Штамм коровьего вируса герпеса типа 1 (внv-1) сncm №1-1213, вакцина, диагностический серологический реактив, способ вакцинации, способ серологического отличия - RU2170254C2
Код документа: RU2170254C2
Чертежи




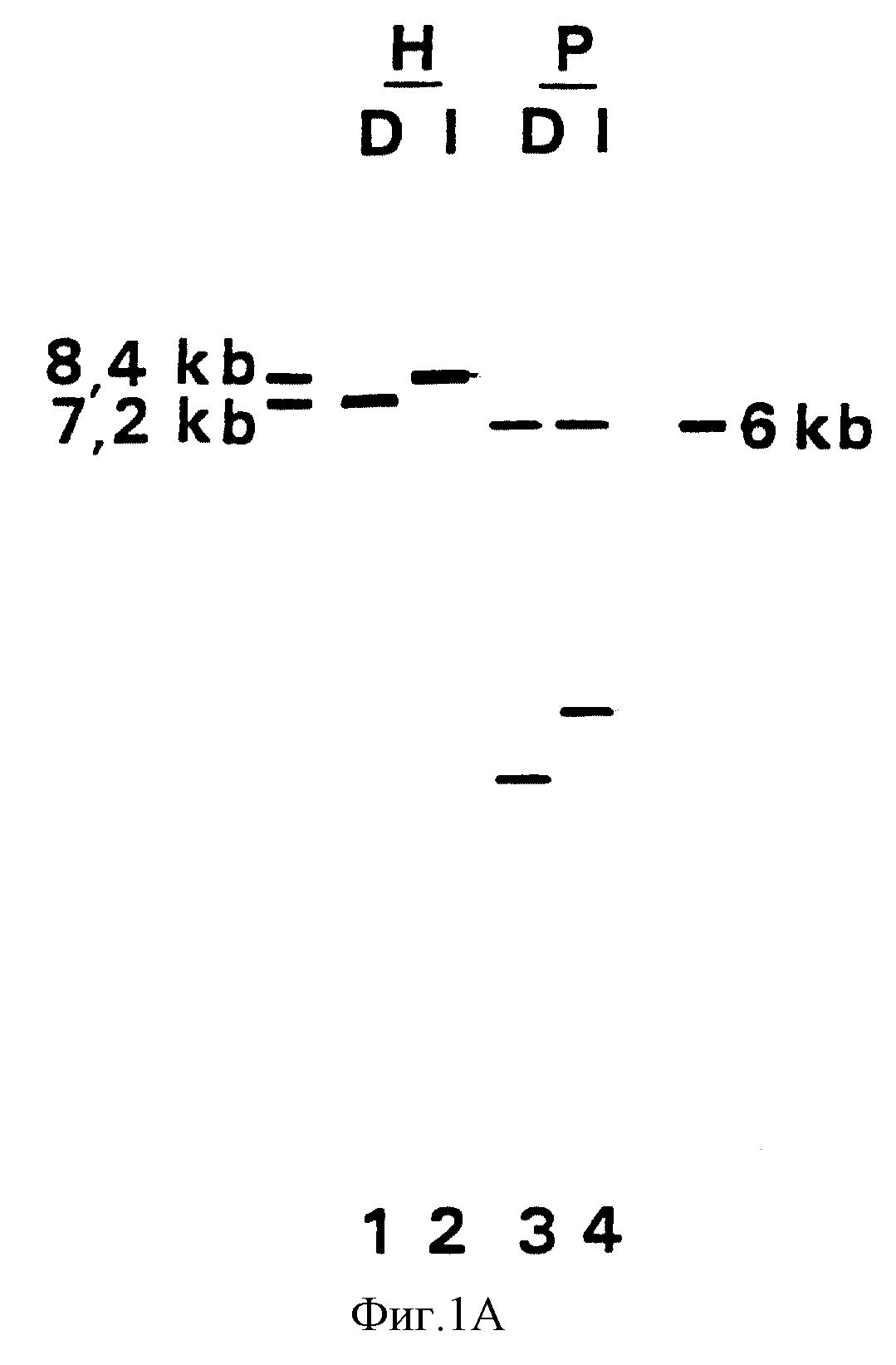



























Описание
Область изобретения
Изобретение относится к областям вакцинации и диагностики в связи с заболеваниями, которые вызываются патогенными бактериями, и включает использование как классических
способов получения живой ослабленной вакцины или инактивированной вакцины, так и современных способов, основанных на ДНК рекомбинантной технологии.
Более конкретно, изобретение относится к живым ослабленным вакцинам и инактивированным вакцинам для защиты животных, особенно крупного рогатого скота, против вируса герпеса типа 1 у коров /BHV-1/, причем эти вакцины таковы, что они не только безопасны и эффективны, но также дают возможность отличить зараженных от незараженных животных в вакцинированной популяции.
Диагностические комплексы и процедуры, которые можно использовать для такого теста на различение зараженных и незараженных животных в вакцинированной популяции, являются также аспектом настоящего изобретения.
Предпосылки
изобретения
BHV-1, включая инфекционный вирус ринотрахеита /IBPV/ и инфекционный пустулезный вирус вульвовагинита /IPVV/, играет важную роль в развитии респираторных заболеваний и нарушении
плодовитости у коров. После острого заражения BH-1 часто остается у носителя в латентной форме. Латентный вирус может вновь активизироваться под влиянием стресса - что может сопровождаться
клиническими проявлениями -, а затем вывестись из организма. Как следствие, зараженный скот можно рассматривать как потенциальный разносчик BHV-1. BHV-1 часто приобретает эндемический характер
приблизительно в 75% голландских ферм. Особенно серологически позитивны к нему старые животные.
Существует ряд инактивированных /"убитых"/ вакцин и множество ослабленных /"живых"/ вакцин для прививки против заражений BHV-1. Инактивированные вакцины готовятся умерщвлением вируса BHV-1, например, теплообработкой, облучением или обработкой этанолом или формалином. Однако это не всегда обеспечивает достаточную защиту. Ослабленные вакцины готовятся большим числом пересевов на гомологичные /коровьи/ или на гетерологичные клетки, такие как клетки свиньи или собаки, и иногда потом вирусы также обрабатываются физически или химически. Таким образом, в геноме вируса развиваются неизвестные мутации/делеции, которые часто снижают свойства вируса, продуцирующие заболевание. Ослабленные живые вакцины дают лучшую защиту, чем инактивированные вакцины, поскольку они дают больше вирусных антигенов иммунной системе хозяина. Другим важным преимуществом живых вакцин является то, что их можно назначать через нос, т.е. на участке, где происходит первое размножение вируса дикого типа после заражения. И все же живые вакцины нуждаются в усовершенствовании. Некоторые живые вакцины все еще обладают абортогенной способностью, что проявляется особенно после внутримышечного назначения. Кроме того, вероятно, все живые вакцины присутствуют в вакцинированной корове в латентном состоянии. И существует вероятность, что, если вакцина не сильно отличается от вируса дикого типа, возможен возврат к вирулентности. Но одна из основных проблем заключается в том, что вакцины BHV-1 не могут помешать заражению вирусами дикого типа. В результате вакцинированный скот может также разносить BHV-1 дикого типа.
Для хорошей программы контроля BHV-1 необходимо иметь эффективную и безопасную вакцину, которую можно отличить от вируса дикого типа, поскольку применение эффективной вакцины может снизить значительно циркуляцию BHV-1, а тест, который может различить вакцину и вирус дикого типа, делает возможным обнаружение /и затем удаление/ инфицированного скота в вакцинированной популяции.
Были разработаны BHV-1 вакцины, которые представлены как более безопасные, чем традиционные вакцины, и отличительные от вируса дикого типа. Был изолирован мутант с делецией тимидинкиназы, который обладает меньшей абортогенностью, становится латентным не так часто и не может реактивироваться. Кроме того, с использованием техники рекомбинантной ДНК была создана BHV-1 вакцина, которая имела делецию в гене для гликопротеина gIII, что делает эту вакцину отличаемой от BHV-1 дикого типа посредством серологических приемов. Однако все же существуют возражения против этих вакцин. С одной стороны, ген тимидинкиназы вовлечен в вирусную репликацию, а снижение репликации ведет к снижению защиты. С другой стороны, гликопротеин gIII важен для выработки защитных антител, что делает менее эффективной вакцину с делецией gIII. Практическая проблема заключается в том, что внутриносовое введение, которое дает наилучшую защиту в случае рекомбинантных вакцин, не разрешено в некоторых странах. Соответственно, существует необходимость создания вакцины, которая была бы столь же безопасна, как и эффективна, и, кроме того, отличима от BHV-1 дикого типа, еще также желательно, чтобы, по меньшей мере, одна из таких вакцин была основана на вирусе, ослабленном с помощью традиционной техники, нежели вирусе, созданном техникой рекомбинантной ДНК.
Теперь, с помощью пересевов в клеточных культурах был получен штамм BHV-1, в котором отсутствует ген гликопротеина gE. Первые результаты наших исследований показывают, что этот ген очень полезен в серологическом разграничении с BHV-1 дикого типа и что он участвует в выражении вирулентности. Поэтому его делеция способствует безопасности и может сделать использование делеций тимидинкиназы ненужным. Гликопротеин gE представляется менее важным для индуцирования защиты, чем гликопротеин gIII. Ослабленный традиционным способом штамм BHV-1, который можно серологически отличить от вируса дикого типа, уникален. Расположение и последовательность ДНК гена gE, описанные здесь в первый раз, не были ранее известны. Не были также известны ни олигонуклеотиды, ни полипептиды и олигопептиды, которые можно получить от них. Проба на серологическое различение на основе gE гена также уникальна.
Важное преимущество этого "традиционного" мутанта с делецией gE /"традиционным" считается использование традиционного метода для изоляции ослабленного вируса/ заключается в том, что его внутриносовое назначение допускается в странах, где это запрещено в отношении рекомбинантных вакцин. Однако с учетом различных мнений по безопасности, кроме этой традиционной вакцины с делецией gE, были также созданы конкретные рекомбинантные варианты. Эти рекомбинантные вакцины также имели делению gE и могут иметь или не иметь делецию гена тимидинкиназы, и также могут использоваться как векторы для экспрессии гетерологичных векторов. Все эти рекомбинантные вакцины можно отличить от вируса дикого типа с помощью того же gE-специфичного теста. Использование стандартного теста для ряда разных вакцин может быть большим преимуществом в борьбе с BHV-1 в международном масштабе. Такой подход не был ранее описан в области BHV-1 вакцин.
Серологический анализ ответной реакции на BHV-1 у скота показал, что важная фракция анти-gE антител направлена против комплекса, сформированного гликопротеином gE и другим гликопротеином BHV-1 - гликопротеином gI. Поэтому серологические тесты, которые могут /также/ демонстрировать присутствие таких комплекс-специфических антител, более чувствительны, чем тесты, которые могут только обнаруживать анти-gE антитела. Скот, вакцинированный одним мутантом с делецией gE, может вырабатывать анти-gI антитела, которые могут помешать обнаружению анти-gI/gE антител. Следовательно, это изобретение также включает вакцину с двойной делецией gI/gE.
Содержание изобретения
Во-первых, это изобретение предусматривает мутант с делецией BHV-1, который имеет делецию в
gE-гене гликопротеина. Слова "деления в" охватывают делецию гена в целом.
Предпочтительный вариант изобретения предполагает мутант BHV-1 с делецией, который имеет делецию в gE-гене гликопротеина, что вызвано процедурой ослабления, такой, как мутант с делецией Difivac-1, описанной ниже.
Другие предпочтительные варианты изобретения заключаются в мутанте с делецией BHV-1, включающем делецию в gE-гене гликопротеина, который был создан технологией рекомбинантной ДНК, такой, как мутанты с делецией IB7 или IB8, описанные ниже.
Другой предпочтительный вариант изобретения состоит из мутанта BHV-1 с двойной делецией, включающего делецию в gE-гене гликопротеина и делецию в gI-гене гликопротеина, такого, как мутант с двойной делецией gI/gE Difivac-1E, описанный ниже.
Кроме того, учитывая максимальную безопасность, в соответствии с изобретением, предпочтителен мутант с делецией BHV-1, который имеет делецию в gE-гене гликопротеина и делецию в гене тимидинкиназы. В изобретение включен также мутант с делецией BHV-1, который имеет делецию в gE-гене гликопротеина, gI-гене гликопротеина, и гене тимидинкиназы.
Изобретение предусматривает композицию вакцины для вакцинации животных, в частности млекопитающих, конкретнее коров, чтобы защитить их против BHV-1, включающую мутант с делецией BHV-1, как описано выше, и подходящий носитель или стимулятор. Указанная композиция может быть композицией живой или инактивированной вакцины.
Далее изобретение предусматривает мутант BHV-1, который имеет делецию в gE-гене гликопротеина и содержит гетерологичный ген, введенный технологией рекомбинантной ДНК. Предпочтительно это касается мутанта BHV-1, который содержит гетерологичный ген, введенный технологией рекомбинантной ДНК, в местоположение gE-гена гликопротеина, и этот гетерологичный ген находится под контролем регуляторных последовательностей gE-гена и может быть связан с частью gE-гена, которая кодирует сигнальный пептид. Такой гетерологичный ген может также быть под контролем другого промотора BHV-1 или под контролем гетерологичного промотора. Если мутант BHV-1 имеет другие делеции в дополнение к делеции в gE-гена гликопротеина, такие, как делеция в гене тимидинкиназы и/или делеция в gI-гене гликопротеина, указанный гетерологичный ген можно также ввести в местоположение этой дополнительной делеции /й/. Множественные вставки - еще одна возможность либо в месте в местоположении одной делеции, либо распределенные по местоположениям нескольких делеции.
Введенный гетерологичный ген предпочтительно кодирует для иммунногенного белка или пептида другого патогена или для цитокина, который стимулирует иммунную реакцию. Примерами подходящих цитокинов являются интерлейкин 2, интерферон-альфа и интерферон-гамма.
Изобретение также предусматривает /живую или инактивированную/ композицию вакцины для вакцинации животных, в частности млекопитающих, конкретнее коров, чтобы защитить их против /различных/ патогенных бактерий, включающую мутант BHV-1, имеющий в себе кодирование гетерологичного гена для иммунногенного белка или пептида этого другого патогена, и подходящий носитель стимулятора. Конечно, защита может относиться более чем к одному патогену, т.е. это многовалентная вакцина, в которой мутант содержит множество гетерологичных генов.
Далее изобретение относится к композиции, включающей рекомбинантную нуклеиновую кислоту, содержащую gE-ген гликопротеина BHV-1, часть этого gE-гена гликопротеина или нуклеотидную последовательность, производную от этого gE-гена гликопротеина. Эта композиция может содержать вектор для клонирования или экспрессии, имеющий в себе вставку рекомбинантной нуклеиновой кислоты, которая включает gE-ген гликопротеина BHV-1, часть этого gE-гена гликопротеина или нуклеотидную последовательность, производную от этого gE-гена гликопротеина.
Изобретение также включает композицию, содержащую гликопротеин gE BHV-1, часть этого гликопротеина gE, пептид, производный от этого гликопротеина gE, или комплекс гликопротеинов gE и gI, и композицию, содержащую антитело, которое специфично для гликопротеина gE BHV-1, части этого гликопротеина gE, пептида, производного от этого гликопротеина gE, или комплекса гликопротеинов gE и gI. "Антитело" означает как препарат поликлонального антитела, так и моноклонального антитела, предпочитаемый для большинства применений. Под терминами "часть гликопротеина gE" и "пептид, производный от гликопротеина gE" понимается обозначение последовательностей gE-специфической аминокислоты, которые обычно имеют длину, по меньшей мере, около 8 аминокислот.
Далее изобретение относится к диагностическим комплексам и процедурам для обнаружения нуклеиновой кислоты BHV-1 в образце, биологическом образце, таком как кровь или сыворотка крови, кровяные клетки, молоко, жидкости тела, такие как слезы, промывающая жидкость легких, носовая жидкость, сперма, в частности семенная жидкость, слюна, мокрота или ткань, в частности нервная ткань, животного, в частности млекопитающего, конкретно коровы, включающим зонд или праймер нуклеиновой кислоты, имеющей нуклеотидную последовательность, полученную от gE-гена гликопротеинa BHV-1, и средства обнаружения, пригодные для реакции на обнаружение нуклеиновой кислоты.
Далее, изобретение относится к диагностическим комплексам и процедурам для обнаружения антител, которые специфичны для BHV-1 в образце, в частности биологическом образце, таком как кровь или сыворотка крови, слюна, мокрота, жидкости тела, такие как слезы, промывающая жидкость легких, носовая жидкость, молоко, или ткань животного, конкретно коровы, включающим гликопротеин gE BHV-1, часть этого гликопротеина gE, пептид, производный от этого гликопротеина gE, или комплекс гликопротеинов gE и gI, и средства обнаружения, пригодные для реакции на обнаружение антитела. Такие диагностические процедуры могут также содержать одно или несколько антител, которые специфичны для гликопротеина gE BHV-1 или специфичны для комплекса гликопротеинов gE и gI BHV-1.
Изобретение также относится к диагностическим процедурам для обнаружения белка BHV-1 в образце, в частности биологическом образце, таком как кровь или сыворотка крови, клетки крови, молоко, жидкости тела, такие как слезы, промывающая жидкость легких, носовая жидкость, сперма, в частности семенная жидкость, слюна, мокрота или ткань, в частности нервная ткань, животного, млекопитающего, конкретнее коровы, включающим одно или несколько антител, которые специфичны для гликопротеина gE BHV-1 или специфичны для комплекса гликопротеинов gE и gI BHV-1, и средства обнаружения, пригодные для анализа на обнаружение белка.
Изобретение далее предусматривает способ определения заражения BHV-1 животного, в частности млекопитающего, конкретно коровы, включающий исследование образца от животного, в частности биологического образца, такого как кровь, или сыворотка крови, клетки крови, сперма, в частности семенная жидкость, слюна, жидкости тела, такие как слезы, промывающая жидкость легких, носовая жидкость, молоко, или ткань, в частности нервная ткань, на присутствие нуклеиновой кислоты, содержащей gE-ген гликопротеина BHV-1, или присутствие гликопротеид gE BHV-1, или присутствие антител, которые специфичны для гликопротеин gE BHV-1 или специфичны для комплекса гликопротеинов gE и gI BHV-1. Образец для исследования может быть взят у животного, которое ранее не вакцинировалось композицией вакцины по изобретению или у животного, которое ранее вакцинировалось препаратом вакцины по изобретению.
Подробное описание
изобретения
Изобретение относится к ряду BHV-1 вакцин как живых, так и инактивированных, у которых общее то, что в них полностью или частично отсутствует гликопротеин gE ген. Эта серия
включает как естественный мутант с gE делецией, так и созданные мутанты с делецией gE, которые могут включать или не включать также делецию гена тимидинкиназы и/или гена gI гликопротеина, и созданные
мутанты с делецией gE, которые используются как векторы для гетерологичных генов. Изобретение далее относится к нуклеотидным последовательностям, кодирующим гликопротеид gE-ген BHV-1, олигонуклеотидам,
производным от этих последовательностей, самому гликопротеину gE, пептидам, которые получены от них, и /моноклональным или поликлональным/ антителам, которые направлены против gE гликопротеина и
пептидов, полученных от них. Изобретение также относится к комплексам гликопротеинов gE и gI BHV-1 и к антителам, направленным против таких комплексов.
Эти материалы в соответствии с
изобретением можно использовать для:
1/ вакцинации скота против заболеваний, вызванных BHV-1, так что можно различить зараженных BHV-1 животных и вакцинированных животных, наряду с
традиционной можно использовать созданную вакцину;
2/ вакцинации скота против как заболеваний BHV-1, так и заболеваний, вызванных другими патогенными бактериями, кодирующие последовательности
которых для защитных антигенов можно ввести в мутанты с делецией BHV-1;
3/ исследования крови, сыворотки, молока или других жидкостей тела скота на серологическое определение или посредством
техники обнаружения нуклеиновой кислоты /например, ЦРП (ЦРП - цепная реакция полимеразы)/, были ли животные заражены BHV-1 дикого типа или были вакцинированы мутантом с делецией gE.
Синтез олигопептидов, полипептидов и гликопротеинов, полученных от кодирующей последовательности гликопротеин gE-гена и гликопротеин gI-гена BHV-1.
Результаты анализа последовательности ДНК, описанного в примерах, гликопротеин gE-гена /фиг. 3А/ и изолированных фрагментов ДНК, которые кодируют для этого гена, делают возможным, используя стандартные молекулярно-биологические процедуры, как синтезировать пептиды gE белка /олиго- или полипептиды/, так и выразить gE белок полностью или большими частями через прокариотный путь /в бактериях/ или через эукариотный путь /например, в мышиных клетках/. Через эти пути можно получить gE-специфический антиген, который может, например, служить для выработки gE-специфических моноклональных антител /MAT/. Кроме того, и gE-специфический антиген /и gE-специфические MAT/ можно использовать в серологических тестах, чтобы сделать различие между животными, вакцинированными вакциной BHV-1 делеции gE и животными, зараженными вирусом BHV-1 дикого типа.
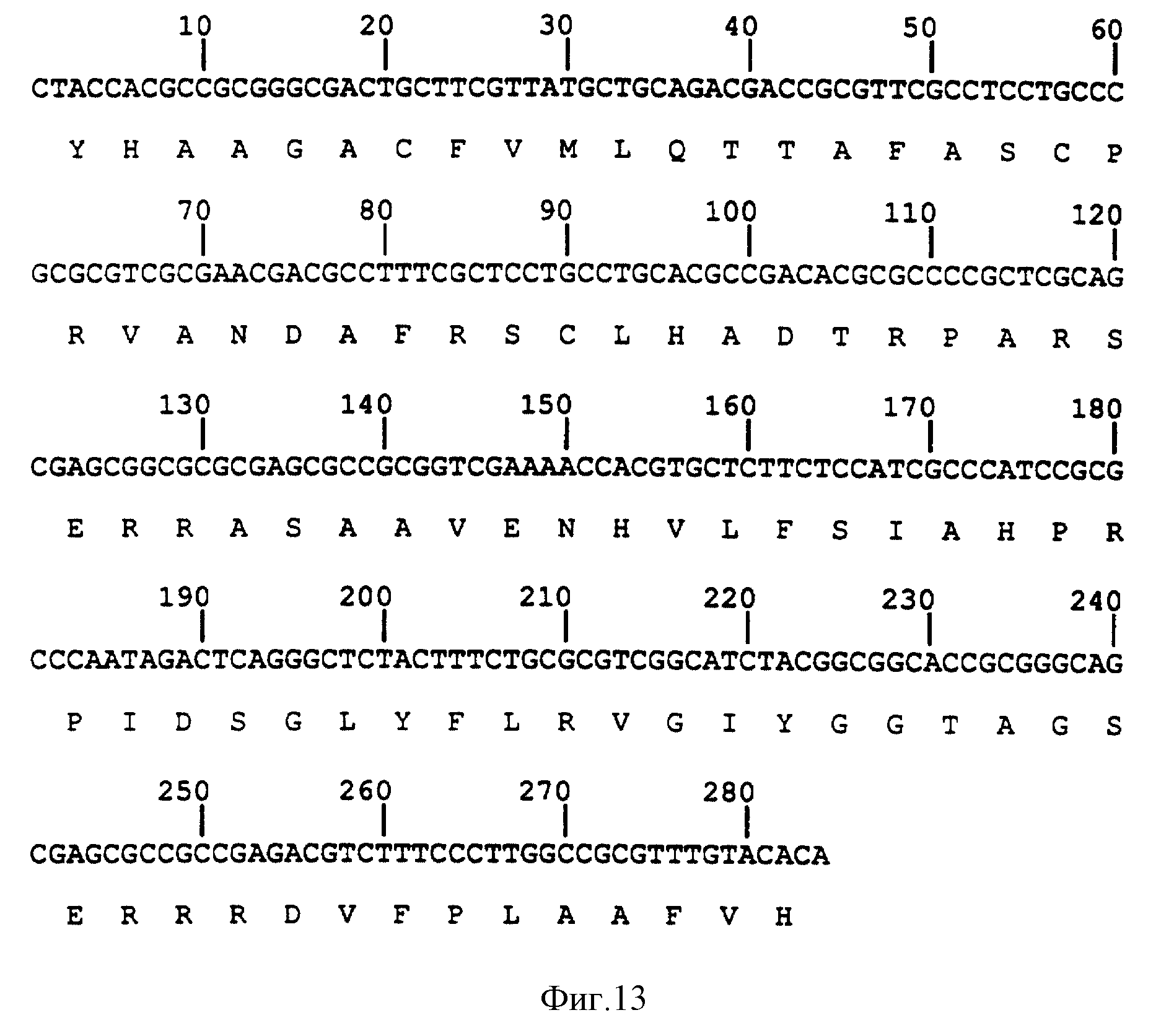
Результаты частичного анализа последовательности ДНК гликопротеина gI гена - описанного в примерах - и изолированные фрагменты ДНК, которые кодируют для этого гена, вместе с эукариотными клетками, выражающими гликопротеин gE, позволяют экспрессию gI/gE комплекса в эукариотных клетках /См. фиг. 13 и 14/. Этот гликопротеиновый комплекс может быть использован, чтобы получить gI/gE специфические моноклональные антитела. gI/gE комплекс можно также использовать как антиген в серологических пробах для дифференциации между животными, вакцинированными одним gE BHV-1 мутантом с делецией или двойным gI/gE BHV-1 мутантом с делецией, и животными, зараженными вирусом BHV-1 дикого типа.
gE-специфические
пептиды
На основе известной кодирующей последовательности белка с помощью автоматического синтезатора можно получить полипептиды не менее чем 40-50 аминокислот. Теперь, когда выявлена
кодирующая последовательность белка gE гликопротеина штамма Lam BHV-1 /фиг. 3A/, можно синтезировать полипептиды этого BHV-1 gE гликопротеина. Такими полипептидами по стандартным методам можно
иммунизировать экспериментальных животных, таких как мышей или кроликов, чтобы выработать gE-специфические антитела. Кроме того, используя эти gE-специфические пептиды, можно также определить
местоположения, где анти-gE антитела реагируют с gE белком /эпитопы/, например, способом PEPSCAN /Geysen и др. , 1984, Proc. Natl, Acad. Sci, США 81, 3998-4002/. gE-специфические олигопептиды можно
также использовать в серологических реакциях, которые демонстрируют анти-gE антитела.
Прокариотная экспрессия gE
Для синтеза gE белка в бактериях /т.е. прокариотной экспрессии
gE/, фрагменты ДНК, которые кодируют для гликопротеина gE или для его частей, должны клонироваться в векторы прокариотной экспрессии. Векторы прокариотной экспрессии представляют собой циркулярные
молекулы ДНК, которые могут сохранять себя в бактерии как отдельная воспроизводящая молекула /плазмида/. Эти векторы экспрессии содержат один или несколько маркерных генов, которые кодируют для
устойчивости к антибиотику, и таким образом позволяют выбор для бактерий с вектором экспрессии. Кроме того, векторы экспрессии включают /часто контролируемый/ участок промотора, за которым фрагменты
ДНК можно лигировать и которые затем выражаются под влиянием промотора. Во многих настоящих векторах прокариотной экспрессии нужный белок выражен в состоянии слияния с так называемым белком-носителем.
В векторе за промотором располагается кодирующая последовательность для белка-носителя, непосредственно к которому можно лигировать нужный фрагмент ДНК. Белки слияния часто более стабильны и/или их
легче распознать и/или изолировать. Уровень состояния устойчивости, которого может достичь определенный слитый белок в определенном бактериальном штамме, зависит от вида слияния и от штамма. Обычно
пробуют различные комбинации.
Эукариотной экспрессии гликопротеин gE-гена
Хотя прокариотная экспрессия белков предлагает некоторые преимущества, белки не имеют модификаций,
таких как гликосилация и других, которые имеют место в эукариотных клетках. В результате этого эукариотически выраженный белок часто является более стабильным антигеном. Для гетерологичной экспрессии
белков в эукариотных клетках, таких как мышиные клетки, используются векторы эукариотной экспрессии. Этими векторами являются плазмиды, которые не только могут размножаться в клетках E.coli, но также
стабильно существуют в эукариотных клетках. В дополнение к прокариотному селективному маркеру они также включают эукариотный селективный маркер. Аналогично векторам прокариотной экспрессии векторы
эукариотной экспрессии содержат участок промотора, за которым можно лигировать желаемые гены. Однако последовательности промотора в эукариотных векторах специфичны для эукариотных клеток. Более того,
в эукариотных векторах слияние с белками-носителями используется очень редко. Эти векторы вводятся в эукариотные клетки посредством стандартного способа трансфекции /Graham и A.I.van der Eb, 1973,
Virology, 52, 456-467/. Кроме эукариотных плазмидных векторов имеются также вирусные векторы, где гетерологичный ген вводится в геном вирусе /напр., ретровирусы, вирусы герпеса и коровьей оспы/.
Следовательно, эукариотные клетки можно заразить рекомбинантными вирусами.
В общем, нельзя предсказать какой вектор и тип клетки наиболее приемлемы для определенного продукта гена. По большей части пробуются несколько комбинаций.
Эукариотная экспрессия и гликопротеина gE и гликопротеина gI
Конечная структура, которой достигает белок, зависит от
последовательности его основной аминокислоты, его укладки, его посттрансляционной модификации и т. д. Важный фактор, от которого зависит структура белка, - его взаимосвязь с одним или несколькими
другими белками. Мы выявили, что BHV-1 гликопротеин gE образует комплекс, по меньшей мере, с одним другим гликопротеином: BHV-1 гликопротеином gI. Первое указание на такой комплекс появилось из наших
результатов с кандидатом анти-gE MAT 1, 51, 67, 75 и 78 /См. таблицу 2/. Эти MAT не реагировали ни с Difivac-1, ни с Lamg E-, а также не опознавали гликопротеин gE-экспрессирующие ЗТЗ
клетки. Однако эти MAT не реагировали с gE-экспрессирующими ЗТЗ клетками после заражения Difivac-1, показывая, что нужны комплементарные факторы, чтобы дать гликопротеину gE нужную антигенную
конформацию для этих МАТ. В некоторых из наших экспериментах по радио-иммунопреципитату с MAT 81 мы выявили копреципитацию белка с молекулярным весом в 63 кД. Ввиду того, что гликопротеин gE вируса
простого герпеса образует комплекс с белком со сравнительным молекулярным весом /HSVI гликопротеин gI/, мы сделали вывод, что BH-1 гликопротеин gE образует комплекс с BHV-1 гомологом гликопротеина gI.
Чтобы изучить этот BHV-1 gE/gI комплекс и получить gE антиген с должной антигенной структурой, мы экспрессировали оба гликопротеина в одной эукариотной клетке. Для этого мы применили те же процедуры,
что описаны для эукариотной экспрессии одного гликопротеина gE. Единственным дополнительным условием было использование векторов экспрессии с различными эукариотными селектируемыми маркерами.
Серологические реакции
Серологические методы для различения скота, вакцинированного Defivac-1 скота, зараженного BH-1 дикого типа на основе антител против gE, предпочтительно
основываются на использовании моноклональных антител, направленных против gE.
Они используются следующим образом:
a/ В соответствии с принципом, описанным Van Oirschot и др.
/Jorn. of Virological Methods, 22, 191-206, 1988/. В этом ELISA (ELISA - блокирующий ферментный иммуносорбентный тест) для обнаружения gI антител против вируса болезни Aujeszky's, антитела проявляются
своим блокирующим действием на реакцию двух MAT, имеющих два разных эпитопа на gI. Тест или реакция проводится следующим образом. Микротитрационные планшеты покрываются MAT 1 в течение ночи при 37oC, после чего они стоят, например, при 4oC или -20oC. Исследуемая сыворотка преинкубируется антигеном в отдельных непокрытых микротитрационных планшетах, например, 2 часа
при 37oC. Планшеты, покрытые МАТ-1, промываются, скажем, 5 раз, после чего на эти планшеты добавляется MAT 2, связанное с пероксидазой хрена обыкновенного /HRPO/. Затем преинкубированные
смеси сыворотка-антиген переносятся на планшеты, в которых находятся два MAT, с последующей инкубацией, например, в течение 1 часа при 37oC. Планшеты промываются и субстрат добавляется к
каждой лунке. После приблизительно 2 часов при комнатной температуре планшеты считываются спектрофотометрически. На каждом планшете имеются четыре отрицательные контрольные сыворотки и четыре серийных
разведения положительной сыворотки. Сыворотка с величиной оптической плотности /ОП/ менее 50% средней величины ОП 4 отрицательных контрольных сывороток, которые исследовались на том же планшете,
считается положительной.
b/ В соответствии с принципом бутерброда непрямого двойного антитела /IDAS/. Здесь микротитровальные планшеты покрываются MAT или поликлональной сывороткой, направленной против gE белка. Инкубация препаратом gE-антигена приводит к тому, что gE связывается с покрытием. Антитела, специфически направленные против gE в исследуемой коровьей сыворотке, последовательно связываются с gE. Эти связанные антитела распознаются антикоровьим иммуноглобулиновым конъюгатом. Антитела в этом конъюгате ковалентно связаны с ферментом пероксидазы. Наконец, связанный конъюгат виден при добавлении хромогенного субстрата. Специфичность реакции проверяется проведением той же процедуры с gE-отрицательным контрольным препаратом вместо препарата gE-антигена. На каждом микротитровальном планшете имеются положительные и отрицательные контрольные сыворотки. Тест считается действительным, если положительные сыворотки дают положительный результат в определенном разбавлении. Сыворотка положительна, если ее ОП выше на 0.2, чем у стандартной отрицательной контрольной сыворотки.
c/ В соответствии с принципом IDAS, описанным под 2, но после инкубации исследуемой сыворотки вместо конъюгата анти-коровьего иммуноглобулина используется анти-gE MAT/HPPO. Можно использовать сыворотку анти-gE пептида или анти-gE поликлональную сыворотку вместо анти-gE MAT. Планшеты промываются и к каждой лунке добавляется хромогенный субстрат. Приблизительно после 2 часов при комнатной температуре планшеты считываются спектрофотометрически. В каждый планшет включены четыре отрицательных контрольных сыворотки и четыре серийных разбавления положительной сыворотки. Сыворотка с величиной ОП менее 50% величины ОП 4-х отрицательных контрольных сывороток, которые исследовались на том же планшете, считается положительной.
d/ В соответствии с принципом блокирующего ферментного иммуносорбентного теста /ELISA/, по которому антиген вируса, который может быть очищенным или неочищенным, наносится покрытием на микротитровальный планшет на всю ночь. На этих планшетах исследуемая сыворотка инкубируется примерно один час или больше при 37oC. После процедуры промывания на планшеты добавляется анти-gE MAT с последующей инкубацией приблизительно 1 час при 37oC. Вместо анти-gE MAT можно использовать сыворотку анти-gE пептида или анти-gE поликлональную сыворотку. Планшеты промываются и к каждой лунке добавляется хромогенный субстрат. После примерно 2 часов при комнатной температуре планшеты считываются спектрофотометрически. На каждом планшете имеются четыре отрицательные контрольные сыворотки и четыре серийных разбавления положительной сыворотки. Сыворотка с величиной ОП менее 50% средней величины ОП 4х отрицательных контрольных сывороток, которые исследовались на том же планшете, считается положительной.
Во всех указанных выше процедурах может использоваться традиционный культивированный антиген вируса, который содержит gE, а также gE-антиген, который выражен через прокариоты или эукариоты. Как вариант, вместо традиционного антигена в указанных диагностических пробах могут использоваться олигопептиды, основанные на BHV-1 gE последовательности. Кроме того, такие олигопептиды можно использовать для разработки так называемого "коровьего" теста в соответствии с принципом, описанным в статье Kemp и др. Science, 241, 1352-1354, 1988. Такой тест будет основываться на связи антигенной последовательности олигопептида антителами, направленными против gE, присутствующего в зараженных животных. Для такого теста олигопептиды должны быть связаны с MAT, направленными против эритроцитов коров.
Анализ на нуклеиновую
кислоту с использованием цепной реакции полимеразы
Олигонуклеотиды /зонды и праймеры/ могут, например, использоваться в цепной реакции полимеразы, чтобы различить вакцинированных и зараженных
животных. Цепная реакция полимеразы /ЦРП/ - технологический прием, посредством которого нуклеиновые кислоты патогена можно в короткое время размножить миллиарды раз /De polymesase kettingreactie. P.F.
Hilderiuk, J.A. Wagenaar, J. W. B. van der Giessen u B.A.M. van der Jeijst, 1990, Fijdschrift voor Dierdeneeskunde deel 115, 1111-1117/. gE олигонуклеотиды можно выбрать таким образом, что в gE
положительном геноме образуется другой продукт, нежели в gE отрицательном геноме. Преимуществом этого является то, что и животное, которое было вакцинировано вакциной с делецией gE, дает положительный
сигнал в тесте на ЦРП. Однако этот подход зависит от присутствия нуклеиновых кислот вируса в образце, например, крови от животного, на которой проводится тест.
После острого заражения BHV-1 имеется большая вероятность, что BHV-1 специфические нуклеиновые кислоты могут быть продемонстрированы в крови, но еще не определилось, будут ли также демонстрироваться в крови нуклеиновые кислоты BHV-1 во время латентного периода.
Использование ВНV-1 в качестве вектора
Для экспрессии гетерологичных генов в BHV-1 геноме необходимо иметь в наличии точную
информацию об участке, куда должен вставляться гетерологичный ген. Основные последовательности не должны нарушать структуру, а для экспрессии гетерологичного гена должны быть регуляторные
последовательности. Гликопротеин gE-ген является подходящим местом для экспрессии гетерологичных генов. gE-ген не является существенным, следовательно, не может быть возражений по замене gE гена
гетерологичным геном. Как следствие этого, гетерологичный ген может так располагаться, что он будет находиться под влиянием регуляторных последовательностей gE гена. Однако нет необходимости
использовать регуляторные последовательности gE-гена. Экспрессия гетерологичных генов может контролироваться другими, т. е. более сильными регуляторными последовательностями различных генов.
Представляется возможным также лигировать гетерологичный ген к /экпортному/ сигнальному пептиду gE гена с тем, чтобы повлиять на секрецию продукта гетерологичного гена. Очевидно, что подробное знание
гена gE и белка gE дает возможность использовать BHV-1 как вектор достаточно контролируемым образом. Разработанные векторы можно серологически отличить от дикого типа. Создание ВНV-1 мутантов, которые
эксцрессируют гетерологичные гены, можно реализовать таким же образом, что и создание мутантов с делецией gE, показанное в примерах. Однако тогда фрагменты делеции следует заменить фрагментом, на
котором гетерологичный ген расположен в местоположении делеции.
Примеры
1/ Изоляция и определение естественного мутанта с делецией gE.
a/ Изоляция
естественного /природного/ мутанта
Геномная ДНК была изолирована из ряда ослабленных традиционным путем вакцин в соответствии со стандартными способами и проанализирована с использованием
рестрикционных ферментов. В частности, мы искали отклонения генома, которые бы подходили, чтобы отличить от вируса BHV-1 дикого типа.
Ослабление было направлено, в частности, на Us участок генома BHV-1, потому что в этом участке - по аналогии с вирусом герпес простой - вероятно располагается ряд генов, кодирующих для несущественных гликопротеинов /Определение гена гликопротеида вируса простого герпеса 1 внутри кластера генов, необязательного для роста в клеточной культуре, R. Longnecker S. Chatterjee, R.J. Whitley u B. Roijrnan (1987) Proc. Natl dead. Sci, 84, 4,03-4307/.
Подборка BHV-1 вакцин из Загребского университета, Югославия /Lugovic и др. Veterinarski Arhiv, 55, 241-245, 1985/, после большого числа пересевов на эмбриональные почечные клетки коровы и эмбриональные трахеальные клетки коровы /Эбтр/, обладала участком отклонения Us, помимо наличия нормального участка Us. Более того, эта вакцина образовывала и большие и малые пятна на Эбтр клетках. Из этой смешанной популяции тремя этапами ограниченного разведения был изолирован вирус с участком Us отклоняющейся формы, с небольшими пятнами в каждом случае. Вирус, изолированный этим путем, исследовался дальше и был назван Difivac-1. Он был депонирован в Институт Пастера, Париж, Франция 27 мая 1992 г, номер депозита 1-1213.
b/ Определение делеции в gE гена в Difivac-1
Для дальнейшего анализа этого отклонения в Us участке геномная ДНК Difivac-1 была изолирована в соответствии со стандартными методами и
подвергнута блот-анализу /фиг. 1A/. Гиридизация этого блота фрагментом Hind III K дикого типа, помеченного 32p подтвердила, что этот фрагмент, расположенный в центре участка Us, короче на
1.0 тысячу пар нуклеотидов /т.п.н./ в Difivac-1. Более того, этим анализом можно было приблизительно определить позицию отсутствующей части /фиг. 1B/. Для дальнейшего анализа этой делеции Us
участок BHV-1 штамма Lam дикого типа был изолирован и клонирован в прокариотные векторы. С этой целью в соответствии со стандартными методами геномная ДНК штамма Lam /фиг. 2A/ была изолирована и
клонирована в векторы pUC18, рACYC и pBP322 /фиг. 2B/. Была составлена физическая карта участка вокруг предполагаемой позиции делеции /фиг. 2C/. Исходя из этой физической карты, в векторах pKUN 19 и
pUC18 /фиг. 2/ были конструктированы субклоны, пригодные для определения последовательности нуклеотидов этой области. Используя эти субклоны, методом Сангера была определена нуклеотидная
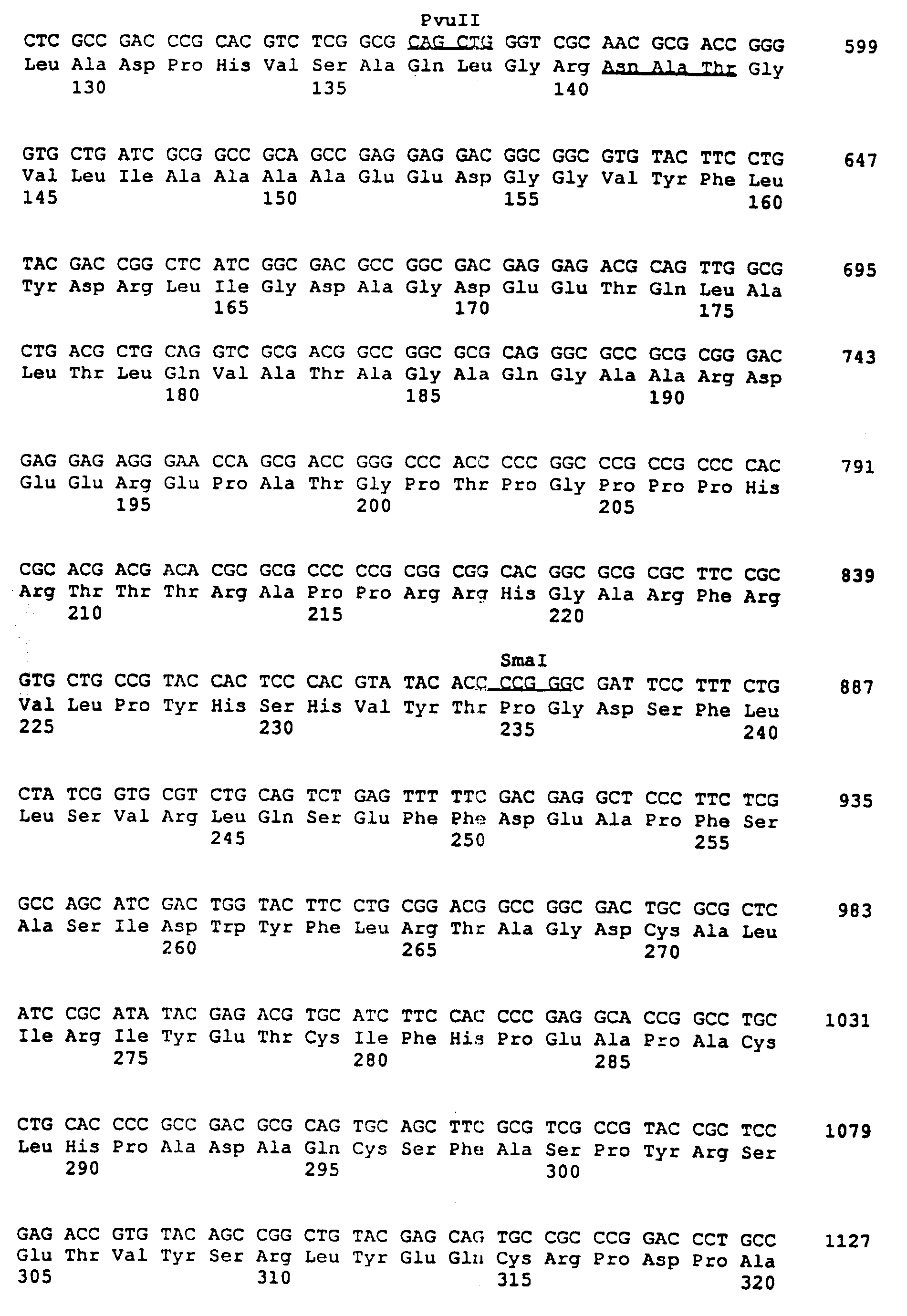
последовательность двух цепей всей области /показано на фиг. 2C/. Эта нуклеотидная последовательность была анализирована с использованием программы PC/ген. Из концептуальной трансляции оказывается,
что нуклеотиды /нт/ 168-1893 кодируют для открытой рамки считывания 575 аминокислот /фиг. 3A/. Дальнейший анализ показал, что эта последовательность аминокислоты имеет характеристики трансмембранного
гликопротеина, как показано на фиг. 3B. Факт таков, что эти первые 26 аминокислот /ак/ опознаются как типично эукариотный экспортный сигнал и область между ак 423 и ак 450 опознается как
трансмембранная область. Кроме того, в этой последовательности оказываются три потенциальных участка N-связанной гликосиляции. Эта предсказанная последовательность аминокислоты демонстрирует близкое
сходство с gE-геном гликопротеина вируса простого герпеса /HSV/; см. фиг. 4A и 4B. Эти и другие сходства оправдывают вывод, что найденный ген является gE гомологом BHV-1. По этой причине ген
называется gE. Чтобы определить, до какой степени этот BHV-1 gE-ген отсутствует в Difivac-1, был изолирован p318 фрагмент. Фрагмент p318 начинается на AluI участке 55 нт до требуемой открытой рамки
считывания BHV-1 gE и заканчивается 133 нт за ней. Геномная ДНК Difivac-1 анализировалась этим p318 фрагментом с использованием блот-гибридизации. Это выявило, что Difivac-1 не содержит p318
обнаруживаемых последовательностей /фиг. 5/. Этот эксперимент подтвердил, что Difivac-1 содержит делецию и четко продемонстрировал, что эта делеция проходит по всему gE гену.
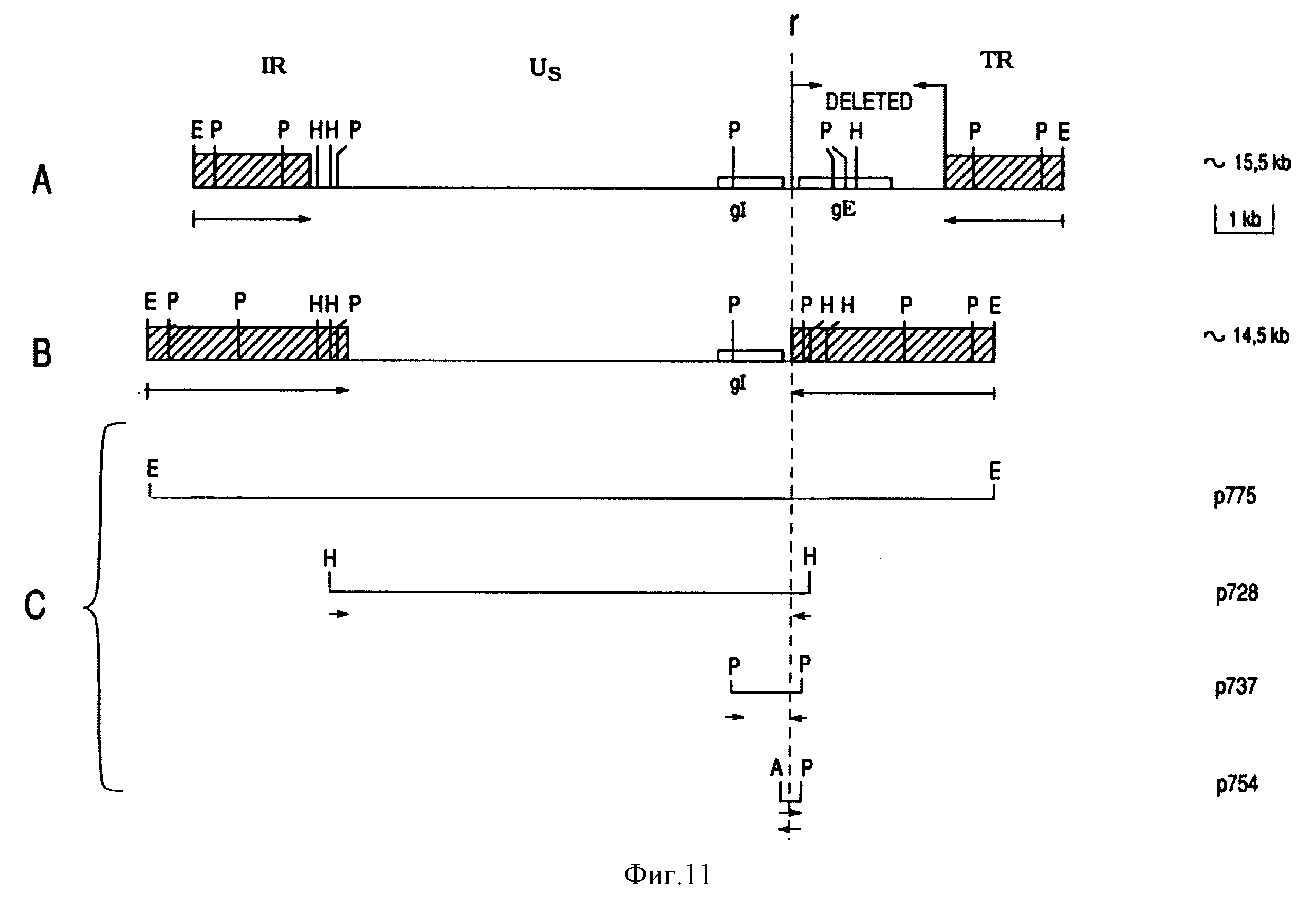
Чтобы определить размер и позицию участка делеции, геномные последовательности, охватывающие Us регион Difivac-1, были клонированы в прокариотные векторы (См. фиг. 11C), фрагмент с 14.5 т.п.н. был клонирован в pACYC вектор и назван p775. HindIII фрагмент в 7.4 т.п.н. был независимо клонирован в pUC18 вектор и назван p728, были изолированы два субклона: фрагмент PstI с 1,4 т. п. н. в клоне p737 и фрагмент AluI - PstI в клоне p754 с 350 т.п.н. Анализ рестрикционного фермента и бло-анализ этих клонов /данные не показаны/ продемонстрировали, что делеция gE в Difivac-1 составляет длину 2,7 т.п. н., начиная сразу от 5' с gE гена и кончая на границе Us участка. Эти 2.7 т. п. н. были замещены дублированием одного сегмента в 1 т.п.н., расположенного в участке Us напротив gE гена. См. фиг. 11B. Чтобы подтвердить результаты этого анализа и определить точную точку рекомбинации, нуклеотидная последовательность большинства вставок клона p754 определялась и сравнивалась с последовательностями дикого типа. См. фиг. 12. Этот анализ показал, что точка рекомбинации расположена у 77 пары основания /по/ верх от начального кодона gE гена.
c/ Оценка безопасности и эффективности Difivac-1
Difivac-1 был опробован на семинедельных телятах, свободных от BHV-1 сероотрицательного специфического патогена. Восемь телят были
вакцинированы через нос 105 TCID50 в 2 мл, из которых по 1 мл распылялось в каждую ноздрю. Восемь семинедельных телят, свободных от BHV-1 сероотрицательного специфического
патогена, помещались в изоляции, им вводилось в нос 2 мл среды культуры, и они служили невакцинированными контрольными животными. Через пять недель после вакцинации вакцинированным и контрольным
телятам капали в нос 107 TCID50 очень вирулентный штамм Iowa BHV-1. Шесть недель спустя после контрольного заражения всем телятам вводился внутримышечно дексаметазон в течение 5
дней, чтобы реактивировать предполагаемый латентный вирус. Клинические проявления, ректальные температуры и рост организма находились под контролем. Вирус изолировался из носовых мазков, и в сыворотке
были определены титры нейтрализующего антитела.
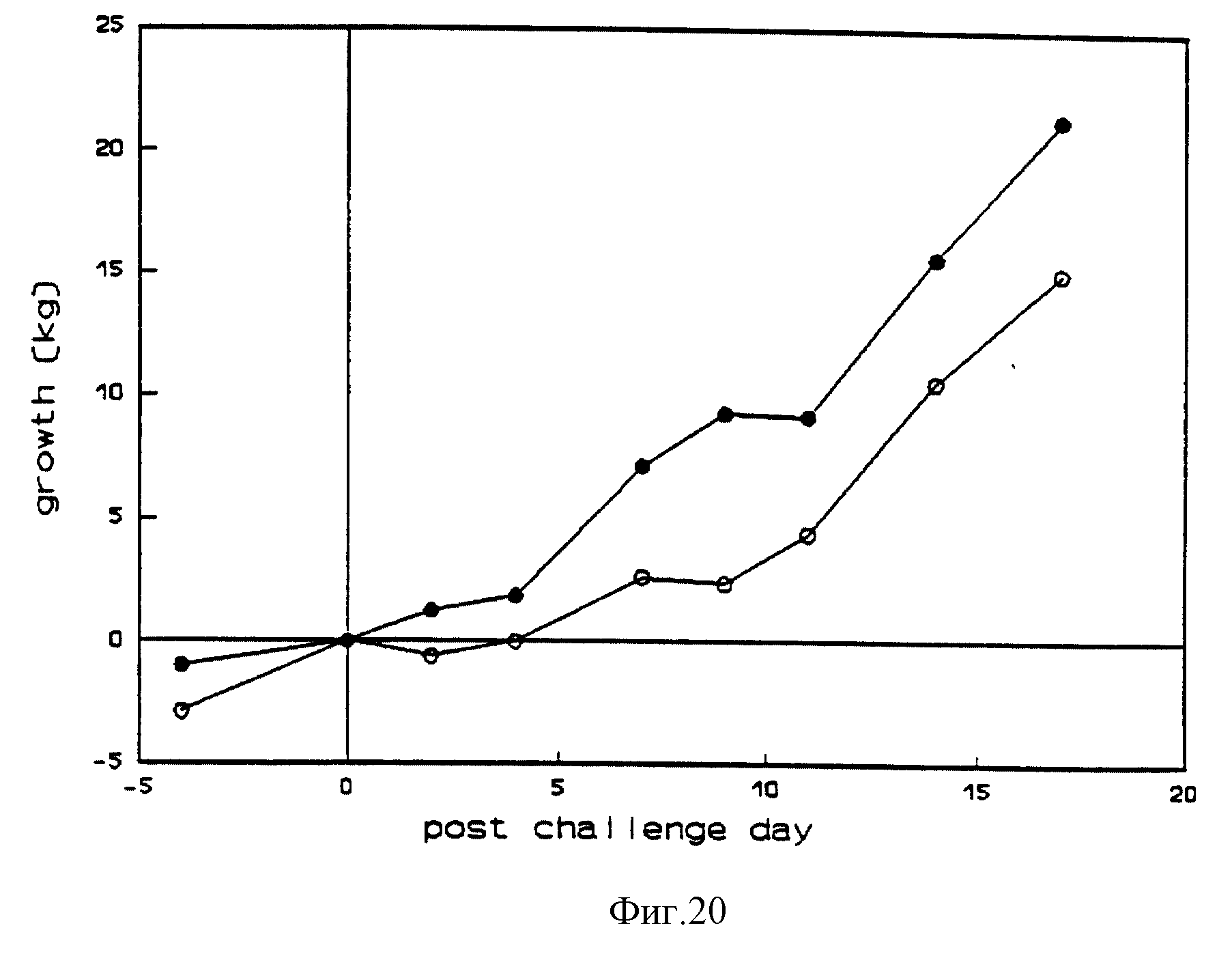
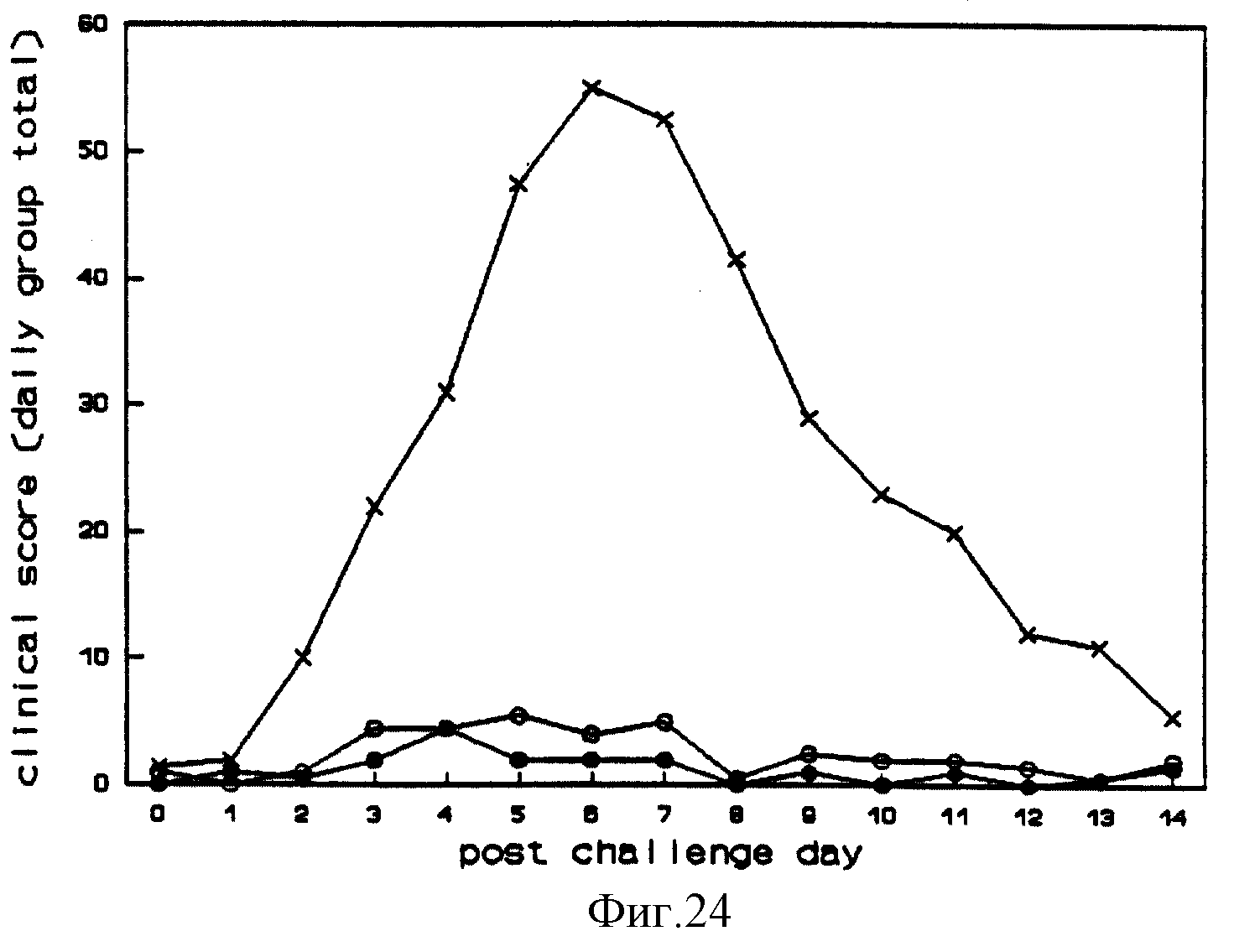
После вакцинации поведение, аппетит, ректальные температуры и скорости роста телят оставались нормальными, но у вакцинированных телят наблюдались некоторые серьезные выделения из носа и повышенное слюноотделение. Нарушений в слизистой оболочке носа не наблюдалось. После вакцинации из носовых мазков выделялся Difivac-1 /фиг. 17/. Все вакцинированные телята вырабатывали нейтрализующие антитела по отношению к BHV-1. После контрольного заражения все не вакцинированные контрольные телята проявляли апатию, потерю аппетита, выделения из носа и глаз, покраснение десен нижней челюсти, сильные нарушения в слизистой оболочке носа в течение 14 дней после контрольного заражения и остановку роста в течение 4 дней. У вакцинированных телят были небольшие, быстро заживающие повреждения слизистой носа и не было остановки роста. Ежедневные клинические проявления, ректальная температура и развитие роста даны на фиг. 18, 19 и 20. После контрольного заражения все телята выделили вирус через нос, но количество и период вирусного выделения были заметно снижены у вакцинированных телят (фиг. 21). Вторичная реакция антитела, развившаяся у вакцинированных и невакцинированных телят, выработала антитела после заражения.
После реактивации вирус контрольного заражения был изолирован от одного вакцинированного теленка и от 5 невакцинированных телят. Difivac-1 нельзя было реактивировать.
Данные результаты демонстрируют, что Difivac-1 не вызывал какого-либо признака заболевания у молодых телят и не подвержен реактивации. Difivac-1 значительно снизил силу заболевания и количество выделения вируса после контрольного заражения.
В заключение можно сказать, что Difivac-1 является безопасной и эффективной вакциной для использования в вакцинации животных против заражений BHV-1.
2/ Построение рекомбинантных мутантов с делецией gE вируса BHV-1
Чтобы иметь в
распоряжении BHV-1 вакцины со способностью дифференциации, которые молекулярно определены лучше, чем Difivac-1, и которые при желании могут содержать делецию, например, в гене тимидинкиназы в
дополнение к делении в gE гене, как дополнение к Difivac-1, были построены рекомбинантные мутанты с делецией gE. Начиная с определенной позиции гликопротеин gE-гена и используя клонированные ДНК
фрагменты, которые фланкируют gE-ген, можно построить фрагмент делеции gE. С использованием стандартной техники /F.L. Graham u van der Eb. 1973, Virolоgy, 52, 456-467/, можно рекомбинировать этот
фрагмент делеции в геноме штамма BHV-1 дикого типа, что дает мутант с делецией gE.
a/ Построение фрагмента с делецией gE
Для построения фрагмента с делецией gE выбирался
фрагмент, у которого, с одной стороны, не доставало полной gE последовательности, и, с другой стороны, который содержал достаточную фланкирующую последовательность, чтобы позволить рекомбинацию с
геномом дикого типа. Был выбран ген на 5' /вверх/ стороне, фрагмента 1.2 т.п. PstI - AsuII, который заканчивается 18 нт до начального кодона gE. Для 3' /вниз/ фрагмента был выбран фрагмент 1.2 т.п.н.
EcoNI-DcaI, который начинается от 2 нт до остановочного кодона gE гена /фиг. 6/.
Для построения фрагмента с делецией gE фрагмент в 1.4 т.п.н. PstI-SmaI от 8.4 т. п. н. HindIII К фрагмента штамма Iam BHV-1, расположенного на 5' стороне gE гена, был субклонирован в SmaI и PstI участок плазмиды pUC18. Этот клон был назван p515. EcoNI-SmaI фрагмент, расположенный на 3' стороне gE и исходящий от 4.1 т.п.н. HindIII-EcoRI клона, был клонирован в уникальный AsuII участок p515. Таким образом построение фрагмента с делецией gE было завершено, и построенный клон был назван p519. Хотя в принципе всю вставку PstI-SmaI p5I9 можно было использовать как фрагмент с делецией gE, это нежелательно. Дело в том, что PstI-SmaI имеет протяженность приблизительно 100-150 пар оснований /по/ в повторяющуюся последовательность, которая фланкирует Us участок. Этот кусок в 100-150 по может рекомбинироваться с повторяющейся последовательностью на другой стороне Us области, где не расположен gE ген, и поэтому может дать нежелательные продукты рекомбинации. По этой причине для эксперимента рекомбинации был выбран PstI-DraI фрагмент, так что 100 по повтора удалены.
b/ Рекомбинация фрагмента с делецией gE с геномом BHV-1 дикого типа
Чтобы осуществить рекомбинацию между построенным фрагментом с делецией gE и геномом BHV-1 дикого типа,
микрограммовые количества двух молекул ДНК вместе трансфицируются на эмбриональные трахеальные клетки коровы /Эбтр/ в соответствии со стандартным методом F.L. Graham u A.J. van der Eb. /1973, Virology,
52, 456-467/. Механизмы клеточной рекомбинации обеспечивают рекомбинацию небольшого процента молекул ДНК /2-4%/, которые включены в клетки. Для выбора рекомбинированных мутантов с делецией gE,
вирусная смесь, сформированная после трансфекции, рассеивается на свежую клеточную культуру Эбтр. В большинстве случаев отдельные популяции вируса, которые развиваются, /пятна/, берут начало от одного
вируса. Для изоляции мутантов BHV-1 штамма Lam с делецией gE были изолированы 230 из этих пятен /бляшек/ и исследованы в соответствии со стандартными иммунологическими методами с помощью специфических
моноклональных антител BHV-1 /MAT/, которые не реагируют с зараженными клетками Difivac-1. Эти MAT направлены против гликопротеина gE. Пять из 230 бляшек не реагировали с этими MAT. ДНК этих 5 бляшек
исследовалась дальше.
c/ ДНК анализ построенных мутантов с делецией gE штамма Lam BHV-1
ДНК препараты 3х /1B7, 1B8, 2H10/ вышеупомянутых 5 кандидатов мутантов с
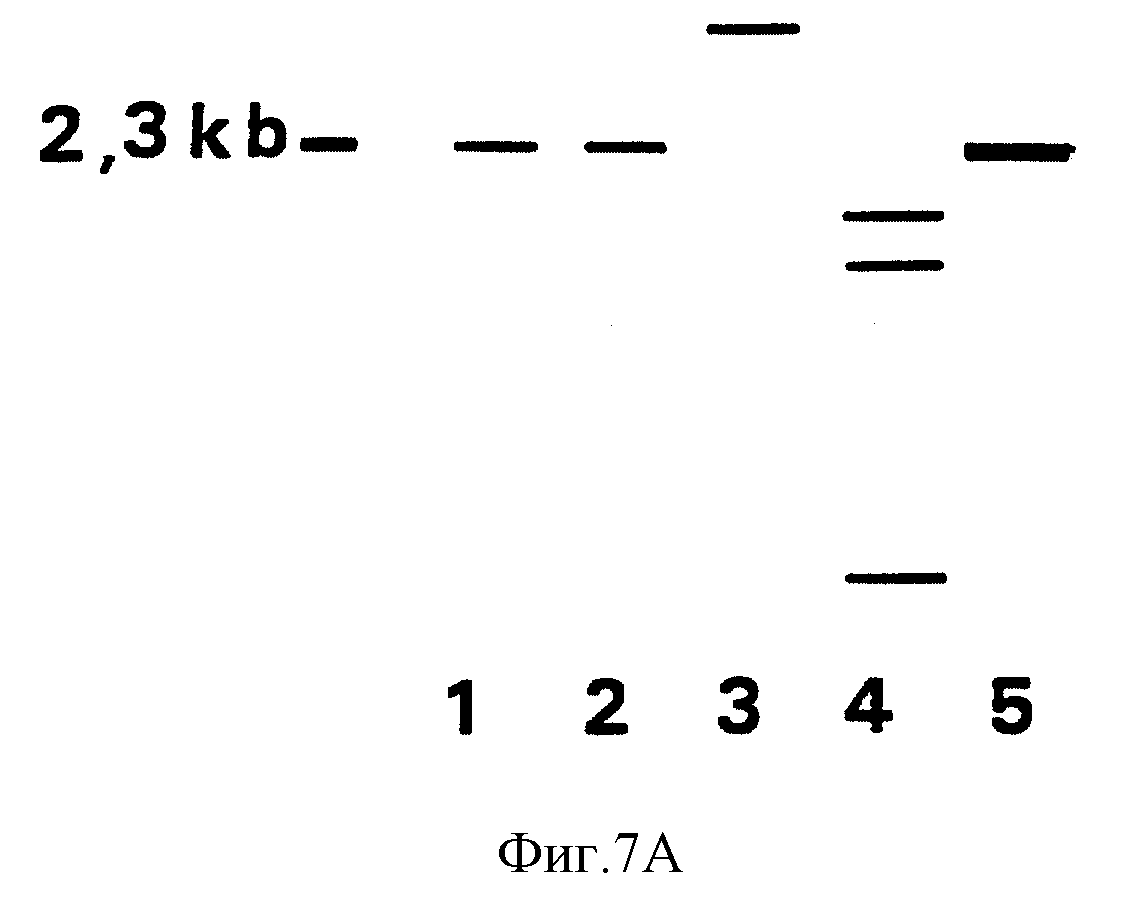
делецией gE исследовались далее с использованием стандартной техники блот-анализа /Sambrook и др. 1989/. Двойное переваривание этих препаратов ДНК с помощью PstI и DraI с последующим гелевым
электрофорезом и блот-гибридизацией фрагментом делеции PstI-DraI в 2.3 т.п.н. в качестве зонда показали, что gE ген генома вирусных популяций 1B7 и 1B8 точно удален желаемым образом; см. фиг. 7A и 7B.
Популяция 2H10 имеет PstI-DraI фрагмент отклонения. Блот-гибридизации с gE-специфическим зондом показывают, что ни в одном из трех препаратов ДНК не располагаются gE последовательности /результаты не
показаны/. Популяции 1B7 и 1B8 вируса BHV-1 являются предполагаемыми рекомбинантными мутантами с делецией gE. Популяция 1B7 вируса BHV-1 тестировалась на свойства вакцины.
d/ Построение мутантов с двойной делецией тимидинкиназы/gE
Поскольку BHV-1 рекомбинантные мутанты с делецией только в одном гене могут быть недостаточно сниженной вирулентности, делеции также
предусмотрены в гене тимидинкиназы /ТК/ BHV-1 штаммов Lam и Harberink. Эти мутанты были построены аналогично используемым для вышеупомянутых мутантов с делецией gE /результаты не показаны/. Эти
мутанты с делецией ТК использовались для построения мутантов с двойной делецией ТК/gE.
e/ Построение мутантов с двойной делецией гликопротеин gI/гликопротеин gE
Поскольку скот,
вакцинированный мутантом с одной делецией gE, может выработать анти-gI антитела, которые могут помешать обнаружению анти gI/gE антител /обсуждается ниже/, мы также изобрели вакцину с двойной делецией
gI/gE. Такой мутант с двойной делецией gI/gE может быть построен с использованием тех же процедур, которые использовались для построения мутанта с одной делецией gE. Анализ частично нуклеотидной
последовательности верхнего конца от фрагмента 1.8 т.п.н. PstI, который охватывает 5' конец gE гена, выявил открытую рамку считывания со значительной гомологией по отношению к gI гомологам, найденным
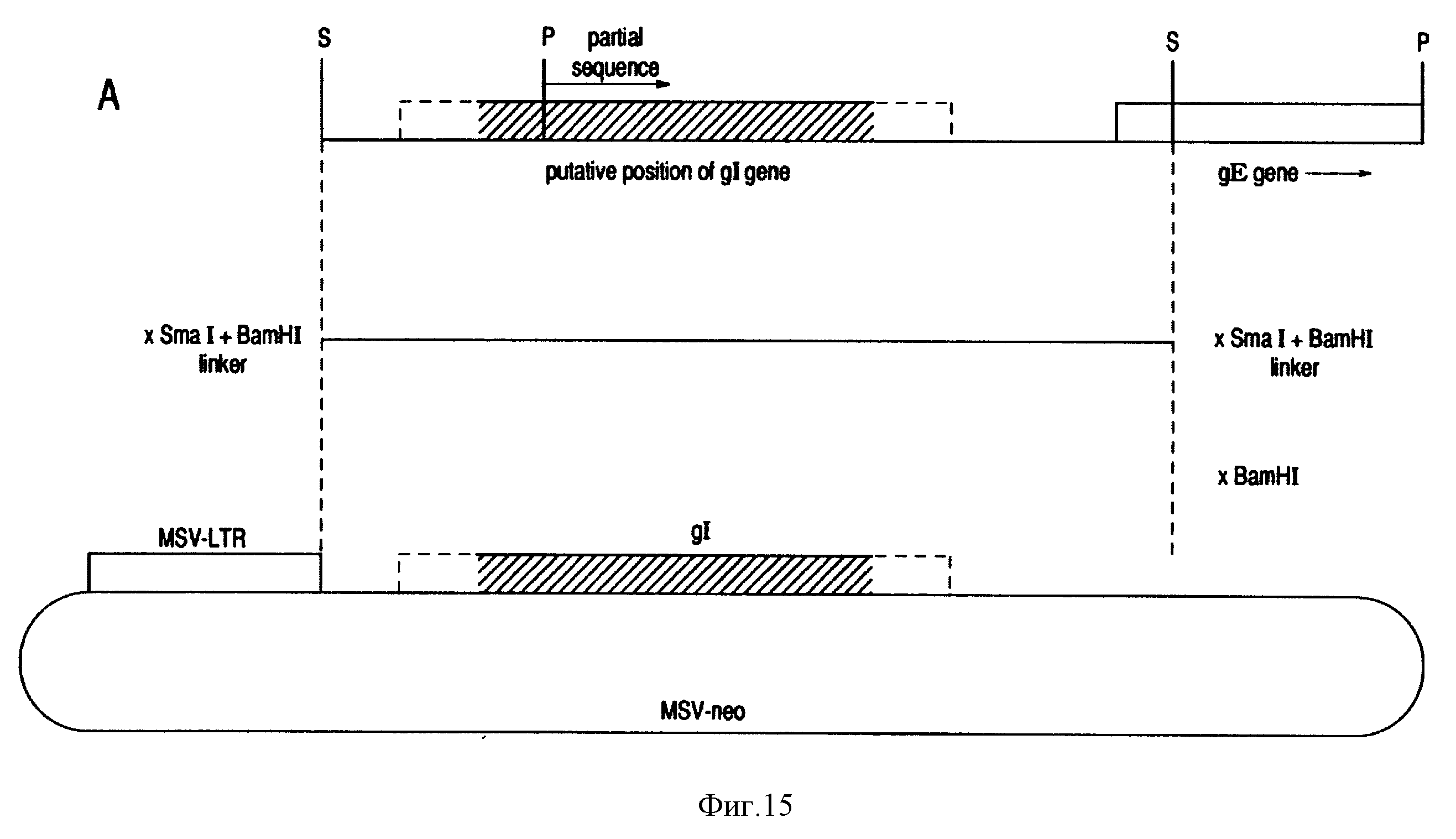
в других вирусах герпеса. См. фиг. 13 и 14. Используя SmaI - PstI фрагмент с 350 по, которых охватывает предполагаемый 5' конец gI гена и фрагмент EcoNI-SmaI, расположенный вниз от gE гена, можно
построить фрагмент делеции gI/gE. Этот фрагмент можно рекомбинировать с геномом дикого типа, чтобы получить мутант с делецией и gI/gE вируса BHV-1. См. фиг. 16. 80-90 аминокислот, которые еще можно
теоретически получить, не смогут выявить антитела, которые могут помешать обнаружению анти-gI/gE антител. Дальнейший анализ последовательности gI гена позволит создать делецию gI, которая охватывает
полную область кодирования gI. Этот мутант с двойной делецией gI/gE был назван Difivac-IE.
e/ Оценка безопасности и эффективности мутантов Lam gE- и Lam gE-,
TK-
Свойства вакцины мутантных штаммов Lam gE- и Lam gE-, TK-, BHV-1 тестировались на семинедельных сероотрицательных свободных от специфических
патогенов телятах. Каждый мутантный штамм распылялся в нос 6 телятам. Каждому теленку давалась общая доза 105 TCID50 в 2 мл среды культуры, из которых 1 мл распылялся в каждую
ноздрю. Еще 6 телятам в нос распылялась среда культуры, свободная от вируса, и это были контрольные невакцинированные телята. Спустя 5 недель после вакцинации все телята, вакцинированные и контрольные,
заражались через нос 107 TCID50 высоко вирулентным штаммом Iowa BHV-1. После вакцинации и после контрольного заражения контролировались клинические проявления, ректальная
температура и вес тела. Брались мазки из носа на определение числа дней выделения вируса из носа.
После вакцинации поведение, аппетит, ректальная температура и рост телят оставались нормальными. У всех вакцинированных телят наблюдались сильные носовые выделения и небольшие нарушения слизистой носа. Вирус можно было изолировать из носа вакцинированных телят приблизительно за 7 дней /Таблица 1/.
После контрольного заражения все невакцинированные контрольные телята проявляли апатию, потерю аппетита, выделения из носа и глаз, покраснение десен нижней челюсти, сильные нарушения слизистой носа и снижение темпа роста. У телят, вакцинированных Lam gE-, TK-, у всех наблюдались выделения из носа и небольшое раздражение слизистой носа. Не у всех телят, вакцинированных Lam gE-, развивались выделения из носа или нарушения слизистой оболочки. У вакцинированных телят не наблюдалось апатии, потери аппетита или других клинических симптомов заболевания. Ректальная температура, рост и клиника после контрольного заражения показаны на фиг. 22, 23 и 24. Невакцинированные телята выделяли вирус из носа в 2 раза дольше вакцинированных /Таблица 1/.
Указанные результаты демонстрируют, что штаммы мутантов Lam gE- и LamgE-, TK- BHV-1 практически не вызывают каких-либо клинических признаков заболевания в молодых телятах. Оба мутантных штамма остановили проявление болезни после контрольного заражения и снизили период выделения вируса через нос на 50%.
Мутантные штаммы Lam gE- и Lam gE-, TK- вируса BHV-1 безопасны и эффективны для использования в качестве вакцины для животных против заражений BHV-1.
3/ Прокариотная экспрессия gE
Для прокариотной экспрессии gE-гена гликопротеина BHV-1 до сих пор использовались векторы экспрессии pGEX /D. B. Smith и K.S. Johnson, Ген 67 /1988/ 31-40/. Код
векторов pGEX для глютатион S-трансферазы /GST/ белка-носителя от Schistoma japonicum, которая находится под влиянием промотора, который индуцируется для экспрессии изопропилтиогалактозидом /IPTG/.
Примером белка слияния GST-gE является продукт построения pGEX-2T600S3 /фиг. 8A/. В этой конструкции с использованием стандартной молекулярно-биологической техники /Sambrook и др. 1989/, за CST геном
был лигирован SmaI фрагмент на 600 по, который кодирует для N-терминального участка в 200 аминокислот gE белка. Эта конструкция предполагается быть в устроенном варианте, и с каждым разом к GST
лигируется фрагмент на 600 по с разной рамкой считывания. Все эти конструкции были введены в штамм DH5 Escherichima coli, индуцированный IPTG и сформированные белки были перенесены на нитроцеллюлозу
после электрофореза полиакриламидным гелем посредством блотирования /wester blatteng/. Иммунологическое обнаружение с помощью анти-GST продемонстрировало, что только правильная рамка считывания /N 3/,
которая кодирует для участка gE белка, ведет к экспрессии четкого белка слияния предполагаемого размера в 27к /GST/ + 20к /gE/ = 47 к. Три из МАБ, изолированные нами, которые не реагируют с Difivac-1,
опознают 47 кД GST - gE белок слияния в блоте; см. фиг. 8B.
4/ Эукариотная экспрессия гликопротеин gE-гена
Для эукариотной экспрессии гликопротеин gE-гена заранее выбирается
вектор pEVHIS. Вектор pEVHIS в качестве эукариотного маркера имеет кодирование HISD гена для гистидинолдегидрогеназы [EC 1.1.1.23] /C. Hartmann и R. Mulligan 1988, Proc. Natl Acad. Sci США, 85,
8047-8051/, которая заставляет клетки перенести токсическую концентрацию гистидинола в 2.5 мМ. Вектор также включает регион промотора непосредственного раннего гена человеческого цитомегаловируса
/HCMV/, с уникальными участками рестрикционного фермента, расположенными за ним. Для создания вектора экспрессии pEVHIS/gE использовался фрагмент, включающий целый кодирующий регион гликопротеид
gE-гена. Он начинается на AluI участке 55 по до предполагаемой открытой рамки считывания gE и кончается на 133 по за ней. Этот регион был клонирован за HCMV промотором вектора pEVHIS, в результате
чего была получена конструкция pEVHIS/gE /фиг. 9/. pEVHIS/gE был амплифицирован в клетках E. coli - DH5α и очищен посредством градиента хлорида цезия /Sambrook и др. 1989/. Эта очищенная ДНК
была трансфицирована на BaLb/C-3T3 в соответствии с методом Graham и van der Eb. Трансформированные клетки отбирались гистидинолом с тем, чтобы изолировать потом двадцать устойчивых к гистидинолу
колоний. Эти колонии исследовались МАТ 81 посредством иммунной реакции монослоя пероксидазы /IPMA/. В четырех колониях был выражен gE белок, из этих четырех колоний 3T3 gE клон 9 использовался, чтобы
изолировать субклон с высокой gE экспрессией. Клон, изолированный этим методом, /названный 3T3 gE 9.5/ использовался для характеристики кандидата в анти-gE моноклональные антитела.
5/ Эукариотная экспрессия и BHV-1 гликопротеина gE, и BHV-1 гликопротеина gI в той же клетке
Чтобы экспрессировать BHV-1 гликопротеин gI в той же клетке, что и BHV-1 гликопротеин gE, мы
вначале определили предполагаемую позицию гена BHV-1 gI. Поскольку ген gI гликопротеина вируса простого герпеса располагается непосредственно выше от gE гена гликопротеида, было предположено, что ген
BHV-1 будет располагаться на соответствующей позиции. Чтобы проверить это, определялась последовательность региона в 283 нуклеотида, расположенных у 1 т.п.н. вверх от начала гена gE BHV-1.
Концептуальная трансляция этого региона показала, что вторая рамка считывания кодирует для последовательности 94 аминокислот, которая гомологична гликопротеину gI вируса простого герпеса /фиг. 13 и
14/. Поскольку гомологичный сегмент находится на протяженности 80 аминокислот от начального кодона, предполагаемое начало открытой рамки считывания gI гена BHV-1 находится приблизительно у 250 нт
вверх от региона последовательности. Из этого следует, что фрагмент SmaI в 1.7 т.п.н., который начинается от 400 нт вверх от региона последовательности, и заканчивается в гене gE, содержит полный
кодирующий регион gI гена BHV-1. Этот SmaI фрагмент в 1.7 т.п.н. был клонирован в эукариотный вектор MSV - neo /См. фиг. 15/. Этот вектор содержит сильный промотор вируса мышиной саркомы и селекторный
ген neo, который кодирует для устойчивости против антибиотика G-418 сульфат Генетицина. Полученная конструкция MSV neo GI была амплифицирована в DH5α клетках E. coli и была трансфицирована в
3T3 gE 9.5 клетки с использованием метода Graham и van der Eb. Трансфицированные клетки отбирались с помощью среды культуры 400 мкг генетицина/мл и устойчивые колонии были изолированы и тестировались
кандидатом анти-gE MAT, который не реагировал с 3T3 gE 9.5 клетками. Из этого мы отобрали клон, который реагировал, например, с MAT 66, как реагирует BHV-1 дикого типа.
6/ Характеристика кандидата анти-gE MAT
Были получены MAT против BHV-1 дикого типа и отобраны по их неспособности реагировать с эмбриональными трахеальными клетками коровы (Эбтр),
инфицированными Difivac-1. Эти MAT исследовались на их реактивность с
a/ мутантом с делецией Lam gE-;
b/ вышеописанным продуктом прокариотной экспрессии в блоте;
c/ вышеописанными gE-экспрессирующими Balb/c-3T3 клетками;
d/ клетками, упомянутыми под c/ и зараженными Difivac-1, и
e/ Balb/c-3T3 клетками, экспрессирующими gE/gI комплекс.
Для тестирования на реактивность под a, c, d и e использовалась иммунная реакция монослоя пероксидазы /IPMA/. Результаты в Таблице 2 показывают, что мы получили MAT, которые направлены против E /номеров 2, 3, 4, 52, 66, 68, 72 и 81/ и MAT /номера 1, 51, 53, 67, 75 и 78/, которые могут быть направлены против конформационных антигенных доменов на gE/gI комплексе. Завершение IPMA для нанесения на карту антигенных доменов, опознанных различными MAT, показало, что, по меньшей мере, 4 антигенных домена присутствуют на гликопротеине gE и что один домен, вероятно, сформирован комплексом gE/gI. Таблица 2.
Обнаружение анти-gE антител у скота, зараженного BHV-1
Чтобы определить, присутствуют ли в сыворотке зараженного скота антитела против gE, было
проведено непрямое блокирование IPMA 16 кандидатами gE-MAT и следующими 8 отобранными сыворотками:
- 2 сыворотками коров, вакцинированных Difivac-1 и зараженных контрольно вирулетным штаммом
Iowa, которые забирались 14 дней спустя после контрольного заражения;
- 2 сыворотками коров, экспериментально зараженных подтипом 1 вируса BHV-1, которые забирались 20 месяцев спустя после
заражения. Одна из коров заражалась подвержением контакту;
- 2 сыворотками коров, экспериментально зараженных подтипом 2 вируса BHV-1, которые забирались 20 месяцев спустя после заражения.
Одна из коров заражалась от контакта;
- сывороткой теленка, свободной от специфического патогена, который был вакцинирован ts мутантной вакциной и спустя 3 недели заражался подтипом 2b вируса
BHV-1, сыворотка забиралась через 7 недель после контрольного заражения;
- сывороткой гнотобиотического теленка, вакцинированного мутантной вакциной и зараженного 3 недели спустя подтипом 2b
вируса BHV-1, сыворотка забиралась через 7 недель после контрольного заражения.
Таблица 2 показывает, что все эти сыворотки содержали антитела против антигенных доменов III и IV на gE и против антигенного домена 1, который, вероятно, расположен на комплексе gE/gI. Мы можем сделать вывод, что gE является подходящим серологическим маркером для различения между зараженным BHV-1 и вакцинированным скотом.
7/ Обнаружение нуклеиновых кислот BHV-1 посредством ЦРП процедуры с использованием gE-специфических праймеров BHV-1
Начиная с определенной нуклеотидной
последовательности gE гена BHV-1, была выбрана пара праймера, подходящая для ЦРП с использованием программы селекции праймера Lowe и др. /Rowe, J. Sharefkin. S. Ai Jand и C.W. Dieffenbach, 1990,
nucleic Acids Res. 18, 1757-1761/. Эти праймеры были названы P3 и P4 и показаны на фиг. 10. Праймеры располагаются на 159 нт в обе стороны и обеспечивают амплификацию фрагмента
200 нт. Используя праймеры P3 и P4 и изолированную ДНК BHV-1, были оптимизированы условия для процедуры ЦРП. Они включали, в частности, варьирование концентрации MgCl2,
концентрации глицерола и условий циклов. Оптимальный подобранный буфер для использования P3 и P4 для амплификации ДНК BHV-1 - 10 мМ Трис pH 8.0, 50 мМ KCl, 0.01% желатин, 2.6 мМ
MgCl2 и 20% глицерол. Оптимальные циклические условия /Perkin Elber Cetus DNA Thermal Cyclek/ для циклов 1-5: 1 мин 98oC, 30 сек 55oC и 45 сек 72oC и для
циклов 6-35: 30 сек 96oC, 30 сек 55oC и 45 сек 72oC. После амплификации ЦРП полученный фрагмент ДНК в 200 нт был подвергнут электрофорезу на 2% агарозном геле,
блотирован на нитроцеллюлозе и затем подвергнут блот-анализу. Проба, помеченная32p dCTP, используемая для блот-анализа, была фрагментом TagI 137 по, который располагался между связующими
участками праймера. /фиг. 10/. После авторадиографии гибридизированных фильтров можно наблюдать полосу в 200 по. С помощью этого пути амплификация только 10 BHV-1 геномов /приблизительно 1,5 •
1015 мкг ДНК/ ведет к хорошо обнаруживаемому сигналу /результат не показан/. Аналогичным образом была разработана процедура ЦРП с использованием праймеров, которые основаны на кодирующей
последовательности гликопротеида gIII BHV-1 /D.R. Fitzpatrick, Babink, Zamb, 1989, Virology, 1973, 46-57/. Для различения между ДНК BHV-1 дикого типа и вакциной мутанта с делецией gE образцы ДНК были
оба подвергнуты анализу gE-специфической ЦРП и gIll-специфической ЦРП. В такой пробе препарат ДНК Difivac-1 был выявлен как gIII положительный и gE отрицательный.
Поскольку обнаружение ДНК BHV-1 в бычьей сперме было бы важно для BHV-1 специфической процедуры ЦРП, была сделана попытка провести gE-специфическую ЦРП на бычьей сперме, зараженной BHV-1. Однако неизвестные компоненты в сперме обладают сильным тормозящим действием на цепную реакцию полимеразы /ЦРП/. Поэтому была разработана процедура изоляции ДНК BHV-1 от бычьей спермы. Чтобы изолировать ДНК от бычьей спермы, 30 мкл спермы инкубировалось 1 мг/мл протеиназы K /пК/ в общем объеме 300 мкл 0.15 NaCl, 0.5% Na-Sarkosil и 40 мМ ДТТ при 60oC. После 1 часа образец охлаждался до комнатной температуры и добавлялось 300 мкл 6М NaI и образец инкубировался 5 минут. Из этой смеси ДНК изолировалась стандартной экстракцией хлороформ/изоамилэтанол и осаждалась 1 объемом изопропанола. Осадок промывался 2.5 М NH4Ac/70% этанолом и повторно суспендировался в 10 мМ Трис pH 7.4, 1 мМ ЭДТК, 0.5% Tween 80 и 0.1 мг/мл пК для второй инкубации в течение 1 часа при 60oC. Этот препарат ДНК можно было непосредственно подвергать цепной реакции полимеразы.
Описание чертежей
Фиг. 1
Блот-анализ /Sodthern blot/ штаммов BH-1 Difivac-1 и Iowa.
A. Авторадиограмма блота Defivac-1 и Iowa геномной ДНК.
В дорожках 1 и 3 ДНК Difivac-1 применялась после переваривания рестрикционного фермента HindIII и PstI соответственно. В дорожках 2 и 4 ДНК Iowa применялась после переваривания рестрикционного фермента HindIII и PstI соответственно. Размер фрагментов указан в тысячах пар нуклеотидов /т.п.н./.
Вирусная ДНК изолировалась центрифугированием среды культуры /70 мл/ роллер-флакон в 450 см2/ с Эбтр клетками, зараженными вирусом в течение 2 часов через 25% /вес/вес/ сахарозную подушку, в 10 мМ Трис pH 7.4. 150 мМ NaCl и 1 мМ ЭТДК при 20 тыс. об/мин в SW 27 роторе ультрацентрифуге Бекмана L5-65. Из полученной таким образом гранулы вируса ДНК изолировалась в соответствии со стандартными методами /J.Sanbroon, Fretsch и T.Maniatis, 1989, Molecular Cloning: Lab. Man, 2-е изд. Cold Spring Harbor Laboratory Press, Naw York). На этой ДНК переваривание рестрикционного фермента проводится ферментами из Boehringer Mannheim в SURE /разрывными буферами, поставляемыми производителем/.
После отделения на 0,7% агарозном геле для горизонтального электрофореза и блотирования на нитроцеллюлозном фильтре /Shleicher & Schuell/ фильтр предварительно гибридизировался 6 часов при 42oC в 50% формамиде, 3 x SSC /1 • SSC = 0.15 М NaCl и 0.015 М Na-цитрата, pH 7.4/, ДНК 50 мкл денатурированной спермы лосося /Sigma/ мл и 0.02% альбумина бычьей сыворотки, 0.02% поливинил пирролидона и 0.02 фиколла и 0.1% а-додецилсульфата /SDS/. Затем гибридизация проводилась добавлением к тому же раствору HindIII K фрагмента, помеченного32p dCTP /Выбор HindIII K фрагмента основывался на клонировании и расщеплении участка на карте ДHК от коровьего герпеса вируса I /Штамм Купера/ Lohn F. Wayfield, Peter L. Good, Holly J. Vanoort, Aephonso R. Campboll и Dand A. Reed, Lournаl of Virology N 1983/ 259-264/. После 12-14 часов гибридизации фильтр промывался 2 часа в 0.1% SDS и 0.1% • SSC /растворе хлорида и цитрата натрия/ при 60oC. HindIII K фрагмент клонировался в pUC18 вектор в соответствии со стандартными процедурами клонирования /Sambrook, Fritsch Maniatis 1989, Moleculal Cloning: Lab. Manual, Inc 2-е изд. Cold Spring Habor Laboratory Press, N.-York. После переваривания Hind III клона HindIII K pUC18 вектор вновь был отделен от HindIII K фрагмента электрофорезом на 0.7% агарозном с низкой температурой плавлении /BRL, Life Technologies, / геле и изолирован от агарозы стандартной фенольной экстракцией и этаноловым осаждением. Изолированный HindIII К фрагмент был помечен меткой произвольной озатравки ДНК Kit 1004.760 от Boehringer Mannheim. Авторадиография гибридизированых фильтров проводилась с экспозицией 36 часов пленкой Kodak XAR при -70oC с использованием отражательного экрана.
B. Физические карты HindIII K фрагмента Iowa в 8.4 т.п.к. и HindIII фрагмента в Difivac-1 в 7.4 т.п.н. Ввиду комиграции PstI фрагментов в 6 т.п. н. и отсутствия PstI фрагмента в 1.8 т.п.н. в Didivac-I делеция должна происходить в заштрихованной области.
Фиг. 2
Субклонирование фрагментов BHV-1 дикого типа вокруг региона без Difivac-1
В A показаны компоненты генома BHV-1: Уникальный длинный регион /Uv/; Уникальный короткий регион /Us/ и два повтора /Iv и Tv/. Эта карта основана на опубликованном анализе штамма Купера /John F. Wayfield P.Y. Good, Holly J. Vanoort, A.R. Campboll и David A.Reed, Journal
of Virology /1983/ 259-264/.
В B показаны фрагменты из региона Us, которые были клонированы в прокариотные векторы: фрагмент EcoRI в 15.2 т.п.н. в pACYC, фрагмент HindIII в 8.4 т. п. н. в pUC18 и фрагмент EcoRI-HindIII в 2.7 и 4.1 т.п.н. в pBP322. Изоляция вирусных фрагментов ДНК проводилась в соответствии с процедурами, которые упомянуты в надписях фиг. 1A. Клонирование этих фрагментов в различные векторы проводилось в соответствии со стандартными процедурами /J. Sambrook, E. F. Fritsch и T.Waniatis 1989. Molecular Cloning: лабораторный протокол, 2-е изд. Cold spring Habor Laboratory Press, N.-York/.
В C показана физическая карта региона, где локализуется предполагаемая делеция в Difivac-1.
В D указаны некоторые субклоны этого региона, которые использовались для дальнейшего анализа. Два PstI фрагмента клонировались в pKUN19 и остальные фрагменты в pUC18.
Фиг. 3
A: Нуклеотидная последовательность
2027 нуклеотидов из Us региона штамма Lam BHV-1 вокруг предполагаемого местоположения, которое было делировано в Difivac-1, как показано на фиг. 2C [от участка опознания ALuI на крайнем
левом до участка опознания HincII на крайнем правом. Нуклеотидная последовательность во вставках субклонов, показанных на фиг. 2, была определена анализом на двух нитях с использованием метода
дидеокси последовательности Sanger и др. /F. Sanger, S. Nicklen и Coulson, 1977, Proc. Natl. Acad. Sci, 74, 5463-5467/. С этой целью использовались процедуры Т7 последовательности Pharmacia в
соответствии с процедурой, определенной производителем. Для радиоактивной пометки был использован [35S] ATP /Amersham/. Анализ последовательности регионов с газовой хроматографией /ГХ/ с
артефактами сжатия повторялся с 7-деаза-dGТР вариантом процедур Pharmacia. Указанная ниже нуклеотидная последовательность в трехбуквенном коде представляет собой последовательность /ак/ аминокислоты
открытой рамки считывания остатков 575 ак, которая была выявлена после концептуальной трансляции нуклеотидной последовательности. Эта трансляция основана на универсальном коде и была определена с
использованием компьютерной программы PC /Ген/ версия 1.03 PC/ген, ноябрь. 1987/. Эта открытая рамка считывания 575 ак начинается с метионина на нт 168 и заканчивается останавливающим кодоном на
нуклеотиде 1893.
Структурный анализ открытой рамки считывания остатков 575 ак также проводился компьютерной программой Pc/ген. Первые 26 ак образуют эукариотный экспортный сигнал, показанный на фиг. "сигнальным пептидом". Со счетом 6.2 расщепление последовательности этого сигнала предполагается между ак 26 и 27. Последовательность 575 ак имеет 3 возможных участка N-связанной гликосиляции /NXT/S/, указанных линией под остатками аминокислоты. В соответствии с методом Pao и Argos между ак 423 и ак 450 имеется трансмембранный регион, показанный на фиг. "трансмембранным витком". Последовательности опознавания /участки/ для рестрикционных ферментов AsuII, SmaI, HindIII и Econi подчеркнуты. Рассчитанный молекулярный вес этого полипептида - 61212.
B: Схематическое представление структурных характеристик открытой рамки считывания вышеупомянутой 575 ак.
Фиг. 4
Аминокислотное сравнение последовательности аминокислоты gE гена
BHB-I с последовательностью аминокислоты gE гена вируса простого герпеса /ВПГ/ и другими E гомологичными генами gI вируса pseudoralies и др. I varicella - zoster (VZV).
- Последовательности, используемые для этого сравнения, брались из следующих публикаций: HSV: Seguence determination and genetiе content od the short unique region in the genome of herpes Simplex virus type I D.Y. Me Geoch, A. Dolan, S. Donald и F.Y. Rixon (1985) Jornal Mol. Biol 181, 1-13, vsv: DNA sequence of the Us component of the varicella - zoster virus genome. A.J. Davidson (1983), EMBO Journal 2.2203-2209. PRV: use of Agt II to usalate genes for 2 ps. rabus v. grey coprotoid, Petnovskis и др. (1986). J. of Virology GO, 1985-193].
Эти последовательности сравнивались с использованием программы анализа последовательности Miltalin/Corpet, 1988, Nucl. Acids Res 16, 10881-10890/.
На A показана диаграмма, на которой все последовательности четырех аминокислот показаны схематически. Здесь предсказанные трансмембранные части /TM/ показаны друг под другом. Кроме предсказанных экспортных сигнальных последовательностей и возможных N-связанных участков гликосиляции /I/ показаны две законсервированные области, в которых относительная позиция остатков цистеина часто остается неизменной /CCC/.
В B показаны результаты Multalin сравнения центрально расположенного региона, богатого цистеином, четырех версий gE. Звездочки показывают идентичные аминокислоты и двоеточия аналогичные аминокислоты.
Фиг. 5
Рисунок фотографий, полученных в блот-анализе Difivac-1 и
Панель A: Геномные ДНК перевариваний рестрикционного фермента Difivaс-1 и Iowa с помощью BstI /1, 2/, EcoPI
/3, 4/ и HindIII /5, 6/, отделенные на 0.7% агарозном геле, блотированные на нитроцеллюлозе и гибридизированные HindIII K фрагментом с пометкой32p штамма Lam BHV-1, в соответствии с
процедурами, определенными в надписях фиг. 1A.
Панель B: Нитроцеллюлозный блот того же геля, что и в A, гибридизированного gE-специфическим зондом p318 BHV-1. Этот зонд включает целый AluI-HincIII регион, показанный на фиг. 2C.
Фиг. 6
Построение фрагмента BHV-I с gE делецией
В A показана позиция gE гена и используемых клонов. Компонентами генома
BHV-I являются: Уникальный длинный /Ui/ регион; Уникальный короткий /Us/ регион и два повтора /IP и TP/. Чтобы получить регион, расположенный на 5' стороне gE гена, PstI-SmaI
фрагмент в 1.4 т.п.н. от HindIII K фрагмента ВНV-1 в 8.4 т. п. н. штамма Lam был субклонирован в участок SmaI и PstI плазмиды PuC18. Этот клон был назван p515, и он показан в B. EcoNI-SmaI фрагмент,
расположенный на 3' стороне gE, исходящий из HindIII-EcoRI клона в 4.1 т. п. н., был клонирован в уникальный AUSII участок p515. Чтобы сделать возможным лигирование остатка EcoNI к остатку ASUII клон
p515 был переварен ASUII, затем обработан ферментом Кленова /Boehringer Mann/ и dCTR, чтобы получить один цитозиновый остаток в остальной части ASUII /Sanbrook и др. 1989/. Этот дополнительный цитозин
показан звездочкой в D. Затем p515 был также переварен ферментом SmaI, после чего EcoRI фрагмент можно было лигировать в этот вектор. Построенный таким образом клон был назван p519.
Фиг. 7
A. Рисунок фотографий, полученных в блот-анализе препаратов ДНК IB7, IB8 и 2H10. Изоляция ДНК, переваривания рестрикционного фермента, блотирование и гибридизация были проведены в
соответствии с процедурами, описанными на фиг. 1A. После двойного переваривания препаратов ДНК 1B7, 1B8 и 2H10 с помощью PstI-DraI фрагменты были отделены на 0.7% агарозном геле и затем блотированы на
нитроцеллюлозном фильтре. Этот фильтр гибридизировался32р dCTP, помеченный в качестве зонда фрагментом делеции PstI-DraI в 2.3 т.п.н. В рядах 1-3 соответственно были отделены образцы 1ВГ,
1B8 и 2H10. В ряде 4 применялась ДНК BHV-1 дикого типа штамма Lam и в ряде 5 - фрагмент делеции в 2.3 т.п.н.
B. Физическая карта EcoRI фрагмента штамма Lam BHV-I в 15,2 т.п.н. Карта показывает позицию участков опознавания PstI, DraI и HindIII и позицию зонда гибридизации, упомянутого в 7A.
Фиг. 8
Прокариотная экспрессия gE BHV-I
Для прокариотной
экспрессии gE BHV-I фрагмент SmaI gE гена в 600 по был слит в трех рамках считывания с кодирующим регионом гена глютатион-S-трансферазы от Schistosoma japonicum в векторе pGEX-2T/ D.B.Smith, K.
Johnson, Gege 67 /1988/ 31-40/. Рекомбинантные молекулы с нужной ориентацией (Syn) фрагмента опознавались посредством анализа рестрикционного фермента с использованием стандартных методов. Клоны DH5
α E.Coli с таким построением слиянием были названы pGEX-2T600SI, PGEX-2T600S 2 и рGЕХ-2T600S3.
A. Диаграмма одной из конструкций pGEX-2T600S. На NH2 стороне региона, который кодирует для продукта слияния GST-gE, располагается регион tac промотора, индуцируемый изопропилтиогалактозидом /ИПТГ/.
B. Изображение фотографий, полученных в блот-анализе всех препаратов белка клеток DH5 α, трансформированных pGEX-2T600S. Ночные культуры клеток DH5 α, трансфицированные конструкциями pGEX-2T600S1, pGEX-2T600S2 и pGEX-2T600S3, пребывали 1/10 в среде Luria - Bertani /LB/ с 50 мкг/мл ампициллина и после 1 часа роста, индуцированного ИПТГ- 5 часов. Эти индуцированные культуры были центрифугированы в течение 5 минут при 6000 х g и включены в 1 х смешанный слой /2% SDS /додецилсульфат натрия/, 10% глицерол, 5% меркаптоэтанол и 0.01% бромофенол голубой/ [1.5 мл культуры включалось в 500 мкл смешанного слоя и нагревались при 95oC 5 минут. Затем 50 мкл на дорожку отделялось на вертикальном 12.5% полиакриламидном геле в соответствии со стандартными процедурами и затем в полусухом состоянии блотировались на нитроцеллюлозный фильтр с использованием системы LKB-miltiphor II Nova Blot при условиях, указанных производителем.
В дорожках M применялся предварительно окрашенный маркерный белок /BRL Life Techol. Inc. 236к, 112к, 71к, 44к, 28к, 18к и 15к/ и в дорожках 1,2,3-препараты общего белка клеток DH5α, трансфицированных тремя соответствующими рамками: pGEX-2T600S1, pGEX-2T600S2 и pGEX-2T600S3.
На панели A можно видеть результат блот-анализа антисыворотки. С этой целью фильтр инкубировался в соответствии со стандартными процедурами /E Hallow, Lane, 1986, Antibodies: Cold Spring Harbor Laboratory, N.-York/ в блокирующем буфере /PBS + 2% молочный порошок и 0.05% Twecn 20/, а затем поликлональной анти-GST сывороткой кролика. Затем фильтр промывался и инкубировался козьей-анти-кроличьей иммуноглобулиновой сывороткой, конъюгированной пероксидазой хрена обыкновенного. Затем связанные козьи антитела иммунохимически обнаруживались хроматогеном /диаминобензидин, хлоронафтол и H2O2/. Продукт слияния GST, который указывается стрелкой, имеет предсказанный размер приблизительно 47 к только в рамке 3.
На панели B можно видеть результат блот-анализа с моноклональным антителом MAT 4, которое опознает gE белок. С этой целью дупло фильтр, как на панели A, был блокирован, инкубирован MAT, промыт и инкубирован козьей-анти-кроличьей иммуноглобулиновой сывороткой, конъюгированной пероксидазой хрена обыкновенного /ПоХО/. Затем связанные кроличьи антитела иммунохимически обнаруживались хромогеном. Полоса, которая видна в дорожке 3 /рамка 3/, составляет 47 т в размере и указана стрелкой.
Фиг. 9
Построение pEVHIS gE плазмиды для эукариотной экспрессии gE гена BGV-I.
Для эукариотной экспрессии gE гена весь регион gE кодирования клонировался в соответствующей ориентации за регионом промотора HCMV вектора экспрессии pEVHIS, используя стандартные процедуры /Sambrook и др. 1989/. С этой целью AIUI фрагмент с 394 по, который начинается на 55 по до открытой рамки считывания gE, был клонирован в pUC18 и назван p201. Затем после переваривания p201 посредством HincII, HincII фрагмент с 1740 по, который включает большую часть gE гена, был клонирован в p201. Это дало плазмиду p318, которая в полилинкере pUC18 включает всю область кодирования gE от AIUI участка на 55 по до начального кодона gE до HincII участка на 133 по за останавливающим кодоном gE. Используя участки рестрикционного фермента в полилинкере вектора, этот фрагмент был вырезан из p318 ферментами BamHI и SphI. Вначале p318 был переварен SphI и затем участок SphI был заполнен использованием полимеразы Кленова и dNTP. После переваривания BamHI вставка в 1.9 т.п.н. была выделена из pUC18 вектора в агарозу с низкой температурой плавления и лигирована в pEVHIS вектор, который был переварен BamHI и ScoRV с этой целью. Полученная таким образом плазмида была названа pEVHIS/gE.
Фиг. 10
Позиция gE-специфических праймеров и зонда для процедуры ЦРП для обнаружения ДНК BHV-1
На схеме показана последовательность нуклеиновой кислоты gE гена
гликопротеина BHV-1 из нуклеотида 1272-2027 [последовательность была взята с фиг. 3] . Праймеры, используемые для gE-специфической процедуры ЦРП, были названы P3 и P4. Участки
связывания праймера для P3 и P4 подчеркнуты. Нуклеотидная последовательность P3-5'-ACG-TGG-TGG-TGC-CAG-TTA-GC-3' /Посл. ID N:2/. Нуклеотидная последовательность P4/дополнительно к связующей последовательности праймера, определенной выше/ -5'-ACC-AAA-CTT-TGA-ACC-CAG-AGC-G-3' /Посл. ID N: 3/. Зонд, который использовался для гибридизации блота для
обнаружения амплифицированной ЦРП ДНК, представлял собой TagI фрагмент в 137 по, расположенный между участками связывания праймера, концы этого фрагмента указаны. Для сравнения с фиг. 3 указаны также
участки HindIII и EconI.
Фиг. 11
Картирование gE делеции Difivac-1
На A показана физическая карта EcoRI фрагмента в 15.5 т.п.н. штамма Lam BHV-1 дикого типа. На B
показана физическая карта EcoRI фрагмента в 14.5 т. п. н. Difivac-1. Оба EcoRI фрагмента покрывают полные уникальные короткие регионы геномов соответствующих вирусов. Позиция gE гена и предполагаемая
позиция gI гена указаны открытыми боксами. Карты A и B расположены таким образом, что фрагменты PstI в 6 т.п.н. внутри каждой карты выравнены. На обоих картах последовательности внутреннего повтора и
концевых повторов были указаны заштрихованными боксами. Стрелки под повторами показывают ориентацию этих последовательностей.
В A была указана часть Us региона, которая отсутствует в штамме Difivac-1.
C показывает позицию клонированных Difivac-1 фрагментов, используемых для картирования gE делеции и получения физической карты> показанной в B. Стрелки под вставками клонов p728, p737 и p754 указывают регионы, которые имели порядок следования, чтобы определить рекомбинационную точку.
Сокращения:
A = AliI, E = EcoRI,
P = PstI, H = HindIII, r - рекомбинационная точка, IR = внутренний повтор, TR = концевой повтор.
Фиг. 12
Определение точной рекомбинационной точки в Us регионе
Difivac-1
Чтобы определить точные границы E делеции, выявленной в штамме Difivac-1, был определен порядок следования клона p754 и концов клонов p728 и p737. Вставки этих клонов были указаны
на фиг. 11. Используемые процедуры последовательности были описаны в пояснениях фиг. 3.
На A была показана последовательность большей части AluI-PatI фрагмента. Эта последовательность начинается в регионе промотора gE гена. Предполагаемый ТАТА бокс подчеркнут. В точке r / = рекомбинационной точке/ этот регион промотора сливается с последовательностью, также выявленной на противоположном участке Us региона, и называется инвертированный повтор. Точная рекомбинационная точка была определена сравнением повтора, выявленного на регионе gE промотора, с копией повтора, выявленного на противоположном участке Us региона. Точка, где эти последовательности расходятся, была указана в B /под I/ обозначением "r". Подобное сравнение было сделано с последовательностью gE промотора, выявленной в Difivac-1, и gE промотора, выявленной в штамме Lam дикого типа. Точка, где эти последовательности расходятся, была показана в B /под II/ и также помечена "r". Выявленные точки рекомбинации те же самые.
Фиг. 13
Анализ частичной последовательности gE гена BHV-1
С использованием PstI клона штамма BHV-1 в 1.8 т.п.к.,
который попадает и в gI, и gE ген BHV-1 /см. фиг. 11/, была определена последовательность 284 нуклеотидов внутри кодирующего региона BHV-1 gI. Используемые процедуры последовательности были описаны в
пояснениях к фиг. 3. Транслируемая последовательность основывалась на универсальном коде версий 1.03 компьютерной программы PC/ген /ноябрь 1987/.
Последовательность аминокислоты, кодируемой второй рамкой считывания, дается в однобуквенном коде ниже нуклеотидной последовательности. Эта последовательность аминокислоты гомологична кодирующему региону других gI гомологов вируса герпеса /См. фиг. 14/.
Фиг. 14
Сравнение аминокислоты частичной последовательности аминокислоты предполагаемого gI гена BHV-1 с соответствующими частями кодирующих регионов
gI гена вируса простого герпеса, др 63 гена вируса (PRV) psudorabies и др. IV гена вируса varecella-joster (vzv).
Последовательность PRV начинается у аминокислоты 82, HSVI последовательность начинается у ак 80 и последовательность vzv начинается у ак 76 их соответствующих кодирующих регионов. Используемая последовательность была опубликована в документах, упомянутых в пояснениях к фиг. 4. Сравнение проводилось с использованием Miltalen компьютерной программы. Звездочки указывают идентичные аминокислоты, и двоеточия указывают аналогичные аминокислоты.
Фиг. 15
Построение MSV neo GI плазмиды для эукариотной экспрессии gI гена BHV-1
На основании сравнения аминокислоты частичной последовательности gI гена BHV-1 была
рассчитана предполагаемая позиция gI гена BHV-1. Основываясь на этом расчете, предполагалось, что SmaI фрагмент в 1.7 т.п.н. должен содержать полный кодирующий регион gE гена BHV-1. Позиция этого SmaI
фрагмента была указана в A. К тупым концам этого SmaI фрагмента в 1.7 т.п.н. были лигированы BamHI линкеры с использованием стандартных процедур. Полученный продукт был переварен BamHI и лигирован в
вектор MSV-neo эукариотной экспрессии. Вектор MSV neo имеет уникальный участок BamHI за MSV-LTR, который обладает сильной активностью промотора. Этот вектор был описан в Rijsewijk и др. 1987 EMBO Y. 6,
127-131.
Фиг. 16
Построение фрагмента двойной делеции gI/gE BHV-1
Позиция гликопротеин gE гена и предполагаемая позиция гликопротеид gI гена в Us регионе
BHV-1 изображена на диаграме A. Заштрихованные блоки показывают повторы, которые граничат с регионом Us. B показывает физическую карту некоторых значительных участков рестрикционного
фермента по отношению к позиции обеих генов. Чтобы построить фрагмент делеций gI/gE, клон pl.r-SmaI/o, содержащий SmaI фрагмент, в 1.7 т.п.н., который охватывает gI ген, будет переварен PstI. Участок
PstI остающейся вставки SmaI-PstI в 350 по будет сделан с затупленными концами с использованием стандартных молекулярно-биологических процедур. EcoNI-SmaI фрагмент /См. фиг. 6В/, изолированный от
фрагмента HindIII-EcoRI в 4.1 т.п.н., описанный на фиг. 6А, также будет сделан с затупленными концами и лигирован к модифицированному PI участку. Это показано на диаграмме в C и D. Из полученного
клона pAIE фрагмент SmaI-DraI в 1.4 т.п.н. может быть изолирован для рекомбинации с ДНК BHV-1 дикого типа.
Сокращения:
E = EcoRI, H = HindIII, S = SmaI, P = PstI, ENI = EcoNI,
D = DraI, kb = тысяча пар нуклеотидов /т.п.н./ и Us = уникальный короткий.
Фиг. 17
Среднее количество вируса, выделяющееся из носа телят после вакцинации
. = = вакцинированные Difivac-1, 0 = не вакцинированные контрольные телята.
Фиг. 18
Средние ежедневные клинические проявления у телят после контрольного заражения вирулентным
штаммом BHV-1, ключевые обозначения, как на фиг. 17.
Фиг. 19
Средняя ректальная температура телят, с контрольным заражением вирулентным штаммом BHV-1, ключевые обозначения,
как на фиг. 17.
Фиг. 20
Средний рост телят после контрольного заражения вирулентным штаммом BHV-1, ключевые обозначения, как на фиг. 17.
Фиг. 21
Среднее количество вируса, выделяющееся из носа телят после контрольного заражения вирулентным штаммом BHV-1, ключевые обозначения, как на фиг. 17.
Фиг. 22
Средняя ректальная
температура телят после контрольного заражения вирулентным штаммом BHV-1
. = вакцинированные Lam gE-, 0 = вакцинированные Lam gE-/TK-,
x =
невакцинированные контрольные телята.
Фиг. 23
Средний рост телят после контрольного заражения вирулентным штаммом BHV-1, ключевые обозначения, как на фиг. 22.
Фиг. 24
Средние ежедневные клинические проявления у телят после контрольного заряжения вирулентным штаммом, ключевые обозначения, как на фиг. 22.
Реферат
Изобретение относится к области вакцинации и диагностики заболеваний К. Р. С. , вызываемых патогенными бактериями, в частности к живым ослабленным и инактивированным вакцинам для защиты животных против вируса герпеса типа 1. Вакцина создана на основе мутанта вируса герпеса BHV-I, имеющего делецию в gЕ-гене гликопротеида, с геном, имеющим соответствующие местоположение в геноме BHV-I, причем указанная делеция позволяет мутанту отличаться серологически от BHV-I дикого типа. Изобретение касается также диагностических наборов для обнаружения нуклеиновой кислоты BHV-I, антител, специфичных для BHV-I, белка BHV-I и способа определения инфекции BHV-I у животных. Разработанные препараты и способ отличаются высокой эффективностью и безопасностью. 5 с.п.ф-лы, 24 ил. 2 табл.
Комментарии