Средства и способы получения высокоаффинных антител - RU2585153C2
Код документа: RU2585153C2
Чертежи

Описание
Изобретение относится к области клеточной биологии. Более конкретно, изобретение относится к области получения антител.
Ex vivo B-клеточные культуры являются важными средствами в биологических и медицинских применениях настоящего времени. Одно из важных применений представляет собой культивирование антитело-продуцирующих клеток с целью получения антител, предпочтительно моноклональных антител. Моноклональные антитела (mAb) представляют собой множество идентичных копий одной молекулы антитела. Среди полезных свойств mAb - их специфичность к одному определенному эпитопу антигена. Эта специфичность дает определенные клинические преимущества для mAb по отношению к стандартным терапиям, при этом с предоставлением пациентам эффективного, хорошо переносимого варианта терапии, имеющей, как правило, слабые побочные эффекты. Кроме того, mAb применяются для биологических и медицинских исследований.
Зрелые B-клетки могут культивироваться in vitro при условиях, которые имитируют некоторые ключевые аспекты реакции зародышевого центра (GC); то есть, активация B-клеток с помощью лиганда CD40 (L) и присутствие цитокинов типа интерлейкина (IL)-4, IL-10 или IL-21. В то время как B-клетки, культивированные с помощью CD40L, IL-2 и IL-4, продуцируют очень малое количество Ig, добавление IL-21 приводит к дифференцировке в плазмациты, сопровождающейся высоким уровнем секреции Ig. Хотя было доказано, что эта система in vitro применялась для изучения некоторых аспектов дифференцировки B-клеток, но как наивные IgD+ B-клетки, так и переключенные IgD- B-клетки памяти фактически дифференцируются в окончательно дифференцированные плазмациты, что сопровождается арестом клеточного цикла, предотвращающего генерирование долгоживущих антигенспецифичных BCR-положительных клеточных линий.
Последние полученные результаты обеспечили понимание того, как многочисленные транскрипционные факторы, включающие B-лимфоцит-индуцированный белок созревания 1 (BLIMP1) и белок B-клеточной лимфомы (BCL)6, контролируют развитие GC B-клеток в окончательно дифференцированные продуцирующие антитела плазмациты.
Было показано, что транскрипционный репрессор BCL6 предотвращает дифференцировку плазмацитов. BCL6 высоко экспрессируется в B-клетках GC, облегчая размножение B-клеток путем отрицательной регуляции p53, и предотвращает преждевременную дифференцировку клеток GC в плазмациты с помощью отрицательной регуляции BLIMP 1.
Улучшенный способ получения продуцирующей антитела плазмабластоподобной B-клетки недавно был описан в WO 2007/067046, который включен в настоящее описание с помощью ссылки. Согласно данному способу количество BCL6 и члена семейства Bcl-2, предпочтительно, Bcl-xL, модулируются в B-клетке, предпочтительно, в B-клетке памяти для получения продуцирующей антитела плазмабластоподобной B-клетки. В WO 2007/067046 оказывается воздействие прямое или косвенное на количество продуктов экспрессии BCL6 и/или Bcl-xL. Предпочтительно, повышены количества обоих продуктов экспрессии, BCL6 и Bcl-xL, в указанной антитело-продуцирующей клетке, так как оба продукта экспрессии вовлечены в стабильность антитело-продуцирующей B-клетки. Указанный Bcl-xL является членом антиапоптотического семейства Bcl-2. Процессы, которые контролируются семейством Bcl-2, которое включает как про-, так и антиапоптотические белки, относятся к митохондриальному пути апоптоза. Данный путь запускается, когда молекулы, изолированные между внешней и внутренней митохондриальной мембраной, высвобождаются в цитозоль благодаря проницаемости внешней митохондриальной мембраны. Члены про-апоптотического семейства могут подразделяться на два класса. Эффекторные молекулы Bax и Bak, которые содержат так называемые Bcl-2-гомологичные домены 3 (BH3), вовлечены в способность проницаемости внешней митохондриальной мембраны путем образования протеолипидных пор; про-апоптотические белки, содержащие только ВН3 (Bad, Bik, Bim, Bid, Hrk, Bmf, bNIP3, Puma и Noxa), функционируют при различных клеточных стрессах с помощью белок-белковых взаимодействий с другими (антиапоптотическими) членами семейства Bcl-2.
Анти-апоптотические члены семейства Bcl-2: Bcl-2, Bcl-xL, Bcl-w, Al и Mcl-1, как правило, интегрированы с внешней митохондиральной мембраной. Они непосредственно связываются и ингибируют про-апоптотические белки Bcl-2 для защиты целостности митохондриальной мембраны.
В таком способе дополнительно предпочтительно, чтобы указанную антитело-продуцирующую плазмабластоподобную B-клетку инкубировали с IL-21 и CD40L. B-клетку, такую как антитело-продуцирующая плазмабластоподобная B-клетка, предпочтительно культивируют в присутствии CD40L, так как он содействует репликации большинства B-клеток. Кроме того, предпочтительно, чтобы STAT3 активировался в указанной антитело-продуцирующей B-клетке. Активации STAT3 можно достичь разнообразными путями. Предпочтительно, STAT3 активируется с помощью предоставления антитело-продуцирующей клетки с помощью цитокина. Цитокины, будучи природно вовлеченными в B-клеточную пролиферацию, очень эффективны в регуляции STAT-белков. Очень эффективные активаторы STAT3 - это IL-2, IL-10, IL-21 и IL-6, а также IL-7, IL-9, IL-15, IL-23 и IL-27, которые, как известно, активируют STAT3. Кроме того, или альтернативно, активация STAT3 осуществляется путем переноса в B-клетку нуклеиновой кислоты, кодирующей мутант STAT3, который придает STAT3 свойство конститутивной активации (Sean A Diehl, Heike Schmidlin, Maho Nagasawa, Simon D van Haren, Mark J Kwakkenbos, Etsuko Yasuda, Tim Beaumont, Ferenc A Scheeren, Hergen Spits STAT3-mediated up-regulation of BLIMP1 is coordinated with BCL6 down-regulation to control human plasma cell differentiation J Immunol 2008 vol. 180 (7) pp. 4805-15).
Наиболее предпочтительно, используется IL-21, так как IL-21 особенно подходит для воздействия на стабильность антитело-продуцирующей плазмабластоподобной B-клетки. Дополнительно к положительной регуляции STAT3, IL-21 способен положительно регулировать экспрессию Blimp-1, даже когда экспрессии Blimp-1 противодействует BCL6. С помощью способа, раскрытого в WO 2007/067046, становится возможным увеличение репликативного жизненного интервала антитело-продуцирующей клетки, так как возможно поддерживать B-клетку в той стадии развития, где происходит репликация. В более ранних ex vivo B-клеточных культурах репликационный жизненный интервал составлял только от нескольких недель до двух месяцев. В течение этого времени культивированные клетки теряют свою способность репликации и умирают. Однако с помощью способа, раскрытого в WO 2007/067046, становится возможным удлинение репликационного жизненного интервала антитело-продуцирующих B-клеток памяти, так чтобы генерировались ex vivo культуры, включающие плазмабластоподобные клетки, которые способны реплицироваться и продуцировать антитело.
Хотя эти способы дают возможность получения антител, которые эффективно направленно воздействуют на интересующий антиген, часто бывает целесообразным улучшение характеристик антитела, таких как аффинность связывания. Таким образом, характеристики связывания, как правило, изменяются путем введения мутаций в кодирующую нуклеиновую кислоту, предпочтительно, в участок, кодирующий CDR, и путем тестирования полученных в результате антител. Однако этот процесс требует больших затрат времени. Таким образом, целесообразна разработка альтернативных способов получения высокоаффинных антител.
Целью настоящего изобретения является предложение способов получения и/или селекции высокоаффинных антител.
В изобретении предлагаются средства и способы получения популяции B-клеток, начиная от данной B-клеточной культуры, и которая будет обладать более высокой способностью связывания, чем исходная B-клеточная культура. Предпочтительно, получают моноклональную B-клеточную популяцию, начиная с моноклональной B-клеточной культуры. В изобретении предлагается простой и изящный путь получения B-клеточных популяций с улучшенной средней способностью связывания без необходимости применения трудоемких методов мутаций.
В изобретении предлагается способ получения антител, специфичных к интересующему антигену, включающий:
а) селекцию B-клетки, способной продуцировать антитело, специфичное к указанному интересующему антигену, или селекцию B-клетки, способной дифференцироваться в B-клетку, которая способна продуцировать антитело, специфичное к указанному интересующему антигену;
b) индуцирование, усиление и/или поддержание экспрессии BCL6 в указанной B-клетке;
c) индуцирование, усиление и/или поддержание экспрессии антиапоптотической нуклеиновой кислоты в указанной B-клетке;
d) обеспечение размножения указанной B-клетки до получения популяции указанных B-клеток;
e) селекцию, по меньшей мере, одной B-клетки из указанной популяции B-клеток, продуцирующих B-клеточный рецептор и/или антитело, со способностью связывания выше, чем средняя способность связывания указанной популяции B-клеток для указанного интересующего антигена;
f) культивирование указанной, по меньшей мере, одной B-клетки до получения популяции B-клеток; и
g) получение антител, продуцированных B-клеточной культурой.
Внутри популяции моноклональных B-клеток, способных продуцировать антитело, специфичное к интересующему антигену, возможна селекция в стадии e) способа согласно изобретению, по меньшей мере, одной B-клетки, необязательно более чем одной, как например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 25 или 50 B-клеток со способностью связывания с указанным интересующим антигеном выше, чем средняя способность связывания указанной популяции B-клеток с указанным интересующим антигеном. Такие B-клетки с более высокой способностью связывания с интересующим антигеном, чем средняя способность связывания популяции B-клеток с указанным интересующим антигеном, в данном документе также называются "высокоаффинными B-клетками". Одна из возможных причин различия в способности связывания между множеством B-клеток в моноклональной популяции B-клеток связана с тем, что экспрессия BCR варьируется между B-клетками в указанной популяции. B-клетка с относительно высоким уровнем экспрессии BCR будет связываться с большим количеством интересующего антигена, чем B-клетка с относительно низким уровнем экспрессии BCR. Однако ожидается, что антитела, продуцированные B-клетками с различным уровнем экспрессии BCR, обладают одинаковой аффинностью связывания. Авторы настоящего изобретения неожиданно обнаружили, что помимо относительно высокого уровня экспрессии BCR, коллекция высокоаффинных B-клеток продуцирует антитела, специфичные к интересующему антигену, которые связываются с указанным антигеном с более высокой аффинностью, чем средняя аффинность антител, продуцируемых указанной популяцией B-клеток. Еще более неожиданно, авторы настоящего изобретения обнаружили, что В-клеточные культуры, полученные способом по изобретению, содержали клетки, которые связывались с антигеном с более высокой аффинностью, чем средняя В-клетка в исходной культуре. Таким образом, могут быть выделены одиночные В-клетки из данной В-клеточной популяции на основе их более высокой способности связывания с использованием способов, известных в данной области, и могут наращиваться с получением новой В-клеточной популяции, по меньшей мере, в течение трех недель. Эти новые В-клетки продуцируют антитела, которые обладают более высокой аффинностью, чем антитела, продуцированные исходной В-клеточной популяцией, из которой были выделены новые В-клетки. Это открытие противоречит ожиданиям, поскольку специалист в данной области ожидает, что после выделения одной В-клетки (субклон) из уже полученной моноклональной популяции указанных B-клеток аффинность к антигену у антитела, продуцированного потомством субклона указанной уже моноклональной В-клеточной популяции, будет возвращаться к средней аффинности для антигена по сравнению со средней аффинностью популяции B-клеток, из которой была селектирована по меньшей мере одна B-клетка.
Таким образом, в одном воплощении в стадии a) способа согласно изобретению предпочтительно селектируют одиночную B-клетку, например, из поликлональной популяции B-клеток. Затем одиночную B-клетку размножают до получения моноклональной популяции B-клеток в стадиях b) - d). Этого достигают, например с использованием способа, описанного в WO 2007/067046, который раскрыт в данном документе ранее. Затем в стадии d) получают моноклональную B-клеточную линию, специфичную к интересующему антигену. В принципе, все B-клетки в моноклональной B-клеточной линии продуцируют по существу одинаковые антитела, специфичные к указанному антигену, хотя между клетками указанной моноклональной В-клеточной линии могут присутствовать небольшие отличия в аффинности к указанному антигену, т.е. некоторые B-клетки в моноклональной популяции продуцируют антитела с аффинностью, которая немного выше, чем средняя аффинность, и некоторые B-клетки в моноклональной популяции продуцируют антитела с немного меньшей аффинностью. Популяция В-клеток становится снова в небольшой степени гетерогенной. В стадии e), из моноклональной В-клеточной линии селектируют, по меньшей мере, одну из таких B-клеток с более высокой аффинностью, чем средняя аффинность. B-клетки стадии f) или B-клетки, селектированные в стадии e), затем культивируют с получением второй, предпочтительно, моноклональной B-клеточной линии. В настоящем изобретении предлагается вариант, что эта вторая, предпочтительно моноклональная B-клеточная линия обладает средней аффинностью, которая выше, чем средняя аффинность исходной моноклональной B-клеточной популяции, полученной в стадии d). Как описано выше, неожиданно обнаружили, что высокая аффинность селектированной B-клетки поддерживается после культивирования, даже если культивирование осуществляется в течение более продолжительного периода времени, вместо возвращения к средней аффинности исходной популяции. Таким образом, вторая моноклональная популяция B-клеток, культивированных в стадии f), обладает более высокой средней аффинностью к антигену, чем моноклональная популяция B-клеток, культивированных в стадии d). Аналогично, аффинность большей части B-клеток во второй моноклональной популяции стадии f) выше, чем аффинность большей части B-клеток в моноклональной популяции стадии d).
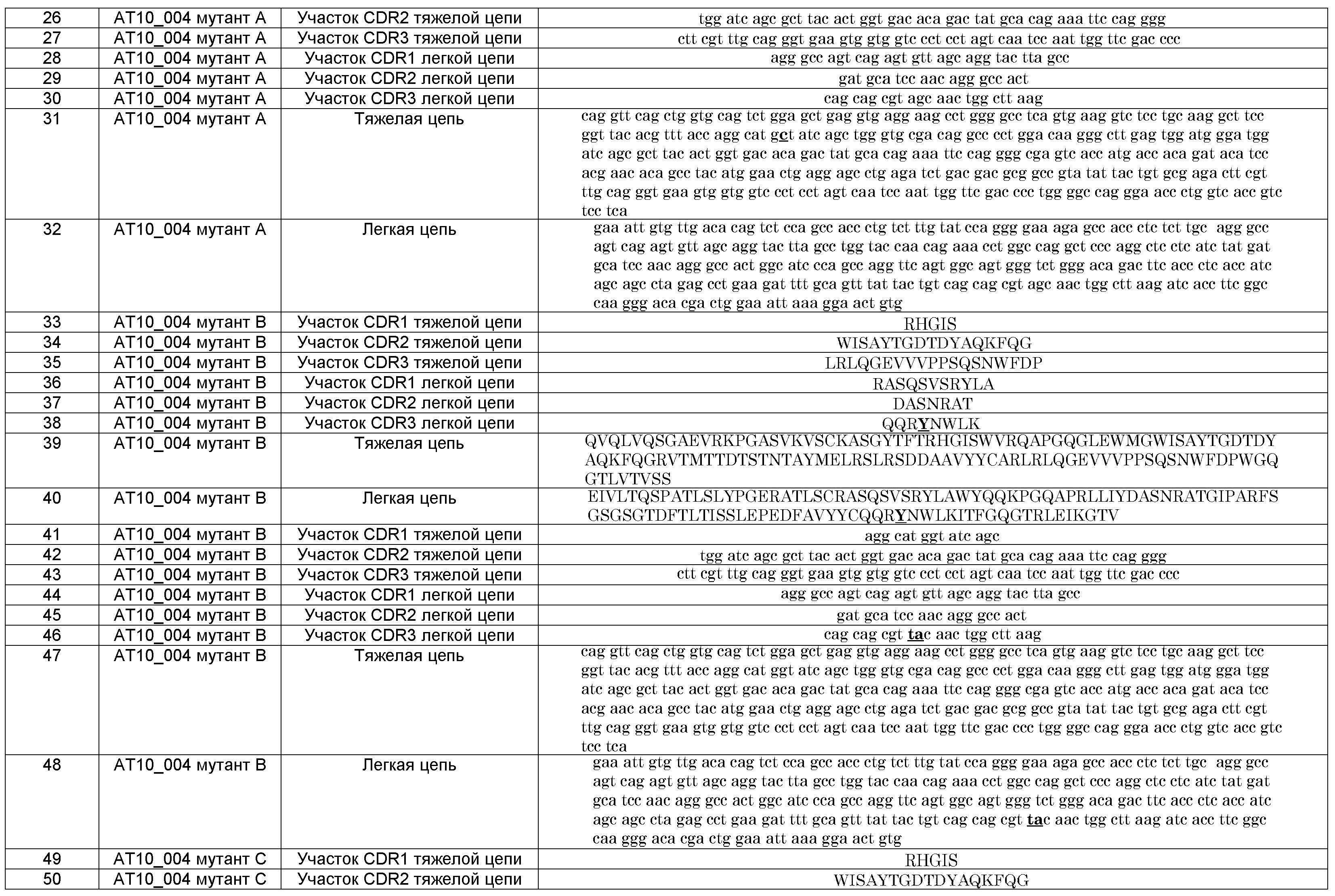
Таким образом, в одном воплощении изобретения предлагается способ получения B-клеточной популяции с повышенной средней аффинностью к интересующему антигену по сравнению с исходной моноклональной B-клеточной популяцией с данной средней аффинностью для указанного интересующего антигена, причем способ включает:
- получение моноклональной B-клеточной популяции, которая специфична к указанному интересующему антигену,
- селекцию, по меньшей мере, одной B-клетки из указанной популяции B-клеток, продуцирующих B-клеточный рецептор и/или антитело, со способностью связывания выше, чем способность связывания указанной популяции B-клеток для указанного интересующего антигена; и
- культивирование указанной, по меньшей мере, одной B-клетки до получения популяции B-клеток.
Кроме того, в изобретении предлагается способ получения антител, специфичных к интересующему антигену, включающий:
а) селекцию одиночной B-клетки, способной продуцировать антитело, специфичное к указанному интересующему антигену, или селекцию B-клетки, способной дифференцироваться в B-клетку, которая способна продуцировать антитело, специфичное к указанному интересующему антигену;
b) индуцирование, усиление и/или поддержание экспрессии BCL6 в указанной B-клетке;
c) индуцирование, усиление и/или поддержание экспрессии антиапоптотической нуклеиновой кислоты в указанной B-клетке;
d) обеспечение размножения указанной B-клетки в первую моноклональную B-клеточную линию;
e) селекцию из указанной первой моноклональной B-клеточной линии, по меньшей мере, одной B-клетки, которая продуцирует B-клеточный рецептор и/или антитело, со способностью связывания с указанным интересующим антигеном, которая выше, чем средняя способность связывания указанной первой моноклональной B-клеточной линии;
f) культивирование указанной, по меньшей мере, одной B-клетки, селектированной в стадии e), во вторую, предпочтительно, моноклональную B-клеточную линию; и
g) получение антител, продуцируемых указанной второй, предпочтительно, моноклональной B-клеточной линией. Получают антитела, которые обладают аффиностью к указанному интересующему антигену, которая выше, чем средняя аффинность к указанному интересующему антигену у антител, продуцированных B-клетками указанной первой моноклональной B-клеточной линии.
В другом воплощении, селектируют более чем одну B-клетку в стадии a) способа по изобретению, например, 2, 3, 4, 5, 10, 15, 25, 50 или 100 B-клеток. Например, B-клетки селектируют из поликлональной популяции B-клеток или из биологического образца. Селектированные B-клетки затем размножают с получением популяции B-клеток в стадиях b) - d), например, с использованием способа, описанного в WO 2007/067046. Таким образом, полученная B-клеточная популяция представляет собой (вторую) поликлональную B-клеточную популяцию. После этого и перед проведением стадии e) способа по изобретению предпочтительно получают моноклональную популяцию B-клеток. Это, например, осуществляется путем селекции одиночной B-клетки из указанной (второй) поликлональной популяции B-клеток с использованием клеточного сортера с активацией флуоресценции или с помощью лимитирующего разведения, которые объяснены в данном документе ниже, и путем размножения указанной селектированной одиночной B-клетки до получения моноклональной популяции В-клеток. Затем проводят стадию e) способа по изобретению, в которой селектируют, по меньшей мере, одну B-клетку с более высокой аффинностью, чем средняя аффинность моноклональной B-клеточной популяции. В стадии f) B-клетка или B-клетки, селектированные в стадии e), затем культивируют до получения второй моноклональной B-клеточной линии, после чего в стадии g) получают антитела, продуцированные указанной второй моноклональной B-клеточной линией.
Способ, описанный в данном документе, дает возможность получения улучшенных высокоаффинных антител, предпочтительно моноклональных антител без использования методов рекомбинантной ДНК. До настоящего изобретения аффинность (моноклональных) антител повышали с использованием таких методов рекомбинантной ДНК. Во-первых, необходимо определить последовательность нуклеиновой кислоты, кодирующей антитело. Затем в последовательность, кодирующую антитело, вводят одну или более мутаций. Затем в клетке необходимо экспрессировать гены, содержащие одну или более мутаций, с последующим продуцированием антител в клетках-продуцентах. Наконец, мутированное антитело необходимо тестировать на предмет его способности связывания с интересующим антигеном с целью определения, получено ли антитело, обладающее улучшенной аффинностью по отношению к указанному антигену по сравнению с не мутированным антителом. Такой процесс улучшения аффинности антитела является сложным и затратным по времени. Способ согласно настоящему изобретению дает возможность получения высокоаффинного антитела в прямом и менее сложном процессе без необходимости молекулярного конструирования.
В одном воплощении изобретения, после стадии селекции, по меньшей мере, одной высокоаффинной B-клетке из указанной уже моноклональной популяции B-клеток (стадия e) способа по изобретению, описанному выше, по меньшей мере, одной указанной высокоаффинной B-клетке дают возможность размножиться до получения популяции B-клеток, предпочтительно, моноклональной B-клеточной линии, и после чего снова осуществляют другую стадию селекции, по меньшей мере, одной высокоаффинной B-клетки из указанной новой популяции B-клеток, предпочтительно, из указанной новой моноклональной B-клеточной линии. Путем повторения стадий, дающих возможность размножения селектированной B-клетки в популяцию и путем селекции, по меньшей мере, одной B-клетки на основе ее способности связывания с антигеном, т.е. повторение стадий d) и e), возможно получить В-клетки, продуцирующие высокоаффинное антитело. Предпочтительно, путем повторения стадий размножения и селекции, описанных выше, возможно увеличение с каждым циклом селекции аффинности антитела, продуцированного полученной B-клеточной популяцией, к интересующему антигену.
Таким образом, предлагается способ, включающий после стадии e) способа по изобретению повторение стадии, дающей возможность размножения, по меньшей мере, одной селектированной высокоаффинной B-клетки до получения популяции B-клеток, предпочтительно, моноклональной B-клеточной линии, и повторение селекции, по меньшей мере, одной высокоаффинной B-клетки, т.е., повторение стадий d) и e) способа по изобретению, по меньшей мере, один раз. Указанные стадии повторяют, например, один раз, но предпочтительно, два раза, три раза, четыре раза, пять раз или даже более раз.
В одном воплощении предлагается способ по изобретению, где указанную, по меньшей мере, одну B-клетку, селектированную в стадии e), культивируют в течение, по меньшей мере, четырех недель. Предпочтительно, указанную, по меньшей мере, одну B-клетку, селектированную в стадии e), культивируют в течение, по меньшей мере, шести недель, более предпочтительно, в течение, по меньшей мере, девяти недель, более предпочтительно, в течение, по меньшей мере, трех месяцев, более предпочтительно, в течение, по меньшей мере, шести месяцев.
Если не быть связанными никакой теорией, то можно считать, что различия аффинности антител по отношению к интересующему антигену в популяции моноклональных B-клеток может быть связано с процессами, опосредованными активацией, Индуцированной Цитидин-Дезаминазой (AID). Антиген-активированные наивные В-клетки и B-клетки памяти в зародышевом центре подвергаются интенсивной пролиферации, сопровождающейся соматическими гипермутациями (SHM) и рекомбинацией на этапе переключения (CSR) генов Ig, опосредованными AID. AID дезаминирует дезоксицитидиновые остатки в генах иммуноглобулинов, которые запускают процесс расхождения антител. Было продемонстрировано в патентной заявке US2008305076, что IL-21 индуцирует экспрессию BLIMP, BCL6 и AID, но не индуцирует напрямую соматическую гипермутацию. Однако авторы настоящего изобретения обнаружили, что AID экспрессируется в B-клетках, которые культивируются согласно способу, описанному в данном документе. Экспрессия AID в (B-клетке, которая разовьется в) антитело-продуцирующей B-клетке дает возможность получения новых иммуноглобулинов, которые несут на себе мутации, которых нет в исходной B-клетке перед трансдукцией с помощью BCL6 и анти-апоптотической нуклеиновой кислоты. Таким образом, культивирование B-клеток, в которых соматическая гипермутация индуцирована экспрессией AID, дает возможность получения вариантов иммуноглобулинов, которые, например, обладают большей или меньшей аффинностью по отношению к интересующему антигену, или которые более стабильны, например, в водном растворе или при условиях повышенного содержания соли, или при любой их комбинации.
При селекции, по меньшей мере, одной высокоаффинной B-клетки из указанной популяции B-клеток, AID все еще продолжает экспрессироваться в указанной селектированной, по меньшей мере, одной B-клетке. Таким образом, после селекции такой B-клетки, AID в указанной B-клетке все еще дает возможность введения мутаций в ген иммуноглобулина потомства указанной B-клетки. Соматические гипермутации в генах иммуноглобулинов происходят предпочтительно в участке CDR3 генов Ig. Мутации, введенные в участок CDR3 иммуноглобулина, наиболее вероятно приведут к уменьшению или к потере аффинности связывания с антигеном у указанного иммуноглобулина, а не к повышению аффинности связывания. Однако авторы настоящего изобретения обнаружили повышение аффинности связывания.
При использовании в данном документе, термин "анти-апоптотическая нуклеиновая кислота" обозначает нуклеиновую кислоту, которая способна замедлять и/или предотвращать апоптоз в B-клетке. Предпочтительно, указанная анти-апоптотическая нуклеиновая кислота способна замедлять и/или предотвращать апоптоз в антитело-продуцирующей B-клетке. Предпочтительно, используется анти-апоптотическая нуклеиновая кислота, которая включает экзогенную нуклеиновую кислоту. Это означает, что либо используется последовательность нуклеиновой кислоты, которая в природе не экспрессируется в B-клетках, либо то, что используется дополнительная копия природной нуклеиновой кислоты, так что экспрессия в полученных B-клетках увеличивается по сравнению с природными B-клетками. В данной области известны различные анти-апоптотические нуклеиновые кислоты, так что доступны различные воплощения. Предпочтительно, используется ген, кодирующий анти-апоптотическую молекулу. Более предпочтительно, используется нуклеиновая кислота, которая является анти-апоптотическим членом семейства Bcl-2, так как анти-апоптотические белки Bcl-2 являются хорошими ингибиторами апоптоза. Множество процессов, которые контролируются семейством Bcl-2 (которое включает как про- так и анти-апоптотические белки), относятся к митохондриальному пути апоптоза, более подробно описанному ниже. Анти-апоптотические члены семейства Bcl-2: Bcl-2, Bcl-xL, Bcl-w, Al и Mcl-1 являются предпочтительными, так как они, как правило, интегрированы с внешней митохондиральной мембраной. Они непосредственно связываются и ингибируют про-апоптотические белки, которые принадлежат семейству Bcl-2, для защиты целостности митохондриальной мембраны.
В особенно предпочтительном воплощении, указанные анти-апоптотические нуклеиновые кислоты кодируют Bcl-xL и/или Mcl-1 и/или функциональную часть Bcl-xL и/или функциональную часть Mcl-1. Комбинация нуклеиновых кислот BCL6 и Bcl-xL, а также комбинация нуклеиновых кислот BCL6 и Mcl-1 конкретно подходит для иммортализации B-клеток и для долгосрочной культуры полученных плазмабластоподобных B-клеток. Наиболее предпочтительно, указанные анти-апоптотические нуклеиновые кислоты кодируют Bcl-xL или его функциональную часть, так как комбинация BCL6 и Bcl-xL стабилизирует B-клетки особенно хорошо.
Функциональная часть Bcl-xL и функциональная часть Mcl-1 определены в данном документе как фрагменты Bcl-xL и Mcl-1, соответственно, которые сохраняют тот же тип анти-апоптотических характеристик, как и полноразмерный Bcl-xL и Mcl-1, соответственно, по качеству (но необязательно по количеству). Функциональные фрагменты Bcl-xL и Mcl-1, как правило, представляют собой более короткие фрагменты Bcl-xL и Mcl-1, которые способны замедлять и/или предотвращать апоптоз в B-клетке. Такие функциональные фрагменты, например, лишены последовательностей, которые не способствуют анти-апоптотической активности Bcl-xL или Mcl-1.
Популяция B-клеток по изобретению предпочтительно представляет собой моноклональную популяцию B-клеток. Примером популяции B-клеток по изобретению является клеточная линия B-клеток, предпочтительно, моноклональных B-клеток. Следовательно, популяция B-клеток по изобретению наиболее предпочтительно представляет собой моноклональную B-клеточную линию. Возможность размножения указанной B-клетки до получения популяции указанных B-клеток осуществляется посредством культивирования указанной B-клетки до получения популяции указанных B-клеток.
Внутри популяции B-клеток, даже в популяции моноклональных B-клеток, способность связывания BCR из B-клеток указанной популяции и способность связывания антител, продуцированных B-клетками указанной популяции, не равны между собой. Наоборот, имеют место вариации в указанной способности связывания. Средняя способность связывания популяции B-клеток определяется в данном документе как средняя способность связывания или средняя аффинность BCR и/или антитела индивидуальных B-клеток в указанной популяции. Средняя аффинность для интересующего антигена у антитела, продуцированного B-клеткой или популяцией B-клеток, определяется в данном документе как среднее значение аффинностей антител, продуцированных всеми индивидуальными В-клетками в указанной популяции, для указанного интересующего антигена. Высокоаффинная B-клетка из популяции B-клеток, предпочтительно, из моноклональной B-клеточной линии, согласно изобретению предпочтительно селектируется из более чем 40% B-клеток популяции, предпочтительно, из моноклональной B-клеточной линии по отношению к способности связывания и/или аффинности, предпочтительно, из более чем 30% B-клеток указанной популяции или из моноклональной B-клеточной линии, более предпочтительно, из более чем 25% B-клеток указанной популяции или из моноклональной B-клеточной линии, более предпочтительно, из более чем 20% B-клеток указанной популяции или из моноклональной B-клеточной линии, более предпочтительно, из более чем 15% B-клеток указанной популяции или из моноклональной B-клеточной линии, более предпочтительно, из более чем 10% B-клеток указанной популяции или из моноклональной B-клеточной линии, более предпочтительно, из более чем 1% B-клеток указанной популяции или из моноклональной B-клеточной линии. В одном воплощении, одну высокоаффинную B-клетку селектируют из более чем 1% B-клеток популяции или из моноклональной B-клеточной линии по отношению к способности связывания и/или к аффинности.
Для интересующего антигена средняя аффинность антитела, продуцированного популяцией B-клеток, предпочтительно, моноклональной B-клеточной линией, культивированной из, по меньшей мере, одной высокоаффинной B-клетки согласно изобретению, предпочтительно составляет, по меньшей мере, в 1,1 раз больше средней аффинности для указанного интересующего антигена популяции B-клеток, из которых, была селектирована, по меньшей мере, одна высокоаффиннная B-клетка, более предпочтительно, по меньшей мере, в 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2, 2,1, 2,2, 2,3, 2,4, 2,5, 3, 3,5, 4, 5, 10, 20, 50, в 100 или более раз больше средней аффинности для указанного интересующего антигена.
Аффинность антитела может быть определена с использованием любого метода, известного специалисту в данной области. Аффинность антитела определяют, например, с использованием твердофазного иммуноферментного анализа (тИФА), Поверхностного Плазмонного Резонанса (такого как Biacore) или Octet (ForteBio). Поверхностный Плазмонный Резонанс (SPR) и Octet представляют собой методы для измерения биомолекулярных взаимодействий в реальном времени в среде без метки. Для SPR, одно из взаимодействующих веществ, например, антитело, иммобилизуют на сенсорной поверхности, другое, например, антиген находится в растворе в свободном состоянии и пропускается над поверхностью. Ассоциацию и диссоциацию измеряют в условных единицах и предпочтительно выводят на сенсограмму. Любое изменение в количестве молекул, связанных с биосенсорной областью, вызывает сдвиг в профиле интерференции, который может быть измерен в реальном времени. При использовании Octet анализируют профиль интерференции белого света, отраженного от двух поверхностей, слоя иммобилизованного белка на биосенсорной области, и от внутреннего эталонного слоя. Связывание между лигандом, иммобилизованным на поверхности биосенсорной области, например, антителом, и белком в растворе, например, в виде интересующего антигена, производит увеличение оптической толщины в биосенсорной области, что приводит к сдвигу длины волны, которая является прямой мерой изменения толщины биологического слоя. тИФА включает иммобилизацию белка, например, интересующего антигена, на поверхности твердой подложки, например, на 96-луночном планшете, и применение образца, который детектируют или оценивают количественно на твердой подложке. Альтернативно, захватывающее антитело фиксируют на поверхности твердой подложки, после чего образец, содержащий белок, который необходимо детектировать или оценить количественно, применяют к иммобилизованному захватывающему антителу, так чтобы у интересующего белка была возможность связаться. Не связавшиеся белки затем смывают. Затем, добавляют к твердой подложке специфическое антитело, конъюгированное с меткой или ферментом (или первичное антитело с последующим вторичным антителом, конъюгированным с меткой или ферментом). Предпочтительно, определяют константу аффинности (Kd) антитела, продуцированного B-клеткой согласно изобретению.
Связывание B-клетки согласно изобретению с интересующим антигеном может быть измерено с использованием метода, известного специалисту в данной области. Например, интересующий антиген метят с помощью, например, флуоресцентной метки. Детектирование связывания может затем определяться с помощью различных методов, среди которых флуоресцентная микроскопия и использование клеточного сортера с возбуждением флуоресценции (FACS). FACS дает возможность разделения клеток в суспензии на основе размера, и при этом детектируют флуоресценцию конъюгированных антител против поверхностных антигенов.
Селекция, по меньшей мере, одной высокоаффинной B-клетки из популяции B-клеток, предпочтительно, из моноклональной B-клеточной линии, может осуществляться с использованием любого метода, известного специалисту в данной области. Селекция, по меньшей мере, одной высокоаффинной B-клетки согласно изобретению осуществляется, например, с помощью клеточного сортинга, например, с использованием FACS (см. выше) или с использованием метода лимитирующего разведения. Лимитирующее разведение включает последовательное разведение суспензии клеток, например, B-клеток, до тех пор, пока в данном объеме не будет присутствовать одна клетка. Затем, способность связывания каждой B-клетки (после размножения одиночной клетки до получения популяции) тестируют на предмет возможности селекции B-клетки, продуцирующей антитела с высокой аффинностью к антигену.
B-клетка, способная продуцировать антитело, определяется как B-клетка, которая способна продуцировать и/или секретировать антитело или его функциональную часть, и/или которая способна развиваться в клетку, которая способна продуцировать и/или секретировать антитело или его функциональную часть.
Функциональная часть антитела определяется как часть, которая обладает, по меньшей мере, одним свойством как у указанного антитела, по качеству, но необязательно по количеству.
Указанная функциональная часть предпочтительно способна связываться с тем же антигеном, что и указанное антитело, хотя и необязательно в такой же степени. Функциональная часть антитела предпочтительно включает однодоменное антитело, одноцепочечное антитело, FAB-фрагмент, нанотело, унитело (unibody), одноцепочечный вариабельный фрагмент (scFv), или F(ab')2-фрагмент.
Частные примеры B-клетки, используемой или селектированной в способе согласно изобретению, включают B-клетки, выделенные из человека, грызуна, кролика, ламы, свиньи, коровы, козы, лошади, примата, шимпанзе, макаки и гориллы. Предпочтительно, указанная B-клетка является человеческой клеткой, мышиной клеткой, кроличьей клеткой, клеткой примата, клеткой шимпанзе, клеткой макаки и/или клеткой ламы. Наиболее предпочтительно, указанная B-клетка является человеческой B-клеткой.
В предпочтительном воплощении, B-клетку памяти селектируют в стадии a) способа, раскрытого в данном документе, например, человеческую B-клетку памяти. В особенно предпочтительном воплощении, указанная B-клетка памяти представляет собой В-клетку памяти периферической крови. B-клетки памяти периферической крови легко получают без особого дискомфорта для индивидуума, из которого их выделяют, и как оказалось, они очень подходят для применения в способе согласно настоящему изобретению.
B-клетка или популяция B-клеток, предпочтительно, моноклональная B-клеточная линия, полученная с помощью способа согласно изобретению, предпочтительно стабильна в течение, по меньшей мере, четырех недель, более предпочтительно, в течение, по меньшей мере, шести недель, более предпочтительно, в течение, по меньшей мере, девяти недель, более предпочтительно, в течение, по меньшей мере, трех месяцев, более предпочтительно, в течение, по меньшей мере, шести месяцев, это означает, что такие B-клетки способны как к репликации, так и к продуцированию антитела, или способны реплицироваться или развиваться в клетку, которая продуцирует антитело в течение указанного периода времени. B-клетки согласно изобретению предпочтительно включают клетки, продуцирующие IgM, или клетки, продуцирующие иммуноглобулины других изотипов типа IgG, или IgA, или IgE, предпочтительно, IgG. B-клетка согласно изобретению особенно подходит для применения в получении антитело-продуцирующей клеточной линии. Высокоаффинные B-клетки или популяция или моноклональная B-клеточная линия высокоаффинных B-клеток согласно изобретению предпочтительно культивируют ex vivo, и антитело предпочтительно собирают для последующего применения. Антитела или их функциональные части, полученные способом согласно изобретению, применяются для широкого спектра нужд, таких как например, терапевтические, профилактические и диагностические применения, а также для исследовательских целей и ex vivo экспериментов. Например, скрининговый анализ осуществляют в случае, где антитела или функциональные части согласно изобретению инкубируют с образцом с целью определения присутствия интересующего антигена.
В одном воплощении, высокоаффинная B-клетка или популяция или моноклональная B-клеточная линия высокоаффинных B-клеток согласно изобретению включает человеческую B-клетку, способную продуцировать человеческое антитело, так как человеческие антитела особенно подходят для терапевтических и/или профилактических применений у человеческих индивидуумов.
Экспрессия BCL6 в B-клетке индуцируется, усиливается и/или поддерживается различными путями. В одном воплощении, предлагается B-клетка, содержащая нуклеиновую кислоту, кодирующую BCL6 или его гомолог. В другом воплощении, предлагается B-клетка, содержащая соединение, способное прямо или косвенно усиливать экспрессию BCL6. Такое соединение предпочтительно включает белок Переносчик Сигнала Активации и Транскрипции 5 (STAT5) или его функциональную часть, производное и/или аналог, и/или последовательность нуклеиновой кислоты, кодирующую его. STAT5 представляет собой переносчик сигнала, способный усиливать экспрессию BCL6. Известны две формы STAT5, STAT5a и STAT5b, которые кодируются двумя различными тандемно расположенными генами. Введение и/или активация STAT5 приводит к повышению уровня BCL6. Следовательно, отрицательная регуляция BCL6 с помощью Blimp-1, по меньшей мере, частично компенсируется положительной регуляцией экспрессии BCL6 с помощью STAT5 или его функциональной части, производного и/или аналога. Следовательно, STAT5 или его функциональная часть, производное и/или аналог способен к прямому воздействию на экспрессию BCL6. Также возможно косвенное воздействие на экспрессию BCL6. Это осуществляется, например, путем регуляции количества соединения, которое в свою очередь способно прямо или косвенно активировать STAT5 и/или регулировать экспрессию STAT5. Следовательно, в одном воплощении экспрессия и/или активность эндогенного и/или экзогенного STAT5 повышается. Это, например, возможно для косвенного повышения экспрессии BCL6 путем культивирования антитело-продуцирующей клетки в присутствии интерлейкина (IL) 2 и/или IL-4 или других цитокинов, которые способны активировать STAT5.
Кроме того, предпочтительно, что в способе согласно изобретению указанные B-клетки в некоторый момент инкубировали вместе с IL-21 и CD40L. B-клетку, такую как антитело-продуцирующая плазмабластоподобная B-клетка, предпочтительно культивируют в присутствии CD40L, так как он содействует репликации большинства B-клеток. Кроме того, предпочтительно, чтобы STAT3 активировался в указанной B-клетке. Наиболее предпочтительно, используется IL-21 для положительной регуляции STAT3, так как IL-21 особенно подходит для воздействия на стабильность B-клетки согласно изобретению. Дополнительно к положительной регуляции STAT3, IL-21 способен положительно регулировать экспрессию Blimp-1, даже когда экспрессии Blimp-1 противодействует BCL6.
В другом воплощении, количество продукта экспрессии Blimp-1 в указанной B-клетке, селектированной в стадии a) способа по изобретению, контролируется прямо или косвенно. Количество продукта экспрессии Blimp-1 может контролироваться различными путями, например, путем регуляции STAT3 или его функциональной части, производного или аналога. STAT3 активируется различными путями. Предпочтительно, STAT3 активируется с помощью предоставления B-клетки согласно изобретению вместе с цитокином. Цитокины, будучи природно вовлеченными в В-клеточную пролиферацию, очень эффективны в регуляции STAT-белков. Очень эффективные активаторы STAT3 - это IL-2, IL-10, IL-21 и IL-6, а также IL-7, IL-9, IL-15, IL-23 и IL-27, которые, как известно, активируют STAT3. Кроме того, Toll-подобные рецепторы (TLR), которые вовлечены во врожденный иммунитет, также способны активировать STAT3. Наиболее предпочтительно, используется IL-21. IL-21 способен положительно регулировать экспрессию Blimp-1, даже когда экспрессии Blimp-1 противодействует BCL6.
Под функциональной частью STAT5 или STAT3 обозначают белковоподобную молекулу, которая обладает такой же способностью - по качеству, но необязательно по количеству - по части воздействия на стабильность антитело-продуцирующей клетки по сравнению с STAT5 или STAT3, соответственно. Функциональная часть белка STAT5 или белка STAT3, например, лишена аминокислот, которые не вовлечены или слабо вовлечены в указанную способность. Производное STAT5 или STAT3 определяется как белок, который был изменен так, что способность указанного белка по части воздействия на стабильность антитело-продуцирующей клетки такая же по качеству, но необязательно по количеству. Производное получают различными путями, например, посредством консервативной аминокислотной замены, где одну аминокислоту заменяют на другую аминокислоту, обладающую, как правило, аналогичными свойствами (размером, гидрофобностью, и т.д.), так что общая функциональность, вероятно, не подвержена существенным воздействиям. Производное, например, включает гибридный белок, такой как гибридный белок STAT5-ER, чья активность зависит от присутствия 4 гидрокситамоксифена (4HT). Аналог STAT5 или STAT3 определяется как молекула, обладающая такой же способностью воздействия на стабильность антитело-продуцирующей клетки по качеству, но необязательно по количеству. Указанный аналог необязательно выделен из указанного белка STAT5 или STAT3.
Способ по изобретению предпочтительно используется для получения клеточной линии высокоаффинных B-клеток, которая стабильна, в течение, по меньшей мере, одной недели, предпочтительно, в течение, по меньшей мере, одного месяца, более предпочтительно, в течение, по меньшей мере, трех месяцев, более предпочтительно, в течение, по меньшей мере, шести месяцев, так чтобы было возможно коммерческое получение высокоаффинного антитела. Предпочтительно, получают стабильную клеточную линию, способную продуцировать моноклональные высокоаффинные антитела. Это предпочтительно осуществляется путем использования B-клеток памяти, которые, например, были выделены из образца путем селекции на CD19 (B-клеточный маркер) и на белок клеточной поверхности IgG и/или CD27 (для маркировки клеток памяти). Кроме того, B-клетку памяти, способную специфически связываться с интересующим антигеном, селектируют, например, в анализе связывания с использованием указанного интересующего антигена. Затем, BCL6 и анти-апоптотическую нуклеиновую кислоту, предпочтительно, Bcl-XL или Mcl-1, со-экспрессируют в указанной B-клетке с получением популяции клеток, специфичных к указанному интересующему антигену. Предпочтительно, селектируют только одну клетку памяти в стадии a) способа, описанного в данном документе, так что получают B-клеточную популяцию согласно изобретению, продуцирующую моноклональные антитела (моноклональная B-клеточная линия).
В одном воплощении, в способе согласно изобретению используют B-клетку, предпочтительно, но необязательно B-клетку памяти, которая происходит из индивидуума, который ранее подвергался воздействию интересующего антигена. Однако это не является необходимым. Также возможно использование B-клетки из индивидуума, который не подвергался воздействию указанного интересующего антигена. Например, используется B-клетка, которая специфична к другому антигену, но демонстрирует кросс-реактивность по отношению к интересующему антигену. В качестве другого примера, использовалась B-клетка, которую селектировали из популяции наивных B-клеток индивидуума. Популяция наивных B-клеток индивидуума может содержать B-клетки, которые демонстрируют реактивность с интересующим антигеном, даже если индивидуум не подвергался воздействию интересующего антигена. Такую B-клетку из популяции наивных B-клеток селектируют, например, с использованием меченого интересующего антигена.
Кроме того, в изобретении предлагаются выделенные или рекомбинантные B-клетки и популяции B-клеток, предпочтительно, моноклональные B-клеточные линии, полученные способом согласно изобретению. Такие высокоаффинные B-клетки предпочтительно стабильны в течение, по меньшей мере, одной недели, предпочтительно, в течение, по меньшей мере, одного месяца, более предпочтительно, в течение, по меньшей мере, трех месяцев, более предпочтительно, в течение, по меньшей мере, шести месяцев, это означает, что B-клетка способна как к репликации, так и к продуцированию антитела или способна реплицироваться или развиваться в клетку, которая продуцирует антитело в течение указанных периодов времени. B-клетки согласно изобретению предпочтительно включают клетки, продуцирующие IgM, или клетки, продуцирующие иммуноглобулины других изотипов типа IgG или IgA, или IgE, предпочтительно, IgG. B-клетка согласно изобретению особенно подходит для применения в получении антитело-продуцирующей клеточной линии. Высокоаффинные B-клетки согласно изобретению предпочтительно культивируют ex vivo, и антитело предпочтительно собирают для последующего применения. Также предлагаются антитела, полученные из B-клетки или из B-клеточной популяции, или из моноклональной клеточной линии согласно изобретению. Высокоаффинные антитела или их функциональные части, полученные способом согласно изобретению, применяются для широкого спектра применений, таких как, например, терапевтические, профилактические и диагностические применения, а также для исследовательских целей и ex vivo экспериментов. Например, анализ скрининга осуществляют в случае, где антитела или функциональные части согласно изобретению инкубируют с образцом с целью определения присутствия интересующего антигена.
B-клетки, полученные способом согласно изобретению, особенно подходят для продуцирования высокоаффинных антител против интересующего антигена. Однако в одном предпочтительном воплощении, гены, кодирующие тяжелые и/или легкие цепи Ig, выделяют из указанной клетки и экспрессируют во второй клетке, такой как, например, клетка клеточной линии яичника китайского хомячка (CHO). Указанная вторая клетка, также называемая в данном документе как клетка-продуцент, предпочтительно адаптирована для коммерческого получения антител. Пролиферация указанной клетки-продуцента приводит к получению клеточной линии-продуцента, способной продуцировать антитело. Предпочтительно, указанная клеточная линия-продуцент подходит для продуцирования соединений для применения у человека. Следовательно, указанная клеточная линия-продуцент предпочтительно не содержит патогенных агентов, таких как патогенные микроорганизмы.
Изобретение дополнительно описано с помощью следующих частных примеров.
Подписи к чертежам
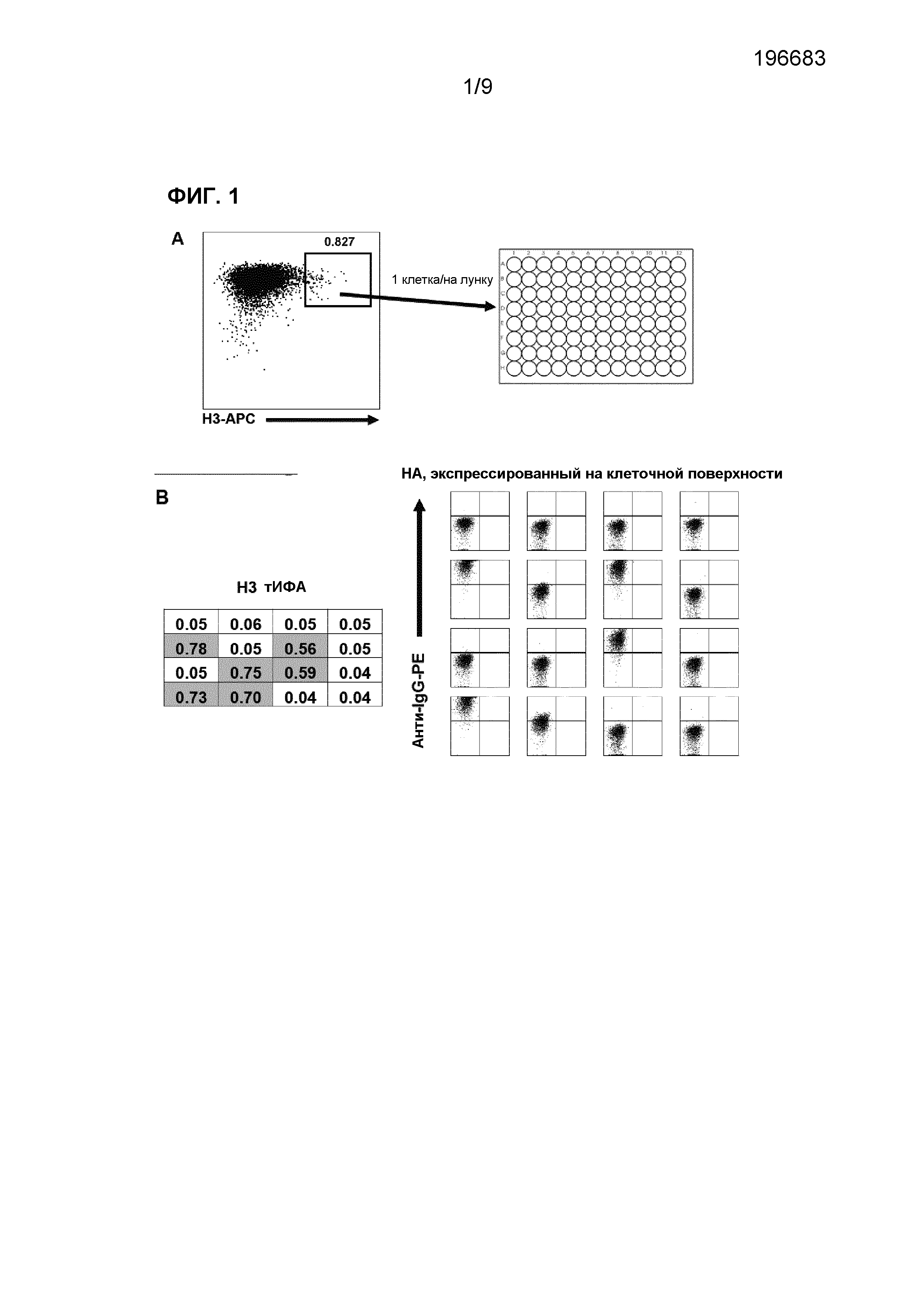
Фиг.1. (A) Связывание меченого белка НА H3 с BCR из H3-специфичных клеток внутри поликлональной В-клеточной популяции. В-клетки, которые связываются с белком H3 с высокой аффинностью, клонировали с помощью клеточного сортинга одиночной клетки. Через 2-3 недели после культивирования надосадочную жидкость культуры скринировали на H3-специфичные антитела. (B) Пример скрининга, осуществляемого для селекции H3-специфичных клонов (левая панель). Скрининг с помощью тИФА. Рекомбинантный белок H3 наносили в виде покрытия на планшет с последующим инкубированием в присутствии надосадочной жидкости культуры. Связывание антитела детектировали с использованием IgG-HRP, связывающего человеческий белок (правая панель). Скрининг на HA, экспрессирующийся на клеточной поверхности. H3N2-инфицированные клетки инкубировали вместе с надосадочной жидкостью В-клеточной культуры. Связывание антитела детектировали с помощью PE-меченого козьего антитела, связывающего человеческий F(ab')2.
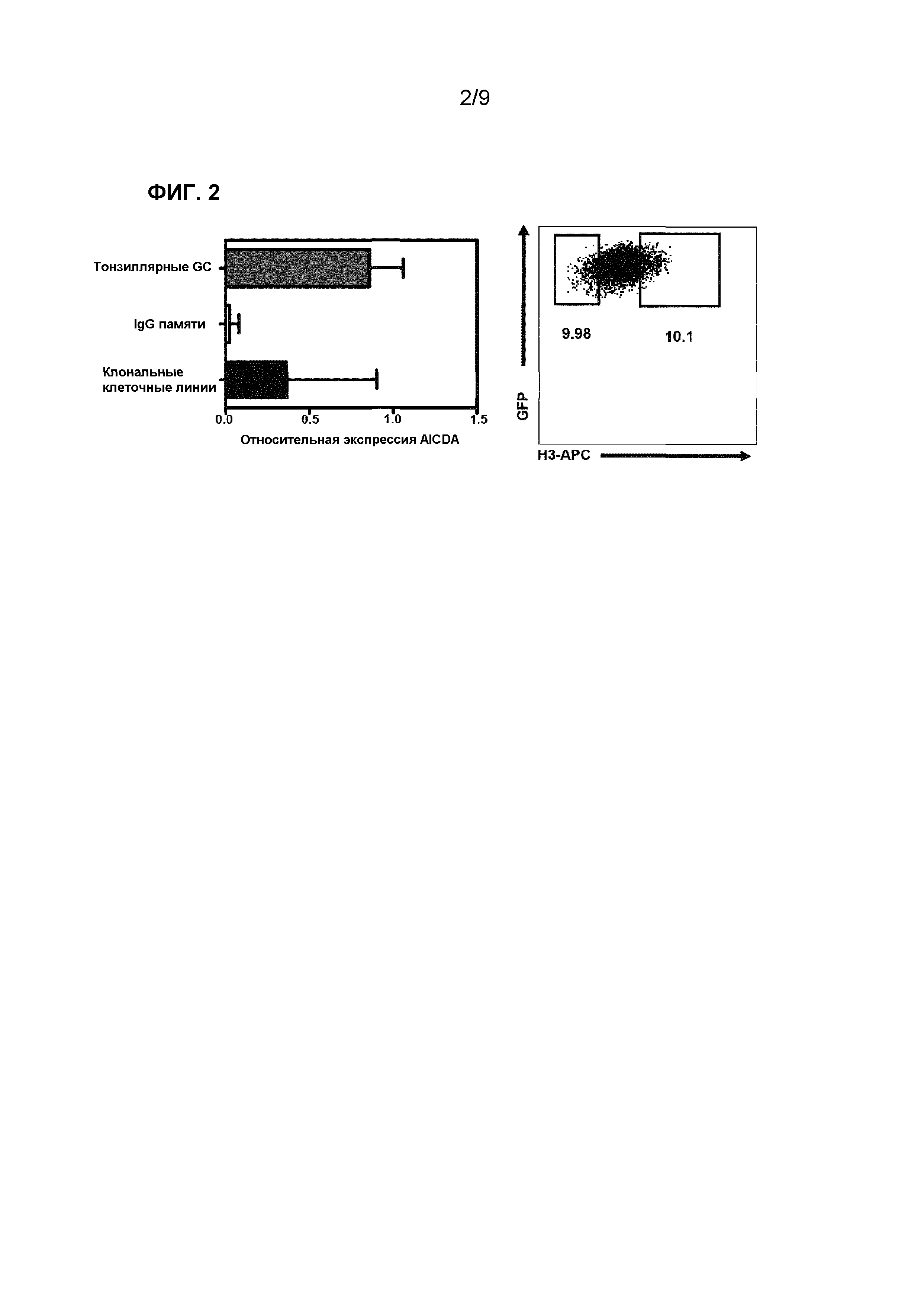
Фиг.2. (Слева) Уровень мРНК AICDA в CD19+CD38+CD20+IgD-тонзиллярных В-клетках GC и в CD19+IgG+CD27+В-клетках памяти периферической крови по сравнению с 23 BCL6- и Bcl-xL-трансдуцированными моноклональными клеточными линиями. (Справа) Селекция субклонов с сильным и слабым связыванием внутри H3-специфичного клона. Обведенные популяции селектировали с помощью клеточного сортинга и далее размножали.
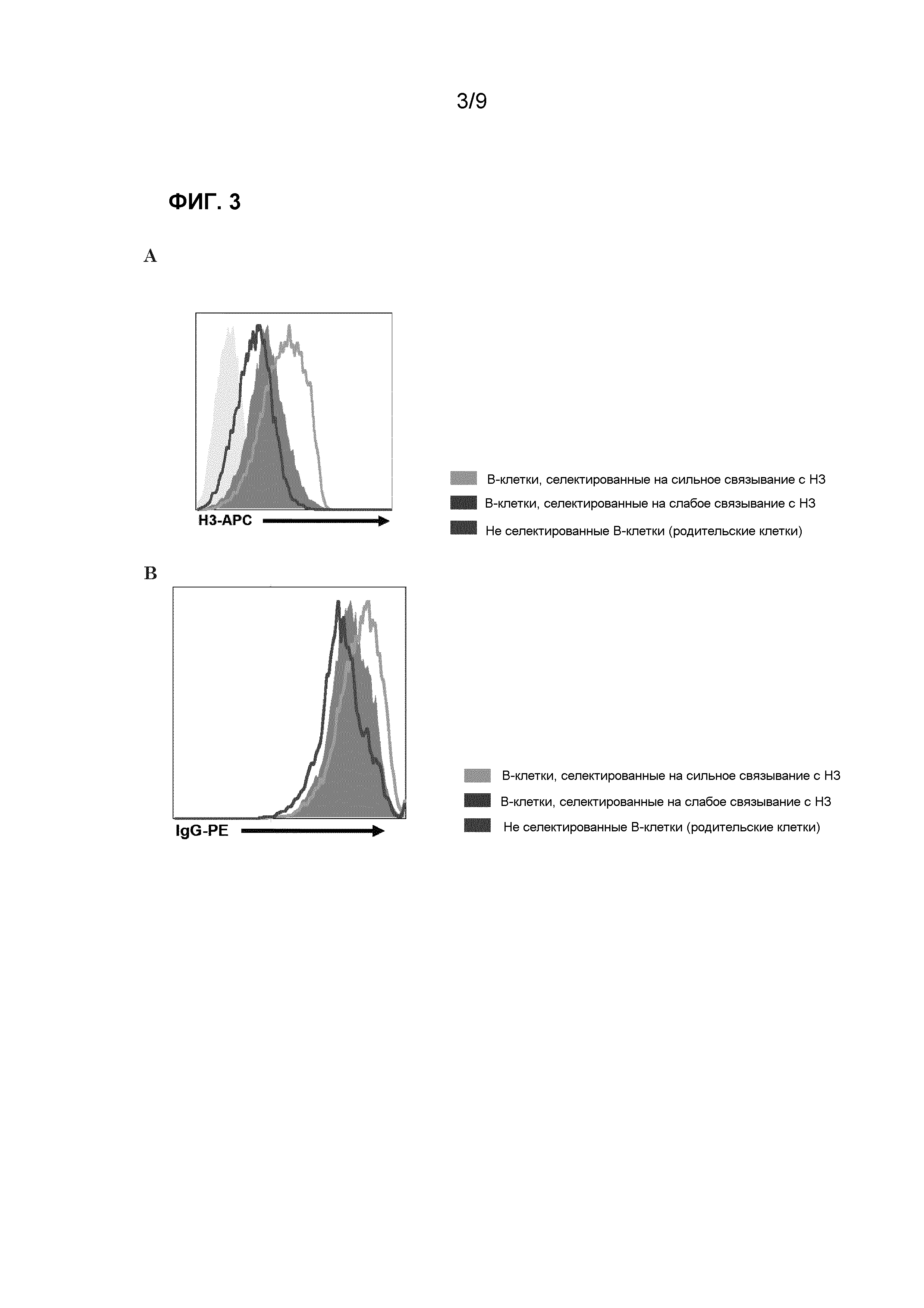
Фиг.3. A) FACS-анализ связывания меченых НА H3 с селектированными клетками через 13 дней после их селекции для более сильного или более слабого связывания BCR H3 из клональной клетки. Поддерживается повышенное или пониженное связывание с H3 и остается стабильным после субклонирования. B) FACS-окрашивание для BCR различных селектированных субпопуляций. Повышенный или пониженный уровень связывания H3 с селектированными популяциями коррелирует с экспрессией BCR на клеточной поверхности этих популяций. Светло-серая линия: В-клетки, селектированные на сильное связывание с H3; закрашенная диаграмма: не селектированные В-клетки (родительские клетки); темно-серая линия: В-клетки, селектированные на слабое связывание H3.
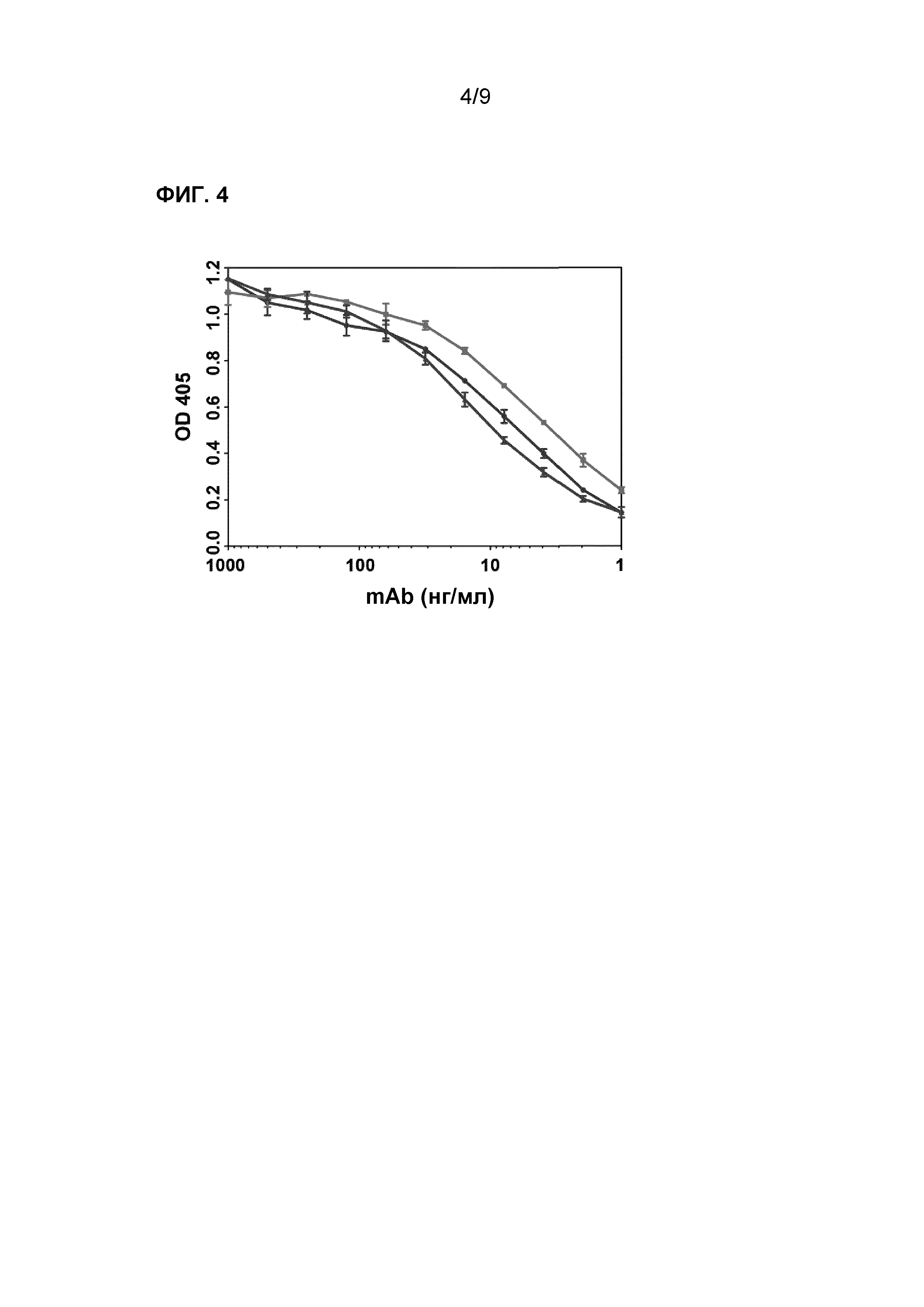
Фиг.4. H3-тИФА надосадочной жидкости культуры различных (суб)популяций. Секретированный IgG из клеток, которые селектировали на более сильное связывание с белком H3-APC, демонстрировал более сильное связывание в H3-тИФА по сравнению с IgG из не подвергнутой клеточному сортингу родительской линии. Верхняя линия: В-клетки, селектированные на сильное связывание с H3; средняя линия: не селектированные В-клетки (родительские клетки); нижняя линия: В-клетки, селектированные на слабое связывание с H3.
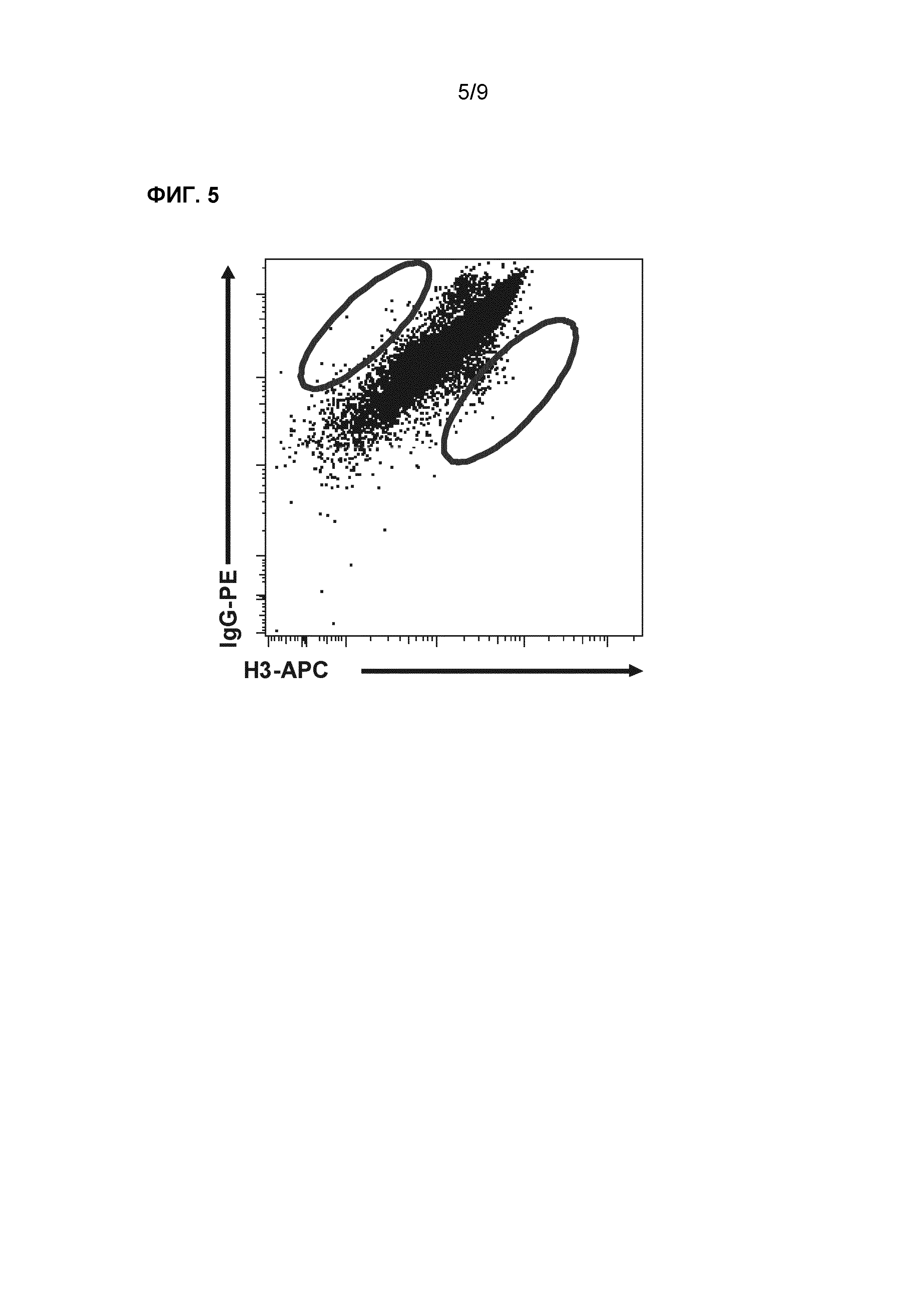
Фиг.5. Селекция субклонов с сильной и слабой аффинностью внутри H3-специфичного клона (AT10_004). Клетки окрашивали с использованием меченого с помощью Alexa-647 антигена НА H3 вместе с антителом IgG-PE. Обведенные популяции селектировали с помощью клеточного сортинга и далее размножали.
Фиг.6. FACS-анализ связывания меченого НА H3 вместе с BCR окрашиванием (как для тяжелой цепие, IgG-PE, так и для легкой цепи, Kappa-PE) с селектированными клетками через 2 недели после третьего раунда селекции на более сильное или более слабое связывание с H3 BCR и с родительскими клетками AT10_004.
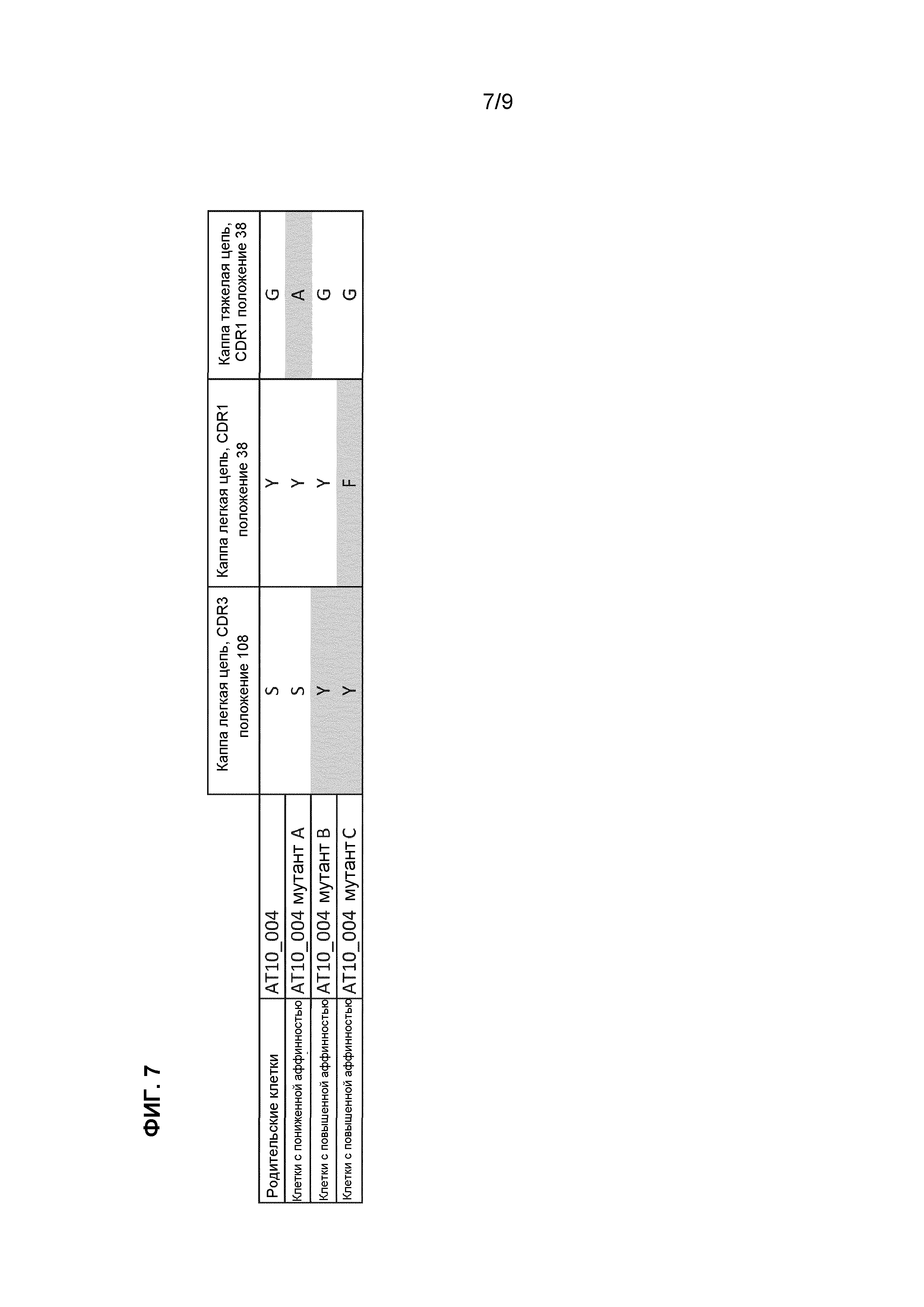
Фиг.7. Краткий обзор аминокислотных замен, которые были обнаружены в селектированных субпопуляциях с повышенной и пониженной аффинностью. Мутации в последовательности AT10_004, которые были ассоциированы с повышенным связыванием с антигеном H3, были включены в последовательность AT10_004, и эти антитела рекомбинантно продуцировали в 239T-клетках и очищали для дальнейшего анализа (AT10_004 мутант В и AT10_004 мутант C).
Фиг.8. SPR-анализ связывания AT10_004-антител с НА H3. Кривые ассоциации антител AT10_004, AT10_004 мутанта A, AT10_004 мутанта В и AT10_004 мутанта C.
Фиг.9. Средняя интенсивность флуоресценции (MFI) связывания вариантов антитела AT10_004 с H3N2-инфицированными клетками в FACS-анализе. Различные концентрации рекомбинантного AT10_004, AT10_004 мутанта B, AT10_004 мутанта С и ритаксимаб (отрицательный контроль) инкубировали с H3N2-инфицированными клетками. Связывание антитела детектировали с помощью PE-меченого козьего антитела, связывающего человеческий F(ab')2. На график наносили среднее значение и стандартную погрешность среднего для MFI от сигнала PE.
ПРИМЕРЫ
Пример 1
Получение моноклонального человеческого антитела, специфичного к гемагглютинину вируса гриппа (НА) H3
Человеческие В-клетки памяти иммортализовали с использованием технологии BCL6/Bcl-xL, описанной у Kwakkenbos et al. (Generation of stable monoclonal antibody-producing В cell receptor-positive human memory В cells by genetic programming. Nature Medicine (2010) vol. 16 (1) pp. 123-8, и в патентной заявке MEANS AND METHODS FOR INFLUENCING THE STABILITY OF ANTIBODY PRODUCING CELLS [WO 2007/067046]). Вкратце, BCL6- и Bcl-xL-трансдуцированные клетки (GFP-положительные) культивировали вместе с L-клетками, экспрессирующими лиганд CD40, и с интерлейкином (IL)-21 перед тем, как клетки, связывающиеся с НА H3, сортировали с использованием анализа клеточного сортинга с активацией флуоресценции (FACS)(Фигура 1A). Белок HA вируса гриппа (Protein Sciences) метили с помощью Alexa Fluor 647 (Molecular Probes) и инкубировали с поликлональными культивированными В-клетками. HA-положительные клетки сортировали по одной клетке на лунку и поддерживали культуру в течение 2-3 недель перед скринингом клонов на предмет связывания с HA с помощью 1) тИФА или 2) связывания с H3-инфицированными клетками (фиг.1B).
Пример 2
Селекция клонов В-клеток с более высокой и с более низкой аффинностью
Так как BCL6 Bcl-XL-трансдуцированные В-клетки экспрессируют фермент Дезаминазу, Индуцированную Активацией (AID, по генной номенклатуре AICDA), как описано у Kwakkenbos et al. (Фиг.2 левая панель и 'Generation of stable monoclonal antibody-producing В cell receptor-positive human memory В cells by genetic programming' Nature Medicine (2010) vol. 16 (1) pp. 123-8), индивидуальная В-клетка может производить нуклеотидные замены в тяжелой и легкой цепи иммуноглобулина. Эти замены могут воздействовать на аффинность связывания клонов с их антигеном. Для определения того, обладают ли субклоны НА H3-связывающего клона различным профилем связывания, H3-специфичный клон снова инкубировали с меченым антигеном НА H3. С использованием FACS, популяцию клеток с сильным связыванием с НА H3 и популяцию клеток со слабым связыванием с НА H3 сортировали (Фигура 2, правая панель) и поддерживали в культуре в течение, по меньшей мере, 13 дней перед сбором и тестированием надосадочной жидкости культуры В-клеток.
Пример 3
Отсортированные В-клетки с высокой и низкой аффинностью связывания с НА H3 стабильно экспрессируют поверхностный иммуноглобулин, но на разном уровне.
Впервые мы охарактеризовали стабильность сортированных клеток с помощью анализа способности связывания В-клеточного рецептора (BCR) с меченым НА H3 с помощью FACS (фиг.3А). Так как клетки, отсортированные как имеющие сильное связывание с НА H3, демонстрировали более высокую способность связывания, мы определили уровень экспрессии поверхностного иммуноглобулина с помощью FACS (фиг.3B). Наблюдали, что клетки, отсортированные как обладающие низкой способностью связывания с HA H3, экспрессировали меньше белка иммуноглобулина на поверхности по сравнению с клетками, отсортированными как обладающие сильным связыванием с HA. Эту более высокую или более низкую экспрессию BCR и связывание BCR с белком НА H3 поддерживали в течение периода времени, и оно становилось еще более выраженными после второго раунда сортинга (данные не показаны).
Пример 4
Аффинность по отношению к НА H3 у антител, выделенных из исходных клеток и из клеток, связывающихся с НА H3 с высокой и низкой аффинностью
Для определения аффинности связывания антител, продуцированных различными НА H3-распознающими В-клетками, анализировали с помощью тИФА надосадочную жидкость культуры после 13 дней культивирования исходных НА H3-связывающих клеток и клеток, связывающихся с НА H3 с высокой аффинностью и низкой аффинностью. НА H3 (1 мкг/мл, Protein Sciences) наносили в виде покрытия непосредственно на планшет перед тем, как лунки инкубировали с различными надосадочными жидкостями культур В-клеток. Связывание человеческих IgG с белком НА H3 детектировали с использованием поликлонального козьего антитела, связывающего человеческий белок, которое было помечено с помощью HRP. Секретированный IgG из клеток, которые селектировали на более сильное связывание с белком H3-APC, демонстрировал более сильное связывание в анализе H3-тИФА по сравнению с IgG из не подвергнутой клеточному сортингу родительской линии (фиг.4).
Пример 5
Объединенное окрашивание с BCR-антигеном для селекции клонов с высокой аффинностью и низкой аффинностью
В примерах 2 и 3 представлено, что с использованием селекции В-клеток внутри гетерогенной субпопуляции моноклонального В-клеточного клона, обладающего наибольшим уровнем связывания с H3, можно селектировать клетки, которые обладают повышенным уровнем экспрессии BCR. Таким образом, когда проводят селекцию только на основе уровня связывания с H3, то клетки, которые обладают повышенной аффинностью к антигену, но низким уровнем экспрессии BCR, могут быть исключены. Для исключения влияния уровня экспрессии иммуноглобулина на селекцию высокоаффинных клонов, осуществляют новые раунды селекции с использованием комбинации антигенного окрашивания (H3-Alexa-647) и окрашивания BCR (фиг.5). Окрашивание BCR осуществляли с использованием антител, которые связываются с тяжелой или легкой цепью BCR. Сильное окрашивание H3 и слабое окрашивание BCR указывает на высокую антигенную аффинность, тогда как слабое окрашивание H3 и сильное окрашивание BCR указывает на низкую антигенную аффинность.
НА H3-специфичный В-клеточный клон (AT10_004) культивировали в течение 2-3 недель до получения миллионов клеток перед осуществлением окрашивания BCR-антигеном. Клетки, которые демонстрировали отклонение антигенной аффинности, как положительное, так и отрицательное, селектировали и сортировали на клеточном сортере. После 3 раундов сортировки и роста, на этих клетках осуществляли анализ FACS для определения различий в антигенном связывании. Клетки, которые сортировали три раза на повышенное или пониженное антигенное связывание, демонстрировали очевидный сдвиг в антигенном связывании по сравнению с не селектированными клетками (фиг.6). Фиг.6 демонстрирует, что поддерживается повышенное или пониженное связывание H3 и остается стабильным после селекции.
Пример 6
Секвенирование BCR из селектированных клеток
Выделяли суммарную РНК с использованием мини-набора реагентов RNeasy® (Qiagen) из В-клеточных культур AT10_004 и AT10_004 мутантов, селектированных на высокую или низкую аффинность, из РНК получали кДНК, осуществляли ПЦР и анализировали последовательность тяжелой и легкой цепи BCR. Мутацию, приводящую к аминокислотной замене в положении 38 (CDRl) Глицина на Аланин в тяжелой цепи, обнаружили для клеток, которые сортировали по пониженной аффинности (далее называется мутант A). Мутации, приводящие к аминокислотным заменам в легкой цепи (по сравнению с родительской последовательностью AT10_004), обнаружили для клеток, сортированных по повышенной аффинности. Анализ последовательности продемонстрировал замену аминокислоты 108 (CDR3) в легкой цепи каппа с Серина на Тирозин (далее называется мутант B). Дополнительную мутацию в положении 38, приводящую к замене Тирозина на Фенилаланин, обнаружили в некоторых последовательностях (далее называется мутант C) (фиг.7 и таблица 1). Для получения рекомбинантного антитела AT10_004 и mAb-мутантов с повышенной аффинностью, В и С, мы клонировали вариабельные участки тяжелой и легкой цепи в одной рамке вместе с константными участками человеческого IgGl и Каппа в основной вектор pcDNA3.1 (Invitrogen) и транзиторно трансфицировали клетки 293T. Очищали рекомбинантное mAb из надосадочных жидкостей культуры с использованием АКТА (GE healthcare).
Пример 7
Анализ поверхностного плазмонного резонанса (SPR)
SPR-анализ осуществляли на системе визуализации IBIS MX96 SPR (IBIS Technologies BV., Enschede, The Netherlands), как описано в (Lokate et al., 2007, J. Am. Chem. Soc. 129:14013-140318). Вкратце, один цикл анализа SPR состоит из одной или нескольких стадий инкубирования, в которых аналиты пропускают над сенсором с покрытием с последующей стадией регенерации, в которой любой связанный аналит удаляют из сенсора. За один эксперимент может осуществляться множество циклов. Последовательные разведения (концентрации в интервале от 0,3 до 10 мкг/мл) AT10_004 и мутантного антитела AT10_004 в конденсирующем буфере (PBS + 0,03% Tween20 + 0,01 мг/мл BSA) иммобилизовали в течение 99 минут на специфичный к человеческому IgG SPR-чип гелевого типа (Ssens, Энсхеде, Нидерланды) с использованием устройства микроспоттера с непрерывным потоком (Wasatch Microfluidics, Солт-Лейк-Сити, Юта, США). После нанесения образца, сенсор промывали три раза с помощью PBS + 0,03% Tween20 (PBST).
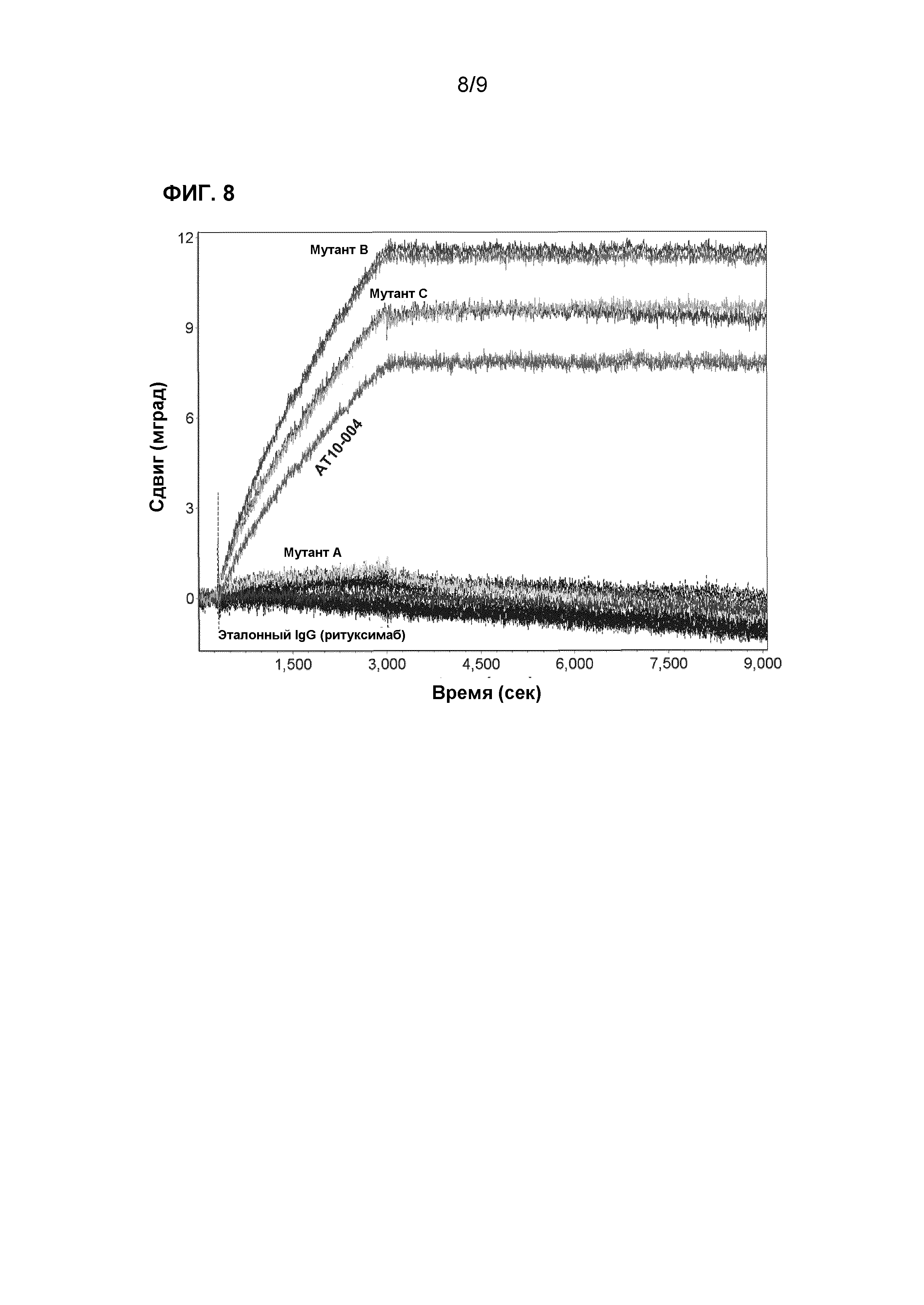
Для блокирования любых не занятых сайтов на SPR-чипе с покрытием в виде анти-IgG, на чип сначала инъецировали неспецифический человеческий IgG (ритуксимаб, 10 мкг/мл в PBST) и инкубировали в течение 45 минут с последующим инкубированием в течение 100 минут вместе с PBST. После этой стадии блокировки проводили два цикла с пустыми инъекциями, каждый состоял из 45-минутных инъекций с пустым аналитическим буфером (1x PBST + 0,01% BSA) с последующим инкубированием в течение 100 минут вместе с PBST. Затем, на сенсор инъецировали 1 мкг/мл рекомбинантного белка НА3 вируса гриппа (из H3N2, Вайоминг, 03/2003, Sino Biological inc., Пекин, Китай) в аналитическом буфере и инкубировали в течение 45 минут. Затем, сенсор промывали с помощью PBST и инкубировали в течение 100 минут (для измерения диссоциации комплекса). Полученные данные анализировали с использованием программного обеспечения Sprint (версия 1.6.8.0, IBIS Technologies BV., Энсхеде, Нидерланды). Константы связывания подбирали с использованием программы Scrubber2 (Biologic Software, Кэмпбелл, Австралия). Фиг.8 демонстрирует, что рекомбинантный HA3 не ассоциирован с AT10_004 мутантом A. Повышенная степень ассоциации HA3 для AT10_004 мутанта В и С, по-видимому, сравнима с не мутированным AT10_004. Константы связывания, полученные для AT10_004 и для каждого мутанта, представлены в таблице 2.
Пример 8
Связывание антитела с клетками, инфицированными вирусом
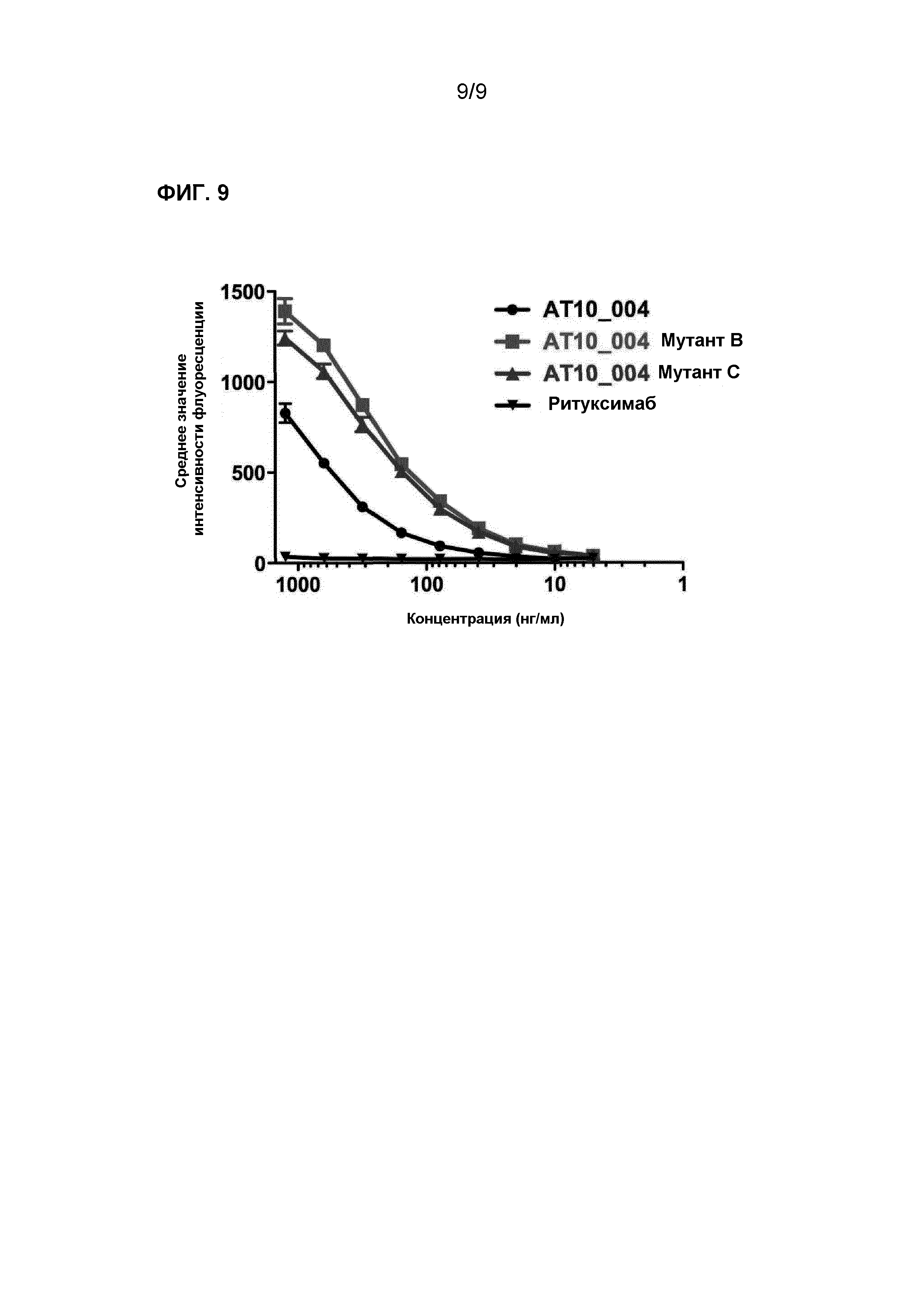
Для тестирования способности связывания AT10_004 и AT10_004-мутантов с клетками, инфицированными вирусом, осуществляли анализ FACS на клетках, инфицированных вирусом гриппа H3N2 (A/Нидерланды/177/2008). Клетки MDCK-SIAT растили в культуральных флаконах T175 до достижения плотности 80-100% в среде DMEM/FCS/PS/G418. Слой клеток промывали 2x с использованием 10 мл PBS, после чего добавляли 15 мл Optimem/PS/G418/триписин. Затем добавляли во флакон 0,5 мл 100,000 TCID50 вируса гриппа, и клетки культивировали при 37°C. Через 24-48 ч клетки промывали 2x с использованием 10 мл PBS и открепляли от пластика с использованием раствора трипсин-EDTA. Клетки считали и замораживали при -150°C до момента применения. Инфицированные клетки размораживали и инкубировали вместе с антителами AT10_004 (мутантными) или с Ритуксимаб (в качестве отрицательного контроля) при нескольких концентрациях в течение 30 минут при 4°C и затем промывали 2x с использованием 150 мкл PBS/2% FCS. Связывание антитела детектировали с использованием антител к человеческому IgG-PE (Southern Biotech) и анализировали на Guava easyCyte 8HT, Millipore. AT10_004 мутанты В и С, оба демонстрировали повышенную интенсивность окрашивания на клетках, инфицированных H3N2, по сравнению с родительским антителом AT10_004 (фиг.9).


Константы подбирали в Scrubber2 с использованием правил от общего подбора до модели взаимодействия 1:1.
Реферат
Изобретение относится к биохимии. Раскрыты in vitro способы получения высокоаффинных антител и антител с низкой аффинностью. Способы включают селекцию В-клетки, способной продуцировать антитело, специфичное к интересующему антигену, или селекцию В-клетки, способной дифференцироваться в указанную В-клетку; предоставление В-клетке нуклеиновой кислоты, кодирующей BCL6; предоставление В-клетке анти-апоптотической нуклеиновой кислоты; культивирование В-клетки до получения первой популяции В-клеток; осуществление антигенного окрашивания и BCR окрашивания В-клеток из первой популяции; селекцию В-клетки, которая обладает высокой или низкой антигенной аффинностью; обеспечение размножения В-клетки до получения второй популяции В-клеток; измерение скорости антигенной ассоциации и скорости диссоциации антител В-клеткой из указанной второй популяции; и селекцию антитела с более высокой или низкой аффинностью для интересующего антигена, чем средняя аффинность первой популяции В-клеток для интересующего антигена. Изобретение представляет улучшенный способ скрининга для выбора антител с высокой или низкой активностью. 2 н. и 9 з.п. ф-лы, 9 ил., 2 табл., 8 пр.
Формула
a) селекцию В-клетки, способной продуцировать антитело, специфичное к указанному интересующему антигену, или селекцию В-клетки, способной дифференцироваться в В-клетку, которая способна продуцировать антитело, специфичное к указанному интересующему антигену, используя анализ связывания;
b) предоставление указанной В-клетке нуклеиновой кислоты, кодирующей BCL6;
c) предоставление указанной В-клетке анти-апоптотической нуклеиновой кислоты;
d) культивирование указанной по меньшей мере одной В-клетки до получения первой популяции В-клеток;
e) осуществление антигенного окрашивания и BCR окрашивания В-клеток из указанной первой популяции;
f) селекцию В-клетки, которая обладает высокой антигенной аффинностью;
g) обеспечение размножения указанной В-клетки до получения второй популяции В-клеток;
h) измерение скорости антигенной ассоциации и скорости диссоциации антител, продуцируемых по меньшей мере одной В-клеткой из указанной второй популяции; и
i) селекцию антитела с более высокой аффинностью для указанного интересующего антигена, чем средняя аффинность указанной первой популяции В-клеток для указанного интересующего антигена.
a) селекцию В-клетки, способной продуцировать антитело, специфичное к указанному интересующему антигену, или селекцию В-клетки, способной дифференцироваться в В-клетку, которая способна продуцировать антитело, специфичное к указанному интересующему антигену, используя анализ связывания;
b) предоставление указанной В-клетке нуклеиновой кислоты, кодирующей BCL6;
c) предоставление указанной В-клетке анти-апоптотической нуклеиновой кислоты;
d) культивирование указанной по меньшей мере одной В-клетки до получения первой популяции В-клеток;
e) осуществление антигенного окрашивания и BCR окрашивания В-клеток из указанной первой популяции;
f) селекцию В-клетки, которая обладает низкой антигенной аффинностью;
g) обеспечение размножения указанной выбранной В-клетки до получения второй популяции В-клеток;
h) измерение скорости антигенной ассоциации и скорости диссоциации антител, продуцируемых по меньшей мере одной В-клеткой из указанной второй популяции; и
i) селекцию антитела с более низкой аффинностью для указанного интересующего антигена, чем средняя аффинность указанной первой популяции В-клеток для указанного интересующего антигена.
Комментарии