Способы лечения боли, обусловленной раком кости, путем введения антагониста фактора роста нервов - RU2389509C2
Код документа: RU2389509C2
Чертежи














Описание
Перекрестная ссылка на родственные заявки
Настоящая заявка имеет приоритет предварительных заявок на патент США № 60/620654, поданной 19 октября 2004 г., и 60/560781, поданной 7 апреля 2004 г., которые полностью включены в данное описание изобретения в качестве ссылки.
Заявление о государственном финансировании исследований или разработок
Настоящее изобретение сделано при поддержке правительства США на основании грантов 5R37-NS23970-16, 5R01-DA11986-05 и 1R01-NS048021-01A1, предоставленных Национальными институтами здравоохранения. Правительство США обладает определенными правами на данное изобретение.
Область техники, к которой относится изобретение
Настоящее изобретение относится к применению антагониста фактора роста нервов (NGF) для профилактики, ослабления или лечения боли, обусловленной раком кости.
Уровень техники
Фактор роста нервов (NGF) был первым идентифицированным нейротрофином, поэтому его роль в развитии и сохранении жизнеспособности периферических и центральных нейронов хорошо исследована. Установлено, что NGF имеет исключительно важное значение для сохранения жизнеспособности в процессе развития периферических симпатических и эмбриональных сенсорных нейронов, а также базальных холинергических нейронов переднего мозга (Smeyne, et al. Nature 368:246-249 (1994); Crowley, et al., Cell 76:1001-1011 (1994)). NGF повышает экспрессию нейропептидов в сенсорных нейронах (Lindsay et al., Nature 337:362-364 (1989)), и его активность опосредована двумя разными мембраносвязанными рецепторами, а именно рецептором тирозинкиназы TrkА и рецептором р75, которые в структурном отношении родственны другим членам семейства рецепторов фактора некроза опухолей (Chao, et al., Science 232:518-521 (1986)).
Помимо воздействия на нервную систему, NGF участвует в процессах, не связанных с нервной системой. Например, установлено, что NGF повышает сосудистую проницаемость у крыс (Otten et al., Eur. J. Pharmacol. 106:199-201 (1984)), усиливает Т- и В-клеточные иммунные реакции (Otten, et al., Proc. Natl. Acad. Sci. USA 86: 10059-10063 (1989)), индуцирует дифференцировку лимфоцитов и пролиферацию мастоцитов (Matsuda et al., Proc. Natl. Acad. Sci. USA 85:6508-6512 (1988); Pearce et al., J. Physiol. 372:379-393 (1986); Bischoff et al., Blood 79:2662-2669 (1992); Horigome et al., J. Biol. Chem. 268:14881-14887 (1993)). Хотя известно, что экзогенно вводимый NGF может оказывать все вышеуказанные действия, необходимо отметить, что эндогенный NGF участвует в указанных процессах in vivo лишь в очень редких случаях (Torcia et al., Cell. 85(3):345-56 (1996)). Поэтому не ясно, каким может быть эффект ингибирования биологической активности эндогенного NGF, если такой эффект вообще будет иметь место.
Фактор роста нейронов продуцируют разные клетки, в том числе мастоциты (Leon et al., Proc. Natl. Acad. Sci. USA 91: 3739-3743 (1994)), B-лимфоциты (Torcia et al., Cell 85:345-356 (1996)), кератиноциты (Di Marco et al., J. Biol. Chem. 268:22838-22846)), клетки гладких мышц (Ueyama et al., J. Hypertens. 11:1061-1065 (1993)), фибробласты (Lindholm et al., Eur. J. Neurosci. 2:795-801 (1990)), эпителиальные клетки бронхов (Kassel et al., Clin. Exp. Allergy 31:1432-40 (2001)), клетки почечного мезангия (Steiner et al., Am. J. Physiol. 261:F792-798 (1991)) и мышечные трубочки скелетных мышц (Schwartz et al., J. Photochem. Photobiol. B66:195-200 (2002)). Рецепторы NGF были обнаружены на клетках разных типов, не относящихся к нервной системе. Например, рецептор TrkA обнаружен на моноцитах, Т- и В-лимфоцитах и мастоцитах человека.
Взаимосвязь между повышенными уровнями NGF и разными воспалительными заболеваниями наблюдается как у людей, так и в нескольких животных моделях. Указанные заболевания включают системную красную волчанку (Bracci-Laudiero et al., Neuroreport 4:563-565 (1993)), рассеянный склероз (Bracci-Laudiero et al., Neurosci. Lett. 147:9-12 (1992)), псориаз (Raychaudhuri et al., Acta Derm. l'enereol. 78:84-86 (1998)), артрит (Falcimi et al., Ann. Rheum. Dis. 55:745-748 (1996)), интерстициальный цистит (Okragly et al., J. Urology 161:438-441 (1991)), астму (Braun et al., Eur. J. Immunol. 28:3240-3251 (1998)), панкреатит и простатит.
Повышенное содержание NGF в периферических тканях также ассоциировано с воспалением и наблюдается в целом ряде форм артрита. В синовиальной оболочке субъектов, страдающих ревматоидным артритом, экспрессированы высокие уровни NGF, в то время как в невоспаленной синовиальной оболочке NGF не был обнаружен (Aloe et al., Arch. Rheum. 35:351-355 (1992)). Аналогичные результаты были получены у крыс, у которых экспериментально индуцировали ревматоидный артрит (Aloe et al., Clin. Exp. Rheumatol. 10:203-204 (1992); Halliday et al., Neurochem. Res. 23:919-22 (1998)). Повышенные уровни NGF были обнаружены у трансгенных мышей, страдающих артритом, наряду с увеличением числа мастоцитов (Aloe et al., Int. J. Tissue Reactions-Exp. Clin. Aspects 15:139-143 (1993)).
Лечение экзогенным NGF усиливает боль и повышает болевую чувствительность. Данное заявление можно проиллюстрировать тем, что инъекция NGF значительно усиливает боль и повышает болевую чувствительность в двух животных моделях (Lewin et al., J. Neurosci. 13:2136-2148 (1993); Amann et al., Pain 64, 323-329 (1996); Andreev et al., Pain 63, 109-115 (1995)) и у человека (Dyck et al., Neurology 48, 501-505 (1997); Petty et al., Annals Neurol. 36, 244-246 (1994)). NGF, по-видимому, воздействует в соответствии с несколькими механизмами, включая индукцию BDNF нейротрофина (Apfel et al., Mol. Cell. Neurosci. 7(2), 134-142 (1996); Michael et al., J. Neurosci. 17, 8476-8490 (1997)), который в свою очередь изменяет обработку болевого сигнала в спинном мозге (Hains et al., Neurosci. Lett. 320(3), 125-8 (2002); Miletic et al., Neurosci. Lett. 319(3), 137-40 (2002); Thompson et al., Proc. Natl. Acad. Sci. USA 96(14), 7714-8 (1999)), вызывая изменение периферических и центральных соединений сенсорных нейронов и других болепередающих нейронов в спинном мозге (Lewin et al., European Journal of Neuroscience 6, 1903-1912 (1994); Thompson et al., Pain 62, 219-231 (1995)), изменение роста аксонов (Lindsay R.M., J. Neurosci. 8(7), 2394-405 (1988)), экспрессию рецептора брадикинина (Peterson et al., Neuroscience 83:161-168 (1998)), изменение экспрессии генов, ответственных за активацию нервов и проводящие пути, такие как ионные каналы (Boettger et al., Brain 125(Pt2), 252-63 (2002); Kerr et al., Neuroreport 12(14), 3077-8 (2001); Gould et al., Brain Res. 854(1-2), 19-29 (2000); Fjell et al., J. Neurophysiol. 81:803-810 (1999), усиление ассоциированного с болью рецептора TRPV1 (Chuang et al., Nature 411 (6840), 957-62 (2001); Shu and Mendell, Neurosci. Lett. 274:159-162 (1999)) и патологические изменения в мышцах (Foster et al., J. Pathol. 197(2), 245-55 (2002)). Многие указанные изменения происходят непосредственно в болепередающих сенсорных нейронах и, по-видимому, не зависят от сопутствующего воспаления. Кроме того, известны по крайней мере два других типа клеток, реагирующих на NGF, которые могут изменять восприятие боли и болевую чувствительность. Вышеуказанные клетки первого типа, мастоцицы, реагируют на NGF утратой зернистости (Yan et al., Clin. Sci. (Lond) 80:565-569 (1991) или, как указано в других исследованиях, увеличивают продуцирование или высвобождение медиаторов совместно с другими агентами (Pearce and Thompson, J. Physiol. 372:379-393 (1986), Kawamoto et al., J. Immunol. 168:6412-6419 (2002)). Как было установлено у крыс, NGF-опосредованные болевые реакции по крайней мере частично обусловлены мастоцитами (Lewin et al., Eur. J. Neurosci. 6:1903-1912 (1994), Woolf, et al., J. Neurosci. 16:2716-2723 (1996)), хотя возможную причастность указанных клеток необходимо продемонстрировать у человека. Кроме того, известно, что первичные симпатические нейроны реагируют на NGF и также участвуют в передаче болевых сигналов (Aley et al., Neuroscience 71:1083-1090 (1996)). Ясно, что устранение иннервации симпатических нейронов изменяет гипералгезию, которая обычно возникает при лечении NGF (Woolf et al., J. Neurosci. 16:2716-2723 (1996)).
В научной литературе описано применение антагонистов NGF, таких как антитело против NGF, при лечении разных типов боли. См., например, заявки на патент США №№ 10/682331, 10/682638, 10/682332 (публикация № 2004/0131615), 10/783730 (публикация № 2004/0253244), 10/745775 (публикация № 2004/0237124), 10/791162, PCT/US03/32089 (WO 04/032870), PCT/US03/32083 (WO 2005/000194), PCT/US03/32113, PCT/US2004/05162 (WO 04/073653), PCT/US03/41252 (WO 04/058184).
Боль, обусловленная раком кости, может быть вызвана у человека первичной опухолью кости или более вероятно метастазами в кость (например, образующимися в результате рака молочной железы, предстательной железы или легкого). См. публикацию Luger et al., Pain 99:397-406 (2002). Созданная модель боли, обусловленной раком кости, у мышей отражает боль от умеренной до сильной, испытываемую человеком в случае рака кости. См. публикации Luger et al., Pain 99:397-406 (2002); Clohisy et al., Clinical Orthopaedics and Related Research 415S:S279-S288 (2003); Schwei et al., J. Neruosci. 19:10886-10897 (1999); Honore et al., Nat. Med. 6:521-529 (2000). В статьях Honore et al. и Schwei et al. указано, что нейрохимическая картина изменений, наблюдаемых в спинном мозге и DRG животных, страдающих раком кости, являются уникальными и отличаются от типичной воспалительной или нейропатической боли, хотя в биохимической картине данной модели могут присутствовать компоненты, аналогичные классической воспалительной и нейропатической боли. Honore et al., Neuroscience 98:585-598 (2000); Schwei et al., J. Neruosci. 19:10886-10897 (1999); Luger et al., Pain 99:397-406 (2002).
Все научные работы, приведенные в данной заявке, в том числе заявки на патент и публикации, полностью включены в настоящее описание изобретения в качестве ссылки.
Сущность изобретения
В основе настоящего изобретения лежит открытие того, что антагонисты NGF, такие как антитело против NGF, позволяют эффективно лечить боль, обусловленную раком кости, включая боль, ассоциированную с метастазированием рака в кость. Такое лечение направлено на устранение одного или нескольких признаков боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, которая описана в настоящей заявке на патент.
Одним объектом настоящего изобретения является способ профилактики или лечения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость (именуемую также “боль, вызываемая метастазами в кость”), путем введение антагониста фактора роста нервов (NGF). В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с нестероидным противовоспалительным средством (NSAID). В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком и NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика. В некоторых вариантах осуществления изобретения антагонист NGF вводят без NSAID.
Другим объектом настоящего изобретения являются способы уменьшения вероятности возникновения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, ослабление боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, временное облегчение боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, и/или замедление появления или развития боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, которые включают введение эффективного количества антагониста NGF. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком и NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика. В некоторых вариантах осуществления изобретения антагонист NGF вводят без NSAID.
В некоторых вариантах осуществления изобретения боль, обусловленная раком кости, возникает вследствие образования в кости раковой опухоли. В некоторых вариантах осуществления изобретения боль, обусловленная раком кости, возникает вследствие остеосаркомы. В некоторых вариантах осуществления изобретения боль, обусловленная раком кости, возникает вследствие метастазирования рака в кость. В некоторых вариантах осуществления изобретения метастазы в кость являются следствием метастазирования в кость рака предстательной железы. В некоторых вариантах осуществления изобретения метастазы в кость являются следствием метастазирования в кость рака молочной железы. В некоторых вариантах осуществления изобретения метастазы в кость являются следствием метастазирования в кость рака легкого. В некоторых вариантах осуществления изобретения метастазы в кость являются следствием метастазирования в кость саркомы. В некоторых вариантах осуществления изобретения метастазы в кость являются следствием метастазирования в кость рака почки. В некоторых вариантах осуществления изобретения метастазы в кость являются следствием метастазирования в кость множественной миеломы. В некоторых вариантах осуществления изобретения подлежащая лечению боль, обусловленная раком, находится в пределах от слабой до умеренной. В некоторых вариантах осуществления изобретения подлежащая лечению боль, обусловленная раком, находится в пределах от умеренной до сильной. В некоторых вариантах осуществления изобретения подлежащая лечению боль, обусловленная раком, является сильной.
Антагонистом NGF, пригодным для применения в способах по настоящему изобретению, является любой агент, который может прямо или косвенно уменьшать биологическую активность NGF. В некоторых вариантах осуществления изобретения антагонист NGF (например, антитело) связывается (физически взаимодействует) с NGF, связывается с рецептором NGF (таким как рецептор trkA и/или р75) и/или ослабляет (препятствует и/или блокирует) передачу сигналов рецептора NGF в нижней области (например, ингибиторы передачи сигнала киназы). Таким образом, в некоторых вариантах осуществления изобретения антагонист NGF связывается (физически взаимодействует) с NGF. В другом варианте осуществления изобретения антагонист NGF связывается с рецептором NGF (таким как рецептор TrkA и/или р75). В других вариантах осуществления изобретения антагонист NGF ослабляет (препятствует и/или блокирует) передачу сигналов рецептора NGF в нижней области (например, ингибиторы передачи сигнала киназы). В других вариантах осуществления изобретения антагонист NGF ингибирует (уменьшает) синтез и/или высвобождение NGF. В другом варианте осуществления изобретения антагонист NGF является иммуноадгезином TrkA. В некоторых вариантах осуществления изобретения антагонист NGF связывается с NGF (таким как NGF человека) и по существу не связывается с родственными нейротрофинами, такими как NT-3, NT-4/5 и/или BDNF. В некоторых вариантах осуществления изобретения антагонист NGF выбирают из одного или нескольких нижеследующих агентов: антитело против NGF, антисмысловая молекула к NGF (включая антисмысловую молекулу к нуклеиновой кислоте, кодирующей NGF), антисмысловая молекула к рецептору NGF (такому как trkA и/или р75) (включая антисмысловую молекулу к нуклеиновой кислоте, кодирующей рецептор NGF), ингибитор NGF, структурный аналог NGF, доминантноотрицательная мутация рецептора TrkA и/или р75, связывающего NGF, антитело против TrkA, антитело против р75 и ингибитор киназы. В другом варианте осуществления изобретения антагонистом NGF является антитело против NGF. В других вариантах осуществления изобретения антитело против NGF является гуманизированным антителом (таким как антитело Е3, описанное в настоящем описании изобретения). В некоторых вариантах осуществления изобретения антителом против NGF является антитело Е3 (описанное в настоящем описании изобретения). В других вариантах осуществления изобретения антитело против NGF содержит один или несколько гипервариабельных участков (комплементоопределяющая область, CDR) антитела Е3 (в частности, один, два, три, четыре, пять или в некоторых вариантах осуществления изобретения все шесть гипервариабельных участков антитела Е3). В других вариантах осуществления изобретения антитело является антителом человека. В некоторых вариантах осуществления изобретения данное антитело содержит три CDR из тяжелой цепи антитела Е3. В некоторых вариантах осуществления изобретения данное антитело содержит три CDR из легкой цепи антитела Е3. В других вариантах осуществления изобретения антитело против NGF содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в таблице 1 (SEQ ID NO:1). В других вариантах осуществления изобретения антитело против NGF содержит аминокислотную последовательность вариабельной области легкой цепи, показанную в таблице 2 (SEQ ID NO:2). В других вариантах осуществления изобретения антитело против NGF содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в таблице 1 (SEQ ID NO:1), и аминокислотную последовательность вариабельной области легкой цепи, показанную в таблице 2 (SEQ ID NO:2). В других вариантах осуществления изобретения антитело содержит модифицированную константную область, в частности иммунологически инертную константную область, которая, например, не запускает комплемент-опосредованный лизис или не стимулирует антителозависимую клеточно-опосредованную цитотоксичность (ADCC). В других вариантах осуществления изобретения константная область модифицирована в соответствии с описанием, приведенным в публикации Eur. J. Immunol. (1999) 29:2613-2624, в заявке РСТ № РСТ/GB99/01441 и/или в заявке на патент Великобритании № 9809951.8.
В некоторых вариантах осуществления изобретения антагонист NGF связывается с NGF. В других вариантах осуществления изобретения антагонистом NGF является антитело, которое специфически связывается с NGF (таким как NGF человека). В других вариантах осуществления изобретения указанное антитело по существу связывается с тем же эпитопом 6 NGF, что и антитело, выбираемое из одного или нескольких нижеследующих моноклональных антител мыши: Mab 911, MAb 912 и MAb 938 (см. публикацию Hongo, et al., Hybridoma 19:215-227 (2000)). В некоторых вариантах осуществления изобретения антагонист NGF связывается с рецептором trkA. Антагонистом NGF может быть моноклональное антитело против NGF человека (против hNGF), которое способно связываться с hNGF и эффективно ингибировать связывание hNGF с TrkA человека (hTrkA) и/или эффективно ингибировать активацию рецептора TrkA человека.
Сродство связывания антитела против NGF с NGF (таким как hNGF) может составлять около 0,10-1,0 нМ, около 0,10-0,80 нМ, около 0,15-0,75 нМ и около 0,18-0,72 нМ. В одном варианте осуществления изобретения сродство связывания составляет примерно 2-22 пМ. В одном варианте осуществления изобретения сродство связывания равно примерно 10 нМ. В других вариантах осуществления изобретения сродство связывания составляет менее примерно 10 нМ. В других вариантах осуществления изобретения сродство связывания равно примерно 0,1 нМ или примерно 0,07 нМ. В других вариантах осуществления изобретения сродство связывания составляет менее примерно 0,1 нМ или менее примерно 0,07 нМ. В других вариантах осуществления изобретения сродство связывания составляет около 100 нМ, около 50 нМ, около 10 нМ, около 1 нМ, около 500 пМ, около 100 пМ, от около 50 пМ до около 2 пМ, около 5 пМ, около 10 пМ, около 15 пМ, около 20 пМ или около 40 пМ. В некоторых вариантах осуществления изобретения сродство связывания составляет около 100 нМ, около 50 нМ, около 10 нМ, около 1 нМ, около 500 пМ, около 100 пМ, около 50 пМ или менее примерно 50 пМ. В некоторых вариантах осуществления изобретения сродство связывания составляет около 100 нМ, около 50 нМ, около 10 нМ, около 1 нМ, около 500 пМ, около 100 пМ или около 50 пМ. В других вариантах осуществления изобретения сродство связывания составляет около 2 пМ, около 5 пМ, около 10 пМ, около 15 пМ, около 20 пМ, около 40 пМ или более примерно 40 пМ. Как хорошо известно в данной области, сродство связывания может быть выражено в виде КD или константы диссоциации, и более высокое сродство связывания соответствует меньшему значению КD. Сродство связывания моноклонального антитела мыши против NGF 911 (Hongo et al., Hybridoma 19:215-227 (2000)) с NGF человека составляет около 10 нМ, и сродство связывания гуманизированного антитела Е3 против NGF (описанного в настоящем описании изобретения) с NGF человека составляет около 0,07 нМ. Значения сродства связывания для антитела 911 и Е3 измеряли, используя их Fab-фрагменты.
Антагонист NGF можно вводить до, во время и/или после диагностирования у субъекта рака кости или метастазирования рака в кость. Антагонист NGF можно вводить любыми способами, известными в данной области, которые включают пероральное, внутривенное, подкожное, внутриартериальное, внутримышечное, интракардиальное, интраспинальное, внутригрудное, внутрибрюшинное, внутрижелудочковое, подъязычное и/или трансдермальное введение. В некоторых вариантах осуществления изобретения антагонистом NGF является антитело против NGF, которое вводят одним или несколькими нижеследующими способами: внутривенно, подкожно, путем ингаляции, внутриартериально, внутримышечно, интракардиально, внутрижелудочково и внутрибрюшинно. Введение может быть системным, например внутривенным, или местным.
В некоторых вариантах осуществления изобретения антагонист NGF вводят в дозе около 0,1-10 мг/кг массы тела и в других вариантах осуществления изобретения антагонист NGF вводят в дозе около 0,3-20 мг/кг массы тела.
Другим объектом настоящего изобретения является композиция для лечения и/или профилактики боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, которая включает эффективное количество антагониста фактора роста нервов (NGF) в комбинации с одним или несколькими фармацевтически приемлемыми эксципиентами. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика или NSAID. В некоторых вариантах осуществления изобретения антагонистом NGF является антитело, которое специфически связывается с молекулой NGF. В других вариантах осуществления изобретения антагонистом NGF является любой антагонист, описанный в настоящем описании изобретения.
Другим объектом настоящего изобретения является набор, предназначенный для использования в любых способах, представленных в данном описании изобретения. В некоторых вариантах осуществления изобретения указанный набор включает любые антагонисты NGF, описанные в настоящем описании изобретения, в комбинации с фармацевтически приемлемым носителем. В других вариантах осуществления изобретения указанный набор дополнительно включает инструкции по использованию антагониста NGF в любых способах, описанных в настоящем описании изобретения.
Краткое описание чертежей
На Фиг.1 изображен график постоянной боли, которую определяли путем измерения спонтанной защитной фиксации и спонтанного подрагивания в течение двухминутного периода наблюдения на 10-й и 14-й день после инъекции саркомы. Определение “незараженный” (Naive) относится к животным, которым не делали никаких инъекций. “Псевдоагент + носитель” (Sham + veh.) относится к животным, которым сначала инъецировали α-минимальную эссенциальную среду в костный мозг бедренной кости и затем инъецировали физиологический раствор. “Саркома + носитель” (Sarc + veh.) относится к животным, которым сначала инъецировали саркому в костный мозг бедренной кости и затем инъецировали физиологический раствор. “Саркома + антитело против NGF” (Sarc + Anti-NGF) относится к животным, которым сначала инъецировали саркому в костный мозг бедренной кости и затем инъецировали антитело против NGF 911.
На Фиг.2 изображен график боли, возникающей в конечности при ходьбе, которую определяли по пользованию конечностью и защитной фиксации при принудительной ходьбе («беличье колесо», ротарод) на 10-й и 14-й день после инъекции саркомы. Определение “незараженный” (Naive) относится к животным, которым не делали никаких инъекций. “Псевдоагент + носитель” (Sham + veh.) относится к животным, которым сначала инъецировали α-минимальную эссенциальную среду в костный мозг бедренной кости и затем инъецировали физиологический раствор. “Саркома + носитель” (Sarc + veh.) относится к животным, которым сначала инъецировали саркому в костный мозг бедренной кости и затем инъецировали физиологический раствор. “Саркома + антитело против NGF” (Sarc + Anti-NGF) относится к животным, которым сначала инъецировали саркому в костный мозг бедренной кости и затем инъецировали антитело против NGF 911.
На Фиг.3 изображен график боли, вызываемой прикосновением, которую определяли по индуцированной пальпацией защитной фиксации и индуцированному пальпацией подрагиванию в течение двухминутного периода наблюдения на 10-й и 14-й день после инъекции саркомы. Определение “незараженный” (Naive) относится к животным, которым не делали никаких инъекций. “Псевдоагент + носитель” (Sham + veh.) относится к животным, которым сначала инъецировали α-минимальную эссенциальную среду в костный мозг бедренной кости и затем инъецировали физиологический раствор. “Саркома + носитель” (Sarc + veh.) относится к животным, которым сначала инъецировали саркому в костный мозг бедренной кости и затем инъецировали физиологический раствор. “Саркома + антитело против NGF” (Sarc + Anti-NGF) относится к животным, которым сначала инъецировали саркому в костный мозг бедренной кости и затем инъецировали антитело против NGF 911.
На Фиг.4 представлены фотографии, показывающие, что антитело против NGF не влияет на развитие болезни в кости на 14-й день (d14) после инъекции опухоли. Животные (n=8), которым вводили псевдоагент и носитель, показаны на фотографиях (а) и (d); животные (n=13), которым инъецировали саркому (GFP-трансфицированные) и носитель, показаны на фотографиях (b) и (е); и животные (n=8), которым инъецировали саркому (GFP-трансфицированные) и антитело против NGF, показаны на фотографиях (с) и (f). На Фиг.4а, 4b и 4с представлены рентгенограммы, показывающие наличие или отсутствие разрушения кости. На Фиг.4d, 4е и 4f представлены фотографии, на которых показано иммунное окрашивание антителом против GFP. Масштаб: шкала 1 мм.
На Фиг.5 представлены фотографии, показывающие, что лечение антителом против NGF не оказывает заметного воздействия на иннервацию сенсорных нейронов в коже. Образцы кожи задней лапы мышей, которым инъецировали саркому, (а, b) и незараженных мышей (с, d) подвергали иммунному окрашиванию пептидом, родственным гену кальцитонина нейропептида (CGRP), который метит немиелинизированные пептидергические сенсорные нервные волокна. На данной фигуре показано иммунное окрашивание CGRP образцов кожи задней лапы мышей (а, n=3), которым инъецировали саркому и носитель, мышей (b, n=8), которым инъецировали саркому и антитело против NGF, незараженных мышей, которым вводили носитель, (с, n=8) и незараженных мышей (d, n=8), которым вводили антитело против NGF. Масштаб: шкала 50 мкм.
На Фиг.6 изображены графики, показывающие, что введение антитела против NGF ослабляет боль, обусловленную раком кости. Время защитной фиксации и число спонтанных подрагиваний конечности, в которую инъецировали саркому, в течение двухминутного периода наблюдения использовали в качестве единицы измерения постоянной боли на 8-й, 10-й, 12-й и 14-й дни после инъекции и удерживания раковых клеток в левой бедренной кости (а, b). Показатели боли, возникающей при движении, включали количественное определение времени защитной фиксации и числа подрагиваний в течение двухминутного периода наблюдения с последующей обычно безболезненной пальпацией бедренной кости, в которую инъецировали саркому (с, d). “#” означает Р<0,05 по сравнению с введением псевдоагента и носителя, и “*” означает Р<0,05 по сравнению с введением саркомы и носителя.
На Фиг.7 изображены графики, показывающие, что введение антитела против NGF не влияет на исходный порог тепловой или механической чувствительности и является более эффективным, чем введение морфина (MS), для ослабления боли, обусловленной раком кости. На Фиг.7а и 7b показана тепловая чувствительность (а, n=8 для незараженных животных, которым вводили носитель, n=8 для незараженных животных, которым вводили антитело против NGF), измеренная в виде латентного периода отдергивания лапы под воздействием теплового раздражителя, и механическая чувствительность (b, n=8 для незараженных животных, которым вводили носитель, n=8 для незараженных животных, которым вводили антитело против NGF), измеренная в виде 50% порога механического раздражения при введении антитела против NGF (10 мг/кг, внутрибрюшинно, каждые 5 дней) у незараженных мышей. На Фиг.7с и 7d показана постоянная боль, которую определяли путем измерения спонтанной защитной фиксации (с) в течение двухминутного периода наблюдения, и боль, возникающая при движении, которую определяли путем измерения времени защитной фиксации (d) в течение двухминутного периода наблюдения с последующей обычно безболезненной пальпацией дистального отдела бедренной кости. На данной фигуре показаны значения спонтанной защитной фиксации (с) и защитной фиксации, вызванной пальпацией, (d) для незараженных мышей, псевдозараженных мышей, которым вводили носитель, мышей, которым инъецировали саркому и вводили носитель, мышей, которым инъецировали саркому и вводили морфин (n=8, внутрибрюшинно, 10 мг/кг за 15 минут до испытания), мышей, которым инъецировали саркому и вводили морфин (n=8, внутрибрюшинно, 30 мг/кг за 15 минут до испытания), и мышей, которым инъецировали саркому и антитело против NGF (n=8, внутрибрюшинно, 10 мг/кг каждые 5 дней с 6-го по 14-й день после инъекции опухоли). Планки погрешностей показывают стандартную ошибку средней (S.E.M.). “#”означает Р<0,05 по сравнению с введением псевдоагента и носителя (n=8); “*” означает Р<0,05 по сравнению с введением саркомы и носителя; “+” означает Р<0,05 по сравнению с введением саркомы и морфина.
На Фиг.8 представлены фотографии, показывающие, что введение антагонистического антитела против NGF позволяет уменьшить нейрохимические изменения и инфильтрацию макрофага в ганглий задних корешков (DRG) спинного мозга пораженных опухолью животных. На Фиг.8а и 8b показано иммунофлуоресцентное окрашивание фактора активации транскрипции 3 (ATF-3) в ипсилатеральном DRG L2 пораженных опухолью животных, которым вводили носитель (а, n=8) и антитело против NGF (b, n=8), через четырнадцать дней после имплантации опухоли. В нижнем блоке показано иммунофлуоресцентное окрашивание CD-68, характеризующее плотность активированных и инфильтрующихся макрофагов вокруг пораженных сенсорных нейронов в ипсилатеральном DRG пораженных опухолью животных, которым вводили носитель (с, n=7) и антитело против NGF (d, n=7). Масштаб: шкала для а-d = 5 мкм.
На Фиг.9 представлены микрофотографии, показывающие уменьшение нейрохимических изменений, ассоциированных с сенсибилизацией центральных нейронов, в результате введения антитела против NGF. На Фиг.9А и 9В показано иммуное окрашивание динорфина в заднем роге спинного мозга мышей, которым инъецировали саркому и вводили носитель (А, n=9), и мышей, которым инъецировали саркому и вводили антитело против NGF (B, n=4). На Фиг.9С и 9D показаны типичные конфокальные изображения с-Fos-экспрессирующих нейронов спинного мозга у мышей, которым инъецировали саркому и вводили носитель, (С, n=4) и у мышей, которым инъецировали саркому и вводили антитело против NGF, (D, n=4) с последующим выполнением обычно безболезненной пальпации конечностей, пораженных опухолью. Масштаб: шкала 150 мкм для А и В; 200 мкм для С и D.
На Фиг.10 изображены графики, показывающие, что лечение антителом против NGF ослабляет боль, обусловленную раком кости вследствие поражения опухолью предстательной железы. Лечение антителом против NGF (внутрибрюшинно, 10 мг/кг на 7-й, 12-й и 17-й день после инъекции опухоли) ослабляло постоянную боль, обусловленную раком кости, начиная с 7-го дня после инъекции опухоли на протяжении всего развития болезни. Время защитной фиксации и число спонтанных подрагиваний бедренной кости, в которую инъецировали АСЕ-1, в течение двухминутного периода наблюдения использовали в качестве единиц измерения постоянной боли (А, В). Антитело против NGF (закрашенный квадрат) значительно уменьшало постоянную боль у животных, которым инъецировали опухоль, по сравнению с животными, которым вводили АСЕ-1 и носитель (незакрашенный квадрат), причем все показатели снижались почти до уровней введения псевдоагента на 9-й день (кружок). Как защитная фиксация, так и подрагивание у животных, которым вводили псевдоагент + носитель, значительно отличались от животных, которым вводили АСЕ-1 + носитель, на протяжении всего развития болезни. Лечение антителом против NGF не влияло на основные тепловые или механические реакции при измерении в виде латентного периода отдергивания лапы под воздействием теплового раздражителя или увеличения порога механического раздражения (С, D). Лечение антителом против NGF вызывало более сильное ослабление постоянной боли на 19-й день, чем при введении 10 мг/кг или 30 мг/кг морфина (внутрибрюшинно, за 15 минут до испытания) (Е, F). Боль, вызываемую движением, измеряли путем количественного определения времени защитной фиксации и числа подрагиваний в течение двухминутного периода наблюдения с последующей обычно безболезненной пальпацией бедренной кости, в которую инъецировали АСЕ-1 (G, H). Планки погрешностей показывают стандартную ошибку средней (S.E.M.). На Фиг.10А-F “#” означает Р<0,05 по сравнению с введением псевдоагента и носителя (n=8); “*” означает Р<0,05 по сравнению с введением АСЕ-1 и носителя; “+” означает Р<0,05 по сравнению с введением АСЕ-1 и морфина. На Фиг.10G и 10Н “*” означает Р<0,01 по сравнению с введением псевдоагента и “#” означает Р<0,01 по сравнению с введением АСЕ-1 и носителя.
На Фиг.11 представлены фотографии, показывающие, что лечение антителом против NGF не влияет на опухолевую массу или вызванное опухолью изменение формы кости. У псевдозараженных животных, которым вводили носитель, (А) рентгенографическими или гистологическими методами (H&E) не было обнаружено (D) разрушения кости на 19-й день, в то время как у животных, которым вводили АСЕ-1 и носитель, (В, Е) и у животных, которым вводили АСЕ-1 и антитело против NGF, (C, F) наблюдался значительный рост опухоли и изменение формы кости при рентгенографическом и гистологическом исследовании. Н = гемопоэтические клетки; Т = опухоль; WB = АСЕ-1-индуцированное образование кости; масштаб: шкала = 1,5 мм.
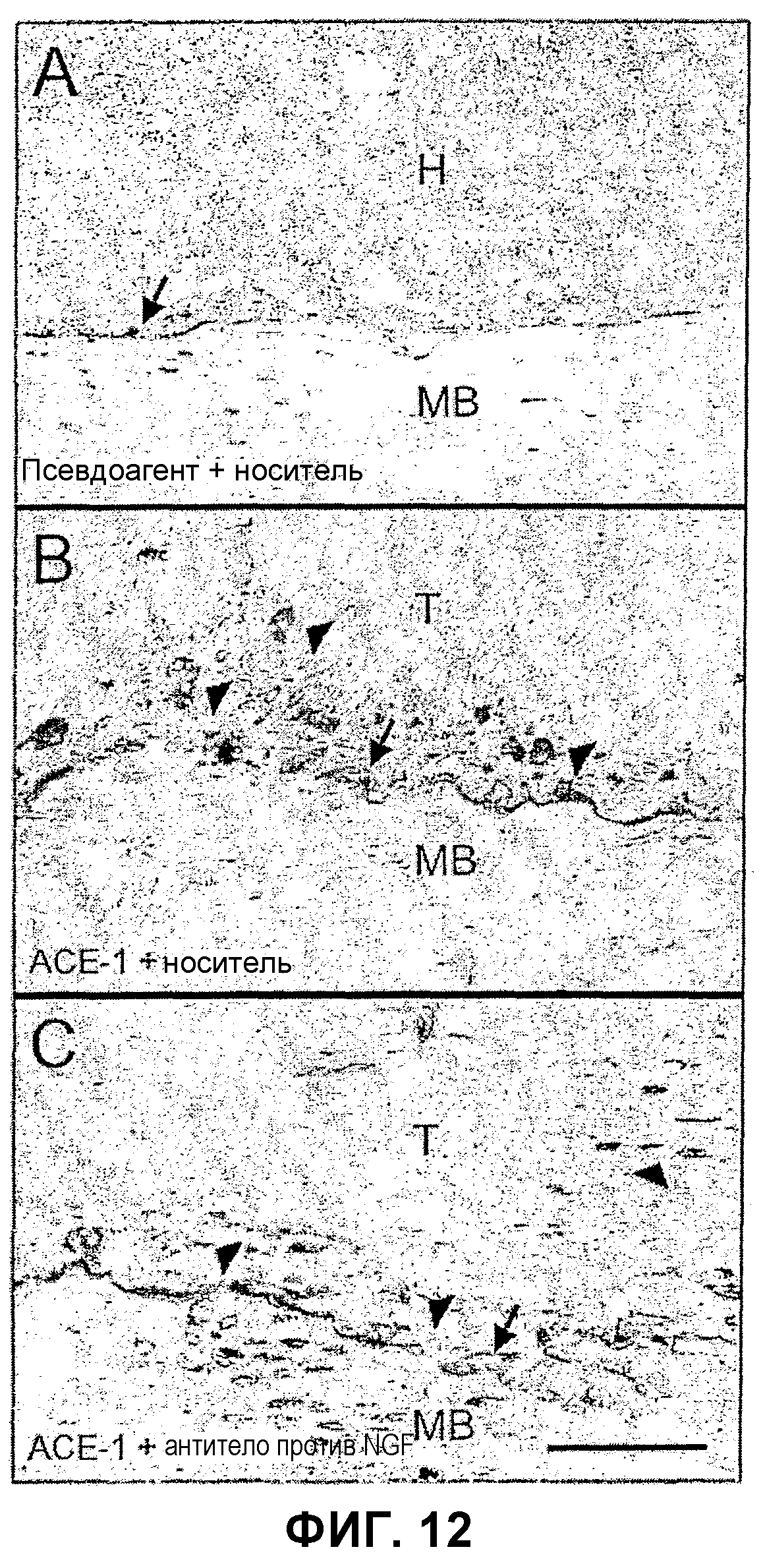
На Фиг.12 представлены изображения, показывающие, что лечение антителом против NGF не вызывает значительного уменьшения индуцированного опухолью остеокластогенеза. TRAP-окрашенные изображения образцов тканей мышей, которым вводили псевдоагент + носитель (А), АСЕ-1 + носитель (В) и АСЕ-1 + антитело против NGF (С), показывают, что пролиферация клеток в данной модели происходит в областях изменения формы кости, индуцированного опухолью, с увеличением числа остеокластов на одном мм2 площади диафизарной интрамедуллярной области у животных, которым вводили антитело против NGF, и у животных, которым вводили носитель, по сравнению с животными, которым вводили псевдоагент и носитель, и незараженными животными, которым вводили носитель. Не наблюдалось каких-либо видимых изменений в гистологических проявлениях остеокластов на поверхности раздела между опухолью и костью или макрофагов в опухоли при сравнении животных, которым вводили антитело против NGF, (С) с животными, которым вводили носитель, (В). У животных, которым вводили псевдоагент и носитель, (А) количество остеокластов, морфология и макрофаги существенно не отличались от незараженных животных. Стрелки = остеокласты; острия стрелок = макрофаги; МВ = минерализованная кость; Н = гемопоэтические клетки; Т = опухоль; масштаб: шкала 50 мкм.
На Фиг.13 представлены фотографии, показывающие, что лечение антителом против NGF не влияет на плотность сенсорных волокон, обладающих иммунореактивностью в отношении пептида, родственного гену кальцитонина (CGRP-IR), в бедренной кости. Не было обнаружено заметных различий в уровнях иммунофлуоресценции или плотности CGRP-IR волокон у животных, которым вводили АСЕ-1 + носитель (А), и у животных, которым вводили АСЕ-1 + антитело против NGF (B). Кроме того, следует отметить, что CGRP-IR волокна сохранялись при лечении антителом против NGF. Т = опухоль; масштаб: шкала 50 мкм.
На Фиг.14 представлены фотографии, показывающие, что лечение антителом против NGF не влияет на плотность сенсорных волокон, обладающих иммунореактивностью в отношении пептида, родственного гену кальцитонина (CGRP-IR), в коже задней лапы. Не было обнаружено заметных различий в уровнях иммунофлуоресценции или плотности CGRP-IR волокон в коже незараженных мышей, которым вводили носитель (А), и незараженных мышей, которым вводили антитело против NGF (B). Аналогичным образом не было обнаружено различий в уровнях иммунофлуоресценции или плотности CGRP-IR нервных волокон у животных, которым вводили АСЕ-1 + носитель (С), и у животных, которым вводили АСЕ-1 + антитело против NGF (D). Кроме того, следует отметить, что не было обнаружено различий в CGRP-IR нервных волокнах у незараженных мышей и мышей, которым инъецировали АСЕ-1 (А, В по сравнению с С, D). Масштаб: шкала 50 мкм.
Подробное описание изобретения
В основе настоящего изобретения лежит открытие того, что терапевтически эффективное количество антагониста NGF, такого как моноклональное антитело против NGF, можно вводить in vivo для лечения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость. Настоящее изобретение сделано в результате исследования модели рака кости у мышей, которое позволило установить, что введение антагонистического антитела против NGF эффективно ослабляет постоянную и возникающую при движении боль, обусловленную раком кости.
Настоящее изобретение относится к способам профилактики или лечения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, у субъекта (являющегося человеком и отличного от человека) путем введения эффективного количества антагониста NGF, такого как антитело против NGF, например моноклональное антитело против NGF человека (против hNGF). В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с нестероидным противовоспалительным средством (NSAID). В некоторых вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика. В некоторых вариантах осуществления изобретения антагонист NGF вводят без NSAID.
Другим объектом настоящего изобретения являются способы ослабления, замедления развития и/или профилактики возникновения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, которые включают введение субъекту эффективного количества антагониста NGF. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика. В некоторых вариантах осуществления изобретения антагонист NGF вводит без NSAID.
Настоящее изобретение относится также к композициям и наборам для лечения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, которые включают антагонист NGF, такой как антитело против NGF, например моноклональное антитело против NGF, предназначенное для использования в любых способах по настоящему изобретению. В некоторых вариантах осуществления изобретения антитело против NGF способно эффективно ингибировать связывание NGF с рецепторами TrkA и/или р75 и/или эффективно ингибировать активацию рецепторов TrkA и/или р75 фактором роста нервов (NGF).
Общие методы
При осуществлении настоящего изобретения за исключением особо оговоренных случаев использованы обычные методы молекулярной биологии (включая методы рекомбинантных ДНК), микробиологии, биологии клетки, биохимии и иммунологии, которые известны в данной области. Такие методы всесторонне описаны в научной литературе, а именно в нижеследующих публикациях Molecular Cloning: A Laboratory Manual, second edition (Sambrook, et al.; 1989) Cold Spring Harbor Press; Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I. Freshney, ed., 1987); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., 1993-8) J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); Current Protocols in Molecular Biology (F.M. Ausubel, et al., eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis, et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et.al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J.D. Capra, eds., Harwood Academic Publishers, 1995).
Определения терминов
Термин “антитело” (взаимозаменяемо используется в форме множественного числа) означает молекулу иммуноглобулина, способную специфически связываться с мишенью, такой как углевод, полинуклеотид, липид, полипептид и т.д., с помощью по крайней мере одного сайта распознавания антигена, расположенного в вариабельной области молекулы иммуноглобулина. В используемом здесь значении в определение данного термина входят не только интактные поликлональные и моноклональные антитела, но и их фрагменты (такие как Fab, Fab', F(ab')2, Fv), одноцепочечные варианты (ScFv), мутанты, слитые белки, включающие антитело, гуманизированные антитела, химерные антитела, двойные антитела, линейные антитела, одноцепочечные антитела, мультиспецифические антитела (например, биспецифические антитела) и любые другие модифицированные конфигурации молекулы иммуноглобулина, которые включают сайт распознавания антигена требуемой специфичности. Указанное антитело включает антитело любого класса, такое как IgG, IgA или IgM (или их подклассы), и антитело, не относящееся к какому-либо определенному классу. В зависимости от аминокислотной последовательности константной области тяжелой цепи антитела иммуноглобулина могут быть распределены в разные классы. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, причем некоторые из указанных классов могут быть подразделены на подклассы (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные области тяжелой цепи, соответствующие разным классам иммуноглобулинов, соответственно именуются альфа, дельта, эпсилон, гамма и мю. Структуры субъединиц и трехмерные конфигурации иммуноглобулинов разных классов хорошо известны.
Термин “моноклональное антитело” означает популяцию гомогенных антител, в которой моноклональное антитело состоит из аминокислот (естественных и синтетических), участвующих в селективном связывании антигена. Популяция моноклональных антител является в высшей степени специфичной, направленно воздействуя на один сайт антигена. В определение термина “моноклональное антитело” входят не только интактные моноклональные антитела и непроцессированные моноклональные антитела, но и их фрагменты (такие как Fab, Fab', F(ab')2, Fv), одноцепочечные варианты (ScFv), мутанты, слитые белки, включающие антитело, гуманизированные моноклональные антитела, химерные моноклональные антитела и любые другие модифицированные конфигурации молекулы иммуноглобулина, которые включают сайт распознавания антигена требуемой специфичности и обладают способностью связываться с антигеном. Моноклональное антитело не имеет ограничений, относящихся к источнику антитела или способу его получения (например, с помощью гибридомы, селекции фага, рекомбинантной экспрессии, трансгенных животных и т.д.).
В используемом здесь значении термин “фактор роста нервов” и “NGF” означает фактор роста нервов и его варианты, которые сохраняют по крайней мере часть активности NGF. В используемом здесь значении NGF включает нативную последовательность NGF всех видов млекопитающих, включая человека, собак, кошек, лошадей или коров.
Термин “рецептор NGF” означает полипептид, который связывается или активируется фактором роста нервов. Рецепторы NGF включают рецептор TrkA и рецептор р75 любых видов млекопитающих, которые включают, не ограничиваясь ими, человека, собак, кошек, лошадей, приматов или коров.
Термин “антагонист NGF” означает любую молекулу, которая блокирует, подавляет или ослабляет (в том числе значительно) биологическую активность NGF, включая пути передачи сигналов в нижней области, опосредуемые NGF, такие как связывание с рецептором и/или индукция реакции клетки на NGF. Термин “антагонист” не предполагает специфического механизма биологического воздействия и включает все возможные прямые или косвенные фармакологические, физиологические и биохимические взаимодействия с NGF или взаимодействие с NGF, его рецептором или при помощи другого механизма, а также все последствия, которые могут быть достигнуты при использовании разных и химически разнообразных композиций. Типичные антагонисты NGF включают, не ограничиваясь ими, антитело против NGF, антисмысловую молекулу к NGF (включая антисмысловую молекулу к нуклеиновой кислоте, кодирующей NGF), ингибитор NGF, структурный аналог NGF, доминантноотрицательную мутацию рецептора TrkA, который связывает NGF, иммуноадгезин TrkA, антитело против TrkA, антитело против р75 и ингибитор киназы. В соответствии с целями настоящего изобретения должно быть хорошо понятно, что в определение термина “антагонист” входят все ранее известные термины, названия, функциональные состояния и характеристики, которые предполагают по существу устранение, ослабление или нейтрализацию в любой значимой степени самого NGF, биологической активности NGF (включая его способность опосредовать любой признак боли, ассоциированной с метастазированием рака в кость) или последствий такой биологической активности. В некоторых вариантах осуществления изобретения антагонист NGF (например, антитело) связывается (физически взаимодействует) с NGF, связывается с рецептором NGF (таким как рецептор trkA и/или рецептор р75), ослабляет (препятствует и/или блокирует) передачу сигналов рецептора NGF в нижней области и/или ингибирует (уменьшает) синтез, продуцирование или высвобождение NGF. В других вариантах осуществления изобретения антагонист NGF связывается с NGF и предотвращает димеризацию и/или аутофосфорилирование рецептора TrkA. В других вариантах осуществления изобретения антагонист NGF ингибирует или уменьшает синтез и/или продуцирование (высвобождение) NGF. В настоящем описании изобретения приведены примеры антагонистов NGF разных типов.
В используемом здесь значении термин “антитело против NGF” означает антитело, которое способно связываться с NGF и ингибировать биологическую активность NGF и/или пути передачи сигналов в нижней области, опосредуемые NGF.
Термин “иммуноадгезин TrkA” означает растворимую химерную молекулу, содержащую фрагмент рецептора TrkA, например внеклеточный домен рецептора TrkA, и последовательность иммуноглобулина, которая сохраняет специфичность связывания рецептора TrkA.
Термин “биологическая активность” NGF обычно означает способность связываться с рецепторами NGF и/или активировать пути передачи сигналов рецептора NGF. Термин “биологическая активность” без каких-либо ограничений имеет одно или несколько нижеследующих значений: способность связываться с рецептором NGF (таким как р75 и/или TrkA); способность стимулировать димеризацию и/или аутофосфорилирование рецептора TrkA; способность активировать путь передачи сигналов рецептора NGF; способность стимулировать дифференцировку, пролиферацию, выживаемость, рост, миграцию и другие изменения физиологии клеток, включая (в случае нейронов, в том числе периферических и центральных нейронов) изменение морфологии, синаптогенеза, синаптической функции нейронов, высвобождение нейротрансмиттера и/или нейропептида и регенерацию после повреждения; и способность опосредовать боль, ассоциированную с метастазированием рака в кость.
В используемом здесь значении термин “лечение” означает способ получения благоприятных или желаемых клинических результатов. В соответствии с целями настоящего изобретения благоприятные или желаемые клинические результаты включают, не ограничиваясь ими, один или несколько нижеследующих аспектов: уменьшение любого проявления боли, в том числе интенсивности боли, ослабление одного или нескольких симптомов боли, обусловленной раком кости (например, боли, ассоциированной с местастазированием рака в кость), включая любой признак боли, обусловленной раком кости (такой как сокращение продолжительности боли и/или уменьшение восприятия или ощущения боли).
Термин “эффективное количество” означает количество, достаточное для достижения благоприятных или желаемых клинических результатов, включая ослабление или уменьшение боли. В соответствии с целями настоящего изобретения эффективное количество антагониста NGF представляет собой количество, достаточное для лечения, ослабления, уменьшения интенсивности или предотвращения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость. В некоторых вариантах осуществления изобретения “эффективное количество” может ослаблять постоянную и/или периодическую боль (включая боль, возникающую при ходьбе и прикосновении) при введении до, во время и/или после метастазирования рака в кость. В некоторых вариантах осуществления изобретения термин “эффективное количество” означает количество, достаточное для замедления развития боли, обусловленной раком кости, включать боль, ассоциированную с метастазированием рака в кость.
Термин “уменьшение проявления” боли означает любые способы ослабления боли (которые могут включать уменьшение потребности в лекарственных средствах и/или сокращение количества других лекарственных средств, обычно применяемых в данных условиях), продолжительности и/или частоты возникновения боли (включая, например, увеличение времени до появления у субъекта боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость). Как должно быть понятно специалистам в данной области, у субъектов может быть разная реакция на лечение, и поэтому, например, “способ уменьшения проявления боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость” предполагает введение субъекту антагониста NGF, описанного в настоящем описании изобретения, с учетом обоснованного ожидания, что такое введение может вызывать уменьшение проявления боли у конкретного субъекта.
Термин “уменьшение интенсивности” боли, обусловленной раком кости (такой как боль, ассоциированная с метастазированием рака в кость), или одного или нескольких симптомов боли, обусловленной раком кости, означает ослабление или улучшение одного или нескольких симптомов боли, обусловленной раком кости, по сравнению с болью, имеющей место без введения антагониста NGF. Термин “уменьшение интенсивности” означает также уменьшение продолжительности симптома.
Термин “временное облегчение” боли, обусловленной раком кости (такой как боль, ассоциированная с метастазированием рака в кость), или одного или нескольких симптомов боли, обусловленной раком кости, означает уменьшение степени проявления одного или нескольких нежелательных клинических признаков боли, обусловленной раком кости, у одного или нескольких субъектов, подвергаемых лечению антагонистом NGF в соответствии с настоящим изобретением.
В используемом здесь значении термин “замедление” развития боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, означает торможение, препятствование, задержку, стабилизацию и/или отсрочку прогрессирования боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость. Подобное замедление может быть различным по времени в зависимости от истории болезни и/или субъекта, подлежащего лечению. Как должно быть очевидно специалисту в данной области, существенное или значительное замедление может в действительности включать предотвращение боли, выражающееся в том, что у субъекта не возникает боль, обусловленная раком кости, включая боль, ассоциированную с метастазированием рака в кость. Способ “замедления” развития симптома является способом, который уменьшает вероятность возникновения симптома в данный период времени и/или уменьшает степень проявления симптома в данный период времени по сравнению с ситуацией, когда данный способ не применяется. Такие сравнения обычно основаны на клинических исследованиях с участием числа субъектов, достаточного для получения статистически значимого результата.
Термин “развитие” или “прогрессирование” боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, означает первоначальное проявление и/или дальнейшее развитие нарушения. Развитие боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, можно обнаружить и определить стандартными клиническими методами, хорошо известными в данной области. Однако развитие может также означать прогрессирование, не поддающееся обнаружению. В соответствии с целями настоящего изобретения развитие или прогрессирование означает биологическое изменение симптомов. Термин “развитие” означает проявление, рецидив и вспышку. В используемом здесь значении термин “вспышка” или “проявление” боли, обусловленной раком кости (такой как боль, ассоциированная с метастазированием рака в кость), означает первоначальное проявление симптомов и/или рецидив.
В используемом здесь значении термин “совместное введение” означает одновременное введение и/или введение в разное время. Совместное введение означает также введение в одном препарате (то есть наличие антагониста NGF и другого лекарственного средства в одной композиции) или введение в разных композициях. В используемом здесь значении термин “совместное введение” означает любой способ одновременного и/или раздельного введения субъекту лекарственного средства и антагониста NGF. Как должно быть понятно из дальнейшего описания настоящего изобретения, антагонист NGF и другое лекарственное средство можно вводить в разных дозах или через разные интервалы времени. Например, антитело против NGF можно вводить один раз в неделю, в то время как другое лекарственное средство можно вводить чаще. Вполне понятно, что антагонист NGF и другое лекарственное средство можно вводить одинаковыми или разными способами введения.
Термин “наркотический аналгетик” означает все натуральные или синтетические лекарственные средства, действующие подобно морфину. Синтетические и полусинтетические наркотические аналгетики являются производными соединений пяти химических классов: фенантрены, фенилгептиламины, фенилпиперидины, морфинаны и бензоморфаны, все из которых входят в определение данного термина. Типичные наркотические аналгетики включают кодеин, дигидрокодеин, диацетилморфин, гидрокодон, гидроморфон, леворфанол, оксиморфон, алфентанил, бупренорфин, буторфанол, фентанил, суфентанил, меперидин, метадон, налбуфин, пропоксифен и пентазоцин или их фармацевтически приемлемые соли.
Термин “NSAID” означает нестероидное противовоспалительное соединение. Нестероидные противовоспалительные соединения классифицируются с учетом их способности ингибировать циклооксигеназу. Циклооксигеназа 1 и циклооксигеназа 2 являются двумя основными изоформами циклооксигеназы, и большинство стандартных NSAID являются смешанными ингибиторами двух указанных изоформ. Большинство стандартных NSAID входят в одну из пяти нижеследующих структурных категорий: (1) производные пропионовой кислоты, такие как ибупрофен, напроксен, напрозин, диклофенак и кетопрофен; (2) производные уксусной кислоты, такие как толметин и слиндак; (3) производные фенамовой кислоты, такие как мефенамовая кислота и меклофенамовая кислота; (4) производные бифенилкарбоновой кислоты, такие как дифлунизал и флуфенизал; и (5) оксикамы, такие как пироксим, судоксикам и изоксикам.
В научной литературе описан другой класс NSAID, которые избирательно ингибируют циклооксигеназу 2. Ингибиторы СOX-2 описаны, например, в патентах США №№ 5616601, 5604260, 5593994, 5550142, 5536752, 5521213, 5475995, 5639780, 5604253, 5552422, 5510368, 5436265, 5409944 и 5130311, которые включены в настоящее описание изобретения в качестве ссылки. Некоторые типичные ингибиторы СОХ-2 включают целекоксиб (SC-58635), DUP-697, флозулид (CGP-28238), мелоксикам, 6-метокси-2-нафтилуксусную кислоту (6-MNA), рофекоксиб, МК-966, набуметон (пролекарство для 6-MNA), нимесулид, NS-398, SC-5766, SC-58215, T-614 или их комбинации.
Термин “субъект” означает млекопитающего, предпочтительно человека. Млекопитающие включают, не ограничиваясь ими, сельскохозяйственных животных, спортивных животных, домашних животных, приматов, лошадей, собак, кошек, мышей и крыс.
Способы по настоящему изобретению
При описании всех способов по настоящему изобретению ссылка на антагонист NGF включает также композиции, содержащие один или несколько указанных агентов. Такие композиции могут дополнительно содержать приемлемые эксципиенты, такие как фармацевтически приемлемые эксципиенты (носители), включая буферы, которые хорошо известны в данной области. Настоящее изобретение может быть использовано отдельно или в сочетании с другими известными способами лечения.
Способы профилактики или лечения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость
Настоящее изобретение предназначено для лечения, замедления развития и/или предотвращения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, у субъекта, являющегося как человеком, так и млекопитающим, отличным от человека. С помощью настоящего изобретения можно улучшить качество жизни субъектов, страдающих раком кости.
Метастазирование рака в кость может быть ассоциировано с образованием или разрушением костной ткани. В некоторых вариантах осуществления изобретения способ по настоящему изобретению применяют для лечения боли, обусловленной раком кости, которая ассоциирована с образованием костной ткани (остеобластическая активность), в частности для лечения боли, обусловленной метастазированием в кость рака предстательной железы. В некоторых вариантах осуществления изобретения способ по настоящему изобретению применяют для лечения боли, обусловленной раком кости, которая ассоциирована с разрушением костной ткани (остеолитическая активность), в частности для лечения боли, обусловленной метастазированием саркомы в кость.
Таким образом, одним объектом настоящего изобретения являются способы лечения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, которые включают введение нуждающемуся субъекту эффективного количества антагониста NGF, такого как антитело против NGF. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком и NSAID. В некоторых вариантах осуществления изобретения количество наркотического аналгетика и/или NSAID, вводимое для ослабления боли, может быть меньше по сравнению с количеством, вводимым без антагониста NGF. Вредное действие наркотического аналгетика и/или NSAID можно уменьшить или устранить при совместном введении с антагонистом NGF. В некоторых вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика. В других вариантах осуществления изобретения антагонист NGF вводят без NSAID. В других вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика и/или NSAID.
Другим объектом настоящего изобретения являются способы профилактики, ослабления и/или предотвращения развития или прогрессирования боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком и NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика. В других вариантах осуществления изобретения антагонист NGDF вводят без NSAID. В других вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика и/или NSAID.
Должно быть понятно, что, хотя в данном описании изобретения особое внимание уделено лечению или профилактике боли, обусловленной раком кости, такой как боль, ассоциированная с метастазированием рака в кость, антагонист NGF можно вводить до возникновения состояния, характеризующегося повышенным риском боли, обусловленной раком кости.
Антагонист NGF можно вводить одновременно с лечением рака кости, таким как облучение и химиотерапия. Антагонист NGF можно также вводить вместе с другими аналгетиками, используемыми для ослабления боли, обусловленной раком кости. Примерами таких аналгетиков являются бисфосфонаты (например, алендронат), габапентин и облучение. Количество таких аналгетиков, вводимых для ослабления боли, обусловленной раком кости, может быть уменьшено по сравнению с количеством, вводимым без антагониста NGF. Вредное действие указанных аналгетиков можно уменьшить или устранить при совместном введении с антагонистом NGF.
В данной области хорошо известны методы диагностики или определения боли. Определение боли может быть произведено на объективной основе, например, путем наблюдения за поведением субъекта, в частности за реакцией на раздражители, выражением лица и подобными признаками. Оценка может быть также произведена на субъективной основе, в частности по определению боли субъектом с использованием шкалы боли разных типов. См., например, публикации Katz et al., Surg. Clin. North. Am. (1999) 79(2):231-52; Caraceni et al., J. Pain Symptom Manage (2002) 23(3):239-55.
Антагонисты NGF
При осуществлении способов по настоящему изобретению используют антагонист NGF, которым является любая молекула, блокирующая, подавляющая или уменьшающая (в том числе значительно) биологическую активность NGF, включая пути передачи сигналов в нижней области, опосредуемые NGF, в частности связывание с рецептором и/или индукцию реакции клетки на NGF. Термин “антагонист” не означает какого-либо специфического механизма биологического действия и скорее предполагает все возможные фармакологические, физиологические и биохимические взаимодействия с NGF и их последствия, которые могут быть достигнуты с помощью разных и химически разнообразных композиций. Типичные антагонисты NGF включают, не ограничиваясь ими, антитело против NGF, антисмысловую молекулу к NGF (включая антисмысловую молекулу к нуклеиновой кислоте, кодирующей NGF), антисмысловую молекулу к рецептору NGF (такому как рецептор ТrkA и/или р75) (включая антисмысловую молекулу к нуклеиновой кислоте, кодирующей рецептор TrkA и/или р75), ингибитор NGF, структурный аналог NGF, доминантноотрицательную мутацию рецептора TrkA, связывающего NGF, иммуноадгезин TrkA, антитело против TrkA, доминантноотрицательную мутацию рецептора р75, связывающего NGF, антитело против р75 и ингибитор киназы. В соответствии с целями настоящего изобретения должно быть хорошо понятно, что термин “антагонист” означает все ранее известные термины, названия, функциональные состояния и характеристики, которые предполагают по существу устранение, ослабление или нейтрализацию в любой значимой степени самого NGF, биологической активности NGF (включая его способность опосредовать любой признак боли, ассоциированной с метастазированием рака в кость) или последствий такой биологической активности. В некоторых вариантах осуществления изобретения антагонист NGF (например, антитело) связывается (физически взаимодействует) с NGF, связывается с рецептором NGF (таким как рецептор TrkA и/или р75) и/или ослабляет (препятствует и/или блокирует) передачу сигналов рецептора NGF в нижней области. Таким образом, в некоторых вариантах осуществления изобретения антагонист NGF связывается (физически взаимодействует) с NGF. В некоторых вариантах осуществления изобретения антагонист NGF является полипептидом, связывающимся с NGF. В некоторых вариантах осуществления изобретения антагонист NGF является пептидом или модифицированным пептидом (таким как NGF-связывающий пептид, слитый с Fc-областью), который описан в публикации РСТ WO 2004/026329. В другом варианте осуществления изобретения антагонист NGF связывается с рецептором NGF (таким как рецептор trkA или р75). В других вариантах осуществления изобретения антагонист NGF уменьшает (препятствует и/или блокирует) передачу сигналов рецептора NGF в нижней области (например, ингибиторы передачи сигналов киназы и ингибиторы путей передачи сигналов в нижней области). В других вариантах осуществления изобретения антагонист NGF ингибирует (уменьшает) синтез и/или высвобождение NGF. В другом варианте осуществления изобретения антагонист NGF не является иммуноадгезином TrkA (то есть отличен от иммуноадгезина TrkA). В другом варианте осуществления изобретения антагонист NGF не является антителом против NGF. В другом варианте осуществления изобретения антагонист NGF не является иммуноадгезином TrkA и антителом против NGF. В некоторых вариантах осуществления изобретения антагонист NGF связывается с NGF (таким как hNGF) и по существу не связывается с родственными нейротрофинами, такими как NT-3, NT4/5 и/или BDNF. В некоторых вариантах осуществления изобретения антагонист NGF не вызывает вредной иммунной реакции. В других вариантах осуществления изобретения антагонист NGF является антителом против NGF. В других вариантах осуществления изобретения антитело против NGF является гуманизированным антителом (таким как антитело Е3, описанное в настоящем описании изобретения). В некоторых вариантах осуществления изобретения антитело против NGF является антителом Е3 (рассмотренным в настоящем описании изобретения). В других вариантах осуществления изобретения антитело против NGF включает один или несколько гипервариабельных участков антитела Е3 (в частности, один, два, три, четыре, пять или в некоторых вариантах осуществления изобретения все шесть гипервариабельных участков антитела Е3). В других вариантах осуществления изобретения антитело является антителом человека. В некоторых вариантах осуществления изобретения антитело является нейтрализующим антителом человека против NGF, описанным в WO 2005/019266. В других вариантах осуществления изобретения антитело против NGF содержит аминокислотную последовательность вариабельной области тяжелой цепи, показанную в таблице 1 (SEQ ID NO:1), и аминокислотную последовательность вариабельной области легкой цепи, показанную в таблице 2 (SEQ ID NO:2). В других вариантах осуществления изобретения антитело содержит модифицированную константную область, в частности иммунологически инертную константную область, которая, например, не запускает комплемент-опосредованный лизис и не стимулирует антителозависимую клеточно-опосредованную цитотоксичность (ADCC). В других вариантах осуществления изобретения константная область модифицирована в соответствии с описанием, приведенным в публикации Eur. J. Immunol. (1999) 29:2613-2624, в заявке на патент РСТ № РСТ/GB99/01441 и/или в заявке на патент Великобритании № 9809951.8.
Антитела против NGF
В некоторых вариантах осуществления изобретения антагонист NGF является антителом против NGF. Антитело против NGF должно выполнять одну или несколько нижеследующих функций: связываться с NGF, ингибировать биологическую активность NGF и/или пути передачи сигналов в нижней области, опосредуемые NGF; (b) предотвращать, ослаблять или лечить любое проявление боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость; (с) блокировать или уменьшать активацию рецептора NGF (включая димеризацию и/или аутофосфорилирование рецептора TrkA); (d) увеличить клиренс NGF; (e) ингибировать (уменьшать) синтез, продуцирование или высвобождение NGF.
Антитела против NGF известны в данной области; см., например, публикации РСТ №№ WO 01/78698, WO 01/64247, патенты США №№ 5844092, 5877016 и 6153189; статьи Hongo et al., Hybridoma, 19:215-227 (2000); Cell. Molec. Biol. 13:559-568 (1993); №№ доступа в банке генов GenBank U39608, U39609, L17078 или L17077.
В некоторых вариантах осуществления изобретения антитело против NGF является гуманизированным моноклональным антителом мыши против NGF, получившим название “антитело Е3” (РСТ WO 04/058184), которое содержит константную область тяжелой цепи IgG2a человека, включающую следующие мутации: А330Р331-S330S331 (нумерация аминокислот произведена в соответствии с последовательностью IgG2a дикого типа; см. статью Eur. J. Immunol. (1999) 29:2613-2624); константную область легкой каппа-цепи и вариабельные области тяжелой и легкой цепи, показанные в таблицах 1 и 2.
Таблица 1. Вариабельная область тяжелой цепи
QVQLQESGPGLVKPSETLSLTCTVSGFSLIGYDLNWIRQPPGKGLEWIGIIWGDGTTDYNSAVKSRVTISKDTSKNQFSLKLSSVTAADTAVYYCARGGYWYATSYYFDYWGQGTLVTVS (SEQ ID NO:1).
Таблица 2. Вариабельная область легкой цепи
DIQMTQSPSSLSASVGDRVTITCRASQSISNNLNWYQQKPGKAPKLLIYYTSRFHSGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQEHTLPYTFGQGTKLEIKRT (SEQ ID NO:2)
Нижеследующие полинуклеотиды, кодирующие вариабельную область тяжелой цепи или вариабельную область легкой цепи депонированы в АТСС 8 января 2003 г.
Вектор Eb.911.3E является полинуклеотидом, кодирующим вариабельную область легкой цепи, показанную в таблице 2; вектор Eb.pur.911.3E является полинуклеотидом, кодирующим вариабельную область легкой цепи, показанную в таблице 2, и вектор Db.911.3E является полинуклеотидом, кодирующим вариабельную область тяжелой цепи, показанную в таблице 1. Указанные полинуклеотиды кодируют также константные области.
Существует по крайней мере два метода определения гипервариабельных участков (CDR): (1) метод, основанный на вариабельности последовательностей в разных видах (см. публикацию Kabat et al., Sequences of Proteins of Immunological Interest, (5th ed., 1991, National Institutes of Health, Bethesda, MD)); и (2) метод, основанный на кристаллографическом исследовании комплексов антиген-антитело (Chothia et al. (1989) Nature 342:877; Al-lazikani et al. (1997) J. Molec. Biol. 273:927-948)). В используемом здесь значении CDR может означать гипервариабельные участки, определенные любым методом или комбинацией обоих методов.
В другом варианте осуществления изобретения антитело против NGF включает один или несколько гипервариабельных участков антитела Е3 (в частности, один, два, три, четыре, пять или в некоторых вариантах осуществления изобретения все шесть гипервариабельных участков антитела Е3). В данной области хорошо известны методы определения гипервариабельных участков. CDR может представлять собой Kabat, Chotia или комбинацию Kabat и Chotia.
Антитела, пригодные для использования в настоящем изобретении, могут включать моноклональные антитела, поликлональные антитела, фрагменты антител (например, Fab, Fab', F(ab')2, Fv, Fc и т.д.), химерные антитела, биспецифические антитела, гетероконъюгатные антитела, одноцепочечные антитела (ScFv), их мутанты, слитые белки, содержащие антитело, гуманизированные антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая имеет сайт распознавания антигена требуемой специфичности, в том числе гликозилированные варианты антител, варианты аминокислотной последовательности антител и ковалентно модифицированные антитела. Указанные антитела могут принадлежать мышам, крысам, человеку или иметь любое другое происхождение (включая химерные или гуманизированные антитела). В соответствии с целями настоящего изобретения антитело взаимодействует с NGF, ингибируя NGF и/или пути передачи сигналов в нижней области, опосредуемые NGF. В одном варианте осуществления изобретения антитело является антителом человека, которое распознает один или несколько эпитопов в NGF человека. В другом варианте осуществления изобретения антитело является антителом мыши или крысы, которое распознает один или несколько эпитопов в NGF человека. В другом варианте осуществления изобретения антитело распознает один или несколько эпитопов в NGF, выбираемом из группы, включающей приматов, собак, кошек, лошадей и коров. В других вариантах осуществления изобретения антитело содержит модифицированную константную область, в частности иммунологически инертную константную область, которая, например, не запускает комплемент-опосредованный лизис или не стимулирует антителозависимую клеточно-опосредованную цитотоксичность (ADCC). Активность ADCC можно определить методами, описанными в патенте США № 5500362. В других вариантах осуществления изобретения константная область модифицирована в соответствии с описанием, приведенным в cтатье Eur. J. Immunol. (1999) 29:2613-2624; в заявке на патент РСТ № РСТ/GB99/01441 и/или в заявке на патент Великобритании № 9809951.8.
Сродство связывания антитела против NGF с NGF (таким как hNGF) может быть равно примерно 0,10-0,80 нМ, примерно 0,15-0,75 нМ и примерно 0,18-0,72 нМ. В одном варианте осуществления изобретения сродство связывания находится в пределах от около 2 пМ до 22 пМ. В некоторых вариантах осуществления изобретения сродство связывания равно примерно 10 нМ. В других вариантах осуществления изобретения сродство связывания составляет менее примерно 10 нМ. В других вариантах осуществления изобретения сродство связывания равно примерно 0,1 нМ или примерно 0,07 нМ. В других вариантах осуществления изобретения сродство связывания составляет менее примерно 0,1 нМ или менее примерно 0,07 нМ. В других вариантах осуществления изобретения сродство связывания составляет около 100 нМ, около 50 нМ, около 10 нМ, около 1 нМ, около 500 пМ, около 100 пМ или от около 50 пМ до около 2 пМ, около 5 пМ, около 10 пМ, около 15 пМ, около 20 пМ или около 40 пМ. В некоторых вариантах осуществления изобретения сродство связывания равно примерно 100 нМ, примерно 50 нМ, примерно 10 нМ, примерно 1 нМ, примерно 500 пМ, примерно 100 пМ, примерно 50 пМ или менее примерно 50 пМ. В некоторых вариантах осуществления изобретения сродство связывания составляет менее примерно 100 нМ, примерно 50 нМ, примерно 10 нМ, примерно 1 нМ, примерно 500 пМ, примерно 100 пМ или примерно 50 пМ. В других вариантах осуществления изобретения сродство связывания равно примерно 2 пМ, примерно 5 пМ, примерно 10 пМ, примерно 15 пМ, примерно 20 пМ, примерно 40 пМ или более примерно 40 пМ.
Одним способом определения сродства связывания антител с NGF является измерение сродства связывания монофункциональных Fab-фрагментов антитела. Монофункциональные Fab-фрагменты можно получить путем расщепления антитела (например, IgG) папаином или рекомбинантной экспрессии. Сродство Fab-фрагмента антитела против NGF можно определить, исследуя поверхностный плазмонный резонанс (система определения поверхностного плазмонного резонанса (SPR) BIAcore3000™, BIAcore, INC, Piscaway NJ). Чипы СМ5 могут быть активированы гидрохлоридом N-этил-N'-(3-диметиламинопропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. NGF человека (или любой другой NGF) разводят в 10 мМ ацетата натрия, рН 4,0, и впрыскивают на активированный чип в концентрации 0,005 мг/мл. При разном времени течения по отдельным каналам чипа могут быть получены два диапазона плотности антигена: 100-200 реакционных единиц (RU) для всестороннего кинетического исследования и 500-600 RU для скрининга. Чип может быть заблокирован этаноламином. Исследования регенерации показывают, что смесь буфера для элюирования компании Pierce (продукт № 21004, Pierce Biotechnology, Rockford, IL) и 4 М раствора NaCl (2:1) позволяет эффективно удалить связанный Fab при сохранении активности hNGF на чипе на протяжении 200 впрыскиваний. В качестве рабочего буфера для выполнения анализов в системе BIAcore используют буфер HBS-EP (0,01 M HEPES, рН 7,4, 0,15 NaCl, 3 мМ EDTA, 0,005% поверхностно-активного вещества Р20). Последовательно разведенные (0,1-10-кратные значения определенной KD) очищенные образцы Fab впрыскивают в течение 1 минуты со скоростью 100 мкл/мин при времени диссоциации до 2 часов. Концентрации белков Fab определяют методом ELISA и/или электрофорезом SDS-PAGE, используя Fab в известной концентрации (определенной при помощи анализа аминокислоты) в качестве эталона. Производят одновременное определение скоростей кинетической ассоциации (kon) и кинетической диссоциации (koff), сравнивая полученные данные с моделью связывания Ленгмюра в отношении 1:1 (Karlsson, R. Roos, H. Fagerstam, L. Petersson, B. (1994). Methods Enzymology 6, 99-110) при помощи программы оценки BIAevaluation. Значения константы равновесной диссоциации (KD) вычисляют в виде koff/kon. Указанный метод пригоден для определения сродства связывания антитела с любым NGF, включая NGF человека, NGF другого позвоночного (млекопитающего в некоторых вариантах осуществления изобретения) (такого как NGF мыши, NGF крысы, NGF примата), а также с другими нейротрофинами, такими как родственные нейротрофины NT3, NT4/5 и/или BDNF.
В некоторых вариантах осуществления изобретения антитело связывается с NGF человека и по существу не связывается с NGF другого вида позвоночных (млекопитающего в некоторых вариантах осуществления изобретения). В некоторых вариантах осуществления изобретения антитело связывается с NGF человека, а также с одним или несколькими NGF другого вида позвоночных (млекопитающего в некоторых вариантах осуществления изобретения). В других вариантах осуществления изобретения антитело связывается с NGF и по существу не взаимодействует с другими нейротрофинами (такими как родственные нейротрофины, NT3, NT4/5 и/или BDNF). В некоторых вариантах осуществления изобретения антитело связывается с NGF, а также по крайней мере с одним другим нейротрофином. В некоторых вариантах осуществления изобретения антитело связывается с NGF млекопитающих, таких как лошадь или собака, и по существу не связывается с NGF млекопитающих других видов.
Эпитоп может представлять собой детерминанту непрерывного или прерывистого типа. В одном варианте осуществления изобретения антитело связывается в основном с теми же эпитопами hNGF, что и антитело, выбираемое из группы, включающей MAb 911, MAb 912 и MAb 938, описанными в публикации Hongo et al., Hybridoma, 19: 215-227 (2000). В другом варианте осуществления изобретения антитело связывается в основном с тем же эпитопом hNGF, что и MAb 911. В другом варианте осуществления изобретения антитело связывается в основном с тем же эпитопом, что и MAb 909. См. приведенную выше публикацию Hongo et al. Например, эпитоп может включать один или несколько остатков К32, К34 и Е35 в вариабельной области 1 (аминокислоты 23-35) hNGF; остатки F79 и Т81 в вариабельной области 4 (аминокислоты 81-88) hNGF; остатки Н84 и К88 в вариабельной области 4; остаток R103 между вариабельной областью 5 (аминокислоты 94-98) hNGF и С-концом (аминокислоты 111-118) hNGF; остаток Е11 в предвариабельной области 1 (аминокислоты 10-23) hNGF; Y52 между вариабельной областью 2 (аминокислоты 40-49) hNGF и вариабельной областью 3 (аминокислоты 59-66) hNGF; остатки L112 и S113 у С-конца hNGF; остатки R59 и R69 в вариабельной области 3 hNGF или остатки V18, V20 и G23 в предвариабельной области 1 hNGF. Кроме того, эпитоп может содержать одну или несколько областей, включающих вариабельную область 1, вариабельную область 3, вариабельную область 4, вариабельную область 5, N-концевую область и/или С-конец hNGF. В другом варианте осуществления изобретения антитело значительно уменьшает доступ растворителя к остатку R103 hNGF. Должно быть понятно, что, хотя вышеописанные эпитопы являются родственными NGF человека, специалист в данной области может сравнить структуры NGF человека с NGF другого вида и идентифицировать эпитопы, подобные данным эпитопам.
В соответствии с одним объектом настоящего изобретения антитела (например, человека, гуманизированные, мыши, химерные), способные ингибировать NGF, можно получить, используя иммуногены, экспрессирующие непроцессированную или процессированную последовательность NGF. В соответствии с другим объектом настоящего изобретения можно использовать иммуноген, включающий клетку, сверхэкспрессирующую NGF. Другим примером приемлемого иммуногена является белок NGF, содержащий непроцессированный NGF или часть белка NGF.
Антитела против NGF могут быть получены любым методом, известным в данной области. Способ и схема иммунизации животного-хозяина обычно соответствуют обычным методам стимуляции и продуцирования антитела, которые будут описаны ниже. Общие методы продуцирования антител человека и мыши известны в данной области и описаны в настоящем описании изобретения.
Считается, что можно использовать любых млекопитающих, включая человека, и их антитело-продуцирующие клетки в качестве основы для получения линий клеток гибридомы млекопитающего, включая человека. Животного-хозяина обычно инокулируют внутрибрюшинно, внутримышечно, перорально, подкожно, интраплантарно и/или интрадермально необходимым количеством иммуногена в соответствии с приведенным описанием.
Гибридомы могут быть получены из лимфоцитов и иммортализованных миеломных клеток обычным методом гибридизации соматических клеток, описанным в публикации Kohler, B. and Milstein, C. (1975) Nature 256:495-497, и модифицированным методом, описанным в публикации Buck, D.W. et al., In Vitro, 18:377-381 (1982). Для гибридизации можно использовать существующие линии миеломных клеток, которые включают, не ограничиваясь ими, Х63-Ag8.653, и линии миеломных клеток, предоставляемые Salk Institute, Cell Distribution Center, San Diego, Calif., USA. Указанный метод обычно включает слияние миеломных клеток и лимфоидных клеток с использованием фузогена, такого как полиэтиленгликоль, или электрического устройства, хорошо известного специалистам в данной области. После слияния клетки отделяют от гибридизирующей среды и выращивают в селективной питательной среде, такой как гипоксантин-аминоптерин-тимидиновая (НАТ) среда, для удаления негибридизированных материнских клеток. Для культивирования гибридом, секретирующих моноклональные антитела, можно использовать любую описанную здесь среду с сывороткой или без сыворотки. В качестве другой альтернативы методу слияния клеток для получения моноклональных антител против NGF по настоящему изобретению можно использовать иммортализованные В-клетки EBV. Гибридомы при желании можно размножить и субклонировать и супернатанты подвергнуть анализу в отношении активности против иммуногена при помощи обычных иммуноанализов (таких как радиоиммуноанализ, иммуноферментный анализ или флуоресцентный иммуноанализ).
Гибридомы, которые могут быть использованы в качестве источника антител, включают все производные, клетки-предшественники материнских гибридом, продуцирующих моноклональные антитела, специфичные к NGF, или их часть.
Гибридомы, продуцирующие такие антитела, могут быть выращены in vitro или in vivo известными методами. Моноклональные антитела могут быть выделены из культуральной среды или жидкостей организма известными методами очистки иммуноглобулина, такими как осаждение сульфатом аммония, гель-электрофорез, диализ, хроматография и ультрафильтрация. Нежелательную активность в случае ее присутствия можно устранить, например, путем пропускания препарата через адсорбенты из иммуногена, прикрепленного к твердой фазе, и элюирования или высвобождения требуемых антител из иммуногена. Иммунизация животного-хозяина при помощи NGF человека или его фрагмента, содержащего аминокислотную последовательность-мишень, конъюгированную с белком, который является иммуногенным для иммунизируемого вида, например гемоцианин лимфы улитки, сывороточный альбумин, бычий тироглобулин или ингибитор соевого трипсина, с использованием бифункционального или производного агента, такого как сложный эфир малеимидобензоилсульфосукцинимида (конъюгирование остатками цистеина), N-гидроксисукцинимид (конъюгирование остатками лизина), глутаральдегид, янтарный ангидрид, SOCl2 или R1N=C=NR, где R и R1 являются разными алкильными группами, позволяет получить популяцию антител (например, моноклональных антител).
При желании представляющее интерес антитело против NGF (моноклональное или поликлональное) может быть секвенировано, после чего полинуклеотидная последовательность может быть клонирована в вектор для экспрессии или размножения. Последовательность, кодирующая представляющее интерес антитело, может находиться в векторе в клетке-хозяине, причем указанная клетка-хозяин затем может быть размножена и заморожена для последующего использования. В качестве альтернативы полинуклеотидную последовательность можно использовать для генетической манипуляции с целью “гуманизации” антитела, для повышения сродства или улучшения других характеристик антитела. Например, можно создать константную область, которая будет в большей степени напоминать константные области человека во избежание иммунной реакции при использовании данного антитела в клинических испытаниях и лечении людей. Может быть желательно генетически манипулировать последовательностью антитела для достижения большего сродства с NGF и большей эффективности ингибирования NGF. Специалисту в данной области должно быть понятно, что в антителе против NGF можно заменить один или несколько полинуклеотидов при сохранении способности связываться с NGF.
“Гуманизированные” антитела обычно представляют собой молекулу, имеющую антигенсвязывающий сайт, выделенный из иммуноглобулина вида, отличного от человека, и сохраняющий структуру иммуноглобулина в молекуле с учетом структуры и/или последовательности иммуноглобулина человека. Антигенсвязывающий сайт может включать полные вариабельные области, слитые с константными областями, или только гипервариабельные участки (CDR), привитые к соответствующим остовным областям в вариабельных областях. Антигенсвязывающие сайты могут быть дикого типа или модифицированы путем замены одной или нескольких аминокислот, например модифицированы для достижения большего сходства с иммуноглобулином человека. Некоторые формы гуманизированных антител сохраняют все последовательности CDR (например, гуманизированное антитело мыши, которое содержит все шесть CDR из антител мыши). Другие формы гуманизированных антител имеют один или несколько CDR (один, два, три, четыре, пять, шесть), которые изменены относительно исходного антитела. В некоторых случаях остатки остовной области (FR) или другие остатки иммуноглобулина человека заменяют соответствующими остатками, отличными от человеческих. Кроме того, гуманизированные антитела могут содержать остатки, которые отсутствуют в антителе реципиента или в антителе донора.
Известны четыре общие стадии гуманизации моноклонального антитела. Указанные стадии включают: (1) определение нуклеотидной и прогнозируемой аминокислотной последовательности вариабельных областей легкой и тяжелой цепи исходного антитела; (2) конструирование гуманизированного антитела, то есть принятие решения относительно того, какая остовная область антитела должна быть использована в процессе гуманизации; (3) выполнение действительных методов гуманизации и (4) трансфекция и экспрессия гуманизированного антитела. См., например, патенты США №№ 4816567, 5807715, 5866692, 6331415, 5530101, 5693761, 5693762, 5585089, 6180370 и 6548640.
В научной литературе описан ряд молекул “гуманизированных” антител, включающих антиген-связывающий сайт, выделенный из иммуноглобулина, отличного от человеческого, в том числе химерные антитела, содержащие V-области грызунов или модифицированные V-области грызунов и ассоциированные с ними гипервариабельные участки (CDR), слитые с константными областями человека. См., например, публикации Winter et al., Nature 349:293-299 (1991), Lobuglio et al., Proc. Nat. Acad. Sci. USA 86:4220-4224 (1989), Shaw et al., J. Immunol. 138:4534-4538 (1987) и Brown et al., Cancer Res. 47:3577-3583 (1987). В других ссылках описаны гипервариабельные участки грызунов, привитые к опорной остовной области (FR) человека до слияния с соответствующей константной областью антитела человека. См., например, публикации Riechmann et al., Nature 332:323-327 (1988), Verhoeyen et al., Science 239: 1534-1536 (1988) и Jones et al., Nature 321:522-525 (1986). В других ссылках описаны гипервариабельные участки грызунов, удерживаемые рекомбинантно связанными остовными областями грызунов. См., например, публикацию европейского патента № 0519596. Указанные “гуманизированные” молекулы конструируют таким образом, чтобы минимизировать нежелательную иммунологическую реакцию в отношении молекул антитела против человека грызунов, которая ограничивает продолжительность и эффективность терапевтического применения указанных частей молекулы в организме реципиента, являющегося человеком. Например, константная область антитела может быть создана так, чтобы она была иммунологически инертной (например, не запускала лизис комплемента). См., например, заявку на патент РСТ № РСТ/GB99/01441; заявку на патент Великобритании № 9809951.8. Другие приемлемые методы гуманизации антител описаны в публикации Daugherty et al., Nucl. Acids Res. 19:2471-2476 (1991), в патентах США №№ 6180377, 6054297, 5997867, 5866692, 6210671 и 6350861 и в публикации РСТ № WO 01/27160. Гуманизация может также включать созревание аффинности. См., например, заявку на патент США № 10/745775 и РСТ/US03/41252.
В качестве другой альтернативы могут быть получены полностью человеческие антитела при использовании коммерчески доступных мышей, экспрессирующих специфические белки иммуноглобулина человека. Трансгенные животные, созданные для индукции более желательной (например, благодаря выработке полностью человеческих антител) или более сильной иммунной реакции, могут быть также использованы для продуцирования гуманизированных или человеческих антител. Примеры такой технологии включают Xenomouse™ компании Abgenix, Inc. (Fremont, CA), HuMAb-Mouse® и TC Mouse™ компании Medarex, Inc. (Princeton, NJ).
В качестве еще одной альтернативы антитела могут быть получены методами рекомбинантных ДНК и экспрессированы любым методом, известным в данной области. В качестве другой альтернативы антитела могут быть получены методами рекомбинантных ДНК путем отображения на фаге. См., например, патенты США №№ 5565332, 5580717, 5733743 и 6265150 и публикацию Winter et al., Annu. Rev. Immunol. 12:433-455 (1994). Альтернативно метод отображения на фаге (McCafferty et al., Nature 348:552-553 (1990)) можно использовать для получения антител и фрагментов антитела человека in vitro из наборов генов вариабельной (V) области иммуноглобулина, выделенных у неиммунизированных доноров. В соответствии с данным методом гены V-области антитела клонируют в рамке считывания основного или минорного гена белка оболочки нитевидного бактериофага, такого как М13 или fd, и отображают в виде функциональных фрагментов антитела на поверхности фаговой частицы. Так как нитевидная частица содержит копию одноцепочечной ДНК генома фага, селекция на основе функциональных свойств антитела также приводит к выбору гена, кодирующего антитело, обладающее указанными свойствами. Таким образом, фаг имитирует некоторые свойства В-клетки. Отображение на фаге может быть произведено разными методами; см., например, Jonhson, Kevin S. and Chiswell, David J., Current Opinion in Structural Biology 3, 564-571 (1993). Для отображения на фаге можно использовать несколько источников сегментов V-гена. Клаксон и др. (Clackson et al., Nature 352:624-628 (1991)) выделили разнообразные антитела против оксазолона из небольшой рандомизированной комбинаторной библиотеки V-генов, полученных из селезенки иммуннизированных мышей. Можно создать набор V-генов, полученных у неиммунизированных доноров, являющихся людьми, и выделить антитела к различным антигенам (включая аутоантигены) методами, описанными в публикациях Mark et al., J. Mol. Biol. 222: 581-597 (1991) или Griffith et al., EMBO J. 12:725-734 (1993). При возникновении естественной иммунной реакции гены антитела быстро накапливают мутации (соматическая сверхмутация). Некоторые внесенные изменения сообщают большую аффинность, и во время последующей антигенной стимуляции происходит предпочтительная репликация и дифференцировка В-клеток, отображающих высокоаффинный поверхностный иммуноглобулин. Естественный процесс можно имитировать с помощью метода, известного как “перестановка цепи”. Marks, et al., Bio/Technol. 10:779-783 (1992)). При выполнении данного метода аффинность “первичных” антител человека, полученных путем отображения на фаге, можно улучшить в результате последовательной замены генов V-области тяжелой и легкой цепи набором естественных вариантов генов V-области, полученных у неиммунизированных доноров. Указанный метод позволяет продуцировать антитела и фрагменты антител с аффинностью в диапазоне пМ-нМ. Стратегия получения очень больших наборов антител на фаге (известных также как “первичные библиотеки”) описана в публикации Waterhouse et al., Nucl. Acids Res. 21:2265-2266 (1993). Для получения антител человека из антител грызунов с образованием антитела человека, обладающего аналогичной аффинностью и специфичностью в отношении исходного антитела грызуна, можно также использовать перестановку генов. В соответствии с данным методом, который также определяется как “импринтинг эпитопа”, ген V-области тяжелой или легкой цепи антител грызунов, полученный методом отображения на фаге, заменяют набором генов V-области человека, создавая химеры грызуна-человека. Выбор на основании антигена позволяет выделить вариабельные области человека, способные восстанавливать функциональный антиген-связывающий сайт, то есть эпитоп определяет (запечатляет) выбор партнера. При повторении данного процесса для замены остальной V-области грызуна получают антитело человека (см. публикацию РСТ № WO 93/06213, опубликованную 1 апреля 1993 г.). В отличие от традиционного метода гуманизации антител человека путем прививки CDR данный метод позволяет получить полностью человеческие антитела, которые не имеют рамки считывания или остатков CDR грызуна.
Вполне понятно, что, хотя приведенное выше описание относится к гуманизированным антителам, рассмотренные общие принципы применимы к специализации антител, предназначенных для использования, например, в организме собак, кошек, приматов, лошадей и коров. Кроме того, должно быть понятно, что можно объединить один или несколько аспектов гуманизации антитела, описанной в настоящем описании изобретения, например прививку CDR, мутацию рамки считывания и мутацию CDR.
Антитела могут быть получены методами рекомбинантных ДНК, в соответствии с которыми антитела и антителопродуцирующие клетки сначала выделяют из организма животных-хозяев, затем получают генную последовательность и используют указанную генную последовательность для рекомбинантной экспрессии антитела в клетках-хозяевах (например, в клетках яичника китайского хомячка (СНО)). Другим приемлемым методом является экспрессия последовательности антитела в растениях (например, в табаке) или трансгенном молоке. В научной литературе описаны методы рекомбинантной экспрессии антител в растениях или молоке. См., например, публикации Peeters et al., Vaccine 19:2756 (2001); Lonberg, N. and D. Huszar Int. Rev. Immunol. 13:65 (1995); и Pollock et al., J. Immunol. Methods 231:147 (1999). В данной области известны методы получения производных антител, например гуманизированных, одноцепочечных и т.д.
Кроме того, для выделения антител, специфичных к NGF, можно использовать методы сортировки с помощью иммуноанализов и проточной цитометрии, такие как клеточная сортировка клеток с возбуждением флуоресценции (FACS).
Антитела могут быть связаны со многими разными носителями. Носители могут быть активными и/или инертными. Примеры хорошо известных носителей включают полипропилен, полистирол, полиэтилен, декстран, найлон, амилазы, стекло, природные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетиты. Носитель может быть растворимым или нерастворимым в соответствии с целями настоящего изобретения. Специалистам в данной области должны быть известны другие приемлемые носители, используемые для связывания антител, либо такие носители могут быть выявлены в результате обычного экспериментирования.
ДНК, кодирующая моноклональные антитела, может быть легко выделена и секвенирована обычными методами (например, при помощи олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи моноклональных антител). Клетки гибридомы служат в качестве предпочтительного источника такой ДНК. Выделенная ДНК может быть помещена в экспрессирующие векторы (такие как экспрессирующие векторы, описанные в публикации РСТ № WO 87/04462), которые затем трансфицируют в клетки-хозяева, такие как клетки E.coli, клетки COS обезьян, клетки яичника китайского хомячка (СНО) или миеломные клетки, которые не продуцируют белок иммуноглобулина, для синтеза моноклональных антител в рекомбинантных клетках-хозяевах. См., например, публикацию РСТ № WO 87/04462. Указанная ДНК может быть также модифицирована, например, путем замены гомологичных последовательностей мышей последовательностью, кодирующей константные области тяжелой и легкой цепи человека, Morrison et al., Proc. Nat. Acad. Sci. 81:6851 (1984), или путем ковалентного связывания с кодирующей последовательностью иммуноглобулина всей или части последовательности, кодирующей полипептид, не являющийся иммуноглобулином. Таким образом получают “химерные” или “гибридные” антитела, обладающие специфичностью связывания моноклонального антитела против NGF, описанного в настоящем описании изобретения.
Антитела против NGF можно исследовать методами, хорошо известными в данной области. Например, одним методом является идентификация эпитопа, с которым связывается антитело, или “картирование эпитопов”. В данной области известно много методов картирования и исследования локализации эпитопов на белках, включая растворение кристаллической структуры комплекса антиген-антитело, анализы конкурентного связывания, анализы экспрессии фрагментов гена и анализы на основе синтетических пептидов, которые описаны, например, в главе 11 публикации Harlow and Lane, Using Antibodies, a Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999. В качестве дополнительного примера можно привести картирование эпитопов, используемое для определения последовательности, с которой связывается антитело против NGF. Картирование эпитопов может быть произведено на коммерческой основе в разных компаниях, например в Pepscan Systems (Edelhertweg 15, 8219 PH Lelystad, The Netherlands). Эпитоп может быть линейным эпитопом, то есть расположенным в одном фрагменте аминокислот, или конформационным эпитопом, образованным трехмерной структурой аминокислот, которые не обязательно могут находиться в одном фрагменте (линейная последовательность первичной структуры). Можно выделить или синтезировать (например, методами рекомбинантных ДНК) пептиды разной длины (например, содержащие по крайней мере 4-6 аминокислот) и использовать их для выполнения анализов связывания с антителом против NGF. В другом примере эпитоп, с которым связывается антитело против NGF, можно определить путем системного скрининга с использованием перекрывающих пептидов, выделенных из последовательности NGF, и определения связывания антителом против NGF. При выполнении анализов экспрессии фрагментов гена производят произвольное или с помощью специфических генетических конструкций фрагментирование открытой рамки считывания, кодирующей NGF, и определяют реактивность экспрессированных фрагментов NGF в отношении испытуемого антитела. Фрагменты гена могут быть получены методом PCR и затем транскрибированы и транслированы в белок in vitro в присутствии меченных радиоактивным изотопом аминокислот. Связывание антитела с меченными радиоактивным изотопом фрагментами NGF определяют методами иммунопреципитации и гель-электрофореза. Некоторые эпитопы можно также идентифицировать, используя большие библиотеки произвольных пептидных последовательностей, отображенных на поверхности фаговых частиц (библиотеки фагов). Альтернативно определенную библиотеку перекрывающих пептидных фрагментов можно исследовать на связывание с испытуемым антителом при выполнении простых анализов связывания. В дополнительном примере можно отметить выполнение мутагенеза антигенсвязывающей области, эксперименты по замене доменов и мутагенез со сканированием аланина для идентификации остатков, требуемых, достаточных и/или необходимых для связывания с эпитопом. Например, эксперименты по замене доменов могут быть выполнены с использованием мутантного NGF, в котором разные фрагменты полипептида NGF заменены последовательностями из близко родственного, но отличного в антигенном отношении белка (такого как другой член семейства белков нейротрофинов). Оценивая связывание антитела с мутантным NGF, можно определить значение определенного фрагмента NGF для связывания с антителом.
Другим методом исследования антитела против NGF являются анализы конкурентного связывания с использованием других антител, которые, как известно, связываются с тем же антигеном, то есть с разными фрагментами NGF; указанные анализы позволяют определить, действительно ли данное антитело против NGF связывается с тем же эпитопом, что и другие антитела. Анализы конкурентного связывания хорошо известны специалистам в данной области. Примеры антител, которые могут быть использованы при выполнении анализов конкурентного связывания в настоящем изобретении, включают MAb 911, 912, 938, описанные в публикации Hongo, et al., Hybridoma 19: 215-227 (2000).
Другие антагонисты NGF
Могут быть использованы антагонисты NGF, не являющиеся антителами против NGF. В некоторых вариантах осуществления изобретения антагонист NGF включает по крайней мере одну антисмысловую молекулу, способную блокировать или уменьшать экспрессию функционального NGF. Нуклеотидные последовательности NGF хорошо известны и могут быть легко получены из общедоступных баз данных. См., например, публикации Borsani et al., Nuc. Acids Res. 1990, 18, 4020; номер доступа NM 002506; Ullrich et al., Nature 303: 821-825 (1983). Можно легко получить антисмысловые олигонуклеотидные молекулы, которые специфически связываются с мРНК NGF без перекрестного взаимодействия с другими полинуклеотидами. Типичные сайты направленного воздействия включают, не ограничиваясь ими, инициирующий кодон, 5'-концевые регуляторные области, кодирующую последовательность и 3'-концевую нетранслируемую область. В некоторых вариантах осуществления изобретения олигонуклеотиды имеют длину от около 10 до 100 нуклеотидов, от около 15 до 50 нуклеотидов, от около 18 до 25 нуклеотидов или больше. Олигонуклеотиды могут включать модификации остова, например фосфортиоатные связи, и 2'-O-сахарные модификации, хорошо известные в данной области. Типичные антисмысловые молекулы включают антисмысловые молекулы NGF, описанные в публикации США № 20010046959; см. также http://www.rna-tec.com/repair.htm.
В других вариантах осуществления изобретения антагонист NGF включает по крайней мере одну антисмысловую молекулу, способную блокировать или уменьшать экспрессию рецептора функционального NGF (такого как TrkA и/или р75). Woolf et al., J. Neurosci. (2001) 21(3):1047-55; Taglialetela et al., J. Neurochem (1996) 66(5): 1826-35. Нуклеотидные последовательности рецепторов TrkA и р75 известны и могут быть легко получены из существующих общедоступных баз данных.
Альтернативно экспрессию и/или высвобождение NGF и/или экспрессию рецепторов NGF можно уменьшить, используя метод удаления генов и методы на основе морфолиноолигонуклеотидов, мРНК или рибозимов, которые хорошо известны в данной области, См. адреса в Интернете
http://www.macalester.edu/~montgomery/RNAi.html;
http://pub32.ezboard.com/fmorpholinosfrm19.showMessage?topicID=6.topic;
http://www.highveld.com/ribozyme.html.
В других вариантах осуществления изобретения антагонист NGF включает по крайней мере один ингибитор NGF. В используемом здесь значении “ингибитор NGF” означает соединение, не являющееся антителом против NGF, которое прямо или косвенно уменьшает, подавляет, нейтрализует или устраняет биологическую активность NGF. Ингибитор NGF должен обладать одной или несколькими нижеследующими характеристиками, в частности, он должен: (а) связываться с NGF и ингибировать биологическую активность NGF и/или пути передачи сигналов в нижней области, опосредуемые NGF; (b) предотвращать, ослаблять или лечить любые проявления боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость; (с) блокировать или уменьшать активацию рецепторов NGF (включая димеризацию и/или аутофосфорилирование рецептора TrkA); (d) увеличивать клиренс NGF; (e) ингибировать (уменьшать) синтез, продуцирование или высвобождение NGF. Типичные ингибиторы NGF включают низкомолекулярные ингибиторы NGF, описанные в публикации США № 20010046959; соединения, ингибирующие связывание NGF с рецептором р75, которые описаны в публикации РСТ № WO 00/69829, и соединение PD90780 [7-(бензоиламино)-4,9-дигидро-4-метил-9-оксо-пиразоло[5,1-b]хиназолин-2-карбоновая кислота], описанное в публикации Colquhoun et al., J. Pharmacol. Exp. Ther. 310(2):505-11 (2004); соединения, ингибирующие связывание NGF с рецепторами TrkA и/или р75, которые описаны в публикации РСТ № WO 98/17278. Дополнительные примеры ингибиторов NGF включают соединения, описанные в публикациях РСТ №№ WO 02/17914 и WO 02/20479 и в патентах США №№ 5342942, 6127401 и 6359130. Другими типичными ингибиторами NGF являются соединения, которые представляют собой конкурентные ингибиторы NGF. См. патент США № 6291247. Кроме того, специалист в данной области может получить другие низкомолекулярные ингибиторы NGF.
В некоторых вариантах осуществления изобретения ингибиторы NGF связываются с NGF. Типичные сайты направленного воздействия (связывания) включают, не ограничиваясь ими, часть NGF, которая связывается с рецептором TrkA и/или р75, и части NGF, которые располагаются рядом с областью связывания с рецептором и частично определяют правильную трехмерную форму части, связывающейся с рецептором. В другом варианте осуществления изобретения ингибитор NGF связывается с рецептором NGF (таким как TrkA и/или р75) и подавляет биологическую активность NGF. Типичные сайты направленного воздействия включают части рецепторов TrkA и/или р75, которые связываются с NGF.
В вариантах осуществления изобретения, включающих использование малых молекул, такая малая молекула может иметь молекулярную массу около 100-20000 дальтон, 500-15000 дальтон или 1000-10000 дальтон. Библиотеки малых молекул являются коммерчески доступными. Малые молекулы можно вводить любыми способами, известными в данной области, которые включают ингаляцию, внутрибрюшинное, внутривенное, внутримышечное, подкожное, интратекальное, интравентрикулярное, пероральное, энтеральное, парентеральное, интраназальное или дермальное введение. Антагонист NGF по настоящему изобретению, являющийся малой молекулой, обычно вводят в количестве 0,1-300 мг/кг массы тела субъекта в виде трех или большего числа доз. Взрослому субъекту с нормальной массой тела можно вводить от 1 мг до 5 г в одной дозе.
В других вариантах осуществления изобретения антагонист NGF включает по крайней мере один структурный аналог NGF. Термин “структурные аналоги NGF” означает в настоящем изобретении соединения, которые имеют 3-мерную структуру, аналогичную части NGF, и связываются с рецептором NGF в физиологических условиях in vitro или in vivo, причем такое связывание по крайней мере частично ингибирует биологическую активность NGF. В одном варианте осуществления изобретения структурный аналог NGF связывается с рецептором TrkA и/или р75. Типичные структурные аналоги NGF включают, не ограничиваясь ими, бициклические пептиды, описанные в публикации РСТ № WO 97/15593; бициклические пептиды, описанные в патенте США № 6291247; циклические соединения, описанные в патенте США № 6017878, и пептиды, выделенные из NGF, которые описаны в публикации РСТ № WO 89/09225. Приемлемые структурные аналоги NGF могут быть также созданы и синтезированы методом молекулярного моделирования связывания с рецептором NGF, описанным, например, в публикации РСТ № WO 98/06048. Структурные аналоги NGF могут быть мономерами или димерами/олигомерами в любой требуемой комбинации одинаковых или разных структур с достижением улучшенного сродства и биологического действия.
Другие варианты осуществления изобретения относятся к антагонисту NGF, включающему по крайней мере один доминантноотрицательный мутант рецептора TrkA и/или р75. Специалист в данной области может получить такие доминантноотрицательные мутанты, например, рецептора TrkA, которые будут связывать NGF и действовать, таким образом, в качестве ловушки для NGF. Однако доминантноотрицательные мутанты не обладают нормальной биологической активностью рецептора TrkA при связывании с NGF. Типичные доминантноотрицательные мутанты включают, не ограничиваясь ими, мутанты, описанные в нижеследующих публикациях: Li et al., Proc. Natl. Acad. Sci. USA 1998, 95, 10884; Eide et al., J. Neurosci. 1996, 16, 3123; Liu et al., J. Neurosci 1997, 17, 8749; Klein et al., Cell 1990, 61, 647; Valenzuela et al., Neuron 1993, 10, 963; Tsoulfas et al., Neuron 1993, 10, 975; and Lamballe et al., EMBO J. 1993, 12, 3083, которые полностью включены в данное описание изобретения в качестве ссылки. Доминантноотрицательные мутанты можно вводить в форме белка или экспрессирующего вектора, обеспечивающего экспрессию доминантноотрицательного мутанта, например мутанта рецептора TrkA, in vivo. Белок или экспрессирующий вектор можно вводить любыми способами, известными в данной области, такими как внутрибрюшинное, внутривенное, внутримышечное, подкожное, интратекальное, интравентрикулярное, пероральное, энтеральное, парентеральное, интраназальное, дермальное введение или ингаляция. Например, экспрессирующие векторы можно вводить локально или системно, включая инъекции, пероральное введение, устройство, выстреливающее частицы, или катетер, и местно. Специалисту в данной области должны быть известны способы введения экспрессирующих векторов для экспрессии экзогенного белка in vivo. См., например, патенты США №№ 6436908, 6413942 и 6376471.
Можно также использовать направленную доставку терапевтических композиций, содержащих антисмысловой полинуклеотид, экспрессирующий вектор или субгеномные полинуклеотиды. Методы доставки ДНК, опосредованные рецепторами, описаны, например, в публикациях Findeis et al., Trends Biotechnol. (1993) 11:202; Chiou et al., Gene Therapeutics: Methods And Applications Of Direct Gene Transfer (J.A. Wolff, ed.) (1994); Wu et al., J. Biol. Chem. (1988) 263-621: Wu et al., J. Biol. Chem. (1994) 269:542; Zenke et al., Proc. Natl Acad. Sci. USA (1990) 87:3655; Wu et al., J. Biol. Chem. (1991) 266:338. Терапевтические композиции, содержащие полинуклеотид, вводят в количестве от около 100 нг до около 200 нг ДНК путем локального введения во время генотерапии. В некоторых вариантах осуществления изобретения во время генотерапии можно также вводить композиции в количестве от около 500 нг до около 50 мг, от около 1 мкг до около 2 мг, от около 5 мкг до около 500 мкг и от около 20 мкг до около 100 мкг ДНК или больше. Терапевтические полинуклеотиды и полипептиды по настоящему изобретению можно вводить с помощью векторов для доставки генов. Вектор для доставки генов может иметь вирусное или невирусное происхождение (см. публикации Jolly, Cancer Gene Therapy (1994) 1:51; Kimura, Human Gene Therapy (1994) 5:845; Connelly, Human Gene Therapy (1995) 1:185; и Kaplitt, Nature Genetics (1994) 6:148). Экспрессию таких кодирующих последовательностей можно индуцировать, используя эндогенные или гетерологичные промоторы и/или энхансеры млекопитающих. Экспрессия кодирующей последовательности может быть конститутивной или регулируемой.
В данной области хорошо известны векторы на основе вирусов, используемые для доставки и экспрессии требуемого полинуклеотида в нужной клетке. Типичные векторы на основе вирусов включают, не ограничиваясь ими, рекомбинантные ретровирусы (см., например, публикации РСТ №№ WO 90/07936, WO 94/03622, WO 93/25698, WO 93/25234, WO 93/11230, WO 93/10218, WO 91/02805, патенты США №№ 5219740 и 4777127, патент Великобритании № 2200651 и европейский патент № 0345242), векторы на основе альфавируса (например, векторы на основе вируса Синдбис, вируса лихорадки леса Семлики (АТСС VR-67, ATCC VR-1247), вируса Росс-ривер (АТСС VR-373, ATCC VR-1246) и вируса венесуэльского энцефалита лошадей (АТСС VR-923, ATCC VR-1250, ATCC VR-1249, ATCC VR-532)) и векторы на основе аденоассоциированного вируса (AAV) (см., например, публикации РСТ №№ WO 94/12649, WO 93/03769, WO 93/19191, WO 94/28938, WO 95/11984 и WO 95/00655). Можно также использовать метод введения ДНК, связанной с убитым аденовирусом, который описан в публикации Curiel, Hum. Gene Ther. (1992) 3:147.
Кроме того, можно использовать невирусные носители, которые включают, не ограничиваясь ими, поликатионную конденсированную ДНК, связанную или не связанную с убитым аденовирусом (см., например, публикацию Curiel, Hum. Gene Ther. (1992) 3:147), ДНК, связанную с лигандом (см., например, публикацию Wu, J. Biol. Chem. (1989) 264:16985), клетки-носители, служащие для доставки в эукариотические клетки (см., например, патент США № 5814482, публикации РСТ №№ WO 95/07994, WO 96/17072, WO 95/30763 и WO 97/42338), нейтрализацию заряда нуклеиновой кислоты или слияние с клеточными мембранами. Можно также использовать депротеинизированную ДНК. Типичные методы введения депротеинизированной ДНК описаны в публикации РСТ № WO 90/11092 и в патенте США № 5580859. Липосомы, способные действовать в качестве носителей для доставки генов, описаны в патенте США № 5422120, публикациях РСТ №№ WO 95/13796, WO 94/23697, WO 91/14445 и европейском патенте № 0524968. Дополнительные методы описаны в публикациях Philip, Mol. Cell Biol. (1994) 14:2411 и Woffendin, Proc. Natl. Acad. Sci. (1994) 91:1581.
Кроме того, очевидно, что экспрессирующий вектор можно использовать для прямой экспрессии любых антагонистов NGF на основе белка, описанных в настоящем описании изобретения (например, антитело против NGF, иммуноадгезин TrkA и т.д.). Например, в данной области известны другие фрагменты рецептора TrkA, способные блокировать (частично или полностью) NGF и/или биологическую активность NGF.
В другом варианте осуществления изобретения антагонист NGF включает по крайней мере один иммуноадгезин TrkA. Иммуноадгезины TrkA в используемом здесь значении означают растворимые химерные молекулы, содержащие внеклеточный домен рецептора TrkA и последовательность иммуноглобулина, которая сохраняет специфичность связывания рецептора TrkA (в основном сохраняет специфичность связывания рецептора trkA) и может связываться с NGF.
В данной области известны иммуноадгезины TrkA, которые, как установлено, блокируют связывание NGF с рецептором TrkA. См., например, патент США № 6153189. Бреннан и др. (Brennan et al.) описали использование иммуноадгезина TrkA в модели послеоперационной боли у крыс. См. публикацию Society for Neuroscience Abstracts 24 (1-2) 880 (1998). В одном варианте осуществления изобретения иммуноадгезин TrkA содержит аминокислотную последовательность рецептора TrkA (или ее часть) из внеклеточного домена TrkA, способного связывать NGF (в некоторых вариантах изобретения аминокислотную последовательность, в основном сохраняющую специфичность связывания рецептора trkA), которая слита с последовательностью иммуноглобулина. В некоторых вариантах осуществления изобретения рецептор TrkA представляет собой последовательность рецептора TrkA человека, и слияние осуществляют с последовательностью константной области иммуноглобулина. В других вариантах осуществления изобретения последовательность константной области иммуноглобулина является последовательностью константной области тяжелой цепи иммуноглобулина. В других вариантах осуществления изобретения объединение двух слияний рецептора TrkA и тяжелой цепи иммуноглобулина (например, путем ковалентного связывания при помощи дисульфидных связей) позволяет получить гомодимерную структуру, подобную иммуноглобулину. Легкая цепь иммуноглобулина может быть далее объединена с одной или обеими химерами рецептора TrkA и иммуноглобулина в димере, связанном дисульфидной связью, с получением гомотримерной или гомотетрамерной структуры. Примеры приемлемых иммуноадгезинов TrkA включают описанные в патенте США № 6153189.
В другом варианте осуществления изобретения антагонист NGF включает по крайней мере одно антитело против TrkA, способное блокировать, подавлять, изменять и/или уменьшать физическое взаимодействие NGF с рецептором TrkA и/или передачу сигналов в нижней области, при этом ослабляется и/или блокируется биологическая активность NGF. В данной области известны антитела против TrkA. Типичные антитела против TrkA включают антитела, описанные в публикациях РСТ №№ WO 97/21732, WO 00/73344, WO 02/15924 и публикации США № 20010046959.
В другом варианте осуществления изобретения антагонист NGF включает по крайней мере одно антитело против р75, способное блокировать, подавлять и/или уменьшать физическое взаимодействие NGF с рецептором р75 и/или передачу сигналов по ходу сигнального пути, при этом ослабляется и/или блокируется биологическая активность NGF.
В другом варианте осуществления изобретения антагонист NGF включает по крайней мере один ингибитор киназы, способный подавлять передачу сигналов киназы по ходу сигнального пути, ассоциированную с активностью рецептора TrkA и/или р75. Типичным ингибитором киназы является К252а или К252b, который известен в данной области и описан в нижеследующих публикациях Knusel et al., J. Neurochem. 59:715-722 (1992); Knusel et al., J. Neurochemistry 57:955-962 (1991); Koizumi et al., J. Neuroscience 8:715-721 (1988); Hirata et al., Chemical Abstracts 111:728, XP00204135, see abstract and 12th Collective Chemical Substance Index, p.34237, c. 3(5-7), (55-60, 66-69), p. 34238, c.1 (41-44), c.2 (25-27, 32-33), p. 3423, c.3 (48-50, 52-53); и в патенте США № 6306849.
Вполне вероятно, что при соответствующем поиске может быть идентифицирован целый ряд антагонистов NGF других категорий.
Идентификация антагонистов NGF
Антитела против NGF и другие антагонисты NGF могут быть идентифицированы или исследованы методами, известными в данной области, для этого можно обнаружить и/или измерить уменьшение, ослабление или нейтрализацию биологической активности NGF. Можно использовать методы, описанные в публикации РСТ WO 04/065560. Для идентификации антагонистов NGF можно использовать также другой метод, например анализ активации рецептора киназы (KIRA), описанный в патентах США №№ 5766863 и 5891650. Анализ типа ELISA пригоден для качественного или количественного измерения активации киназы путем измерения аутофосфорилирования домена киназы в протеин-тирозин-киназе рецептора (далее именуемая “rPTK”), например рецептора TrkA, а также для идентификации и исследования потенциальных антагонистов выбранной rTPK, например TrkA. Первая стадия данного анализа включает фосфорилирование домена киназы в рецепторе киназы, например в рецепторе TrkA, который находится в клеточной мембране эукариотической клетки. Указанный рецептор может быть эндогенным рецептором или нуклеиновой кислотой, кодирующей данный рецептор, либо в клетку может быть введена рецепторная конструкция. Первую твердую фазу (например, лунку в первом аналитическом планшете) обычно сенсибилизируют по существу гомогенной популяцией таких клеток (обычно линия клеток млекопитающего) с достижением сцепления клеток с твердой фазой. Клетки часто являются адгезивными и поэтому естественно сцепляются с первой твердой фазой. Используемая “рецепторная конструкция” обычно включает слияние рецептора киназы и сигнального полипептида. Сигнальный полипептид распознается захватывающим агентом, часто иммобилизованным антителом, при выполнении части ELISA данного анализа. Затем в лунки со сцепленными клетками добавляют анализируемое вещество, такое как антитело-кандидат против NGF или другие антагонисты NGF, при этом рецептор тирозинкиназы (например, рецептор TrkA) подвергается воздействию (или контактирует с) NGF и анализируемым веществом. Данный анализ позволяет идентифицировать антитела (или другие антагонисты NGF), которые ингибируют активацию TrkA лигандом NGF. Клетки, подвергнутые воздействию NGF и анализируемого вещества, солюбилизируют, используя лизирующий буфер (содержащий солюбилизирующий детергент), осторожно перемешивают и получают клеточный лизат, который подвергают анализу методом ELISA без концентрирования или очистки клеточного лизата.
Полученный таким образом клеточный лизат затем анализируют методом ELISA. На первой стадии анализа ELISA вторую твердую фазу (обычно лунку титрационного микропланшета для ELISA) сенсибилизируют захватывающим агентом (часто иммобилизованным антителом), который специфически связывается с рецептором тирозинкиназы или в случае рецепторной конструкции с сигнальным полипептидом. Сенсибилизацию второй твердой фазы выполняют таким образом, чтобы захватывающий агент сцеплялся со второй твердой фазой. Захватывающим агентом обычно является моноклональное антитело, но, как описано в примерах, приведенных в настоящем описании изобретения, можно также использовать поликлональные антитела. Полученный клеточный лизат затем подвергают воздействию или осуществляют контактирование со сцепляющим захватывающим агентом, в результате чего рецептор или рецепторная конструкция сцепляется (или захватывается) второй твердой фазой. Затем выполняют стадию промывки для удаления несвязанного клеточного лизата, получая при этом захваченный рецептор или рецепторную конструкцию. Сцепленный или захваченный рецептор или рецепторную конструкцию затем подвергают воздействию или осуществляют контактирование с антителом против фосфотирозина, которое позволяет идентифицировать остатки фосфорилированного тирозина в рецепторе тирозинкиназы. В одном варианте осуществления изобретения антитело против фосфотирозина конъюгировано (прямо или непрямо) с ферментом, катализирующим изменение цвета нерадиоактивного красителя. Фосфорилирование рецептора можно измерить с учетом последующего изменения цвета красителя. Фермент может быть связан прямо с антителом против фосфотирозина, или с антителом против фосфотирозина может быть конъюгирована конъюгирующая молекула (например, биотин), после чего фермент может быть связан с антителом против фосфотирозина при помощи данной конъюгирующей молекулы. И, наконец, по изменению цвета красителя измеряют связывание антитела против фосфотирозина с рецептором или рецепторной конструкцией.
Антагонисты NGF можно также идентифицировать путем культивирования агента-кандидата с NGF и исследования одной или нескольких нижеследующих характеристик: (а) связывание с NGF и ингибирование биологической активности NGF и/или путей передачи сигналов в нижней области, опосредуемой NGF; (b) предотвращение, ослабление или лечение любого проявления боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость; (с) блокирование или уменьшение активности рецептора NGF (включая димеризацию и/или аутофосфорилирование рецептора TrkA); (d) увеличение клиренса NGF; (e) ингибирование (уменьшение) синтеза, продуцирования или высвобождения NGF. В некоторых вариантах осуществления изобретения антагонист NGF идентифицируют, культивируя агент-кандидат с NGF и контролируя связывание и происходящее уменьшение или нейтрализацию биологической активности NGF. Анализ связывания может быть выполнен с использованием очищенных полипептидов NGF или клеток, которые естественным путем экспрессируют полипептиды NGF или трансфицированы для экспрессии полипептидов NGF. В одном варианте осуществления изобретения анализ связывания является конкурентно-связывающим анализом, при выполнении которого можно определить способность антитела-кандидата конкурировать с известным антагонистом NGF за связывание с NGF. Указанный анализ может быть выполнен разными методами, в том числе методом ELISA. В других вариантах осуществления изобретения антагонист NGF идентифицируют, культивируя агент-кандидат с NGF и контролируя происходящее ингибирование димеризации и/или аутофосфорилирования рецептора TrkA.
После первоначальной идентификации активность антагонистического антитела-кандидата против NGF может быть подтверждена при помощи биоанализов, которые, как известно, позволяют исследовать целенаправленную биологическую активность. Альтернативно биоанализы можно использовать для прямого скрининга кандидатов. Например, NGF стимулирует ряд морфологически распознаваемых изменений в иммунореактивных клетках. Указанные изменения включают, не ограничиваясь ими, стимуляцию дифференцировки клеток РС12 и усиление роста невритов из таких клеток (Greene and Tischler, Proc. Nat. Acad. Sci. USA 73:2424-2428 (1976); Urfer et al., Biochem. 36:4775-4781 (1997); Tsoulfas et al., Neuron 10:975-990 (1993)), стимуляцию роста невритов из эксплантатов сенсорного и симпатического ганглия (Levi-Montalcini, R. and Angeletti, P. Nerve growth factor. Physiol. Rev. 48, 534-569, 1968) и стимуляцию выживаемости NGF-зависимых нейронов, таких как нейроны ганглия первичных задних корешков, ганглия тройничного нерва или симпатического ганглия (см., например, публикации Chun & Patterson, Dev. Biol. 75:705-711, (1977); Buchman & Davies, Development 118:989-1001, (1993). Таким образом, анализ ингибирования биологической активности NGF включает культивирование NGF-иммунореактивных клеток вместе с NGF и анализируемым веществом, таким как антитело-кандидат против NGF или антагонист-кандидат NGF. По истечении соответствующего периода времени анализируют реакцию клеток (дифференцировку клеток, рост невритов или выживаемость клеток).
Способность антагониста-кандидата NGF блокировать или нейтрализовать биологическую активность NGF можно также определить, контролируя способность агента-кандидата ингибировать NGF-опосредованную выживаемость ганглия первичных задних корешков крысы в соответствии с описанием, приведенным в публикации Hongo et al., Hybridoma 19:215-227 (2000).
Композиции, предназначенные для применения в способах по настоящему изобретению
Композиции, применяемые в способах по настоящему изобретению, включают эффективное количество антагониста NGF (такого как антитело против NGF) и в некоторых вариантах осуществления изобретения дополнительно фармацевтически приемлемый эксципиент. В некоторых вариантах осуществления изобретения вышеуказанная композиция предназначена для применения в любых способах по настоящему изобретению. Примеры таких композиций и способы их получения описаны также в предшествующем разделе и ниже. В одном варианте осуществления изобретения композиция включает антагонист NGF. В другом варианте осуществления изобретения композиция включает один или несколько антагонистов NGF. В другом варианте осуществления изобретения композиция включает один или несколько антагонистов NGF, выбираемых из нижеследующей группы: антагонист (например, антитело), связывающийся (физически взаимодействующий) с NGF, антагонист, связывающийся с рецептором NGF (таким как рецептор TrkA и/или р75), и антагонист, ослабляющий (препятствующий и/или блокирующий) передачу сигналов рецептора NGF в нижней области. В других вариантах осуществления изобретения композиция включает любой антагонист NGF, который не является иммуноадгезином TrkA (то есть отличный от иммуноадгезина TrkA). В других вариантах осуществления изобретения композиция включает любой антагонист NGF, отличный от антитела против NGF. В других вариантах осуществления изобретения композиция включает любой антагонист NGF, отличный от иммуноадгезина TrkA и антитела против NGF. В других вариантах осуществления изобретения антагонист NGF ингибирует (уменьшает) синтез, продуцирование или высвобождение NGF. В некоторых вариантах осуществления изобретения антагонист NGF связывается с NGF и по существу не взаимодействует с родственными нейротрофинами (такими как NT3, NT4/5 и/или BDNF). В некоторых вариантах осуществления изобретения антагонист NGF не вызывает вредной иммунной реакции. В некоторых вариантах осуществления изобретения антагонист NGF выбирают из группы, включающей антитело против NGF, антисмысловую молекулу к NGF (включая антисмысловую молекулу к нуклеиновой кислоте, кодирующей NGF), антисмысловую молекулу к рецептору NGF (такому как TrkA и/или р75), ингибитор NGF, структурный аналог NGF, доминантноотрицательную мутацию рецептора TrkA, связывающего NGF, иммуноадгезин TrkA, антитело против TrkA, антитело против р75 и ингибитор киназы. В другом варианте осуществления изобретения антагонист NGF является антителом против NGF. В других вариантах осуществления изобретения антитело против NGF распознает NGF человека. В некоторых вариантах осуществления изобретения антитело против NGF является антителом человека. В других вариантах осуществления изобретения антитело против NGF является гуманизированным антителом (таким как антитело Е3, описанное в настоящем описании изобретения). В другом варианте осуществления изобретения антитело против NGF содержит константную область, которая не запускает нежелательную иммунную реакцию, такую как антитело-опосредованный лизис или ADCC. В других вариантах осуществления изобретения антитело против NGF включает один или несколько гипервариабельных участков антитела Е3 (в частности, один, два, три, четыре, пять или в некоторых вариантах осуществления изобретения все шесть гипервариабельных участков антитела Е3).
Понятно, что указанные композиции могут содержать более одного антагониста NGF. Например, композиция может содержать несколько членов класса антагонистов NGF (например, смесь антител против NGF, распознающих разные эпитопы NGF), а также членов разных классов антагонистов NGF (например, антитело против NGF и ингибитор NGF). Другие типичные композиции содержат несколько антител против NGF, распознающих одинаковые эпитопы, разные виды антител против NGF, которые связываются с разными эпитопами NGF, или разные ингибиторы NGF.
Композиция, используемая в настоящем изобретении, дополнительно может содержать фармацевтически приемлемые носители, эксципиенты или стабилизаторы (Remington: The Science and Practice of Pharmacy 20th Ed. (2000) Lippincott Williams and Wilkins, Ed. K.E. Hoover) в форме лиофилизированных препаратов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы должны быть нетоксичными для реципиентов в используемых дозах и концентрациях и могут включать буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включающие аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония, хлорид гексаметония, хлорид бензалкония, хлорид бензетония; феноловый, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол, резорцинол, циклогексанол, 3-пентанол и m-крезол); низкомолекулярные полипептиды (менее примерно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстраны; хелатообразователи, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионогенные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG). Фармацевтически приемлемые эксципиенты описаны далее в настоящем описании изобретения.
Антагонист NGF и содержащие его композиции можно также использовать в сочетании с другими лекарственными средствами, которые усиливают и/или дополняют его эффективность. В некоторых вариантах осуществления изобретения другим лекарственным средством является наркотический аналгетик. В некоторых вариантах осуществления изобретения другим лекарственным средством является NSAID. В некоторых вариантах осуществления изобретения указанные лекарственные средства не являются наркотическими аналгетиками. В некоторых вариантах осуществления изобретения указанные лекарственные средства не являются NSAID.
Наборы
Настоящее изобретение относится также к наборам, предназначенным для использования в способах по данному изобретению. Наборы по настоящему изобретению включают один или несколько контейнеров, содержащих антагонист NGF (такой как антитело, в частности гуманизированное антитело Е3, описанное в настоящем описании изобретения), и в некоторых вариантах осуществления изобретения дополнительно включают инструкции по использованию данного набора в соответствии с любыми способами по настоящему изобретению. В некоторых вариантах осуществления изобретения антагонист NGF является любым антагонистом NGF, описанным в настоящем описании изобретения. В других вариантах осуществления изобретения указанный набор включает антагонист NGF, который не является иммуноадгезином TrkA (то есть отличный от иммуноадгезина TrkA). В других вариантах осуществления изобретения набор включает антагонист NGF, отличный от антитела против NGF. В других вариантах осуществления изобретения набор включает любой антагонист NGF, отличный от иммуноадгезина TrkA и антитела против NGF. В некоторых вариантах осуществления изобретения набор включает атитело против NGF (такое как антитело Е3, описанное в настоящем описании изобретения). В других вариантах осуществления изобретения набор включает антитело против NGF, содержащее один или несколько гипервариабельных участков антитела Е3 (в частности, один, два, три, четыре, пять или в некоторых вариантах осуществления изобретения все шесть гипервариабельных участков антитела Е3). В некоторых вариантах осуществления изобретения набор включает наркотический аналгетик. В некоторых вариантах осуществления изобретения набор включает NSAID. В некоторых вариантах осуществления изобретения набор не включает наркотический аналгетик. В некоторых вариантах осуществления изобретения набор не включает NSAID. В некоторых вариантах осуществления изобретения прилагаемые инструкции содержат описание введения антагониста NGF для лечения, ослабления или предотвращения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость, в соответствии с любыми способами, описанными в настоящем описании изобретения. В инструкции далее должно быть указано, что лечению с помощью данного набора подлежат субъекты, у которых выявлена боль, обусловленная раком кости, включая боль, ассоциированную с метастазированием рака в кость, или субъекты с повышенным риском возникновения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость. В других вариантах осуществления изобретения инструкция включает описание введения антагониста NGF для лечения, предотвращения и/или ослабления боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость. В других вариантах осуществления изобретения инструкции включают описание введения антагониста NGF субъекту с повышенным риском возникновения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят вместе с наркотическим аналгетиком и NSAID. В некоторых вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика. В некоторых вариантах осуществления изобретения антагонист NGF вводят без NSAID.
Инструкции, относящиеся к применению антагониста NGF, обычно включают информацию о дозировке, схеме применения и способе введения для проведения необходимого лечения. Контейнеры могут содержать лекарственное средство в дозах на один прием, нерасфасованное лекарственное средство (например, упаковку лекарственного средства для многократного приема) или лекарственное средство в разделенных унифицированных дозах. Инструкции, прилагаемые к наборам по настоящему изобретению, обычно напечатаны на этикетке или вкладыше (например, на листе бумаги, вложенном в набор), но также приемлемы машинно-читаемые инструкции (например, инструкции на магнитном или оптическом диске).
На этикетке или вкладыше должно быть указано, что данная композиция предназначена для лечения, ослабления и/или предотвращения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость. Инструкции могут быть предназначены для осуществления любых способов, описанных в настоящем описании изобретения.
Наборы по настоящему изобретению находятся в приемлемой упаковке. Приемлемые упаковки включают, не ограничиваясь ими, флаконы, бутылки, банки, гибкие упаковки (например, запечатанные миларные или пластиковые пакеты) и тому подобные. В объем настоящего изобретения входят также упаковки, предназначенные для использования в комбинации со специальным устройством, таким как ингалятор, устройство для введения в нос (например, аэрозольный ингалятор) или устройство для вливания, такое как мини-насос. Набор может иметь стерильное входное отверстие (например, контейнер может представлять собой баллон или флакон с раствором для внутривенного вливания, снабженный пробкой, прокалываемой иглой для подкожных инъекций). Контейнер может также иметь стерильное входное отверстие (например, контейнер может представлять собой баллон или флакон с раствором для внутривенного вливания, снабженный пробкой, прокалываемой иглой для подкожных инъекций). По крайней мере один активный агент в композиции является антагонистом NGF, таким как антитело против NGF. Контейнер может далее содержать второй фармацевтически активный агент.
Наборы могут необязательно включать дополнительные компоненты, такие как буферы и разъясняющую информацию. Набор обычно включает контейнер и прилагаемую к нему этикетку или вкладыш.
Некоторые варианты осуществления изобретения относятся к изделию, включающему содержимое вышеописанных наборов. В некоторых вариантах осуществления изобретения наборы включают антагонист NGF (такой как антитело против NGF) вместе с информацией, описывающей применения данного набора для лечения боли, обусловленной раком кости, включая боль, ассоциированную с метастазированием рака в кость.
Введение антагониста NGF и оценка лечения
Антагонист NGF можно вводить субъекту любым приемлемым способом. Например, антагонист NGF можно вводить перорально, внутривенно, подъязычно, подкожно, внутриартериально, внутрисуставно, в мочевой пузырь (например, при помощи шприца для промывания мочевого пузыря), внутримышечно, интракардиально, интраторакально, внутрибрюшинно, интравентрикулярно, путем ингаляции, в виде суппозитория и трансдермально. Антагонист NGF можно вводить перорально, например, в форме таблеток, пастилок, капсул, эликсиров, суспензий, сиропов, облаток, леденцов, жевательной резинки или в подобных формах, которые получают методами, известными в данной области. Специалисту в данной области должно быть очевидно, что примеры, приведенные в данном описании изобретения, не ограничивают настоящее изобретение, а лишь иллюстрируют существующие методы.
В некоторых вариантах осуществления изобретения антагонист NGF, такой как антитело против NGF, вводят субъекту известными методами, такими как внутривенное введение, например, в виде болюса или непрерывного вливания в течение определенного периода времени, внутримышечное, внутрибрюшинное, интрацереброспинальное, подкожное, внутрисуставное, интратекальное, пероральное введение, путем ингаляции или местно. Для введения можно использовать коммерчески доступные распылители для жидких препаратов, в том числе струйные распылители и ультразвуковые распылители. Жидкие препараты можно распылять сразу, а лиофилизированный порошок сначала восстанавливают и затем распыляют. Альтернативно антагонист NGF можно распылять в виде аэрозоля при помощи дозирующего ингалятора или ингалировать в виде лиофилизированного или измельченного порошка.
В одном варианте осуществления изобретения антагонист NGF вводят методами сайтспецифической или направленной локальной доставки. Примеры методов сайтспецифической или направленной локальной доставки включают разные имплантируемые депо, содержащие антагонист NGF, или катетеры для локальной доставки, такие как катетеры для вливания, постоянный катетер или игольчатый катетер, синтетические трансплантаты, адвентициальные повязки, шунты и стенты или другие имплантируемые устройства, сайтспецифические носители, прямые инъекции или прямое наложение. См., например, публикацию РСТ № WO 00/53211 и патент США № 5981568.
Для введения можно использовать разные препараты, содержащие антагонист NGF (такой как антитело против NGF). В некоторых вариантах осуществления изобретения антагонист NGF можно вводить в чистом виде. В некоторых вариантах осуществления изобретения антагонист NGF является антителом против NGF и может входить в состав разных препаратов, включая препараты, содержащие фармацевтически приемлемый эксципиент. Фармацевтически приемлемые эксципиенты известны в данной области и представляют собой относительно инертные вещества, облегчающие введение фармакологически эффективного вещества. Например, эксципиент может сообщать нужную форму или консистенцию или действовать в качестве разбавителя. Приемлемые эксципиенты включают, не ограничиваясь ими, стабилизаторы, смачивающие вещества и эмульгаторы, соли для изменения осмотической концентрации раствора, инкапсулирующие агенты, буферы и усилители проникновения через кожу. Эксципиенты, а также препараты для парентерального и непарентерального введения лекарственного средства представлены в публикации Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing (2000).
В некоторых вариантах осуществления изобретения указанные лекарственные средства получают в форме, пригодной для введения путем инъекции (например, внутрибрюшинной, внутривенной, подкожной, внутримышечной инъекции и т.д.). Поэтому указанные лекарственные средства могут быть объединены с фармацевтически приемлемыми носителями, такими как физиологический раствор, раствор Рингера, раствор декстрозы и тому подобные. Определенная схема приема лекарственного средства, то есть доза, время введения и повторное введение, зависит от конкретного субъекта и его истории болезни.
Антитело против NGF можно вводить любым приемлемым способом, в том числе в виде инъекции (например, внутрибрюшинно, внутривенно, подкожно, внутримышечно и т.д.). Антитела против NGF можно также вводить путем ингаляции, описанной в настоящем описании изобретения. Начальная доза для введения антител против NGF обычно составляет около 2 мг/кг. В соответствии с целями настоящего изобретения обычная суточная доза может составлять от около 0,1 мкг/кг до 3 мкг/кг, до 30 мкг/кг, до 300 мкг/кг, до 3 мг/кг, до 30 мг/кг до 100 мг/кг или больше в зависимости от факторов, указанных в настоящем описании изобретения. При повторном введении на протяжении нескольких дней или более продолжительного срока в зависимости от состояния субъекта лечение продолжают до желаемого подавления симптомов или до достижения достаточной терапевтической концентрации, необходимой для ослабления боли, ассоциированной с метастазированием рака в кость. Типичная схема приема лекарственного средства включает введение начальной дозы, равной примерно 2 мг/кг, с последующим введением еженедельной поддерживающей дозы, равной примерно 1 мг/кг антитела против NGF, или с последующим введением поддерживающей дозы, равной 1 мг/кг, через одну неделю. Однако можно использовать другие схемы приема лекарственного средства в зависимости от характера фармакокинетического распада, которого хотел бы достичь лечащий врач. Например, лекарственное средство можно вводить один-четыре раза в неделю. В некоторых вариантах осуществления изобретения можно использовать дозу в пределах от около 3 мкг/мг до около 2 мг/кг (в частности, около 3 мкг/мг, около 10 мкг/мг, около 30 мкг/мг, около 100 мкг/мг, около 300 мкг/мг, около 1 мг/кг и около 2 мг/кг). В некоторых вариантах осуществления изобретения лекарственное средство можно вводить один раз в неделю, один раз в 2 недели, один раз в 4 недели, один раз в 5 недель, один раз в 6 недель, один раз в 7 недель, один раз в 8 недель, один раз в 9 недель или один раз в 10 недель; либо один раз месяц, один раз в 2 месяца, один раз в 3 месяца или реже. Прогресс, достигаемый при данном лечении, можно легко контролировать при помощи обычных методов и анализов. Схема приема лекарственного средства (включая использованные антагонисты NGF) может изменяться с течением времени.
Антагонист NGF, не являющийся антителом (в некоторых вариантах осуществления изобретения), обычно вводят в количестве, равном около 0,1-300 мг/кг массы тела субъекта, в виде одной-трех доз или в соответствии с описанием, приведенным в настоящем описании изобретения. В некоторых вариантах осуществления изобретения взрослому субъекту с нормальной массой тела можно вводить дозы, равные 0,3-5,00 мг/кг. Определенная схема приема лекарственного средства, то есть доза, время введения и повторность введения, зависит от конкретного субъекта и его истории болезни, а также от свойств лекарственных средств (таких как период полувыведения лекарственного средства и других факторов, хорошо известных в данной области).
В соответствии с целями настоящего изобретения соответствующая доза антагониста NGF зависит от используемых антагонистов NGF (или содержащих их композиций), типа и тяжести боли, подлежащей лечению, профилактической или терапевтической цели введения данного лекарственного средства, предшествующего лечения, клинической истории болезни субъекта, реакции на данное лекарственное средство и решения лечащего врача. Антагонист NGF, такой как антитело против NGF, обычно вводят до достижения дозы, позволяющей получить требуемый результат.
При определении дозы обычно учитывают эмпирические данные, такие как период полувыведения. Например, для увеличения периода полувыведения антитела и предотвращения разрушения данного антитела иммунной системой хозяина можно использовать антитела, совместимые с иммунной системой человека, в частности гуманизированные антитела или полностью человеческие антитела. Частоту введения можно определить и отрегулировать в процессе лечения с учетом результатов лечения, подавления, ослабления и/или замедления появления боли. Альтернативно можно использовать препараты на основе антител против NGF пролонгированного действия. В данной области известны разные препараты и устройства для пролонгированного высвобождения лекарственного средства.
В одном варианте осуществления изобретения дозы антагониста NGF могут быть определены эмпирически для субъектов, которым один или несколько раз вводили антагонист NGF (такой как антитело). Субъектам обычно вводят увеличивающиеся дозы антагониста NGF, например антитела против NGF. Эффективность антагониста NGF можно определить по индикации боли.
Антагонист NGF в соответствии со способом по настоящему изобретению можно вводить постоянно или периодически в зависимости, например, от физиологического состояния реципиента, терапевтической или профилактической цели введения и других факторов, известных квалифицированным лечащим врачам. Антагонист NGF (например, если антагонистом NGF является антитело против NGF) можно вводить постоянно в течение заранее определенного периода времени или в виде серии доз, вводимых через некоторые промежутки времени, например до, во время или после возникновения боли; во время, до или после возникновения боли; во время и после возникновения боли; до и во время возникновения боли; до, во время или после возникновения боли. Антагонист NGF можно вводить до, во время и/или после метастазирования рака в кость и любого другого события, вызывающего возникновение боли, ассоциированной с метастазированием рака в кость.
В некоторых вариантах осуществления изобретения можно вводить несколько антагонистов NGF, таких как антитело. Антагонисты могут быть одинаковыми или разными. Можно вводить по крайней мере один, по крайней мере два, по крайней мере три, по крайней мере четыре, по крайней мере пять разных антагонистов NGF. Такие антагонисты NGF обычно обладают комплементарным действием, но не оказывают вредного воздействия друг на друга. Антагонисты NGF можно также использовать в сочетании с другими лекарственными средствами, служащими для усиления и/или дополнения эффективности данных агентов. В некоторых вариантах осуществления изобретения антагонист NGF вводят без наркотического аналгетика. В некоторых вариантах осуществления изобретения антагонист NGF вводят без NSAIS.
Терапевтические препараты, содержащие антагонист NGF (такой как антитело), используемый по настоящему изобретению, получают с возможностью хранения, смешивая антитело, характеризующееся требуемой степенью чистоты, с необязательными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington, The Science and Practice 20th Ed. Mack Publishing (2000)) в форме лиофилизированных препаратов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы должны быть нетоксичными для реципиентов в используемых дозах и концентрациях и могут включать буферы, такие как фосфат, цитрат и другие органические кислоты; соли, такие как хлорид натрия; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония, хлорид гексаметония, хлорид бензалкония, хлорид бензетония, феноловый, бутиловый или бензиловый спирт, алкилпарабены, такие как метил- или пропилпарабен, катехол, резорцинол, циклогексанол, 3-пентанол и м-крезол); низкомолекулярные (менее примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатообразователи, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например Zn-белковые комплексы) и/или неионогенные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG).
Липосомы, содержащие антагонист NGF (такой как антитело), получают методами, известными в данной области, такими как описанные в публикациях Epstein et al., Proc. Natl. Acad. Sci. USA 82: 3688 (1985); Hwang et al., Proc. Natl. Acad. Sci. USA 77:4030 (1980); и в патентах США №№ 4485045 и 4544545. Липосомы с более продолжительным временем циркуляции в крови описаны в патенте США № 5013556. Особенно пригодные липосомы могут быть получены методом упаривания с обращенной фазой с использованием липидной композиции, содержащей фосфатидилхолин, холестерин и PEG-производный фосфатидилэтаноламин (PEG-PE). Липосомы экструдируют через фильтры с определенным размером отверстий, получая при этом липосомы с требуемым диаметром.
Активные ингредиенты могут быть также заключены в микрокапсулы, полученные, например, методами коацервации или межфазной полимеризации, такие как гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакралатные микрокапсулы, в коллоидные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методы описаны в публикации Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing (2000).
Могут быть получены препараты пролонгированного действия. Приемлемые примеры препаратов пролонгированного действия включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, которые имеют определенную форму, например, пленок или микрокапсул. Примеры матриц пролонгированного действия включают сложные полиэфиры, гидрогели (например, поли-2-гидрокси-этилметакрилат или поливиниловый спирт), полиактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и 7-этил-L-глутамата, неразлагающийся этиленвинилацетат, разлагающиеся сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (инъецируемые микросферы из сополимера молочной кислоты и гликолевой кислоты и ацетата лейпролида), изобутират ацетата сахарозы и поли-D-(-)-3-гидроксимасляную кислоту.
Препараты, предназначенные для введения in vivo, должны быть стерильными. Указанные препараты можно легко стерилизовать, например, путем фильтрации через стерильные фильтрующие мембраны. Терапевтические композиции, содержащие антитело против NGF, обычно помещают в контейнер со стерильным входным отверстием, например в баллон или флакон с раствором для внутривенного вливания, который снабжен пробкой, прокалываемой иглой для подкожных инъекций.
Композиции по настоящему изобретению могут быть получены в виде единичных дозированных форм, таких как таблетки, пилюли, капсулы, порошки, гранулы, растворы, суспензии или суппозитории, предназначенных для перорального, парентерального или ректального введения либо для введения путем ингаляции или инсуффляции.
Для получения твердых композиций, таких как таблетки, основной активный ингредиент смешивают с фармацевтическим носителем, например с обычными таблетирующими ингредиентами, такими как кукурузный крахмал, лактоза, сахароза, сорбит, тальк, стеариновая кислота, стеарат магния, дикальцийфосфат или камеди, и другими фармацевтическими разбавителями, такими как вода, с образованием твердой предварительно полученной композиции, содержащей гомогенную смесь соединения по настоящему изобретению или его нетоксичную фармацевтически приемлемую соль. Определение предварительно полученной композиции как гомогенной означает, что активный ингредиент равномерно диспергирован в такой композиции, которая может быть легко разделена на одинаковые единичные дозированные формы, такие как таблетки, пилюли и капсулы. Твердую предварительно полученную композицию затем делят на единичные дозированные формы вышеописанного типа, содержащие около 0,1-500 мг активного ингредиента по настоящему изобретению. Таблетки или пилюли, полученные из композиции по настоящему изобретению, могут быть покрыты оболочкой или обработаны каким-либо другим образом с образованием единичной дозированной формы, обеспечивающей пролонгированное действие. Например, таблетка или пилюля может содержать внутренний и наружный дозированный компонент, причем второй компонент может служить оболочкой для первого компонента. Оба компонента могут быть разделены энтеросолюбильным слоем, препятствующим распаду таблетки в желудке и обеспечивающим прохождение интактного внутреннего компонента в двенадцатиперстную кишку или пролонгированное высвобождение. Для изготовления таких энтеросолюбильных слоев или покрытий можно использовать разные вещества, которые включают ряд полимерных кислот и смесей полимерных кислот с такими веществами, как шеллак, цетиловый спирт и ацетат целлюлозы.
Приемлемые поверхностно-активные вещества включают, в частности, неионогенные вещества, такие как полиоксиэтиленсорбиты (например, Tween™ 20, 40, 60, 80 или 85) и другие сорбиты (например, Span™ 20, 40, 60, 80 или 85). Композиции с поверхностно-активным веществом обычно включают 0,05-5% или 0,1-2,5% поверхностно-активного вещества. Вполне очевидно, что в указанные композиции при необходимости могут быть добавлены другие ингредиенты, например маннит и другие фармацевтически приемлемые носители.
Приемлемые эмульсии могут быть получены с использованием коммерчески доступных жировых эмульсий, таких как интралипид™, липозим™, инфонутрол™, липофундин™ и липифизан™. Активный ингредиент может быть растворен в предварительно смешанной эмульсии или альтернативно может быть растворен в масле (например, соевом, сафлоровом, хлопковом, кунжутном, кукурузном или миндальном масле), и эмульсию получают в результате смешивания с фосфолипидом (таким как яичные фосфолипиды, соевые фосфолипиды или соевый лецитин) и водой. Совершенно очевидно, что в эмульсию для повышения тонуса могут быть добавлены другие ингредиенты, например глицерин или глюкоза. Приемлемые эмульсии обычно содержат до 20% масла, например от 5% до 20%. Жировая эмульсия может включать капли жира размером 0,1-1,0 мкм, в частности 0,1-0,5 мкм, и иметь значение рН в пределах от 5,5 до 8,0.
Эмульсионные композиции могут быть получены при смешивании антагониста фактора роста нервов с интралипидом™ или его компонентами (соевое масло, яичные фосфолипиды, глицерин и вода).
Композиции для ингаляции или инсуффляции включают растворы и суспензии в фармацевтически приемлемых водных или органических растворителях или их смесях и порошки. Жидкие или твердые композиции могут содержать вышеуказанные фармацевтически приемлемые эксципиенты. В некоторых вариантах осуществления изобретения указанные композиции вводят в дыхательные пути через рот или нос для достижения местного или системного действия. Композиции в предпочтительно стерильных фармацевтически приемлемых растворителях можно распылять с помощью газов. Нуждающийся субъект может вдыхать распыляемые растворы непосредственно из распыляющего устройства, либо распыляющее устройство может быть присоединено к маске для лица, палатке или устройству для дыхания под положительным давлением. Композиции в виде раствора, суспензии или порошка можно вводить предпочтительно перорально или назально из устройств, доставляющих препарат соответствующим образом.
Эффективность лечения можно определить методами, хорошо известными в данной области.
Примеры
Нижеследующие примеры приведены только для иллюстрации и не ограничивают объем изобретения.
Пример 1
Эффективность моноклонального антитела против NGF при лечении боли, ассоциированной с метастазированием рака в кость
Для определения эффективности лечения антителом против NGF 911 (моноклональное антитело мыши; см. публикацию Hongo et al., Hybridoma 19:215-227 (2000)) была использована модель боли, обусловленной раком кости, у мышей. Указанная модель боли, обусловленной раком кости, у мышей была получена в результате интрамедуллярной инъекции клеток остеолитической саркомы в бедренную кость мыши, после чего отверстие от иглы заполняли амальгамой для зубной пломбы для удерживания опухоли в кости. См. публикации Schwei et al., Neuroscience 19:10886-10897 (1999) и Luger et al., Pain 99:397-406 (2002). Эксперименты выполняли с использованием взрослых самцов мышей С3Н/HeJ (Jackson Laboratories, Bar Harbor, ME). В 0-й день мышей анестезировали пентобарбиталом натрия (50 мг/кг, внутрибрюшинно (i.p.)) и выполняли артротомию. Иглу вводили в медуллярный канал, создавая проход для клеток саркомы. Затем с помощью пневматического высокоскоростного наконечника для бормашины делали углубление. Помимо незараженных животных (n=5) использовали псевдозараженных животных (n=5), которым инъецировали α-минимальную основную среду (20 мкл, Sigma, St. Louis. MO) в интрамедуллярную область бедренной кости (получившую название “псевдоагент”), в то время как животным, зараженным саркомой (n=5 для каждого испытанного состояния), инъецировали среду, содержащую 105 2472 клетки остеолитической саркомы (получившую название “саркома”) (20 мкл, АТСС, Rockville, MD). У всех животных место инъекции закрывали амальгамой для зубной пломбы для удерживания клеток или инъецированной среды в интрамедуллярном канале и затем промывали стерильной водой (гипотонический раствор). На место разреза накладывали скобки. Скобки удаляли на 5-й день, чтобы они не мешали определению поведенческой реакции. Второй группе животных, которым инъецировали саркому, вводили антитело против NGF (10 мг/кг, внутрибрюшинно) на 6-й и 13-й день.
Поведенческий анализ. Поведение животных, связанное с болью, исследовали на 10-й и 14-й день после имплантации опухоли. Поведение животных исследовали в результате выполнения нижеследующих испытаний: постоянная боль (спонтанная защитная фиксация и подрагивание); боль, возникающая при ходьбе (использование конечности или ротарод), и боль, возникающая при движении (защитная фиксация и подрагивание, вызываемые пальпацией). Животных помещали в прозрачный пластиковый ящик для наблюдения с полом из металлической сетки, где они находились в течение 30 минут. После адаптации у животных определяли спонтанную защитную фиксацию, спонтанное подрагивание, использование конечности во время нормальной ходьбы на открытом пространстве и спонтанную защитную фиксацию во время принудительной ходьбы. Защитную фиксацию и подрагивание, вызываемые пальпацией, измеряли после двухминутной обычно безболезненной пальпации дистального отдела бедренной кости у животных, которым инъецировали саркому и псевдоагент.
Число спонтанных подрагиваний и время защитной фиксации, характерные для ноцицептивного поведения, регистрировали одновременно в течение двухминутного периода наблюдения. Защитную фиксацию определяли как время нахождения задней лапы в приподнятом положении, в то время как боль при ходьбе и подрагивания определяли как число поднятий конечности животным.
Нормальное использование конечности во время спонтанной ходьбы оценивали по шкале от 5 до 0: (5) нормальное использование и (0) полное отсутствие использования конечности.
Защитную фиксацию при принудительной ходьбе определяли с помощью ротарода (Columbus Instruments, Columbus, OH). Ротарод имеет вращающийся валик и оснащен органами управления скоростью, ускорением и восприятием. Животных помещали на валик с 4-кратной скоростью вращения, ускорением 8,0 и восприятием 2,5. Защитную фиксацию при принудительной ходьбе оценивали по шкале 5-0: (5) нормальное использование и (0) полное отсутствие использования конечности.
После обычно безболезненной пальпации дистального отдела бедренной кости, выполняемой каждую секунду в течение 2 минут, животных помещали в ящик для наблюдения и в течение еще 2 минут измеряли защитную фиксацию и подрагивание, вызванные пальпацией.
Лечение антителом против NGF. На 6-й и 13-й день животным, которым инъецировали саркому, внутрибрюшинно (i.p.) инъецировали антитело против NGF 911 в количестве 10 мг/кг (саркома + антитело против NGF, n=5) или животным, которым инъецировали саркому и псевдоагент, внутрибрюшинно инъецировали физиологический раствор (псевдоагент + носитель или саркома + носитель, n=5 для каждого состояния). У всех животных анализировали поведение на 10-й и 14-й день.
Оценка поведения при постоянной боли. Как показано на Фиг.1, у животных, которым инъецировали саркому (с физиологическим раствором), возникало поведение, характерное для постоянной боли, которое определяли по спонтанной защитной фиксации и спонтанному подрагиванию (для обоих показателей р<0,05, ANOVA) по сравнению с животными, которым инъецировали псевдоагент (с физиологическим раствором). На Фиг.1 также показано, что внутрибрюшинное введение антитела против NGF 911 значительно уменьшало спонтанную защитную фиксацию и спонтанное подрагивание у мышей, которым инъецировали саркому, на 10-й и 14-й день после имплантации саркомы по сравнению с введением физиологического раствора мышам, которым инъецировали саркому (р<0,05, ANOVA, как для спонтанной защитной фиксации, так и для спонтанного подрагивания). Полученные результаты свидетельствуют о том, что антитело против NGF 911 ослабляет постоянную боль у мышей, которым инъецировали саркому.
Оценка поведения при боли, возникающей при ходьбе. Как показано на Фиг.2, у животных, которым инъецировали саркому (с физиологическим раствором), возникало поведение, характерное для боли при ходьбе, которое определяли по использованию конечности и защитной фиксации при принудительной ходьбе (ротарод) (для обоих показателей р<0,05, ANOVA), по сравнению с животными, которым инъецировали псевдоагент (с физиологическим раствором). На Фиг.2 также показано, что внутрибрюшинное введение антитела против NGF 911 значительно повышало оценку использования конечности (близко к нормальной) и оценку защитной фиксации при принудительной ходьбе у мышей, которым инъецировали саркому, на 10-й и 14-й день после имплантации саркомы по сравнению с введением физиологического раствора мышам, которым инъецировали саркому (р<0,05, ANOVA, как для использования конечности, так и для защитной фиксации при принудительной ходьбе). Полученные результаты свидетельствуют о том, что антитело против NGF 911 ослабляет боль при ходьбе у мышей, которым инъецировали саркому.
Оценка поведения при боли, вызванной прикосновением. Как показано на Фиг.3, у животных, которым инъецировали саркому (с физиологическим раствором), возникало поведение, характерное для боли, вызываемой прикосновением, которое определяли по защитной фиксации и подрагиванию, индуцированным пальпацией (для обоих показателей р<0,05, ANOVA), по сравнению с животными, которым инъецировали псевдоагент (с физиологическим раствором). На Фиг.3 также показано, что внутрибрюшинное введение антитела против NGF 911 значительно уменьшает защитную фиксацию и подрагивание, вызванные пальпацией, у мышей, которым вводили саркому, на 10-й и 14-й день после имплантации саркомы по сравнению с введением физиологического раствора мышам, которым инъецировали саркому (р<0,05, ANOVA, как для защитной фиксации, так и подрагивания, вызванным пальпацией). Полученные результаты свидетельствуют о том, что антитело против NGF 911 уменьшает боль, вызванную прикосновением, у мышей, которым инъецировали саркому.
Пример 2
Эффективность моноклонального антитела против NGF при лечении боли, обусловленной раком кости, и уменьшении нескольких нейрохимических изменений, ассоциированных с сенсибилизацией периферических и центральных нейронов в ганглии задних корешков и спинном мозге
Применяемые методы
Животные. Эксперименты выполняли с использованием в общей сложности 158 взрослых самцов мышей С3Н/HeJ (Jackson Laboratories, Bar Harbor, ME) с массой тела 20-25 г. Мышей содержали в соответствии с требованиями Национальных институтов здравоохранения в специальных стерильных условиях в стерилизованных клетках при 22°С с 12-часовым циклом чередования света и темноты и давали стерилизованную в автоклаве пищу и воду ad libitum.
Культивирование и инъецирование опухолевых клеток. Полученные клетки остеолитической саркомы мышей (NCTC 2472, ATCC, Rockville, MD) устойчиво транфицировали зеленым флуоресцирующим белком (GFP) в соответствии с описанием, приведенным в публикации Sabino et al., Cancer Res. 62:7343-9 (2002).
Опухолевые клетки инъецировали в соответствии с описанием, приведенным в научной литературе. См. публикации Honore et al., Nat. Med. 6:521-8 (2000); Honore et al., Neuroscience 98:585-598 (2000); Luger et al., Cancer Research 61:4038-4047 (2001). Мышей анастезировали пентобарбиталом натрия (50 мг/кг, внутрибрюшинно) и выполняли артротормию, обнажая мыщелок дистального отдела бедренной кости. В интрамедуллярную область бедренной кости мыши инъецировали забуференный стерильный физиологический раствор Хэнка (HBSS, Sigma Chemical Co., St. Louis, MO; 20 мкл; псевдоагент, n=40) или среду, содержащую 105 клеток остеолитической саркомы мышей (20 мкл, NCTC 2472, ATCC, Rockville, MD; саркома, n=90), место инъекции закрывали амальгамой для зубной пломбы (Dentsply, Milford, DE) и промывали стерильной фильтрованной водой. Эксперимент выполняли в течение 14 дней, так как именно в течение данного периода времени опухоль все еще удерживается в кости, в максимальной степени проявляется поведение, характерное для боли, обусловленной раком, и наблюдаются максимальные изменения в экспрессии нейрохимических маркеров сенсибилизации периферических и центральных нейронов. Для контрольного анализа нейрохимических изменений и гистологии кости использовали псевдозараженных животных, так как незараженные животные существенно не отличались в поведенческом, нейрохимическом или гистологическом отношении.
Лечение антителом против NGF. Для оценки воздействия лечения антителом против NGF на поведение, обусловленное болью, нейрохимические изменения, рост опухоли и разрушение кости мышам вводили антитело против NGF (моноклональное антитело 911, описанное в публикации Hongo, et al., Hybridoma 19:215-227 (2000)) (10 мг/кг/каждые 5 дней, внутрибрюшинно); введение начинали через 6 дней после инъекции, когда были обнаружены первые признаки разрушения кости, и заканчивали через 14 дней после инъекции, когда наблюдалось значительное разрушение кости и поведение, обусловленное болью. Дозы, использованные в настоящем исследовании, не вызывали у незараженных мышей вредных эффектов, таких как гипоальгезия. Общее состояние здоровья мышей контролировали путем регистрации массы тела в начале и конце экспериментов.
Мышей произвольно распределяли в экспериментальные группы и один раз в неделю вводили или стерильный физиологический раствор (псевдоагент + носитель: n=28; саркома + носитель: n=35; 1,4 мкг/г/каждые 5 дней, внутрибрюшинно), или антитело против NGF (псевдоагент + антитело против NGF: n=4; саркома + антитело против NGF: n=23, 10 мг/кг/каждые 5 дней, внутрибрюшинно). Для сравнения поведения мышей при введении антитела против NGF и сульфата морфина мышам вводили дозу морфина за 15 минут до исследования поведения (незараженные мыши: n=6; псевдоагент + носитель: n=8; саркома + носитель: n=8; саркома + антитело против NGF: n=8; саркома + морфин, 10 мг/кг, внутрибрюшинно: n=8; саркома + морфин, 30 мг/кг, внутрибрюшинно: n=8). Для испытания тепловой и механической чувствительности и оценки иннервации кожи задней лапы мышей распределяли в две экспериментальные группы и вводили или стерильный физиологический раствор (незараженные мыши + носитель: n=11), или антитело против NGF (незараженные мыши + антитело против NGF: n=11, 10 мг/кг/каждые 5 дней, внутрибрюшинно) один раз в неделю в течение 2 недель.
Исследование антитела против NGF. Антагонистическое антитело против NGF (моноклональное антитело 911) позволяет эффективно блокировать связывание NGF c рецепторами NGF TrkA и р75, ингибировать аутофосфорилирование TrkA и блокировать NGF-зависимое выживание сенсорных нейронов ганглия задних корешков. Hongo, et al., Hybridoma 19:215-227 (2000).
Эвтаназия и обработка ткани. Мышей умерщвляли на 14-й день после инъекции опухоли и производили обработку тканей для выполнения иммуногистохимического анализа спинного мозга, ганглия задних корешков (DRG) и кожи задней лапы в соответствии с описанием, приведенным в научной литературе. Honore et al., Nat. Med. 6:521-8 (2000); Luger et al., Cancer Research 61:4038-4047 (2001). У мышей производили обычно безболезненную механическую стимуляцию инъецированного колена за 1,5 часа до эвтаназии для индукции экспрессии с-Fos. Honore et al., Neuroscience 98:585-598 (2000); Hunt et al., Nature 328:632-634 (1987). После вышеуказанной манипуляции мышей умерщвляли с помощью СО2 и перфузировали в сердце 12 мл 0,1 М физиологического раствора с фосфатным буфером (PBS) и затем 25 мл раствора 4% формальдегида/12,5% пикриновой кислоты.
Удаляли сегменты спинного мозга (L2-L4), DRG (L1-L5) и кожу подошвы, фиксировали в фиксаторе для перфузии и подвергали криозащите в 30% сахарозы в течение 24 часов. На санном микротоме изготавливали последовательные срезы замороженного спинного мозга и кожи толщиной 60 мкм, помещали в PBS и обрабатывали в виде свободно плавающих срезов. Последовательные срезы DRG толщиной 15 мкм изготавливали на криостате и оттаивали для обработки на покрытых желатином предметных стеклах.
Изготовленные срезы DRG, спинного мозга и кожи подошвы промывали в PBS, инкубировали в блокирующем растворе (3% нормальной сыворотки осла (NDS), 0,3% тритона Х-100 в PBS) в течение 1 часа и затем инкубировали в течение ночи в первичном антителе. Срезы спинного мозга подвергали иммунному окрашиванию для обнаружения белка с-Fos (1:2000, Oncogene Research, San Diego, CA) и динорфина (поликлональное антитело против динорфина морской свинки, 1:1000, Neuromics, Minneapolis, MN). Срезы DRG подвергали иммунному окрашиванию для обнаружения фактора активации транскрипции 3 (ATF-3) (поликлональное антитело против ATF-3 кролика, 1:500, Santa Cruz Biotechnologies, Santa Cruz, CA) и CD68 (ED-1; поликлональное антитело против CD68 крысы, 1:5000, Serotec, Raleigh, NC). Срезы кожи подвергали иммунному окрашиванию для обнаружения пептида, генетически родственного кальцитонину (CGRP) (1:15000, Sigma, St. Louis, MO), тирозингидроксилазы (ТОН) (поликлональное антитело против ТОН кролика, 1:2000, Chemicon, Temecula, CA) и нейрофиламента Н (клон RT97) (поликлональное антитело против RT-97 кролика, 1:2500, Chemicon, Temecula, CA).
После инкубации в первичном антителе срезы промывали в PBS и затем инкубировали в растворе вторичного антитела в течение 3 часов. Вторичные антитела, конъюгированные с Су3 или биотином (Jackson ImmunoResearch, West Grove, PA), использовали в отношении соответственно 1:600 или 1:500. Для обнаружения вторичных антител, конъюгированных с биотином, после инкубации в растворе вторичного антитела срезы промывали в PBS и инкубировали в стрептавидине, конъюгированном с Су3 (1:4000; Jackson ImmunoResearch) в течение 45 минут. Для подтверждения специфичности первичных антител использовали контрольные образцы, в которых отсутствовало первичное антитело, или выполняли предварительную адсорбцию соответствующим синтетическим пептидом. После иммунного окрашивания срезы спинного мозга и кожи подошв помещали на покрытые желатином предметные стекла. Срезы кожи, спинного мозга и DRG дегидратировали в градиентах спирта (70, 90, 100%), очищали в ксилоле и наносили на покровные стекла DPX (Fluka, Switzerland).
После выполнения радиологического исследования на 14-й день правую (внутренний контрольный образец) и левую (пораженную опухолью) бедренную кость фиксировали в пикриновой кислоте и 4% формалине при 4°С в течение ночи и декальцифицировали в 10% EDTA (Sigma, St. Louis, MO) в течение не более 14 дней. Затем кости погружали в парафин. Изготавливали срезы боковой плоскости бедренной кости толщиной 5 мкм и окрашивали устойчивой к тартрату кислой фосфатазой (TRAP), гематоксилином и эозином (H&E) для визуализации гистологических признаков нормального костного мозга, опухоли, остеокластов и макрофагов. Для визуализации клеток саркомы при помощи флуоресцентной микроскопии срезы бедренной кости толщиной 5 мкм окрашивали антителом против зеленого флуоресцирующего белка (GFP) (антитело против GFP кролика, 1:6000, Molecular Probes, Eugene, OR). Окрашивание GFP выполняли с использованием системы TSA-Plus Cyanine 3 (PerkinElmer Life Sciences, Inc., Boston, MA), ранее описанной в публикации Sevcik et al., Pain 111: 169-80 (2004).
Иммуногистохимический анализ псевдозараженной и раковой бедренной кости выполняли с использованием декальцифицированных, погруженных в парафин последовательных срезов толщиной 14 мкм. Для амплификации Су3-меченных антител использовали систему амплификации сигнала тирамина (TSA)(Perkin Elmer life Sciences, Boston, MA). Эндогенные пероксидазы блокировали, инкубируя срезы в 2% пероксиде водорода в течение 1 часа. Затем срезы трижды промывали PBS в течение 10 минут и блокировали в ТSA-блокирующем буфере в течение 1 часа. После удаления блокирующего буфера добавляли первичную антисыворотку и инкубировали при комнатной температуре в течение ночи. Афферентные немиелинизированные и слабо миелинизированные сенсорные нервные волокна метили кроличьим поликлональным антителом против пептида, генетически родственного кальцитонину (CGRP) (1:15000, Sigma). Срезы трижды промывали в промывочном буфере TSA в течение 10 минут и затем в течение 45 минут инкубировали в стрептавидине HRP (1:4000). Затем срезы трижды промывали промывочным буфером ТSA в течение 10 минут. Бедренные кости подвергали воздействию тирамина, конъюгированного с CY3, (1:600) в течение 7 минут, дважды промывали промывочным буфером ТSA и один раз PBS. И наконец, срызы подвергали воздушной сушке, дегидратировали в градиенте спирта (70, 90 и 100%), очищали в ксилоле и обрабатывали DPX (Fluka).
Рентгенографический анализ пролиферации остеокласов и макрофагов в кости. Рентгенограммы (Faxitron X-ray Corl., Wheeling, IL) рассеченной бедренной кости получали на 14-й день для оптимальной оценки разрушения кости. Изображения были получены на пленке для маммографии Kodak Min-R 2000 (Eastman Kodak Co., Rochester, NY; регистрирующая экспозиция: 7 секунд, 21 кВ). Степень разрушения бедренной кости, индуцированной опухолью, оценивали рентгенологическим методом в боковой плоскости изображений цельной кости при 5-кратном увеличении по шкале 0-5 (0, нормальная кость без признаков разрушения и 5, бикортикальное разрушение кости по всей толщине). Honore et al., Nat. Med. 6:521-8 (2000); Honore et al., Neuroscience 98:585-598 (2000); Luger et al., Cancer Research 61:4038-4047 (2001).
Пролиферацию остеокластов и макрофагов, ассоциированных с опухолью, (ТАМ) измеряли путем количественного определения числа TRAP+ остеокластов или ТАМ на TRAP-окрашенных срезах бедренной кости в соответствии с описанием, приведенным в научной литературе. Honore et al., Nat. Med. 6:521-8 (2000); Honore et al., Neuroscience 98:585-598 (2000). Макрофаги, ассоциированные с опухолью, отделяли гистологически от остеокластов на срезах бедренной кости, окрашенных TRAP, в виде TRAP+ клеток, которые были свободно и многомерно диспергированы в опухолевой массе. Макрофаги в кости активировались под воздействием выделяемых опухолью факторов, стимулирующих клетки, и на появление в клетках активированных ТАМ указывали очень неправильная форма, множественная пластинчатость и фагоцитарные вакуоли. Остеокласты гистологически отделяли в виде TRAP+ клеток, которые были тесно связаны с участками резорбции кости. Указанные клетки являются многоядерными и располагаются вдоль кортикального слоя кости и губчатой кости. Результаты выражены в виде среднего числа остеокластов на мм или ТАМ на мм2.
Количественное определение роста опухоли. Изображения бедренной кости, содержащей GFP-экспрессирующие клетки саркомы, были получены с использованием желтого эмиссионного фильтра пропускания нижних частот длиной 515 нм в флуоресцентном микроскопе Nikon E600, оснащенном цифровой камерой SPOT II, при помощи программного обеспечения для получения изображений SPOT (Diagnostic Instruments, Sterling Heights, MI). Общую площадь интрамедуллярной области и процентное значение интрамедуллярной области, занимаемой опухолью, высчитывали при помощи программы обработки изображений Image Pro Plus, версия 3.0 (Media Cybernetics, Silver Spring, MD). Sabino et al., Cancer Res. 62:7343-9 (2002); Sevcik et al., Pain 111: 169-80 (2004). Характеристики опухоли, образуемой клетками саркомы, трансфицированными GFP, такие как скорость роста, скорость резорбции кости и способность индуцировать боль, обусловленную раком кости, были во временном, поведенческом и физическом отношении идентичны нетрансфицированным клеткам саркомы. Sabino et al., Cancer Res. 62:7343-9 (2002).
Количественное определение сенсорных нервных волокон в кости. Число сенсорных нервных волокон определяли в соответствии с описанием, приведенным в научной литературе. Mach et al., Neuroscience 113:155-66 (2002). Производили количественное определение числа CGRP-положительных волокон в трех отделах кости (проксимальный, дистальный и диафизарный) и в трех тканях кости (надкостница, минерализованная кость и костный мозг). При выполнении данного анализа использовали только нервные волокна длиной более 30 мкм. Анализировали по шесть срезов, полученных у каждого животного, при этом подсчитываемые волокна были экспрессированы на всей площади кости.
Количественное определение спинного мозга, ганглия задних корешков и кожи задней лапы. Срезы спинного мозга, DRG и ткани кожи метили флуоресцентным красителем и анализировали при помощи системы получения изображений конфокального микроскопа MRC 1024 (Bio-Rad, Philadelphia, РA) или цифровой камеры SPOT II в флуоресцентном микроскопе Olympus BX-60 с использованием программного обеспечения получения изображений SPOT (Diagnostic Instruments, Inc).
Число нейронов DRG, экспрессирующих фактор активации транскрипции 3 (ATF-3), подсчитывали при 200-кратном увеличении с 1 см2 окулярной сеткой. Общее число нейронов (мелких, средних и крупных) определяли, подсчитывая меченые и немеченые тела нейронов (немеченые клеточные тела характеризуются фоновым мечением, которое можно наблюдать через родаминовый или FITC фильтр); результаты были выражены в виде процентного значения от общего числа нейронов, обладающих иммунореактивностью в отношении ATF-3 (IR). Во избежание двойного подсчета тел нейронов подсчет производили на каждом четвертом последовательном срезе для каждого маркера. Для количественного определения активированных или инфильтрующихся макрофагов в DRG с помощью камеры SPOT были получены полутоновые изображения как минимум на четырех ипсилатеральных и контралатеральных срезах DRG каждого животного, которые анализировали с использованием программного обеспечения обработки изображений Image Pro Plus, версия 3.0 (Media Cybernetics). На каждом изображении были определены области DRG, содержащие только тела сенсорных нейронов (исключая периферический нерв). Во время просмотра изображений на мониторе верхний и нижних пороги полутонового представления изображений были заданы таким образом, чтобы на фоне выделенных DRG были различимы только специфические профили CD68-IR клеток. Число профилей клеток подсчитывали автоматически на каждом срезе. Выходной сигнал камеры SPOT был калиброван программным обеспечением Image Pro Plus таким образом, что можно было определить фактическую площадь каждого выделенного участка на полученных изображениях. Значения профилей CD68-IR клеток на срезах и выделенные площади суммировали для каждого животного и результаты выражали в виде общего числа профилей CD68-IR клеток на единицу площади (мм2).
Количественное определение срезов спинного мозга производили на уровне поясничного отдела L2-L4, так как данные сегменты спинного мозга получают значительный афферентный входной сигнал от ганглия задних корешков на уровне L2-L3, который является основным ганглием, направляющим афферентный входной сигнал в бедренную кость мыши. Edoff et al., Cell & Tissue Research 299:193-200 (2000); Molander C., J. Comp. Neurol. 260:246-255 (1987); Puigdellivol-Sanchez A et al., the Anatomical Record 260:180-188 (2000); Puigdellivol-Sanchez A et al., Neurosci. Lett. 251:169-172 (1998). Количественное определение динорфина на срезах спинного мозга производили на 4 произвольно выбранных фронтальных срезах спинного мозга на уровне L2-L4 каждого животного. Число динорфин-IR нейронов в слоях III-IV спинного мозга подсчитывали при 100-кратном увеличении и выражали в виде среднего числа нейронов на 60 мкм срезе L2-L4 каждого животного. Число с-Fos-иммунореактивных нейронов подсчитывали в слоях III-IV заднего рога на 8 произвольно выбранных фронтальных срезах спинного мозга на уровне L3-L4 каждого животного. Для определения иммунореактивности в отношении с-Fos порог профиля иммунофлуоресценции ядра был задан в три раза больше среднего уровня фоновой иммунофлуоресценции среза ткани. Результаты представлены в виде среднего числа с-Foc-IR нейронов на одном срезе спинного мозга.
Количественное определение плотности эпидермальной иннервации выполняли на 4 произвольно выбранных срезах кожи подошвы задней лапы одного животного. Общее число CGRP, TOH и RT97-IR нервных волокон подсчитывали при 200-кратном увеличении. В соответствии с правилами подсчета производили подсчет только отдельных интраэпидермальных волокон, а не многочисленных ответвлений одного волокна. McCarthy et al., Neurology 45:1848-55 (1995). Общую длину эпидермиса на всех срезах измеряли с использованием 1 см2 окулярной сетки. Подсчитывали только нервные волокна, имеющие длину, равную по крайней мере 25 мкм, и прорастающие в поверхностный эпидермис. Результаты представлены в виде среднего числа интраэпидермальных нервных волокон на мм длины одного животного.
Поведенческий анализ. Поведение мышей, характерное для боли, обусловленной раком кости, исследовали через 10 и 14 дней после инъекций псевдоагента или опухоли, когда такое поведение является особенно выраженным и позволяет произвести оценку эффективности лечения антителом против NGF. Антитело против NGF сравнивали с морфином (Baxter, Deerfield, IL; 10 мг/кг, внутрибрюшинно), который вводили за 15 минут до выполнения поведенческого анализа для гарантии исследования животных в пределах терапевтического действия лекарственного средства. Hasselstrom et al., Pharmacology & Toxicology 79:40-6 (1996).
Мышей исследовали также через 8, 10, 12 и 14 дней после инъекций опухоли или псевдоагента для оценки эффективности лечения антителом против NGF (10 мг/кг/каждые 5 дней, внутрибрюшинно), направленного на ослабление боли в процессе прогрессирования болезни. Животных наблюдали в течение 2 минут и анализировали постоянную и вызванную пальпацией боль, обусловленную раком кости, в соответствии с описанием, приведенным в научной литературе. Luger et al., Pain 99:397-406 (2002); Sabino et al., Cancer Res. 62:7343-9 (2002); Sabino et al., International Journal of Cancer 104:550-558 (2003). В качестве измерения постоянной боли регистрировали число подрагиваний и время защитной фиксации задней лапы, так как такие измерения отображают поведение субъектов, страдающих раком кости, которые стараются защитить или приподнять конечность, пораженную опухолью. В использованной модели боль, возникающую при движении, которую вызывали пальпацией конечности с инъецированной саркомой, оценивали при помощи ранее обоснованных испытаний. Luger et al., Cancer Research 61:4038-4047 (2001); Sabimo et al., International J. of Cancer 104:550-558 (2003); Sevcik et al., Pain 111:169-80 (2004). Поведение, характерное для боли, вызванной пальпацией, анализировали, подвергая животных обычно безболезненной пальпации конечности, в которую инъецировали опухоль или псевдоагент, в течение двух минут до исследования. Luger et al., Cancer Research 61:4038-4047 (2001); Sevcik et al., Pain 111:169-80 (2004). Мышей наблюдали в течение 2 минут и фиксировали число подрагиваний и время защитной фиксации. Анализы поведения, вызванного пальпацией, отражали клиническое состояние субъектов, испытывающих боль, обусловленную раком кости, после обычно безболезненного движения конечности, пораженной опухолью.
После адаптации в течение 15 минут у незараженных животных и незараженных животных, которым вводили антитело против NGF, измеряли тепловую и механическую чувствительность для оценки возможного изменения нормального порога болевой чувствительности после лечения антителом против NGF. Тепловую чувствительность измеряли при помощи теплового стимулятора лап (University of California, San Diego, CA). Интенсивность излучаемого тепла регулировали таким образом, чтобы незараженные животные реагировали на тепло поднятием задней лапы примерно через девять секунд после начала нагревания. Choi et al., Life Sci. 73:471-85 (2003). Мышей оставляли на 5 минут для восстановления между испытаниями. Одно испытание состояло из 4 измерений задней лапы, при этом самый продолжительный латентный период отбрасывали и результаты остальных 3 измерений усредняли. Механическую чувствительность измеряли при помощи ранее обоснованного метода. Chaplan et al., J. Neuroscience Methods 53:55-63 (1994). К задней лапе животных прикрепляли нити Фрея (Stoelting Co., Wood Dale, IL) и определяли порог отдергивания, увеличивая и уменьшая интенсивность раздражения в виде силы, эквивалентной 0,2-15,1 грамма. Положительную реакцию отмечали в том случае, если животное быстро отдергивало лапу.
Анализ методом RT-PCR уровней мРНК NGF в линии клеток 2472. Общую РНК из трех образцов ткани мыши или 2472 клеток саркомы получали с помощью микронабора RNeasy (Qiagen) в соответствии с инструкциями изготовителя и производили количественное определение РНК, используя реагент рибогрин (Molecular Probes). Выполняли двухстадийную реакцию RT-PCR, используя набор для RT-PCR TaqMan Gold (Applied Biosystems). РНК подвергали обратной транскрипции с помощью произвольных гексамеров и кДНК амплифицировали с помощью набора затравок/зондов, специфичных для NGF (muNGF-187F; GGGCTGGATGGCATGCT (SEQ ID NO:3), muNGF-256R: GCGTCCTTGGCAAAACCTT (SEQ ID NO:4), muNGF-208T: CCAAGCTCACCTCAGTGTCTGGGCC (SEQ ID NO:5)). Образцы, полученные на стадии обратной транскрипции, анализировали в двух экземплярах и нормализовали до получения полной РНК.
Статистический анализ. Для выполнения статистических анализов использовали пакет компьютерных программ для выполнения статистических расчетов SPSS, версия 11 (SPSS, Chicago, IL). Для анализа повторно измеренных данных использовали линейную регрессионную модель, которая позволяет объединять данные, измеренные в разные периоды времени, включать постоянные и изменяемые во времени коварианты и оценивать отдельные скорости изменения. Каждую зависимую переменную сравнивали в группах при помощи непараметрического анализа изменчивости (Kruskal-Wallis). Вслед за анализами Kruskal-Wallis выполняли непараметрические сравнения между парами групп при помощи U-критерия Mann-Whitney. Результаты считались статистически значимыми при Р<0,05. Во всех случаях исследователю не был известен экспериментальный статус каждого животного.
Результаты
Введение антитела против NGF не влияло на прогрессирование болезни или инфильтрацию макрофагов в кость. Воздействие лечения антителом против NGF на разрушение кости, пролиферацию остеокластов и рост опухоли исследовали на 14-й день после инъекции опухоли. У мышей, которым инъецировали псевдоагент, не наблюдалось значительного разрушения кости (оценка кости 0,9±0,4; Фиг.4а), пролиферации остеокластов (4,6±0,4 остеокласта/мм) или роста опухоли (Фиг.4d) при выполнении рентгенологических, TRAP и H&E/GFP анализов по сравнению с мышами, которым инъецировали саркому. У мышей, которым вводили саркому + носитель, происходили обширное разрушение кости, определяемое по множественным участкам просветления на рентгенограмме (оценка кости 3,5±0,2; Фиг.4b), значительное увеличение числа остеокластов (4,0±0,7 остеокласта/мм), при этом опухоль полностью заполняла интрамедуллярную область (100±0,0% интрамедуллярной области; Фиг.4е). Лечение мышей, пораженных опухолью, антителом против NGF с 6-го по 14-й день после инъекции опухоли не вызывало существенного изменения резорбции кости (3,1±0,6; Фиг.4с), не уменьшало пролиферацию остеокластов, индуцированную саркомой, (3,5±0,1 остеокласта/мм) или рост опухоли (98,0±0,9% интрамедуллярной области; Фиг.4f) по сравнению с животными, которым вводили саркому + носитель.
Через четырнадцать дней после инъекции опухоли у мышей, которым вводили саркому + носитель, было отмечено повышение ТАМ (39,8±12,6 ТАМ/мм2) по сравнению с контрольными мышами, которым вводили псевдоагент + носитель (0,0±0,0 ТАМ/мм2). Лечение мышей, которым инъецировали саркому, (29,5±7,3 ТАМ/мм2) антителом против NGF существенно не изменяло инфильтрацию ТАМ, наблюдаемую у мышей, которым вводили саркому + носитель.
Лечение антителом против NGF не влияло на иннервацию сенсорных или симпатических нейронов в кости или коже. Тонко миелинизированные или немиелинизированные пептидергические сенсорные нервные волокна метили антителом против пептида, родственного гену кальцитонина (CGRP). CGRP-иммунореактивные нервные волокна были обнаружены во всей кости (надкостница, минерализованная кость и костный мозг) как у незараженных мышей, которым вводили носитель, (12,2±0,3 волокна/мм), так и у незараженных мышей, которым вводили антитело против NGF (13,0±0,8 волокна/мм).
Тонко миелинизированные или немиелинизированные пептидергические сенсорные нервные волокна (CGRP-IR), крупные миелинизированные сенсорные волокна (RT97-IR) и норадренергические симпатические нервные волокна (ТОН-IR) анализировали в коже подошвы задней лапы при помощи антител против CGRP, RT-97 и ТОН. Не наблюдалось существенных различий между интенсивностью или плотностью CGRP-положительных волокон в образцах кожи задней лапы животных, которым вводили саркому + носитель, (12,0±0,8 волокна/мм) и животных, которым вводили саркому + антитело против NHF (12,5±0,6 волокна/мм) (фиг.5а и 5b). Аналогичным образом не наблюдалось различий в интенсивности и плотности CGRP-положительных волокон между незараженными мышами, которым вводили носитель (Фиг.5с, n=8), и незараженными мышами, которым вводили антитело против NGF (Фиг.5d, n=8). Не изменялось число нервных волокон, экспрессирующих CGRP у мышей, которым инъецировали саркому, и у незараженных мышей (а, b по сравнению с с, d). Не были обнаружены также различия в плотности и интенсивности RT97-положительных и ТОН-положительных волокон у животных, которым вводили саркому + носитель, (7,3±0,7 RT97+ волокна/мм; 3,1±0,7 ТОН+ волокна/мм) и у животных, которым вводили саркому + антитело против NGF (7,3±0,7 RT97+ волокна/мм; 3,6±0,7 ТОН+ волокна/мм). Аналогичным образом не было обнаружено существенных различий между интенсивностью или плотностью CGRP-положительных волокон в образцах кожи задней лапы незараженных мышей, которым вводили носитель, (12,5±0,5 волокна/мм) и незараженных мышей, которым вводили антитело против NGF (11,9±0,7 волокна/мм). Различия в плотности и интенсивности RT97-положительных и ТОН-положительных волокон не были также обнаружены у незараженных животных, которым вводили носитель, (10,4±0,4 RT97+ волокна/мм; 3,4±0,4 ТОН+ волокна/мм) и у незараженных животных, которым вводили антитело против NGF (11,9±0,7 RT97+ волокна/мм; 3,0±0,8 ТОН+ волокна/мм). Не наблюдалось существенных различий между интенсивностью или плотностью СGRP-, RT97- или ТОН-положительных волокон в образцах кожи животных, которым вводили саркому + носитель, и животных, которым вводили саркому + антитело против NGF, по сравнению с незараженными животными, которым вводили носитель, и незараженными животными, которым вводили антитело против NGF.
Лечение антителом против NGF значительно улучшало поведение, характерное для боли, обусловленной раком кости. Мыши, которым вводили саркому + носитель, характеризовались большим временем защитной фиксации по сравнению с контрольными мышами, которым вводили псевдоагент + носитель (Фиг.6а). Кроме того, мыши, которым вводили саркому + носитель, характеризовались большим числом подрагиваний по сравнению с контрольными мышами, которым вводили псевдоагент + носитель (Фиг.6b). Введение антитела против NGF (с 6-го по 14-й день) мышам, которым инъецировали саркому, значительно ослабляло спонтанную защитную фиксацию по сравнению с мышами, которым вводили саркому + носитель (Фиг.6а). Кроме того, лечение антителом против NGF значительно уменьшало спонтанное подрагивание у мышей, которым инъецировали саркому (Фиг.6b).
Боль, вызываемую движением, анализировали путем измерения реакций, индуцируемых пальпацией. Мыши, которым вводили саркому + носитель, характеризовались большим временем защитной фиксации после пальпации по сравнению с контрольными мышами, которым вводили псевдоагент + носитель (Фиг.6с). Мыши, которым вводили саркому + носитель, также характеризовались большим числом подрагиваний после пальпации по сравнению с контрольными мышами, которым вводили псевдоагент + носитель (Фиг.6d). Лечение антителом против NGF мышей, которым инъецировали саркому, значительно уменьшало вызываемую пальпацией защитную фиксацию (Фиг.6с) и вызываемое пальпацией подрагивание (Фиг.6d). В предварительных исследованиях не наблюдалось значительных различий в поведении или побочных эффектов у псевдозараженных животных, которым вводили носитель или антитело против NGF.
На Фиг.6 показано, что лечение антителом против NGF (n=8) с 6-го по 14-й день после инъекции опухоли (треугольник) значительно улучшало поведение, характерное для постоянной боли и боли, вызванной пальпацией, на 10-й, 12-й и 14-й день по сравнению с мышами, которым вводили саркому + носитель, (n=8) (квадрат) и существенно уменьшало уровни псевдоагента на 10-й день для всех показателей (ромб). Во все периоды времени мыши, которым вводили псевдоагент + носитель, (n=8) значительно отличались от мышей, которым вводили саркому + носитель. Таким образом, лечение антителом против NGF (10 мг/кг, внутрибрюшинно, каждые 5 дней) улучшало поведение, характерное для постоянной боли и вызываемой движением боли, обусловленной раком кости, на протяжении всего развития болезни.
Лечение антителом против NGF не влияло на исходные пороги тепловой или механической чувствительности и было сравнимо с эффективностью морфина в отношении ослабления боли, обусловленной раком кости. Не наблюдалось значительного увеличения латентного периода при отдергивании лапы вследствие теплового раздражения или увеличения порога механического раздражения при введении антитела против NGF по сравнению с нормальными порогами болевой чувствительности. Лечение антителом против NGF не влияло на нормальную тепловую реакцию (Фиг.7а) по сравнению с незараженными мышами, которым вводили носитель, или на нормальную механическую реакцию (Фиг.7b) по сравнению с незараженными мышами, которым вводили носитель.
Животных исследовали для сравнения эффективности сульфата морфина (MS) с антителом против NGF в отношении ослабления боли, обусловленной раком кости. Оценка поведения на 10-й и 14-й день показала, что животные, которым вводили саркому + носитель, характеризовались статистически более продолжительным временем защитной фиксации (Фиг.7с) и более продолжительным временем защитной фиксации вследствие пальпации (Фиг.7d) конечности, в которую инъецировали саркому, по сравнению с животными, которым вводили псевдоагент + носитель. Лечение антителом против NGF (10 мг/кг/каждые 5 дней, внутрибрюшинно) или сульфатом морфина (10 мг/кг или 30 мг/кг, внутрибрюшинно) значительно уменьшало постоянную или вызванную движением боль на 10-й и 14-й день после инъекции опухоли (Фиг.7с, 7d) по сравнению с мышами, которым вводили саркому + носитель. Лечение антителом против NGF более эффективно ослабляло боль, обусловленную раком кости, по сравнению с морфином в дозах, равных 10 мг/кг или 30 мг/кг (Р<0,05 по сравнению с мышами, которым вводили саркому + антитело против NGF).
Лечение антителом против NGF модулировало изменения периферических нейронов, индуцированные раком кости в DRG. Ранее было установлено, что в модели повреждения периферических нервов повышается уровень фактора активации транскрипции 3 (ATF-3), который относится к семейству ATF/CREB. Tsujino et al., Molecular & Cellular Neurosciences 15:170-82 (2000). Уровни ATF-3 повышаются в телах сенсорных и двигательных нейронов и, как известно, метят поврежденные нейроны. Наблюдалось значительное увеличение процентного содержания ATF-3-иммунореактивных нейронов в DRG L2, расположенном ипсилатерально по отношению к бедренной кости, в которую инъецировали саркому, (14,0±5,9% от общего числа нейронов в ATF-3, экспрессируемом L2; Фиг.8а) по сравнению с животными, которым вводили псевдоагент + носитель (1,6±0,5% от общего числа нейронов в ATF-3, экспрессируемом L2). Лечение антителом против NGF значительно уменьшало экспрессию ATF-3 (2,6±1,0% от общего числа нейронов в ATF-3, экспрессируемом L2; Фиг.8b) через 14 дней после инъекции опухоли.
Установлено, что в результате повреждения периферических нервов повышается инфильтрация макрофагов. Abbadie et al., Proc. Natl. Acad. USA 100:7947-52 (2003); Myers et al., Exp. Neurol. 141:94-101 (1996); Tofaris et al., J. Neurosci. 22:6696-703 (2002). Антитело против CD68 (ED-1), липосомного белка, экспрессируемого активированными макрофагами ткани, использовали для оценки инфильтрации макрофагов у мышей, которым инъецировали саркому. Наблюдалось увеличение числа CD68-иммунореактивных нейронов в ипсилатеральном DRG мышей, которым вводили саркому + носитель, (119,6±12,1 клеточного профиля/ипсилатеральный DRG L2; Фиг.8с) по сравнению с мышами, которым вводили псевдоагент + носитель (80,6±6,0 клеточных профилей/ипсилатеральный DRG L2). Лечение антителом против NGF значительно уменьшало число CD68-иммунореактивных нейронов в ипсилатеральном DRG (92,0±9,9 клеточного профиля/ипсилатеральный DRG L2; Фиг.8d) у мышей, которым инъецировали саркому, свидетельствуя о значительном уменьшении числа активированных и инфильтрующихся макрофагов в ипсилатеральном DRG L2 животных, пораженных опухолью.
Лечение антителом против NGF модулировало изменения центральных нейронов, индуцированные раком кости в спинном мозге. Установлено, что экспрессия динорфина способствует возникновению хронической боли. Vanderah et al., Pain 92:5-9 (2001). Кроме того, установлено, что экспрессия динорфина повышается в заднем роге спинного мозга в некоторых случаях постоянной боли. Iadarola, et al., Brain Res. 455:205-212 (1988); Noguchi et al., Molecular Brain Research 10:227-233 (1991); Schwei et al., J. Neurosci. 19:10886-97 (1999). У мышей, которым вводили псевдоагент + носитель, небольшое число спинномозговых нейронов экспрессировало динорфин в глубоком спинномозговом слое (2,3±1,1 динорфин-IR нейрона/отдел L3-L4). В отличие от этого мыши, которым вводили саркому + носитель, экспрессировали значительно больше динорфин-IR нейронов (6,0±0,5 динорфин-IR нейрона/отдел L3-L4; Фиг.9А). Лечение антителом против NGF значительно уменьшало экспрессию динорфина (2,0±0,6 динорфин-IR нейрона/отдел L3-L4; Фиг.9В) у мышей, которым инъецировали саркому.
Лечение антителом против NGF предотвращало активацию предраннего гена. Экспрессию с-Fos в глубине заднего рога (слои III-VI) использовали в качестве маркера сенсибилизации центральных нейронов в случае боли, обусловленной саркомой кости. Honore et al., Nat. Med. 6: 521-8 (2000); Honore et al., Neuroscience 98:585-598 (2000); Luger et al., Cancer Research 61:4038-4047 (2001); Schwei et al., J. Neurosci. 19:10886-97 (1999). Нормальная безболезненная пальпация псевдозараженных животных вызывала минимальную экспрессию с-Fos в глубоких слоях. Sabino et al., Cancer Res. 62:7343-9 (2002). Мыши, которым вводили саркому + носитель, характеризовались увеличенным числом с-Fos-IR нейронов (27,7±4,9; с-Fos-IR нейроны/отдел L3-L4; Фиг.9С), и лечение антителом против NGF значительно уменьшало указанную экспрессию (11,1±1,9; с-Fos-IR нейроны/отдел L3-L4; Фиг.9D).
Результаты RT-PCR. Для установления клеток саркомы в качестве возможного источника NGF, в клетках 2472, выращенных в культуре, определяли уровень мРНК NGF методом RT-PCR. Указанные уровни сравнивали с несколькими нормальными тканями мыши, а также с уровнем мРНК NGF, полученной из слюнной железы самца мыши, которая является источником аберрантно высокого уровня экзокринного NGF. Как показано в приведенной ниже таблице 3, клетки 2472 саркомы содержали in vitro легко обнаруживаемую мРНК NGF. Данный уровень находится в пределах содержания мРНК NGF, полученной из нормальных тканей, экспрессирующих высокие уровни мРНК NGF, таких как радужная оболочка. Shelton et al., Proc. Natl. Acad. Sci. USA 81:7951-5 (1984). Однако указанный уровень на несколько порядков величины ниже уровня мРНК NGF, присутствующей в слюнной железе самца мыши.
Пример 3
Воздействие моноклонального антитела против NGF на лечение боли, обусловленной раком кости, в модели рака кости у мышей, созданной в результате интрамедуллярной инъекции опухолевых клеток остеобластической карциномы предстательной железы в бедренную кость
Методы
Модель боли, обусловленной раком кости вследствие карциномы предстательной железы, у мышей. Модель боли, обусловленной раком кости вследствие карциномы предстательной железы, у мышей использовали для оценки эффективности лечения антителом против NGF 911 (моноклональное антитело мыши; см. публикацию Hongo, et al., Hybridoma 19:215-227 (2000)). Клетки остеобластической карциномы собаки (ACE-1, предоставленные д-ром Томасом Дж. Розолом, Ohio State University) сохраняли в поддерживающей среде и инъецировали в соответствии с описанием, приведенным в научной литературе. Sabino et al., Cancer Res. 62:7343-7349, 2002; Honore et al., Nature Medicine 6:521-528. 2000; Honore et al., Prog. Brain Res. 129:389-397, 2000; Luger et al., Cancer Research 61:4038-4047, 2001. Клетки АСЕ-1 выращивали в среде при 37°С и 5% СО2. Клетки выращивали в колбах Т75 (7,5 см2) и пассировали при 80-90% конфлюэнтности дважды в неделю. В данном исследовании были использованы только пассажи 3-11. В 0-й день мышей анестезировали пентобарбиталом натрия (50 мг/кг, внутрибрюшинно) и выполняли артротомию, обнажая мыщелок дистального отдела бедренной кости. В интрамедуллярную область бедренной кости мыши инъецировали забуференный стерильный физиологический раствор Хэнка (HBSS, Sigma Chemical Co., St. Louis, MO; 20 мкг; псевдоагент, n=7) или среду, содержащую 105 клеток АСЕ-1 остеобластической карциномы собаки (20 мкл, АСЕ-1, n=60), место инъекции закрывали амальгамой для зубной пломбы (Dentsply, Milford, DE) и промывали стерильной фильтрованной водой. Эксперименты выполняли с использованием в общей сложности 89 взрослых бестимусных самцов мышей с мутацией по гену nude в возрасте 8-10 недель (Harlan Laboratories, Madison, WI) с массой тела 20-32 г. Мышей содержали в соответствии с требованиями Национальных институтов здравоохранения в специальных стерильных условиях в стерилизованных клетках при 22°С с 12-часовым циклом чередования света и темноты и давали стерилизованную в автоклаве пищу и воду ad libitum.
Исследование завершали на 19-й день после инъекции, так как именно в данный период времени опухоль все еще удерживается в кости, в максимальной степени проявляется поведение, характерное для боли, обусловленной раком, и происходит изменение формы кости, индуцированное опухолью. Для контрольного анализа при исследовании поведения и выполнении гистологических и иммуногистохимических исследований кости использовали псевдозараженных животных, так как поведение незараженных животных существенно не отличалось от поведения псевдозараженных животных через 9 дней после инъекции опухоли.
Лечение антителом против NGF или морфином. На 7-й, 12-й и 17-й день после инъекции опухоли животным, которым инъецировали АСЕ-1, внутрибрюшинно (i.p.) вводили антитело против NGF 911 в количестве 10 мг/кг (АСЕ-1 + антитело против NGF, n=9); животным, которым инъецировали АСЕ-1, внутрибрюшинно вводили физиологический раствор (АСЕ-1 + носитель, n=21; 1,4 мкг/кг) и животным, которым инъецировали псевдоагент, внутрибрюшинно вводили физиологический раствор (псевдоагент + носитель, n=7). Поведение всех животных анализировали в период между 7-м и 19-м днями.
Для сравнения поведенческой реакции на антитело против NGF и сульфат морфина мышам вводили дозу морфина за 15 минут до исследования поведения (незараженные мыши: n=6; псевдозараженные мыши: n=7; АСЕ-1 + носитель: n=7; АСЕ-1 + антитело против NGF: n=7; АСЕ-1 + морфин в дозе 10 мг/кг, подкожно: n=8; АСЕ-1 + морфин в дозе 30 мг/кг, подкожно: n=8). Для испытания тепловой и механической чувствительности и оценки иннервации кожи задней лапы незараженных мышей распределяли в две экспериментальные группы и вводили или стерильный физиологический раствор (незараженные мыши + носитель: n=8), или антитело против NGF (незараженные мыши + антитело против NGF: n=8, 10 мг/кг, внутрибрюшинно).
Поведенческий анализ. У животных исследовали поведение, обусловленное болью, до имплантации опухоли или инъекции псевдоагента и на 7-й, 9-й, 11-й, 13-й, 15-й, 17-й и 19-й день после имплантации опухоли или инъекции псевдоагента. Поведение животных исследовали с помощью нижеследующих испытаний: постоянная боль (спонтанная защитная фиксация и подрагивание) и боль, вызванная движением (защитная фиксация и подрагивание, вызванные пальпацией). Животных помещали в прозрачный пластиковый ящик для наблюдения с полом из металлической сетки, где они находились в течение 30 минут. После адаптации у животных определяли спонтанную защитную фиксацию и спонтанное подрагивание. Защитную фиксацию и подрагивание, вызванные пальпацией, измеряли через 2 минуты после обычно безболезненной пальпации дистального отдела бедренной кости у животных, которым инъецировали АСЕ-1 и псевдоагент. Указанные испытания выполняли аналогично примерам 1 и 2.
Эвтаназия и обработка ткани. Мышей умерщвляли через 19 дней после инъекции опухоли и производили обработку тканей для выполнения иммуногистохимического анализа бедренной кости и кожи задней лапы в соответствии с описанием, приведенным в научной литературе. Honore et al., Prog. Brain Res. 129:389-397, 2000; Honore et al., Nat. Med. 6:521-8 (2000); Luger et al., Cancer Research 61:4038-4047 (2001). Мышей умерщвляли с помощью СО2 и перфузировали в сердце 12 мл 0,1 М физиологического раствора с фосфатным буфером (PBS) и затем 25 мл раствора 4% формальдегида/ 12,5% пикриновой кислоты.
Удаляли кожу подошвы задней лапы, фиксировали в фиксаторе для перфузии и подвергали криозащите в 30% сахарозы в течение 24 часов. На санном микротоме изготавливали последовательные срезы кожи толщиной 60 мкм, помещали в PBS и обрабатывали в виде свободно плавающих срезов. Полученные срезы кожи подошвы быстро промывали в PBS и инкубировали в блокирующем растворе (3% нормальной сыворотки осла (NDS), 0,3% тритона Х-100 в PBS) в течение 1 часа и затем инкубировали в течение ночи в первичном антителе. Срезы кожи подвергали иммунному окрашиванию для обнаружения пептида, родственного гену кальцитонина (CGRP) (1:15000; Sigma, St.Louis, MO), тирозингидроксилазы (ТОН) (поликлональное антитело против ТОН кролика, 1:2000, Chemocon, Tenecula, CA) и нейрофиламента Н (клон RT97) (поликлональное антитело против RT-97 кролика, 1:2500, Chemicon, Temecula, CA).
После инкубации в первичном антителе срезы промывали в PBS и затем инкубировали в растворе вторичного антитела в течение 3 часов. Вторичные антитела, конъюгированные с Су3 или биотином (Jackson ImmunoResearch, West Grove, PA), использовали в отношении соответственно 1:600 или 1:500. Для обнаружения вторичных антител, конъюгированных с биотином, срезы промывали в PBS и инкубировали в стрептавидине, конъюгированном с Су3 (1:4000; Jackson ImmunoResearch) в течение 45 минут. Для подтверждения специфичности первичных антител использовали контрольные образцы, в которых отсутствовало первичное антитело, или выполняли предварительную адсорбцию соответствующим синтетическим пептидом. После иммунного окрашивания срезы кожи подошвы промывали и помещали на покрытые желатином предметные стекла. Срезы кожи дегидратировали в градиентах спирта (70, 90, 100%), очищали в ксилоле и наносили на покровные стекла с DPX (Fluka, Buchs, Switzerland).
После выполнения рентгенологического исследования на 19-й день после инъекции правую (внутренний контрольный образец) и левую (пораженную опухолью) бедренную кость фиксировали в пикриновой кислоте и 4% формалине при 4°С в течение ночи и декальцифицировали в 10% EDTA (Sigma) в течение не более 14 дней. Затем кости погружали в парафин. Изготавливали срезы боковой плоскости бедренной кости толщиной 5 мкм и окрашивали устойчивой к тартрату кислой фосфатазой (TRAP), гематоксилином и эозином (H&E) для визуализации гистологических признаков нормального костного мозга, опухоли, остеокластов, остеобластов и макрофагов (Ms).
Иммуногистохимический анализ псевдозараженной и раковой бедренной кости выполняли с использованием декальцифицированных, погруженных в парафин последовательных срезов толщиной 14 мкм. Эндогенные пероксидазы блокировали, инкубируя срезы в 2% пероксиде водорода в течение 1 часа. Затем срезы трижды промывали PBS в течение 10 минут и блокировали в ТSA-блокирующем буфере (система TSA-Plus Cyanine 3, PerkinElmer Life Sciences, Inc., Boston, MA) в течение 1 часа. После удаления блокирующего буфера добавляли первичную антисыворотку и инкубировали при комнатной температуре в течение ночи. Афферентные немиелинизированные и слабо миелинизированные сенсорные нервные волокна метили антителом против пептида, родственного гену поликлонального антитела против кальцитонина кролика (CGRP) (1:15000, Sigma). Срезы трижды промывали в промывочном буфере TSA в течение 10 минут и затем в течение 45 минут инкубировали в стрептавидине HRP (1:4000). Затем срезы трижды промывали промывочным буфером ТSA в течение 10 минут. Бедренные кости подвергали воздействию тирамина, конъюгированного с CY3, (1:600) из системы TSA-Plus Cyanine 3 в течение 7 минут, дважды промывали промывочным буфером ТSA и один раз PBS. И наконец, срезы подвергали воздушной сушке, дегидратировали в градиенте спирта (70, 90 и 100%), очищали в ксилоле и обрабатывали DPX (Fluka).
Рентгенографический анализ кости. Рентгенограммы (Faxitron X-ray Corр., Wheeling, IL) рассеченной бедренной кости получали на 19-й день для оценки образования и разрушения кости. Изображения были получены на пленке для маммографии Kodak Min-R 2000 (Eastman Kodak Co., Rochester, NY; регистрирующая экспозиция: 7 секунд, 21 кВп). Анализ плотности кости использовали для рентгенографической оценки изменения формы кости под воздействием опухоли на изображениях боковой плоскости цельной кости при 5-кратном увеличении. Бедренные кости, пораженные опухолью и не пораженные опухолью (n=8 для незараженных мышей, которым вводили носитель, мышей, которым вводили псевдоагент + носитель, мышей, которым вводили АСЕ-1 + носитель, и мышей, которым вводили АСЕ-1 + антитело против NGF), анализировали при помощи 'ImageJ (Research Services Branch, National Institute of Mental Health, Bethesda, MD)' в соответствии с описанием, приведенным в научной литературе. Corey et al., Prostate 52:20-33, 2002. Для построения калибровочной кривой использовали рентгеновскую пленку для переноса изображения и стандартную покадровую табличку (Eastman Kodak Co.). ImageJ использовали для измерения оптической плотности, которую затем преобразовывали в пропускание следующим образом: пропускание = 1/(антилогарифм10[оптическая плотность]). Указанные данные определяли с негативного изображения, поэтому пропускание является прямым представлением плотности кости. Для получения субнасыщенных рентгенограмм бедренной кости использовали сканер HP ScanJet 7400c, при этом показания регистрировали в двух повторностях для каждой бедренной кости. Результаты представлены в виде среднего значения нормализованного пропускания ± стандартная ошибка.
Гистологический анализ остеобластов, остеокластов и макрофагов, роста опухоли и изменения формы кости. Пролиферацию остеобластов анализировали путем количественного определения числа остеобластов, непосредственно соприкасающихся с участками индуцированного опухолью образования новой кости, находящимися внутри бедренной кости и кортикального слоя кости на протяжении всей диафизарной интрамедуллярной области у незараженных мышей, псевдозараженных мышей и мышей, пораженных опухолью. Диафизарная интрамедуллярная область, проходящая от проксимальной части дистальной трабекулы до дистальной части проксимальной трабекулы, была выбрана для количественного определения с учетом того, что основное активное изменение формы кости происходит именно на этом участке. Остеобласты были идентифицированы как клетки, непосредственно соприкасающиеся с вновь образованной костной структурой, расположенные в виде типичного эпителиального слоя кубовидных или цилиндрических клеток и соединенные друг с другом тонкими выростами, обнаруживаемыми при сильном увеличении (200-кратное или более сильное увеличение). Результаты представлены в виде числа остеобластов/мм2 диафизарной интрамедуллярной области для незараженных мышей, псевдозараженных мышей и мышей, пораженных опухолью. Пролиферацию остеокластов измеряли путем количественного определения TRAP+ остеокластов на поверхности раздела между костью и опухолью и на поверхности раздела между нормальным костным мозгом и костью для незараженных мышей, псевдозараженных мышей и мышей, которым инъецировали АСЕ-1, на TRAP-окрашенных срезах бедренной кости в соответствии с описанием, приведенным в научной литературе. Honore et al., Nat. Med. 6:521-528 (2000). Остеокласты являются гистологически дифференцированными клетками, представляющими собой TRAP+ клетки, которые непосредственно связаны с участками резорбции кости. Указанные клетки являются многоядерными клетками, обнаруживаемыми в лакуне Хаушипа вдоль кортикального слоя кости и губчатой кости. Fawcett, D.W.; A Textbook of Histology. In: D. Dreibelbis (ed.), Bone, 11 edition, pp.211-213. Philadelphya, РA: W.B. Saunders Company, 1986. Пролиферацию макрофагов (Ms) измеряли путем количественного определения числа TRAP+ клеток, диспергированных в опухоли и нормальном костном мозге, не ассоциированном с эндостальной поверхностью минерализованной кости. Макрофаги внутри кости активируются под воздействием высвобождаемых опухолью факторов, которые стимулируют клетки, и на появление в клетках указанных активированных макрофагов указывает очень неправильная форма, множественная пластинчатость и фагоцитарные вакуоли. Результаты выражены в виде среднего числа остеокластов на мм2 или Ms на мм2 диафизарной интрамедуллярной области.
Изображения бедренной кости, содержащей клетки АСЕ-1, были получены при помощи светлопольной микроскопии в флуоресцентном микроскопе Nikon E600, оснащенном цифровой камерой SPOT II, с использованием программного обеспечения получения изображений SPOT (Diagnostic Instruments, Sterling Heights, MI). Общая площадь интрамедуллярной области и процентное значение интрамедуллярной области, занимаемой опухолью, образование кости и оставшиеся гемопоэтические клетки высчитывали при помощи программного обеспечения Image Pro Plus, версия 3.0 (Media Cybernetics, Silver Spring, MD). Sabino et al., Cancer Res. 62:7343-7349, 2002; Sevcik et al., Pain 111:169-180, 2004. Образование кости анализировали, используя те же срезы бедренной кости, окрашенные H&E, которые были использованы для количественного определения роста опухоли. Срезы бедренной кости исследовали в поляризованном свете для идентификации областей образования сетчатой и слоистой кости. Изображения областей образования сетчатой кости были получены с помощью цифровой камеры SPOT II и подвергнуты количественному определению с использованием программного обеспечения Image Pro Plus, версия 3.0. Результаты представлены в виде процентного значения площади опухоли, индуцированного опухолью образования кости и оставшихся гемопоэтических клеток от общей площади интрамедуллярной области.
Количественное определение сенсорных волокон в кости и коже. Число сенсорных нервных волокон определяли в соответствии с описанием, приведенным в научной литературе. Mach et al., Neuroscience 113:155-166, 2002. Число CGRP-иммунореактивных волокон в трех отделах кости (проксимальный, дистальный и диафизарный) и в трех тканях кости (надкостница, минерализованная кость и костный мозг) идентифицировали при помощи конфокальной системы получения изображений MRC-1024 (Bio-Rad, Richmond, CA), оснащенной объективом с 20-кратным увеличением. Подсчет нервных волокон производили в результате исследования шести срезов бедренной кости у одной мыши под флуоресцентным микроскопом Olympus BH-2. При выполнении данного анализа исследовали только нервные волокна длиной более 30 мкм. Для измерения общей площади поверхности (мм2) каждой кости анализировали те же срезы бедренной кости, в которых производили подсчет нервных волокон. Общую площадь кости измеряли на цифровых изображениях срезов бедренной кости, полученных при помощи цифровой камеры SPOT II и программного обеспечения Image Pro Plus, версия 3.0. Результаты представлены в виде числа волокон, подсчитанных на общей площади кости.
Количественное определение плотности эпидермальной иннервации производили на 4 произвольно выбранных срезах кожи подошвы задней лапы одной мыши. Общее число CGRP-, ТОН- и RT97-иммунореактивных нервных волокон подсчитывали при 200-кратном увеличении. В соответствии с правилами подсчета производили подсчет только отдельных интраэпидермальных волокон, а не многочисленных ответвлений одного волокна. McCarthy et al., Neurology 45:1848-55 (1995). Общую длину эпидермиса на всех срезах измеряли с использованием 1 см2 окулярной сетки. Подсчитывали только нервные волокна, имеющие длину, равную по крайней мере 30 мкм, и прорастающие в эпидермис. Результаты представлены в виде среднего числа интраэпидермальных нервных волокон на мм длины у одной мыши.
Анализ методом RC-PCR уровней мРНК NGF в клетках АСЕ-1. Общую РНК из головного мозга собаки или опухолевых клеток АСЕ-1 предстательной железы собаки получали с помощью микронабора RNeasy (Qiagen) в соответствии с инструкциями изготовителя и производили количественное определение РНК, используя реагент рибогрин (Molecular Probes). Выполняли двухстадийную реакцию RT-PCR, используя набор для RT-PCR TaqMan Gold (Applied Biosystems). РНК подвергали обратной транскрипции с помощью произвольных гексамеров и кДНК амплифицировали с помощью набора затравок/зондов, специфичных для NGF (LB041: AACAGGACTCACAGGAGCAA (SEQ ID NO:6), LB042: CGGCACTTGGTCTCAAAGAA (SEQ ID NO:7) и LB045: AATGTTCACCTCTCCCAGCACCATCA (SEQ ID NO:8)). Образцы, полученные на стадии обратной транскрипции, анализировали в двух экземплярах и нормализовали до получения полной РНК.
Статистический анализ. Для выполнения статистических анализов использовали пакет компьютерных программ для статистической обработки данных Statview (SAS Institute, Inc., Cary, NC). Для сравнения результатов поведенческого анализа, результатов гистологического исследования кости и иммуногистохимических измерений в экспериментальных группах использовали однонаправленный метод ANOVA. Для многократных сравнений использовали критерий PLSD Фишера (защищенная наименьшая значимая разница). Уровень значимости был задан равным Р<0,05. Исследователю, выполнявшему поведенческий, иммуногистохимический анализ и производившему балльную оценку изменения формы кости, не был известен экспериментальный статус каждого животного.
Результаты
Лечение антителом против NGF ослабляло боль, обусловленную раком кости, в большей степени, чем сульфат морфина, но не влияло на исходные пороги тепловой или механической чувствительности. Постоянную боль анализировали, измеряя спонтанную защитную фиксацию и подрагивание в течение 2-минутного периода времени. У мышей, которым вводили АСЕ-1 + носитель, наблюдалась более продолжительная защитная фиксация (7,7±0,8 сек, 19-й день) по сравнению с контрольными мышами, которым вводили псевдоагент + носитель (0,6±0,3 сек, 19-й день, Фиг.10А). Кроме того, у мышей, которым вводили АСЕ-1 + носитель, было отмечено большее число подрагиваний (11,9±1,2, 19-й день) по сравнению с контрольными мышами, которым вводили псевдоагент + носитель (1,0±0,4, 19-й день, Фиг.10В). Введение антитела против NGF мышам, которым инъецировали АСЕ-1, значительно уменьшало спонтанную защитную фиксацию (1,2±0,4 сек, 19-й день) по сравнению с мышами, которым вводили АСЕ-1 + носитель (Фиг.10А). Лечение антителом против NGF также значительно уменьшало спонтанное подрагивание у мышей, которым инъецировали АСЕ-1, (2,1±0,7, 19-й день) по сравнению с мышами, которым вводили АСЕ-1 + носитель (Фиг.10В). В предварительных исследованиях не наблюдалось существенных различий в поведении или побочных эффектов у псевдозараженных контрольных животных, которым вводили носитель или антитело против NGF.
Лечение антителом против NGF не влияло на нормальную тепловую реакцию (10,2±0,4 сек, 19-й день) по сравнению с незараженными мышами, которым вводили носитель (11,2±0,4 сек, 19-й день, Фиг.10С) или на нормальную механическую реакцию (5,4±0,3 г, 19-й день) по сравнению с незараженными мышами, которым вводили носитель (5,2±0,4 г, 19-й день, Фиг.10D).
Животных исследовали для сравнения эффективности сульфата морфина (MS) и антитела против NGF в отношении ослабления боли, обусловленной раком кости. Оценка поведения на 11-й и 19-й день после инъекции опухоли показала, что животные, которым вводили АСЕ-1 + носитель, характеризовались статистически более продолжительным временем защитной фиксации инъецированной конечности (соответственно 6,0±1,0 и 7,6±1,2 сек, 11-й и 19-й день) по сравнению с мышами, которым вводили псевдоагент + носитель (соответственно 0,4±0,2 и 0,6±0,3, 11-й и 19-й день, Фиг.10Е). У мышей, которым вводили АСЕ-1 + носитель, наблюдалось статистически большее число подрагиваний инъецированной конечности (соответственно 8,6±1,2 и 11,7±1,7, 11-й и 19-й день) по сравнению с мышами, которым вводили псевдоагент + носитель (соответственно 0,7±0,3 и 1,0±0,4, 11-й и 19-й день, Фиг.10F). Защитная фиксация при постоянной боли значительно уменьшилась при продолжительном лечении антителом против NGF (соответственно 2,1±1,1 и 1,4±0,4 сек, 11-й и 19-й день), сульфатом морфина в дозе 10 мг/кг (соответственно 3,5±0,3 и 4,0±0,5 сек, 11-й и 19-й день) или сульфатом морфина в дозе 30 мг/кг (соответственно 2,2±0,3 и 2,0±0,4 сек, 11-й и 19-й день) по сравнению с мышами, которым вводили АСЕ-1 + носитель (Фиг.10Е). Подрагивание при постоянной боли также существенно уменьшилось при продолжительном лечении антителом против NGF (соответственно 3,4±1,7 и 2,6±0,6, 11-й и 19-й день), сульфатом морфина в дозе 10 мг/кг (соответственно 5,6±0,5 и 6,8±0,7, 11-й и 19-й день) или сульфатом морфина в дозе 30 мг/кг (соответственно 3,6±0,5 и 3,5±0,7, 11-й и 19-й день) по сравнению с мышами, которым вводили АСЕ-1 + носитель (Фиг.10F). Антитело против NGF более эффективно улучшало поведение, характерное для боли, обусловленной раком кости, чем сульфат морфина в дозе 10 мг/кг. Не было обнаружено различий в конечной массе тела у мышей, которым вводили псевдоагент + носитель (27±1 г), у мышей, которым вводили АСЕ-1 + носитель (27±1 г), и у мышей, которым вводили АСЕ-1 + антитело против NGF (26±1 г). В выполненных исследованиях не наблюдалось существенных различий в поведении или побочных эффектов, таких как атаксия, болезненность или вялость, у животных, которым вводили носитель или антитело против NGF.
Лечение антителом против NGF ослабляло вызванную прикосновением боль, обусловленную раком кости. Оценивали поведение, характерное для боли, вызванной прикосновением. Защитную фиксацию и подрагивание, индуцированные пальпацией, измеряли после 2 минут обычно безболезненной пальпации дистального отдела бедренной кости у животных, которым инъецировали АСЕ-1 и псевдоагент. Как показано на Фиг.10G и 10Н, поведение животных на 7-й день после инъекции АСЕ-1 (с физиологическим раствором) было характерно для возникновения боли, вызываемой прикосновением, о чем свидетельствовала индуцированная пальпацией защитная фиксация (Фиг.10G) и индуцированное пальпацией подрагивание (Фиг.10Н) (для обоих показателей р<0,01, ANOVA), по сравнению с животными, которым инъецировали псевдоагент (с физиологическим раствором). На Фиг.10G и 10Н также показано, что внутрибрюшинное введение антитела против NGF 911 значительно уменьшало индуцированную пальпацией защитную фиксацию (Фиг.10G) и индуцированное пальпацией подрагивание (Фиг.10Н) у мышей, которым инъецировали АСЕ-1, с 11-го по 19-й день после имплантации опухоли АСЕ-1 по сравнению с введением физиологического раствора мышам, которым инъецировали АСЕ-1 (р<0,01, ANOVA, как для индуцированной пальпацией защитной фиксации, так и для индуцированного пальпацией подрагивания). Полученные результаты показывают, что антитело против NGF 911 ослабляет вызванную прикосновением боль у мышей, которым инъецировали АСЕ-1.
Лечение антителом против NGF не влияло на показатели развития болезни или индуцированное опухолью образование кости. Влияние лечения антителом против NGF на образование и разрушение кости, рост опухоли (Фиг.11) и пролиферацию остеокластов (Фиг.12) исследовали через 19 дней после инъекции опухоли (приведенная ниже таблица 4). У мышей, которым инъецировали псевдоагент, отсутствовало значительное изменение формы кости (значение нормализованного пропускания 115±2%) (Фиг.11А), пролиферация остеокластов во всей интрамедуллярной области (16±10 остеокластов/мм2 площади диафизарной интрамедуллярной области) (Фиг.12А) или опухолевых клеток (0±0%) (Фиг.11D) по результатам рентгенологических, TRAP и H&E анализов по сравнению с мышами, которым инъецировали АСЕ-1. У мышей, которым вводили АСЕ-1 + носитель, наблюдалось обширное образование и разрушение кости, но почти эквивалентное имеющему место в случае образования множественных диафизарных межклеточных мостиков и рентгенопрозрачности (значение нормализованного пропускания 109±5%) (Фиг.11В), значительное увеличение числа остеокластов (Фиг.12В) и остеоблатов по всей площади диафизарной интрамедуллярной области (47±3 остеокласта/мм2 и 127±7 остеобластов/мм2), при этом опухоль заполняла большую часть интрамедуллярной области (60±7% интрамедуллярной области) (Фиг.11Е). Лечение мышей, пораженных опухолью, антителом против NGF, начиная с 7-го дня после инъекции опухоли, не вызывало значительного изменения формы кости (значение нормализованного пропускания 106±9%) (Фиг.11С), уменьшения остеокластов, индуцированных АСЕ-1 (Фиг.12С), пролиферации остеобластов по всей площади диафизарной интрамедуллярной области (47±5 остеокластов/мм2 и 118±15 остеобластов/мм2) или роста опухоли (57±6% интрамедуллярной области) по сравнению с животными, которым вводили АСЕ-1 + носитель (Фиг.11F).
Таблица 4. Гистологическое и рентгенологическое количественное определение изменения формы кости и развития опухоли при введении животным, которым инъецировали АСЕ-1, антитела против NGF и носителя
Через девятнадцать дней после инъекции опухоли у мышей, которым вводили АСЕ-11 + носитель, наблюдалось увеличение макрофагов (Ms) (27±2 макрофага/мм2 площади диафизарной интрамедуллярной области) по сравнению с контрольными мышами, которым вводили псевдоагент + носитель (2±1 макрофаг/мм2). Лечение антителом против NGF мышей, которым инъецировали АСЕ-1, (24±3 макрофага/мм2) существенно не изменяло инфильтрацию макрофагов, как показано для мышей, которым вводили АСЕ-1 + носитель (таблица 4).
Лечение антителом против NGF не оказывало заметного влияния на иннервацию сенсорных или симпатических нейронов в кости или коже. Тонко миелинизированные или немиелинизированные пептидергические сенсорные нервные волокна (CGRP-IR), крупные миелинизированные сенсорные волокна (RT97-IR) и норадренергические симпатические нервные волокна (ТОН-IR) анализировали в бедренной кости, в которую инъецировали АСЕ-1, или в коже подошвы задней лапы иммуногистохимическим методом с использованием соответственно антител против CGRP, RT-97 и ТОН. CGRP-IR нервные волокна были обнаружены во всей кости (надкостница, минерализованная кость, костный мозг и опухоль) у животных, которым вводили АСЕ-1 + носитель, (23,5±1,9 волокна/мм2) и у животных, которым вводили АСЕ-1 + антитело против NGF (24,0±1,9 волокна/мм2), а также у животных, которым вводили псевдоагент + носитель, (28,2±1,5 волокна/мм2) и у незараженных животных, которым вводили носитель, (24,6±2,4 волокна/мм2) или у незараженных животных, которым вводили антитело против NGF (23,1±1,9 волокна/мм2) (Фиг.14). Не было обнаружено существенного различия между интенсивностью или плотностью CGRP-IR волокон в образцах кожи задней лапы, полученных у животных, которым вводили АСЕ-1 + носитель, (13,9±0,5 волокна/мм) и у животных, которым вводили АСЕ-1 + антитело против NGF (15,2±0,7 волокна/мм) (Фиг.13А и В). Аналогичным образом не было обнаружено существенного различия между интенсивностью или плотностью CGRP-IR волокон в образцах кожи задней лапы, полученных у незараженных животных, которым вводили носитель, (14,4±0,4 волокна/мм) и у незараженных животных, которым вводили антитело против NGF (14,2±1,3 волокна/мм) (Фиг.14А и В). Различия в плотности и интенсивности RT97-IR и ТОН-IR волокон не были также обнаружены у незараженных животных, которым вводили носитель, (4,2±2,2 RT97+ волокна/мм; 16,0±2,7 ТОН+ волокна/мм) и у незараженных животных, которым вводили антитело против NGF (8,0±0,6 RT97+ волокна/мм; 12,8±1,1 ТОН+ волокна/мм). Не было обнаружено существенных различий между интенсивностью или плотностью CGRP, RT97 или ТОН-иммунореактивных волокон в образцах кожи животных, которым вводили АСЕ-1 + носитель и АСЕ-1 + антитело против NGF, по сравнению с незараженными животными, которым вводили носитель, и незараженными животными, которым вводили антитело против NGF.
Уровень экспрессии мРНК в клетках АСЕ-1. Производили сравнение экспрессии NGF в головном мозге собаки и в клетках АСЕ-1. Анализировали пять независимых образцов АСЕ-1, и во всех образцах экспрессия NGF были ниже уровня обнаружения методом PCR. NGF в головном мозге собаки превышал пороговое значение в цикле 35,2 из 40 циклов эксперимента, в то время как NGF в образцах АСЕ-1 не превышал порогового значения после 40 циклов. Таким образом, экспрессия мРНК NGF в образцах АСЕ-1 была по крайней мере в 27,8 раза меньше экспрессии в головном мозге.
Пример 4
Аналгезирующие эффекты антитела Е3 против NGF у субъектов, испытывающих боль от умеренной до сильной вследствие метастазирования в кость рака предстательной железы или молочной железы
При выполнении рандомизированного, контролируемого плацебо исследования двойным слепым методом аналгезирующие эффекты (включая время до возникновения боли, время до максимального проявления боли, продолжительность и ослабление боли, измеряемые по визуальной аналоговой шкале (VAS)) внутривенно вводимых доз (100 мкг/кг, 300 мкг/кг или 1000 мкг/кг) антитела против NGF Е3 сравнивали с эффектом введения плацебо у субъектов, испытывающих боль от умеренной до сильной вследствие метастазирования в кость рака предстательной железы или молочной железы. В исследовании участвовали взрослые мужчины и женщины (в возрасте от 35 до 75 лет), испытывающие боль от умеренной до сильной вследствие метастазирования в кость рака предстательной железы или молочной железы. Во время скрининга субъектов просили регистрировать степень проявления боли четыре раза в день, а также применение других аналгетиков в течение 14 дней до введения антитела Е3 против NGF.
В исследовании участвовали двести восемьдесят субъектов. Антитело Е3 против NGF вводили утром в 1-й и 29-й день с последующей регистрацией в течение двух недель основного уровня боли, других применяемых аналгетиков и вредных эффектов. Двести восемьдесят субъектов были распределены в четыре группы по семьдесят субъектов в каждой группе. Субъекты в каждой группе получали плацебо, 100 мкг/кг, 300 мкг/кг или 1000 мкг/кг антитела Е3 против NGF.
Аналгезирующие эффекты оценивали четыре раза в день на протяжении четырнадцати дней до введения и в течение шести месяцев после введения антитела Е3. Результат определяли в виде изменения, произошедшего по сравнению с исходным показателем во время скрининга (средний уровень боли в течение четырнадцати дней до введения плацебо или антитела Е3). Любое уменьшение оценок боли и/или сокращение применения других аналгетиков в одной или нескольких группах субъектов, которые получали антитело Е3 против NGF, по сравнению с плацебо свидетельствовало об эффективности лечения антителом Е3 против NGF.
Несмотря на то что настоящее изобретение подробно описано со ссылкой на примеры, приведенные для простоты понимания, описание изобретения и примеры не следует рассматривать как ограничение объема изобретения.
Реферат
Группа изобретений относится к медицине, а именно к онкологии, и может быть использована при лечении боли, обусловленной раком кости. Способ заключается во введении пациенту эффективного количества антагониста фактора роста нервов (NGF), представляющего собой антитело против NGF. Набор включает в себя антагонист NGF в виде антитела против NGF и инструкции по его использованию. Изобретения позволяют эффективно купировать указанный болевой синдром, как постоянный, так и вызываемый движением, за счет высокого сродства антитела к NGF без влияния при этом на исходные пороги тепловой и механической чувствительности. 2 н. и 18 з.п. ф-лы, 14 ил., 4 табл.
Комментарии