Коллекция и способы ее применения - RU2569187C2
Код документа: RU2569187C2
Чертежи







































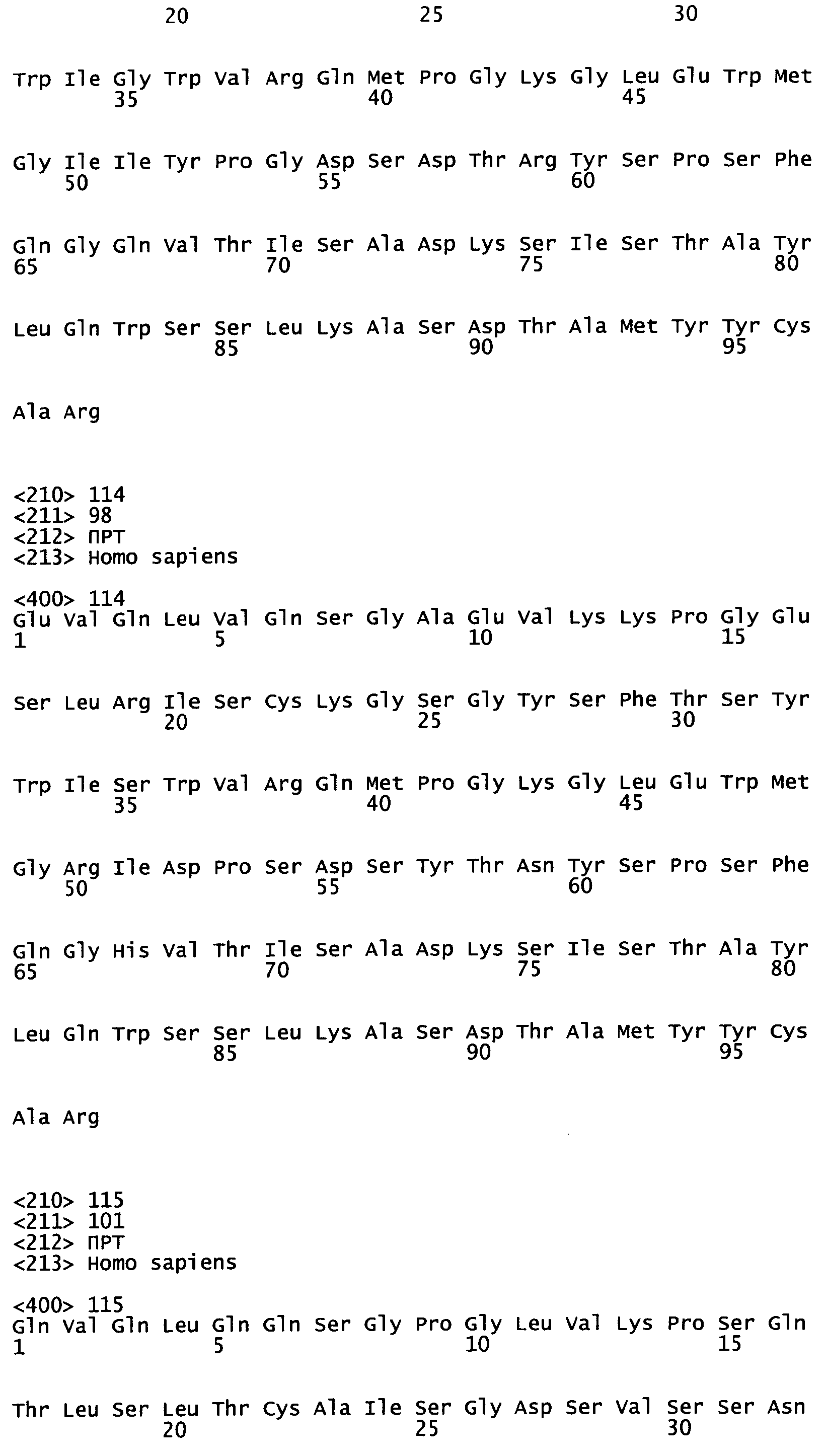

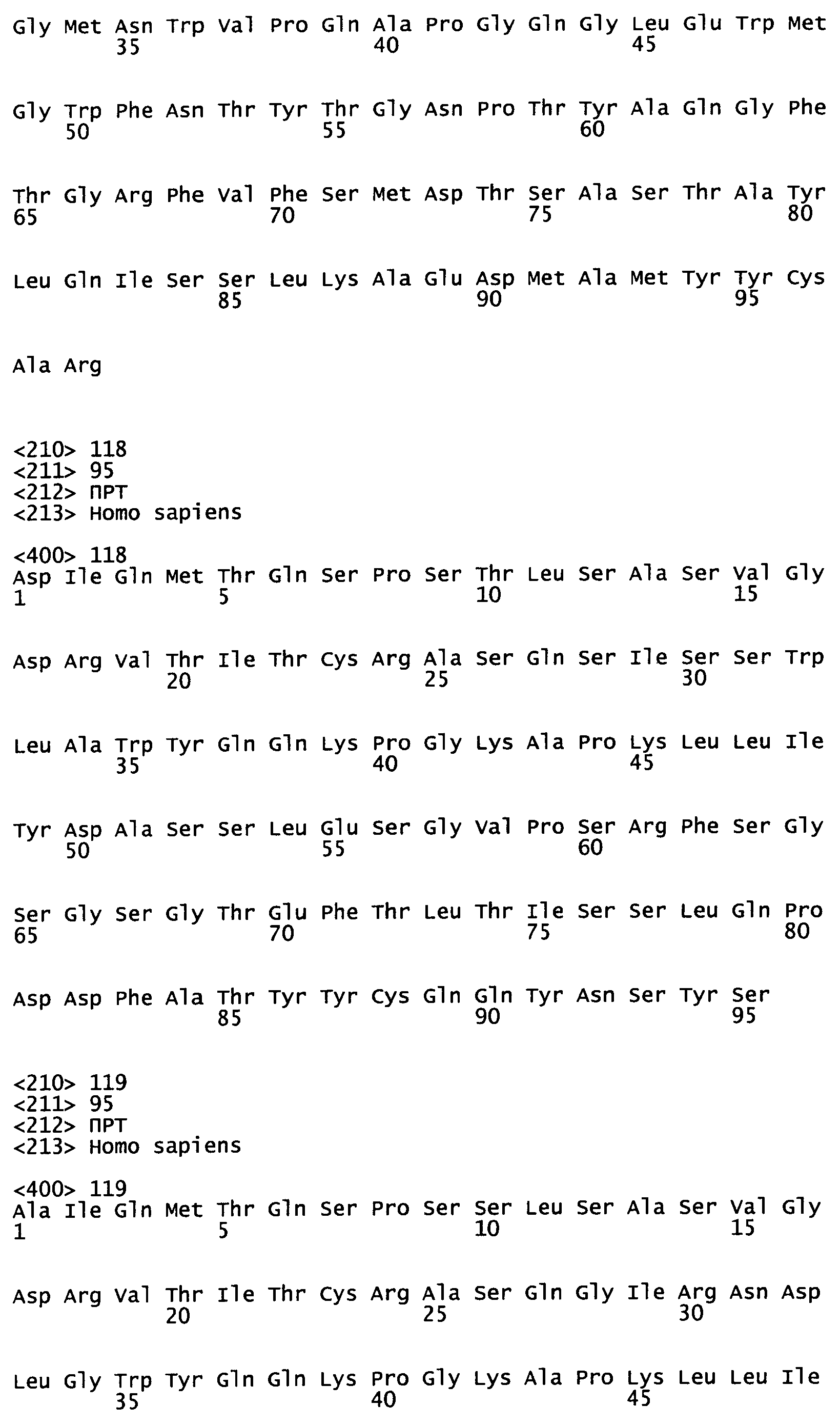





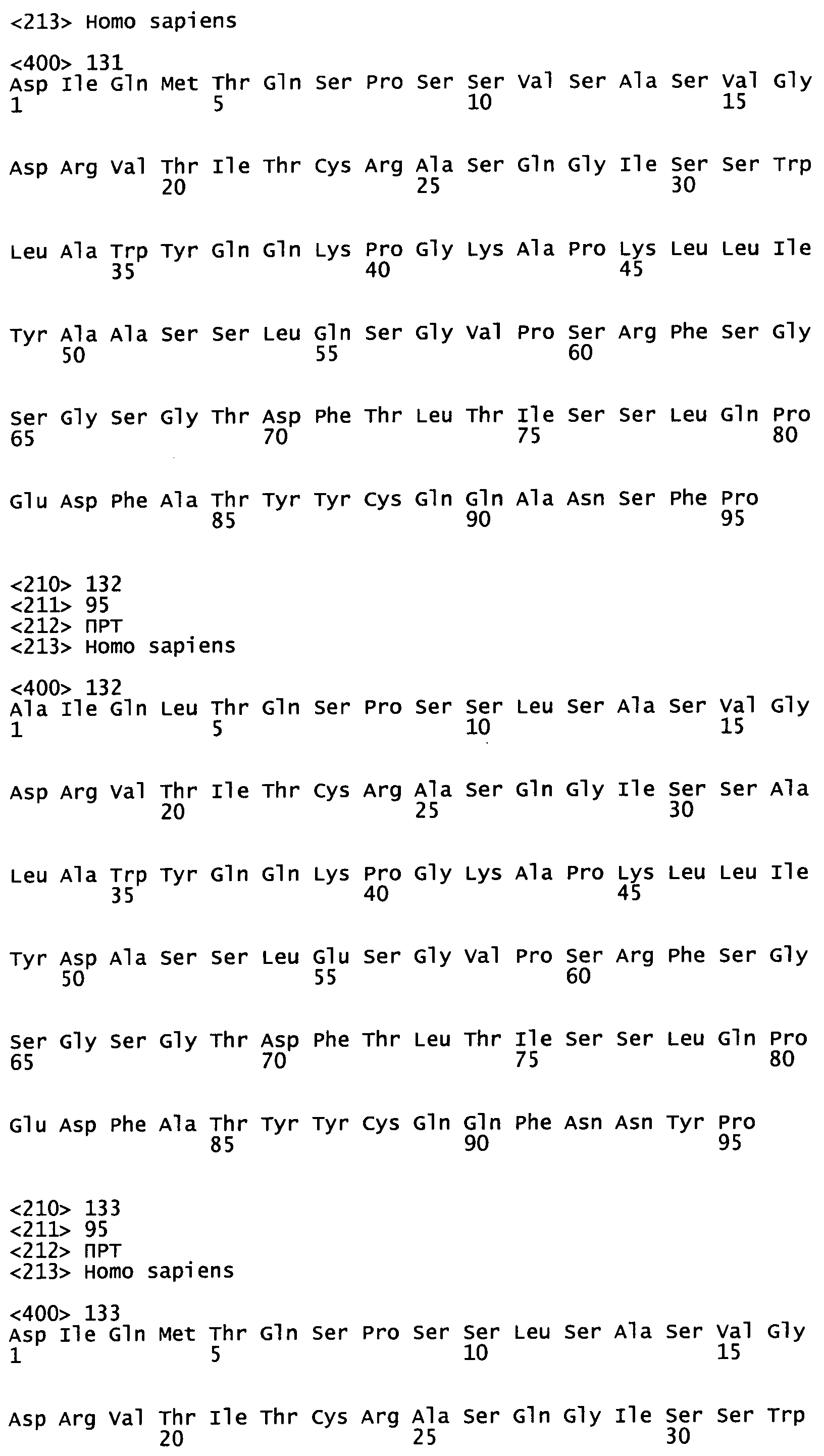



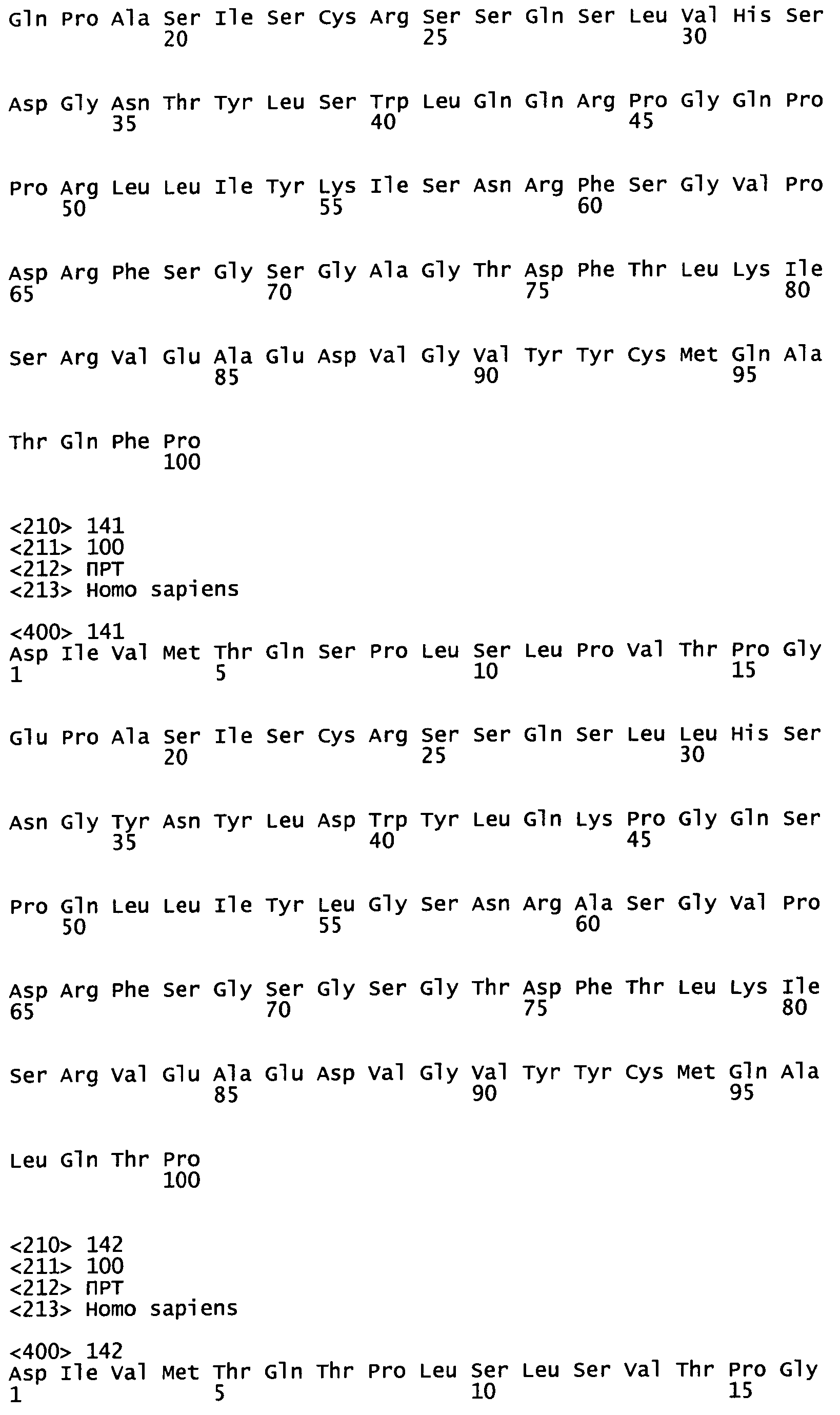

























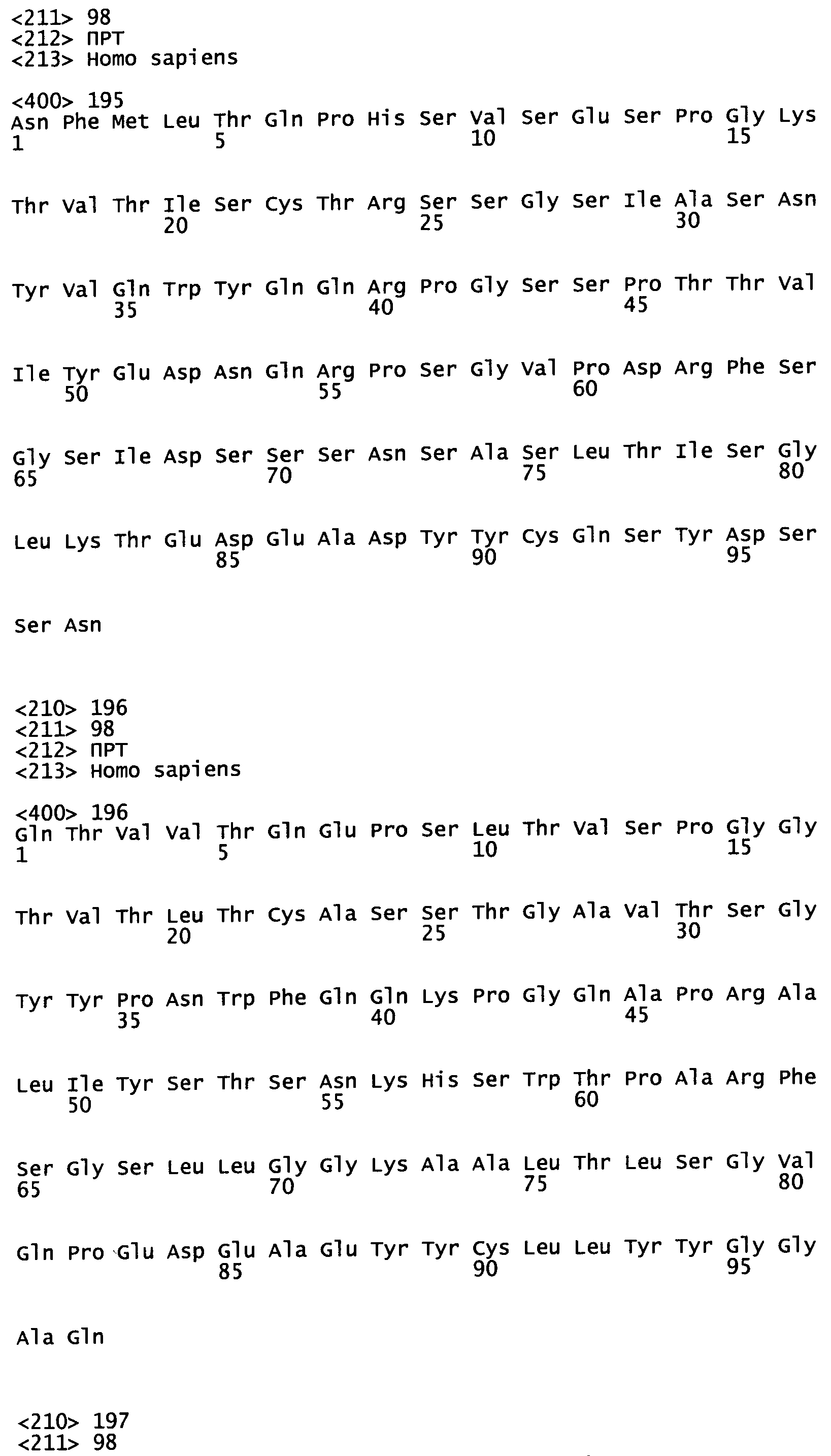































































Описание
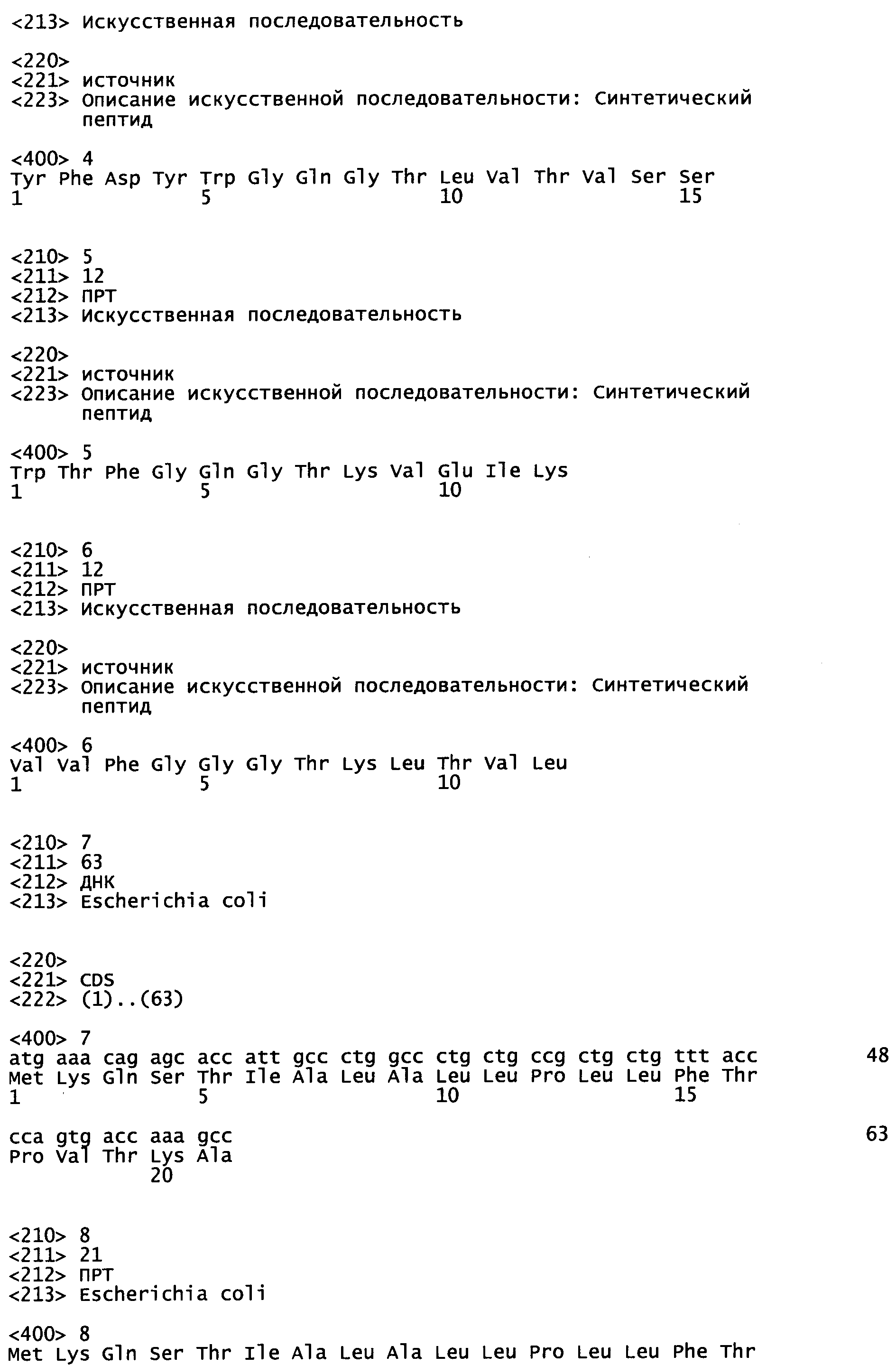
В данной заявке заявлен приоритет согласно США предварительной заявке, серийный номер 61/299401, поданной 29 января 2010 г. и США предварительной заявке, серийный номер 61/182350, поданной 29 мая 2009 г., которые включены в данную заявку посредством ссылки во всей своей полноте.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Достижения в разработке фармацевтических средств, особенно в области терапевтических антител, быстро обеспечивают и/или улучшают лечение многих заболеваний. Эти достижения, благодаря охвату новых целевых областей и обеспечения новых механизмов действия, приводят к значительному улучшению качества жизни пациентов даже с самыми тяжелыми и сложными заболеваниями. Одна из проблем для системы здравоохранения, s целом, и пациентов, в частности, заключается в том, что стоимость новых лекарственных средств, обеспечиваемых этими фармацевтическими достижениями, также быстро увеличивается. Высокая стоимость является результатом капиталовложений, необходимых для разработки фармацевтических средств, особенно антител, которые в настоящее время превышают один биллион долларов на рыночный продукт. Высокий риск неудачи при разработке и очень продолжительные сроки разработки делают эти вложения неизбежными. С момента идентификации потенциального терапевтического антитела и до тех пор, когда оно достигнет рынка и сможет приносить пользу пациентам, может пройти более пятнадцати лет. Каждая стадия разработки от идентификации, доклинической, клинической до выхода на рынок пестрит проблемами и рисками. Фармацевтические компании постоянно оценивают, как уменьшить стоимость разработки путем снижения сроков и рисков неудачи для того, чтобы быстро предложить пациентам наиболее эффективные лекарственные средства и для того, чтобы сделать их более доступными.
В следующем описании представлено важное достижение, которое позволяет быстрее идентифицировать оптимальные терапевтические антитела для лечения, возможно, любого заболевания. Терапевтические антитела-кандидаты должны удовлетворять целому ряду критериев разработки для продвижения на рынок, таких как долговременная стабильность и высокие выходы экспрессии. Описанное достижение увеличивает вероятность и скорость идентификации антитела, которое может удовлетворять всем строгим критериям разработки с самого начала. Полученное в результате антитело будет менее дорогостоящим при производстве и будет эффективным и безопасным при лечении многочисленных заболеваний.
Хорошо известный способ идентификации терапевтических антител осуществляется посредством применения технологии фагового дисплея. В фаговом дисплее используют вирусоподобные частицы, которые выращивают в бактериях для экпонирования антител. Одно из преимуществ этой технологии заключается в том, что используемые библиотеки являются огромными, вплоть до 1×1010 антител, которые можно быстро протестировать на связывание с любой мишенью, связанной с любым заболеванием. См., например, Knappik et al., (2000), "Fully synthetic human combinatorial antibody libraries (HuCAL) based on modular consensus frameworks and CDRs randomized with trinucleotides", J. Mol. Вiоl. 11; 296(1):57-86. Преимущество работы с такими большими количествами заключается в том, что результат скрининга против мишени может выявить сотни антител, которые связываются с терапевтической мишенью и которые все могут быть терапевтически значимыми. Проблема, однако, заключается в том, что часто только несколько из этих антител можно разрабатывать, что означает, что они могут удовлетворять всем строгим критериям, требующимся для продвижения на рынок.
Для того чтобы для новой фаговой дисплейной библиотеки быстро сократить сроки идентификации и уменьшить неотъемлемые риски, данная библиотека должна содержать антитела, имеющие свойства, которые необходимы для селекции и клинической разработки и которые приводят к безопасному и эффективному лечению пациентов. Такие свойства включают: 1) высокие уровни фагового дисплея, с тем чтобы все без исключения антитела коллекции можно было протестировать против мишени, представляющей интерес; 2) высокие уровни экспрессии, обеспечивающие эффективное воспроизводство антитела или фрагмента; 3) высокую термостабильность, с тем чтобы антитело могло достичь пациента в эффективной форме; 4) высокую стабильность в сыворотке, с тем чтобы антитело могло сохраняться в организме в течение терапевтически значимого времени; 5) низкий риск иммуногенности, увеличивающей тем самым безопасность, и 5) высокое разнообразие, с тем чтобы одну библиотеку можно было использовать для идентификации антител против любой терапевтической мишени.
Библиотека, которая по существу имитирует человеческую иммунную систему, должна представлять собой чрезвычайно полезное или даже оптимальное решение. Человеческая иммунная система состоит из антител, кодируемых генами эмбрионального типа. Антитела, в частности, содержат вариабельную область тяжелой цепи и вариабельную область легкой цепи. Имеются приблизительно 50 генов вариабельных областей тяжелых цепей эмбрионального типа и приблизительно 50 генов вариабельных областей легких цепей эмбрионального типа, которые при объединении обеспечивают создание примерно 2500 комбинаций пар различных вариабельных областей тяжелых и легких цепей. Считают, что у людей продуцируются все 2500 этих комбинаций. Однако обнаружили, что некоторые вариабельные области тяжелых цепей, вариабельные области легких цепей и/или комбинации (пары) вариабельных областей тяжелых и легких цепей экспрессируются на более высоком уровне, чем другие. Было высказано предположение, что должна быть причина, почему некоторые цепи экспрессируются больше, чем другие, и если это так, то высоко экспрессируемые гены эмбрионального типа могут обладать благоприятными функциональными свойствами. Поэтому один из путей получения библиотеки антител, имеющих благоприятные функциональные свойства, заключается в создании библиотеки, содержащей избыток вариабельных областей тяжелых цепей, вариабельных областей легких цепей и/или пар вариабельных областей тяжелых цепей и вариабельных областей легких цепей эмбрионального типа из иммунного репертуара человека.
Кроме того, считают, что генные последовательности эмбрионального типа, присутствующие у людей, имеют очень низкую иммуногенность, поэтому, по очевидным причинам, эти последовательности можно имитировать в рекомбинантных антителах с целью снижения риска иммуногенности.
Были предприняты разные подходы для оценки спариваний генов вариабельных областей тяжелых и легких цепей эмбрионального типа, преобладающих в иммунном репертуаре человека. См. de Wildt et at., Analysis of heavy and light chain pairings indicates that receptor editing shapes the human antibody repertoire, J Mol Biol. 22;285(3):895-901 (January 1999), который включен посредством ссылки во всей своей полноте. Wildt с соавт.брал образцы крови от людей-доноров, сортировал IgG+В-клетки, которые претерпевали соматическую гипермутацию, амплифицировал кДНК с помощью ПЦР (полимеразная цепная реакция), секвенировал каждую кДНК и выравнивал каждую последовательность с известными человеческими генами вариабельных доменов эмбрионального типа. Wildt с соавт.наблюдал, что только несколько генов эмбрионального типа доминировали в иммунном репертуаре и что обычно экспрессирующиеся сегменты генов тяжелых и легких цепей часто спариваются.
Также были предприняты попытки поддержания спариваний вариабельных доменов тяжелых и легких цепей индивидуальных В-клеток. Например, были описаны библиотеки "когнатных пар" вариабельных доменов. См. Meijer et al., Isolation of human antibody repertoires with preservation of the natural heavy and light chain pairing, J Mol Biol., 358(3):764-72 (May 5 2006); и WO 2005042774. Библиотеки согласно методам, описанным в Meijer et al., были созданы из индивидуальных В-клеток иммунизированного хозяина. Как правило, В-клетки сортируют с помощью FACS (клеточный сортер с возбуждением флуоресценции) таким образом, чтобы отобрать CDSS4' В-клетки, которые представляют собой соматически гипермутировавшие клетки, амплифицируют их кДНК с помощью ПЦР и продукты генов антител встраивают в Fab-векторы для селекции. Такие библиотеки когнатных пар также включают ограничения. Например, хозяева, обеспечивающие В-клетки, обычно иммунизированы, и сортированные популяции В-клеток были гипермутированными, поэтому полученные библиотеки ориентированы на конкретный иммуноген.
Кроме того, были предприняты попытки применения основных вариабельных областей тяжелых цепей или вариабельных областей легких цепей для создания библиотеки. Например, в Shi et al., "De Novo Selection of High-Affinity Antibodies from Synthetic Fab Libraries Displayed on Phage as pIX Fusion Proteins; J Mol Biol., 397(2):385-96 (26 марта 2010 г.) (который не считается предшествующим уровнем техники в отношении настоящего изобретения) и в соответствующей патентной завке WO 2009085462; и WO 2006014498, белковые последовательности вариабельной области тяжелой цепи или вариабельной области легкой цепи эмбрионального типа были включены в библиотеки на основании частоты использования в иммунном репертуаре человека.
Были также предприняты дополнительные попытки, с помощью которых включали специфическую пару эмбрионального типа в библиотеку. Например, в WO 1999020749 описана библиотека, в которой ее члены содержат тяжелые цепи, имеющие каноническую структуру гипервариабельной петли, кодируемой человеческим генным сегментом DP-47 тяжелой цепи эмбрионального типа (IGHV3-23), и/или каркасные участки, кодируемые геном эмбрионального типа, и/или легкие цепи, имеющие каноническую структуру гипервариабельной петли, кодируемые человеческим генным сегментом 02/012 легкой цепи эмбрионального типа (IGKV1-39/1D-39), и/или каркасные участки, кодируемые геном эмбрионального типа.
С использованием дополнительных подходов были созданы библиотеки непосредственно или происходящие из В-клеток. Например, Glanville et al., Precise Determination of the Diversity of a Combinatorial Antibody Library Gives Insight into the Human Immunoglobulin Repertoire, Proc NatI Acad Sci 1;106(48):20216-21 (December 2009) (который не считается предшествующим уровнем техники в отношении настоящего изобретения), в котором описана библиотека антител, построенных на разнообразии 654 человеческих донорских репертуаров иммуноглобулинов М (IgM). В частности, кДНК V-генов тяжелых и легких цепей от 654 человек-доноров раздельно амплифицировали с помощью ПЦР (выделяя пару вариабельных областей тяжелых и легких цепей) и домены тяжелых и легких цепей затем подвергали случайной реассоциации. В WO 2003052416 описано выделение В-клеток из хозяина, демонстрирующего выраженный ответ на интересующий патоген, возникающий либо в результате инфекции микроорганизмом, либо в результате лечения вакциной. В WO 2003052416, кДНК, кодирующую CDR3-участок вариабельных областей, секвенировали и конструировали фрагменты антитела, содержащие доминантные CDR3. В WO 2009100896 описано выделение В-клеток из иммунизированного хозяина, где кДНК, кодирующие вариабельные области тяжелых и легких цепей, секвенировали и определяли относительное количество неспаренных последовательностей вариабельных областей тяжелых и вариабельных областей легких цепей. В WO 2009100896 (который не считается предшествующим уровнем техники в отношении настоящего изобретения), синтезировали библиотеки, содержащие случайно рекомбинирующие вариабельные тяжелые и вариабельные легкие цепи, где антитела были специфичны к одному иммуногену. Их краткое описание и дополнительные подходы можно найти в Fun et al., Synthetic antibodies as therapeutics, Expert Opin Biol Ther., 7(1):73-87 (January 2007).
Следовательно, существует потребность в коллекции антител или их фрагментов, которые включают пары генов вариабельных областей тяжелых цепей и вариабельных областей легких цепей эмбрионального типа, экспрессирующиеся в иммунном репертуаре человека, которые имеют подходящие биофизические свойства, приводящие к созданию легко разрабатываемых антител, которые безопасны и эффективны при применении у пациентов. Настоящее изобретение удовлетворяет этим и другим потребностям.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложено полезное решение проблемы эффективной идентификации антител против любой мишени, которые могут разрабатываться, являются безопасными и эффективными при применении у пациентов.
В наиболее общем смысле авторы изобретения начали с идеи, что библиотека антител, которая имитирует человеческую иммунную систему в основных путях, может быть наиболее предпочтительной. С одной стороны, авторы изобретения решили имитировать человеческую иммунную систему путем включения оптимальных генных последовательностей эмбрионального типа из иммунного репертуара человека в антитела. По существу, в некоторых воплощениях антитела библиотеки содержат участки, например каркасные участки, которые являются эмбриональными по последовательности. Использование последовательностей эмбрионального типа должно резко уменьшить риск иммуногенности рекомбинантных антител для терапевтического применения у пациентов.
Кроме того, авторы изобретения исходили из гипотезы, что пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, присутствующие в большом количестве в иммунном репертуаре человека, по-видимому, имеют предпочтительные биофизические свойства, которые приведут к более эффективной клинической разработке и увеличению безопасности и эффективности полученных антител при применении у пациентов. В качестве основы, каждая В-клетка кодирует одно антитело, и каждое антитело содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи. Каждую вариабельную область тяжелой цепи и вариабельную область легкой цепи антитела можно выравнивать с последовательностями эмбрионального типа для того, чтобы определить происхождение антитела, означающее, какой ген эмбрионального типа кодирует вариабельную область тяжелой цепи и вариабельную область легкой цепи. Поэтому для каждого антитела вариабельная область тяжелой цепи и вариабельная область легкой цепи содержит пару эмбрионального типа, например, VH3-23, спаренную с VK1-5.
Для того чтобы подтвердить гипотезу, что основные пары генов эмбрионального типа, по-видимому, имеют предпочтительные биофизические свойства, первая стадия заключалась в идентификации пар генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, присутствующих в иммунном репертуаре человека. Это делали посредством тщательного поиска в общедоступной литературе и анализа В-клеток от человека-хозяина. На следующей стадии первоначальные данные объединяли, анализировали и пары вариабельной области тяжелой цепи и вариабельной области легкой цепи эмбрионального типа, присутствующие в иммунном репертуаре человека, ранжировали по их распространенности. Из этих данных ясно, что определенные пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи присутствовали чаще, чем другие в иммунном репертуаре человека.
Кроме того, авторы изобретения считают, что определенные пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи могут по-разному экспрессироваться в "наивных" В-клетках ("необученных" антигеном, от англ. antigen inexperienced) по сравнению с "обученными" антигеном (antigen experienced) В-клетками, поэтому полученные данные анализировали на основании развития или дифференцировки выбранных В-клеток. Из этого анализа ясно, что определенные пары генов эмбрионального типа по-разному экспрессируются в популяциях "наивных" В-клеток по сравнению с популяциями "обученных" антигеном В-клеток.
На следующей стадии следовало определить, какие пары белков эмбрионального типа необходимо протестировать, поскольку существует примерно 2500 пар в иммунном репертуаре человека. Один из способов состоял в том, чтобы протестировать пары белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые чаще всего встречаются в иммунном репертуаре человека, например, см. Таблицу 18. Можно, например, выбрать верхние четыре сотни пар для тестирования или выбрать пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, экспрессирующиеся выше определенной пороговой концентрации. Такой подход обычно требует синтеза и тестирования большого числа последовательностей пар белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи; поэтому такой подход может быть не очень эффективным.
В качестве альтернативного подхода авторы изобретения выбрали субпоппуляцию пар вариабельных областей тяжелой цепи и вариабельных областей легкой цепи эмбрионального типа, которые являются репрезентативными, точно воспроизводит или охватывает большую часть преимущественно экспрессирующихся пар из иммунного репертуара человека. Этот подход основан, частично, на наблюдении, что небольшое число генов эмбрионального типа вариабельной области тяжелой, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепей является доминантным в иммунном репертуаре человека. Этот феномен описан в Wildt et al. at 895-896. Wildt с соавт.также утверждает, что обычно экспрессирующиеся генные сегменты тяжелых и легких цепей часто спариваются, и наблюдал, что половина выбранных спариваний соответствовала лишь пяти парам эмбрионального типа. Поэтому небольшое число преимущественно экспрессирующихся генов эмбрионального типа тяжелых и легких цепей (неспаренных) можно комбинировать с образованием группы пар, которые являются типичными для иммунного репертуара человека.
Этот подход осуществляли следующим образом. Собранные данные и дополнительные данные (идентифицирующие только экспрессию VH или VL, не связанных пар) анализировали для определения экспрессии генов эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи в иммунном репертуаре человека. На следующей стадии преимущественно экспрессирующиеся белковые последовательности эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи (не пары) оценивали для определения их биофизических свойств, имеющих отношение к разработке. Оценивали белковые последовательности эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи in silica в отношении следующих свойств: (1) длина CDR, (2) изоэлектрическая точка (рl) (предпочтительная изоэлектрическая точка составляет 8 или выше, которая должна обеспечивать стабильность в нейтральном буфере препарата), (3) посттрансляционные модификации (РТМ) (конкретно, N-связанные сайты гликозилирования (N×S или N×T) или химические модификации, такие как отщепление Asp (часто в DP), (4) Asp изомеризация (DD, DG), (5) дезамидирование (NS, NG), которое встречается in vivo (в сыворотке) или при хранении в буфере препарата и приводит к потере антительного связывания), (6) присутствие метионинов в CDR (могут окисляться при воздействии растворителя), (7) присутствие неспаренных цистеинов (будут образовывать дисульфидные связи с другими неспаренными цистеинами, что приводит таким образом к сшиванию белков и/или более низким уровням экспрессии), (8) отклонения от эмбрионального типа, (9) присутствие возможных Т-клеточных эпитопов и (10) теоретическая склонность к агрегации.
На следующей стадии пары эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи, имеющие предпочтительные биофизические характеристики, комбинировали с образованием пар вариабельной области тяжелой цепи и вариабельной области легкой цепи. Эта субпопуляция пар, которая представляет, точно воспроизводит или охватывает большую часть преимущественно экспрессирующихся пар иммунного репертуара человека, как показано в Таблице 23. Это делали путем синтеза генов эмбрионального типа вариабельных областей тяжелой и легкой цепи, комбинирования их в пары, экспрессирования пар в виде белка и тестирования каждого для идентификации их биофизических свойств. Тестировали следующие свойства: (1) относительный уровень экспонирования на фаге в Fab-формате, (2) относительный уровень экспрессии в Fab-формате, например, в Е.соli; (3) термостабильность в Fab-формате; (4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате; (5) относительный уровень экспрессии в IgG-формате; (6) стабильность в сыворотке крупного рогатого скота в IgG-формате.
После того как пары белков эмбрионального типа, имеющие предпочтительные биофизические свойства, были идентифицированы, конструировали коллекции для включения этих пар. Аспект настоящего изобретения относится к коллекции антител или функциональных фрагментов, содержащих пары генов эмбрионального типа вариабельных областей тяжелой и легкой цепи, имеющие подходящие свойства, которые увеличивают способность к разработке, но исключающих пары генов эмбрионального типа вариабельных областей тяжелой и легкой цепи, не имеющие таких свойств, даже если они преимущественно экспрессируются в иммунном репертуаре человека. Таким путем конструировали коллекцию с целью исключения комбинаций или пар вариабельных областей тяжелой и легкой цепи, которые встречаются в природе (из 2500 пар) и которые не имеют подходящих функциональных свойств. Например, VH4-34 часто встречается в иммунном репертуаре человека, как показано в Таблице 20, но также известно, что антитела, происходящие из этого гена тяжелой цепи эмбрионального типа, являются цитотоксичными для В-клеток, поэтому антитела, происходящие из этого гена, можно исключить из конструирования библиотеки. См. Bhat et al., Rapid cytotoxicity of human В lymphocytes induced by VH4-34 (VH4.21) gene-encoded monoclonal antibodies, Clin Exp Immunol., 105(1); 183-90 (July 1996).
В некоторых воплощениях коллекции по настоящему изобретению включают антитела, содержащие большое число функционально ппредпочтительных комбинаций или пар вариабельных областей тяжелой и легкой цепи, так что антитела из данных коллекций являются совершенно разными, образуя таким образом коллекцию, которую можно использовать для идентификации антител против любой терапевтической мишени.
Такие коллекции преодолевают многие проблемы из предшествующего уровня техники. Например, когнатная библиотека, происходящая из В-клеток, не соответствует этой концепции, так как спаривания класса VH и VL, присутствующие в такой библиотеке, идентичны спариваниям классов, присутствующих в образце В-клеток. Если взять достаточно большой образец В-клеток, то в нем будет присутствовать каждая из комбинаций спариваний класса приблизительно 50 VH и 50 VL (2500). Всестороннее тестирование пар VH и VL в настоящем описании показало, что многие пары генов VH и VL эмбрионального типа лишены свойств, которые позволяют их разработку в клинике. Поэтому такие когнатные библиотеки содержат много пар VH и VL, которые, по-видимому, не могут разрабатываться. Поэтому желательно создавать библиотеки с большим разнообразием, содержащие только пары класса VH и VL, имеющие подходящие функциональные свойства, но с подходом когнатной библиотеки это невозможно.
Кроме того, в некоторых воплощениях пары генов эмбрионального типа, содержащиеся в коллекции, основаны на образцах "наивных" или "необученных" антигеном В-клеток, поэтому представленные пары генов эмбрионального типа не ориентированы на конкретный иммуноген, и коллекции могут быть лучше при скрининге против любого иммуногена.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг.1 показаны результаты ИФА (твердофазный иммуноферментный анализ) экспрессии анти-Fd после периплазматической экстракции антитела с тяжелой цепью VH3-23 (верхняя панель) и тяжелой цепью VH1-69 (нижняя панель), где каждое антитело несет одну из трех модифицированных сигнальных последовательностей phoA, содержащих С-концевые сайты рестрикции Aflll (VLS), Nhel (VLA) и Avrll (VLG), по сравнению с сигнальной последовательностью дикого типа (ТКА). В группе VH3-23 все модифицированные сигнальные последовательности phoA поддерживали уровни экспрессии в диапазоне дикого типа (ТКА).
На Фиг.2 показаны результаты ИФА экспрессии анти-Fd после периплазматической экстракции антитела с легкой цепью VK1-39 (левая верхняя панель), с легкой цепью VK3-11 (правая верхняя панель), с легкой цепью VL1-40 (левая нижняя панель) и с легкой цепью VL3-1 (правая нижняя панель), где каждое антитело несет одну из трех модифицированных сигнальных последовательностей ompA, содержащих С-концевые сайты рестрикции Ndel (AYG), Ndel (AYA) и BsiWI (TYA), по сравнению с сигнальной последовательностью дикого типа (AQA). Модифицированные сигнальные последовательности ompA, содержащие С-концевые сайты рестрикции, и сигнальную последовательность дикого типа тестировали с использованием Vк и Vλ Fab-фрагментов. Сигнальная последовательность, включающая Ndel (AYA), демонстрировала постоянно такую же или даже более высокую экспрессию, чем дикий тип (AQA).
На Фиг.3 показаны сайты рестрикции, выбранные для включения в С-конец сигнальных последовательностей phoA и ompA E.coli, как подробно описано в Примерах 1-1.3, и включены сигнальные последовательности вокруг CDR3 и их соответствующие ориентации. На этой Фигуре, при изображении сигнальных последовательностей Е.соli, также представлены С-концевые сайты рестрикции, выбранные для включения в лидерные последовательности человеческой тяжелой цепи и каппа-цепи, для применения в экспрессии IgG, как подробно описано в Примере 1.5.
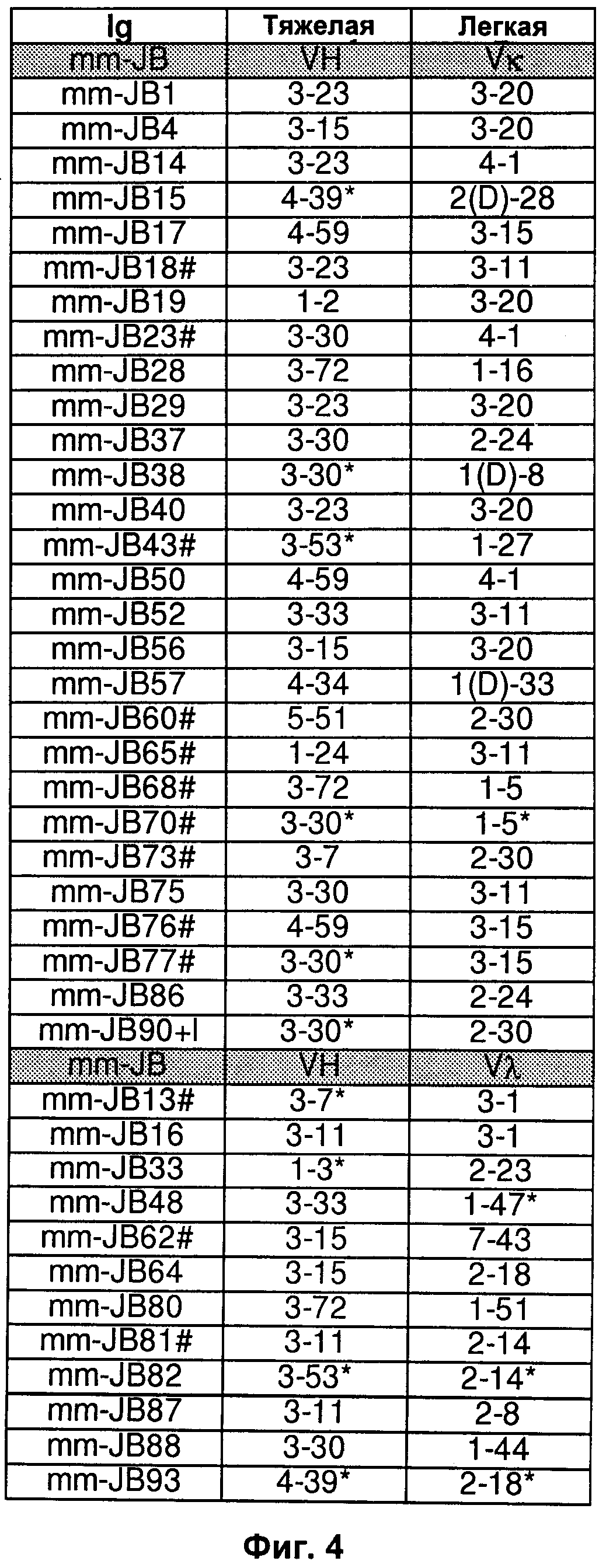
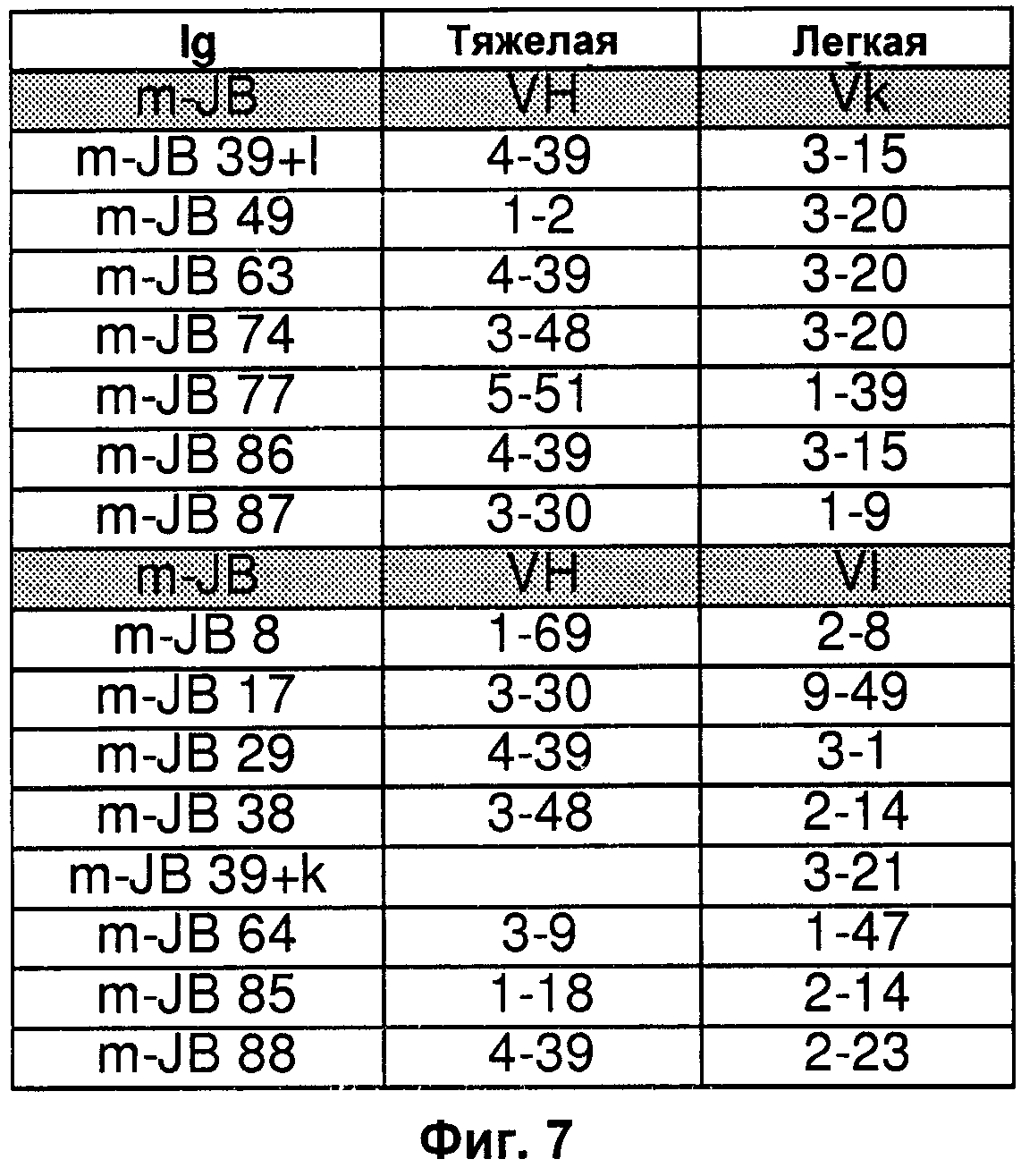
На Фиг.4-9 показаны пары генов VH/VL эмбрионального типа В-клеток, выделенные и описанные в Tsuiji M. et al. (2006).
На Фиг.10-12 показаны пары генов VH/VL эмбрионального типа В-клеток, выделенные и описанные в Tiller Т. et al. (2007).
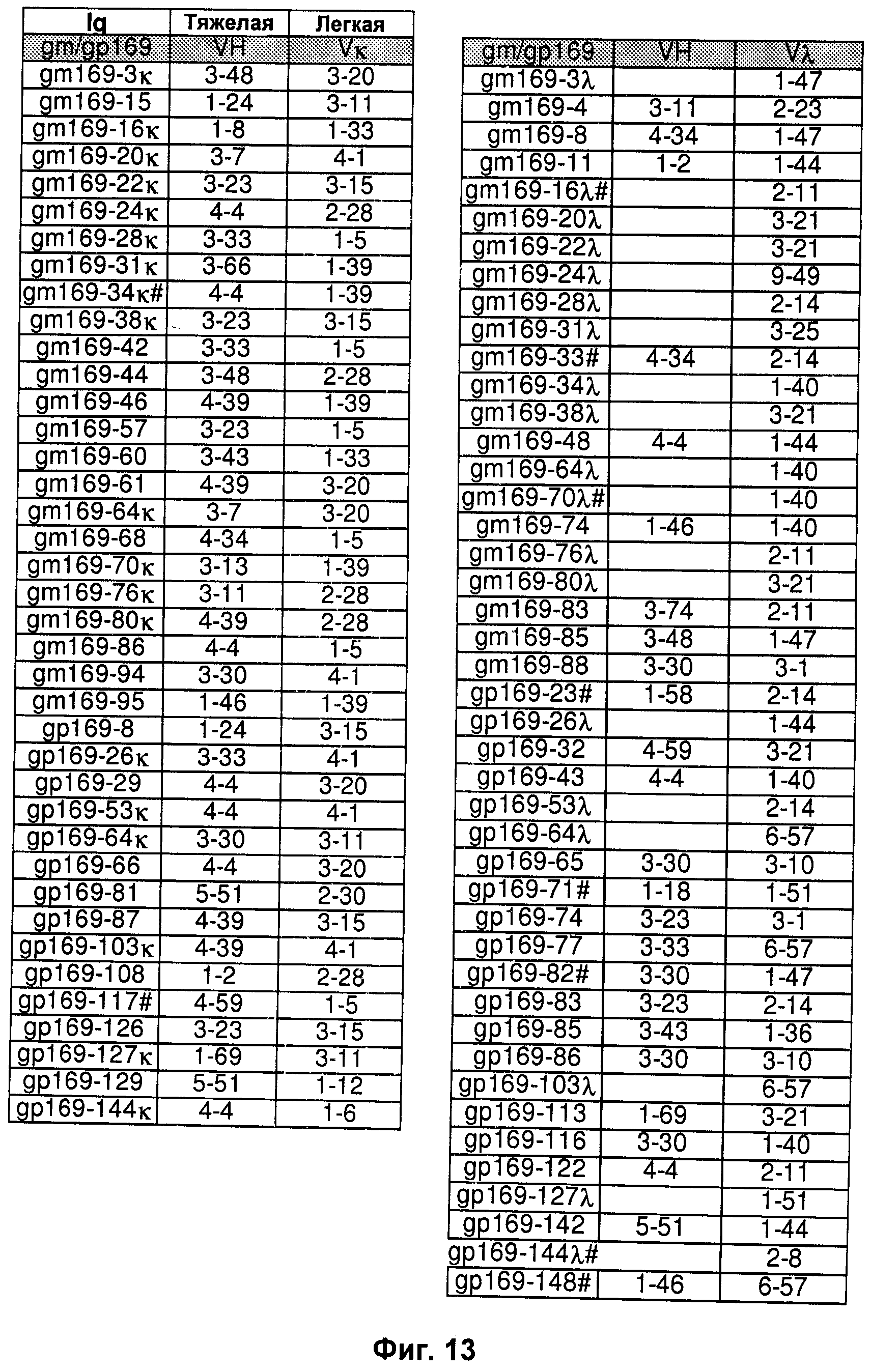
На Фиг.13-17 показаны пары генов VH/VL эмбрионального типа В-клеток, выделенные и описанные в Mietzner В. et al. (2008).
На Фиг.18-20 показаны пары генов VH/VL эмбрионального типа В-клеток, выделенные и описанные в Wardemann H. et al. (2003).
На Фиг.21-23 показаны пары генов VH/VL эмбрионального типа В-клеток, выделенные и описанные в Yurasov S. et al. (2005).
На Фиг.24-26 показаны пары генов VH/VL эмбрионального типа В-клеток, выделенные и описанные в Yurasov S. et al. (2006).
На Фиг.27 показана ПЦР-стратегия, используемая для амплификации кДНК одиночных отсортированных зрелых "наивных" (mn, от англ. mature naive) В-клеток и антитело-секретирующих клеток (asc), выделенных от человека-хозяина, как подробно описано в Примере 2.2.
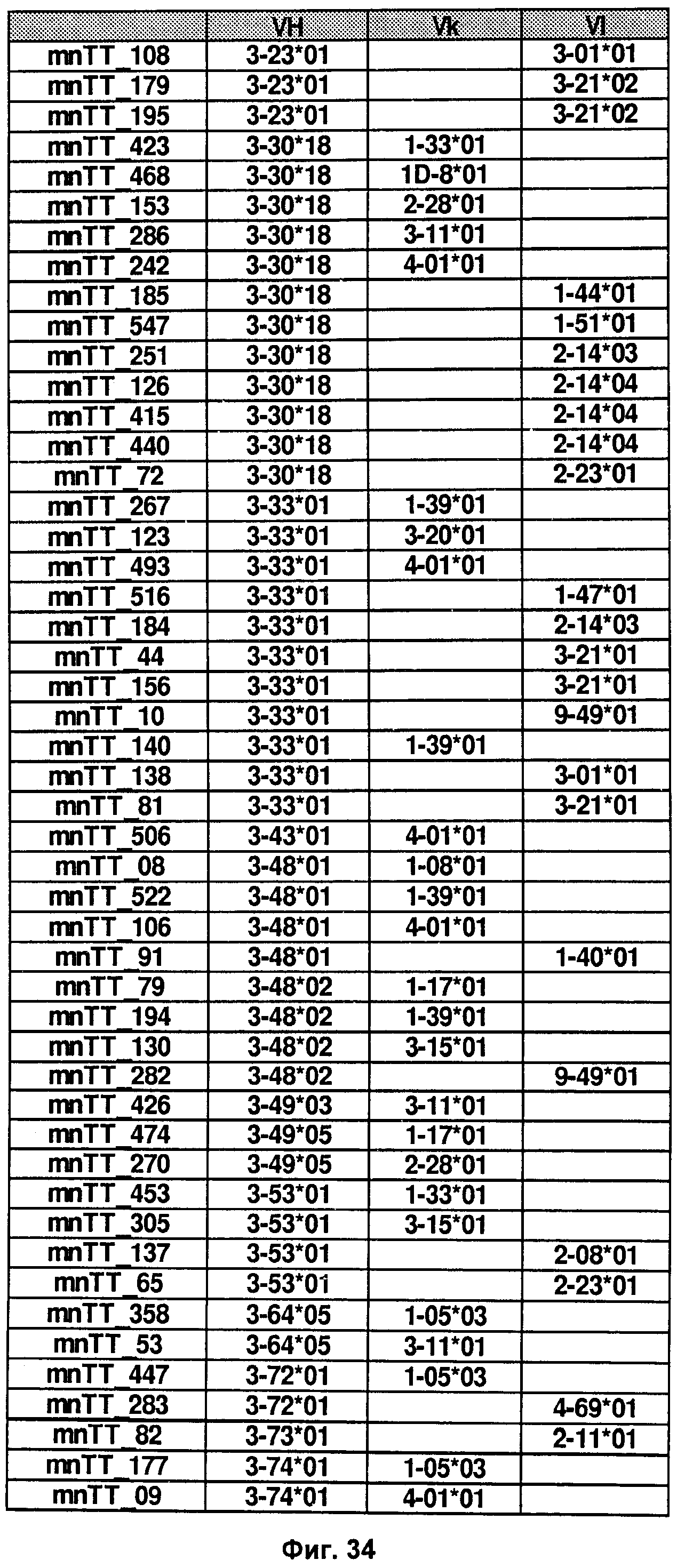
На Фиг.28-36 показаны пары VH/VL из В-клеток, выделенных из человеческого образца, как подробно описано в Примере 2.2.
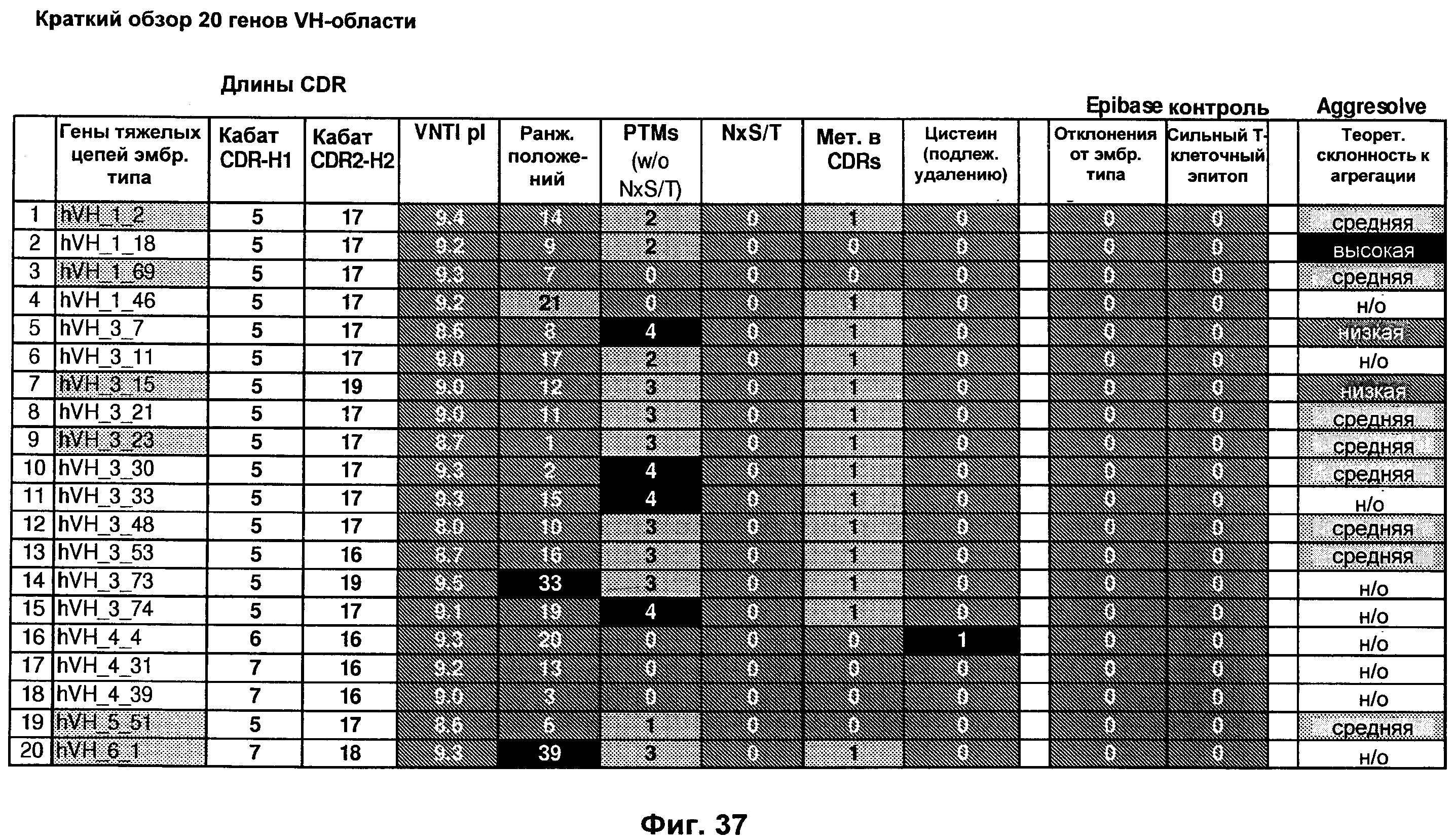
На Фиг.37 показаны 20 генов VH эмбрионального типа, выбранных для синтеза, комбинирования и функциональной характеристики, как подробно описано в Примерах 4-4.1. На Фиг.также показаны результаты in silico анализа каждого гена эмбрионального типа, где рl представляет изоэлектрическую точку, PTMs представляют посттрансляционные модификации в участках, определяющих комплементарность, как описано в данной заявке, N×S/T представляют М-связанные сайты гликозилирования, и Met в CDR представляют метионины.
На Фиг.38 показаны 8 генов Vλ и 12 генов Vk эмбрионального типа, выбранные для синтеза, комбинирования и функциональной характеристики, как подробно описано в Примерах 4-4.1. На Фиг.также показаны результаты in silico анализа каждого гена эмбрионального типа, где рl представляет изоэлектрическую точку, PTMs представляют посттрансляционные модификации в участках, определяющих комплементарность, как описано в данной заявке, N×S/T представляют М-связанные сайты гликозилирования, и Met в CDR представляют метионины.
На Фиг.39 показаны пары VH/Vк из объединенных данных из Примеров 2.1, показанных на Фиг.4-26, и Примера 2.2, показанного на Фиг.28-36. Числовые данные представляют количество каждой пары генов VH/Vк эмбрионального типа из индивидуальной В-клетки, идентифицированной в объединенных данных. На оси Y показаны гены VH эмбрионального типа, располагающиеся сверху (VH3-23) вниз (VH3-20) по частоте экспрессии в объединенных данных. На оси Х показаны гены Vк эмбрионального типа, располагающиеся слева (IGKV3-20) направо (IGKV1D-17) по частоте экспрессии в объединенных данных. Число 1358 представляет собой число проанализированных В-клеток.
На Фиг.40 показаны пары VH/Vλ из объединенных данных из Примеров 2.1, показанных на Фиг.4-26, и Примера 2.2, показанного на Фиг.28-36. Числовые данные представляют число каждой пары генов VH/Vλ эмбрионального типа из индивидуальной В-клетки, идентифицированной в объединенных данных. На оси Y показаны гены VH эмбрионального типа, располагающиеся сверху (VH3-23) вниз (VH3-20) по частоте экспрессии в объединенных данных. На оси Х показаны гены Vλ эмбрионального типа, располагающиеся слева (IGLV2-14) направо (IGLV4-60) по частоте экспрессии в объединенных данных. Число 779 представляет собой число проанализированных В-клеток.
На Фиг.41 показаны пары VH/Vк из объединенных данных из Примеров 2.1 показанных на Фиг.4-26, и Примера 2.2, показанного на Фиг.28-36, но включающих только популяции "необученных" антигеном В-клеток из незрелых В-клеток, новые эмигрирующие В-клетки и зрелые "наивные" В-клетки с целью идентификации пар VH/Vк, выраженных в иммунном репертуаре интактного (naive) человека. Числовые данные представляет число каждой пары генов VH/VL эмбрионального типа из индивидуальной В-клетки, идентифицированной в объединенных данных. На оси Y показаны гены VH эмбрионального типа, располагающиеся сверху (VH3-23) вниз (VH3-20) по частоте экспрессии в объединенных данных. На оси Х показаны гены Vк эмбрионального типа, располагающиеся слева (IGKV3-20) направо (IGKV1D-17) по частоте экспрессии в объединенных данных. Число 888 представляет собой число проанализированных В-клеток.
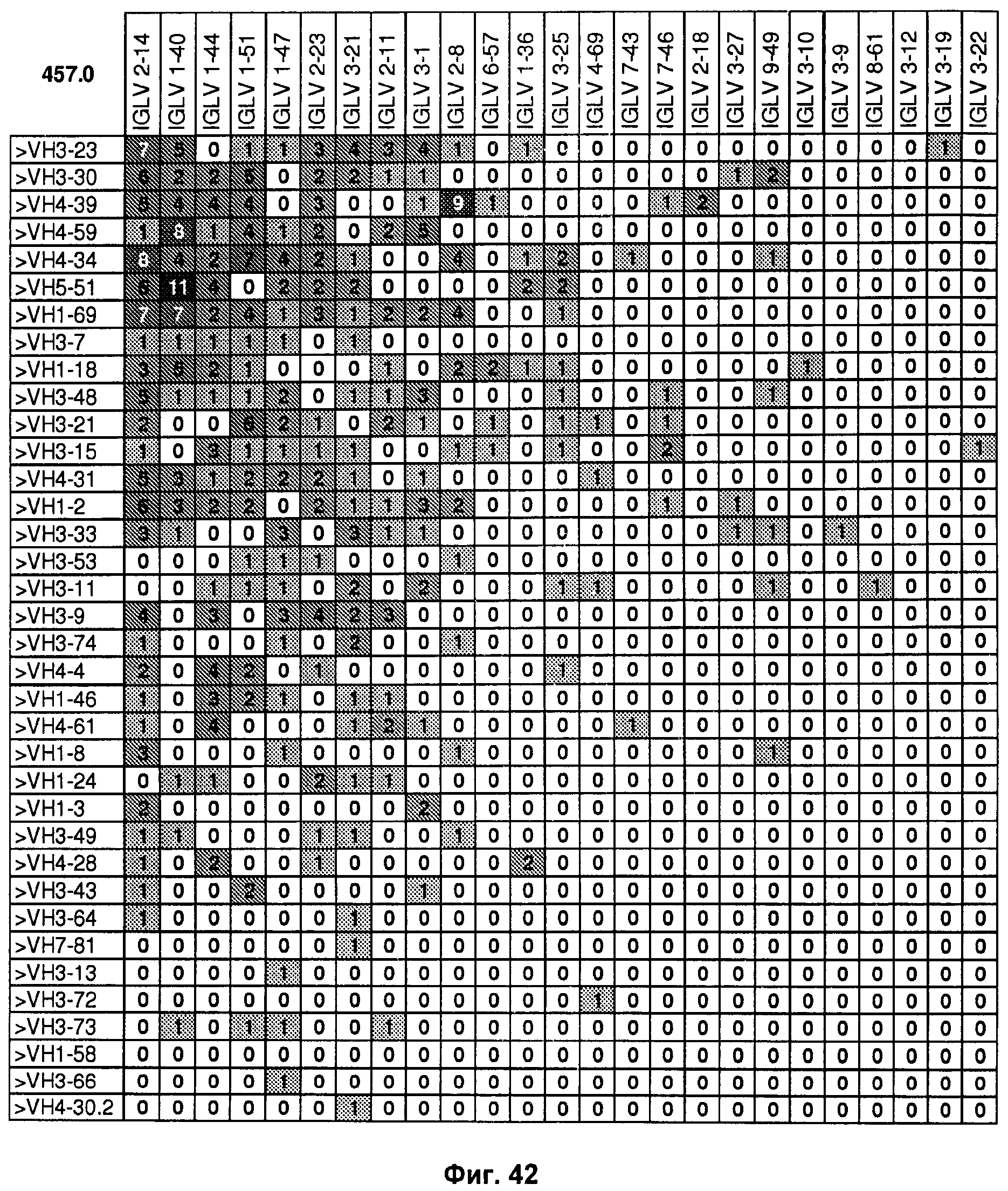
На Фиг.42 показаны пары VH/Vλ из объединенных данных из Примеров 2.1, показанных на Фиг.4-26, и Примера 2.2, показанного на Фиг.28-36, но включающих только популяции "необученных" антигеном В-клеток из незрелых В-клеток, новые эмигрирующие В-клетки и зрелые "наивные" В-клетки с целью идентификации пар VH/Vλ, выраженных в иммунном репертуаре интактного человека. Числовые данные представляет число каждой пары генов VH/Vλ эмбрионального типа из индивидуальной В-клетки, идентифицированной в объединенных данных. На оси Y показаны гены VH эмбрионального типа, располагающиеся сверху (VH3-23) вниз (VH3-20) по частоте экспрессии в объединенных данных. На оси Х показаны гены Vλ эмбрионального типа, располагающиеся слева (IGLV2-14) направо (IGLV4-60) по частоте экспрессии в объединенных данных. Число 457 представляет собой число проанализированных В-клеток.
На Фиг.43 показаны пары VH/Vк из объединенных данных из Примеров 2.1, показанных на Фиг.4-26, и Примера 2.2, показанного на Фиг.28-36, но включающих только популяции "обученных" антигеном В-клеток из IgG антитело-секретирующих клеток, и IgM и IgG В-клетки памяти. Числовые данные представляет число каждой пары генов VH/Vк эмбрионального типа из индивидуальной В-клетки, идентифицированной в объединенных данных. На оси Y показаны гены VH эмбрионального типа, располагающиеся сверху (VH3-23) вниз (VH3-20) по частоте экспрессии в объединенных данных. На оси Х показаны гены Vк эмбрионального типа, располагающиеся слева (IGKV3-20) направо (IGKV1D-17) по частоте экспрессии в объединенных данных. Число 470 представляет собой число проанализированных В-клеток.
На Фиг.44 показаны пары VH/Vλ из объединенных данных из Примеров 2.1, показанных на Фиг.4-26, и Примера 2.2, показанного на Фиг.28-36, но включающих только популяции "обученных" антигеном В-клеток из IgG антитело-секретирующих клеток, и IgM и IgG В-клетки памяти. Числовые данные представляет число каждой пары генов VH/Vλ эмбрионального типа из индивидуальной В-клетки, идентифицированной в объединенных данных. На оси показаны Y гены VH эмбрионального типа, располагающиеся сверху (VH3-23) вниз (VH3-20) по частоте экспрессии в объединенных данных. На оси Х показаны гены Vλ эмбрионального типа, располагающиеся слева (IGLV2-14) направо (IGLV4-60) по частоте экспрессии в объединенных данных. Число 322 представляет собой число проанализированных В-клеток.
На Фиг.45А-С показаны аминокислотные последовательности, кодируемые генами VH эмбрионального типа, как описано в Tomlinson et al., (1992), "The Repertoire of Human Germline Vh Sequences Reveals about Fifty Groups of Vh Segments with Different Hypervariable Loop" J. Mol. Biol. 227, 776-798; Matsuda et al. (1998), "The complete nucleotide sequence of the human immunoglobulin heavy chain variable region locus" J Exp Med 188(11):2151 -62; и LeFranc MP (2001) "Nomenclature of the human immunoglobulin heavy (IGH) genes." Exp Clin Immunogenet. 18(2): 100-16.
На Фиг.46А-С показаны аминокислотные последовательности, кодируемые генами Vк эмбрионального типа, как описано в Schable and Zachau (1993), "The variable genes of the human immunoglobulin kappa locus", Biol. Chem Hoppe Seyler. 374(11); 1001-22; Brensing-Kuppers et al. (1997), "The human immunoglobulin kappa locus on yeast artificial chromosomes (YACs)" Gene. 191 (2): 173-81; Kawasaki et al. (2001), "Evolutionary dynamics of the human immunoglobulin kappa locus and the germline repertoire of the Vkappa genes" Eur J Immunol 31 (4): 1017-28; и Lefranc MP (2001) "Nomenclature of the human immunoglobulin kappa (IGK) genes" Exp Clin Immunogenet, 18, 161-174.
На Фиг.47А-В показаны аминокислотные последовательности, кодируемые генами Vλ эмбрионального типа, как описано у Kawasaki et al., (1997) "One-Megabase Sequence Analysis of the Human immunoglobulin lambda Gene Locus" Genome Research 7(3):250-61; Frippiat et al., (1995) "Organization of the human immunoglobulin lambda light-chain locus on chromosome 22q11.2" Hum. Mol. Genet., 4, 983-991; и LeFranc MP (2001) "Nomenclature of the human immunoglobulin lambda (IGL) genes. Exp Clin lmmunogenet.;18:242-254.
На Фиг.48 показан трицистронный фаговый дисплейный вектор pJPd1.
На Фиг.49 показан экспрессирующий вектор pJPx1 Fab.
На Фиг.50 показан экспрессирующий вектор рМх11 (pMORPHXH) Fab.
На Фиг.51 показан дисплейный вектор pMORPHSO Fab.
На Фиг.52 показан экспрессирующий вектор pJP_hJgG1f для вариабельной области тяжелой цепи IgG.
На Фиг.53 показан экспрессирующий вектор pJP_h_lg_kappa для вариабельной области к-легкой цепи IgG.
На Фиг.54 показан экспрессирующий вектор pJP_h_lg_lambda2 для вариабельной области λ-легкой цепи IgG.
На Фиг.55 показаны относительные уровни экспонирования Fab для 400 протестированных пар генов VH/VL эмбрионального типа. Более высокие значения указывают на более высокие уровни экспонирования.
На Фиг.56 показаны относительные уровни экспрессии Fab для 400 протестированных пар генов VH/VL эмбрионального типа. Более высокие значения указывают на более высокие уровни экспрессии Fab.
На Фиг.57 показаны данные по термостабильности для 400 протестированных пар генов VH/VL эмбрионального типа в Fab-формате. Числа 60 и 70 указывают на пары VH/VL, которые стабильны в течение 45 мин при 60 или 70°С в тестируемой системе. Число 4 указывает на термонестабильные пары, и bg указывает на низкие уровни экспрессии.
На Фиг.58 показаны данные по стабильности в сыворотке крупного рогатого скота для 400 протестированных пар генов VH/VL эмбрионального типа в Fab-формате. S означает стабильные, и U - нестабильные в тестируемых условиях.
На Фиг.59 показаны данные по стабильности в мышиной сыворотке для 400 протестированных пар генов VH/VL эмбрионального типа в Fab-формате. S означает стабильные, и U - нестабильные в тестируемых условиях.
На Фиг.60 показаны относительные уровни экспрессии IgG для 400 протестированных пар генов VH/VL эмбрионального типа. Более высокие значения указывают на более высокие уровни экспрессии IgG1.
На Фиг.61 показаны данные по стабильности в сыворотке для 400 протестированных пар генов VH/VL эмбрионального типа в IgG-формате. S означает стабильные, и U - нестабильные в тестируемых условиях.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
Для того чтобы облегчить понимание изобретения, предложены следующие определения и иллюстрации.
Основные термины
Термины "примерно" или "приблизительно" в контексте числовых значений и диапазонов относятся к значениям или диапазонам, которые приблизительно равны или близки к таким представленным значениям или диапазонам, что изобретение может осуществляться как и предполагается, таким как имеющие желательное значение или процент гомологии последовательности, как очевидно специалисту из идей, содержащихся в данной заявке. Это является следствием, по меньшей мере частично, варьирующих культуральных условий и вариабельности биологических систем. Таким образом, эти термины охватывают значения за пределами значений, полученных из систематической погрешности. Эти термины выражают ясно то, что подразумевается. Как правило, "примерно" охватывает ±10% представленного значения. Поэтому термин "примерно" можно использовать для описания диапазона.
Все диапазоны, представленные в данной заявке в кратком изложении сущности и описании изобретения, включают все числа или значения около них или между числами диапазона. Все диапазоны по изобретению точно определяют и представляют все целые числа, десятичные числа и дробные величины в данном диапазоне.
Термин "субъект" включает человека и животных, не являющихся человеком. Животные, не являющиеся человеком, включают всех позвоночных, например, млекопитающих и не млекопитающих, таких как приматы, не являющиеся человеком, овцы, собаки, коровы, цыплята, земноводные и пресмыкающиеся. За исключением указанных случаев, термины "пациент" или "субъект" используют в данной заявке взаимозаменяемо.
Термин "лечение" включает введение композиций или антител для предупреждения или замедления появления симптомов, осложнений или биохимических признаков заболевания, смягчения симптомов или прекращения или ингибирования дальнейшего развития заболевания, состояния или расстройства. Поэтому лечение охватывает, но не ограничивается этим, "излечение". Лечение может быть профилактическим (для предупреждения или замедления начала заболевания или для предупреждения или замедления проявления его клинических или субклинических симптомов) или терапевтической супрессией или смягчением симптомов после проявления заболевания.
"База данных или читаемый носитель", как использовано в данной заявке, относится к любому формату для хранения данных о последовательностях и, таким образом, к любой коллекции информации, такой как файл базы данных, поисковая таблица, сводная таблица Excel или тому подобное. В некоторых воплощениях базу данных хранят в электронной форме, такой как машиночитаемое запоминающее устройство. Оно включает носители информации, такие как сервер, клиент, жесткий диск, CD, DVD, персональное информационное устройство, такое как Palm Pilot, ленточный носитель, zip диск, встроенный в компьютер ROM (постоянное запоминающее устройство) или Интернет или всемирная паутина. Другие средства для хранения файлов, доступные для компьютера, очевидны специалисту в данной области техники.
"In silico" относится к манипуляциям, анализу и конструкциям, выполняемым на компьютере, но которые могут быть выполнены на бумаге или в уме.
Антитела и их свойства
Используемый в данной заявке термин "антитело" включает целые антитела. Антитело может быть поликлональным, аффинноочищенным поликлональным, моноклональным, полностью человеческим, антителом мыши или грызуна, химерным, верблюжьим или гуманизированным антителом. Антитело может относиться к любому классу антител, такому как IgG, IgG1, IgG2, IgG3, IgG4, IgA (включая человеческие подклассы 1дА1 и lgA2), IgD, IgE, IgG или IgM. Встречающееся в природе "антитело" представляет собой гликопротеин, содержащий по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями.
Используемый в данной заявке термин "его функциональный фрагмент" включает любой антиген-связывающий фрагмент, такой как Fab, F(ab')2, Fab', Fv, scFv, одиночные цепи, которые включают Fc-участок, нанотела и другие антителоподобные структуры, имеющие остовы, отличные от каркасных участков вариабельных областей. Термин "его функциональный фрагмент" включает, но не ограничивается этим, любой функциональный участок антитела, где функция включает связывание иммуногена или эффекторную функцию.
Используемый в данной заявке термин "аффинность" относится к силе взаимодействия между антителом и антигеном в антигенных сайтах. В пределах каждого антигенного сайта вариабельная область "плеча" антитела взаимодействует посредством нековалентных сил с антигеном в многочисленных сайтах; чем больше взаимодействий, тем сильнее аффинность. Используемый в данной заявке термин "высокая аффинность" для антитела или его функционального фрагмента, такого как IgG антитело, относится к антителу, имеющему KD 10-8 М или меньше, 10-9 М или меньше, или 10-10 М или меньше, или 10-11 М или меньше для целевого антигена. Однако связывание с "высокой аффинностью" может варьировать для других изотипов антител. Например, связывание с "высокой аффинностью" для IgM изотипа относится к антителу, имеющему KD 10-7 М или меньше, или 10-8 М или меньше.
Используемый в данной заявке термин "Кассоц." или "Ка" предназначен для обозначения скорости ассоциации конкретного антитело-антигенного взаимодействия, где термин "Кдис." или "Kd", используемый в данной заявке, относится к скорости диссоциации конкретного антитело-антигенного взаимодействия. Термин "KD", используемый в данной заявке, предназначен для обозначения константы диссоциации, которая получена из отношения Kd к Ка (т.е. Kd/Ka) и выражается в виде молярной концентрации (М). Значения KD для антител можно определить с использованием методов, хорошо известных в данной области техники. Способ определения KD антитела осуществляют с использованием поверхностного плазменного резонанса или с использованием биосенсорной системы, такой как система Biacore®.
Термины "кросс-блок", "кросс-блокированные" и "кросс-блокирование", используемые в данной заявке взаимозаменяемо, означают способность антитела или другого связывающего агента препятствовать связыванию других антител или связывающих агентов с той же мишенью в стандартном анализе конкурентного связывания. Способность или степень, с которой антитело или другой связывающий агент могут препятствовать связыванию другого антитела или связывающей молекулы с той же мишенью, и поэтому можно ли указанный кросс-блок согласно изобретению определить с использованием стандартных анализов конкурентного связывания. Один подходящий анализ включает применение технологии Biacore (например, с использованием прибора BIAcore 3000 (Biacore, Uppsala, Швеция)), при котором можно измерять степень взаимодействий с использованием технологии поверхностного плазмонного резонанса. В другом анализе для измерения кросс-блокирования используют подход на основе ИФА.
Термин "эпитоп" означает белковую детерминанту, способную к специфическому связыванию с антителом. Эпитопы обычно состоят из химически активных поверхностных группировок молекул, таких как аминокислоты или сахарные боковые цепи, и обычно имеют специфические трехмерные структурные характеристики, а также специфические зарядные характеристики. Конформационные и неконформационные эпитопы отличаются тем, что связывание с первыми, но не с последними, не происходит в присутствии денатурирующих растворителей.
Термин "химерное антитело" представляет собой молекулу антитела, в которой константная область или ее часть изменена, замещена или заменена таким образом, чтобы антиген-связывающий сайт (вариабельная область) был связан с константной областью другого или измененного класса, эффекторной функцией и/или видом.
Термин "изотип" относится к классу антител (например, IgM, IgE, IgG такому как IgG1 или IgG4), который обеспечивается генами константной области тяжелой цепи. Изотип также включает модифицированные версии одного из этих классов, где были сделаны модификации для изменения Fc-функции, например, для увеличения или уменьшения эффекторных функций или связывания с Fc-рецепторами.
Термин "эмбриональный тип" означает последовательность нуклеиновой кислоты, кодирующая антитела или их функциональные фрагменты, которые передаются от родителей потомству.
Термин "белковая последовательность эмбрионального типа" означает:
а) аминокислотную последовательность вариабельной области антитела или его функционального фрагмента, кодируемую геном эмбрионального типа, б) аминокислотную последовательность, кодируемую модифицированной последовательностью нуклеиновой кислоты, кодирующей вариабельную область антитела или его функционального фрагмента, имеющего такую же аминокислотную последовательность вариабельной области антитела или его функционального фрагмента, кодируемого геном эмбрионального типа, где последовательность нуклеиновой кислоты модифицирована, например, посредством оптимизации кодонов, добавления желательных сайтов рестрикции, оптимизированного GC-содержания, удаления нежелательных сайтов сплайсинга или удаления мотивов нестабильности мРНК, или в) аминокислотную последовательность, кодируемую геном эмбрионального типа, но с точечными мутациями в аминокислотной последовательности, такими как удаление нежелательного цистеина или введение желательных сайтов рестрикции, например Bbsl, или которые являются результатом ошибок синтеза, амплификации или клонирования.
Термин "генная последовательность эмбрионального типа" означает: а) последовательность нуклеиновой кислоты гена эмбрионального типа, кодирующую вариабельную область антитела или его функциональный фрагмент, или б) модифицированную последовательность нуклеиновой кислоты, кодирующую вариабельную область антитела или его функционального фрагмента, имеющего такую же аминокислотную последовательность, как вариабельная область антитела, кодируемого геном эмбрионального типа, где последовательность нуклеиновой кислоты модифицирована, например, посредством оптимизации кодонов, добавления желательных сайтов рестрикции, оптимизированного GC-содержания, удаления нежелательных сайтов сплайсинга или удаления мотивов нестабильности мРНК.
Термин "пара(ы) генов эмбрионального типа" означает пару последовательностей нуклеиновых кислот и их соответствующий ген эмбрионального типа, кодирующий вариабельную область тяжелой цепи и вариабельную область легкой цепи антитела или его функционального фрагмента. Например, пара генов эмбрионального типа может представлять собой VH3-23/VK1-5, где антитело, кодируемое VH3-23/VK1-5, содержит вариабельную область тяжелой цепи или ее часть, кодируемую геном VH3-23 эмбрионального типа, и вариабельную область легкой цепи или ее часть, кодируемую геном Vк1-5 эмбрионального типа.
Термин "пара белков эмбрионального типа" означает антитело или его функциональный фрагмент, где вариабельная область тяжелой цепи или ее часть и вариабельная область легкой цепи или ее часть а) кодируются специфическим геном эмбрионального типа или б) кодируются модифицированной последовательностью нуклеиновой кислоты, кодирующей вариабельную область антитела или его функционального фрагмента, имеющего такую же аминокислотную последовательность, как вариабельная область антитела, кодируемого специфическим геном эмбрионального типа, где последовательность нуклеиновой кислоты модифицирована, например, посредством оптимизации кодонов, добавления желательных сайтов рестрикции, оптимизированного GC-содержания, удаления нежелательных сайтов сплайсинга или удаления мотивов нестабильности мРНК, или в) каждая содержит аминокислотную последовательность, кодируемую геном эмбрионального типа, но с точечными мутациями в аминокислотной последовательности, например, с целью удаления нежелательного цистеина или введения желательных сайтов рестрикции, например Bbsl, или которые являются результатом ошибок в синтезе, амплификации или клонировании. Например, пара белков эмбрионального типа может представлять собой антитело или функциональный фрагмент, кодируемый VH3-23/VK1-5, где антитело содержит вариабельную область тяжелой цепи или ее часть, кодируемую геном эмбрионального типа VH3-23, и вариабельную область легкой цепи или ее часть, кодируемую геном эмбрионального типа Vid-5. "Пара белков эмбрионального типа" включает конструкции, полученные в Примере 5, которые содержат:
а) для VH: лидерную последовательность (модифицированную phoA, включающую RE сайт Nhel, как показано на Фиг.3); FR1, CDR1, FR2, CDR2 и FR3 эмбрионального типа (включающие RE сайт BssHII, как показано на Фиг.3); CDR-H3 (WGGDGFYAMDY) антитела 4D5, как использовано в Ewert S. et al., J. Mol. Biol. (2003) 325, 531-553; и JH4 FR4 (включающий RE сайт Xhol/Sall как показано на Фиг.3);
б) для Vк: лидерную последовательность (оmрА, включающую RE сайт Ndel, как показано на Фиг.3); FR1, CDR1, FR2, CDR2 и FR3 эмбрионального типа (включающие RE сайт Bbsl, как показано на Фиг.3), каппа-подобный CDR-L3 (QQHYTTPPT) согласно Ewert S. et al., J. Mol. Biol. (2003) 325, 531-553; и Jk1 FR4 (включающий RE сайт Kpnl, как показано на Фиг.3); и
в) для Vλ: лидерную последовательность (оmрА, включающую RE сайт Ndel, как показано на Фиг.3); FR1, CDR1, FR2, CDR2 и FR3 эмбрионального типа (включающие RE сайт Bbsl, как показано на Фиг.3), лямбда-подобный CDR-L3 (QSYDSSLSGV5) согласно Ewert S. et al., J. Mol. Biol. (2003) 325, 531-553; и JI2/3 FR4 (включающий RE сайт Kpnl, как показано на Фиг.3).
Каждую из этих конструкций синтезировали, экспрессировали и тестировали в виде Fab и IgG, как описано в Примерах 6 и 7, в отношении следующих функциональных свойств: а) относительный уровень экспонирования после получения фагов и фаговый ИФА в Fab-формате; б) относительные уровни экспрессии Fab после продукции Fab в Е.соli, лизиса клеток Е.соli и ИФА-детекции продуцируемого Fab; в) термостабильность Fab после продукции Fab в Е.соli, лизиса клеток Е.соli и ИФА-детекции неденатурированного Fab после инкубации при повышенных температурах; г) стабильность Fab из лизатов Е.соli в сыворотке крупного рогатого скота/мышей посредством ИФА-детекции неденатурированного Fab после инкубации в сыворотке крупного рогатого скота/мышей; д) относительные уровни экспрессии человеческого IgG1 после продукции IgG1 в клетках млекопитающих и ИФА-детекции секретируемого IgG1 из супернатантов клеточных культур; и е) стабильность человеческого IgG1 в сыворотке крупного рогатого скота, как определено посредством ИФА-детекции неденатурированного Fab после инкубации в сыворотке крупного рогатого скота/мышей.
Термин "по существу белковая последовательность эмбрионального типа" означает аминокислотную последовательность, кодируемую геном эмбрионального типа, но с точечными мутациями в аминокислотной последовательности, например, с целью удаления нежелательного цистеина или введения желательных сайтов рестрикции, например Bbsl, или которые являются результатом ошибок синтеза, амплификации или клонирования.
"Гены эмбрионального типа" представляют собой нуклеиновые кислоты генов эмбрионального типа, кодирующих антитела или их функциональные фрагменты, раскрытые в следующих публикациях, для VH: Tomlinson et al., (1992), "The Repertoire of Human Germline Vh Sequences Reveals about Fifty Groups of Vh Segments with Different Hypervariable Loop" J. Mol. Biol. 227, 776-798; Matsuda et al. (1998), "The complete nucleotide sequence of the human immunoglobulin heavy chain variable region locus" J Exp Med 188(11):2151-62; и LeFranc MP (2001) "Nomenclature of the human immunoglobulin heavy (IGH) genes". Exp Clin Immunogenet. 18(2): 100-16; для Vλ: Kawasaki et al., (1997) "One-Megabase Sequence Analysis of the Human immunoglobulin lambda Gene Locus" Genome Research 7(3):250-61; Frippiat et al., (1995) "Organization of the human immunoglobulin lambda light-chain locus on chromosome 22q11.2" Hum. Mol. Genet, 4, 983-991; и LeFranc MP (2001) "Nomenclature of the human immunoglobulin lambda (IGL) genes. Exp Clin Immunogenet.; 18:242-254; и для Vк: Schable and Zachau (1993), "The variable genes of the human immunoglobulin kappa locus", Biol. Chem Hoppe Seyler. 374(11): 1001-22; Brensing-KQppers et al. (1997), "The human immunoglobulin kappa locus on yeast artificial chromosomes (YACs)" Gene. 191 (2): 173-81; Kawasaki et ai. (2001), "Evolutionary dynamics of the human immunoglobulin kappa locus and the germline repertoire of the Vkappa genes" Eur J Immunol 31 (4):1017-28; и Lefranc MP (2001) "Nomenclature of the human immunoglobulin kappa (IGK) genes" Exp Clin Immunogenet., 18, 161-174, которые все включены в данную заявку посредством ссылки во всей своей полноте.
Последовательности JH4 для вариабельной области тяжелой цепи, Jк1 для вариабельной области к-легкой цепи и JA2/3 для вариабельных областей Л-легкой цепи описаны в следующих публикациях: Scaviner et al., (1999), "Protein displays of the human immunoglobulin heavy, kappa and lambda variable and joining regions" Exp Clin Immunogenet. 1б(4):234-40; для JH: Ravetch et al., (1981), "Structure of the human immunoglobulin mu locus: characterization of embryonic and rearranged J and D genes". Cell 27 (3 pt 2): 583-91; для JK: Hieter et al. (1982), "Evolution of human immunoglobulin kappa J region genes". J Biol Chem 257(3):1516-22; для JL: Kawasaki et al., (1997) "One-Megabase Sequence Analysis of the Human immunoglobulin lambda Gene Locus" Genome Research 7(3):250-61, которые все включены в данную заявку посредством ссылки во всей своей полноте. Последовательность JH4 представляет собой (YFDYWGQGTLVTVSS); последовательность Jк1 представляет собой (WTFGQGTKVEIK); и последовательность JA2/3 представляет собой (WFGGGTKLTVL).
Термин "позиционно-зависимая частота использования аминокислот" относится к вероятности наличия конкретной аминокислотной последовательности в данном положении в полипептиде. В настоящем изобретении позиционно-зависимая частота использования аминокислот определяли для перестроенных аминокислотных последовательностей, классифицированных с помощью индивидуального гена эмбрионального типа. Это обеспечивает индивидуальную, точную конструкцию CDR в контексте его природного эмбрионального типа.
Термин "вариабельный домен/область (VH или VL)" означает область иммуноглобулина, которая содержит один или более доменов Ig, по существу кодируемых любой из нуклеиновых кислот VL (включающих Vk и Vλ), VH, JL (включающих Jк и JA) и JH, которые составляют генетические локусы легкой цепи (включающие к и А) и тяжелой цепи иммуноглобулина, соответственно. Вариабельная область (VL и VH) легкой или тяжелой цепи состоит из "каркасного" или "FR" участка с рассеянными тремя гипервариабельными участками, называемыми "участками, определяющими комплементарность" или "CDRs". Протяженность каркасного участка и CDRs были точно определены (см. Kabat, 1991, J. Immunol., 147, 915-920.; Chothia & Lesk, 1987, J. Mol. В/о/. 196: 901-917; Chothia et al., 1989, Nature 342: 877-883; AI-Lazikani et al., 1997, J. Mol. Biol. 273: 927-948). Каркасные участки антитела, т.е. комбинированные каркасные участки, составляющие легкие и тяжелые цепи, служат для позиционирования и выравнивания CDRs, которые в первую очередь ответственны за связывание с антигеном.
Термин "каркасный участок" означает вариабельный домен антитела, как определено Kabat с соавт.(1991), в виде части вариабельного домена, которая служит каркасом для антиген-связывающих петель этого вариабельного домена. Примеры каркасных участков включают FR1, FR2, FR3 и FR4 либо вариабельной области тяжелой, либо вариабельной области легкой цепи.
Термин "участок, определяющий комплементарность" или "CDR" означает антигенсвязывающие петли антитела, как описано Kabat с соавт.(1991). Каждый из двух вариабельных доменов Fv-фрагмента антитела содержит три CDRs. Участки, определяющие комплементарность, включают CDR1, CDR2 и CDR3 либо вариабельной области тяжелой, либо вариабельной области легкой цепи.
"Предпочтительная пара класса VH и VL" означает такие пары класса VH и VL, которые являются предпочтительными в иммунном репертуаре, например, в иммунном репертуаре человека, согласно пороговой совокупности критериев. Например, пары VH-VL, которые часто встречаются, или имеют предпочтительные биофизические свойства, такие как низкая иммуногенность; стабильность; легко экспонируются и/или экспрессируются, или пары VH-VL, которые появляются в концентрации по меньшей мере 0,05% в образце из примерно 2500 В-клеток человека. Пары класса VH и VL, предпочтительные в иммунном репертуаре человека, могут иметь предпочтительные характеристики по сравнению с другими парами класса VH и VL.
Термин "наивный" означает "необученный" антигеном.
Термин "наивная В-клетка" означает В-клетку, в которой нуклеиновые кислоты, кодирующие антитела или их функциональные фрагменты, не подвергались соматической гипермутации, поэтому считаются содержащими нуклеиновые кислоты генов эмбрионального типа, с наличием перестройки генных сегментов V(D)J. Популяции В-клеток, считающихся "наивными", представляют собой незрелые В-клетки, новые эмигрирующие В-клетки и зрелые "наивные" В-клетки.
Термин "иммунный репертуар интактного человека" означает репертуар нуклеиновых кислот, выделенных из "необученных" антигеном В-клеток из иммунной системы человека, где нуклеиновые кислоты, кодирующие антитела или их функциональные фрагменты, не подвергались соматической гипермутации, поэтому рассматриваются как содержащие нуклеиновые кислоты генов эмбрионального типа, с наличием перегруппировки генных сегментов V(D)J. Репертуар может представлять собой репертуар индивидуума или популяции. Настоящее изобретение подходит для определения иммунного репертуара отдельного индивидуума при условии получения достаточного количества В-клеток. Предпочтительно, иммунный репертуар получают от множества индивидуумов во избежание деформации выборки.
Термин "иммунный репертуар человека" означает репертуар нуклеиновых кислот, выделенных из В-клеток иммунной системы человека. Репертуар может представлять собой репертуар индивидуума или популяции и может состоять из "наивных" В-клеток и/или "необученных" антигеном В-клеток. Настоящее изобретение подходит для определения иммунного репертуара отдельного индивидуума при условии получения достаточного количества В-клеток. Предпочтительно, иммунный репертуар получают от множества индивидуумов во избежание деформации выборки.
"Антиген" и "иммуноген" определяют как любую молекулу, которая специфически связывается антителом.
Термин "специфический в отношении иммуногена" означает специфическую ассоциацию между антителом и соответствующей молекулой.
"Диверсификация CDR" или "диверсифицированные CDR", как использовано в данной заявке, представляют собой модификацию аминокислотных последовательностей с CDRs любым подходящим способом. CDRs представляют собой, как известно, иммуногенсвязывающие участки, поэтому имеющие коллекции, содержащие члены, представляющие большое разнообразие в пределах CDRs, которое увеличивает вероятность того, что коллекция будет содержать антитела или его фрагменты, имеющие специфичность и оптимальные свойства в отношении любого иммуногена. Разнообразие получают путем варьирования аминокислотного состава одного или более CDRs. Этого можно достичь любыми способами, известными специалисту в данной области, включая способы, описанные в данной заявке.
"Коллекция синтетических нуклеиновых кислот, кодирующих антитела или их фрагменты", означает, что все нуклеиновые кислоты, которые кодируют антитело или его фрагмент, являются синтетическими, но не относятся к другим нуклеиновым кислотам, таким как векторы, которые могут быть функциональным образом связаны с такими синтетическими нуклеиновыми кислотами.
Термины, используемые в контексте молекулярной биологии
Термин "синтез" или "синтезировали" означает синтез гена, где последовательности нуклеиновых кислот синтезируют в физическую ДНК, содержащую полинуклеотиды. Стандартный синтез ДНК включает однонуклеотидный синтез, где получают одноцепочечные олигонуклеотиды и затем перекрывающиеся олигонуклеотиды лигируют с использованием ПЦР-подобной сборки. Компании, такие как Sloning (Puchheim, Германия), Geneart (Regensburg, Германия), DNA2.0 (Menio Park, CA, США) и Genscript (Piscataway, NJ, США) предоставляют технологию синтеза генов. Sloning, например, использует набор заранее подготовленных двухцепочечных триплетных нуклеотидов, которые отжигают и последовательно лигируют.
Термин "синтетический" описывает молекулу, которую получают посредством синтеза или синтезируют.
Термин "коллекция" или "библиотека" означает по меньшей мере два члена. Термин "член" включает, но не ограничивается этим, нуклеиновые кислоты, кодирующие антитела или их фрагменты, или сами антитела или их фрагменты.
Термин "хозяин" относится к любому хозяину, включающему млекопитающее, такое как человек, мышь или грызун, мыши, крысы, белки, бурундуки, суслики, дикообразы, бобры, хомяки, песчанки, морские свинки, кролики, собаки, кошки, коровы или лошади.
Термин "нуклеиновая кислота" используют в данной заявке взаимозаменяемо с термином "полинуклеотид" и относится к дезоксирибонуклеотидам или рибонуклеотидам и их полимерам, как в одно-, так и в двухцепочечной форме. Термин охватывает нуклеиновые кислоты, содержащие известные нуклеотидные аналоги или модифицированные скелетные остатки или связи, которые являются синтетическими, встречающимися в природе и не встречающимися в природе, которые имеют сходные связывающие свойства с исходной нуклеиновой кислотой. Примеры таких аналогов включают, без ограничения, фосфоротиоаты, фосфорамидаты, метилфосфонаты, хиральные метилфосфонаты, 2-0-метилрибонуклеотиды и пептид-нуклеиновые кислоты (PNAs).
Если не указано иное, конкретная последовательность нуклеиновой кислоты также косвенно охватывает их консервативно модифицированные варианты (например, замены на основе вырождености кодонов) и комплементарные последовательности, а также косвенно указанную последовательность. В частности, как указано ниже, замены на основе вырождености кодонов могут достигаться путем создания последовательностей, в которых третье положение одного или более выбранных (или всех) кодонов заменено остатком со смешанным основанием (mixed-base) и/или дезоксиинозиновым остатком (Batzer et al., Nucleic Acid Res. 19:5081, 1991; Ohtsuka e/ al., J. Biol. Chem. 260:2605-2608, 1985; и Rossolini et аl., Mоl. Сеll. Probes 8:91-98, 1994).
Термин "функциональным образом связанный" относится к функциональной взаимосвязи между двумя или более полинуклеотидными (например, ДНК) сегментами. Как правило, это относится к функциональной взаимосвязи транскрипционной регуляторной последовательности с транскрипционной последовательностью. Например, промоторная или энхансерная последовательность функциональным образом связана с кодирующей последовательностью, если она стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке-хозяине или другой эспрессирующей системе. Как правило, промоторные транскрипционные регуляторные последовательности, которые функциональным образом связаны с транскрибируемой последовательностью, физически примыкают к транскрибируемой последовательности, т.е. они являются цис-действующими. Однако некоторые транскрипционные регуляторные последовательности, такие как энхансеры, не обязательно должны быть физически смежными или расположенными в непосредственной близости к кодирующим последовательностям, транскрипцию которых они усиливают.
Используемый в данной заявке термин "кодон-оптимизированный" или "оптимизация кодонов" означает, что нуклеотидная последовательность была изменена с целью кодирования аминокислотной последовательности с использованием кодонов, которые являются предпочтительными в продуцирующей клетке или организме. Оптимизированную нуклеотидную последовательность конструируют для сохранения аминокислотной последовательности, первоначально кодируемой исходной нуклеотидной последовательностью. Кроме того, можно сконструировать нуклеотидную последовательность, полностью или насколько возможно лишенных ингибиторных мотивов, сайтов сплайсинга, мотивов нестабильности мРНК и нежелательных сайтов рестрикции. Она также может быть оптимизирована в отношении GC-содержания, желательных сайтов рестрикции и других параметров. Последовательности можно оптимизировать в отношении экспрессии в разных хозяевах, включая бактериальные или эукариотические клетки. Аминокислотные последовательности, кодируемые оптимизированными нуклеотидными последовательностями, также называются оптимизированными.
Термин "аминокислота" относится к природным и синтетическим аминокислотам, а также аминокислотным аналогам и аминокислотным миметикам, которые функционируют способом, аналогичным природным аминокислотам. К природным аминокислотам относятся аминокислоты, кодируемые генетическим кодом, а также аминокислоты, которые впоследствии модифицируются, например, гидроксипролин, y-карбоксиглутамат и 0-фосфосерин. Аминокислотные аналоги относятся к соединениям, которые имеют такую же основную химическую структуру, как и природная аминокислота, т.е. альфа-углерод, который связан с водородом, карбоксильную группу, аминогруппу и R-группу, например, гомосерин, норлейцин, метионинсульфоксид, метионинметилсульфоний. Такие аналоги имеют модифицированные R-группы (например, норлейцин) или модифицированные пептидные скелеты, но сохраняют такую же основную химическую структуру, как и природная аминокислота. Аминокислотные миметики относятся к химическим соединениям, которые имеют структуру, отличную от общей химической структуры аминокислоты, но которые функционируют таким же образом, как природные аминокислоты.
Термины "полипептид" и "белок", используемые взаимозаменяемо в данной заявке, относятся к полимеру из аминокислотных остатков. Данные термины используются для аминокислотных полимеров, в которых один или более аминокислотных остатков представляют собой искусственный химический миметик соответствующей природной аминокислоты, а также природных аминокислотных полимеров и неприродных аминокислотных полимеров. Если не указано иное, конкретная полипептидная последовательность также косвенно охватывает ее консервативно модифицированные варианты.
Термин "консервативно модифицированный вариант" относится как к аминокислотной, так и к нуклеиновокислотной последовательности. В отношении конкретных последовательностей нуклеиновых кислот, консервативно модифицированные варианты относятся к таким нуклеиновым кислотам, которые кодируют идентичные или по существу идентичные аминокислотные последовательности, или если нуклеиновая кислота не кодирует аминокислотную последовательность, то по существу к идентичным последовательностям. Из-за вырожденности генетического кода большое число функционально идентичных нуклеиновых кислот кодирует любой данный белок. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в любом положении, где аланин уточняется кодоном, этот кодон можно заменить на любой из соответствующих описанных кодонов без изменения кодируемого полипептида. Такие варианты нуклеиновых кислот представляют собой "молчащие варианты", которые представляют собой один вид консервативно модифицированных вариантов. Каждая последовательность нуклеиновой кислоты в данной заявке, которая кодирует полипептид, также описывает любой возможный молчащий вариант нуклеиновой кислоты. Специалист поймет, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно является единственным кодоном для метионина, и TGG, который обычно является единственным кодоном для триптофана), который можно модифицировать с получением функционально идентичной молекулы. Соответственно, каждый молчащий вариант нуклеиновой кислоты, которая кодирует полипептид, подразумевается в каждой описанной последовательности.
Для полипептидных последовательностей, "консервативно модифицированные варианты" включают индивидуальные замены, делеции или добавления к полипептидной последовательности, которые приводят к замене аминокислоты на химически подобную аминокислоту. Таблицы консервативных замен, представляющие функционально подобные аминокислоты, хорошо известны в данной области. Такие консервативно модифицированные варианты являются дополнительными и не исключают полиморфных вариантов, межвидовых гомологов и аллелей по изобретению. Следующие восемь групп содержат аминокислоты, которые являются консервативными заменами друг для друга: 1) Аланин (А), Глицин (G); 2) Аспарагиновая кислота(О), Глутаминовая кислота (Е); 3) Аспарагин (N), Глутамин (Q); 4) Аргинин (R), Лизин (К); 5) Изолейцин (I), Лейцин (L), Метионин (М), Валин (V); 6) Фенилаланин (F), Тирозин (Y), Триптофан (W); 7) Серин (S), Треонин (Т); и 8) Цистеин (С), Метионин (М) (см., например, Creighton, Proteins (1984)). В некоторых воплощениях термин "консервативные модификации последовательности" используют для обозначения модификаций аминокислот, которые не оказывают существенного влияния или не изменяют связывающие характеристики антитела, содержащего аминокислотную последовательность.
Термины "идентичный" или "процент идентичности" в контексте двух или более нуклеиновых кислот или полипептидных последовательностей относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми. Две последовательности являются "по существу идентичными", если две последовательности имеют определенный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т.е. 60% идентичности, возможно 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% идентичности в определенной области, или, когда не определено, во всей последовательности), при сравнении и выравнивании для максимального соответствия в пределах окна сравнения или предполагаемой области, как измерено с использованием одного из следующих алгоритмов сравнения последовательностей или выравнивания вручную и визуальной оценки. В некоторых случаях идентичность находится в пределах области, которая имеет длину по меньшей мере примерно 50 нуклеотидов (или 10 аминокислот), или более предпочтительно в пределах области, которая имеет длину от 100 до 500 или 1000 или более нуклеотидов (или 20, 50, 200 или более аминокислот). Для сравнения последовательностей, обычно одна последовательность действует в качестве ссылочной последовательности, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей, тестируемую и ссылочную последовательности вводят в компьютер, задают координаты подпоследовательности, если необходимо, и задают параметры программы алгоритма последовательностей. Можно использовать параметры программы по умолчанию или устанавливать альтернативные параметры. Затем алгоритм сравнения последовательностей рассчитывает процент идентичности последовательностей для тестируемых последовательностей относительно последовательности сравнения на основании параметров программы.
"Окно сравнения", используемое в данной заявке, включает ссылку на сегмент из любого количества смежных положений, выбранных из группы, состоящей из количества от 20 до 600, обычно от примерно 50 до примерно 200, чаще всего от примерно 100 до примерно 150, в котором последовательность можно сравнивать с последовательностью из того же количества смежных положений после оптимального выравнивания этих двух последовательностей. Методы выравнивания последовательностей для их сравнения хорошо известны в области техники. Оптимальное выравнивание последовательностей для их сравнения можно проводить, например, с помощью алгоритма локальной гомологии (Smith and Waterman (1970) Adv. Appl. Math. 2:482c), с помощью алгоритма выравнивания областей гомологии (Needleman and Wunsch, J. Mol. В/о/. 48:443, 1970), с помощью метода поиска сходства (Pearson and Lipman, Proc. Nat'1. Acad. Sci. USA 85:2444, 1988), с помощью компьютерных реализации этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wl), или с помощью выравнивания вручную и визуального оценки (см., например, Brent et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (2003)).
Два примера алгоритмов, которые подходят для определения процента идентичности последовательностей и сходства последовательностей, представляют собой алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul etal., Nucl. Acids Res. 25:3389-3402, 1977; и Altschul et al., J. Mol. В/о/. 215:403-410, 1990, соответственно. Программное обеспечение для выполнения BLAST анализов общедоступно через Национальный центр биотехнологической информации (National Center for Biotechnology Information). Этот алгоритм включает, в первую очередь, идентификацию пар последовательностей с высоким сходством (HSPs, от англ. high scoring sequence pairs) путем идентификации коротких слов длиной W в последовательности запроса (query), которые либо совпадают, либо удовлетворяют некоторому положительному пороговому значению Т при выравнивании со словом такой же длины в последовательности из базы данных. Т называют пороговым счетом соседства слова (neighborhood word score threshold) (Altschul et al., supra). Эти удачные попытки с исходным словом соседства действуют как семена для инициации поисков содержащих его более длинных HSPs. Удачные попытки со словом распространяются в обоих направлениях вдоль каждой последовательности до тех пор, пока общий счет выравнивания может увеличиваться. Общий счет вычисляют, используя для нуклеотидных последовательностей параметры М ("награда" для пары за совпадение остатков; всегда>0) и N ("штраф" за несовпадение остатков; всегда<0). Для аминокислотных последовательностей, для вычисления общего счета используют матрицу замен. Расширение попадания слов в каждом направлении прекращается, когда: суммарный счет выравнивания снижается на величину Х от его максимально достигнутого значения; суммарный счет достигает нуля или ниже из-за накопления одного или более выравниваний негативно-оцененных остатков; или при достижении конца любой последовательности. Параметры W, Т и Х алгоритма BLAST определяют чувствительность и скорость выравнивания. Программа BLASTN (для нуклеотидных последовательностей) использует по умолчанию длину слова (W) 11, ожидание (Е) или 10, М=5, N=-4 и сравнение обеих цепей. Для аминокислотных последовательностей программа BLASTP использует по умолчанию длину слова 3 и ожидание (Е) 10 и матрицу замен BLOSUM62 (см. Henikoffand Henikoff, Proc. Natl. Acad. Sci. USA 89:10915, 1989) выравнивания (В) 50, ожидание (Е) 10, М=5, N=-4 и сравнение обеих цепей.
Алгоритм BLAST также выполняет статистический анализ сходства между двумя последовательностями (см., например, Karlin and Altschul, Proc. Natl. Acad. Sci. USA 90:5873-5787, 1993). Одним из показателей сходства, обеспечиваемого алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), которая позволяет судить о вероятности совпадения между двумя нуклеотидными или аминокислотными последовательностями, которое происходит случайно. Например, нуклеиновую кислоту считают сходной с ссылочной последовательностью, если наименьшая суммарная вероятность при сравнении тестируемой нуклеиновой кислоты с ссылочной нуклеиновой кислотой составляет менее чем примерно 0,2, более предпочтительно менее чем примерно 0,01, и наиболее предпочтительно менее чем примерно 0.001.
Процент идентичности между двумя аминокислотными последовательностями можно также определить с использованием алгоритма Е. Meyers и W. Miller (Comput. Appl. Biosci., 4:11-17, 1988), который был включен в программу ALIGN (версия 2.0), используя таблицу РАМ 120 весов остатков, штраф за длину пробела 12 и штраф за пробел 4. Кроме того, процент идентичности между двумя аминокислотными последовательностями можно определить с использованием алгоритма Needleman и Wunsch (J. Mol. Biol. 48:444-453, 1970), который был включен в программу GAP в пакете программного обеспечения GCG (доступна на сайте afwww.qca.com). используя либо матрицу Blossom 62, либо матрицу РАМ250 и вес за пробел 16, 14, 12, 10, 8, 6 или 4, и вес за длину 1, 2, 3, 4, 5 или 6.
Помимо процента идентичности последовательностей, указанного выше, другим признаком того, что две последовательности нуклеиновой кислоты или полипептиды по существу идентичны, является то, что полипептид, кодируемый первой нуклеиновой кислотой, иммунологически перекрестие реагирует с антителами, полученными против полипептида, кодируемого второй нуклеиновой кислотой, как описано ниже. Таким образом, полипептид обычно по существу идентичен второму полипептиду, например, если два пептида отличаются только консервативными заменами. Другим признаком того, что две последовательности нуклеиновых кислот по существу идентичны, является то, что две молекулы или их комплементы гибридизуются друг с другом в жестких условиях, как описано ниже. Еще одним признаком того, что две последовательности нуклеиновых кислот по существу идентичны, является то, что одинаковые праймеры можно использовать для амплификации последовательности.
Термин "рекомбинантная клетка-хозяин" (или просто "клетка-хозяин") относится к клетке, в которую встроен рекомбинантный экспрессирующий вектор. Следует понимать, что такие термины предназначены для обозначения не только конкретной клетки-субъекта, но и потомства такой клетки. Поскольку некоторые модификации могут происходить в последующих поколениях в результате либо мутации, либо влияния окружающей среды, такое потомство фактически не может быть идентично родительской клетке, но все еще включено в объем термина "клетка-хозяин", используемого в данной заявке. Типичными клетками-хозяевами являются прокариотические клетки (такие как бактериальные, включая, но не ограничиваясь этим, E.coli) или эукариотические (которые включают клетки дрожжей, млекопитающих и более).
Термин "вектор" предназначен для обозначения полинуклеотидной молекулы, способной переносить другой полинуклеотид, с которым он связан. Предпочтительными векторами являются векторы, способные к автономной репликации и/или экспрессии нуклеиновых кислот, с которыми они связаны. Векторы, способные направлять экспрессию нуклеиновых кислот, с которыми они функциональным образом связаны, называются в данной заявке "экспрессирующими векторами". Один тип вектора представляет собой "плазмиду", которая относится к кольцевой двухцепочечной ДНК петле, в которую могут быть лигированы дополнительные сегменты ДНК. Другой тип вектора представляет собой вирусный вектор, где дополнительные сегменты ДНК можно лигировать в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальную точку начала репликация и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина и, таким образом, реплицироваться вместе с геномом хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функциональным образом связаны. Такие векторы называются в данной заявке "рекомбинантными экспрессирующими векторами" (или просто "экспрессирующими векторами"). В общем, экспрессирующие векторы, используемые в методах рекомбинантных ДНК, часто находятся в форме плазмид. В настоящем описании "плазмида" и "вектор" могут использоваться взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора. Однако предполагается, что изобретение включает и другие формы экспрессирующих векторов, такие как вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
Векторы, как правило, включают прокариотический репликон, который может включать прокариотический промотор, способный направлять экспрессию (транскрипцию и трансляцию) VH- и/или VL-кодирующих гомологов в бактериальной клетке-хозяине, такой как Eschehchia со//, трансформированной им. Промотор представляет собой элемент, контролирующий экспрессию, образованный ДНК последовательностью, которая обеспечивает связывание РНК-полимеразы и осуществление транскрипции. Промоторные последовательности, совместимые с бактериальными хозяевами, как правило, представлены в плазмидных векторах, содержащих подходящие сайты рестрикции для вставки ДНК-сегмента. Примеры таких плазмидных векторов включают коммерчески доступные pUC8, pUC9, pBR322 и pBR329, pPL и рКК223.
"Дисплейный вектор" включает последовательность ДНК, обладающую способностью к прямой репликации и поддержанию рекомбинантной молекулы ДНК вне хромосомы в клетке-хозяине, такой как бактериальная клетка-хозяин, трансформированная им. Такие последовательности ДНК хорошо известны в данной области. Дисплейные векторы могут представлять собой, например, фаговые векторы или фагмидные векторы, происходящие из нитевидного бактериофага класса fd, M13 или fl. Такие векторы способны облегчать экспонирование включенного белка, например связывающего белка или его фрагмента, на поверхности нитевидного бактериофага. Дисплейные векторы, подходящие для экспонирования на фаге, рибосомах, ДНК, бактериальных клетках или эукариотических клетках, например клетках дрожжей или млекопитающих, также хорошо известны в данной области, например, как вирусные векторы или векторы, кодирующие химерные белки.
Сайты рестрикции, которые являются "уникальными", представляют собой сайты рестрикции, которые существуют или появляются только один раз на данной молекуле нуклеиновой кислоты.
Коллекции и способы их получения и применения
В настоящем описании представлены коллекции антител или их функциональных фрагментов, которые могут быть использованы в идентификации терапевтического антитела против любой мишени, где антитела могут клинически разрабатываться, являются безопасными и эффективными у пациентов. В качестве предпосылки, авторы изобретения предположили, что пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, распространенные в иммунном репертуаре человека, по-видимому, имеют предпочтительные биофизические свойства, которые будут приводить к более эффективной разработке и увеличению безопасности и эффективности полученных антител у пациентов. Каждая В-клетка кодирует одно антитело, и каждое антитело содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи. Каждую вариабельную область тяжелой цепи и вариабельную область легкой цепи антитела можно выравнивать с генной последовательностью эмбрионального типа с целью определения происхождения антитела, содержания, из которого был образован ген эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи. Поэтому для каждого антитела можно сказать, что вариабельная область тяжелой цепи и вариабельная область легкой цепи содержат пару генов эмбрионального типа, например VH3-23, спаренный с VK1-5. Такие предпочтительные биофизические свойства могут включать: а) высокий относительный уровень экспонирования в Fab-формате; б) высокие относительные уровни экспрессии Fab; в) термостабильность Fab; г) стабильность Fab в сыворотке крупного рогатого скота/мышей; д) высокие относительные уровни экспрессии человеческого IgG1; и е) стабильность человеческого IgG1 в сыворотке крупного рогатого скота.
Для того чтобы доказать гипотезу, что пары генов эмбрионального типа, по-видимому, имеют предпочтительные биофизические свойства, первая стадия заключалась в идентификации пар генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, экспрессирующихся в иммунном репертуаре человека. В некоторых аспектах настоящее изобретение включает способ получения коллекции синтетических антител или их функциональных фрагментов, включающий стадию получения данных о парах генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, присутствующих в иммунном репертуаре человека. В некоторых воплощениях данные получены из общедоступной литературы, в которой предложены пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи. Как правило, в соответствующей общедоступной литературе представлены следующие способы: В-клетки выделяли из людей-доноров, В-клетки сортировали с целью определения стадии их развития или дифференцировки, получали и амплифицировали кДНК, представляющую ДНК, кодирующую антитело из каждой В-клетки, кДНК секвенировали, кДНК, кодирующие вариабельную область тяжелой цепи и вариабельную область легкой цепи, выравнивали с известными генными последовательностями эмбрионального типа и определяли пары генов эмбрионального типа из каждой В-клетки. В некоторых воплощениях данные получали в результате отбора образцов и выделения человеческих В-клеток, которые включали способ, аналогичный способу, используемому в литературе. В этих аспектах способ получения коллекции синтетических антител или их функциональных фрагментов включает стадию получения данных о парах генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, присутствующих в иммунном репертуаре человека; где стадия получения дополнительно включает стадии: аа) выделение человеческих В-клеток из образца; аб) получение кДНК из В-клеток; ав) ПЦР-амплификация кДНК из В-клеток; аг) секвенирование ПЦР-продуктов; и ад) идентификация генов эмбрионального типа ПЦР-продуктов. Оба набора данных представляли пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые присутствуют в иммунном репертуаре человека.
На следующей стадии первоначальные данные собирали, анализировали и пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, присутствующие в иммунном репертуаре человека, располагали в соответствии с уровнем экспрессии. В этих аспектах настоящее изобретение включает способ получения коллекции антител или их функциональных фрагментов, включающий идентификацию пар генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека.
Пары генов эмбрионального типа, преимущественно экспрессирующиеся в иммунном репертуаре человека
Исходя из этих данных, стало ясно, что в иммунном репертуаре человека некоторые пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи присутствуют чаще, чем другие. Поскольку эти главные пары, как ожидается, имеют лучшие биофизические свойства, то аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, происходящих из пар генов эмбрионального типа, которые выделяются в иммунном репертуаре человека, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов происходят из пар генов эмбрионального типа, которые выделяются в иммунном репертуаре человека. В других аспектах настоящее изобретение включает коллекции синтетических антител или их функциональных фрагментов, содержащие по существу белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат по существу белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека. В других аспектах настоящее изобретение включает коллекции синтетических антител или их функциональных фрагментов, содержащие белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека. В некоторых аспектах настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пар белков эмбрионального типа, где указанные гены эмбрионального типа преимущественно экспрессируются в иммунном репертуаре человека.
В некоторых воплощениях настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, где указанные каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи состоят по существу из белковых последовательностей эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека. В некоторых воплощениях антитела или их функциональные фрагменты состоят по существу из пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека, где в некоторых воплощениях один или более CDRs состоят по существу из белковых последовательностей эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека. В некоторых воплощениях настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, где указанные каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи состоят из белковых последовательностей эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека. В некоторых воплощениях антитела или их функциональные фрагменты состоят из пар белков эмбрионального типа, которые кодируются парами генами эмбрионального типа, преимущественно экспрессирующимися в иммунном репертуаре человека, где в некоторых воплощениях один или более CDRs состоят по существу из белковых последовательностей эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека. В некоторых воплощениях большая часть или по существу все антитела или их функциональные фрагменты из данных коллекций содержат белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека.
В некоторых воплощениях пары генов эмбрионального типа, которые часто встречаются или преимущественно экспрессируются в иммунном репертуаре человека, экспрессируются в концентрации по меньшей мере 0,05% в иммунном репертуаре человека, по меньшей мере 0,09% в иммунном репертуаре человека; по меньшей мере 0,14% в иммунном репертуаре человека; по меньшей мере 0,19% в иммунном репертуаре человека; по меньшей мере 0,23% в иммунном репертуаре человека; по меньшей мере 0,28% в иммунном репертуаре человека; по меньшей мере 0,33% в иммунном репертуаре человека; по меньшей мере 0,37% в иммунном репертуаре человека; по меньшей мере 0,42% в иммунном репертуаре человека; по меньшей мере 0,47% в иммунном репертуаре человека; по меньшей мере 0,51% в иммунном репертуаре человека; по меньшей мере 0,56% в иммунном репертуаре человека; по меньшей мере 0,61% в иммунном репертуаре человека; по меньшей мере 0,66% в иммунном репертуаре человека; по меньшей мере 0,70% в иммунном репертуаре человека; по меньшей мере 0,84% в иммунном репертуаре человека; по меньшей мере 0,89% в иммунном репертуаре человека; по меньшей мере 0,94% в иммунном репертуаре человека; по меньшей мере 1,03% в иммунном репертуаре человека; по меньшей мере 1,12% в иммунном репертуаре человека; по меньшей мере 1,17% в иммунном репертуаре человека; или по меньшей мере 1,26% в иммунном репертуаре человека.
Дополнительный аспект настоящего изобретения состоит в возможности использовать коллекцию в идентификации антител или их функциональных фрагментов против любого иммуногена. Предположили, что создание коллекций по меньшей мере с двумя парами белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека, могло бы обеспечить разнообразие в пределах коллекции, особенно в пределах определяющих комплементарность областей антител из коллекции, в отношении длины CDR и разнообразия конформаций или канонических структур. Это позволяет использовать коллекции по настоящему изобретению в идентификации антител или их функциональных фрагментов против любого иммуногена. Поэтому некоторые аспекты изобретения включают коллекции, содержащие антитела или их функциональные фрагменты, содержащие по меньшей мере две различные пары белков эмбрионального типа; по меньшей мере три различные пары белков эмбрионального типа; по меньшей мере четыре различные пары белков эмбрионального типа; по меньшей мере пять различных пар белков эмбрионального типа; по меньшей мере шесть различных пар белков эмбрионального типа; по меньшей мере семь различных пар белков эмбрионального типа; по меньшей мере восемь различных пар белков эмбрионального типа; по меньшей мере девять различных пар белков эмбрионального типа; по меньшей мере десять различных пар белков эмбрионального типа; по меньшей мере одиннадцать различных пар белков эмбрионального типа; по меньшей мере двенадцать различных пар белков эмбрионального типа; по меньшей мере тринадцать различных пар белков эмбрионального типа; по меньшей мере четырнадцать различных пар белков эмбрионального типа; по меньшей мере пятнадцать различных пар белков эмбрионального типа; по меньшей мере шестнадцать различных пар белков эмбрионального типа; по меньшей мере семнадцать различных пар белков эмбрионального типа; по меньшей мере восемнадцать различных пар белков эмбрионального типа; по меньшей мере девятнадцать различных пар белков эмбрионального типа; по меньшей мере двадцать различных пар белков эмбрионального типа; по меньшей мере 21 различную пару белков эмбрионального типа; по меньшей мере 22 различные пары белков эмбрионального типа; по меньшей мере 23 различные пары белков эмбрионального типа; по меньшей мере 24 различные пары белков эмбрионального типа; по меньшей мере 25 различных пар белков эмбрионального типа; по меньшей мере 26 различных пар белков эмбрионального типа; по меньшей мере 27 различных пар белков эмбрионального типа; по меньшей мере 28 различных белков эмбрионального типа вариабельных областей тяжелых цепей; по меньшей мере 29 различных последовательностей пар белков эмбрионального типа; по меньшей мере 30 различных пар белков эмбрионального типа; по меньшей мере 31 различную пару белков эмбрионального типа; по меньшей мере 32 различные пары белков эмбрионального типа; по меньшей мере 33 различные пары белков эмбрионального типа; по меньшей мере 34 различные пары белков эмбрионального типа; по меньшей мере 35 различных пар белков эмбрионального типа; по меньшей мере 36 различных пар белков эмбрионального типа; по меньшей мере 37 различных пар белков эмбрионального типа; по меньшей мере 38 различных пар белков эмбрионального типа; по меньшей мере 39 различных пар белков эмбрионального типа; по меньшей мере 40 различных пар белков эмбрионального типа; по меньшей мере 41 различную пару белков эмбрионального типа; по меньшей мере 42 различные пары белков эмбрионального типа; по меньшей мере 43 различные пары белков эмбрионального типа; по меньшей мере 44 различных белков эмбрионального типа вариабельных областей тяжелых цепей; по меньшей мере 45 различных последовательностей пар белков эмбрионального типа; по меньшей мере 46 различных пар белков эмбрионального типа; по меньшей мере 47 различных пар белков эмбрионального типа; по меньшей мере 48 различных пар белков эмбрионального типа; по меньшей мере 49 различных пар белков эмбрионального типа; или по меньшей мере 50 различных пар белков эмбрионального типа, выбранных из преимущественно экспрессирующихся пар белков эмбрионального типа иммунного репертуара человека.
В некоторых воплощениях коллекции содержат каркасные участки вариабельной области тяжелой и вариабельной области легкой цепи, содержащие одну или более пар белков эмбрионального типа, выбранных из пар генов эмбрионального типа, показанных в Таблице 18.
В некоторых воплощениях настоящее изобретение включает выделенное антитело или его функциональный фрагмент, содержащие вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, содержащий FR1, CDR1, FR2, CDR2 и FR3, содержащие белковые последовательности эмбрионального типа пары генов эмбрионального типа, где пара генов эмбрионального типа выбрана из пар генов эмбрионального типа из Таблицы 18.
Пары генов эмбрионального типа, преимущественно экспрессирующиеся в иммунном репертуаре интактного человека
Также предполагают, что некоторые пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи могут по-разному экспрессироваться в "наивных" В-клетках ("необученных" антигеном) по сравнению с "обученными" антигеном В-клетками, поэтому данные анализировали на основании развития или дифференцировки выбранных В-клеток. Коллекции, содержащие пары белков эмбрионального типа пар генов эмбрионального типа, по-разному экспрессирующиеся в "наивных" В-клетках, могут быть выгодными для выбора антител или их функциональных фрагментов против любого иммуногена. Поэтому аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, происходящие из пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов происходят из пар генов эмбрионального типа, которые преимущественно экспрессируется в иммунном репертуаре интактного человека. В других аспектах настоящее изобретение включает коллекции синтетических антител или их функциональных фрагментов, содержащие по существу белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат по существу белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека. В других аспектах настоящее изобретение включает коллекции синтетических антител или их функциональных фрагментов, содержащие белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека. В некоторых аспектах настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа и где указанные белковые последовательности эмбрионального типа содержат пары белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные пары белков эмбрионального типа кодируются парами генов эмбрионального типа, преимущественно экспрессирующимися в иммунном репертуаре интактного человека.
В некоторых воплощениях настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, где указанные каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи состоят по существу из белковых последовательностей эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека. В некоторых воплощениях антитела или их функциональные фрагменты состоят по существу из пар белков эмбрионального типа, кодируемых парами генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека. В некоторых воплощениях настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, где указанные каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи состоят из белковых последовательностей эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека. В некоторых воплощениях антитела или их функциональные фрагменты состоят из пар белков эмбрионального типа, которые кодируются парами генов эмбрионального типа, преимущественно экспрессирующимися в иммунном репертуаре интактного человека. В некоторых воплощениях большинство или по существу все антитела или их функциональные фрагменты из коллекций содержат белковые последовательности эмбрионального типа пар генов эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека.
В некоторых воплощениях пары генов эмбрионального типа, которые часто встречаются или преимущественно экспрессируются в иммунном репертуаре интактного человека, экспрессируются в концентрации по меньшей мере 0,07% в иммунном репертуаре интактного человека, по меньшей мере 0,15% в иммунном репертуаре интактного человека; по меньшей мере 0,22% в иммунном репертуаре интактного человека по меньшей мере 0,30% в иммунном репертуаре интактного человека по меньшей мере 0,37% в иммунном репертуаре интактного человека по меньшей мере 0,45% в иммунном репертуаре интактного человека по меньшей мере 0,52% в иммунном репертуаре интактного человека по меньшей мере 0,59% в иммунном репертуаре интактного человека по меньшей мере 0,67% в иммунном репертуаре интактного человека по меньшей мере 0,74% в иммунном репертуаре интактного человека по меньшей мере 0,82% в иммунном репертуаре интактного человека по меньшей мере 0,89% в иммунном репертуаре интактного человека по меньшей мере 0,97% в иммунном репертуаре интактного человека по меньшей мере 1,19% в иммунном репертуаре интактного человека; или по меньшей мере 1,56% в иммунном репертуаре интактного человека.
Дополнительный аспект настоящего изобретения состоит в возможности использовать коллекции в идентификации антител или их функциональных фрагментов против любого иммуногена. Предположили, что создание коллекций по меньшей мере с двумя парами белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые кодируются парами генов эмбрионального типа, преимущественно экспрессирующимися в иммунном репертуаре интактного человека, могло бы обеспечить разнообразие в пределах коллекции, особенно в пределах определяющих комплементарность областей антител из коллекции, в отношении длины CDR и разнообразия конформаций или канонических структур. Это позволяет использовать коллекции по настоящему изобретению в идентификации антител или их функциональных фрагментов против любого иммуногена. Поэтому некоторые аспекты изобретения включают коллекции, содержащие антитела или их функциональные фрагменты, содержащие по меньшей мере две различные пары белков эмбрионального типа; по меньшей мере три различные пары белков эмбрионального типа; по меньшей мере четыре различные пары белков эмбрионального типа; по меньшей мере пять различных пар белков эмбрионального типа; по меньшей мере шесть различных пар белков эмбрионального типа; по меньшей мере семь различных пар белков эмбрионального типа; по меньшей мере восемь различных пар белков эмбрионального типа; по меньшей мере девять различных пар белков эмбрионального типа; по меньшей мере десять различных пар белков эмбрионального типа; по меньшей мере одиннадцать различных пар белков эмбрионального типа; по меньшей мере двенадцать различных пар белков эмбрионального типа; по меньшей мере тринадцать различных пар белков эмбрионального типа; по меньшей мере четырнадцать различных пар белков эмбрионального типа; по меньшей мере пятнадцать различных пар белков эмбрионального типа; по меньшей мере шестнадцать различных пар белков эмбрионального типа; по меньшей мере семнадцать различных пар белков эмбрионального типа; по меньшей мере восемнадцать различных пар белков эмбрионального типа; по меньшей мере девятнадцать различных пар белков эмбрионального типа; по меньшей мере двадцать различных пар белков эмбрионального типа; по меньшей мере 21 различную пару белков эмбрионального типа; по меньшей мере 22 различные пары белков эмбрионального типа; по меньшей мере 23 различные пары белков эмбрионального типа; по меньшей мере 24 различные пары белков эмбрионального типа; по меньшей мере 25 различных пар белков эмбрионального типа; по меньшей мере 26 различных пар белков эмбрионального типа; по меньшей мере 27 различных пар белков эмбрионального типа; по меньшей мере 28 различных белков эмбрионального типа вариабельных областей тяжелых цепей; по меньшей мере 29 различных последовательностей пар белков эмбрионального типа; по меньшей мере 30 различных пар белков эмбрионального типа; по меньшей мере 31 различную пару белков эмбрионального типа; по меньшей мере 32 различные пары белков эмбрионального типа; по меньшей мере 33 различные пары белков эмбрионального типа; по меньшей мере 34 различные пары белков эмбрионального типа; по меньшей мере 35 различных пар белков эмбрионального типа; по меньшей мере 36 различных пар белков эмбрионального типа; по меньшей мере 37 различных пар белков эмбрионального типа; по меньшей мере 38 различных пар белков эмбрионального типа; по меньшей мере 39 различных пар белков эмбрионального типа; по меньшей мере 40 различных пар белков эмбрионального типа; по меньшей мере 41 различную пару белков эмбрионального типа; по меньшей мере 42 различные пары белков эмбрионального типа; по меньшей мере 43 различные пары белков эмбрионального типа; по меньшей мере 44 различных белков эмбрионального типа вариабельных областей тяжелых цепей; по меньшей мере 45 различных последовательностей пар белков эмбрионального типа; по меньшей мере 46 различных пар белков эмбрионального типа; по меньшей мере 47 различных пар белков эмбрионального типа; по меньшей мере 48 различных пар белков эмбрионального типа; по меньшей мере 49 различных пар белков эмбрионального типа; по меньшей мере 49 различных пар белков эмбрионального типа; или по меньшей мере 50 различных пар белков эмбрионального типа.
В некоторых воплощениях коллекции содержат каркасные участки вариабельной области тяжелой и вариабельной области легкой цепи, содержащие одну или более пар белков эмбрионального типа, выбранных из пар генов эмбрионального типа из Таблицы 19.
В некоторых воплощениях настоящее изобретение включает выделенное антитело или его функциональный фрагмент, содержащие вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, содержащие FR1, CDR1, FR2, CDR2 и FR3, содержащие белковые последовательности эмбрионального типа, содержащие пару белков эмбрионального типа, где указанная пара белков эмбрионального типа выбрана из пар генов эмбрионального типа из Таблицы 19.
Гены эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи, преимущественно экспрессирующиеся в иммунном репертуаре человека
На следующей стадии объединенные данные и дополнительные данные анализировали для определения экспрессии генов эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи в иммунном репертуаре человека. Поэтому дополнительные аспекты настоящего изобретения включают способы получения коллекции антител или их функциональных фрагментов, включающие стадию идентификации генов эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека. Один способ сделать это заключается в ранжировании генов эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи на основании уровня их экспрессии.
Антитела, содержащие белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, кодируемые генами эмбрионального типа, преимущественно экспрессирующимися в иммунном репертуаре человека, по-видимому, имеют предпочтительные биофизические свойства, которые улучшают разработку, безопастность и эффективность у пациентов. Поэтому аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, происходящие из генов эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов происходят из генов эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека. В других аспектах настоящее изобретение включает коллекции синтетических антител или их функциональных фрагментов, содержащие по существу белковые последовательности эмбрионального типа генов эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат по существу белковые последовательности эмбрионального типа генов эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре интактного человека. В других аспектах настоящее изобретение включает коллекции синтетических антител или их функциональных фрагментов, содержащие белковые последовательности эмбрионального типа генов эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат белковые последовательности эмбрионального типа генов эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека. В некоторых аспектах настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа, где указанные белковые последовательности эмбрионального типа кодируются генами эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека.
В некоторых воплощениях настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, где указанные каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи состоят по существу из белковых последовательностей эмбрионального типа генов эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека. В некоторых воплощениях антитела или их функциональные фрагменты состоят по существу из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, кодируемых генами эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека. В некоторых воплощениях настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, где указанные каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи состоят из белковых последовательностей эмбрионального типа генов эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, которые преимущественно экспрессируются в иммунном репертуаре человека. В некоторых воплощениях антитела или их функциональные фрагменты состоят из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи, кодируемых генами эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре человека. В некоторых воплощениях большинство или по существу все антитела или их функциональные фрагменты из коллекций содержат белковые последовательности эмбрионального типа, кодируемые генами эмбрионального типа, которые преимущественно экспрессируются в иммунном репертуаре интактного человека.
В некоторых воплощениях гены вариабельной области тяжелой цепи эмбрионального типа, которые часто встречаются или преимущественно экспрессируются в иммунном репертуаре человека, экспрессируются в концентрации по меньшей мере 0,1% в иммунном репертуаре человека; по меньшей мере 0,2% в иммунном репертуаре человека; по меньшей мере 0,3% в иммунном репертуаре человека; по меньшей мере 0,4% в иммунном репертуаре человека; по меньшей мере 0,5% в иммунном репертуаре человека; по меньшей мере 0,6% в иммунном репертуаре человека; по меньшей мере 1,0% в иммунном репертуаре человека; по меньшей мере 1,6% в иммунном репертуаре человека; по меньшей мере 2,1% в иммунном репертуаре человека; по меньшей мере 2,2% в иммунном репертуаре человека; по меньшей мере 2,6% в иммунном репертуаре человека; по меньшей мере 2,7% в иммунном репертуаре человека; по меньшей мере 3,0% в иммунном репертуаре человека; по меньшей мере 3,2% в иммунном репертуаре человека; по меньшей мере 3,3% в иммунном репертуаре человека; по меньшей мере 4,0% в иммунном репертуаре человека; по меньшей мере 4,1% в иммунном репертуаре человека; по меньшей мере 4,5% в иммунном репертуаре человека; по меньшей мере 4,6% в иммунном репертуаре человека; по меньшей мере 5,3% в иммунном репертуаре человека; по меньшей мере 5,8% в иммунном репертуаре человека; по меньшей мере 6,8% в иммунном репертуаре человека; по меньшей мере 7,6% в иммунном репертуаре человека; по меньшей мере 8,0% в иммунном репертуаре человека или по меньшей мере 10,6% в иммунном репертуаре человека.
В некоторых воплощениях гены вариабельной области к-легкой цепи эмбрионального типа, которые часто встречаются или преимущественно экспрессируются в иммунном репертуаре человека, экспрессируются в концентрации по меньшей мере 0,1% в иммунном репертуаре человека; по меньшей мере 0,2% в иммунном репертуаре человека; по меньшей мере 0,3% в иммунном репертуаре человека; по меньшей мере 0,4% в иммунном репертуаре человека; по меньшей мере 0,5% в иммунном репертуаре человека; по меньшей мере 0,7% в иммунном репертуаре человека; по меньшей мере 1,0% в иммунном репертуаре человека; по меньшей мере 1,1% в иммунном репертуаре человека; по меньшей мере 1,3% в иммунном репертуаре человека; по меньшей мере1.9% в иммунном репертуаре человека; по меньшей мере 2,2% в иммунном репертуаре человека; по меньшей мере 2,4% в иммунном репертуаре человека; по меньшей мере 2,6% в иммунном репертуаре человека; по меньшей мере 4,6% в иммунном репертуаре человека; по меньшей мере 6,0% в иммунном репертуаре человека; по меньшей мере 7,6% в иммунном репертуаре человека; по меньшей мере 8,5% в иммунном репертуаре человека; по меньшей мере 11,1% в иммунном репертуаре человека; по меньшей мере 11,2% в иммунном репертуаре человека; по меньшей мере 14,2% в иммунном репертуаре человека; или по меньшей мере 16,2% в иммунном репертуаре человека
В некоторых воплощениях гены эмбрионального типа вариабельной области λ-легкой цепи, которые часто встречаются или преимущественно экспрессируются в иммунном репертуаре человека, экспрессируюся в концентрации по меньшей мере 0,1% в иммунном репертуаре человека; по меньшей мере 0,3% в иммунном репертуаре человека; по меньшей мере 0,5% в иммунном репертуаре человека; по меньшей мере 0,6% в иммунном репертуаре человека; по меньшей мере 1,0% в иммунном репертуаре человека; по меньшей мере 1,2% в иммунном репертуаре человека; по меньшей мере 1,5% в иммунном репертуаре человека; по меньшей мере 1,7% в иммунном репертуаре человека; по меньшей мере 4,5% в иммунном репертуаре человека; по меньшей мере 5,1% в иммунном репертуаре человека; по меньшей мере 5,3% в иммунном репертуаре человека; по меньшей мере 6,5% в иммунном репертуаре человека; по меньшей мере 8,1% в иммунном репертуаре человека; по меньшей мере 10,0% в иммунном репертуаре человека; по меньшей мере 11,3% в иммунном репертуаре человека; или по меньшей мере 18,1% в иммунном репертуаре человека
Дополнительный аспект настоящего изобретения состоит в возможности использовать коллекции в идентификации антител или их функциональных фрагментов против любого иммуногена. Предположили, что создание коллекций с одним или более генами вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепью эмбрионального типа, преимущественно экспрессирующимися в иммунном репертуаре человека, могло бы обеспечить разнообразие в пределах коллекции, особенно в длине CDR и конформациях или канонических структурах, которые позволяют использовать данные коллекции в идентификации антител или их функциональных фрагментов против любого иммуногена. Воплощения настоящего изобретения включают коллекции, содержащие антитела или их функциональные фрагменты, содержащие по меньшей мере две различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере три различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере четыре различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере пять различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере шесть различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере семь различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере восемь различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере девять различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере десять различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере одиннадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере двенадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере тринадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере четырнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере пятнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере шестнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере семнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере восемнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере девятнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере двадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 21 различную белковую последовательность эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 22 различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 23 различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 24 различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 25 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 26 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 27 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 28 различных вариабельных областей тяжелой цепи белка эмбрионального типа; по меньшей мере 29 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 30 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 31 различную белковую последовательность эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 32 различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 33 различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 34 различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 35 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 36 различных белковых последовательностей эмбрионального типа вариабельной области тяжелей цепи; по меньшей мере 37 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 38 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 39 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 40 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 41 различную белковую последовательность эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 42 различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 43 различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 44 различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 45 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 46 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 47 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 48 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 49 различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи.
Воплощения настоящего изобретения включают коллекции, содержащие антитела или их функциональные фрагменты, содержащие по меньшей мере две белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере три различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере четыре различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере пять различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере шесть различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере семь различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере восемь различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере девять различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере десять различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере одиннадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере двенадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере тринадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере четырнадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере пятнадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере шестнадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере семнадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере восемнадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере девятнадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере двадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 21 различную белковую последовательность эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 22 различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 23 различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 24 различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 25 различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 26 различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 27 различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 28 различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 29 различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 30 различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 31 различную белковую последовательность вариабельной области к-легкой цепи эмбрионального типа; по меньшей мере 32 различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 33 различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 34 различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере 35 различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи.
Воплощения настоящего изобретения включают коллекции, содержащие антитела или их функциональные фрагменты, содержащие по меньшей мере две различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере три различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере четыре различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере пять различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере шесть различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере семь различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере восемь различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере девять различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере десять различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере одиннадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере двенадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере тринадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере четырнадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере пятнадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере шестнадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере семнадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере восемнадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере девятнадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере двадцать различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 21 различную белковую последовательность эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 22 различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 23 различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 24 различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 25 различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 26 различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 27 различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 28 различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 29 различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 30 различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 31 различную белковую последовательность эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 32 различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере 33 различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи.
В некоторых воплощениях коллекции содержат антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой и вариабельной области легкой цепи, где указанные каркасные участки содержат одну или более белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, выбранных из группы, состоящей из: IGHV3-23; IGHV3-30; IGHV4-39; IGHV4-34; IGHV4-59; IGHV1-69; IGHV5-51; IGHV3-7; IGHV1-18; IGHV3-48; IGHV3-15; IGHV3-21; IGHV1-2; IGHV3-33; IGHV4-31; IGHV3-53; IGHV3-11; IGHV3-9; IGHV4-4; IGHV1-46; IGHV3-74; IGHV1-24; IGHV4-61; IGHV1-8; IGHV1-3; IGHV3-49; IGHV3-43; IGHV4-28; IGHV3-64 и IGHV7-81.
В некоторых воплощениях коллекции содержат антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой и вариабельной области легкой цепи, где указанные каркасные участки содержат одну или более белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи, выбранных из группы, состоящей из: IGKV3-20; IGKV1-39/1D-39; IGKV1-5; IGKV3-15; IGKV4-1; IGKV3-11; IGKV2-28/2D-28; IGKV1-33/1D-33; IGKV2-30; IGKV1-9; IGKV1-17; IGKV1-27; IGKV1-8; IGKV1-16; IGKV1-6; IGKV1-12; IGKV2D-29; IGKV1-13; IGKV1D-8HlGKV2-24.
В некоторых воплощениях коллекции содержат антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой и вариабельной области легкой цепи, где указанные каркасные участки содержат одну или более белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи, выбранных из группы, состоящей из: IGLV2-14; IGLV1-40; IGLV1-44; IGLV1-51; IGLV2-23; IGLV3-21; IGLV1-47; IGLV3-1; IGLV2-11; IGLV2-8; IGLV6-57; IGLV3-25; IGLV7-46; IGLV1-36; IGLV7-43; IGLV9-49; IGLV4-69; IGLV2-18; IGLV3-10 и IGLV3-27.
В некоторых воплощениях настоящее изобретение включает выделенное антитело или его функциональный фрагмент, содержащие FR1, CDR1, FR2, CDR2 и FR3, содержащие белковые последовательности эмбрионального типа, выбранные из группы, состоящей из: IGHV3-23; IGHV3-30; IGHV4-39; IGHV4-34; IGHV4-59; IGHV1-69; IGHV5-51; IGHV3-7; IGHV1-18; IGHV3-48; IGHV3-15; IGHV3-21; IGHV1-2; IGHV3-33; IGHV4-31; IGHV3-53; IGHV3-11; IGHV3-9; IGHV4-4; IGHV1-46; IGHV3-74; IGHV1-24; IGHV4-61; IGHV1-8; IGHV1-3; IGHV3-49; IGHV3-43; IGHV4-28; IGHV3-64; IGHW-81; IGKV3-20; IGKV1-39/1D-39; IGKV1-5; IGKV3-15; IGKV4-1; IGKV3-11; IGKV2-28/2D-28; IGKV1-33/1D-33; IGKV2-30; IGKV1-9; IGKV1-17; IGKV1-27; IGKV1-8; IGKV1-16; IGKV1-6; IGKV1-12; IGKV2D-29; IGKV1-13; IGKV1D-8; IGKV2-24; IGLV2-14; IGLV1-40; IGLV1-44; IGLV1-51; IGLV2-23; IGLV3-21; IGLV1-47; IGLV3-1; IGLV2-11; IGLV2-8; IGLV6-57; IGLV3-25; IGLV7-46; IGLV1-36; IGLV7-43; IGLV9-49; IGLV4-69; IGLV2-18; IGLV3-10;и IGLV3-27.
Гены эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи, имеющие предпочтительные биофизические свойства
На следующей стадии оценивали главные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи для определения их биофизических свойств, связанных с разработкой. Белковые последовательности эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи оценивали in silico в отношении следующих свойств: длина CDR, изоэлектрическая точка (рl), где предпочтительная изоэлектрическая точка составляет 7,5 или выше, поскольку это должно обеспечивать стабильность в стандартном буфере для приготовления препаратов с рН от 5,5 до 7, посттрансляционные модификации (РТМ) в участках, определяющих комплементарность (конкретно, N-связанных сайтах гликозилирования (N×S или N×T), или химические модификации, такие как отщепление Asp (часто в DP), Asp изомеризация (DD, DG), дезамидирование (NS, NG), которое может происходить in vivo (в сыворотке) или при хранении в буфере для приготовления препаратов и приводит к потере связывания антитела), присутствие метионинов в CDRs (могут окисляться при воздействии растворителя), присутствие неспаренных цистеинов (будут образовывать дисульфидные связи с любым другим неспаренным цистеином, приводя таким образом к сшиванию белков и/или более низким уровням экспрессии), отклонения от эмбрионального типа, присутствие потенциальных Т-клеточных эпитопов и теоретическая склонность к агрегации.
В некоторых воплощениях настоящее изобретение включает способ получения коллекции синтетических антител или их функциональных фрагментов, включающий стадии: а) идентификации белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, обладающих следующими свойствами: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; и 6) изоэлектрическую точку по меньшей мере 7,5; и б) создания коллекции антител или их функциональных фрагментов, содержащей последовательности генов эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, идентифицированные в (а)
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, происходящих из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих одно или более из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов происходят из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих такие свойства.
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, содержащие по существу белковые последовательности эмбрионального типа белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих одно или более из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат по существу белковые последовательности эмбрионального типа из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих такие свойства.
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, содержащие белковые последовательности эмбрионального типа белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих одно или более из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи белка эмбрионального типа, имеющего такие свойства.
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, содержащие белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, не содержащие неспаренные цистеины.
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, содержащие белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, содержащие четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; три или меньше посттрансляционных модификаций в участках, определяющих комплементарность; две или меньше посттрансляционных модификаций в участках, определяющих комплементарность; одну или меньше посттрансляционных модификаций в участках, определяющих комплементарность или никаких посттрансляционных модификаций в участках, определяющих комплементарность.
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, происходящих из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих изоэлектрическую точку по меньшей мере 7,5; по меньшей мере 8,0; по меньшей мере 8,5; по меньшей мере 9 или по меньшей мере 9,5.
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, содержащие каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющие одно или более из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, содержащие по меньшей мере два из следующих свойств:
1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющие по меньшей мере четыре из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющие по меньшей мере четыре из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях коллекции синтетических антител или их фунциональные фрагменты содержат белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющие по меньшей мере пять из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях коллекции синтетических антител или их фунциональные фрагменты содержат белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющие следующие свойства: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; и 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки состоят по существу из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих одно или более из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях антитела или их функциональные фрагменты состоят по существу из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих одно или более из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки состоят из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих одно или более из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях антитела или их функциональные фрагменты состоят из белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющих одно или более из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях большинство или по существу все антитела или их функциональные фрагменты из коллекций содержат белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, имеющие одно или более из следующих свойств: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
Некоторые воплощения включают коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа, где указанные белковые последовательности эмбрионального типа имеют следующие свойства: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; и 5) изоэлектрическую точку по меньшей мере 7,5.
Некоторые воплощения включают коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа, где указанные белковые последовательности эмбрионального типа имеют следующее свойство: 1) один или меньше неспаренных цистеинов.
Некоторые воплощения включают коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа, где указанные белковые последовательности эмбрионального типа имеют следующие свойства: 1) один или меньше неспаренных цистеинов; 2) один или меньше потенциальных Т-клеточных эпитопов.
Некоторые воплощения включают коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа, где указанные белковые последовательности эмбрионального типа имеют следующие свойства: 1) один или меньше неспаренных цистеинов; 2) один или меньше потенциальных Т-клеточных эпитопов; и 3) изоэлектрическую точку по меньшей мере 7,5.
Некоторые воплощения включают коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа, где указанные белковые последовательности эмбрионального типа имеют следующие свойства: 1) два или меньше метионинов в участках, определяющих комплементарность; 2) один или меньше неспаренных цистеинов; 3) один или меньше потенциальных Т-клеточных эпитопов; и 4) изоэлектрическую точку по меньшей мере 7,5.
Дополнительный аспект настоящего изобретения состоит в возможности использовать коллекции в идентификации антител или их функциональных фрагментов против любого иммуногена. Предположили, что создание коллекций с одной или более белковыми последовательностями эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи могло бы обеспечить разнообразие в пределах коллекции, особенно в длине CDR и конформациях или канонических структурах, таким образом позволяя использовать коллекцию в идентификации антител или их функциональных фрагментов против любого иммуногена.
Воплощения настоящего изобретения включают коллекции, содержащие антитела или их функциональные фрагменты, содержащие по меньшей мере две различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере три различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере четыре различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере пять различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере шесть различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере семь различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере восемь различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере девять различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере десять различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере одиннадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере двенадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере тринадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере четырнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере пятнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере шестнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере семнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере восемнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, по меньшей мере девятнадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи или по меньшей мере двадцать различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, имеющие следующие свойства: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
Воплощения настоящего изобретения включают коллекции, содержащие антитела или их функциональные фрагменты, содержащие по меньшей мере две различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере три различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере четыре различные белковые последовательности эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере пять различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере шесть различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере семь различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере восемь различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере девять различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере десять различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере одиннадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи; по меньшей мере двенадцать различных белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи, обладающих следующими свойствами: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
Воплощения настоящего изобретения включают коллекции, содержащие антитела или их функциональные фрагменты, содержащие по меньшей мере две различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере три различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере четыре различные белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере пять различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере шесть различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере семь различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи; по меньшей мере восемь различных белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи, обладающих следующими свойствами: 1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность; 2) два или меньше метионинов в участках, определяющих комплементарность; 3) один или меньше неспаренных цистеинов; 4) один или меньше потенциальных Т-клеточных эпитопов; 5) среднюю или низкую склонность к агрегации; или 6) изоэлектрическую точку по меньшей мере 7,5.
В некоторых воплощениях коллекции содержат антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат одну или более белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи, выбранных из группы, состоящей из: IGHV1-2; IGHV1-18; IGHV1-69; IGHV1-46; IGHV3-7; IGHV3-11; IGHV3-15; IGHV3-21; IGHV3-23; IGHV3-30; IGHV3-33; IGHV3-48; IGHV3-53; IGHV3-73; IGH3-74; IGHV4-4; IGHV4-31; IGHV4-39; IGHV 5-51 и IGHV6-1.
В некоторых воплощениях коллекции содержат антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат одну или более белковых последовательностей эмбрионального типа вариабельной области к-легкой цепи, выбранных из группы, состоящей из: IGKV1-5; IGKV1-6; IGKV1-9; IGKV1-12; IGKV1-16; IGKV1-17; IGKV1-27; IGKV1-39; IGKV2-30; IGKV3-11; IGKV3-15 и IGKV3-20.
В некоторых воплощениях коллекции содержат антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат одну или более белковых последовательностей эмбрионального типа вариабельной области λ-легкой цепи, выбранных из группы, состоящей из: IGLV1-40; IGLV1-47; IGLV1-51; IGLV2-11; IGLV2-23; IGLV2-14; IGLV3-1 и IGLV3-21.
В некоторых воплощениях настоящее изобретение включает выделенное антитело или его функциональный фрагмент, содержащие FR1, CDR1, FR2, CDR2 и FR3, содержащие белковые последовательности эмбрионального типа, выбранные из группы, состоящей из: IGHV1 -2; IGHV1 -18; IGHV1 -69; IGHV1-46; IGHV3-7; IGHV3-11; IGHV3-15; IGHV3-21; IGHV3-23; IGHV3-30; IGHV3-33; IGHV3-48; IGHV3-53; IGHV3-73; IGH3-74; IGHV4-4; IGHV4-31; IGHV4-39; IGHV 5-51; IGHV6-1; IGKV1-5; IGKV1-6; IGKV1-9; IGKV1-12; IGKV1-16; IGKV1-17; IGKV1-27; IGKV1-39; IGKV2-30; IGKV3-11; IGKV3-15; IGKV3-20; IGLV1-40; IGLV1-47; IGLV1-51; IGLV2-11; IGLV2-23; IGLV2-14; IGLV3-1 и IGLV3-21.
Пары генов эмбрионального типа, имеющие предпочтительные биофизические свойства
На следующей стадии нужно было определить, какие пары белков эмбрионального типа нужно было исследовать, поскольку имеется примерно 2500 пар в иммунном репертуаре человека. Один способ заключался в тестировании пар белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые встречаются преимущественно в иммунном репертуаре человека, например, см. Таблицу 18. Например, можно выбрать верхние четыреста пар для тестирования или выбрать пары белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, экспрессирующиеся выше определенной пороговой концентрации. Поэтому аспекты настоящего изобретения включают способы получения коллекции синтетических антител или их функциональных фрагментов, где стадия получения дополнительно включает стадию идентификации пар генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, экспрессирующихся в концентрации по меньшей мере 0,05% в иммунном репертуаре человека; образования антител или их функциональных фрагментов, содержащих идентифицированные пары белков эмбрионального типа; и оценку следующих свойств указанной пары белков эмбрионального типа: 1) относительный уровень экспонирования в Fab-формате; 2) уровень экспрессии в Fab-формате; 3) термостабильность при 60°С или больше в Fab-формате в течение по меньшей мере 45 минут; 4) стабильность в сыворотке крупного рогатого скота в Fab-формате в течение более 10 суток при 37°С; 5) уровень экспрессии в IgG-формате; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
Этот подход требует синтеза и исследования большого числа последовательностей пар белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи; поэтому такой подход не может быть очень эффективным.
В качестве альтернативного подхода авторы изобретения выбрали субпопуляцию пар вариабельной области тяжелой цепи и вариабельной области легкой цепи эмбрионального типа, которая представляет, точно воспроизводит или охватывает большую часть преимущественно экспрессирующихся пар из иммунного репертуара человека. Этот подход был основан частично на наблюдении, что небольшое число генов эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи является доминантным в иммунном репертуаре человека. Wildt с соавт.на с.895-896 описывает это явление. Wildt с соавт.также сообщает, что обычно экспрессирующиеся генные сегменты тяжелой и легкой цепи часто спарены, и обнаружил, что половина тестируемых пар соответствует только пяти парам эмбрионального типа. Поэтому небольшое число преимущественно экспрессирующихся генов эмбрионального типа тяжелой и легкой цепи (неспаренных) можно комбинировать с получением группы пар, которая является типичной для иммунного репертуара человека.
Поэтому аспекты настоящего изобретения включают коллекции антител или их фрагментов, содержащие пары белков эмбрионального типа, которые представляют, точно воспроизводят или охватывают большую часть преимущественно экспрессирующихся пар генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи иммунного репертуара человека или иммунного репертуара интактного человека. Как описано ниже, подход авторов изобретения дает в результате коллекции, содержащие антитела или их фрагменты, которые могут в полной мере разрабатываться, поскольку пары белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи сначала тестируют в отношении предпочтительных биофизических свойств и затем конструируют коллекции для включения пар белков эмбрионального типа, имеющих одно или более из этих предпочтительных биофизических свойств.
Аспекты настоящего изобретения включают способы получения коллекции антител или их функциональных фрагментов, включающие стадию идентификации пар белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, имеющих одно или более из следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab pMx11_FH VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых аспектах настоящее изобретение включает способы получения коллекции антител или их функциональных фрагментов, включающие создание коллекции антител или их функциональных фрагментов, содержащей каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные один или более каркасных участков содержат белковые последовательности эмбрионального типа пар белков эмбрионального типа, где указанные пары белков эмбрионального типа имеют одно или более из следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С. В некоторых воплощениях участки FR1, FR2 и FR3 содержат белковые последовательности эмбрионального типа.
В некоторых воплощениях участки FR1, FR2 и FR3 содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа.
В некоторых воплощениях антитела или их функциональные фрагменты содержат одну или более определяющих комплементарность областей, содержащих белковые последовательности эмбрионального типа. В некоторых воплощениях антитела или их функциональные фрагменты содержат одну или более определяющих комплементарность областей, содержащих белковые последовательности эмбрионального типа пары белков эмбрионального типа. В некоторых воплощениях участки CDR1 и CDR2 содержат белковые последовательности эмбрионального типа. В некоторых воплощениях участки CDR1 и CDR2 содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа. В некоторых воплощениях участки FR1, CDR1, FR2, CDR2 и FR3 содержат белковые последовательности эмбрионального типа. В некоторых воплощениях участки FR1, CDR1, FR2, CDR2 и FR3 содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа. В некоторых воплощениях участок FR4 содержит участок JH4 тяжелой цепи. В некоторых воплощениях участок FR4 содержит участок Jk1 легкой цепи. В некоторых воплощениях участок FR4 содержит участок JA2/3 легкой цепи.
В других воплощениях настоящее изобретение включает способы получения коллекций антител или их функциональных фрагментов, включающие получение коллекции, где получение дополнительно включает стадии синтеза нуклеиновых кислот, кодирующих антитела или их функциональные фрагменты; клонирование нуклеиновых кислот в вектор; и экспрессирование антител или их функциональных фрагментов.
Как только преимущественно экспрессирующаяся или репрезентативная группа пар белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи была синтезирована и протестирована, можно было конструировать коллекции, включающие пары белков эмбрионального типа, имеющие предпочтительные биофизические свойства.
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, происходящих из пар белков эмбрионального типа, имеющих одно или более из следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов происходят из пар белков эмбрионального типа, имеющих такие свойства.
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, содержащие по существу белковые последовательности эмбрионального типа пар белков эмбрионального типа, имеющих одно или более из следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат по существу белковые последовательности эмбрионального типа пар белков эмбрионального типа, имеющих такие свойства.
Аспекты настоящего изобретения включают коллекции синтетических антител или их функциональных фрагментов, содержащие белковые последовательности эмбрионального типа пар белков эмбрионального типа, имеющих одно или более из следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С, где в некоторых воплощениях один или более каркасных участков и/или определяющих комплементарность областей антител или их функциональных фрагментов содержат белковые последовательности эмбрионального типа пар белков эмбрионального типа, имеющих такие свойства.
В некоторых аспектах коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пар белков эмбрионального типа, имеющих одно или более следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLAVI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях каркасные участки, содержащие белковые последовательности эмбрионального типа пары белков эмбрионального типа, содержат участки FR1, FR2 и FR3. В некоторых воплощениях антитела или их функциональные фрагменты содержат одну или более определяющих комплементарность областей, содержащих белковые последовательности эмбрионального типа. В некоторых воплощениях антитела или их функциональные фрагменты содержат одну или более определяющих комплементарность областей, содержащих белковые последовательности эмбрионального типа пары белков эмбрионального типа. В некоторых воплощениях участки CDR1 и CDR2 содержат белковые последовательности эмбрионального типа. В некоторых воплощениях участки CDR1 и CDR2 содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа. В некоторых воплощениях участки FR1, CDR1, FR2, CDR2 и FR3 содержат белковые последовательности эмбрионального типа. В некоторых воплощениях участки FR1, CDR1, FR2, CDR2 и FR3 содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа. В некоторых воплощениях участок FR4 содержит участок JH4 тяжелой цепи. В некоторых воплощениях участок FR4 содержит участок JK1 легкой цепи. В некоторых воплощениях участок FR4 содержит участок JA2/3 легкой цепи.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей по меньшей мере два из следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей по меньшей мере три из следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей по меньшей мере четыре из следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей по меньшей мере пять из следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей следующие свойства: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLAVI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки состоят по существу из белковых последовательностей эмбрионального типа пары белков эмбрионального типа, имеющей одно или более следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях антитела или их функциональные фрагменты состоят по существу из белковых последовательностей эмбрионального типа пары белков эмбрионального типа, имеющей одно или более следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLAVI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекции синтетических антител или их функциональных фрагментов содержат каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки состоят из белковых последовательностей эмбрионального типа пары белков эмбрионального типа, имеющей одно или более следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях антитела или их функциональные фрагменты состоят из белковых последовательностей эмбрионального типа пары белков эмбрионального типа, имеющей одно или более следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLAVI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях большинство или по существу все антитела или их функциональные фрагменты из коллекций содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей одно или более следующих свойств: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых аспектах антитела или их функциональные фрагменты из коллекций содержат белковые последовательности эмбрионального типа пар белков эмбрионального типа, имеющих одно или более из следующих свойств:
1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях антитела или их функциональные фрагменты состоят по существу из белковых последовательностей эмбрионального типа пары белков эмбрионального типа, имеющей следующие свойства: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях антитела или их функциональные фрагменты состоят из белковых последовательностей эмбрионального типа пары белков эмбрионального типа, имеющей следующие свойства: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекция синтетических антител или их функциональных фрагментов содержит каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки вариабельной области тяжелой цепи и каркасные участки вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, где указанная пара белков эмбрионального типа обладает следующими свойствами: 1) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; и 2) стабильность в сыворотке в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекция синтетических антител или их функциональных фрагментов содержит каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки вариабельной области тяжелой цепи и каркасные участки вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, где указанная пара белков эмбрионального типа обладает следующими свойствами: 1) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 2) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 3) стабильность в сыворотке в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекция синтетических антител или их функциональных фрагментов содержит каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки вариабельной области тяжелой цепи и каркасные участки вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, где указанная пара белков эмбрионального типа обладает следующими свойствами: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs; 2) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 3) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более, чем десяти суток при 37°С; и 4) стабильность в сыворотке в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях коллекция синтетических антител или их функциональных фрагментов содержит каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки вариабельной области тяжелой цепи и каркасные участки вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, где указанная пара белков эмбрионального типа обладает следующими свойствами: 1) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 2) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 3) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 4) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR003080; и 5) стабильность в сыворотке в IgG-формате в течение четырнадцати суток при 37°С.
В других воплощениях коллекции по настоящему изобретению содержат коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки вариабельной области тяжелой цепи и каркасные участки вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, где указанная пара белков эмбрионального типа обладает следующими свойствами: 1) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 2) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 3) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR003080; и 4) стабильность в сыворотке в IgG-формате в течение четырнадцати суток при 37°С.
В других воплощениях коллекции по настоящему изобретению и/или способы получения таких коллекций включают антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей относительный уровень экспонирования в Fab-формате по меньшей мере 0,1 по сравнению с контролем; по меньшей мере 0,2 по сравнению с контролем; по меньшей мере 0,3 по сравнению с контролем; по меньшей мере 0,4 по сравнению с контролем;
по меньшей мере 0,5 по сравнению с контролем; по меньшей мере 0,6 по сравнению с контролем; по меньшей мере 0,7 по сравнению с контролем; по меньшей мере 0,8 по сравнению с контролем; по меньшей мере 0,9 по сравнению с контролем; по меньшей мере 1,0 по сравнению с контролем; по меньшей мере 1,1 по сравнению с контролем; по меньшей мере 1,2 по сравнению с контролем; по меньшей мере 1,3 по сравнению с контролем; по меньшей мере 1,4 по сравнению с контролем; по меньшей мере 1,5 по сравнению с контролем; по меньшей мере 1,6 по сравнению с контролем; по меньшей мере 1,7 по сравнению с контролем; по меньшей мере 1,8 по сравнению с контролем; по меньшей мере 1,9 по сравнению с контролем; по меньшей мере 2,0 по сравнению с контролем; по меньшей мере 2,1 по сравнению с контролем; по меньшей мере 2,2 по сравнению с контролем; по меньшей мере 2,3 по сравнению с контролем; по меньшей мере 2,4 по сравнению с контролем; по меньшей мере 2,5 по сравнению с контролем; по меньшей мере 2,6 по сравнению с контролем; по меньшей мере 2,7 по сравнению с контролем; по меньшей мере 2,8 по сравнению с контролем; по меньшей мере 2,9 по сравнению с контролем; по меньшей мере 3,0 по сравнению с контролем; по меньшей мере 3,2 по сравнению с контролем; по меньшей мере 3,3 по сравнению с контролем; по меньшей мере 3,4 по сравнению с контролем; по меньшей мере 3,5 по сравнению с контролем; по меньшей мере 3,6 по сравнению с контролем; по меньшей мере 3,7 по сравнению с контролем; по меньшей мере 3,8 по сравнению с контролем; по меньшей мере 4,1 по сравнению с контролем; по меньшей мере 4,3 по сравнению с контролем; по меньшей мере 4,4 по сравнению с контролем; по меньшей мере 4,5 по сравнению с контролем; по меньшей мере 4,6 по сравнению с контролем; по меньшей мере 4,7 по сравнению с контролем; по меньшей мере 5,0 по сравнению с контролем; по меньшей мере 5,1 по сравнению с контролем; по меньшей мере 5,2 по сравнению с контролем; по меньшей мере 5,4 по сравнению с контролем; по меньшей мере 5,5 по сравнению с контролем; по меньшей мере 5,7 по сравнению с контролем; по меньшей мере 5,9 по сравнению с контролем; по меньшей мере 6,0 по сравнению с контролем; по меньшей мере 6,1 по сравнению с контролем; по меньшей мере 6,3 по сравнению с контролем; по меньшей мере 6,4 по сравнению с контролем; по меньшей мере 6,7 по сравнению с контролем; по меньшей мере 6,9 по сравнению с контролем; по меньшей мере 7,0 по сравнению с контролем; по меньшей мере 7,1 по сравнению с контролем; по меньшей мере 7,2 по сравнению с контролем; по меньшей мере 7,3 по сравнению с контролем; по меньшей мере 7,4 по сравнению с контролем; по меньшей мере 8,1 по сравнению с контролем; по меньшей мере 8,2 по сравнению с контролем; по меньшей мере 8,3 по сравнению с контролем; по меньшей мере 8,4 по сравнению с контролем; по меньшей мере 8,5 по сравнению с контролем; по меньшей мере 8,6 по сравнению с контролем; по меньшей мере 8,7 по сравнению с контролем; по меньшей мере 8,8 по сравнению с контролем; по меньшей мере 8,9 по сравнению с контролем; по меньшей мере 9,1 по сравнению с контролем; по меньшей мере 9,2 по сравнению с контролем; по меньшей мере 9,3 по сравнению с контролем; по меньшей мере 9,4 по сравнению с контролем; по меньшей мере 9,5 по сравнению с контролем; по меньшей мере 9,7 по сравнению с контролем; по меньшей мере 9,8 по сравнению с контролем; по меньшей мере 10,0 по сравнению с контролем; по меньшей мере 10,2 по сравнению с контролем; по меньшей мере 10,3 по сравнению с контролем; по меньшей мере 10,5 по сравнению с контролем; по меньшей мере 10,6 по сравнению с контролем; по меньшей мере 10,7 по сравнению с контролем; по меньшей мере 10,8 по сравнению с контролем; по меньшей мере 11,0 по сравнению с контролем; по меньшей мере 11,2 по сравнению с контролем; по меньшей мере 11,3 по сравнению с контролем; по меньшей мере 11,5 по сравнению с контролем; по меньшей мере 11,7 по сравнению с контролем; по меньшей мере 11,8 по сравнению с контролем; по меньшей мере 12,1 по сравнению с контролем; по меньшей мере 12,3 по сравнению с контролем; по меньшей мере 12,4 по сравнению с контролем; по меньшей мере 12,9 по сравнению с контролем; по меньшей мере 13,0 по сравнению с контролем; по меньшей мере 13,6 по сравнению с контролем; по меньшей мере 14,4 по сравнению с контролем; по меньшей мере 14,5 по сравнению с контролем; по меньшей мере 16,1 по сравнению с контролем; по меньшей мере 16,6 по сравнению с контролем; по меньшей мере 16,7 по сравнению с контролем; по меньшей мере 17,1 по сравнению с контролем; по меньшей мере 19,4 по сравнению с контролем; по меньшей мере 27,3 по сравнению с контролем; или по меньшей мере 29,0 по сравнению с контролем.
В некоторых воплощениях коллекция синтетических антител или их функциональных фрагментов содержит каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки вариабельной области тяжелой цепи и каркасные участки вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, где указанная пара белков эмбрионального типа имеет относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 10% выбранных Fabs;
значение не ниже 15% выбранных Fabs; значение не ниже 20% выбранных Fabs; значение не ниже 25% выбранных Fabs; значение не ниже 30% выбранных Fabs; значение не ниже 35% выбранных Fabs; значение не ниже 40% выбранных Fabs; значение не ниже 45% выбранных Fabs; значение не ниже 50% выбранных Fabs; значение не ниже 55% выбранных Fabs; значение не ниже 60% выбранных Fabs; значение не ниже 65% выбранных Fabs;
значение не ниже 70% выбранных Fabs; значение не ниже 75% выбранных Fabs; значение не ниже 80% выбранных Fabs; значение не ниже 85% выбранных Fabs; или значение не ниже 90% выбранных Fabs.
В других воплощениях коллекции по настоящему изобретению и/или способы получения таких коллекций включают антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей относительный уровень экспрессии в Fab-формате по меньшей мере 0,1 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; по меньшей мере 0,2 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; по меньшей мере 0,3 по сравнению с Fab VH1-69 VLA__VI1-40 AYA; по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; по меньшей мере 0,5 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; по меньшей мере 0,6 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; по меньшей мере 0,7 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; по меньшей мере 0,8 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; по меньшей мере 0,9 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; по меньшей мере 1,0 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; no меньшей мере 1,1 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; по меньшей мере 1,2 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; или по меньшей мере 1,3 по сравнению с Fab VH1-69 VLA.VI1-40 AYA.
В других воплощениях коллекции по настоящему изобретению и/или способы получения таких коллекций включают антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей термостабильность при 70°С или более в течение по меньшей мере 45 минут в Fab-формате; или имеющей термостабильность при 80°С или более в течение по меньшей мере 45 минут в Fab-формате.
В других воплощениях коллекции по настоящему изобретению и/или способы получения таких коллекций включают антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, имеющей относительный уровень экспрессии в IgG-формате по меньшей мере 0,1 по сравнению с MOR03080; по меньшей мере 0,2 по сравнению с MOR03080; по меньшей мере 0,3 по сравнению с MOR03080; по меньшей мере 0,4 по сравнению с MOR03080; по меньшей мере 0,5 по сравнению с MOR03080; по меньшей мере 0,6 по сравнению с MOR03080; по меньшей мере 0,7 по сравнению с MOR03080; по меньшей мере 0,8 по сравнению с MOR03080; по меньшей мере 0,9 по сравнению с MOR03080; по меньшей мере 1,0 по сравнению с MOR03080; по меньшей мере 1,1 по сравнению с MOR03080; по меньшей мере 1,2 по сравнению с MOR03080; по меньшей мере 1,3 по сравнению с MOR03080; по меньшей мере 1,4 по сравнению с MOR03080no меньшей мере 1,5 по сравнению с MOR03080; по меньшей мере 1,6 по сравнению с MOR03080; по меньшей мере 1,7 по сравнению с MOR03080; по меньшей мере 1,8 по сравнению с MOR03080; по меньшей мере 1,9 по сравнению с MOR03080.
В некоторых аспектах настоящее изобретение включает коллекции и способы получения или применения коллекций антител или их функциональных фрагментов, содержащих одну или более определяющих комплементарность областей, содержащих белковые последовательности эмбрионального типа, по существу последовательности эмбрионального типа или последовательности, происходящие из белковых последовательностей эмбрионального типа. В некоторых воплощениях антитела или их функциональные фрагменты содержат CDR1 и CDR2, содержащие белковые последовательности эмбрионального типа. В некоторых воплощениях антитела или их функциональные фрагменты содержат CDR1 и CDR2, содержащие белковые последовательности эмбрионального типа пары белков эмбрионального типа.
В некоторых аспектах один или более каркасных участков содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа. Как в некоторых аспектах, FR4 выбрана из группы, состоящей из JH4, Jк1 и JA2/3. Как показано на Фиг.45А-47 В, белковые последовательности эмбрионального типа содержат только FR1-FR3. Поэтому в некоторых аспектах, когда указанные каркасные участки вариабельной области тяжелой цепи и каркасные участки вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа, FR1, FR2 и/или FR3 содержат белковые последовательности эмбрионального типа. В некоторых аспектах один или более каркасных участков содержат белковые последовательности эмбрионального типа, что способствует разнообразию одной или более областей, определяющих комплементарность. В некоторых воплощениях настоящее изобретение включает коллекции и способы продуцирования и получения указанных коллекций синтетических антител или их функциональных фрагментов, содержащих диверсифицированный участок HCDR3. В некоторых воплощениях настоящее изобретение включает коллекции и способы получения и применения указанных коллекций синтетических антител или их функциональных фрагментов, содержащих диверсифицированный участок LCDR3.
Дополнительный аспект настоящего изобретения состоит в возможности использовать коллекции в идентификации антител или их функциональных фрагментов против любого иммуногена. Предположили, что создание коллекций по меньшей мере с двумя парами белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, обладающими вышеупомянутыми функциональными свойствами, будет обеспечивать разнообразие в пределах коллекции, особенно в пределах определяющих комплементарность областей антител из коллекции, в отношении длины CDR и разнообразия конформаций или канонических структур. Это позволяет использовать коллекции по настоящему изобретению в идентификации антител или их функциональных фрагментов против любого иммуногена.
Некоторые воплощения изобретения включают коллекции, содержащие антитела или их функциональные фрагменты, содержащие каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковую последовательность эмбрионального типа пары генов эмбрионального типа, имеющую следующие свойства: 1) относительный уровень экспонирования в Fab, представляющий собой значение не ниже 75% выбранных Fabs; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С, где указанная коллекция содержит антитела или их функциональные фрагменты, содержащие по меньшей мере две различные пары белков эмбрионального типа; по меньшей мере три различные пары белков эмбрионального типа; по меньшей мере четыре различные пары белков эмбрионального типа; по меньшей мере пять различных пар белков эмбрионального типа; по меньшей мере шесть различных пар белков эмбрионального типа; по меньшей мере семь различных пар белков эмбрионального типа; по меньшей мере восемь различных пар белков эмбрионального типа; по меньшей мере девять различных пар белков эмбрионального типа; по меньшей мере десять различных пар белков эмбрионального типа; по меньшей мере одиннадцать различных пар белков эмбрионального типа; по меньшей мере двенадцать различных пар белков эмбрионального типа; по меньшей мере тринадцать различных пар белков эмбрионального типа; по меньшей мере четырнадцать различных пар белков эмбрионального типа; по меньшей мере пятнадцать различных пар белков эмбрионального типа; по меньшей мере шестнадцать различных пар белков эмбрионального типа; по меньшей мере семнадцать различных пар белков эмбрионального типа; по меньшей мере восемнадцать различных пар белков эмбрионального типа; по меньшей мере девятнадцать различных пар белков эмбрионального типа; по меньшей мере двадцать различных пар белков эмбрионального типа; по меньшей мере 21 различную пару белков эмбрионального типа; по меньшей мере 22 различные пары белков эмбрионального типа; по меньшей мере 23 различные пары белков эмбрионального типа; по меньшей мере 24 различные пары белков эмбрионального типа; по меньшей мере 25 различных пар белков эмбрионального типа; по меньшей мере 26 различных пар белков эмбрионального типа; по меньшей мере 27 различных пар белков эмбрионального типа; по меньшей мере 28 различных белков эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 29 различных последовательностей пар белков эмбрионального типа; по меньшей мере 30 различных пар белков эмбрионального типа; по меньшей мере 31 различную пару белков эмбрионального типа; по меньшей мере 32 различные пары белков эмбрионального типа по меньшей мере 33 различные пары белков эмбрионального типа по меньшей мере 34 различные пары белков эмбрионального типа по меньшей мере 35 различных пар белков эмбрионального типа по меньшей мере 36 различных пар белков эмбрионального типа по меньшей мере 37 различных пар белков эмбрионального типа по меньшей мере 38 различных пар белков эмбрионального типа по меньшей мере 39 различных пар белков эмбрионального типа по меньшей мере 40 различных пар белков эмбрионального типа; по меньшей мере 41 различную пару белков эмбрионального типа; по меньшей мере 42 различные пары белков эмбрионального типа; по меньшей мере 43 различные пары белков эмбрионального типа; по меньшей мере 44 различных белка эмбрионального типа вариабельной области тяжелой цепи; по меньшей мере 45 различных последовательностей пар белков эмбрионального типа; по меньшей мере 46 различных пар белков эмбрионального типа; по меньшей мере 47 различных пар белков эмбрионального типа; по меньшей мере 48 различных пар белков эмбрионального типа; по меньшей мере 49 различных пар белков эмбрионального типа или по меньшей мере 50 различных пар белков эмбрионального типа.
Антитела или их функциональные фрагменты, содержащие генные последовательности эмбрионального типа
Кроме того, предположили, что использование белковых последовательностей эмбрионального типа должно снизить риск иммуногенности антител при введении пациентам. Поэтому аспекты настоящего изобретения включают коллекции и способы получения и применения указанных коллекций синтетических антител или их функциональных фрагментов, содержащих каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки содержат белковые последовательности эмбрионального типа. В некоторых воплощениях каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи антител или их функциональных фрагментов содержат по существу последовательности эмбрионального типа. В некоторых воплощениях каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи антител или их функциональных фрагментов происходят из последовательностей эмбрионального типа. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат участки FR1, FR2, FR3 и FR4, содержащие белковые последовательности эмбрионального типа, по существу последовательности эмбрионального типа или происходят из белковых последовательностей эмбрионального типа. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат FR1, FR2, FR3, содержащие белковые последовательности эмбрионального типа репрезентативной пары белков эмбрионального типа. В некоторых воплощениях используемая участок FR4 представляет собой JH4 для вариабельной области тяжелой цепи, Jk1 для вариабельной области к- легкой цепи и JA2/3 для вариабельной области λ-легкой цепи.
Также, поскольку использование белковых последовательностей эмбрионального типа должно снижать риск иммуногенности антител при введении пациентам, некоторые аспекты настоящего изобретения включают коллекции и способы получения или применения коллекций антител или их функциональных фрагментов, содержащих одну или более определяющих комплементарность областей, содержащих белковые последовательности эмбрионального типа, по существу последовательности эмбрионального типа или происходящие из белковых последовательностей эмбрионального типа. В некоторых воплощениях антитела или их функциональные фрагменты содержат CDR1 и CDR2, содержащие белковые последовательности эмбрионального типа. В некоторых воплощениях антитела или их функциональные фрагменты содержат CDR1 и CDR2, содержащие белковые последовательности эмбрионального типа пары белков эмбрионального типа.
В некоторых аспектах один или более каркасных участков содержат белковые последовательности эмбрионального типа, обеспечивающие разнообразие одной или более областей, определяющих комплементарность. В некоторых воплощениях настоящее изобретение включает коллекции и способы продуцирования и получения указанных коллекций синтетических антител или их функциональных фрагментов, содержащих диверсифицированный участок HCDR3. В некоторых воплощениях настоящее изобретение включает коллекции и способы получения и применения указанных коллекций синтетических антител или их функциональных фрагментов, содержащих диверсифицированный участок LCDR3. CDRs можно конструировать с помощью методов, хорошо известных в данной области, включая описанные в Knappik et al. 2000; WO 97/08320; WO 2008053275; WO 2009036379 WO 2007056441; WO 2009114815, которые все включены посредством ссылки во всей своей полноте.
Кроме того, для получения коллекций, содержащих антитела или их функциональные фрагменты, имеющие низкий риск иммуногенности, в некоторых аспектах коллекция по настоящему изобретению и способы ее получения и использования включают антитела или их функциональные фрагменты, содержащие человеческие последовательности.
В некоторых аспектах коллекция по изобретению содержит по меньшей мере 1×104; по меньшей мере 1×105; по меньшей мере 1×106; по меньшей мере 1×107; по меньшей мере 1×108; по меньшей мере 1×109; по меньшей мере 1×1010; или по меньшей мере 1×1011 последовательностей нуклеиновой кислоты, кодирующих антитела или их функциональные фрагменты, или антител или их функциональных фрагментов.
В некоторых аспектах антитела или их функциональные фрагменты из коллекций являются синтетическими.
В некоторых аспектах коллекции содержат нуклеиновые кислоты, кодирующие антитела или их функциональные фрагменты.
Дополнительные воплощения настоящего изобретения
В некоторых аспектах настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи, где указанные каркасные участки вариабельной области тяжелой цепи и каркасные участки вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа,
где указанная пара белков эмбрионального типа обладает следующими свойствами:
1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs;
2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA;
3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате;
4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С;
5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и
6) стабильность в сыворотке в IgG-формате в течение четырнадцати суток при 37°С;
где указанная коллекция антител или их функциональных фрагментов содержит белковые последовательности эмбрионального типа по меньшей мере двух различных пар белков эмбрионального типа, и
где указанная пара белков эмбрионального типа кодируется парой генов эмбрионального типа.
В некоторых воплощениях настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, где указанные каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи состоят по существу из белковых последовательностей эмбрионального типа пар белков эмбрионального типа, имеющих следующие свойства:
1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs;
2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA;
3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате;
4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С;
5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и
6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, где указанные каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи состоят из белковых последовательностей эмбрионального типа пар белков эмбрионального типа, имеющих следующие свойства:
1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs;
2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA;
3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате;
4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С;
5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и
6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в иммунном репертуаре человека в концентрации по меньшей мере 0,05%. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,23% в иммунном репертуаре человека. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,51% в иммунном репертуаре человека. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,07% в иммунном репертуаре интактного человека. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,52% в иммунном репертуаре интактного человека. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,88% в иммунном репертуаре интактного человека. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат человеческие последовательности. В некоторых воплощениях указанная коллекция антител или их функциональных фрагментов содержит белковые последовательности эмбрионального типа по меньшей мере семнадцати различных пар белков эмбрионального типа.
В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат одну или более определяющих комплементарность областей, содержащих белковые последовательности эмбрионального типа. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат участки FR1, CDR1, FR2, CDR2 и FR3, содержащие белковые последовательности эмбрионального типа. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат участок FR4, выбранный из группы, состоящей из JH4, Jк1 и JA2/3. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат диверсифицированный участок HCDR3. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат диверсифицированный участок LCDR3.
В некоторых воплощениях коллекция содержит 1×104 антител или их функциональных фрагментов. В некоторых воплощениях указанные пары белков эмбрионального типа имеют относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 60% выбранных Fabs. В некоторых воплощениях указанные пары белков эмбрионального типа имеют уровень экспрессии в Fab-формате по меньшей мере 0,6 по сравнению с Fab VH1-69 VLA_VI1-40 AYA. В некоторых воплощениях указанные пары белков эмбрионального типа имеют термостабильность при 70°С или более в течение по меньшей мере 45 минут в Fab-формате. В некоторых воплощениях указанные пары белков эмбрионального типа имеют уровень экспрессии в IgG-формате по меньшей мере 0,6 по сравнению с MOR03080.
В некоторых воплощениях каркасные участки вариабельной области тяжелой и вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа выбранной из группы, состоящей из: IGHV3-23/IGKV1-5; IGHV3-23/IGKV3-20; IGHV4-39/IGKV3-15; IGHV3-23/IGKV3-15; IGHV4-39/IGKV1-39/1D-39; IGHV1-18/IGKV3-20; IGHV3-30/IGKV3-20; IGHV4-39/IGKV1-5; IGHV1-69/IGKV1-39/1D-39; IGHV5-51/IGLV1-40; IGHV4-39/IGKV3-20; IGHV3-23/IGLV2-14; IGHV4-39/IGLV3-21; IGHV3-23/IGKV1-39/1D-39; IGHV3-30/IGKV1-39/1D-39; IGHV1-69/IGKV3-20; IGHV3-48/IGKV3-20; IGHV1-2/IGKV3-20; IGHV3-30/IGKV4-1; IGHV5-51/IGLV2-14; IGHV5-51/IGKV3-20; IGHV3-7/IGKV1-39/1D-39; IGHV3-7/IGKV1-5; IGHV3-15/IGKV3-20; IGHV4-39/IGLV2-14; IGHV3-23/IGKV3-11; IGHV3-30/IGKV1-5; IGHV3-30/IGKV3-15; IGHV3-21/IGKV1-5; IGHV3-21/IGKV3-15; IGHV3-30/IGLV1-51; IGHV3-21/IGLV1-51 и IGHV1-69/IGKV3-11.
В некоторых воплощениях указанные функциональные фрагменты указанных антител выбраны из группы, состоящей из Fab, F(ab')2, Fab', Fv и scFv.
В некоторых аспектах настоящее изобретение включает коллекцию нуклеиновых кислот, кодирующих описанные коллекции антител. В некоторых аспектах настоящее изобретение включает вектор, содержащий нуклеиновые кислоты, кодирующие описанные коллекции антител. В некоторых аспектах настоящее изобретение включает рекомбинантную клетку-хозяина, содержащую нуклеиновые кислоты, кодирующие описанные коллекции антител. В некоторых воплощениях рекомбинантная клетка-хозяин является прокариотической или эукариотической. В некоторых воплощениях рекомбинантная клетка-хозяин представляет собой клетку Е.соli или млекопитающего.
В некоторых аспектах настоящее изобретение включает коллекцию синтетических антител или их функциональных фрагментов, содержащую каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи,
где указанные каркасные участки содержат белковые последовательности эмбрионального типа,
где указанные белковые последовательности эмбрионального типа имеют следующие свойства:
1) четыре или меньше посттрансляционных модификаций в участках, определяющих комплементарность;
2) два или меньше метионинов в участках, определяющих комплементарность;
3) один или меньше неспаренных цистеинов;
4) один или меньше потенциальных Т-клеточных эпитопов;
5) среднюю или низкую склонность к агрегации; и
6) изоэлектрическую точку по меньшей мере 7,5; и
где указанная коллекция антител или их функциональных фрагментов содержит по меньшей мере две различные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи,
где указанная белковая последовательность эмбрионального типа кодируется генной последовательностью эмбрионального типа.
В некоторых воплощениях указанные генные последовательности эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи присутствует в концентрации по меньшей мере 0,5% в иммунном репертуаре человека. В некоторых воплощениях указанная коллекция антител или их функциональных фрагментов содержит по меньшей мере пять различных белковых последовательностей эмбрионального типа вариабельной области тяжелой цепи. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат человеческие последовательности. В некоторых воплощениях указанные генные последовательности эмбрионального типа вариабельной области тяжелой цепи или вариабельной области легкой цепи присутствует в концентрации по меньшей мере 5,0% в иммунном репертуаре человека. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат одну или более определяющих комплементарность областей, содержащих белковые последовательности эмбрионального типа. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат FR1, CDR1, FR2, CDR2 и FR3, содержащие белковые последовательности эмбрионального типа. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат участок FR4, выбранный из группы, состоящей из JH4, Jк1 и JA2/3. В некоторых воплощениях указанные антитела или их функциональные фрагменты также содержат диверсифицированный участок HCDR3. В некоторых воплощениях указанные антитела или их функциональные фрагменты также содержат диверсифицированный участок LCDR3. В некоторых воплощениях коллекция содержит 1×104 антител или их функциональных фрагментов.
В некоторых воплощениях указанные белковые последовательности эмбрионального типа вариабельной области тяжелой цепи выбраны из группы, состоящей из: IGHV3-23; IGHV3-30; IGHV4-39; IGHV4-34; IGHV4-59; IGHV1-69; IGHV5-51; IGHV3-7; IGHV1-18; IGHV3-48; IGHV3-15; IGHV3-21; IGHV1-2; IGHV3-33; IGHV4-31; IGHV3-53; IGHV3-11; IGHV3-9; IGHV4-4; IGHV1-46; IGHV3-74; IGHV1-24; IGHV4-61; IGHV1-8; IGHV1-3; IGHV3-49; IGHV3-43; IGHV4-28; IGHV3-64nlGHV7-81.
В некоторых воплощениях белковые последовательности эмбрионального типа вариабельной области к-легкой цепи выбраны из группы, состоящей из: IGKV3-20; IGKV1-39/1D-39; IGKV1-5; IGKV3-15; IGKV4-1; IGKV3-11; IGKV2-28/2D-28; IGKV1-33/1D-33; IGKV2-30; IGKV1-9; IGKV1-17; IGKV1-27; IGKV1-8; IGKV1-16; IGKV1-6; IGKV1-12; IGKV2D-29; IGKV1-13; IGKV1D-8 и IGKV2-24.
В некоторых воплощениях белковые последовательности эмбрионального типа вариабельной области λ-легкой цепи выбраны из группы, состоящей из: IGLV2-14; IGLV1-40; IGLV1-44; IGLV1-51; IGLV2-23; IGLV3-21; IGLV1-47; IGLV3-1; IGLV2-11; IGLV2-8; IGLV6-57; IGLV3-25; IGLV7-46; IGLV1-36; IGLV7-43; IGLV9-49; IGLV4-69; IGLV2-18; IGLV3-10 и IGLV3-27.
В некоторых аспектах настоящее изобретение включает способ получения описанных коллекций синтетических антител или их функциональных фрагментов. В некоторых воплощениях стадии получения дополнительно включают создание коллекции антител или их функциональных фрагментов, содержащих каркасные участки вариабельной области тяжелой цепи и вариабельной области легкой цепи,
где указанные каркасные участки вариабельной области тяжелой цепи и каркасные участки вариабельной области легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа,
где указанная пара белков эмбрионального типа обладает следующими свойствами:
1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs;
2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA;
3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате;
4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С;
5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и
6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С;
где указанная коллекция антител или их функциональных фрагментов содержит по меньшей мере две различные пары белков эмбрионального типа.
В некоторых воплощениях стадия получения дополнительно включает стадии:
а) получения данных о парах генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, присутствующих в иммунном репертуаре человека;
б) идентификации пар белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, обладающих следующими свойствами:
1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fabs;
2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab pMx11_FH VH1-69 VLA_VI1-40 AYA;
3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате;
4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С;
5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR3080; и
6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С; и
в) создание коллекции антител или их функциональных фрагментов, содержащих белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи пар белков эмбрионального типа, идентифицированных на стадии (б).
В некоторых воплощениях стадия (б) дополнительно включает стадии:
(ба) идентификации пар генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, присутствующих в концентрации по меньшей мере 0,05% в иммунном репертуаре человека;
бб) создания антител или их функциональных фрагментов, содержащих пары белков эмбрионального типа, идентифицированные на стадии (ба); и
бв) оценки следующих свойств указанной пары белков эмбрионального типа:
1) относительный уровень экспонирования в Fab-формате;
2) уровень экспрессии в Fab-формате;
3) термостабильность при 60°С или более в Fab-формате;
4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С;
5) уровень экспрессии в IgG-формате; и
6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°С.
В некоторых воплощениях стадия (а) дополнительно включает следующие стадии:
аа) выделение человеческих В-клеток из образца;
аб) получение кДНК из В-клеток;
ав) ПЦР-амплификация кДНК из В-клеток;
аг) секвенирование ПЦР-продуктов;
ад) идентификация генов эмбрионального типа из каждого ПЦР-продукта. В некоторых воплощениях стадия создания коллекции дополнительно включает следующие стадии;.
ва) синтез нуклеиновых кислот, кодирующих антитела или их функциональные фрагменты;
вб) клонирование нуклеиновых кислот в вектор;
вв) экспрессирование антител или их функциональных фрагментов. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,05% в иммунном репертуаре человека. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,23% в иммунном репертуаре человека. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,51% в иммунном репертуаре человека. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,07% в иммунном репертуаре интактного человека. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,52% в иммунном репертуаре интактного человека. В некоторых воплощениях указанные пары генов эмбрионального типа присутствуют в концентрации по меньшей мере 0,88% в иммунном репертуаре интактного человека. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат человеческие последовательности. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат белковые последовательности эмбрионального типа по меньшей мере семнадцати различных пар белков эмбрионального типа. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат одну или более определяющих комплементарность областей, содержащих белковые последовательности эмбрионального типа.
В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат FR1, CDR1, FR2, CDR2 и FR3, содержащие белковые последовательности эмбрионального типа. В некоторых воплощениях указанные антитела или их функциональные фрагменты содержат участок FR4, выбранный из группы, состоящей из JH4, Jк1 и JA2/3. В некоторых воплощениях антитела или их функциональные фрагменты содержат диверсифицированный участок HCDR3. В некоторых воплощениях антитела или их функциональные фрагменты содержат диверсифицированный участок LCDR3. В некоторых воплощениях коллекция содержит 1×104 антител или их функциональных фрагментов.
В некоторых воплощениях указанные пары белков эмбрионального типа имеют относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 60% выбранных Fabs. В некоторых воплощениях указанные пары белков эмбрионального типа имеют уровень экспрессии в Fab-формате по меньшей мере 0,6 по сравнению с Fab VH1-69 VLA_VI1-40 AYA. В некоторых воплощениях указанные пары белков эмбрионального типа имеют термостабильность при 70°С или более в течение по меньшей мере 45 минут в Fab-формате. В некоторых воплощениях указанные пары белков эмбрионального типа имеют уровень экспрессии в IgG-формате по меньшей мере 0,6 по сравнению с MOR03080.
В некоторых воплощениях каркасные участки вариабельной области тяжелой цепи и легкой цепи содержат белковые последовательности эмбрионального типа пары белков эмбрионального типа, выбранной из группы, состоящей из: IGHV3-23/IGKV1-5; IGHV3-23/IGKV3-20; IGHV4-39/IGKV3-15; IGHV3-23/IGKV3-15; IGHV4-59/IGKV1-39/1D-39; IGHV4-39/IGKV1-39/1D-39; IGHV4-59/IGKV3-20; IGHV1-18/IGKV3-20; IGHV3-30/IGKV3-20; IGHV4-39/IGKV1-5; IGHV1-69/IGKV1-39/1D-39; IGHV5-51/IGLV1-40; IGHV3-23/IGKV4-1; IGHV4-39/IGKV3-20; IGHV3-23/IGLV2-14; IGHV4-39/IGLV3-21; IGHV3-23/IGKV1-39/1D-39; IGHV3-30/IGKV1-39/1D-39; IGHV3-30/IGKV3-11; IGHV1-69/IGKV3-20; IGHV3-48/IGKV3-20; IGHV1-2/IGKV3-20; IGHV3-30/IGKV4-1; IGHV5-51/IGLV2-14; IGHV4-59/IGKV4-1; IGHV5-51/IGKV3-20; IGHV3-7/IGKV1-39/1D-39; IGHV3-7/IGKV1-5; IGHV3-15/IGKV3-20; IGHV4-39/IGLV2-14; IGHV4-39/IGLV2-8; IGHV3-23/IGKV3-11; IGHV3-30/IGKV1-5; IGHV3-30/IGKV3-15; IGHV3-21/IGKV1-5; IGHV3-21/IGKV3-15; IGHV3-30/IGLV1-51; IGHV3-21/IGLV1-51; IGHV3-53/IGLV1-44; IGHV4-59/IGKV3-15; IGHV5-51/IGKV4-1; IGHV1-69/IGKV4-1 и IGHV1-69/IGKV3-11.
В некоторых аспектах указанные функциональные фрагменты указанных антител выбраны из группы, состоящей из Fab, F(ab')2, Fab', Fv и scFv.
В некоторых аспектах настоящее изобретение включает выделенную нуклеиновую кислоту, кодирующую сигнальную или лидерную последовательность, содержащую С-концевой сайт рестрикции. В некоторых воплощениях сайт рестрикции представляет собой Nhel. В некоторых воплощениях сигнальная или лидерная последовательность содержит рhоА или лидерную последовательность тяжелой цепи человека. В некоторых воплощениях сайт рестрикции представляет собой Ndel. В некоторых воплощениях сигнальная последовательность содержит оmрА или лидерную последовательность каппа-цепи человека.
В некоторых аспектах настоящее изобретение включает вектор, содержащий нуклеиновые кислоты, кодирующие сигнальную или лидерную последовательность, содержащую С-концевой сайт рестрикции. В некоторых аспектах настоящее изобретение включает клетку-хозяина, содержащую вектор. В некоторых воплощениях клетка-хозяин является прокариотической или эукариотической. В некоторых воплощениях клетка-хозяин представляет собой клетку E.соli. В некоторых воплощениях клетка-хозяин представляет собой клетку млекопитающего.
В некоторых аспектах настоящее изобретение включает выделенное антитело или его функциональный фрагмент, содержащие FR1, CDR1, FR2, CDR2 и FR3, содержащие белковые последовательности эмбрионального типа пары белков эмбрионального типа,
где указанная пара белков эмбрионального типа выбрана из группы, состоящей из: IGHV3-23/IGKV1-5; IGHV3-23/IGKV3-20; IGHV4-39/IGKV3-15; IGHV3-23/IGKV3-15; IGHV4-39/IGKV1-39/1D-39; IGHV1-18/IGKV3-20; IGHV3-30/IGKV3-20; IGHV4-39/IGKV1-5; IGHV1-69/IGKV1-39/1D-39; IGHV5-51/IGLV1-40; IGHV4-39/IGKV3-20; IGHV3-23/IGLV2-14; IGHV4-39/IGLV3-21; IGHV3-23/IGKV1-39/1 D-39; IGHV3-30/IGKV1-39/1D-39; IGHV1-69/IGKV3-20; IGHV3-48/IGKV3-20; IGHV1-2/IGKV3-20; IGHV3-30/IGKV4-1; IGHV5-51/IGLV2-14; IGHV5-51/IGKV3-20; IGHV3-7/IGKV1-39/1 D-39; IGHV3-7/IGKV1-5; IGHV3-15/IGKV3-20; IGHV4-39/IGLV2-14; IGHV3-23/IGKV3-11; IGHV3-30/IGKV1-5; IGHV3-30/IGKV3-15; IGHV3-21/IGKV1-5; IGHV3-21/IGKV3-15; IGHV3-30/IGLV1-51; IGHV3-21/IGLV1-51 и IGHV1-69/I.
В одном аспекте в настоящем описании описаны коллекции антител, содержащие каркасные участки вариабельной области тяжелой и легкой цепи, содержащие последовательности эмбрионального типа, конкретно FR1.
Ожидается, что имеющиеся каркасные участки эмбрионального типа снижают риск иммуногенности антител при введении пациентам. Сайты рестрикции, однако, должны использоваться для того, чтобы сделать возможным стандартное клонирование нуклеиновых кислот, кодирующих коллекции антител, в дисплейные и/или экспрессирующие векторы, с тем чтоы антитела моли быть подвергнуты скринингу против иммуногенов. Раньше сайты рестрикции, используемые для клонирования, часто располагали в пределах каркасных участков, таким образом модифицируя последовательность нуклеиновой кислоты от эмбрионального типа. Для того чтобы гарантировать, что по меньшей мере каркасный участок 1 (FR1) каждого из антител по настоящему изобретению сохраняет последовательность эмбрионального типа, в пределах FR1 не должно быть никаких неприродных сайтов рестрикции. Поэтому аспект настоящего изобретения относится к включению сайта рестрикции в пределах С-конца прокариотических сигнальных последовательностей и человеческой лидерной последовательности, конкретно, в пределах трех С-концевых остатков. Кроме того, сигнальная последовательность и лидерная последовательность, содержащие сайт рестрикции, должны быть функциональными и обеспечивать хорошие уровни экспонирования и экспрессии антител или их фрагментов как в прокариотических системах экспрессии, так и в системах экспрессии у млекопитающих.
В некоторых аспектах настоящее изобретение включает выделенную нуклеиновую кислоту, кодирующую сигнальную или лидерную последовательность, содержащую С-концевой сайт рестрикции. В некоторых воплощениях сайт рестрикции представляет собой Nhel или Ndel. В некоторых воплощениях сигнальная или лидерная последовательность содержит phoA или лидерную последовательность тяжелой цепи человека. В некоторых воплощениях сигнальная или лидерная последовательность содержит оmрА или лидерную последовательность каппа-цепи человека. В некоторых аспектах настоящее изобретение включает вектор, содержащий выделенную нуклеиновую кислоту, кодирующую сигнальную или лидерную последовательность, содержащую С-концевой сайт рестрикции. В некоторых аспектах настоящее изобретение включает клетку-хозяина, содержащую выделенную нуклеиновую кислоту, кодирующую сигнальную или лидерную последовательность, содержащую С-концевой сайт рестрикции, или вектор, содержащий выделенную нуклеиновую кислоту, кодирующую сигнальную или лидерную последовательность, содержащую С-концевой сайт рестрикции. В некоторых воплощениях клетка-хозяин является прокариотической, например E.соli, или эукариотической, например, клеткой млекопитающего.
Настоящее описание является первым описанием, в котором раскрыта концепция, что пары класса VH и VL, которые являются наиболее распространенными в иммунном репертуаре интактного человека, по-видимому, имеют предпочтительные характеристики, такие как большая стабильность и более низкая иммуногенность. Настоящее описание также является первым описанием, которое включает эту концепцию в конструирование коллекции и использует полный синтез гена для получения таких коллекций. Настоящее описание обеспечивает способы идентификации пар класса VH и VL в "наивных" и "обученных" антигеном иммунных репертуарах человека, определения пар класса VH и VL, которые являются наиболее распространенными, и затем создания коллекций, содержащих эти пары класса VH и VL. Более конкретно, коллекции по настоящему изобретению содержат наиболее распространенные и/или предпочтительные пары класса VH и VL с высоко диверсифицироваными CDRs. Эта стратегия увеличивает вероятность того, что коллекции содержат антитела или их фрагменты против любого иммуногена, которые являются стабильными, имеют низкую иммуногенность и высокую аффинность в отношении специфического антигена. Результат состоит в резко повышенной вероятности, что коллекции содержат высокоэффективные антитела или их фрагменты против любого иммуногена, которые можно использовать для терапевтических или диагностических целей.
Соответственно, последовательности нуклеиновых кислот (или их выбранная часть), кодирующие антитела или их фрагменты, полученные из "наивных" В-клеток их хозяев-людей, можно секвенировать. Из этих данных о последовательностях можно идентифицировать спаривания цепей класса VH/VL семейства эмбрионального типа, представленные в иммунном репертуаре. На основании определенных критериев, таких как распространенность и/или предпочтительные биофизические свойства, выбирают пары классов тяжелых и легких цепей для включения в коллекции. Коллекции затем можно синтезировать посредством генного синтеза. В некоторых воплощениях синтетические коллекции содержат по существу каркасные участки VH и VL эмбрионального типа, где CDRs диверсифицированы, или только один CDR из VH и/или VL диверсифицирован.
С использованием последовательностей ДНК, полученных из "наивных" В-клеток хозяев-людей в качестве "матрицы", настоящее описание предоставляет способы идентификации наиболее распространенных пар VH и VL. После того, как была выявлена относительная распространенность спариваний класса VH и VL, может быть получена высокоразнообразная коллекция антител или их фрагментов, содержащих наиболее распространенные и/или предпочтительные спаривания класса VH и VL. Используя эту информацию, специалист может создать высокую степень разнообразия без потери ключевых преимуществ, присущих наиболее распространенным и/или предпочтительным комбинациям пар класса VH и VL. До настоящего изобретения никто не выявлял наиболее распространенные и/или предпочтительные пары класса VH и VL или не пытался использовать эти знания в методах создания библиотеки. Поэтому такой подход обеспечивает исчерпывающие коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, которые представляют иммунную систему интактного человека.
Используя способы конструирования и экспонирования коллекции, описанные в данной заявке, можно создавать коллекции с огромным разнообразием, так как некоторые воплощения включают коллекции по меньшей мере1×1010 членов.
В некоторых аспектах настоящее изобретение включает векторы и клетки-хозяева, содержащие описанные коллекции нуклеиновых кислот.
В некоторых аспектах настоящее изобретение включает способы получения таких коллекций.
В некоторых воплощениях "наивные" последовательности ДНК, типичные для иммунного репертуара человека, получают на отдельной стадии и хранят в базе данных; поэтому конструкцию коллекции можно легко модифицировать, оптимизировать и адаптировать к требованиям заказчика in silico (с помощью компьютерного моделирования), учитывая уровень соответствия требованиям заказчика, который обычно может быть реализован в синтетической библиотеке.
В некоторых аспектах настоящее изобретение включает способы идентификации антител или их фрагментов с использованием описанных коллекций.
В некоторых аспектах настоящее изобретение относится к коллекциям или библиотекам, кодирующим антитела или их фрагменты, содержащим белковые последовательности эмбрионального типа, кодируемые семействами и/или генами VH и VL эмбрионального типа, которые являются наиболее распространенными и/или предпочтительными в иммунном репертуаре. В некоторых воплощениях нуклеиновые кислоты, кодирующие антитела или их фрагменты, являются эмбриональными, по существу эмбриональными или их кодон-оптимизированными вариантами. Такие коллекции или библиотеки могут содержать семейства и/или гены VH и VL эмбрионального типа, имеющие предпочтительные биофизические свойства, включая высокое экспонирование на фаге; высокую экспрессию в Е.соli в Fab-формате; высокую экспрессию в клетках млекопитающих в IgG-формате; высокую термостабильность;
стабильность в сыворотке; низкую тенденцию к агрегации (т.е. высокую растворимость); и низкий риск иммуногенности. В некоторых воплощениях коллекции или библиотеки могут содержать семейства и/или гены VH и VL эмбрионального типа, существующие в иммунном репертуаре интактного человека. Родственные воплощения включают способы получения и применения таких коллекций.
В некоторых аспектах настоящее изобретение относится к коллекциям или библиотекам, кодирующим антитела или их фрагменты, содержащие белковые последовательности эмбрионального типа, кодируемые семействами и/или генами VH и VL эмбрионального типа, наиболее распространенными и/или предпочтительными в иммунном репертуаре наряду с парами класса VH/VL, наиболее распространенными и/или предпочтительными в иммунном репертуаре. В некоторых воплощениях нуклеиновые кислоты, кодирующие антитела или их фрагменты, являются эмбриональными, по существу эмбриональными или их кодон-оптимизированными вариантами. Такие коллекции или библиотеки могут содержать белковые последовательности эмбрионального типа, кодируемые семействами и/или генами VH и VL эмбрионального типа и/или парами класса VH/VL, имеющими предпочтительные биофизические свойства, включая высокое экспонирование на фаге; высокую экспрессию в Е.соli в Fab-формате; высокую экспрессию в клетках млекопитающих в IgG-формате; высокую термостабильность; стабильность в сыворотке; низкую тенденцию к агрегации (т.е. высокую растворимость); и низкий риск иммуногенности. В некоторых воплощениях коллекции или библиотеки могут содержать белковые последовательности эмбрионального типа, кодируемые семействами и/или генами VH и VL эмбрионального типа и/или парами класса VH/VL, которые существуют в иммунном репертуаре интактного человека. Родственные воплощения включают способы получения и применения таких коллекций.
Соответственно, настоящее изобретение включает коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, по существу типичные для иммунного репертуара, где каждое антитело или его фрагмент содержит пару класса VH/VL, где пара класса VH/VL, по существу типичная для иммунного репертуара, является такой, что каждая пара класса VH/VL, присутствующая в коллекции, представляет собой пару класса VH/VL, присутствующую в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре. Иммунный репертуар может принадлежать индивидууму или популяции, и может быть "наивным". Такой иммунный репертуар можно определить как иммунный репертуар из пар класса VH/VL по меньшей мере в 1×105 В-клеток от индивидуума; пар класса VH/VL по меньшей мере в 1×105 В-клеток из популяции индивидуумов; или пар класса VH/VL, присутствующих по меньшей мере в 1×105 антител, например. Иммунный репертуар может представлять собой иммунный репертуар "наивных" В-клеток или "обученных" антигеном В-клеток. Индивидуум или популяция может представлять собой человека или быть человеческой. Иммунный репертуар можно определить путем анализа общедоступной базы данных и/или литературы.
В некоторых воплощениях нуклеиновые кислоты, кодирующие антитела или их фрагменты, являются синтетическими, такими как полученные посредством полного синтеза генов. В родственных воплощениях нуклеиновые кислоты представляют собой последовательности эмбрионального типа; по существу последовательности эмбрионального типа; или кодон-оптимизированные варианты эмбрионального типа или по существу последовательности эмбрионального типа. В некоторых воплощениях по меньшей мере один из CDRs является высоко диверсифицированным.
В некоторых воплощениях коллекция по настоящему изобретению содержит антитела или их фрагменты, где FR1, FR2 и FR3 и из VH, и из VL содержат белковые последовательности эмбрионального типа пар класса VH и VL, имеющих предпочтительные характеристики. Наиболее предпочтительно, коллекция по настоящему изобретению содержит антитела или их фрагменты, где FR1, FR2 и FR3 и из VH, и из VL содержат белковые последовательности эмбрионального типа пар класса VH и VL, имеющих предпочтительные характеристики, где CDR3 и из VH, и из VL являются высоко диверсифицированными.
В родственных воплощениях коллекцию указанных нуклеиновых кислот, кодирующих антитела или их фрагменты, клонируют в вектор. Подходящие векторы известны в данной области техники и включают дисплейные векторы, такие как фаговые дисплейные векторы, плазмидные векторы, фагмидные векторы, экспрессирующие векторы, включая бактериальный экспрессирующий вектор или экспрессирующий вектор млекопитающего. В другом родственном воплощении коллекцию или коллекцию, клонированную в векторы, трансформируют в клетки-хозяева. Таким образом, изобретение включает коллекцию клеток-хозяев. Подходящие клетки-хозяева включают прокариотические клетки-хозяева (такие как Е.соli) и эукариотические клетки-хозяева (такие как клетки-хозяева млекопитающих).
В другом воплощении изобретение представляет собой базу данных, содержащую пары класса VH/VL из примерно 1345 "наивных" человеческих В-клеток или из общедоступных последовательностей на читаемом носителе. Такая база данных полезна для разработки и конструирования коллекции и библиотек по изобретению.
Изобретение также включает способы получения коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, по существу типичные для иммунного репертуара. Иммунный репертуар может представлять собой иммунный репертуар "наивных" В-клеток или "обученных" антигеном В-клеток человека или людей. Такой способ может включать следующие стадии: (а) идентификацию пар класса VH/VL, присутствующих в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре; (б) синтез коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH/VL, присутствующие в концентрации по меньшей мере 0,05%, по меньшей мере1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре.
Стадию идентификации можно осуществлять различными путями. Например, идентификация пар класса VH/VL может включать выделение "наивных" В-клеток от одного или более хозяев-людей и определение пар класса VH/VL в каждой В-клетке путем выделения и секвенирования ДНК, мРНК или кДНК, кодирующих пары класса VH/VL, или путем зондирования с помощью одного или более нуклеиновокислотных зондов, специфичных к каждому VH и VL, и затем анализа пар класса VH/VL. В альтернативном или дополнительном воплощении пары класса VH/VL могут быть определены из уже существующих баз данных, таких как базы данных по последовательностям антител. В альтернативном или дополнительном воплощении пары класса VH/VL могут быть идентифицированы из литературы. Таким образом, в одном воплощении изобретение включает получение нуклеиновокислотных последовательностей антител (либо уже существующих, либо полученных de novo), определение пар класса VH/VL посредством выравнивания последовательностей и сопоставление таких последовательностей для идентификации пар класса VH/VL, присутствующих в иммунном репертуаре.
В некоторых воплощениях способ используют для создания коллекций, в которых большинство членов имеет предпочтительные биофизические свойства, которые облегчают продукцию и экспрессию антител или их фрагментов (например, на фаге или из клеток) и дают растворимые и термостабильные антитела. Более конкретно, такие свойства включают: (1) эффективное экспонирование на фаге; (2) эффективное экспонирование на клетках млекопитающих (3) значительную экспрессию в Е.соli в Fab-формате;
(4) значительную экспрессию в клетках млекопитающих в IgG-формате; (5) термостабильность; (6) растворимость; и (7) низкую иммуногенность. Посредством определения пар класса VH/VL, имеющих некоторые или все эти свойства, можно затем конструировать коллекцию, в которой большинство членов имеет такие биофизические свойства, например, посредством синтеза только нуклеиновых кислот с такими свойствами. Соответственно, изобретение включает такую коллекцию и способы получения такой коллекции.
В некоторых воплощениях синтезированные нуклеиновые кислоты являются эмбриональными, по существу эмбриональными или их кодон- оптимизированными вариантами. Изменение можно вводить по меньшей мере в один участок, определяющий комплементарность (CDR). Любой CDR является подходящим, особенно CDR3. Предпочтительно, изменение последовательности, добавленное в CDR, ограничивается последовательностями в рамке считывания и свободно от цистеинов и стоп-кодонов, обеспечивая таким образом правильную экспрессию всех членов библиотеки.
После того, как нуклеиновые кислоты были синтезированы, их можно клонировать в вектор (такой как дисплейный вектор, фаговый дисплейный вектор; фагмидовый вектор; или экспрессирующий вектор млекопитающих) и можно трансформировать в клетку-хозяина. Подходящие клетки-хозяева включают прокариотические клетки-хозяева (например, E.coli) и эукариотические клетки-хозяева (например, клетки-хозяева млекопитающих).
В других воплощених в изобретении предложены способы идентификации антител, специфических в отношении иммуногена. Такие способы могут включать в одном воплощении идентификацию пар класса VH/VL, присутствующих в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре;
синтез коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH/VL, присутствующие в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре; экспонирование или экспрессирование антитела или его фрагмента из коллекции; скрининг коллекции против специфического иммуногена; и выбор по меньшей мере одного антитела или его фрагмента, специфического в отношении указанного иммуногена. Поскольку способы и коллекции по изобретению можно конструировать с учетом предпочтительных биофизических свойств, настоящее изобретение особенно полезно для идентификации антитела или фрагмента антитела для лечения заболевания или состояния посредством создания коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты с такими предпочтительными свойствами, и скрининга против специфического иммуногена для идентификации антител, связывающихся с таким иммуногеном.
В некоторых аспектах изобретение относится к коллекциям или библиотекам антител или их фрагментов, содержащих пары VH/VL. В некоторых воплощениях коллекции или библиотеки антител или их фрагментов содержат белковые последовательности эмбрионального типа, кодируемые семействами и/или генами VH и VL эмбрионального типа, которые распространены в иммунном репертуаре. В некоторых воплощениях изобретение относится к коллекциям или библиотекам антител или их фрагментам, содержащим белковые последовательности эмбрионального типа, кодируемые семействами и/или генами VH и VL эмбрионального типа, имеющими определенные предпочтительные биофизические характеристики. В некоторых воплощениях семейства и/или гены VH и VL эмбрионального типа являются такими, которые встречаются в природе в иммунном репертуаре и находятся среди наиболее многочисленных и распространенных в репертуаре. В некоторых воплощениях коллекции или библиотеки антител или их фрагментов содержат белковые последовательности эмбрионального типа, кодируемые семействами и/или генами VH и VL эмбрионального типа. В некоторых воплощениях коллекции или библиотеки антител или их фрагментов содержат каркасные участки и/или участки CDR из эмбриональных, по существу эмбриональных или кодон-оптимизированных семейств и/или генов VH и VL. В некоторых воплощениях коллекции или библиотеки антител или их фрагментов содержат белковые последовательности эмбрионального типа, кодируемые семействами и/или генами VH и VL эмбрионального типа, которые являются синтетическими, сконструированными посредством тотального генного синтеза. В некоторых воплощениях коллекции или библиотеки антител или их фрагментов содержат участки семейств и/или генов VH и VL эмбрионального типа, которые являются синтетическими, сконструированными посредством тотального генного синтеза. В некоторых воплощениях коллекции или библиотеки антител или их фрагментов содержат белковые последовательности эмбрионального типа, кодируемые семействами и/или генами VH и VL эмбрионального типа, имеющими предпочтительные биофизические свойства, которые облегчают скрининг и дальнейшую разработку антител, особенно в терапевтическом контексте. Предпочтительные биофизические свойства включают, но не ограничиваются этим, то, что: (1) они хорошо экспонируются на фаге в Fab-формате, (2) они хорошо экспонируются на клетках млекопитающих в IgG-формате (3) они экспрессируются в высоких количествах в Fab-формате, например, в Е.coli, и IgG-форматах, например, в клетках млекопитающих, (4) являются термодинамически стабильными; (5) имеют высокую стабильность в сыворотке, (6) имеют низкую тенденцию к агрегации (т.е. высокую растворимость); и (7) имеют низкий риск иммуногенности.
В других аспектах коллекции по настоящему изобретению содержат антитела или их фрагменты, содержащие белковые последовательности эмбрионального типа предпочтительных пар класса VH и VL. Коллекции по настоящему изобретению предпочтительно содержат антитела или их фрагменты, где один или более каркасных участков содержат белковые последовательности эмбрионального типа, кодируемые парами класса VH и VL, имеющими предпочтительные характеристики, особенно где FR1, FR2 и FR3 и из VH, и из VL содержат белковые последовательности эмбрионального типа пар класса VH и VL, имеющих предпочтительные характеристики. CDRs могут быть высоко диверсифицированы. Предпочтительно, CDR3 и из VH, и из VL являются высоко диверсифицированными. В некоторых воплощениях CDR1 и CDR2 из VH и/или VL являются эмбриональными или по существу эмбриональными по последовательности.
Эта стратегия увеличивает вероятность того, что коллекции по настоящему изобретению содержат антитела или их фрагменты против любого иммуногена, которые могут разрабатываться для терапевтического применения, поскольку большинство антител или их фрагментов, присутствующих в коллекциях, содержат последовательности эмбрионального типа пар VH и VL, имеющих упомянутые выше предпочтительные характеристики. Выбранные антитела также будут иметь низкую иммуногенность и высокую аффинность к специфическому антигену. Результат заключается в существенно увеличенной вероятности того, что антитела или их фрагменты, которые селектированы из описанных коллекций, являются высоко эффективными против любого иммуногена и могут разрабатываться для терапевтических или диагностических целей.
Такие коллекции преодолевают многие проблемы из предшествующего уровня техники. Например, в когнатной библиотеке, происходящей из В-кпеток, спаривания класса VH и VL, присутствующие в библиотеке, зависят от спариваний класса, присутствующих в образце. Если взять достаточно большой образец В-клеток, то будет присутствовать каждая из комбинаций спариваний класса приблизительно 50 VH и 50 VL (примерно 2500). Присутствие такого количества пар класса VH и VL может быть сравнимо с фоновым шумом. Может быть желательным создание библиотек с большим разнообразием, содержащих только самые распространенные пары класса VH и VL, но, учитывая когнатный характер библиотеки, это не возможно.
В дополнение к этому, в некоторых воплощениях последовательности ДНК, на которых основаны коллекции, получены их образцов "наивных" В-клеток, которые "не обучены" антигеном, поэтому экспрессирующиеся члены не смещены в сторону конкретного иммуногена, и данные коллекции могут быть использованы для скрининга против любого иммуногена.
Соответственно, последовательности нуклеиновых кислот (или их выбранный участок), кодирующие антитела или их фрагменты, полученные из "наивных" ("необученных" антигеном) В-клеток от хозяев-людей, можно секвенировать. Из этих данных о последовательностях можно идентифицировать спаривания класса VH/VL цепей семейства эмбрионального типа, преимущественно представленные в иммунном репертуаре. На основании определенных критериев, таких как распространенность, можно выбрать пары класса тяжелых и легких цепей для включения в коллекции. Затем коллекции можно синтезировать посредством генного синтеза. В некоторых воплощениях синтетические коллекции содержат по существу каркасные участки VH и VL эмбрионального типа, где CDRs диверсифицированы.
С использованием последовательностей ДНК, полученных, например, из "наивных" ("необученных" антигеном) В-клеток от хозяев-людей или из общедоступных баз данных или литературы в качестве "матрицы", в настоящем описании предложены способы идентификации наиболее распространенных семейств и/или генов и/или пар класса VH и VL эмбрионального типа. Как только была выявлена относительная распространенность семейств и/или генов и/или пар класса VH и VL эмбрионального типа, антитела или их фрагменты, содержащие белковые последовательности эмбрионального типа, кодируемые семействами и/или генами и/или парами класса VH и VL эмбрионального типа, можно протестировать в отношении следующих предпочтительных характеристик: (1) они хорошо экспонируются на фаге в Fab- формате, (2) они экспрессируются в высоких количествах и в растворенной форме в Fab-формате и IgG-форматах, (3) и они термодинамически стабильны. Посредством тестирования белковых последовательностей эмбрионального типа, кодируемых наиболее распространенными семействами и/или генами и/или парами класса VH и VL эмбрионального типа, можно идентифицировать те из них, которые имеют предпочтительные характеристики. Используя эту информацию, специалист может создать высокую степень разнообразия без ухудшения ключевых преимуществ, относящихся к наиболее распространенным комбинациям семейств и/или генов и/или пар класса VH и VL эмбрионального типа.
Используя способы конструирования и экспонирования коллекции, описанные в данной заявке, можно создать коллекции с большим разнообразием, поскольку некоторые воплощения включают коллекции из по меньшей мере 1×1010 членов.
Настоящее описание относится в основном к коллекциям синтетических антител, содержащим пары класса VH и VL, имеющие наиболее предпочтительные характеристики. В некоторых воплощениях коллекции содержат белковые последовательности эмбрионального типа, кодируемые семействами VH и VL, представленными парой данного класса.
Настоящее описание относится в основном к коллекциям синтетических антител, содержащих одну или более пар класса VH и VL, имеющим предпочтительные характеристики. В некоторых аспектах коллекции содержат белковые последовательности эмбрионального типа, кодируемые семействами VH и VL, представленными парой класса.
В некоторых аспектах в настоящем описании предложены способы идентификации генов VH и VL эмбрионального типа, которые являются наиболее распространенными в иммунном репертуаре, тестирования антител, имеющих последовательности наиболее распространенных генов VH и VL эмбрионального типа для идентификации генов VH и VL эмбрионального типа, имеющих предпочтительные характеристики, и затем создания коллекций, содержащих предпочтительные классы VH и VL. В настоящем описании предложены способы идентификации пар класса VH и VL в иммунном репертуаре человека, который может быть интактным, определения пар класса VH и VL, которые являются наиболее распространенными, тестирования пар класса VH и VL для идентификации пар класса VH и VL, имеющих предпочтительные характеристики, и затем создания коллекций, содержащих предпочтительные пары класса VH и VL и/или антитела, происходящие из предпочтительных генов VH и VL эмбрионального типа. Как только VH и VL и/или пары класса VH и VL идентифицированы, определяют их соответствующие последовательности эмбрионального типа, так что последовательности эмбрионального типа могут быть включены в конструируемую коллекцию.
В некоторых аспектах в настоящем описании предложены векторы и клетки-хозяева, содержащие описанные коллекции нуклеиновых кислот.
В некоторых аспектах в настоящем описании предложены способы получения таких коллекций.
В некоторых воплощениях последовательности ДНК, представляющие иммунный репертуар человека, получают на отдельной стадии и хранят в базе данных; поэтому конструкцию коллекции можно легко модифицировать, оптимизировать и адаптировать к требованиям заказчика in silica (с помощью компьютерного моделирования), учитывая уровень соответствия требованиям заказчика, который обычно может быть реализован в синтетической библиотеке.
В некоторых аспектах в настоящем описании предложены способы идентификации антител или их фрагментов с использованием описанных коллекций.
Способы, нуклеиновые кислоты, белки, вектор, клетка-хозяин
В одном аспекте в настоящем описании предложены коллекции нуклеиновых кислот, полученных посредством тотального синтеза генов. Технология генного синтеза значительно продвинулась в последние годы и можно создавать очень большие коллекции нуклеиновых кислот.Следующие компании предлагают услуги по такому синтезу: Entelechon (Regensburg, Германия), Geneart (Regensburg, Германия) и Sloning Biotechnology (Puchheim, Германия). Для создания коллекции компанией генного синтеза могут быть представлены последовательности каждого члена коллекции.
В некоторых воплощениях в настоящем описании предложены коллекцию синтетических нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащую пары класса VH и VL, присутствующие в концентрации по меньшей мере 0,05% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток. В другом воплощении коллекции по настоящему изобретению содержат пары класса VH и VL, присутствующие в концентрации по меньшей мере 1% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток. В другом воплощении коллекции по настоящему изобретению содержат пары класса VH и VL, присутствующие в концентрации по меньшей мере 1,5% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток. В другом воплощении коллекции по настоящему изобретению содержат пары класса VH и VL, присутствующие в концентрации по меньшей мере 2% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток. В другом воплощении коллекции по настоящему изобретению содержат пары класса VH и VL, присутствующие в концентрации по меньшей мере 3% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток. В другом воплощении коллекции по настоящему изобретению содержат пары класса VH и VL, присутствующие в концентрации по меньшей мере 4% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток. В другом воплощении коллекции по настоящему изобретению содержат пары класса VH и VL, присутствующие в концентрации по меньшей мере 5% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток.
В некоторых воплощениях в настоящем описании предложены коллекции, где пары класса VH и VL идентифицированы в В-клетках, выделенных из человеческого организма. В некоторых воплощениях В-клетки являются "наивными". В некоторых воплощениях коллекции нуклеиновых кислот кодируют антитела или их фрагменты, содержащие каркасные участки VH и VL эмбрионального типа. В предпочтительном воплощении коллекции нуклеиновых кислот синтезируют с включением каркасных участков VH и VL эмбрионального типа с диверсифицироваными CDRs. Каркасные участки эмбрионального типа желательны, поскольку антитела или их фрагменты, содержащие каркасные участки эмбрионального типа, вероятно, не являются иммуногенными.
Используя способы конструирования и экспонирования коллекции, описанные в данной заявке, можно получать коллекции с большим разнообразием, поскольку некоторые воплощения включают коллекции из по меньшей мере 1×104 последовательностей нуклеиновых кислот, некоторые воплощения включают коллекции из по меньшей мере 1×105, 106, 107, 108, 109, 1010, 1011 или 1012 последовательностей нуклеиновых кислот.Такое разнообразие получают посредством синтеза коллекций, содержащих члены, содержащие распространенные пары класса VH и VL с диверсифицироваными CDRs.
Коллекции по настоящему изобретению сконструированы на основании данных о последовательностях, по существу представляющих иммунный репертуар. В некоторых воплощениях данные о последовательностях получают посредством поиска общедоступных перечней последовательностей иммуноглобулинов. Например, можно осуществлять поиск в NCBI, используя Ig-Blast, или в общедоступной литературе. Начиная с 2005 г., база данных содержала по меньшей мере 25000 перегруппированных последовательностей человеческих антител в FASTA формате. Из 22500 элементов, 13235 представляли последовательности VH, 1506 представляли Vк, и 2259 представляли Vλ. Последовательности VH, Vк и Vλ можно классифицировать в соответствующие семейства и/или гены эмбрионального типа. Поскольку некоторые из Ig-Blast включают последовательности целых антител, из базы данных по последовательностям можно определить точные семейства и/или гены эмбрионального типа для каждого из спариваний доменов класса VH и VL. При использовании этого подхода, значение каждого семейства и/или гена VH и VL эмбрионального типа, и/или семейства и/или гена каждой пары доменов класса VH и VL эмбрионального типа может быть легко определено специалистом в данной области. Выбор, какой из VH и VL и/или какую из пар класса VH и VL включить в библиотеку, можно осуществлять различными способами. В некоторых воплощениях для включения в коллекцию или библиотеку выбирают VH и VL с наибольшей распространенностью. В некоторых воплощениях VH и VL, имеющие предпочтительные биофизические свойства, выбирают для включения в коллекцию или библиотеку. В некоторых воплощениях пары класса VH и VL, имеющие наибольшую распространенность, выбирают для включения в коллекцию или библиотеку. В некоторых воплощениях пары класса VH и VL, имеющие предпочтительные биофизические свойства, выбирают для включения в коллекцию или библиотеку. В некоторых воплощениях и VH, и VL, имеющие наибольшую распространенность и/или предпочтительные биофизические свойства, и/или пары класса VH и VL, имеющие наибольшую распространенность, и/или пары класса VH и VL, имеющие предпочтительные биофизические свойства, выбирают для включения в коллекцию или библиотеку.
Одним из. недостатков такого подхода является то, что общедоступные базы данных часто наполнены последовательностями антител, полученных против специфических иммуногенов, поэтому последовательности имеют отклонения. Кроме того, в большинстве баз данных последовательности тяжелой и легкой цепи не связаны, поэтому спаривание класса VH и VL не может быть идентифицировано.
В некоторых воплощениях последовательности нуклеиновых кислот получены путем сбора В-клеток от одного или более хозяев; выделения ДНК из В-клеток и предпочтительно секвенирования ДНК. Предпочтительно, В-клетки являются "наивными". Образцы из В-клеток собирают от одного или более людей-доноров. Ниже приведен метод, который может быть использован для выделения В-клеток. В-лимфоциты в состоянии покоя (В-клетки) выделяют из селезенки, используя отрицательный отбор против других типов клеток с помощью моноклональных антител анти-СD43 и анти-Мас-1/CDHb, например, с помощью магнитных микрогранул. Эта стратегия истощает не В-клетки из смешанной популяции спленоцитов и основана на том факте, что большинство зрелых лейкоцитов, за исключением покоящихся В-клеток селезенки, экспрессируют CD43 (в действительности, экспрессия CD43 была продемонстрирована на незрелых В-клетках, плазматических клетках и некоторых зрелых клетках, в дополнение к гранулоцитам, моноцитам, макрофагам, тромбоцитам, клеткам естественным киллерам (NK), тимоцитам и периферическим CD8+и большинству CD4+Т-клеток). Анти-Мас-1/CD11b микрогранулы включены в отрицательную селекцию для улучшения удаления миелоидных клеток. Выделение В-клеток можно автоматизировать с использованием AutoMACS автоматизированного сортировщика клеток с помощью магнитных шариков (Miltenyi Biotec). Как оценено с помощью флуоресцентного анализа В220+клеток, такое выделение обычно дает приблизительно 4×10е7 В-клеток на селезенку с чистотой более 95%. См. также Miltenyi S, Muller W, Weichel W, and Radbruch A. (1990) Cytometry 11(2), 231-238.
Число выделенных В-клеток по существу представляет иммунный репертуар. В некоторых воплощениях по меньшей мере 1х104 В-клеток выделяют из хозяина, более предпочтительно по меньшей мере 105 В-клеток;
более предпочтительно по меньшей мере 106 В-клеток; наиболее предпочтительно 107 В-клеток выделяют из хозяина.
ДНК, кодирующую антитела и их фрагменты из каждой В-клетки, выделяют и амплифицируют, например, тяжелые и легкие цепи связаны посредством ПЦР реакции. ДНК предпочтительно секвенируют.Секвенированная ДНК может представлять собой кДНК, полученную из мРНК В-клеток. Экстракция мРНК из эукариотических клеток, таких как В-клетки, является хорошо известной технологической процедурой. Существуют многочисленные протоколы и имеются коммерческие наборы. Такие как система выделения мРНК PolyATtract® (Promega, Madison, WI, США) или различные наборы RNeasy и Oligotex DirectmRNA (оба из Qiagen, Hilden, Германия). Во многих этих методах используют polyA-хвост эукариотических мРНК, например, посредством аффинной очистки на олиго (dT) матрицах, таких как олиго (dT) целлюлоза.
кДНК можно селективно амплифицировать из выделенной мРНК с помощью обратной транскрипции с использованием специфических праймеров, затем стандартной ПЦР. Специфические праймеры используют для амплификации нуклеиновых кислот вариабельных доменов тяжелых и легких цепей. См. Cancer Surv. 1997; 30:21-44, J Clin. Pathol. 1994; 47:493-6, J. Clin. Pathol. 1990; 43:888-90 или Mol. Pathol. 2002 April; 55(2): 98-101.
ДНК, кодирующую вариабельные домены тяжелой и легкой цепи из одной В-клетки, хранят вместе, так что можно идентифицировать спаривание вариабельных доменов класса тяжелых и легких цепей. Методы выделения нуклеиновых кислот, кодирующих спаривание вариабельных доменов из индивидуальной В-клетки, хорошо известны в данной области. См., например, WO 01/92291; WO 92/15678; WO 93/03151, WO 2005/042774; Mullinax RL et al„ 1992 Biotechniques 12:6 864-868; Chapal, N. etal. 1997 Biotechniques 23, 518-524, Embleton MJ et al., 1992 Nucleic Acids Res. 20:15, 3831-3837; Coronella, J.A. et al. 2000 Nucleic Acids Res. 28:20, E85; Thirion S et al., 1996 European Journal of Cancer Prevention 5:6 507-511; и Wang, X et al. 2000 J. Immunol. Methods 20, 217-225.
Эти методы можно использовать отдельно или в комбинации с другими методами. Например, если последовательности вариабельных доменов тяжелой и легкой цепи из большого образца не идентифицированы успешно вместе с их соответствующими В-клетками, то можно выполнить следующий способ с целью идентификации точного домена пары класса вариабельной области тяжелой и вариабельной области легкой цепи. Выполняют ПЦР на одной клетке (single cell PCR) для каждой отдельной В-клетки.
Предпочтительно, ДНК из каждой В-клетки секвенируют.Существуют различные компании, которые способны секвенировать полные геномы, такие как Helicos BioSciences Corporation (Cambridge, MA, США). С помощью своей технологии True Single Molecule Sequencing™, Helicos способна осуществлять прямое секвенирование одиночных молекул ДНК или РНК с высокой скоростью и эффективностью. Другие компании, способные выполнять аналогичную работу по секвенированию, включают Illumina (San Diego, CA, США; система Solexa) и Roche (Basel, CH; система 454). Никаких стадий клонирования не требуется перед секвенированием.
В другом аспекте в описании предложены способы идентификации семейства эмбрионального типа пар вариабельных доменов тяжелой и легкой цепи, присутствующих в иммунном репертуаре. Все антитела или их фрагменты можно проследить до их семейства эмбрионального типа с использованием способов, известных специалисту в данной области. Анализируя последовательность нуклеиновой кислоты, кодирующей антитело или его фрагмент, можно определить семейство эмбрионального типа как для VH, так и для VL с помощью способов, известных специалисту в данной области. Например, Wildt et. al, (1999) собирали В-клетки от 3 пациентов и идентифицировали 365 спариваний класса VH и VL. РНК из каждой В-клетки использовали для синтеза кДНК, и кДНК, кодирующую VH- и VL-области, амплифицировали с помощью ПЦР и секвенировали. Как показано на Фиг.1 Wildt, некоторые классы VH и VL спаривались чаще, чем другие, например, VH3-8 сVк3-1, Vк3-19, Vк4-1, Vλ2-3 или Vλ1-2, и VH3-9 с Vк3-1, Vк3-3 или Vλ1-5.
В другом аспекте в описании предложены способы конструирования диверсифицированных участков, определяющих комплементарность, перед синтезированием коллекции. CDRs можно конструировать способами, хорошо известными в данной области, включая описанные в Knappik et al. 2000;
В другом аспекте в описании предложены способы селекции спариваний класса вариабельных доменов, которые желательно включить в коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты. В некоторых воплощениях синтезируют коллекцию нуклеиновых кислот, содержащих все пары класса VH и VL доменов, идентифицированные описанными способами.
Кроме того, распространенность пар класса VH и VL можно определить с помощью различных статистических тестов. В самой простой форме просто подсчитывают отдельные пары класса VH и VL. Более сложные статистические тесты могут учитывать различные другие параметры. В качестве неограничивающих примеров следующие статистические тесты и ссылки могут служить руководством в качестве примеров многочисленных подходов, сделанных в таком или подобном анализе: Байесовская оценка сжатия (Bayesian Shrinkage Estimation) (см., например, Biometrics 59 (2003): 476-486), DADA (Цифровой анализ распространенности кДНК, см., например, ВМС Genomics 2002, 3:7), линейное моделирование (Pacific Symposium on Biocomputing, 1999, 4:41-52) и различные методы кластеризации (ВМС ВМС Bioinformatics 2006, 7:397, Fourth IEEE International Conference on Data Mining (ICDM'04), pp.403-406).
В других аспектах в настоящем описании предложена коллекция векторов, содержащих коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты. В некоторых воплощениях векторы содержат экспрессирующие векторы, дисплейные векторы, фаговые дисплейные векторы или фагмидные векторы.
Эукариотические экспрессирующие векторы хорошо известны в данной области, а также имеются в продаже. Как правило, предложенные векторы содержат удобные сайты рестрикции для вставки желательной ДНК. Примеры таких векторов включают pSVL и pKSV-10, pBPV-1/PML2d и pTDT1 (ATCC, №31255).
В других аспектах в настоящем описании предложена коллекция клеток-хозяев, трансформированных описанной коллекцией векторов. Клетки-хозяева могут быть либо прокариотическими, либо эукариотическими. Бактериальные клетки представляют собой предпочтительные прокариотические клетки-хозяева и обычно представляют собой штамм Escherichia соli (Е.coli), такой как, например, E.coli штамм DH5, доступный от Bethesda Research Laboratories, Inc., Bethesda, Md. Предпочтительные эукариотические клетки-хозяева включает клетки дрожжей и млекопитающих, в том числе мышей и грызунов, предпочтительно клетки позвоночных, такие как клетки, полученные из клеточной линии мыши, крысы, обезьяны или человека.
Введение векторов в клетки-хозяева можно осуществить различными способами трансформации или трансфекции, известными специалистам в данной области, включая преципитацию фосфатом кальция, электропорацию, микроинъекцию, слияние с липосомами, слияние с помощью "теней" эритроцитов (RBC ghost fusion), слияние протопластов, вирусную инфекцию и тому подобное. Получение моноклональных полноразмерных антител, Fab-фрагментов, Fv-фрагментов и scFv-фрагментов хорошо известно.
Трансформацию соответствующих клеток-хозяев рекомбинантной молекулой ДНК выполняют способами, которые обычно зависят от типа используемого вектора. Что касается трансформации прокариотических клеток-хозяев, см., например, Cohen et al., Proceedings National Academy of Science, USA, Vol.69, P. 2110 (1972); и Maniatis et al., Molecular Cloning, a Laboratory Manual, Cold spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1982). Что касается трансформации клеток позвоночных ретровирусными векторами, содержащими рДНК, см., например, Sorge et al., Mol. Cell. В/о/., 4:1730-1737 (1984); Graham et al., Virol., 52:456 (1973); и Wigler et al., Proceedings National Academy of Sciences, USA, Vol.76, P. 1373-1376 (1979).
В другом аспекте в описании предложены набор или база данных, содержащие данные о последовательностях, иллюстрирующие нуклеиновые кислоты, кодирующие антитела или их фрагменты, содержащие нуклеиновые кислоты, присутствующие в образце из по меньшей мере 1×105 "наивных" человеческих В-клеток, где указанные данные о последовательностях находятся на считываемом носителе.
В другом аспекте в описании предложен способ получения коллекции синтетических нуклеиновых кислот, кодирующих антитела или их фрагменты, включающий синтез коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH и VL, присутствующие в концентрации по меньшей мере 0,05% пар класса VH и VL, находящихся в образце из по меньшей мере примерно 2500 В-клеток. В некоторых воплощениях в описании предложен способ получения коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, по существу представляющие иммунный репертуар, включающий: (а) идентификацию пар класса VH/VL, присутствующих в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре; (б) синтез коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH/VL в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре. В некоторых воплощениях идентификация пар класса VH/VL включает: (1) выделение В-клеток из одного или более хозяев-людей; (2) определение пар класса VH/VL в каждой В-клетке способом, выбранным из: (А) выделение и секвенирование ДНК, мРНК или кДНК, кодирующей пары класса VH/VL; или (В) исследование с помощью одного или более нуклеиновокислотных зондов, специфичных к каждому VH и VL; и (3) анализ пар класса VH/VL. В некоторых воплощениях идентификация пар класса VH/VL включает: (1) получение последовательности нуклеиновых кислот антител; (2) определение пар класса VH/VL посредством выравнивания последовательностей; (3) сопоставление таких последовательностей из по меньшей мере 100 антител для идентификации пар класса VH/VL, присутствующих в иммунном репертуаре. В некоторых воплощениях способы включают селекцию пар класса VH/VL, демонстрирующих по меньшей мере одно биофизическое свойство, выбранное из группы, состоящей из: (1) эффективного экспонирования на фаге; (2) эффективного экспонирования на клетках млекопитающих; (3) значительной экспрессии в E.coli в Fab-формате;
(4) значительной экспрессии в клетках млекопитающих в IgG-формате; (5) термостабильности; (6) растворимости; и (7) низкой иммуногенности; и синтезирование коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, демонстрирующие по меньшей мере одно из указанных биофизических свойств. В некоторых воплощениях коллекция нуклеиновых кислот, кодирующих антитела или их фрагменты, является эмбриональной, по существу эмбриональной или кодон-оптимизированными вариантами нуклеиновых кислот эмбрионального типа. В некоторых воплощениях во время синтеза коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH/VL, изменение последовательности вводят по меньшей мере в один участок, определяющий комплементарность (CDR). В некоторых воплощениях изменение последовательности ограничивается последовательностями без стоп-кодонов. В некоторых воплощениях способы дополнительно включают клонирование коллекции нуклеиновых кислот в вектор. В некоторых воплощениях вектор выбран из группы, состоящей из: (1) дисплейного вектора, (2) фагового дисплейного вектора; (3) фагмидного вектора; и (4) экспрессирующего вектора млекопитающих. В некоторых воплощениях способы дополнительно включают трансформацию в клетку-хозяина. В некоторых воплощениях клетки-хозяева выбраны из группы, состоящей из: (1) прокариотических клеток-хозяев; (2) эукариотических клеток-хозяев: (3) клеток-хозяев Е.соli; и (4) клеток-хозяев млекопитающих.
Некоторые воплощения также включают встраивание указанных нуклеиновых кислот в коллекцию векторов и трансформормирование/ трансфицирование клетки-хозяина и экспонирование антител или их фрагментов. В некоторых воплощениях векторы представляют собой экспрессирующие векторы, дисплейные векторы, такие как фагмидный вектор. Некоторые воплощения также включают трансфицирование указанных векторов в подходящую клетку-хозяина. В некоторых воплощениях клетка-хозяин является прокариотической, такой как Е.соli, или эукариотической, такой как клетка млекопитающего.
В другом аспекте в описании предложен способ идентификации антитела или фрагментов антитела, специфических в отношении иммуногена, включающий стадии синтеза коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH и VL, присутствующие в концентрации по меньшей мере 0,05% пар класса VH и VL, находящихся в образце из по меньшей мере примерно 2500 В-клеток; скрининг коллекции против специфического иммуногена; и выбор одного или более антител или их фрагментов, специфических в отношении указанного иммуногена. Некоторые воплощения включают способ идентификации антитела или фрагментов антитела, специфических в отношении иммуногена, включающий стадии: (а) идентификации пар класса VH/VL, присутствующих в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре; (б) синтеза коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH/VL, присутствующие в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре; (в) экспонирования или экспрессирования антитела или его фрагмента из коллекции; (г) скрининга коллекции против специфического иммуногена; и (г) выбора по меньшей мере одного антитела или его фрагмента, специфического в отношении указанного иммуногена. Некоторые воплощения включают способ идентификации антитела или фрагмента антитела для лечения заболевания или состояния, включающий стадии: (а) идентификации пар класса VH/VL, присутствующих в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре; (б) идентификации пар класса VH/VL, демонстрирующих по меньшей мере одно биофизическое свойство, выбранное из группы, состоящей из: (1) эффективного экспонирования на фаге; (2) эффективного экспонирования на клетках млекопитающих; (3) значительной экспрессии в Е.соli в Fab-формате; (4) значительной экспрессии в клетках млекопитающих в IgG-формате; (5) термостабильности; (6) растворимости; и (7) низкой иммуногенности; (в) синтеза коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH/VL, присутствующие в концентрации по меньшей мере 0,05%, по меньшей мере 1% или по меньшей мере 2% пар класса VH/VL в иммунном репертуаре и демонстрирующие по меньшей мере одно биофизическое свойство из (1) - (7); (г) экспонирования или экспрессирования антитела или его фрагмента из коллекции; (д) скрининга коллекции против специфического иммуногена, ассоциированного с данным заболеванием или состоянием; и (е) выбора по меньшей мере одного антитела или его фрагмента, специфического в отношении указанного иммуногена.
В некоторых воплощениях В-клетки выделены из хозяина-человека. В некоторых воплощениях В-клетки являются "наивными". В некоторых воплощениях пары класса VH и VL, находящиеся в образце из по меньшей мере 1 Х примерно 2500 В-клеток, идентифицируют посредством способа, включающего сбор "наивных" В-клеток от одного или более хозяев-людей; выделение ДНК из собранных В-клеток; и анализ выделенной ДНК. В некоторых воплощениях стадия анализа ДНК включает секвенирование ДНК. В некоторых воплощениях стадия анализа ДНК дополнительно включает определение частоты, с которой каждая пара класса VH и VL находится в образце.
Некоторые воплощения также включают встраивание указанных нуклеиновых кислот в коллекцию векторов и трансформацию/трансфицирование в клетку-хозяина и экспонирование антител или их фрагментов.
В другом аспекте в описании предложен способ идентификации антитела или фрагментов антитела, специфических в отношении иммуногена, включающий стадии синтеза коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH и VL, присутствующие в концентрации по меньшей мере 0,05% пар класса VH и VL, находящихся в образце из по меньшей мере примерно 2500 В-клеток; скрининг коллекции против специфического иммуногена; и выбор одного или более антител или их фрагментов, специфических в отношении указанного иммуногена.
В некоторых воплощениях В-кпетки выделены из хозяина-человека. В некоторых воплощениях В-клетки являются "наивными". В некоторых воплощениях пары класса VH и VL в образце из по меньшей мере 2500 В-клеток определены посредством способа, включающего сбор В-клеток от одного или более хозяев-людей; выделение ДНК из собранных В-клеток; и анализ выделенной ДНК. В некоторых воплощениях стадия анализа ДНК включает секвенирование ДНК. В некоторых воплощениях стадия анализа ДНК дополнительно включает определение частоты, с которой каждая пара класса VH и VL находится в образце.
В некоторых воплощениях коллекция экспонируется перед тестированием/скринингом с использованием фагового, дрожжевого, рибосомального, бактериального или эукариотического дисплея. В некоторых воплощениях коллекция экспонируется на прокариотических или эукариотических клетках. В некоторых воплощениях коллекция экспонируется в Fab- или IgG-формате или другом формате, известном специалисту в данной области техники.
Скрининг можно выполнять с использованием одного из способов, хорошо известных в данной области техники, таких как фаговый дисплей, селетивно инфекционный фаг, полисомная технология для скрининга связывания и аналитические системы для ферментативной активности или стабильности белка. Многие такие способы известны специалистам и в качестве примерных ссылок предложены следующие: Valle RP, Curr. Opin. Drug Discov. Devel. 2003 Mar; 6(2): 197-203; Ackermann BL Expert Rev. Proteomics. 2007 Apr; 4(2):175-86; и Anderson KS J Proteome Res. 2005 Jul-Aug; 4(4):1123-33.
В одном воплощении скрининговые анализы выполняют так, что связывание лиганда антителом приводит к детектируемому сигналу, либо прямо, либо опосредованно. Такие сигналы включают, например, получение комплекса, образование продукта каталитической реакции, высвобождение или поглощение энергии и тому подобное. Клетки из популяции, подвергнутые трансформации с помощью рекомбинантной ДНК, можно клонировать с получением моноклональных колоний, например. Клетки, образующие эти колонии, можно собрать, лизировать и их ДНК содержимое исследовать на присутствие рекомбинантной ДНК с использованием способа, известного в данной области, например, как описано в Southern, J. Mol. Biol., 98:503 (1975) или Berent etal., Biotech. 3:208 (1985).
Биофизические свойства
Изобретение также включает коллекции и способы получения таких коллекций, в которых пары класса VH/VL имеют желательные биофизические свойства. Предпочтительные и желательные биофизические свойства включают более высокую стабильность, более высокий уровень экспрессии и низкую тенденцию к агрегации.
Подходящие биофизические свойства облегчают применение коллекции на различных стадиях. Например, скрининг коллекции облегчается, если антитела или их фрагменты являются растворимыми и не агрегируют, и хорошо экспрессируются при скрининге фона, такого как фаг.Дальнейшая разработка антитела, например, для тестирования на животных и терапевтических применений, облегчается такими свойствами антител, как растворимость, термостабильность, высокие уровни экспрессии (особенно в виде IgG в клетках млекопитающих) и низкая иммуногенность.
Для того чтобы все или по меньшей мере большинство антител или их фрагментов имели такие предпочтительные биофизические свойства, пары класса VH/VL могут быть подвергнуты скринингу заранее для определения того, какие пары класса демонстрируют эти свойства. Библиотеку затем конструируют посредством синтеза нуклеиновых кислот, кодирующих только антитела с такими предпочтительными биофизическими свойствами.
Конечно, не все пары класса VH/VL будут демонстрировать все биофизические свойства в равной степени, и средний специалист определит, какие свойства являются более подходящими и/или баланс каждого из свойств перед определением того, какие пары класса VH/VL следует синтезировать.
Таким образом, в некоторых аспектах в настоящем изобретении предложена библиотека синтетических антител, подвергнутая селекции в отношении комбинаций VH/VL, которые эффективно экспонируются, например, на поверхности фага или в других дисплейных технологиях. Предпочтительно все, в основном все или по существу все комбинации VH/VL эффективно экспонируются. Эффективность экспонирования можно измерить посредством "сэндвич"-фагового ИФА, как описано в настоящем изобретении.
В других аспектах в настоящем изобретении предложена библиотека синтетических антител, подвергнутая селекции в отношении комбинаций VH-VL, которые хорошо экспрессируются в E.coli в Fab-формате. Предпочтительно все, в основном все или по существу все комбинации VH/VL хорошо экспрессируются в Е coli в Fab-формате. Экспрессию в Fab-формате в E.coli можно определить количественно и она составляет предпочтительно более 2 мг/л, более 5 мг/л, более 10 мг/л или более 15 мг/л в бактериальной культуре. В некоторых аспектах все VH-VL экспрессируются на уровне более чем 2 мг/л, по существу все комбинации VH-VL экспрессируются на уровне более чем 5 мг/л, большинство комбинаций VH-VL экспрессируются на уровне более чем 10 мг/л в бактериальной культуре и/или по меньшей мере две, по меньшей мере три, по меньшей мере четыре или по меньшей мере пять комбинаций VH-VL экспрессируются на уровне более чем 15 мг/л в бактериальный культуре.
В некоторых аспектах в настоящем изобретении предложена библиотека синтетических антител, подвергнутая селекции в отношении комбинаций VH-VL, хорошо экспрессирующихся в системе млекопитающих в IgG-формате. Подавляющее большинство терапевтических биологических препаратов на основе антител, имеющихся на рынке в настоящее время, находится в IgG-формате по многим причинам: (1) период полувыведения молекул IgG из организма человека является очень высоким (примерно 3 недели) из-за взаимодействия IgG с неонатальным рецептором (FcRn); (2) молекулы IgG являются высоко растворимыми, термодинамически стабильными и относительно устойчивыми к протеазам в крови; и (3) IgG обладают ADCC (антитело-зависимая клеточная цитотоксичность) и/или CDC (комплемент-зависимая цитотоксичность) активностью, которые требуются для элиминации опухолевых клеток. Экспрессия конкретной VL/VH-комбинации в Fab-формате не обязательно коррелирует с экспрессией той же VL/VH-комбинации в IgG-формате, и поэтому экспрессия и растворимость комбинаций VL/VH в IgG-формате также являются важными независимыми факторами.
Система млекопитающих может включать, например, суспензионную культуру млекопитающих, культуру прикрепленных клеток млекопитающих, клетки НКВ11, клетки PERC.6 или клетки СНО. Предпочтительно, все, в основном все или по существу все комбинации VH/VL хорошо экспрессиются в системе млекопитающих в IgG-формате. В некоторых аспектах в настоящем изобретении предложена библиотека синтетических человеческих антител, где все комбинации VH-VL экспрессируются на уровне более чем 10 мг/л в системе млекопитающих в IgG-формате, где по существу все комбинации VH-VL экспрессируются на уровне более чем 15 мг/л в системе млекопитающих в IgG-формате; где большинство комбинаций VH-VL экспрессируются на уровне более чем 20 мг/л в системе млекопитающих в IgG-формате; и/или по меньшей мере три, по меньшей мере четыре или по меньшей мере пять комбинаций VH-VL экспрессируются на уровне более чем 25 мг/л в системе млекопитающих в IgG-формате.
В некоторых аспектах в настоящем изобретении предложена библиотека синтетических антител, подвергнутая селекции в отношении комбинаций VH/VL, которые являются термостабильными. Предпочтительно, все, в основном все или по существу все комбинации являются термостабильными с Т.пл., равной по меньшей мере 68, 70, 72, 74 или 76°С. Термостабильность можно измерять, как описано в данной заявке. В некоторых аспектах в настоящем изобретении предложена библиотека синтетических человеческих антител, где по существу все комбинации VH-VL имеют Т.пл. более 68°С; по существу все комбинации VH-VL имеют Т.пл. более 70°С или более 72°С; большинство комбинаций VH-VL имеют Т.пл. более 74°С; и/или многие комбинации VH-VL имеют Т.пл. более 76°С. В некоторых аспектах по меньшей мере три, по меньшей мере четыре или по меньшей мере пять комбинаций VH-VL имеют Т.пл. более 70°С.
В некоторых аспектах в настоящем изобретении предложена библиотека синтетических антител, подвергнутая селекции в отношении комбинаций VH-VL, которые являются растворимыми, т.е. не склонны к агрегации. Растворимость может быть определена, например, посредством подходящего сворачивания и характеристик экспрессии тестируемого Fab в бактериальном хозяине или IgG1 в эукариотическом хозяине или агрегации после очистки, как определено посредством аналитической гель-фильтрации.
Низкая иммуногенность может быть предсказана или определена непосредственно способами, известными в данной области, а также о ней можно судить по факту, что данные пары класса VH/VL являются наиболее распространенными в репертуаре; и используемые белковые последовательности представляют собой по существу белковые последовательности эмбрионального типа.
Как описано в данной заявке, данные о последовательностях антител могут быть получены из В-клеток, например, "наивных" В-клеток, общедоступных баз данных и/или литературы. Каждую из последовательностей антител можно выравнивать относительно наиболее близкого семейства и/или гена эмбрионального типа. Из этих данных можно определить семейства и/или гены VH и VL эмбрионального типа и/или пары класса VH/VL, которые являются наиболее распространенные.
Как только определены семейства и/или гены VH и VL эмбрионального типа и/или пары класса VH/VL, которые являются наиболее распространенными, можно выбрать, какое семейство и/или гены VH и VL эмбрионального типа и/или пары класса VH/VL следует тестировать в отношении предпочтительных биофизических свойств. Один подход заключается в ранжировании семейств и/или генов VH и VL эмбрионального типа в соответствии с распространенностью и последующее тестирование семейств и/или генов VH и VL эмбрионального типа, которые являются наиболее распространенными, например, верхних 20 наиболее распространенных семейств и/или генов VH и VL эмбрионального типа. Кроме того, можно комбинировать верхние 20 наиболее распространенных семейств и/или генов VH и VL эмбрионального типа, что дает, например, 400 комбинаций VH и VL, и тестировать их в отношении предпочтительных биофизических свойств. Кроме того или дополнительно, можно тестировать пары класса VH/VL, которые являются наиболее распространенными, в отношении предпочтительных биофизических свойств.
Предпочтительные биофизические свойства включают, но не ограничиваются этим, то, что: (1) они хорошо экспонируются на фаге в Fab-формате, (2) они хорошо экспонируются на клетках млекопитающих в IgG-формате, (3) они экспрессируются в высоких количествах в Fab-формате, например, in E.соli, и IgG-форматах, например, в клетках млекопитающих, (4) являются термодинамически стабильными; (5) имеют высокую стабильность в сыворотке, (6) имеют низкую тенденцию к агрегации (т.е. высокую растворимость); и (7) имеют низкий риск иммуногенности.
В некоторых аспектах настоящее изобретение включает коллекцию синтетических нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH и VL, присутствующие в концентрации по меньшей мере 0,5% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток. В некоторых воплощениях пары класса VH и VL присутствуют в концентрации по меньшей мере 1% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток. В некоторых воплощениях пары класса VH и VL присутствуют в концентрации по меньшей мере 2% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105В-клеток. В некоторых воплощениях В-клетки выделяют из хозяина-человека. В некоторых воплощениях В-клетки являются "наивными". В некоторых аспектах коллекция содержит нуклеиновые кислоты, кодирующие антитела или их фрагменты, содержащие каркасные участки VH и VL эмбрионального типа. В некоторых воплощениях коллекция содержит по меньшей мере 1×104последовательностей нуклеиновых кислот, по меньшей мере 1×106последовательностей нуклеиновых кислот; по меньшей мере 1×108последовательностей нуклеиновых кислот, по меньшей мере 1×1010последовательностей нуклеиновых кислот или по меньшей мере 1×1011последовательностей нуклеиновых кислот.
В некоторых аспектах настоящее изобретение включает набор, содержащий данные о последовательностях, иллюстрирующие нуклеиновые кислоты, кодирующие антитела или их фрагменты, содержащие нуклеиновые кислоты, присутствующие в образце из по меньшей мере 1×105 "наивных" В-клеток человека, где указанные данные о последовательностях находятся на считываемом носителе. В некоторых воплощениях изобретение включает коллекцию нуклеиновых кислот, кодирующих антитела или их функциональные фрагменты. В некоторых воплощениях вектор представляет собой фаговый дисплейный вектор. В некоторых воплощениях вектор представляет собой фагмидный вектор. В некоторых аспектах настоящее изобретение включает клетки-хозяева, трансформированные с помощью векторов коллекции, которые содержат коллекцию нуклеиновых кислот, кодирующих антитела или их функциональные фрагменты. В некоторых воплощениях клетки-хозяева являются прокариотическими. В некоторых воплощениях клетки-хозяева представляют собой Е.соli. В некоторых воплощениях клетки-хозяева являются эукариотическими. В некоторых воплощениях клетки-хозяева представляют собой клетки млекопитающих.
В некоторых аспектах настоящее изобретение включает способ получения коллекции синтетических нуклеиновых кислот, кодирующих антитела или их фрагменты, включающий: синтез коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH и VL, присутствующие в концентрации по меньшей мере 0,5% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×105 В-клеток. В некоторых воплощениях В-клетки выделяют из хозяина-человека. В некоторых воплощениях В-клетки являются "наивными". В некоторых воплощениях пары класса VH и VL, находящиеся в образце из по меньшей мере 1×105 В-клеток, идентифицируют с помощью способа, включающего:
аа) сбор "наивных" В-клеток от одного или более хозяев-людей;
аб) выделение ДНК из В-клеток, собранных на стадии (аа); и
ав) анализ ДНК, выделенной на стадии (аб).
В некоторых воплощениях стадия анализа ДНК включает секвенирование ДНК. В некоторых воплощениях стадия анализа ДНК дополнительно включает определение частоты, с которой каждая пара класса VH и VL находится в образце. В некоторых воплощениях способы дополнительно включают встраивание нуклеиновых кислот в коллекцию векторов. В некоторых воплощениях вектор представляет собой экспрессирующий вектор. В некоторых воплощениях вектор представляет собой дисплейный вектор. В некоторых воплощениях дисплейный вектор представляет собой фагмидный вектор. В некоторых воплощениях способ дополнительно включает трансфицирование указанных векторов в подходящую клетку-хозяина. В некоторых воплощениях клетка-хозяин является прокариотической. В некоторых воплощениях клетка-хозяин представляет собой Е.coli. В некоторых воплощениях клетка-хозяин является эукариотической. В некоторых воплощениях клетка-хозяин представляет собой клетку млекопитающих.
В некоторых аспектах настоящее изобретение включает способ идентификации антитела или фрагментов антитела, специфических в отношении иммуногена, включающий стадии: а) синтеза коллекции нуклеиновых кислот, кодирующих антитела или их фрагменты, содержащие пары класса VH и VL, присутствующие в концентрации по меньшей мере 0,5% пар класса VH и VL, имеющихся в образце из по меньшей мере 1×10 В-клеток; б) скрининга коллекции против специфического иммуногена; и в) выбора одного или более антител или их фрагментов, специфических в отношении указанного иммуногена. В некоторых воплощениях В-клетки выделены из хозяина-человека. В некоторых воплощениях В-клетки являются "наивными". В некоторых воплощениях пары класса VH и VL, находящиеся в образце из по меньшей мере 1×105 В-клеток определены с помощью способа, включающего:
аа) сбор "наивных" В-клеток от одного или более хозяев-людей;
аб) выделение ДНК из В-клеток, собранных на стадии (аа); и
ав) анализ ДНК, выделенной на стадии (аб).
В некоторых воплощениях стадия анализа ДНК включает секвенирование ДНК. В некоторых воплощениях стадия анализа ДНК дополнительно включает определение частоты, с которой каждая пара класса VH и VL находится в образце. В некоторых воплощениях стадия синтеза коллекции дополнительно включает встраивание указанных нуклеиновых кислот в коллекцию векторов. В некоторых воплощениях способ дополнительно включает трансфицирование указанных векторов в подходящую клетку-хозяина. В некоторых воплощениях способ дополнительно включает экспонирование указанную коллекции.
Примеры
Пример 1: Получение сайтов рестрикции на С-конце прокариотической сигнальной___последовательности и человеческой лидерной последовательности, обеспечивающие полноразмерные участки FR1 эмбрионального типа
В одном аспекте в настоящем описании описаны коллекции антител, содержащие каркасные участки, содержащие белковые последовательности эмбрионального типа, конкретно FR1. Ожидают, что имеющиеся последовательности эмбрионального типа будут снижать риск иммуногенности антител при введении человеку. Однако следует использовать совместимые сайты рестрикции, позволяющие осуществлять стандартное клонирование нуклеиновых кислот, кодирующих коллекции антител, в дисплейные и/или экспрессирующие векторы, с тем чтобы антитела можно было подвергнуть скринингу против иммуногенов. Раньше сайты рестрикции, используемые для клонирования, часто размещали в пределах каркасных участков, таким образом, модифицируя нуклеиновую кислоту и/или аминокислотную последовательность в направлении от эмбрионального типа. Для обеспечения того, чтобы по меньшей мере каркасный участок 1 (FR1) каждого из антител по настоящему изобретению сохранял белковую последовательность эмбрионального типа, не должно быть неприродных сайтов рестрикции в пределах FR1. Поэтому аспект настоящего изобретения заключается во включении идентичного или по меньшей мере совместимого сайта рестрикции в пределах С-конца прокариотических сигнальных последовательностей и человеческой лидерной последовательности, конкретно в пределах трех С-концевых остатков. Кроме того, сигнальная последовательность и лидерная последовательность, содержащие идентичный или совместимый сайт рестрикции, должны быть функциональными и позволять хорошие уровни экспонирования и эспрессии антител или их фрагментов как в прокариотических экспрессирующих системах, так и в экспрессирующих системах млекопитающих.
Пример 1.1: Анализ распространенных аминокислотных остатков на С-конце сигнальных последовательностях Е.соli
Ниже описан выбор сайтов рестрикции, подлежащих включению в С-конец сигнальной последовательности как оmрА, так и phoA Е.соli, а также оценка функциональности полученных сигнальных последовательностей.
На первой стадии анализировали общие аминокислотные остатки в трех С-концевых аминокислотах сигнальных последовательностей (с -3 до -1) и получали консенсусную последовательность, как показано в Таблице 1. См. Chou et al., Prediction of protein signal sequences, Protein Pept. Sci. 3(6):615-22 (December 2002).
Таблица 1: Консенсусные последовательности в трех С-концевых аминокислотах сигнальных последовательностей:
В положении -3 наблюдали преимущественно аминокислоты A, S, V и Т. В положении -2 наблюдали преимущественно аминокислоты L, А, S и Q. В положении -1 наблюдали преимущественно аминокислоты A, G и S.
Пример 1.2: Выбор сайта рестрикции для сигнальной последовательности (phoA) E.coli тяжелой цепи
После сравнения консенсусных последовательностей, показанных в Таблице 1, с известными сайтами рестрикции, следующие три сайта рестрикции: Aflll, Nhel и Avrll выбрали для встраивания в С-конец phoA и затем исследовали уровни экспрессии. Важно отметить, что изменение нуклеотидных последовательностей дикого типа на модифицированные нуклеотидные последовательности также приводит к изменению аминокислотных последовательностей. Последовательности нуклеиновых кислот выбранных сайтов рестрикции и соответствующие аминокислотные последовательности показаны в Таблице 2.
В качестве контроля исследовали уровни экспрессии сигнальной последовательности phoA дикого типа. Нуклеиновокислотные и аминокислотные последовательности сигнальной последовательности phoA дикого типа, включая 3 С-концевые последовательности, показаны в Таблице 3.

Для оценки уровней экспрессии сайты рестрикции, показанные в Таблице 2, включали в сигнальную последовательность phoA, таким образом модифицируя аминокислотную последовательность дикого типа. Полученные сигнальные последовательности использовали для экспрессии Fab-фрагментов, содержащих: белковые последовательности эмбрионального типа a) VH3-23 или б) VH1-69. Эти гены эмбрионального типа выбрали, так как известно, что они стабильны и хорошо экспрессируются. В обе последовательности генов эмбрионального типа VH3-23 и VH1-69 встраивали CDR-H3 (WGGDGFYAMDY) из антитела 4D5 и последовательность гена JH4 эмбрионального типа использовали для FR4. Антитело 4D5 описано в (PDB entry 1FVC; Carter, P., Presta, L, German, С.M., Ridgway, J. В., Henner, D., Wong, W. L. et al. (1992); Humanization Biophysical Properties of Human Antibody Domains 551 of an anti-pi 85HER2 antibody for human cancer therapy. Proc. NatI Acad. Sci. USA, 89, 4285-4289. Создавали плазмиды на основе pMORPHXH (показан на Фиг.50), содержащие: а) сигнальные последовательности phoA, содержащие С-концевые сайты рестрикции и аминокислотные последовательности из Таблицы 2, б) VH последовательности VH3-23 и VH1-69, включающие CDR-H3 и JH4, как описано выше, и в) стабильную и хорошо экспрессирующуюся легкую цепь из MOR03207. Все гены создавали в Geneart (Regensburg, Германия).
Экспрессию и периплазматический транспорт измеряли, выполняя анти-Fd ИФА после периплазматической экстракции. Результаты ИФА экспрессии анти-Fd после периплазматической экстракции с использованием буфера BBS показаны на Фиг.1. Как показано, в группе VH3-23 сигнальные последовательности, включающие С-концевые сайты рестрикции: Aflll (VLS), Nhel (VLA) и Avrll (VLG), поддерживали уровни экспрессии в диапазоне дикого типа (ТКА) с Nhel (VLA), более эффективным, чем дикий тип (ТКА).
Кроме того, экспрессию Fab в Е.соli выполняли после культивирования в течение ночи в колбах при встряхивании и уровни продукции Fab определяли после очистки Fab посредством аффинной хроматографии и замены буфера. Результаты показаны в Таблице 4.
Как показано, сигнальные последовательности, включающие С-концевые сайты рестрикции Aflll (VLS), Nhel (VLA) и Avrll (VLG), по сравнению с диким типом (ТКА) экспрессируют сходные количества Fab.
На основании приведенных выше данных, сайт рестрикции Nhel (VLA) выбрали для включения в сигнальные последовательности тяжелой цепи (рпоА). Нуклеиновокислотные и аминокислотные последовательности модифицированной сигнальной последовательности Nhel (VLA) phoA показаны в Таблице 5.

Пример 1.3: Выбор сайта рестрикции для сигнальной последовательности Е.соli(оmрА) каппа- и лямбда-легкой цепи:
Способ, аналогичный описанному в Примерах 1.2, использовали для выбора сайтов рестрикции, подлежащих включению в С-концы сигнальных последовательностей (ompA) легких цепей и для каппа, и для лямбда.
После сравнения консенсусных последовательностей, показанных в Таблице 1, с известными сайтами рестрикции, следующие сайты рестрикции:
Ndel (AYG), Ndel (AYA) и BsiWI (TYA) выбрали для включения в С-конец ompA, таким образом модифицируя также аминокислотные последовательности, и затем исследовали уровни экспрессии. Последовательности выбранных сайтов рестрикции показаны в Таблице 6.
В качестве контроля исследовали уровни экспрессии сигнальной последовательности ompA дикого типа. Нуклеиновокислотные и аминокислотные последовательности сигнальной последовательности ompA дикого типа, включая 3 С-концевые последовательности, показаны в Таблице 7.

Для оценки уровней экспрессии сайты рестрикции, показанные в Таблице 6, включали в сигнальные последовательности ompA. Полученные модифицированные сигнальные последовательности использовали для экспрессии Fab-фрагментов, содержащих последовательности генов эмбрионального типа: а) КаппаЧ 012 (IGKV1-39), б) КарраЗ L6 (IGKV3-11) или в) Лямбда1 V1-13 (IGLV1-40). Эти гены эмбрионального типа выбрали, так как известно, что они стабильны и хорошо экспрессируются. В а) Каппа1 012 (IGKV1-39) и б) Карра3 L6 (IGKV3-11), включали участок CDR-L3: QQHYTTPPT (для каппа), в в) Лямбда1 V1-13 (IGLV1-40) включали участок CDR-L3: QSYDSSLSGW (для лямбда), и в а)-в), использовали последовательность гена эмбрионального типа Jk1 в качестве FR4 для каппа-легкой цепи; и последовательность гена эмбрионального типа JI2/3 использовали в качестве FR4 для лямбда-легкой цепи. Получали плазмиды pMORPHXH (показаны на Фиг.50), содержащие: а) сигнальные последовательности оmрА, содержащие С-концевые сайты рестрикции из Таблицы 6, б) последовательности VL эмбрионального типа Каппа1 012 (IGKV1 -39), б) КарраЗ L6 (IGKV3-11), или в) Лямбда1 V1-13 (IGLV1-40), включающие CDR-L3 и FR4, как описано выше, и в) конструкцию IGHVH3-23 ТКА в качестве тяжелой цепи, описанную в Примере 1.2. Все гены получали в Geneart (Regensburg, Германия).
Экспрессию и периплазматический транспорт измеряли посредством продуцирования Fab в течение ночи в Е.соli, периплазматической экстракции и ИФА анти-Fd после периплазматической экстракции. Результаты ИФА экспрессии анти-Fd после периплазматической экстракции с использованием буфера BBS показаны на Фиг.2. Как видно, сигнальная последовательность, включающая Ndel (AYA), демонстрирует такую же или даже лучшую экспрессию, чем дикий тип (AQA).
Кроме того, экспрессию Fab в Е.соli определяли после ночного культивирования в колбах при встряхивании и уровни продукции Fab определяли посредством аффинной хроматографии и замены буфера. Результаты показаны в Таблице 8.
На основании вышеприведенных данных, сайт рестрикции Ndel (AYA) выбрали для включения в сигнальные последовательности (ompA) каппа и лямбда. Нуклеиновокислотные и аминокислотные последовательности модифицированной сигнальной последовательности Ndel (AYA) ompaA показаны в Таблице 9.

Пример 1.4: Оценка эффективности экспонирования Fab-фрагментов с помощью сигнальных последовательностей в фаговом дисплее
Как описано в Примерах 1.2 и 1.3, были выбраны следующие сайты рестрикции для включения в С-концы сигнальных последовательностей Fab и лидерные последовательности IgG:
Вариабельные области тяжелой цепи (рпоА и лидер тяжелой цепи): Nhel (VLA)
Вариабельные области легкой цепи (к и А) (оmрА и каппа лидер): Ndel (AYA)
На Фиг.3 показаны выбранные сайты рестрикции и соответствующие аминокислотные последовательности.
Для того чтобы показать, что эти модифицированные сигнальные последовательности способствуют эффективному транспорту и продукции Fab-фрагментов, создавали векторные конструкции, включающие выбранные сигнальные последовательности в трицистронные дисплейные векторы, которые кодируют VH, VL и pill (фаговый оболочечный белок pill, используемый для фагового дисплея). Это делали для того, чтобы подтвердить, что такие векторы, содержащие выбранные сигнальные последовательности, способны обеспечивать полезные уровни фагового дисплея. Создавали трицистронные векторные конструкции pJPd1 (показаные на Фиг.48), содержащие VH из генов эмбрионального типа VH3-23 или VH1-69, или VL из генов эмбрионального типа VL1-40, VK3-11 или VK1-39, и выбранные сайты рестрикции тяжелой цепи (ompА): Nhel (VLA) и ompА дикого типа (ТКА) в качестве контроля, или выбранные сайты рестрикции легкой цепи (ompA): Ndel (AYA) и оmрА дикого типа (AQA), в качестве контроля. Кроме того, в качестве контролей получали трицистронные векторные конструкции pMORPHSO (показаны на Фиг.51), содержащие то же. Относительные уровни экспонирования показаны в Таблице 10.
н/д - не делали
Как показано, векторы pJPd1, включающие выбранные сигнальные последовательности, давали сравнимые относительные уровни экспонирования, как и векторы рМОРРН30; и превосходные относительные уровни экспонирования по сравнению с векторами pMORPHSO векторы, при использовании гиперфага в качестве вспомогательного фага для фаговой продукции. Поэтому векторы pJPd1, включающие модифицированные сигнальные последовательности, должны хорошо работать для селекции антител или их функциональных фрагментов против целевых антигенов посредством фагового дисплея.
Примеры 1.2-1.4 описывают необходимые инструменты для создания, экспрессии и экспонирования коллекций антител или функциональных фрагментов по настоящему изобретению, поскольку они описывают сигнальные последовательности и лидерные последовательности, содержащие сайты рестрикции, которые обеспечивают участки FR1 с белковыми последовательностями эмбрионального типа, и описывают векторные остовы, полезные для включения описанных коллекций антител или их функциональных фрагментов в систему селекции с помощью фагового дисплея, или экспрессирующие системы млекопитающих для идентификации антител против любого иммуногена. Более того, сигнальные последовательности несут сайты рестрикции, которые обеспечивают полную совместимость с Fab-фаговым дисплеем и экспрессирующими плазмидами и соответствующими IgG экспрессирующими плазмидами.
Пример 1.5: Тестирование лидерных последовательностей тяжелой цепи человека и каппа для экспрессии IgG. содержащих выбранные С-концевые сайты рестрикции
Для того чтобы осуществить легкий переход от Fab, экспрессирующихся в Е.соli, к IgG-форматам, экспрессирующимся в системе млекопитающих, создавали человеческие лидерные последовательности для легкой цепи IgG (лидерная последовательность каппа-цепи человека) и тяжелой цепи IgG (лидерная последовательность тяжелой цепи человека), чтобы они содержали те же сайты рестрикции, что и С-концы сигнальных последовательностей оmрА (Ndel (AYA)) и phoA (Nhel (VLA)), таким образом модифицируя также некоторые из трех С-концевых аминокислотных последовательностей.
Перенос VH из плазмиды для экспрессии Fab в Е.соli в IgG-экспрессирующий вектор млекопитающих можно выполнить с использованием описанного сайта рестрикции Nhel, который находится: (а) на С-конце сигнальной последовательности phoA, а также (б) в соответствующем положении на С-конце лидерной последовательности тяжелой цепи человека. Для того чтобы выполнить это, три концевые аминокислоты сигнальной последовательности phoA модифицировали (из ТКА в VLA), и С-конец лидерной последовательности тяжелой цепи человека адаптировали путем замены аминокислотной последовательности дикого типа (с -3 до -1) с VLS на рпоА-совместимый VLA.
Перенос VL из плазмиды для экспрессии Fab в Е.соli в IgG-экспрессирующий вектор млекопитающих можно выполнить с использованием описанного сайта рестрикции Ndel, который находится (а) на С-конце оmрА сигнальной последовательности, а также (б) в соответствующем положении на С-конце лидерной последовательности каппа-цепи человека. Для того чтобы выполнить это, три концевые аминокислоты сигнальной последовательности оmрА модифицировали (с AQA в AYA), и С-конец лидерной последовательности каппа-цепи человека адаптировали путем замены аминокислотной последовательности дикого типа (с -3 до -1) с AYG на оmрА-совместимый AYA. Лидерные последовательности дикого типа и модифицированные лидерные последовательности тяжелой цепи человека и каппа-цепи человека показаны в Таблице 11.

Для того чтобы показать, что эти модифицированные сигнальные/лидерные последовательности способствуют эффективному транспорту и продукции человеческого белка 1д01, и модифицированную лидерную последовательность тяжелой цепи человека, и модифицированную лидерную последовательность каппа-цепи человека клонировали в плазмиды pJP_lS (показаны на Фиг.52-54) для экспрессии полноразмерного человеческого IgG1 в системе млекопитающих, заменяя лидерные последовательности дикого типа. Полученные экспрессирующие векторы, содержащие модифицированные лидерные последовательности, гены вариабельной области, как показано в Таблице 12, и константные области для каппа- или лямбда-легких цепей и тяжелых цепей трансфицировали в клетки НКВ11, и человеческий IgG1 очищали из супернатантов клеточных культур через несколько суток после трансфекции с использованием протеин А-хроматографии. Содержание IgG1 определяли после очистки и замены буфера. Выходы экспрессии показаны в Таблице 12.
Все протестированных конструкций экспрессируют высокие количества человеческого IgG1, что указывает на то, что модифицированные лидерные последовательности поддерживают уровни экспрессии. Выбранные модифицированные лидерные последовательности: (а) дают высокие выходы белка IgG в соответствии с используемой векторной системой, (б) обеспечивают полную совместимость для переключения антительных форматов, векторов и экспрессирующих систем между прокариотическими системами и системами млекопитающих, и (в) расположены в сигнальных/лидерных последовательностях, поддерживающих посредством этого полные последовательности FR1 эмбрионального типа.
Пример 2: Идентификация наиболее распространенных пар VH/VL в человеческом репертуаре
Аспект настоящего изобретения представляет собой коллекцию или библиотеку антител или их функциональных фрагментов, содержащих белковые последовательности эмбрионального типа пар генов эмбрионального типа, наиболее распространенных в иммунном репертуаре человека, где каждое антитело или его функциональные фрагменты содержат белковые последовательности эмбрионального типа соответствующей пары белков эмбрионального типа, где пары белков эмбрионального типа, выбранные для включения в коллекцию, имеют биофизические свойства, которые увеличивают вероятность того, что каждое из антител или их функциональных фрагментов, выбранных из коллекции, могут разрабатываться в клинике и коммерчески успешны. Для того чтобы создать такую коллекцию, нужно было оценить многочисленные критерии.
Как правило, осуществляли следующие стадии: идентифицировали преобладающие пары генов эмбрионального типа из иммунного репертуара человека; синтезировали кДНК преобладающих пар генов эмбрионального типа из иммунного репертуара человека и клонировали в различные векторные окружения и получали антитела или их функциональные фрагменты; антитела или их функциональные фрагменты, содержащие белковые последовательности эмбрионального типа преобладающих пар генов эмбрионального типа, функционально тестировали для определения их биофизических свойств; и сравнивали биофизические свойства антител или их функциональных фрагментов, содержащих соответствующие пары белков эмбрионального типа; затем выбирали субпопуляцию пар белков эмбрионального типа для включения в коллекцию. В некоторых воплощениях белковые последовательности эмбрионального типа выбранных пар белков эмбрионального типа действуют в качестве каркасов. В таких воплощениях каркасы содержат белковые последовательности эмбрионального типа выбранных пар белков эмбрионального типа, где оба VH и VL содержат белковые последовательности эмбрионального типа соответствующей пары по меньшей мере в FR1, CDR1, FR2, CDR2 и FR3. В конкретных воплощениях CDR3 может быть подвергнут диверсификации. В конкретных воплощениях FR4 является фиксированным, например, для VH можно использовать последовательность JH4, для каппа VL можно использовать, последовательность Jк1 и для лямбда VL можно использовать JI2/3.
Пример 2.1: Определение частоты использования генов эмбрионального типа пар VH/VL
Для того чтобы идентифицировать преобладающие пары генов эмбрионального типа VH/VL из иммунного репертуара человека, анализировали общедоступные данные и брали образцы человеческих В-клеток. В качестве первой стадии рассматривали общедоступные данные для обнаружения статей, в которых описаны пары генов эмбрионального типа VH/VL, выделенных из В-клеток. Как уже упоминалось, многие общедоступные базы данных представляют последовательности антител, однако многие представляют лишь последовательности одного из вариабельных доменов, VH или VL, но редко представляют связь пар генов эмбрионального типа VH/VL. Следующие статьи были определены и проанализированы подробно:
Wardemann H. et al. (2003) Science 301, 1374-1377, и любые вспомогательные таблицы; Yurasov S. et al. (2005) J. Exp.Med. 201, 703-712 и любые вспомогательные таблицы; Tsuiji М. et al. (2006) J. Exp.Med. 203, 393-401, и любые вспомогательные таблицы; Yurasov S. et al. (2006) J. Exp.Med. 203, 2255-2262, и любые вспомогательные таблицы, Tiller Т. et al. (2007) Immunity 26, 205-213, и любые вспомогательные таблицы, и Mietzner В. et al. (2008) PNAS 105, 9727-9732, и любые вспомогательные таблицы.
На Фиг.4-9 показаны пары VH/VL из В-клеток, выделенных в Tsuiji М. et al. (2006).
На Фиг.10-12 показаны пары VH/VL из В-клеток, выделенных в Tiller Т. et al. (2007).
На Фиг.13-17 показаны пары VH/VL из В-клеток, выделенных в Mietzner В. et al. (2008).
На Фиг.18-20 показаны пары VH/VL из В-клеток, выделенных в Wardemann H. et al. (2003).
На Фиг.21-23 показаны пары VH/VL из В-клеток, выделенных и идентифицированных в Yurasov S. et al. (2005).
На Фиг.24-26 показаны пары VH/VL из В-клеток, выделенных в Yurasov S. et al. (2006).
Дополнительные данные о парах VH/VL идентифицировали из образца человеческих В-клеток, как описано ниже.
Пример 2.2: Определение частоты использования пар генов VH/VL из человеческого образца
Для того чтобы получить дополнительные данные о частоте использования пар генов эмбрионального типа VH/VL, PBMCs (мононуклеарные клетки периферической крови) выделяли из человеческого организма. PBMCs сортировали, кДНК из В-клеток амплифицировали с использованием ПЦР, ДНК из В-клеток секвенировали и затем последовательности анализировали с помощью IgBLAST (NCBI) для идентификации пар генов эмбрионального типа VH/VL из каждой В-клетки.
Пример 2.2(а): Выделение и сортировка человеческих мононуклеарных клеток периферической крови (PBMCs)
Общие способы выделения и сортировки человеческих PBMCs из венозной крови и мононуклеарных клеток из костного мозга описаны в Tiller et al. JIM 2008. PBMCs выделяли следующим образом. 40 мл венозной крови собирали от донора-человека (через 7 суток после вакцинации Pandemrix™ (H1N1 вакцина GlaxoSmithKline)) в 4х Li-Heparin пробирки для сбора крови (Sarstedt) (10 мл в каждую). Содержимое каждой системы monovette объединяли в одном 50 мл Falcon (в общем 40 мл) и затем добавляли 100 мкл RosetteSep (StemCell technologies) (2,5 мкл/мл), хорошо перемешивали на шейкере (5 об/мин) и инкубировали при комнатной температуре в течение 30 мин. Комбинацию кровь/RosettaSep разбавляли равным объемом 1х PBS (Invitrogen). Добавляли 15 мл FicollPaque (GE Healthcare) в новые 50 мл конические пробирки и 20 мл разведенной крови наслаивали на FicollPaque, в общей сложности (4 пробирки: каждая с 15 мл FicollPaque+20 мл крови). Пробирки центрифугировали на центрифуге в течение 30 мин при 400д (1400 об/мин на лабораторной центрифуге sigma) при комнатной температуре без торможения. После центрифугирования обогащенные PBMCs образовывали слой на границе между плазмой и FicollPaque. PBMCs извлекали из каждой пробирки с помощью пипетки и переносили в новую 50 мл пробирку. PBMCs промывали путем разведения до 40 мл буфером FACS (PBS, 3% FCS) и центрифугировали на центрифуге в течение 10 мин при 1250 об/мин при 4°С.PBMCs подсчитывали с помощью трипанового синего (10 мкл образца+90 мкл трипанового синего (разбавленного PBS 1:10)).
PBMCs окрашивали путем ресуспендирования в примерно 5 мл ледяного буфера FACS. Аликвоты клеток готовили для окрашивания. Флуорофор готовили для тестирования. Аликвоты центрифугировали при 1250 об/мин, 4°С, и супернатант отбрасывали. Антитела для окрашивания добавляли к клеточным осадкам в соответствии со схемой, описанной в Таблице 13.
Затем клетки пропускали через сетчатый фильтр для клеток на FACS пробирках (Eppendorf) во избежание засорения цитометра. Клетки помещали на лед и держали в темноте.
Клетки подвергали одиночной сортировке в соответствии с клеточным поверхностным маркером фенотипа, представляющего интерес.Например, антитело-секретирующие клетки представляют собой CD19+CD20lowCD27hiCD38hi, и зрелые "наивные" В-клетки представляют собой CD19+CD27negCD10neglgM+. Присутствие клеточных поверхностных маркеров определяли с использованием мышиных антител против человека (AbD: CD19, CD27, CD38, CD-20 и CDIO) (Becton Dickinson: IgM). Клетки сортировали на основании прямого светорассеяния относительно бокового светорассеяния (канал для живых клеток с двойным распознаванием) в 96-луночные планшеты для ПЦР на одной клетке (Eppendorf), содержащие 4 мкл 0,5×PBS, 10 мМ DTT, 8 ед. RNAsin (Promega) с использованием FACS Aria.
ПЦР планшеты готовили, как показано в Таблице 14.
После сортировки каждый планшет немедленно запечатывали в фольгу (BioRad) и помещали на сухой лед. После окончания сортировки клеток все планшеты замораживали при -80°С.
Пример 2.2(б): ПЦР-амплификация ДНК В-клеток человека
PBMCs выделяли и сортировали, как описано в Примере 2.2(а). Транскрипты Ig гена одиночных отсортированных зрелых "наивных" (mn) В-клеток и антитело-секретирующих клеток (asc) затем амплифицировали с помощью ПЦР для определения спариваний генов эмбрионального типа VH/VL. Общие методы ПЦР-амплификации кДНК В-клеток и полезные для этого праймеры описаны в Tiller et al. J Immunol Methods, 2008.
Общая стратегия ПЦР показана на Фиг.27. Используемые специфические праймеры показаны в Таблице 15.

кДНК из отдельных отсортированных зрелых "наивных" (mn) В-клеток и антитело-секретирующих клеток (asc) синтезировали следующим образом. Сначала готовили RHP-Mix, RT-Mix и RT-Mix на льду. RHP-Mix готовили следующим образом: 115 мкл случайных гексамерных праймеров (Roche) (300 нг/мкл), 115 мкл NP-40 (Sigma) (10%), 35 мкл RNAsin и 542 мкл воды. RT-Mix готовили следующим образом: 660 мкл 5х RT буфера, 110 мкл dNTP (Invitrogen) (25 мМ каждый), 450 мкл воды, 220 мкл 0,1 М DTT (дитиотреитол), 44 мкл RNAsin (Promega), 55 мкл Superscript III (обратная транскриптаза) (Invitrogen).
Затем планшет помещали на сухой лед и добавляли 3,5 мкл RHP-Mix. Планшет покрывали фольгой и инкубировали при 68°С в водяной бане в течение 1 мин. Затем планшет помещали на обычный лед. После этого добавляли 7 мкл RT-Mix и лунки закрывали алюминиевой фольгой. Программу RT-Амплификации запускали при следующих температурах и в течение следующих периодов времени: 42°С в течение 5', 25°С в течение 10', 50°С в течение 60', 94°С в течение 5' и 4°С и останавливали. кДНК хранили при -20°С.
Гнездовую ПЦР (nested PCR) выполняли следующим образом. Транскрипты человеческих генов IgH, Igk и IgL V амплифицировали с помощью ПЦР независимо. 3,5 мкл кДНК использовали в качестве матрицы. Все ПЦР реакции выполняли в 96-луночных планшетах в общем объеме 40 мкл на лунку. Для каждого планшета готовили 3 реакционные пробирки, каждая с: 1154 мкл воды, 150 мкл 10х буфера, 16 мкл dNTP, 5 мкл 5' смеси праймеров, 5 мкл 3' праймера и 7 мкл HotStar Taq (Qiagen). Все реакции гнездовой ПЦР с геноспецифическими праймерами или смесями праймеров выполняли с 3,5 мкл неочищенного первого ПЦР-продукта. Каждый раунд ПЦР выполняли, как показано в Таблице 16.
Затем 3 мкл аликвоты продуктов вторых ПЦР пропускали на 2%-ном агарозном геле, содержащем бромистый этидий в буфере 1х ТВЕ с равным количеством загрузочного буфера в течение 45 мин при 150 В. Полосы ДНК визуализировали под УФ светом. Ожидаемые размеры ПЦР-продуктов составляли приблизительно 450 п.н. для Igy, 510 п.н. для 1дк и 405 п.н. для IgA.
4 мкл ПЦР-продуктов VH, Vк и VL (с выравниванием, соответствующим VH или VL продукту) объединяли с 16 мкл ddl-bO (дважды дистиллированная вода) в 96-луночных планшетах и переносили в Eurofins MWG Орегоп, Ebersberg, Германия, для секвенирования планшета. Секвенирующие праймеры были представлены при 10 пмоль/мкл (концентрат 50 пмоль/мкл, разведение 1:5) и показаны в Таблице 17.
Результаты секвенирования анализировали с помощью IgBLAST (NCBI) для идентификации генов эмбрионального типа VH, VK и VL, показанных на Фиг.28А-С (28-36).
Пример 2.3 Пары генов эмбрионального типа VH/VL. идентифицированные в иммунном репертуаре человека
Данные о парах генов эмбрионального типа VH/VL, установленные из общедоступной литературы, как описано в Примере 2.1 и показано на Фиг.4-26, объединили с данными, выявленными из человеческого образца, как описано в Примере 2.2 и показано на Фиг.28-36.
Объединенные данные проанализировали и представили в виде ранжирования в Таблице 18, т.е. ранжирования концентрации (%) пар генов эмбрионального типа VH/VL, идентифицированных в иммунном репертуаре человека.
Таблица 18: Частота использования пар генов эмбрионального типа VH/VL в иммунном репертуаре человека
Пол.: представляет положение относительного ранжирования пар VH/VL, как определено по процентному содержанию (%) каждой пары VH/VL из общего образца.
N=2137
Пример 2.4 Частота использования пар генов эмбрионального типа VH/VL в "наивном" иммунном репертуаре человека
Кроме того, объединенные данные, содержащие данные о парах генов эмбрионального типа VH/VL, установленные из общедоступной литературы, как описано в Примере 2.1 и показано на Фиг.4-26, и данные о парах генов эмбрионального типа VH/VL, выявленные из человеческого образца, как описано в Примере 2.2 и показано на Фиг.28-36, анализировали для того, чтобы определить частоту использования генов эмбрионального типа в репертуаре интактного человека. Важно различать "наивные", "необученные" антигеном, и "обученные" антигеном популяции В-клеток. "Наивные", "необученные" антигеном В-клетки включают, но не ограничиваются этим, незрелые В-клетки, новые эмигрирующие В-клетки и зрелые "наивные" В-клетки, где последовательности антител относятся еще к эмбриональному типу. "Обученные" антигеном В-клетки включают, но не ограничиваются этим, клетки, секретирующие IgG антитело, и В-клетки памяти IgM и IgG, где большинство антител содержат соматические гипермутации.
Считается, что различные пары генов эмбрионального типа сверхпредставлены между двумя популяциями В-клеток: "наивных", "необученных" антигеном В-клеток и по сравнению с "обученными" антигеном популяциями В-клеток. Аспект настоящего изобретения заключается в создании коллекции антител или их функциональных фрагментов, которые могут быть использованы для идентификации антител или их функциональных фрагментов против любого иммуногена, поэтому может быть предпочтительным получение коллекции, содержащей пары белков эмбрионального типа VH/VL, преимущественно экспрессирующиеся в интактном, "необученном" антигеном иммунном репертуаре.
Для идентификации пар генов эмбрионального типа VH/VL, преимущественно экспрессирующихся в интактном, "необученном" антигеном иммунном репертуаре, объединенные данные из общедоступной литературы, как описано в Примере 2.1 и показано на Фиг.4-26, и данные о парах генов эмбрионального типа VH/VL, выявленные из человеческого образца, как описано в Примере 2.2 и показано на Фиг.28-36, анализировали для разделения популяций "необученных" антигеном В-клеток: незрелых В-клеток, новых эмигрирующих В-клеток и зрелых "наивных" В-клеток от популяций "обученных" антигеном В-клеток. Ранжирование пар генов эмбрионального типа VH/VL, представляющих иммунный репертуар интактного человека, показано в Таблице 19.
Таблица 19
N=1345
Пример 3: Определение частоты использования генов эмбрионального типа VH и VL
Обзор Таблиц 18-19 из Примеров 2.3-4 показывает, что небольшое число генов эмбрионального типа VH, Vк и VA являются доминантным в иммунном репертуаре человека и в иммунном репертуаре интактного человека по сравнению с общим числом генов эмбрионального типа. Wildt с соавт. На с.895-896 также описал это явление. Wildt с соавт. также описал, что часто экспрессирующиеся генные сегменты тяжелой и легкой цепи часто спариваются, и наблюдал, что половина тестируемых спариваний соответствовала только пяти парам генов эмбрионального типа VH/VL.
Кроме того, объединенные данные оценивали для идентификации генов эмбрионального типа VH, Vк и VA, которые независимо высоко экспрессируются в иммунном репертуаре человека. Поэтому данные о парах генов эмбрионального типа VH/VL, идентифицированных из общедоступной литературы, как описано в Примере 2.1 и показано на Фиг.4-26; парах генов эмбрионального типа VH/VL, идентифицированных из человеческого образца, как описано в Примере 2.2 и показано на Фиг.28-36, и дополнительные литературные ссылки, которые включали экспрессию неспаренных генов эмбрионального типа VH и/или VL (см. Brezinschek H.P. et al. (1997) J. Clin. Invest. 99, 2488, Demaison С et al. (1995) Immunogenetics 42, 342 и Foster S.J. et al. (1997) J. Clin. Invest. 99, 1614, эти статьи включены посредством ссылки во всей своей полноте) объединяли и ранжировали для того, чтобы определить, какие гены эмбрионального типа VH, Vк и Vλ экспрессировались в иммунном репертуаре человека в наибольшем количестве. В Таблице 20 показано ранжирование по распространенности генов эмбрионального типа VH, Vλ и Vк.
Таблица 20: Частота использования неспаренных генов эмбрионального типа VH, Vк и Vλ в иммунном репертуаре человека
При сравнении Таблицы 20, показывающей распространенность несвязанных генов эмбрионального типа VH, Vλ и Vк, и Таблиц 18-19, показывающих распространенность связанных пар генов эмбрионального типа VH/VL в иммунном репертуаре человека и иммунном репертуаре интактного человека, стало очевидно, что многие из генов эмбрионального типа VH, Vλ и Vк, которые в значительной степени представлены, при оценке независимо от связи или спаривания, были также в значительной степени представлены при оценке спариваний VH/VL.
Это наблюдение подтверждено графиками, представленными на Фиг.39-40, которые показывают пары генов эмбрионального типа VH/VL иммунного репертуара человека, и Фиг.41-42, которые показывают пары генов эмбрионального типа VH/VL иммунного репертуара интактного человека. На этих графических материалах показано фактическое число каждой пары генов эмбрионального типа VH/VL, идентифицированной из объединенных данных, нанесенных на матрицу, где ось Y представляет ранжирование генов эмбрионального типа VH, и ось Х представляет ранжирование генов эмбрионального типа VL.
Пример 4: Селекция спариваний генов эмбрионального типа VH/VL для дальнейшей оценки биофизических свойств
На следующей стадии нужно было определить, какие пары белков эмбрионального типа следует тестировать, поскольку имеется примерно 2500 пар в иммунном репертуаре человека, и цель авторов изобретения заключалась в идентификации того, какие пары белков эмбрионального типа имеют предпочтительные биофизические свойства, которые могли бы помочь при селекции и разработке. Один способ заключается в тестировании пар белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые встречаются наиболее часто в иммунном репертуаре человека, например, см. Таблицу 18. Можно, например, выбрать верхние четыреста пар для тестирования или выбрать пары генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, экспрессирующиеся выше определенной пороговой концентрации. Этот подход обычно требует синтеза и тестирования большого числа последовательностей пар белков эмбрионального типа вариабельных областей тяжелой цепи и вариабельных областей легкой цепи; поэтому такой подход не может быть очень эффективным.
В качестве альтернативного подхода, авторы изобретения выбрали субпопуляцию пар эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые представляют, точно воспроизводят или охватывают большую часть преимущественно экспрессирующихся пар из иммунного репертуара человека. Этот подход основан частично на приведенном выше наблюдении, что небольшое число генов эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи преобладает в иммунном репертуаре человека. Поэтому небольшое число преимущественно экспрессирующихся генов эмбрионального типа (неспаренных) тяжелой и легкой цепи можно комбинировать для создания группы пар, которые являются типичными для иммунного репертуара человека.
Этот подход осуществляют следующим образом. В Примере 3 определяли экспрессию генов эмбрионального типа вариабельной области тяжелой цепи, вариабельной области к-легкой цепи и вариабельной области λ-легкой цепи. На следующей стадии выполняли анализ in silico значимых генов эмбрионального типа VH, Vλ и Vк, где оценивали по меньшей мере следующие факторы: длину CDR, изоэлектрическую точку (р!) (предпочтительная изоэлектрическая точка составляет 7,5 или выше, поскольку она должна обеспечивать стабильность в стандартном буфере для приготовления препаратов при рН от 5,5 до 7), посттрансляционные модификации (РТМ) (конкретно, М-связанные сайты гликозилирования (N×S или NxT) или химические модификации, такие как отщепление Asp (часто в DP), изомеризация Asp (DD, DG), дезамидирование (NS, NG), которые могут происходить in vivo (в сыворотке) или при хранении в буфере для приготовления препаратов и приводят к потере связывания антител), присутствие метионинов в CDRs (могут окисляться при воздействии растворителя), присутствие неспаренных цистеинов (будут образовывать дисульфидные связи с любым другим неспаренным цистеином, таким образом приводя к сшиванию белков и/или более низким уровням экспрессии), отклонения от эмбрионального типа, присутствие возможных Т-клеточных эпитопов и теоретическую склонность к агрегации. Выбранные данные из анализа in silico показаны на Фиг.37-38.
На основании анализа in silico наиболее распространенных генов эмбрионального типа VH, Vλ и Vк, выбрали их субпопуляцию для синтеза, комбинирования и последующего функционального тестирования, эта субпопуляция показана на Фиг.37-38. Как показано, не все наиболее распространенные гены эмбрионального типа VH, Vλ и Vк были выбраны для дальнейшего тестирования. Из наиболее распространенных генов эмбрионального типа VH, как показано в Таблице 20, не были выбраны IGHV4-34, IGHV4-59 и IGHV3-9. Вместо этого, как показано на Фиг.37-38 и в Таблице 21, выбрали IGHV3-74, IGHV3-73 и IGHV6-1. В целом, выбрали 20 генов эмбрионального типа VH. Из наиболее распространенных генов эмбрионального типа Vк, как показано в Таблице 20, не были выбраны IGKV4-1, IGKV2-28/2D-28, IGKV1-33/1D-33 и IGKV1-8. В целом, выбрали 12 генов эмбрионального типа Vк. Из наиболее распространенных генов эмбрионального типа VH, как показано в Таблице 20, не был выбран IGLV1-44. В целом, выбрали 8 генов эмбрионального типа Vλ.
В Таблице 21 снова показано ранжирование частоты использования генов эмбрионального типа VH, Vк и Vλ из иммунного репертуара человека и жирный шрифт и подчеркивание показывает гены эмбрионального типа, которые выбраны для дальнейшего функционального тестирования.

Пример 4.1: Рекомбинация часто встречающихся генов эмбрионального типа VH. Vк и Vλ для получения представления о наиболее распространенных парах VH/VL в иммунном репертуаре человека
Как обсуждалось выше и показано в Таблице 21 и на Фиг.39-40, 41-42, выбраны 20 VH, 12 Vк и 8 Vλ, когда при комбинировании точно воспроизводят или охватывают большую часть преимущественно экспрессирующихся пар генов эмбрионального типа VH/VL из иммунного репертуара человека и иммунного репертуара интактного человека.
В качестве следующей стадии выбранные 20 VH, 12 Vк и 8 Vλ генов эмбрионального типа VH, Vк и Vλ синтезировали и комбинировали для получения 400 пар генов эмбрионального типа VH/VL, которые точно воспроизводят или охватывают большую часть преимущественно экспрессирующихся пар генов эмбрионального типа VH/VL в иммунном репертуаре человека. Затем 400 пар генов эмбрионального типа VH/VL тестировали в отношении их биофизических свойств.
В Таблице 22 показаны выбранные гены эмбрионального типа VH, Vк и Vλ, подлежащие комбинированию для получения 400 пар генов эмбрионального типа VH/VL.

Для того чтобы показать, что 400 пар генов эмбрионального типа VH/VL, образованных для функционального тестирования, фактически точно воспроизводят или охватывают большую часть преимущественно экспрессирующихся пар генов эмбрионального типа VH/VL в иммунном репертуаре человека, Таблицу 18 воспроизводят ниже в видеТаблицы 23, где 400 пар VH/VL, которые были протестированы, выделены жирным шрифтом и подчеркнуты.















Пример 4.2
Как обсуждалось в Примере 2.4, важно различать "наивные", "необученные" антигеном и "обученные" антигеном популяции В-клеток. Поэтому Таблица 19, в которой показано ранжирование пар генов эмбрионального типа VH/VL, идентифицированных в иммунном репертуаре интактного человека, воспроизводят в виде Таблицы 24, где 400 пар генов эмбрионального типа VH/VL, которые синтезировали и комбинировали для дальнейшего функционального тестирования, выделены жирным шрифтом и подчеркнуты.


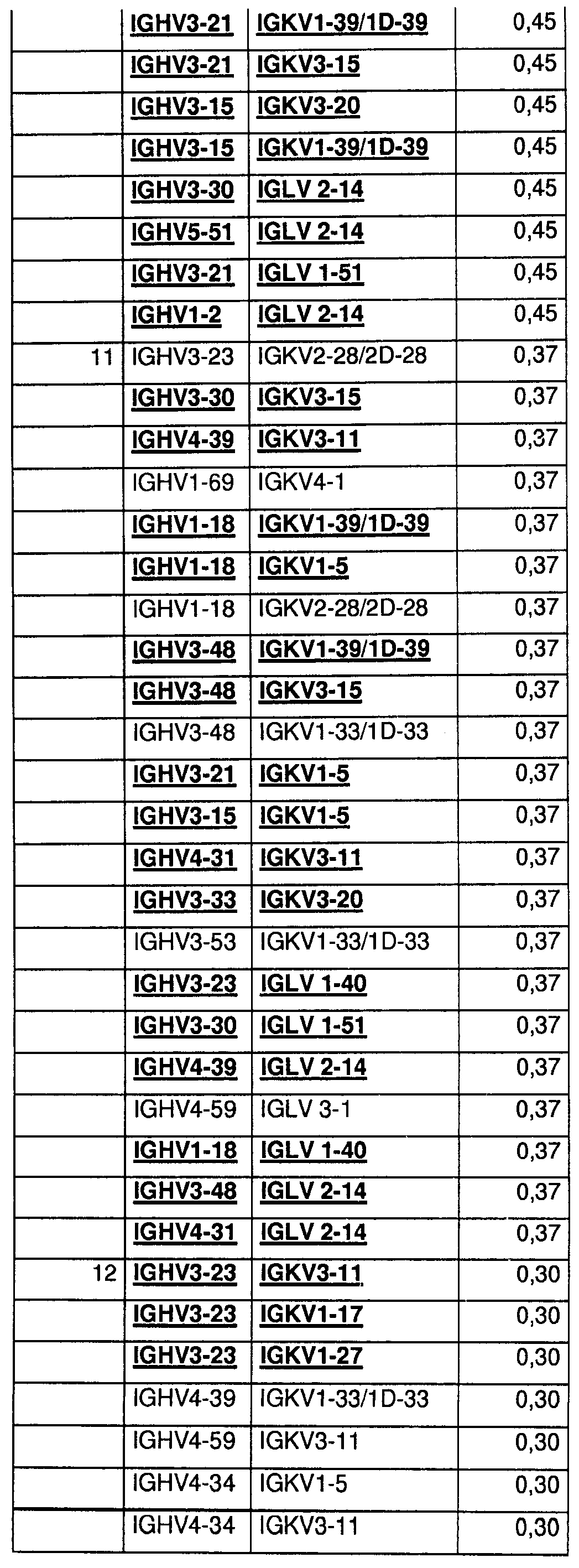
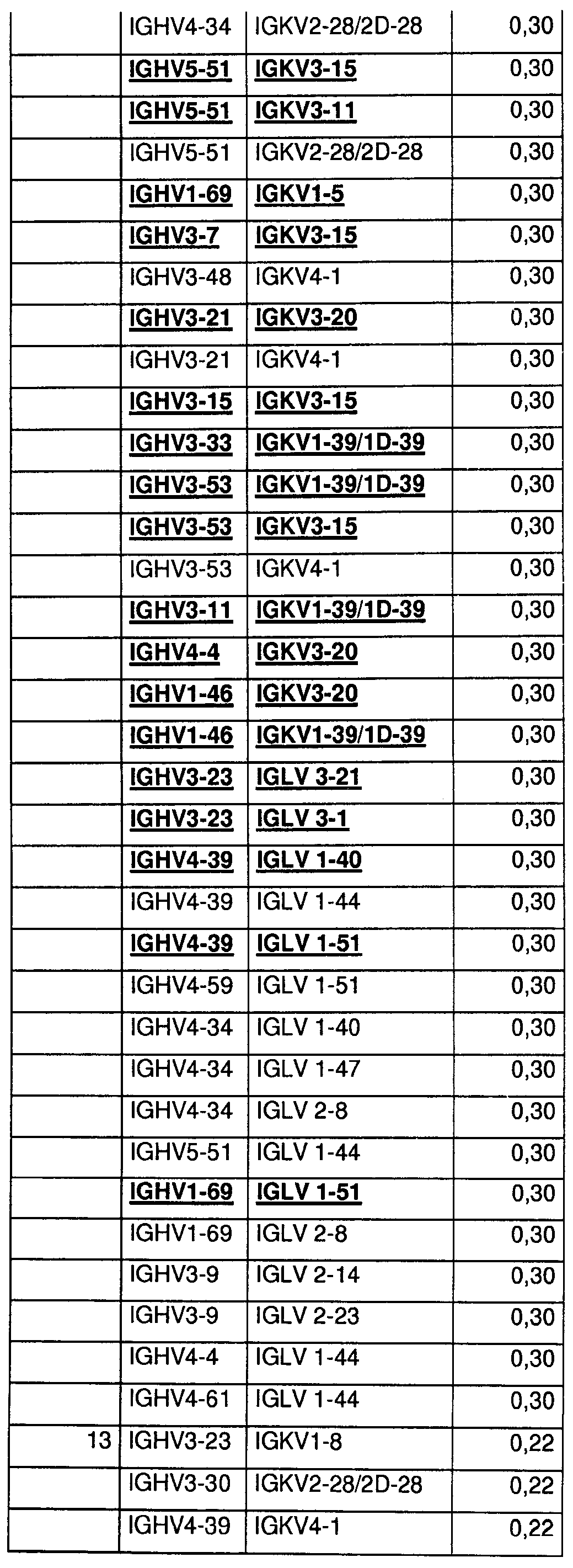




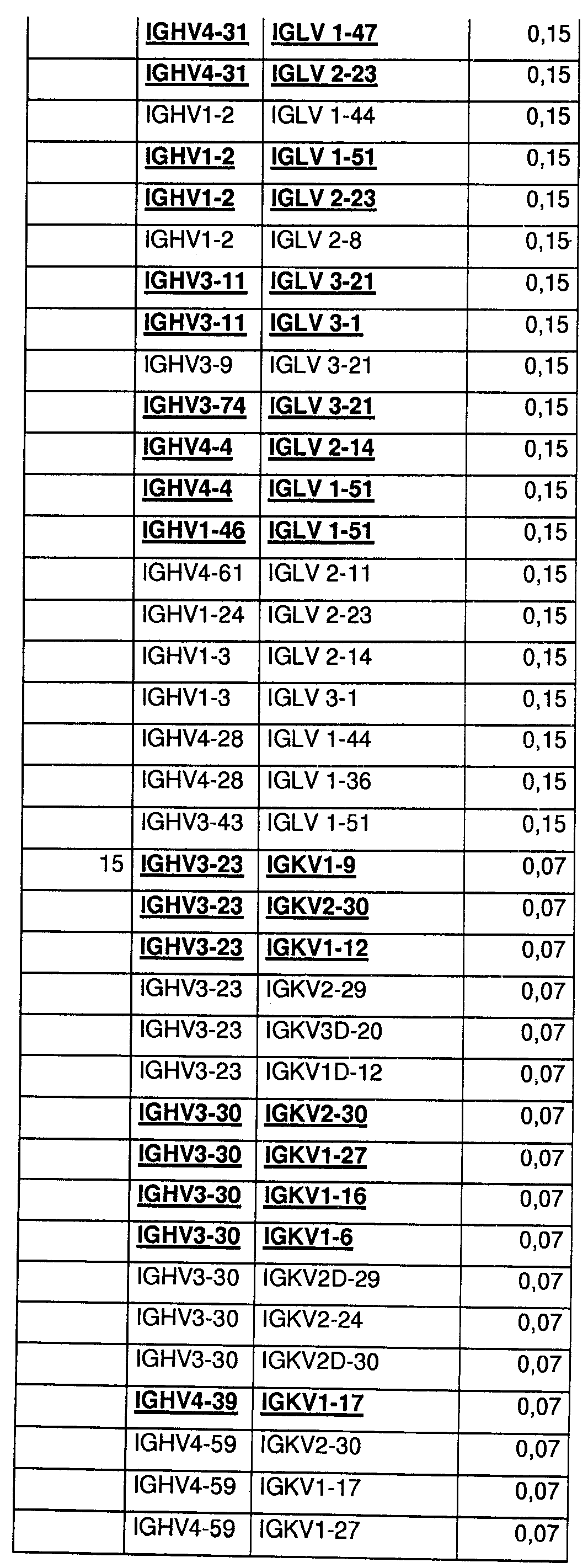



Пример 5: Получение генов эмбрионального типа для функционального анализа
В качестве следующей стадии гены эмбрионального типа VH, Vλ и Vк, выбранные для комбинирования и последующего тестирования, как показано в Таблице 25, послали в Geneart (Regensburg, Германия) для оптимизации кодонов, соответствующей экспрессии в Е.соli (нейтральной для экспрессии в млекопитающих) и синтеза.
Последовательности генов эмбрионального типа каждого из генов эмбрионального типа VH, Vλ и Vк показаны на Фиг.45-47. Каждую последовательность генов эмбрионального типа синтезировали для того, чтобы включить следующее:
а) для VH: лидерная последовательность (модифицированная сигнальная последовательность phoA, включающая сайт рестрикции Nhel, как показано на Фиг.3); FR1, CDR1, FR2, CDR2 и FR3 эмбрионального типа (включающие сайт рестрикции BssHII, как показано на Фиг.3); CDR-H3 (WGGDGFYAMDY) из антитела 4D5, как использовано в Ewert S. et al., J. Mol. Biol. (2003) 325, 531-553; и JH4 FR4 (включающий RE сайт Xhol/Sall, как показано на Фиг.3);
б) для Vk: лидерная последовательность (модифицированная сигнальная последовательность оmрА, включающая сайт рестрикции Ndel, как показано на Фиг.3); FR1, CDR1, FR2, CDR2 и FR3 эмбрионального типа (включающие сайт рестрикции Bbsl, как показано на Фиг.3), каппа-подобный CDR-L3 (QQHYTTPPT) согласно Ewert S. et al„ J. Mol. Biol. (2003) 325, 531-553; и Jk1 FR4 (включающий RE сайт Kpnl, как показано на Фиг.3);
в) для Vλ: лидерная последовательность (модифицированная сигнальная последовательность оmрА, включающая сайт рестрикции Ndel, как показано на Фиг.3); FR1, CDR1, FR2, CDR2 и FR3 эмбрионального типа (включающие сайт рестрикции Bbsl, как показано на Фиг.3), лямбда-подобный CDR-L3 (QSYDSSLSGV5) согласно Ewert S. et al„ J. Moi. Biol. (2003) 325, 531-553; и JI2/3 FR4 (включающий RE сайт Kpnl, как показано на Фиг.3).
Пример 6: Функциональное тестирование пар генов эмбрионального типа VH/VL. типичных для иммунного репертуара человека
Затем 400 пар генов эмбрионального типа VH/VL тестировали в отношении следующих свойств: а) относительный уровень экспонирования после продукции фага и фагового ИФА в Fab-формате; б) относительные уровни экспрессии Fab после продукции Fab в Е.соli, лизис клеток Е.соli и ИФА-детекция продуцируемого Fab; в) термостабильность Fab после продукции Fab в Е.соli, лизис клеток £. со//' и ИФА-детекция неденатурированного Fab после инкубации при повышенных температурах; г) стабильность Fab из лизатов Е.соli в сыворотке крупного рогатого скота/мышей посредством ИФА-детекции неденатурированного Fab после инкубации в сыворотке крупного рогатого скота/мышей; д) относительные уровни экспрессии человеческого IgG1 после продукции IgG1 в клетках млекопитающих и ИФА-детекции секретируемого IgG1 из супернатантов клеточных культур; и е) стабильность человеческого IgG1 в сыворотке крупного рогатого скота, как определено посредством ИФА-детекции неденатурированного Fab после инкубации в сыворотке крупного рогатого скота/мышей.
Пример 6.1: Создание пула Fab. экспонируемого на фаге. для функциональной характеристики
Антитело или фрагменты антитела, синтезированные в Примере 5, показанные в Таблице 25, клонировали в трицистронный Fab дисплейный вектор pJPcH (Фиг.48) для функционального тестирования. Создавали пулы Fab, которые содержали комбинации каждого из основных генов, 20 VH, комбинированных с 8 Vλ и 12 Vк, которые давали 400 комбинаций, которые представляли подавляющее большинство из наиболее распространенных пар генов эмбрионального типа VH/VL из иммунного репертуара человека, как
показано в Таблице 18 и на Фиг.39-40.
Фаг, содержащий вышеуказанные пары генов, продуцировали в небольшом масштабе, используя 96-луночные планшеты. Эталонный планшет готовили, наполняя каждую из лунок 2х средой YT/CAM/TET/Gluc и внося клоны из 400 комбинаций VH/VL, где pMORPH30_Vk3-11_AQA / VH3-23_TKA или pMORPH30_Vk3-11_AYA / VH3-23_VLA (pMORPH30 показана на Фиг.51) использовали в качестве контроля. Планшеты инкубировали в течение ночи при 37°С при встряхивании. Эталонные планшеты хранили при конечной концентрации глицерина 15% и замораживали при -80°С.
Дополнительные 96-луночные планшеты готовили для продукции фагов, используя 2×YT/CAM/TET/Gluc в качестве среды и внося клоны из эталонных планшетов, описанных выше. Планшеты инкубировали при 37°С в течение примерно 2-4 ч при встряхивании при 400 об/мин до достижения ООбООнм
примерно 0,5.
Планшеты инфицировали 5 мкл вспомогательного фага на лунку (НурегрПаде; PROGEN; 1×1012 БОЕ/мл (бляшкообразующая единица). Планшеты инкубировали при 37°С в течение 45 мин без встряхивания и затем в течение 60 мин при встряхивании при 400 об/мин. Бактерии собирали центрифугированием при 2200д в течение 5 мин при 4°С.
Супернатанты, содержащие вспомогательный фаг, отбрасывали и инфицированные осадки Е.соli ресуспендировали с помощью 2xYT/Cam/TET/Kan/IPTG без глюкозы. Ресуспендированные осадки переносили в новый 96-луночный планшет с глубокими лунками, предварительно заполненный 2×YT/Cm/TET/Kan/IPTG. Планшеты инкубировали в течение ночи при 22°С, при встряхивании. Фаг-содержащие супернатанты собирали центрифугированием и отбрасывали клетки E.соli и дебрис.
Пример 6.2: Оценка ранжирования фагового дисплея Fab с использованием ИФА
Фаговые супернатанты, приготовленные, как описано в Примере 6,1, использовали для ранжирования фагового дисплея Fab с помощью фагового ИФА.
Дисплей Fab-фрагментов оценивали в фаговом ИФА с использованием двух различных захватывающих (capture) антител:
(1) Анти-М13 антитело (Amersham #27-9420-01) использовали для захвата фаговых частиц с помощью главного оболочечного белка д8р; поэтому можно определить титр фага.
(2) Использовали анти-Fd антитело (Binding Site #PC075), которое связывается с экспонируемым Fab; поэтому захватываются только фаги, экспонирующие Fabs, содержащие основные гены.
Соответствующие захватывающие антитела иммобилизовали на черных 96-луночных планшетах Maxisorp™ путем распределения 100 мкл раствора антител в концентрации 7,5 мкг/мл для анти-М13 антитела и в концентрации 1,0 мкг/мл для анти-Fd антитела в различные лунки, запечатывания планшета ламинированной фольгой и инкубирования в течение ночи при 4°С.На следующие сутки планшеты дважды промывали TBST и каждую лунку блокировали 300 мкл CTBST в течение 1 ч при комнатной температуре.
Оба фаговых супернатанта и эталонные образцы переносили для детекции следующим образом. Блокированные ИФА планшеты промывали дважды TBST. 100 мкл соответствующим образом разведенных фаговых супернатантов в CTBST переносили из планшетов для разведения в покрытые планшеты для ИФА, инкубировали в течение 1-2 ч при комнатной температуре и промывали 5х TBST. Добавляли 100 мкл/лунку конъюгата анти-М13 с пероксидазой (Amersham), разведенного 1:5000 в CTBST, и инкубировали в течение 1-2 ч при комнатной температуре. Рабочий раствор Quanta Blu (Pierce) готовили путем смешивания 1 части (например 0,5 мл) раствора перекиси с 9 частями (например 4,5 мл) раствора субстрата и уравновешивали при комнатной температуре в течение по меньшей мере 30 мин. ИФА планшеты промывали 5х TBST, добавляли 100 мкл/лунку рабочего раствора QuantaBlu. Флуоресценцию измеряли после инкубации в течение примерно 2 мин (возбуждение: 320 нм, излучение: 430 нм) и далее с интервалами 5 мин.
Оценку данных ИФА выполняли следующим образом: калибровочные кривые получали с использованием препарата эталонного фага HuCAL GOLD (VH3 каппа+лямбда) и рассчитывали титры фаговых супернатантов и контролей. Для каждого образца титр по анти-Fd делили на титр по анти-М13 (анти-pVIII), полученное соотношение представляло собой относительный уровень экспонирования (relative display rate).
Поскольку относительный уровень экспонирования Fab рассчитывали с использованием внутреннего стандарта (HuCAL GOLD препарат фага VH3 каппа+лямбда), который не является общедоступным. Относительный уровень экспонирования оценивали в виде ранжирования. Посредством ранжирования относительных значений экспонирования специалист может воспроизводить вышеупомянутый способ, используя любой контроль. Например, каждая пара белков эмбрионального типа демонстрирует количество по сравнению с контролем. Поэтому пары белков эмбрионального типа, имеющие наиболее высокий относительный уровень экспонирования по сравнению с контролем, также имеют наиболее высокий относительный уровень экспонирования по сравнению с любым контролем, несмотря на то, что конкретный относительный уровень экспонирования, по-видимому, будет отличаться. Поэтому ранжирование значений создавали с использованием относительных данных экспонирования, показаных на Фиг.55, также показанных в Таблице 32, где данные располагались от самых высоких до самых низких значений. Это ранжирование показано в Таблице 26. Из этих пар белков эмбрионального типа, имеющих относительный уровень экспонирования в пределах верхних 10%, 20%, 30%, 45%, 50%, 55%, 60%, 70%, 75%, 80% и 90%, можно идентифицировать тестируемые Fabs.
Конкретно, из 400 протестированных пар относительные значения экспонирования были получены для 196 пар, см. Таблицу 26. Поэтому специалист может точно определить, какие пары белков эмбрионального типа находятся в пределах верхних 10%, 20%, 30%, 45%, 50%, 55%, 60%, 70%, 75%, 80% или 90% тестируемых Fabs. Например, верхние 75% включают пары белков эмбрионального типа, ранжированные как №№1-147 в Таблице 26.



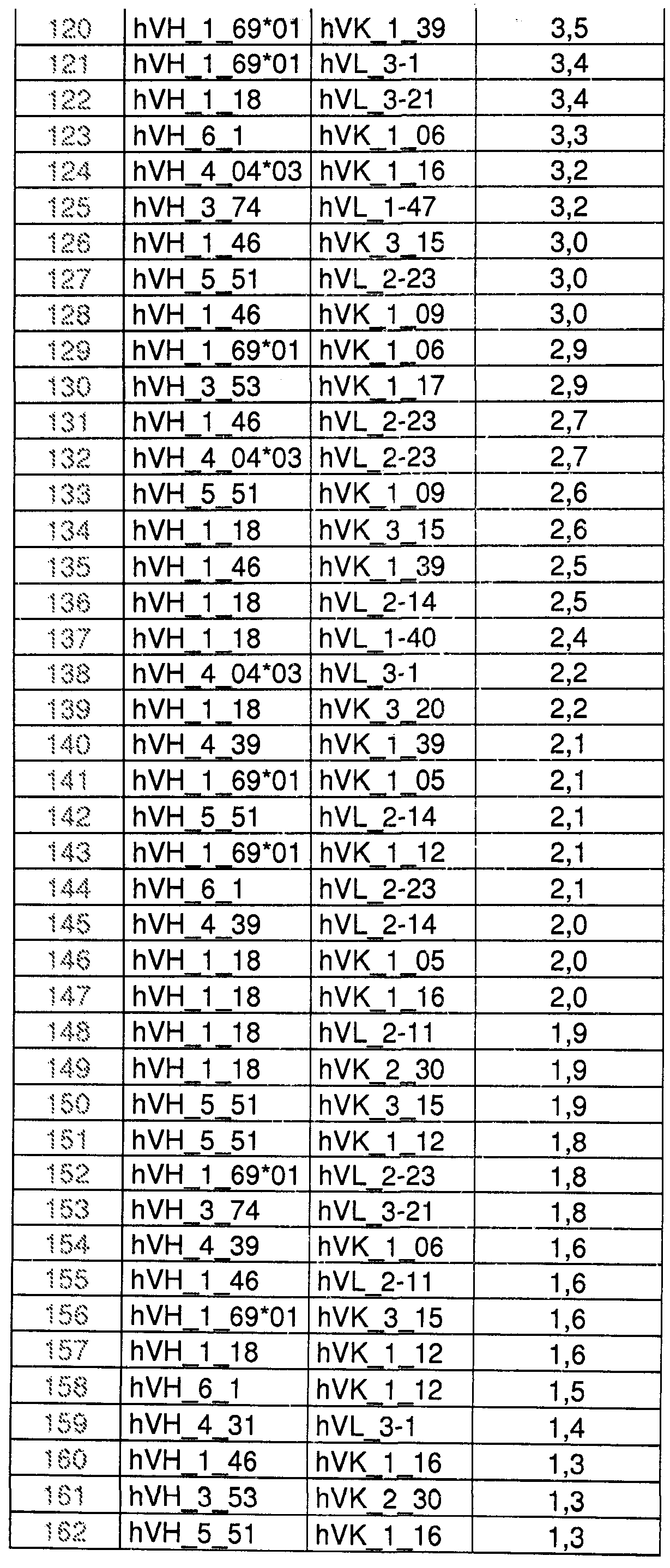

На Фиг.55 показаны относительные уровни дисплея для большинства 400 пар генов эмбрионального типа VH/VL.
Пример 6.3: ИФА скрининг 400 комбинаций VH/VL для определения уровня экспрессии Fab в лизатах Е.соli
Эталонные планшеты (МР, Masterplates) инокулировали выбранными клонами, трансформированными пулами комбинаций VH/VL в Fab-экспрессирующем векторе pJP×1 (показаны на Фиг.49) в 2YT/Cam/1%Gluc среде на лунку. Эти планшеты инкубировали при 37°С в течение ночи при встряхивании. Планшеты для экспрессии (ЕР) инокулировали с помощью 2,5 мкл культур из MPs в 2YT/Cam/0,1%глюкозы на лунку. Контроли (см. Таблицу 27) инокулировали из глицериновых концентратов. Эти планшеты инкубировали в течение 6 часов при 37°С при встряхивании, затем экспрессию Fab индуцировали путем добавления IPTG (изопропилтиогалактозид) и инкубировали при 22°С в течение ночи при встряхивании. Лизаты клеток E.coli получали путем добавления буфера борная кислота/ЕОТА/лизоцим к EPs (1 ч инкубации при 22°С, встряхивание) и бактериальные лизаты затем блокировали с помощью 12,5% MPBST, встряхивая по меньшей мере в течение 30 мин при комнатной температуре. Лизаты Е.соli из планшетов для экспрессии разбавляли соответственно в 0,5% MPBS и использовали в следующем
анализе.
В Таблице 27 показаны используемые немеченые покрывающие (coating) антитела и АР-меченые детектирующие антитела.
В Таблице 28 описаны используемые контроли.
Таблица 28:
Скрининг ИФА включал следующие стадии: Покрытие 384 лунок планшета MaxiSorp с помощью Fd-специфических антител против IgG человека, разведенных в PBS, и инкубацию в течение ночи при 4°С. На следующие сутки планшеты промывали 2×PBST и блокировали путем добавления (5% сухого молока в PBS) в каждую лунку и инкубировали в течение 1-2 ч при к.т. при встряхивании. Затем планшеты снова промывали PBST и добавляли предварительно блокированные лизаты Е.соli, разбавляя в 0,5% MPBS, и инкубировали в течение 1 ч при встряхивании при к.т. Также добавляли контроли #3 и #5. Планшеты затем промывали PBST, и АР-меченое детектирующее антитело разбавляли в 0,5% MPBS. Добавляли разведенное детектирующее антитело и затем инкубировали в течение 1 ч при к.т.при осторожном встряхивании. Сигнал определяли следующим образом:
промывали лунки TBST и добавляли 20 мкл AttoPhos (разведенный 1:5 в ddHzO) и считывали при 5 мин и 7-8 мин с использованием Tecan (infiniTe F200), программа PrimeScreen.
Относительные уровни экспрессии Fab рассчитывали путем деления ИФА сигнала соответствующей пары VH/VL на ИФА сигнал эталонного Fab pMx11FH VH1-69 VLA_VI1-40 AYA. Таким образом, одинаково высокие ИФА сигналы дают относительный уровень экспрессии Fab, равный 1. Эталонный Fab экспрессируется в плазмидах pMORPHXH (показаны на Фиг.50), содержащих: а) модифицированную сигнальную последовательность тяжелой цепи phoA, содержащую С-концевой сайт рестрикции Nhel; б) модифицированную сигнальную последовательность легкой цепи оmрА, содержащую С-концевой сайт рестрикции Nhel; в) белковые последовательности эмбрионального типа вариабельной области тяжелой цепи гена эмбрионального типа VH1-69* 01, как показано на Фиг.45А, г) белковые последовательности эмбрионального типа вариабельной области легкой цепи гена эмбрионального типа IGLV1-40, как показано на Фиг.47А; д) включение CDR-H3 (WGGDGFYAMDY) из антитела hu4D5-8, и белковой последовательности эмбрионального типа JH4 для FR4 тяжелой цепи; е) включение участка CDR-L3 (QSYDSSLSGV5) и белковой последовательности эмбрионального типа JI2/3 для FR4 легкой цепи. hu4D5-8 описан в Carter P. et al. (1992) "Humanization of an anti-pi 85Her2 antibody for human cancer therapy" Proc. Natl. Acad. Sci. USA 89, 4285-4289). Все гены созданы в Geneart (Regensburg, Германия).
Результаты показаны на Фиг.56.
Пример 6.4: ИФА скрининг 400 комбинаций VH/VL для определения термостабильности Fab в BEL лизатах
Планшеты для экспрессии готовили, как в Примере 6.3. Разведенные лизаты Е.соli из планшетов для экспрессии инкубировали при различных температурах в течение 45 минут и использовали в следующем анализе.
В Таблице 29 показаны используемые немеченые покрывающие антитела и АР-меченые детектирующее антитела.
ИФА скрининг включал следующие стадии: на 384 лунок планшета MaxiSorp наносили покрывающие антитела (см. таблицу выше), разведенные в PBS. Планшеты инкубировали в течение ночи при 4°С.На следующей день планшеты промывали PBST и блокировали путем добавления 5% MPBS в каждую лунку и инкубировали в течение 1-2 ч при к.т.при встряхивании. Затем разведенные лизаты Е.соli из планшетов для экспрессии распределяли в четыре 96-луночных ПЦР-планшета (каждый примерно 40 мкл) и подвергали воздействию различных температур (4°С (на льду), 60°С, 70°С, 80°С и затем на льду) в ПЦР-циклере, каждая температура в течение 45 мин. Блокированные 384-луночные планшеты промывали PBST, затем в планшеты добавляли предварительно инкубированные Fab лизаты. Планшеты затем инкубировали 1 ч при к.т.при встряхивании. Планшеты промывали PBST, АР-меченые детектирующие антитела разбавляли в 0,5% MPBS. Добавляли 20 мкл/лунку разведенных детектирующих антител и инкубировали в течение 1 ч при к.т.при осторожном встряхивании. Сигнал определяли следующим образом:
промывали лунки TBST и во все лунки добавляли AttoPhos (разведенный 1:5 в ddH2O). Сигнал считывали в различные временные точки (от 5 мин до 10 мин), используя Tecan (infiniTe F200), программу PimeScreen.
Результаты показаны на Фиг.57.
Пример 6.5: ИФА скрининг 400 комбинаций VH/VL для определения сывороточной стабильности Fab в лизатах Е.соli
Планшеты для экспрессии готовили, как в Примере 6.3. Лизаты Е.соli, содержащие Fab, разбавляли и инкубировали в сыворотке крупного рогатого скота и мышей с использованием следующих стадий: лизаты Е.соli из планшетов для экспрессии разбавляли в 50% сыворотке (общий объем 100 мкл), добавляли 1:1000 Cam для предотвращения роста бактерий и лизаты разделяли на два 96-луночных планшета и оба планшета замораживали. Первый планшет размораживали и инкубировали при 37°С в течение 12-13 суток. Второй планшет хранили при -80°С до проведения ИФА (0 суток инкубации при 37°С).
В Таблице 30 показаны используемые немеченые покрывающие антитела и АР-меченые детектирующие антитела.
На 11-е или 12-е сутки на 384 лунок планшета MaxiSorp наносили 20 мкл покрывающего антитела, разведенного в PBS. Планшеты инкубировали в течение ночи при 4°С.На следующие сутки планшеты промывали PBST и блокировали путем добавления 5% MPBS в каждую лунку и инкубировали в течение 1-2 ч при к.т.при встряхивании. Затем блокированные 384-луночные планшеты промывали PBST. Лизаты Е.соli в сыворотке из образцов при -80°С и 37°С переносили в покрытые ИФА планшеты и инкубировали в течение 1 часа при к.т. при встряхивании. Планшеты промывали PBST и АР-меченые детектирующие антитела разбавляли в 0,5% MPBS. Добавляли АР-меченые детектирующие антитела и планшет инкубировали в течение 1 ч при к.т. при встряхивании. Сигнал идентифицировали следующим образом: промывали лунки TBST и во все лунки добавляли AttoPhos (разведенный 1:5 в ddHzO). Сигнал считывали в различные временные точки (от 5 мин до 10 мин), используя Tecan (infiniTe F200), программу PrimeScreen.
Результаты тестирования стабильности в сыворотке крупного рогатого скота показаны на Фиг.58. Результаты тестирования стабильности в мышиной сыворотке показаны на Фиг.59.
Пример 7: Создание iqg для оценки биофизических свойств
Для создания 400 пар генов эмбрионального типа VH/VL, 20 генов вариабельной области тяжелой цепи субклонировали в вектор экспрессии человеческого IgG1 pJP_hIgG1, показанный на Фиг.52. Одновременно 12 генов вариабельной области каппа-цепи субклонировали в вектор экспрессии каппа-легкой цепи млекопитающих pJP_hlgkappa, показанный на Фиг.53, и 8 генов вариабельной области лямбда-цепи субклонировали в вектор экспрессии лямбда-легкой цепи млекопитающих pJPJ-ilglambda, показанный на Фиг.54.
Путем котрансфекции каждой экспрессирующей плазмиды для тяжелой цепи и легкой цепи все 400 пар VH/VL могут быть получены отдельно посредством клонирования только 40 экспрессирующих конструкций. Таким образом, все 20 конструкций тяжелой цепи котрансфицировали с каждой из экспрессирующих конструкций для легкой цепи в клетки HEK.EBNA. Человеческий IgG1 собирали или определяли через несколько суток после трансфекции из супернатантов клеточных культур.
Пример 7.1: Ранжирование экспрессии IgG
Одним из критереев для выбора спариваний VH/VL, подлежащих включению в библиотеку, является уровень экспрессии 400 различных спариваний VH/VL в IgG-формате. Уровень экспрессии каждого спаривания VH/VL в формате человеческого IgG1 оценивали с помощью "сэндвич"-ИФА. Поэтому все 400 комбинаций VH/VL в формате человеческого IgG1 трансфицировали в клетки HEK.EBNA и экспрессировали в небольшом масштабе. Супернатанты клеточной культуры собирали через несколько суток и оценивали уровень IgG.
Выполняли следующую процедуру. 384-луночные планшеты MaxiSorpTM покрывали Fcγ-пан R10Z8E9 мышиным антителом против человеческого IgG при 2,5 мкг/мл в PBS. Планшеты инкубировали в течение ночи при 4°С.Планшеты промывали PBST. Планшеты блокировали 5% БСА (бычий сывороточный альбумин) или 1х Chemiblocker в PBST и инкубировали в течение 1 ч при к.т. при встряхивании и снова промывали PBST. Супернатанты, экспрессирующие IgG, разбавляли в 2,5% БСА-PBST, и разбавленные образцы добавляли к блокированному и промытому ИФА планшету. Использовали следующие контроли: пустой супернатант и супернатанты со слабо экспрессирующими антителами, умеренно экспрессирующими антителами и высоко экспрессирующими антителами. Планшеты инкубировали в течение 2 ч при комнатной температуре при встряхивании. Планшеты затем промывали TBST. Добавляли соответственно разведенный Fcγ-пан R10Z8E9 конъюгат мышиного антитела против человеческого IgG с биотином в 1% БСА-TBST. Планшеты инкубировали в течение 1 ч при при комнатной температуре. Планшеты промывали TBST. Добавляли стрептавидин-АР, разведенный 1:2000 в 0,5% БСА-TBST, и планшеты инкубировали в течение 1 ч при при комнатной температуре при встряхивании. Планшеты промывали TBST. Добавляли флуоресцентный субстрат AttoPhosTM (приготовленный согласно инструкциям производителя), разведенный в TBST непосредственно перед использованием. Через 5 и 10 мин измеряли флуоресценцию с помощью микропланшет-ридера Тесап.
Относительные уровни экспрессии IgG1 рассчитывали путем деления ИФА сигнала соответствующей пары VH/VL на ИФА сигнал эталонного IgG1 MOR03080 (показанный в Таблице 31). Таким образом, одинаково высокие ИФА сигналы дают относительный уровень экспрессии IgG1, равный 1.

Результаты показаны на Фиг.60.
Пример 7.2: Ранжирование стабильности lqG1 в сыворотке
Одним из критериев для выбора спариваний вариабельных областей тяжелых и вариабельных областей легких цепей, подлежащих включению в библиотеку, является стабильность в сыворотке 400 различных спариваний вариабельных областей тяжелых и вариабельных областей легких цепей в IgG-формате. Стабильность в сыворотке каждого супернатанта IgG антитела оценивали путем инкубации в 50% мышиной сыворотке в течение 14 суток и последующего "сэндвич"-ИФА с мышиным антителом против человеческого IgG (СН2), клон R10Z8E9. Снова все 400 комбинаций VH/VL в формате человеческого IgG1 трансфицировали в клетки HEK.EBNA и экспрессировали в небольшом масштабе. Супернатанты клеточных культур собирали через несколько суток и IgG в супернатанте тестировали на стабильность в сыворотке.
Выполняли следующую процедуру. 384-луночный планшет MaxiSorpTM покрывали Fcy-пан R10Z8E9 мышиным антителом против человеческого IgG при 2,5 мкг/мл в PBS. Планшеты инкубировали в течение ночи при 4°С.Планшеты промывали PBST и затем блокировали 5% БСА-PBST или 1х Chemiblocker в течение 1 ч при комнатной температуре при встряхивании. Планшеты промывали PBST. Супернатанты клеточной культуры, содержащие IgG1, разбавляли: а) в 2,5% БСА-PBST и б) в 50% мышиной сыворотке и инкубировали при 37°С в течение по меньшей мере14 суток, и эти образцы добавляли в блокированный и промытый ИФА планшет.Использовали следующие контроли: пустой супернатант и Супернатанты со слабо экспрессирующими антителами, умеренно экспрессирующими антителами и высоко экспрессирующими антителами. Планшеты инкубировали в течение 2 ч при комнатной температуре при встряхивании. Планшеты промывали TBST. Добавляли Fcy-пан R10Z8E9 конъюгат мышиного антитела против человеческого IgG с биотином, разведенный до 0,8 мкг/мл в 1% БСА-TBST. Планшеты инкубировали в течение 1 ч при комнатной температуре. Планшеты промывали TBST. Добавляли стрептавидин-АР, разведенный 1:2000 в 0,5% БСА-TBST. Планшеты инкубировали в течение 1 ч при комнатной температуре при встряхивании. Планшеты промывали TBST. Добавляли флуоресцентный субстрат AttoPhosTM (приготовленный согласно инструкциям производителя), разведенный 1:5 в TBST непосредственно перед использованием. Через 5 и 10 мин измеряли флуоресценцию с помощью микропланшет-ридера Тесап.
Результаты показаны на Фиг.61.
Пример 8: Выбор пар VH/VL с предпочтительными биофизическими свойствами для включения в коллекцию
После тестирования 400 пар генов эмбрионального типа VH/VL в отношении следующих свойств: а) относительный уровень экспонирования после продукции фага и фагового ИФА в Fab-формате; б) относительные уровни экспрессии Fab после продукции Fab в Е.соli, лизиса клеток E.coli и ИФА-детекции продуцируемого Fab; в) термостабильность Fab после продукции Fab в E.coli, лизиса клеток Е coli и ИФА-детекции неденатурированного Fab после инкубации при повышенных температурах; г) стабильность Fab из лизатов E.coli в сыворотке крупного рогатого скота/мышей посредством ИФА-детекции неденатурированного Fab после инкубации в сыворотке крупного рогатого скота/мышей; д) относительные уровни экспрессии человеческого IgG1 после продукции IgG1 в клетках млекопитающих и ИФА-детекции секретируемого IgG1 в супернатантах клеточных культур; и е) стабильность человеческого IgG1 в сыворотке крупного рогатого скота, как определено посредством ИФА-детекции неденатурированного Fab после инкубации в сыворотке крупного рогатого скота/мышей; затем следующая стадия заключалась в выборе пар эмбрионального типа VH/VL, подлежащих включению в коллекцию.
Результаты функционального тестирования для каждой пары белков эмбрионального типа VH/VL показаны в Таблице 32.


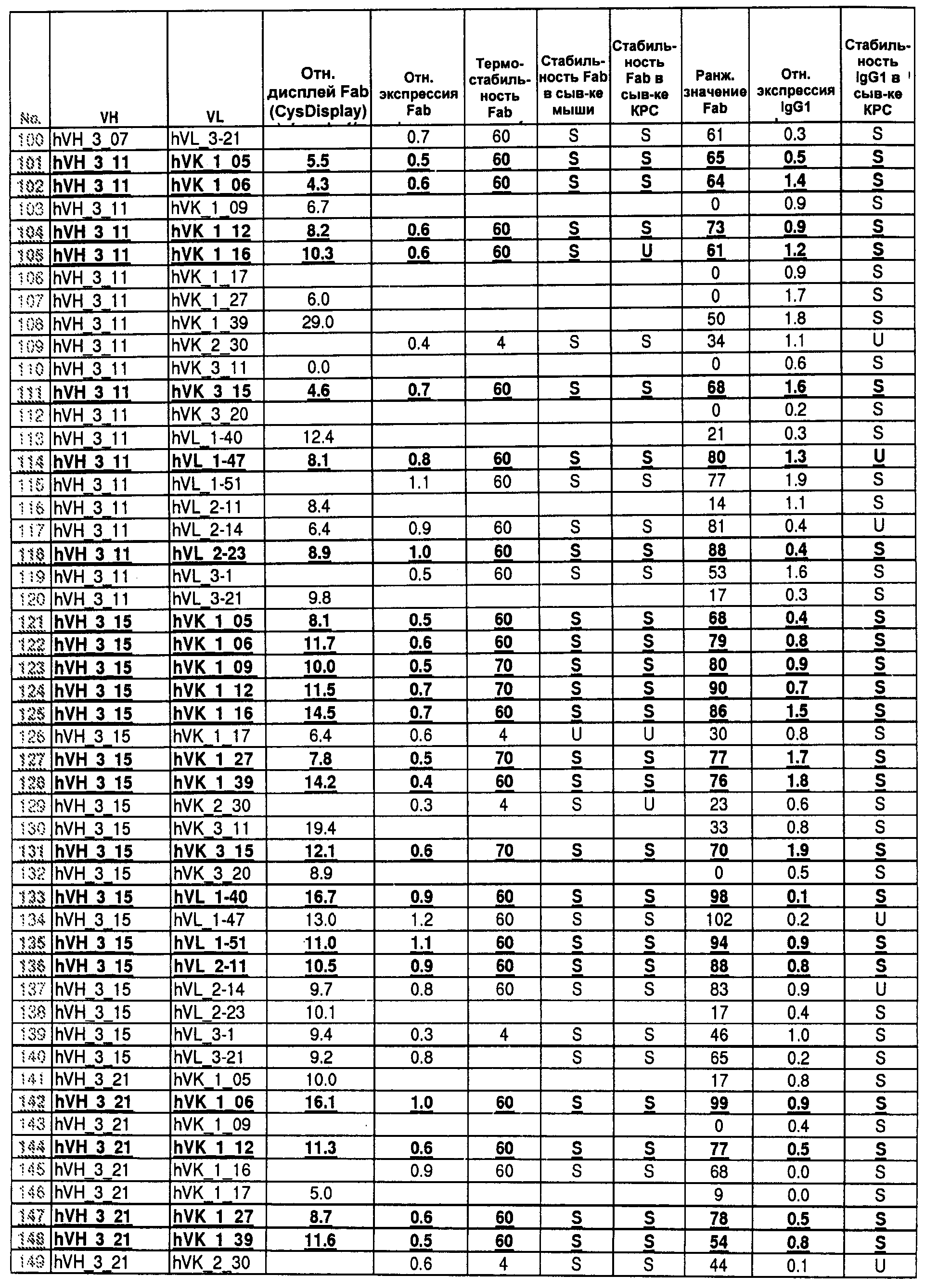





Как описано в предыдущих примерах, преобладающие гены эмбрионального типа VH и VL и преобладающие пары генов эмбрионального типа VH/VL идентифицировали из иммунного репертуара человека и иммунного репертуара интактного человека, затем преобладающие белковые последовательности эмбрионального типа VH и VL анализировали in silico для того, чтобы выявить белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, имеющие предпочтительные биофизические свойства. Как показано в Таблице 21 и на Фиг.37-38, обычно верхние 20 VH, верхние 8 Vλ и верхние 12 Vк выбирали для синтеза, комбинирования и последующего функционального анализа. Генные последовательности эмбрионального типа синтезировали и затем комбинировали с целью создания 400 пар белков эмбрионального типа, которые являются типичными для распространенных пар генов эмбрионального типа, находящихся в иммунном репертуаре, где каждая из вариабельных областей имеет предпочтительные биофизические свойства, как определено in silico. 400 пар белков эмбрионального типа VH/VL тестировали в отношении следующих свойств: а) относительный уровень экспонирования после продукции фага и фагового ИФА в Fab-формате; б) относительные уровни экспрессии Fab после продукции Fab в Е.соli, лизиса клеток Е.соli и ИФА-детекции продуцируемого Fab; в) термостабильность Fab после продукции Fab в Е.соli, лизиса клеток Е.соli и ИФА-детекции неденатурированного Fab после инкубации при повышенных температурах; г) стабильность Fab из лизатов Е.соli в сыворотке крупного рогатого скота/мышей посредством ИФА-детекции неденатурированных Fab после инкубации в сыворотке крупного рогатого скота/мышей; д) относительные уровни экспрессии человеческого IgG1 после продукции IgG1 в клетках млекопитающих и ИФА-детекции секретируемых IgG1 из супернатантов клеточных культур; и е) стабильность человеческого IgG1 в сыворотке крупного рогатого скота, как определено посредством ИФА-детекции неденатурированного Fab после инкубации в сыворотке крупного рогатого скота/мышей.
Используя данные, представленные в Таблице 32, специалист в данной области сможет легко идентифицировать пары белков эмбрионального типа, имеющие предпочтительные биофизические свойства.
Как правило, пары белков эмбрионального типа, имеющие пороговое значение для каждого функционального свойства, выбирали для включения в коллекции. Например, в некоторых воплощениях пары белков эмбрионального типа, имеющие все из следующих свойств, были выбраны для включения в коллекцию: 1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fab; 2) уровень экспрессии в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_VI1-40 AYA; 3) термостабильность при 60°С или более в течение по меньшей мере 45 минут в Fab-формате; 4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°С; 5) уровень экспрессии в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и 6) стабильность в сыворотке в IgG-формате в течение четырнадцати суток при 37°С. В Таблице 32 показаны жирным шрифтом и подчеркнуты пары белков эмбрионального типа, имеющие все эти функциональные свойства.
Однако как описано выше, пары белков эмбрионального типа, имеющие одно или более функциональных свойств, можно выбрать для включения в коллекции. В данной заявке создали комплексное ранжирование 400 тестируемых пар белков эмбрионального типа, так что каждую пару белков эмбрионального типа можно ранжировать относительно другой с учетом значения каждого из тестируемых функциональных свойств. Это позволило авторам изобретения выбрать одну или более пар белков эмбрионального типа, имеющих одно или более или все перечисленные функциональные свойства. В некоторых воплощениях коллекции содержат все пары белков эмбрионального типа, имеющие вышеупомянутые характеристики. В некоторых воплощениях коллекция содержит пары белков эмбрионального типа, имеющие самый высокий совокупный балл для 400 протестированных пар. В некоторых воплощениях пары белков эмбрионального типа, имеющие совокупные баллы в пределах верхних 10%, верхних 20% или верхних 30% из 400 протестированных пар, выбрали для включения в коллекции.
Следует понимать, что описание, конкретные примеры и данные, несмотря на указание конкретных воплощений, даны в качестве иллюстрации и не предназначены для ограничения настоящего изобретения. Различные изменения и модификации в пределах настоящего изобретения будут очевидны специалисту из обсуждения, описания и данных, содержащихся в данной заявке, и поэтому рассматриваются как часть изобретения.
Реферат
Настоящее изобретение относится к области иммунологии. Предложена коллекция антител (или их функциональных фрагментов), содержащих эмбриональные каркасные участки. Кроме того, рассмотрены коллекции нуклеиновых кислот, экспрессирующих векторов и рекомбинантных клеток-хозяев, а также способ получения коллекции антител по изобретению. Данное изобретение позволяет определить пары классов VH и VL в иммунном репертуаре человека для коллекции антител по изобретению, которые являются наиболее распространенными и имеют предпочтительные биофизические свойства. Предложенное решение может найти дальнейшее применение в скрининге терапевтических антител. 5 н. и 42 з.п. ф-лы, 61 ил., 32 табл., 8 пр.
Формула
1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fab;
2) уровень экспрессии в клетке Е. coli в Fab-формате по меньшей мере 0,4 по сравнению с Fab VH1-69 VLA_Vl1-40 AYA, последовательности которых представлены в Таблице 5, Таблице 9, на Фиг. 45А и Фиг. 47А;
3) термостабильность при 60°C или более в течение по меньшей мере 45 минут в Fab-формате;
4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°C;
5) уровень экспрессии в клетке HEK.EBNA в IgG-формате по меньшей мере 0,4 по сравнению с MOR03080; и
6) стабильность в сыворотке в IgG-формате в течение четырнадцати суток при 37°C;
и где указанные пары белков эмбрионального типа кодируются парой генов эмбрионального типа и выбраны
из
IGHV1-18 (SEQ ID NO: 65)/IGKV1-05 (SEQ ID NO: 118); IGHV1-18 (SEQ ID NO: 65)/IGLV2-23 (SEQ ID NO: 174); IGHV1-46 (SEQ ID NO: 68)/IGKV1-09 (SEQ ID NO: 121); IGHV1-46 (SEQ ID NO: 68)/IGKV1-39 (SEQ ID NO: 129); IGHV1-46 (SEQ ID NO: 68)/IGKV3-15 (SEQ ID NO: 153); IGHV1-69*01 (SEQ ID NO: 70)/IGKV1-05 (SEQ ID NO: 118); IGHV3-07 (SEQ ID NO: 77)/IGKV1-09 (SEQ ID NO: 121); IGHV3-07 (SEQ ID NO: 77)/IGKV1-12 (SEQ ID NO: 122); IGHV3-07 (SEQ ID NO: 77)/IGKV1-27 (SEQ ID NO: 126); IGHV3-07 (SEQ ID NO: 77)/IGKV3-15 (SEQ ID NO: 153); IGHV3-07 (SEQ ID NO: 77)/IGLV1-47 (SEQ ID NO: 167); IGHV3-07 (SEQ ID NO: 77)/IGLV2-23 (SEQ ID NO: 174); IGHV3-07 (SEQ ID NO: 77)/IGLV3-01 (SEQ ID NO: 176); IGHV3-11 (SEQ ID NO: 79)/IGKV1-05 (SEQ ID NO: 118); IGHV3-11 (SEQ ID NO: 79)/IGKV1-06 (SEQ ID NO: 119); IGHV3-11 (SEQ ID NO: 79)/IGKV1-12 (SEQ ID NO: 122); IGHV3-11 (SEQ ID NO: 79)/IGKV1-16 (SEQ ID NO: 124); IGHV3-11 (SEQ ID NO: 79)/IGKV3-15 (SEQ ID NO: 153); IGHV3-11 (SEQ ID NO: 79)/IGLV1-47 (SEQ ID NO: 167); IGHV3-11 (SEQ ID NO: 79)/IGLV2-23 (SEQ ID NO: 174); IGHV3-15 (SEQ ID NO: 81)/IGKV1-05 (SEQ ID NO: 118); IGHV3-15 (SEQ ID NO: 81)/IGKV1-06 (SEQ ID NO: 119); IGHV3-15 (SEQ ID NO: 81)/IGKV1-09 (SEQ ID NO: 121); IGHV3-15 (SEQ ID NO: 81)/IGKV1-12 (SEQ ID NO: 122); IGHV3-15 (SEQ ID NO: 81)/IGKV1-16 (SEQ ID NO: 124); IGHV3-15 (SEQ ID NO: 81)/IGKV1-27 (SEQ ID NO: 126); IGHV3-15 (SEQ ID NO: 81)/IGKV1-39 (SEQ ID NO: 129); IGHV3-15 (SEQ ID NO: 81)/IGKV3-15 (SEQ ID NO: 153); IGHV3-15 (SEQ ID NO: 81)/IGLV1-40 (SEQ ID NO: 165); IGHV3-15 (SEQ ID NO: 81)/IGLV1-51 (SEQ ID NO: 169); IGHV3-15 (SEQ ID NO: 81)/IGLV2-11 (SEQ ID NO: 171); IGHV3-21 (SEQ ID NO: 84)/IGKV1-06 (SEQ ID NO: 119); IGHV3-21 (SEQ ID NO: 84)/IGKV1-12 (SEQ ID NO: 122); IGHV3-21 (SEQ ID NO: 84)/IGKV1-27 (SEQ ID NO: 126); IGHV3-21 (SEQ ID NO: 84)/IGKV1-39 (SEQ ID NO: 129); IGHV3-21 (SEQ ID NO: 84)/IGLV2-14 (SEQ ID NO: 172); IGHV3-21 (SEQ ID NO: 84)/IGLV2-23 (SEQ ID NO: 174); IGHV3-30 (SEQ ID NO: 86)/IGLV2-23 (SEQ ID NO: 174); IGHV3-30 (SEQ ID NO: 86)/IGLV3-1 (SEQ ID NO: 176); IGHV3-33 (SEQ ID NO: 89)/IGKV3-15 (SEQ ID NO: 153); IGHV3-33 (SEQ ID NO: 89)/IGLV2-23 (SEQ ID NO: 174); IGHV3-48 (SEQ ID NO: 93)/IGKV1-27 (SEQ ID NO: 126); IGHV3-53 (SEQ ID NO: 95)/IGKV1-09 (SEQ ID NO: 121); IGHV3-53 (SEQ ID NO: 95)/IGKV1-12 (SEQ ID NO: 122); IGHV3-53 (SEQ ID NO: 95)/IGLV1-51 (SEQ ID NO: 169); IGHV3-53 (SEQ ID NO: 95)/IGLV2-23 (SEQ ID NO: 174); IGHV3-53 (SEQ ID NO: 95)/IGLV3-1 (SEQ ID NO: 176); IGHV3-74 (SEQ ID NO: 100)/IGKV1-06 (SEQ ID NO: 119); IGHV3-74 (SEQ ID NO: 100)/IGKV1-09 (SEQ ID NO: 121); IGHV3-74 (SEQ ID NO: 100)/IGKV1-27 (SEQ ID NO: 126); IGHV3-74 (SEQ ID NO: 100)/IGKV3-15 (SEQ ID NO: 153); IGHV3-74 (SEQ ID NO: 100)/IGLV1-51 (SEQ ID NO: 169); IGHV4-04 (SEQ ID NO: 102)/IGLV2-23 (SEQ ID NO: 174); IGHV4-04(SEQ ID NO: 102)/IGLV3-1 (SEQ ID NO: 176); IGHV4-04 (SEQ ID NO: 102)/IGLV3-21 (SEQ ID NO: 182); IGHV4-39 (SEQ ID NO: 109)/IGLV2-14 (SEQ ID NO: 172); IGHV4-39 (SEQ ID NO: 109)/IGLV3-1 (SEQ ID NO: 176); IGHV5-51 (SEQ ID NO: 113)/IGKV1-09 (SEQ ID NO: 121); IGHV5-51 (SEQ ID NO: 113)/IGLV2-23 (SEQ ID NO: 174); IGHV5-51 (SEQ ID NO: 113)/IGLV3-1 (SEQ ID NO: 176); IGHV6-1 (SEQ ID NO: 115)/IGKV1-06 (SEQ ID NO: 119) и IGHV6-1 (SEQ ID NO: 115)/IGLV2-23 (SEQ ID NO: 174).
а) определения пар генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, присутствующих в иммунном репертуаре человека;
б) идентификации пар белков эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, обладающих следующими свойствами:
1) относительный уровень экспонирования в Fab-формате, представляющий собой значение не ниже 75% выбранных Fab;
2) уровень экспрессии в клетке Е. coli в Fab-формате по меньшей мере 0,4 по сравнению с Fab pMx11_FH VH1-69 VLA_Vl1-40 AYA, последовательности которых представлены в Таблице 5, Таблице 9, на Фиг. 45А и Фиг. 47А;
3) термостабильность при 60°C или более в течение по меньшей мере 45 минут в Fab-формате;
4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°C;
5) уровень экспрессии в клетке HEK.EBNA в IgG-формате по меньшей мере 0,4 по сравнению с MOR3080; и
6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°C; и
в) создание коллекции антител или их функциональных фрагментов, содержащих белковые последовательности эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи пар белков эмбрионального типа, идентифицированных на стадии (б).
ба) идентификации пар генов эмбрионального типа вариабельной области тяжелой цепи и вариабельной области легкой цепи, присутствующих в концентрации по меньшей мере 0,05% в иммунном репертуаре человека;
бб) создания антител или их функциональных фрагментов, содержащих пары белков эмбрионального типа, идентифицированные на стадии ба); и
бв) оценки следующих свойств указанной пары белков эмбрионального типа:
1) относительный уровень экспонирования в Fab-формате;
2) уровень экспрессии в Fab-формате;
3) термостабильность при 60°C или больше в Fab-формате;
4) стабильность в сыворотке крупного рогатого скота или мышей в Fab-формате в течение более десяти суток при 37°C;
5) уровень экспрессии в IgG-формате; и
6) стабильность в сыворотке крупного рогатого скота в IgG-формате в течение четырнадцати суток при 37°C.
аа) выделение человеческих В-клеток из образца;
аб) получение кДНК из В-клеток;
ав) ПЦР(полимеразная цепная реакция)-амплификация кДНК из В-клеток;
аг) секвенирование ПЦР-продуктов;
ад) идентификация генов эмбрионального типа каждого ПЦР-продукта.
ва) синтез нуклеиновых кислот, кодирующих антитела или их функциональные фрагменты;
вб) клонирование нуклеиновых кислот в вектор; и
вв) экспрессирование антител или их функциональных фрагментов.