Днк-фрагмент, кодирующий нейротропный фактор мозга (bdnf), белок bdnf и способ его получения - RU2131926C1
Код документа: RU2131926C1
Чертежи




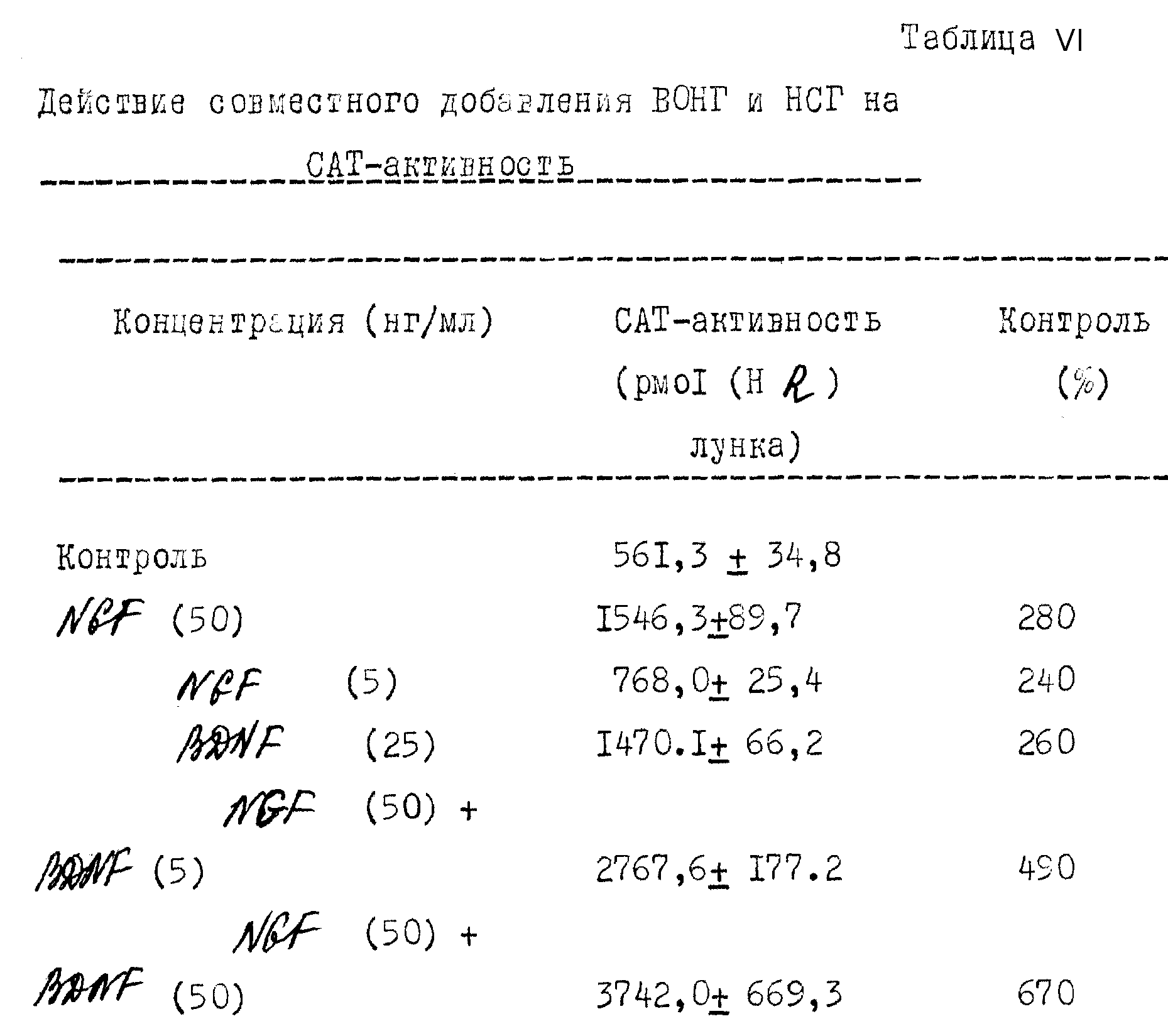
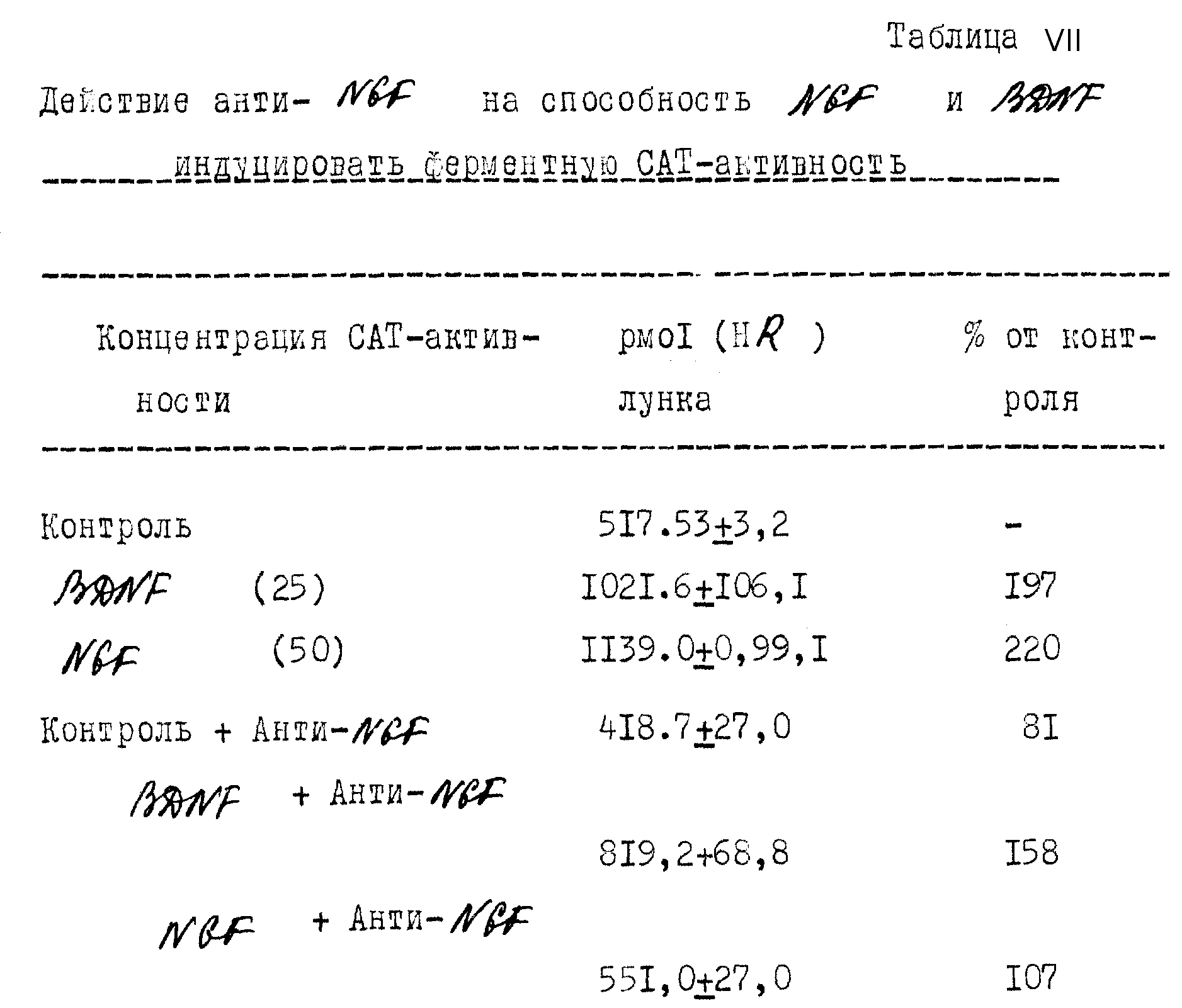



































Описание
1. Введение.
Изобретение касается нуклеиновых последовательностей, кодирующих мозговой нейротропный фактор (BDNF), в основном чистого протеина, пептидных фрагментов или производных, полученных от него в больших количествах, а также к антителам, направленным против BDNF-протеина, пептидных фрагментов или производных. Дополнительно изобретение касается генов, которые являются членами вновь определенного генного семейства BDNF/NGF, и продуктов этих генов. Изобретение также относится к фармацевтическим композициям, включающим эффективные количества BDNF генных продуктов или альтернативно антителам, направленным против BDNF генных продуктов, и к способам диагностики и лечения множества неврологических заболеваний и расстройств, включая болезнь Альцхаймера и болезнь Паркинсона, а также, в частности, BDNF генные продукты по изобретению ценны в диагностике и терапии расстройств допаминергических нейронов, таких как болезнь Паркинсона, а также нарушений сенсорных нейронов и дегенеративных заболеваний сетчатки.
2. Предпосылки изобретения.
2.1 Гибель нейронных клеток и роль нейротропных факторов в развитии нервной системы.
В начальный период развития во многих частях нервной системы позвоночных присутствует гораздо больше нейронов, чем обнаруживают у взрослых животных. Периоды раннего развития характеризуются волнами естественно происходящей гибели нейронных клеток (Carr и Simpson, 1978, J. Comp. Neurol., 182: 727 - 740; Cohan и др., 1984, Science 225: 1258 - 1265). Выживание, дифференциация и созревание развивающихся нейронов может регулироваться скорее факторами окружающей среды или "эпигенетическими" факторами, чем строгой наследственной генетической программой. Например, экспериментальные манипулирования куриными эмбрионами показали, что трансплантация или экстирпация периферийных "мишенных полей", таких как зачаток конечности или глаз, на ранних стадиях развития цыпленка могут привести к соответствующему увеличению или снижению соответственно количества сенсорных, симпатических или двигательных нейронов, смежных увеличенному или уменьшенному мишенному полю (Hamburger, 1934, J. Exp. Zool 68, 449; Hollyday и Hamburger, 1976, J. Comp. Neurol 170: 311 - 321; Landmesser и Pilar, 1976, J. Cell Biol., 68: 357 - 374). Мишенное поле может поддерживать только ограниченное количество нейронов, и нормальной частью процесса развития может быть отбор лишнего количества нейронов, чтобы приспособить к нему "нейтротропную" мощность мишенной ткани. Открытие и выделение протеина, названного теперь фактором нервного роста (NGF), привело к молекулярной гипотезе о том, как мишень может быть способна к регулированию количества нейронов, которые выживают и иннервируют эту ткань (Levi-Montalcini и др. , 1968, Physiol. Rev., 48: 524 - 569; Thoenen и Barde, 1980, Physiol. Rev., 60: 1284 - 1335).
Теперь хорошо установлено, по меньшей мере в периферийной нервной системе, что нейронные мишенные ткани синтезируют и освобождают ограниченные количества различных нейротропных молекул, которые являются критическими для выживания специфических типов нейронов (Korsching и Thoenen, 1983, Proc. Nath. Acad. Sci, США, 80: 3513 - 3516; Neumann и др., 1984, EMBO, 3: 3183 - 3189; Shelton и Reichardt, 1984, Proc. Nath. Acad. Sci, США, 81: 7051 - 7955; Korsching и Thoenen, 1985, Neurosci, Lett. 54: 201 - 205).
2.2 Фактор нервного роста.
Фактор нервного роста является пока наиболее полно охарактеризованным из этих нейротропных молекул, и как "ин витро", так и "ин виво" было показано, что он необходим для выживания симпатических и происходящих от неврального гребня сенсорных нейронов на стадии раннего развития как цыплят, так и крыс (Levi-Montalcini и Angeletti, 1963, Develop. Biol. 7: 653 - 659; Levi-Montalcini и др., 1968, Physiol. Rev. 48: 524 - 569). Инъекции очищенного NGF в развивающиеся куриные эмбрионы обнаружили, что NGF вызывает массивную гиперплазию и гипертрофию сенсорных нейронов спинного мозга и симпатических нейронов (Levi-Montalcini и Booker, 1960, Proc. Nath. Acad. Sci, США, 46: 373 - 384; Hamburger и др., 1981, J. Neurosci 1: 60 - 71). Наоборот, удаление или разрушение эндогенного NGF путем ежедневных инъекций антиNGF-антител неонатальным крысам было связано в итоге с деструкцией симпатической нервной системы (Levi-Montalcini и Booker, 1960, Proc.Nath.Acad.Sci. США, 46: 384-391; Levi-Montalcini и Angeletti, 1966, Pharmacol.Rev. 18: 619-628). Применение NGF антител даже наиболее ранней стадии развития либо путем внутриматочных инъекций антител или путем пассивного трансплацентарного переноса материнских антител привело к существенной потере происходящих от неврального гребня сенсорных нейронов, таких как спинальные и дорсомедиальные тройничные сенсорные нейроны (Goedert и др., 1984, Proc.Natl. Acad. Sci. США 81: 1580-1584; Gorin и Johnson, 1979, Proc.Nath.Acad.Sci. США, 76: 5382-5386). До недавнего времени почти все исследования NGF фокусировались на его роли в периферийной нервной системе, но теперь очевидно, что NGF также влияет на развитие и поддержание специфических популяций нейронов в центральной нервной системе (Thoenen и др., 1987, Rev.Physiol.Biochem.Pharmacol. 109: 145 - 178; Whittmore и Senger, 1987, Brain Res. Rev. 12: 439 - 464).
Удачное открытие больших количеств протеина NGF в подчелюстной железе взрослого самца мыши (Cohen, 1960, Proc.Nath.Acad.Sci. США 46: 302 - 311) и еще более раннее открытие высоких уровней NGF в змеином яде (Cohen и Levi-Montalcini, 1956, Proc. Nath.Acad.Sci. США 42: 571 - 574) дало достаточные количества NGF для проведения исследований, касающихся физиологии, химии белков и позднее молекулярной биологии NGF (т.е. для молекулярного клонирования NGF и его рецептора). Функция больших количеств NGF в подчелюстной железе взрослого самца крысы остается неясной; однако понятно, что такой богатый источник NGF не может играть какую-либо роль в развитии и поддержании периферийной или центральной нервной системы. В мишенных тканях, иннервированных чувствительными к NGF нейронами (это нейроны, для которых было установлено, что они зависят от NGF для их выживания, для обладания высокой аффинности NGF-рецепторами для интернализации и ретроградной транспортировки NGF с высокой специфичностью), устойчивое состояние измеримых уровней NGF было крайне низким, в районе пикограммов или нанограммов на грамм тканей, по сравнению с тысячекратно более высокими уровнями в ткани подчелюстной слюнной железы взрослого самца крысы. NGF не был обнаружен на каком-либо ощутимом уровне в сыворотке крови и поэтому, очевидно, не является циркулирующим фактором роста или гормоном (Suda и др., 1978, Proc.Nath.Acad.Sci. США 75: 4042 - 4046).
В дополнение к важному открытию основного источника NGF-протеина в мышиной подчелюстной железе развитие чувствительных, надежных и эффективных биологических экспериментов сыграло основную роль в объяснении биологии и биохимии NGF. Ганглии дорсального корня (DRG) развивающегося куриного эмбриона были одними из самых первых нейронных типов, у которых была обнаружена способность реагировать на NGF "ин витро". Культуры эксплантантов E8 - E12 DRG цыпленка в сгустке плазмы и, позднее, обогащенные нейронами культуры DRG цыпленка оказались очень полезными в биоэкспериментах на NGF-активность (например, во время его очистки) и для исследований "ин витро" биологии NGF (Levi-Montalcini и др., 1954, Cancer Res. 4: 49 - 57; Levi-Montalcini и Angeletti, 1963, Develop-Biol 1: 653 - 659; Greene, 1977, Develop.Biol. 58: 96, там же, 58: 106). Тот факт, что путем диссекции может быть получено более 40 DRG из единственного куриного эмбриона привел к широкому распространению использования NGF-биоэкспериментов во многих лабораториях.
В дополнение к доступности больших количеств NGF-протеина и эффективным системам опытов на NGF третьим основным фактором, который сделал важный вклад в наше понимание биологии NGF, является относительная легкость, с которой антитела к NGF могут быть получены от гвинейских свиней, кроликов, овец и т. д.; представляется, что мышиный NGF является высокоиммуногенным. Cohen (1960, Proc.Nath. Acad.Sci. США 46: 302 - 311) получал антитела к NGF, выделявшимся им из подчелюстной железы мыши, и показал вместе с Леви-Монтальчини и Букером (1960, Proc.Nath.Acad.Sci. США, 46: 384 - 391), что эти антитела вызывают деструкцию симпатических ганглиев или "иммуносимпатектомию" при ежедневном введении их новорожденным крысам (Levi-Montalcini и Angeletti, 1966, Pharmacol.Rev. 18: 619 - 628).
Обилие NGF-протеина позволяет определить первичную последовательность сравнительно традиционной белковой химией (Angeletti и Bradshaw, 1971, Proc. Nath.Acad.Sci. США, 68: 2417 - 2420). Ген NGF был клонирован от многих видов животных, включая мышь (Scott и др., 1983, Nature, 302 538 - 540), человека (Ullrich и др., 1983, Nature 303-821-825), корову и цыпленка (Meier и др., 1986, EMBO 5: 1489 - 1493) и крысу (Whittmore и др., 1988, J. Neurosci.Res. 20-402-410) с использованием в основном традиционной молекулярной биологии, основанной на доступности белковой последовательности мышиного NGF для создания подходящих олигонуклеотидных зондов. Доступность NGF также значительно облегчила исследования NGF рецептора, что в конечном счете привело к молекулярному клонированию NGF рецепторов человека и крысы (Johnson и др., 1986, Cell, 47, 545-554; Radeke и др., 1987, Nature 325: 593-597).
Теперь хорошо известно, что NGF не является везде присутствующим нейротропным фактором. В периферийной нервной системе NGF, вероятно, не является фактором выживания парасимпатических нейронов, происходящих от невральной плакоды сенсорных нейронов или кишечных нейронов, как определено в исследованиях "ин витро" и "ин виво". Более того, NGF, вероятно, не является фактором выживания для развивающихся двигательных нейронов (Oppenheim, 1982, J. Comp. Nunol 210: 174 - 189), хотя эти нейроны экспрессируют по меньшей мере слабоаффинную форму NGF-рецептора во время своего развития (Raivich и др., 1985, EMBO, 4: 637 - 644). Отсутствие действия NGF на эти типы нейронов побудило к поиску других нейротропных факторов, особенно факторов, которые бы поддерживали выживание двигательных нейронов спинного мозга и/или парасимпатических нейронов силиарного ганглия.
2.3. Другие нейротропные факторы.
В последние десятилетия были многочисленные сообщения о нейротропной активности в экстрактах самых разных тканей и в кондиционированной культурирующей среде многих различных клеточных типов. Почти во всех случаях, однако, прогресс в очистке и характеризирования этих активностей был затруднен тем, что такие активности присутствуют в крайне малых количествах, в пределах от пико- до нанограммов на грамм ткани.
Кроме того, в то время как адекватные биоэксперименты были поставлены для
периферийных нейронов, создание надежных, повторимых и специфичных экспериментов для нейронов центральной нервной
системы оказалось проблематичным. Тогда как отдельные типы периферийных нейронов
обнаруживаются как дискретные, легко подвергающиеся диссекции ганглии, нейроны центральной нервной системы являются
неизменно высокогетерогенными в их распределении. Так, для идентификации или
обогащения отдельных классов ЦНС-нейронов требуются специфичные маркеры. Прогресс в получении таких маркеров, например
антител, направленных против клеточной поверхности, или цитоскелетных
компонентов, или специфичных гистологических красок, был очень ограничен. Соответственно оказалось крайне трудным процессом
характеризование нейтропных факторов, которые:
(I) не так обильны,
как NGF, (II) трудны для проведения эксперимента и (III) не имеются в достаточных количествах для получения антител.
2.3.1. Сравнение мозгового нейтропного фактора (BDNF) и фактора нервного роста.
Нейротропная активность, способная поддерживать выживание нейронов ганглия дорсального корня куриного эмбриона "ин витро" была идентифицирована в "кондиционированной среде", в которой культурировали крысиные C-6 глиомные клетки (Barde и др., 1978, Nature 274: 818). Эта активность была нейтрализована антителами к мышиному NGF, предполагая присутствие другого нейротропного фактора в кондиционированной среде. О сходных активностях, которые не могли быть блокированы NGF-антителами, сообщалось впоследствии в культурах нормальных астроглиальных клеток головного мозга взрослой крысы (Lindsay 1979, Nature 282: 80-82 Lindsay и др., 1982, Brain Res 243:329-343) и в экстрактах головного мозга развивающейся и взрослой крысы (Bardl и др., 1980, Proc. Nath. Acad. Sci. США 77: 1199-1203) и спинном мозге развивающегося и зрелого цыпленка (Lindsay и Peters 1984, Neurosci 12: 45-51). Однако ни в одном из случаев не были изолированы или идентифицированы активные факторы, и остается не ясным, относятся наблюдающиеся активности к одному и тому же или к нескольким факторам.
Использовав свиной мозг в качестве исходного материала, Barde и др. (1982, ЕМВО, 1: 549-553) сообщил о факторе, названном теперь мозговым нейротропным фактором (BDNF), который способствовал выживанию нейтронов ганглия дорсального корня от E10/E11 куриных эмбрионов. Было обнаружено, что нейротропная активность располагается в высокощелочном белке (с изоэлектрической точкой p1 более 10.1), который мигрировал при электрофорезе на геле с додецил-сульфатом натрия (SDS) в виде единственной полосы молекулярного веса 12,3 кД. Очищенный фактор оценивался 1,4 • 106, но выход был очень мал, всего лишь приблизительно 1 мкг BDNF, очищенного из 1,5 кг свиного мозга. Кроме того, поскольку последней стадией процесса очистки был гелевый электрофорез, активность BDNF не могла быть полностью вторично ренатурирована из-за присутствия остаточного SDS (Barde и Thoenen, 1985, в "Hormones and Cell Regulation", том 9, изд. Дюмон и др., Элсвир саенс паблишерз, стр. 385-390). Было отмечено, что высокощелочная природа и молекулярный размер BDNF были очень сходными со свойствами мономера NGF. Однако BDNF показал свойства, которые отличаются от известных свойств NGF в том, что (а) в биоэксперименте на ганглии дорсального корня цыпленка антитела к NGF не дали видимого эффекта воздействия на биологическую активность BDNF, (б) действия BDNF и NGF оказались дополняющими и (в) в отличие от NGF было обнаружено, что BDNF не оказывает влияния на выживание симпатических нейронов E12 куриного эмбриона. В дополнение во время ранних исследований мозгового экстракта наблюдалось, что нейротропная активность в этих источниках действует на сенсорные нейроны на более поздних стадиях развития, чем активность, связанная с NGF. С использованием диссоциированных культур нейронов куриного эмбриона, культурированных на поликатионном субстрате, таком как полилизин или полиорнитин, было обнаружено, что BDNF поддерживает выживание более чем 30% E10-E11 (десятого или одиннадцатого дня эмбрионального развития) нейронов ганглия дорсального корня, но слабо действует на выживание тех же нейронов при E6 (Barde и др. , 1980, Proc. Nath. Acad. Sci. США, 77: 1199-1203, выше). При сходных условиях сообщалось, что NGF поддерживает выживание 30-40 процентов E6 DRG-нейронов. Любопытно, что позднее было обнаружено, что при культурировании на субстрате, покрытом внеклеточным матричным гликопротеином ламинином, как NGF так и BDNF поддерживали выживание примерно 50% DRG-нейронов от куриных эмбрионов возраста E6 - E12 (Lindsay и др., 1985, Develop. Biol. 112: 319-328). В более позднем исследовании было обнаружено, что действия NGF и BDNF являются дополняющими, когда оба присутствуют в насыщающих концентрациях.
Ранние исследования Леви-Монтальчини (1966, Harvey Lectures 60: 217-259) нейронной специфичности NGF позволяли предположить, что NGF не является вездесущим нейротропным фактором даже для сенсорных нейронов, поскольку NGF не оказал действия на нейроны определенных черепных сенсорных ганглиев цыпленка, особенно на нодозный ганглий десятого черепного нерва. Позднее "ин виво" исследования (Johnson и др., 1980, Science 210: 916-918; Pearson и др. , 1983, Develop. Biol. 96: 32-36) показали, лишение NGF во время эмбриогенеза не оказало влияния на выживание нейронов в большинстве черепных сенсорных ганглиев крыс, тогда как подобная обработка сильно уменьшила число нейронов в сенсорных ганглиях, происходящих от нервного гребня. Более детальные исследования "ин витро" (Lindsay и Bhrer, 1985, Develop Biol 112: 30-48; Davies и Lindsay, 1985, Develop. Biol. III: 62-72, Lindsay и др., 1985, J. Cell Sci. Suppl 115-129) ясно указывали, что NGF поддерживает выживание большинства неврального гребня сенсорных нейронов, но не оказывает видимого влияния на выживание черепных сенсорных нейронов, происходящих от невральных плакод.
Первым свидетельством нейронной специфичности BDNF, отличающейся от активности NGF, была демонстрация "ин витро" того, что очищенный BDNF поддерживает выживание 40-50% сенсорных нейронов, диссоциированных из происходящего от невральной плакоды нодозного ганглия куриного эмбриона возраста E6, E9 или E12 (Lindsay и др., 1982, J. Cell. Sci. Suppl 3: 115-129). NGF не оказал при этом видимого действия на эти нейроны как сам, так и в сочетании с BDNF. Позднее было показано в исследованиях культур эксплантантов, что BDNF поддерживает выживание и отрастание нейритов от сенсорных ганглиев, происходящих от невральных плакод, включая височный, коленчатый и вентролатеральный тройничный ганглии (Davies и др., 1986, J. Neurosci. 6: 1897-1904), ни в одном из которых не была обнаружена чувствительность к NGF. Во всех вышеуказанных исследованиях антитела к NGF не дали эффекта на наблюдаемой активности BDNF. В дополнение к этим эффектам на культурированных нейронах из периферийных ганглиев было обнаружено, что BDNF стимулирует выживание и нейронную дифференциацию клеток, культурированных из неврального гребня перепелки (Kolcheim и Gcandreau, 1988, Develop. Brain Res. 41: 79-86).
До настоящего изобретения невозможность получить достаточное количество BDNF для иммунизации препятствовала производству антиBDNF-антител для сравнения с антителами против NGF по их действию на нейронные популяции и не позволяла осуществить BDNF/NGF эксперименты по их перекрестной нейтрализации. Два недавних исследования BDNF (Kalcheim и др., 1987, ЕМВО 6: 2873; Hofer и Barde, 1988, Nature 331: 261-262) показали, однако, физиологическую роль BDNF в развитии птичьей периферийной нервной системы. Если в яйце между E3/E4 DRG и их ЦНС мишенью в невральной трубке было установлено механическое препятствие, то наблюдалась гибель многих DRG-нейронов (Kalcheim и Le Douarin, 1986, Develop. Biol. 116: 451-466). Было постулировано, что эта нейронная гибель может происходить вследствие лишения ЦНС-(невральной трубки)-нейтропного фактора. Впоследствии наблюдали, что BDNF, прикрепленный к покрытой ламинином сиаластиковой мембране, смог предотвратить эту клеточную гибель (Kalcheim и др., 1987, ЕМВО 6: 2871). Было обнаружено, что инъекции в развивающиеся яйца перепелки снизили естественно происходящую клеточную гибель в нодозных ганглиях, эффект, который не наблюдался с NGF (Hofer и Barde 1988, Nature 331: 261-262). В дополнение к этому эффекту на периферийных сенсорных нейронах происхождения как от неврального гребня, так и от невральных плакод было обнаружено, что BDNF поддерживает выживание развивающихся ЦНС-нейронов. Джонсон и др. (1986, J. Neurosci 6: 3031-3938) представил данные, которые указывают, что BDNF поддерживает выживание клеток ганглиев сетчатки, культурированных из крысиных эмбрионов E17. Это согласуется с предыдущими исследованиями, которые показали, что кондиционированные среды и мозговые экстракты, приготовленные из мишенных областей клеток ганглиев сетчатки, поддерживают выживание этих нейронов (McCaffery и др., 1982, Ex Brain Res. 48: 37-386; Sarthy и др., 1983, J. Neurosci 3: 2532-2544; Turner и др., 1983, Dev. Brain Res. 6: 77-83).
В дополнение к этому действию на выживание развивающихся нейронов в культуре было показано, что BDNF действует на культурированные взрослые нейроны периферийной и центральной нервной системы. BDNF как и NGF стимулировал регенерацию аксонов от DRG-нейронов взрослой крысы в культуре (Lindsay, 1988, J. Neurosci 8: 2394-2405), хотя взрослые сенсорные нейроны не требовали нейротропных факторов для своего поддержания "ин витро" в течение более 3 или 4 недель. Кроме того, в культурах сетчатки взрослой крысы, наблюдалось, что BDNF способствует выживанию и аксонному удлинению от ганглиевых клеток сетчатки (Thonos и др., 19 Eur. J. Neurosci 1: 19-26). Сравнение биологического действия NDF и BDNF представлено в табл. I.
2.3.2. Нейронные мишени мозгового нейротропного фактора.
Было обнаружено, что сенсорные нейроны периферийных нервных ганглиев происходят от двух отличных временных эмбриологических образований, а именно от неврального гребня и невральных плакод. Невральный гребень дает начало нейронам и сателлитным клеткам автономных ганглиев и сенсорным ганглиям спинного мозга, т. е. DRG. Вклад неврального гребня и невральных плакод в образование сенсорных ганглиев черепных нервов изучался с использованием перепелка (цыпленок химерной трансплантационной системы согласно Le Douarin, 1973, Develop. Biol 20:217 - 222; Noden, 1987, Develop. Biol 67: 313 - 329; Narayanan и Narayanan, 1980, Anat. Rec. 196: 71 - 82; Ayer - Le Lievre и Le Douarin, 1982, Develop. Biol 94: 291 - 310; D'Amico - Maratel и Noden, 1983, Am J. Anat. 166: 445 - 468). Как указано в обзоре Линдсея и др. (1985, J.Cell. Sci. Supp. 3: 115 - 129), теперь полагают, по меньшей мере в отношении птиц, что нейроны дистальных ганглиев восьмого и десятого черепных нервов (коленчатый, височный и нодозный ганглии соответственно) и нейроны вестибулоакустического комплекса восьмого черепного нерва происходят исключительно от невральных плакод. Тройничный ганглий пятого черепного нерва содержит нейроны как гребневого, так и плакодного происхождения (с нейронами плакодного происхождения, преобладающими в вентролатеральном полюсе максилло-мандибулярной доли), тогда как сателлитные клетки всех черепных ганглиев имеют полностью гребневое происхождение.
Из экспериментов "ин витро" с использованием как эксплантанта, так и диссоциированных обогащенных нейронами культур сенсорных нейронов нервов спинного мозга и черепных нервов было обнаружено, что сенсорные нейроны, происходящие от неврального гребня, реагируют на NGF, напротив нейроны плакодного происхождения (включая нейроны вентролатеральной части тройничного ганглия и всю нейронную популяцию вестибулярного, коленчатого, височного и нодозного ганглиев) в основном были нечувствительны к NGF во время эмбрионального развития. Несмотря на их различия в потребности и чувствительности к NGF как плакодные, так и гребневые сенсорные нейроны, как было обнаружено, реагируют (табл. I) на способствующую выживанию и отрастанию нейритов активность BDNF (Lindsay и др., 1985, J.Cell. Sci Supp. 3:115 - 129; Lindsay и др. , 1985, Develop Biol 112: 319-328; Kalcheim и Geandreau, 1988, Develop. Brain Res. 41: 79-86) Тебар и Бард (Tebar и Barde, 1988, J.Neurosci 8: 3337-3342) исследовали связывающие параметры радиоактивного меченного BDNF к нейронам ганглия дорсального корня куриного эмбриона; эти результаты согласуются с существованием двух классов BDNF рецепторов, одного - с высокой аффинностью к BDNF и другого - низкой аффинностью. На симпатических нейронах не наблюдалось рецепторов с высокой аффинностью.
Известные нейронные мишени BDNF далее рассматривались в обзоре Берда и др. (1987, Prog. Brain Res. 71: 185-189). До настоящего изобретения, идентификация клеток, синтезирующих BDNF, была невозможна вследствие отсутствия нуклеиновокислотных или антительных зондов, специфичных для BDNF. Попытки приготовить поли- или моноклональные антитела к BDNF были безуспешными. Эта неудача в получении антител воспрепятствовала молекулярному клонированию BDNF детерминированию физиологического действия, создаваемого лишением развивающихся нейронов BDNF "ин виво", коническому определению BDNF в такнях с использованием иммуноопытов и локализации BDNF с использованием иммуноцитохимии.
Список реагирующих и нереагирующих на BDNF нейронов (Barde и др., 1987, Prog. Brain Res. 71: 185-189).
A. Реагирующие нейроны.
I. Сенсорные нейроны цыпленка происхождения от неврального гребня в:
а) ганглии дорсального
корня,
б) шейном ганглии,
в) дорсомедиальном тройничном ганглии,
г) мезенцефальном тройничном нуклеусе (Dav'us и др., 1986, Nature 319: 497-499).
II.
Сенсорные нейроны цыпленка происхождения от эктодермальной плакоды в:
а) нодозном ганглии,
б) вестибулярном ганглии,
в) височном ганглии,
г) коленчатом ганглии,
д) вентролатеральном тройничном ганглии.
III. Клетки ганглия сетчатки крысы.
B. Нереагирующие нейроны.
I. Симпатические нейроны крысы и цыпленка.
II. Парасимпатические цилиарные нейроны цыпленка.
3. Краткое содержание изобретения.
Настоящее изобретение касается нуклеиновокислотных последовательностей, кодирующих мозговой нейротропный фактор (BDNF) в основном чистого протеина, а также пептидных фрагментов или производных, производимых от него в большом количестве, и антител, направленных против протеина BDNF, пептидных фрагментов или производных. Настоящее изобретение обеспечивает впервые достаточные количества BDNF, позволяющие осуществить производство антиBDNF-антител и поддержать диагностическое и терапевтическое применение BDNF.
В различных выполнениях изобретения BDNF нуклеиновые кислоты, протеины, пептиды, производные или антитела по изобретению могут быть использованы в методах для диагностики и лечения разнообразных неврологических заболеваний и расстройств и, в частности, в диагностике и лечении нарушений, связанных с сенсорными нейронами, а также дегенерации сетчатки. Кроме того, в специфических выполнениях изобретения нуклеиновые кислоты BDNF и BDNF-генные продукты могут быть использованы для диагностики и лечения нейробластомной опухоли, болезни Паркинсона и болезни Альцхаймера. BDNF-генные продукты могут быть также использованы для того, чтобы облегчить инкорпорацию имплантантов в нервную ткань или альтернативно для способствования нервной регенерации после повреждений, вызванных травмой, инфарктом, инфекцией или хирургическим вмешательством (послеоперационно) в дополнительных специфических выполнениях изобретения.
Изобретение также относится к фармацевтическим композициям, содержащим эффективные количества BDNF-генных продуктов или альтернативно антител, направленных против BDNF-генных продуктов, которые могут быть использованы в диагностике или лечении различных неврологических болезней и расстройств.
Дополнительно, путем обеспечения полной нуклеотидной последовательности BDNF настоящее изобретение позволяет провести сравнение BDNF- и NGF-генов, при этом идентифицируя гомологичные области и определяя BDNF/NGF-генное семейство. Соответственно настоящее изобретение касается способа идентифицирования дополнительных членов BDNF/NGF-генного семейства. В специфическом выполнении этот способ по изобретению используется для идентифицирования нового, не-BDNF и не-NGF члена генного семейства BDNF/NGF. Изобретение далее обеспечивает получение дополнительных членов генного семейства BDNF/NGF, идентифицированных согласно раскрытому способу, и также их генные продукты.
3.1. Сокращения и их значение.
BDNF - мозговой нейротропный фактор.
hBDNF - человеческий BDNF.
САТ - холин-ацетилтрансфераза.
ЦНС - центральная нервная система.
DRG - ганглий (или ганглии) дорсального корня.
ЭДТК - этилен-диамин-тетрауксусная кислота.
NGF - фактор нервного роста.
PBS - буферированный фосфатом физиологический раствор.
PCS - реакция цепной полимеризации.
RNG - периферийная нервная система.
SDS -натрия додецил-сульфат.
Tris - три-/гидроксиметил/аминометан.
4. Описание чертежей.
Фиг. 1 - нуклеотидная последовательность и выведенная из нее аминоксилотная последовательность свиной prepro-BDNF кДНК. На этом чертеже показана полная последовательность двух перекрывающих кДНК клонов. Подчеркнуты пептидные последовательности, полученные при микросеквенировании (табл. II).
Фиг. 2 - сравнение последовательностей NGR и BDNF. Области с более чем двумя аминокислотами, обнаруживаемыми в идентичных положениях, отмечены. Эти последовательности начинаются с первых аминокислот зрелых протеинов и кончаются с последними аминокислотами перед стоп-кодонами. 51 аминокислота - общие для BDNF и различных NGF (Schwarz и др., 1989, J.Neurochem. 52: 1203-1209), включая шесть цистеиновых остатков.
Фиг. 3 - авторадиография саузерн-блоттинга EcoRI разреза геномной ДНК человека, обезьяны, крысы, мыши, собаки, коровы, кролика, цыпленка и дрожжей, гибридизированной к меченным 32 P NGF и BDNF-зондам.
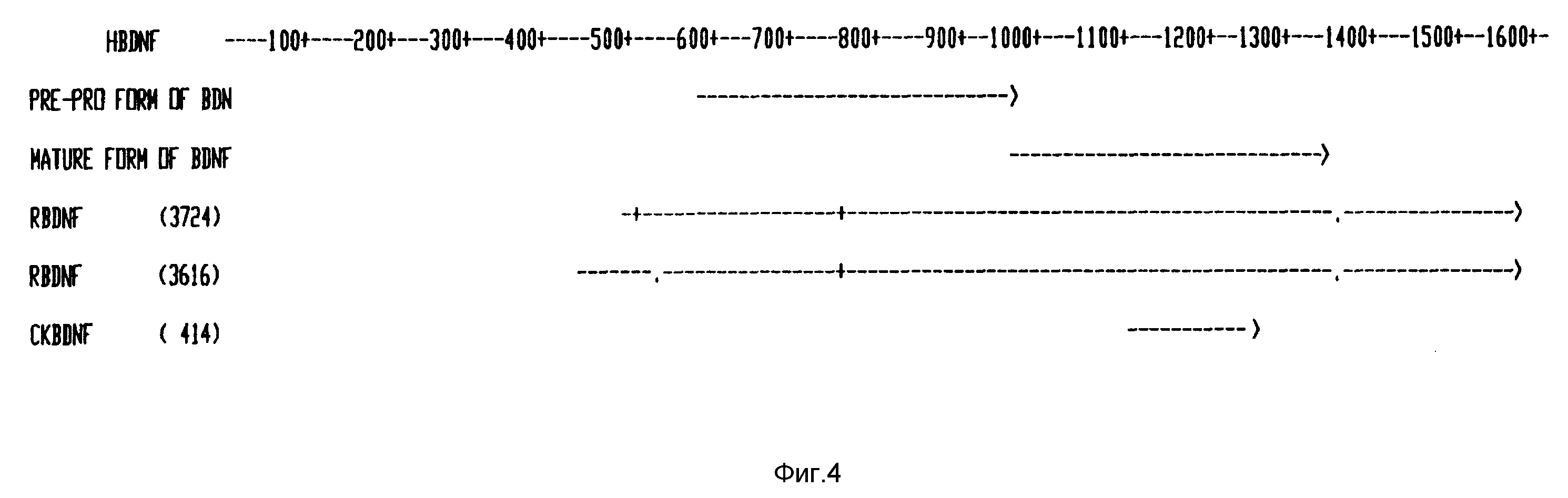
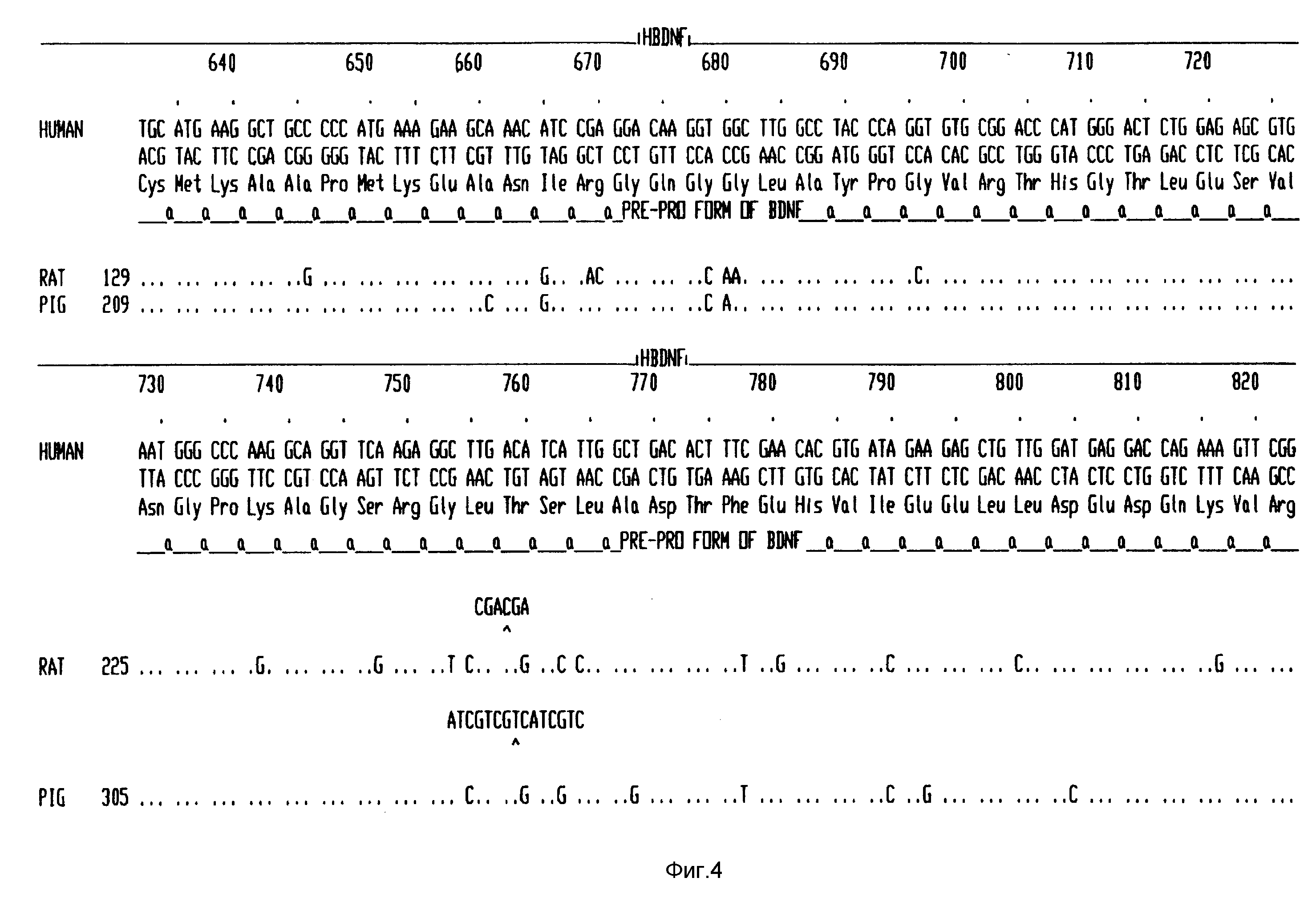
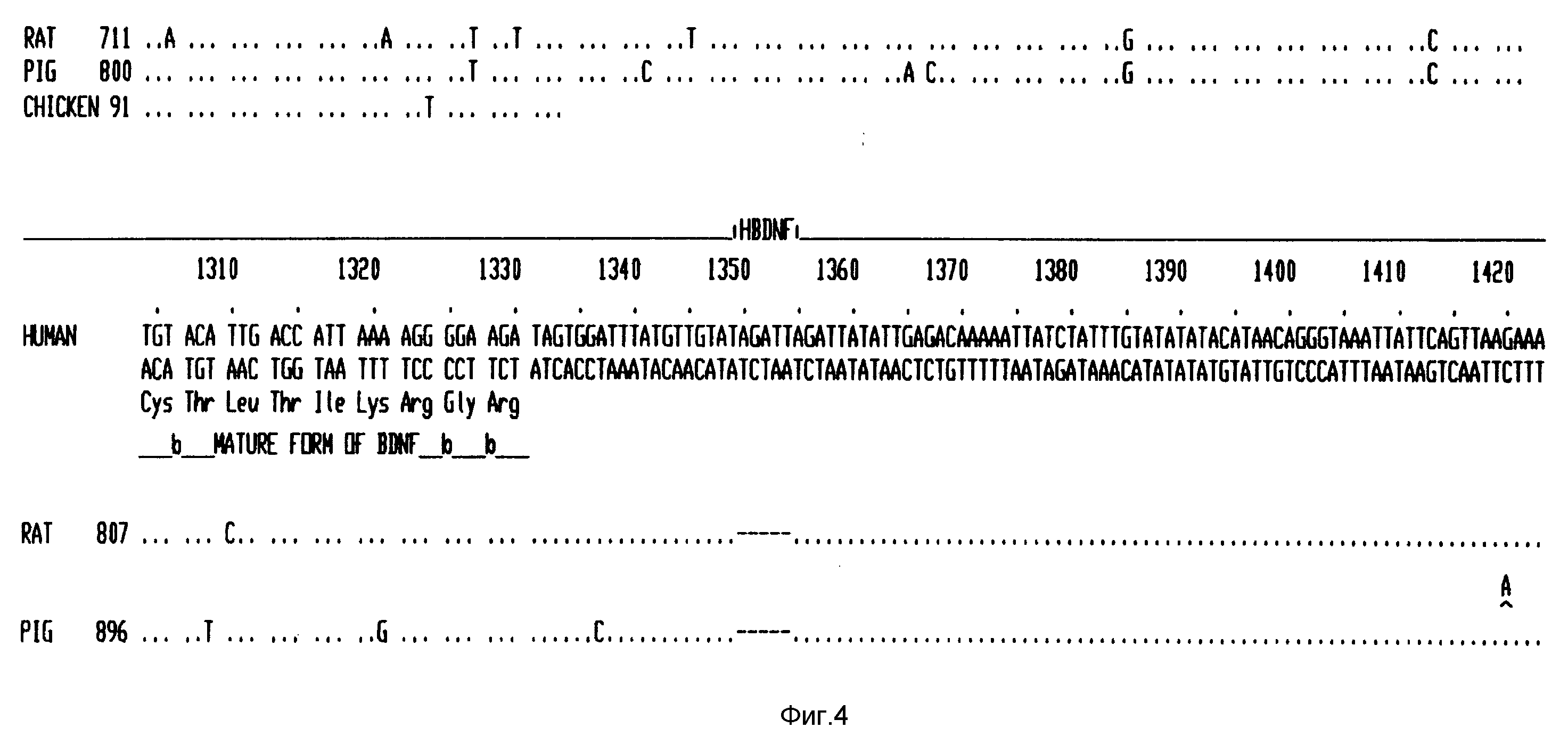
Фиг. 4 - последовательность кДНК человеческого BDNF и выведенная из нее аминокислотная последовательность, и сравнение последовательностей ДНК свиньи, крысы и цыпленка.
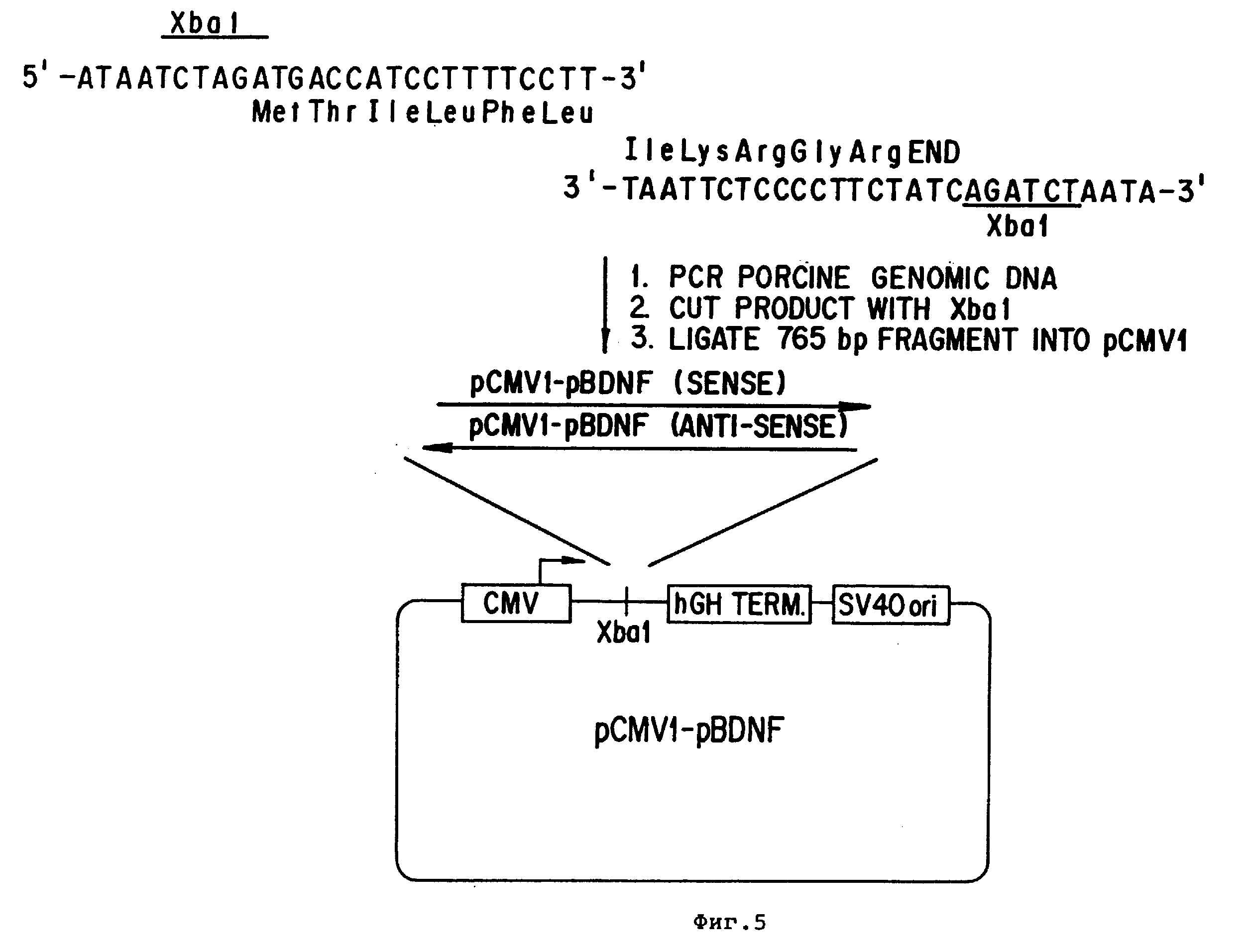
Фиг.5 - BDNF-экспрессионная плазмида pCMVI-pBDNF.
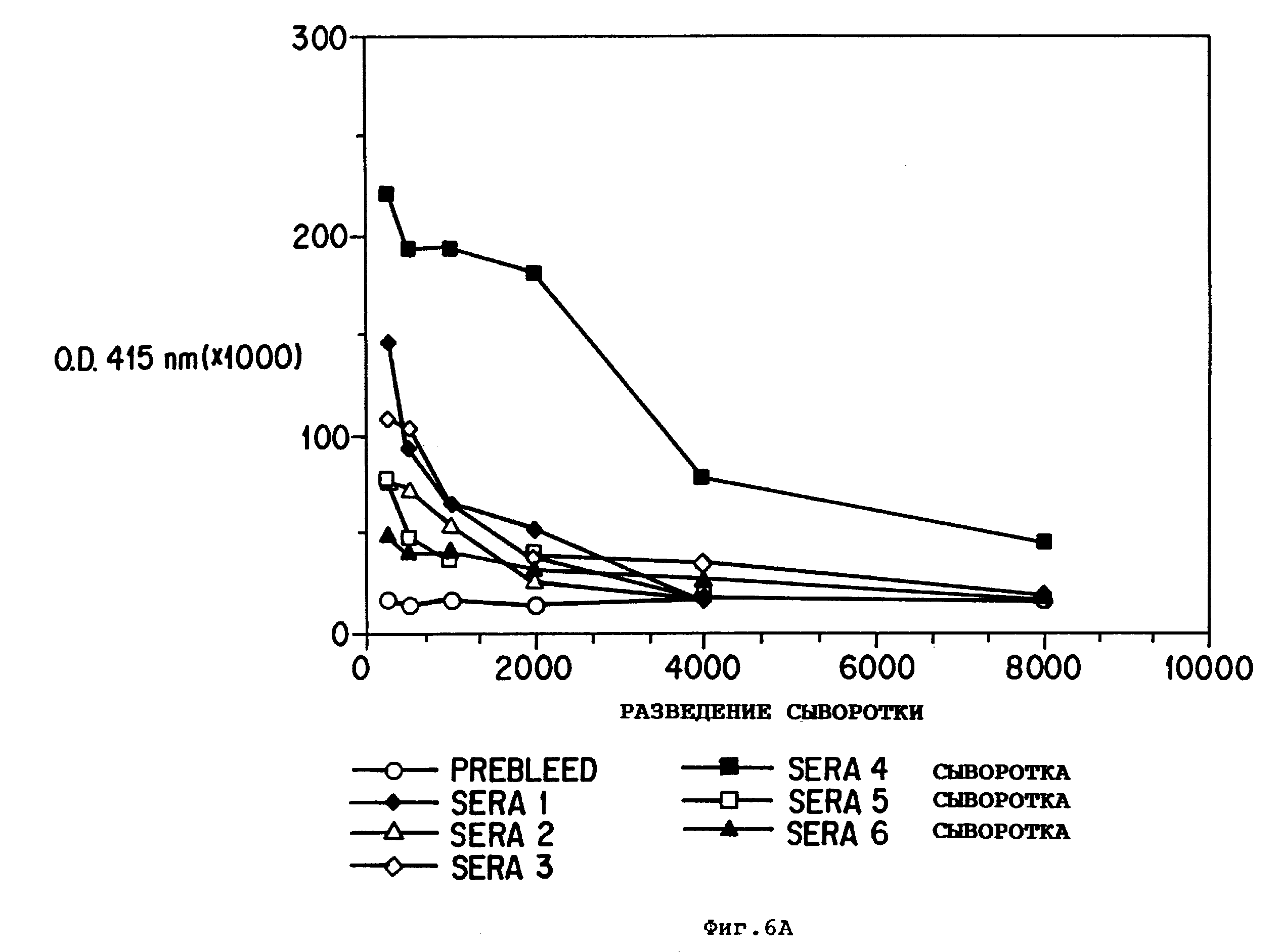
Фиг.6: (A) - результаты ELI SA-детерминирования связывания антисывороток к B5 пептиду с использованием серийных разведений антисывороток;
(B) - количественное
определение связывания при разведении 1:500 различных антисывороток к 50 нг BDNF.
Фиг. 7 - результаты иммуногистохимического окрашивания на тирозингидроксилазу в BDNF-обработанной (прерывистые линии) и контроль (сплошные линии) культур вентрального среднего мозга.
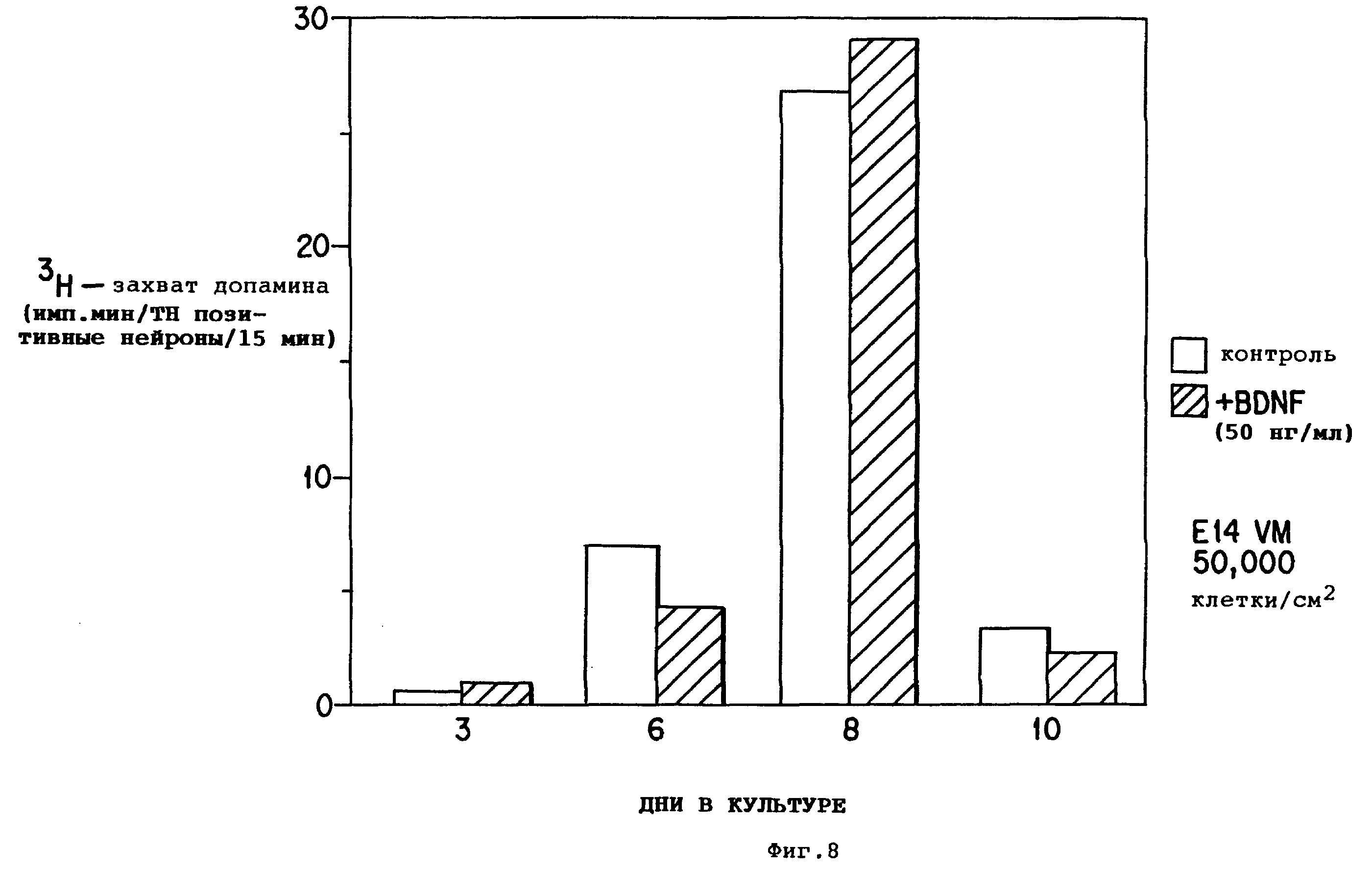
Фиг.8 - потребление допамина культурами вентрального среднего мозга. Культуры с BDNF отмечены прерывистыми линиями, контрольные культуры показаны сплошными линиями.
Фиг.9: (A) - действие BDNF на несколько CAT положительных клеток в
культурах холинергических нейронов переднего
мозга;
(B) - культуры холинергических нейронов переднего мозга при плотности 260,000 клеток (черная полоса) или 150,000 клеток на лунку
(прерывистая полоса) обрабатывали 150 нг/мл NGF. Было
сравнено количество CAT иммуноположительных клеток среди NGF-обработанных и необработанных клеток.
Фиг.10 - изменения количества CAT энзимной активности, в пикомолях субстрата, катализируемого за минуту, в виде функции концентрации BDNF в культурах холинергических нейронов переднего мозга.
Фиг. 11 - культуры астроглиальных клеток, примерно с 60%-ным слиянием, были обработаны: (A) - фактором эпидермального роста или (B) - BDNF в течение 42 часов, а затем инкубированы с [3H] метилтимидином. Количество присоединенного [3H] измеряли относительно концентраций EGF и BDNF (EGF - фактор эпидерм. роста).
Фиг. 12 - саузерн-блоттинг EcoRI-разрезанных BDNF/NGF цыпленка, мыши, крысы, гибридизированных к зонду RIB/20. Положения NGF и BDNF геномных EcoRI фрагментов указаны как N и B соответственно.
Фиг.13 - сравнение последовательностей BDNF и NGF с новым членом генного семейства BDNF/NGF, идентифицированным путем PCR-реакции из мышиной ДНК с праймерами Box 3/Box 4: новый ген (обозначенный здесь M3-4), а также известный как Нейротропин-3 (NT-3), показывающий последовательность только кодирующей нити, а также выведенная аминокислотная последовательность в сравнении со зрелым мышиным NGF и зрелым свиным, крысиным, мышиным или человеческим BDNF. Тире обозначают позиции, в которых использована делеция одного кодона в NGF для оптимизации сравнения относительно DBNF и M3/4. Подчеркнутое указывает на совместимости в аминокислотной последовательности и/или консервативные аминокислотные замещения.
Фиг. 14 - назерн-блоттинг РНК из различных клеточных линий из опухоли человека, гибридизированной к BDNF человека зонду.
Фиг. 15 - эффект деполяризации уровней BDNF-мРНК на нейроны аммонова рога. (A) - экспрессия BDNF мРНК по времени в первичных культурах нейронов аммонова рога в присутствии 50 мМ KCl. Общую клеточную РНК экстрагировали из 0,5 • 106 клеток, гликозилировали и анализировали посредством электрофореза на 1,3% агарозном геле (Biziere, K. Coyle, T. Neurosei., 1978, Lett, 8:303; McGeer и др. , 1978, Brain Res., 139:381). РНК, перенесенную на фильтры Hybond N, гидридизировали 32P-меченым кРНК зондом (специфическая активность, 109 срм/мг), специфичным для мышиного BDNF и полученным сбегающей транскрипцией "ин витро". Две верхние полосы соответствуют (4 и 1,5 кВ) BDNF-мРНК; две полосы для BDNF (4 кб и 1,5 кб) являются рибонуклеазо-A-резистентными и представляют два различных транскрипта, которые, однако, регулируются одинаковым образом, а нижняя полоса (700 бр) соответствует 10 пикограммам стандартного выхода короткой BDNF-мРНК, который был добавлен к образцам перед экстракцией РНК.
(B) - кальциевая зависимость. Нейроны инкубировали в течение 3 часов в нормальной культурирующей среде (СМ), в модифицированной среде без кальция (-Ca) или в среде с 10 м нифедипина (NIF). Где указано (+K), там добавляли 50 мМ KCl.
Фиг.16 - повышение уровней NSF-мРНК у нейронов аммонова рога посредством калия и каиновой кислоты. Нейроны инкубировали в течение 3 ч в контрольной среде (C) или в присутствии 50 мМ RCl (+K) или 25 мМ каиновой кислоты (KA). РНК экстрагировали и определяли уровни NGF-мРНК количественной PCR-реакцией (II). Оценки даны: среднее ±.
Фиг.17 - кривая реакции на дозу, показывающая действие каиновой кислоты. Нейроны аммонова рога инкубировали в течение 3 часов в присутствии различных концентраций каиновой кислоты. РНК экстрагировали и анализировали, как описано на фиг.16. Оценки представляют среднее ± SEM трех экспериментов.
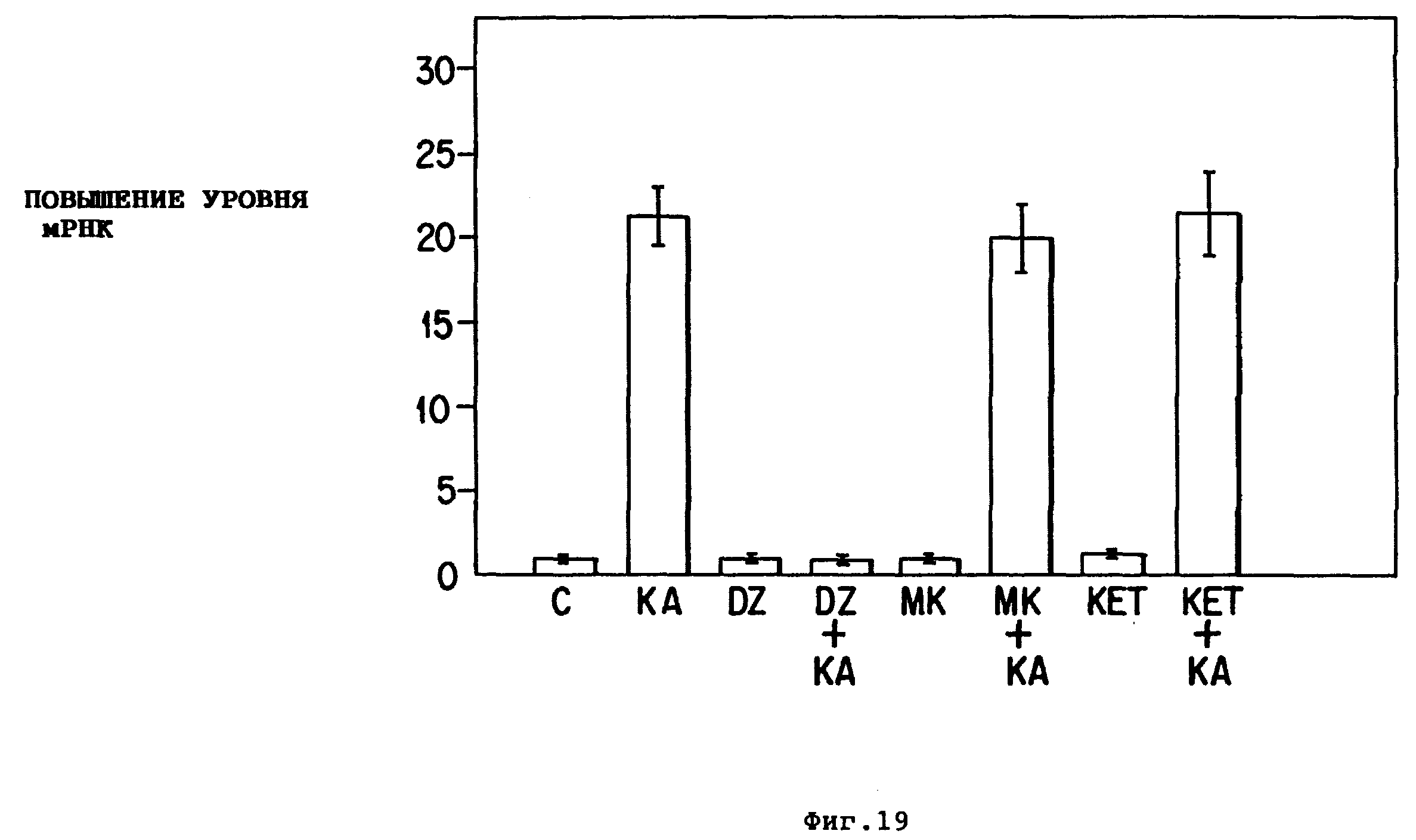
Фиг. 18 - временная зависимость повышения уровней BDNF- и NGF-мРНК вследствие обработки каиновой кислотой. Общую клеточную РНК экстрагировали и анализировали, как указано на фиг.16, из аммонова рога (A) или кортекса (B) в указанное время (в часах) после внутрибрюшинной инъекции каиновой кислоты (12 мг/кг). Одну разовую дозу диацепама (10 мг/кг) вводили инъекцией через 90 мин после каиновой кислоты (две полосы для BDNF (4 кб и 165 кб) являются рибонуклеазо-A-резистентными и представляют два различных транскрипта, которые, однако, регулируются схожим образом). Показаны значения, соответствующие 1,5 кб BDNF-мРНК (кружок) и 1,3 кб NGF-мРНК (закрашенный кружок). Похожие повышения уровней наблюдались для 4 кб BDNF-мРНК. Данные на фиг. значения представляют среднее ± SEM от 3-4 экспериментов.
Фиг. 19. Действие антиконвульсантов на вызванную каиновой кислотой экспрессию BDNF-мРНК. Крыс инъецировали внутрибрюшинно диацепамом (10 мг/кг) (DZ); MK-801 (1 мг/кг) (MK) или кетамином (20 мг/кг) (KET) за 15 мин до инъекции каиновой кислоты (12 мг/кг) (+KA) или физиологического раствора (контроль). Тремя часами позже ткань аммонова рога использовали для приготовления общей РНК, как указано на фиг.16. Значения представляют среднее ± SEM трех экспериментов.
Фиг.20 - перегородочные клеточные культуры.
(A) - фазово-контрастная микрография роста клеток в культуре в зависимости от времени. Клетки помещали на чашку с плотностью 1,3 • 105 клеток на см2 и поддерживали в 5 HS/NS, как описано в тексте. Указанные моменты времени - A, B - 1 день, C, D - 2 дня и E, F - 4 дня.
Специфичные для клеточного типа маркеры использовали для идентификации популяций клеток, ACL E гистохимически окрашенных нейронов (C, H) и NGF рецептор иммуноположительных нейронов (1). Масштаб шкалы = 25 мм.
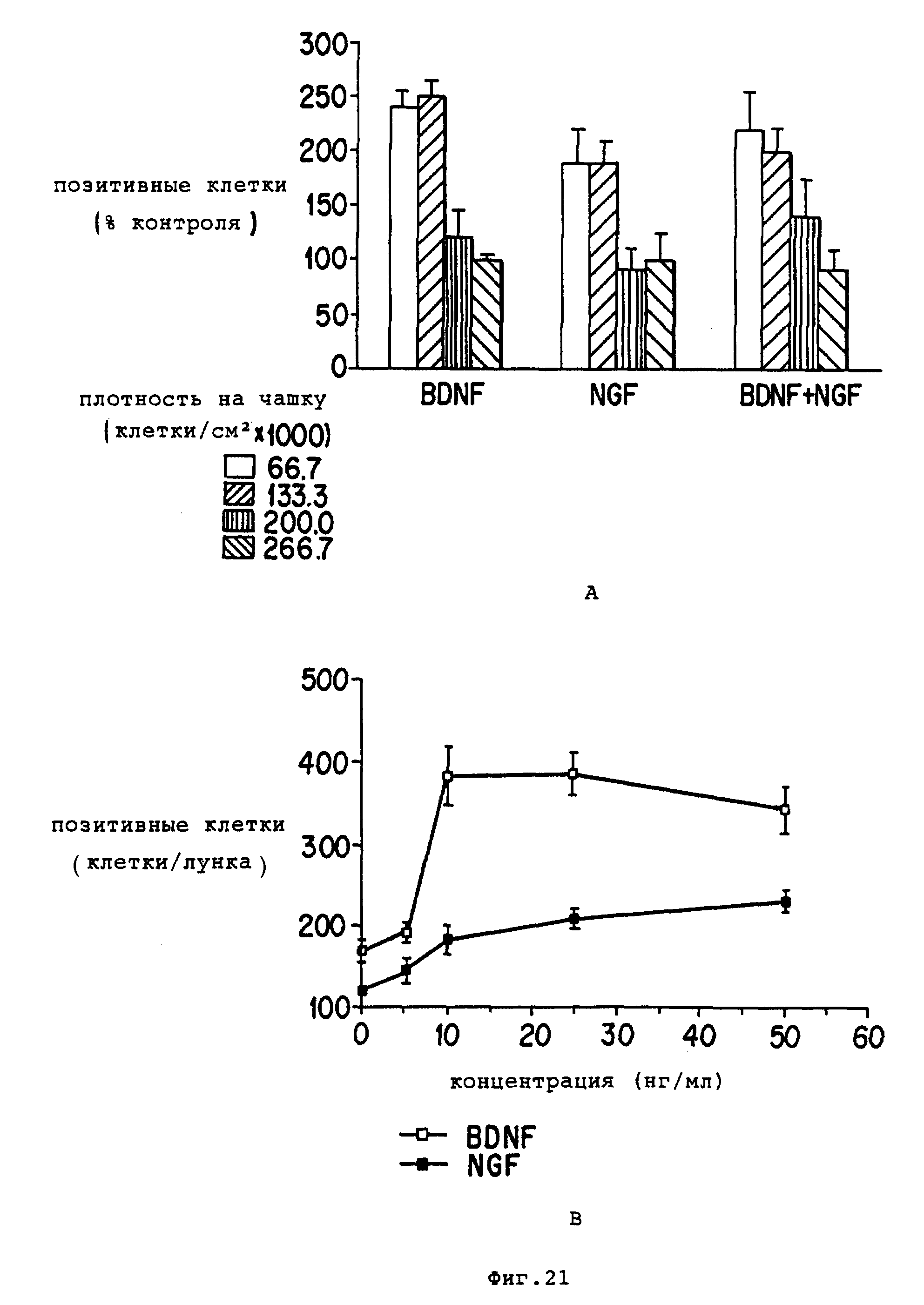
Фиг. 21 - cравнение реакции диссоциированных перегородочных клеток на обработку BDNF и NGF на основе AchE клеточного количества.
(A) - cравнение реакции на BDNF (25 нг/мл), NGF (50 нг/мл) или сочетание обоих лигандов при различной чашечной плотности.
(B) - cравнение реакции холинергических нейронов на разные концентрации BDNF и NGF (от 0 до 50 нг/мл). Клетки, на которых были получены эти результаты, выращивали в течение 11-12 дней в серосодержащей среде. Данные представляют среднее ± SEM от 6-9 определений.
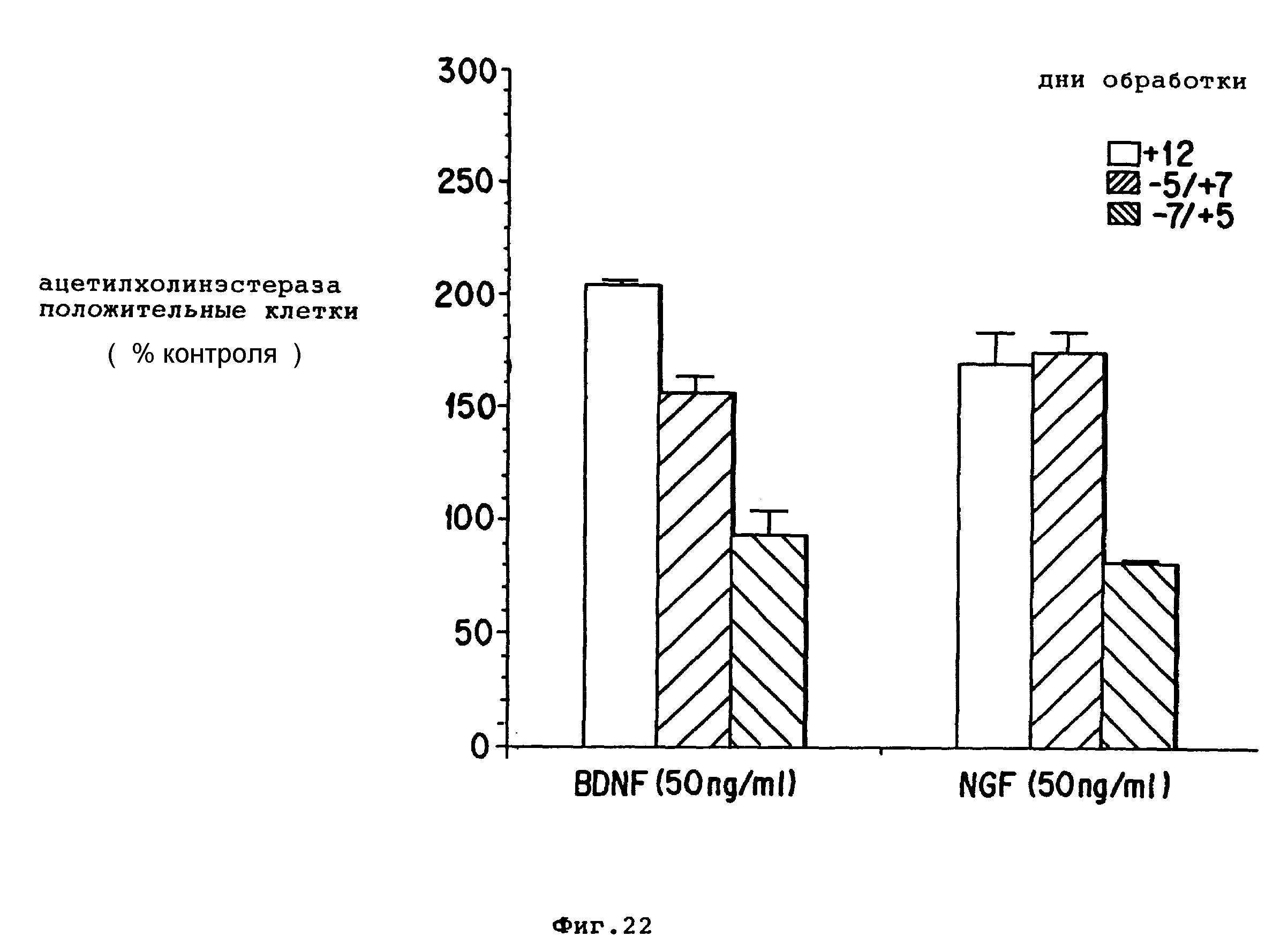
Фиг. 22 - полосовой график изображает действие задержки добавления BDNF или NGF к диссоциированным перегородочным культурам, BDNF (50 нг/мл) и NGF (50 нг/мл) добавляли к диссоциированным клеточным культурам с задержкой 5-6 часов (+12), 5 дней (-5/+7) или 7 дней (-7/+5). Культуры поддерживали в течение 12 дней. Клетки подсчитывали на основе положительного гистохимического окрашивания. Результаты выражены в виде среднего ± SEM от 4-5 определений.
Фиг. 23 - полосовой график, изображающий способность BDNF или NGF регулировать количество NGF-рецепторных иммуноположительных клеток. Иммунное исследование для NGF-рецептора было проведено с использованием моноклонального антитела 192- IgG при разведении 121000. Диссоциированные перегородочные клетки помещали на чашку с плотностью 1,3 • 105 на см2 и непрерывно обрабатывали в течение 12 дней. Сравнение сделано между насыщающей дозой NGF и разными дозами BDNF (от 0 до 100 нг/мл). Изображенные результаты представляют среднее ±SEM при n = 4.
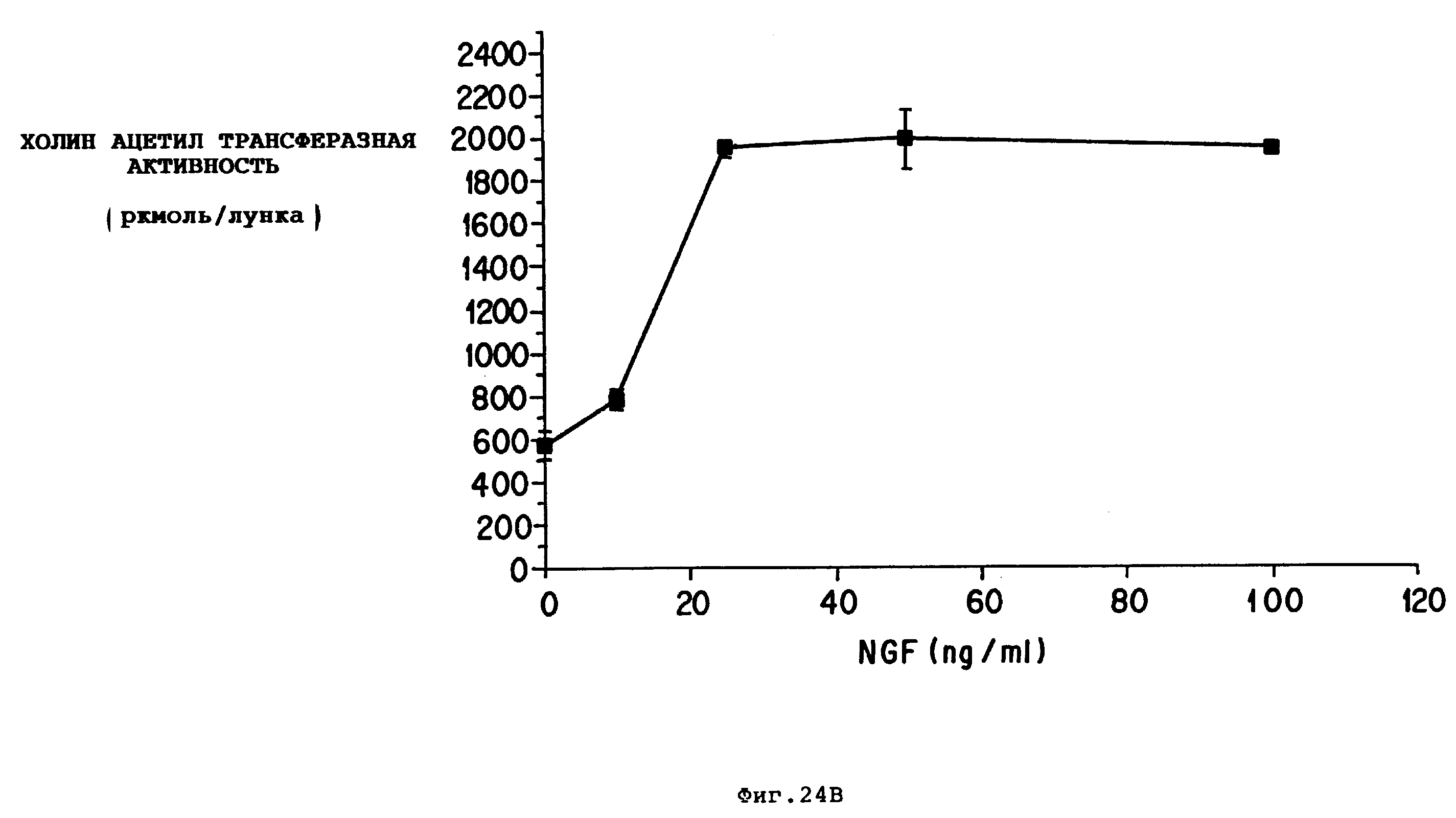
Фиг. 24 - кривая реакции на дозу перегородочных холинергических нейронов на дозу CAT вызывающих активностей BDNF (A) и NGF (B). Культуры помещали в чашки с покрытыми полиорнитином и ламинином 6 мм лунками и поддерживали в течение 12 дней в 5 HS/NS. Клетки подвергали воздействию BDNF или NGF через 5-6 часов после помещения на чашку. Факторы заменяли каждые 3 дня, когда меняли среду, показанные результаты представляют среднее ± SEM из 5-6 определений.
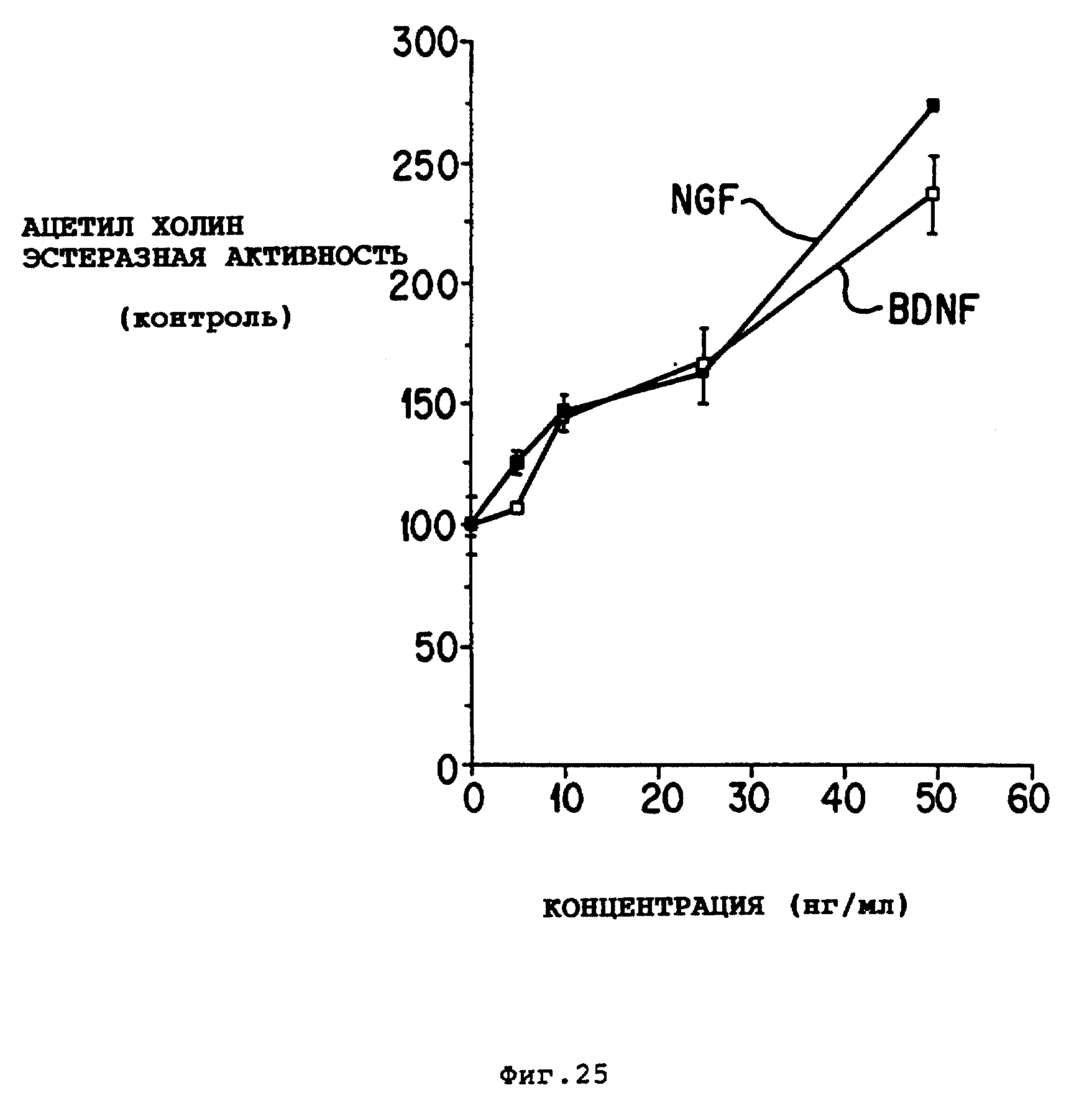
Фиг. 25 - зависимость реакции от доз BDNF и NGF на AChE энзимную активность. Перегородочные холинергические нейроны крысы выращивали в течение 12 дней в серосодержащей среде с гормональными добавками. Клетки обрабатывали BDNF (открытые квадратиками) и NGF (сплошные квадратики), как описано для фиг. 25. Результаты представляют среднее ± SEM при n 6-9, AChE активность в необработанных культурах была 16,4 ± 2 нмоль/Hr/лунка.
Фиг. 26 - временная зависимость повышения CAT энзимной активности, вызванной BDNF по сравнению с NGF. Определение требуемого времени для стимулирования CAT-активности проводили на клетках, помещенных на чашку с плотностью 2,3•105 клеток на см2, которые выращивали различные периоды времени в серосодержащей среде с гормональными добавками и экзогенными факторами. Клетки вначале подвергали воздействию BDNF (открытые квадратики) или NGF (сплошные квадратики) через 5-6 часов после помещения клеток на чашку. Результаты представляют проценты активности, определенной в обработанных культурах против необработанных культур (n = 6, BDNF 12 дней n = 3).
Фиг. 27 - cравнение действия глиальных клеток на способность BDNF вызывать CAT энзимную активность.
Перегородочные клетки помещали на чашку с плотностью 2,3• 105 клеток на см2. Через 5-6 часов после периода засева среду заменяли либо свободной от серы средой или серосодержащей средой, в каждом случае содержавшей 1% NS, как указано в тексте описания. На 24 часа добавляли цитозинарабинозид (1 uM) с целью дальнейшего снижения числа астрацитов. Эти культуры затем поддерживали в течение 12 дней. Результаты представлены в виде среднего ± SEM из 4 определений.
Фиг. 28 - графическое полосовое изображение действия обработки BDNF и NGF на уровень высокоаффинного потребления холина. Исследование потребления холина было проведено на клетках, поддерживаемых в течение 11 дней, после посева с плотностью 1,3 • 105 клеток на см2. BDNF (50 нг/мл) или NGF (50 нг/мл) присутствовали во время всего периода культивирования. Данные представляют среднее ± SEM из 5 определений для BDNF и 10 для NGF.
Фиг. 29 - разрешение листа SDS с35S-мечеными продуктами реакции от ферментного расщепления человеческого мозгового нейротропного фактора трипсином и эндопротеиназой Arg-G.
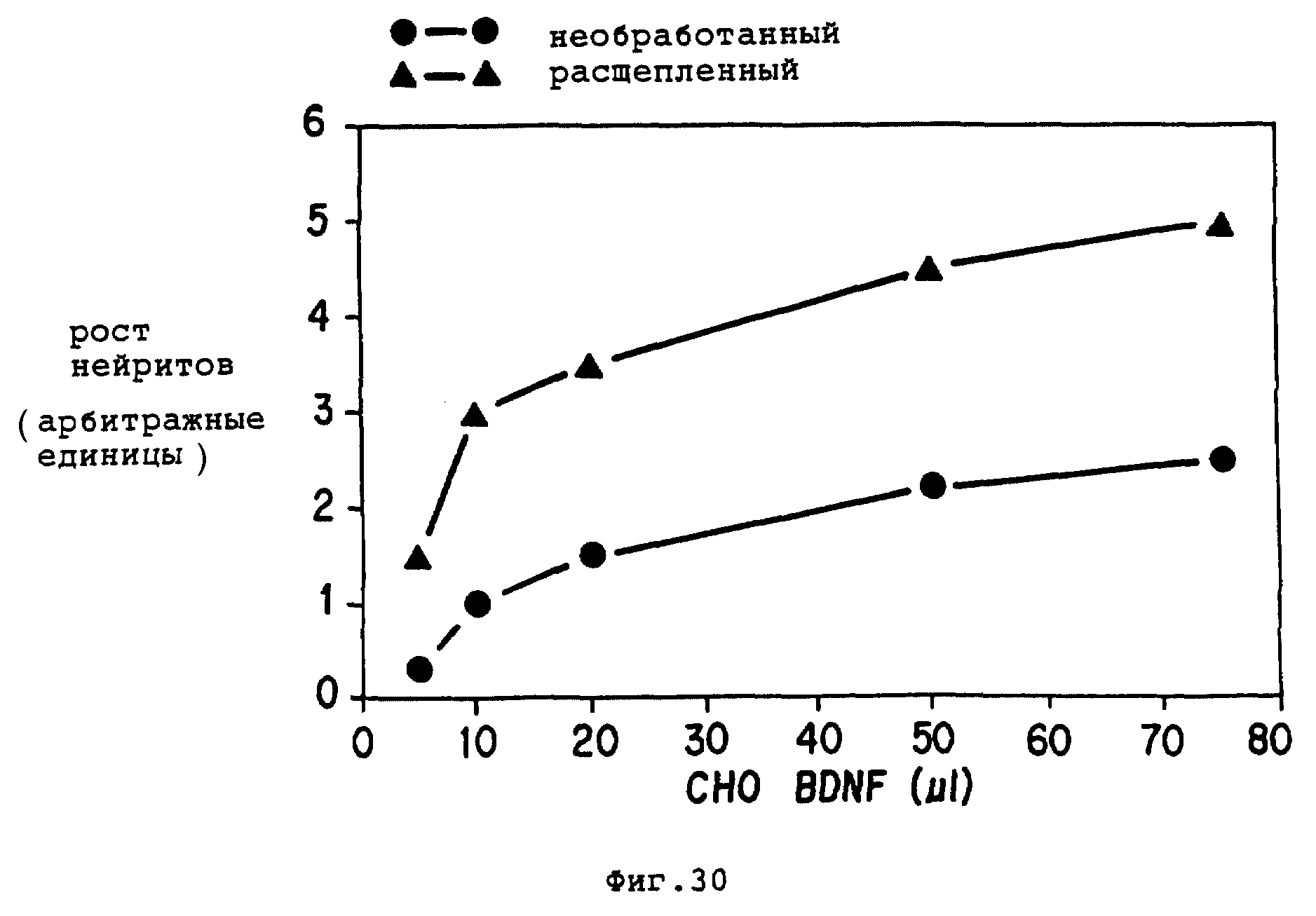
Фиг. 30 - графическое представление DRG биоэксперимента, сравнивающего необработанные CHO-hBDNF и расщепленные эндопротеиназой CHO-hBDNF. Нейтритное отрастание оценено по шкале от 0 до 5, представляющей максимальную биоактивность.
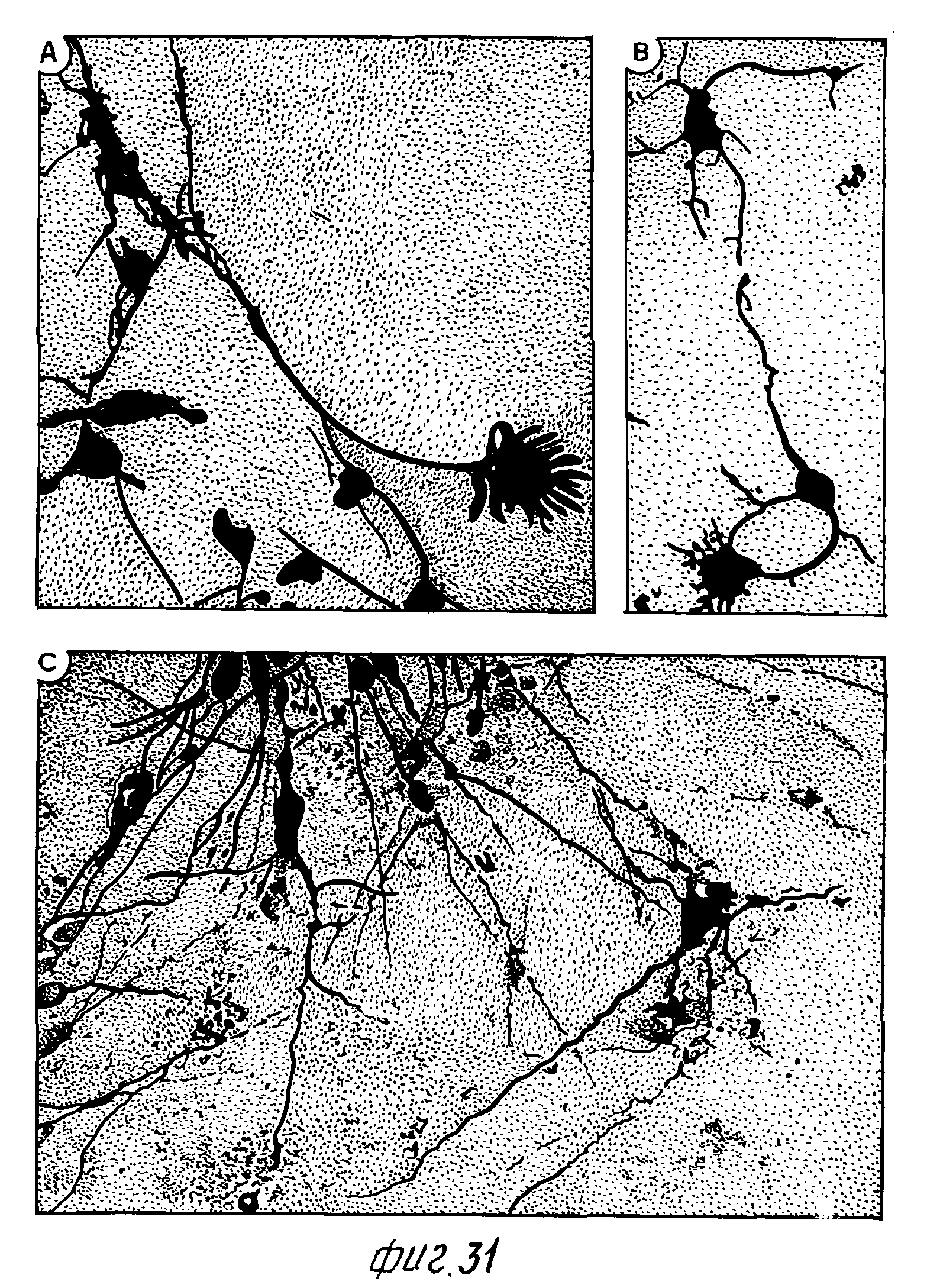
Фиг. 31 - фотомикрографии сильного увеличения 6-дневных культур E14 крысиных вентральных мезенцефальных клеток, показывающих различную морфологию TH+ нейронов, некоторые из них - с экстенсивным нейтритным ростом, и крайне развитыми конусами роста (A, B). Культуры были созданы и окрашены, как описано для фиг. 32. Масштаб шкалы = 25 мкМ.
Фиг. 32 - фотомикрографии, фазоконтрастные (A) и в светлом поле (B, C) диссоциированных культур E14 крысиных вентральных мезенцефальных клеток, окрашенных через 9 дней в культуре с моноклональными антителами к тирозингидроксилазе (TH).
A и B. Фотографии, фазоконтрастные и в ярком поле, того же поля, показывающие низкое содержание TH+-нейронов в этих культурах. Только один TH+-нейрон (стрелка) виден в поле, содержащем много фазоярких нейронов с длинными нейритами. Очевидны несколько ненейронных клеток, посредством морфологии либо посредством окрашивания на GFAP (не показано).
C. Фотография яркого поля, показывающая два TH+-нейрона (маленькие стрелки) в поле, содержащем примерно 98 фазоярких клеток нейронной морфологии. Масштаб шкалы = 100 1М.
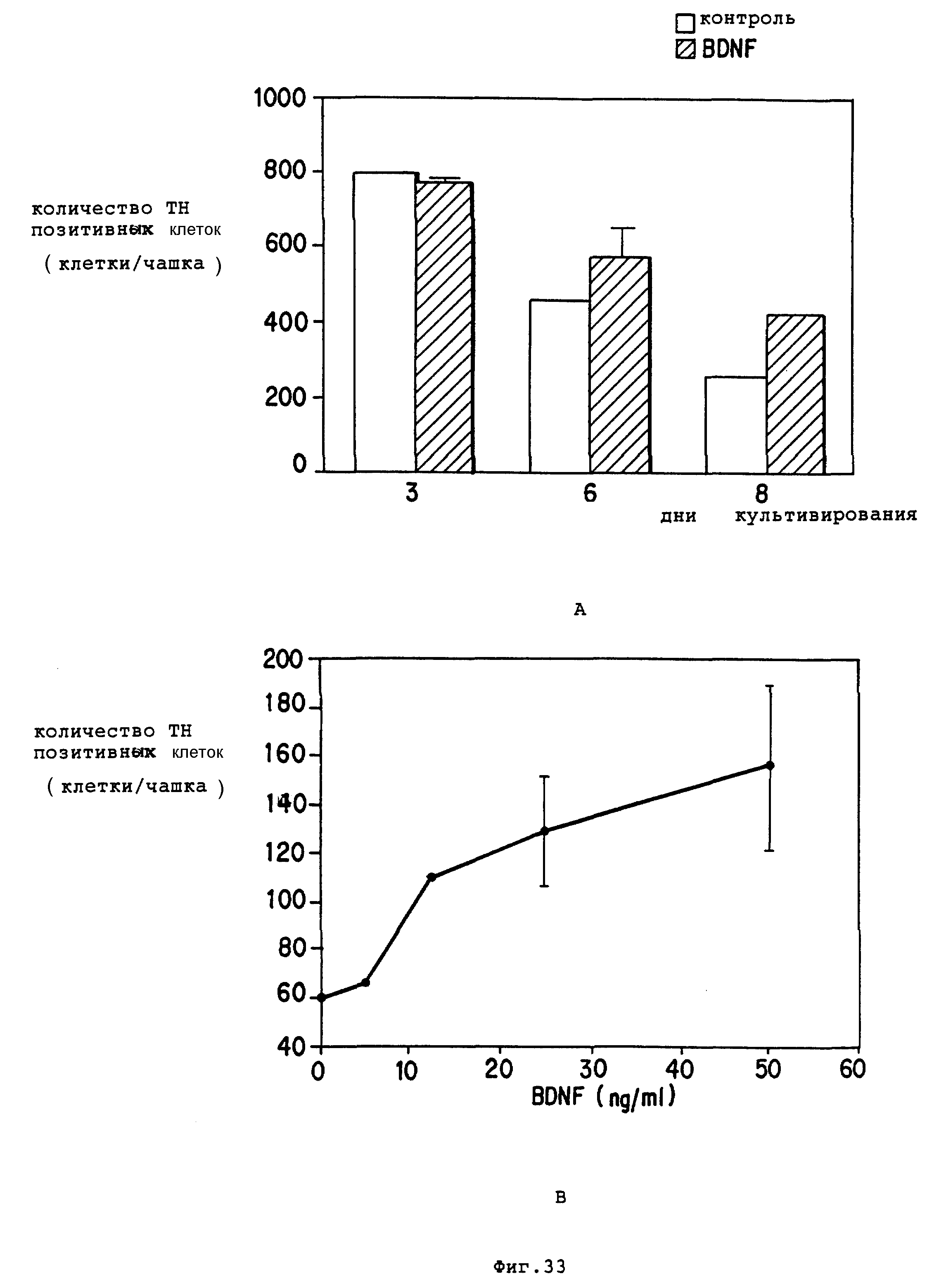
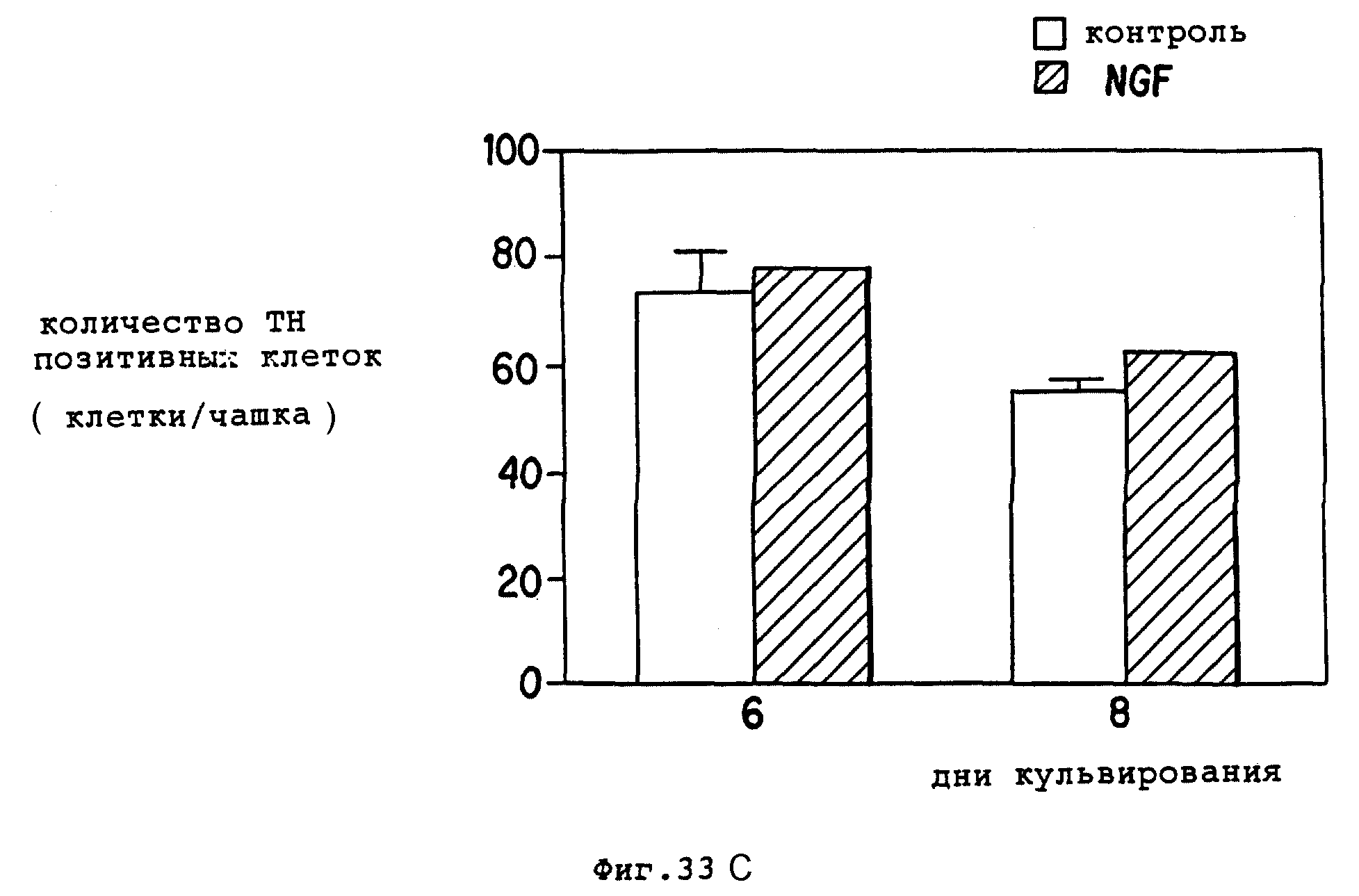
Фиг. 33 - BDNF способствует выживанию TH+, допаминергических мезенцефальных нейронов в культуре, зависимым от дозы образом, который очевиден при периодах культурирования 3 дней.
A. Сравнение количества TH+-нейронов в культурах E14 крысиных вентральных мезенцефальных клеток, поддерживаемых BDNF (сплошные линии) или без BDNF (открытые полосы). В обработанных культурах BDNF (50 нг/мл) добавляли один раз на второй день культурирования. На третий день не было обнаружено разницы между контролем и обработанными культурами, но количество TH+-нейронов в BDNF обработанных культурах было в 1,8 раз выше, чем в контроле на 8-й день. Оценки даны по среднему из двух образцов.
B. Действие BDNF на повышение выживания TH+-нейронов зависит от доз. Количество TH+-нейронов определяли в двойных культурах через 8 дней в культуре при отсутствии BDNF или же среда содержала повышающиеся концентрации BDNF, добавляемого на второй день.
C. NGF не оказал влияния на выживание TH+ -нейронов.
Для оценки специфичности BDNF культуры также выращивали в присутствии или отсутствии NGF (50 нг/мл) в течение 8 дней и анализировали на TH+-клетки, NGF добавленный на второй день, не повысил выживания TH+-нейронов в культурах, проверенных на шестой или восьмой день.
Результаты даны средние по двум чашкам.
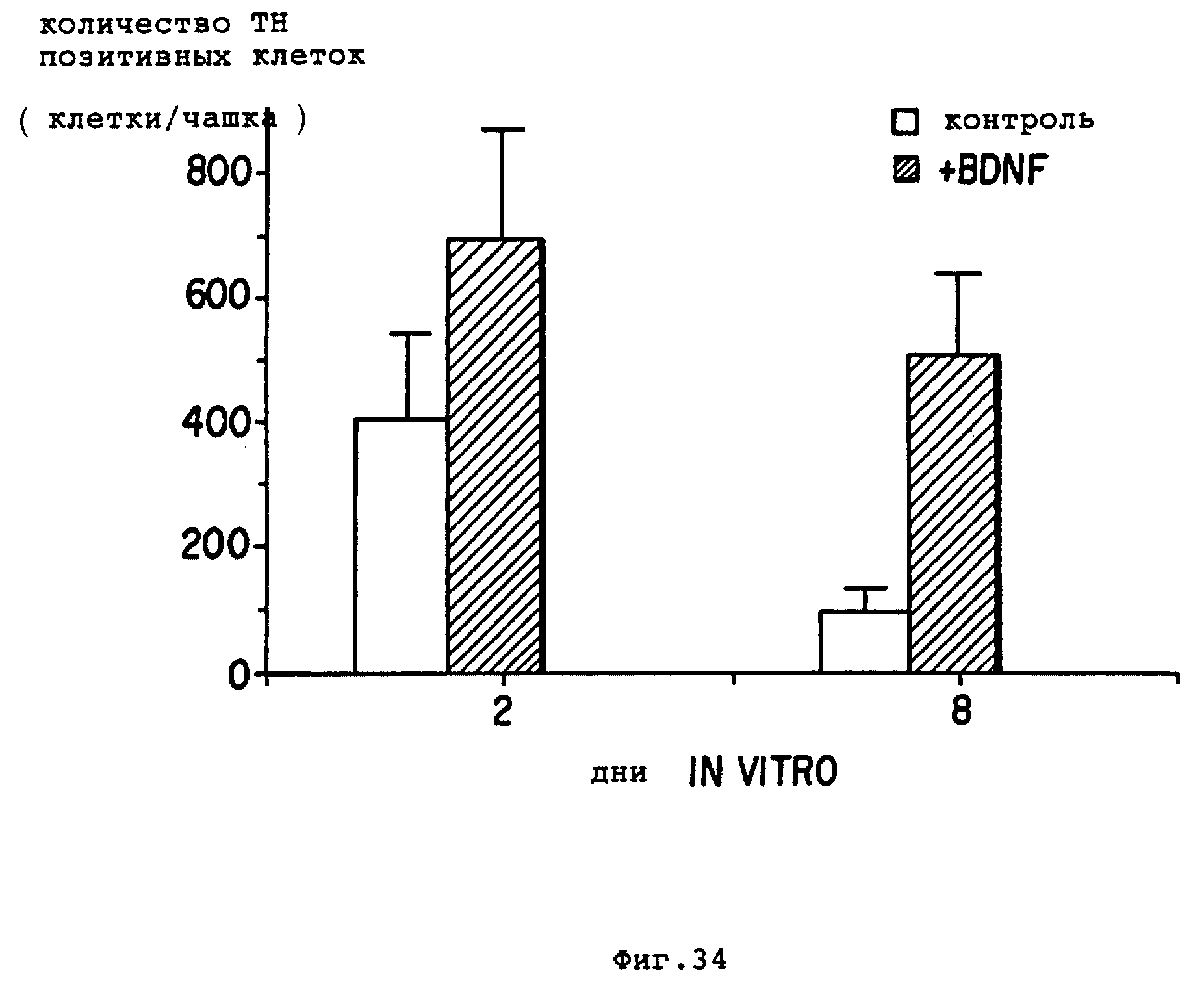
Фиг. 34 - повторные дозы BDNF еще более повышают выживание TH+ -нейронов по сравнению с единственным добавлением BDNF на второй день. Культуры приготавливали, как описано в тексте. Рекомбинантный человеческий BDNF очищали из супернатанта COS M5 клеток. Количество TH+-клеток на чашку показано для контроля (незаштрихованные полосы) и для обработанных BDNF (заштрихованные полосы) на 2-й и 8-й день ин витро.
Фиг. 35 - отсроченное добавление BDNF не увеличило количества TH+-нейронов в той же степени, как добавление BDNF на второй день.
Культуры приготавливали, как описано в тексте, за исключением времени добавления экзогенного BDNF. Все культуры были переключены на свободную от серы среду на второй день, и добавляли BDNF (50 нг/мл, BDNF свиного головного мозга) единственным добавлением либо на второй день, пятый и седьмой (СД2, СД5, СД7). На шестой, восьмой и десятый дни культивирования количество TH+-нейронов определяли в двойных культурах. В этом эксперименте, единственное добавление BDNF на второй день привело к трехкратному увеличению количества TH+ -нейронов на десятый день. Когда BDNF добавляли с задержкой, на пятый день, увеличение количества TH+-нейронов по сравнению с контролем на десятый день было менее чем двукратным, и это различие в количестве TH+-клеток между обработанными и контрольными культурами не возрастало далее с продлением культурирования. Контроль: открытые полосы, BDNF добавление на второй день - пунктир, BDNF добавление на 5 день - сплошная линия, BDNF добавления на 7 день - диагональные прерывистые линии.
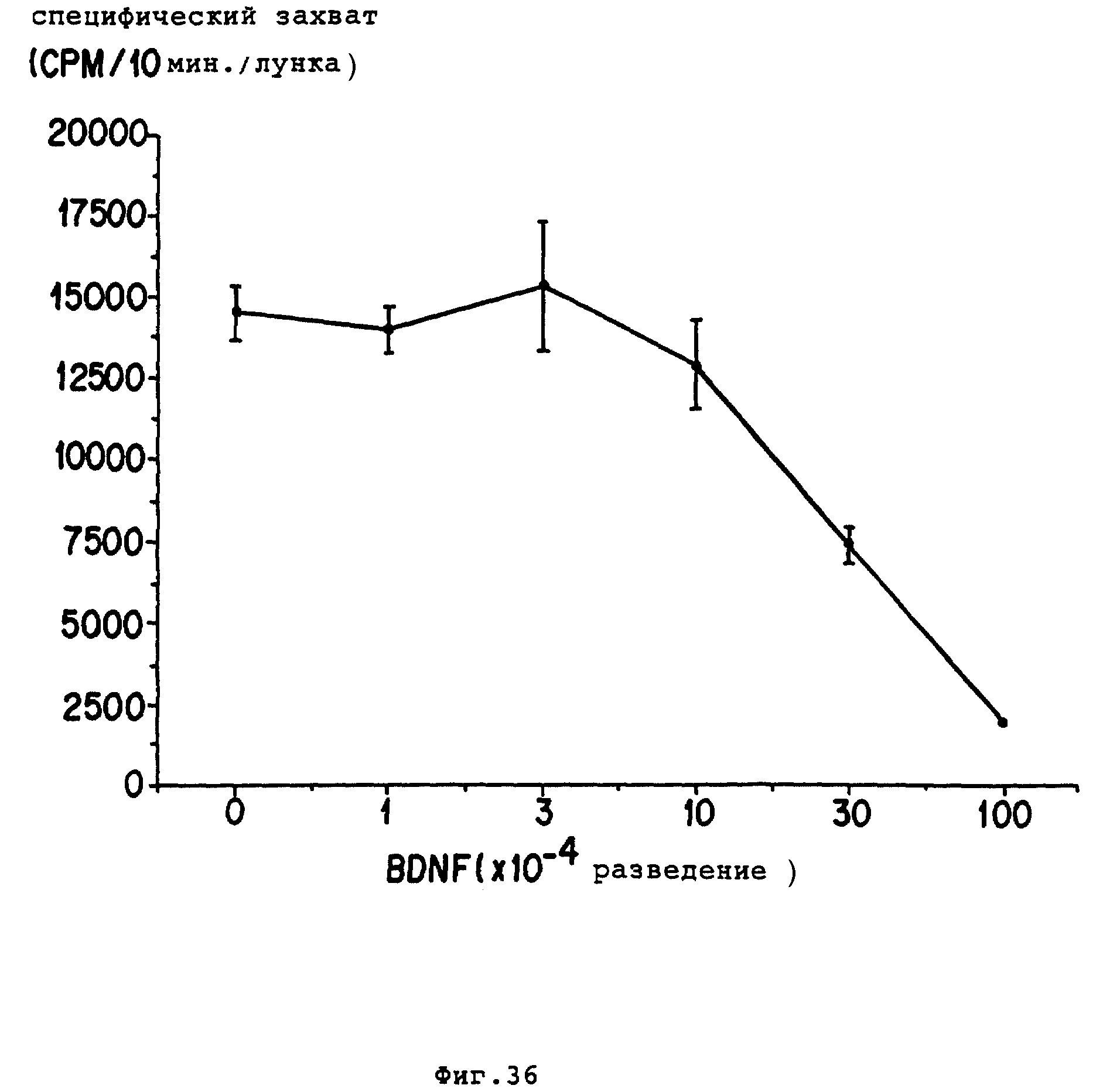
Фиг. 36 - зависимые от дозы реакции BDNF на потребление GABA высокой аффинности в культурах аммонова рога. Частично очищенный nBDNF (ротофорочищенный из COSM5 супернатантов) добавляли к культурам при различных разбавлениях. Было обнаружено, что при разведении 1 • 10-2 BDN5 проявляет максимальную активность в содействии отрастанию нейритов в E8 ганглиях дорсального корня цыпленка. Потребление3H-GABA высокой аффиности измеряли через 8 дней после обработки BDNF "ин витро".
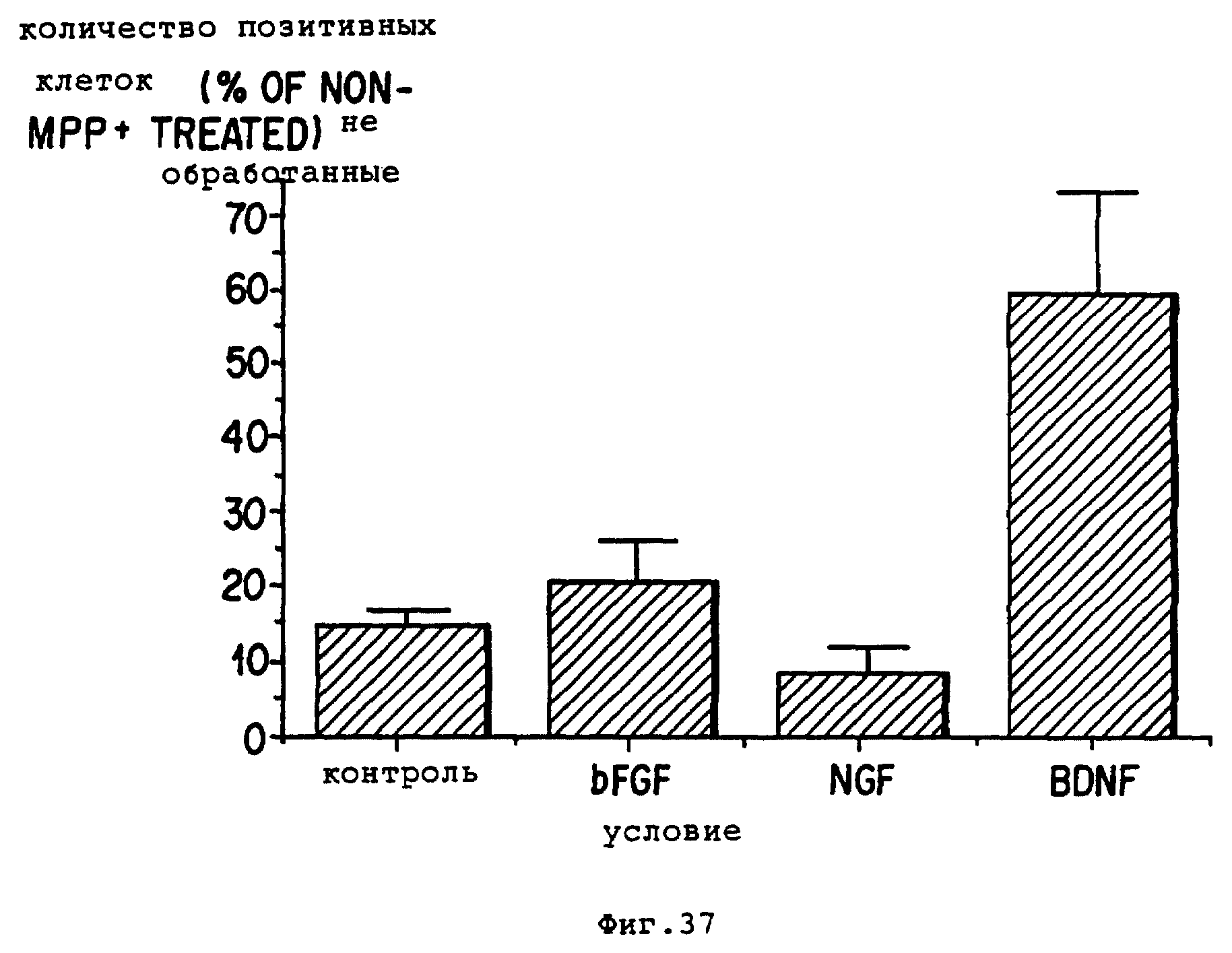
Фиг. 37 - BDNF защищает в культуре нигральные
допаминергические нейроны от нейротоксического воздействия MPP+. Культуры
создавали из E14 крысиных эмбрионов. Через 24 часа ин витро культуральную среду меняли на свободный от серы состав.
Через 3 дня в культуре, эти культуры поделили на четыре группы, по 6
35-миллиметровых чашек. Одну группу держали в качестве контроля, а другие получили:
(I) фактор роста основных фибробластов
10 нг/мл (BFGF бычьего головного мозга), Boerninger-Mannheim (II)
мышиный NGF 50 нг/мл; и (III) BDNF 50 нг/мл. Культуры держали еще 24 часа, а после этого по 3 чашки из каждой группы получали по 1 мМ
MPP+ (Rescarch Biochemicals, Inc. Natick, MA) на 48 часов. В
конце этого эксперимента, длившегося в целом 6 дней, все чашки обработали на TH иммунореактивность и в каждой группе было определено
количество TH+-клеток. Представленные данные показывают
количество TH+-нейронов после MPP+-обработки, которое было рассчитано в процентах от количества TH+
-нейронов в подобных культурах, не получивших MPP+. Все
значения представляют среднее ± SEM тройных культур.
5. Подробное описание изобретения.
Настоящее изобретение касается нуклеиновокислотных последовательностей, кодирующих мозговой нейротропный фактор (BDNF), а также BDNF-протеина, пептидных фрагментов и дериватов (производных), получаемых в большом количестве с использованием этих нуклеиновокислотных последовательностей. Кроме того, изобретение касается фармакологических композиций и терапевтических применений BDNF, впервые обеспечивая средства для генерирования достаточных количеств в основном чистого BDNF для клинического применения. Изобретение также относится к антителам, направленным против BDNF или его фрагментов, обеспечивая способ генерирования достаточного количества иммуногена. Далее, благодаря обеспечению возможности сравнить нуклеиновокислотные последовательности BDNF и NGF настоящее изобретение позволяет провести идентификацию гомологичных областей нуклеиновокислотых последовательностей BDNF и NGF, определяя тем самым генное семейство BDNF/NGF, изобретение обеспечивает способ идентифицирования и выделения дополнительных членов этого генного семейства.
Для большей ясности описания, но не с целью ограничения объема изобретения оно далее описывается в следующих разделах:
(I) очистка BDNF,
(II) BDNF-биоэксперименты,
(III) микросеквенирование BDNF-протеина,
(IV) клонирование ДНК, кодирующей BDNF,
(V) экспрессия BDNF,
(VI)
BDNF-гены и протеины,
(VII) генерирование
антиBDNF-антител,
(VIII) идентифицирование дополнительных членов генного семейства BDNF/NGF,
(IX) полезность (возможности
применения) изобретения,
(X) фармацевтические
композиции.
5.1. Очистка мозгового нейротропного фактора
Для идентифицирования BDNF-кодирующей нуклеиновой кислоты
микрограммы BDNF-протеина могут быть получены из ткани для
обеспечения возможности определения аминокислотных последовательностей, которые затем могут быть использованы для создания олигонуклеотидных
зондов. Крайняя редкость BDNF-протеина практически
ограничивает количество аминокислотной последовательности, которое может быть определено. BDNF может быть приготовлен из свиного мозга способами,
описанными у Барда и др. (1982, ЕМВО, 1:549-553) или
у Хофера и Барда (1988, Nature, 331: 261-262).
Предпочтительно BDNF может быть приготовлен согласно нижеследующему подробному описанию метода, который представлен в качестве примера, а не для ограничения объема изобретения; различные модификации могут быть использованы специалистом в данной области. Вследствие редкости BDNF в процессе очистки предпочтительно использовать примерно шесть килограммов мозговой ткани. Мозговая ткань может быть гомогенизирована в натрий-фосфатном буфере при концентрации натрия фосфата 0,2 М и pH примерно 6, содержащем 1 мМ ЭДТКи 1 мМ свежедобавленного фенилметансульфонил-фторида, так что соотношение мозговой ткани к жидкости составляет примерно 1 кг мозга к 2 литрам жидкости. Затем может быть использована соляная кислота для того, чтобы довести pH смеси до примерно 4, и смесь затем может перемешиваться примерно в течение 2 часов при 4oC. Смесь затем можно центрифугировать в течение 25 минут при 20000g. После центрифугирования собирают супернатант, доводят его до pH примерно 6 с использованием натрия гидроксида, и затем перемешивают с 1 литром предварительно набухшей карбоксиметилцеллюлозы (на 6 кг мозговой ткани), которую уравновешивают 0,1 М раствором натрия фосфата до pH 6. После нескольких промывок в целом примерно 20 литрами 0,1 М натрия фосфата, pH 6, кашицу выливают в колонну и промывают тем же буфером, содержащим 0,13 М NaCl, предпочтительно до утра. Активные фракции, идентифицированные посредством чувствительного к BDNF биоэксперимента (см. раздел 5,2) М NaCl и в последующем повергаться диализу с несколькими сменами примерно 5 литров 5 мМ калия фосфата при pH 6,8. Диализованные фракции затем могут быть помещены в гидроксиапатитовую колонну, имеющую объем слоя около 20 мл на каждый килограмм исходной мозговой ткани; гидроксиапатитовая колонна должна быть предварительно уравновешена 5 мМ калия фосфатом с pH 6,8 перед закладкой образца. Эта колонна затем может быть элюирована с линейным градиентом от 500 мл 5 мМ калия фосфата до 500 мл 700 мМ калия фосфата, оба с pH около 6,8. BDNF-активность оптимально элюируется примерно при 500 мМ концентрации калия фосфата. Полученные активные фракции могут быть затем доведены до конечной молярности порядка 700 мМ калия фосфата и помещены в фенил-сефарозную колонну (с объемом слоя около 5 мл на каждые 6 кг исходной мозговой ткани), уравновешенную раствором 700 мМ калия фосфата с pH 6,8. После промывки примерно 40 мл тем же буфером BDNF-активность может быть элюирована 0,1 М калия фосфатом, pH 6,8, и затем BDNF-активность может быть диализирована в дистиллированной воде, а затем лиофилизирована. Лиофилизированный материал затем может быть помещен в буфер для SDS-гелевого электрофореза образца, содержащего 0,1% SDS, но предпочтительно не содержащий меркаптоэтанола, и затем помещен в SDS-гель, с градиентом (линейным) акриламида от 10 до 25%. После завершения электрофоретического разделения, гель может быть окрашен в течение примерно 10 минут COOMASSIE BLUE, а затем обесцвечивают примерно 20 минут. Полоса, мигрирующая на уровне цитохромного-с-маркера может вырезана и электрофоретически элюирована из геля. SDS может быть удален в основном как описано Вебером и Кутером (Weber и Kuter, 1971, J. Biol. Chem. 246: 4504-4509). Следует заметить что удаление SDS в общем происходит не полностью. Процесс очистки BDNF для микросеквенирования был модифицирован согласно Хоферу и Барду (1988, Nature, 331: 261-262), так что он не использует на последней стадии очистки гелевый электрофорез.
5.2. Биоисследования мозгового нейротропного фактора.
Согласно настоящему изобретению может быть использована любая система, которая качественно или количественно обнаруживает BDNF-активность. Такие биоэксперименты могут использоваться для идентифицирования или/и измерения активности природного или рекомбинантного BDNF.
Любое биоисследование на BDNF, известное по уровню техники, может быть здесь использовано. Например, DRG нейроны (ганглия дорсального корня куриного эмбриона) могут быть использованы в таком BDNF-исследовании, как описано у Барда и др. (1980, Proc. Nath. Acad. Sci. США, 77: 1199-1203).
В частности, могут быть собраны ганглии дорсального корня 6-14-дневных куриных эмбрионов, предпочтительно 10-12-дневных, с использованием диссекции, известной в этой области, и немедленно помещены в небольшой объем среды F14 (сделанной из порошка GIBCO F-12, с добавлениям по Фогелю и др. - Vogel 1972, Proc. Nath. Acad. Sci. США, 69: 3180-3184), которую следует заменять в конце диссекции на буферный, фосфатный, свободный от ионов Ca2+ и Mg2+ физиологический раствор (PBS), содержащий около 0,1% трипсина. Примерно после 20 мин инкубирования при 37oC эти ганглии могут быть центрифугированы и дважды промыты средой F14, содержащей около 10% (об./об.) инактивированной теплом лошадиной сыворотки. Эти ганглии затем могут быть диссоциированы мягким тритурированием (около 10-15 аспираций) с использованием малой, с диаметром около 1 мм, силиконизированной пипетки Пастера. Оставшиеся комочки ткани могут быть затем предпочтительно удалены путем пропускания клеточной суспензии через нейлоновое сито, имеющее поры размером около 40 мкм. Клеточную суспензию теперь можно предварительно поместить на чашку из пластиковой ткани примерно на 210 минут, во время этого периода большинство не-нейронных клеток должны прилипнуть к пластиковой поверхности чашки, оставляя обогащенную нейронами популяцию клеток в суспензии. Клетки затем можно разбавить до концентрации примерно 5 • 103 клеток на миллиметр чашечной среды (предпочтительно, среды F14 с добавлением 10 об./об.% инактивированной теплом лошадиной сыворотки вместе с антибиотиками), и их можно поместить на чашки, покрытые полиорнитиновой или полиорнитин/ламининовой тканью (см. ниже).
Альтернативно, но не для ограничения BDNF-биоэксперименты, которые относительно нечувствительны к NGF, могут при некоторых обстоятельствах превосходить системы, к которым относится описанная выше DGR-система, способная в определенных условиях дать реакцию на BDNF и на NGF. Относительно специфичная к BDNF система должна бы включать культуры ганглиев сетчатки, а также культуры нейронов, происходящих из невральных плакод.
Перинатальные клетки сетчатки могут быть культурированы согласно методам, описанным у Джонсона и др. (Johnson, 1986, J. Neurosci. 6: 3031-3038). Например, сетчатки могут удаляться у перинатальных животных (для крыс термин "перинатальный" относится к 17-му эмбриональному дню и далее до постнатальных щенков возраста 48 часов), затем сетчатку промывают в PBS и инкубируют в PBS, содержащем примерно 0,05-0,1% трипсина, примерно на 15 минут примерно при 37oC. После протеолитического переваривания сетчатки могут быть промыты в среде F14, содержащей около 10 об./об.% инактивированной теплом лошадиной сыворотки (F14 - HS). Сетчатки затем могут быть диссоциированы мягким пипеттированием в небольшом объеме (около 1-10 мл) свежей F14- HS. Недиссоциированной ткани позволяют осесть и оставшиеся клетки и среду вытягивают пипеткой для культуры.
Альтернативно, BDNF биоэксперименты по изобретению могут использовать клетки, происходящие из невральных плакод. Эксплантанты черепных нервов эмбриона, например вентролатеральной части тройничного ганглия или вестибулярного, коленчатого, височного или нодозного ганглия, могут культивироваться в коллаген-геле, покрытом культурирующей средой, как описано у Дэвиса и др. (Davies, 1986, J., Neurosci, 6: 1897-1904) или диссоциированными согласно Барду и др. (Barde, Proc. Nath. Acad. Sci. 1980, 77: 1199-1203).
Вследствие того, что было установлено, что реакция на BDNF может быть десятикратно усилена путем замены субстрата для роста - от полиорнитина на ламинин-полиорнитин (Barde и др., 1987, Prog. Brain Res. 71: 185-189), эксперименты с BDNF проводят предпочтительно на ламининсодержащих субстратах. Поверхности для культурирования могут быть приготовлены, например, путем (I) покрытия поверхности на 8-10 часов стерильным раствором полиорнитина; (II) промывкой несколько раз стерильной водой, и (III) покрытием поверхности на примерно два часа ламинином с концентрацией приблизительно 25 мкг/мл в PBS (Johnson и др., 1986, J. Neurosci, 6: 3031-3038).
В любой биоисследовательской системе по изобретению известными в этой области методами может быть установлена кривая зависимости от дозы BDNF. При использовании диссоциированных цыплячьих сенсорных ганглиев было обнаружено половинное от максимального выживание при концентрации очищенного BDNF примерно 5 нг/мл, и максимальное выживание при концентрации между 10 и 20 нг/мл очищенного BDNF (Barde и др., 1987, Prog. Brain Res. 71: 185-189).
5.3. Микросеквенирование протеина мозгового нейротропного фактора.
BDNF-протеин, приготовленный из мозга, может быть секвенирован; однако следует подчеркнуть, что крайняя редкость этого протеина не позволяет надежно получить существенную часть последовательности BDNF-протеина. Этот протеин может быть секвенирован непосредственно или вначале расщеплен любой протеазой или другим известным в этой области соединением, включая, но не ограничиваясь этим, Staphylococcus aurues V8, трипсин и цианоген-бромид. Пептиды могут быть секвенированы с помощью автоматизированной деградации Эдмана на микросеквенсере с газовой фазой, согласно методу Huvick и др. (1981, J. Biol Chem. 256: 7990-7997) и Hunkapillar и др. (1983, Methods Enzymol. 91: 227-236). Обнаружение фенилтиогидантоин-аминокислот может затем быть осуществлено согласно Lottspeich (1985, Chromatography 326: 321-327). Перекрывающие фрагменты аминокислотной последовательности могут быть определены и затем использованы для выведения более длинных отрезков смежной последовательности.
5.4. Клонирование ДНК, кодирующей мозговой нейроптропный фактор.
Редкость BDNF сильно препятствует использованию стандартных приемов клонирования гена BDNF. Например, если имеющаяся белковая последовательность была использована для генерирования комплементарно меченного олигонуклеотидного зонда, а этот зонд был использован для скрининга библиотек кДНК, полученных из тканей, которые предположительно синтезируют BDNF, то количество положительных клонов будет, вероятно, непостижимо малым. Настоящее изобретение обеспечивает клонирование гена BDNF комбинацией приемов, включая очистку подходящего количества протеинов BDNF, микросеквенирование протеина BDNF, получение олигонуклеотидного зонда, создание кДНК-библиотеки, амплификацию, основанную на полученной аминокислотной последовательности BDNF и, наконец, селекцию на ген BDNF. Этот способ по изобретению в предпочтительном варианте предусматривает амплифицирование последовательностей нуклеиновых кислот, полученных из ткани, посредством цепной реакции полимеразы (PCR) (Saiki и др., 1985, Science 230: 1350-1354) для того, чтобы расширить количество BDNF-последовательностей, имеющихся для клонирования. Ниже приведено детальное описание этого способа.
Прежде всего, аминокислотная последовательность, полученная из очищенного BDNF-протеина, может быть использована для дедукции олигонуклеотидных праймеров для их использования в PCR-реакции. Вследствие дегенерации генетического кода, в котором несколько нуклеиновокислотных триплетов могут означать одну и ту же аминокислоту, несколько олигонуклеотидов следует синтезировать для данной аминокислотной последовательности, чтобы обеспечивать множество потенциальных комбинаций нуклеотидных последовательностей; полученные олигонуклеотиды упоминаются как дегенерированные праймеры.
Цепная реакция полимеразы требует наличия как праймера со смысловой нитью, так и антисмыслового праймера. Соответственно, аминокислотная последовательность может быть использована в качестве праймера для одной нити ДНК, а второй праймер, гомологичный обычно встречающейся последовательности ДНК, такой как отрезок тимидиновых остатков, полученных реверс-транскрипцией полиаденозиновых хвостов мРНК, может использоваться как праймер для второй нити ДНК. Эти праймеры затем могут быть использованы в цепной реакции полимеразы с нуклеиновокислотной матрицей, которая предположительно содержит кодирующие BDNF-последовательности, такой как геномная ДНК или предпочтительно кДНК, приготовленная из мРНК, собранных из ткани, предположительно синтезирующей BDNF. Продукты ДНК-реакции могут затем анализироваться электрофорезом для того, чтобы определить, имеет ли продукт ДНК-реакции молекулярный размер, сходный с ожидаемым размером гена BDNF и предпочтительно нуклеотидным секвенированием.
Однако вследствие использования двух дегенерированных праймеров в PCR, которая повышает вероятность амплифицирования нуклеиновокислотных последовательностей, которые не кодируют BDNF, предпочтительный вариант способа предусматривает использование только одного дегенерированного праймера, а другой праймер при этом соответствует точной BDNF-последовательности. Чтобы идентифицировать точную BDNF-последовательность, аминокислотная последовательность, определенная с использованием очищенного протеина BDNF, может быть использована для конструирования обоих дегенерированных праймеров - как смыслового, так и антисмыслового. Продукт ДНК-реакции, получаемый от использования этих праймеров в PCR-реакции, используя в качестве матрицы кодирующую BDNF нуклеиновую кислоту, будет фрагментом нуклеиновой кислоты, кодирующим аминокислотный отрезок, используемый для создания праймеров, и он может быть предсказуемого размера (т.е. как минимум, количеством основных пар по длине, определяемым количеством аминокислотных остатков, помноженным на три). Результаты секвенирования продукта ДНК-реакции могут быть сравнены с подтвержденной аминокислотной последовательностью, чтобы подтвердить, что амплифицированная нуклеиновокислотная последовательность может в действительности кодировать BDNF пептидный фрагмент. Хотя может быть использован любой способ нуклеиновокислотного секвенирования, известный в данной области, предпочтительно метод дидеоксинуклеотидной терминации цепи (Sanger и др. , 1979, Proc. Nath. Acad. Sci. США, 72: 3918-3921). Секвенирование может быть завершено с использованием гель-очищенного или предпочтительно клонированного продукта ДНК реакции. Последовательность продукта ДНК реакции может быть затем использована для создания олигонуклеотидного праймера, соответствующего точной последовательности, кодирующей BDNF. Этот праймер может быть затем использован вместе со вторым праймером, который может быть дегенерированным, для расширения количества BDNF-последовательности сверх того, что представлено вначале определенным фрагментом точной последовательности. Например, но не для ограничения, смысловой праймер может соответствовать точной нуклеотидной последовательности BDNF, тогда как антисмысловой праймер может быть дегенерированным праймером, гомологичным области последовательности, которая находится ниже секвенированного фрагмента, например полиаденозиновой части мРНК, как обратно считанной в кДНК. Затем может возникнуть необходимость использовать аналогичный способ, чтобы восстановить последовательность, выше секвенированного фрагмента; например антисмысловой праймер может соответствовать точной нуклеотидной последовательности BDNF, а смысловой праймер может быть дегенерированным праймером, гомологичным области последовательности BDNF выше секвенированного фрагмента, например 5' полиаденозиновый конец может быть добавлен к 5'-концу кДНК с использованием терминальной диоксинуклеотид-трансферазы. Соответственно последовательность всего гена BDNF или мРНК также могут быть получены таким синтезом.
Продукты реакции ДНК могут клонироваться с использованием любого известного в этой области способа. Может быть использовано большое количество известных систем вектор - хозяин. Возможные векторы включают, не с целью ограничения космиды, плазмиды или модифицированные вирусы, и векторная система должна быть совместима с клеткой-хозяином. Такими векторами являются, например, но не для ограничения, бактериофаги, такие как производные лямбды, или плазмиды, такие как pBR322, pUC или Bluescript (Стратаген)-плазмидные производные. Рекомбинантные молекулы могут быть введены в клетку-хозяина путем трансформации, трансфекции, инфекции, электропрорации и т.д.
Ген BDNF вставляется в клонирующий вектор, который может быть использован для трансформации, трансфекции или инфекции соответствующих клеток-хозяев так, чтобы генерировалось множество копий генных последовательностей. Это может быть осуществлено путем лигирования фрагмента ДНК в клонирующий вектор, который имеет комплементарные липкие концы. Однако, если комплементарные сайты рестрикции, используемые для фрагментирования ДНК, не присутствуют в клонирующем векторе, то концы молекул ДНК могут быть модифицированы энзиматически. Может оказаться выгодным вставить сайты рестрикции эндонуклеазного расщепления в олигонуклеотидные праймеры, используемые в цепной реакции полимеризации для того, чтобы облегчить инсерцию в векторы. Альтернативно любой желаемый сайт может быть получен путем лигирования нуклеотидных последовательностей (линкеров) к концам ДНК; эти лигированные линкеры могут включать специфичные, синтезированные химически олигонуклеотиды, кодирующие последовательности узнавания рестрикционных эндонуклеаз. В альтернативном выполнении способа расщепленный вектор и ген BDHF могут быть модифицированы посредством гомополимерного наращивания.
В специфических выполнениях трансформирование клеток-хозяев рекомбинантными молекулами ДНК, которые внедряют выделенный ген BDHF, кДНК или синтезированная последовательность ДНК позволяет генерирование множественных копий гена. Таким образом, этот ген может быть получен в больших количествах посредством выращивания трансформантов, выделения рекомбинантных молекул ДНК из этих трансформантов и, если необходимо, восстановление вставленного гена из выделенной рекомбинантной ДНК.
Согласно предпочтительному выполнению изобретения, когда уже генерирован происходящий от кДНК клон, кодирующий BDHF, геномный клон, кодирующий BDHF, может быть выделен с использованием стандартной, известной технологии. Например, меченый нуклеотиновокислотный зонд может быть получен от BDHF клона и использован для скрининга библиотеки геномных ДНК нуклеиновокислотной гибридизацией с использованием, например, способа Benton и Davies (1977, Science, 196: 180), примененного ими для библиотеки бактериофагов, и способа Grunstein и Hogness (1975, Proc. Nath. Acad. Sci. США, 72: 3961-3965) для плазмидных библиотек. Восстановленные клоны могут затем анализироваться картированием рестрикционных фрагментов и секвенированием согласно хорошо известным методам.
Далее из кДНК-библиотеки могут идентифицироваться дополнительные кДНК-клоны с использованием последовательностей, полученных по изобретению.
5.5. Экспрессия гена мозгового нейротропного фактора.
Нуклеотидная последовательность, кодирующая протеин BDNF или его часть, может быть вставлена в подходящий экспрессионный вектор, т.е. вектор, который содержит необходимые элементы для транскрипции и трансляции вставленной протеинкодирующей последовательности. Необходимые транскрипционные и трансляционные сигналы могут быть также предоставлены природным геном BDNF и/или его фланкирующими областями. Множество систем вектор - хозяин может быть использовано для экспрессии протеинкодирующей последовательности. В эти системы входят, например, но не с целью ограничения, системы клеток млекопитающих, инфицированные вирусом (например, вирусом коровьей оспы, аденовирусом и т.д.); системы клеток насекомых, инфицированных вирусом (например, бакуловирусом); микроорганизмы, такие как дрожжи, содержащие дрожжевые векторы, или бактерии, трансформированные бактериофаговой ДНК, плазмидной ДНК или космидной ДНК. Экспрессионные элементы этих векторов различаются по своей силе и специфичности. В зависимости от используемой системы вектор - хозяин может быть использован любой из многих подходящих транскрипционных и трансляционных элементов.
Любой из ранее описанных методов для инсерции ДНК-фрагментов в вектор может быть использован для конструирования экспрессионных векторов, содержащих химерный ген, состоящий из подходящих сигналов, управляющих транскрипцией (трансляцией) и кодирующей протеин последовательности. Эти способы могут включать рекомбинантную ин витро ДНК и технологию синтеза, а также рекомбинации ин виво (генетическая рекомбинация).
Экспрессия нуклеиновокислотной последовательности, кодирующей BDNF-протеин или пептидный фрагмент, может регулироваться второй нуклеотиновокислотной последовательностью так, что BDNF-протеин или пептид экспрессируется в хозяине, трансформированном рекомбинантной молекулярной ДНК. Например, экспрессия BDNF может управляться любым промоторным/энтахансерным элементом, известным в этой области. Промоторы, которые могут быть использованы для управления BDNF-экспрессией, включают, например, но не с целью ограничения, раннюю промоторную область SV40 (Bernoist и Chambon, 1981, Nature, 290: 304-310), промотор, содержащий в 3' длинном конце повторение вируса саркомы Рауса (Rous) (Yamamoto и др., 1980, Cell, 22: 787-797), промотор тимидин киназы герпеса (Wagner и др., 1981, Proc. Nath. Acad. Sci США, 78: 1440-1445), регуляторные последовательности гена металлотионина (Brinster и др., Nature, 296: 39-42); прокариотические векторы экспрессии, такие как бета-лактамазы промотор (Villa-Kamaroff и др., 1978, Proc. Nath. Acad. Sci. США, 75: 3727-3731), или tac-промотор (DeBoer и др., 1983, Proc. Natl. Acad. Sci. США, 80: 21-25), смотри также в "Сайентифик Американ" статью "Useful proteins from reconebinant bacteria" (S.A., 1980, 242: 74-94), растительные векторы экспрессии, включающие промоторную область нопалин-синтетазы (Herrera-Estrella и др., Nature, 303: 209-213) или промотор 35S РНК вируса мозаичной болезни цветной капусты (Gardner и др., 1981, Nucl. Acids Res. 9: 2871) и промотор для фотосинтезового энзима рибулозо-бифосфат-карбоксилаза (Herrera-Estrella, 1984, Nature, 310: 115-120), промоторные элементы из дрожжей или других грибков, таких как Gal4 промотор, ADC (алкогольдегидрогеназа) промотор, PGK (фосфоглицеринкиназа) промотор, промотор щелочной фосфатазы, и следующие области, управляющие транскрипцией у животных, которые проявляют тканевую специфичность и использовались в трансгенных животных: контролирующая область гена эластазы-1, активная в панкреатических ацинарных клетках (Swift и др., 1984, Cell, 38: 639-646; Ornitz и др. , 1986, Cold Spring Harbor Symp. Quant. Biol 50: 399-409; MacDonald, 1987, Hepatology, 7: 425-515); контролирующая область гена инсулина, которая активна в панкреатических бета-клетках (Hanahan, 1985, Nature 315: 115-122), контролирующая область гена иммуноглобулина, которая активна в лимфоидных клетках (Grosschedl и др., 1984, Cell, 38: 647-658; Adams и др., 1985, Nature 318: 533-538; Alexander и др., 1987, Mol. Cell. Biol 7: 1436-1444), контролирующая область мышиного вируса опухоли молочной железы, которая активна в семенниках, груди, лимфоидных образованиях и помете (Leder и др., 1986, Cell, 45: 485-495), контролирующая область гена альбумина, которая активна в печени (Pinkert и др., 1987, Genes and Devel, 1: 268-276), контрольная область альфа-фетопротеинового гена, которая активна в печени (Krumlauf и др., 1985, Mol. Cell. Biol 5: 1639-1648; Наммег и др., 1987, Science 235: 53-58); область управления гена альфа-1-антитрипсина, которая активна в печени (Kelsey и др., 1987, Genes and Develop 1: 161-171), управляющая область гена бета-глобина), которая активна в миелоидных клетках (Morgan и др., 1985, Nature 315: 338-340; Kollias и др., 1986, Cell, 46: 89-94); область управления геном миелиносновного протеина, которая активна в олигодендроцитах головного мозга (Readhead и др., 1987, Cell, 48: 703-712); область управления геном миозиналегкой цепи-2, которая активна в скелетных мышцах (Sani, 1985, Nature 314: 283-286), и контрольная область гена, гонадотропного, освобождающего гормон, которая активна в гипоталамусе (Mason и др., 1986, Science 234: 1372-1378).
Экспрессионные векторы, содержащие инсекции гена BDNF могут быть идентифицированы тремя основными подходами: (a) гибридизацией ДНК-ДНК, (b) присутствием или отсутствием функций "маркерного" гена, и (c) экспрессией вставленных последовательностей. При первом подходе присутствие чужеродного гена, инсертированного в эксперссионный вектор, может быть обнаружено путем гибридизации ДНК-ДНК с использованием зондов, содержащих последовательности, которые гомологичны инсертированному гену BDNF. При втором подходе система рекомбинантный вектор - хозяин может быть идентифицирована и подвергнута селекции на основе присутствия или отсутствия определенных функций "маркерного" гена (например, тимидинкиназная активность, резистивность к антибиотикам, трансформационный фенотип, образование окклюзивных тел у бакуловирусов и т. д. ), вызванных инсерцией чужеродных генов в вектор. Например, если ген BDNF инсертирован внутри маркерного гена, в его последовательности в векторе, то рекомбинанты, содержащие инсерт BDNF, могут быть идентифицированы посредством отсутствия функции маркерного гена. В третьем подходе рекомбинантные экспрессионные векторы могут быть идентифицированы путем экспериментального исследования продукта чужеродного гена, экспрессируемого рекомбинантом. Такие исследования могут быть основаны, например, на физических или функциональных свойствах продукта BDNF-гена в биоэкспериментальной системе, как описано выше, в разделе 5.2.
Когда конкретная рекомбинантная молекула ДНК идентифицирована и выделена, для ее размножения могут быть использованы несколько известных способов. Когда созданы подходящая система хозяина и условия роста, то экспрессионные векторы могут быть размножены и получены в большом количестве. Как ранее объяснялось, могут быть использованы экспрессионные векторы, например, но не с целью ограничения, следующего типа: человеческие или животные вирусы, такие как вирус коровьей оспы или аденовирус; вирусы насекомых, такие как бакуловирусы; дрожжевые векторы; бактериофаговые векторы (например, лямбда); и векторы плазмидной и космидной ДНК, и это далеко не все.
Дополнительно, штамм клеток-хозяев может быть выбран таким, что он модулирует экспрессию инсертированных последовательностей, или модифицирует и продуцирует генный продукт желаемым специфичным образом. Экспрессия от определенных промоторов может быть усилена в присутствии определенных индукторов; так, можно управлять экспрессией полученного генной инженерией BDNF-протеина. Кроме того, различные клеточные хозяева обладают характерными и специфичными механизмами для трансляционного и посттрансляционного процессинга и модифицирования (например, гликозилированием, расщеплением) белков. Могут быть выбраны подходящие клеточные линии или системы хозяев для обеспечения желаемой модификации и процессинга экспрессируемого чужеродного белка. Например, для производства продукта в виде негликозилированного сердцевинного белка может использоваться экспрессия в бактериальной системе. Экспрессия в дрожжах даст гликозилированный продукт. Экспрессия в клетках млекопитающих может быть использована для обеспечения "нативной" гликозиляции гетерологичного белка BDNF. Кроме того, различные экспрессионные системы вектор - хозяин могут влиять на реакции процессинга, такие как протеолитическое расщепление, до различной степени.
В специфическом выполнении изобретения ДНК, кодирующая prepro BDNF может клонироваться в плазмиду рСМУ, амплифицироваться и затем использоваться для трансфекции COS-клеток по кальциево-фосфатному методу (Chen и Okayama, 1987, Mol. Cell. Biol, 7: 2745-2752); BDNF-активность затем может быть собрана из культуральной клеточной среды (см. пример раздела 10, ниже).
5.5.1. Идентифицирование и очистка экспрессированного продукта.
Когда идентифицирован рекомбинант, который экспрессирует ген BDNF, генный продукт следует проанализировать. Это может быть осуществлено экспериментами на основе физических или функциональных свойств продукта.
Когда идентифицирован белок BDNF, он может быть выделен и очищен стандартными методами, предусматривающими хроматографию (например, ионный обмен, аффинность и фильтрационная колоночная хроматография), центрифугирование, дифференциальную растворимость или любой другой стандартный метод для очистки протеинов. Функциональные свойства могут быть оценены и использованием любого известного эксперимента на BDNF, включая, например, но не с целью ограничения, использование ганглиев дорсального корня куриного эмбриона, клетки сетчатки перинатальной крысы или нейроны, происходящие от невральных плакод.
Важно, что способы, используемые для приготовления BDNF из ткани головного мозга, продуцируют BDNF, который не полностью активен из-за присутствия остаточного SDS, поскольку эти способы включают на последней стадии гелевый электрофорез с SDS (Barde и Thoenen и др., 1985, в "Hormones and Cells Regulation", т. 9, из. Дюмон и др. Элсвир Сайенс паблишерз, стр. 385-390). В отличие от известных способов настоящее изобретение позволяет выделять BDNF, который продуцируется от рекомбинантных нуклеиновокислотных молекул и который свободен от SDS и поэтому обладает полной активностью. Например, но не с целью ограничения, антиBDNF-антитела по изобретению (такие как антитела, направленные против B5-33 аминокислотного фрагмента свиного BDNF, описанные в разделе 11, ниже) могут быть использованы для собирания рекомбинантного BDNF иммунопреципитацией или аффинной хроматографией, продуцируя при этом свободные от детергента, полностью активные молекулы BDNF.
В другом выполнении изобретения prepro BDNF может быть конвертирован в активный зрелый BDNF энзиматически с использованием, например, эндопротеиназы Arg-C (см. пример, раздел 17, ниже).
5.6. Гены и белки мозгового нейротропного фактора.
С использованием описанных выше методов (а также см. примеры разделов 6 и 9, ниже) определяли соответствующие нуклеиновокислотные последовательности и дедукцией получали соответствующие аминокислотные последовательности. Определялась кДНК-последовательность свиного BDNF, которая изображена на фиг. 1. Была определена BDNF-последовательность геномной ДНК человека, которая изображена на фиг. 5, на которой представлены также последовательности ДНК от свиньи, крысы и цыпленка. Каждая из этих последовательностей или их функциональные эквиваленты могут быть использованы в соответствии с изобретением.
Дополнительно, изобретение относится к генам BDNF и его белкам, выделенным от свиных, бычьих, лошадиных, птичьих, кошачьих или собачьих, так же как и приматных источников и любых других видов животных, у которых существует BDNF-активность. Изобретение далее направлено на субпоследовательности BDNF нуклеиновых кислот, включающие по меньшей мере десять нуклеотидов, причем эти субпоследовательности содержат гибридизируемые части BDNF-последовательности, которые использованы, например, в экспериментах по нуклеиновокислотной гибридизации, анализах Саузерн- и назерн-блоттингом и т. д. Изобретение также обеспечивает BDNF-протеин, а также его фрагменты и производные, согласно аминокислотным последовательностям, представленным на фиг. 1 и 5, или их функциональным эквивалентам. Изобретение также обеспечивает фрагменты или производные BDNF-протеинов, которые включают антигенный детерминант (или детерминанты), или которые являются функционально активными. Как здесь использовано, выражение "функционально активный" означает обладание положительной активностью в опытах на известные функции BDNF, например в опытах на DRG куриного эмбриона.
Например, нуклеиновокислотные последовательности, описанные на фиг. 1 и 5, могут быть изменены замещениями, добавлениями или делециями, которые обеспечивают функционально эквивалентные молекулы. Вследствие дегенерации нуклеотидных кодирующих последовательностей на практике по изобретению могут быть использованы другие последовательности ДНК, которые кодируют в основном те же самые аминокислотые последовательности, как изображено на фиг. 1 и 5. Они включают, например, но не с целью ограничения, нуклеотидные последовательности, включающие целиком или часть BDNF-генов, изображенных на фиг. 1 и 5, которые изменены замещением различных кодонов, которые кодируют функционально эквивалентный аминокислотный остаток внутри этой последовательности, продуцируя таким образом молчащую мутацию. Аналогично BDNF-протеины или их фрагменты и производные по изобретению включают, например, но не с целью ограничения, те из них, которые содержат, как первичная аминокислотная последовательность, целиком или часть аминокислотной последовательности, в основном, как изображено на фиг. 1 и 5, включая измененные последовательности, у которых внутри последовательности одни аминокислотные остатки заменены функционально эквивалентными аминокислотными остатками, приводящими к молчащей мутации. Например, одна или более аминокислот могут быть заменены другими аминокислотами сходной полярности, которые действуют как функциональный эквивалент, приводя к молчащей мутации. Замещения аминокислоты внутри последовательности может быть выбрано из других членов того класса, к которому принадлежит заменяемая аминокислота. Например, неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин. Нейтральные полярные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глютамин. Положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин. Отрицательно заряженные (кислые) аминокислоты включают аспартовую кислоту и глютамовую кислоту. В объем изобретения также входят BDNF-протеины или их фрагменты и производные, которые дифференциально модифицированы во время или после транслирования, например при гликозилировании, протеолиитческом расщеплении, лигировании к молекуле антитела или другому клеточному лиганду и т.д.
Дополнительно данный BDNF может мутировать ин витро или ин виво, для создания и/или разрушения последовательностей трансляции, инициации и/или терминирования, или для создания вариаций в кодирующих областях и/или для формирования новых сайтов рестрикции эндонуклеазами, или для разрушения существующих рестрикционных сайтов, чтобы облегчить дальнейшую модификацию ин витро. Может быть использована любая известная специалисту технология мутагенеза, включая, например, но не с целью ограничения ин витро сайт-направленный мутагенез (Hutchinson и др., 1978, J. Biol. Chem 253: 6551), использование TAB® линкеров ("Фармация") и т.д.
5.7. Генерирование антител, направленных против мозгового нейротропного фактора.
Согласно изобретению BDNF-протеин или его фрагменты и производные могут быть использованы в качестве иммуногена для генерирования антиBDNF-антител. Предыдущие попытки продуцирования антиBDNF-иммунного ответа были безуспешными, возможно вследствие того, что имелись малые количества очищенного BDNF. Путем обеспечения производства относительно больших количеств BDNF-протеина с использованием рекомбинантной технологии для протеинового синтеза (основанной на BDNF нуклеионовокислотных последовательностях по изобретению), теперь решена проблема получения достаточных количеств BDNF.
Для дальнейшего увеличения вероятности производства иммунной реакции против BDNF аминокислотная последовательность BDNF может быть проанализирована, чтобы идентифицировать те части молекулы, которые могут быть связаны с повышенной способностью к иммунизации. Например, аминокислотная последовательность может быть подвергнута компьютерному анализу для идентифицирования эпитопов поверхности по методу Хоппа и Вудса (Hopp и Woods, 1981, Proc. Nath. Acad. Sci. США, 78: 3824-3828), который был успешно применен для идентифицирования антигенных пептидов поверхностного антигена гепатита B, альтернативно могут сравниваться выведенные аминокислотные последовательности BDNF от различных видов животных и идентифицироваться относительно негомологичные области; эти негомологичные области будут скорее всего иммуногенными у разных видов животных.
Для приготовления моноклональных антител, направленных против BDNF, может быть использована любая технология, которая обеспечивает производство молекул антител посредством устойчивых в культуре клеточных линий. Например, в объем настоящего изобретения входят: технология гибридомы, разработанная впервые Kohler и Milstein (1975, Nature, 256: 495-497), технология триомы - человеческой B-клеточной гибридомы (Kozbor и др., 1983, Immunology Today 4: 72) и технология EBV-гибридомы для получения человеческих моноклональных антител (Cole и др., 1985, в "Monoclonal Antibodies and Cancer Therapy", изд. Алан Р. Лисс, Инк., стр. 77 - 96) и им подобные.
Моноклональными антителами для терапевтического применения могут быть человеческие моноклональные антитела или химерные человеко-мышиные (или от других видов животных) моноклональные антитела. Человеческие моноклональные антитела могут быть получены любой из многочисленных технологий, известных специалисту (например, Teng и др., 1983, Proc. Nath. Acad. Sci. США, 80: 7308 - 7312; Kazbor и др., 1983, Immunology today 4: 72- 79; Olsson и др., 1982, Meth. Enzymol, 92 : 3 - 16). Химерные молекулы антител могут быть приготовлены с содержанием мышиного антигенсвязывающего домена с человеческими постоянными областями (Morrison и др., 1984, Proc. Nath. Acad. Sci. США, 81: 6851; Takeda и др., 1985, Nature, 314: 452).
Различные известные специалисту приемы могут быть использованы для производства поликлональных антител к эпитопам BDNF. Для производства антител, различные животные-хозяева могут быть иммунизированы путем инъекции BDNF-протеина, или его фрагмента, или производного, включая, например, но не с целью ограничения, такие виды, как кролики, мыши и крысы и т.д. Могут быть использованы различные адъюванты для усиления иммунологической реакции, зависящей от вида хозяина, и включающие, например, но не с целью ограничения, адъювант Фрейнда (Freund) полный или неполный, минеральные гели, такие как гидроксид алюминия; поверхностно-активные субстанции, такие как изолецитин, плюроновые полиолы, полианионы, пептиды, масляные эмульсии, "heyhole limpet" изоцианин, динитрофенол, и потенциально полезные человеческие адъюванты, такие как BCG (Bacille Calmetle-Guerin) и Corynebakterium parvum.
Молекулярный клон антитела к эпитопу BDNF может быть приготовлен по известным технологиям. Для конструирования нуклеиновокислотных последовательностей, которые кодируют моноклональную молекулу антитела или ее антигенсвязывающую область, может быть использована методология рекомбинантной ДНК.
Молекулы антител могут быть очищены по известным методам, например иммуноабсорбционной или иммуноаффинной хроматографией, а также такими хроматографическими методами, как HPLC (высокопроизводительная жидкостная хроматография), или их сочетанием.
Фрагменты антител, которые содержат идиотип молекулы, могут быть генерированы по известной технологии. Например, но не с целью ограничения такие фрагменты включают: F(ab')2 фрагмент, который может быть получен перевариванием пепсином молекулы антитела; Fab' фрагменты, которые могут быть генерированы редуцированием дисульфидных мостиков фрагмента F(ab')2, а также 2 Fab и Fab фрагменты, которые могут генерироваться путем обрабтки молекулы антитела папаином и редуцирующим агентом.
Пример раздела II описывает приготовление поликлональной антисыворотки, направленной против B5-33 пептидного фрагмента BDNF-протеина.
5.8. Идентификация дополнительных членов генного семейства.
Анализ последовательности гена, кодирующего BDNF, и дедукция его аминокислотной последовательности обнаружили, что этот протеин имеет много структурных совпадений с NGF (фиг. 2). Первичная последовательность зрелого BDNF, так же как и общая структура и возможный режим процессинга от протеина-предшественника, служит свидетельством того, что гены NGF и BDNF могли быть извлечены из общего родительского гена внутри области зрелых полипептидов, если введено только три разрыва в NGF-последовательности для оптимизации совпадения, всего 51 идентичные аминокислоты являются общими у ранее известных NGF от многих видов животных, и у свиного и человеческого BDNF. Эти идентичности включают все шесть цистеиновых остатков, предполагая тем самым, что NGF-белки и BDNF разделяют очень похожую вторичную структуру. Кроме того, можно видеть четыре сегмента из шести или более аминокислот, в которых NGF белки от всех перечисленных выше видов животных и свиной BDNF либо идентичны, либо отличаются не более чем одной консервативной аминокислотной заменой. Таким образом, будет разумно заключить, что NGF и BDNF видимо, являются очень родственными членами одного генного семейства.
Рациональный поиск дополнительных членов этого BDNF/NGF генного семейства может быть проведен с использованием подхода, который извлекает выгоду из неожиданного существования консервированных сегментов с сильной гомологией между NGF и BDNF. Например, дополнительные члены BDNF генного семейства могут быть идентифицированы путем селекции из множества нуклеиновокислотных последовательностей тех последовательностей, которые гомологичны как BDNF, так и NGF, дальнейшим идентифицированием из выбранных таким образом последовательностей тех из них, которые содержат также нуклеиновокислотные последовательности, которые являются негомологичными последовательностями NGF и BDNF. Выражение "негомологичный" здесь может употребляться для обозначения области, которая содержит по меньшей мере около шести смежных нуклеотидов, в которой по меньшей мере два примерно нуклеотида отличаются от последовательности NGF и BDFN.
Предпочтительное выполнение изобретения предусматривает следующий способ. В соответствии с каждым из четырех консервированных сегментов, описанных выше, может быть синтезирована группа дегенерированных олигонуклеотидов, представляющих все возможные кодирующие последовательности для аминокислот, обнаруживаемых как в BDNF, так и в NGF через шесть смежных кодонов. Пронумеровав их по отношению к аминоконцу зрелых полипептидов (так, что His 134 prepro BDNF рассматривается как HisI в зрелом протеине), эти четыре консервированных сегмента ("боксы") могут быть охарактеризованы таким образом, как показано в табл. II (нумерация относительно человеческого зрелого протеина; последовательность ДНК изображена на фиг. 1).
Синтетическая олигонуклеотиды, полученные от пар последовательностей из боксов, показанных в табл. II (см. в конце описания), могут быть использованы в качестве праймеров для амплифицирования реакцией PCR-последовательностей из источников (РНК или ДНК), представляющего потенциальный интерес. Он может включать мРНК, или кДНК, или геномную ДНК от любого эукариота, который бы мог экспрессировать полипептид, близкородственный BDNF или NGF. Путем проведения всего шести PCR реакций (а именно: праймер из бокса 1 с праймером из бокса 2; бокс 1 с боксом 3; бокс 1 с боксом 4; бокс 2 с боксом 3; бокс 2 с боксом 4; и бокс 3 с боксом 4) можно обнаружить ген или генный продукт, разделяющий любые два из четырех указанных сегментов консервированной последовательности с NGF и BDNF. Если хотят синтезировать несколько различных дегенерированных праймеров для каждого бокса, то все еще можно провести полный поиск со сравнительно небольшим количеством PCR реакций. Можно также изменять строгость условий гибридизации, используемых в примировании PCR реакций, чтобы обеспечить большую или меньшую степень сходства между неизвестным геном и NGF или BDNF. Если сегмент ранее неизвестного члена генного семейства NGF/BDNF успешно амплифицировался, этот сегмент может быть молекулярно клонирован и секвенирован, и использован в качестве зонда для выделения полной кДНК или геномного клона. Это, в свою очередь, позволит определение полной нуклеотидной последовательности неизвестного гена, анализа его экспрессии и производство его белкового продукта для функционального анализа.
Описанный выше подход был использован для идентифицирования нового гена, родственного BDNF и NGF, описанного в примере раздела 13, ниже.
Дополнительно, настоящее изобретение обеспечивает использование гомологий последовательностей BDNF/NGF для создания новых рекомбинантных молекул, которые являются членами генного семейства BDNF/NGF, но которые в природе не встречаются. Например, но не с целью ограничения, рекомбинантная молекула может быть сконструирована согласно изобретению с включением частей как NGF, так и BDNF-генов. Такая молекула могла бы проявлять свойства, связанные и с BDNF, и с NGF, и показать новый профиль биологической активности, включая агонистов, так же как и антагонистов. Первичная последовательность BDNF и NGF может быть также использована для предсказания третичной структуры молекул с использованием компьютерного моделирования (Hoop и Woods, 1981, Proc. Nath. Acad.Sci. США, 78: 3824 - 3828); BDNF/NGF химерные рекомбинантные гены могут быть созданы с учетом корреляции меду третичной структурой и биологической функцией. Аналогично, химерные гены, включающие части любого одного или нескольких членов генного семейства BDNF/NGF, включая новый член, описанный в разделе 13, могут быть сконструированы таким образом.
5.9. Использование изобретения.
Настоящее изобретение касается нуклеиновокислотной последовательности BDNF и в основном чистого протеина, пептидных фрагментов или производных, полученных от него. BDNF может быть теперь впервые получена в количествах, достаточных для диагностического и терапевтического применения. Подобным образом в диагностике и терапии могут быть впервые использованы антитела против BDNF, полученные как результат изобретения, а также BDNF нуклеиновокислотные зонды. Для большинства назначений предпочтительно использовать гены BDNF или генные продукты от одних и тех же видов для диагностических или терапевтических целей, хотя в специфичных выполнениях изобретения может оказаться полезным кросс-видовое применение BDNF.
5.9.1. Диагностические применения.
Настоящее изобретение, которое относится к нуклеиновым кислотам, кодирующим BDNF, и к протеинам, пептидным фрагментам или производным, полученным от него, а также к антителам, направленным против BDNF-протеина, пептидов или производных, которые могут быть использованы для диагностики болезней и расстройств нервной системы, которые могут быть связаны с изменениями в паттерне BDNF-экспрессии.
В различных выполнениях изобретения гены BDNF и относящиеся к ним нуклеиновокислотные последовательности и субпоследовательности, включая комплементарные последовательности, могут быть использованы в исследованиях диагностической гибридизации. BDNF нуклеиновокислотные последовательности или субпоследовательности от них, включающие около 15 нуклеотидов, могут быть использованы в качестве гибридизационных зондов. Гибридизационные эксперименты могут использоваться для обнаружения, прогноза, диагноза или мониторинга состояний, расстройств или болезней, связанных с изменениями в BDNF-экспрессии, включая, в частности, условия, приводящие к повреждению сенсорных нейронов и дегенерации нейронов сетчатки. Такие болезни и условия включают, например, но не с целью ограничения, травмы центральной нервной системы, инфаркты, инфекцию, дегенеративные нервные заболевания, злокачественные болезни или послеоперационные изменения, включая, например, но не с целью ограничения, болезнь Альцхаймера, болезнь Паркинсона, хорею Хантингтона и дегенеративные болезни сетчатки. Например, общая РНК в образце ткани пациента может быть исследована на присутствие BDNF-мРНК, причем снижение количества BDNF-мРНК указывает на нейронную дегенерацию. Относительно высокие уровни содержания BDNF мРНК были идентифицированы в клетках нейробластомной опухоли (см.раздел 14, ниже); соответственно в частном выполнении изобретения обнаружение повышенных уровней мРНК с использованием BDNF нуклеиновокислотных зондов может быть использовано для диагностики нейробластомной опухоли. Крайней низкие уровни содержания BDNF, синтезируемые в большинстве тканей, делают BDNF-мРНК особенно полезным маркером опухоли.
В альтернативных выполнениях изобретения антитела, направленные против BDNF-протеина, пептидных фрагментов или производных, могут быть использованы для диагностики болезней и расстройств нервной системы, включая, в частности, сенсорные расстройства и дегенеративные болезни сетчатки, так же как те расстройства и заболевания, которые перечислены выше. Антитела по изобретению могут быть использованы, например, в технологии гибридизации "ин ситу" (in situ - на месте) на образцах ткани, полученных от пациента, при необходимости такой оценки. В другом примере антитела по изобретению могут быть использованы в процедурах ELISA для обнаружения и/или измерения количеств BDNF, присутствующих в образцах ткани или жидкости; аналогично антитела по изобретению могут быть использованы в анализе вестерн-блоттингом для обнаружения и и/или измерения BDNF, присутствующего в тканевых или жидких образцах. Антитела по изобретению, которые связываются с BDNF в ELISA и Вестерн-блоттинге, описаны в разделе II (см. ниже).
В других выполнениях изобретения BDNF-протеин, пептидные фрагменты или производные могут быть использованы для диагностики и расстройств нервной системы. В частном выполнении, например, но не с целью ограничения, меченый BDNF-протеин или меченые пептидные фрагменты могут быть использованы для идентифицирования тканей или клеток, которые экспрессируют BDNF-рецептор для идентификации аберраций экспрессии BDNF-рецептора, и, следовательно, потенциальных отклонений от нормы тканевой или клеточной реакции на BDNF.
5.9.2. Терапевтическое применение.
Настоящее изобретение, которое относится к нуклеиновым кислотам, кодирующим BDNF, к его протеинам, пептидным фрагментам или производным, а также к антителам, направленным против BDNF-протеина, пептидов или производных, может быть использовано для лечения болезней и расстройств нервной системы, которые могут быть связаны с изменениями в паттерне BDNF-экспрессии или которые могут возникать от воздействия BDNF или антител против BDNF.
В различных выполнениях изобретения BDNF-протеин, пептидные фрагменты или его производные могут вводиться пациентам, у которых нервная система была повреждена травмой, хирургией, ишемией, инфекцией, болезнями обмена веществ, пищевой недостаточностью, злокачественными болезнями или токсическими веществами. Изобретение, в частности, может быть использовано для устранения условий, в которых возникает повреждение сенсорных нейронов или клеток ганглия сетчатки путем введения эффективных терапевтических количеств BDNF-протеина или пептидных фрагментов или производных. В различных специфических выполнениях изобретения BDNF может вводиться местно к сенсорным нейронам, которые были рассечены, включая, например, но не с целью ограничения, нейроны ганглиев дорсального корня или в любой из следующих тканей: коленчатый, височный и нодозный ганглии; вестибулоакустический комплекс восьмого черепного нерва; вентролатеральный полюс максилломандибулярной доли тройничного ганглия; и мезенцефальный тройничный нуклеус (ядро). Может быть желательным вводить родственные BDNF-пептиды или BDNF-протеин посредством абсорбции на мембране, например, сиаластиковой мембране, которая может быть имплантирована вблизи поврежденного нерва. Настоящее изобретение может быть также использовано, например, для ускорения восстановления пациентов, страдающих диабетической нейропатией, например множественной мононейропатией. В других выполнениях изобретения BDNF-протеины или его пептидные фрагменты или производные могут быть использованы для лечения врожденных состояний и нейродегенеративных расстройств, включая, например, но не с целью ограничения, болезнь Альцхаймера, болезнь Паркинсона, синдромы Паркинсон-плюс (при которых паркинсоновые симптомы происходят от дегенерации допаминергических нейронов), такие как прогрессивный сверхнуклеарный паралич (синдром Стил-Ричардсона-Ольшанского), атрофия мозжечкового мозга (Olivopontocerebellar Artophy), синдром Шай-Дрэгера (Shy-Drager) - множественная системная атрофия, и деменциальный комплекс Гуманового Паркинсонизма (Guamanian Parkinsonism), а также хорея Хантингтона; в частности, изобретение может быть использовано для лечения врожденных или нейродегенеративных расстройств, связанных с дисфункцией сенсорных нервов и дегенеративными болезнями сетчатки. Например, BDNF-протеин, или пептидные фрагменты, или производные по изобретению могут быть использованы в лечении наследственной спастической параплегии с дегенерацией сетчатки (синдромы Кьелина и Бернарда-Шольца), пигментоз сетчатки, болезнь Старгардта, синдром Ашера (пигментоз сетчатки с врожденной потерей слуха) и синдром Рефсума (пигментоз сетчатки, наследственная потеря слуха и полинейропатия), но это только часть. Возможно, что дефект синтеза BDNF или реакции на него могут быть причиной синдромов, характеризующихся сочетанием дегенерации сетчатки и других сенсорных дисфункций.
В специфическом выполнении изобретения введение BDNF-протеина, или пептидных фрагментов, или его производных может быть использовано в соединении с хирургическим имплантированием ткани при лечении болезни Альцхаймера и/или болезни Паркинсона. Как обсуждается ниже в разделах 12 и 18, BDNF может быть использован для содействия выживанию допаминергических; "субстанция нигра" нейронов зависимым от дозы образом (фиг. 34), поддерживая использование BDNF в лечении расстройств ЦНС-допаминергических нейронов, включая, например, но не с целью ограничения, болезнь Паркинсона (особенно в свете данных, представленных в разделе 20, ниже, которые показывают, что BDNF может быть использован для предотвращения нейротоксичности, вызываемой МРР, токсином, который связан с вызыванием паркинсоноподобного синдрома). Дополнительно наблюдалось, что BDNF поддерживает выживание холинергических нейронов центральной нервной системы (см. ниже разделы 12 и 16) и, в частности, холинергических нейронов базального переднего мозга, указывая тем самым, что BDNF может быть полезным при лечении расстройств, вовлекающих холинергические нейроны, включая, например, но не с целью ограничения, болезнь Альцхаймера. Было показано, что примерно 35% пациентов с болезнью Паркинсона страдают деменцией Альцхаймерового типа; BDNF, полученный согласно изобретению, может оказаться полезным как единственный терапевтический агент этого комплекса болезней. Подобным образом, BDNF, полученный согласно изобретению, может быть использован терапевтически для лечения болезни Альцхаймера в соединении с синдромом Дауна. BDNF, полученный по изобретению, может быть использован при лечении различных деменций так же, как врожденных расстройств обучения. Было также обнаружено, что BDNF вероятно подавляет пролиферацию астроглиальных клеток, поддерживая тем самым применение BDNF для уменьшения рубцовых образований в ЦНС (например, после хирургии, травмы или инфаркта), а также применение BDNF при лечении опухолей ЦНС астроглиального происхождения. В еще одном выполнении изобретения BDNF может быть использован для регулировки экспрессии NGF-рецептора, и поэтому он может с выгодой вводиться до или совместно с NGF пациенту при необходимости такого лечения.
Как приведено в качестве примера в разделе 15, внизу, введение каиновой кислоты может быть использовано для вызывания усиления экспрессии BDNF (и в меньшей степени экспрессии NGF) в нейронах, включая нейроны аммонова рога, а также кортекса. Наблюдалось, что (см. раздел 15, ниже) ингибирование не-NMDA рецепторов блокирует это, вызванное каиновой кислотой усиление экспрессии BDNF. Соответственно экспрессия родственных BDNF пептидов и самого BDNF по изобретению может быть вызвана ин витро или ин виво путем каиновой кислоты или родственных ей соединений, или карбахола, гистамина, или брадикинина, или родственных им соединений, которые, как установлено, обладают сходным действием ин витро или ин виво или посредством агонистов не-NMDA глютаматных рецепторов, или других лекарств, которые усиливают или ослабляют действие ацетилхолина, гистамина или брадикинина. Вызвать экспрессию NGF можно также путем введения каиновой кислоты или родственных ей молекул или агонистов не-NMDA рецепторов.
В других выполнениях изобретения BDNF-протеин, его фрагменты или производные могут быть использованы в сочетании с другими цитокинами для достижения желаемого нейротропного эффекта. Например, но не с целью ограничения, согласно изобретению BDNF может использоваться вместе с NGF или с экстрактом скелетных мышц для достижения синергической стимуляции роста сенсорных нейронов, где выражение "синергической" следует понимать так, что действие комбинации BDNF-протеина, его пептидных фрагментов или производных, со вторым агентом дает больший эффект, чем то же количество каждого вещества по отдельности. Предусматривается, что BDNF может функционировать синергетически с другими, происходящими от ЦНС пептидными факторами, которые еще предстоит полностью охарактеризовать, на рост, развитие и выживание широкой разновидности нейронных субпопуляций в центральной нервной системе.
Далее предусматривается, что на основе полной характеристики молекулы BDNF новые пептидные фрагменты, производные или мутанты BDNF могут быть созданы так, чтобы они были способны действовать как антагонисты некоторых или всех биологических функций BDNF. Такие антагонисты BDNF могут быть полезны при селективном извлечении сенсорных нейронов, например при лечении синдромов хронической боли.
В еще других выполнениях изобретения антитела, направленные против BDNF-протеина, или его пептидных фрагментов, или производных, могут вводиться пациентам, страдающим разнообразными неврологическими расстройствами и болезнями и которые нуждаются в таком лечении. Например, в таком лечении могут нуждаться пациенты, страдающие от перепроизводства BDNF. Антитела против BDNF могут быть использованы для предотвращения аберрантной регенерации сенсорных нейронов (например, послеоперационно) или, как обсуждалось выше, при лечении синдромов хронической боли. В свете высоких уровней содержания BDNF мРНК в нейробластомной ткани возможно, что BDNF служит фактором роста аутокриновой опухоли для нейробластомы; соответственно, терапевтически могут вводиться антитела против BDNF для регрессии опухоли в специфическом выполнении изобретения.
5.10. Фармацевтические композиции.
Активные композиции по изобретению, которые могут включать целиком или части генного продукта BDNF, включая протеин, пептидные фрагменты или производные, или антитела (или фрагменты антител), направленные против BDNF-протеина, или его пептидных фрагментов, или его производных, или могут содержать комбинацию BDNF со вторым агентом, таким как NGF, или экстракт скелетных мышц, могут вводиться в любом биосовместимом фармацевтическом носителе, включая, например, но не с целью ограничения, физиологический раствор, буферный физ. раствор, декстрозу и воду.
BDNF-протеин, его пептидный фрагмент или производное могут включать аминокислотную последовательность или ее субпоследовательность, в основном такую, как изображено на фиг. 1 или 5: может быть предпочтительным использование BDNF-протеина, включающего, в частности, целиком или часть аминокислотной последовательности от примерно аминокислоты 134 до примерно аминокислоты 252, как изображено на фиг. 1, или функционально эквивалентную последовательность, если эта последовательность предпочтительно включает функциональную часть BDNF-молекулы, BDNF может быть получен от последовательностей, соответствующих генам BDNF от любых подходящих видов животных, включая, например, но не с целью ограничения, такие виды как человек, свинья, крыса, цыпленок, корова, собака, овца, коза, кошка, кролик и т.д.
То количество BDNF-протеина, пептидного фрагмента, производного или антитела, которое окажется эффективным при лечении конкретного расстройства или состояния, будет зависеть от природы этого расстройства или состояния и может быть определено стандартными клиническими методами. Когда возможно, желательно определить кривую зависимости реакции от дозы и фармацевтическую композицию по изобретению сначала ин витро, например в BDNF биоэкспериментальных системах, описанных выше, а затем на полезной животной модели перед тем, как проверять действие композиции на людях. На основании данных, полученных ин витро, в специфическом выполнении изобретения фармацевтическая композиция, эффективно содействующая выживанию сенсорных нейронов, может обеспечить местную концентрацию протеина BDNF между примерно 5 и 25 нг/мл и предпочтительно между 10 и 20 нг/мл. В дополнительном специфическом выполнении изобретения фармацевтическая композиция, эффективно способствующая росту и выживанию дипаминергических или холинергических нейронов, может обеспечивать местную концентрацию протеина BDNF примерно между 10 и 100 нг/мл.
Способы введения включают, например, но не с целью ограничения, внутрикожный, внутримышечный, внутрибрюшной, внутривенный, подкожный, оральный и интраназальный способы. В дополнение может быть желательным введение фармацевтической композиции по изобретению в центральную нервную систему любым подходящим путем, включая интравентрикулярную или интратекальную инъекцию; интравентрикулярная инъекция может быть облегчена посредством внутрижелудочного катетера, прикрепленного, например, к резервуару, такому как резервуар Омайя (Оммауа).
Далее, может быть желательным введение фармацевтической композиции по изобретению локально в области, нуждающейся в лечении; это может быть достигнуто, например, но не с целью ограничения, местной инфузией во время хирургической операции, путем инъекции, посредством катетера или имплантированием, причем указанный имплантант может быть пористым, или непористым, или желатиноподобным материалом, включая мембраны, такие как сиаластиковые мембраны, или волокна.
Изобретение также обеспечивает фармацевтические композиции, включающие BDNF-протеины, пептидные фрагменты или производные, вводимые посредством липосом, микрочастиц или микрокапсул. В различных выполнениях изобретения может быть полезным использование таких композиций для достижения поддерживаемого высвобождения BDNF или родственных BDNF продуктов.
Предусмотрено, что имеется возможность введения клеток, активно продуцирующих BDNF, родственных BDNF веществ, антагонистов BDNF или антител против BDNF в зоны, нуждающиеся в повышении или снижении концентраций BDNF.
6. Пример. Молекулярное клонирование и характеризирование кДНК свиного мозгового нейротропного фактора.
Крайняя редкость протеина BDNF в тканях препятствует использованию стандартных стратегий клонирования
гена BDNF. Вместо этого, ограниченное количество
данных протеиновых последовательностей расширяют с использованием технологии амплифицирования ДНК следующим образом:
(I) микрограммные
количества BDNF очищали из килограммов свиного головного
мозга,
(II) очищенный BDNF анализировали технологией белкового микросеквенирования и определяли аминокислотную последовательность
отрезка из 36 аминокислотных остатков,
(III)
основанные на определенной в (II) аминокислотной последовательности, олигонуклеотиды синтезируются и используются в качестве праймеров в цепной
реакции полимеразы, с использованием свиной, superior
colliculus кДНК в качестве матрицы для того, чтобы амплифицировать ДНК, кодирующую определенный аминокислотный фрагмент,
(IV)
секвенировали продукт ДНК-реакции по (III), и
(V)
синтезировали соответствующие олигонуклеотидные праймеры и использовали их в цепной реакции полимеразы со свиной superior colliculus кДНК
для генерирования перекрывающих фрагментов ДНК, представляющих
BDNF мРНК, выше, так же как и ниже последовательностей, кодирующих исходный фрагмент из 36 аминокислот. Таким образом в двух
перекрывающих фрагментах клонировалась полная кодирующая область для
свиного BDNF.
Детали каждого из этих этапов, так же как и дальнейшая характеристика гена BDNF, даны ниже.
6.1. Материалы и методы.
6.1.1. Очистки BDNF из свиного мозга.
Очистку BDNF из свиного головного мозга осуществляли посредством процедуры, в основном такой, как описано у Хофера и Барда (Hofer и Barde, 1988, Nature, 331: 261-262).
Шесть килограммов свиного головногомозга гомогенизировали с помощью гомогенизатора "Ультра-Турракс" в 0,2 M натриево-фосфатном буфере с pH 6,0, содержащем 1 мМ ЭДТК и свежедобавленным фенилметансульфонилфторидом (1 мМ) в отношении - на 1 кг мозга 2 литра буфера. pH супернатанта приводили к pH 4,0 с помощью 1н. HCl и перемешивали в течение 2 часов при 4o C. После центрифугирования в течение 25 минут при 20 тыс. "g" соединенные супернатанты (доведенные до pH 6,0 посредством добавления 1н. NaOH), соответствующие трем килограммам мозга, перемешивали в течение 2 часов с 1 литром предварительно набухшей карбокси-метил-целлюлозы, которые уравновешивали 0,1 M натрий-фосфатом до pH 6,0. После двух промывок в общем 20 литрами 0,1 M натрий фосфата с pH 6, 0 эту кашицу выливали в колонну и промывали до утра в том же буфере, содержащем 0,13 M NaCl. Активные фракции элюировали с фосфатным буфером, содержащим 0,5 M NaCl, а затем диализовали против 2 х 2 л 5 мМ калия фосфата с pH 6,8. Диализованные фракции от процессинга 2 х 3 кг исходного материала были помещены в 130 мл слой гидроксиапатитовой колонны, который предварительно уравновесили 5 мМ калия фосфатом с pH 6,8. Колонну затем элюировали с линейным градиентом от 500 мл 5 мМ до 500 мл 700 мМ калия фосфатом с pH 6,8, причем BDNF-активность элюировали приблизительно 500 мМ калия фосфатом (см. Barde и др., 1982, ЕМВО, 1:549-553). Добавлением 1,0 M калия фосфата полученные активные фракции довели до конечной молярности 700 мМ калия фосфата и помещали в 5 мл фенил-сефарозную колонну, уравновешенную 700 мМ калия фосфата с pH 6,8. После промывки 40 мл того же самого буфера BDNF-активность элюировали 0,1 M калия фосфата с pH 6,8, диализовали против дистиллированной воды и лиофилизировали. Лиофилизированный материал помещали в буфер для SDS-гелевого электрофореза образца, содержащий 0,1% SDS, но без меркаптоэтанола, как описано у Барда и др. (1982, ЕМВО, 1:549-553), а затем помещают на SDS-гель, включающий акриламид с линеарным (скорее, чем экспоненциальным) градиентом от 10 до 25%. После завершения электрофоретического разделения гель кратковременно окрашивали (10 мин) "Коомасси блю", обесцвечивали 20 минут, а полосу, мигрирующую на уровне цитохрома "c", вырезали и элюировали из геля. SDS удаляли как описано у Барда и др. (1982, ЕМВО, 1: 549-553). Процедура очистки, которая эвентуально приводила к микросеквенированию BDNF была модифицирована согласно Хоферу и Барду (1988, Nature 331: 261-262) и не использует теперь гелевый электрофорез на последней стадии очистки.
6.1.2. Секвенирование белка.
BDNF либо секвенировали непосредственно (55 pmol как определено аминокислотным анализом с начальным выходом в 40 pmol для амино-терминального гистидина), или расщепляли следующим образом: 5-10 мкг BDNF (из пяти разных приготовлений) расщепляли согласно следующей процедуре. V8: 1 мкг S. aureus V8 (Майлс) добавляли к 5 мкг BDNF 0,2 NH4CO3, pH 8,0, содержащем 10% ацетонитрила (общий объем 50 мкг), и инкубировали до утра при комнатной температуре. Трипсин: 1 мкг ТРСК-обработанного трипсина (бычья поджелудочная железа, Сигма, тип XIII) добавляли к 8 мкг BDNF в Трис-HCl (0,1 М, pH 8,0), содержащей 10 мМ CaCl2 (общий объем: 40 мкл) и инкубировали до утра при 37oC. Цианоген-бромид (CNBr): 10 мкг BDNF инкубировали в течение 3 часов при комнатной температуре (общий объем: 60 мкл) с 10% (вес./об.) CNBr в 70% (об./об.) муравьиной кислоты (конечные концентрации). После добавления 500 мкл H2O в конце реакции образец концентрировали до 50 мкл в "спид-ваке" (скоростной вакуум-выпариватель). 50 мкл Трис-HCl (1,0 М с pH 8,0) было добавлено вместе с 5 мкл бета-меркаптоэтанола, и образец инкубировали до утра при 37oC. После добавления 5 мкл йодометана образец выпаривали до сухого состояния в спид-ваке. Было обнаружено, что после CNBr расщепления необходимо провести восстановление и алкилирование BDNF: без восстановления не получали фрагменты, как обнаружил электрофорез на SDS-геле Swank-Munkres и HPLC (1971, Anal, Biochem 39: 462 - 477). Это указывает, что в BDNF присутствуют дисульфидные мостики, устроенные таким образом, что ни одно из расщеплений не освобождает пептиды. После всех расщеплений высушенные образцы ресуспендировали в 0,1% трифторуксусной кислоте (TFA) и поместили в С8 микробор HPLC колонну обратной фазы ("Applied Biosystems") и пептиды элюировали со скоростью 0,1 мл/мин с использованием 60 мин линейного градиента 0 - 60% ацетонитрила в 0,1% TFA. Детекцию осуществляли при 214 нм с помощью Waters 441 UV детектора. Эти пептиды секвенировали автоматизированной деградацией Эдмана на газофазовом микросеквенаторе (модель 470А, "Applied Biosystems"), согласно Хевику (Hunkapil, 1981, J. Biol. Chem. 256: 7990 - 7997) и Хункапиллару (Hunkapillar, 1983, Methods Enzymol. 91: 227 - 236). Обнаружение РТН аминокислот проводили как описано Лоттшпайхом (Lottspeich, J. Chromatogr 326: 321 - 327).
6.1.3. Приготовление ДНК-матриц.
Свиная геномная ДНК выделялась по методу Herrmann и Frischlauf (1987, Methods Enzymol. 152: 180 - 183).
Для приготовления кДНК из шести граммов superior colliculus свиного головного мозга получали общую РНК. Образец ткани получали ди секцией и замораживали в жидком азоте на местной бойне. Для экстракции РНК использовали стандартные методы (Okayama и др., 1987, Methods Enzymol. 154: 3 - 28). 80 мкг общей РНК транскрибировали с обратной транскриптазой вируса мышиной лейкемией Moloney (BRh, согласно инструкциям производителя, за исключением добавления 1 мкл RNAsin и без актиномицина D), с использованием праймерной смеси CGGATCCGAATTCTGCAGTTTTTTTTTTTT C A, G или G в качестве терминального 3' нуклеотида (oligo 3, созданный для совместимости с 3'поли-A отрезком и содержащий сайты узнавания для Bam HI, EcoRI и Pst I).
6.1.4. Цепная реакция полимеразы.
Цепную реакцию полимеразы осуществляли согласно методу, описанному у Сайки и др. (Saiki, 1985, Science 230: 1350 - 1354).
6.2. Результаты и обсуждение.
6.2.1. Результаты микросеквенирования белка.
Экспериментально определенная пептидная последовательность BDNF.
N-терминал - HSDPARRGELSV
V8 - XVTAADKKTAVD
V8
- KVPVSKGQLKQYFYE
CNBr - XGGTVTVLEKVP (V) (S)
CNBr - GYTKEGXRGIXRGI
Trypsin
- (T)AVDMSGGTVTVLEK
Trypsin - ALTMDSK
Соединенная последовательность:
VTAADKKTAVDMSGGTVTVLEKVPVSKGQLKQYFYE подчеркнутые аминокислоты представляют последовательности, кодированные
олигонуклеотидными праймерами, использованными в реакции PCR (см. раздел 6.1.2, выше).
Четыре смежных сегмента аминокислотных последовательностей определяли с использованием деградации Эдмана, которую
можно было использовать для выведения 36 аминокислот, представляющих приблизительно
одну треть всей аминокислотной последовательности.
6.2.2. Синтез олигонуклеотидов и использование PCR реакции для получения ДНК, кодирующей аминокислотный фрагмент.
Два полностью дегенерированных 17-мерных олигонуклеотидных праймера синтезировали химически на основе кодирующих последовательностей для сегментов из шести аминокислот около аминотерминального и карбоксил-терминального концов, соответственно, фрагмента из 36 аминокислотных остатков, описанного в разделе 6.2.1, выше. В частности, были химически синтезированы две раздельные смеси 17-мерных олигонуклеотидов (соответственно всем возможным кодонам и созданным oligo 1 и oligo 2) на основе последовательности AADKKT (смысловая) и KQYFUE (антисмысловая) (подчеркнуты в табл. II), и по 150 рмо1 каждого праймера было добавлено к 1 мкг свиной геномной ДНК-матрицы. Было осуществлено 35 циклов амплификации с использованием реакции PCR согласно указаниям производителя (GeneAmpTM, Perkin-Elmer Cetus). Денатурация длилась при 94o одну минуту, праймер ренатурировали при 45o в течение 2 минут, и расширяли при 72oC в течение 2 мин. ДНК-полосу предсказанного размера (101 бр) вырезали из 3%-ного агарозного геля (окрашенного этидиум-бромидом) и ассимметрично амплифицировали как описано Иннисом и др. (Innis, 1988, Proc. Nath. Acad. Sci. США, 85: 9436 - 9440) со 100-кратным избытком либо oligo 1, либо oligo 2.
6.2.3. Нуклеотидная последовательность фрагмента кДНК.
Полученные смысловые и антисмысловые фрагменты кДНК секвенировали методом дидеоксинуклеотидного цепного терминирования (Sanger и др., 1979, Proc. Nath. Acad. Sci. США, 72: 3918 - 3921) с использованием 32P-меченных на конце олигонуклеотидов 1 и 2 в качестве праймеров. Наблюдавшаяся нуклеотидная последовательность содержала только открытую рамку считывания, непрерванную кодоном терминирования цепи. Выведенная аминокислотная последовательность для этой открытой рамки считывания полностью соответствовала действительной аминокислотной последовательности, определенной для этой области свиного BDNF.
6.2.4. Клонирование всей кДНК свиного BDNF.
Полную кодирующую область для свиного BDNF молекулярно
клонировали в двух перекрывающих сегментах. Чтобы получить 3' участок
(относительно смысловой нити) кДНК BDNF 30-мерный олигонуклеотидный праймер, содержащий 21 основание точной последовательности
смысловой нити BDNF из области, описанной выше, синтезировали для того,
чтобы использовать его в качестве "смыслового праймера". Нуклеотидная последовательность этого праймера была oligo 4, (5')
CGGATCCGAATTCTGCAGTTTTTTTTTTTX
(подчеркнута последовательность,
соответствующая смысловой нити BDNF от позиции 643 до 633 на фиг. 1, кодирующая аминокислотную последовательность
Thr-Ala-Val-Asp-Met-Ser-Gly
(первые два основания - кодона Gly).
Дополнительные 9 нуклеотидов у 5'-конца этого праймера были включены для обеспечения сайтов расщепления рестрикционными
эндонуклеазами для Spe 1 и Sal 1 при использовании в молекулярном клонировании.
Был создан тройной дегененированный 31-мерный олигонуклеотидный праймер, комплементарный отрезку поли-А, предваряемому
любым нуклеотидом (T, C или G) на смысловой нити кДНК, для того, чтобы содержать
сайты узнавания для рестрикционных эндонуклеаз Bam HI, Eco RI и Pst I. Последовательность этого "антисмыслового"
праймера (oligo 3) была (5') CGGATCCGAATTCTGCAGTTTTTTTTTTTTX (3'), где X = A, C или G.
Синтетические олигонуклеотидные праймеры использовали для амплифицирования последовательностей от свиной superior
colliculus кДНК, приготовление которой описано выше, путем PCR. В частности, 3'
амплифицированная ДНК была получена с использованием 10 л реакционной смеси для обратной транскрипции, 150 рмо1
смыслового праймера (oligo 4) и150 рмо1 oligo 3 в качестве антисмыслового праймера в
PCR реакции. Саузерн-блот-анализ осуществляли на амплифицированных продуктах ДНК; и полосу, дающую гибридизационный
сигнал с 32Р-меченым концом олигонуклеотидом
AAGGATCCTGCAGTTGGCCTTTCGAGAGGC (oligo 5, используемый в качестве антисмыслового праймера в 5' реакции, описанной ниже, и содержащий
последовательности узнавания для Bam HI и Pst I) вырезали,
экстрагировали методом "glassmilk" (ген clean, переваренный Eco RI и Sall, клонировали в Bluescript Sk+-плазмиду (Стратаген) и
секвенировали).
Чтобы получить остаток (выше или 5' участок) BDNF кодирующий последовательности, кДНК приготавливали как описано выше, и к 5' концам добавляли поли-A хвосты с использованием терминальной деоксинуклеотидной трансферазы. Ту же смесь, из трех 31-мерных олигонуклеотидов, каждый из которых содержал отрезок из I2 последовательных T остатков, описанный выше, использовали для получения праймера, комплементарного этим добавленным поли-A хвостам. Был синтезирован единственный 30-мерный олигонуклеотидный праймер, содержащий 17 оснований, соответствующих комплементарной нити кодирующей BDNF-последовательности, с сайтами узнавания для рестрикционных эндонуклеаз BamHI и PstI. Последовательность этого праймера была следующей: (5')AAGGATCCTGCAGTTGGCCTTTCGAGACGG (3') (oligo 5, см. выше) - подчеркнута область, комплементарная смысловой нити кодирующей последовательности BDNF от позиции 709 до 693 на фиг. 1. Эта последовательность соответствует сегменту, кодирующему аминокислотную последовательность Pro (последние два основания кодона) Val-Ser-Lys-Gly-Gln. Эти праймеры добавляли к поли-A-хвостатой ДНК, и проводили, как выше, амплификацию реакцией PCR. Продукты реакции вырезали Pst I и клонировали в Bluescript вектор и определяли нуклеотидную последовательность методом дидеоксинуклеотидного цепного терминирования.
6.2.5. Нуклеотидная последовательность свиной кДНК BDNF.
Соединенная нуклеотидная последовательность, определенная из переклонов прекрывающей свиной BDNF кДНК, показана на фиг. 1 вместе с выведенной аминокислотной последовательностью. Эта последовательность включает открытую рамку считывания для полипептида из 252 аминокислот. Идентификация инициаторного Мет кодона (АТГ) основана на присутствии двух смежных кодонов терминации цепи (ТАГ-ТГА) в той же рамке считывания, начиная 36 парами оснований выше. Аминотерминал свиного BDNF, определенный прямым анализом последовательности на очищенном протеине, соответствует остатку His B4 этого полипептида. Так, данные секвенирования показывают, что зрелый BDNF получен процессингом от предшествующего полипептида. Непосредственно перед остатком His 134 расположена последовательность Arg-Val-Arg-Arg. Такие последовательности из одного основного аминокислотного остатка, за которым следует нейтральный остаток и затем еще два основных остатка, являются мишенными сайтами для протеолитического процессинга полипептидов-предшественников. Выведенная аминокислотная последовательность зрелого BDNF предсказывает протеин из 119 аминокислот (с молекулярным весом 13,511 Дальтон) с щелочным зарядом (р1 = 9,99), свойствами, согласующимися с предварительной характеристикой BDNF посредством оценки биологически активного фактора после его фракционирования посредством двухмерного гелевого электрофореза. Аминокислотная последовательность частей BDNF, определенная белковым микросеквенированием (всего 64 аминокислотных остатка), находится в полном согласии с аминокислотной последовательностью, выведенной из нуклеотидной последовательности кДНК клонов (подчеркнуто на фиг. 1). Эта последовательность предшествующего полипептида согласуется с процессингом BDNF по меньшей мере на двух этапах: во-первых, сигнальный пептид, вероятно из 18 остатков, должен расщепляться от аминотерминала с последующим расщеплением между Arg 133 и His 164 для освобождения зрелого полипептида. Если эта модель верна, тогда предшественника следует обозначить как BDNF.
7. Пример. Ген BDNF отличается от гена NGF у различных видов позвоночных.
7.1. Материалы и методы.
7.1.1. Приготовление NGF и BDNF ДНК-зондов.
От фирмы
"Бритиш Биотехнолоджиз Лимитед" была получена плазмида, содержащая синтетический ген,
кодирующий зрелый человеческий NGF, с включением с последовательностью, кодирующей нормальный человеческий NGF,
несколько консервированных кодонных замещений для введения удобных рестрикционных
сайтов расщепления эндонуклеазами. Пара 18-мерных олигонуклеотидных праймеров была синтезирована для обеспечения
амплификации 270 бр сегмента этого гена, соответствующего кодирующей области для
аминокислотных остатков 9 - III, с помощью PCR. Для того чтобы получить меченый ДНК-зонд, проводили 10 циклов PCR
реакции с 32Р-dCTP. BDNF-зонд был получен сходными процедурами, за исключением того,
что амплификацию проводили с использованием свиной геномной ДНК в качестве исходной матрицы, и амплифицированный
сегмент соответствовал кодирующей области для аминокислот 28 - III зрелого BDNF.
Комплементарный нитевой ("антисмысловой") праймер, соответствующий области с аминокислотами 106 - III, был синтезирован
с последовательностью (5') ACATACACAGGAAGTGTG (3'). Олигонуклеотидный праймер
смысловой нити был приготовлен для области из аминокислотных остатков 28 - 33
(5') GCAGTGGACATGTCGGGT (3').
Саузерн-блот-гибридизация (Southern, 1975, J. Mol. Biol. 98: 508 - 517) с этими двумя зондами проводили в строгих условиях в 2 x SSC при 68oC.
7.1.2. Секвенирование генов BDNF от различных видов животных.
Те же два 18-мерных олигонуклеотида, захватывающие кодирующую область для аминокислот 28-III зрелого свиного BDNF, описанные выше, использовали в качестве праймеров для амплифицирования, в стандартных условиях PCR, 252 бр сегментов геномной ДНК свиньи, крысы, цыпленка и человека, и полученные ДНК реакционные продукты секвенировали посредством метода дидеоксинуклеотидного цепного терминирования (Sanger и др., 1979, Proc. Natl. Acad. Sci. США, 72: 3918 - 3921). В некоторых случаях полосу амплифицированной ДНК вырезали и экстрагировали после агарозного гелевого электрофореза и реамплифицировали перед секвенированием. В других случаях реамплификация была необязательна.
7.2. Результаты и обсуждение.
До настоящего изобретения BDNF-протеин был очищен только от свиньи. Важнейшее значение имела однозначная демонстрация того, что BDNF это не просто бета-субъединица свиного фактора нервного роста (бета-NGF, или просто NGF, как здесь он упоминается), очистка и молекулярное клонирование которого до сих пор отсутствовали в сообщениях. Это было особенно важно вследствие того, что ранее сообщалось о физических свойствах свиного BDNF, что они, фактически, идентичны свойствам бета-NGF мономера из многих видов животных. Отсутствие нейтрализующих антител против свиного BDNF и отсутствие ранее аминокислотной или нуклеотидной последовательности свиного BDNF не позволяло определить точную взаимосвязь между BDNF и NGF. Можно было понять, что наблюдаемые различия в биологической активности между BDNF и NGF могли, например, просто отражать различия у NGF между свиньей и другими видами животных (например, мыши), или же происходить от дифференциальной модификации молекулы NGF протеина в различных тканях (например, свиной мозг против слюнной железы мыши), или вследствие модификаций, случайно введенных в протеин на какой-то стадии (или стадиях) во время очистки свиного мозга.
Если бы было установлено, что BDNF четко отличается от NGF, было бы важно определить, присутствует ли ген BDNF у других видов, особенно у человека. До настоящего изобретения не было информации на эту тему, поскольку BDNF был очищен только от одного вида. Присутствие нейротропной активности, которая, вероятно, отличалась от NGF во многих сырых экстрактах и кондиционированных средах, еще не подразумевало существования субстанции, которая бы была идентичной или в основном эквивалентной свиному BDNF.
Сравнение предсказанной аминокислотной последовательности свиного BDNF с известными последовательностями NDF от нескольких видов животных (человек, корова, гвинейская свинья, мышь, цыпленок и змея) показало что BDNF значительно менее тесно связан с любым NGF позвоночных, чем NGF разных видов друг с другом (фиг. 2). Неожиданной особенностью первичной структуры зрелого BDNF является его схожесть с такой структурой NGF с только тремя разрывами, введенными в последовательности для оптимизирования совместимости, имеется 51 идентичность, общая для различных NGF (от змеи до человека) и BDNF (фиг. 2). Важно, что эти идентичности включают все шесть цистеиновых остатков. Хотя еще не известно точное устройство дисульфидных мостиков BDNF, ясно, что такие мостики имеются (табл. II, пояснения). 3 триптофановых и 2 фенилаланиновых остатка, обнаруживаемых в BDNF, находятся в идентичных позициях у NGF. Следует также отметить, что 6 остатков аспартовой кислоты (из 7 у BDNF) и 7 валинов (из 9) присутствуют в идентичных позициях у NGF млекопитающих и BDNF. Эти пять аминокислот дают примерно половину аминокислотных идентичностей между двумя протеинами. Наоборот, имеется несколько резких отличий между BDNF и всеми NGF. В дополнение к уже упомянутым трем разрывам, также имеется 21 позиция, в которой аминокислоты являются идентичными во всех NGF, но различаются у BDNF.
Большая часть последовательности предшественника не родственна последовательности NGF с 2 исключениями: предполагаемая секреторная сигнальная последовательность BDNF показывает 5 аминокислотных идентичностей (из 18 аминокислот) и общую удивительную похожесть с сигнальной последовательностью мышиного NGF, где, как было продемонстрировано, расщепление происходит после аланинового остатка, находящегося в позиции 18 метионина, инициирующего трансляцию (Эдвардз и др., 1988, Mol. Cell. Biol. 8: 2456-2464). Выглядит вероятным, что аланин, также находящийся в позиции 18 у BDNF, представляет потенциальный сайт расщепления для удаления сигнальной последовательности BDNF. Другая схожесть с NGF начинается у единственной консенсусной последовательности N-гликозилирования, соответствуя аспарагину 126. Этот аспарагин расположен за восемь аминокислот перед сайтом расщепления, дающим начало зрелому BDNF. То же устройство обнаружено у нескольких NGF; так же как последовательность Arg-X-Basic-Arg в качестве последних 4 аминокислот предшественника (Шварц и др., 1989, J.Neurochem. 52:1203-1209).
Доказательство того, что NGF и BDNF кодированы различными генами у разных видов позвоночных, было получено путем приготовления ДНК зондов из молекулярно клонированных человеческого NGF и свиного BDNF и проведением саузернблот гибридизации с геномной ДНК, переваренной рестрикционной эндонуклеазой EcoRI. Геномные ДНК анализировали из следующих: человек, обезьяна, крыса, мышь, собака, корова, кролик, цыпленок и дрожжи. ДНК переваривали EcoRI и анализировали саузернблоттингом, на двойных фильтрах с 32P-меченым зондом человеческого NGF и свиного BDNF. С каждым зондом у всех испытанных организмов за исключением дрожжей наблюдались единственные полосы. В большинстве случаев полосы, гибридизирующие с NGF и BDNF-зондами, в любом организме имели разную подвижность при электрофорезе, хотя в некоторых случаях (например, мышиная ДНК), EcoRI фрагменты, гибридизирующие кNGF и BDNF-зондам, были примерно одного размера, и не могли быть разделены при использованных условиях электрофореза (фиг. 3).
Часть кодирующих последовательностей для зрелого BDNF была амплифицирована PCR из геномной ДНК свиньи, крысы, цыпленка и человека, и нуклеотидные последовательности определялись. Анализ последовательности ДНК амплифицированной области от свиной геномной ДНК точно подтвердили результаты по последовательности, полученные молекулярным клонированием от свиной мозговой кДНК. Были также определены геномные последовательности крысы, человека и цыпленка BDNF для 252 бр сегмента (фиг. 5). Следует заметить, что у крысы и человека выведенная аминокислотная последовательность для области по меньшей мере из аминокислот от 28-III была идентичной последовательности свиного BDNF, хотя у разных видов наблюдалось несколько нуклеотидных различий (например, консервативные изменения в третьей позиции кодона). У цыпленка в этой области наблюдалось единственное аминокислотное замещение; остаток 61 зрелого протеина, это лизин у цыпленка по сравнению с метионином у BDNF млекопитающих. Данные анализа последовательности вместе с данными эксперимента по саузерн-блот-гибридизации обеспечивают однозначное доказательство того, что BDNF кодирован высококонсервированным геном, отличающимся от гена, кодирующего NGF.
8. Пример. Экспрессия BDNF РНК в нервных и других тканях.
8.1. Материалы и методы.
8.1.1. Приготовление РНК.
Общую РНК экстрагировали от взрослых самок мышей согласно Окаяме и др. (1987, Methods Enzymol, 154: 3-28).
Вкратце, замороженные ткани гомогенизировали в 5,5 М гуанидинтиоцианате, лизат центрифугировали для удаления мусора, и супернатант, перенесенный на подложку из трифторацетата цезия, доводили до плотности 1,51 мкг/мл. После 24 ч центрифугирования при 125000g в sw-27 бакет-роторе (Бекманн) РНК ресуспендировали и преципитировали этанолом и 8 М ацетатом аммония и хранили при -70oC. Электрофорез проводили согласно Леражу и др. (Lehrach, 1977, Biochemestry 16:4743-4571) на 1,3% агарозоформальдегидных гелях. РНК переносили на нелоновые мембраны (Hybond-N, Amerscham) и гибридизовали до утра при 62oC в 1 мл 2 х SSC, содержащем 50% формамида с 32P-кРНК зондом мышиного BDNF (107 срм, см.ниже). Промывка длилась 60 мин при 65oC в 0,1 х SSC. После промывки блок инкубировали в течение 60 мин при комнатной температуре с 0,1 мкг/мл РНКазы A (Фармация) и пленку экспонировали при -70oC (с интенсифицирующим экраном) в течение 48 часов.
8.1.2. Приготовление кРНК зонда.
Библиотеку мышиной мозговой кДНК скринировали с двумя независимыми BDNF олигонуклеотидами. Двойные положительные клоны выделяли и субклонировали в EcoRI сайт Bluescript SK+ плазмиды (Стратаген), определяли нуклеотидную последовательность, соответствующую нуклеотидам 350-829 свиной последовательности (см. фиг. 1). В этой последовательности было обнаружено всего только 4 аминокислотных отличия между мышиным и свиным BDNF, указывая на значительную степень консервирования этого домена между свиным и мышиным BDNF. Единственный зонд нитевой РНК был приготовлен с использованием этой матрицы и T3 полимеразы (Промега). Специфическая активность этого зонда составила 108 срм/мкг.
8.2. Результаты и обсуждение.
Для оценки экспрессии BDNF мРНК в нервной по сравнению с другими тканями использовали назерн-блот-анализ. Назерн-блот-анализ осуществляли с использованием мышиных тканей, позволяющих более быстрый процесс экстракции РНК, чем свиные ткани. 32P-кДНК зонд обнаружил сигнал примерно 1,45 кб в мозге (фиг. 4) и спинном мозге (данные не показаны). Важно, что не было обнаружено сигнала ни в одной другой ткани, включая почки, кишечник, легкие, печень, селезенку, сердце и мышцы (фиг. 4). Допуская, что размер свиной мРНК подобен размеру мРНК мыши, показанная на фиг. 2 последовательность кДНК представляет более 80% полной мРНК последовательности.
Важным наблюдением в трансдифференцировке физиологии BDNF является то, что мРНК, кодирующая этот протеин, обнаруживалась только в центральной нервной системе, и не обнаруживалась в семи других, не нервных тканях, BDNF мРНК находилась не только в головном мозге, но также и в спинном мозге, и в superior colliculus (последовательность, представленная на фиг. 1, получена полностью из кДНК-матрицы superior colliculus). Эти данные подтверждают идею, что BDNF является нейротропным фактором мишенного происхождения, и что реагирующие на BDNF-нейроны либо принадлежат ЦНС, либо непосредственно связаны со структурами ЦНС. Действительно, все нейроны, которые, как известно, реагируют на BDNF, либо проникают в ЦНС, как нейроны дорсального корня или черепных сенсорных ганглиев (Линдсей, 1985, Дев.Биол. 112: 319-328; Дейвис, 1986, Джорнал Нейросайенс, 6:1897-1904), или являются ЦНС-нейронами, как клетки ганглия сетчатки (Джонсон, 1986, Дж. Нейрос, 6:3031-3038). Понятно, необходимо еще провести подробные исследования, чтобы изучить точное распределение мест синтеза BDNF в центральной нервной системе, но уже теперь очевидно, что распределение BDNF мРНК очень отличается от распределения NGF мРНК, обнаруживаемой во многих не-ЦНС тканях (Хойманн, 1984, ЕМВО 3: 3183-3189; Шелтон, 1984, Proc. Nath. Acad. Sci. США, 81: 7951-7955).
9. Пример. Молекулярное клонирование и характеристика человеческого и крысиного BDNF-генов.
9.1. Материалы и методы.
9.1.1. Библиотеки геномной ДНК и кДНК.
Библиотека кДНК сетчатки взрослого человека в лямбда-ZAPII была получена от Стратагена. Библиотека геномной ДНК плаценты человека в EMBL3/SP6/T7 была получена от Клонтеха. Библиотека кДНК мозга человеческого плода в λ gtII была получена от Клонтеха. Библиотека крысиной геномной ДНК в EMBL3/SP6/T7 была получена от Клонтеха. Обе геномные библиотеки были подготовлены частичным перевариванием геномной ДНК рестрикционной нуклеазой Sau3A и лигированием в BamHI сайт вектора. Библиотека кДНК крысиного мозга была получена в лямбда-ZAPII от Стратагена.
9.1.2. Приготовление BDNF ДНК зондов.
32P-меченные BDNF ДНК зонды приготавливали с использованием тех же самых олигонуклеотидных праймеров, описанных в разделе 7.1.1, выше, в реакции PCR с человеческой геномной ДНК, чтобы амплифицировать кодирующую область для остатков 28-III человеческого BDNF. Параллельно был получен специфичный для крысиного BDNF 32P-меченый зонд, с использованием крысиной геномной ДНК в качестве матрицы для PCR реакции.
9.1.3. Скрининг библиотек.
Лямбда-фаговые библиотеки подвергали скринингу по стандартным методикам (Бентон и Дейвис, 1977, Сайенс, 196: 180-182; Маниатис и др., 1978, Cell, 15: 687-701), гибридизируя в 50%-ном формальдегиде с декстран-сульфатом и раствором Денхардта при 42oC. Фильтры предварительно гибридизировали при 42oC в 50%-ном формамиде. 5хSSCPE 10% Денхардта, 0,5 мг/мл ДНК спермы лосося, 0,1% SDS и 10% декстран-сульфата. Гибридизацию осуществляли в том же буфере за исключением того, что раствора Денхардта было 2%. ДНК спермы лосося было 0,1 мг/мл, а SDS дестрана сульфат отсутствовали. После гибридизации фильтры промывали при 68oC. Зонд человеческого BDNF использовали для скрининга человеческой геномной ДНК-библиотеки и библиотеки кДНКсетчатки; зонд крысиного BDNF использовали для скрининга крысиной геномной ДНК библиотеки и библиотеки кДНК мозга. Библиотеки также скринировали с зондами человеческого и крысиного NGF приготовленными как описано выше в разделе 7.1.1.
9.2. Результаты и обсуждение.
По меньшей мере 670 тыс. пятен было скринировано от каждой библиотеки. Положительными человеческими клонами считались такие из них, которые гибридизировались с зондом человеческого BDNF, но не гибридизировались с описанным выше зондом человеческого NGF. BDNF геномные клоны были получены от человеческой и от крысиной библиотек с частотой, согласующейся с наличием одной копии BDNF-гена на гаплоидный геном. Для крысиной и человеческой библиотек было скринировано приблизительно миллион бляшек. Три положительные были получены из крысиной геномной библиотеки и одна - из человеческой геномной библиотеки. Положительные клоны из библиотек кДНК человеческой сетчатки и крысиного мозга были получены с частотой, согласующейся с тем, что ген экспрессируется на очень низких уровнях; в кДНК крысиного мозга было идентифицировано два положительных клона из 670 тыс.; в библиотеке кДНК человеческой сетчатки был идентифицирован один положительный клон из 670 тысяч. Не было обнаружено положительных клонов из 670 тысяч от библиотеки ДНК, приготовленной из мозга человеческого плода. Нуклеотидное секвенирование проводили на человеческой BDNF кДНК и геномных клонах с использованием синтетических олигонуклеотидных праймеров, представляющих точные последовательности из человеческой и крысиной BDNF кодирующей области, как определено в разделе 7.2.1, выше. Длиннейший полученный клон человеческой кДНК имел инсерт примерно от 1,6 до 1,8 кбр и, как ожидалось, содержал точную последовательность части человеческого BDNF, определенного после прямого амплифицирования из человеческой геномной ДНК, как описано в разделе 7.2, выше. Подробный анализ последовательности этого кДНК (фиг. 5) клона обнаружил открытую рамку считывания, кодирующую полипептид из 247 аминокислот, похожий, но не идентичный полнодлинному предшественнику свиного BDNF. Внутри области, соответствующей зрелому BDNF-полипептиду (например, от кодона для His 134 до кодона цепного терминирования) не было обнаружено различий в выведенной аминокислотной последовательности; все нуклеотидные замещения между свиньей и человеком были консервативными по отношению к кодирующей специфичности. Остаток полипептида - предполагаемого предшественника BDNF показал некоторые различия у аминокислотных последовательностей человека и свиньи, наиболее заметно отсутствие в гене человеческого BDNF пяти из шести последовательных Ser кодонов, имеющихся у свиньи, приводящее к несколько более короткому полипептиду (247 против 252 а.а.).
Библиотеки, приготовленные в векторе EMBL3, имеют инсерты в 10-23 кбр чужеродной ДНК. Точный размер инсерта в детально проанализированном клоне человеческого геномного BDNF не был определен. Однако этот клон, содержащий единственный фрагмент рестрикционной эндонуклеазы EcoRI, приблизительно в 4 кбр, который гибридизировался с меченым BDNF-зондом, использовали для скрининга библиотеки. Этот фрагмент имеет ожидаемый размер основанный на результатах саузерн-блот-гибридизации человеческой геномной ДНК со свиным BDNF-зондом, описанным выше. Анализ последовательности проводили на человеческом клоне с использованием синтетических олигонуклеотидов, представляющих кДНК последовательность для примирования ДНК синтеза от бактериофаговой ДНК матрицы. Было обнаружено, что последовательность кодирующей последовательности для предполагаемого предшественника человеческого BDNF является идентичной последовательности в клоне человеческой кДНК, за исключением единственного нуклеотидного замещения в prepro-области, соответствующей аминокислотной замене метионина (АТГ) на валин (ГТГ) у нуклеотида 785, см. фиг. 5. Это изменение может отражать полиморфизм в человеческом геноме. Как в случае человеческого NGF гена, не было обнаружено ни одного нитрона внутри кодирующей последовательности для предполагаемого человеческого BDNF. Данные по крысиной кДНК-последовательности также представлены на фиг. 5.
10. Пример. Экспрессия рекомбинантного BDNF.
10.1. Материалы и методы.
10.1.1. Приготовление вектора экспрессии BDNF.
Последовательность, соответствующая свиному prepro BDNF была приготовлена с
использованием
олигонуклеотидных праймеров (по 150 pmoI каждый):
ATAATCTAGATGACCATCCTTTTCCTT (смысловой нити)
ATAATCTAGACTATCTTCCCCTCTTAAT (антисмысловой)
в реакции PCR и с
использованием
1 мкг свиной геномной матрицы (каждый праймер имеет добавленный сайт). Реакцию амплификации осуществляли как описано за исключением того, что температура ренатурации составляла 50oC. После
переваривания ХбаI амплифицированную ДНК лигировали в ХбаI сайт плазмиды pCMV для продуцирования pCMV1-pBDNF-1 (-1 означает смысловую ориентацию на фиг. 6 и
соответствует
COS+ в табл. III), и плазмиду вводили в бактерию XL-1 посредством электропорации.
10.1.2. Экспрессия BDNF в COS-клетках.
pCMVI-pBDNF ДНК из положительных клонов (проверенных гибридизацией с oligo 5, фиг. 2) была вырезана из ХбаI и PstI. Размер полученных продуктов позволил определить ориентацию инсертов, и обе плазмиды были использованы для трансфекции COS клеток по кальциево-фосфатному методу (Чен и др., 1987, Молекулярная клеточная биология, 7: 2745-2752), и ростовую среду собирали через 24 часа. BDNF-активность испытывали в биоэксперименте на ганглии дорсального корня куриного эмбриона, подробно обсуждавшегося выше.
10.2. Результаты и обсуждение.
Сенсорные нейроны спинного мозга цыпленка E8 помещали на чашку с плотностью 6 тысяч на лунку, инкубировали 24 часа и подсчитывали через 24 часа (Линдсей и др., 1985, развитие биологии, 112: 319-328). Оценки представляют среднее из трех определений ± стандартная девиация. BDNF и NGE использовались при 1 нг/мл концентрациях, при которых наблюдалось максимальное выживание при каждом факторе. COS+ относится к клеткам, трансфецированным плазмидами, содержащими BDNF инсерт в смысловой ориентации, а COS- относится к клеткам, трансфецированным плазмидами, содержащими BDNF инсерты в обратной ориентации. COS относится к нетрансфецированным клеткам. При разбавлениях более 1:20, не наблюдалось выживания выше контроля в COS- и COS кондиционированных средах. Во всех эспериментах без NGF моноклональные антитела против NGF (Коршинг и др., 1988, Proc. Nath. Acad. Sci. США, 80: 3513-3516) использовали при концентрации 1 мкг/мл.
Как показано в табл. III, только среда, полученная от культур COS клеток, несущих pCMVI -pBDNF плазмиду в смысловой ориентации, была связана с выживанием сенсорных нейронов цыпленка сверх контрольных значений. Поэтому вывод - рекомбинантный BDNF является биологически активным. Далее, добавление BDNF, выделенного из свиного мозга, не повышало выживание сенсорных нейронов значительно выше уровней выживания, достигнутых только одним рекомбинантным BDNF, тем самым указывая, что рекомбинантный BDNF способен насыщать BDNF-рецепторы. Наблюдалось также, что рекомбинантный BDNF обладает дополняющим и синергическим действием, способствующим выживанию сенсорных нейронов цыпленка, при использовании его вместе с NGF.
11. Пример. Генерирование антител к BDNF.
11.1. Материалы и методы.
Поликлональную антисыворотку, специфичную для BDNF, обозначенную sera4, генерировали в NZ белого кролика иммунизацией синтетическим пептидом.
11.1.1. Синтез пептидов и их соединение с носителем.
Пептид, состоящий из 34 аминокислотных остатков, обозначенный B5 был синтезирован конвенциональными (обычными) методами. Этот пептид имеет
следующую
аминокислотную последовательность, соответствующую 33 остаткам зрелого BDNF (остатки 153-185 полной свиной prepro BDNF последовательности, показанной на фиг. 1) с дополнительным цистеиновым
остатком у
аминотерминала (показанного прописным шрифтом) для обеспечения присоединения к протеину-носителю с использованием м-малеимидо бензойной кислоты -N- гидроксисункцинимида (MBS) по
желанию:
Cys-Val-Thr-Ala-Ala-Asp-Lys-Lys-Thr-Ala- Val-Asp-Met-Ser-Gly-Gly-Thr-Val-Thr- Val-Leu-Glu-Lys-Val-Pro-Val-Ser-Lys-Gly- Gln-Leu-Lys-Gln-Tyr
B5 пептид был соединен с бычьим
сывороточным
альбумином с использованием бис-диазо-бензидина (BDB). Свежий BDB был приготовлен путем растворения 46 мг бензидин-HСl. 35 мг NaNO2 растворяли в 1,0 мл H2O и
добавляли его к
раствору бензидина, перемешивали 1 час при 4oC.
21 мг BSA растворяли в 3,0 мл буфера борат-CaCl2, pH 9,0. Примерно 15 мг B5 пептида растворяли в 1,5 мл борат-NaCl буфера, pH 6,0. Раствор пептида добавляли в раствор BSA и помещали на лед, 1,0 мл BOB добавляли к BSA-пептидному раствору, и реакционную смесь инкубировали при перемешивании в течение 2 часов при 4oC; pH наблюдали при этом и поддерживали на уровне 9,0 путем добавления небольших количеств 0,5 М NaOH, как требовалось. Реакцию прекращали добавлением 0,2 мл 1% буферированного фенолом раствора. Излишек реагентов удаляли диализом против буферированного фосфатом физиологического раствора (PBS).
11.1.2. Иммунизация.
Всего
шесть кроликов было
иммунизировано в соответствии со следующими процедурами:
кролики 1 и 4 - пептидом B5, спаренным (соединенным) у его C-конца к BSA с использованием BDB;
кролики 2
и 3 - пептидом B5,
соединенным у его N-конца к BSA с использованием MBS;
кролики 5 и 6 - пептидом B5, смешанным с порошкообразной нитроцеллюлозой.
Во всех случаях на первую иммунизацию использовали 1 мг иммуногена (100 мкг B5/500 мкг нитроцеллюлозы для кроликов 5 и 6) в 0,5 мл PBS + 0,5 мл полного адъюванта Фрейнда. Эту смесь вводили подкожной инъекцией в несколько мест на спине. Вторую иммунизацию проводили тремя неделями позже, и она была идентичной первой, за исключением того, что вместо полного адъюванта Фрейда использовали неполный адъювант его же. Последующие инъекции осуществляли с интервалом в 4-6 недель. Через 1 неделю после иммунизации у кроликов брали кровь и обычно проверяли антисыворотку на связывание к чистому B5 пептиду в эксперименте на иммуносорбент, связанный с энзимом (ELISA).
11.1.3. Обнаружение связывания антител с BDNF.
100 мкг антигена (B5 пептид) в H2O добавляли в лунки чашек для микотитрования и оставляли до утра высохнуть, затем кратковременно промывали водой и блокировали 100 мкг 1%-ного желатина в течение 30 мин при комнатной температуре. Лунки промывали трижды дистиллированной водой, а затем добавляли 100 мкг антисыворотки и оставляли при 4oC до утра. Лунки затем промывали трижды в PBS/0,05% тритон X-100, после чего в лунки добавляли 100 мкг пероксидазного меченого антикроличьего разведения иммунопробы (1:1000) и инкубировали при комнатной температуре в течение трех часов. Лунки промывали дважды, и добавляли 100 мкг раствора ABTS (10 мг ABTS "Сигма" растворяли в 10 мл 0,1 М натрия цитрата pH 4,0 плюс 10 мкг H2O2) и инкубировали примерно 5 минут, пока не появится цвет. Реакцию прекращали добавлением 10 мкг 1% NaN3. Образцы разводили 1:5 водой и измеряли оптическую плотность при 415 нм.
11.2. Результаты и обсуждение.
Антисыворотка от кролика 4 (зега 4) показала наивысший титр (фиг. 7A) и была использована в дальнейших экспериментах. Антитела из сыворотки 4 были частично выделены преципитацией с фосфатом аммония; и к части антисыворотки медленно добавляли равный объем насыщенного сульфата аммония при перемешивании, и раствор перемешивали еще 15 минут, а затем центрифугировали при ускорении 2000g. Дебрис промывали дважды в 50% насыщенном сульфате аммония, затем снова растворяли в PBS в объеме, соответствующем исходному объему сыворотки. Сульфат аммония удаляли диализом при нескольких заменах PBS. Диализованный раствор аликвотировали в 1,0 мл объемах и подвергали лиофилизации с использованием спидвака. Образец антител сыворотки 4 ресуспендировали в 0,5 мл воды и испытывали на реактивность с пептидом B5 посредством ELISA. Реакция была обнаружена при разведении 1:4000.
Поликлональные антитела к синтетическому пептиду (B5), соответствующему фрагменту из 33 аминокислот свиного BDNF, генерировали иммунизацией кроликов как описано выше. Сыворотка 4, показавшая наиболее высокий титр против синтетического пептида, показала реактивность с очищенным BDNF из свиного мозга посредством ELISA (фиг. 7B). Слабая реактивность была также обнаружена при иммуноблоттинге (данные не показаны). Однако антисыворотка была неспособна блокировать активность BDNF в биоэксперименте на сенсорных нейронах дорсального корня куриного эмбриона.
12. Пример. Новые биологические действия BDHF.
Следующие наблюдения указывают, что BDNF способен:
(I) поддерживать выживание и вызывание полностью
дифференцированного состояния
допаминергических нейронов ЦНС;
(II) поддерживать выживание холинергических нейронов ЦНС; и
(III) подавлять пролиферацию астроглиальных клеток. Эти
биологические действия BDNF ранее
не описывались. Поскольку допаминергические нейроны, холинергические нейроны и астроглиальные клетки могут быть связаны нервными болезнями или расстройствами, BDNF
может оказаться полезным при лечении
невропатологий, включающих эти клеточные популяции.
12.1. Материалы и методы.
12.1.1. Методы культурирования допаминергических нейронов substantia nigra.
Вентральный средний мозг извлекали диссекцией из мозгов крысиных эмбрионов различного возраста от 13 дня эмбрионального развития (E13) до 15 эмбрионального
дня. Обычно в каждом эксперименте
использовали образцы от двух пометов. Диссекционный раствор имел следующий состав:
NaCl 138,8 мМ; KCl 2,7 мМ; Na2HPO4 • 7H2O 8,0 мМ; KH2
PO4 1,5 мM; глюкоза 6 мг/мл; BSA 0,1 мг/мл, pH 7,4. Этот раствор приготавливали с последующей стерилизацией через фильтр с порами 0,2 мкм. Диссекцию
осуществляли в нестерильных условиях.
Когда ткань была извлечена из всех мозгов, остаток процедуры проводили в стерильных условиях. Фрагменты ткани помещали в 35 мм культуральную чашку и измельчали
маленькими ножницами. Два миллилитра
питательной среды F-12, содержащей 0,125% трипсина, добавляли затем к ткани и инкубировали при 37oC. В конце этого периода инкубации к кашице добавляли
DNAse 1 так, что конечная
концентрация составляла 80 нг/мл. Осуществляли другую идентичную инкубацию, и тканевую кашицу впоследствии добавляли в 8,0 мл ростовой среды, состоящей из минимальной
необходимой среды (МЕМ) с
добавлением 2 мМ глютамина, 6 мг/мл глюкозы, 5 ед/мл стрептомицина и 7,5% сыворотки плода теленка (FCS). Образец центрифугировали на центрифужном столике при комнатной
температуре и 500 об/мин в
течение 5 мин. Среду извлекали всасыванием, и 2 мл ростовой среды добавляли к клеточному дебрису. Отожженной пипеткой с отверстием 1 мм тритурировали клетки восемь раз.
Оставшимся клеточным фрагментом
позволили осесть под действием силы тяжести и взяли небольшую аликвоту супернатанта для оценки количества клеток путем подсчета на гемоситометре. После определения
клеточной плотности эти клетки
помещали на чашки тканевого культурирования с плотностью 50 тыс. клеток на см2.
Культуральные чашки приготавливали за день до диссекции. Тканевые чашки (24 лунки, 2 см2 на лунку) были предварительно покрыты полиорнитином (молекулярный вес 30000 - 70000 г/моль) в количестве 0,5 мг/мл при комнатной температуре в течение 3 часов. Чашки обильно промывали водой и впоследствии обрабатывали мышиным ламинином, 5 мкг/мл при комнатной температуре в течение 3 часов. Чашки затем промывали водой как выше и инкубировали до утра при 37oC в увлажненной атмосфере, состоящей из 5% CO2, 95% воздуха в присутствии ростовой среды. Среду на чашках удаляли на следующий день и заменяли свежей ростовой средой.
Когда клетки были помещены на культуральные чашки, клетки ставили в инкубатор при 37oC и 5% CO2 (95% воздуха) в течение 24 часов. Культуральную среду заменяли на свободный от серы состав (SFM) 1:1 (об./об.): смесь среды ВЕМ (Basal eagle Medium) и питательной среды F-12 с глюкозой (33 мМ), глютамином (2 мМ), NaHCO3 (15 мМ), HEPES (10 мМ) с добавлением инсулина (25 мкг/мл), трансферина (100 мкг/мл, путресина (60 мкг/мл), прогестерона (20 нМ), селенита натрия (30 нМ), пенициллина (5 ед/мл), стрептомицина (5 мг/мл) и T3 (30 нМ). В некоторых экспериментах очищенный BDNF добавляли к культурам после замены среды на SFM на 2 день культурирования.
Растворы, использовавшиеся для культурирования допаминергических нейронов, приготавливали с использованием воды, взятой из водной системы реагента Милли-Q. Состав культуральной тканевой среды был получен от лабораторий ЖИБКО (Gibco, Santa Clara, California), так же как и сыворотка плода теленка (номер партии 43 N 1086) и мышиный ламинин. Все другие компоненты среды были закуплены у "Сигама Кемикал" (St. Louis, MO) и были тестированного сорта на клеточных культурах. Полиорнитин и DNAseI были также получены от Сигама. Трипсин был получен от Worthington (Freehold, NJ), номер партии 3667. Коммерческие химикаты были аналитической чистоты и закуплены у Бейкер Кемикал (Phillipsburg, NJ). BDNF, использовавшийся в этих экспериментах, был очищен из свиного мозга др-ом Бардом согласно его методике (У.А. Barde, 1982, см. выше).
12.1.12. Методы иммуноцитохимического окрашивания культур из вентрального среднего мозга.
Фиксирующие растворы приготавливали свежими для каждого эксперимента. Для окрашивания тирозин-гидроксилазой (TH) фиксатором был 4,0% параформальдегид в фосфатном буфере Соренсона. Буфер Соренсона приготавливали путем добавления 0,2 М раствора KH2PO4 к 0,2 М NaHPO4, пока pH не достигло 7,3. Параформальдегид в последующем добавляли к раствору и кратковременно нагревали, чтобы он растворился, и охлаждали до комнатной температуры перед использованием.
Для начала процедуры, культуральную среду удаляли из культуральных чашек осторожным всасыванием, и осторожно добавляли в чашку подходящий фиксирующий раствор. Проводили инкубирование при комнатной температуре в течение 20 минут. Затем следовали три промывки в фосфатном буфере Соренсона, по 5 мин каждая, с легким вращением. Клетки затем инкубировали в гасящем растворе в течение 30 мин при комнатной температуре с легким вращением. Гасящий раствор для окрашиваемых культур для ТН, состоял из фосфатного буфера Соренсона, содержащего 2% нормальной лошадиной сыворотки. Затем, культуры инкубировали в пермеабилизационном (для улучшения проницаемости) буфере при комнатной температуре в течение 30 минут при легком вращении. Для культур, окрашиваемых для ТН, раствор состоял из буфера Соренсона с 0,2% сапонина и 1,5% нормальной лошадиной сыворотки. После стадии повышения проницаемости, культуры инкубировали в присутствии первичных антител до утра при 4oC. Антителами против крысиного ТН были мышиные моноклональные антитела изотипа IgG 2a. Их использовали в концентрации 40 мкг/мл в растворе из 10 мМ NaPO4, 50 мМ NaCl, 0,2% сапонина и pH 7,5. После инкубирования первичного антитела культуры промывали 5 раз по 15 минут каждый раз в соответствующем пермеабилизационном буфере. Затем культуры инкубировали со вторичным антителом, конъюгированным к биотину, а именно - с биотинированным лошадиным антимышиным IgG. Это инкубирование осуществляли при комнатной температуре в течение двух часов с легким вращением. Далее следовали промывки, идентичные описанным выше, и затем культуры инкубировали в присутствии предварительно сформированного авидин-биотинированного комплекса хреновой пероксидазы (реагент ABC, лаборатории Вектор, Burlingame, CA), приготовленного согласно инструкциям производителя. Через 30 мин инкубирования при комнатной температуре с легким вращением эти культуры промывали, как описано выше. Культуры затем инкубировали с 55 мМ Трис-HCl pH 7, 3, содержащим 0,5 мг/мл диаминобензидина и 0,01% перекиси водорода. Реакционному продукту позволяли развиваться в течение 2-5 минут, после чего раствор удаляли и культуры промывали несколько раз ледяным PBS. Затем определяли количество положительных клеток на см2.
Параформальдегид и глютаральдегид были получены от Fluka Chemical. Наборы "Vectastain", содержащие нормальную сыворотку (используемую как блокирующий агент), биотинированный аффинно-очищенный антииммуногемоглобин, авидин DH, и биотинированный HRP-H были закуплены у Вектор. Диаминобензидин был получен от BRL (Gaithersberg, MD).
12.1.3. Методы, использованные для измерения потребления 3H-допамина в культурах вентрального среднего мозга.
Исследование потребления 3H-допамина (3H-DA) осуществляли как описано Дал Тосо и др. (Dal Toso, 1988, J. Neurosci 8: 733-745) с небольшими модификациями. Буфер для этого имел следующий состав: NaCl 136,8 мМ; KCl 2, 7 мМ; Na2HPO4 • 7H2O 8,0 мM; KH2PO4 1,5 мМ; глюкоза 5,0 мМ; CaCl2 1,0 мМ; MgSO4 1,0 мМ; аскорбиновая кислота 0,1 мМ; pargyline 0,1 мМ с pH 7,4. Когда необходимо, к этому буферу добавляют 5,0 мкМ BZT (benzotropine mesylate).
Клетки промывали один раз с предварительно нагретым (37oC) буфером и помещали в 2 см2 лунки по 0,4 мл этого буфера. Культуры затем предварительно инкубировали в течение 5 мин при 37oC. В конце этой предварительной инкубации в этот буфер (250 нM, 40 Ci/мМ) добавляли 0,1 мл 3H-ДА так, что конечная концентрация H-ДА в буферу составляла 50 нM. Культуры инкубировали при 37oC в течение 15 минут, с последующими четырьмя промывками при 4oC по 0,5 мл каждая с описанным буфером. Две дополнительные промывки осуществляли с ледяным PBS (10 мМ NaPO4, 150 мМ NaCl, pH 7,6). После того как была завершена последняя промывка к клеткам на 2 см2 лунки добавляли по 0,2 мл 0,5н. NaOH и оставляли стоять при комнатной температуре в течение двух часов. Затем собирали экстракт NaOH и подсчитывали в сцинтилляционном счетчике ("Паккард", LS 500 TD) с 10 мл сцинтилляционной жидкости "ультимагольд". Специфическое определение потребления осуществляли путем прекращения потребления в присутствии 5 мкM BZT. Обычно оно составляло 70-90% общего потребления.
3H-ДА был получен от NEN (Бостон, МА). Аскорбат, паргилин, BZT и глюкозу получили от Сигама (C-Луис, МО). Ультимагольд сцинтилляционная жидкость была закуплена у Паккарда (Сетерлинг, VA).
12.1.4. Методы получения культур холинергических нейронов базального переднего мозга.
Первичные культуры холинергических клеток базального переднего мозга были получены от крыс 17 эмбрионального дня. Конкретно, холинергические нейроны, использовавшиеся в этом исследовании, были получены из медиального перегородочного нуклеуса и нуклеуса диагональной полосы Брока. Эта нейронная популяция первично проникает в аммонов рог. Диссоциированные смешанные культуры (нейроны и глия) получали следующим образом.
Перегородочную область высвобождали диссекцией из окружающей тканей и удаляли мозг плода. Куски ткани затем собирали, измельчали ножницами и обрабатывали в течение 20 мин при 37oC 12,5%-ным трипсином. Трипсин инактивировали разведением в чашечной среде (среда МЕМ, модифицированная Дульбекко (ДМЕМ), которая содержала 1% пенициллина и стрептомицина, 5% лошадиной сыворотки и 1% N3-гормональной добавки). Суспензию (единственную) клеток получали тритурированием переваренных тканевых фрагментов с отожженной пастеровской пипеткой. Диссоциированные клетки подсчитывали на гемоцитометре и помещали на чашки при соответствующей плотности в чашечную среду. К культурам через 5-6 часов после посева добавляли тестовые лиганды, а затем клетки выращивали ин витро в течение десяти дней со сменой среды каждые три дня.
12.1.5. Испытания с холин-ацетил-трансферазой.
После обработки клетки использовали либо в экспериментах на фермент CAT (холин-ацетилтрансфераза) или проводили иммуногистохимическое окрашивание на CAT по следующей методике. Моноклональные антитела к CAT были закуплены от Берингер Маннгейм Биокемикал Ко. В конце эксперимента клетки дважды промывали ДМЕМ. Культуры фиксировали двухстадийной процедурой; 50 мкл 4%-ного параформальдегида добавляли к 50 мкл ДМЕМ для десятиминутного инкубирования. Этот раствор удаляли и заменяли 100 мкл 4%-ного формальдегида, и инкубацию продолжали 30 мин при комнатной температуре. После фиксирования клетки промывали трижды буферированным фосфатом физиологическим раствором (PBS) и делали их проницаемыми 30 минутным инкубированием с сапонином 0,5 мг/мл. Детергент удаляли тремя промывками PBS и на 30 минут добавляли блокирующий раствор 5% нормальной сыворотки кролика. Первичное антитело добавляли при разведении 1: 3 в 1% нормальной сыворотке кролика, после удаления блокирующего раствора, и культуры инкубировали до утра при 4oC. Этот раствор, содержащий первичное антитело, удаляли промывкой PBS. Связанный иммуноглобулин обнаруживали методом Vectastain "ABC". Диаминобензидина тетрагидрохлорид (ДАВ) использовали в качестве субстрата для реакции пероксидазы, которую обычно проводили в течение 1-5 минут. Реакцию прекращали ополаскиванием культур дважды 0,1 М трис-HCl pH 7.2. Культуры хранили в 50 мМ трис, pH 7,6, содержащем 0,15 М NaCl, 42% глицерина и 0,15% Lephiran (Pierce Chemical Co., Rockvilli, IL) при 4oC.
12.1.6. Метод генерирования очищенных культур астроглиальных клеток.
Очищенные глиальные культуры приготавливали в основном методом Мак Карти и ДеВелли (McCarthy, K.D. и Dwellis, J. 1980, J. Cell. Biol 85, 890-902) из аммонова рога крысы первого - второго постнатального дня. Аммоновы рога удаляли у пяти щенков и измельчали ножницами. Кусочки ткани затем переваривали в 2 mls 0,125% трипсина в течение 20 мин при 37oC. Протеазу инактивировали разведением, чашечной средой (10% сыворотки плода теленка (Жибко), ДМЕМ, 0,5% пенициллина (5000 мкг/мл) и 0,5% глютамина). Единственная клеточная суспензия была получена пропусканием переваренных фрагментов ткани через пастеровскую пипетку соперетяжкой. Клетки пелетировали центрифугированием при 900 об/мин в течение 5 мин, ресуспендировали в чашечной среде и затем подсчитывали на земс-цитометре. Единственная клеточная суспензия была разделена на три, каждую - в емкость для тканевого культурирования площадью 75 см2, и клетки выращивали примерно до 80%-ного слияния. Клетки затем субкультурировали методом трипсинизации, подобным только что описанному. Глиальные клетки подсчитывали и помещали на чашку с плотностью 10 тыс. клеток на 0,9 см2.
12.2. Результаты.
12.2.1. Действие BDNF на тирозин-гидроксилазу, присутствующую в культурах вентрального среднего мозга.
Иммуноцитохимическое окрашивание, как оно описано в разделе 12.1.2., выше, использовали для оценки действия BDNF на количество клеток, положительных на тирозин-гидроксилазу (TH) (фиг. 8). Максимальное увеличение более чем на 200% контроля наблюдалось на 8-й день у BDNF-стимулированных культур клеток вентрального среднего мозга. Раньше, на 3-й день у BDNF-стимулированных культур наблюдалось очень слабое увеличение.
12.2.2. Действие BDNF на потребление допамина культурами вентрального среднего мозга.
3H-допамина (3H-ДА) потребление было измерено по методу Даль Тасо и др. (1988, J. Neurosci 8:733-745) с незначительными модификациями, как описано в разделе 12.1.3, выше. Небольшое увеличение усвоения допамина наблюдалось у BDNF-стимулированных культурах вентрального среднего мозга на 8-й день культурирования (фиг. 9).
12.2.3. Действие BDNF на экспрессию холинацетилтрансферазы холинергическими нейронами переднего мозга.
На фиг.10(A) изображен эффект BDNF на количество CAT-положительных клеток после периода роста в 12 дней ин витро. 5,9-кратное увеличение количества CAT-клеток наблюдалось с добавлением 100 нг/мл BDNF, а рассчитанное значение для EC50 составило 10 нг/мл. В качестве положительного контроля использовали культуры с плотностью 260 тысяч (черная полоса) или 150 тысяч (прерывистая полоса) клеток на лунку, обрабатывали тем же образом, но с NGF (фиг.10B). Плотность 260 тысяч клеток на лунку соответствовала плотности, использовавшейся при исследовании BDNF. Это увеличение количества CAT-иммуноположительных клеток является сходным увеличению, о котором было сообщено ранее. Потенциальная способность BDNF действовать на холинергические нейроны была также проверена путем измерения CAT-энзимной активности (F.Fonnum; I. Neurochemistry, 1975, 24: 407-409). На фиг.11 изображены изменения в CAT, полученные обработкой BDNF. В этом случае было достигнуто 1,8-кратное увеличение с 100 нг/мл BDNF и EC 50 был рассчитан в 61 нг/мл.
12.2.4. Действие BDNF или EGF на культуры астроглиальных клеток.
Было продемонстрировано, что астроциты типа II обладают высокой аффинностью с рецепторами множества нейротрансмиттеров и нейропептидов. Так, астроциты способны реагировать на сигналы нейронного происхождения. По этой причине, а также вследствие того, что астроциты типа II представляют клеточный компонент в первичных культурах, испытывали возможный прямой эффект BDNF на глиальные клетки. Эти клетки держали четыре дня ин витро перед добавлением факторов роста, чтобы процент их слияния достиг приблизительно 60%, а затем обрабатывали 42 часа EGF и BDNF. После 18 часов инкубации в среде присутствовал [3H] метилтимидин. Действие EGF показано на фиг.12A. Как сообщалось ранее, обнаружено, что EGF является митогенным для астроцитов. Максимальная реакция наблюдалась с 10 нг/мл EGF, который дал 5,2-кратное увеличение инкорпорирования [3H] метилтимидина. На фиг.12(B) изображено влияние BDNF на инкорпорирование/усвоение [3H] 3H-метилтимидина. Реакция на BDNF представляется двухфазной: очень малые дозы (0,1 нг/мл) дали слабое повышение усвоения тимидина; тогда как дозы выше 1 нг/мл BDNF задержали усвоение [3H] метилтимидина. В дозе 5 нг/мл BDNF дал ингибирование 24%, предполагая тем самым уменьшение степени пролиферации глиальных клеток во время периода обработки.
12.3. Обсуждение.
Эти эксперименты ин витро ясно показывают, что BDNF поддерживает выживание или вызывает полностью дифференцированное состояние допаминергических нейронов развивающейся крысиной субстанции нигра, как показано окрашиванием на тирозин-гидроксилазу, и потребление допамина в культурах этих нейронов, взятых из среднего мозга крысиного эмбриона. Поскольку именно эти нейроны дегенерируют при болезни Паркинсона, существует значительная вероятность того, что BDNF обладает терапевтическим потенциалом при лечении болезни Паркинсона либо посредством уменьшения потерь нейронов, либо путем повышения уровней содержания тирозингидроксилазы (энзим, ограничивающий степень синтеза допамина), либо, возможно, обоими путями.
Кроме того, как и фактор нервного роста NGF, BDNF обладает действием на выживание холинергических нейронов крысиного базального переднего мозга, как показано увеличением CAT-окрашивания, повышенной CAT-активностью и усиленным ацетилхолинэстеразным окрашиванием в культурах медиального перегородочного нуклеуса крысиного эмбриона и нуклеуса диагональной полосы Брока. Соответственно, сам BDNF или в сочетании с NGF может быть полезен при лечении заболеваний или расстройств, которые затрагивают холинергические нейроны базального переднего мозга, включая, например, болезнь Альцхаймера.
13. Пример. Идентифицирование нового гена в генном семействе BDNF/NGF.
Метод идентифицирования новых членов генного семейства NGF/BDNF с использованием PCR с дегенерированными олигонуклеотидами, основанными на сегментах консервации аминокислотной последовательности между NGF и BDNF (боксы 1-4, см. раздел 5.8), сначала испытывали путем определения того, могут ли обе пары таких праймеров быть использованы для амплифицирования как гена NGF, так и гена BDNF из геномной ДНК нескольких видов животных. Этот метод затем был использован для идентифицирования нового гена, который разделяет гомологию с NGF и BDNF во всех отмеченных четырех боксах гомологии.
13.1. Материалы и методы.
13.1.1. Цепная реакция полимеразы.
Реакцию PCR осуществляли в основном как описано в разделе 6, см. выше.
13.2. Результаты.
13.2.1. Амплификация последовательностей NGF и BDNF из геномной ДНК.
Дегенерированные синтетические
олигонуклеотидные праймеры были синтезированы согласно участкам бокс 1 и бокс 2 консервированной аминокислотной последовательности между NGF и BDNF
(см. раздел 5.8, выше) и использовались в
PCR-реакции с крысиной геномной ДНК в качестве матрицы. Точные праймерные последовательности были следующими (позиции дегенерации со смесью из двух или более
оснований, включенных на стадии
олигонуклеотидного синтеза, показаны в скобках; подчеркнуты хвосты с множественными сайтами расщепления рестрикционными эндонуклеазами, которые обеспечивают облегчение
лигирования к вектору на
последующем этапе клонирования; A = аденин, G = гуанидин; C = цитозин, T = тимин и N = смесь из A, C, G, T):
бокс 1 (смысловой), праймер IB:
5'-GACTCGAGTCGACTCGGTGTG(C,T)GACAG(C,
T)(A,G)T(C,T,A)AG-3'
бокс 2 (антисмысловой), праймер 2C:
5'-CCAAGCTTCTAGAATTCCA(C,T)TT(N)GT(C,T)TC(A,G)(A,T)A(A,G) AA(A,G),TA
300 нг
смеси каждого дегенерированного
праймера добавляли к 500 нг крысиной геномной ДНК в 100 микролитрах стандартной реакционной смеси для PCR. Было проведено 35 циклов, каждый состоящий из инкубирования в
течение 1 мин при 94oC, 2 мин при 43oC и 2 мин при 72oC. Ожидаемый размер продукта PCR амплификации BDNF- или NGF-гена с использованием этих праймеров составляет 175
пар оснований, включая
два 17-мерных "Хвоста" (подчеркнуто), включенных для удобства в последующие стадии клонирования. Электрофорез реакционной смеси на 8% полиакриламида (5% глицерина) дал основную
полосу
амплифицированной ДНК ожидавшегося размера, в 175 пар оснований.
13.2.2. Обнаружение последовательностей, комплементарных BDNF/NGF зонду в геномных ДНК различных видов животных.
Полосу в 175 бр удаляли из акриламидного геля электроэлюированием и амплифицировали далее во второй PCR реакции семь циклов в идентичных условиях реакции, как и в первой PCR за исключением того, что концентрации dGTP, dATP и TTP были снижены до 50 мкМ каждая, и вместо намеченного dCTP использовали метку альфа 32P-dCTP. Радиомеченый ДНК-продукт выделяли из реакционных компонентов хроматографией на размерной колонне. Этот зонд, обозначенныйnRIB/2C" (для крысиной ДНК, амплифицированной от праймеров 1B и 2C), использовали затем для обнаружения комплементарных последовательностей в геномных ДНК различных позвоночных (результаты, полученные с крысой, мышью и цыпленком, показаны на фиг.13), после переваривания с EcoRI рестрикционной эндонуклеазой и блоттинга к нитроцеллюлозе по методу саузерн-гибридизации на EcoRI-переваренных геномных ДНК, как показано на фиг.13.
Размеры EcoRI фрагментов геномной ДНК, содержащей последовательности NGF и BDNF, были определены в контролях на параллельных блотах с использованием радиомеченого человеческого NGF и BDNF хондов, приготовленных реакцией PCR из клонированных генов. Положение NGF и BDNF геномных EcoRI фрагментов указаны на фиг. 13 как N и B соответственно. Результаты похожего анализа представлены на фиг.3, и эквивалентные результаты были получены с использованием человеческого BDNF-зонда как со свиным BDNF-зондом на фиг.3. Как указано выше, NGF и BDNF виды каждый гибридизировали к единственным EcoRI фрагментам в различных геномных ДНК позвоночных. Например, в крысиной ДНК BDNF-проба обнаружила полосу примерно 8,8 кб, тогда как NGF-проба обнаружила полосу примерно 10,4 кб.
Как показано на фиг.13, у каждого испытанного вида (данные показаны для цыпленка, мыши и крысы) зонд RIB/2C гибридизировался к полосе ДНК, неотличимой от полосы ДНК, идентифицированной NGF зондом, так же как и к полосе ДНК, неотличимой от ДНК, идентифицированной BDNF-зондом (у мыши, NGF и BDNF-геномные EcoRI фрагменты имеют ту же самую электрофоретическую мобильность, соответствующую примерно 11,5 - 12,0 кб). Это демонстрирует, что дегенерированные олигонуклеотидные праймеры 1B и 2C могут быть использованы для амплифицирования последовательностей как от NGF, так и BDNF-генов. Следует заметить, что в некоторых случаях наблюдались дополнительные полосы в геномном саузерн-блоте, гибридизированном зондом RIB/2C. Например, в EcoRI-переваренной мышиной геномной ДНК наблюдались, по меньшей мере, две дополнительные полосы (меченные X1 и X2, размером приблизительно 19,0 кб и 1,5 кб соответственно), которые не отвечали ни NGF, ни BDNF. Подобным образом по меньшей мере две дополнительные полосы наблюдались в гибридизации к крысиной ДНК (X1, X2 примерно 7,3 и 1,2 кб соответственно) и по меньшей мере одну ДНК цыпленка (X, приблизительно 2,6 кб). В некоторых случаях наблюдались также дополнительные полосы, не помеченные на этом чертеже. Присутствие таких полос, не относящихся ни к NGF, ни к BDNF, предполагают вероятное существование дополнительного члена (или членов) генного семейства. Аналогично, при использовании других комплектов праймерных пар и геномных ДНК матриц (данные не показаны) обнаруживали полосы, ясно отличающиеся от известных полос, содержащих BDNF и NGF генные последовательности.
13.2.3. Идентифицирование нового гена, родственного BDNF и NGF.
Конкретное испытание гипотезы о том, что новый ген, связанный с NGF и BDNF,
мог бы быть идентифицирован посредством реакции PCR
с использованием дегенерированных олигонуклеотидных праймеров, проводилось с использованием праймеров для бокса 3 и бокса 4 (см. раздел 5.8, выше)
и мышиной геномной ДНК в качестве матрицы.
Дегенерированные праймеры были синтезированы следующих последовательностей:
бокс 3 (смысловой):
5'-GGGGATCCGCGGITG (T,C)(G,A) GIGGIAT(T,C,
A)GA3'
бокс 4 (антисмысловой):
5'-TCGAATTCTAGATIC (T,G) IAT(AG)AAIC(T,G)ICCA-3',
где G - гуанидин, A - аденин, C - цитозин, Т - тимин и I - инозин; смеси из более чем одного
основания в единственной позиции в
нуклеотиде показаны в скобках. Следует отметить, что инозин использован в некоторых позициях, соответствующих третьему ("шаткому") основанию кодона, чтобы позволить
дегенерацию генетического кода.
Также возможно использовать смесь из четырех конвенциональных (обычных) оснований ДНК вместо инозина, и с такими праймерами были получены, в сущности, идентичные
результаты.
С дегенерированной бокс 3/бокс 4 праймерной парой, PCR от геномных NGF- и BDNF-последовательностей в мышиной ДНК, как ожидалось, должна была амплифицировать сегмент размером примерно в 90 бр. Используя праймеры, показанные выше, PCR-реакцию проводили в четыре цикла с температурой ренатурации 45oC, а затем еще в 31 цикл с температурой ренатурации 49oC. Продукты анализировали гелевым электрофорезом, и наблюдали основную полосу ожидавшегося размера. У мыши ген NGF содержит сайт расщепления для рестрикционной эндонуклеазы Hind II в области между боксом 3 и боксом 4, тогда как ген BDNF содержит сайт расщепления для энзима ApaI в этой области. Поэтому ожидалось, что переваривание продукта PCR - амплификации с Hind II и ApaI устранит последовательности NGF и BDNF из основной полосы продукта. Однако, когда PCR-продукт был переварен полностью с этими двумя ферментами рестрикции осталась в амплифицированной ДНК. Это позволяет предположить, что в дополнение к генам NGF и BDNF был амплифицирован по меньшей мере один новый ген.
13.2.4. Характеристика нового члена генного семейства.
Резистивный к перевариванию продукт PCR-реакции элюировали из геля и использовали в качестве матрицы в ассиметричных PCR-реакциях, в которых один из исходных дегенерированных праймеров присуствовал в 10-100-кратном молярном избытке по сравнению с другим праймером. Это ассиметричное амплифицирование позволило произвести однонитевые ДНК-матрицы, подходящие для секвенирования методом цепного терминирования. Анализ последовательности нового гена (обозначенного здесь N 3/4, но также и упоминаемого как "нейротропин-3") был расширен далее путем PCR амплификации последовательности между точным праймером, расположенным между боксами 3 и 4, и поли-A последовательностью у 3'-конца транскрипта гена, используя стратегию "быстрой амплификации концов кДНК" (RACE), описанной M.A.Trohman, M.K.Dush т G.R.Martin, Proc. Natt. Acad. Sci. США, 85: 8998 - 9002 (1988). Результаты секвенирования ДНК обнаружили, что был амплифицирован новый ген, содержащий открытую рамку считывания, способную кодировать полипептид, отличающийся от NGF и BDNF, но близко родственный по своей аминокислотной последовательности к обоим (фиг. 14).
Предварительное подтверждение того, что этот новый ген кодирует нейротропный фактор, было получено путем определения его паттерна экспрессии в крысиных тканях путем назерн-блотгибридизации. Этот анализ показал, что этот новый ген экспрессируется гораздо более сильно в мозговой ткани, чем в любой другой исследованной ткани.
13.3. Обсуждение.
Как показано выше (см. фиг.13), ДНК-зонд, полученный PCR-амплификацией с использованием праймеров для боксов 1 и 2, с крысиной геномной ДНК в качестве матрицы (RIB/2C) гибридизировался к новым полосам в дополнение к полосам, которые, как было известно, содержали NGF- и BDNF-генные последовательности в EcoRI-переваренной ДНК каждого исследованного вида животного. Подобный анализ был проведен с использованием радиоактивно меченного зонда для нового гена, амплифицированного с использованием праймеров бокса 3 и 4 на мышиной геномной ДНК (М3/4). В каждом случае зонд N 3/4 гибридизировался к единственной основной полосе EcoRI-переваренной геномной ДНК в отличие от полос, содержащих последовательности BDNF и NGF. Следует заметить, что EcoRI фрагменты геномной ДНК мыши, крысы и цыпленка, у которых наблюдалась гибридизация с зондом M3/4, во всех случаях совпадают с одной из новых полос, полученных гибридизацией с зондом RIB/2C. Этими фрагментами являются: 19,0 кб EcoRI-фрагмент в крысиной ДНК (XI на фиг. 12), 7,3 кб фрагмент в крысиной ДНК (XI на фиг. 12) и 2,6 кб полоса в ДНК цыпленка (X на фиг. 12). Это предполагает, что части одного и того же гена были амплифицированы из крысиной ДНК с использованием праймерной пары 1B/2C и из мышиной ДНК с праймерами 3 и 4. Таким образом, по меньшей мере один новый ген разделяет гомологию с генами NGF и BDNF во всех четырех боксах гомологии, отмеченных выше.
Идея гомологичных "боксов" между NGF, BDNF и дополнительными членами этого генного семейства (например, М3/4, известный также как NT-3) была выражена здесь на первичной аминокислотной последовательности, и методами идентифицирования новых генов семейства. Однако важно иметь в виду, что, вероятно, имеет место дополнительное вовлечение вторичной и третичной структуры этих нейротропных факторов, их взаимодействие с специфичными рецепторами для рациональной конструкции новых молекул с потенциальной терапевтической ценностью.
Например, у NGF имеется 6 цистеиновых остатков, и все они, как было показано, вовлечены в образование дисульфидных мостиков. Пронумеровав их от N-терминала к C-терминалу, как цис 1 - цис 6, имеем дисульфидные мостики цис 1-цис-4, цис 5 и цис 3 - цис 6. Позиции всех шести цистеиновых остатков консервированы между NGF и BDNF, и позиции трех цистеиновых остатков в участке "M3/4" выстраиваются точно с цис 4, цис 5 и цис 6 NGF и BDNF. Это предполагает наличие тесной связи вторичной структуры всех членов генного семейства, которая в основном определяется консервированными цис-остатками. Следует заметить, что боксы гомологии, указанные для BDNF и NDF, покрывают 5 из шести цис-остатков (цис 1 в боксе 1, цис 2 в боксе 2, цис 3 в боксе 3, цис 5 и цис 6 - в боксе 4). Это поддерживает ту идею, что эти цис-остатки и их непосредственные соседи играют важную роль в определении общей структуры нейротропных факторов. Эти структурные детерминанты для специфического взаимодействия с рецепторами высокой аффинности для каждого из нейротропных факторов предположительно располагаются в уникальных частях каждой молекулы.
Соответственно новые химерные гены могут быть получены посредством рекомбинации между членами семейства (например, посредством ин витро рекомбинации или посредством прямого генного синтеза) в любом из уже описанных четырех боксов гомологии или где-нибудь еще в молекуле. Похоже, что такие химерные белки имеют сходную вторичную структуру вследствие консервирования цис-остатков и других аминокислотных остатков, но может обладать новыми биологическими свойствами. Например, BDNF/NGF химерный протеин может быть бифункциональным с точки зрения взаимодействия с рецепторами BDNF и NGF. Химерные протеины могут также отличаться от родительской молекулы (или молекул) в отношении образования двухмерной структуры и другими физико-химическими свойствами. Химерные протеины могут также функционировать как антагонисты любой родительской молекулы.
Активные фрагменты BDNF/NGF могут быть использованы для создания других членов семейства на основе знания критичных "сердцевинных" областей для соответствующего складывания плюс информации, касающейся того, какие области требуются для типоспецифичного взаимодействия с рецепторами.
Сравнение новых
членов семейств с уже известными может быть использовано для обнаружения новых боксов гомологии, которые могут помочь в
проведении поиска дополнительных членов генного семейства BDNF/NGF. Например,
сравнение "M3/4" с BDNF обнаруживает некоторые полезные боксы гомологии. Один из них, представляющий особый интерес,
включает четвертый консервированный цис-остаток, единственный отсутствующий в
описанных выше боксах 1-4. Это действительно довольно длинный сегмент идентичности или консервированного аминокислотного
замещения, разделяемого BDNF и "M3/4", который включает этот цис-остаток, а
именно:
His-Trp-Asn-Ser-Gln-Cys-/Arg или Lys/-Thr/Thr или Ser/-Gln-/Ser или Thr/-Tyr-Val-Arg -Ala-Leu-Thr
Внутри этой области можно выбрать по меньшей мере два бокса гомологии для
синтезирования полезных дегенерированных олигонуклеотидных праймеров (например, His-Trp-Asn-Ser-Gln-Cys требует 96-кратную
дегенерацию для 18-мерного параймера, или 48-кратную для 17-мерного),
полезным боксом был бы также и Tyr-Val-Arg-Ala-Leu-Thr.
14. Пример. Повышенная экспрессия BDNF в клетках нейробластомы.
14.1. Материалы и методы.
14.1.1. Клеточные линии.
CHP100, CHP126, CHP134, CHP234, LANI, LAN5, NB9, SV5V, V79, FO1, Bu2, HOI, HL60 и COL320 - клеточные линии, поддерживаемые в лаборатории доктора Фреда Альта (Dr. Fred Alt), который представил РНК для назернблоттинга на фиг. 15. Все клеточные линии - из опухолей человека. CHP100-линия клеток нейроэпителиомы, CHP126, CHP134, CHP234, LANI, LAN5, NB9 и SV5V - линии клеток нейробластомы, V79 - линия клеток ретинобластомы, FOI, BU2, HOI - линии клеток меланомы, HL60 - линия клеток промиелоцитной лейкемии, COL320 - линия клеток нейроэндокринной карциномы толстой кишки.
14.1.2. Приготовление РНК.
Приготовление РНК и осуществление назерн-блоттинга было, в сущности, таким, как описано в разделе 8.1.1, см. выше, с использованием зонда человеческой кДНК полной длины. 10 мкг общей РНК в каждой дорожке геля было использовано для производства назерн-блоттинга (фиг. 15) за исключением того, что РНК в LANI была менее загружена, а для SV5V использовали один микрограмм поли (A)+РНК.
14.2. Результаты.
На фиг. 15 изображены результаты назерн-блот-анализа зонда BDNF, гибридизированного к панели образцов РНК, полученной от многих человеческих клеточных линий. Значительное количество РНК, гибридизированной к BDNF-зонду, было обнаружено в клеточных линиях CHP234 и LAN5, и меньшие количества были обнаружены у CHP126 и CHP134. Все положительные линии происходили из нейробластомных опухолей человека.
15. Пример. Зависимое от активности регулирование BDNF и NGF-РНК в крысином аммоновом роге осуществляется при участии не-NMDA глютаматных рецепторов.
15.1. Материалы и методы.
15.1.1. Обработка крыс каиновой кислотой.
Крысы обычно получали диацепам (valium) через 90 минут после внутрибрюшной инъекции каиновой кислоты в количестве 12 мг/кг живой массы для подавления двигательной активности. Как показано на фиг. 19, valium, данный после каиновой кислоты, не влиял на дальнейшее повышение уровней BDNF и NGF-мРНК. Крысы, не получавшие valium, показали сходные увеличения уровней BDNF- и NGF-мРКН в аммоновом роге через 3 часа после каиновой кислоты в сравнении с животными, получившими каиновую кислоту, а затем valium.
15.1.2. Приготовление культур клеток аммонова мозга.
Аммонов мозг брали у крысиных эмбрионов E17, извлекали диссекцией и инкубировали в течение 20 мин при 37oC в фосфатном буферном физиологическом растворе (PBS) без ионов кальция и магния, но содержащем 10 мМ глюкозы, 1 мг/мл альбумина, 6 мкг/мл ДНКазы и 1 мг/мл папаина. После промывки раствором без папаина клетки аммонова рога осторожно диссоциировали отожженной пастеровской пипеткой. Клетки собирали центрифугированием на малой скорости, ресуспендировали в ДМЕМ, добавляли 10% сыворотки плода теленка и засевали на пластиковые культуральные чашки с плотностью 0,5 • 106 клеток на 35 мм, предварительно покрывавшиеся поли-ДЛ-орнитином (0,5 мг/мл) и ламинином (5 г/мл). Через три часа после посева, среду заменяли на свободную от серы среду, которая содержала добавки, как описано Bruwer и Cotman (Brain Res. 497: 65, 1989), но без глютамата. Нейроны оставались жизнеспособными в течение трех недель в культуре и обычно использовались для экспериментов на седьмой день после посева.
15.1.3. Амплифицирование РНК.
Общую клеточную РНК экстрагировали как описано Chomcynsky и Sacci (Anal. Biochem 1987, 162: 156-159) из 0,5 • 106 клеток после добавления укороченного РНК восстановительного стандарта (30 fg). NGF мРНК и кРНК были совместно амплифицированы в соединенной в одной пробирке реакции обратной транскрипции/цепной реакции полимеразы (RT/PCR), содержащей 1/5 экстрагированной РНК, 1 х RT/PCR буфер (10 мМ Трис-HCl pH 8,3, 50 мМ KCl, 1,5 мМ MgCl2, 0,1 мг/мл желатина, 0,1% тритона X-100), 0,25 мМ dNTP, 0,1 мкМ каждого 5' и 3' праймеров, 5 ед. РНКазы (Promega), 3,2 ед АМУ-обратной транскриптазы (Life Science) и 2 ед Taq полимеразы (Genofit) в общем объеме 25 мкл. Эту смесь покрыли минеральным маслом, инкубировали в течение 30 минут при 41oC, нагревали до 92oC 60 с, ренатурирование праймера - при 55oC 60 с, и праймерное расширение - при 72oC 60 с). Продукты амплифицирования (203 бр для NGF мРНК и 153 бр для восстановительного стандарта) разделяли на 3% Nusieve/Agarose 3: 1 геле (FMC Bioproducts), делали щелочной блоттинг к мембране Hybond N plus (Ameriham) и гибридизировали как описано (Heumann и Thoenen 1986, J.Biol.Chem. 261:9246; Lindbold и др., 1988, J.Biol.Chem. 263: 16348). Для абсолютного количественного подсчета, известные количества транскрибированной ин витро NGF мРНК и восстановительный стандарт совместно амплифицировали в параллельных реакциях, недавно подобный метод был описан Wang и др. (PNAS, 86: 9717-9721, 1989).
15.2. Результаты и обсуждение.
BDNF и NGF являются членами генного семейства примерно с 50%-ной аминокислотной идентичностью (Leibrock и др., 1989, Nature, 341: 149). Эти молекулы проявляют строго консервированные домены. Содержащиеся внутри этих доменов шесть цистеиновых остатков наиболее вероятно вовлечены в стабилизацию трехмерной структуры этих молекул, которая необходима для их биологической активности. Однако у NGF и BDNF имеются также отличающиеся домены, которые определяют их разную нейронную специфичность (Линдсей и др., 1985, Dev. Biol. 112: 319; Джонсон и др., 1986, J. Neurosci, 6: 3031; Hofer и Barde, 1988, Nature 331: 261; Rodriguez-Tebor и др., 1989, Dev. Biol. 136 : 296). Кроме того, различия между этими двумя нейротропными молекулами также указаны местами их синтезирования, NGF экспрессируется как на периферии (Korsching, S. и Thoenen, H., 1983, Proc. Nath. Acad. Sci. США, 80: 3513; Ebendal и др., 1983, Exp. Cell Res. 148: 311; Heumann и др., 1984, EMBO, 3: 3183; Dhelton и Reichardt, 1984, Proc. Nath. Acad. Sci. США, 81: 7951) так и в центральной нервной системе (Korsching и др., 1985, EMBO, 4: 1389; Shelton и Reichardt, 1986, Proc. Nath. Acad. Sci. США, 83: 2714; Whittemore, 1986, Proc. Nath. Acad. Sci. США, 83: 817; Large и др., 1986, Science, 234: 352).
Плотность иннервации нейронами, реагирующими на NGF, отражает уровни содержания NGF в соответствующих мишенных тканях (Korsching и Thoenen, 1983, Proc. Nath. Acad. Sci. США, 80: 3513; Ebendal, 1983, Exp. Cell Res. 148: 311; Heumann и др., 1984, EMBO, 3: 3183; Dhelton и Reichardt, 1984, Proc. Nath. Acad. Sci, США, 81: 7951; Korsching и др., 1985, EMBO, 4: 1389; Shelton и Reichardt, 1986, Proc. Nath. Acad. Sci, США, 83: 2714; Whittemore и др., 1986, Proc. Nath. Acad. Sci. США, 83: 817; Larg и др., 1986, Science, 234: 352). В отличие от NGF BDNF в основном экспрессируется в ЦНС, в нейронах, и уровни содержания BDNF мРНК значительно выше, чем уровни NGF мРНК, например примерно в 50 раз в аммоновом роге. В периферийной нервной системе NGF синтезируется различными типами не-нервных клеток (Bandtlow и др., 1987, EMBO, 6: 891), тогда как в головном мозге NGF в основном локализуется в нейронах, как продемонстрировано посредством ин-ситу гибридизацией (Rennert и Heinrich 1986, Biochem. Biophys. Res. Commun 138: 813; Ayer-LeLivre и др., 1988, Science 240 : 1339; Whittmore и др. 1988, J. Neurosci. Res. 20 : 403). Однако культурированные астроциты типа I так же, как было показано, продуцировали существенные количества НСГ (Линдсей, 1979, Watuu 282: 80; Фурукава и др., 1987, Biochem. Biophys. Res. Com. 142: 395). Относительный вклад нейронов и астроцитов в NGF, синтезируемых в головном мозге, пока еще неизвестен.
С учетом того что основная экспрессия BDNF и NGF-мРНК происходит в центральной нервной системе, были исследованы следующие вопросы - может ли нейронная активность повлиять на уровни содержания этих двух РНК, и если это так, то какой трансмиттер(ы) мог бы участвовать в регуляции. В первой серии экспериментов, были приготовлены нейронные культуры из аммонова рога крысиных эмбрионов E17. Как показано на фиг. 16A, деполяризация нейронов аммонова мозга высоким содержанием (50 мМ) калия приводит к повышению BDNF-мРНК. Максимальные уровни были достигнуты между 3 и 6 часами после повышения концентрации калия. Сообщаемое калием повышение BDNF-мРНК могло быть заторможено путем удаления ионов кальция из среды и также снижалось путем ингибирования поступления кальция посредством нифедипина, блокирующего путь поступления кальция (см. фиг. 16B).
Вследствие того что используемые культуры аммонова рога состояли из смешанной популяции нейронов, проявляющих различные паттерны трансмиттерной и рецепторной экспрессии, здесь излучали действия различных физиологических и синтетических рецепторных агонистов на BDNF и NGF-экспрессию. Результаты, представленные в табл. IV, показывают, что из всех испытанных субстанций каиновая кислота - агонист глютаматного рецептора (Monaghan и др., 1989, Annual. Rev. Pharmacol. Toxicol. 29: 365) - дала до сих пор наибольшее повышение BDNF-мРНК в нейронах аммонова рога.
Таблица IV
Действие различных рецепторных агонистов на
экспрессию BDNF-мРНК в культурируемых нейронах.
Добавление - % от контроля
Ничего - 100 ± 5
Карбахол (50 мкМ) - 220 ± 25
Карбахол (50 мкМ) +
атропин (10 мкМ) - 85 ± 15
Никотин (100 мкМ)
- 110 ± 7
Гистамин (50 мкМ) - 150 ± 12
Серотонин (100 мкМ) - 95 + 6
Допамин (100 мкМ) - 115
± 7
Норепинефрин (25 мкМ) - 75 ± 10
Субстанция "П" (1 мкМ) - 85 ± 9
Соматостатин (1 мкМ) - 110 ± 8
Брадикинин (1 мкМ) - 155 ±
15
Каиновая кислота (25 мкМ) - 1365 ± 70
NMDA (25 мкМ) - 105 ± 10
Напротив, другие молекулы, такие как карбахол (мускарина рецепторный агонист) и в меньшей
степени гистамин и брадикинин, BDNF-мРНК повышали слегка, хотя и
заметно (табл. IV). Вследствие того что уровни NGF-мРНК в нейронах аммонова рога были очень низкими, здесь был создан количественный
метод цепной реакции полимеразы, пригодный для определения
изменений в NGF-мРНК. С использованием этого метода установлено, что уровни BDNF-мРНК, как уровни NGF-мРНК, также повышались калием и
каиновой кислотой в нейронах аммонова рога (фиг. 17).
Как показано на фиг. 18, максимальное повышение BDNF-мРНК в нейронах аммонова рога было получено с каиновой кислотой в концентрации примерно 25 мкм. Дальнейшее повышение ее концентрации приводило к снижению уровней BDNF-мРНК, что скорее всего отражает токсическое действие высоких концентраций каиновой кислоты на нейроны аммонова рога (фиг. 18). Ранее сообщалось о передаваемой глютаматным рецептором нейротоксичности (Choi и др., 1987, J. Neurosci, 7: 357; Rothman и Olney, 1987, Trends Neurosci, 10: 299; Choi, 1988, Neuron, 1: 623) на различных ЦНС-нейронах после применения аналогов возбуждающего аминокислотного глютамата. Однако концентрации каиновой кислоты, необходимые для усиления экспрессии BDNF- и NGF-мРНК в нейронах аммонова рога, могут быть четко отделены от концентраций, приводящих к нейротоксичности.
Для того чтобы исследовать действие каиновой кислоты более детально, здесь изучали, может ли быть блокировано повышение уровней BDNF-мРНК каким-либо из известных глютамат-рецепторных антагонистов (Monaghan и др., 1989, Annual Rev. Pharmacol. Toxicol. 29: 265). Кинуреновая кислота, глютамат-рецепторный антагонист широкого спектра, так же как и CNAX конкурентный ингибитор не-NDA рецепторов (Minaghan и др., 1989, Annual Rev. Pharmacol. Toxicol. 29: 365), полностью блокировала сообщенное каиновой кислотой повышение BDNF-мРНК в нейронах аммонова рога (табл. V).
Таблица V
Действие антагонистов к различным глютаматным рецепторам на
вызванную каиновой кислотой экспрессию BDNF-мРНК.
Добавление - % от контроля
ничего - 100 ± 60
каиновая кислота (25 мкМ) - 1250 ± 60
кинуреновая кислота (1 мкМ) - 70 ± 6
кинуреновая кислота (1
мкМ) + каиновая кислота (25 мкМ) - 84 ± 7
CNQX (10 мкМ) - 109 ± 6
CNQX (10 мкМ) + каиновая
кислота (25 мкМ) - 105 ± 7
MK-801 (5 мкМ) - 95 ± 5
MK-801 (5 мкМ) + каиновая кислота (25 мкМ) - 1130 ± 80
С другой стороны, MK 801, который специфически
блокирует NMDA рецепторы, был неэффективен при блокировании повышения
BDNF-мРНК в этих нейронах. Кроме того, сам NMDA не изменял уровней BDNF-мРНК (табл. IV). Это показывает, что каиновая кислота
действует непосредственно через свои рецепторы и что это действие
происходит не из-за освобождения эндогенного глютамата (который также действует на NMDA рецепторы). Таким образом, можно заключить,
что каиновая кислота ин витро повышает уровни BDNF -мРНК
посредством не-NMDA глютаматных рецепторов.
Чтобы оценить физиологическое значение этих ин витро наблюдений, изучали, действуют ли подобные механизмы также ин виво. Обрабатывали взрослых крыс Wistar обоего пола, весом 180-200 г, 12 мг/кг каиновой кислоты. После различных периодов времени определяли изменения в уровнях BDNF- и NGF-мРНК в аммоновом роге и кортексе посредством назерн-блот-анализа. Фиг. 19 показывает, что каиновая кислота вызывает повышение уровней BDNF- и NGF-мРНК в обоих областях головного мозга. Повышение BDNF-мРНК было существенно выше, чем повышение NGF-мРНК. Кроме того, еще после 24 часов за введением каиновой кислоты, уровни мРНК оставались повышенными. Изменения BDNF- и NGF-мРНК по времени показали, что максимальное повышение в аммоновом роге достигалось примерно через 3 часа после введения каиновой кислоты (фиг. 19). Интересно отметить, что перед повышением в аммоновом роге уровней BDNF- и NGF-мРНК происходило повышение C-fos-мРНК: эти два явления могут быть связаны случайно, как недавно было показано в случае седалищного нерва после его повреждения (Hengerer и др. , 1990, Proc. Nath. Acad. Sci США, 87:3899). Кроме того, имеется задержка повышения как BDNF-, так и NGF-мРНК в церебральном кортексе по сравнению с повышением в аммоновом роге, указывающая на то, что сигналы, приводящие к усилению экспрессии BDNF и NGF распространяются от аммонова рога в другие области мозга. Эти наблюдения согласуются с предыдущими сообщениями. (Morgan и др., 1987, Sciece 237: 192), показывая, что увеличение c-fos, вызываемое конвульсантным метразолем, происходит вначале в аммоновом роге, а затем в кортексе.
Предшествующие исследования обеспечили доказательство того, что NMDA-рецепторы вовлечены в регуляцию мРНК аммонова рога, таких как NGFI-A (Cole и др. , 1989, Нат ге, 340: 474), поэтому здесь изучалось, могли ли или нет NMDA-рецепторные антагонисты MK 801 и кетамин блокировать повышение BDNF и NGF-мРНК ин виво.
Однако, подтверждая результаты, полученные ин витро, МК 801 не ингибировал вызванное каиновой кислотой повышение BDNF-мРНК в аммоновом мозге, хотя он эффективно подавлял судороги, происходящие от активации NMDA-рецептора (фиг. 20). Скорее всего, эта активация NMDA-рецептора происходит от эндогенного глютамата, освобождаемого каиновой кислотой (Бизьер и Койл, Нейросайенс, 1988, Lett, 8: 303; и др., 1978, Brain. Res. 139: 381). Аналогично, повышение NGF-мРНК в аммоновом роге после его обработки каиновой кислотой не предотвращалось ни МК 80К, ни кетамином.
В предыдущем исследовании (Gall и Isackson 1989, Science 245:758), было показано, что электролитические повреждения, вызывающие лимбические судороги, повышают NGF-мРНК в крысином аммоновом роге. Однако настоящие результаты, полученные с МК 801 и каиновой кислотой, показали четкое размежевание между вызыванием судорог и повышениями BDNF- и NGF-экспрессии. Обработка крыс диацепамом (valium) перед инъекциями каиновой кислоты полностью блокировала повышение BDNF- и NGF-мРНК в крысином аммоновом роге (фиг. 20). Блокирующий эффект диацепама на повышение BDNF- и NGF-мРНК скорее всего происходит от подавления нейронной активности посредством ингибиторной GABA-эргической системы. Однако через 90 минут после введения каиновой кислоты ( то есть, приблизительно, через 30 минут после начала конвульсий), повышение BDNF- и NGF-мРНК не могло быть больше блокировано диацепамом, указывая, что относительно короткого периода увеличенной активности может хватить для инициирования каскада событий, ведущих к усилению экспрессии мРНК двух нейротропных факторов.
Настоящее исследование показало, что не-NMDA рецепторы могут быть вовлечены в регуляцию BDNF-NGF-мРНК в аммоновом роге, отличая таким образом этот регуляторный механизм от ранее описанного Коулем и др. (Cole, 1989, Nature, 304: 474), который считает, что мРНК аммонова рога, кодирующие ранние гены, по-видимому, регулируются NMDA-субтипами лютаматных рецепторов. Сообщалось, что некоторые из действий каиновой кислоты на нейроны могут также передаваться посредством глютаматных рецепторов quisqualate-типа (Monaghan и др., 1989, Annual. Rev. Pharmacol. Toxicol. 29: 365).
В заключение, настоящее исследование показало, что нейронная активность регулирует уровни мРНК, кодирующих нейротропные факторы BDNF и NGF в крысином аммоновом роге и кортексе. Из всех испытанных веществ каиновая кислота, действуя посредством не-NMDA глютаматных рецепторов, вызывала повышение BDNF- и NGF-мРНК как ин виво, так и в культурах нейронов аммонова рога. Напротив, на периферии не было свидетельств тому, что NGF-синтез в не-нейронных клетках регулируется посредством обычных трансмиттерных субстанций или нейтропептидами, освобождаемыми иннервирующими нейронами (реагирующими на NGF) (Barde и др. , 1984, J.Cell. Biol, 99:839; Helleveg и др., Exp. Cell.Res. 179: 18), BDNF в основном экспрессируется в центральной нервной системе и имеется по меньшей мере частичное перекрывание между центральными путями с использованием глютамата в качестве нейротрансмиттера и областями, которые, как показано, богаты BDNF- и NGF-мPHK (Hofer и др., EMBO - в печати; Monaghan и др., 1989, Annual. Rev. Pharmacol. Toxicol. 29: 365). В частности, имеется удивительная схожесть между распределением NGF-мРНК, как показано гибридизированием "ин ситу" (Hofer, EMBO, в печати) и каиновокислотных рецепторов, проявленных посредством авторадиографии в аммоновом роге, церебральном кортексе и мозжечке (Monaghan и др., 1989, Annual. Rev. Pharm. Toxicol 29: 365). Эти наблюдения совместимы с интерпретацией того, что глютамат представляет собой физиологический трансмиттер, регулирующий BDNF- и NGF-мРНК в ЦНС.
16. Пример. Мозговой нейротропный фактор повышает выживание и дифференцирование функций у крысиных перегородочных холинергических нейронов в культуре.
16.1. Материалы и методы.
16.1.1. Приготовление диссоциированных клеток и условия клеточного культурирования.
Перегородочную область у крыс (Sprague-Dawley) после 17 дней беременности (Е17 - 17-й эмбриональный день) освобождали диссекцией от окружающей ткани. Фрагменты ткани собирали, трижды промывали Нам F-10 и затем переносили на 35 мм чашку для культурирования ткани и измельчали. Суспензию из отдельных клеток делали инкубированием ткани с 0,25% трипсина в течение 20 минут при 37oC. После инактивирования трипсина в течение 5 минут инкубировали при комнатной температуре в ростовой среде (см. ниже), содержащей 50 ug/мл ДНКазы типа 1 ("СИГМА"), и клетки диссоциировали повторяющимся прохождением фрагментов через суженный кончик пастеровской пипетки. Диссоциированные клетки затем центрифугировали при 500g в течение 45 секунд, супернатант удаляли и рецентрифугировали. Свободный клеточный дебрис ресуспендировали и соединяли в ростовой среде, а клеточный выход определяли с использованием гемоцитометра. Наконец, эти клетки засевали в 6-миллиметровые лунки, покрытые полиорнитином (10 мкг/мл) и ламинином (10 мкг/мл). Проверяли клеточную жизнеспособность после 24 часов в культуре посредством способности клеток не пропускать в себя трипан синий (trypan blue).
Нормальная ростовая среда 5HS/N3 для культур, составленных из нейронов и глин, содержала: 5% (об./об.) лошадиной сыворотки ("Жибко"), 1% добавок N 3 (об. /об. ) (Romijn и др., 1982, Dev. Brain Res. 2: 583-589), 0,5% (об/об) глютамина (200 мМ, Жибко) и 0,25% (об./об.) пенициллин-стрептомицина (10000 ед/мл и 10000 мкг/мл соответственно) (Жибко) в модифицированной Дульбекко среде Игл (ДМЕМ). Обогащенные нейронами культуры были приготовлены путем замены ростовой среды через: 5 - 6 часов после посева, на ДМЕМ, содержащую: 1% N3 добавок, 0,5% глютамина и 0,25% пенициллина и стрептомицина. В обоих условиях использовали обработку цитозин-арабинозой (1 мкМ на 24 часа) для ограничения пролиферации глиальных клеток.
16.1.2 Исследование холин-ацетилтрансферазной активности (САТ).
Ростовую среду удаляли из культуры промыванием клеток дважды 100 мкл PBS. Клетки лизировали одним циклом замораживание-оттаивание и 15 минутным инкубированием при 37oC в 50 мМ КH2РО4 pH 6,7, содержащем 200 мМ NaCl и 0,25% (об. /об. ) Тритон Х-100. Удаляли два микролитра клеточного дизата и испытывали на CAT-активность согласно процедуре микро-фоннум (micro-Fonnum) (Fonnum, 1975, J.Neurochem, 24: 407-409). Конечный состав вещества был: 0,2 мМ [14C] ацетил-CoA (NEN, 54,4 мCi/ммоль), 300 мМ NaCl, 8 мМ холин-бромида, 20 мМ этилен-диамин-тетрауксусной кислоты и 0,1 мМ неостигмина в 50 мМ NaH2PO4 (с pH 7,4) буфере. При этих концентрациях фермента и субстрата, ферментативная реакция была линейной в течение 90 - 120 минут. Специфичность индуцирования холин-ацетилтрансферазы испытывалась путем добавления специфичного ингибитора CAT-активности, N-гидроксиэтил-4-/1-нафтилвинил/пиридиума (HNP) во время эксперимента (White H.L. и Cavallito, C.I., 1970, J. Neurochem, 17: 1579-1589).
16.1.3. Исследование ацетилхолиноэстеразной активности (AChE).
Уровень присутствующей в лизатах AChE измеряли по методам Поттера (Potter, 1967, J.Pharmacology and Experimental Therapeutics 156: 500-506) и Джонсона и Рассела (Johnson и Rassel, 1975, Anal. Biochem. 64: 229-238) с несколькими модификациями.
Лизаты приготавливали в 50 мМ KH2PO4 буфере с pH 6,7, содержащем 200 мМ NaCl и 0,25 (об./об.) Тритона X-100, соединяли их [3H] ацетилхолин-иодидом (NEN, NEN-113, 73,3 мCi/ммоль), этопропазином (0,1 мМ) в 50 мМ с pH 7,0. После 15 мин инкубации при 37oC реакцию прекращали добавлением 10 мкл стоп-буфера, содержащего: 1 М хлоруксусной кислоты, 0,5 М NaOH и 2,0 М NaCl. Специфическую AChE активность определяли в присутствии 1,0 (10-6) М неостигмина.
16.1.4. Измерение высокоаффинного усвоения холина.
Усвоение холина посредством высокой аффинности Na+-зависимого механизма исследовали в основном по методу Вака и Пилар (Vaca и Pilar 1979 J.Gen. Physiol 73: 605-628). Клетки промывали один раз 100 мкл F-10(Haм) с последующим ополаскиванием 100 мкл раствором Tyrodes. Через 10 минут после деполяризации, вызванной раствором Tyrodes, содержащим 25 мМ калия, клетки инкубировали в течение 20 минут при комнатной температуре с [3H]холином (NEN, NEN-109, 0,11 мкМ, 86,7 Ci/ммоль) в нормальном Na+ или содержащем ионы Li+ растворе Tyrodes. Реакция усвоения прекращалась двойным промыванием клеток PBS. Аккумулированный холин экстрагировали путем инкубирования культур по меньшей мере в течение 20 минут с 100 мкл холодного ацетона, содержащего 1Н муравьиной кислоты. Растворимую фракцию затем удаляли и подсчитывали. Специфическим усвоением холина является разница между общим и Na+ зависимым усвоением холина.
16.1.5. Гистохимическое окрашивание на ацетилхолинэстеразу.
Холинергические клетки идентифицировали путем их гистохимического окрашивания на ацетилхолинэстеразу посредством модифицированного метода Geneser-Jensen и Blackstadt (1971, Z. Zellforsch. 114: 460-481). После фиксирования культур в 4%-ном параформальдегиде эти клетки инкубировали 5-6 дней при 4oC в присутствии AChE субстратного раствора, имеющего следующий состав: 4 мМ ацетил-литий-холин иода, 2 мМ сульфата меди, 10 мМ глицина и 10 мкг/мл желатина в 50 мМ ацетатном буфере с pH 5,0. Проявление (визуализацию) продукта реакции осуществляли, как было описано ранее (Hartikka, Hefti, 1988, J. Neurosci. 8: 2967-2985).
16.1.6. Иммуногистохимическое окрашивание на NGF-рецептор.
Культуры промывали дважды ДМЕМ и затем фиксировали 4%-ным параформальдегидом. Клетки затем обрабатывали 3-4 часа натриевофосфатным буфером с pH 7,4, содержащим 5% бычьего сывороточного альбумина, 0,02% тритона х-100 и 5% сахарозы (Hartikka, 1988, J.Neurosci 8: 2967-2985) NGF-R обнаруживали с использованием моноклонального антитела 192-IgG (Taniuchi и Johnson 1985, J Cell Biol 101: 1100-1106; Chandler и др., 1984, J. Biol. Chem. 259:6882-6889) и при разведении 1:1000 в буфере, содержащем 5% лошадиной сыворотки. Культуры инкубировали с разведенным антителом в течение 15-18 часов при 4oC. Связанный мышиный иммуноглобулин обнаруживали с использованием биотинированного лошадиного антимышиного IgG (1:200 Vector). Иммунореактивные клетки идентифицировали с использованием 3',3'-диаминобензидина в качестве субстрата для связанной пероксидазы.
16.1.7. Очистка BDNF и NGF.
Очистку BDNF из головного мозга свиньи и NGF из подчелюстной железы самца мыши проводили согласно ранее опубликованным процедурам (Barde и др., 1982, EMBO, 1:549-553; Lindsay и др., 1985, Dev. Biol. 112: 319-328). Рекомбинантный BDNF, использованный для части этих исследований, был охарактеризован ранее (Maisonpierre и др., 1990, Scince 247: 1446-1451).
16.2. Результаты.
Культуры перегородочных клеток дают отрастание нейритов на покрытых полиорнитин-ламинином лунках, при этом многие из клеток проявляют характерную фазояркую сому в 24 часа (фиг. 21A и B). Продолжавшееся отрастание нейритов привело к прогрессивному повышению распространения клеточного контакта на 2-й и 4-й дни ин витро (сравни фиг. 21C, D с фиг. 21E и F). Даже после 4 дней в культуре имелось очень мало не-нейронных клеток, что отмечено по отсутствию уплощенных фазотемных клеток. Для оценки действия BDNF на перегородочные холинергические нейроны в отношении их выживания в культуре использовали гистохимическое окрашивание на AChE и иммуногистохимическое окрашивание на AChE-рецептор. Типичные морфологии AChE-положительных нейронов показаны на фиг. 21G, H. Несколько NGF-рецептор-положительных нейронов показано на фиг. 21I.
BDNF дал 2,4-кратное увеличение количества AChE-положительных клеток в культурах низкой плотности (определяемой как плотность клеток между 66,700 - 133,300 клеток на квадратный сантиметр) перегородочных клеток крысиного эмбриона, как показано на фиг. 22A. При той же клеточной плотности действие NGF было слегка ниже (1,9-кратное). Обнаружение того, что BDNF как и NGF, может продуцировать увеличение количества AChE-положительных нейронов в перегородочных культурах, побудило нас изучить, действуют ли эти два фактора на сходные или различные нейронные популяции. Как показано на третьей панели фиг. 22A, совместное добавление насыщающих уровней BDNF и NGF не приводит к большему увеличению количества AChE положительных нейронов, чем с каждым фактором в отдельности. В отношении нейронного выживания, эти результаты предполагают, что холинергические нейроны в крысиных перегородочных культурах, реагирующих на BDNF и NGF должны быть тесно перекрывающихся популяций.
Действие BDNF на выживание AChE-положительных нейронов зависело от плотности, при которой росли клетки (фиг. 22A). При высоких плотностях посева (от 200000 до 266000 клеток на см2) ни BDNF сам, ни в сочетании с NGF, не производил повышения количества AChE-экспрессирующих нейронов. Это может происходить из-за повышенных эндогенных уровней нейротропной активности или, альтернативно, - от способствующего выживанию повышенного междуклеточного (клетка - клетка) контакта, происходящего при высоких клеточных плотностях.
При низкой клеточной плотности количество AChE-положительных клеток, повышенное как функция концентрации BDNF с максимальной реакцией, наблюдалось при дозе 10 нг/мл, которая увеличила количество AChE-положительных клеток на лунку от контрольных значений 169,2±12,9 до 388±26,2 (2,5-кратное увеличение) в обработанных культурах. В сравнении, максимальная реакция на NGF наблюдалась при 50 нг/мл и дала повышение AChE-положительных клеток на лунку от 121,0±7,6 до 231, 7±12,9 (1, 9-кратное увеличение). Горизонтальный участок ("плато") в кривой, графически отображающей реакцию клеток, кажется стабильной для BDNF и NGF в том, что нет уменьшения количества AChE-положительных нейронов при дозах высотой порядка 100 нг/мл.
Очевидно, что в культурах с низкой плотностью клеток и BDNF, и NGF способны увеличить количество клеток, обнаруженных AChE гистохимией. Возможно, что это увеличение происходит из-за спасения холинергических клеток, погибающих в культуре вследствие отсутствия ростового фактора, для пополнения предшествующих клеток или для индукции холинергического маркера, AChE. В попытке дифференцировать эти возможности были проведены позже дополнительные эксперименты (фиг. 23) на сериях культур, поддерживаемых всего в течение 12 дней. Одну группу культур, обрабатываемых через 5-6 часов после посева, поддерживали в присутствии BDNF или NGF все 12 дней. Вторую группу культур делали без ростовых факторов в течение 5 дней, а затем обрабатывали BDNF-или NGF в течение последних семи дней (-5/+7). При сравнении этих двух групп культур реакция на BDNF или NGF, добавленные после 5 дней, была в основном такой же, которая наблюдалась, когда клетки росли постоянно в присутствии этих факторов (сравни сплошные и штрихованные полосы на фиг. 23). Результаты были на удивление разными, однако, в третьей группе культур, в которой BDNF или NGF присутствовали только последние 5 дней (-7/+5). При этих условиях ни NGF, ни BDNF не были способны повысить количество AChE-положительных клеток. В других экспериментах было установлено, что экспонирование перегородочных нейронов NGF или BDNF на 3-4 дня недостаточно для производства большого увеличения AChE энзимной активности и, таким образом, для обнаружения клеток AChE окрашиванием. Поэтому отсутствие обнаружимого увеличения количества AChE-положительных нейронов в "-7/+5" культурах происходило не из-за того, что время экспозиции ростовому фактору было недостаточным для обеспечения индукции AChE-положительных клеток.
Ин виво было показано (Montero и Hefti, 1988, J. Neurosci 8: 2986-2999; Higgins и др. , 1989, Nueron 3: 247-256), так же как и ин витро, что NGF способен регулировать уровни его собственного рецептора, как измерено на уровне мРНК и на уровне белка. Поскольку было обнаружено, что BDNF проявляет похожие эффекты с NGF в отношении индукции холинергического фенотипа и клеточного выживания в перегородочных культурах, испытывалась потенциальная возможность того, что BDNF может регулировать уровни NGF-рецептора, как определено количеством IgG-192 иммуноположительных клеток. При 10 нг/мл BDNF произвел трехкратное увеличение количества NGF-рецепторных иммунопозитивных клеток, как показано на фиг. 24. Интересно, что при более высоких концентрациях (25-100 нг/мл) действие BDNF прогрессивно уменьшалось. Таким образом, ответ на дозу в этом случае заметно отличался от наблюдавшегося действия BDNF при других параметрах на эти культуры, например, на количество AChE-положительных клеток. В качестве положительного контроля эти клетки обрабатывали NGF при концентрации 50 нг/мл, что также приводило к трехкратному увеличению количества положительных клеток. Поэтому на основе IgG-окрашивания BDNF оказывает приблизительно равный эффект по сравнению с NGF на регулирование повышения экспрессии NGF-рецептора.
В дополнение к действию на выживание клеток исследовали возможность того, что BDNF мог способствовать холинергическим фенотипным чертам клеток. Линейное повышение САТ активности было обнаружено в перегородочных культурах, выращенных в присутствии возрастающих количеств BDNF до 50 нг/мл, когда реакция насыщалась на уровне 1,8-кратного увеличения по сравнению с контрольными значениями, как показано на фиг. 25. Индуцирование САТ активности в параллельных культурах, обработанных NGF, достигло плато при 25 нг/мл, представляя 3,4-кратное увеличение сверх контрольных значений. Реакция на BDNF или NGF не показала значительного снижения в три раза концентрации, достаточной для производства насыщающего эффекта. Индуцирование САТ-активности BDNF или NGF не повлияло на уровень HNP-независимой трансферазной активности (HNP=N-гидроксиэтил-4-(1-нафтилвинил/пиридиум).
В свете открытия, что BDNF, как и NGF, дает повышение САТ-активности в перегородочных культурах, изучалось реагирование клеток на совместное обрабатывание обоими факторами (табл. VI). Для этих экспериментов использовали две концентрации BDNF, 5 и 25 нг/мл, которые увеличивали САТ-активность на 1,4 и 2,6 раза соответственно. Когда с BDNF соединяли NGF (50 нг/мл), который сам увеличивал САТ-активность в 2,8 раза, уровень САТ-активности отразил по меньшей мере дополнительный эффект (табл. VI). При использовавшихся концентрациях BDNF чисто дополнительный эффект от NGF привел к увеличению соответственно в 4,2 и 5,4 раза сверх контрольных значений. Наблюдавшиеся значения действительно были несколько выше, чем просто дополнительный эффект.
Перегородочные клетки выращивали в течение 12 дней в 5HS/N3 в присутствии указанных концентраций трофических факторов, САТ-активность определяли как описано. Значения представляют среднее ± SEM из 4-5 определений.
Была проверена возможность того, что биологическая реакция, наблюдаемая при обработке BDNF, происходит от BDNF-зависимого освобождения эндогенного NGF. Чтобы изучить этот вопрос, для блокирования любой связанной с NGF реакции использовали моноклональное антитело против NGF (антитело 27/21, Korsching и Thoenen 1983). Как показано в табл. VII, действие NGF на САТ-активность было значительно снижено антиNGF, тогда как индуцированная BDNF-реакция осталась в основном без изменений.
Следует заметить, однако, что основной уровень САТ-активности был снижен примерно на 20% в культурах, обработанных только антиNGF, по сравнению с необработанными культурами (контролем). Тем не менее, наблюдавшееся повышение САТ-активности в перегородочных культурах, вызываемое BDNF, кажется прямым эффектом, не требующим посредника в виде повышения эндогенного NGF.
После 12-дневного периода роста в среде 5HS/N3 в присутствии указанных концентраций трофического фактора, перегородочные клетки (посеянные с плотностью 2, 3•105 клеток на см2) были собраны. САТ-активность определяли как описано в разделе экспериментальные методы, представленные значения - это среднее ± SEM из 6 определений.
Также была исследована возможность того, что AChE-активность равным образом регулируется вместе с САТ-активностью посредством BDNF или NGF (фиг. 26). В отличие от САТ-активности дозовая реакция AChE на BDNF или NGF кажется совсем похожей на то, как ферментная активность повышается линейно вплоть до концентрации 50 нг/мл. При этой концентрации NGF и BDNF повышали AChE-активность на 274% и 234% соответственно по сравнению с необработанными контрольными образцами.
Исследовалось изменение BDNF и NGF вызванного повышения САТ-активности по времени (фиг. 27). САТ-активность в культурах, обработанных 50 нг/мл BDNF, повышалась до 170% контрольных значений за три дня. Это увеличение продолжалось до шестого дня, когда оно вышло на "плато" на уровне 2,5-кратного повышения контрольных значений в течение всего остального периода исследований. Напротив, САТ-реакция на NGF (50 нг/мл) была более быстрой, достигнув на третий день 2, 5-кратного превышения контрольных значений. Увеличение САТ-активности далее продолжалось линейно, достигнув 3,2-кратного превышения контрольных значений после 12 дней.
Было показано, что астроциты, выращиваемые некоторое время в культуре, синтезируют различные нейротропные активности в дополнение к NGF (Lindsay, 1979, Nature 280: 80-82: Lindsay и др., 1982, Brain Res. 243: 329-343; Alderson и др. , 1989, Dev. Brain. Res 48: 229-241). Хотя мы исключили возможность того, что настоящие наблюдения на BDNF вызваны повышением уровней NGF, понятно, что BDNF может действовать непрямым образом, посредством регулирования экспрессии других нейротропных факторов, особенно в глиальных клетках. Чтобы оценить возможность того, что не-нейроные клетки могли бы повлиять на действие BDNF на САТ-активность в перегородочных культурах, сравнивали реакцию холинергических нейронов на BDNF в смешанных глиально-нейронных культурах и обогащенных нейронами культурах (фиг. 28). В обогащенных нейронами культурах реакция САТ-активности на BDNF проявила колоколообразную кривую; максимальное увеличение было достигнуто при дозе 5 нг/мл, а при концентрации 25 нг/мл имелось значительное снижение энзиматической активности (p > 0,001, по сравнению с 15-25 нг/мл BDNF). Подобно результату, показанному на фиг. 25, САТ-реакция на BDNF в присутствии слившегося глиального слоя была линейной в этом случае - вплоть до высшей испытанной дозы, 25 нг/мл BDNF.
В присутствии не-нейронных клеток требовались более высокие уровни BDNF для производства эквивалентного повышения САТ-активности по сравнению с активностью, обнаруженной в аналогично обработанных обогащенных нейронами культурах. Тем не менее, способность BDNF максимально стимулировать САТ-активность в перегородочных холинергических нейронах была, в основном, одинаковой в присутствии или в отсутствие глиальных клеток.
На основании САТ- и AChE-активности теперь очевидно, что BDNF может действовать подобно NGF в усилении холинергических фенотипных маркеров. Чтобы исследовать это далее, действие BDNF на уровень Na+-зависимое, высокоаффинное усвоение холина сравнивали с действием NGF (фиг. 29). Клетки выращивали 12 дней в присутствии 50 нг/мл BDNF и они аккумулировали холин до уровня в 3,8 раза выше уровня контроля. В параллельных культурах NGF (25 нг/мл) вызывал 2,3-кратное увеличение аккмулирования холина.
16.3. Обсуждение.
По сравнению с установленными эффектами на периферийных нейронах (Barde и др, 1982, EMBO J., 1:549-553; Lindsay и др., 1985, Dev. Biol., 112: 319-328; Davies и др. , 1986, J. Neurosci 6: 1897-1904; Hofer и Barde, 1988, Nature, 331: 262-263) нейронная специфичность BDNF в центральной нервной системе хуже охарактеризована. До сих пор единственно известными реагирующими ЦНС-нейронами является небольшая их субпопуляция Thr-1 положительные клетки ганлия сетчатки, впервые идентифицированные в культурах сетчатки крысиного эмбриона E17 (Johnson и др., 1986, J.Neurosci 6: 3031-3038). Дальнейшие исследования также установили действие BDNF на взрослые клетки ганглия сетчатки в эксплантатных культурах (Thanos и др., 1989, Eur. J. Neuro 1. 19-26).
В данном исследовании показано, что BDNF повышает выживание в культуре холинергических нейронов из перегородки крысиных эмбрионов, как показано AchE гистохимическим окрашиванием, увеличивает экспрессию различных холинергических фенотипных маркеров, т.е. энзиматическую активность AchE и САТ, и высокоаффинное усвоение холина. Дополнительно было обнаружено, что BDNF повышает экспрессию NGF-рецептора в перегородочных культурах.
В начальных экспериментах было обнаружено, что BDNF увеличивает количество AChE-положительных нейронов в E17 перегородочных культурах примерно вдвое. Что касается NGF (Hartikka и Hefti; 1988, J. Neurosci 8: 2967-2985), то этот эффект был наиболее очевиден при низких плотностях клеток. Интересно, что совместное добавление NGF и BDNF было неэффективным в отношении дальнейшего увеличения количества AchE положительных клеток сверх уровня, наблюдавшегося в присутствии каждого фактора роста по отдельности. Это наблюдение подтверждает, что BDNF и NGF действуют в основном на ту же самую популяцию перегородочных холенергических нейронов.
Как вначале сообщалось для NGF (Hefti и др., 1985, Neuroscience, 1:55-68), BDNF не повышал выживание холинергических нейронов в культурах, сделанных с относительно высокой клеточной плотностью. Было обнаружено в виде пропорции посеянных клеток, что выживание холинергических нейронов выше при высокой клеточной плотности, чем при низкой клеточной плотности, даже в отсутствие любого нейротропного фактора. Имеется несколько возможных объяснений этому, включая повышенный контакт клеток между собой, благоприятное действие увеличенного количества не-нейронных клеток или более чем пропорциональное повышение эндогенной нейротропной активности. В отношении последней возможности не было обнаружено намека на существенное повышение уровней эндогенного NGF, поскольку добавление излишка антиNGF к необработанным культурам высокой плотности только понизило основную САТ-активность на 20%. В таких культурах эндогенный NGF был способен произвести 3,4-кратное повышение САТ-активности.
Как наблюдали на уровне протеина и на уровне мРНК, во многих сообщениях указывается, что NGF может регулировать в сторону повышения экспрессию своего собственного рецептора как у нейронов в культуре, так и ин виво. В данном исследовании использовалось моноклональное антитело IgG-192 для обнаружения NGF-рецептора. Это моноклональное антитело было охарактеризовано как на сенсорных крысиных нейронах, так и на крысином головном мозге (Taniuchi и Johnson, 1985, J.Cell Biol 101:1100-1106). Подтвердив обнаруженное Хартикка и Хефти явление, что NGF может повышать уровень своего рецептора в перегородочных культурах, что отмечается по увеличению количества IgG-192 иммуноположительных клеток, мы дополнительно нашли, что BDNF увеличивает количество IgG-192 иммуноположительных клеток до трехкратного. Реакция на BDNF была двухфазной в том, что при высоких концентрациях в пределах 25-100 нг/мл BDNF не дал столь значительного увеличения количества положительных клеток, как при 10 нг/мл. Интересно, что этот тип реакции полностью отличался от того, как BDNF вызывал увеличение количества AChE-положительных нейронов. В последнем случае при высоких концентрациях BDNF не наблюдалось снижения максимального эффекта от BDNF. Следует заметить, недавно было показано, что низкоаффинный NGF-рецептор также связывают с подобной аффинностью, предполагая тем самым, что NGF- и BDNF-рецепторы низкой аффинности могут быть идентичными (Rodriguez - Tebar и др., 1990, Binding of Brain-Derived Neurotrophic Factor to the Nerve Gravth Factor Receptor Neuron, 4: 487-492).
Возможное сходство механизмов действия BDNF и NGF предполагалось согласно открытию того, что BDNF равно с NGF вызывал AChE-активность. Хотя также было обнаружено, что BDNF способствует выживанию холинергических нейронов в сходной степени по сравнению с NGF, действие BDNF на САТ-активность не было столь большим, как действие NGF. В нескольких экспериментах максимальная индукция САТ посредством BDNF была в пределах 1,8-2,6 раз, тогда как значения более 3-кратных обычно наблюдали с NGF. Таким образом, хотя может быть имеется некоторое сходство механизма действия BDNF и NGF, вполне возможно, что имеются тонкие различия в экспрессии и регуляции их соответствующих рецепторов в соединении этих рецепторов со вторым посредником (посредниками), необходимым для индуцирования САТ-активности.
Исследование изменений по времени поддержало предположение, что различия в уровне индуцирования САТ-активности BDNF и NGF могут представлять различные регуляторные пути. Перегородочные нейроны в культуре реагировали на BDNF повышением САТ-активности до 6-го дня, когда реакция выходила на "плато". Напротив, индукция САТ-активности в ответ на NGF показала отлогий линейный подъем от 3-го до 12-го дня. Прекращение повышения индуцирования САТ-активности на 6-й день в обработанных BDNF культурах может указывать на развитийные изменения в экспрессии либо BDNF-рецепторов, либо требуемого компонента на пути реакции. Учитывая, что количество холинергических нейронов (AChE-положительных нейронов), обнаруживаемых в высокоплотных культурах на 12-й день, не зависит от экзогенного BDNF или NGF, кажется маловероятным, что дифференциальная гибель нейронных клеток происходит вследствие различий в изменении по времени, которые были обнаружены при индуцировании САТ-активности этими двумя ростовыми факторами.
В начальных экспериментах, включающих BDNF-обработку холинергических клеток базального переднего мозга, не делалось предположений, касающихся первичных клеточных типов, нейронного или глиального, отвечающих на этот нейротропный фактор. Хотя исследования, проводившиеся на высокообогащенных культурах периферийных нейронов, доказывают, что BDNF оказывает свое действие непосредственно на нейроны (Линдсей и др., Dev.Biol.112: 319 - 328; Линдсей, 1988, J.Neurosci 7: 2394 - 2405), можно предположить, что нейронные эффекты BDNF, видимые в настоящем исследовании, могли вызываться через первичное действие BDNF на астроглию или другие нейронные клетки. Однако было обнаружено, что BDNF равноэффективен в индуцировании САТ-активности в присутствии слившегося монослоя преимущественно астроглиальных клеток или в культурах, значительно лишенных глиальных клеток, обработкой митотическим ингибитором, цитозин-арабинозой. В этих экспериментах имелось любопытное наблюдение - форма кривых зависимости от доз BDNF отчетливо различалось в двух случаях. В присутствии глиального монослоя реакция на BDNF была линейной с увеличением концентрации BDNF. В обогащенных нейронами культурах реакция на BDNF в зависимости от его дозы сдвинулось влево. Одно возможное объяснение этих результатов состоит в том, что глия сама может экспрессировать BDNF-рецептор, снижая таким образом эффективную концентрацию лиганда, имеющегося для рецептора, который локализуется в нейронах. Альтерантивно подавляющее действие астроцитов могло быть непрямым, независимым от эффекта на концентрацию лиганда или рецепторную экспрессию, и передаваться, например, посредством значительно усиленной деградации BDNF. О сходных наблюдениях сообщали Хартикка и Хефти (1988, J.Neurosce, 8: 2967 - 2985), а также Хонеггер и Ленуар (1982, Dev.Brain.Res. 3: 229-238). Было продемонстрировано, что глиальные клетки различных областей мозга могут оказывать значительное влияние на морфологию связанных с ними нейронов (Prochiantz и др., 1979, Proc. Natt. Acad. Sci. США, 76: 5387 - 5391; Prochiantz и др., 1981, Nature, 293-570-572). Таким образом, перегородочная глия может обладать присущими ей свойствами либо мембранной, либо секторной природы, которые могут действовать в направлении подавления экспрессии холинергического фенотипа. Регуляторные свойства астроцитов из аммонова рога теперь были исследованы при похожих условиях опыта.
На основании представленных данных очевидно, что и BDNF, и NGF, оба способны увеличивать функцию перегородочных холинергических нейронов. Интересно провести изучение физиологической вероятности того, что два высокогомологичных нейротропных фактора действуют на единственную нейронную популяцию. Имелось какое-то предположение, что действия BDNF и NGF могут вначале перекрываться в процессе раннего развития периферийных нейронов, особенно на субпопуляции нейронов ганглия дорсального корня или их предшественников, и затем позднее разделяться и действовать независимо на разные популяции (Lindsay и др., 1985, Dev. Biol. 112: 319 - 328: Ersberger и Rohrer, 1988, Dev. Biol., 126: 420 - 432; Barde 1989, Neuron 2: 1525 - 1534).
Это, однако, не было точно установлено с использованием соответствующих маркеров для определения суб-популяций сенсорных нейронов. Поскольку настоящее исследование сфокусировано на эмбрионных перегородочных культурах в единственной временной точке развития, E17, есть возможность того, что разделительный феномен такого вида может происходить при более поздних стадиях развития в перегородке. Таким образом, могут оказаться интересными временные и пространственные различия в реакции перегородочных холинергических нейронов на BDNF и NGF.
Теперь было бы интересно исследовать действие BDNF на холинергические нейроны базального переднего мозга ин виво, где теперь имеется свидетельство тому, что NGF из аммонова рога может вовлекаться в нормальное развитие и поддержание холинергических нейронов переднего мозга ин виво. Будет интересно установить, играет или нет BDNF подобную роль ин виво. Недавнее открытие того, что BDNF мРНК особенно обильна в аммоновом роге, подтверждает такую роль BDNF.
17. Пример. Энзиматическое превращение BDNF в активный, зрелый BDNF.
17.1. Материалы и методы.
Коммерчески доступная эндопротеиназа Arg-C (очищенная из подчелюстных желез мыши) была использована для энзиматического превращения большой предшествующей формы hBDNF (prepro BDNF) в биологически активный зрелый протеин. Это энзиматическое расщепление было совершено реакцией ин витро. Субстратом в этой энзиматической реакции был prepro BDNF, синтезированный с CHO-DG 44 клетках, которые были выделены в клеточную культуральную среду и собраны. Культуральная среда состояла из среды F-12 (Haм), свободной от нуклеозидов, с добавлением 1% сыворотки плода теленка (FBS) и по 1% пенициллина и стрептомицина. Реакцию проводили при 37oC в течение 5 минут с использованием 5 ед. энзима и 50 мкл клеток CHO-DG44 клеточного супернатанта, содержащего prepro BDNF.
При этих условиях было осуществлено расщепление prepro BDNF в зрелый hBDNF.
17.2. Результаты и обсуждение.
Prepro-форму рекомбинантного hBDNF (приблизительно 31000 Дальтон) полностью превращали в зрелую биоактивную форму hBDNF (приблизительно 12000 Дальтон) ин витро с использованием эндопротеиназы Arg-l. Рекомбинантный нейротропный фактор человеческого мозга был успешно продуцирован в клетках млекопитающего (CHO-DG 44). Ген hBDNF был стабильно интегрирован в геном CHO-клеток и амплифицирован с использованием метотраксатной стратегии амплифицирования. Рекомбинантный hBDNF секретировался в среду и был биологически активен. Метаболическое мечение с последующим электрофорезом на SDS-полиакриламидном геле (15%) продемонстрировало, что большинство синтезированного [35S] -меченого hBDNF продукта, секретированного в среду, мигрировало в виде prepro-формы с молекулярным весом 31000 (фиг. 30, дорожка 2). [35S]-меченые протеины, секретированные из клеток CHO-DG 44 дикого типа, показаны на фиг. 30, дорожка 1. Чтобы генерировать преимущественно зрелую форму hBDNF (молекулярного веса 12000), создали специальную ин витро-стратегию для энзиматического расщепления prepro-формы hBDNF.
Продукты трипсинового переваривания показаны на фиг. 30, дорожки 3 - 6. Трипсин бычьей поджелудочной железы (закупленный у Вортингтона), свободный от химотрипсина, растворяли в фосфатно-буферном физиологическом растворе и в раствор добавляли 50 ul аликвоты [35S]-меченого CHO-клеточного hBDNF. Использовавшиеся концентрации трипсина были 25, 50, 75, и 100 ug/мл. [35 S]-меченые продукты реакции показаны на фиг. 30, дорожки 3-6 соответственно. Реакцию энзиматического расщепления проводили при 37oC в течение 5 минут. Постепенное снижение интенсивности мечения у 31000-prepro-hBDNF наблюдалось с его соответствующим увеличением для 18500 - продукта. Зрелая форма hBDNF (молекулярный вес 12000) не генерировалась при этих условиях. Фиг. 30, дорожка 7 показывает, что [35S]-меченые продукты реакции после энзиматического переваривания CHO-клеточного hBDNF эндопротеиназой Arg-C, выделенной из подчелюстной железы мыши и полученной от фирмы Берингер Мангейм Кемикалс, Инкю. 100 ед. этого фермента было восстановлено из лиофилизированного приготовления с 100 ul воды Milli-Q и хранилось при -20oC. Для энзиматического переваривания CHO-клеточного hBDNF, использовали 5 единиц (5 ul) энзима и 50 ul [35S]-меченого протеина из CHO-клеточного супернатанта, содержащего prepro hBDNF (фиг. 30, дорожка 2). Как видно на фиг. 30, дорожка 7, 5 единиц эндопротеиназы Arg-C были способны превратить prepro-форму hBDNF (31000 м.в. ). в зрелую форму hBDNF (12000 м.в.) за 5 минут при 37oC.
Энзиматическая обработка эндопротеиназой Arg-C супернатантов CHO-клеток, экспрессирующих hBDNF повышала уровень биологической активности BDNF по отношению к необработанным супернатантам. Супернатанты от CHO-DG44 клеток, экспрессировавших hBDNF, обрабатывали в течение 5 мин эндопротеиназой Arg-C при 37oC. 200 ul супернатанта обрабатывали 20 единицами энзима и как обработанный, так и необработанный hBDNF исследовали на физиологическую активность с использованием эксплантантов E8 ганглия дорсального корня цыпленка. Через 24 часа после добавления hBDNF оценивали (качественно) отрастание нейритов. Наблюдалось, что в соответствии с вышеизложенным обработанные эндопротеиназой образцы BDNF были значительно более биоактивны, чем контрольные (необработанные) образцы hBDNF. Концентрационная кривая по этим данным показана на фиг. 31.
В заключение, очищенная эндопротеиназа Arg-C может быть использована для генерирования зрелой биоактивной формы рекомбинантного нейротропного фактора из человеческого мозга из неполностью процессированных молекул предшественника (то есть, про-BDNF). Этот новый метод позволит эффективное широкомасштабное производство биоактивного человеческого BDNF из культуральной системы клеток млекопитающих. Например, можно иммобилизовать эндопротеиназу Arg-C на твердой матрице и включить стадию эффективной колонной хроматографии.
18. Пример. Мозговой нейротропный фактор способствует выживанию и созреванию допаминергических нейронов крысиного вентрального среднего мозга.
18.1. Материалы и методы.
18.1.1. Клеточные культуры.
Диссоциированные культуры создали путем энзиматической и механической диссоциации ткани вентрального среднего мозга, извлеченных диссекцией из крысиных эмбрионов возраста E14-E15. Обычно ткань, собранную от двух или трех пометов крысиных эмбрионов от одновременно осемененных крыс вида Spraque-Dawley трипсинизировали (0,125%, Вортингтон) в среде F-12 (Жибко) в течение 20 минут при 37oC. После промывания в ростовой среде (МЕМ, содержащая следующие добавки: глютамин 2 мМ, глюкоза 6 мг/мл, пенициллин G 0,5 ед/мл, стрептомицин 5 мкг/мл, сыворотка плода теленка 7,5%) ткань кратковременно центрифугировали при низких оборотах в течение 5 минут и дебрис диссоциировали тритурацией. Через еще 1-2 минуты для того, чтобы недисперсные клетки осели, единственную клеточную суспензию засевали на 35 мм чашки (предварительно покрытые полиорнитином и ламинином; Линдсей и др., 1985, Dev. Biol. 112: 319), содержавшие ростовую среду с плотностью 5 • 104 клеток на кв.см. После инкубации до утра в ростовой среде, чтобы позволить клеткам прикрепиться, клетки культивировали в присутствии или отсутствие BDNF в свободной от сыворотки определенной среде, описанной Боттенштайном и Сато (Bottenstein и Sato 1979, Proc. Nath. Acad. Sci США 76: 514), за исключением того, что инсулин был включен при концентрации 20 нг/мл. Для визуализации допаминергических клеток культуры фиксировали 4%-ным параформальдегидом, длительно промывали, делали проницаемыми 0,02%-ным сапонином в буфере Соренсена с 1,5% лошадиной сыворотки, и окрашивали моноклональным антителом на крысиный ТН (фирма Берингер, Маннгейм). Связанное первичное антитело было визуализировано с использованием набора Вектасталин АВС (Лаборатории "Вектор").
18.1.2. Измерение усвоения допамина.
Культуры приготавливали как описано в 18.1.1 выше, и выращивали указанное количество дней либо в присутствии, либо в отсутствие свиного BDNF 50 нг/мл. Усвоение допамина определяли в тройных культурах в указанные временные моменты. Клетки предварительно промывали в специальном буфере следующего состава: 136,8 мМ NaCl, 2,7 мМ KCl, 8 мМ Na2HPO4•7H2O, 1,5 мМ KH2PO4, 5 мМ глюкозы, 1 мМ CaCl2, 1 мМ MgSO4, 0,1 мМ аскорбата и 0,1 мМ паргилина с pH 7,4. Образцы, подлежащие измерению на не-нейронное усвоение 3H-допамина, содержали в указанном буфере бензотропин (BZT) при концентрации 5 мкМ. После промывания клетки предварительно нагревали до 37oC на пять минут в свежем буфере, и теперь 3H-допамин (NEN, 40 Ci/ммоль) был добавлен до конечной концентрации 50 нМ. Культуры затем инкубировали при 37oC в течение 15 минут, раствор удаляли, и клетки помещали на лед, четырежды промывали описанным выше ледяным буфером. Клетки собирали посредством добавления 0,5н. NaOH к культуральным чашкам, и щелочной экстракт подсчитывали в сцинтилляционном счетчике Бекмана с 15 млс Ультима Гольд (сцинтилляционной жидкости). Специфичный усвоенный нейронами допамин был определен тем, что усвоение (срм), наблюдавшееся в отсутствие BZT, было меньше, чем наблюдалось в присутствии BZT. Обычно, между 70-90% общего усвоения было способно ингибироваться посредством BZT. В реплицированных культурах в каждый момент времени количество TH+-нейронов определяли иммуноцитохимическим окрашиванием, и показанные результаты представляют усвоение, нормализованное на основании TH+-базиса.
18.1.3. Временная экспрессия BDNF.
COS M5 клетки трансфецировали вектором plD M8, содержащим последовательность, кодирующую человеческий BDNF. Через 72 часа после трансфецирования собирали 50 мл супернатанта культуры и диализовали против 6 М мочевины. Диализованный супернатант затем соединяли с амфолинами (pH 3,5 - 10 BioRad) на 4,5 часа. Собирали фракции, диализовали против 25 мМ NaPO4 буфера с pH 7,6 с использованием диализной трубки, которую предварительно промывали в BSA (0,5 мг/мл), чтобы предотвратить неспецифичную абсорбцию. Фракции исследовали на активность, способствующую отрастанию нейритов в культурах E8 эксплантов ганглиев дорсального корня цыпленка. Активные фракции были собраны в пул. Анализ активного пула посредством SDS PAGE и последующего серебряного окрашивания (Wray и др., 1981, Anal. Biochem 118: 197) на 8-18% геле обнаружил, что имеется слабая полоса 68 кД, соответствующая BSA, и единственная полоса примерно 12 кД, соответствующая тому, что ранее наблюдалось для свиного BDNF (Barde и др., 1982, ЕМВО, 1:549) (данные не показаны). В сравнительных биоэкспериментах, проводившихся на эксплантантах ганглиев куриного дорсального корня, нодозном и симпатическом, для выявления специфической активности и нейронной специфичности очищенного рекомбинантного человеческого BDNF результаты оказались сходными с очищенным свиным BDNF.
18.1.4. Производство hBDNF из трансфецированных CHO-клеток.
CHO-DG44 клетки были любезно подарены д-ром L.Chasin (Колумбийский университет, США). Эти клетки бедны дигидрофолат-редуктазой (dhfr-) (Urlaub и Chasin 1980, Proc. Nath. Acad. Sci США 77: 4216-4220), CHO-DG44 клетки поддерживали в среде F-12 (Нам) с 10% сыворотки плода теленка, 1% пенициллина и стрептомицина и 2 мМ глютамина. Клетки пассировали дважды в неделю и обычно проверяли на загрязнение микоплазмой. Вся плазмидная ДНК была очищена двойным цезий-хлоридным электрофорезом перед трансфекцией. Ген человеческого мозгового нейротропного фактора субклонировали в экспрессионный вектор млекопитающих pCDM8 для генерирования pC8hB, как описывалось ранее (Maisonpierre и др., 1960, Science 247: 1446-1451). Ген ослабленной дигидрофолат-редуктазы, p410, был любезно предоставлен др-ом L. Chasin. Эти две генные конструкции были совместно трансфецированы с CHO-DG 44 клетки посредством электропорации (рассмотренной Shigekawa и Dawer, 1988, BioTechniques 6: 742-751).
Примерно 1•106 клеток трансфецировали 20 ug pC8hB и 0,2 ug p410. Через 48 часов после трансфекции, CHO-DG-44 клетки расщепляли в селекционной среде, состоящей из свободной от нуклеозидов среды F-12 (Нам) без тимидина и гипоксантина; -HT, плюс 10% диализованной сыворотки плода теленка и 1% пенициллина и стрептомицина.
Отдельные колонии выделяли примерно через 10 дней после помещения клеток в свободную от нуклеозидов селекционую среду. Индивидуально отобранные колонии, резистентные к росту в свободной от нуклеозидов среде F-12 (-HT) обрабатывали 0,02, или 0,05, или 0,1 мкМ метотрексата для индуцирования амплифицирования гена (Alt и др., 1978, J. Biol Chem 253: 1357-1370). Резистентные к МТХ (метотрексату) колонии культурировали в виде пулов и далее амплифицировали с 1,5 или 2,0, или 2,5 мкМ МТХ. Была выделена единственная колония, резистентная к 2,5 мкМ МТХ и отобрана для масштабного культурирования и производства hBDNF. Анализ этого клона саузерн-блоттингом (клон DGZ 1000-B-2,5) показал примерно 50-кратную амплификацию BDNF-гена относительно клеток CHO-DG44 дикого типа. Биоэксперименты с эксплантантами ганглиев дорсального корня куриного эмбриона E8 (DRG) были использованы для оценки того, что этот клон продуцировал примерно 9,5 ug hBDNF/мл (не показано). Рекомбинантный hBDNF производили в большом масштабе посредством культурирования DGZ 1000-B-3-2,5 клеток в 480 см2 роллер-флаконах. Среду собирали приблизительно через четыре дня после того, как эти клетки достигали слияния. Очистку BDNF из культурального супернатанта осуществляли как указано выше, для COS-супернатанта.
18.2. Результаты и обсуждение.
Характеризирование молекулярных сигналов, контролирующих нейронное выживание и специфичность мишенной иннервации, представляет фундаментальный интерес не только потому, что это поможет понять, как сформирована нервная система, но также вследствие того, что это, вероятно, приведет к лучшему пониманию механизмов, вовлеченных в поддержание и ремонт зрелых нейронов в подходящих синаптических конфигурациях. Поскольку широко признано, что выживание и поддержание нейронов зависит от специфичных, происходящих из мишенных клеток нейротропных факторов (Леви-Монтальчини, и Ангелетти, 1968, Physiol. Rev. 48: 534; Thoenen и Barde, 1980, Physiol, Rev. 60: 1284; Purves, 1988, в "Body and Brain", Harward University Preis, Cambridge, Barde, 1989, Heuron 2:1525; Lindsay 1988, The Making of the Nervous Systems, Oxford, p. 148), то очень мало таких молекул было до сих пор полностью охарактеризовано. До сих пор было показано, что только два нейротропных фактора, фактор нервного роста (NGF) и мозговой нейротропный фактор (BDNF) селективно поддерживают разные нейронные популяции ин виво (Леви-Монтальчини и Ангелетти, 1968, Physiol. Rev. 48: 534; Barde, 1989, Neuron, 2: 1525; Leibrock и др., 1989, Nature, 341: 149).
Нейронные клеточные культуры широко использовались для проведения биоэкспериментов для идентификации нейтропной активности в клетках и тканевых экстрактах (Леви-Монтальчини и Ангелетти, 1968, Physiol. Rev. 50: 1284; Lindsay, 1988, The Making of the Nervous System. Oxford, Varon и Adler, 1981, Adv. Cell Neurobiol, 2: 118) и для определения нейронной специфичности (или специфичностей) очищенных нейротропных факторов (Ebendal, 1989, в "Ner growth Factors", John Wiley и др., 1982, EMBO J., 1: 549; Davies и др., 1986, J. Neurosci., 6: 1897; Lindsay и др., 1985, Dev. Biol, 112: 319).
До сих пор большинство исследований концентрировались на нейротропных потребностях различных классов нейронов периферийной нервной системы. Доступность NGF сделала его пока что наиболее хорошо охарактеризованным нейротропным фактором: было показано, что NGF /ин витро и ин виво/ необходим для выживания и поддержания нейронных субпопуляций как периферийной, так и центральной нервной системы (Леви-Монтальчини и Ангелетти, 1968, Physiol. Rev. 48: 534; Thoenen и Barde, 1980, Physiol Rev., 60: 1284, Whittemore и Seiger, 1987, Brain Res. Rev. 12: 439, Snider и Johnson, 1989, Ann. Neurol. 26: 489; Heffi и др., 1989, Neurobiology of Aging 10: 515; Martinez и др., 1987, Brain Res. 412: 295; Takei и др., 1988, J. Neurochem., 51: 1118; Hartikka и Hiffi, 1988, J. Neurosci. 8: 2967).
Трудности в идентифицировании нейротропных ростовых факторов для неспецифичных нейронов центральной нервной системы вызвала ограниченная доступность очищенных препаратов нейротропных факторов, отличных от NGF, а также трудность приготовления гомогенных препаратов специфических нейронных популяций. Два недавних наблюдения понудили нас исследовать действия BDNF на выживание ЦНС-(нигральных) допаминергических нейронов. Прежде всего, теперь ясно, что ген BDNF экспрессируется в ЦНС, и на сравнительно более высоких уровнях, чем NGF-ген (Leibrock и др., 1989, Nature 341: 149). Во-вторых, сообщалось, что протеин, частично очищенный из бычьего стриатума (Striatum) мишени нигральных допаминергических нейронов, - с характеристиками, очевидно сходными с характеристиками BDNF, может повышать ин витро выживание допаминергических нейронов, приготовленных из среднего мозга (DalToso и др., 1988, J. Neurosci. 8:733).
Фиг. 32A иллюстрирует типичный вид диссоциированных клеток вентрального среднего мозга после девяти дней в культуре.
Фактически все из этих клеток имели фазояркие перикарионы и длинные отростки. Явно было несколько клеток фибробластовой морфологии, но не было обнаружено астроглиальных клеток, когда культуры окрасили с антителами на астроглиальный маркер, - глиальный фибриллярный кислородный протеин (GFAP; Bignami и др. , 1972, Brain Res. 43:429). Для визуализации допаминергических нейронов в этих культурах иммуноцитохимическое окрашивание проводили с моноклональными антителами к TH. Как ожидалось, количество TH+-нейронов было небольшим (стрелки, фиг. 32B, 32C) и изменялось в зависимости от времени в культуре, между 0,1 - 0,5% от посеянных клеток. Среди TH+-клеток были отмечены различные нейронные морфологии, как показало на фиг. 31.
Во всех культурах неизменно имелось постепенное уменьшение количества TH+-клеток (фиг. 34A, 35) после первых 3 - 4 дней ин витро. При 8 дн. в культуре, например, количество TH+-клеток в контрольных культурах составляло только 25% от того, что было найдено в подобных культурах на второй день (фиг. 35). Однако в культурах, обработанных BDNF, утрата TH+-клеток была значительно снижена по сравнению с контролем. Во все исследованные моменты после 8 дней ин витро количество TH+-клеток в обработанных BDNF культурах было больше, чем в необработанных контрольных культурах, например, в 1,8 раз выше после единственного добавления BDNF (фиг. 34A) и в 5 раз выше после множественных добавлений (фиг. 35). Даже при добавлении только однажды в культуры, поддерживаемые 8 дней, действие BDNF зависело от его дозы (фиг. 34B). В соответствии с предыдущими сообщениями (Dal Toso и др., 1988, J. Neurosci. 8: 733; Knusel и др., 1990, J.Neurosci; 10: 558) NGF (50 нг/мл) не оказал действия на количество TH+-клеток (фиг. 34C).
Как дальнейший путь изучения действия BDNF на допаминергические нейроны в этих культурах изучали способность TH+ -клеток усваивать 3H-допамин. Как показано в табл. VIII, способность усвоения допамина (нормализованная по TH+-нейрону) заметно увеличилась сверх 8 дней в культуре, но затем падала. Было обнаружено, однако, что BDNF не изменял способность TH+-клеток усваивать допамин, поскольку та же временная зависимость и сходные значения были получены в присутствии или отсутствие BDNF (табл. VIII). Из этого результата была предложена гипотеза, что BDNF, вероятно, действует непосредственно, способствуя выживанию допаминергических нейронов в культурах среднего мозга, в отличие от индуцирования экспрессии допаминергического фенотипа в нейронах, которые необязательно требуют BDNF для выживания. Для того чтобы исследовать это дальше, проводили эксперименты, в которых количество TH+-нейронов определяли в культуре, в которой добавление BDNF вначале задерживали на несколько дней. Обнаружили, что, когда добавление BDNF было задержано до 5-го дня, а количество TH+-клеток определяли на 6, 8 и 10 дни, в этих культурах было видно меньше допаминергических клеток по сравнению с параллельными культурами, в которых BDNF присутствовал со 2-го дня (фиг. 36). Хотя и задержанное добавление BDNF ясно повысило количество TH+-нейронов по сравнению с контролем, однако при изучении в эквивалентные моменты времени было обнаружено, что задержанное добавление никогда не спасало столько нейтронов, как добавление BDNF на 2-й день.
Вместе взятые, эти результаты свидетельствуют против возможности того, что BDNF действует только в плане повышения TH-генной экспрессии в фиксированном количестве клеток, и, предполагают, что BDNF проявляет свое действие посредством повышения выживания допаминергических нейронов, которые иначе погибают, когда этот нейротропный фактор не добавляют к культурам на ранней стадии.
Несколько сообщений касалось стимулирования созревания и увеличения выживания ин витро допаминергических нейронов среднего мозга либо экстрактами из мишенной ткани, или различными нейротропными факторами (Dal Toso и др., 1988, J.Neurosci 8: 733; Knusel и др., 1990, J.Neurosci, 10: 558; Prochiantz и др. , Proc. Natl. Acad. Sci. США, 75: 5387; DiPorzio и др., 1980, Nature, 288: 370; Prochiantz и др., 1981, Nature 293: 570; Denis-Donini и др., 1983, J.Neurosci. 3: 2292).
До сих пор, однако, не было сообщений о полностью очищенной молекуле, непосредственно или селективно поддерживающей выживание TH+-нейронов в полном отсутствии глиальных клеток или усиленном клеточном делении (как наблюдалось, например, с I GF-1 или FGF, Dal Toso и др., 1988, J.Neurosci 8:733). В соответствии с предыдущими сообщениями, в которых использовали ткани раннего мышиного эмбриона (Dal Toso и др., 1988, J.Neurosci, 8:733), Е14 клетки крысиного центрального среднего мозга культурировали в свободной от сыворотки, определенной среде, и эти клетки оказались в основном свободными от астроцитов или фибробластов. Использованная в этом исследовании относительно низкая клеточная плотность порядка 50 000 клеток на см2 снизила эффекты любой возможной эндогенной нейротропной активности и позволила провести более точный подсчет клеток. С учетом представленных здесь результатов, представляется, что нейротропный фактор, частично очищенный из бычьего стриатума Даль Тосо с сотрудниками (1988, J.Neurosci, 8: 733), вероятно, является BDNF. Это предполагает, что BDNF продуцируется в стриатуме и усваивается концами допаминергических нейронов, простирающихся от субстанции нигра. До сих пор, однако, не обнаружено, чтобы мРНК для BDNF (или для других членов семейства нейротропинов) было много в ткани стриатума посредством назерн-блот-анализов.
Ясно, что добавление BDNF даже в случае множественных добавлений, не приводит к полному выживанию нигральных допаминергических нейронов во время исследованного периода культурирования. Этот феномен уже наблюдался при условиях эксперимента, очень сходных с нашими, проведенного Даль Тосо и др. (1988, J. Neurosci, 8: 733), который показал, что эта гибель может быть связана с использовавшейся клеточной плотностью; при более высоких клеточных плотностях (в 4 раза больше использованных здесь) можно было видеть большее количество катехоламин-флуоресцентных клеток, и с спонтанное исчезновение этих клеток снижалось. Возможно, межклеточный контакт необходим для долгосрочного выживания этих клеток.
Дальнейшие исследования, особенно такие, которые направлены на изучение действия BDNF на развитие и поддержание допаминергических нейронов вентрального среднего мозга ин виво, будут необходимы для определения физиологического смысла действий BDNF, описанных здесь. Учитывая специфичную гибель нигральных допаминергических нейронов при болезни Паркинсона, особенно желательно обнаружить нейтропный фактор, который избирательно воздействует на эти клетки. Так, данное открытие того, что BDNF поддерживает выживание этих нейронов, клеток, регулирующих на NGF, побуждает к последующим экспериментам на животных, которые позволят определить, может ли BDNF защитить допаминергические нейроны от нейротоксического действия 6-гидроксидопамина или МРТР (1-метил-4-фенил-1,2,3,6-тетрагидропиридин). Такие исследования могут привести к новому терапевтическому подходу к паркинсонизму.
19. Пример. BDNF производит дозозависимое ингибирование усвоения гамма-аминомасляной кислоты.
19.1. Материалы и методы.
19.1.1. Клеточные культуры аммонова рога.
Аммонов рог удаляли диссекцией у E18-E19 крысиных эмбрионов крыс вида Sprague-Dawley и собирали в среде F10. Ткани измельчали, дважды промывали средой F10 (Жибко) и трипсинизировали 0,25%-ным трипсином (Жибко) в течение 20 минут при 37oC. Трипсин интактировали добавлением содержащей сыворотку среды, состоящей из минимально необходимой среды (MEM) с добавлением сыворотки плода теленка (FCS, 10%), глютамина (2 мМ), пенициллина (25 ед/мл) и стрептомицина (25 ед/мл). Диссоциированные клетки получали путем осторожной тритурации, собирали и центрифугировали при низких оборотах (500 об/мин) в течение 30 с. Центрифугирование повторяли дважды, и клеточный дебрис затем ресуспендировали в содержащей сыворотку среде. Клетки затем засевали на 6 мм лунки или 35 мм чашки, которые были покрыты полиорнитином (10 ug/мл) и ламинином (10 ug/мл). В большинстве экспериментов клетки засевали с низкой плотностью 71000 клеток на см2. Через пять или шесть часов после посева клеток среду заменяли на среду, свободную от сыворотки, содержащую 1% N3 и пенициллин-стрептомицин (25 ед/мл и 25 ug/мл соответственно), при этом добавлялся BDNF. Среду заменяли каждые три-четыре дня с добавлением каждый раз заново BDNF.
19.1.2. Измерение высокоэффективного усвоения гамма-аминомасляной кислоты (GABA).
Высокоэффективное GABA усвоение измеряли с использованием усовершенствованной процедуры Tomozawa и Appel (1986, Brain Res. 399: III-124). Клетки промывали в специальном буфере, содержащем 140 мМ NaCl, 2,6 мМ KCl, 1 мМ KH2PO4, 1 мМ Na2HPO4, 6 мг/мл глюкозы, 1 мМ MgCl2, 1 мМ CaCl2 и 0,1% BSA. После промывания клетки инкубировали с указанным буфером в течение 5 минут при 37oC. 3H-GABA (NSN, NET-19IX, III. 4 Ci/ммоль) затем добавляли при конечной концентрации 12 мМ, и проводили инкубирование при 37oC в течение 10 минут. Клетки затем держали на льду, и трижды промывали указанным буфером. Клетки инкубировали с 0,14 NaOH в течение 2 часов при комнатной температуре, и подсчитывали в экстракте3H-GABA. Обнаружено, что усвоение 3H-GABA было линейным по меньшей мере до 30 минут. Усвоение GABA в не-нейронных клетках ингибировалось добавлением 2 мМ B-аланина, тогда как усвоение, специфичное для нейронов, было проверено путем ингибирования нипекотиновой кислоты в количестве 1 мМ.
19.2. Результаты и обсуждение.
Мы недавно обнаружили признаки высокого содержания BDNF и обогащенных нейронами культур из аммонова рога, но не в астроцитах аммонова рога. Эти данные подтверждают, что BDNF локализуется в нейронах аммонова рога. Чтобы проверить действие BDNF на нейронах аммонова рога в культуре, клетки обрабатывали различными концентрациями BDNF (ротофор-очищенных из COS-супернатантов). В конце 8 дней периода обработки измеряли высокоаффинное GABA-усвоение в нейронах. Как показано на фиг.37, BDNF производил зависимое от дозы ингибирование GABA-усвоения. Таким образом, BDNF может воздействовать на выживание и/или фенотипичную экспрессию GABA-эргических нейронов. BDNF не оказал никакого эффекта на глютаматсодержащие нейроны (как оценено в измерениях усвоения глютамата), ни на уровни нейрофиламентного протеина. Таким образом, действие BDNF на культуры аммонова рога представляется специфичным для GABA-эргических нейронов.
20. Пример. BDNF оказывает защитное действие от токсических эффектов MPP+.
20.1. Материалы и методы.
20.1.1. Измерение действия нейротропных факторов на MPP+-обработанные клетки SH-SV5V.
Клетки SH-SV5V засевали в 24-лунковые чашки с плотностью 1•105 клеток на лунку. Через 24 часа после посева клетки обрабатывали нейтротропными факторами. Через 24 часа после обработки нейротропными факторами клетки обрабатывали MPP+ (10 uM). Через 48 часов после добавления MPP+, жизнеспособные клетки подсчитывали посредством исключения трипаном синим. Использовавшимися нейтротропными факторами были mNGF-COS (разведение 1:5), hBDNF-COS (1:5 разведение), очищенный rCNTF из E.coli (25 нг/мл), очищенный в FGF из бычьего мозга (25 нг/мл), rNT3-COS (разведение 1:5), и очищенный hEGF (25 нг/мл).
20.1.2. Измерение действий нейротропных факторов на MPP+-обработанных культурах вентрального среднего мозга.
Культуры диссоциированных нейронов вентрального среднего мозга создавали из E14 крысиных эмбрионов согласно установленным процедурам (см. раздел 18, выше). Через 48 часов после создания культур их обрабатывали соответствующими нейротропными агентами, как указано в описании к фиг.37.
20.2. Результаты и обсуждение.
Когда BDNF, NGF, CNTF, NT-3, вFGF и FGF раздельно испытывались на способность защищать клетки от 1-метил-4-фенил-пиридиновой (MPP+) токсичности, то, как измерено опытом на исключение трипана синего, только BDNF и NGF проявили значительную защитную активность против MPP+-токсичности (табл. IX и X).
Таблица IX
Защита клеток SH-SY5V от MPP+-токсичности посредством BDNF и NGF
Результаты: - Количество жизнеспособных
Обработка - клеток (% жизнеспособных)
мнимое
трансфецирование
(COS-Mock) - 0,4 • 104 (5%)
BDNF-COS
(1:5) - 1,2 • 105 (67%)
NGF-COS (1:5) - 1,7 • 105 (84%)
CNTF - 0,6 • 104 (14%)
NT - 3 - 0,7 • 104
(20%)
bFGF - 0,4 • 104 (11%)
EGF - 0,1 • 104 (3%)
Таблица X
Влияние концентрации BDNF и NGF на MPP+-токсичность
Обработка - % жизнеспособных клеток
COS-MOCK (1:2) - 13
(1:5) - 5
(1:10) - 3
(1:20) - 6
(1:50) - 7
(1:100) - 2
COS-BDNF (1:2)
- 73
(1:5) - 67
(1:10) - 65
(1:20) - 42
(1:50) - 30
(1:100) - 25
COS-NGF (1:2) - 88
(1:5) - 84
(1:10) - 63
(1:20)
- 50
(1:50) - 47
(1:100) - 34
В культурах вентрального среднего мозга данные показывают, что
BDNF проявляет значительное защитное действие против MPP+-токсичности,
тогда как bFGF проявляет меньший эффект (фиг.38). Было обнаружено, что обработка MPP+ снижает количество
положительных на тирозин-гидроксилазу нейронов на 85% в контрольных культурах и
только на 40% в культурах, предварительно обработанных BDNF (перед экспозицией MPP+). Предполагается, что
MPP+ является активным метаболитом для токсина MPTP, который, как
наблюдалось, вызывает паркинсоноподобный синдром ин виво, что является принятой модельной системой для болезни Паркbнсона.
Таким образом, защитное действие BDNF и NT-3 указывает, что эти соединения
могут оказаться полезными при лечении болезни Паркинсона или для предотвращения неврологического повреждения в результате
воздействия токсина.
21 Депонирование микроорганизмов.
Следующие рекомбинантные бактериофаг и плазмидная ДНК были депонированы 30 августа 1989 в американской
коллекции типов культур, 12301 Parklain Drive, Rockville, Maryland 20852:
phBDNF -C-1 - 40648
λ hBDNF -G-1 - 40649
Настоящее изобретение не ограничивается
приведенными в описании специфическими выполнениями. В действительности, различные
усовершенствования изобретения в дополнение к описанным выполнениям будут очевидны для специалиста из предыдущего
описания и сопровождающих фигур. Такие усовершенствования входят в объем прилагаемых
пунктов формулы изобретения.
Различные цитированные публикации включены в описание путем ссылки во всей их полноте.
Реферат
Изобретение относится к биотехнологии, генной инженерии. Получен белок BDNF с нейротропной активностью путем культивирования клеток млекопитающих, в частности Cos-клеток, трансформированных вектором экспрессии PCMYI-PBDNF, вектор содержит фрагмент ДНК, кодирующий указанный белок. Определена нуклеотидная последовательность фрагмента ДНК человека, крысы, свиньи и цыпленка. Белок BDNF имеет молекулярную массу 12 кД, изоэлектрическую точку 9,9 и способен поддерживать выживание нейронов ганглиев орального корня, допаминергических и холинергических нейронов центральной нервной системы и ингибировать дифференциацию астроглиальных клеток. Белок BDNF может быть использован для диагностики и лечения различных неврологических нарушений. 3 с. и 3 з.п. ф-лы, 10 табл., 37 ил.
Формула
His Ser Asp Pro Ala Arg Arg Gly Glu Leu Ser Val Cys Asp Ser Ile Ser Glu Trp Val Thr Ala Ala Asp Lys Lys Thr Ala Val Asp Met Ser Gly Gly Thr Val Thr Val Leu Glu Lys Val Pro Val Ser Lys Gly Gln Leu Lys Gln Tyr Phe Tyr Glu Thr Lys Cys Asn Pro Met Gly Tyr Thr Lys Glu Gly Cys Arg Gly Ile Asp Lys Arg His Trp Asn Ser Gln Cys Arg Thr Thr Gln Ser Tyr Val Arg Ala Leu Thr Met Asp Ser Lys Lys Arg Ile Gly Trp Arg Phe Ile Arg Ile Asp Thr Ser Cys Val Cys Thr Leu Thr Ile Lys Arg Gly Arg,
установленный на основе кодирующей ДНК-последовательности, мол.массой 12 кД, изоэлектрической точкой 9.9 и способностью поддерживать выживание нейронов ганглиев орального корня, допаминергических и холинергических нейронов центральной нервной системы и ингибировать дифференциацию астроглиальных клеток.
Комментарии