Выделенный белок фактора роста кератиноцита (kgf), рекомбинантная молекула днк, вектор, способ конструирования клетки-хозяина, способ получения белка, фармацевтическая композиция - RU2250213C2
Код документа: RU2250213C2
Чертежи










Описание
Область изобретения.
Настоящее изобретение относится к факторам роста, особенно к выделению фактора роста, полипептида, аналогичного семейству факторов, включающему известные факторы роста фибробласта (FCF).
Настоящее изобретение относится также к конструированию сегментов комплементарной ДНК (cДНК) из информационной РНК (мРНК), кодирующей новый фактор роста. Кроме того, настоящее изобретение относится к синтезу продуктов таких ДНК сегментов рекомбинантными клетками, а также к производству и использованию некоторых других новых продуктов, получаемых в результате идентификации и клонирования ДНК, кодирующих такой фактор роста.
Сокращения, используемые в заявке
аFСF - фактор роста кислотного фибробласта
bFСF - фактор роста основного фибробласта
ЕСF - эпидермальный фактор роста
HCA - гепарин-Сефарозная сродственная хроматография
кb – килооснования
кДа – килодальтоны
КСF - фактор роста кератиноцита
NаДоdSО4/РАGЕ электрофорез в системе додецилсульфат натрия/SДS/полиакриламидный гель
RР-НРLС - обратимо-фазная хроматография высокого разрешения
ТGFа - трансформирующий фактор роста а.
Предпосылка создания изобретения
Факторы роста являются важными медиаторами межклеточной коммуникации. Такие мощные молекулы обычно выделяются клеткой одного типа и оказывают влияние на пролиферацию клеток другого типа (см. ссылку I-1 в Экспериментальном разделе I, ниже). Интерес к факторам роста усилился в результате получения свидетельств их потенциального участия в неоплазии (ссылка II-2 в Экспериментальном разделе II, ниже). У-sis трансформирующий ген вируса саркомы обезьян кодирует протеин гомологичный В цепи фактора роста - производного тромбоцита (I-1, I-2) Кроме этого, ряд онкогенов является гомологом генов, кодирующих рецепторы фактора роста (I-1). Таким образом, углубленное понимание путей сигнальной трансдукции, усиленной факторами роста или их рецепторами, по-видимому, может обеспечить проникновение в механизмы роста как нормальных, так и злокачественных клеток.
Известное семейство факторов роста, оказывающее влияние на клетки соединительной ткани, включает фактор роста кислотного фибробласта (аFcF), фактор роста основного фибробласта (bFСF) и родственные продукты hst и int-2 онкогенов.
Кроме того, известно, что некоторые факторы роста, включающие следующие разновидности, обладают гепарин-связующими свойствами aFcF (I-20, I-21), bFcF (I-19, I-20), фактор, стимулирующий гранулоцит/макрофаговую колонию (I-1), и интерлейкин 3 (I-1). Каждый из таких полипептидных факторов продуцируется стромальными клетками (I-1, I-2, I-25). По-видимому, такие факторы отлагаются во внеклеточной матрице или на протеогликанах, покрывающих поверхность стромальной клетки (I-1, I-25). Постулируется, что их хранение, выделение и контактирование с конкретными клетками-мишенями регулируется таким взаимодействием (I-25, I-28).
Однако хорошо известно, что большое число человеческих злокачественных образований является производными эпителиальных тканей (I-5). Были описаны эффекторы пролиферации эпителиальных клеток из мезенхимальных тканей (I-1, I-2, I-3), однако их молекулярная идентичность и структуры установлены не были. В свете дефицита сведений о мезинхимальных факторах роста, влияющих на эпителиальные клетки, совершенно понятно, что имеется необходимость в способах, композициях и биологических анализах, которые обеспечили бы углубленное знание и анализ механизмов регуляции пролиферации эпителиальных клеток и, особенно, необходимость в новых диагностических и терапевтических средствах, содержащих такие факторы.
Настоящее изобретение предусматривает применение методов выделения протеина и технологий на основе рекомбинантной ДНК для удовлетворения указанных потребностей и для создания средства продуцирования протеиновых факторов мезенхимального происхождения, которые, по-видимому, относятся к процессам пролиферации эпителиальных клеток и которые не могут быть получены иным путем. Настоящее изобретение также предусматривает применение механизмов действия таких факторов в отношении процессов роста эпителиальных клеток.
Сущность изобретения
Настоящее изобретение относится к разработке технологий выделения протеина и рекомбинантной ДНК, которые включают получение новых протеинов фактора роста, влияющих на эпителиальные клетки, которые не содержат других пептидных факторов. В сферу изобретения входят также новые сегменты ДНК и методы биологического анализа.
Главным образом настоящее изобретение относится к новому протеину, обладающему структурными и/или функциональными характеристиками известного семейства факторов роста, которое включает фактор роста кислотного фибробласта (аFCF), фактор роста основного фибробласта (bFBF) и родственные продукты hst и nit–2 - онкогенов. Такой новый член семейства FcF полипептидов сохраняет гепарин - связующие свойства FcF, но, кроме того, обладает уникальной специфичностью в отношении клетки-мишени. Такой фактор роста, по-видимому, специфичен по отношению к эпителиальным клеткам и проявляет особенную активность в отношении кератиноцитов. Поэтому такой новый фактор обозначен, как "фактор роста кератиноцита" (КСF). Несмотря на отсутствие активности в отношении фибробластов, поскольку такой фактор является шестым известным членом FCF полипептидного семейства, КСF может быть также обозначен, как FCF - 6.
В связи с этим, настоящее изобретение частично относится к очищенным КСF или КСF-подобным протеинам и способам получения таких протеинов. Такие очищенные факторы могут быть получены культивацией человеческих клеток, которые по своей природе секретируют такие протеины и применением методов выделения согласно практическому аспекту настоящего изобретения. Такие протеины могут использоваться для биохимических и биологических исследований, приводящих, например, к выделению сегментов ДНК, кодирующих KGF или КGF-подобные полипептиды.
Настоящее изобретение также относится к таким сегментам ДНК, которые кодируют KGF или КCF-подобные протеины. Согласно принципиальному воплощению настоящее изобретение относится к ДНК-сегментам, которые кодируют КGF-родственные продукты, состоящим из: человеческих сДНК клонов 32 или 49, полученных из полиаденилированной РНК экстрагированной из клеточной линии М426 человеческого эмбрионного легочного фибробласта; рекомбинантов и мутантов таких клонов; и родственных сегментов ДНК, которые могут детектироваться гибридизацией с любым из указанных выше сегментов человеческой ДНК; причем такие родственные сегменты кодируют КCF-подобные протеины или их части.
При практической реализации одного из воплощений настоящего изобретения ДНК-сегменты изобретения способны экспрессироваться в клетки подходящего хозяина в результате чего продуцируются КСF или КCF-подобные протеины. Настоящее изобретение также относится к мРНК, продуцированным в результате транскрипции чувствительных тяжей сегментов ДНК настоящего изобретения.
Согласно другому воплощению настоящее изобретение относится к молекуле рекомбинантной ДНК, включающей вектор и ДНК настоящего изобретения. Такие рекомбинантные молекулы могут быть представлены молекулами, включающими KCF сДНК и любую из следующих векторных ДНК: вектор, клонирующий бактериофаг λ (в качестве примера которого можно привести λ рСЕУ9); плазмидный вектор, устанавливающий последовательность ДНК (например, вариант рУС) вектор экспрессии бактериального гена (например, РКК233-2); или вектор экспрессии гена млекопитающего (такой как рММТ).
Согласно еще одному воплощению настоящее изобретение включает клетку, предпочтительно, клетку млекопитающего, трансформированную с помощью ДНК изобретения. Кроме этого, настоящее изобретение включает клетки, например, клетки насекомых, дрожжевые клетки и бактериальные клетки разновидностей Escherichia coli u B. subtilis, трансформированные ДНК настоящего изобретения. Согласно другому решению такого аспекта изобретения трансформирующая ДНК способна экспрессироваться в клетке, в результате чего в ней повышается количество KCF или КСF-подобного протеина закодированного такой ДНК.
Первичный продукт трансляции KCF, предсказанный на основании его сДНК последовательности, содержит N-терминальный гидрофобный участок, который вероятно служит сигнальной последовательностью для секреции и отсутствует в зрелой молекуле KCF. Согласно наиболее предпочтительному воплощению генно-экспрессионного аспекта изобретения клетка, трансформированная с помощью ДНК изобретения, секретирует протеин закодированный этой ДНК в такой форме (транкатированной), которая секретируется клетками человеческого эмбрионного легочного фибробласта.
Кроме этого, настоящее изобретение предусматривает KCF или КСF-подобные протеины, продуцированные экспрессией ДНК изобретения, или трансляцией РНК изобретения. Предпочтительно когда такие протеины находятся в секретированной форме (например, при отсутствии сигнальной последовательности). Такие протеиновые факторы могут использоваться для функциональных исследований и могут быть очищены для дополнительного структурного и функционального анализа, такого как качественный и количественный анализ на связывание рецептора.
Кроме этого, возможность продуцирования больших количеств такого нового фактора роста рекомбинантными методами позволяет осуществить испытание на его клиническую применимость в тех ситуациях, когда особенно важна специфическая стимуляция роста эпителиальных клеток. В соответствии с этим настоящее изобретение включает фармацевтические композиции, содержащие KCF или КСF-подобные полипептиды предназначенные для использования при лечении болезненных состояний включающем, например, заживление ран от ожогов или стимуляцию трансплатированной роговичной ткани.
В соответствии с таким воплощением настоящего изобретения новые КCF-подобные протеины представляют собой протеиновые продукты "немодифицированных" ДНК и мРНК настоящего изобретения, или модифицированные либо полученные методами генной инженерии протеиновые продукты. В результате инженерных мутаций в последовательностях ДНК модифицированные КCF-подобные протеины приобретают одно или более отличий в аминокислотной последовательности от соответствующих природных протеинов "дикого типа". В соответствии с одним из воплощений такого аспекта настоящего изобретения модифицированные КCF-подобные протеины будут включать "химерные" молекулы, содержащие сегменты аминокислотных последовательностей КCF, и, по крайней мере, один другой член FCF пептидного семейства.
Результаты аналогичных успешных подходов с использованием других пептидных факторов, обладающих аналогичными свойствами, позволяют заключить, что разработка таких химерных KCF-подобных полипептидов должна привести к улучшенным формам КCF-подобных пептидов "второго поколения", предназначенным для клинических целей. Так, например, такие модифицированные КСF-подобные продукты могут иметь меньший размер, могут быть более устойчивыми, более мощными и/или их получение может быть более легким или менее дорогостоящим.
Настоящее изобретение также охватывает новые методы биоанализа для определения экспрессии в человеческих клетках мРНК и протеинов, полученных из генов родственных сегментов ДНК изобретения. Согласно одному из таких воплощений ДНК настоящего изобретения могут использоваться в качестве зондов на определение уровней содержания стационарных состояний или кинетики индукции родственных мРНК. Доступность КСF-родственных cДНК клонов позволяет определять те случаи, когда аномальная экспрессия такого фактора роста имеет место в клинических состояниях, характеризующихся избыточным ростом эпителиальных клеток, включающих дисплазию и неоплазию (например, псориаз, злокачественные или доброкачественные опухоли).
Настоящее изобретение также предусматривает новые антитела для пептида закодированного ДНК сегментов настоящего изобретения. Согласно такому воплощению изобретения такие антитела могут быть моноклонального или поликлонального происхождения и их вырабатывают с помощью КСF-родственных полипептидов из природных, рекомбинантных или синтетических химических источников.
Антитела настоящего изобретения специфически связываются с KCF или КСF-подобным протеином, который включает последовательность такого пептида, предпочтительно в том случае, когда такой протеин находится в его природной (биологически активной) конформации. Такие антитела могут использоваться для детекции или очистки KCF либо они могут применяться для детекции или очистки KCF или KCF-подобных протеиновых факторов. Согласно наиболее предпочтительному воплощению этого аспекта изобретения антитела нейтрализуют промотирующую рост активность КСF, что позволяет осуществлять механистические исследования и, в конечном счете, проводить терапию клинических состояний, включающих избыточные уровни содержания КCF.
Краткое описание чертежей
На рис.I-1 представлены результаты гепарин-Сефарозной сродственной хроматографии кондиционированной среды из м426 человеческих эмбрионных фибробластов, показывающие, что более 90% митогенной активности мышинных кератиноцитов (BAIB/MK) элюируется 0,6М NаСl.
На рис.I-2 проиллюстрированы результаты дополнительной очистки митогена из человеческих фибробластов с использованием HPIC и адсорбционной матрицы. На фотографии (А) показан профиль обратимо-фазной (C4) HPIC BAIB/MK миогенной активности. На фотографии (В) представлены данные электрофорезного (NаДоdSО4/PAGE) анализа фракций, выбранных со стадии хроматографии С4, показанной на фотографии А, демонстрирующие тот факт, что пиковые HPIC фракции содержат единственную полосу на геле, окрашенном серебром. На фотографии (С) представлен график синтеза ДНК в клетках BAIB/MK, под действием фракций проанализированных на фотографии В, причем полученные результаты показывают, что относительная митогенная активность хорошо корредирует с интенсивностью протеиновой полосы в профиле активности.
На рис.I-3 показана альтернативная стадия с помощью очистки RP-HPIC, с использованием ситовой хроматографии с операциями на колонке (ТSК C3000SW GIaSPac) в водном растворе со значением рН, близким к значению физиологического раствора, в результате чего основной пик митогенной активности обнаруживается в BAIB/MK биологическом анализе.
На рис.I-4 приведено сравнение синтезов ДНК в BAIB/MK при воздействии ТSК-очищенного митогена и других факторов роста.
На рис.I-5 представлены данные сравнения роста клеток BAIB/MK в химически установленной среде при воздействии различных комбинаций факторов роста.
В таблице 1-1 суммированы результаты различных стадий очистки, демонстрирующие, что ситовая хроматография обеспечивает значительно лучшую регенерацию активности, чем использование адсорбционной RР-НPIС.
В таблице 1-2 суммированы данные по специфичности действия различных факторов роста в отношении клетки-мишени, демонстрирующие, что новый выделенный фактор обладает сильным митогенным действием на кератиноциты (BAIB/MK) и в резком отличие от этого не обнаруживает детектируемого действия на фибробласты или эндотелиальные клетки подкожной вены человека.
На рис.II-1 представлена нуклеотидная последовательность и установленная аминокислотная последовательность KCF сДНК, а также идентификация РНК транскрибированных из KCF гена. На фотографии (А) схематично представлены клоны сДНК человеческого KCF. На фотографии (В) изображен KCF СДНК нуклеотид и предсказанные аминокислотные последовательности. (C) - идентификация матриц РНК KCF генов анализом на окрашивание, по Нортерну.
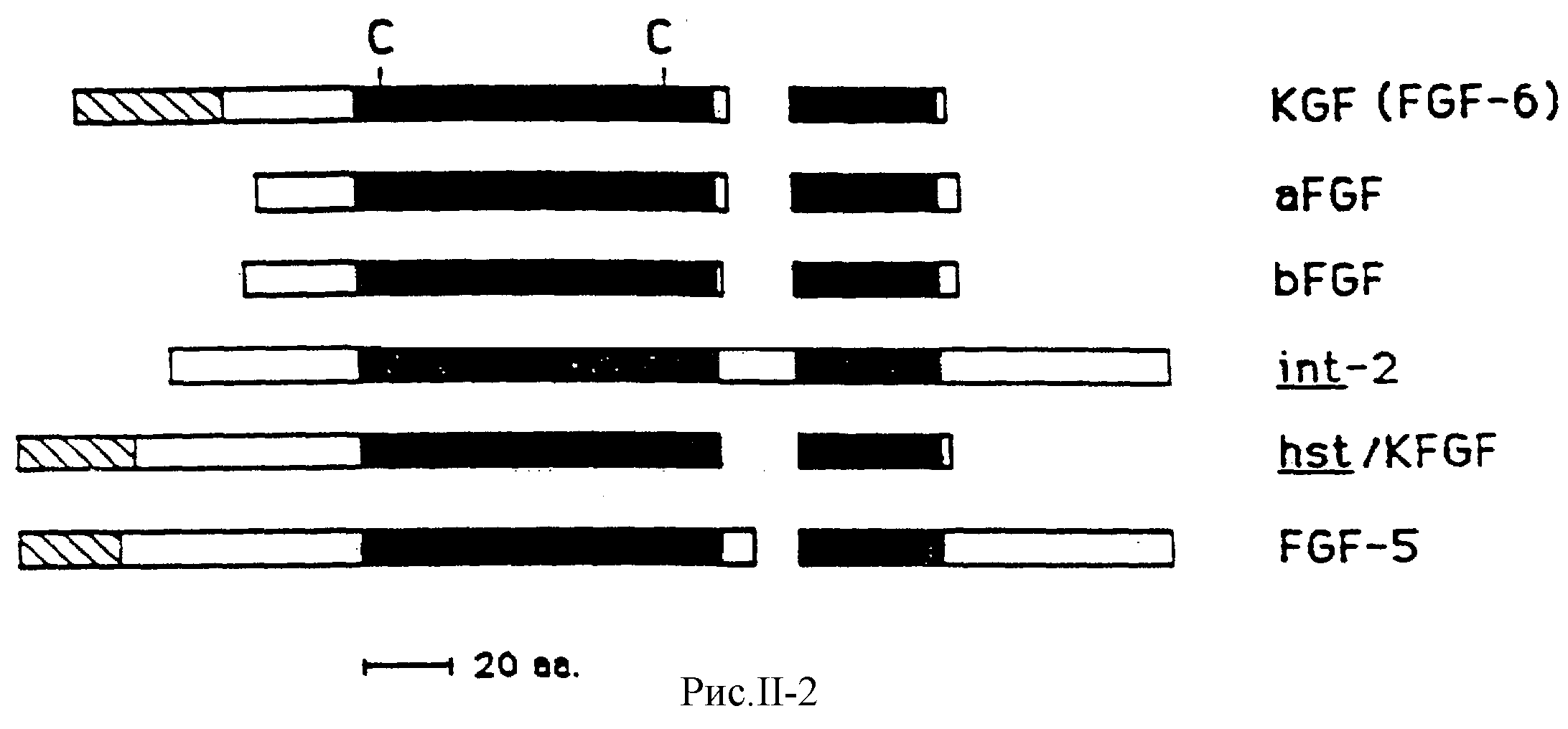
На рис.II-2 представлены данные топологического сравнения семейства FCF родственных молекул, включая KCF, причем особое внимание уделено тем протеиновым областям, которые разделяют высокую гомологию, предполагаемым сигнальным пептидным последовательностям и двум консервированным цистеиновым остаткам.
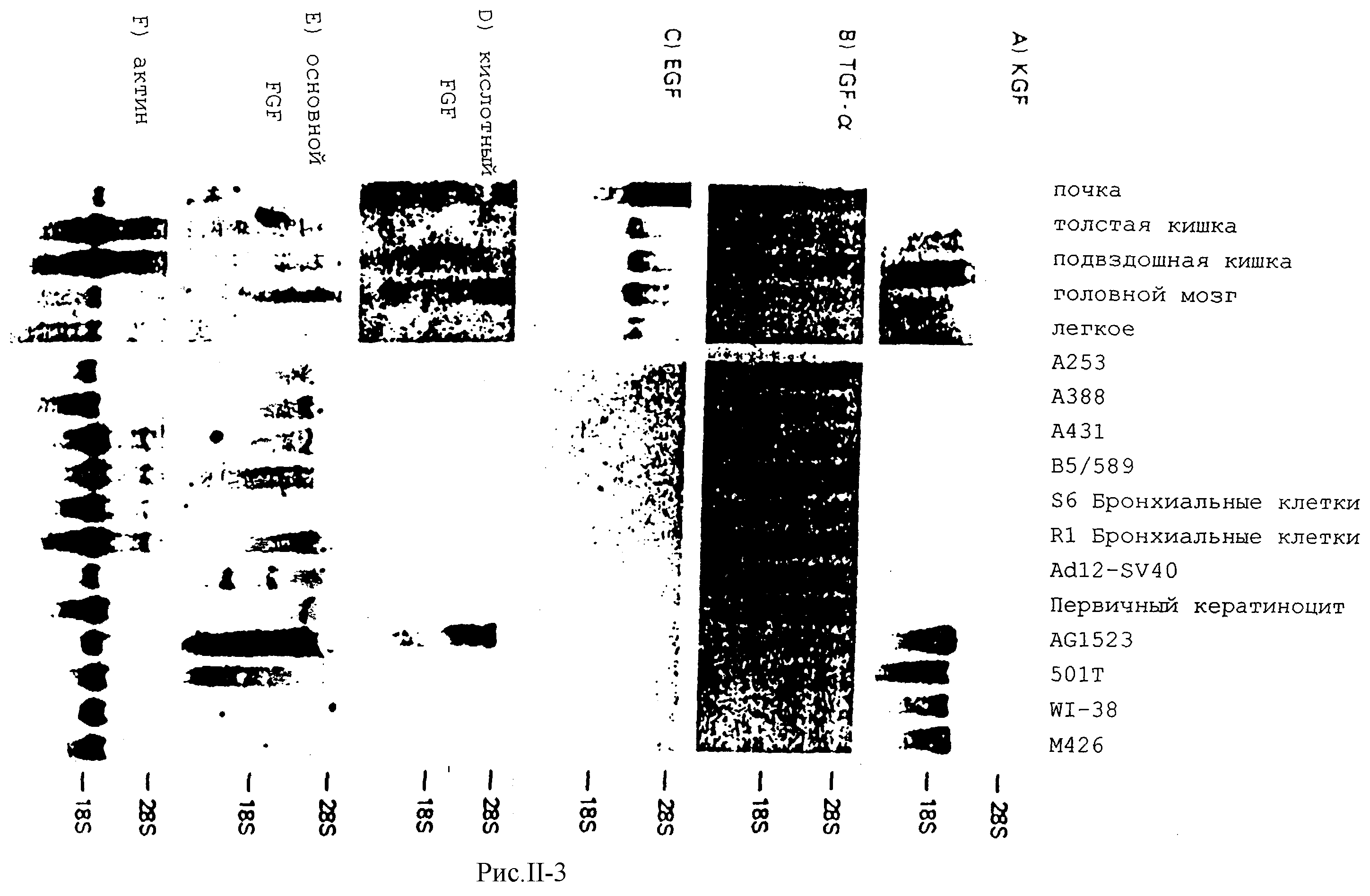
На рис.II-3 представлены данные анализа (окрашивание по Нортерну) экспрессии KCF-родственной мРНК в отобранных нормальных человеческих клеточных линиях и тканях, которые позволяют установить, что одна матрица размером 2,4 кb присутствует в РНК из человеческих эмбрионных легочных фибробластов и из фибробластов кожи взрослых особей, тогда как матрица не была обнаружена в (В5/589) эпителиальной или (НА83) глиальных клеточных линиях, или в первичных культурах эндотелиальных клеток подкожной вены человека.
В таблице II-1 суммированы данные сравнения влияния гепарина на митогенную активность KCF с соответствующим влиянием на другие факторы роста, причем показано, что введение тимидина в ДНК с помощью клеток BAIB/MK при воздействии КCF ингиибируется гепарином в отличие от активностей как aFCF, так и bFCF, которые повышаются при такой обработке.
Описание конкретных воплощений.
Настоящее изобретение частично относится к очищенным KCF- или KCF-подобным протеинам и способам получения таких протеинов. Главное воплощение такого аспекта настоящего изобретения относится к гомогенному KCF, характеризующемуся кажущимся молекулярным весом порядка 28 кДа на основании миграции в NаДоdSО4/РАСЕ, движением одним пиком при анализе обратимо-фазной хроматографией высокого разрешения и удельной активностью по крайней мере 3,4×104 единиц на мг, предпочтительно, по крайней мере 3,2×105, единиц на мг, причем одна единица активности определяется как количество, вызывающее половину максимально возможной стимуляции синтеза ДНК в некоторых эпителиальных (кератиноцит) клетках в стандартных условиях анализа указанных ниже.
Для идентификации новых факторов роста специфичных к эпителиальным клеткам, клональную кератиноцитную клеточную линию мышей разновидности ВАIВ/с, обозначенную как ВАIВ/MK (I-6), использовали в качестве индикаторной клетки для детекции таких факторов. Такие клетки зависимы от их роста на экзогенном источнике митогена эпителиальной клетки даже в среде, содержащей сыворотку (I-6). Разработка химически определенной среды для таких клеток делает возможным демонстрацию того факта, что для пролиферации BAIB/MK требуется два главных митогенных пути. Один из них предполагает участие инсулиноподобного фактора роста I (или инсулина в высокой концентрации), а другой - эпидермального фактора роста (ЕСF), трансформирующего фактора роста а (ТСFа), фактора роста кислотного фибробласта (АFСF) или фактора роста основного фибробласта (bFВF) (I-7).
С использованием BAIB/MK в качестве прототипной эпителиальной клеточной линии и NIH/3ТЗ в качестве ее фибробластовой противочасти, кондиционированную среду из различных человеческих клеточных линий анализировали на наличие новых специфичных митогенов эпителиальных клеток. Такие методы биологического анализа настоящего изобретения позволяют осуществлять очистку до гомогенности одного из таких новых факторов роста, выделяемого человеческой эмбрионной легочной фибробластовой линией и обозначенного как фактор роста кератин опита (KCF).
Вкратце, биоанализ на KCF-подобную активность в стандартных условиях включает следующие стадии:
(I) Мышинные кератиноциты (BAIB/MK клетки) выращивают в культуре до слияния и затем выдерживают в течение 24-72 часов в среде не содержащей сыворотки;
(II) После добавления испытуемых образцов стимуляцию синтеза ДНК определяют путем введения3 Н-тимидина в кислотно-осаждаемую ДНК.
С целью определения специфичности действия митогенного фактора роста в отношении клетки-мишени, стимуляцию синтеза ДНК, выраженную отношением стимулированного синтеза к фоновому внедрению тимидина в отсутствие добавленного испытуемого образца, можно сравнить с аналогичной стимуляцией наблюдаемой в клетках отличных от кератиноцитов в аналогичных условиях анализа. В рамках такого сравнения было установлено, что митогенная активность KCF имеет ярко выраженную специфичность в отношении кератиноцитов в отличие от специфичности в отношении фибробластов (по крайней мере, 500-кратное увеличение стимуляции) и наблюдается меньшая, но значительно (по крайней мере, 50-кратная) более высокая активность по отношению к кератинопитам, чем по отношению к другим типам эпителиальных клеток (см. таблицу I-2 и раздел Материалы и методы исследования в Экспериментальном разделе I для более детального ознакомления со стандартными условиями биоанализа).
В результате использования метода продуцирования KCF, включающего культурирование клеток и выделение митогенной активности, причем такой метод включает ультрафильтрацию, гепарин-Сефарозную сродственную хроматографию (НSАС) и адсорбционную обратимофазную жидкостную хроматографию высокого разрешения (RР-НРIC), или согласно другому варианту HPIC на молекулярных ситах (TSK-HPIC), согласно настоящему изобретению было выделено количество препарата, достаточное для достаточного охарактеризования физических и биологических свойств такой молекулы.
Таким образом, способ получения KCF из таких продуцирующих клеток, как М426 человеческие эмбрионные фибробласты (I-8), включают, например, следующие стадии:
(I) Приготовление кондиционированной среды (например, 10 л) с использованием монослойных культур, полученных циклической обработкой в системе от среды, не содержащей сыворотки, до среды, содержащей ее, и хранение не содержащего сыворотки урожая при -70°С до последующего использования.
(II) Концентрирование ультрафильрацией с использованием мембран с молекулярным весом 10 кДа в несколько последовательных стадий при разбавлении в буффере (с целью облегчения удаления низкомолекулярных материалов), с последующим необязательным хранением при -70°С.
(III) Хроматографию по сродству на гепарине, связанном с полимерной подложкой (например, Сефарозой) при элюировании градиентом, повышающей концентрации NaCl.
(IV) Концентрирование до 10-20 кратного уменьшения объема с помощью мелкомасштабных ультрафильтрационных устройств с запором из материала с молекулярным весом 10 кДа (например, микроконцентратор Центрикон-10 от Амикон) и хранение при -70°С.
Следующая стадия процесса очистки включает приведенные ниже стадию (V) или (VI):
(V) Обратимо-фазная HPIC активных фракции (0,6М NаСl) с предыдущей стадии HSAC в системе органических растворителей, или
(VI) Молекулярно-ситовая HPIC (например, на ТSK-С3000 Глас-Пак колонке фирмы ЛКБ) в водном буфере при значении рН, близком к физиологическому (например, Трис-HCI, рН 6.8/0,5М NaCl) с последующим хранением при -70°С.
Препарат, полученный на стадии TSK (VI) имеет практически ту же чистоту, что и препарат, полученный с использованием RР-HPIC, согласно анализу серебрянного окрашивания NаДоdSO4/РAGЕ (данные не приведены). Однако использование ТSK обеспечивает значительно лучшую регенерацию активности (таблица I). Кроме этого, очищенный методом TSK материал обладал более высокой удельной активностью, чем материал, полученный методом RР-HPIC, KCF, полученный по описанной выше методике с участием ТSК, стимулирует синтез ДНК в эпителиальных клетках в субнаномольных концентрациях, но не индуцирует внедрение тимидина в ДНК фибробластов или эндотелиальных клеток при сравнимых или более высоких концентрациях (вплоть до 5 нМ). Такая активность чувствительна к кислотам, теплу и растворителям, используемым на стадии RP-HPIC (см. данные по чувствительности в разделе у Экспериментальной части, а также дополнительные подробности способа получения).
С использованием стандартной методологии, хорошо известной из литературы, была установлена аминокислотная последовательность для положений 2-13 из амино-окончания очищенного KCF: Asn-Asp-Met-Thr-Pro-Glu-Gln-Met-Ala-Thr-Asn-Val (см. Экспериментальный раздел I).
Настоящее изобретение также включает ДНК сегменты, кодирующие KCF и KCF-подобные полипептиды. Примерами ДНК настоящего изобретения могут служить следующие материалы: клоны 32-49 человеческой сДНК, полученные из полиаденилированных РНК, экстрагированных из клеточной линии М426 человеческого эмбрионного легочного фибробласта; рекомбинанты и метанты таких клонов; а также родственные ДНК сегменты, которые могут детектироваться гибридизацией в такие сегменты ДНК.
Как указывается в Экспериментальном разделе II, с целью поиска клонов сДНК, соответствующих известной части аминокислотной последовательности KCF, создавали две партии олегонуклеотидных зондов, базируясь на всех возможных нуклеотидных последовательностях, кодирующих девятую аминокислотную последовательность, Аsn-Аsn-Меt-Тhr-Рro-Glu-Gln-Met-Ala. Библиотеку сДНК конструировали в клонирующем векторе сДНК, λрСЕУ9, с использованием полиаденилированной РНК экстрагированной из клеточной линии М426 человеческого эмбирионного легочного фибробласта, которая представляла собой исходный источник фактора роста. В результате скрининга библиотеки (9×105 бляшек) с помощью32 Р-меченных олигонуклеотидов идентифицировали 88 бляшек, которые гибридизировали с обоими зондами.
Из 10 очищенных от бляшек клонов, которые подвергали анализу, один, обозначенный как клон 49, имел вставку сДНК в 3,5 кb, тогда как остальные имели вставки в интервале 1,8-2,1 кb. Анализ более мелких клонов позволил обнаружить несколько общих ограничительных сайтов, а установление аминокислотной последовательности одного из представителей мелких клонов, обозначенного, как клон 32, совместно с клоном 49, показано, что они представляют собой перекрывающиеся сДНК (рис.II-IA). Рассмотрение двух таких сДНК позволило установить наличие непрерывной последовательности в 3,8 кb, содержащей полную KCF кодирующую последовательность. Нуклеотидная последовательность чувствительного тяжа ДНК и закодированная предсказанная последовательность первичного протеина представлены для композитной KCF сДНК последовательности полной длины на рис.II-IB.
Такие ДНК, сДНК клоны 32 и 49, а также рекомбинантные формы таких сегментов, включающие полную KCF кодирующую последовательность, являются наиболее предпочтительными ДИК настоящего изобретения.
Из последовательности сДНК ясно следует, что продукты трансляции первичного KCF и hst содержат гидрофобные N-терминальные участки, которые, по-видимому, являются сигнальными последовательностями на основании схожести таких последовательностей в большом числе других протеинов. В соответствии с этим, такой Н-терминальный участок не присутствует в молекуле очищенного зрелого КСF, которая секретируется человеческими эмбрионными фибробластами.
Кроме того, KCF разделяет со всеми другими членами FСF семейства два основных участка гомологии, аминокислоты 65-156 и 162-189 в предсказанной последовательности KCF, которые разделены негомологическими сериями аминокислот различной длины у различных членов семейства. Последовательность очищенной формы KCF содержит пять цистеиновых остатков, два из которых сохраняются во всем семействе FСF-родственных протеинов. В последовательности KCF встречаются пять пар основных остатков. Та же картина наблюдается и у других членов семейства FCF.
Специалисту в данной области должно быть понятно, что в результате использования ДНК и РНК настоящего изобретения в методах гибридизации (таких как анализ геномных человеческих сДНК пятнением по Сaутерну), особенно наиболее предпочтительных ДНК перечисленных выше, без нежелательного экспериментирования можно осуществить скрининг геномной или сДНК библиотек с целью отыскивания других KCF подобных ДНК, охватываемых сферой настоящего изобретения. Кроме того, в результате такого использования ДНК настоящего изобретения, генетические маркеры, ассоциированные с КСF геном, такие как рестрикционные фрагменты длины полиморфизмов (PFIP), могут быть идентифицированы и связаны с унаследованными клиническими состояниями, включающими такие или близкие к ним гены.
Настоящее изобретение также включает модифицированные формы KCF ДНК. В соответствии с главным воплощением такого аспекта изобретения такие модифицированные ДНК кодируют KCF-подобные протеины, включающие сегменты аминокислотных последовательностей KCF, по крайней мере, один другой член семейства пептидов FCF, Так, например, поскольку не имеется существенной N-терминальной гомологии между секретированной формой KCF и аналогичными положениями в других FСF-росдственных протеинах, полипептиды с новыми структурными и функциональными свойствами могут быть созданы прививкой ДНК сегментов кодирующих N-терминальные сегменты другого полипептида в семействе FCF на сегмент ДНК КCF вместо его обычной NН2-терминальной последовательности.
Полипептидные химеры, продуцированные такими модифицированными ДНК, используются для определения достаточности NH2-терминального участка КСF в объяснении его уникальной специфичности в отношении клетки-мишени. Исследование таких химер должно также обеспечить информацию относительно природы участков, определяющей различное влияние гепарина на их биологическую активность.
Разумеется, что применимость такого подхода была уже подтверждена успешной генной инженерией и экспрессированием химерной молекулы, в которой около 40 аминокислот от NH2-окончания секретированной формы KCF (начиная от цисаминоокончания зрелого KCF, под номером 32 на рис.II-1 и кончая arg, остатком 78 КСF) связано со 140 аминокислотами СО2-окончания оболочки аFСF (начиная с остатка 39, arg и продолжаясь до С-окончания аFСF кодирующей последовательности. Такой химерный продукт превосходит кератиноциты, например KCF, по отношению к клетке-мишени, но уступает в восприимчивости к гепарину, что делает его более похожим на аFCF, а не на KCF. Такой новый КСF-подобный фактор роста может иметь преимущества в клинической практике, где желательно применение питедиально-специфичного фактора роста в присутствии гепарина, общепринятого антикоагулянта. Дополнительные детали конструкции такой химерной молекулы и свойства полипептида описаны в Экспериментальном разделе II.
Другие ДНК настоящего изобретения включают следующие молекулы рекомбинантной ДНК, содержащие KCF сДНК и любую из следующих векторных ДНК; клонирующий вектор бактериофага λ (λрСЕУ9), вектор устанавливающей аминокислотную последовательность плазмиды ДНК (вариант рУС), вектор бактериальной экспрессии (рКК233-2), вектор экспрессии млекопитающих (рММТ/neo). Такие рекомбинантные ДНК представлены примерами конструкций в Экспериментальных разделах.
Наиболее предпочтительные рекомбинантные молекулы включают: молекулы, содержащие последовательность, кодирующую секретированную форму KCF, и вектор бактериальной экспрессии (например, рКК233-2) или сДНК, кодирующую весь продукт первичной трансляций (включая NH2-терминальный сигнальный пептид), а также вектор экспрессии млекопитающего (например, рМMMT) способный экспрессировать вставленную ДНК в клетках млекопитающих (например, NIH/3Т3).
Конструкция рекомбинантных ДНК, содержащая КСF-ДНК и вектор бактериальной экспрессии, описана в экспериментальном разделе II. КСF сДНК была экспрессирована с продуцированием полипептида в E. coli путем помещения его кодирующей последовательности под контролем гибридного trk промотора в плазмидный экспрессионный вектор рКК233-2/(II-31).
Конструирование рекомбинантной ДНК, содержащей КСF-ДНК и вектор млекопитающего, способный к экспрессии вставленной ДНК в культивированных человеческих или животных клетках, можно осуществлять по стандартной технологии генной экспрессии с использованием хорошо известных методов, применимых для экспрессирования такого относительно простого полипептида. Одно конкретное воплощение рекомбинантной ДНК такого аспекта настоящего изобретения, включающее вектор млекопитающего рММТ, описано ниже в разделе, касающемся рекомбинантных клеток настоящего изобретения.
ДНК и РНК с чувствительными цепями настоящего изобретения могут использоваться совместно с методами продуцирования протеинов для получения большого количества практически чистых KCF или КСF-подобных протеинов. Полученный таким образом практически чистый протеин КСF может применяться, с использованием хорошо известных методик, в диагностических анализах на определение наличия рецепторов для такого протеина в различных образцах ткани и жидкостной среды тела.
В соответствии со сказанным, настоящее изобретение также охватывает клетку, предпочтительно клетку бактерии или млекопитающего, трансформированную с помощью ДНК изобретения, в которой трансформирующая ДНК способна к экспрессированию. Согласно предпочтительному воплощению такого аспекта изобретения клетка трансформированная ДНК изобретения производит протеин КCF в полностью митогенной форме. Наиболее предпочтительно, когда такие протеины находятся в секретированной форме (например, у них отсутствует сигнальная последовательность). Такие протеиновые факторы могут использоваться для функциональных исследований и могут быть очищены для дополнительного биохимического и функционального анализа, например качественный и количественный анализ на связывание рецептора.
Рекомбинантные E. coli клетки конструировали в бактериальном экспрессионном векторе, рКК233-2, для получения KCF, как это подробно описано в Экспериментальном разделе II. Некоторые рекомбинантные бактериальные клоны испытывали на продуцирование протеина обычными мелкомасштабными методами. Все испытанные рекомбинанты синтезировали протеины, о чем свидетельствовали антитела, возникающие у аминотерминального KCF пептида (см. ниже). Один рекомбинант выращивали в одном литре культуры, которая продуцировала рекомбинантный KCF, который эффективно стимулировали внедрение тимидина в ДНК клепки к ВАIВ/MK кератиноцита, но проявляя лишь незначительную активность в отношении NIH/3Т3 фибробластов. Половина максимальной стимуляции клеток BAIB/MK в стандартном биоанализе на керетиноцит достигалась при концентрации 2-5 нг/мл, по сравнению с концентрацией 10-15 кг/мл для КCF очищенного от клеток М426.
Один литр бактериальных клеток давал примерно 50 мкг моноочищенного рекомбинантного KCF. Специалисту в области генной экспрессии должно быть ясно, что такой первоначальный выход может быть значительно улучшен без нежелательного дополнительного экспериментирования путем применения различных известных приемов рекомбинантной ДНК технологии.
Рекомбинантные клетки млекопитающего (NIН/3Т3 также были сконструированы с использованием полной KCF сДНК последовательности (включая NН2-терминальный сигнальный пептид) и вектора рММТ/nео, который несет мышинный металлотиониновый (ММТ) промотор и селективный маркерный ген на устойчивость к неомицину. Такие клетки оценивали на продуцирование KCF. Особенно на секрецию зрелой формы (отсутствие сигнального пептида) продуцируемой человеческими фибробластами с использованием биоанализов настоящего изобретения. Такая комбинация вектора и клетки-хозяина были с успехом использованы для экспрессирования некоторых других аналогичных рекомбинантных полипептидов, включая цепи факторов роста А и В, являющихся производными тромбоцитов (РДСЕ, II-32). Таким образом, специалисту в данной области должно быть ясно, что высокие выходы рекомбинантного KCF могут быть достигнуты таким образом с использованием указанных выше рекомбинантных ДНК и трансформированных клеток настоящего изобретения.
Крупномасштабное производство может использоваться для клинических испытаний в условиях, требующих специфической стимуляции роста эпителиальных клеток. Материалы и методы получения фармацевтических композиций для локального применения полипептидов (на коже или роговице глаза) или их системного применения хорошо известны в данной области и они могут быть легко приспособлены для применения КСF и KCF-подобных пептидов без дополнительного экспериментирования.
Настоящее изобретение также охватывает новые антитела против пептида закодированного сегментом ДНК изобретения. Такое воплощение изобретения представлено примерами нескольких видов антител, которые распознают KCF. Они были получены с использованием стандартных методов хорошо известных в данной области экспериментальной иммунологии, как указывается в экспериментальном разделе II. Такие антитела включают: моноклональные антитела, вырабатываемые мышами против неповрежденного очищенного протеина из человеческих фибробластов; поликлональные антитела, вырабатываемые кроликами против синтетических пептидов с последовательностями на основе аминокислотных последовательностей, предсказанных из последовательности KCF сДНК (примеры которых может служить пептид с последовательностью остатков KCF 32-45, а именно, NДМТРЕОМАТNVR (с использованием стандартного однобуквенного кода для аминокислотных последовательностей, см. фиг.II-1)); поликлональные антитела, вырабатываемые кроликами против как природно-секретированного KCF из человеческих фибробластов, так и рекомбинантного КCF, продуцированного в E. coli; (см. выше). Все испытанные антитела распознают рекомбинант, а также встречающийся в природе КCF, как с помощью твердофазного (ЕIISА) анализа, так и анализа пятнением по Вестерну. Некоторые антитела, которые представляют собой предпочтительные антитела настоящего изобретения, предназначены для нейтрализации митогенной активности KCF в BAIB/MK биоанализе.
Фрагменты антител настоящего изобретения, такие как Fab или F/аb/’, которые сохраняют антигенсвязывающую активность и могут быть получены способами хорошо известными в данной области также охватываются сферой настоящего изобретения. Кроме этого, настоящее изобретение охватывает фармацевтические композиции антител изобретения или их активные фрагменты, которые могут быть получены с использованием материалов и методов, предназначенных для получения фармацевтических композиций для применения полипептидов, которые хорошо известны в данной области и могут быть легко адаптированы к применению КСF и КCF-подобных пептидов без дополнительного экспериментирования.
Такие антитела и их активные фрагменты могут использоваться, например, для детекции KCF в биоанализах или для очистки протеиновых факторов. Они могут также применяться в методах, хорошо известных в данной области для выделения рецептора на KCF, который, как указывается в Экспериментальном разделе II, по-видимому, отличается от рецепторов всех других известных факторов роста.
Такие предпочтительные антитела и фрагменты, а также их фармацевтические комбинации, которые нейтрализуют митогенную активность КCF в отношении эпителиальных клеток, о чем свидетельствует BAIB/MK анализ, могут использоваться для лечения клинических состояний, характеризующихся избыточным ростом эпителиальных клеток, включающих дисплазию и неоплазию (например, псориаз, или злокачественные либо доброкачественные опухоли).
Настоящее изобретение также охватывает новые методы биоанализа для детекции генов родственных ДНK настоящего изобретения. В некоторых примерах воплощения изобретения, ДНК настоящего изобретения использовали в качестве зондов для определения промежуточных уровней содержания родственных мРНК. Методы такого биоанализа настоящего изобретения с использованием KCF ДНК, а также стандартные методы окрашивания по Нортону подробно описаны в Экспериментальном разделе II.
Специалист в данной области без дополнительных экспериментов сумеет понять, что такие методы могут быть легко применены для анализа генной экспрессии для KCF подобного протеина как в изолированных клетках, так и в различных тканях. Такие биоанализы могут, например, использоваться для идентификации различных классов опухолевых клеток или генетических дефектов в процессах эпителиального роста.
Предполагается, что средний специалист в данной области, используя предыдущее описание и следуя методам, описанным ниже в Экспериментальных разделах, сможет применить настоящее изобретение в его наиболее полном контексте. Материал, изложенный в Экспериментальных разделах, если особо не оговорено, приведен в целях иллюстрации и никоим образом не ограничивает прилагаемую формулу изобретения.
Экспериментальный раздел I
Идентификация и охарактеризование нового фактора роста, специфичного в отношении эпителиальных клеток.
В этом разделе описывается экспериментальная работа, приводящая к идентификации фактора роста, специфичного в отношении эпителиальных клеток в кондиционированной среде клеточной линии человеческого эмбрионного легочного фибробласта. Фактор, условно названный фактором роста кератиноцита (KCF) из-за его доминирующей активности в отношении клеток такого типа, очищали до гомогенности комбинацией методов ультрафильтрации, гепарин-Сефарозной сродственной хроматографии и гидрофобной хроматограции на С4 обратимо-фазной HPIC колонке согласно методам настоящего изобретения. Было обнаружено, что KCF лабилен в отношении действия кислот и тепла и состоит из одной полипептидной цепи с кажущимся молекулярным весом примерно 28000 дальтонов. Очищенный KCF представляет собой мощный митоген в отношении эпителиальных клеток, способный стимулировать синтез ДНK в находящихся в покое ВАIВ/МК эпидермальных кератиноцитах с более чем 500-кратным повышением активности при 0,1 нМ и максимумом при 1,0 нМ. Отсутствие митогенной активности как в отношении фибробластов, так и эндотелиальных клеток указывает на то, что KCF обладает специфичностью в отношении клетки-мишени, которая отличается от предварительной охарактеризованного фактора роста. Установление аминокислотной последовательности микрометодами позволило обнаружить аминотерминальную последовательность, не содержащую значительной гомологии с любым известным протеином. Выделение такого нового фактора роста человеческими эмбрионными фибробластами указывает, на то, что KСF играет определенную роль в мезенхимальной стимуляции пролиферации нормальной эпителиальной клетки.
Способ и материалы
Приготовление кондиционированной среды.
Ранний перенос М426 человеческих эмбрионных фибробластов (1-8) высевали в Т-образных склянках площадью 175 см2 и выращивали в течение 10-14 дней в Дульбекко модифицированной среде Игла (ДМЕМ, CIBCO) дополненной 10% телячей сывороткой (CIBCO). После завершения операции монослои в течение недели подвергали циклической обработке в диапазоне сред от содержащей до не содержащей сыворотки, причем последняя среда содержала только ДМЕМ. Клетки дважды промывали 5 мл фосфатного буфферного раствора перед добавлением 20 мл ДМЕМ. Через 72 часа культивированные жидкости собирали и заменяли на 35 мл среды, содержащей сыворотку. Кондиционированную среду хранили при -70°С до последующего использования.
Ультрафильтрация.
Примерно 10 л кондиционированной среды оттаивали, предварительно фильтроали через 0,5 микронный фильтр (Миллипор НАWР 142 50) и концентрировали до объема 200 мл с использованием кассетной системы (Миллипор ХХ42 00K 60) и кассетного запора, имеющего молекулярный вес 10 КДа Миллипор РТСС 000 05). После концентрирования образец подвергали последовательному разбавлению одним литром 20 мМ Триc-HCl, рН 7.5/0,3М NaCl, причем каждую стадию такого разбавления сопровождали стадией ультрафильтрации с использованием системы Пелликон. Активность, регенерированную в ретентате, сразу применяли на гепарин-Сефарозной смоле или хранили при -70°С.
Гепарин-Сефарозное сродство
Хроматография (НSAC).
Ретентат со стадии ультрафильтрации загружали на гепарин-Сефарозную смолу (Фармация), которую уравновешивали в 20 мМ Трис-HCI, рН 7.5/0,3М NаСl. Смолу интенсивно промывали до возвращения оптической плотности к фоновому значению и затем подвергали воздействию линейного градиента с увеличивающимися концентрациями NаСl. После удаления аликвот из фракций для биоанализа с внедрением тимидина, отобранные фракции концентрировали до уменьшения объема в 10-20 раз с помощью микроконцентратора Центрикон-10 (Амикон) и хранили при -70°С.
Обратимо-фазная HPIC (RP-НРIС).
Активные фракции (0,6м NаСl слив) со стадии HSАС оттаивали, сливали и дополнительно концентрировали с помощью Центрикона-10 до конечного объема ≤200 мкл. Образец загружали в колонку Видак С4 HPIC (Группа разделения, Гесперия, СА), которую уравновешивали в 0,1% трифторуксусной кислоте (ТFА, Флюка) 20% ацетонитрила (Байкер, сорт HPIC) и элюировали линейным градиентом увеличивающихся концентрацией ацетонитрила. Аликвоты для биоанализа немедленно разбавляли 10-кратным избытком 50 мкг/мл BSIA (Фракция У, Сигма) 20 мМ TPc-HCI pH 7.5. Оставшуюся часть образца сушили в системе Спид-Вак (Савант) в препарате для структурного анализа.
Молекулярно-ситовая HPIC.
Примерно 50 мкл дважды концентрировали гепарин-Сефарозных фракций загружали в ТSK-С30000 Глас-Пак колонку (ЛКБ), которую уравновешивали в 20 мМ Трис-HCI, pH 6, 8/0,5M NaCl. Образец элюировали в указанный буфер с линейной скоростью 0,4 мл/мин. После удаления аликвот для биоанализа, фракции хранили при -70°С.
NаДоdSO4 - Полиакриламидный гель электрофорез (NаДоdSO4/РАСЕ)
Полиакриламидные гели получали в присутствии NаДоdSО4 согласно методике Лазммли (I-9). Образцы в течение 3 минут кипятили в присутствии 2,5% 2-меркаптоэтанола (об. об.). Гели фиксировали и окрашивали серебром (I-10) с использованием реагентов и методики согласно БиоРад. Молекулярновесовые маркеры получали от фирмы Фармации.
Стимуляция синтеза ДНК.
96-пластин для микротитрования (Фалкон №3596) предварительно покрывали человеческим фибронектином (Коллаборэйтив Рисерч) с плотностью 1 мкг/см2 перед засеванием клетками BAIB/MK. После этого клетки выдерживали в течение 24-72 часов в среде, не содержащей сыворотки и содержащей 5 мкг/мл трансферина (Коллаборэйтив Рисерч) и 30 нМ Nа2SeО3 (Бэйкер).
Внедрение3Н-тимидина (конечная концентрация 5 мкС (мл, NЕN) в ДНК измеряли в течение 6-чаоового периода, начинающегося через 16 ч после добавления образцов. Анализ прекращали промыванием клеток, охлажденным на льду фосфатно-буферным раствором и двойным промыванием 5%-ным раствором трихлоруксусной кислоты. Осадок повторно растворяли в 0,25М NаОН, переносили в жидкую сцинтилляционную жидкость (Биофлуор, NЕN) и проводили подсчет.
За стимуляцией синтеза ДНК следи в соответствии с методикой, описанной выше для клеток ВАIВ/MK с использованием большого числа других клеточных линий. NIH/3T3 фибробласты (I-11) получали от Национального Института здоровья, тогда как бронхиальные эпителиальные клетки резусных обезьян CC1208 (I-12) получали из Американской коллекции типовых культур. Клеточную линию эпителия грудной железы человека В5/589, приготовленную согласно описанному в ссылке (I-13), получали от Марта Стампфер (Калифорнийский Университет, Беркли). Клетки грудной железы выращивали в RРМI 1640 дополненной 10% сыворотки телячьего плода и 4 нг/мл ЕСF. После выдерживания в бессывороточных условиях основная среда представляла собой ДМЕМ. Первичные культуры эндотелиальных клеток подкожной вены человека готовили и выдерживали в соответствии с описанным в ссылке (I-14). Фактор эпидермального роста и инсулин получали из Коллаборэйтив Рисерч. Кислотный FCF и bFСF получали из Калифорния Биотехнолоджи, Инк. Рекомбинантный ТСFа получали из Генетикс Инк. Среду и сыворотку получали либо от ГИБКО, Биофлюидс, Инк. или среда представляла собой среду NIН.
Анализ на пролиферацию.
Чаши для культивирования диаметром 35 мм предварительно последовательно покрывали поли-Д-лизином (20 мкг/см3 (Сигма) и человеческим фибропектином и затем засеевали 2,5×104 клеток BAIB/MK. Основная среда представляла собой смесь в соотношении 1:1 минимальной существенной среды Игла с низким содержанием Ca2+ и среды FI2 Хамса, дополненную 5 мкг/мл трансферина, 30 нМ Nа2SеО3 и 0,2 мМ этаноламина (Сигма). Среду заменяли каждые 2-3 дня. Через 10 дней клетки закрепляли в формалине (Фишер Сайнтифик Ко.) и Окрашивали с помощью Гиемза (Фишер Сайнтифик Ко.).
Микроопределение последовательности протеина.
Примерно 40 мкг (150 пмоль) протеина из активных фракций с колонки С4 повторно растворяли в 50% ТFА и загружали в устройство для газофазного определения аминокислотной последовательности протеина фирмы Эпплайл Биосистемс. Проводили двадцать циклов деградации по Эдману и идентификацию аминокислотных производных осуществляли с помощью автоматизированной HPIC /Модель 120A, Эпплайл Биосистемс/.
Результаты.
Детекция и выделение фактора роста.
Предварительный скрининг кондиционированной среды из различных клеточных линий показал, что среда из некоторых фибробластовых линий содержит митогенную активность, детектируемую как на клетках ВАIВ/MK, так и NIН/3Т3. В то время как кипячение разрушает активность у ВАIВ/MK, митогенная активность клеток NIH/3Т3 остается незатронутой. Основываясь на известных данных о том, что ЕСF (I-15) и ТСFа (I-16) являются термически стабильными материалами, разумно предположить, что ВАIВ/MK миогенная активность связана с наличием агента отличного от известных факторов роста эпителиальных клеток.
Линию человеческого эмбрионного легочного фибробласта М426 выбирали в качестве наиболее продуктивного источника такой активности для очистки факторов роста. Ультрафильтрация с помощью системы Пелликон обеспечивает удобный путь уменьшения объема образца до значения, подходящего для последующей хроматографии. В ходе разработки схемы очистки испытывали различные комбинации ситовой, ионообменной хроматографии и хроматографии с изоэлектрической фокусировкой. Однако в результате получали недопустимо низкие выходы. С другой стороны - стороны гепарин-Сефарозная сродственная хроматография (НSAС), которую использовали для очистки других факторов роста (I-17 I-22), оказалась полезной в качестве ранней стадии очистки настоящего изобретения. Хотя оценки регенерированной удельной активности на этой стадии были неточными из-за возможного присутствия других факторов, кажущийся выход активности составил 50-70% с соответствующим обогащением примерно в 1000 раз.
Как показано на рис.I-1, более 90% ВAIВ/МК митогенной активности элюируется с колонки HSAC с помощью 0,6М NaCl. Такой пик активности не связан с какой-либо активностью на клетках NIН/3Т3 Сданные не показаны). Значительно меньший пик ВАIВ-MK митогенной активности проявляется в присутствии 0,8-1,2М NaCl.
В связи с воспроизводимостью образца НSАС, активные фракции могут быть идентифицированы на основе профиля градиента и оптической активности. 10-20 кратное концентрирование с помощью Центрикона-10 оказалось существенным для стабильности, которая могла сохраняться при -70°С в течение нескольких месяцев.
Окончательную очистку осуществляли методом RP-HPIC на колонке С4 Видак, представляющим собой препаративный метод анализа установления аминокислотной последовательности. Поскольку выход активности со стадии С4 обычной составляет лишь несколько процентов, такая потеря может быть приписана применяемым растворителям. В других экспериментах 1-часовое экспонирование системой 0,1% ТFА/50% ацетонитрил при комнатной температуре понижает митогенную активность препарата до 98%. Как показано на рис.I-2, был получен единственный пик ВАIВ/MK стимуляторной активности, что согласуется с ответливым пиком в профиле оптической плотности. Пиковые фракции дают единственную полосу в ходе NаДоdSO4/РАСЕ и серебрянного окрашивания геля (рис.I-2В), а относительная митогенная активность каждой испытанной фракции (рис.I-2C) хорошо коррелирует с интенсивностью полос в профиле активности.
Стадия очистки альтернативная описанной выше методике HPIC с использованием ситовой хроматографии в присутствии TSK С3000 ГласПак колонки в водном растворе со знамением рН, близким к физиологическому, давала в анализе BAIB/MK основной пик активности (рис.I-3). Такой препарат имел практически такую же чистоту, что и препарат, полученный со стадии RP-HPIC, согласно данным по серебрянному окрашиванию NаДоdSО4/РАСЕ (данные не показаны), но обеспечивал значительно лучшую регенерацию активности (Таблица I-1). TSK-очищенный материал использовали общепринятым способом для биологических исследований, поскольку он имел более высокую удельную активность.
Для обоих типов очищенных препаратов (т.е. очищенного методом HPIC или с помощью молекулярных сит), профиль митогенной активности был связан с отчетливой полосой на NаДоdSО4/РАСЕ, которая не различина для обоих препаратов.
Физические и биологические характеристики фактора роста.
Очищенный фактор имел молекулярный вес порядка 28 КДа на основе данных NаДоdSО4/РАСЕ в восстановительных (рис.I-2) и невосстановительных условиях (данные не приведены). Это значение хорошо согласуется с положением элюирования на двух колонках различного размера в тех растворителях, которые, как ожидалось, сохраняют природную конформацию (TSK-С3000-SW, фиг.I-3, и супероза-12, данные не приведены). Из этих данных следует, что митоген, по-видимому, состоит из одной полипептидной цепочки с молекулярным весом 25-30 кДа.
Термическая и кислотная лабильность митогенной активности были продемонстрированы с использованием ВАIВ/MK биоанализа на митогенезис. Хотя активность не изменялась за 10 мин инкубирования при 50°С. Она понижалась на 68% через 10 мин при 60°С и не детектировалась через 3 ч при 100°С. Воздействие 0,5 М уксусной кислоты в течение 60 мин при комнатной температуре приводило в результате к понижению активности до 14% от контрольного значения. В целях сравнения следует отметить, что митогенная активность известного фактора роста, ЕСF, не уменьшается при любой из таких обработок. Кривая доза-отклик для очищенного фактора роста, изображенная на рис.I-4 иллюстрирует тот факт, что всего лишь 0,1 нМ приводит к детектируемой стимуляции синтеза ДНК. Таким образом, интервал активности сравним с интервалом для других факторов роста проанализированных к настоящему времени. В интервале концентраций 0,1-1,0 нМ наблюдается линейная зависимость, причем максимальная 600-кратная стимуляция наблюдается при концентрации 1,0 нМ. Новый фактор индуцирует более высокий уровень максимального внедрения тимидина, чем ЕСF, аFСF или bFСF в ВАIВ/MK кератиноциты (рис.I-4).
Выраженная специфичность в отношении клетки-мишени такого фактора демонстрируется сравнением его активности на большем числе типов клеток с активностью других факторов роста, которые, как известно, обладают митогенной активностью в отношении эпителиальных клеток.
Как показано в таблице I-2, новый выделенный фактор оказывает сильное митогенное действие на ВАIВ/MK, но также индуцирует демонстрируемое внедрение тимидина в ДНК других испытанных эпителиальных клеток. В отличие от этого, такой фактор не обладает детектируемым митогенным эффектом на мышинных или человеческих (данные не приведены) фибробластах или эндотелиальных клетках подкожной вены человека.
В целях сравнения следует отметить, что ни один из других известных факторов роста преимущественно не стимулирует кератиноцита. ТСFа и ЕСF обладают мощной активность в отношении фибробластов, тогда как FCF являются митогенно-активными в отношении эндотелиальных клеток и фибробластов (I-2). Из-за специфичности в отношении эпителиальных клеток и особенно, чувствительности в отношении кератиноцитов, новый митоген был условно обозначен, как фактор роста кератиноцита (KCF).
С целью установления того факта, что KCF способен не только стимулировать синтез ДНК, но и поддерживать длительный рост клеток, оценивали способность клеток BAIВ/MK расти в полностью определенной, не содержащей сыворотки среде, дополненной фактором роста. Как показано на рис.I-5, KCF служит отличным заменителем ЕСF, но не инсулина (или инсулино-подобного фактора роста I) в такой химически определенной среде. Таким образом, КСF, по-видимому, действует по главному сигнальному маршруту, конкурируя с ECF, аFСF и bFСF за пролиферацию клеток ВAIВ/MK.
Микроопределение аминокислотной последовательности обнаруживает уникальную N-терминальную аминокислотную последовательность КСF. С целью дополнительного охарактеризования фактора роста, примерно 150 пмоля С4 у очищенного материала подвергали анализу на аминокислотную последовательность. Одну последовательность детектировали при особом вникании на циклы 2-13: Х-Аsn-Asp-Met-Тhr-Рrо-Glu-Gln-Met-Ala-Thr-Аsn-УAI. Высокий финовый шум затрудняет отнесение для первой позиции, которая, поэтому, указана, как Х.
Компьютерный поиск с использованием программы FАSТР/I-24/ показал, что N-терминальная аминокислотная последовательность КСF не обладает значительной гомологией с любым из протеинов в банке данных Национального биомедицинского исследовательского фонда, что подтверждает новизну такого эпителиального фактора роста.
Обсуждение.
Исследования, описанные в настоящем Экспериментальном разделе, позволили идентифицировать человеческий фактор роста, который обладает уникальной специфичностью в отношении эпителиальных клеток. В результате использования ультрафильтрации, НSАС и RР-НРIС или TK ситовой хроматографии согласно настоящему изобретению, было выделено количество препарата, достаточное для детального охарактеризования физических и биологических свойств такой молекулы.
Единственная окрашенная серебром полоса, соответствующая молекулярному весу около 28.000 дальтонов детектируется в активных фракциях из RP-HPIC и интенсивность такой полосы пропорциональна уровню митогенной активности таких фракций. Полоса неразличима с полосой, полученной методом RP-HPIC наблюдается в активных фракциях, полученных ТSК хроматографией. Очищенный протеин стимулирует синтез ДНК в эпителиальных клетках при субнаномолярных концентрациях, но не способен индуцировать какое-либо внедрение тимидина в фибробласты или эндотелиальные клетки при сравнимых или более высоких концентрациях (вплоть до 5 нМ). Такая выраженная специфиность в отношении клетки-мишени совместно с единственной новой N-терминальной аминокислотной последовательностью определенной в очищенной молекуле, приводит к выводу о том, что KCF представляет собой новый факт роста.
В химически определенной среде очищенный фактор способен дополнять инсулиноподобный фактор роста I (потребности в инсулине клеток ВАIВ/MK и поэтому должен действовать по пути сигнальной трансдукции, конкурируя с ECF, TCFa и FСF. Кроме этого, новый фактор обладает большей мощностью, чем любой из известных митогенов эпителиальных клеток при стимуляции внедрения тимидина в клетки ВАIВ/MK. Предварительные данные свидетельствуют о том, что такой фактор также способен поддерживать пролиферацию вторичных культур человеческих кератиноцитов (данные не представлены).
В ходе очистки КСF манипуляции с ним и его хранение являются проблематичными операциями. Помимо присущей ему лабильности в отношении действия тепла и кислот, этот фактор неустойчив в отношении лиофилизации или диализа. После операции НSAС происходит полная потеря активности за 24 ч несмотря на использование протеинов-носителей, гепарина, ингибиторов протеазы, силиконизированных трубок или хранения при 4 или -20°С. Лишь концентрирование образца на этой стадии может сохранить его активность.
Кроме этого, с целью переноса высушенного, очищенного фактора необходимо использовать сильную кислоту или детергент, соответствующие адсорбционной тенденции или нерастворимости. Таким образом, для сохранения активности очищенный фактор выдерживают в растворе при высокой концентрации при -70°С, где он остается стабильным в течение нескольких месяцев.
Способность КCF связывать гепарин может представлять собой функламентальное свойство такого фактора, которое переносится на его функцию in vivo. Факторы роста с гепаринсвязующими свойствами включают aFCF (I-20 - I-22), bFСF (I-19, I-22), фактор, стимулирующий гранулоцит/макрофаговую колонию и интерлейкин 3 (I-25). Каждый из таких факторов производится стромальными клетками (I-25 - I-27). Такие факторы, по-видимому, отлагаются в межклеточной матрице, или на протеогликанах, покрывающих поверхность стромальной клетки (I-25, I-28). Было постулировано, что их хранение, выделение и контактирование со специфическими клетками-мишенями регулируется такими взаимодействиями (I-25, I-28). Хотя эффекторы пролиферации эпительной клетки мезенхимального происхождения также были описаны (I-29 - I-31) их идентичность не установлена. Гепаринсвязующие свойства, выделение стромальными клетками человеческого эмбрионного фибробласта и тропизм эпителиальных клеток наделяет KCF всеми свойствами, ожидаемыми для такого паракринового медиатора роста нормальных эпителиальных клеток.
Неполная аминокислотная последовательность, установленная для такого нового фактора роста, дает возможность молекулярного клонирования его кодирующей последовательности и определения его структурной взаимосвязи с известным семейством факторов роста, как это описано ниже в Экспериментальном разделе II.
Литературные ссылки по Экспертиментальному разделу I.
References for experimental section I
I-1. James, R. and Bradshaw, R.A. (1984) Ann.
Rev. Biochem. 53, 259-292.
I-2. Doolittle, R.F., Hunkapiller, M.W.,
Hood, L.E., Devare, S.G., Robbins, K.C.,
Aaronson, S.A. and Antoniades, H.M.
(1983) Science 221, 275-277.
I-3. Waterfield, M.D., Scrace, G.J., Whittle,
N., Strooband, P., Johnson, A.,
Wasteton, A., Westermark, В., Heldin,
C.-H., Huang, J.S. and Deuel, T.F.
(1983.) Nature 304, 35-39.
I-4. Hunter, Т. and Cooper, J.A. (1985) Ann.
Rev. Biochem. 54, 897-930.
I-5. Wright, N. and Allison, M. (1984) The
Biology of Epithelial Cell Populations (Oxford
University Press, New York) Vol. 1, pp.
3-5.
I-6. Weissman, B.E. and Aaronson, S.A. (1983)
Cell 32, 599-606.
I-7. Falco, J.P., Taylor, W.G., DiFiore,
P.P., Weissnan, B.E., and Aaronson, S.A.
(1988) Oncogene 2, 573-578.
I-8. Aaronson, S.A. and Todaro, G.J. (1968)
Virology 36, 254-261.
I-9. Laemmli, U.K. (1970) Nature 227, 680-685.
I-10. Merril, C.R., Goldman, D., Sedman, S.A.
and Ebert, M.H. (1981) Science 211,
1437-1438.
I-11. Jainchill, J.L., Aaronson, S.A. and
Todaro, G.J. (1969) J. Virol. 4, 549-553.
I-12. Caputo, J.L., Hay, R.J. and Williams,
C.D. (979) In Vitro 15, 222-223.
I-13. Staaipfer, M.R. and Bartley, J.C. (1985)
Proc. Natl. Acad. Sci. USA 82, 2394-2398.
I-14. Sharefkin, J.B., Fairchild, K.D., Albus,
R.A., Cruess, D.F. and Rich, N.M. (1986)
J. Surgical Res. 41, 463-472.
I-15. Cohen, S. (1962) J. Biol. Chem. 237, 1555-1562.
I-16. DeLarco, J.E. and Todaro, G.J. (1978)
Proc. Nail. Acad. Sci. USA 75, 4001-4005.
I-17. Raines, E.W. and Ross, R. (1982) J.Biol.
Chem. 257, 5154-5160.
I-18. Shing, Y., Folkman, J., Sullivan, R.,
Butterfield, C., Murray, J. and
Klagsburn, M. (1984) Science 223, 1296-1299.
I-19. Gospodarowics, D., Cheng, J., Lui, G.-
M., Baird, A. and Bohlen, P. (1984)
Proc. Natl. Acad. Sci. USA 81, 6963-6967.
I-20. Maciag, Т., Hehlman, Т., Friesel, R. and
Schreiber, A.B. (1984) Science 225,
932-935.
I-21. Conn, G. and Hatcher, V.B. (1984)
Biochem. Biophys. Res. Comm. 124, 262-268.
I-22. Lobb, R.R. and Fett, J.W. (1984)
Biochemistry 23, 6295-6299.
I-23. Bradford, H. (1976) Anal. Biochem. 72, 248-254.
Экспериментальный раздел II
Последовательность сДНК нового специфического фактора роста эпителиальных клеток, представляющего новый член семейства FСF.
Исследование результатов, приведенные в предыдущем Экспериментальном разделе I, позволили идентифицировать и очистить новый гепаринсвязующий фактор роста, обозначенный как фактор роста кератиноцита (КCF), который проявляет особую активность в отношении кератиноцитов и, по-видимому, является специфичным в отношении эпителиальных клеток. В настоящем втором Экспериментальном разделе описывается выделение и охарактетеризование сДНК клонов, кодирующих КСF, с использованием синтетических олигонуклеотидов, основываясь на экспериментально установленной NH2-терминальной аминокислотной последовательности, в качестве гибридизационных зондов. В результате анализа аминокислотного состава нуклеотида идентифицирован открытый каркас считывания размером в 582 bр, который может кодировать полипептид из 194 аминокислот, на 41-33% идентичный гепаринсвязующим факторам роста кислотного и основного фибробласта (FCF), и родственные продукты hst и int-2 онкогенов. KCF матрицы генной РНК экспрессирует в нормальных фибробластах как эмбрионного, так и взрослого происхождения, но не в эпителиальных, эндотелиальных или глиальных клетках. Таким образом, KCF обычно экспрессируется механизмом, это указывает на его роль в регуляции проилиферации эпителиальных клеток.
Материалы и методы исследования.
Выделение клонов сДНК. Очистка и определение аминокислотной последовательности N-окончания KCF описано ранее (см. Экспериментальный раздел I и ссылку II-3). Сливы (50 пмолей) деоксиолигонуклеотидов, описанных в разделе "Результаты" метили по окончанию 5’ с использованием 83 пмолей -32Р-АТР (3000 Ci) ммоль, Амершам и 10 единиц Т4 полинуклеотидной киназы. Рекомбинантный фаг, несущий клоны сДНК, представлял собой реплику, высеенную на нитроцеллюлозные фильтры и гибридизированную с32P-меченными деоксилигонуклеотидами в 20% формамида, 10% сульфата декстрана, 10 мМ Трис-НСl (рН 7.5), 8х SSС, 5xДенхардт и 50 мкг/мл ДНК денатурированной спермы лосося, в течение ночи при 42°С. Фильтры промывали в 0,5х SSС, 0,1% SДS при 50°С и экспонировали на пленке Кодак Х-0мат АR.
Определение аминокислотной последовательности ДНК.
Нуклеотидную последовательность сДНК KCF определяли методом обрыва дидеокси цепи (II-26) перекрывающихся ограничительных фрагментов субклонированных в рУС векторы (II-27).
Конструирование бактериального экспрессионного вектора для KCF сДНК.
KCF сДНК кодирующую зрелую секретированную форму полипептида помещали под контролем гибридного trk промотора в вектор экспрессии плазмиды рКК233-2 (II-31) согласно следующей методике. Конкретную длину KCF cДНК, которая содержала информацию для кодирования молекулы зрелой KCF (т.е. без его сигнального пептида) усиливали с использованием метода цепной реакции полимеразы (PCR, II-32). Фрагмент направленно вставляли между двумя сайтами в вектор, сайтом Ncol, с затупленными концами в результате SI нуклеазного переваривания и сайтом Hind III, с использованием стандартных методов рекомбинантной ДНК технологии. Окончания KCF сДНК, полученного методом PCR, были следующими;
окончание 5’ было тупым и начиналось с ATG кодона, после чего шел кодон TGC для цисостатка, номер 32, который представлял собой аминотерминальный остаток зрелой формы KCF (см. рис.II-1) и затем полная KCF кодирующая последовательность. Стоп-кодон ТДА и следующие за ними четыре основания, TTGC, также включены в 3’ окончание сДНК. Затравка, используемая в методе PCR для направления синтеза ДНК в желаемое положение на окончании 3’ сДНК включает Нind III сайт для внедрения усиленной сДНК в векторную ДНК.
Продуцирование антител к KCF и KCF-родственным пептидом.
Моноклональные антитела вырабатывали в мышах против неповрежденного очищенного протеина человеческих фибробластов с использованием 5 или более подкожных инъекций. Испытуемые образцы крови подвергали скринингу с помощью твердофазного (ELISA) анализа с использованием высокоочищенного KCF из человеческих эпителиальных клеток в качестве антигена. Гибридом готовили обычными методами и надосадочные жидкости подвергали скринингу с помощью анализа ЕLISА. с целью детекции КСF-реакционноспособных антител.
Положительные клоны серийно субклонировали обычными методами и отобранные субклоны выращивали в виде опухолей у мышей с целью продуцирования больших количеств антител. Антитела очищали от асцитных жидкостей с использованием стандартных приемов (например, с помощью гидроксиапатита или иммуносродственных смол).
Поликлональные антитела против синтетического пептида выращивали в кроликах следующими стандартными методами. Пептиды получали твердофазной технологией и соединяли с тироглобулином по реакции с глутаровым альдегидом. Производили серийные подкожные инъекции и образцы крови подвергали скринингу анализом ЕLISА, а также другими методами, включая анализа пятнения по Вестерну и биоанализа на митогенезис. IgC иммуноглобулины выделяли методом хроматографии по сродству с использованием иммобилизованного протеина С.
Поликлональные антитела выращивали в кроликах против как природно секретированного KCF из человеческих фибробластов, так и рекомбинантного КCF продуцированного в E. coli (см. следующий раздел) с использованием следующей последовательности операции:
I) Начальную инъекций и первую иммунизацию осуществляли в паховых лимфотических узлах,
II) последующие иммунизации проводили внутримышечно.
Скрининг испытуемых образцов осуществляли с использованием анализа ЕLISA и анализа на окрашивание по Вестерну, а также биоанализа на митогенезис, а IgС очищали по методике, описанной выше для антител к синтетическим пептидам.
Результаты
Выделение сДНК клонов, кодирующих новый фактор роста.
С целью поиска сДНК клонов, соответствующих KCF кодирующей последовательности, получали две группы олигонуклеотидов с длинами в 26 оснований, основываясь на девятой аминокислотной последовательности, Asn-Asp-Met-Thr-Pro-Glu-Gln-Met-Ala, как это было установлено микроустановлением аминокислотной последовательности очищенного KCF (см. Экспериментальный раздел I и ссылку II-3). Одна нуклеотидная группа содержала смесь всех из 256 возможных кодирующих последовательностей для девяти аминокислот, тогда как другой содержал инозиновые остатки в дегенерированном третьем положении кодонов для Тhr и Pro.
Такая последняя конструкция понижает число возможных кодирующих последовательностей в группе до 16. Инозин в антикодоне тРНК может образовывать водородные связи с А, С или У (II-4), а олигонуклеотиды, которые содержат деоксинозин, как было показано, может эффективно гибридизироваться с соответствующей сДНК (II-5).
сДHK библиотеку конструировали в клонирующем векторе сДHК, рСЕУ9 (II-6) с использованием полиаденилированной РНК, экстрагированной из клеточной линии М426 человеческого эмбрионного легочного фибробласта (II-7), исходного источника фактора роста. В результате скрининга библиотеки (9×10 тромбоцитов) с помощью32Р-меченных 26-mer олигонуклеотидов, идентифицировали 88 тромбоцитов, которые гибридизировали с обеими группами олигонуклеотидных зондов.
Охарактеризование и установление аминокислотной последовательности выбранных клонов сДНК. Из 10 очищенных от тромбоцитов клонов, подвергнутых анализу, один, обозначенный как клон 49, имел сДНК вставку размером 3,5 кb, тогда как оставшиеся клоны имели вставки размерами 1,8-2,1 кb.
Анализ мелких клонов позволил обнаружить несколько общих рестрикционных сайтов. Установление аминокислотной последовательности представителя мелких клонов, обозначенного как клон 32, совместно с клоном 49, продемонстрировало тот факт, что они представляют собой перекрывающиеся сДНК (рис.II-IA). В то время как клон 49 подвергали действию затравки из поли(А) отростка информации, клон 32 возникает в ходе конструирования библиотеки гибридизацией олиго (dТ) затравки с А-обогащенной последовательностью в 3’ некодирующей области KCF мРНК.
Описание последовательности кодирующей полипептид KCF.
Исследование двух таких сДНК (клоны 32 и 49) позволило установить непрерыную последовательность из 3.85 кb, содержащую полную КCF кодирующую последовательность (рис.II-IВ). АТC, по-видимому, являющий инициирующим кодоном, находится в положении 446 нуклеотида, создавая каркас открытого считывания из 582-оснований, который заканчивается на ТАA обрывающем кодоне в положении 1030. Такой каркас открытого считывания может кодировать полипептид из 194 аминокислот с расчитанным молекулярным весом 22,512 дальтонов.
Боковая последовательность AТС кодона не соответствует предполагаемой GСС(G/A)ССАТСG согласованности для оптимального инициирования эквариотными рибосомами (II-8), однако в направлении вверх от АТС кодона имеется три нуклеотида. В этом положении А представляет собой наиболее высокосохраненный нуклеотид в такой согласованности. Такому АТС кодону предшествует 85 нуклеотидов расположенных вверх от ТСА стоп-кодана в рамках того же каркаса считывания.
Аминокислотная последовательность 19, которая гомологична экспериментально определенному NН2 -окончанию очищенного KCF начинается с 32 аминокислот, расположенных вниз от предполагаемого инициирующего кодона. Предсказанная и экспериментально установленная аминокислотные последовательности полностью согласуются друг с другом в результате чего может быть проведено недвусмысленное отнесение.
С целью поиска гомологии между KCF и любым известным протеином был осуществлен компьютерный поиск в банке данных Национального Биомедицинского исследовательского фонда с использованием программы FASТР Липмана и Пирсона (II-9). В результате такого подхода была выявлена поразительная родственность между предсказанной структурой KCF и структурой кислотного и основного FСF, а также родственных hst и int-2 закодированных протеинов.
Экспрессия матриц мРНК КCF гена в человеческих клетках.
В предварительных попытках исследования экспрессии KCF мРНК в человеческих клетках зонд, охватывающий большую часть KCF кодирующей последовательности (зонд А, рис.II-IA), позволил детектировать с помощью анализа пятнением по Нордерну одну матрицу размером 2,4 кb из всей М426 РНК (рис.II-IC). Эта величина значительно меньше, чем длина композитной сДНК последовательности, 3,85 кb.
Однако при скрининге поли(А)-селекционированной М426 РНК, была установлена дополнительная матрица размером примерно 5 кb. Кроме этого, зонд на основе нетранслированной области клона 49 от окончания 3’ до клона 32 (зонд В, рис.II-IA), гибридизируется только с большей частью информации (рис.II-IC). Таким образом, видно, что KCF ген подвергается транскрипции только в тех участках, которые касаются альтернативных РНК. Два других члена семейства FСF гена, bFСF (II-29) и int 2-(II-30), также экспрессируют большое число РНК, важность которых требует дополнительного выяснения.
С целью исследования нормальной функциональной роли KCF исследовали экспрессию его матрицы в большем числе человеческих клеточных линий и тканей. Как показано на рис.II-3, доминирующая матрицы КСF размером 2,4 кb определяется в каждой из нескольких стромальных фибробластовых линий, являющихся производными эпителиальных тканей из эмбрионных, новорожденных и взрослых источников, но не из эпителиальных клеточных линий обычного происиождения. Такую матрицу также детектировали в РНК экстрагированной из нормальных взрослых почек и органов желудочно-кишечного тракта, но не из легких или мозга. Выраженная спефифичность KCF РНК экспрессии в стромальных клетках эпителиальных тканей указывает на то, что такой фактор играет нормальную роль в мезенхимальной стимуляции роста эпителиальных клеток.
В целях сравнения мРНК других факторов роста с известной активностью в отношении эпителиальных клеток также анализировали в тех же тканях, что перечислены выше. Среди проанализированных эпителиальных и стромальных клеточных линий не было обнаружено согласующейся картины экспрессии аFСF или bFСF матриц (рис.II-3). ЕСF матрица не экспрессировала каких-либо одинаковых клеточных линий и наблюдалась лишь в почках среди различных тканей. Наконец ТСFа не детектировался в какой-либо из стромальных фибробластовых линий и экспрессировался с различными уровнями в каждой из эпителиальных клеточных линий. Его также обнаруживали в низких количествах в почках среди изученных тканей С фиг.II-3).
Ингибирование KCF митогенной активности гепарином. Было обнаружено, что гепарин значительно повышает митогенную активность аFСF в отношении большого числа клеток-мишеней в культуре и стабилизирует его в отношении термической дезактивации (II-21, II-22). Несмотря на прочное присоединение к bFСF гепарин оказывает минимальное влияние на его митогенную активность (II-22). Основываясь на родственности КCF к FСF изучали влияние гепарина на митогенную активность KCF. Как показано в таблице II-1 внедрение тимидина клетками ВАIВ/MK под действием KCF 16-кратно ингибируется при включении гепарина в культурную среду. В отличии от этого, активности aFCF и bFСF в результате такой обработки повышались.
Продуцированне анти-КСF антител.
Некоторые виды антител, которые распознают KCF или КОF-подобные полипептиды были получены с использованием стандартных методов хорошо известных в области экспериментальной иммунологии и суммированных выше в разделе. Методы. Они включают: моноклональные антитела, выращенные в мышах против целого, очищенного протеина из человеческих фиброблатов; поликлональные антитела, выращенные в кроликах против синтетических пептидов с последовательностями на основе аминокислотных последовательностей, предсказанных из последовательности KCF сДНК поликлональные антитела, выращенные в кроликах против как естественно секретированного KCF человеческого фиброблаета, так и рекомбинантного КСF продуцированного в Е. соli (см. следующий раздел).
Были очищены моноклональные тела из трех различных гибридов. Все три таких антитела распознают рекомбинант, а также встречающийся в природе KCF с помощью твердофазного (ЕLISА) анализа. Ни один из них не вступает в перекрестные реакции с KCF в условиях денатурации (согласно пятнению по Вестерну) и не нейтрализует митогенную активность KCF в биоанализе ВAIВ/MK.
Поликлональные антитела получали с использованием синтетического пептида с аминокислотной последовательностью NДMTРЕОМАТNУ, соответствующей остаткам 32-44 в KCF (см. рис.II-1), плюс R (аrg) остаток вместо действительного аsр остатка, закодированного сДHК в положении 45. asp остаток возможно гликозилирован в природном KCF полипептиде и поэтому представляет собой аrg в данных по аминокислотной последовательности, полученных непосредственно для такого полипептида (см. раздел "Обсуждение"). Поликлональные антитела генерированные с помощью такого синтетического пептида распознают как встречающийся в природе, так и рекомбинантный KCF в анализах ЕLISА и пятнения по Вестерну, при уровне чувствительности, по крайней мере, 10 нг протеина. Однако такие антитела не нейтрализуют митогенную активность КCF в биоанализе ВАIВ/MK.
Поликлональная антисыворотка против целого природного KCF протеина распознает КСF в анализах ЕLISА и пятнения по Вестерну. Такие антитела также ингибируют митогенную активность KCF в биоанализе BAIB/MK.
Экспрессия КCF сДНК в Е. соli.
KCF сДНК подвергали экспрессии с целью продуцирования полипептида в E. coli путем помещения его кодирующей последовательности под контролем гибридного trk промотора (включающего элементы trp и Iac промоторов) в плазмиду рКК233-2 (II-31). Для совершения такой операции, специальную длину KCF сДНК, которая содержала информацию для кодирования зрелой молекулы KCF (т.е. без его сигнальной последовательности) усиливали с использованием метода полимеразной цепной реакции (II-32). Фрагмент направленно вставляли между двумя сайтами в вектор, а именно между сайтом Ncol, полученный затуплением окончаний в результате SI нуклеазного переваривания и сайтом Hind III, с использованием методов рекомбинантной ДНК. В выбранных рекомбинантах осуществляли установление аминокислотной последовательности на их сДНК 5’ окончаниях, что способствовало корректному выравниванию АТG инициирующего кодона с регуляторными элементами trk промотора.
Некоторые рекомбинанты испытывали на продуцирование протеина с помощью обычных микрометодов. Клоны выращивали в мидэкспоненциальной фазе (ОД595-0,5), обработанной 1 мМ изопропил β -Д-тиогалактопиранозида (I С) в течение 90 минут и клеточные экстракты переносили на SДS-полиакриламидные гели для анализа окрашиванием по Вестерну. Все испытанные рекомбинанты синтезировали протеин, который распознавался антителами против аминотерминального KCF пептида. Выбирали один рекомбинант, который демонстрировал наивысшую индукцию из IPTC с целью последующего анализа протеина.
Один литр бактерий выращивали в NZУ бульоне, содержащем 50 мкг/мл ампициллина и 12,5 мкг/мл тетрациклина до значения ОД595-0,5 и обрабатывали в течение 90 минут с помощью IPTC. Клетки собирали центрифугированием ресуспендировали в 50 мМ фосфата натрия (рН 7.3), 0,2 М NaCl и лизировали с помощью ультразвука. Клеточный дебрис удаляли центрифугированием и лизат непосредственно применяли в колонке с гепарин-Сефаразой.
Как было установлено анализом пятнения по Вестерну и определениием митогенной активности в кератиноцитах, рекомбинантный KCF алюировали в 0,5-0, 6 М NaCl. Последующая очистка НSАС материала с помощью Моно-(FPIC) колонки (Фармация) давала препарат KCF с чистотой ≥90%, о которой судили с помощью электрофоретического анализа с использованием SДS-полиакриламидных гелей и окрашивания серебром.
Рекомбинантный KCF эффективно стимулирует внедрение тимидина в ВАIВ/MK кератиноцитные клетки, но лишь незначительно активен в отношении фибробластов NIН/3Т3. Половинная от максимального значения стимуляции клеток ВАIВ/МК в стандартном кератиноцитном анализе достигалась при концентрации 2-5 нг/мл, по сравнению с концентрацией 10-15 нг/мл для KCF очищенного от клеток М426. Один литр бактериальных клеток давал примерно 50 мкг Моноочищенного рекомбинантного KCF.
Конструирование химеры, содержащей KCF и аFСF последовательности.
Проведенные выше исследования показали, что KCF обладает двумя выраженными характеристиками, которые могут быть закодированы определенными частями или областями полипептидной последовательности, что, как известно, имеет место в кодирующих последовательностях других полифункциональных полипептидов. С целью испытания такой возможности, сегмент химерной ДНК кодирующей NН2-терминальную последовательность KCF привитого на ядро С-окончания аFСF конструировали следующим образом. Сайт реетрикционного энзима San ЗAI (САТС) на 5’ окончании KCF сДНК внутри кодонов для остатков 78 и 79 (аrg и tle соответственно; см. рис.II-1) разрезали и присоединяли к гомологическому сайту в аFСF cДНК внутри кодонов для аминокислот 39 (аrg) и 40. Окончания 3’ и 5’ такой химерной ДНК соединяли с векторной ДНK плазмиды рКК233-2 тем же способом, что использовали для внедрения KCF сДНК кодирующего секретированную форму полипептида (см. раздел "Методы" приведенный выше).
В том случае когда рекомбинантные клетки E. coli конструировали с использованием вектора несущего химеру, а экспрессионные испытания проводили в соответствии с описанным выше для зрелого KCF, получали новый продукт со свойствами, как KCF так и аFСF. Пептид обогащали с помощью гепарин-Сефарозной хроматографии и было обнаружено, что предпочтение в качестве клетки-мишени отдается кератиноцитам, подобно KCF, при минимальной активности в отношении фибробластов (NIН/3Т3). Однако митогенная активность такого химерного полипептида теряет восприимчивость к ингибированию гепарином, т.е. характеристику, которая присуща аFCF, а не KCF. Фактически, митогенная активность кератиноцитов в действительности усиливается гепарином, как это имеет место в случае аFСF. Таким образом, согласно практическому воплощению настоящего изобретения пептидные области, ответственные за специфичность в отношении клетки-мишени и чувствительность в отношении гепарина, обладают выраженной индивидуальностью и могут быть легко выделены из KCF.
Обсуждение.
Эксперименты, описанные в этом разделе, иллюстрируют практическую реализацию нескольких главных воплощений настоящего изобретения. Они включают выделение сДНК кодирующих КСF, экспрессирование таких сДНК в рекомбинантных клетках, продуцирование различных антител реакционноспособных в отношении KCF и конструирование и экспрессию химерной сДНК, кодирующей новый фактор роста с аминокислотными последовательностями и родственными функциональностями присущими как KCF, так и аFСF. Следующие ниже пункты, относящиеся к таким воплощениям, могут улучшить понимание настоящего изобретения.
Последовательность, предсказанная на основании KCF сДHK, согласуется с аминокислотной последовательностью, определенной из очищенной формы KCF секретированной человеческими фибробластами. Кроме этого, такая последовательность предполагает потенциальное объяснение положений, где определенное отнесение аминокислот не может быть сделано прямым установлением аминокислотной последовательности. Остатки 32 и 46, предсказанные из сДHK последовательности, являются цистеинами, а гидролизованные производные немодифицированных цистеиновых остатков не детектируются с помощью деградации по Эдману. Предсказанная аминокислотная последовательность KCF содержит также один потенциальный N-связанный сайт гликозиляции (Аsn-Х-Ser/Тhr) из остатков 45-47. Если Аsn 45 гликозилирован, то он не может быть детектирован используемыми методами установления аминокислотной последовательности. Фактически, KCF мигрирует в виде широкой полосы на NаДоdSО4/PACE при более высоком молекулярном весе, чем предсказано для очищенного протеина. Это может быть отнесено за счет гликозилирования.
FCF представляют собой гепаринсвязующие митогены с широким спектром специфичности в отношении клетки-мишени (II-10). Нst ген были идентифицирован как трансформирующий ген из желудочной опухоли человека (II-11), по соседству с обычной желудочной тканью (II-12) и на саркомы Капоши (II-13), с помощью стандартного NIH/ТЗТ трансфекционного анализа. Продукт int-2 гена нормально экспрессируется в ходе эмбриогеноза мышей (II-14) и c отклонениями от нормы после провирусной интеграции вируса опухоли молочной железы мышей (II-15).
KCF представляет собой шестой член семейства факторов роста фибробластов подлежащего идентификации (II-28). Хотя название FСF-6 по-видимому не подходит из-за того, что KCF не обладает активностью в отношении фибробластов, такая номенклатура может также использоваться для такого фактора роста с целью отражения его структурной взаимосвязи с семейством FСF. Поскольку все ранее охарактеризованные факторы роста либо исключают эпителиальные клетки в качестве мишеней или включают их в ряд чувствительных клеток-мишеней, высокоспецифичная природа KCF митогенной активности в отношении эпителиальных клеток и, в особенности чувствительность к кератиноцитам, делают этот фактор уникальным.
Согласно проведенным исследованиям экспрессия матрицы KCF, по-видимому, является специфичной в отношении стромальных клеток эпителиальных тканей, что предполагает ее участие в пролиферации нормальных эпителиальных клеток. Доступность KCF сДНК клона позволяет определять те случаи, когда анормальная экспрессия такого фактора роста может принимать участие в клинических состояниях, характеризующихся дисплазией и/или неоплазией эпителиальных клеток. Кроме того, возможность получения больших количеств такого нового фактора роста с помощью рекомбинантной технологии позволяет осуществлять испытание его клинической применимости в тех случаях, когда особую важность имеет специфический рост эпитеальных клеток.
Трассировка KCF последовательности пятью другими протеинами семейства FСF обнаруживает два основных участка гомологии, охватываемых аминокислотами 65-156 и 162-189 в предсказанной последовательности KCF, которые разделены короткими, негомологическими сериями аминокислот различной длины в различных членах семейства (риc.II-2). В случае int-2 длина такой последовательности составляет 17 остатков, тогда как в случае hst, два таких гомологических участка являются соприкасающимися. В KCF такая последовательность состоит из пяти аминокислот. В регулируемых областях KCF аминокислотная последовательность была на 44% идентична int-2 (мышиная), на 39% идентична bFСF (человеческий), на 30% идентична aFCF (человеческая) и на 33% идентична hst (человеческая). В этой же области все шесть протеинов были идентичны 19% остатков, а при осуществлении консервативных замещений они проявляли 28% гомологию.
Как показано на рис.II-2 аминоокончания таких родственных протеинов были негомологичными и имели разную длину. Первичный KCF и hst трансляционные продукты содержат гидрофобные N-терминальные области, которые, по-видимому, служат сигнальными последовательностями (II-16). Такое предположение подтверждается также тем фактом, что такая N-терминальная область отсутствует у молекулы зрелого KCF (рис.II-18). В отличии от этого FСF синтезируются без сигнальных пептидов (II-10). Int-2-протеин содержит нетипично короткую область N-терминальных гидрофобных остатков II-17), однако она неизвестна, если протеин секретирован. Более того, int-2-протеин содержит длинное C-терминальное удлинение, сравнимое с другими членами семейства.
Очищенный KCF содержит пять цистеиновых остатков, два из которых сохраняются во всем семействе FCF-родственных протеинов (рис.II-2). Следует также отметить пять пар основных остатков последовательности KCF. Та же картина наблюдается у других членов семейства FCF и это обстоятельство следует учитывать в их взаимодействии с гепарином (II-18). Двухосновные сайты также являются обычными мишенями в отношении протеолитического воздействия и такое воздействие может быть причиной микрогетерогенности, наблюдаемой в одинаковых KCF препаратах (неопубликованные данные).
KCF сДНК последовательность обогащена AT по ее длине, но особенно это проявляется в 3’ нетранслированной области, где содержание AT составляет 70% по сравнению с 60% в предполагаемой кодирующей последовательности и 63% в 5’ нетранслированной области 3’ нетранслированная область содержит большое число ATТТА последовательностей, которые, как предполагается, принимают участие в селективной деградации промежуточно экспрессированных, неустойчивых РНК (II-19). Классический сигнал ААТААА полиаденилирования отсутствует, однако детектируются две вариантные последовательности ААТТАА и ААТАСА (II-20), содержащие 24 и 19 нуклеотидов, соответственно вверх от поли(А) последовательности на 3’ окончании сДНК.
Было предположено, что влияние гепарина на кислотный FCF связан либо со стабилизацией активной конформации фактора роста или с образованием тройного комплекса с кислотным FCF и его рецептором (II-21, II-22). Если это так, то гепарин может стабилизировать конформацию KCF, которая не столь активна, как свободная молекула или образовывать прочный комплекс, способный эффективно взаимодействовать с его рецептором.
Хотя способность связывать гепарин отражает структурное подобие KCF и FСF, различие в специфичностях в отношении клетки-мишени у этих родственных митогенов является значительным. FCF индуцируют деление большинства нетерминально дифференцированных клеток как эмбрионного мезодермального, так и нейтроэктодермального происхождения. Помимо фибробластов и мышечных эндотелиальных тканей мишени мезобермального происхождения в культуре включают миобласты, хонцроциты и остеобласты (II-23). FCF также проявляют митогенность в отношении глиальных астроцитов и нейтробластов (II-24).
Продукт онкогена, выделенный из саркомы Калоши, который идентичен hst, также стимулирует пролиферацию NIH/3T3 и капиллярных эндотелиальных клеток (II-25). К настоящему времени, KCF индуцированный мутогениз наблюдался лишь в эпителиальных клетках и было также продемонстрировано отсутствие какой-либо детектируемой активности в фибробластах или эндотелиальных клетках (см. Экспериментальный раздел I, II-3). Поэтому, представляется вероятным, что KCF действует через другой клеточный поверхностный рецептор, чем FCF.
Не наблюдается значительной N-терминальной гомологии между KCF и другими FCF родственными протеинами. Так, например, была создана конструкция из химерных молекул между KCF и прототипом FCF с целью установления достаточности KCF- N-терминальной области для его уникальной специфичности в отношении клетки-мишени. Результаты исследования такой рекомбинантной полипептидной последовательности показали, что N-терминальная область KCF существенно кодирует клеточное предпочтение в пользу кератиноцитов, тогда как восприимчивость КСF к гепарину закодирована где-то в области ядра С-окончания, которое заменено последовательностями аFСF. Такой новый KCF-подобный фактор роста может иметь преимущества при клиническом применении, где желательно применение эпителиально-специфичного фактора роста в присутствии гепарина, традиционно применяемого антикоагулянта. Дополнительное изучение химер также должно обеспечить информацию о природе конкретных областей ядра С-окончания, которые обеспечивают различные эффекты гепарина на их биологическую активность.
References for experimental II
II-1. Janes, R. and Bradshaw, R.A. (1984)
Ann. Rev. Biochem. 53, 259-292.
II-2. Sporn, M.В. and Todaro, G.J. (1980) N.
Eng. J. Med. 303, 878-880.
II-3. Rubin, J.S., Osada, H., Finch, P.W.,
Taylor, W. G., Rudikoff, S. and
Aaronson, S.A. (1989) Proc. Netl. Acad. Sci.
USA (in press).
II-4. Crick, F.H.C. (1966) J. Mol. Biol. 19,
548-555.
II-5. Ohtsuka, E., Matsuki, S., Ikehara, M.,
Takashi, Y. and Matsubara, K. (1985) J.
Biol. Chem. 260, 2605-2608.
II-6. Miki, Т., Matsui, Т., Heidaran, M. and
Aaronson, S.A. (1989) Proc. Natl. Acad. Sci.
(in press).
II-7. Aaronson, S.A. and Todaro, G.J. (1968)
Virology 36, 254-261.
II-8. Kozak, M. (1987) Nucl. Acids Res. 13,
8125-8148.
II-9. Lipman, D.J. and Pearson, R.W. (1985)
Science 227, 1435-1441.
II-10. Thomas, К. (1987) FASEB J. 1, 434-440.
II-11. Taira, M., Yoshida, Т., Miyagawa, К.,
Sakamoto, H., Terada, M. and Sugimara,
T. (1987) Proc. Natl. Acad. Sci. USA 84,
2980-2984.
II-12. Yoshida, Т., Miyagava, К., Odagiri, H.,
Sakamoto, H., Little, P.F.R., Terada,
M. and Sugimara, T. (1987) Proc. Natl. Acad.
Sci. USA 84, 7305-7309.
II-13. Delli-Bovi, P. and Basillico, С. (1987)
Proc. Natl. Acad. Sci. USA 84, 5660-5664.
II-14. Jakobovits, A., Shackleford, -. М.,
Varmus, Н. Е. and Martin, G. R. (1986)
Proc. Natl. Acad. Sci. USA 83, 7806-7810.
II-15. Peters, G., Brookes, S. and Dickson, S.
(1983) Cell 33, 364-377.
II-16. von Heijne, G. (1986) Nucl. Acids Res. 14,
4683-4690.
II-17. Moore, R. Casey, G., Brookes, S., Dixon,
M., Peters, G. and Dickson, C. (1986)
EMBO J. 5, 919-924.
II-18. Schwarzbauer, J.E., Tamkum, J.M.,
Lemischka, I.R. and Hynes, R.O. (1983)
Cell 35, 421-431.
II-19. Shaw, G. and Kamen, R. (1986) Cell 46,
659-667.
II-20. Birnsteil, M.L., Busslinger, M. and
Strub, K. (1985) Cell 41, 349-359.
II-21. Schrieber, А.В., Kenny, J., Kowalski,
W., Friesel, J., Mehlman, T. and Maciag,
T. (1985) Proc Natl. Acad. Sci. USA 82,
6138-6142.
II-22. Gospodarowizc, O. and Cheng, J. (1986)
J. Cell Physiol. 128, 475-485.
II-23. Thomas, К.A. and Giminez-Gallego, G.
(1986) Trends Biochem. Soc. 11, 81-84.
II-24. Gensburger С., Labourdette, J. and
Sensembrenner, M. (1987) FEBS Lett. 217, 1-5.
II-25. Delli-Bovi, P, Curatola, АН., Kern, F.
G., Greco, A., Ittman, M. and Basilico,
C. (1987) Cell 50, 729-737.
II-26. Sanger, F., Nicklen, S. and Coulson, A.
R. (1977) Proc. Natl. Acad. Sci. USA 74, 5463
5467.
II-27. Yanisch-Perron, C., Vieira, J. and
Messing, J. (1985) Gene 33, 103-119.
II-28. Zhan, X., Bates, В., Нu, X. and
Goldfarb, M. (1988) Mol. Cell. Biol. 8,
3487-3495
II-29. Abrahams J. A., Mergia, A., Whang, J.L.,
Tumolo, A., Friedman, J., Hjerrild, K.
A., Gospodarowicz, D. and Fiddes, J.C.
(1986) Science 233, 545-548.
II-30. Mansour, S.L. and Martin, G.R. (1988)
EMBO J. 1, 2035-2041.
II-31. Amman, E. and Brosius, J. (1985) Gene 40,
183.
II-32. Sakai, R.К., Scharf, S., Faloona, F.,
Mullis, K.B., Norn, G.T.. Eriich, H.A.
and Arnheim, N. (1985) Science 230, 1350- 1354.
II-33. Bectman, P.M., Betsholtz, С. Heldin, C-
H., Westermark, B. DiMarco, E., DiFiore,
P.P., Robbins, K.C. and Aaronson, S.A.
(1988) Science 241, 1346-1349.

Регенерации вычисляли, полагая что вся митогенная активность в исходном материале была обусловлена изолированным фактором.
Сравнение максимального внедрения тимидина, стимулированного KGF и другими факторами роста, в целом ряде линий клеток, выраженное в виде кратности увеличения стимуляции по сравнению с фоновым значением.
Эти данные представляют собой итог четырех различных экспериментов.
Рисунок I-1. Хроматография гепарин-сефарозного сродства кондиционированной среды из М426 человеческих эмбриональных фибробластов. Приблизительно 150 мл ультрафильтрационного ретентата, извлеченного из литров М426 кондиционированной среды наносили в течение 1 часа на гепарин-сефарозную колонку (объем слоя 6 мл). После промывки колонки 150 мл уравновешивающего буффера, 20 мМ Тris-HCl, рН 7,5/0,3 М NаСl, удержанный протеин (<5% от общего протеина в ретентате) вымывали элюентом с модифицированным линейным градиентом увеличения концентрации NаСl. Объем пробы был 3,8 мл, а скорость потока при градиентном элюировании составляла 108 мл/ч. Для проведения анализа на внедрение3H-тимидина в BALB/MK клетки, который описан в Методах, два мкл указанных фракций переносили в ячейки для микротитрования, вмещающие конечный объем 0,2 мл.
Рисунок I-2. (А) Обратимо-фазовая C2HPLC митогенная активность ВАLВ/МК. Активные фракции, элюированные 0-6М NaCL из гепарин-сефарозы, обрабатывали центриконом - 10 и тотчас же наносили на С4Уydаc колонку (4,6×250 мм), которая была уравновешена в 0,1% трифторуксусной кислоте 20% ацетонитриле (ACN). После промывки колонки 4 мл уравновешивающего буфера, пробу вымывали элюентом с модифицированным линейным градиентом увеличения % АСN. Объем пробы был 0,2 мл, а скорость потока составляла 0,5 мл/мин. Для проведения анализа внедрения3H-тимидина в клетки BALB/MK, аликвоты разбавляли сразу 10-кратно 50 мкг/мл бычьим сывороточным альбумином/ 20 мМ Tris-HCl, рН 7,5, и проводили анализ при окончательном 200-кратном разбавлении. (В) NаДоdSО4/РАGЕ анализ фракций, отобранных после C4 хроматографии, приведен на фотоснимке А. Половицу каждой фракции высушивали, повторно растворяли в NаДоdSО4 (2-меркапто-этаноле, денатурировали нагреванием и проводили электрофорез в 14%-ном полиакрилоамидном геле, который впоследствии окрашивали серебром. Расположение каждого молекулярного веса отмечено стрелкой (масса в кДа). (С) Синтез ДHK в клетках ВАLВ/MK под действием фракций проанализирован на фотоснимке. В. Активность выражена в виде кратности увеличения стимуляции по сравнению с фоновым значением, которое составляло 100 срm.
Рисунок I-3. HPIC (TSK 3000 SW) хроматография BALВ/МК митогенной активности на молекулярных ситах. Приблизительно 50 мкл центрикон-обработанного, 0,6 М NaCl слива из НSАС наносили на LKВ Glas Рас TSK G-3000Sw колонку (8×300 мм), предварительно уравновешенную в 20 мМ Tris HCl, рН 6,8/0,5 М NaCl, и элюировали в виде фракций по 0,2 мл со скоростью потока 0,4 мл/мин. Для проведения анализа на внедрение3H-тимидина в ВАLВ/MK клетки, аликвоты объемом 2 мкл переносили в ячейки для микротитрования. Нахождение молекулярных весов при элюировании (масс в кДа) указаны стрелками.
Рисунок I-4. Сравнение BALB/MK ДНК синтеза под воздействием ТSK-очищенного митогена и других факторов роста. Внедрение3Н-тимидина в ДНК, нерастворимую в трихлоруксусной кислоте, выраженное как кратность увеличения стимуляции по сравнению с фоновым значением, было измерено как зависимость от концентрации указанных факторов роста. Значения фона в отсутствие образцов составляли 150 срm. Результаты представляют средние значения двух независимых экспериментов. Отклонения от средних величин в каждом эксперименте не превышали 10%. ТSК-очищенный митоген, •------•; EGF, Δ------Δ, аFGF, □------□, вFGF, о--------o.
Рисунок I-5. Сравнительный рост ВАLВ/MK клеток в химически определенной среде под воздействием различных сочетаний факторов роста. Культуры высевали с плотностью 2,5×104 клеток на одну чашку Петри (диаметр 35 мм), предварительно покрытую поли-Д-лизином/фибронектином в 1:1 смеси минимальной основной среды Eagle и среды Ham F12, дополненной трансферрином, Nа2SeО3, этаноламином и факторами роста, указанными ниже. Через 10 дней чашки фиксировали и окрашивали Giemsa.
Расшифровка:
а) не содержит фактора роста,
в) только ЕGF,
с) только инсулин, d) только KGF, е) EGF + инсулин. Конечные концентрации факторов роста были следующие: EGS, 20 нг/мд, инсулин, 10 мг/мл и КGF, 40 нг/мд.
Клетки высевали в чашках для микротитрования, культивировали до слияния в среде, содержащей сыворотку, а затем, перед добавлением образца, помещали на 24-72 часа в среду, не содержащую сыворотку. Анализы на митогенез проводили по методикам, описанным выше (смотреть экспериментальную часть, ранее описанные в II-3). Если указано, что гепарин был введен в питательную среду, то его конечная концентрация составляла 20 мкг/мл. Концентрация всех факторов роста составляла 50 нг/мл. Результаты представляют кратность увеличения стимуляции внедрения3H-тимицина в указанную испытуемую клетку в присутствии (+) или отсутствии (-) гепарина. Каждое значение представляет собой средний результат из двух независимых экспериментов, в которых каждая точка представляет, в свою очередь, среднее значение двойных анализов.
Рисунок II-1. Нуклеотидная последовательность и дедуцированная аминокислотная последовательность KGF сДНК, и идентификация записей KGF генов. (А) Схематическое представление клонов человеческого KGF сДНК. Перекрывающиеся рСЕУ9 клоны 32 и 49, используемые в определении последовательности, приведены выше в виде диаграммы полной структуры, в которой нетранслированные области изображены линией, а кодирующая последовательность изображена прямоугольником. Штрихованная область обозначает последовательности сигнального пептида, а незаштрихованная область - завершенный протеин. Указаны местоположения выбранных ограничений. (B)KGF сДHК нуклеотид и предсказанные аминокислотные последовательности. Нуклеотиды пронумерованы справа; аминокислоты пронумерованы с начала до конца. Подчеркнута пептидная последовательность с N-окончанием. Гидрофобная область с N-окончанием выделена курсивом. Участок возможного аспарагин связанного гликозилирования отмечен линией, проведенной сверху. Сигналы различного полиаденилирования, ААТТАА и AАТАСА, находящиеся у 3’ окончания, заключены в прямоугольник. (С) Идентификация КGF мРHK анализом Northerm blot. Ланы а и с, поли(А)-селекционированная М426 РНК; лан d, общеклеточная М426 РНК. Фильтры гибридизировали с32Р-меченным 695 (Gp BamH1) Всl I фрагментом из клона 32 (зонд А, рис.II-IА), ланы а и в, или с 541 вр Ара I/Есо RI фрагментом из клона 49 (зонд В, рис.II-IA), ланы с и d.
Рисунок II-2. Топологическое сравнение FGF семейства родственных молекул. Темными прямоугольниками обозначены две области протеина, которые разделяют высокую гомологию. Заштрихованные прямоугольники показывают предполагаемые последовательности сигнальных пептидов. Приведены местонахождения двух сохранившихся остатков цистеина (С).
Рисунок II-3. Northern blot анализ KGF мРНК в линиях нормальных человеческих клеток и тканях, и сравнение с мРHK экспрессии других факторов роста с известной активностью на эпителиальных клетках. Все клеточные РН-кислоты выделяли градиентным центрифугированием с использованием трифторацетата цезия. 10 мкг РНК денатурировали и проводили электрофорез в 1%-ных формальдегидных гелях. После мягкой щелочной денатурации (50 мМ NаОН в течение 30 минут) РНК переносили на нитроцеллюлозные фильтры, используя в качестве конвектанта I M ацетат аммония. Фильтры гибридизировали в32Р-меченный сДНК, зонд, содержащий 647 вр Еcо RI фрагмент у 5’ окончания КGF-кодирующей последовательности (А) или в аналогичные зонды из других ДH-кислот фактора роста. Были использованы следующие виды клеток человека: раковые клетки сквамозного эпителия (А253, А388 и A431); эпителиальные клетки молочной железы (S6 и R1); кератиноциты, законсервированные Аd12-Sv40; первичные человеческие кератиноциты; фибробласты новорожденной крайней плоти (AG 1523); фибробласты развитой кожи (50IT); и фибробласты эмбрионального легкого (WI-38 и М426).
Реферат
Изобретение относится к биотехнологии, конкретно к факторам роста эпителиальных клеток, и может быть использовано для получения нового фактора роста кератиноцитов (KGF). Белок KGF получают путем культивирования рекомбинантной клетки-хозяина, трансформированной вектором, содержащим молекулу ДНК, которая кодирует аминокислотную последовательность белка KGF. Полученный белок KGF в составе фармацевтической композиции используют для стимуляции пролиферации эпителиальных клеток. Изобретение позволяет получить белок KGF, обладающий специфической митогенной активностью 3,4·104единиц на мг белка в отношении кератиноцитных клеток. 7 н. и 45 з.п. ф-лы, 3 табл., 14 ил.

Комментарии