полученные с FLAG-содержащей версией bscCD19×CD3.
Очистка bscCD19×CD3 из супернатанта трансфектированных СНО-клеток позволяла получить 4 мг/л супернатанта культуры. Очищали bsc-Ат с использованием его С-концевого гистидинового "хвоста" с помощью аффинной хроматографии на Ni-NTA-колонке, как описано ранее (1). Элюировали bsc-Ат из Ni-NTA-колонки в виде отдельного пика при концентрации 200 нМ имидазола. Для анализа очистки bsc-Ат проводили ДСН-ПААГ согласно методу Laemmli (26) с использованием 12%-ного геля с последующим окрашиванием кумасси бриллиантовым голубым R250. Результаты ДСН-ПААГ-анализа (фиг.1) свидетельствуют об ожидаемом размере bsc-Ат (60 кДа).
Пример 3: Особенности связывания bscCD19×CD3
Особенности связывания bsc-Ат к CD3 и CD19 оценивали с помощью проточного цитометрического анализа на CD-позитивных клетках линии Jurkat, человеческих РВМС и нескольких различных CD19-позитивных линиях клеток В-клеточной лимфомы, включая Blin I, SKW6.4, Daudi, BJAB и Raji. CD19-Позитивные линии В-клеток Daudi, Raji, BJAB (лимфома Беркитта), SKW6.4 (человеческие В-клетки, трансформированные вирусом Эпштейна-Барра (EVB) и Blin I (пре-В-клеточная линия) использовали для проточного цитометрического анализа и в опытах по оценке высвобождения хрома. Jurkat представляет собой CD3-позитивную Т-клеточную линию; BL60 и плазмацитомные клеточные линии NCI и L363 представляют собой линии, негативные по обеим поверхностным молекулам CD3 и CD19. Клеточные линии культивировали в полной среде RPMI 1640 (фирма Biochron) с 10%-ной ФТС (фирма GIBCO).
1·106 Клеток промывали ЗФР, ресуспендировали в 200 мкл ЗФР с 10%-ным верниммуном (фирма Centeon, Marburg, Германия) и 0,1%-ным NaN3 и инкубировали в течение 30 мин при 4°С. После стадии центрифугирования (100×g, 5 мин) клетки инкубировали в 50 мкл bscCD19×CD3 (200 мкг/мл в ЗФР с 10%-ным верниммуном и 0,1%-ным NaN3) в течение 30 мин при 4°С. Клетки промывали дважды в ЗФР. Для выявления bsc-Ат использовали конъюгированное с ФИТЦ (флуоресцинизотиоцианат) антитело к His-метке (фирма Dianova). He обладающее такими же характеристиками bsc-Ат 17-1AxCD3, полученное в той же экспрессионной системе, что и bscCD19×CD3, или одно антитело к His-метке служили в качестве негативных контролей. Проточную цитометрию проводили с использованием клеточного сортера с возбуждением флуоресценции типа Becton Dickinson FACScan. He обнаружено никакого связывания с поверхностью клеток линии BL60, которые не экспрессировали ни CD19, ни CD3 (фиг.2).
Пример 4: Цитотоксическая активность bscCD19×CD3 в отношении CD19-позитивных клеток лимфомы
В опытах по высвобождению51Сr установлено, что антитело bscCD19×CD3 обладает высокой токсичностью в отношении нескольких клеточных линий лимфомы (фиг.3). Человеческие мононуклеары периферической крови (РВМС) в качестве клеток-эффекторов выделяли из свежих лейкоцитных пленок случайных доноров с использованием центрифугирования с помощью Lymphoprep™ (фирма Nycomed) с последующими стадиями центрифугирования при 100×g для удаления тромбоцитов. CD19-позитивные В-клетки элиминировали с помощью Dynabeads® V-450 CD19 (фирма Dynal). Уменьшенные (истощенные) популяции клеток анализировали с помощью проточной цитометрии (Becton Dickinson), которая показала 99%-ное истощение популяции CD19-позитивных клеток. РВМС инкубировали в течение ночи при 37°С, 5% СО2, в качестве клеток-мишеней использовали линии CD19-позитивных В-клеток (Raji, Blin I, Daudi, BJAB, SKW6.4). Цитотоксичность измеряли согласно стандартному опыту по высвобождению хрома в круглодонных 96-луночных планшетах (фирма Nunc), используя полную среду RPMI 1640 (фирма Biochrom) с 10%-ной ФТС (фирма GIBCO).
Нестимулированные РВМС добавляли в объеме 80 мкл среды в каждую лунку, содержащую 20 мкл различных концентраций bsc-Ат. Затем добавляли 100 мкл меченых с помощью51 Сr клеток-мишеней (1·104), планшеты центрифугировали в течение 3 мин при 100×g и инкубировали в течение 4 ч при 37°С, 5% СО2. После дополнительной стадии центрифугирования 50 мкл супернатанта удаляли и анализировали в отношении высвободившего51Сr в счетчике гамма-излучения (TopCount, Canberra Packard).
Спонтанное высвобождение оценивали путем инкубации клеток-мишеней без клеток-эффекторов или антител, и максимальное высвобождение определяли путем инкубации клеток-мишеней с 10%-ным Тритоном Х-100. Инкубация клеток-мишеней с bsc-Ат без клеток-эффекторов не привела к выявляемому лизису. Процент удельного лизиса рассчитывали как неспецифическое высвобождение (%)=[(cрm, высвобождение в условиях эксперимента)-(cpm, спонтанное высвобождение)]/[(сpm, максимальное высвобождение)-(сpm, спонтанное высвобождение)]·100. Все опыты проводили в трех повторностях. СКО (среднее квадратичное отклонение) по трем повторностям во всех экспериментах составляло менее 6%. Для моделирования условий in vivo в качестве клеток-эффекторов использовали нестимулированные РВМС из здоровых доноров. Без использования какого-либо протокола предварительной стимуляции Т-клеток наблюдали быструю индукцию цитотоксичности через 4 ч. Взятое в качестве контроля bsc-антитело с другой специфичностью в отношении опухолей (bscl7-1AxCD3), но полученное с помощью такой же системы, что и антитело bscCD19×CD3, проявило литическую активность, не существенно превышающую фоновый уровень среды. Кроме того, никакой цитотоксической активности не обнаружено при использовании в качестве клеток-мишеней плазмацитомных линий клеток NCI и L363, которые не экспрессируют CD19 (фиг.4). В конкурентных анализах с использованием возрастающих количеств CD-19-специфического родительского моноклонального антитела HD37 цитотоксическая активность bscCD19×CD3 может быть практически полностью заблокирована (фиг.5). Эти контрольные варианты показали, что опосредованные bscCD19×CD3 цитотоксические действия являются антигеспецифическими. Для получения дополнительной информации о молекулярных механизмах нейтрализации с использованием bscCD19×CD3 CD19-позитивных клеток мишеней была предпринята попытка блокировать опосредованную bscCD19×CD3 цитотоксичность с помощью ЭГТК. Как видно из фиг.6, цитотоксическая активность bscCD19×CD3 может быть полностью заблокирована при использовании ЭГТК, что свидетельствует о том, что специфический лизис более вероятно представляет собой опосредованное Т-клетками действие (вероятно, через путь метаболизма перфорина), а не прямое (т.е. индуцирующее апоптоз) действие самого антитела. С использованием нестимулированных Т-клеток даже при использовании концентраций антитела ниже 1 нг/мл наблюдалось выраженное цитотоксическое действие в отношении клеток линии Blin I (фиг.7). Даже при относительно низком соотношении Е:Т (5:1; 2,5:1) и очень низких концентрациях антитела 10-100 пг/мл bscCD19×CD3-антитело могло быстро индуцировать специфическую цитотоксическую активность в отношении нестимулированных Т-клеток (фиг.7). В противоположность этому обычное биспецифическое антитело CD19×CD3, полученное с помощью гибридо-гибридомного метода (5-7, 27), в этих условиях не проявило заметной цитотоксической активности даже при концентрации до 3000 нг/мл (фиг.7). Это обычное биспецифическое антитело нуждалось в дополнительной предварительной стимуляции Т-клеток в высоких концентрациях антитела вплоть до примерно 100 нг/мл для того, чтобы индуцировать специфическую цитотоксичность в отношении Т-клеток (данные не приведены), что согласуется с литературными данными (5-7, 27).
Пример 5: Элиминация первичных (злокачественных) В-клеток с помощью аутологичных Т-клеток в результате цитотоксической активности bscCD19×CD3
Для того чтобы оценить цитотоксическую активность bscCD19×CD3 в отношении первичных злокачественных В-клеток, мононуклеары периферической крови (РВМС) пациентов, страдающих B-CLL (обусловленный В-клетками хронический лимфатический лейкоз), выделяли с помощью центрифугирования в градиенте плотности с использованием Фиколла. Эти клетки затем культивировали в присутствии bscCD19×CD3 в течение 5 дней при 37°С/5% СО2 в среде RPMI 1640, дополненной 10%-ной ФТС, необязательно с 60 ед./мл IL-2, или культивировали без bscCD19×CD3. Проточный цитометрический анализ показал, что PBL (лимфоциты периферической крови) этой конкретной страдающей не-ходжкинской лимфомой (NHL) пациентки (которой затем системно вводили bscCD19×CD3; см. пример 7) содержали 92,6% CD19-позитивных В-клеток (т.е. клеток-мишеней) и 7,4% СD3-позитивных Т-лимфоцитов (т.е. клеток-эффекторов) при соотношении CD4/CD8 Т-клеток 2,6 : 4,8. Основное большинство этих CD19-позитивных В-клеток представляло собой злокачественные клетки. Высевали 3·106 PBL/мл на лунку, в объем среды 1 мл, на 24-луночный планшет для культуры ткани. В качестве негативных контролей использовали культуральную среду плюс IL-2 и культуральную среду плюс IL-2 с не обладающим такими же характеристиками биспецифическим одноцепочечным антителом bscl7-1AxCD3 (1) в концентрации 0,5 мкг/мл. Как видно из фиг.9, никакого истощения CD19-позитивных клеток не было обнаружено в этих условиях при инкубации в течение 5 дней. Однако когда добавляли bscCD19×CD3 в концентрации 0,5 мкг/мл или 0,05 мкг/мл (как в присутствии 11-2, так и без него), почти все CD 19-позитивные В-клетки были уничтожены. Культивируемые клетки в это время состояли в основном из Т-лимфоцитов при соотношении CD4/CD8 Т-клеток примерно от 1:2 до 1:3. Этот факт демонстрирует исключительную направленность цитотоксического действия bscCD19×CD3 в отношении CD19-позитивных В-клеток, поскольку общее истощение первичных В-клеток аутологичными Т-клетками может быть индуцировано только при концентрации 50 нг/мл при чрезвычайно нежелательном начальном соотношении клеток-эффекторов и клеток-мишеней ниже, чем 1:10, даже без IL2 или другого типа дополнительной Т-клеточной стимуляции.
Пример 6: Очистка bscCD19×CD3 для терапевтических целей
BscCD19×CD3 получали из клеток яичника китайского хомячка (СНО), стабильно трансфпектированных экспрессионным вектором (pEF-DHFR; см. пример 2), кодирующим bscCD19×CD3, и дополнительно гексагистидиновую метку и FLAG-метку. Клетки выращивали в бессывороточной среде (фирма Rencyte) в ферментере с системой полых волокон (фирма Unisyn). 500 мл Супернатанта клеточной культуры собирали и стерилизовали фильтрацией через фильтры с размером пор 0,2 мкм (типа AcroCap; Pall Gelman).
Выявляли bscCD19×CD3 и оценивали количественно Вестрен-блоттингом с использованием мышиного IgG к FLAG (фирма Sigma) и козьего антитела к IgG мыши, конъюгированного с щелочной фосфатазой (фирма Sigma). Обнаружение проводили хемолюминисцентным методом с использованием системы BCIP/NBT (фирма Devitron). Концентрации протеина определяли с помощью метода Брадфорда (фирма Biorad), используя бычий IgG (фирма Biorad) в качестве стандарта протеина. Чистоту колоночной фракции определяли с помощью восстанавливающего электрофореза в 4-12%-ном градиенте полиакриламидного геля (ПААГ) с додецилсульфатом натрия (ДСН), бис/трис, применяя систему MOPS-буфера (фирма Novex).
Для очистки bscCD19×CD3 до гомогенного состояния была необходима катионообменная хроматография, кобальтовая хелатная аффинная хроматография и на конечной стадии гель-фильтрация. Эти стадии очистки проводили с использованием стандартных протоколов (см. ниже). Схема процесса очистки приведена на фиг.10.
Катионообменная хроматография: супернатант клеточной культуры, полученный при использовании СНО-клеток, смешивали с двумя объемами буфера С (30 мМ морфолинэтансульфоновая кислота [MES], 20 мМ NaCl, 3 мМ ЭДТК, 0,3 мМ гидрохлорид бензамидина, рН 5,5) и пропускали через 70-миллиметровую катионообменную сефарозную колонку типа SP Sepharose Fast Flow (фирма Pharmacia) со скоростью потока 20 мл/мин. Колонку уравновешивали буфером А (20 мМ MES, 20 мМ NaCl, рН 5,8). После промывки буфером А, взятом в объеме, составляющем 5 объемов колонки, bscCD19×CD3 элюировали с помощью ступенчатого градиента 45%-ного буфера В (20 мМ MES, 1M NaCl, рН 5,8) в буфере А. Элюат получали, используя 0,046 объема 1M трис/НС1-буфера, рН 8,5, содержащего 47 мМ имидазол, и затем подвергали стерилизации фильтрацией (0,2 мкм; тип AcroCap). Типичный профиль элюции, полученной при использовании катионообменной хроматографии, приведен на фиг.12. BscCD19×CD3 входило во фракцию 6.
Очистка кобальтовой хелатной аффинной хроматографией: элюат, полученный с катионообменной колонки, пропускали при скорости потока 2,5 мл/мин через 10-миллиметровую сефарозную хелатирующую колонку типа Chelating Sepharose Fast Flow (фирма Pharmacia), уравновешенную буфером ОА (50 мМ Na2HPO4, 400 мМ NaCl, pH 8,0). Колонка была предварительно уравновешена 0,1М раствором хлорида кобальта. После промывки объемом, соответствующим 33 объемам колонки, буфера ОА, буфера А (50 мМ Na2HPO4, 400 мМ NaCl, 500 мМ имидазол, pH 6,4) и градиента от 0 до 12% буфера В (50 мМ Nа2НРO4, 400 мМ NaCl, 500 мМ имидазол, pH 6,4) в буфере А, bscCD19×CD3 элюировали в одно стадию с помощью 30 мл 100%-ного буфера В. Элюат подвергали стерилизации фильтрацией с последующим примерно 10-кратным концентрированием в устройстве типа MacroSep (фирма Pall Gelman; предел пропускающей способности 10 кДа). Типичный профиль элюирования для кобальтовой хелатной аффинной хроматографии приведен на фиг.13. BscCD19xCD3 было обнаружено в 7 фракции.
Гель-фильтрация: концентрированный элюат, полученный из колонки для кобальтовой хелатной аффинной хроматографии, загружали со скоростью потока 0,75 мл/мин на 124-миллиметровую колонку типа High Load Superdex 200 (фирма Pharmacia; чистота "prep"), уравновешенную забуференным фосфатом физиологическим раствором (фирма Gibco). BscCD19×CD3 элюировалось во фракции с молекулярной массой, соответствующей примерно 55 кДа (фиг.14, фракция №7). Полученную в результате гель-фильтрации фракцию, содержащую bscCD19×CD3, дополняли 5%-ным человеческим сывороточным альбумином (фирма Behring) с последующей стерилизацией фильтрацией через фильтр с размером пор 0,1 мкм (фирма Millex, тип Millipore).
Количество bscCD19×CD3 в супернатанте клеточной культуры и различные активные колоночные фракции по данным ДСН-ПААГ-анализа приведены на фиг 11. BscCD19×CD3 представлял собой основную протеиновую полосу в супернатантах клеточной культуры (полоса 2). В антителе к CD19×CD3 с высокой степенью очистки, которое можно применять для терапии человека, не обнаружено никаких выявляемых примесей (фиг.11, полоса 5).
Пример 7: Клиническое применение bscCD19×CD3 для лечения пациентки с В-клеточной лимфомой
Из сострадания пациентку (А-В, женщина 1937 г. р.), страдающую обусловленным В-клетками хроническим лимфатическим лейкозом, лечили биспецифическим одноцепочечным антителом bscCD19×CD3 в щадящем режиме.
Анамнез пациентки:
У пациентки B-CLL был обнаружен в 1992 г. В момент первоначального диагноза болезнью были поражены различные области лимфатических узлов и селезенка; кроме того, были обнаружены гемолитическая анемия аутоиммунного происхождения и дефицит иммуноглобулина. У пациентки были узлы, которые легко контролировались на фоне нормального функционирования щитовидной железы путем обработки карбимазолом (2,5 мг/день).
Пациентка прошла несколько циклов химиотерапии хлорамбуцилом и преднизоном в течение 1992-1994 гг. Из-за прогрессивного развития болезни лечение было изменено и были назначены циклофосфамид, доксорубицин, винкристин и преднизон (CHOP, 8 циклов) и в течение более года наблюдалась стадия ремиссии. После нового рецидива было проведено еще 6 циклов CHOP, с последующим лечением хлорамбуцилом и преднизоном и один курс лечения одним хлорамбуцилом, которые не привели ни к какому улучшению состояния. В декабре 1998 г. для того, чтобы контролировать прогрессирующую спленомегалию у пациентки, было проведено облучение селезенки. У пациентки обнаружено выраженное подавление функции костного мозга с многочисленными инфекционными осложнениями. Из-за анемии и тромбоцитопении требовались частые трансфузии эритроцитов и замещение тромбоцитов.
Из-за наличия развитой стадии болезни и поражения функции костного мозга применение более сильнодействующей химиотерапии с использованием высоких доз не было показано для этой пациентки. Лечение с использованием антитела к CD20 ритуксимаба не применялось, поскольку к настоящему времени отсутствуют данные об эффективности ритуксимаба в отношении B-CLL.
FACS-анализ показал, что 95% клеток периферической крови пациентки представляли собой CD19-позитивные клетки и 77% клеток экспрессировали антиген CD20. Инкубация клеток периферической крови пациентки с bscCD19×CD3 привела к выраженному уменьшению CD19-позитивных В-клеток (см. пример 5). После этого лечащие врачи решили подвергнуть пациентку лечению новым bscCD19×CD3 в щадящем режиме. Пациентка была проинформирована в деталях о новизне соединения и о потенциальном риске и возможной пользе такого лечения. Она полностью поняла объяснения и дала письменное согласие на такое лечение в щадящем режиме.
Описание клинического применения
Перед началом лечения пациентку подвергли клиническому обследованию и обширным диагностическим процедурам для подтверждения степени болезни и исключения каких-либо дополнительных факторов риска. У пациентки наблюдалось четкое клиническое состояние с явлениями анемии, тромбоцитопении и потери веса, но не обнаружено какого-либо сердечно-сосудистого нарушения и других осложнений, препятствующих применению bscCD19×CD3. В течение ночи перед первым днем лечения пациентка страдала от связанной с мигренью головной боли. Для введения bscCD19×CD3 пациентка была помещена в больничную палату, где она находилась под постоянным внимательным присмотром для того, чтобы гарантировать немедленное лечение любого непредвиденного случая, который может произойти. Для предупреждения каких-либо обусловленных цитокинами острых реакций и осложнений при лизисе опухоли пациентка получила профилактические IV дозы по 2 мг клемастина (Tavegil®) и 200 мг циметидина (Tagamet®), а также 300 мг аллопуринола и 20 мг омепразола (Antra®).
Во время лечения и во время периода реабилитации проводили алкализацию и гепаринизацию. Кроме того, пациентка получала все необходимое симптоматическое лечение.
Образцы крови брали до и во время периода лечения лекарственным средством с определением биохимических, гематологических и иммунологических параметров.
Первая обработка bscCD19×CD3 (14 апреля 1999 г.)
Пациентка получила первую дозу, включающую 3 мкг bscCD19×CD3, в виде 20-минутной инфузии в изотоническом фосфатом буфере, содержащем 5%-ный человеческий сывороточный альбумин (ЧСА). В процессе инфузии у пациентки не было обнаружено никаких отрицательных реакций. Примерно через 1 ч после инфузии у пациентки был озноб примерно в течение 5 мин с последующей испариной, умеренное понижение кровяного давления примерно на 10 мм рт.ст. и умеренное повышение температуры тела (+0,5°С) в течение нескольких часов. Кроме того, ее головная боль несколько усилилась. Пациентка получила еще 2 мг Tavegil® и 200 мг Tagamet®, 250 мг преднизолона (Solu-Decortin® ) и 50 мг петидина (Dolantin®). Все симптомы исчезли без последствий в этот же день.
Вторая обработка bscCD19×CD3 (15 апреля 1999 г.)
Вторая обработка 10 мкг bscCD19×CD3 была проведена через день в таких же условиях. Примерно через 1 ч после инфузии у пациентки наблюдался выраженный озноб, лихорадка (39,2°С), небольшая гипервентиляция легких и признаки гипотензии. Пациентка получила 2 мг Tavegil, 200 мг Tagamet и 300 мг Solu-Decortin и 15 мг пиритрамида (Dipidolor®). Для стабилизации сердечно-сосудистой функции пациентке была сделана инфузия допамина и изменен объем. После такого лечения симптомы значительно уменьшились. Несмотря на это пациентка была переведена на ночь в кардиологическое отделение, где гарантированно соответствующим образом контролировались параметры жизненно важных функций и было гарантировано немедленное вмешательство в случае их нарушения. На следующее утро пациентка была переведена в обычную палату без каких-либо дополнительных осложнений.
В течение трех последующих дней у пациентки сохранялась субфибрильная температура (примерно 37,2°С) и развился небольшой плевральный выпот через день после обработки второй дозой (16 апреля 1999 г.) и умеренный отек нижних конечностей (18 апреля 1999 г.). Сердечно-сосудистая функция осталось стабильной и лабораторные оценки показали отсутствие каких-либо заметных опасных изменений, за исключением повышения уровня γ-глутамилтрансферазы после второй обработки bscCD19×CD3 (фиг.15).
Поскольку лечение bscCD19×CD3 хорошо переносилось пациенткой и побочные воздействия могли контролироваться симптоматическим лечением, лечение пациентки новым bscCD19×CD3 по изобретению было продолжено.
Клиническая и иммунологическая эффективность bscCD19×CD3
Результаты клинического опыта
Ультразвуковое обследование селезенки и пяти брюшных и подмышечных лимфатических узлов проводили через 1 день и через 4 дня после введения второй дозы bscCD19×CD3. Уже через 1 день после введения дозы 10 мкг (16 апреля 1999 г.) обнаружено уменьшение размера лимфатических узлов, а также селезенки примерно на 20% по сравнению с исходным состоянием. Это наблюдение было подтверждено при втором ультразвуковом обследовании 19 апреля 1999 г. Вес селезенки уменьшился на 350 г (с 1630 г в исходном состоянии до 1280 г 19 апреля 1999 г.) (таблица 1; фиг.16).
Результаты гематологического обследования:
Количество лейкоцитов, включая главным образом злокачественные В-клетки, уменьшилось в процессе лечения и в последующие дни (таблица 2; фиг.17). С-реактивный протеин (CRP) представляет собой протеин острой фазы реакции, который отражает Т-клеточную активацию и воздействие противоспалительных цитокинов. Его уровень заметно повысился после введения 10 мкг bscCD19×CD3, а затем непрерывно уменьшался в течение 3 последующих дней наблюдения (таблица 2; фиг.18).
Результаты иммунологического обследования
Уровень сывороточных цитокинов, который отражает острый иммунологический ответ на введение соединения, измеряли до и через различные периоды времени после введения нового соединения по изобретению. Уровни в сыворотке цитокинов и растворимого рецептора IL-2 оценивали количественно с помощью анализа ELISA согласно инструкциям производителя.
Уровень фактора некроза опухоли TNF-α заметно повысился в зависимости от дозы в первый час после введения bscCD19×CD3 (фиг.19).
Также обнаружено заметное и зависящее от дозы повышение уровня интерлейкина 6 (IL-6) и интерлейкина 8 (IL-8). Их максимальные уровни обнаружены через 2-4 ч после введения bscCD19×CD3 (фиг.20, 21). Через несколько часов концентрации всех цитокинов вернулись к исходному уровню.
Уровень растворимого рецептора IL-2 был повышен уже в исходном состоянии, что может быть объяснено большим количеством злокачественных В-клеток, экспрессирующих рецептор IL-2. После введения нового bscCD19×CD3 наблюдали повышение уровня растворимого рецептора IL-2, что свидетельствует об активации эффекторных клеток (фиг.22).
Заключение
Введение нового bscCD19×CD3 безопасно для пациентки, страдающей рефракторным B-CLL. Переносимость bscCD19×CD3 в дозах 3 мкг и 10 мкг оказалась удовлетворительной и могла контролироваться профилактическими мероприятиями и симптоматическим лечением.
Применение нового bscCD19×CD3 вызвало уменьшение у пациентки ранее увеличенной селезенки и лимфатических узлов, что подтверждено ультразвуковым обследованием. Поскольку увеличение селезенки и лимфатических узлов вызвано инфильтрацией злокачественных В-клеток, то уменьшение размеров отражает разрушение злокачественных В-клеток в результате введения bscCD19×CD3.
В явном отличие от любых других биспецифических антител CD19×CD3, известных в данной области, биспецифическое антитело CD19×CD3 по изобретению (bscCD19×CD3) проявляет клиническую эффективность в отношении обусловленной В-клетками не-ходжкинской лимфомы, что видно по уменьшению лимфоидных органов, инфильтрованных злокачественными В-клетками. Очень важно, что для bscCD19×CD3 доказана клиническая эффективность в неожиданно низких дозах, которые хорошо переносимы при системном введении. Таким образом, клиническая эффективность bscCD19×CD3 подтверждает его чрезвычайно высокую цитотоксическую активность, определенную in vitro.
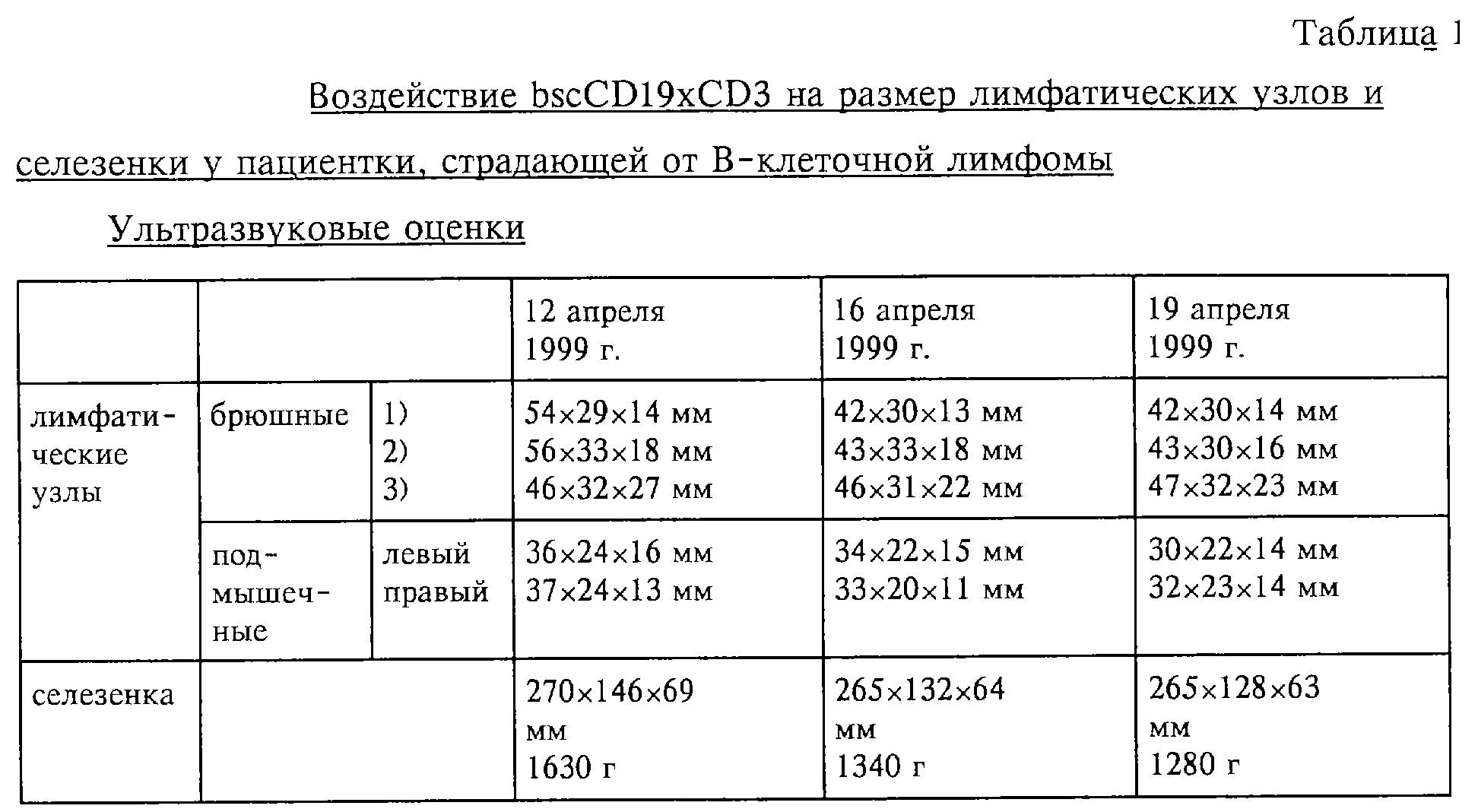
Были определены размеры трех брюшных лимфатических узлов, одного левого и одного правого подмышечного лимфатических узлов и селезенки с помощью сонографии с использованием устройства типа Toshiba SSA100.
Размеры оценивали в трех направлениях в мм. Вес селезенки рассчитывали на основе данных о ее размерах и ультразвуковой плотности.

Уровни в крови гамма-глутамилтрансферазы (GGT), лактат-дегидрогеназы (LDH) и С-реактивного протеина (CRP) определяли стандартными методами клинической биохимии и выражали в виде ед./мл (GGT), ед./л (LDH) и мг/дл (CRP). Количество лейкоцитов выражали в виде Гч./л, а количество лимфоцитов в виде процента от общего количества лейкоцитов. В первой колонке представлены данные об исходных уровнях на 14 апреля 1999 г. до начала лечения. Во второй колонке приведены данные о реакции на обработку 3 мкг bscCD19×CD3 (которая была проведена 14 апреля 1999 г.), полученные 15 апреля 1999 г. Данные о реакции на вторую обработку 10 мкг соединения, полученные в этот же день, приведены в третьей колонке. Уровни в крови селектрируемых маркеров на четвертый день после обработки лекарственным средством приведены в четырех последних колонках.
Список литературы
1. Mack, Proc. Natl. Acad. Sci. USA 92 (1995), 7021-5.
2. Gianni, N Engl. J. Med. 336 (1997), 1290-7.
3. Urba, J. Natl. Cancer Inst. Monogr. (1990), 29-37.
4. Fisher, Cancer (1994).
5. Bohlen, Blood 82 (1993), 1803-121.
6. Bohlen, Cancer Res 53 (1993), 18:4310-4.
7. Bohlen, Cancer Res 57 (1997), 1704-9.
8. Haagen, Clin Exp Immunol 90 (1992), 368-75.
9. Haagen, Cancer Immunol Immunother. 39 (1994), 391-6.
10. Haagen, Blood 84 (1994), 556-63.
11. Haagen, Blood 85 (1995), 3208-12.
12. Weiner, Leuk Lymphoma 16 (1995), 199-207.
13. Csoka, Leukemia 10 (1996), 1765-72.
14. Uckun, Proc. Natl. Acad. Sci. USA 85 (1988), 8603-7.
15. Staerz, Proc. Natl. Acad. Sci. USA 83 (1986), 1453-7.
16. Lanzavecchia, EurJ Immunol 17 (1987), 105-11.
17. Mallender, J Biol Chem 269 (1994), 199-206.
18. Gruber, J Immunol 152 (1994), 5368-74.
19. Kostelny, J Immunol 148 (1992), 1547-53.
20. Mack, J Immunol 158 (1997), 3965-70.
21. Kufer, Cancer Immunol Immunother 45 (1997), 193-7.
22. Pezzutto, J Immunol 138 (1987), 2793-9.
23. Orlandi, Proc. Natl. Acad. Sci. USA 86 (1989), 3833-7.
24. Dubel, J Immunol Methods 175 (1994), 89-95.
25. Traunecker, Embo J 10 (1991), 3655-9.
26. Laemmli, Nature 227 (1970), 680-5.
27. Bohlen, J Immunol Methods 173 (1994), 55-62.
28. Demanet, Int J Cancer Suppl 7 (1992), 67-8.
29. De, J Hematother 4 (1995), 433-7.
30. Haagen, Leuk Lymphoma 19 (1995), 381-93.
31. Anderson, Blood 80 (1992), 2826-34.
32. Zhu, Int J Cancer 62 (1995), 319-24.
33. Hartmann, Blood 89(1997), 2042-7.
34. Valone, J Clin Oncol 13 (1995), 2281-92.
35. Valone, J Hematother 4 (1995), 471-5.
36. Bolhuis, Int J Cancer Suppi 7 (1992), 78-81.
37. Canevari, J Natl Cancer Inst 87 (1995), 1463-9.
38. Nitta, Lancet 335 (1990), 368-71.
39. Yokota, Cancer Res 52 (1992), 3402-8.
40. Weiner, J Immunol 152 (1994), 2385-92.
41. Maloney, Blood 84 (1994), 2457-66.
42. Reff, Blood 83 (1994), 435-45.
43. Kipriyanov, Int. J. Cancer 77 (1998), 763-772.







Реферат
Изобретение относится к медицинской промышленности и касается полипептидов, специфичных в отношении CD19 и CD3, и их применения. Изобретение включает одноцепочечный многофункциональный полипептид, специфичный в отношении С19 и CD3 антигенов формулы: VLCD19 - VHCD19 - VHCD3 - VLCDS, полинуклеотид, кодирующий этот полипептид, экспрессирующий вектор, штамм культуры клеток, способ получения полипептида, фармацевтическую композицию для лечения злокачественных заболеваний В-клеток, диагностическую композицию, способ выявления активаторов или ингибиторов Т-клеточной активации или стимуляции, способ приготовления фармацевтической композиции, предназначенной для лечения злокачественных заболеваний В-клеток, способ лечения злокачественных заболеваний В-клеток или истощения В-клеток и способ замедления развития патологического состояния. Преимущество изобретения заключается в разработке методов, пригодных для лечения опосредованных В-клетками болезней. 11 с. и 17 з.п. ф-лы, 22 ил., 2 табл.
Комментарии