Библиотеки только-hc-cdr3 с уменьшенной комбинаторной избыточностью и оптимизированным распределением длины петли - RU2745452C2
Код документа: RU2745452C2
Чертежи






















Описание
Настоящее изобретение относится к библиотеке векторов, содержащих человеческие области HC-CDR3 различной длины, где разнообразие указанной библиотеки сфокусировано только на области HC-CDR3, и разнообразие оптимизировано таким образом, что избыточность уменьшена для коротких петель HC-CDR3, и перекрывание вариантов области HC-CDR3 для больших длин петель увеличено. Библиотека по настоящему изобретению экспонирована на фаге для отбора против антигенов-мишеней.
Уровень техники для изобретения
Несмотря на недавнее развитие платформ для исследования in vivo, обеспечивающих полностью человеческие антитела (Green, 2014), библиотеки рекомбинантных антител продолжают представлять важный вспомогательный способ, в частности, для сложных мишеней, попытки исследований для которых in vivo были неудачными? или их было невозможно провести из-за характера антигена. Библиотеки рекомбинантных антител описаны во множестве топологий и форматов (Mondon et al., 2008). Библиотеки обычно конструируют комбинаторным способом, случайным образом последовательно комбинируя более комплексные варианты внутри вплоть до шести областей петель, определяющих комплементарность областей (CDR). Наибольшее разнообразие, как правило, вводят в область CDR3 вариабельного домена тяжелой цепи (HC) (HC-CDR3), наиболее вариабельного и важного фрагмента CDR, присутствующего в природных антителах (Tonegawa, 1983; Chothia et al., 1989).
Для областей HC-CDR3 распределение длин петель (процент каждой длины петли, присутствующей в библиотеке), реализованное в библиотеках рекомбинантных антител, обычно отражает распределение, наблюдаемое в природных антителах, которое представляет собой распределение, для которого показано приблизительно колоколообразное распределение с максимумом в районе петель HC-CDR3 длиной 12 (Zemlin et al., 2003). За немногими исключениями (например, Fellouse et al, 2007; Mahon et al., 2013), библиотеки рекомбинантных антител разрабатывали, чтобы следовать (приблизительно) этому колоколообразному распределению. Это имеет важные последствия, когда библиотеку высокой сложности (с общей сложностью от 109 до 1010 или выше) разрабатывают комбинаторным образом. Варианты более коротких петель HC-CDR3 могут избыточно присутствовать (практически все варианты присутствуют, или даже присутствуют несколько раз), относительно вариантов длинных петель HC-CDR3, поскольку для последних присутствует только небольшая доля всех возможных вариантов. Использование постоянного распределения длин для длины петли HC-CDR3 (все длины HC-CDR3 присутствуют в равных долях в библиотеке) дополнительно увеличивает избыточность более коротких петель HC-CDR3 (их процентная доля выше по сравнению с колоколообразным распределением, наблюдаемым для природных антител), незначительно увеличивает общее перекрывание возможных вариантов для длинных петель HC-CDR3 и уменьшает перекрывание для петель HC-CDR3 в диапазоне средней длины.
Общее количество антител, уже одобренных в качестве лекарственных средств или находящихся в клинической разработке, постоянно увеличивается. Поиск в базе данных ChEMBL (https://www.ebi.ac.uk/chembl/) показал, что распределение длин их HC-CDR3 имеет выраженный максимум на длине HC-CDR3 10, что отличается от сглаженного колоколообразного распределения длин, наблюдаемого для природных человеческих антител, но также от наблюдаемого для мышиных антител. Петли HC-CDR3 длиной 10 должны, таким образом, быть представлены особенно хорошо в библиотеке, нацеленной на выделение кандидатов для терапевтических антител. Является вероятным, что антитела с более короткими петлями HC-CDR3 экспрессируются хорошо и проявляют более низкую тенденцию к агрегации, что является важными характеристиками для успешной разработки продукта.
Несмотря на то, что библиотеки только-HC-CDR3 получены в различных контекстах (Barbas et al., 1992; Braunagel et al., 1997; Pini et al., 1998; Hoet et al., 2005; Silacci et al., 2005; Mahon et al., 2013; US 2006/0257937A1), многие библиотеки рекомбинантных антител вносят разнообразие не только в область HC-CDR3, но также в одну или несколько из пяти других областей CDR (например, Knappik et al., 2000; Prassler et al., 2013). Затем разнообразие, представленное в различных областях CDR, комбинируют, полностью случайным образом, в ходе клонирования библиотеки, обычно начиная с области CDR с наименьшим общим разнообразием. За исключением коротких петель HC-CDR3, где может существовать некоторая избыточность и могут присутствовать дубликаты, каждый вариант области HC-CDR3 следует считать уникальным, присутствующим только один раз в библиотеке. Как следствие, каждый вариант петли HC-CDR3 становится «ассоциированным» с полностью случайной комбинацией вариантов других областей CDR, без какого-либо структурного или функционального отбора по совместимости. По сравнению с ситуацией, когда другие области CDR представлены зародышевыми последовательностями или одной консенсусной последовательностью (например, для области CDR3 легкой цепи), в данном случае не существует преимущества наличия конкретного варианта HC-CDR3, комбинированного со случайной выборкой вариантов других (от одной до пяти) областей CDR. Библиотека только-HC-CDR3 должна, таким образом, функционировать так же хорошо или даже лучше по сравнению с библиотекой с дополнительным разнообразием. Единственным исключением являются короткие петли HC-CDR3, для которых, из-за избыточности (присутствия вариантов в более чем одной копии в библиотеке), можно исследовать очень ограниченное количество комбинаций изменчивости в других CDR, т.е. один и тот же вариант HC-CDR3 может присутствовать множество раз, каждый раз с различной комбинацией вариантов в других областях CDR. Однако, чтобы комбинировать вариант HC-CDR3 только с 10 из этих комбинаций, уровень дупликации области HC-CDR3 должен составлять около 10. Даже для коротких петель HC-CDR3 это может повлечь за собой увеличение доли этой длины петли в библиотеке в 10 раз, что непрактично для большинства длин петли HC-CDR3. Например, петлю HC-CDR3 конкретной длины, представляющую небольшой процент из общей библиотеки, необходимо представить в относительно высокой двузначной процентной доле, чтобы эффективно исследовать дополнительное разнообразие, присутствующее в библиотеке, например, в LC-CDR3. В то время как этого уже трудно достичь для одной длины петли HC-CDR3, является невозможным получение библиотеки, в которой варианты для всех длин петли HC-CDR3 эффективно скомбинированы даже с ограниченным количеством вариантов в другой области CDR. В одном случае (Mahon et al., 2013) эффективность библиотеки только-HC-CDR3 сравнивали с соответствующей библиотекой HC-CDR3- и -LC-CDR3. Для библиотеки только-HC-CDR3 показаны превосходящие свойства; однако авторы не полностью признали «комбинаторный эффект», обеспечивающий преимущества библиотеки только-HC-CDR3, но приписали лучшую эффективность библиотеки HC-CDR3 возможным случаям структурной несовместимости между разнообразием LC-CDR3 и HC-CDR3 в библиотеке HC-CDR3-и-LC-CDR3.
Библиотеки рекомбинантных антител, в которых дизайн разнообразия HC-CDR3 основан на частотах аминокислот в каждом положении, наблюдаемых в природных антителах, получали либо с использованием стандартных вырожденных олигонуклеотидов (например, Philibert et al., 2007), позволяющих только приближенное представление желательного распределения аминокислот и образующих нежелательные кодоны Cys и стоп-кодоны, либо посредством олигонуклеотидов, разнообразие в которые введено посредством смесей тримерных блоков, кодирующих аминокислоты (Braunagel et al., 1997; Knappik et al., 2000; Prassler et al., 2013, Mahon et al., 2013; патентные заявки США 2006/0257937A1, EP1979378B1).
Однако ни в одном из этих примеров не принимают во внимание комбинаторный эффект, который относится к количеству различных вариантов, фактически присутствующих в библиотеке для конкретной длины HC-CDR3, представляющих определенную долю общей библиотеки, по сравнению с теоретически возможным количеством вариантов, как определено дизайном библиотеки. При наличие колоколообразного «подобного природному» распределения длин петли HC-CDR3, комбинаторный эффект приводит к избыточному представлению вариантов коротких петель HC-CDR3 и очень небольшому перекрыванию более длинных петель HC-CDR3. В US 2006/0257937A1 описаны только виды дизайна библиотеки, перекрывающей ограниченный диапазон длин петли HC-CDR3 (8, 10, 13, 14, 15, 17, 18, 19), и аминокислотный состав в положениях петли HC-CDR3 либо соответствует фиксированной эквимолярной смеси 19 различных аминокислот, либо ограничен фиксированной смесью небольшого количества аминокислот для конкретного положения, для всех без исключения длин петли HC-CDR3. В EP1979378B1 описан дизайн библиотеки, в котором длины петли HC-CDR3 разделены на три различных диапазона длин, где каждый диапазон имеет определенный аминокислотный состав (называемый фактором разнообразия). Фактор разнообразия, представляющий аминокислотный состав всех петель HC-CDR3 в конкретном диапазоне длин для различных положений петли HC-CDR3, содержит положения по Kabat 95-102. Для каждого положения или диапазона положений внутри HC-CDR3, фактор разнообразия приписывает конкретные частоты подгруппе аминокислот, в то время как все остальные аминокислоты (за исключением Cys) включены с фиксированной частотой, за исключением положений 101 и 102, в которых присутствует только подгруппа аминокислот. Этот дизайн, таким образом, образует огромное количество теоретически возможных вариантов, поскольку все аминокислоты (за исключением Cys) присутствуют, с различными частотами, почти во всех положениях петли HC-CDR3 и для всех длин петли HC-CDR3. Даже для петель HC-CDR3 в диапазоне средней длины (например, длиной 9, 10, 11), фактическое количество вариантов, присутствующих в библиотеке общей сложностью 1010, представляет только долю всех возможных вариантов в соответствии с этим дизайном.
Библиотеки рекомбинантных человеческих антител, включающие разнообразие синтетических CDR3 вплоть до суммарной общей сложности приблизительно 1012, получены (Knappik et al., 2000, Prassler et al., 2011), и доказано их успешное практическое применение (отбор антител против конкретной мишени), возможно также, благодаря их огромному размеру. Однако получение библиотек такого размера требует очень значительных усилий, а также имеет высокую экономическую стоимость.
Таким образом, существует необходимость дизайна библиотек человеческих антител с оптимизированными свойствами, т.е. с высокой вероятностью отбора хороших клонов-кандидатов для дальнейшей разработки терапевтических антител, которые можно получать с приемлемой трудоемкостью экспериментов и с приемлемой экономической стоимостью.
Краткое описание чертежей
Фигура 1: Схема нумерацииKabat для области CDR3 тяжелой цепи для длин петли от 5 до 20. Указана область HC-CDR3, и остатки, составляющие часть области HC-CDR3, выделены серым. В положениях по Kabat 92, 93, 94 и 103, 104, предшествующих области HC-CDR3 и следующих за ней, соответственно, указаны наиболее часто наблюдаемые аминокислоты (CAR и WG).
Фигура 2: Распределение длин петли HC-CDR3, наблюдаемое в природных антителах.
Фигура 3: Последовательность одноцепочечного каркаса, включающего балластный элемент, заменяющий область HC-CDR3. Расщепление с использованием PstI/StyI удаляет балласт и позволяет вставку олигонуклеотидов, содержащих разнообразие HC-CDR3. Уникальные участки рестрикции обозначены подчеркиванием в последовательности. Показана также локализация областей CDR1 и CDR3 вариабельной легкой цепи.
Фигура 4: Схематическое представление, показывающее тяжелую цепь одноцепочечного каркаса после вставки олигонуклеотидов, содержащих разнообразие HC-CDR3.
Фигура 5: Таблица кодирующих кодоны тримерных блоков, используемых для получения разнообразия в положениях с более, чем одной аминокислотой в олигонуклеотидах, содержащих разнообразие HC-CDR3.
Фигура 6: Схематическое представление фагмидного вектора BaseVector_VH3_VK1_v22
Фигура 7: Схематическое представление фагмидного вектора BaseVector_VH3_VK1_v22 с указанными уникальными участками рестрикции.
Фигура 8: Последовательности HC-CDR3 и длина петли HC-CDR3 одобренных или разработанных для клиники терапевтических антител.
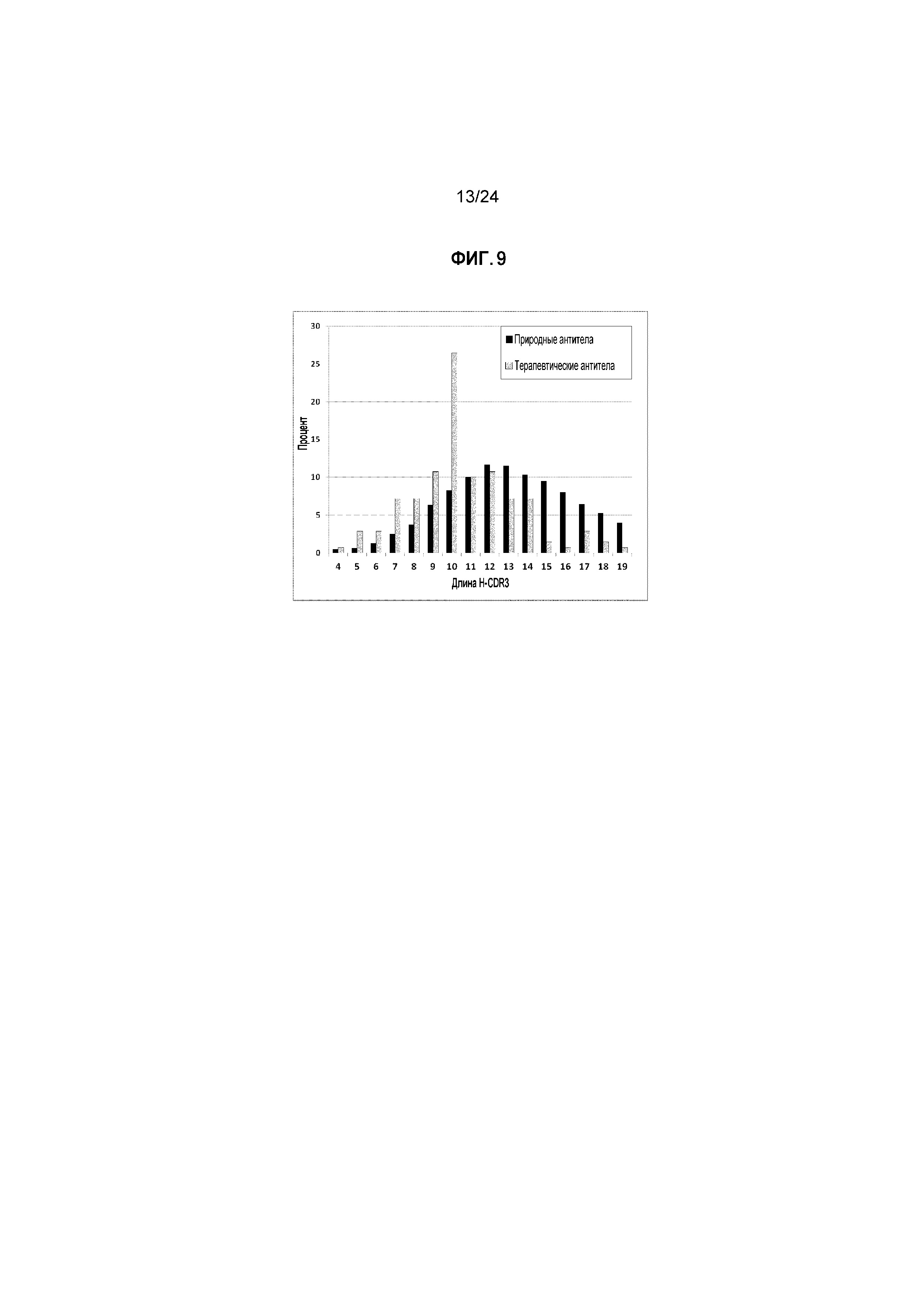
Фигура 9: Сравнение распределения длин петли HC-CDR3 одобренных или разработанных для клиники терапевтических антител и природных антител. Процентные значения для природных антител повторно нормализованы (всего до 100%) в диапазоне длин от 4 до 19.
Фигура 10: Сравнение распределения длин петли HC-CDR3 одобренных или разработанных для клиники терапевтических антител, природных антител, и распределения длин петли в оптимизированном дизайне библиотеки. Затем процентные значения для природных антител повторно нормализовали (всего до 100%) в диапазоне длин от 5 до 19.
Фигура 11: Олигонуклеотид, кодирующий разнообразие HC-CDR3 для петли длиной 15. Для положений с присутствием более чем одной аминокислоты, показана смесь тримерных блоков, включая относительные частоты индивидуальных тримерных блоков. Части олигонуклеотида, содержащие участки рестрикции PstI или StyI, выделены серым.
Фигура 12 (A-D): Выделение специфических для BSA клонов M13-scFv из библиотеки.
A) Проводили три цикла пэннинга, и относительное обогащение представлено как соотношение ВХОДА/ВЫХОДА (всего трансд.ед. x105); B) Специфичность поликлональных смесей фагов (подбиблиотек, состоящих из элюированных фагов из I-III циклов отбора), тестировали посредством фагового ELISA на BSA и нескольких неродственных антигенах: C) Анализ ELISA на BSA для отдельных клонов, выделенных после III цикла; пунктирной линией показан рассчитанный порог отсечения, использованный для определения специфичности (OD=0,133); D) Специфичность 12 положительных клонов тестировали посредством фагового ELISA на BSA и нескольких неродственных антигенах.
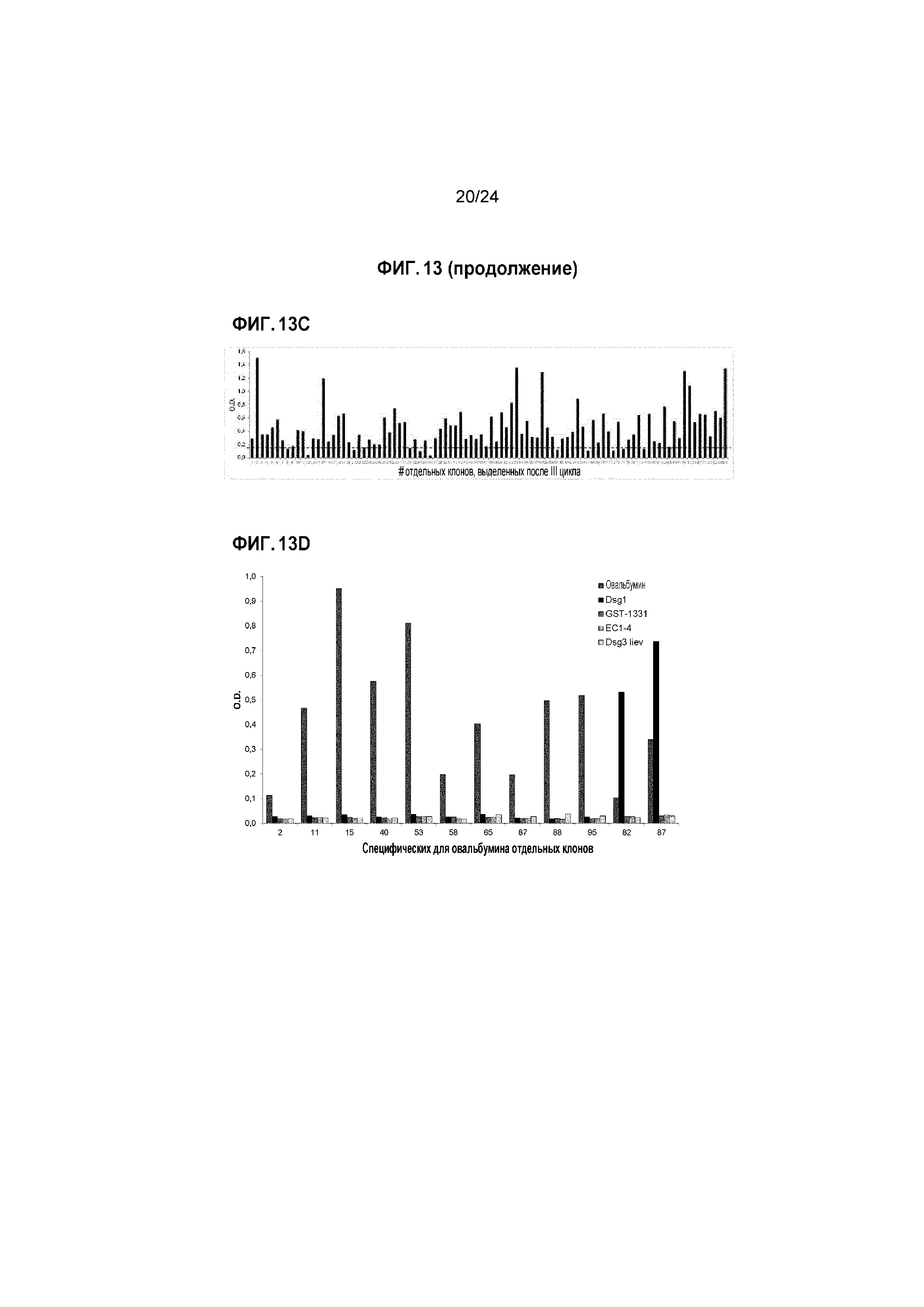
Фигура 13 (A-D). Выделение специфических для овальбумина (OVA) клонов M13-scFv из библиотеки.
A) Проводили три цикла пэннинга, и относительное обогащение представлено как соотношение ВХОДА/ВЫХОДА (всего трансд.ед. x105); B) Специфичность поликлональных смесей фагов (подбиблиотек, состоящих из элюированных фагов из I-III циклов отбора), тестировали посредством фагового ELISA на OVA и нескольких неродственных антигенах; C) Анализ ELISA на OVA для отдельных клонов, выделенных после III цикла; пунктирной линией показан рассчитанный порог отсечения, использованный для определения специфичности (OD=0,166); D) Специфичность 10 положительных клонов тестировали посредством фагового ELISA на OVA и нескольких неродственных антигенах.
Фигура 14 (A-D). Выделение специфических для Dsg1 клонов M13-scFv из библиотеки.
A) Проводили три цикла пэннинга, и относительное обогащение представлено как соотношение ВХОДА/ВЫХОДА (всего трансд.ед. x105); B) Специфичность поликлональных смесей фагов (подбиблиотек, состоящих из элюированных фагов из I-III циклов отбора), тестировали посредством фагового ELISA в лунках, предварительно покрытых коммерчески доступным Dsg1, и на нескольких неродственных антигенах; C) Анализ ELISA в предварительно покрытых Dsg1 лунках для отдельных клонов, выделенных после III цикла; пунктирной линией показан рассчитанный порог отсечения, использованный для определения специфичности (OD=0,102); D) Специфичность 10 положительных клонов тестировали посредством фагового ELISA на Dsg1 и нескольких неродственных антигенах.
Фигура 15 (A-D). Выделение специфических для FGFR4 клонов M13-scFv из библиотеки.
A) Проводили три цикла пэннинга, и относительное обогащение представлено как соотношение ВХОДА/ВЫХОДА (всего трансд.ед. x105); B) Специфичность поликлональных смесей фагов (подбиблиотек, состоящих из элюированных фагов из I-III циклов отбора), тестировали посредством фагового ELISA на FGFR-4и нескольких неродственных антигенах; C) Анализ ELISA на FGFR-4 для отдельных клонов, выделенных после III цикла; пунктирной линией показан рассчитанный порог отсечения, использованный для определения специфичности (OD=0,134); D) Специфичность 20 положительных клонов тестировали посредством фагового ELISA на FGFR-4 и нескольких неродственных антигенах.
Определения
Если не определено иное, все термины из данной области, условные обозначения и другая научная терминология, применяемые в настоящем документе, имеют такие же значения, какие являются общепринятыми для специалиста в области, к которой относится это описание. В некоторых случаях, термины с общепринятыми значениями определены в настоящем описании для ясности и/или в качестве справочного материала; таким образом, включение таких определений не следует рассматривать как представление существенных отличий от общепринятых в данной области.
Термин «сложность библиотеки» в настоящем описании относится к общему количеству различных вариантов, присутствующих в библиотеке, независимо от длины петли HC-CDR3.
Термин «разнообразие» в настоящем описании относится к присутствию более, чем одной аминокислоты в одном или нескольких положениях.
Термин «избыточность» в настоящем описании относится к среднему количеству раз, которое варианты для петли HC-CDR3 с определенной длиной представлены в библиотеке.
Под термином «только-HC-CDR3 библиотека» авторы настоящего изобретения подразумевают библиотеку, которая имеет изменчивость только внутри области HC-CDR3 и в положении 94 по Kabat, предшествующем области HC-CDR3, и которая не имеет изменчивости в других пяти областях CDR, HC-CDR1, HC-CDR2, LC-CDR1, LC-CDR2 и LC-CDR3.
Термин «фрагмент антитела» или «функциональный фрагмент», в рамках изобретения включает любой антигенсвязывающий фрагмент, такой как Fab, F(ab′)2, Fab′, Fv, scFv, одиночные цепи, включающие часть Fc, наноантитела и другие подобные антителам структуры, имеющие каркасы, отличные от каркаса вариабельных областей. Термин «функциональный фрагмент» включает, но без ограничения, любую часть антитела, сохраняющую способность связывать представляющий интерес антиген.
Термин «зародышевая» в настоящем описании относится к последовательности нуклеиновой кислоты, кодирующей антитела или их функциональные фрагменты, передаваемой от родителя к потомству.
Термин «комбинация вариабельной тяжелой цепи и вариабельной легкой цепи» или «комбинация VH/VL» в настоящем описании относится к комбинации одной вариабельной тяжелой цепи и одной вариабельной легкой цепи. Антитело или функциональный фрагмент антитела содержит по меньшей мере одну вариабельную тяжелую цепь, связанную с вариабельной легкой цепью, формирующие область связывания антигена.
Термин «вариабельный домен», вариабельный домен легкой цепи (VL) или вариабельный домен тяжелой цепи (VH), в настоящем описании относится к области иммуноглобулина, которая находится в контакте с антигеном и содержит три гипервариабельные области, обозначенные как «определяющие комплементарность области» или «CDR» (Kabat, 1983; Chothia & Lesk, 1987).
HC-CDR3 и LC-CDR3 в настоящем описании относятся к третьим определяющим комплементарность областям вариабельного домена тяжелой цепи и вариабельного домена легкой цепи, соответственно.
Термин «номенклатура Kabat» в настоящем описании относится к схеме нумерации остатков доменов VL или VH, как определено в Kabat, 1983, и схематически показано для домена VH области CDR3 на фиг. 1. Нумерация остатков для домена VH и петли HC-CDR3 относится к номенклатуре Kabat на протяжении настоящего описания.
Термин «вариант» в настоящем описании относится к антителу или фрагменту антитела с аминокислотной последовательностью, которая отличается от аминокислотной последовательности всех других антител или фрагментов антител в библиотеке.
Термин «оптимизированный по кодонному составу» или «оптимизация по кодонному составу» в настоящем описании относится к нуклеотидной последовательности, измененной таким образом, чтобы кодированная аминокислотная последовательность оставалось такой же, в то время как кодоны, кодирующие индивидуальные аминокислоты, были изменены таким образом, чтобы оптимизировать экспрессию кодированного белка в конкретном хозяине, например, бактериальных клетках.
Термин «библиотека», в рамках изобретения включает, но без ограничения, библиотеки фагового дисплея, рибосомного дисплея, бактериального дисплея, дрожжевого дисплея и дисплея у млекопитающих. В предпочтительном варианте осуществления настоящего изобретения используют библиотеку фагового дисплея.
Термин «вектор для дисплея», в рамках изобретения, включает последовательность ДНК, имеющую способность к прямой репликации и поддержанию рекомбинантной молекулы ДНК вне хромосомы в клетке-хозяине, такой как бактериальная клетка-хозяин, трансформированная им. Такие последовательности ДНК хорошо известны в данной области.
В соответствии с настоящим изобретением, векторы для дисплея могут, например, представлять собой фаговые векторы или фагмидные векторы, происходящие из класса нитевидного бактериофага fd, M13 или fl. Такие векторы являются способными содействовать экспонированию белка на поверхности нитевидного бактериофага.
Термин «родственные антителу пептиды» в настоящем описании относится к пептидам, которые содержат структурные домены, происходящие из антитела, и могут содержать один или несколько доменов антитела, которые могут являться ковалентно связанными, связанными дисульфидными связями или ассоциированными в форме комплекса.
Термин «генетические упаковки», в рамках изобретения, относится к поддающейся репликации упаковке генетического дисплея, в которой частица экспонирует полипептид на своей поверхности. Упаковка может представлять собой бактериофаг, экспонирующий антигенсвязывающий домен на своей поверхности. Когда антигенсвязывающий домен соответствует родственному антителу пептиду, этот тип упаковки называют фаговым антителом.
Описание изобретения
Изобретение относится к коллекции различных областей HC-CDR3 различных человеческих антител с оптимизированными свойствами, где разнообразие сконструировано посредством способа, который позволяет, при желательной общей сложности библиотеки, изменение распределения длины петли HC-CDR3 в библиотеке и изменение разнообразия аминокислот в каждом положении в каждой петле HC-CDR3.
В частности, по настоящему изобретению получают библиотеку человеческих антител с оптимизированными свойствами: уменьшенной комбинаторной избыточностью (присутствием дуплицированных вариантов) посредством оптимизации распределения длин петли HC-CDR3, полученную с приемлемой трудоемкостью экспериментов и с низкой экономической стоимостью.
Указанные преимущества неожиданно получены посредством ограничения разнообразия только петлей HC-CDR3 и положением, предшествующим области HC-CDR3, таким образом, что избыточность уменьшается до приемлемого уровня (менее 2) для всех длин петли HC-CDR3, и варианты для длин петли HC-CDR3 от 9 до 11 особенно хорошо представлены в библиотеке.
Библиотека по настоящему изобретению, таким образом, имеет преимущество особенно хорошего представления длин петли HC-CDR3, часто наблюдаемых в одобренных терапевтических антителах или антителах, находящихся в клинической разработке.
С использованием способа оптимизации дизайна библиотеки, описанного в примере 4, желательное значение порога избыточности (присутствия дуплицированных вариантов) можно корректировать для каждой длины петли HC-CDR3.
В то же самое время перекрывание для каждой длины петли HC-CDR3 (долю от всех возможных вариантов для конкретной длины петли HC-CDR3, фактически присутствующую в библиотеке) можно оптимизировать для одной или нескольких длин петли HC-CDR3, представляющих особенный интерес, в пределах, требуемых суммарной общей сложностью библиотеки.
Таким образом, целью настоящего изобретения является получение библиотеки векторов или генетических упаковок с общей сложностью C, которые экспонируют и экспрессируют или содержат члены разнообразного семейства родственных антителу пептидов, полипептидов или белков, и совместно экспонируют и экспрессируют или содержат по меньшей мере часть разнообразия семейства антител, где векторы или генетические упаковки содержат последовательности ДНК, кодирующие область HC-CDR3, и положение, предшествующее области HC-CDR3, имеющие следующую последовательность:
ZX1YnX3X4X5,
где:
C=1,3×1010
Z соответствует положению 94 по Kabat;
X1 соответствует положению 95 по Kabat;
n представляет собой целое число от 3 до 11;
Y соответствует положениям 96-98 HC по Kabat (n=3) или положениям 96-99 HC по Kabat (n=4), или положениям HC 96-100 по Kabat (n=5), или положениям 96-100a HC по Kabat (n=6), или положениям 96-100b HC по Kabat (n=7), или положениям 96-100c HC по Kabat (n=8), или положениям 96-100d HC по Kabat (n=9), или положениям 96-100e HC по Kabat (n=10), или положениям 96-100f HC по Kabat (n=11);
X3 соответствует положению 99 HC по Kabat (n=3) или положению 100 HC по Kabat (n=4), или положению 100a HC по Kabat (n=5), или положению 100b HC по Kabat (n=6), или положению 100c HC по Kabat (n=7), или положению 100d HC по Kabat (n=8), или положению 100e HC по Kabat (n=9), или положению 100f HC по Kabat (n=10), или положению 100g HC по Kabat (n=11);
X4 соответствует положению 101 HC по Kabat;
X5 соответствует положению 102 HC по Kabat;
отличающиеся тем, что процентная доля p(L) (L=n+4) каждой области ZX1YnX3X4X5присутствует в библиотеке в соответствии со значениями, приведенными в таблице 2C;
и положения Z, X1, X2, X3, X4, X5 и Yn (n=3-11) для каждой области HC-CDR3 длиной L=n+4 заняты определенными аминокислотами, в соответствии с относительной частотой, описанной в таблицах 3A-3I.
В предпочтительном варианте осуществления, указанные родственные антителу пептиды, полипептиды или белки происходят из человека.
В следующем варианте осуществления указанные родственные антителу пептиды, полипептиды или белки происходят из кошки или собаки.
В следующем предпочтительном варианте осуществления в библиотеке векторов или генетических упаковок по настоящему изобретению, длины петель HC-CDR3 лежат в диапазоне от 9 до 11. В одном варианте осуществления настоящего изобретения, родственные антителу пептиды, полипептиды или белки представляют собой антитела или их фрагменты, выбранные из антитела, содержащего один или несколько константных доменов, одноцепочечного антитела, фрагмента FAB, антитела только с тяжелой цепью или только вариабельного домена тяжелой цепи.
Предпочтительно, указанное антитело или его фрагменты представляют собой одноцепочечное антитело.
В следующем варианте осуществления, родственные антителу пептиды, полипептиды или белки в соответствии с настоящим изобретением содержат зародышевые вариабельные фрагменты человеческого антитела.
В следующем варианте осуществления настоящего изобретения указанную область только HC-CDR3 вводят в константный одноцепочечный каркас, характеризующийся зародышевыми вариабельными доменами человеческой тяжелой и легкой цепи, где область CDR3 легкой цепи имеет длину 9.
Указанные последовательности представляют аминокислоты, наиболее часто наблюдаемые в каждом из девяти положений CDR3 легкой цепи в природных человеческих антителах.
Использование зародышевых последовательностей вариабельного домена в одноцепочечном каркасе обеспечивает преимущества, поскольку эти последовательности не содержат соматических мутаций и, таким образом, ожидают, что они являются менее иммуногенными, что уменьшает вероятность последующего наблюдения ответа человеческих антител против человеческих антител в ходе клинического тестирования у субъектов-людей.
В предпочтительном варианте осуществления настоящего изобретения, родственные антителу пептиды, полипептиды или белки содержат вариабельный домен VK1 легкой цепи антитела человека, содержащий человеческие зародышевые последовательности, и вариабельный домен VH3 тяжелой цепи антитела человека, содержащий человеческие зародышевые последовательности.
Предпочтительно, указанный вариабельный домен VK1 легкой цепи каппа содержит человеческие зародышевые последовательности SEQ ID N. 3 и SEQ ID N. 4, область CDR3 легкой цепи содержит последовательность SEQ ID N. 5, и вариабельный домен VH3 тяжелой цепи содержит человеческие зародышевые последовательности SEQ ID N. 1 и SEQ ID N. 2.
В следующем предпочтительном варианте осуществления, в библиотеке векторов или генетических упаковок в соответствии с настоящим изобретением, вариабельный домен VH3 тяжелой цепи, содержащий человеческую зародышевую последовательность, соединен с вариабельным доменом VK1 легкой цепи каппа антитела человека, содержащим человеческую зародышевую последовательность, с использованием линкера из SEQ ID N. 6.
В соответствии со следующим вариантом осуществления, основной вектор, используемый для получения библиотеки по настоящему изобретению, имеет SEQ ID N. 8.
В следующем варианте осуществления, библиотеку векторов или генетических упаковок в соответствии с настоящим изобретением используют для отбора антител против антигенов-мишеней.
Предпочтительно, указанная библиотека экспонирована на фаге для отбора против антигенов-мишеней.
В соответствии с настоящим изобретением, библиотека векторов или генетических упаковок имеет разнообразие, ограниченное областью HC-CDR3 и положением, предшествующим HC-CDR3.
Библиотека, включающая разнообразие HC-CDR3, и способы по настоящему изобретению в настоящее время более полно описаны в следующих примерах. Однако, следует отметить, что такие примеры приведены с целью иллюстрации, а не ограничения.
Примеры
Пример 1. Анализ изменчивости последовательности и длины вариабельных доменов из природных антител
Данные высокопроизводительного секвенирования нового поколения для областей петли HC-CDR3 загружали из архива NCBI SRA SRR400158 (Ippolito et al, 2012) и исследовали длину и аминокислотный состав кодированных петель HC-CDR3 (фиг. 2), следуя номенклатуре Kabat (фиг. 1). Распределение длин петли HC-CDR3 имело максимум на длине петли 12 (фигура 2). Аминокислотный состав в каждом положении петли для длин петли HC-CDR3 от 7 до 15 из природных антител показан в таблицах 1a-1i.
Таблица 1a. Аминокислотный состав для петель HC-CDR3 длиной 7 в природных антителах. Под каждым положением в петле (верхний ряд, номенклатура Kabat, положения HC-CDR3 обозначены серым) показаны наблюдаемые частоты в процентах для каждой аминокислоты в этом положении. Аминокислоты с наблюдаемыми частотами менее чем 0,05% не показаны.

Таблица 1b. Аминокислотный состав для петель HC-CDR3 длиной 8 в природных антителах. Под каждым положением в петле (верхний ряд, номенклатура Kabat, положения HC-CDR3 обозначены серым) показаны наблюдаемые частоты в процентах для каждой аминокислоты в этом положении. Аминокислоты с наблюдаемыми частотами менее чем 0,05%, не показаны.
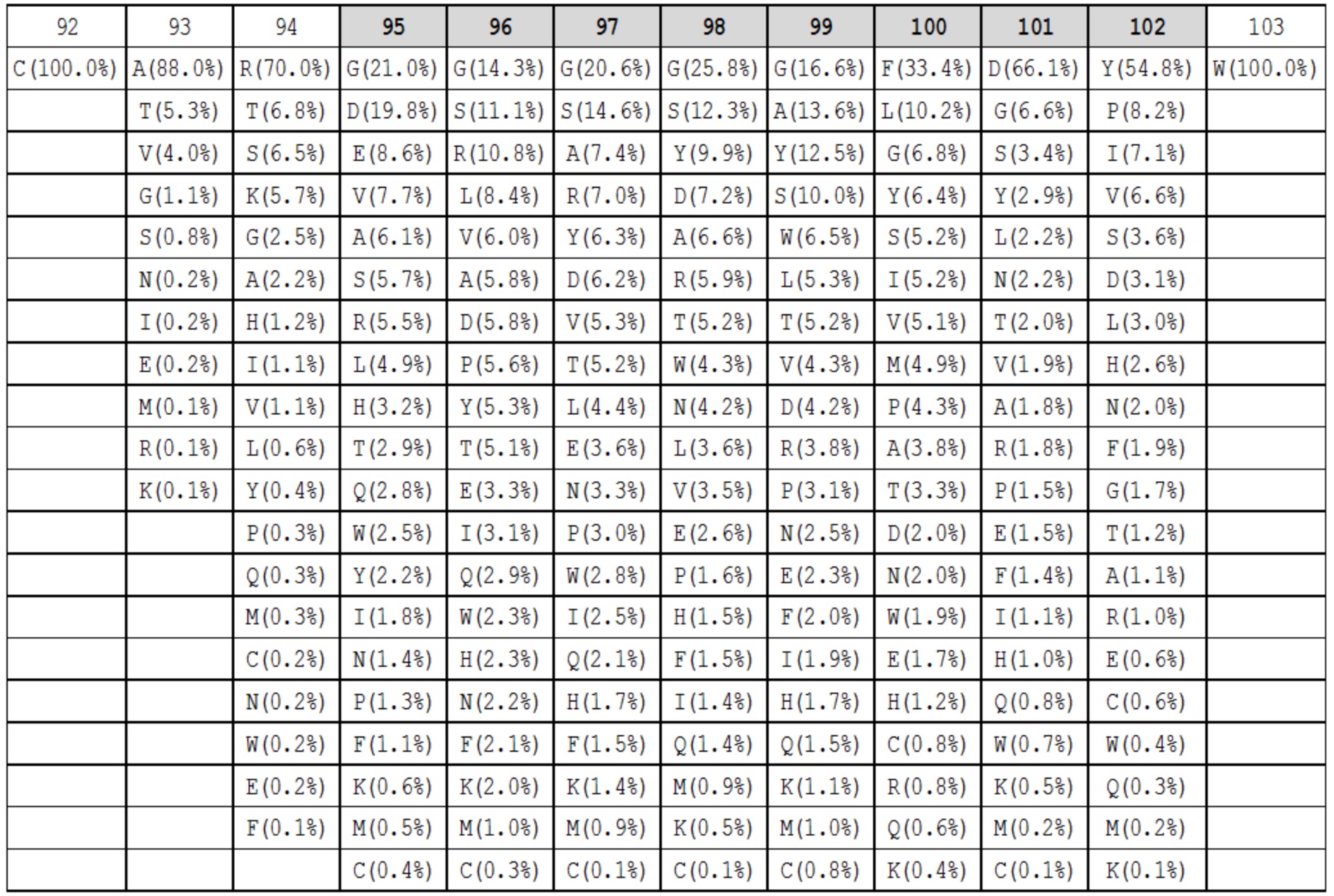
Таблица 1c. Аминокислотный состав для петель HC-CDR3 длиной 9 в природных антителах. Под каждым положением в петле (верхний ряд, номенклатура Kabat, положения HC-CDR3 обозначены серым) показаны наблюдаемые частоты в процентах для каждой аминокислоты в этом положении. Аминокислоты с наблюдаемыми частотами менее чем 0,05%, не показаны.
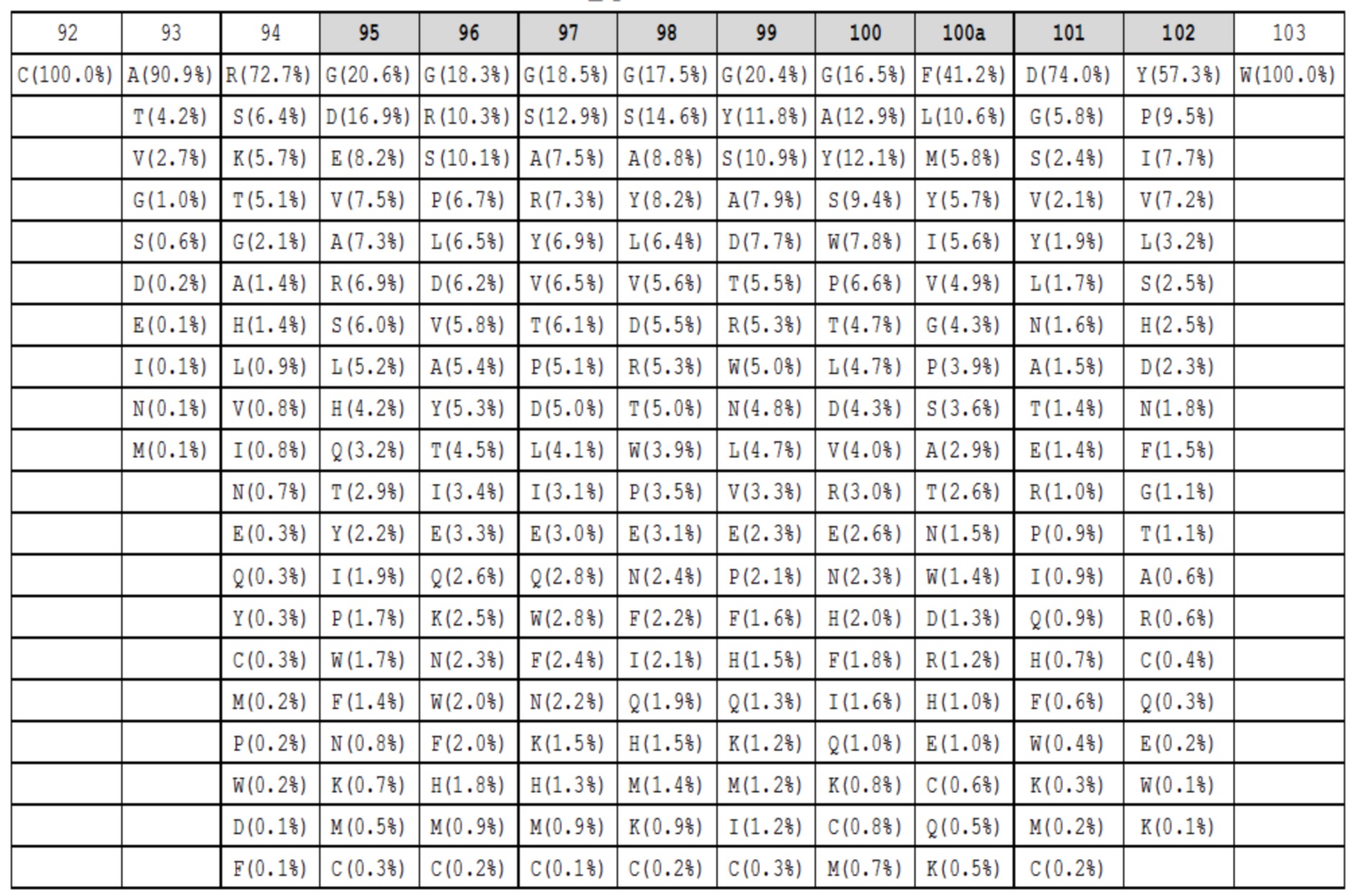
Таблица 1d. Аминокислотный состав для петель HC-CDR3 длиной 10 в природных антителах. Под каждым положением в петле (верхний ряд, номенклатура Kabat, положения HC-CDR3 обозначены серым) показаны наблюдаемые частоты в процентах для каждой аминокислоты в этом положении. Аминокислоты с наблюдаемыми частотами менее чем 0,05%, не показаны.

Таблица 1e. Аминокислотный состав для петель HC-CDR3 длиной 11 в природных антителах. Под каждым положением в петле (верхний ряд, номенклатура Kabat, положения HC-CDR3 обозначены серым) показаны наблюдаемые частоты в процентах для каждой аминокислоты в этом положении. Аминокислоты с наблюдаемыми частотами менее чем 0,05%, не показаны.
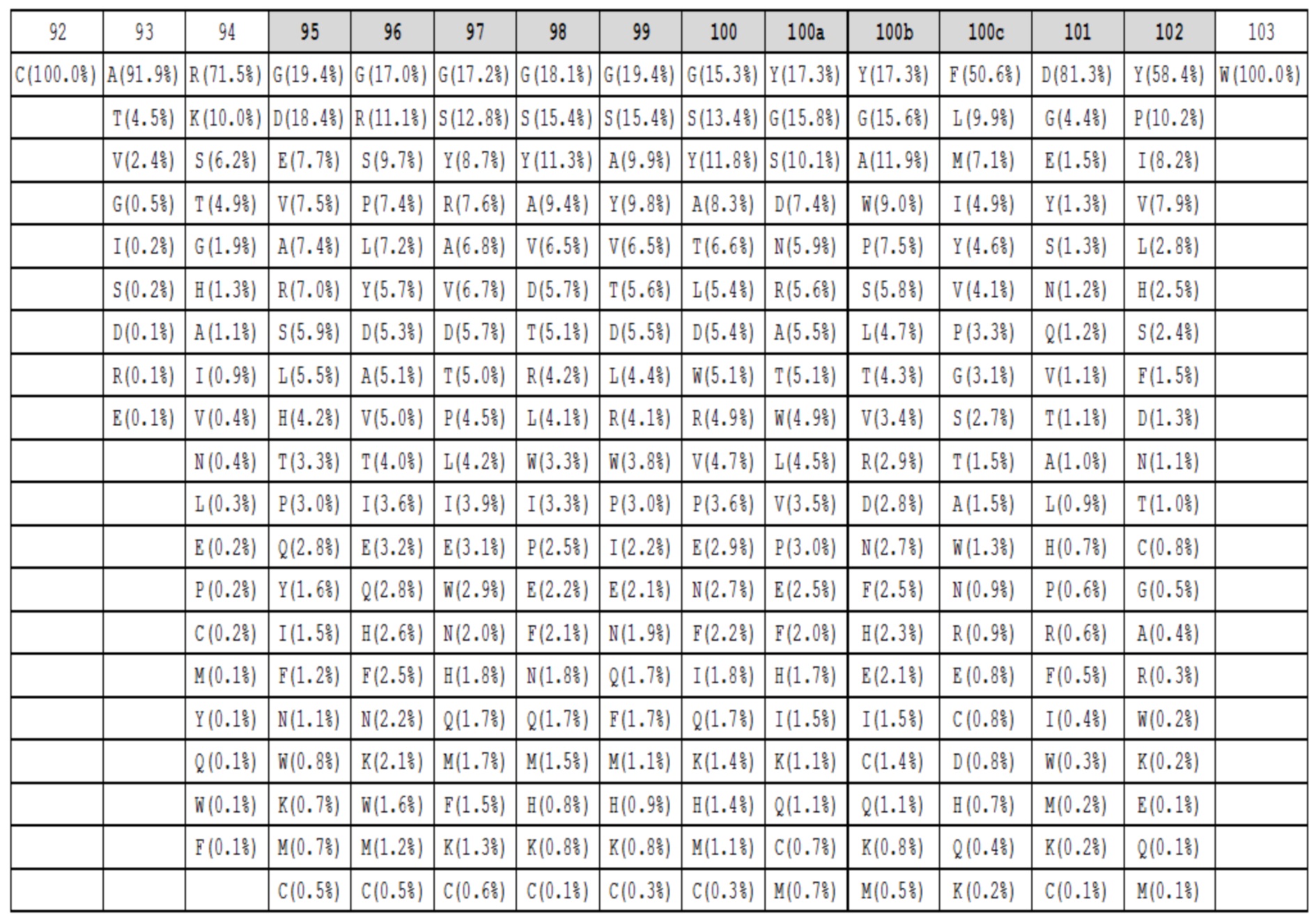
Таблица 1f. Аминокислотный состав для петель HC-CDR3 длиной 12 в природных антителах. Под каждым положением в петле (верхний ряд, номенклатура Kabat, положения HC-CDR3 обозначены серым) показаны наблюдаемые частоты в процентах для каждой аминокислоты в этом положении. Аминокислоты с наблюдаемыми частотами менее чем 0,05%, не показаны.
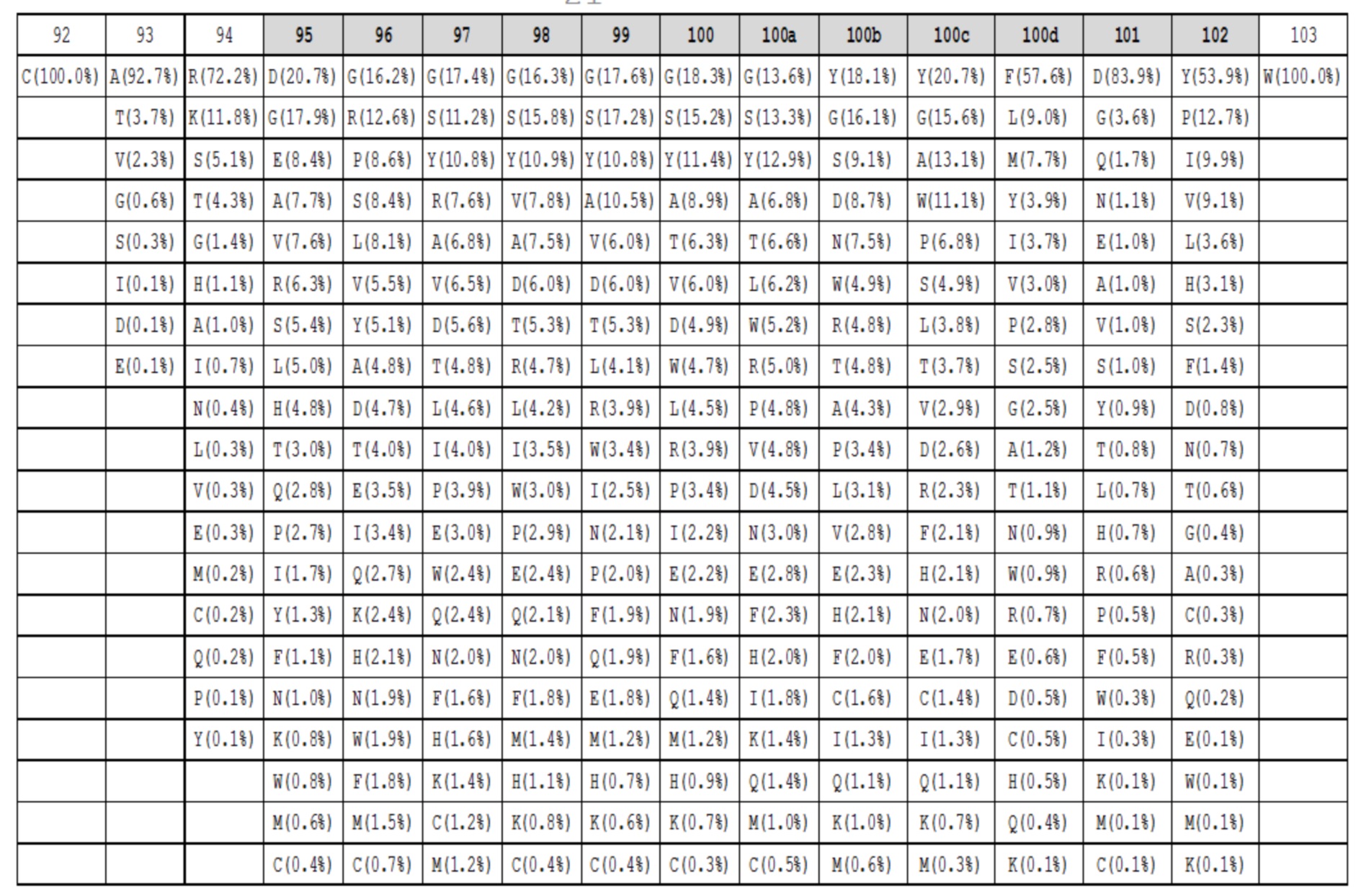
Таблица 1g. Аминокислотный состав для петель HC-CDR3 длиной 13 в природных антителах. Под каждым положением в петле (верхний ряд, номенклатура Kabat, положения HC-CDR3 обозначены серым) показаны наблюдаемые частоты в процентах для каждой аминокислоты в этом положении. Аминокислоты с наблюдаемыми частотами менее чем 0,05%, не показаны.
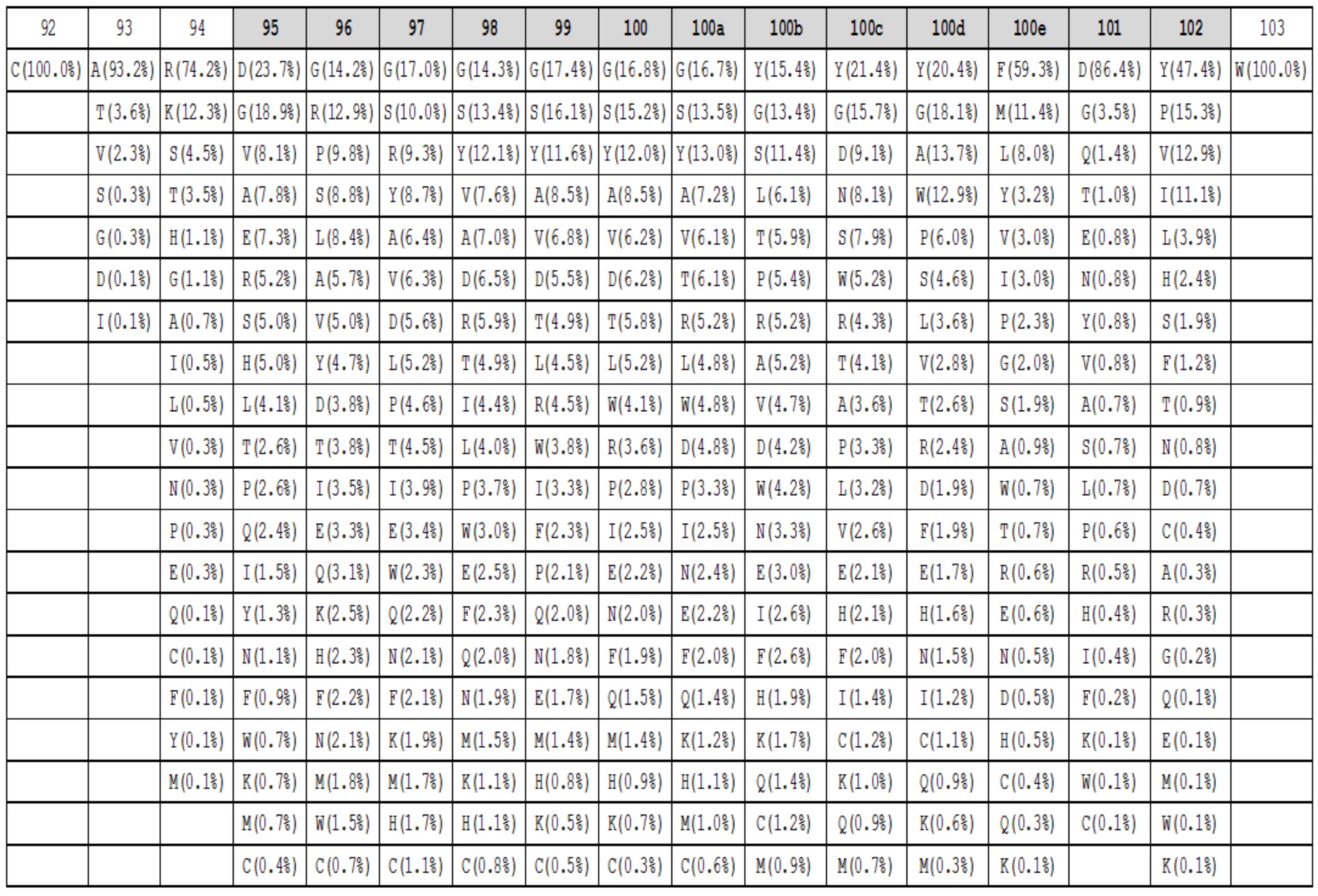
Таблица 1h. Аминокислотный состав для петель HC-CDR3 длиной 14 в природных антителах. Под каждым положением в петле (верхний ряд, номенклатура Kabat, положения HC-CDR3 обозначены серым) показаны наблюдаемые частоты в процентах для каждой аминокислоты в этом положении. Аминокислоты с наблюдаемыми частотами менее чем 0,05%, не показаны.

Таблица 1i. Аминокислотный состав для петель HC-CDR3 длиной 15 в природных антителах. Под каждым положением в петле (верхний ряд, номенклатура Kabat, положения HC-CDR3 обозначены серым) показаны наблюдаемые частоты в процентах для каждой аминокислоты в этом положении. Аминокислоты с наблюдаемыми частотами менее чем 0,05%, не показаны.

Пример 2. Дизайн одноцепочечного каркаса зародышевые VH-линкер-VL
Библиотеки фагового дисплея антител на основе одноцепочечных антител успешно разработаны для большого спектра топологий библиотек (Mondon et al., 2008), и этот формат, таким образом, выбран в качестве формата антитела, в который следует вставлять разнообразие HC-CDR3. Спаривание вариабельных доменов VH3/VK1 является одной из наиболее часто наблюдаемых комбинаций VH/VL в природных антителах (Huang et al., 1996; de Wildt et al., 1999; DeKosky et. al., 2013). Ее наблюдали также с высокой частотой в библиотеке рекомбинантных одноцепочечных антител из природных вариабельных доменов (Glanville et al., 2010), она имеет хорошую термостабильность, а также эффективно экспрессируется (Ewert et al., JMB 2003). Для сборки одноцепочечного каркаса VH3/VK1 выбраны человеческие зародышевые последовательности антител, поскольку зародышевые последовательности имеют преимущественные свойства, такие как отсутствие соматических мутаций, которые могут вызывать иммуногенность при присутствии в терапевтическом антителе, и присущая им устойчивость к присутствию разнообразных областей HC-CDR3. Каркасную аминокислотную последовательность вариабельного домена тяжелой цепи собирали из транслированной зародышевой последовательности M99660 из Genbank (SEQ ID 1) и транслированной зародышевой последовательности J00256 из Genbank (фрагмент J4) (SEQ ID 2). В этом каркасе область HC-CDR3, включая несколько фланкирующих аминокислот, представлена балластным фрагментом, содержащим уникальный участок EcoRV, позволяющий удаление нерасщепленного вектора в ходе клонирования библиотеки (фигура 3). Балластный фрагмент содержит участок PstI на 5'-конце и участок StyI на 3'-конце, позволяющие вырезание фрагмента в ходе получения библиотеки и клонирвание разнообразных олигонуклеотидов, кодирующих HC-CDR3 (фигура 4). PstI (CTGCAG) и StyI (CCWWGG) выбраны, поскольку только немногие комбинации кодирующих кодоны тримерных блоков (фигура 5), используемых для синтеза олигонуклеотидов, кодирующих разнообразие HC-CDR3, образуют участки узнавания PstI и StyI. Тримерные блоки His и Gln (CAT и CAG), таким образом, впоследствии исключали из дизайна олигонуклеотиов, кодирующих разнообразие HC-CDR3. Каркасную аминокислотную последовательность вариабельного домена легкой цепи собирали из транслированной зародышевой последовательности X93627 из Genbank (SEQ ID 3) и зародышевой последовательности J00242 из Genbank (фрагмент VK-1 J1) (SEQ ID 4). Выбрана длина петли LC-CDR3 9 аминокислот, наиболее часто наблюдаемая длина в комбинациях VH3/VK1 тяжелой цепи/легкой цепи в природных антителах (DeKosky et al., 2013). Аминокислотная последовательность петли LC-CDR3 представлена наиболее часто наблюдаемыми аминокислотами в петлях LC-CDR3 длиной 9 в комбинациях VH3/VK1 (SEQ ID 5). В аминокислотной последовательности петли LC-CDR3 инициирующий кодон из фрагмента VK-1 J1 (TGG), кодирующий Trp, заменен на CTG (Leu), наиболее часто наблюдаемую аминокислоту в этом положении в петлях LC-CDR3 длиной 9 в природных антителах с комбинациями VH3/VK1 (DeKosky et al., 2013). Дополнительные уникальные участки рестрикции вводили выше и ниже области легкой цепи LC-CDR1 и LC-CDR3 (фигура 3). Затем одноцепочечный каркас собирали в одноцепочечной топологии VH/VL посредством соединительного линкера GGGGSGGGGSGGGGS (SEQ ID 6) (фигура 3), нежелательные участки рестрикции удаляли посредством выбора альтернативных кодонов, и полученную нуклеотидную последовательность подвергали оптимизации кодонного состава для экспрессии в E.coli (SEQ ID 7).
Пример 3. Дизайн и получение вектора для дисплея BaseVector_VH3_VK1_v22, позволяющего получение библиотеки одноцепочечных только-HC-CDR3
Вектор для фагового дисплея для конструирования библиотеки основан на векторе pCANTAB6, производном вектора pCANTAB5 (McCafferty et al., 1994). Последовательность pCANTAB6 реконструировали, начиная с записи U14321 в Genbank, вводя модификации, описанные в McCafferty et al., 1994. Участок клонирования scFv в pCANTAB6 заменяли на одноцепочечный каркас VH3-линкер-VK1, включающий балластный фрагмент (фигура 6). Введение нескольких замен отдельных оснований для удаления нежелательных участков рестрикции и восстановления последовательности точки начала репликации, перечисленных ниже, таким образом, привело к получению последовательности BaseVector_VH3_VK1_v22 (SEQ ID 8).
Состав вектора для дисплея BaseVector_VH3_VK1_v22:
Положения 1-2334: из U14321 со следующими модификациями:
320 C->T удаляет участок XhoI
617 G->C удаляет участок PvuI
1379 T->C делает последовательность точки начала репликации идентичной другим фагмидным векторам
2328 C->G удаляет участок StyI
Положения 2335-3366: одноцепочечный каркас VH3-линкер-VK1 (SEQ ID 7).
Положения 3367-3447: фрагмент, соединяющий C-конец вариабельного домена легкой цепи с белком pIII, как показано в McCafferty et al., 1994.
Положения 3448-5540: из U14321 со следующими модификациями:
4029 G->A удаляет участок BamHI
4788 A -> C удаляет участок PvuI
Схематическая топология BaseVector_VH3_VK1_v22 с вставленным одноцепочечным каркасом показана на фигуре 6. Вырезание балластной области с использованием PstI и StyI позволяет клонирование олигонуклеотидов, кодирующих разнообразие HC-CDR3. Дополнительные уникальные участки рестрикции позволяют вырезание полного одноцепочечного антитела, отдельных вариабельных легких или тяжелых цепей, или введение дополнительного разнообразия в вариабельный домен легкой цепи (фигура 7). Затем BaseVector_VH3_VK1_v22 синтезировали и собирали стандартными способами (Genscript Corporation).
Пример 4. Оптимизация распределения длин петли HC-CDR3 и оптимизация разнообразия аминокислот по положениям внутри каждой петли HC-CDR3.
His и Gln исключали из дизайна разнообразия HC-CDR3, чтобы исключить образование участков PstI и StyI внутри петли HC-CDR3 из-за тримерных блоков His и Gln. Это исключение являлось приемлемым, поскольку выявлено, что в составе последовательностей HC-CDR3 природного антитела His и Gln, как правило, наблюдали только с небольшой частотой (таблицы 1a -1i). Cys исключали в любом положении петли HC-CDR3, чтобы избежать формирования межмолекулярных дисульфидных мостиков посредством неспаренных остатков Cys. Met, подверженный окислению, также исключали в любом из положений. Met, как правило, присутствует только с очень низкой частотой в природных последовательностях HC-CDR3 (таблицы 1a-1i), за исключением положения, предшествующего положению 101. Положение 101 всегда сохраняли фиксированным как Asp.
Дизайн разнообразия петли HC-CDR3 для длины петли от 7 до 15 (распределения длин петли и разнообразия аминокислот по положениям) оптимизировали с использованием приложения для работы с электронными таблицами. В этом приложении можно корректировать процентная доля для каждой длины петли HC-CDR3 в библиотеке и разнообразие (количество различных аминокислот) в каждом из положений HC-CDR3 (включая также положение 94, предшествующее петле HC-CDR3) для каждой длины петли HC-CDR3. Затем приложение рассчитывает, для каждой длины петли HC-CDR3, теоретически возможное количество вариантов, количество фактически присутствующих клонов (= процентная доля конкретной длины петли HC-CDR3, умноженная на общую сложность библиотеки), оценка Пуассона доли из всех теоретически возможных вариантов, фактически присутствующей, фактическое количество различных вариантов, присутствующих в соответствии с оценкой Пуассона, и избыточность (количество раз, которое в среднем присутствует каждый вариант).
Оценка Пуассона: 1 - e(-1 * N/M),
где
N=количество клонов с петлей HC-CDR3 длиной L в библиотеке со сложностью C
M=количество теоретически возможных вариантов для петли HC-CDR3 длиной L с конкретным составом аминокислот в каждом положении петли
Исходно количество различных аминокислот в каждом положении петли HC-CDR3 устанавливали на 16 (все аминокислоты, за исключением Cys, Gln, His и Met), кроме положений 101 и 102 (5 и 8 различных аминокислот, соответственно), и исходная процентная доля каждой длины петли HC-CDR3 представляла собой распределение длин петли HC-CDR3, наблюдаемое в природных антителах для длин петли от 7 до 15, повторно нормализованное до 100%. В этой конфигурации более короткие петли HC-CDR3 представлены слишком часто, и для длин петли HC-CDR3 от 10 до 15, менее чем 1% от теоретически возможных вариантов присутствует в библиотеке (таблица 2A). Для петель HC-CDR3 длиной 10, по которым обогащены терапевтические антитела или антитела в клинической разработке (фиг. 8-9), только очень малая доля из всех возможных вариантов фактически присутствует в библиотеке.
Таблица 2A. Схема дизайна библиотеки только-HC-CDR3 с общей сложностью 1,3×1010, с распределением длин петли HC-CDR3, подобным распределению природных аминокислот, с 16 различными аминокислотами в гипервариабельных положениях, 5 различными аминокислотами в положении 101, и 8 различными аминокислотами в положении 102. Положения, считаемые гипервариабельными, имеют подчеркнутое количество различных аминокислот. Положение 94 по Kabat не изменяли.
Уменьшение количества различных аминокислот от 16 до 8 в гипервариабельных положениях приводит к получению библиотеки, в которой петли HC-CDR3 с длиной вплоть до 11 хорошо представлены, однако, в то же самое время, избыточность (среднее количество раз, которое представлен вариант) сильно увеличена (Таблица 2B).
Таблица 2B. Схема дизайна библиотеки только-HC-CDR3 с общей сложностью 1,3×1010, с распределением длин петли HC-CDR3, подобным распределению природных аминокислот, с 8 различными аминокислотами в гипервариабельных положениях, постоянной аминокислотой в положении 101 и 8 различными аминокислотами в положении 102. Положения, считаемые гипервариабельными, имеют подчеркнутое количество различных аминокислот. Положение 94 по Kabat не изменяли.
Коррекция процентных значений для распределения длин петли HC-CDR3, введение дополнительной изменчивости в положении 94 для петель HC-CDR3 длиной от 7 до 11, постепенное уменьшение количества различных аминокислот, присутствовующих в гипервариабельных положениях для более длинных петель HC-CDR3 и уменьшение количества различных аминокислот в положении 102 и положении, предшествующем положению 101, обеспечивает дизайн библиотеки с преимущественными свойствами (Таблица 2C). Избыточность является значительно уменьшенной для коротких петель HC-CDR3, покрытие вариантов для длин петли от 9 до 11 является высоким, и более длинные петли HC-CDR3 также представлены относительно хорошо (Таблица 2C). Оптимизированное распределение длин петли HC-CDR3 показано на фигуре 10 вместе с распределением длин петли HC-CDR3, наблюдаемым в природных антителах и с распределением длин петли HC-CDR3 в терапевтическ антителах, уже одобренных или находящихся в клинической разработке.
Таблица 2C. Схема дизайна библиотеки только-HC-CDR3 с общей сложностью 1,3×1010, с оптимизированным распределением длин петли HC-CDR3 и с оптимизированной изменчивостью в положении 94 по Kabat и в каждом положении каждой петли HC-CDR3. Положения, считаемые гипервариабельными, имеют подчеркнутое количество различных аминокислот.
Пример 5. Дизайн олигонуклеотидов в соответствии с оптимизированным дизайном библиотеки
Аминокислотный состав для каждой длины петли HC-CDR3 компилировали на основании оптимизированного дизайна библиотеки, показанного в таблице 2C. В каждом положении и для каждой длины петли HC-CDR3 количество аминокислот, присутствующих в таблице 2C, выбирали из наиболее часто наблюдаемых аминокислот в природных антителах для этой длины петли HC-CDR3 и положения в соответствии с таблицами 1a-1i, и процентные значения повторно нормализовали до 100. Аминокислотный состав и частоты в процентах для HC-CDR3 длиной от 7 до 15 в соответствии с оптимизированным дизайном библиотеки, показаны в таблицах 3A-3I. Например, для петли HC-CDR3 длиной 7 в таблице 2C указано, что 3 различные аминокислоты должны присутствовать в положении 94. 3 наиболее часто наблюдаемые аминокислоты в природных петлях HC-CDR3 длиной 7 представляют собой Arg 63,4%, Ser 8,9% и Thr 7,3% (Таблица 1a). После повторной нормализации до 100%, процентными значениями для этих аминокислот становятся Arg 79,6%, Ser 11,2% и Thr 9,2%.
Таблица 3A. Оптимизированный аминокислотный состав библиотеки для петли HC-CDR3 с длиной L=7. Для каждого положения различные присутствующие аминокислоты показаны вместе с их относительными частотами. Для увеличения общей изменчивости, положение 94, предшествующее петле HC-CDR3, также изменяли для петли HC-CDR3 длиной 7.
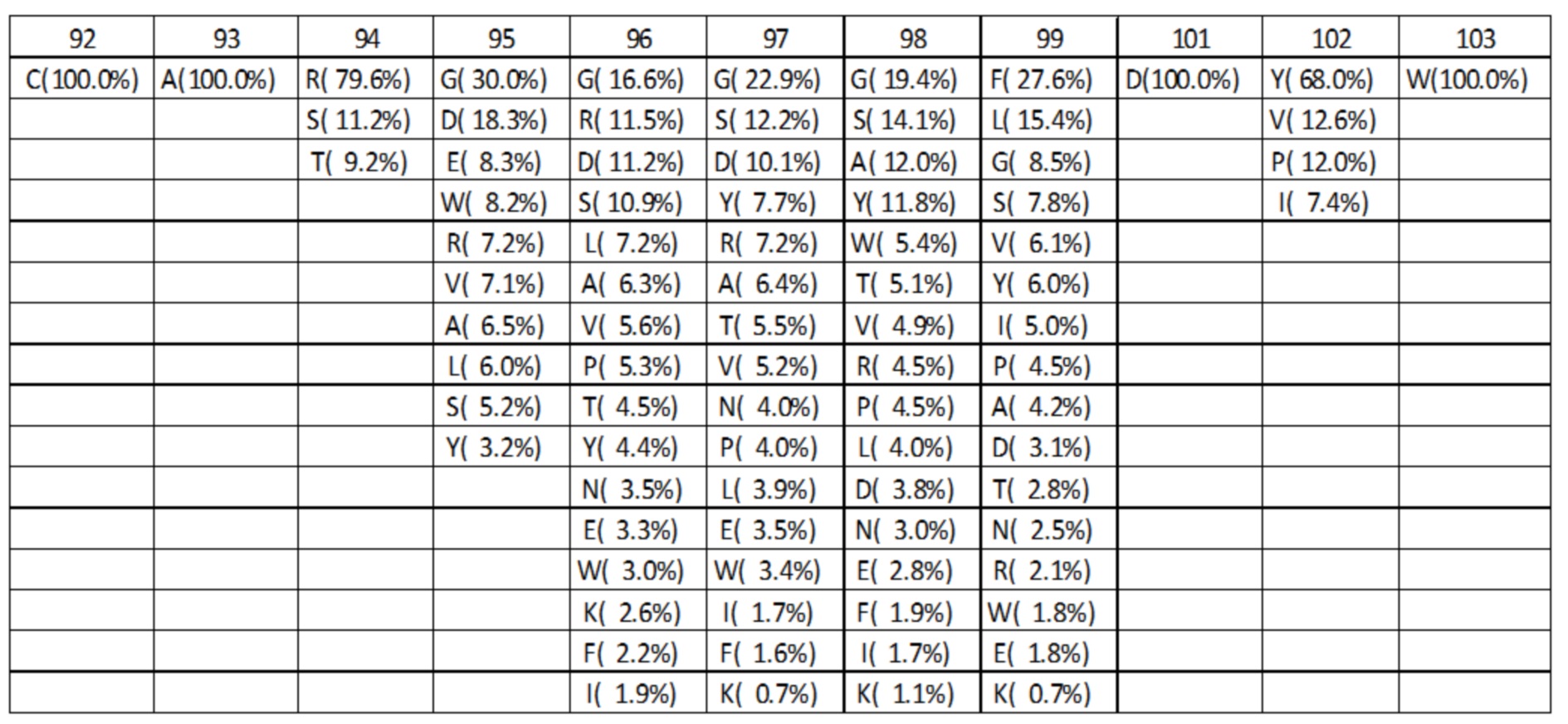
Таблица 3B. Оптимизированный аминокислотный состав библиотеки для петли HC-CDR3 с длиной L=8. Для каждого положения различные присутствующие аминокислоты показаны вместе с их относительными частотами. Для увеличения общей изменчивости, положение 94, предшествующее петле HC-CDR3, также изменяли для петли HC-CDR3 длиной 8.

Таблица 3C. Оптимизированный аминокислотный состав библиотеки для петли HC-CDR3 с длиной L=9. Для каждого положения различные присутствующие аминокислоты показаны вместе с их относительными частотами. Для увеличения общей изменчивости, положение 94, предшествующее петле HC-CDR3, также изменяли для петли HC-CDR3 длиной 9.

Таблица 3D. Оптимизированный аминокислотный состав библиотеки для петли HC-CDR3 с длиной L=10. Для каждого положения различные присутствующие аминокислоты показаны вместе с их относительными частотами. Для увеличения общей изменчивости, положение 94, предшествующее петле HC-CDR3, также изменяли для петли HC-CDR3 длиной 10.

Таблица 3E. Оптимизированный аминокислотный состав библиотеки для петли HC-CDR3 с длиной L=11. Для каждого положения различные присутствующие аминокислоты показаны вместе с их относительными частотами. Для увеличения общей изменчивости, положение 94, предшествующее петле HC-CDR3, также изменяли для петли HC-CDR3 длиной 11.

Таблица 3F. Оптимизированный аминокислотный состав библиотеки для петли HC-CDR3 с длиной L=12. Для каждого положения различные присутствующие аминокислоты показаны вместе с их относительными частотами.
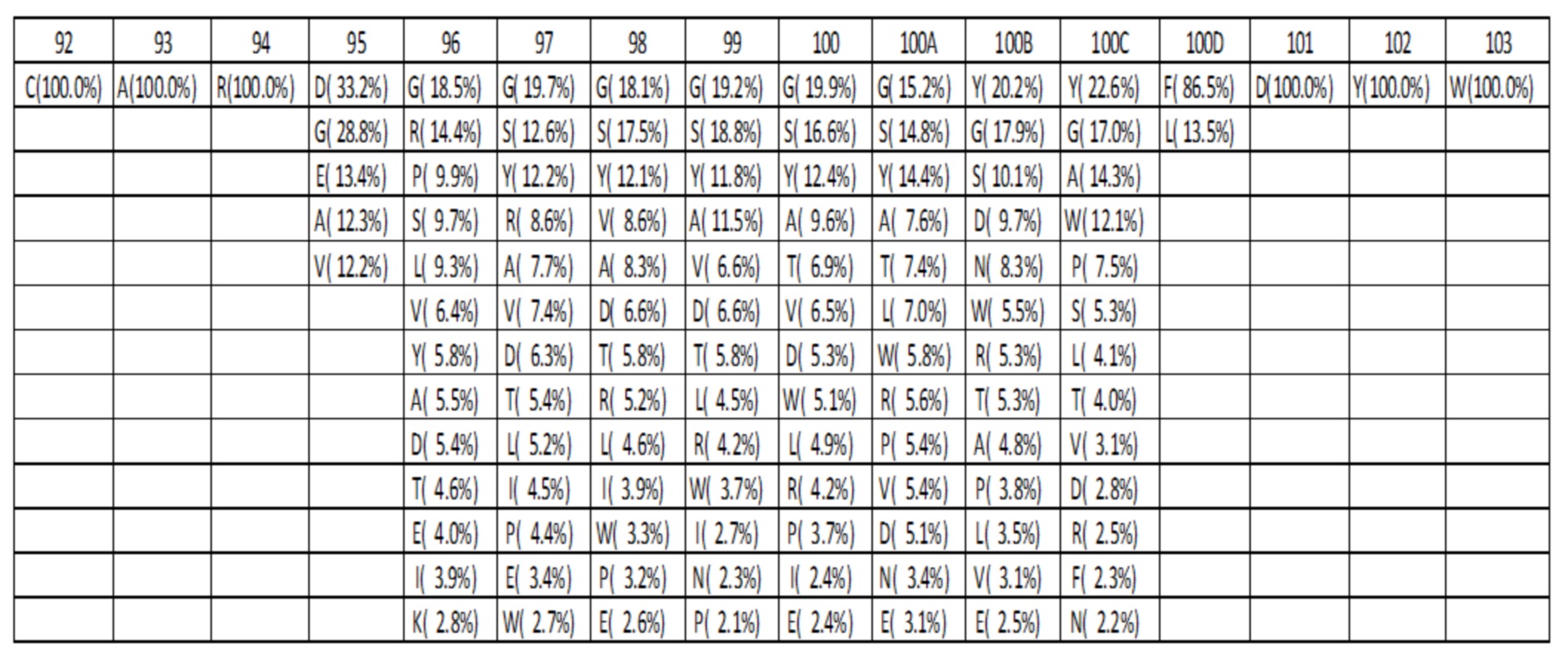
Таблица 3G. Оптимизированный аминокислотный состав библиотеки для петли HC-CDR3 с длиной L=13. Для каждого положения различные присутствующие аминокислоты показаны вместе с их относительными частотами.

Таблица 3H. Оптимизированный аминокислотный состав библиотеки для петли HC-CDR3 с длиной L=14. Для каждого положения различные присутствующие аминокислоты показаны вместе с их относительными частотами.
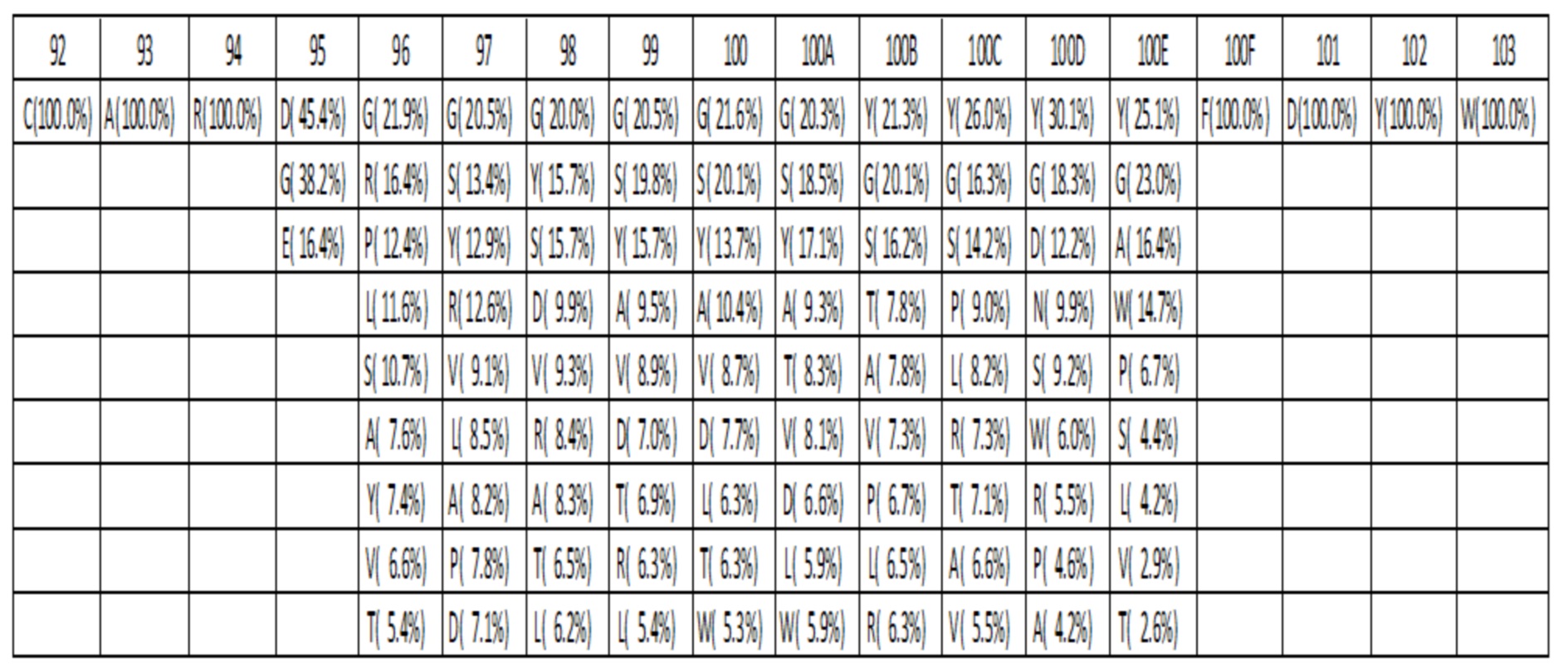
Таблица 3I. Оптимизированный аминокислотный состав библиотеки для петли HC-CDR3 с длиной L=15. Для каждого положения различные присутствующие аминокислоты показаны вместе с их относительными частотами.

Затем для каждой длины петли HC-CDR3 конструировали олигонуклеотиды на основании значений в таблицах 3A-I. В положениях с более чем одной аминокислотой, соответствующие тримерные блоки из фигуры 5выбирали и включали в дизайн с желательной процентной долей в форме смеси. Использование тримерных блоков, кодирующих полные кодоны, обеспечивает преимущества, поскольку они позволяют получать аминокислотный состав и частоты, показанные в таблицах 3A-I, в каждом положении каждой петли HC-CDR3, без образования нежелательных стоп-кодонов или включения нежелательных аминокислот, как было бы в случае стандартных вырожденных олигонуклеотидов. Константные части олигонуклеотидов (положения только с 1 присутствующей аминокислотой или части, необходимые для клонирования) вместо этого разрабатывают для синтеза посредством стандартного синтеза олигонуклеотидов. Дизайн олигонуклеотидов для петли HC-CDR3 длиной 15 показан на фигуре 11. Олигонуклеотиды для других длин петли HC-CDR3 разрабатывали эквивалентным образом с использованием значений, приведенных в таблицах 3A-I. Олигонуклеотиды синтезировали в EllaBiotech с использованием их технологии тримерных блоков.
Пример 6. Получение разнообразия библиотеки посредством вставки олигонуклеотидов, содержащих разнообразие HC-CDR3, в BaseVector_VH3_VK1_22
На первой стадии одноцепочечные олигонуклеотиды, кодирующие разнообразие петли HC-CDR3, подвергали удлинению праймеров (получению двухцепочечных олигонуклеотидов) с использованием праймера Sty_rev_1 (SEQ ID 9) и ДНК-полимеразы Herculase II-Fusion (Agilent, кат.#600679), и следующих условий: денатурация при 98°C в течение 35 с, гибридизация при 47°C в течение 15 с, удлинение в течение 15 с при 47°C и 65°C для олигонуклеотидов, кодирующих петли длиной от 7 до 11 и от 12 до 15, соответственно. Затем полученные двухцепочечные олигонуклеотиды подвергали амплификации с использованием праймеров PstI_for_1 (SEQ ID 10) и Sty_rev_2 (SEQ ID 11) и с использованием ДНК-полимеразы Herculase II-Fusion (Agilent, кат.#600679) в следующих условиях: денатурация при 95°C в течение 15 с, гибридизация при 52°C в течение 15 с, удлинение при 72°C в течение 15 с, повтор в течение 16 циклов.
После амплификации, олигонуклеотиды очищали посредством набора для удаления нуклеотидов Qiaquick (Qiagen, кат.# 28304), подвергали расщеплению с использованием ферментов рестрикции PstI/StyI и лигировали в расщепленный PstI/StyI фагмидный вектор в соотношении вставка:вектор 1:6 (эффективность трансформации в диапазоне 5×107/108 клонов/мкг), отдельно для каждой длины петли HC-CDR3. Затем полученным продуктом лигирования трансформировали электро-компетентные клетки XL1blue-MRF' (50 нг вектора в 50 мкл клеток), рассевали на чашки Bioassay 23×23 см с 2XTY-агаром и выращивали в течение ночи при 37°C. На следующие сутки клетки собирали с планшетов в 2XTYAG/глицерин 17% и хранили при -80°C. Для достижения желательного распределения длин петли HC-CDR3 и общей сложности библиотеки 1,3×1010, эффективность трансформации проверяли регулярно, повторяя цикл сбора клонов, пока не достигали желательного распределения длины петли HC-CDR3 и сложности. Затем посредством пулирования клонов совместно из всех индивидуальных циклов сбора получали конечную библиотеку.
Для получения библиотеки в формате фагов, 6 мл пулированных бактерий инокулировали в 4л 2XTY/ампициллин/2% глюкоза с начальной OD600=0,1 всего для 8,4×1010 бактерий, представляющих приблизительно 6,5-кратную сложность библиотеки. После достижения OD600=0,5, клетки подвергали суперинфекции с использованием фага-помощника M13K07 при MOI=10, среду меняли на 2XTY/ампициллин/канамицин, и клетки инкубировали при встряхивании в течение ночи при 30°C. Полученную культуру клеток центрифугировали, и супернатант, содержащий библиотеку фагов, подвергали двум стадиям преципитации с PEG (добавления 20%PEG8000/NaCl 2,5M до 3/10 объема) с последующим ресуспендированием фагов в 1X TE для стадии дальнейшей очистки в градиенте CsCl. Затем популяцию фагов собирали из градиента и подвергали диализу в течение ночи против 2л 1X TE, чтобы убрать CsCl, концентрацию фагов в препарате определяли по TU, БОЕ и PP/мл и хранили в TBE1X, глицерин 15%, NaN3 0,02%.
Пример 7. Идентификация кандидатов, избирательно связывающих бычий сывороточный альбумин (BSA)
Поскольку на нескольких стадиях отбора стандартный блокирующий буфер изготовлен из 2% молока, и таким образом, содержит много бычьего альбумина, авторы настоящего изобретения заключили, что этот растворимый BSA может конкурировать с рекомбинантным белком BSA - мишенью, которым покрыт пластик для отбора. Следовательно, для отбора фагов на этой мишени, реагент блокирующий буфер с 1% казеином (Roche) использовали вместо стандартного блокирующего буфера. Кроме того, чтобы избежать отбора специфических фагов против NaN3, присутствующего в растворе BSA, использованном для покрытия, использовали реагент блокирующий буфер, содержащий 0,02% NaN3, так чтобы фаги, потенциально специфические для NaN3, оставались в растворе и удалялись при отмывке.
Для этой мишени 2,1×1012 TU (фагов) из библиотеки инкубировали с очищенным BSA, нанесенным в качестве покрытия на иммунологические пробирки, в конечном объеме 3 мл, в концентрации 50 мкг/мл (1-й цикл) и масштабировали с понижением до 30 и 15 мкг/мл, соответственно, в двух следующих циклах отбора.
Соотношение входа/выхода представляет собой показатель обогащения специфических клонов, фактически, оно обычно является очень высоким в первом цикле отбора, и быстро уменьшается в следующих циклах, когда популяция фагов, элюированных в предыдущем цикле, все больше обогащается специфическими фагами. Для этой мишени соотношение входа/выхода позволяло предполагать возможное обогащение специфических клонов от 1-го цикла до 3-го цикла пэннинга (фигура 12A). Поликлональные фаги тестировали посредством фагового ELISA на BSA и нескольких неродственных антигенах. Клоны M13-scFv против BSA присутствовали в 2-м и 3-м циклах отбора (фигура 12B). Неспецифическое связывание с полученными из коммерческих источников антигенами (BP180 и коллагеном VII), нанесенными в качестве покрытия в лунки, возможно, было обусловлено блокирующими реагентами, содержащими BSA, использованными производителем (MBL) (фигура 12B). Процент клонов против BSA в 3-м цикле отбора составлял 50 из 89 (56%) (фигура 12C). Все 12 анализированных положительных клонов являлись специфическими для BSA (фигура 12D).
Пример 8. Идентификация кандидатов, избирательно связывающих овальбумин (OVA)
Для этой мишени отбор положительных фагов из 9,3×1012 TU в качестве общего количества клонов проводили посредством нанесения OVA в качестве покрытия на иммунологические пробирки в концентрации 50 мкг/мл (1-й цикл), затем масштабировали с понижением до 30 и 10 мкг/мл в двух следующих циклах отбора, соответственно. Соотношение входа/выхода позволяло предполагать возможное обогащение специфических клонов от 1-го цикла до 2-го и 3-го циклов пэннинга (фигура 13A). Поликлональные фаги тестировали посредством фагового ELISA на OVA и нескольких неродственных антигенах. Клоны M13-scFv против OVA присутствовали во II и III циклах отбора без какого-либо неспецифического связывания с другими антигенами (фигура 13B). Процент клонов против OVA в третьем цикле отбора составлял 81 из 94 (86%) (фигура 13C). Анализ 10 положительных клонов показал, что все они являлись специфическими для OVA (фигура 13D).
Пример 9. Идентификация кандидатов, избирательно связывающих десмоглеин 1 (Dsg1)
Отбор из библиотеки проводили с использованием десяти предварительно покрытых Dsg1 лунок (набор для ELISA MBL) и 9,3 x1012 TU в 1600 мкл содержащего библиотеку буфера (160 мкл/лунку). Концентрация антигена в покрытии неизвестна. Соотношение входа/выхода позволяло предполагать возможное обогащение специфических клонов от 1-го цикла до 3-го цикла пэннинга (фигура 14A). Поликлональные фаги тестировали посредством фагового ELISA в предварительно покрытых Dsg1 лунках и на нескольких неродственных антигенах. M13-scFv против Dsg1 присутствовали во 2-м и 3-м циклах отбора без какого-либо неспецифического связывания с другими антигенами, за исключением Dsg3 и полученных из Dsg3 конструкций (EC1-2) (фигура 14B). Детектированная перекрестная реакционная способность, вероятно, была обусловлена высокой гомологией последовательности между Dsg1 и 3. Процент клонов против Dsg1 в 3-м цикле отбора составлял 86 из 94 (91%) (фигура 14C). Анализ 8 положительных клонов показал, что все они являлись специфическими для Dsg1 (фигура 14D).
Пример 10. Идентификация кандидатов, избирательно связывающих рецептор 4 фактора роста фибробластов (FGFR4)
Отбор из библиотеки проводили с использованием 1600 мкл библиотеки (160 мкл/лунку) в 10 лунках микропланшета для титрования, покрытых FGFR-4 в концентрации 10 мкг/мл. Все три цикла отбора проводили в лунках, покрытых при одинаковой концентрации FGFR-4. Поскольку рекомбинантный белок FGFR-4 представляет собой химерный белок, слитый с FC IgG1 человека, и чтобы уменьшить количество отобранных фагов против FC, первый цикл проводили с использованием другого рекомбинантного белка, слитого с FC (8 мкг/мл Dsg3-FC). Соотношение входа/выхода позволяло предполагать возможное обогащение специфических клонов от I цикла до III цикла пэннинга (фигура 15A). Поликлональные фаги тестировали посредством фагового ELISA на FGFR-4 и нескольких неродственных антигенах. Клоны M13-scFv против FGFR-4 присутствовали во 2-м и 3-м циклах отбора без какого-либо неспецифического связывания с другими антигенами (фигура 15B). Следует отметить, что конкурентный отбор с использованием Dsg3-FC не исключил все фаги против FC. Фактически, поликлональные смеси вступали в реакцию как с FGFR-4, так и с Dsg3-FC (фигура 15B). Процент клонов против FGFR-4-FC в 3-м цикле отбора составлял 72 из 77 (94%) (фигура 15C). Двадцать анализированных положительных клонов связывались с FGFR-4, и четыре вступали в слабую реакцию также с Dsg3-FC (фигура 15D). Таким образом, с использованием конкурентного биопэннинга с Dsg3-FC достигали уменьшения количества клонов против FC.
Результаты отбора на четырех мишенях обобщены в таблице 4.
Таблица 4 - Обобщение результатов скрининга библиотеки на четырех различных мишенях для проверки. A) Количество и процент фагов, положительно отобранных против каждой мишени; B) Специфичность в отношении связывания со специфической мишенью или другими неродственными рекомбинантными белками, и количество различных последовательностей, обнаруженных для каждой мишени (см. ниже).
A
B
Для всех четырех антигенов-мишеней, при отборах успешно получили большое количество клонов и, что важно, клонов, узнающих антиген-мишень с высокой специфичностью.
Пример 11. Идентификация последовательности ДНК HC-CDR3 отобранных фагов
Как указано в панели B таблицы 4, ряд клонов секвенировали для каждой мишени, и последовательности анализировали и выравнивали, как показано в таблице 5 (см. ниже).
В случае BSA, 5 клонов секвенировали, и для всех из них показали одинаковую последовательность HC-CDR3. В отличие от этого, все 10 секвенированных положительных для OVA клонов представляли собой уникальные клоны и соответствовали по меньшей мере четырем различным семействам клонов. Также, в случае Dsg1, 8 положительных клонов секвенировали, и все из них являлись специфическими, и 5 из 8 являлись уникальными, с наличием трех различных семейств клонов.
Наконец, 10 специфических для FGFR-4 клонов секвенировали и показали, что семь имеют идентичные последовательности. Идентифицировали три различных семейства клонов.
Аффинность клона M13-scFv (кл. 33) для его мишени FGFR-4 определяли посредством фагового ELISA и обнаружили, что она лежит в наномолярном диапазоне (8,7×10-8 M). Это значение при сравнении имеет преимущество над значениями, опубликованными в Pfizer (Mahon et al. J.Mol:Biol. 405, 1712, 2013), где обнаружены аффинности между 105 и 457 нМ для фагов, отобранных с использованием разработанной авторами библиотеки только-HC-CDR3.
Таблица 5. Анализ последовательности для подгруппы клонов, выделенных в ходе отборов. Для каждой мишени секвенировали 5-10 клонов.
Заключение
По полученным результатам, дизайн библиотеки являлся удачным для всех четырех тестированных антигенов-мишеней. Помимо полученной высокой специфичности для мишеней (Таблица 4), библиотека обеспечивает также множество отдельных семейств последовательностей клонов для каждой мишени. Важно, что полученные результаты также отражают основной принцип дизайна библиотеки с большинством последовательностей, имеющих HC-CDR3 длиной 11. Кроме того, присутствие ряда отобранных клонов только с небольшим количеством (часто только 1) различных аминокислот подтверждает, что библиотека обеспечивает большое перекрывание всех вариантов петли HC-CDR3 для наиболее важной длины HC-CDR3 от 9 до 11, как реализовано в дизайне.
Список литературы
- Barbas CF 3rd, Bain JD, Hoekstra DM, Lerner RA. Semisynthetic combinatorial antibody libraries: a chemical solution to the diversity problem. Proc Natl Acad Sci U S A. 1992 May 15;89(10):4457-61.
- Braunagel M, Little M. Construction of a semisynthetic antibody library using trinucleotide oligos. Nucleic Acids Res. 1997 Nov 15;25(22):4690-1.
- Chothia C, Lesk AM. Canonical structures for the hypervariable regions of immunoglobulins. J Mol Biol. 1987 Aug 20;196(4):901-17.
- DeKosky BJ, Ippolito GC, Deschner RP, Lavinder JJ, Wine Y, Rawlings BM, Varadarajan N, Giesecke C, Dörner T, Andrews SF, Wilson PC, Hunicke-Smith SP, Willson CG, Ellington AD, Georgiou G.: High-throughput sequencing of the paired human immunoglobulin heavy and light chain repertoire. Nat Biotechnol. 2013 Feb;31(2):166-9.
- de Wildt RM1, Hoet RM, van Venrooij WJ, Tomlinson IM, Winter G. Analysis of heavy and light chain pairings indicates that receptor editing shapes the human antibody repertoire. J Mol Biol. 1999 Jan 22;285(3):895-901.
- Ewert S1, Huber T, Honegger A, Plückthun A. Biophysical properties of human antibody variable domains. Mol Biol. 2003 Jan 17;325(3):531-53.
- Fellouse FA, Esaki K, Birtalan S, Raptis D, Cancasci VJ, Koide A, Jhurani P, Vasser M, Wiesmann C, Kossiakoff AA, Koide S, Sidhu SS. High-throughput generation of synthetic antibodies from highly functional minimalist phage-displayed libraries. J Mol Biol. 2007 Nov 2;373(4):924-40.
- Glanville J1, Zhai W, Berka J, Telman D, Huerta G, Mehta GR, Ni I, Mei L, Sundar PD, Day GM, Cox D, Rajpal A, Pons J. Precise determination of the diversity of a combinatorial antibody library gives insight into the human immunoglobulin repertoire. Proc Natl Acad Sci U S A. 2009 Dec 1;106(48):20216-21.
- Green LL. Transgenic mouse strains as platforms for the successful discovery and development of human therapeutic monoclonal antibodies. Curr Drug Discov Technol. 2014 Mar;11(1):74-84.
- Hoet RM, Cohen EH, Kent RB, Rookey K, Schoonbroodt S, Hogan S, Rem L, Frans N, Daukandt M, Pieters H, van Hegelsom R, Neer NC, Nastri HG, Rondon IJ, Leeds JA, Hufton SE, Huang L, Kashin I, Devlin M, Kuang G, Steukers M, Viswanathan M, Nixon AE, Sexton DJ, Hoogenboom HR, Ladner RC. Generation of high-affinity human antibodies by combining donor-derived and synthetic complementarity-determining-region diversity. Nat Biotechnol. 2005 Mar;23(3):344-8.
- Huang, S. C., Jiang, R., Glas, A. M. & Milner, E. C. Non-stochastic utilization of Ig V region genes in unselected human peripheral B cells. Mol. Immunol. 1996 33, 553-560.
- Ippolito GC1, Hoi KH, Reddy ST, Carroll SM, Ge X, Rogosch T, Zemlin M, Shultz LD, Ellington AD, Vandenberg CL, Georgiou G. Antibody repertoires in humanized NOD-scid-IL2Rγ(null) mice and human B cells reveals human-like diversification and tolerance checkpoints in the mouse. PLoS One. 2012;7(4):e35497.
- Kabat EA. Sequence of proteins of immunological interest. Bethesda: National Institute of Health; 1991, Vol.1.Fifth Ed.
- Knappik A, Ge L, Honegger A, Pack P, Fischer M, Wellnhofer G, Hoess A, Wölle J, Plückthun A, Virnekäs B. Fully synthetic human combinatorial antibody libraries (HuCAL) based on modular consensus frameworks and CDRs randomized with trinucleotides. J Mol Biol. 2000 Feb 11;296(1):57-86.
- Mahon CM, Lambert MA, Glanville J, Wade JM, Fennell BJ, Krebs MR, Armellino D, Yang S, Liu X, O'Sullivan CM, Autin B, Oficjalska K, Bloom L, Paulsen J, Gill D, Damelin M, Cunningham O, Finlay WJ.: Comprehensive interrogation of a minimalist synthetic CDR-H3 library and its ability to generate antibodies with therapeutic potential. J Mol Biol. 2013 May 27;425(10):1712-30.
- McCafferty J1, Fitzgerald KJ, Earnshaw J, Chiswell DJ, Link J, Smith R, Kenten J. Selection and rapid purification of murine antibody fragments that bind a transition-state analog by phage display. Appl Biochem Biotechnol. 1994 May-Jun;47(2-3):157-71
- Mondon P, Dubreuil O, Bouayadi K, Kharrat H. Human antibody libraries: a race to engineer and explore a larger diversity. Front Biosci. 2008 Jan 1;13:1117-29.
- Pini A, Viti F, Santucci A, Carnemolla B, Zardi L, Neri P, Neri D. Design and use of a phage display library. Human antibodies with subnanomolar affinity against a marker of angiogenesis eluted from a two-dimensional gel. J Biol Chem. 1998 Aug 21;273(34):21769-76.
- Philibert P, Stoessel A, Wang W, Sibler AP, Bec N, Larroque C, Saven JG, Courtête J, Weiss E, Martineau P.: A focused antibody library for selecting scFvs expressed at high levels in the cytoplasm. BMC Biotechnol. 2007 Nov 22;7:81.
- Prassler J, Thiel S, Pracht C, Polzer A, Peters S, Bauer M, Nörenberg S, Stark Y, Kölln J, Popp A, Urlinger S, Enzelberger M. HuCAL PLATINUM, a synthetic Fab library optimized for sequence diversity and superior performance in mammalian expression systems. J Mol Biol. 2011 Oct 14;413(1):261-78.
- Silacci, M., Brack, S., Schirru, G., Marlind, J., Ettorre, A., Merlo, A., Viti, F., Neri, D.: Design, construction, and characterization of a large synthetic human antibody phage display library. Proteomics. 2005 Jun;5(9):2340-50.
- Tonegawa S. Somatic generation of antibody diversity. Nature. 1983 Apr 14;302(5909):575-81.
- Zemlin M, Klinger M, Link J, Zemlin C, Bauer K, Engler JA, Schroeder HW Jr, Kirkham PM.: Expressed murine and human CDR-H3 intervals of equal length exhibit distinct repertoires that differ in their amino acid composition and predicted range of structures. J Mol Biol. 2003 Dec 5;334(4):733-49.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Italfarmaco
<120> Библиотека антител
<150> EP16187884.8
<151> 2016-09-08
<160> 168
<170> BiSSAP 1.2
<210> 1
<211> 97
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> Транслированный зародышевый фрагмент V3-23 VH3 тяжелой цепи
<400> 1
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala
<210> 2
<211> 11
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> Транслированный зародышевый фрагмент J4 тяжелой цепи
<400> 2
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 3
<211> 88
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> Транслированный зародышевый фрагмент VK1 легкой цепи каппа
<400> 3
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys
85
<210> 4
<211> 11
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> Транслированный зародышевый фрагмент J1 VK1 легкой цепи каппа
<400> 4
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
1 5 10
<210> 5
<211> 9
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> CDR3 легкой цепи каппа
<400> 5
Gln Gln Ser Tyr Ser Thr Pro Leu Thr
1 5
<210> 6
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> линкер VH/VL
<400> 6
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 7
<211> 1032
<212> ДНК
<213> Homo sapiens
<220>
<221> источник
<222> 1..1032
<223> /тип_молекулы="необозначенная ДНК"
/примечание="Каркас, включающий балласт"
/организм="Homo sapiens"
<400> 7
gaagttcagc tgctggaatc tggtggtggt ctggttcagc cgggtggttc tctgcgtctg 60
tcttgcgctg cttctggttt caccttctct tcttacgcta tgtcttgggt tcgtcaggct 120
ccgggtaaag gtctggaatg ggtttctgct atctctggtt ctggtggttc tacctactac 180
gctgactctg ttaaaggtcg tttcaccatc tctcgtgaca actctaaaaa caccctgtac 240
ctgcaaatga actctctgcg tgctgaagac actgcagggc actaaatatg taacacactc 300
aatatcaaca tgacctcaaa cacaggctct tacaaaggta gaagaaattt tagttatgga 360
aaattgagct atgctaattg ttcccatagt ggaagtttga actgaagtcg tgcgcagaac 420
atcaagggca gtagaaactt tctatatcac gcaaggacat cgatatcgaa gcccgtaccg 480
tgagaacttt ttcagtacgg caaagtatac taggcctatt gcccttttcg taacttgtgc 540
gtattctctt tcatcactgt tcacaaccag taccttgtcc tcaaaaggtc atcacgttta 600
tttaaattcc cattcgaaag gcatacatcg tagtgccaag gcacactcgt taccgtctca 660
agtggtggcg gaggatccgg aggaggtggc tctggaggtg gcggttcaga catccagatg 720
acccagtctc cgtcttctct gtctgctagc gttggcgatc gtgttaccat cacctgccgt 780
gcttctcagt ctatctcttc ttacctgaac tggtatcagc agaaacccgg gaaagctccg 840
aaactgctga tctacgctgc ttcttctctt cagtctggtg ttccgtctcg tttctctggt 900
tctggttctg gcaccgactt caccctgacc atctcgagcc ttcagccgga agacttcgct 960
acctactact gccagcagtc ttactctacc ccgctgacct tcggtcaggg taccaaagtt 1020
gaaatcaaac gt 1032
<210> 8
<211> 5540
<212> ДНК
<213> Homo sapiens
<220>
<221> источник
<222> 1..5540
<223> /тип_молекулы="другая ДНК"
/примечание="Base vector VH3_VK1_v22"
/организм="Homo sapiens"
<400> 8
gacgaaaggg cctcgtgata cgcctatttt tataggttaa tgtcatgata ataatggttt 60
cttagacgtc aggtggcact tttcggggaa atgtgcgcgg aacccctatt tgtttatttt 120
tctaaataca ttcaaatatg tatccgctca tgagacaata accctgataa atgcttcaat 180
aatattgaaa aaggaagagt atgagtattc aacatttccg tgtcgccctt attccctttt 240
ttgcggcatt ttgccttcct gtttttgctc acccagaaac gctggtgaaa gtaaaagatg 300
ctgaagatca gttgggtgcc cgagtgggtt acatcgaact ggatctcaac agcggtaaga 360
tccttgagag ttttcgcccc gaagaacgtt ttccaatgat gagcactttt aaagttctgc 420
tatgtggcgc ggtattatcc cgtattgacg ccgggcaaga gcaactcggt cgccgcatac 480
actattctca gaatgacttg gttgagtact caccagtcac agaaaagcat cttacggatg 540
gcatgacagt aagagaatta tgcagtgctg ccataaccat gagtgataac actgcggcca 600
acttacttct gacaaccatc ggaggaccga aggagctaac cgcttttttg cacaacatgg 660
gggatcatgt aactcgcctt gatcgttggg aaccggagct gaatgaagcc ataccaaacg 720
acgagcgtga caccacgatg cctgtagcaa tggcaacaac gttgcgcaaa ctattaactg 780
gcgaactact tactctagct tcccggcaac aattaataga ctggatggag gcggataaag 840
ttgcaggacc acttctgcgc tcggcccttc cggctggctg gtttattgct gataaatctg 900
gagccggtga gcgtgggtct cgcggtatca ttgcagcact ggggccagat ggtaagccct 960
cccgtatcgt agttatctac acgacgggga gtcaggcaac tatggatgaa cgaaatagac 1020
agatcgctga gataggtgcc tcactgatta agcattggta actgtcagac caagtttact 1080
catatatact ttagattgat ttaaaacttc atttttaatt taaaaggatc taggtgaaga 1140
tcctttttga taatctcatg accaaaatcc cttaacgtga gttttcgttc cactgagcgt 1200
cagaccccgt agaaaagatc aaaggatctt cttgagatcc tttttttctg cgcgtaatct 1260
gctgcttgca aacaaaaaaa ccaccgctac cagcggtggt ttgtttgccg gatcaagagc 1320
taccaactct ttttccgaag gtaactggct tcagcagagc gcagatacca aatactgtcc 1380
ttctagtgta gccgtagtta ggccaccact tcaagaactc tgtagcaccg cctacatacc 1440
tcgctctgct aatcctgtta ccagtggctg ctgccagtgg cgataagtcg tgtcttaccg 1500
ggttggactc aagacgatag ttaccggata aggcgcagcg gtcgggctga acggggggtt 1560
cgtgcataca gcccagcttg gagcgaacga cctacaccga actgagatac ctacagcgtg 1620
agcattgaga aagcgccacg cttcccgaag ggagaaaggc ggacaggtat ccggtaagcg 1680
gcagggtcgg aacaggagag cgcacgaggg agcttccagg gggaaacgcc tggtatcttt 1740
atagtcctgt cgggtttcgc cacctctgac ttgagcgtcg atttttgtga tgctcgtcag 1800
gggggcggag cctatggaaa aacgccagca acgcggcctt tttacggttc ctggcctttt 1860
gctggccttt tgctcacatg ttctttcctg cgttatcccc tgattctgtg gataaccgta 1920
ttaccgcctt tgagtgagct gataccgctc gccgcagccg aacgaccgag cgcagcgagt 1980
cagtgagcga ggaagcggaa gagcgcccaa tacgcaaacc gcctctcccc gcgcgttggc 2040
cgattcatta atgcagctgg cacgacaggt ttcccgactg gaaagcgggc agtgagcgca 2100
acgcaattaa tgtgagttag ctcactcatt aggcacccca ggctttacac tttatgcttc 2160
cggctcgtat gttgtgtgga attgtgagcg gataacaatt tcacacagga aacagctatg 2220
accatgatta cgccaagctt tggagccttt tttttggaga ttttcaacgt gaaaaaatta 2280
ttattcgcaa ttcctttagt tgttcctttc tatgcggccc agccggcgat ggccgaagtt 2340
cagctgctgg aatctggtgg tggtctggtt cagccgggtg gttctctgcg tctgtcttgc 2400
gctgcttctg gtttcacctt ctcttcttac gctatgtctt gggttcgtca ggctccgggt 2460
aaaggtctgg aatgggtttc tgctatctct ggttctggtg gttctaccta ctacgctgac 2520
tctgttaaag gtcgtttcac catctctcgt gacaactcta aaaacaccct gtacctgcaa 2580
atgaactctc tgcgtgctga agacactgca gggcactaaa tatgtaacac actcaatatc 2640
aacatgacct caaacacagg ctcttacaaa ggtagaagaa attttagtta tggaaaattg 2700
agctatgcta attgttccca tagtggaagt ttgaactgaa gtcgtgcgca gaacatcaag 2760
ggcagtagaa actttctata tcacgcaagg acatcgatat cgaagcccgt accgtgagaa 2820
ctttttcagt acggcaaagt atactaggcc tattgccctt ttcgtaactt gtgcgtattc 2880
tctttcatca ctgttcacaa ccagtacctt gtcctcaaaa ggtcatcacg tttatttaaa 2940
ttcccattcg aaaggcatac atcgtagtgc caaggcacac tcgttaccgt ctcaagtggt 3000
ggcggaggat ccggaggagg tggctctgga ggtggcggtt cagacatcca gatgacccag 3060
tctccgtctt ctctgtctgc tagcgttggc gatcgtgtta ccatcacctg ccgtgcttct 3120
cagtctatct cttcttacct gaactggtat cagcagaaac ccgggaaagc tccgaaactg 3180
ctgatctacg ctgcttcttc tcttcagtct ggtgttccgt ctcgtttctc tggttctggt 3240
tctggcaccg acttcaccct gaccatctcg agccttcagc cggaagactt cgctacctac 3300
tactgccagc agtcttactc taccccgctg accttcggtc agggtaccaa agttgaaatc 3360
aaacgtgcgg ccgcacatca tcatcaccat cacggggccg cagaacaaaa actcatctca 3420
gaagaggatc tgaatggggc cgcatagact gttgaaagtt gtttagcaaa acctcataca 3480
gaaaattcat ttactaacgt ctggaaagac gacaaaactt tagatcgtta cgctaactat 3540
gagggctgtc tgtggaatgc tacaggcgtt gtggtttgta ctggtgacga aactcagtgt 3600
tacggtacat gggttcctat tgggcttgct atccctgaaa atgagggtgg tggctctgag 3660
ggtggcggtt ctgagggtgg cggttctgag ggtggcggta ctaaacctcc tgagtacggt 3720
gatacaccta ttccgggcta tacttatatc aaccctctcg acggcactta tccgcctggt 3780
actgagcaaa accccgctaa tcctaatcct tctcttgagg agtctcagcc tcttaatact 3840
ttcatgtttc agaataatag gttccgaaat aggcagggtg cattaactgt ttatacgggc 3900
actgttactc aaggcactga ccccgttaaa acttattacc agtacactcc tgtatcatca 3960
aaagccatgt atgacgctta ctggaacggt aaattcagag actgcgcttt ccattctggc 4020
tttaatgaag atccattcgt ttgtgaatat caaggccaat cgtctgacct gcctcaacct 4080
cctgtcaatg ctggcggcgg ctctggtggt ggttctggtg gcggctctga gggtggcggc 4140
tctgagggtg gcggttctga gggtggcggc tctgagggtg gcggttccgg tggcggctcc 4200
ggttccggtg attttgatta tgaaaaaatg gcaaacgcta ataagggggc tatgaccgaa 4260
aatgccgatg aaaacgcgct acagtctgac gctaaaggca aacttgattc tgtcgctact 4320
gattacggtg ctgctatcga tggtttcatt ggtgacgttt ccggccttgc taatggtaat 4380
ggtgctactg gtgattttgc tggctctaat tcccaaatgg ctcaagtcgg tgacggtgat 4440
aattcacctt taatgaataa tttccgtcaa tatttacctt ctttgcctca gtcggttgaa 4500
tgtcgccctt atgtctttgg cgctggtaaa ccatatgaat tttctattga ttgtgacaaa 4560
ataaacttat tccgtggtgt ctttgcgttt cttttatatg ttgccacctt tatgtatgta 4620
ttttcgacgt ttgctaacat actgcgtaat aaggagtctt aataagaatt cactggccgt 4680
cgttttacaa cgtcgtgact gggaaaaccc tggcgttacc caacttaatc gccttgcagc 4740
acatccccct ttcgccagct ggcgtaatag cgaagaggcc cgcaccgctc gcccttccca 4800
acagttgcgc agcctgaatg gcgaatggcg cctgatgcgg tattttctcc ttacgcatct 4860
gtgcggtatt tcacaccgca tataaattgt aaacgttaat attttgttaa aattcgcgtt 4920
aaatttttgt taaatcagct cattttttaa ccaataggcc gaaatcggca aaatccctta 4980
taaatcaaaa gaatagcccg agatagggtt gagtgttgtt ccagtttgga acaagagtcc 5040
actattaaag aacgtggact ccaacgtcaa agggcgaaaa accgtctatc agggcgatgg 5100
cccactacgt gaaccatcac ccaaatcaag ttttttgggg tcgaggtgcc gtaaagcact 5160
aaatcggaac cctaaaggga gcccccgatt tagagcttga cggggaaagc cggcgaacgt 5220
ggcgagaaag gaagggaaga aagcgaaagg agcgggcgct agggcgctgg caagtgtagc 5280
ggtcacgctg cgcgtaacca ccacacccgc cgcgcttaat gcgccgctac agggcgcgta 5340
ctatggttgc tttgacgggt gcagtctcag tacaatctgc tctgatgccg catagttaag 5400
ccagccccga cacccgccaa cacccgctga cgcgccctga cgggcttgtc tgctcccggc 5460
atccgcttac agacaagctg tgaccgtctc cgggagctgc atgtgtcaga ggttttcacc 5520
gtcatcaccg aaacgcgcga 5540
<210> 9
<211> 15
<212> ДНК
<213> Homo sapiens
<220>
<221> источник
<222> 1..15
<223> /тип_молекулы="другая ДНК"
/примечание="Sty_rev_1"
/организм="Homo sapiens"
<400> 9
ccgaccttgg cccca 15
<210> 10
<211> 27
<212> ДНК
<213> Homo sapiens
<220>
<221> источник
<222> 1..27
<223> /тип_молекулы="другая ДНК"
/примечание="праймер PstI_for _1"
/организм="Homo sapiens"
<400> 10
cgaaaagcac ctgcagtgta ttactgc 27
<210> 11
<211> 19
<212> ДНК
<213> Homo sapiens
<220>
<221> источник
<222> 1..19
<223> /тип_молекулы="другая ДНК"
/примечание="праймер Sty_rev_2"
/организм="Homo sapiens"
<400> 11
ctgaccgacc tgggcccca 19
<210> 12
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 12
Cys Ala Arg Asp Pro Leu Asp Ile Tyr Ser Trp Gly Tyr Phe Asp Tyr
1 5 10 15
Trp
<210> 13
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 13
Cys Ala Val Val Thr Gly Val Leu Trp Gly Phe Asp Tyr Trp
1 5 10
<210> 14
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 14
Cys Ala Lys Asp Tyr Ala Ser Gly Tyr Gly Tyr Phe Asp Tyr Trp
1 5 10 15
<210> 15
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 15
Cys Ala Lys Asp Tyr Ala Ser Gly Trp Gly Tyr Phe Asp Tyr Trp
1 5 10 15
<210> 16
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 16
Cys Ala Lys Asp Phe Gly Arg Gly Tyr Gly Tyr Phe Asp Tyr Trp
1 5 10 15
<210> 17
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 17
Cys Ala Lys Asp Phe Arg Ser Gly Tyr Gly Tyr Phe Asp Tyr Trp
1 5 10 15
<210> 18
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 18
Cys Ala Gln Asp Val Arg Arg Gly Cys Gly Tyr Phe Asp Tyr Trp
1 5 10 15
<210> 19
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 19
Cys Ala Lys Asp Val Ala Arg Gly Tyr Gly Tyr Phe Asp Tyr Trp
1 5 10 15
<210> 20
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 20
Cys Ala Lys Asp Val Trp Arg Gly Tyr Gly Tyr Phe Asp Tyr Trp
1 5 10 15
<210> 21
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 21
Cys Ala Lys Asp Val Gly Arg Gly Val Gly Tyr Phe Asp Tyr Trp
1 5 10 15
<210> 22
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 22
Cys Ala Lys Val Val Gly Gly Val Leu Tyr Ala Phe Asp Tyr Trp
1 5 10 15
<210> 23
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 23
Cys Ala Arg Gly Ala Gly Tyr Tyr Gly Leu Pro Val Tyr Ala Phe Asp
1 5 10 15
Tyr Trp
<210> 24
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 24
Cys Ala Arg Gly Ala Gly Tyr Ser Gly Leu Pro Val Tyr Ala Phe Asp
1 5 10 15
Tyr Trp
<210> 25
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 25
Cys Ala Arg Gly Ala Gly Tyr Leu Gly Tyr Pro Val Tyr Ala Phe Asp
1 5 10 15
Tyr Trp
<210> 26
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 26
Cys Ala Arg Ala Asn Trp Ala Ser Ser Tyr Glu Phe Asp Tyr Trp
1 5 10 15
<210> 27
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 27
Cys Ala Arg Val Gly Arg Tyr Gly Tyr Tyr Asp Ala Ala Phe Asp Tyr
1 5 10 15
Trp
<210> 28
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 28
Cys Ala Arg Glu Gly Asp Pro Asp Leu Tyr Phe Phe Asp Tyr
1 5 10
<210> 29
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 29
Cys Ala Arg Gly Tyr Ala Trp Pro Ser Trp Phe Asp Tyr Trp
1 5 10
<210> 30
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 30
Cys Ala Arg Glu Trp Trp Asp Tyr Leu Tyr Leu Asp Tyr Trp
1 5 10
<210> 31
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 31
Cys Ala Ser Gly Asp Tyr Trp Gly Gln Gly
1 5 10
<210> 32
<211> 11
<212> БЕЛОК
<213> Homo sapiens
<400> 32
Cys Ala Thr Asn Asp Asp Tyr Trp Gly Gln Gly
1 5 10
<210> 33
<211> 12
<212> БЕЛОК
<213> Homo sapiens
<400> 33
Cys Ala Thr Glu Leu Phe Asp Leu Trp Gly Gln Gly
1 5 10
<210> 34
<211> 12
<212> БЕЛОК
<213> Homo sapiens
<400> 34
Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Arg Gly
1 5 10
<210> 35
<211> 12
<212> БЕЛОК
<213> Homo sapiens
<400> 35
Cys Val Lys Pro Gly Gly Asp Tyr Trp Gly Gln Gly
1 5 10
<210> 36
<211> 12
<212> БЕЛОК
<213> Homo sapiens
<400> 36
Cys Lys Arg Ser Tyr Tyr Gly His Trp Gly His Gly
1 5 10
<210> 37
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 37
Cys Lys Thr His Gly Ser His Asp Asn Arg Gly Gln Gly
1 5 10
<210> 38
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 38
Cys Glu Arg Glu Gly Gly Val Asn Asn Trp Gly Gln Gly
1 5 10
<210> 39
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 39
Cys Ala Arg Ser Pro Ser Gly Phe Asn Trp Gly Gln Gly
1 5 10
<210> 40
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 40
Cys Ala Arg Arg Gln Leu Tyr Phe Asp Tyr Trp Gly Gln Gly
1 5 10
<210> 41
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 41
Cys Ala Gln Asn Leu Glu Ile Pro Arg Thr Phe Gly Gln Gly
1 5 10
<210> 42
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 42
Cys Ala Arg Gly Gly Gly Val Phe Asp Tyr Trp Gly Gln Gly
1 5 10
<210> 43
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 43
Cys Ala Gln Asn Leu Glu Ile Pro Arg Thr Phe Gly Gln Gly
1 5 10
<210> 44
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 44
Cys Ala Ser Leu Tyr Ser Leu Pro Val Tyr Trp Gly Gln Gly
1 5 10
<210> 45
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 45
Cys Ala Lys Pro Phe Pro Tyr Phe Asp Tyr Trp Gly Gln Gly
1 5 10
<210> 46
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 46
Cys Ala Arg Val Thr Asp Ala Phe Asp Ile Trp Gly Gln Gly
1 5 10
<210> 47
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 47
Cys Ala Lys Gly Arg Asp Ser Phe Asp Ile Trp Gly Gln Gly
1 5 10
<210> 48
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 48
Cys Ala Arg Asp Pro Ile Ala Ala Gly Tyr Trp Gly Gln Gly
1 5 10
<210> 49
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 49
Cys Ala Arg Asn Trp Met Asn Phe Asp Tyr Trp Gly Gln Gly
1 5 10
<210> 50
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 50
Cys Val Arg Ile Gly Glu Asp Ala Leu Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 51
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 51
Cys Ser Arg Asp Tyr Gly Tyr Tyr Phe Asp Phe Trp Gly Gln Gly
1 5 10 15
<210> 52
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 52
Cys Ala Asn Tyr Gly Asn Tyr Trp Phe Ala Tyr Trp Gly Gln Gly
1 5 10 15
<210> 53
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 53
Cys Ala Arg Trp Gly Arg Val Phe Phe Asp Trp Trp Gly Gln Gly
1 5 10 15
<210> 54
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 54
Cys Thr Gly Tyr Tyr Ala Asp Ala Met Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 55
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 55
Cys Ala Arg His Leu His Gly Ser Phe Ala Ser Trp Gly Gln Gly
1 5 10 15
<210> 56
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 56
Cys Ala Arg Asp Arg Tyr Tyr Gly Pro Glu Met Trp Gly Gln Gly
1 5 10 15
<210> 57
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 57
Cys Ala Arg Asp Asn Trp Asp Ala Met Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 58
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 58
Cys Ala Ser Trp Ile Ser Asp Phe Phe Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 59
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 59
Cys Gly Arg Ser Lys Arg Asp Tyr Phe Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 60
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 60
Cys Ala Lys Asp Ser Asn Trp Gly Asn Phe Asp Leu Trp Gly Arg Gly
1 5 10 15
<210> 61
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 61
Cys Ala Thr Gly Asp Thr Thr Tyr Lys Phe Asp Phe Trp Gly Gln Gly
1 5 10 15
<210> 62
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 62
Cys Ala Arg Asp Leu Arg Thr Gly Pro Phe Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 63
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 63
Cys Ala Arg Thr Gly Ser Ser Gly Tyr Phe Asp Phe Trp Gly Gln Gly
1 5 10 15
<210> 64
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 64
Cys Ala Arg Asn Tyr Val Gly Ser Ile Phe Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 65
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 65
Cys Ala Arg Gln Met Gly Tyr Trp His Phe Asp Leu Trp Gly Arg Gly
1 5 10 15
<210> 66
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 66
Cys Ala Arg Gly Tyr Asn Trp Asn Tyr Phe Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 67
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 67
Cys Ala Arg Met Arg Lys Gly Tyr Ala Met Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 68
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 68
Cys Ala Arg Glu Ser Tyr Tyr Gly Phe Thr Ser Tyr Trp Gly Gln Gly
1 5 10 15
<210> 69
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 69
Cys Ser Arg Pro Thr Met Leu Pro Trp Phe Ala Tyr Trp Gly Gln
1 5 10 15
<210> 70
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 70
Cys Ala Arg Asp Lys Trp Thr Trp Tyr Phe Asp Leu Trp Gly Arg Gly
1 5 10 15
<210> 71
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 71
Cys Ser Arg Gly Ile Pro Gly Tyr Ala Met Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 72
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 72
Cys Ala Arg Glu Leu Gly Arg Arg Tyr Phe Asp Leu Trp Gly Arg Gly
1 5 10 15
<210> 73
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 73
Cys Ala Arg Tyr Asp Tyr Asn Tyr Ala Met Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 74
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 74
Cys Ala Arg Asp Tyr Gly Asp Tyr Gly Met Asp Tyr Trp Gly Gln Gly
1 5 10 15
<210> 75
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 75
Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 76
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 76
Cys Ala Arg Gly Glu Gly Asn Tyr Ala Trp Phe Ala Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 77
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 77
Cys Val Arg Pro Leu Tyr Asp Tyr Tyr Ala Met Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 78
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 78
Cys Ala Arg Asn Val Phe Asp Gly Tyr Trp Leu Val Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 79
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 79
Cys Ala Arg Glu Gly His Thr Ala Ala Pro Phe Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 80
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 80
Cys Ala Arg Gly Gly Tyr Asp Gly Arg Gly Phe Asp Tyr Trp Gly Ser
1 5 10 15
Gly
<210> 81
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 81
Cys Ala Arg Leu Asp Gly Tyr Tyr Phe Gly Phe Ala Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 82
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 82
Cys Thr Arg Asp Arg Gly Leu Arg Phe Tyr Phe Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 83
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 83
Cys Ala Arg Arg Arg Asn Trp Gly Asn Ala Phe Asp Ile Trp Gly Gln
1 5 10 15
Gly
<210> 84
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 84
Cys Ala Arg Arg Gly Pro Tyr Asn Trp Tyr Phe Asp Val Trp Gly Gln
1 5 10 15
Gly
<210> 85
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 85
Cys Ala Arg Tyr Asp Gly Ile Tyr Gly Glu Leu Asp Phe Trp Gly Gln
1 5 10 15
Gly
<210> 86
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 86
Cys Ala Arg Gly Phe Gly Gly Ser Tyr Gly Phe Ala Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 87
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 87
Cys Ala Arg Asp Tyr Asp Tyr Asp Val Gly Met Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 88
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 88
Cys Ala Arg Pro Asp Gly Asn Tyr Trp Tyr Phe Asp Val Trp Gly Gln
1 5 10 15
Gly
<210> 89
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 89
Cys Ala Arg His Gly Asp Asp Pro Ala Trp Phe Ala Tyr Trp Gly Gln
1 5 10 15
<210> 90
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 90
Cys Ser Thr Ile Phe Gly Val Val Thr Asn Phe Asp Asn Trp Gly Gln
1 5 10 15
Gly
<210> 91
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 91
Cys Ala Pro Arg Tyr Ser Ser Ser Trp Tyr Leu Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 92
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 92
Cys Ala Lys Val Ala Val Ala Gly Thr His Phe Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 93
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 93
Cys Ala Arg Asn Arg Tyr Asp Pro Pro Trp Phe Val Asp Trp Gly Gln
1 5 10 15
Gly
<210> 94
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 94
Cys Ala Arg His Arg Ser Gly Tyr Phe Ser Met Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 95
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 95
Cys Ala Arg Glu Ala Arg Gly Ser Tyr Ala Phe Asp Ile Trp Gly Gln
1 5 10 15
Gly
<210> 96
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 96
Cys Ala Arg Arg Asp Tyr Asp Leu Asp Tyr Phe Asp Ser Trp Gly Gln
1 5 10 15
Gly
<210> 97
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 97
Cys Ala Arg Tyr Asp Tyr Phe Thr Gly Thr Gly Val Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 98
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 98
Cys Ala Gly Asp Gly Tyr Tyr Pro Tyr Ala Met Asp Asn Trp Gly Gln
1 5 10 15
Gly
<210> 99
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 99
Cys Gly Arg His Ser Asp Gly Asn Phe Ala Phe Gly Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 100
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 100
Cys Ala Arg Asp Met Ile Phe Asn Phe Tyr Phe Asp Val Trp Gly Gln
1 5 10 15
Gly
<210> 101
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 101
Cys Thr Arg Thr Thr Leu Ile Ser Val Tyr Phe Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 102
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 102
Cys Ala Thr Tyr Arg Ser Tyr Val Thr Pro Leu Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 103
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 103
Cys Ala Lys Phe Arg Gln Tyr Ser Gly Gly Phe Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 104
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 104
Cys Ala Arg Leu Gly Asn Phe Tyr Tyr Gly Met Asp Val Trp Gly Gln
1 5 10 15
Gly
<210> 105
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 105
Cys Ala Arg Gly Leu Trp Gly Tyr Tyr Ala Leu Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 106
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 106
Cys Ala Arg Gln Leu Trp Gly Tyr Tyr Ala Leu Asp Ile Trp Gly Gln
1 5 10 15
Gly
<210> 107
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 107
Cys Ala Arg Leu Arg Asn Trp Asp Glu Pro Met Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 108
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 108
Cys Ala Arg Arg Asp Gly Asn Tyr Gly Trp Phe Ala Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 109
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 109
Cys Ala Arg Arg Gly Asp Ser Met Ile Thr Thr Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 110
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 110
Cys Val Arg Tyr Tyr Gly Tyr Asp Glu Ala Met Asp Tyr Trp Gly Gln
1 5 10 15
Gly
<210> 111
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 111
Cys Val Lys Gly Gly Tyr Tyr Gly His Trp Tyr Phe Asp Val Trp Gly
1 5 10 15
Ala Gly
<210> 112
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 112
Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 113
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 113
Cys Ala Arg Gly Gly Arg Asp Phe Gly Asp Ser Phe Asp Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 114
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 114
Cys Ala Ser Thr Leu Gly Leu Val Leu Asp Ala Met Asp Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 115
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 115
Cys Ala Arg Leu Ser Pro Gly Gly Tyr Tyr Val Met Asp Ala Trp Gly
1 5 10 15
Gln Gly
<210> 116
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 116
Cys Ala Arg Asp Gly Gly Ile Ala Ala Pro Gly Pro Asp Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 117
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 117
Cys Ala Ile Val Gly Ser Phe Ser Pro Leu Thr Leu Gly Leu Trp Gly
1 5 10 15
Gln Gly
<210> 118
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 118
Cys Ser Arg Ser Arg Gly Lys Asn Glu Ala Trp Phe Ala Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 119
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 119
Cys Ala Arg Ser Thr Met Ile Thr Asn Tyr Val Met Asp Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 120
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 120
Cys Ala Arg Val Ser Ile Phe Gly Val Gly Thr Phe Asp Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 121
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 121
Cys Ala Arg Gly Gly Gly Ser Ile Tyr Tyr Ala Met Asp Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 122
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 122
Cys Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 123
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 123
Cys Ala Arg Ala Asn Asp Gly Val Tyr Tyr Ala Met Asp Tyr Trp Gly
1 5 10 15
Gln Gly
<210> 124
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 124
Cys Gly Arg Glu Gly Ile Arg Tyr Tyr Gly Leu Leu Gly Asp Tyr Trp
1 5 10 15
Gly Gln Gly
<210> 125
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 125
Cys Ala Arg Asp Asp Ser Ser Asp Trp Asp Ala Lys Phe Asn Leu Trp
1 5 10 15
Gly Gln Gly
<210> 126
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 126
Cys Ala Arg Asp Arg Gly Gly Asp Tyr Tyr Tyr Gly Met Asp Val Trp
1 5 10 15
Gly Gln Gly
<210> 127
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 127
Cys Ala Lys Ile Leu Gly Ala Gly Arg Gly Trp Tyr Phe Asp Leu Trp
1 5 10 15
Gly Lys Gly
<210> 128
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 128
Cys Ala Arg Glu Ser Arg Val Ser Phe Glu Ala Ala Met Asp Tyr Trp
1 5 10 15
Gly Gln Gly
<210> 129
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 129
Cys Ala Arg Glu Pro Glu Lys Phe Asp Ser Asp Asp Ser Asp Val Trp
1 5 10 15
Gly Arg Gly
<210> 130
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 130
Cys Ala Arg Ser Thr Tyr Val Gly Gly Asp Trp Gln Phe Asp Val Trp
1 5 10 15
Gly Lys Gly
<210> 131
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 131
Cys Ala Arg Gly Ser His Tyr Phe Gly His Trp His Phe Ala Val Trp
1 5 10 15
Gly Gln Gly
<210> 132
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 132
Cys Ala Arg Ile Arg Val Gly Pro Ser Gly Gly Ala Phe Asp Tyr Trp
1 5 10 15
Gly Gln Gly
<210> 133
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 133
Cys Ala Arg Glu Asp Tyr Tyr Asp Asn Ser Tyr Tyr Phe Asp Tyr Trp
1 5 10 15
Gly Gln Gly
<210> 134
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 134
Cys Ala Arg Gly Gly Tyr Asp Phe Trp Ser Gly Tyr Phe Asp Tyr Trp
1 5 10 15
Gly Gln Gly
<210> 135
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 135
Cys Ala Arg Ser Thr Tyr Tyr Gly Gly Asp Trp Tyr Phe Asn Val Trp
1 5 10 15
Gly Ala Gly
<210> 136
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 136
Cys Ala Arg Gly Gly Tyr Asp Gly Trp Asp Tyr Ala Ile Asp Tyr Trp
1 5 10 15
Gly Gln Gly
<210> 137
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 137
Cys Ala Arg Ser Thr Tyr Tyr Gly Gly Asp Trp Tyr Phe Asp Val Trp
1 5 10 15
Gly Gln Gly
<210> 138
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 138
Cys Ala Lys Asp Lys Ile Leu Trp Phe Gly Glu Pro Val Phe Asp Tyr
1 5 10 15
Trp Gly Gln Gly
20
<210> 139
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 139
Cys Ala Arg Ala Asp Tyr Tyr Gly Ser Asp Tyr Val Lys Phe Asp Tyr
1 5 10 15
Trp Gly Gln Gly
20
<210> 140
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 140
Cys Ala Arg Glu Lys Asp Asn Tyr Ala Thr Gly Ala Trp Phe Ala Tyr
1 5 10 15
Trp Gly Gln Gly
20
<210> 141
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 141
Cys Ala Arg Glu Lys Asp Asn Tyr Ala Thr Gly Ala Trp Phe Ala Tyr
1 5 10 15
Trp Gly Gln Gly
20
<210> 142
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 142
Cys Thr Arg Asp Gly Tyr Ser Ser Gly Arg His Tyr Gly Met Asp Val
1 5 10 15
Trp Gly Gln Gly
20
<210> 143
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 143
Cys Ala Arg Asp Gly Asp Tyr Gly Ser Ser Tyr Gly Ala Met Asp Tyr
1 5 10 15
Trp Gly Gln Gly
20
<210> 144
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 144
Cys Ala Arg Val Val Tyr Tyr Ser Asn Ser Tyr Trp Tyr Phe Asp Val
1 5 10 15
Trp Gly Thr Gly
20
<210> 145
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 145
Cys Ala Arg Asp Ser Ser Ser Ser Trp Ala Arg Trp Phe Phe Asp Leu
1 5 10 15
Trp Gly Arg Gly
20
<210> 146
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 146
Cys Ser Ala Ser Tyr Tyr Arg Tyr Asp Val Gly Ala Trp Phe Ala Tyr
1 5 10 15
Trp Gly Gln Gly
20
<210> 147
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 147
Cys Ala Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu
1 5 10 15
Trp Gly Arg Gly
20
<210> 148
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 148
Cys Ala Lys Tyr Pro His Tyr Tyr Gly Ser Ser His Trp Tyr Phe Asp
1 5 10 15
Val Trp Gly Gln Gly
20
<210> 149
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 149
Cys Ala Arg Asp His Asp Phe Arg Ser Gly Tyr Glu Gly Trp Phe Asp
1 5 10 15
Pro Trp Gly Gln Gly
20
<210> 150
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 150
Cys Ala Arg Val Tyr Ser Ser Gly Trp His Val Ser Asp Tyr Phe Asp
1 5 10 15
Tyr Trp Gly Gln Gly
20
<210> 151
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 151
Cys Thr Arg Asn Gly Asp Tyr Tyr Val Ser Ser Gly Asp Ala Met Asp
1 5 10 15
Tyr Trp Gly Gln Gly
20
<210> 152
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 152
Cys Ala Arg Phe Ser His Phe Ser Gly Ser Asn Tyr Asp Tyr Phe Asp
1 5 10 15
Tyr Trp Gly Gln Gly
20
<210> 153
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 153
Cys Ala Arg Ile Pro Ser Gly Ser Tyr Tyr Tyr Asp Tyr Asp Met Asp
1 5 10 15
Val Trp Gly Gln Gly
20
<210> 154
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 154
Cys Ala Arg His Ser Gly Tyr Gly Thr His Trp Gly Val Leu Phe Ala
1 5 10 15
Tyr Trp Gly Gln Gly
20
<210> 155
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 155
Cys Ala Arg Glu Gly Val Tyr His Asp Tyr Asp Asp Tyr Ala Met Asp
1 5 10 15
Tyr Trp Gly Gln Gly
20
<210> 156
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 156
Cys Ala Arg Leu Gly Tyr Asp Asp Ile Tyr Asp Asp Trp Tyr Phe Asp
1 5 10 15
Val Trp Gly Gln Gly
20
<210> 157
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 157
Cys Ala Arg Gly Glu Leu Pro Tyr Tyr Arg Met Ser Lys Val Met Asp
1 5 10 15
Val Trp Gly Gln Gly
20
<210> 158
<211> 22
<212> БЕЛОК
<213> Homo sapiens
<400> 158
Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr Tyr Ala Met
1 5 10 15
Asp Tyr Trp Gly Gln Gly
20
<210> 159
<211> 22
<212> БЕЛОК
<213> Homo sapiens
<400> 159
Cys Ala Arg Gly Tyr Tyr Asp Ile Leu Thr Gly Tyr Tyr Tyr Tyr Phe
1 5 10 15
Asp Tyr Trp Gly Gln Gly
20
<210> 160
<211> 23
<212> БЕЛОК
<213> Homo sapiens
<400> 160
Cys Ala Lys Asp Leu Gly Trp Ser Asp Ser Tyr Tyr Tyr Tyr Tyr Gly
1 5 10 15
Met Asp Val Trp Gly Gln Gly
20
<210> 161
<211> 23
<212> БЕЛОК
<213> Homo sapiens
<400> 161
Cys Ala Arg Gly Arg Tyr Ser Gly Ser Gly Ser Tyr Tyr Asn Asp Trp
1 5 10 15
Phe Asp Pro Trp Gly Gln Gly
20
<210> 162
<211> 24
<212> БЕЛОК
<213> Homo sapiens
<400> 162
Cys Ala Arg Gly Lys Gly Asn Thr His Lys Pro Tyr Gly Tyr Val Arg
1 5 10 15
Tyr Phe Asp Val Trp Gly Gln Gly
20
<210> 163
<211> 24
<212> БЕЛОК
<213> Homo sapiens
<400> 163
Cys Ala Arg Asp His Tyr Gly Ser Gly Val His His Tyr Phe Tyr Tyr
1 5 10 15
Gly Leu Asp Val Trp Gly Gln Gly
20
<210> 164
<211> 24
<212> БЕЛОК
<213> Homo sapiens
<400> 164
Cys Ala Arg Gln Ser Thr Tyr Tyr Tyr Gly Ser Gly Asn Tyr Tyr Gly
1 5 10 15
Trp Phe Asp Arg Trp Asp Gln Gly
20
<210> 165
<211> 24
<212> БЕЛОК
<213> Homo sapiens
<400> 165
Cys Ala Arg Pro Val Arg Ser Arg Trp Leu Gln Leu Gly Leu Glu Asp
1 5 10 15
Ala Phe His Ile Trp Gly Gln Gly
20
<210> 166
<211> 25
<212> БЕЛОК
<213> Homo sapiens
<400> 166
Cys Ala Arg Glu Asn Leu Asp Asn Ser Gly Tyr Thr Tyr Tyr Phe Ser
1 5 10 15
Gly Trp Phe Asp Pro Trp Gly Gln Gly
20 25
<210> 167
<211> 25
<212> БЕЛОК
<213> Homo sapiens
<400> 167
Cys Val Arg Asp Tyr Tyr Asp Ile Leu Thr Tyr Asp Tyr Ile His Tyr
1 5 10 15
Trp Tyr Phe Asp Leu Trp Gly Arg Gly
20 25
<210> 168
<211> 26
<212> БЕЛОК
<213> Homo sapiens
<400> 168
Cys Ala Ala Ala Gly Val Arg Ala Glu Asp Gly Val Arg Arg Thr Leu
1 5 10 15
Pro Ser Glu Tyr Thr Phe Trp Gly Gln Gly
20 25
<---
Реферат
Изобретение относится к области биотехнологии, конкретно к библиотеке векторов, содержащих человеческие области HC-CDR3 различной длины, где разнообразие указанной библиотеки сфокусировано областью HC-CDR3. Полученная библиотека оптимизирована таким образом, что избыточность уменьшена для коротких петель HC-CDR3, а перекрывание вариантов области HC-CDR3 для больших длин петель увеличено, что позволяет использовать полученную библиотеку для эффективного отбора рекомбинантных антител против антигенов-мишеней. 9 з.п. ф-лы, 5 табл., 11 пр., 15 ил.
Комментарии