Условно реплицирующийся цитомегаловирус в качестве вакцины против cmv - RU2611198C2
Код документа: RU2611198C2
Чертежи























































































Описание
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к способам индукции иммунного ответа на цитомегаловирус (CMV) с использованием генетически модифицированного CMV, который является условно дефектным по репликации. Настоящее изобретение также относится к CMV, который был изменен рекомбинантными методами для создания возможности внешнего контроля над вирусной репликацией. Композиции, содержащие дефектный по репликации CMV, также охвачены настоящим изобретением.
УРОВЕНЬ ТЕХНИКИ
Цитомегаловирус (CMV), также известный как вирус герпеса человека 5 типа (HHV-5), представляет собой вирус герпеса, классифицируемый как член подсемейства бета семейства вирусов герпеса. Согласно данным Центра по контролю и профилактике заболеваний, CMV инфекция распространена в человеческой популяции практически повсеместно, по оценкам им инфицированы 40-80% взрослого населения США. Вирус распространяется преимущественно через биологические жидкости и часто передается от беременных женщин плоду или новорожденному. У большинства людей CMV инфекция протекает в латентной форме, хотя активация вируса может приводить к высокой температуре, ознобу, повышенной утомляемости, головным болям, тошноте и спленомегалии.
При том, что в большинстве случаев CMV инфекция у человека протекает бессимптомно, CMV инфекция у лиц с иммунодефицитом (таких как ВИЧ-положительные пациенты, пациенты с аллогенными трансплантатами и онкологические пациенты) или у лиц с незрелой иммунной системой (таких как новорожденные) может быть чревата особенно большими проблемами (Mocarski et al., Cytomegalovirus, в "Field Virology", 2701-2772, под редакцией: Knipes и Howley, 2007). CMV инфекция у таких лиц может вызывать тяжелые клинические проявления, включая пневмонию, гепатит, энцефалит, колит, увеит, ретинит, слепоту и невропатию, среди прочих болезненных состояний. Кроме того, CMV инфекция во время беременности является основной причиной врожденных дефектов детей (Adler, 2008 J. Clin Virol, 41: 231; Arvin et al., 2004 Clin Infect Dis, 39: 233; Revello et al., 2008 J Med Virol, 80: 1415). CMV инфицирует различные клетки in vivo, включая моноциты, макрофаги, дендритные клетки, нейтрофилы, эндотелиальные клетки, эпителиальные клетки, фибробласты, нейроны, гладкомышечные клетки, гепатоциты и стромальные клетки (Plachter et al. 1996, Adv. Virus Res. 46: 195). Хотя клинические изоляты CMV реплицируются в клетках различных типов, лабораторные штаммы AD169 (Elek & Stern, 1974, Lancet 1: 1) и Towne (Plotkin et al., 1975, Infect. Immun. 12: 521) реплицируются почти исключительно в фибробластах (Hahn et al., 2004, J. Virol. 78: 10023). Ограничение по тропизму, являющееся результатом серийных пассажей и в конечном итоге приспособления вируса к фибробластам, является обусловленным маркером аттенуации (Gerna et al., 2005, J. Gen. Virol. 86: 275; Gerna et al., 2002, J. Gen Virol. 83: 1993; Gerna et al., 2003, J. Gen Virol. 84: 1431; Dargan et al., 2010, J. Gen Virol. 91: 1535). Мутации, вызывающие утрату тропизма к эпителиальным клеткам, эндотелиальным клеткам, лейкоцитам и дендритным клеткам у лабораторных штаммов человеческого CMV, были картированы в трех открытых рамках считывания (ORF): UL128, UL130 и UL131 (Hahn et al., 2004, J. Virol. 78: 10023; Wang and Shenk, 2005 J. Virol. 79: 10330; Wang and Shenk, 2005 Proc Natl Acad Sci USA. 102: 18153). Биохимические и восстановительные исследования показали, что UL128, UL130 и UL131 собираются в каркасную структуру gH/gL, образуя пентамерный gH-комплекс (Wang and Shenk, 2005 Proc Natl Acad Sci USA, 102: 1815; Ryckman et al., 2008 J. Virol. 82: 60). Восстановление этого комплекса в вирионах восстанавливает тропизм вирусов к эпителиальным клеткам в лабораторных штаммах (Wang and Shenk, 2005 J. Virol. 79: 10330).
Утрату тропизма к эндотелиальным и эпителиальным клеткам считали недостатком у таких ранее изученных вакцин, как Towne (Gerna et al., 2002, J. Gen Virol. 83: 1993; Gerna et al., 2003, J. Gen Virol. 84: 1431). Нейтрализующие антитела в сыворотке людей, естественным образом инфицированных CMV, обладали повышенной более чем в 15 раз активностью, препятствующей проникновению вируса в эпителиальные клетки, по сравнению с активностью, препятствующей проникновению в фибробласты (Cui et al., 2008 Vaccine 26: 5760). У людей с первичной инфекцией быстро вырабатываются нейтрализующие антитела, препятствующие проникновению вируса в эпителиальные и эндотелиальные клетки, но медленно вырабатываются нейтрализующие антитела, препятствующие проникновению в фибробласты (Gerna et al., 2008 J. Gen. Virol. 89: 853). Более того, нейтрализующая активность, препятствующая проникновению вируса в эпителиальные и эндотелиальные клетки, отсутствует в иммунной сыворотке людей, получавших вакцину Towne (Cui et al., 2008 Vaccine 26: 5760). Совсем недавно была описана панель человеческих моноклональных антител от четырех доноров, инфицированных HCMV, и более активные нейтрализующие клоны из панели узнавали антигены пентамерного gH-комплекса (Macagno et al., 2010 J. Virol. 84: 1005).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к условно дефектному по репликации CMV (rdCMV) и к применению rdCMV в композициях и способах для лечения и/или снижения вероятности CMV инфекции или патологии, связанной с такой инфекцией, у пациента. Описанный здесь rdCMV содержит нуклеиновую кислоту, кодирующую один или более слитых белков, включающих необходимый белок, слитый с дестабилизирующим белком. В отсутствие стабилизатора слитый белок разрушается. Таким образом, rdCMV можно выращивать в тканевой культуре в условиях, допускающих репликацию (то есть, в присутствии стабилизатора), но репликация уменьшается, и предпочтительно предотвращается, при введении пациенту (в отсутствие стабилизатора).
Один вариант осуществления настоящего изобретения относится к условно дефектному по репликации CMV. rdCMV содержит нуклеиновую кислоту, кодирующую один или более слитых белков, включающих необходимый белок, слитый с дестабилизирующим белком. Нуклеиновые кислоты, кодирующие необходимый белок дикого типа, больше не присутствуют в rdCMV, и, таким образом, слитый белок является необходимым для вирусной репликации. В предпочтительных вариантах осуществления необходимые белки выбирают из группы, состоящей из IE1/2, UL51, UL52, UL79 и UL84, и дестабилизирующий белок представляет собой FKBP или его производное.
Другой вариант осуществления настоящего изобретения относится к композиции, содержащей выделенный rdCMV и фармацевтически приемлемый носитель. Композиция может дополнительно содержать адъювант, включая, но без ограничения, адъювант ISCOMATRIX® и алюминий-фосфатный адъювант.
Другой вариант осуществления настоящего изобретения относится к применению композиции rdCMV для индукции иммунного ответа против CMV у пациента. Пациентов можно лечить профилактически или терапевтически путем введения rdCMV по настоящему изобретению. Профилактическое лечение обеспечивает достаточный защитный иммунитет для снижения вероятности или тяжести CMV инфекции, включая первичные инфекции, рецидивирующие инфекции (то есть, те, которые возникают в результате реактивации латентного CMV) и суперинфекции (то есть, те, которые возникают в результате инфицирования штаммом CMV, отличающимся от того, которым пациент был инфицирован ранее). В конкретных вариантах осуществления женщин детородного возраста, особенно девочек-подростков, вакцинируют для снижения вероятности CMV инфекции (как первичной, так и рецидивирующей или суперинфекции) во время беременности и, таким образом, снижения вероятности передачи CMV плоду. Терапевтическое лечение можно проводить для снижения продолжительности/тяжести текущей CMV инфекции.
Другой вариант осуществления настоящего изобретения относится к способам получения rdCMV по изобретению, включающим размножение rdCMV в эпителиальных клетках, таких как клетки ARPE-19 (ATCC, регистрационный № CRL-2302), в присутствии Shield-1. В некоторых вариантах осуществления rdCMV размножают в эпителиальных клетках на микроносителях или других системах клеточных культур высокой плотности.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1A-1C демонстрируют схематическую диаграмму конструкции штамма CMV с восстановленной экспрессией пентамерного gH-комплекса. (A) Стратегия получения самоудаляемой бактериальной искусственной хромосомы (BAC) для манипуляций с вирусным геномом AD169. (B) Исправление мутации со сдвигом рамки в UL131 для восстановления его экспрессии. (C) Замена GFP геном cre-рекомбиназы для создания самоудаляемой BAC CMV.
Фиг. 2A-2D демонстрируют влияние общепринятых способов инактивации на иммуногенность gH-комплекса. Использовали γ-облучение (A, B) и β-пропиолактон (BPL) (C, D) для инактивации экспрессирующего gH-комплекс CMV. Кинетику инактивации определяли анализом бляшкообразования (A, C), а иммуногенность определяли, оценивая сыворотку мышей, которым вводили CMV, на нейтрализующую активность против проникновения вируса в эпителиальные клетки (B, D).
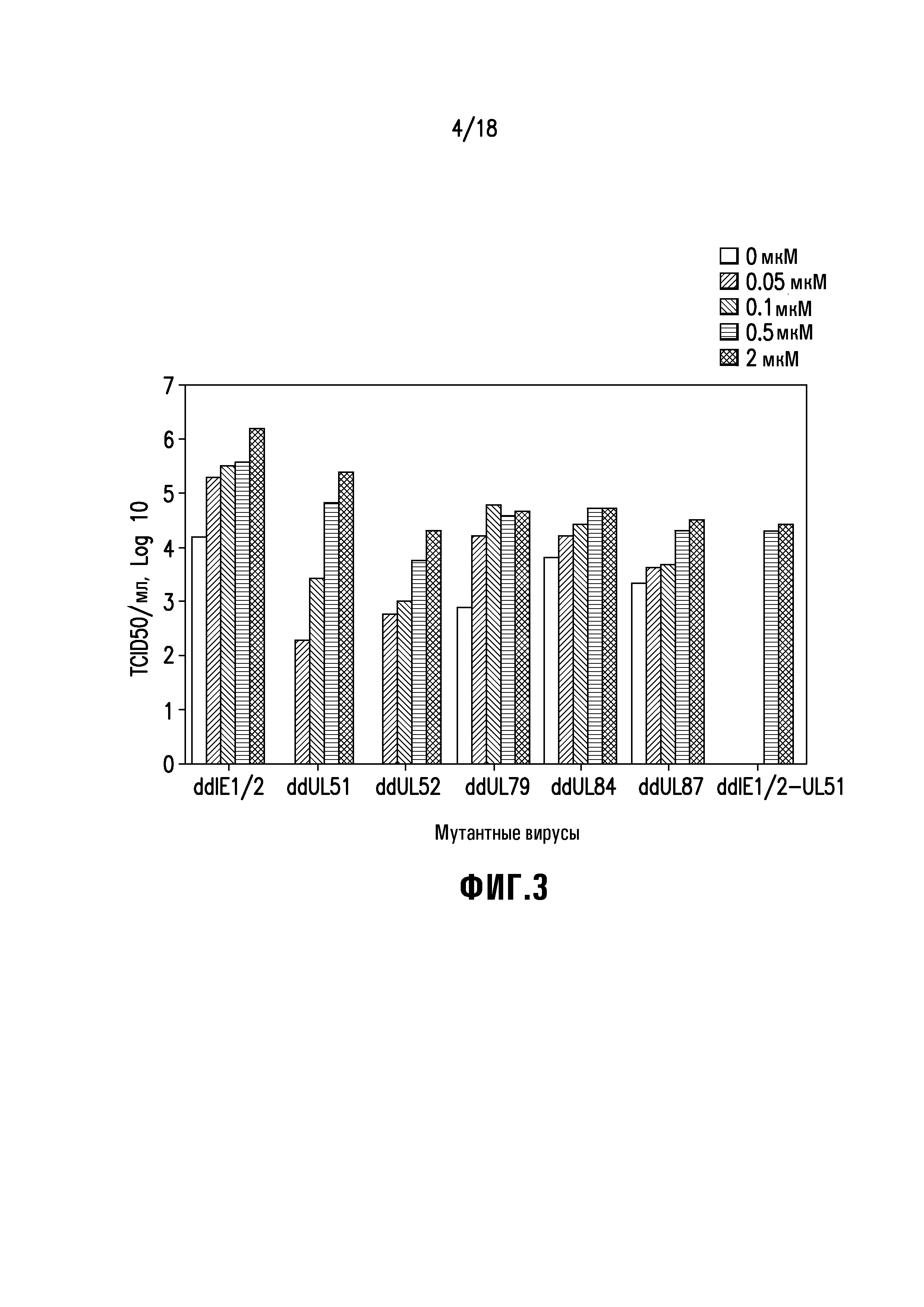
Фиг. 3 демонстрирует зависимую от концентрации Shield-1 продукцию вирусного потомства экспрессирующего gH-комплекс CMV с различными необходимыми белками, слитыми с производным FKBP. Клетки ARPE-19 инфицировали вирусами rdCMV при множественности заражения 0,01 БОЕ/клетку в течение 1 часа, дважды промывали свежей средой и инкубировали в ростовой среде, содержащей 0, 0,05, 0,1 0,5 или 2 мкМ Shield-1. Через семь дней после инфицирования внеклеточный вирус собирали, и титры вируса определяли анализом TCID50 на клетках ARPE-19 в присутствии 2 мкМ Shield-1.
Фиг. 4A-4D демонстрируют кинетику роста rdCMV в клетках ARPE-19. Клетки инфицировали вирусами, содержащими слитые белки с (A) IE1/2, (B) UL51, (C) IE1/2-UL51, или (D) родительским вирусом beMAD при множественности заражения 0,01 БОЕ/клетку. Через один час клетки дважды промывали свежей средой и инкубировали в отсутствие (незакрашенные кружки) или в присутствии (закрашенные кружки) 2 мкМ Shield-1. Внеклеточный вирус собирали в указанные моменты времени после инфицирования, и инфекционный вирус количественно определяли анализом TCID50 на клетках ARPE-19 в среде, содержащей 2 мкМ Shield-1.
Фиг. 5A-5E. Кинетика роста rdCMV IE1/2-UL51 в клетках различного типа. Клетки (A) MRC-5, (B) HUVEC, (C) AoSMC, (D) SKMC, (E) CCF-STTG1 инфицировали вирусом rdCMV и инкубировали в течение одного часа. Клетки дважды промывали свежей средой, а затем инкубировали в отсутствие (незакрашенные кружки) или в присутствии (закрашенные кружки) 2 мкМ Shield-1. Внеклеточный вирус собирали в указанные моменты времени после инфицирования, и инфекционный вирус количественно определяли анализом TCID50 на клетках ARPE-19 в среде, содержащей 2 мкМ Shield-1.
Фиг. 6A-6C. Анализ иммуногенности rdCMV IE1/2-UL51 на мышах, кроликах и макаках-резусах. (A) Мышей иммунизировали в недели 0 и 4 beMAD (незакрашенные кружки) или rdCMV IE1/2-UL51 (закрашенные кружки). (B) Кроликов иммунизировали в недели 0, 3 и 8, используя 10 мкг beMAD или указанных rdCMV. (C) Макак-резусов иммунизировали в недели 0 и 8, используя 100 мкг beMAD или rdCMV IE1/2-UL51. В каждом случае образцы сыворотки собирали и анализировали с помощью микроанализа нейтрализации CMV на клетках ARPE-19. Линии указывают среднее геометрическое титров нейтрализующих антител (NT50) в каждой группе.
Фиг. 7 представляет протяженные во времени титры нейтрализующих антител у макак-резусов, вакцинированных вирусом с двойным слиянием IE1/2-UL51. Группы макак-резусов (n=5) вакцинировали указанными дозами вакцин или препаратов в недели 0, 8 и 24 (показано красными треугольниками), в то время как одна группа получала gb/mf59 (30 мг/дозу) в недели 0, 4 и 24. Иммунные сыворотки собирали в указанные моменты времени и оценивали в анализе на нейтрализацию вируса. GMT титров NT50 вычерчивали на протяжении времени со стандартной ошибкой для группы. AAHS: аморфный гидроксифосфат-сульфат алюминия; IMX: ISCOMATRIX; HNS: базовый буфер.
Фиг. 8A-8D представляют результаты анализа ELISPOT на IFN-γ у макак-резусов, вакцинированных вирусом с двойным слиянием IE1/2-UL51 при использовании либо 100 мкг (A), либо 10 мкг (B-D) на дозу. Адъювант либо не использовали (A-B), либо использовали AAHS (C) или ISCOMATRIX (D). PBMC стимулировали пулами пептидов, представляющих антигены HCMV. Серые столбики представляют GMT для каждого антигена в группе (n=5). Частота положительного ответа для каждого антигена указана сверху каждого антигена в панелях.
Фиг. 9A-9B демонстрируют, что вакцинация вирусом с двойным слиянием IE1/2-UL51 способна вызывать T-клеточные ответы как CD8+ (A), так и CD4+ (B) фенотипов у макак-резусов. PBMC собирали от обезьян, получавших дозу либо 100 мкг, либо 10 мкг вакцины с ISCOMATRIX® в качестве адъюванта. PBMC стимулировали пулами пептидов, представляющих антигены HCMV, с последующим окрашиванием на IFN-γ и CD4+/CD8+ поверхностные T-клеточные маркеры. Данные представлены в виде количества положительных по CD4+/CD8+, положительных по IFN-γ клеток на миллион PBMC. Линии представляют значения среднего геометрического титров (GMT) в группе, получавшей одну и ту же вакцину (n=5). Цифры в нижней части графиков представляют собой GMT в обеих вакцинированных группах (n=10). CMV: очищенный вирус; SEB: митоген, использованный в качестве положительного контроля; IMX: ISCOMATRIX.
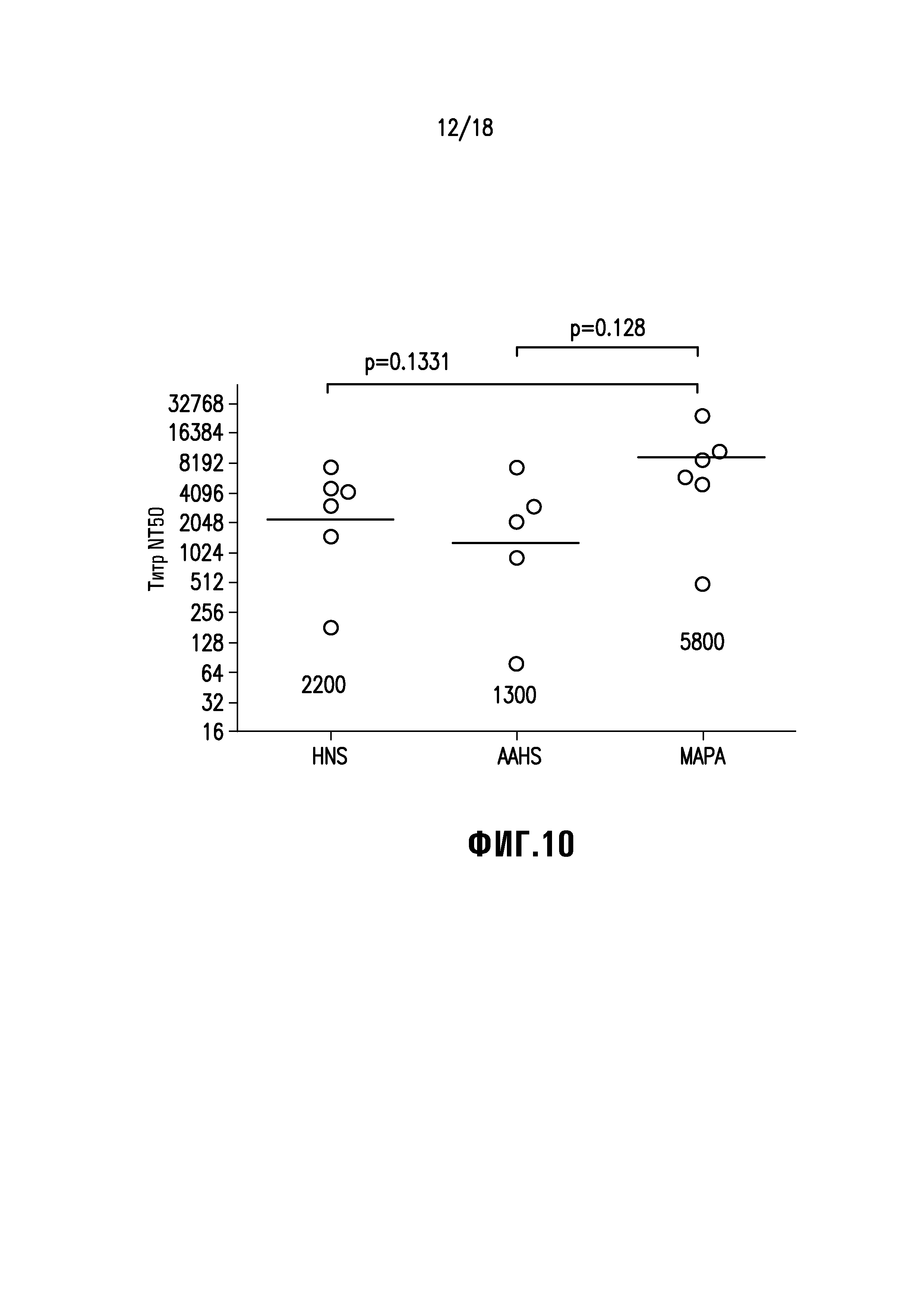
Фиг. 10 демонстрирует, что алюминий-фосфатный адъювант Merck (MAPA) способен увеличивать титры нейтрализующих антител у обезьян. Обезьян резусов иммунизировали дозой 30 мкг вакцины вируса с двойным слиянием, сформулированной в HNS (базовом буфере), AAHS или MAPA, в недели 0 и 8. Образцы сыворотки собирали в неделю 12 и оценивали в отношении титров нейтрализующих антител. Линии представляют значения геометрического среднего для группы.
Фиг. 11A-11B демонстрируют стабильность экспрессирующего gH CMV в сбалансированном солевом растворе Хэнка (HBSS) при различных температурах. (A) Образцы CMV в HBSS хранили при указанных значениях температуры в течение 4 дней до того, как измеряли стабильность вируса CMV при помощи анализа на проникновение вируса. (B) Величины EC50 рассчитывали для образцов, используя результаты анализа на проникновение вируса. * указывает на то, что EC50 невозможно рассчитать в результате полной потери инфекционности.
Фиг. 12A-12B демонстрируют влияние pH на стабильность экспрессирующего gH CMV при комнатной температуре. (A) Образцы CMV в буферах с различным значением pH хранили при комнатной температуре в течение 4 дней до того, как была измерена стабильность вируса CMV при помощи анализа на проникновение вируса. (B) Величины EC50 рассчитывали для образцов, используя результаты анализа на проникновение вируса.
Фиг. 13A-13B демонстрируют влияние мочевины самой по себе или в сочетании с хлоридом натрия на стабильность экспрессирующего gH вируса CMV. (A) 2% мочевину отдельно или в сочетании с 150 мМ NaCl добавляли к CMV в 25 мМ гистидиновом буфере, pH 6, при комнатной температуре на 4 дня перед тем, как стабильность вируса CMV измеряли при помощи анализа на проникновение вируса. (B) Величины EC50 рассчитывали для образцов, используя результаты анализа на проникновение вируса.
Фиг. 14A-14B демонстрируют влияние ионной силы на стабильность экспрессирующего gH вируса CMV. (A) NaCl в возрастающих концентрациях (0 мМ, 75 мМ, 150 мМ и 320 мМ NaCl) добавляли к CMV в 25 мМ гистидиновом буфере, pH 6, при комнатной температуре на 4 дня перед тем, как стабильность вируса CMV измеряли при помощи анализа на проникновение вируса. (B) Величины EC50 рассчитывали для образцов, используя результаты анализа на проникновение вируса.
Фиг. 15 демонстрирует влияние криопротекторов на стабильность экспрессирующего gH вируса CMV при проведении циклов замораживания-размораживания. Указанные криопротекторы добавляли к CMV в 25 мМ гистидиновом буфере, pH 6, и проводили три цикла замораживания-размораживания перед тем, как стабильность вируса CMV измеряли при помощи анализа на проникновение вируса. Величины EC50 рассчитывали для образцов, используя результаты анализа на проникновение вируса.
Фиг. 16A-16B демонстрируют влияние температуры хранения на индукцию нейтрализующих CMV антител при исследовании иммуногенности на мышах. Мышей иммунизировали в день 0, и делали бустерную инъекцию в день 21 с последующим забором крови в день 28. Сыворотку мышей тестировали на нейтрализующие антитела против экспрессирующего gH CMV с использованием клеток ARPE-19. Титры NT50 получали путем аппроксимации нелинейной кривой. (A) Образцы CMV хранили при различных значениях температуры в течение 3 месяцев до изучения иммуногенности. (B) Образцы CMV хранили при различных значениях температуры в течение 8 часов после размораживания и до изучения иммуногенности.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к условно дефектному по репликации CMV (rdCMV) и к применению rdCMV в композициях и способах для лечения и/или снижения вероятности CMV инфекции или патологии, связанной с такой инфекцией, у пациента. Описанный здесь rdCMV содержит нуклеиновую кислоту, кодирующую один или более слитых белков, включающих необходимый белок, слитый с дестабилизирующим белком, вместо необходимого белка дикого типа. В отсутствие стабилизатора слитый белок разрушается аппаратом клетки-хозяина. В присутствии стабилизатора слитый белок стабилизируется и не разрушается.
Подходящие слитые белки для использования по настоящему изобретению в достаточной степени сохраняют необходимую белковую активность для облегчения вирусной репликации в клетке-хозяине в присутствии стабилизатора и вызывают уменьшение (предпочтительно уменьшение более чем на 50%, 75%, 90%, 95% или 99%) репликации CMV в отсутствие стабилизатора. Предпочтительно, необходимый белок для использования в слитом белке является неструктурным белком и, таким образом, не упаковывается в вирионы rdCMV. Подходящие необходимые белки, определенные здесь, включают белки CMV, кодируемые необходимыми генами IE1/2, UL51, UL52, UL79 и UL84.
Пример дестабилизирующего белка и стабилизатора описан в патентной публикации США 2009/0215169, в которой раскрыты композиции, системы и способы модуляции стабильности белков при помощи малых молекул. Вкратце, проводят слияние белка с белком, влияющим на стабильность, FKBP или его производным. Добавленная экзогенно проникающая в клетки малая молекула, Shield-1 (Shld-1), взаимодействует с FKBP или его производным и стабилизирует слитый белок. В отсутствие Shield-1 FKBP или его производное направляет разрушение слитого белка аппаратом клетки-хозяина.
В одном из вариантов осуществления настоящего изобретения необходимый белок CMV слит с FKBP или его производным. В присутствии Shield-1 слитый белок стабилизируется. Однако в отсутствие Shield-1 FKBP или его производное направляет разрушение слитого белка аппаратом клетки-хозяина.
В отсутствие слитого белка репликация rdCMV уменьшается (предпочтительно, на более чем 50%, 75%, 90%, 95% или 99% по сравнению с CMV, не содержащим дестабилизированный необходимый белок) или предотвращается.
Рекомбинантный вирус, используемый в способе по изобретению, также экспонирует иммуногенный пентамерный gH-комплекс на вирионе.
Варианты осуществления также включают рекомбинантный CMV или его композиции, описанные здесь, или вакцину, содержащую или состоящую из указанного CMV, или композиции (i) для применения в, (ii) для применения в качестве лекарственного средства для, или (iii) для применения в получении лекарственного средства для: (a) терапии (например, человеческого организма); (b) медицины; (c) ингибирования репликации CMV; (d) лечения или профилактики CMV инфекции или (e) лечения, профилактики или отсрочки начала или прогрессирования связанного с CMV заболевания(-ий). В этих вариантах применения рекомбинантный CMV, его композиции и/или вакцины, содержащие или состоящие из указанного CMV или композиций, необязательно можно использовать в сочетании с одним или более противовирусными средствами (например, противовирусными соединениями или противовирусными иммуноглобулинами; комбинированными вакцинами, описанными ниже).
Используемый здесь термин "индуцировать иммунный ответ" относится к способности условно дефектного по репликации CMV вызывать иммунный ответ у пациента, предпочтительно млекопитающего, более предпочтительно человека, которому его вводят, при этом ответ включает, но без ограничения, производство элементов (таких как антитела), которые специфически связывают и предпочтительно нейтрализуют CMV и/или вызывают активацию T-клеток. "Защитный иммунный ответ" представляет собой иммунный ответ, который снижает вероятность того, что пациент получит CMV инфекцию (включая первичную, рецидивирующую и/или суперинфекцию), и/или облегчает по меньшей мере одну патологию, связанную с CMV инфекцией, и/или снижает тяжесть/продолжительность CMV инфекции.
Используемый здесь термин "иммунологически эффективное количество" означает количество иммуногена, способное при введении пациенту вызвать иммунный ответ против CMV, который может защитить пациента от CMV инфекции (включая первичную, рецидивирующую и/или суперинфекцию), и/или облегчить по меньшей мере одну патологию, связанную с CMV инфекцией, и/или снизить тяжесть/продолжительность CMV инфекции у пациента. Количество должно быть достаточным, чтобы значительно снизить вероятность или тяжесть CMV инфекции. Животные модели, известные в данной области, можно использовать для оценки защитного эффекта от введения иммуногена. Например, иммунные сыворотки или иммунные Т-клетки от индивидуумов, которым вводили иммуноген, можно анализировать на нейтрализующую способность антител или цитотоксических T-клеток или цитокин-продуцирующую способность иммунных T-клеток. Анализы, обычно используемые для такой оценки, включают, но без ограничения, анализ на нейтрализацию вируса, анализ ELISA на вирусные антигены, анализ ELISA на цитокин интерферон-гамма, анализ ELISPOT на интерферон-гамма, внутриклеточное окрашивание различных цитокинов (ICS) и анализ на цитотоксичность с высвобождением51хрома. Животные модели со стимуляцией антигеном также можно использовать для определения иммунологически эффективного количества иммуногена.
Используемый здесь термин "условно дефектный по репликации вирус" относится к вирусным частицам, которые способны реплицироваться в определенных условиях среды, но не в других. В предпочтительных вариантах осуществления вирус делают условно дефектным по репликации вирусом путем дестабилизации одного или более белков, необходимых для вирусный репликации. Нуклеиновые кислоты, кодирующие недестабилизированные необходимые белки дикого типа, больше не присутствуют в условно дефектном по репликации вирусе. В условиях, в которых один или более необходимых белков дестабилизированы, вирусная репликация сокращается предпочтительно на более чем 50%, 75%, 90%, 95%, 99% или 100% по сравнению с репликацией вируса без дестабилизированных необходимых белков. Однако в условиях, которые стабилизируют дестабилизированные необходимые белки, вирусная репликация может происходить на уровне, предпочтительно по меньшей мере 75%, 80%, 90%, 95%, 99% или 100% от уровня репликации CMV, не содержащего дестабилизированный необходимый белок. В более предпочтительных вариантах осуществления один или более из необходимых белков дестабилизируют путем слияния с дестабилизирующим белком, таким как FKBP или его производное. Такие слитые белки могут быть стабилизированы в присутствии стабилизатора, такого как Shield-1. Используемый здесь термин "rdCMV" относится к условно дефектному по репликации цитомегаловирусу.
В предпочтительных вариантах осуществления иммунный ответ, индуцируемый дефектным по репликации вирусом, в сравнении с его естественным вирусным аналогом является таким же или практически таким же по степени и/или широте. В других предпочтительных вариантах осуществления морфология дефектного по репликации вируса при электронно-микроскопическом анализе является неотличимой или практически такой же, как у его естественного вирусного аналога.
Используемый здесь термин "FKBP" относится к дестабилизирующему белку с SEQ ID NO: 11. Слитые белки, содержащие FKBP, разрушаются аппаратом клетки-хозяина. Используемый здесь термин "производное FKBP" относится к белку FKBP, или его части, который был изменен в результате одной или более аминокислотных замен, делеций и/или добавлений. Производные FKBP при слиянии с белком сохраняют практически все из дестабилизирующих свойств FKBP и также сохраняют практически полностью способность FKBP стабилизироваться Shield-1. Предпочтительные производные FKBP имеют одну или более из следующих замен в указанных положениях аминокислот F15S, V24A, H25R, F36V, E60G, M66T, R71G, D100G, D100N, E102G, K105I и L106P. Производное FKBP, имеющее замены F36V и L106P (SEQ ID NO: 12), является особенно предпочтительным. В предпочтительных вариантах осуществления нуклеиновая кислота, кодирующая FKBP или производное FKBP, содержит по меньшей мере некоторые кодоны, которые обычно не используются в эндогенном FKBP у человека. Это уменьшает вероятность того, что FKBP или производное FKBP в слитом белке подвергнется реаранжировке или рекомбинации со своим аналогом в геноме человека. Последовательность нуклеиновой кислоты SEQ ID NO: 13 кодирует SEQ ID NO: 12 с использованием таких кодонов.
Используемые здесь термины "Shield-1" или "Shld1" означают синтетическую малую молекулу, которая связывается с FKBP дикого типа и его производными и выступает в качестве стабилизатора. Связывание с производным F36V примерно в 1000 раз более прочное, чем с FKBP дикого типа (Clackson et al., 1998, PNAS 95: 10437-42). Shield-1 можно синтезировать (в основном, как описано в статьях Holt et al., 1993, J. Am. Chem. Soc. 115: 9925-38 и Yang et al., 2000, J. Med. Chem. 43: 1135-42 и Grimley et al., 2008, Bioorganic & Medicinal Chemistry Letters 18: 759) или приобретать у Cheminpharma LLC (Farmington, CT) или Clontech Laboratories, INC. (Mountain View, CA). Соли Shield-1 также можно использовать в способах по изобретению. Shield-1 имеет следующую структуру:

Используемые здесь термины "слитый" или "слитый белок" относятся к двум полипептидам, скомбинированным в рамке считывания в виде частей одной непрерывной последовательности аминокислот. Слияние может быть прямым, так, что между полипептидами нет дополнительных аминокислотных остатков, или опосредованным, при котором существует небольшой аминокислотный линкер для повышения активности или дополнительной функциональности. В предпочтительных вариантах осуществления слияние является прямым.
Используемые здесь термины "пентамерный gH-комплекс" или "gH-комплекс" означают комплекс из пяти вирусных белков на поверхности вириона CMV. Комплекс состоит из белков, кодируемых UL128, UL130 и UL131 и собранных в каркасную структуру gH/gL (Wang and Shenk, 2005 Proc Natl Acad Sci USA. 102: 1815; Ryckman et al., 2008 J. Virol. 82: 60). Последовательности белков комплекса из штамма AD169 CMV указаны под регистрационными номерами GenBank NP_783797.1 (UL128), NP_040067 (UL130), CAA35294.1 (UL131), NP_040009 (gH, также известный как UL75) и NP 783793 (gL, также известный как UL115). Некоторые аттенуированные штаммы CMV имеют одну или более мутаций в UL131, так, что белок не экспрессируется, и, следовательно, gH-комплекс не образуется. В таких случаях UL131 должен быть восстановлен (при помощи способов, таких как те, что описаны в статье Wang and Shenk, 2005 J. Virol. 79: 10330), таким образом, чтобы gH-комплекс экспрессировался в rdCMV по изобретению. Вирусы по настоящему изобретению экспрессируют пять вирусных белков, которые образуют пентамерный gH-комплекс, и пентамерный gH-комплекс располагается на оболочке вируса.
Используемый здесь термин "необходимый белок" относится к вирусному белку, который необходим для вирусной репликации in vivo и в тканевой культуре. Примеры необходимых белков CMV включают, но без ограничения, IE1/2, UL37x1, UL44, UL51, UL52, UL53, UL56, UL77, UL79, UL84, UL87 и UL105.
Используемый здесь термин "дестабилизированный необходимый белок" относится к необходимому белку, который экспрессируется и выполняет свои функции в вирусной репликации и разрушается в отсутствие стабилизатора. В предпочтительных вариантах осуществления необходимый белок слит с дестабилизирующим белком, таким как FKBP или его производное. В нормальных условиях роста (то есть, без присутствия стабилизатора) слитый белок экспрессируется, но разрушается аппаратом клетки-хозяина. Разрушение не позволяет необходимому белку функционировать в вирусной репликации, и, таким образом, имеет место функциональный нокаут необходимого белка. В условиях, когда стабилизатор, такой как Shield-1, присутствует, слитый белок стабилизируется и может выполнять свои функции на уровне, который может поддерживать вирусную репликацию, то есть, предпочтительно по меньшей мере 75%, 80%, 90%, 95%, 99% или 100% от уровня репликации CMV, который не содержит дестабилизированный необходимый белок.
Дефектный по репликации CMV
В способах по настоящему изобретению используют дефектный по репликации CMV (rdCMV), который экспрессирует пентамерный gH-комплекс. Любой аттенуированный вирус CMV, который экспрессирует пентамерный gH-комплекс, можно сделать дефектным по репликации с помощью способов по изобретению. В одном варианте осуществления аттенуированный CMV представляет собой AD169, в котором была восстановлена экспрессия gH-комплекса вследствие исправления мутации в гене UL131 (см. пример 1).
Условно дефектные по репликации вирусы представляют собой мутанты, в которых один или более необходимых вирусных белков заменены дестабилизированными аналогами необходимых белков. Дестабилизированный аналог кодируется нуклеиновой кислотой, которая кодирует слитый белок, состоящий из необходимого белка и дестабилизирующего белка. Дестабилизированный необходимый белок способен функционировать, поддерживая вирусную репликацию только в присутствии стабилизатора. В предпочтительных вариантах осуществления способы, описанные в патентной публикации США 2009/0215169, используют для придания условно дефектного по репликации фенотипа экспрессирующему пентамерный gH-комплекс CMV. Вкратце, проводят слияние одного или более белков, необходимых для репликации CMV, с дестабилизирующим белком, FKBP или производным FKBP. Нуклеиновые кислоты, кодирующие необходимый белок дикого типа, больше не присутствуют в rdCMV. В присутствии экзогенно добавленного проникающего в клетки низкомолекулярного стабилизатора, Shield-1 (Shld-1), слитый белок стабилизируется, и необходимый белок может функционировать, поддерживая вирусную репликацию. Репликация rdCMV в присутствии стабилизатора предпочтительно составляет по меньшей мере 75%, 80%, 90%, 95%, 99% или 100% от уровня репликации CMV, который не содержит дестабилизирующий слитый белок (например, родительского аттенуированного CMV, использованного для конструирования rdCMV). В отсутствие Shield-1 дестабилизирующий белок из слитого белка является причиной того, что слитый белок в значительной степени разрушается аппаратом клетки-хозяина. В отсутствие или в присутствии минимальных количеств необходимого белка CMV не может реплицироваться в степени, достаточной, чтобы вызывать или поддерживать CMV инфекцию у пациента. Репликация rdCMV в отсутствие стабилизатора не происходит или сокращается предпочтительно более чем на 50%, 75%, 90%, 95% или 99% по сравнению с репликацией CMV, который не содержит дестабилизирующий слитый белок (например, родительского аттенуированного CMV, использованного для конструирования rdCMV).
При помощи методов рекомбинантной ДНК, хорошо известных в данной области, нуклеиновую кислоту, кодирующую белок, необходимый для репликации CMV и/или возникновения/поддержания CMV инфекции, присоединяют к нуклеиновой кислоте, которая кодирует FKBP или его производное. Кодируемый слитый белок содержит FKBP, или производное FKBP, слитый в рамке считывания с необходимым белком. Кодируемый слитый белок стабилен в присутствии Shield-1. Однако кодируемый слитый белок дестабилизируется в отсутствие Shield-1 и подлежит разрушению. В предпочтительных вариантах осуществления FKBP представляет собой SEQ ID NO: 11. В других предпочтительных вариантах осуществления производное FKBP представляет собой FKBP, содержащий одну или более аминокислотных замен, выбранных из группы, состоящей из: F15S, V24A, H25R, F36V, E60G, M66T, R71G, D100G, D100N, E102G, K105I и L106P. В более предпочтительном варианте осуществления производное FKBP содержит замены F36V и/или L106P (SEQ ID NO: 12). В более предпочтительном варианте осуществления производное FKBP кодируется SEQ ID NO: 13.
Необходимые белки, намеченные для дестабилизации в результате слияния с FKBP или его производным, 1) являются необходимыми для вирусной репликации, 2) способны выдерживать слияние с дестабилизирующим белком без существенного нарушения функции необходимого белка, и 3) способны выдерживать вставку нуклеиновой кислоты, кодирующей FKBP или его производное, на 5' или 3'-конце вирусной ORF, кодирующей необходимый белок, без существенного нарушения ORF других окружающих вирусных генов. В предпочтительных вариантах осуществления необходимые белки, намеченные для дестабилизации в результате слияния с FKBP или a его производным, представляют собой неструктурные белки и, как таковые, имеют меньшую вероятность быть упакованными в вирионы рекомбинантного CMV. В таблице 1 приведены гены CMV, соответствующие вышеперечисленным критериям.
Настоящее изобретение охватывает rdCMV, который содержит слитые белки, состоящие из необходимого белка или его производного, слитого с дестабилизирующим белком. Производные необходимого белка содержат одну или более из аминокислотных замен, добавлений и/или делеций относительно необходимого белка дикого типа, хотя все еще могут обеспечивать активность необходимого белка по меньшей мере в достаточной степени, чтобы поддерживать вирусную репликацию в присутствии Shield-1. Примеры измерения вирусной активности приведены в примерах ниже. Можно использовать методы, известные в данной области, для определения степени различия между интересующим необходимым белком CMV и его производным. В одном варианте осуществления для определения родства используют идентичность последовательности. Производные по изобретению будут предпочтительно на по меньшей мере 85% идентичными, по меньшей мере 90% идентичными, по меньшей мере 95% идентичными, по меньшей мере 97% идентичными, по меньшей мере 99% идентичными базовой последовательности. Процент идентичности определяют как количество идентичных остатков, деленное на общее количество остатков и умноженное на 100. Если последовательности при выравнивании имеют разную длину (вследствие разрывов или удлинений), в расчетах используют длину самой длинной последовательности, представляющую значение общей длины.
В некоторых вариантах осуществления один или более из необходимых для вирусной репликации вирусных белков, намеченных для дестабилизации, выбирают из группы, состоящей из IE1/2, UL51, UL52, UL84, UL79, UL87, UL37x1, UL77 и UL53 или их производных. В конкретном варианте осуществления один или более из необходимых для вирусной репликации вирусных белков, намеченных для дестабилизации, выбирают из группы, состоящей из IE1/2, UL51, UL52, UL84, UL79, UL87. В более конкретном варианте осуществления один или более из необходимых для вирусной репликации вирусных белков, намеченных для дестабилизации, выбирают из группы, состоящей из IE1/2, UL51, UL52, UL79 и UL84.
Более чем один необходимый белок может быть дестабилизирован путем слияния с FKBP или его производным. В некоторых вариантах осуществления необходимые белки функционируют на разных стадиях репликации и/или инфицирования CMV (включая, но без ограничения, предраннюю, раннюю или позднюю стадии). В предпочтительных вариантах осуществления сочетание необходимых для вирусной репликации вирусных белков, намеченных для дестабилизации, выбирают из группы, состоящей из IE1/2 и UL51, IE1/2 и UL52, IE1/2 и UL79, IE1/2 и UL84, UL84 и UL51, а также UL84 и UL52. В более предпочтительном варианте осуществления IE1/2 и UL51 выбирают для дестабилизации в одном и том же рекомбинантном CMV. В наиболее предпочтительном варианте осуществления слитый белок, содержащий IE1/2, представляет собой SEQ ID NO: 1, и слитый белок, содержащий UL51, представляет собой SEQ ID NO: 3. SEQ ID NO: 1 и 3 могут кодироваться SEQ ID NO: 2 и 4, соответственно. Геном rdCMV с дестабилизированными IE1/2 и UL51 приведен в SEQ ID NO: 14.
Можно проводить слияние FKBP или его производного с необходимым белком либо напрямую, либо опосредованно. В предпочтительных вариантах осуществления проводят слияние FKBP или его производного с необходимым белком напрямую.
Можно проводить слияние FKBP или его производного с необходимым белком либо на N-, либо на C-конце необходимого белка. В предпочтительных вариантах осуществления проводят слияние FKBP на N-конце необходимого белка.
Можно проводить слияние более чем одного FKBP или его производного с необходимым белком. В вариантах осуществления, в которых более чем один FKBP или его производное слиты с необходимым белком, все из отдельных FKBP или их производных могут быть одинаковыми или разными. В предпочтительных вариантах осуществления один FKBP, или его производное, слит с необходимым белком.
Дополнительные способы инактивации
В некоторых вариантах осуществления rdCMV, описанный выше, дополнительно инактивируют путем химической или физической инактивации. Примеры такой инактивации включают термическую обработку, инкубацию с формальдегидом, β-пропиолактоном (BPL), или бинарным этиленимином (BEI), или гамма-облучение. Предпочтительные способы не нарушают или существенно не нарушают иммуногенность, включая, но без ограничения, иммуногенность, индуцируемую пентамерным gH-комплексом. Как таковой, иммунный ответ, вызываемый CMV, который был дополнительно инактивирован, сохраняется или в значительной степени сохраняется по сравнению с ответом на rdCMV, не подвергнутый дополнительной инактивации. В предпочтительных вариантах осуществления способность дополнительно инактивированного CMV вызывать продукцию нейтрализующих антител сопоставима с такой способностью у rdCMV, не подвергнутого дополнительной инактивации. Режим инактивации с использованием одного из, или сочетания, химических или физических методов определяют эмпирически для обеспечения иммуногенности CMV, включая пентамерный gH-комплекс.
Оценка вирусной репликации
Специалист в данной области может использовать тесты на вирусную репликацию для определения полезности конкретного необходимого белка, слитого с FKBP или его производным. Поскольку экспрессия гена/функция кодируемого продукта не должна быть существенным образом затронута в результате присоединения FKBP или его производного к необходимому белку в присутствии Shield-1, rdCMV в присутствии Shield-1 должен реплицироваться со скоростью, сопоставимой со скоростью репликации родительского CMV (предпочтительно на уровне по меньшей мере 75%, 80%, 90%, 95%, 99% или 100% от уровня родительского вируса). Репликация rdCMV существенно отличается от репликации родительского CMV в отсутствие Shield-1 (уменьшается предпочтительно более чем на 50%, 75%, 90%, 95%, 99% или 100% по сравнению с репликацией CMV, который не содержит дестабилизирующий слитый белок).
В предпочтительных вариантах осуществления rdCMV в присутствии по меньшей мере 2 мкМ Shield-1 реплицируется предпочтительно на уровне по меньшей мере 90%, более предпочтительно по меньшей мере 95%, наиболее предпочтительно по меньшей мере 99% от уровня, на котором реплицируется не-rdCMV.
В одном варианте осуществления композиция, содержащая rdCMV по изобретению, имеет титр вируса по меньшей мере 105 БОЕ/мл, более предпочтительно по меньшей мере 107БОЕ/мл, в присутствии по меньшей мере 2 мкМ Shield-1.
Напротив, rdCMV не должен существенно реплицироваться в отсутствие Shield-1. О качестве механизма репликационного дефекта судят по тому, насколько строгим является контроль в условиях, запрещенных для вирусной репликации, то есть, каковы инфекционные титры вирионов потомства в таких условиях. rdCMV по настоящему изобретению практически не способен реплицироваться (как в клеточной культуре, так и в организме пациента) без присутствия Shield-1. Его репликация в клетках ARPE-19 и других типах первичных клеток человека является условной, и молярная концентрация Shield-1 выше 0,1 мкМ, предпочтительно по меньшей мере 2 мкМ, в культуральной среде необходима для поддержания вирусной репликации.
В одном варианте осуществления композиция, содержащая rdCMV по изобретению, имеет вирусный титр менее 2 БОЕ/мл, более предпочтительно менее 1 БОЕ/мл, в отсутствие Shield-1. Способы оценки репликации CMV можно использовать, чтобы оценивать репликацию rdCMV либо в отсутствие, либо в присутствии Shield-1. Однако в предпочтительных вариантах осуществления используют TCID50.
В другом варианте осуществления титры rdCMV определяют при помощи анализа для определения 50% инфекционной дозы для тканевой культуры (TCID50). Вкратце, этот анализ с разбавлением позволяет рассчитать количество вируса, необходимое для гибели 50% инфицированных хозяев. Клетки-хозяева (например, клетки ARPE-19) высевают, и к ним добавляют вирус в серийных разведениях. После инкубации определяют и регистрируют процент гибели клеток (то есть, инфицированных клеток) для каждого разведения вируса. Результаты используют, чтобы математически рассчитать TCID50.
В другом варианте осуществления титры rdCMV определяют при помощи анализа бляшкообразования. Анализ на образование вирусных бляшек позволяет определять количество бляшкообразующих единиц (БОЕ) в образце вируса. Вкратце, слившийся монослой клеток-хозяев (например, клеток ARPE-19) инфицируют rdCMV в различных разведениях и покрывают полутвердой средой, такой как агар или карбоксиметилцеллюлоза, для предотвращения беспорядочного распространения вирусной инфекции. Вирусная бляшка образуется, когда вирус инфицирует клетку в фиксированном клеточном монослое. Инфицированные вирусом клетки будут лизироваться и распространять инфекцию в соседние клетки, и цикл инфицирование-лизис будет повторяться. Область инфицированных клеток образует бляшку (область инфекции, окруженную неинфицированными клетками), которую можно увидеть визуально или с помощью оптического микроскопа. Бляшки подсчитывают, и результаты в сочетании с коэффициентом разведения, используемым для конкретной чашки, используют для расчета количества бляшкообразующих единиц на единицу объема образца (БОЕ/мл). Полученное значение БОЕ/мл соответствует количеству инфекционных частиц в образце, и оно основано на предположении, что каждая образованная бляшка представляет одну инфекционную вирусную частицу.
В другом варианте осуществления модель на мышах hu-SCID используют для оценки способности rdCMV реплицироваться in vivo. Вкратце, кусочки тканей плода человека (например, тимуса и печени) хирургическим путем имплантируют в почечную капсулу мышей SCID. rdCMV инокулируют через 2-3 месяца, когда человеческие ткани становятся васкуляризированными. Вирусные титры оценивают через 3-4 недели после инокуляции в анализах на бляшкообразование. Эксперименты на животных можно проводить в отсутствие или в присутствии Shield-1.
Оценка иммунного ответа
Введение rdCMV по изобретению пациенту вызывает иммунный ответ на CMV, предпочтительно защитный иммунный ответ, который способен лечить и/или уменьшать вероятность инфекции CMV или патологии, связанной с такой инфекцией, у пациента. Иммунный ответ возникает, по меньшей мере частично, из-за пентамерного gH-комплекса.
Иммунный ответ, вызванный rdCMV, можно оценивать с помощью методов, известных в данной области.
Животные модели, известные в данной области, можно использовать для оценки защитного эффекта от введения rdCMV. В одном варианте осуществления иммунную сыворотку от индивидуумов, которым вводили rdCMV, можно анализировать на нейтрализующую способность, включая, но без ограничения, блокирование прикрепления вируса или проникновения вируса в клетку-хозяина. В других вариантах осуществления T-клетки от индивидуумов, которым вводили rdCMV, можно анализировать на способность продуцировать цитокины, включая, но без ограничения, интерферон-гамма, в присутствии интересующего антигена. Животные модели со стимуляцией антигеном также можно использовать для определения иммунологически эффективного количества иммуногена.
Нейтрализация вируса означает, что специфичные для вируса антитела способны препятствовать проникновению и/или репликации вируса в культурах. Общепринятым анализом для измерения нейтрализующей активности является анализ на уменьшение бляшкообразования. Анализы на нейтрализацию в данном изобретении означают титрование сыворотки, способной блокировать проникновение вируса в клетки. Титры NT50 определяют как реципрокные разведения сыворотки, необходимые для блокирования 50% внесенного вируса в анализах на нейтрализацию вируса. Титры NT50 получают из аппроксимации нелинейной логистической четырехпараметрической кривой.
Получение дефектного по репликации CMV
Настоящее изобретение относится к способам получения rdCMV. rdCMV по изобретению размножают в присутствии стабилизатора, такого как Shield-1, на эпителиальных клетках, предпочтительно человеческих эпителиальных клетках, и более предпочтительно человеческих пигментированных эпителиальных клетках сетчатки. В дополнительных вариантах осуществления человеческие пигментированные эпителиальные клетки сетчатки представляют собой клетки ARPE-19, депонированные в Американской коллекции типовых культур (ATCC) с регистрационным № CRL-2302. В некоторых вариантах осуществления Shield-1 присутствует в средах для тканевых культур в концентрации по меньшей мере 0,5 мкМ. В предпочтительных вариантах осуществления Shield-1 присутствует в средах для тканевых культур в концентрации по меньшей мере 2,0 мкМ.
В некоторых вариантах осуществления клетки, используемые для размножения rdCMV, выращивают на микроносителях. Микроноситель представляет собой поддерживающую матрицу, позволяющую расти прикрепляющимся клеткам во вращающихся колбах или биореакторах (таких как микрогравитационные биореакторы с вращающимися стенами). Микроносители, как правило, представляют собой 125-250-мкм сферы, имеющие плотность, которая позволяет им поддерживаться в суспензии при осторожном перемешивании. Микроносители могут быть выполнены из различных материалов, включая, но без ограничения, DEAE-декстран, стекло, полистирольный пластик, акриламид и коллаген. Микроносители могут иметь различный химический состав поверхности, включая, но без ограничения, белки внеклеточного матрикса, рекомбинантные белки, пептиды и заряженные молекулы. Можно также использовать другие системы клеточных культур высокой плотности, такие как системы Corning HyperFlask® и HyperStack®.
Бесклеточную среду для тканевых культур можно собирать и выделять из нее rdCMV. Вирусные частицы CMV имеют примерно 200 нм в диаметре и могут быть отделены от других белков, присутствующих в собранной среде, с помощью методов, известных в данной области, включая, но без ограничения, ультрацентрифугирование в градиенте плотности или в "подушке" из 20% сорбита. Содержание белка в вакцинах можно определять анализом по Брэдфорду.
Для контроля репликации rdCMV можно использовать Shield-1 в сочетании с FKBP. После того, как выращивание нужного количества вирусов в клетках тканевой культуры будет завершено, способность к репликации перестает быть нужной. Shield-1 выводят из rdCMV, чтобы сделать вирус дефицитным по репликации (например, для введения пациенту). В одном варианте осуществления rdCMV очищают от Shield-1 промыванием один или более раз. В другом варианте осуществления rdCMV очищают от Shield-1 ультрацентрифугированием. В другом варианте осуществления rdCMV очищают от Shield-1 диафильтрацией. Как правило, для очистки вирусных частиц используют диафильтрацию. В одном варианте осуществления используют фильтры с размером пор примерно 750 килодальтон, через которые может проходить только Shield-1.
После очистки rdCMV от Shield-1 в композиции rdCMV может оставаться небольшое количество остаточного Shield-1. В одном варианте осуществления уровень Shld-1 в композиции CMV после очистки по меньшей мере в 100 раз ниже уровня, необходимого для поддержания репликации в тканевой культуре. В другом варианте осуществления уровень Shld-1 в композиции CMV после очистки составляет 0,1 мкМ или менее. В другом варианте осуществления уровень Shld-1 в композиции CMV после очистки не поддается определению.
Определение уровней Shield-1 в композиции можно проводить при помощи ЖХ/МС (жидкостная хроматография/масс-спектроскопия) или ВЭЖХ/МС (высокоэффективная жидкостная хроматография/масс-спектроскопия) анализов. Данные методы сочетают возможности физического разделения ЖХ или ВЭЖХ с возможностями анализа масс и позволяют обнаруживать интересующие химические вещества в комплексных смесях.
Адъюванты
Адъюванты представляют собой вещества, которые способны помогать иммуногену вызывать иммунный ответ. Адъюванты могут иметь различные механизмы действия, например, один или более из следующих: увеличение биологического или иммунологического времени полураспада антигена; улучшение доставки антигена к антигенпредставляющим клеткам; улучшение процессинга и представления антигена антигенпредставляющими клетками; возможность щадящего режима дозирования и индукция выработки иммуномодулирующих цитокинов (Vogel, 2000, Clin Infect Dis 30: S266). В некоторых вариантах осуществления композиции по изобретению содержат rdCMV и адъювант.
Можно использовать адъюванты различных типов, чтобы способствовать развитию иммунного ответа. Примеры конкретных адъювантов включают гидроксид алюминия; фосфат алюминия, гидроксифосфат алюминия, адъювант аморфный гидроксифосфат-сульфат алюминия (AAHSA) или другие соли алюминия; фосфат кальция; мотивы CpG ДНК; монофосфориллипид A; холерный токсин; термолабильный токсин E. coli; коклюшный токсин; мурамилдипептид; неполный адъювант Фрейнда; MF59; SAF; иммуностимулирующие комплексы; липосомы; биоразлагаемые микросферы; сапонины; неионные блок-сополимеры; аналоги мурамилпептида; полифосфазен; синтетические полинуклеотиды; IFN-γ; IL-2; IL-12 и ISCOMS. (Vogel, 2000, Clin Infect Dis 30: S266; Klein et al., 2000, J Pharm Sci 89: 311; Rimmelzwaan et al., 2001, Vaccine 19: 1180; Kersten, 2003, Vaccine 21: 915; O'Hagen, 2001, Curr. Drug Target Infect. Disord. 1: 273.)
В некоторых вариантах осуществления адъюванты на масляной основе, включая, но без ограничения, неполный адъювант Фрейнда и MF59, не используют в композициях по изобретению.
В других вариантах осуществления адъюванты в виде частиц, включая, но без ограничения, адъювант ISCOMATRIX® и/или алюминий-фосфатный адъювант, используют в композициях по изобретению.
Фармацевтические композиции
Еще одной особенностью изобретения является применение рекомбинантного CMV, описанного здесь, в композиции, предпочтительно иммуногенной композиции или вакцине, для лечения пациентов с CMV инфекцией и/или снижения вероятности CMV инфекции. Соответственно, композиция содержит фармацевтически приемлемый носитель. "Фармацевтически приемлемый носитель" означает жидкий наполнитель, разбавитель или инкапсулирующее вещество, которые можно безопасно использовать для системного введения. В зависимости от конкретного способа введения можно использовать разнообразные фармацевтически приемлемые носители, хорошо известные в данной области. Эти носители можно выбирать из группы, включающей сахара, крахмалы, целлюлозу и ее производные, солод, желатин, тальк, сульфат кальция, растительные масла, синтетические масла, полиолы, альгиновую кислоту, фосфатные буферные растворы, включая фосфатно-солевой буфер, эмульгаторы, изотонический солевой раствор и апирогенную воду. В частности, фармацевтически приемлемые носители могут содержать различные компоненты, такие как буфер, стерильная вода для инъекций, нормальный солевой раствор или фосфатно-солевой буфер, сахароза, гистидин, соли и полисорбат. Такие термины, как "физиологически приемлемый", "разбавитель" или "эксципиент", можно использовать взаимозаменяемо.
Способы получения вакцинных препаратов раскрыты, например, в книге "New Generation Vaccines" (1997, Levine et al., Marcel Dekker, Inc. New York, Basel, Hong Kong), включенной в данный документ посредством ссылки.
Препараты
В некоторых вариантах осуществления rdCMV по изобретению вводят пациенту, чтобы вызвать иммунный ответ. Желательно минимизировать или избегать потери активности композиции rdCMV во время хранения иммуногенной композиции. Условия для достижения этой цели включают, но без ограничения, (1) устойчивую стабильность при хранении, (2) устойчивость к стрессу циклов замораживания-размораживания, (3) стабильность при температурах окружающей среды на срок до недели, (4) сохранение иммуногенности, (5) совместимость со стратегией использования адъювантов. Условия, которые влияют на стабильность rdCMV, включают, но без ограничения, pH буфера, ионную силу буфера, наличие/отсутствие конкретных эксципиентов и температуру. Композиции содержат буферы для повышения стабильности очищенных вирусных частиц rdCMV, подходящих в качестве вакцинной композиции.
Сохранение целостности вирусных частиц можно оценивать с помощью анализов на иммуногенность, проводимых на мышах, и/или анализов на проникновение вируса. Способность вируса к проникновению зависит от целостности и функций вирусных гликопротеинов, включая пентамерный gH-комплекс. Пентамерный gH-комплекс также в значительной степени обеспечивает иммуногенность rdCMV, таким образом, эти два свойства связаны.
В некоторых вариантах осуществления rdCMV хранят в буфере, содержащем 15-35 мМ гистидина и 100-200 мМ NaCl при значении pH от 5 до 7. В более конкретном варианте осуществления буфер содержит 25 мМ гистидина и 150 мМ NaCl при pH 6.
В других вариантах осуществления можно добавлять сахара для обеспечения дополнительной стабильности, например, полиолы (включая, но без ограничения, маннит и сорбит); моносахариды (включая, но без ограничения, глюкозу, маннозу, галактозу и фруктозу); дисахариды (включая, но без ограничения, лактозу, мальтозу, мальтозу, сахарозу, лактулозу и трегалозу) и трисахариды (включая, но без ограничения, рафинозу и мелизитозу). В более конкретном варианте осуществления сахар представляет собой сахарозу. В еще более конкретном варианте осуществления сахароза составляет 5-15%.
В предпочтительных вариантах осуществления rdCMV хранят в буфере, содержащем 25 мМ гистидина, 150 мМ NaCl, 9% сахарозы при pH 6.
Введение
rdCMV, описанный здесь, можно формулировать и вводить пациенту, используя руководство, предоставленное здесь, наряду с методами, хорошо известными в данной области. Руководство для фармацевтического введения, как правило, можно найти, например, в книгах "Vaccines" под редакцией Plotkin и Orenstein, W.B. Sanders Company, 1999; "Remington's Pharmaceutical Sciences" 20-е издание, под редакцией Gennaro, Mack Publishing, 2000; и "Modern Pharmaceutics" 2-е издание, под редакцией Banker и Rhodes, Marcel Dekker, Inc., 1990.
Вакцины можно вводить различными способами, например, подкожным, внутримышечным, внутривенным, на слизистые оболочки, парентеральным, чрескожным или внутрикожным. Подкожное и внутримышечное введение можно выполнять с использованием, например, игл или безыгольных шприцев. В одном из вариантов осуществления вакцину по изобретению вводят внутримышечно. Чрескожную или внутрикожную доставку можно выполнять при помощи шприца с иглой для внутрикожных инъекций или передовых устройств, таких как микронные иглы или пластыри с микрочипами.
Композиции, описанные здесь, можно вводить способом, совместимым с дозированной препаративной формой, и в таком количестве, которое иммунологически эффективно для лечения и/или снижения вероятности CMV инфекции (включая первичную, рецидивирующую и/или суперинфекцию). Доза, вводимая пациенту, в контексте настоящего изобретения, должна быть достаточной для оказания благотворного влияния на пациента с течением времени, например, для снижения уровня CMV инфекции, облегчения симптомов заболевания, связанного с CMV инфекцией, и/или уменьшения продолжительности и/или тяжести CMV инфекции, или для снижения вероятности CMV инфекции (включая первичную, рецидивирующую и/или суперинфекцию).
Подходящие режимы дозирования могут быть легко определены специалистами в данной области и предпочтительно определены с учетом факторов, хорошо известных в данной области, включая возраст, массу тела, пол и медицинское состояние пациента, способ введения, желаемый эффект и конкретную используемую композицию. При определении эффективного количества rdCMV, которое нужно вводить для лечения или профилактики CMV, врач может оценивать циркулирующие уровни вируса в плазме, прогрессирование заболевания и/или продукцию анти-CMV антител. Доза для вакцинной композиции находится в диапазоне от 103 до 1012 бляшкообразующих единиц (БОЕ). В различных вариантах осуществления используют дозы в диапазоне от 104 до 1010 БОЕ, от 105 до 109 БОЕ, от 106 до 108 БОЕ или любую дозу в этих указанных диапазонах. При введении более одной вакцины (то есть, в комбинированных вакцинах) количество каждого действующего вещества вакцины находится в описанном для него диапазоне.
Вакцинную композицию можно вводить одной дозой или в многодозовом формате. Вакцины можно готовить с адъювантом за несколько часов или дней до введений, в зависимости от определения стабилизирующего буфера(-ов) и подходящего адъювантного состава. Вакцины можно вводить в объемах, которые обычно практикуются, в диапазоне от 0,1 мл до 0,5 мл.
Выбор времени дозирования зависит от факторов, хорошо известных в данной области. После первоначального введения одну или более дополнительных доз можно вводить для поддержания и/или повышения титров антител и T-клеточного иммунитета. Дополнительные бустерные введения могут потребоваться для поддержания защитных уровней иммунных ответов, которые находят отражение в титрах антител и показателях T-клеточного иммунитета, определяемых, например, анализом ELISPOT. Уровни таких иммунных ответов зависят от клинических исследований.
В случае комбинированных вакцинаций, все иммуногены можно вводить вместе в одной композиции или отдельно в разных композициях. rdCMV, описанный здесь, вводят одновременно с одним или более из желаемых иммуногенов. Термин "одновременно" означает не только введение терапевтических средств точно в одно и то же время, он скорее означает, что rdCMV, описанный здесь, и другой желаемый иммуноген(-ы) вводят субъекту в такой последовательности и в таком временном интервале, что они могут действовать совместно, принося больше пользы, чем если бы их вводили иным образом. Например, все терапевтические средства можно вводить одновременно или последовательно в любом порядке в различные моменты времени; однако, если их не вводить одновременно, то они должны быть введены достаточно близко по времени, чтобы обеспечить желаемый терапевтический эффект. Каждое терапевтическое средство можно вводить отдельно, в любой подходящей форме и любым подходящим способом введения.
Популяция пациентов
Термин "пациент" относится к млекопитающему, которое может быть инфицировано CMV. В предпочтительном варианте осуществления пациентом является человек. Пациент может получать профилактическое или терапевтическое лечение. Профилактическое лечение обеспечивает достаточный защитный иммунитет для снижения вероятности или тяжести CMV инфекции, включая первичные инфекции, рецидивирующие инфекции (то есть, те, которые возникают в результате реактивации латентного CMV) и суперинфекции (то есть, те, которые возникают в результате инфицирования штаммом CMV, отличающимся от того, которым пациент был инфицирован ранее). Терапевтическое лечение можно проводить для снижения тяжести CMV инфекции или снижения вероятности/тяжести рецидивирующей или суперинфекции.
Лечение можно проводить с использованием фармацевтической композиции, содержащей rdCMV, описанный здесь. Фармацевтические композиции можно вводить всем слоям населения, особенно тем лицам, которые имеют повышенный риск получения CMV инфекции (как первичной, так и рецидивирующей или суперинфекции), или для которых CMV инфекция была бы особенно проблематичной (например, лицам с иммунной недостаточностью, пациентам с трансплантатами или беременным женщинам). В одном варианте осуществления женщин детородного возраста, особенно девочек-подростков, вакцинируют для снижения вероятности CMV инфекции (как первичной, так и рецидивирующей или суперинфекции) во время беременности.
Нуждающиеся в лечении включают тех, которые уже имеют инфекцию, а также тех, которые предрасположены к инфекции или для которых было бы желательным снижение вероятности получения инфекции. Лечение может облегчать симптомы заболевания, связанного с CMV инфекцией, и/или уменьшать продолжительность и/или тяжесть CMV инфекции, включая инфекцию вследствие реактивации латентного CMV.
Лица с повышенным риском получения CMV инфекции (как первичной, так и рецидивирующей или суперинфекции) включают пациентов с ослабленным иммунитетом или пациентов, которым предстоит терапия, приводящая к ослаблению иммунитета (например, проходящих химиотерапию или радиотерапию вследствие онкологического заболевания или принимающих подавляющие иммунитет лекарственные средства). Используемый здесь термин "ослабленный иммунитет" относится к иммунной системе, которая в меньшей степени способна бороться с инфекциями из-за того, что иммунный ответ неправильно функционирует или не функционирует на уровне, соответствующем уровню у нормального здорового взрослого человека. Примерами пациентов с ослабленным иммунитетом являются младенцы, дети младшего возраста, пожилые, беременные или пациенты с заболеванием, которое влияет на функцию иммунной системы, таким как ВИЧ-инфекция или СПИД.
ПРИМЕРЫ
Ниже приведены примеры для дальнейшей иллюстрации различных отличительных признаков настоящего изобретения. Примеры также иллюстрируют полезную методологию для практического применения изобретения. Данные примеры не ограничивают заявленное изобретение.
Пример 1: Восстановление пентамерного gH-комплекса
Инфекционный клон CMV с бактериальной искусственной хромосомой был сконструирован так, что был закодирован вирион, экспрессирующий пентамерный gH-комплекс, состоящий из UL128, UL130 и UL131, собранных в каркасную структуру gH/gL.
Штамм AD169 CMV первоначально был выделен из аденоидов 7-летней девочки (Elek and Stern, 1974, Lancet, 1: 1). Было проведено 58 пассажей вируса в человеческих фибробластах различных типов для аттенуации вируса (Neff et al., 1979, Proc Soc Exp Biol Med, 160: 32), с последними 5 пассажами в человеческих фибробластах WI-38. Этот пассированный вариант вируса AD169, названный в данном исследовании Merck AD169 (MAD169), использовали в качестве родительского вируса для конструирования инфекционного BAC-клона. Ни родительский вирус AD169, ни пассированный вариант вируса MAD169 не экспрессировали UL131 или пентамерный gH-комплекс.
MAD169 использовали в качестве родительского вируса для конструирования инфекционного клона с бактериальной искусственной хромосомой (BAC). BAC-вектор представляет собой молекулярный инструмент, позволяющий проводить генетические манипуляции с фрагментом ДНК большого размера, таким как геном CMV (~230 т.п.н.), в E. coli. Элемент BAC вместе с маркерным геном GFP встраивали сразу после стоп-кодона открытой рамки считывания US28 (между ORF US28 и US29 в вирусном геноме) с сайтом LoxP, созданным на обоих концах фрагмента (фиг.1A). Вкратце, фрагмент ДНК, содержащий экспрессионную кассету GFP, фланкированную двумя сайтами loxP, и последовательности US28-US29 CMV, синтезировали и клонировали в вектор pBeloBAC11. BAC-вектор линеаризовали ферментом рестрикции Pme I и совместно трансфицировали в клетки MRC-5 с ДНК MAD169, экстрагированной из очищенных вирионов. Рекомбинантные варианты, идентифицированные по экспрессии зеленого флуоресцентного белка, были очищены путем образования бляшек. После одного цикла амплификации кольцевую форму вирусного генома извлекали из инфицированных клеток и вводили электропорацией в клетки DH10 E. coli. Проводили скрининг бактериальных колоний методом ПЦР на присутствие областей US28 и US29. Колонии-кандидаты дополнительно изучали рестрикционным анализом с использованием EcoR I, EcoR V, Hind III, Spe I и Bam HI. После скрининга один клон, bMAD-GFP, продемонстрировал одинаковую картину рестрикции с родительским вирусом MAD169.
Мутацию со сдвигом рамки в первом экзоне UL131, являющуюся причиной отсутствия тропизма к эпителиальным клеткам в MAD169, исправляли генетически в E. coli (фиг.1B). В частности, один адениновый нуклеотид (н) из 7-н A-фрагмента в гене UL131 был делетирован (фиг.1B). Делеции 1 н было достаточно для восстановления тропизма к эпителиальным и эндотелиальным клеткам за счет UL131, и, таким образом, пентамерный gH-комплекс вновь экспрессировался. Экспрессию подтверждали методами ELISA и вестерн-блоттинга (данные не представлены). Данный клон был дополнительно модифицирован путем удаления сегмента BAC посредством рекомбинации LoxP/Cre. ДНК BAC трансфицировали в клетки ARPE-19, человеческие пигментированные эпителиальные клетки сетчатки (ATCC, регистрационный № CRL-2302), для восстановления инфекционного вируса (фиг.1C). Полученный инфекционный вирус, названный BAC-производным вирусом MAD169 с тропизмом к эпителиальным клеткам (beMAD), отличался от MAD169 только по двум локусам, (1) ORF UL131, в которой один адениновый нуклеотид был делетирован, и (2) сайтом LoxP из 34 п. о., вставленным между ORF US28 и US29 (см. таблицу 2).
Геном BAC-клона beMAD был полностью секвенирован. Общая структура генома beMAD идентична той, о которой сообщалось для варианта AD169 ATCC (GenBank, регистрационный № X17403), которая состоит из двух уникальных областей, уникальной длинной (UL) и уникальной короткой (US). Каждая уникальная область ограничена двумя последовательностями повторов, концевым повтором длинной (TRL)-внутренним повтором длинной (IRL), концевым повтором короткой (TRS)-внутренним повтором короткой (IRS). Кинетики роста пассированного варианта MAD169 и производного вируса beMAD были неотличимы в клетках MRC-5, линии клеток человеческих фибробластов (ATCC, регистрационный № CCL-171) (данные не представлены). Поскольку gH-комплекс не является необходимым для роста на фибробластах, различия в экспрессии gH-комплекса между MAD169 и beMAD не имеют значения.
Пример 2: Влияние обычных методов инактивации на gH-комплекс
Изучали влияние двух общепринятых методов вирусной инактивации, γ-облучения и β-пропиолактона (BPL), на CMV, экспрессирующий gH.
γ-Облучение проводили на лиофилизированных вирионах. Вакцинный препарат рекомбинантного CMV в концентрации 0,15 мг/мл в HNS (25 мМ гистидина, 150 мМ NaCl, 9% масс./об. сахарозы, pH 6,0) лиофилизировали с использованием консервативного цикла лиофилизации (замораживание при -50°C и первичное высушивание при -35°C в течение ~30 часов с последующим вторичным высушиванием при 25°C в течение 6 часов), получая сухой порошок. Вакцину лиофилизировали в 3-мл стеклянных флаконах, внося по 0,5 мл в каждый флакон. В конце лиофилизации флаконы закрывали пробками в атмосфере азота, и образцы вынимали, наклеивали этикетки, укупоривали обжимом и хранили при -70°C до проведения гамма-облучения. Флаконы облучали в облучателе с Co до достижения необходимой дозы облучения.
Для обработки BPL маточный раствор BPL добавляли к неочищенному вирусному культуральному супернатанту, полученному после роста на клетках ARPE-19, до достижения конечных концентраций 0,01% или 0,1% (по объему). Реакцию останавливали добавлением тиосульфата натрия в различные моменты времени. Затем обработанные BPL экспрессирующие gH CMV очищали ультрацентрифугированием.
Кинетику инактивации для обоих способов определяли анализом бляшкообразования на клетках ARPE-19. Вкратце, получали серийные разведения вирусных образцов в PBS, и 0,1 мл использовали для инокуляции в каждую лунку 6-луночного планшета, в котором находились высеянные клетки ARPE-19. Планшеты инкубировали при 37°C в течение 1 часа перед добавлением по 6 мл в лунку покрывающей среды, содержащей 0,5% агарозы. Планшеты инкубировали в течение 18 дней при 37°C. Для визуализации бляшек в каждую лунку добавляли примерно 0,5 мл раствора MTT в концентрации 5 мг/мл (тиазолил синий тетразолий бромид, Sigma M5655). Планшеты инкубировали при 37°C в течение 2-4 часов, и бляшки подсчитывали с помощью устройства для просмотра с подсветкой (фиг.2A и 2C).
Иммуногенность инактивированного экспрессирующего gH CMV анализировали путем определения титров нейтрализующих антител у мышей. Вкратце, самок мышей Balb/c (n=10) иммунизировали 2,5 мкг CMV на дозу в недели 0 и 3. Сыворотки собирали в неделю 4 и оценивали на нейтрализующую активность против проникновения вируса в эпителиальные клетки. Титр нейтрализующих антител (NT50) определяли как реципрокное разведение сыворотки, приводящее к 50% уменьшению проникновения вируса в эпителиальные клетки по сравнению с отрицательным контролем. Результаты изучения иммуногенности на мышах продемонстрировали, что оба общепринятых способа инактивации отрицательно влияли на титры нейтрализующих антител, выработанных в ответ на экспрессирующий gH CMV (фиг.2B и 2D). Снижение титров NT50 коррелировало с продолжительностью обработки γ-облучением или BPL. Длительная обработка делала CMV, экспрессирующий пентамерный gH-комплекс, более похожим на родительский AD169 CMV в отношении иммуногенности у мышей. Аналогичные результаты были получены на кроликах и макаках-резусах при тестировании вакцин, инактивированных γ-облучением или BPL (данные не представлены). Эти наблюдения показали, что пентамерный gH-комплекс чувствителен к обоим методам инактивации при выбранных условиях инактивации.
Пример 3: Конструирование и скрининг слитых продуктов FKBP-необходимый белок
CMV конструировали с использованием в качестве основы аттенуированного штамма AD169 с восстановленным тропизмом к эпителиальным клеткам, который в то же время является условно дефектным по репликации. Для восстановления тропизма к эпителиальным клеткам использовали способы, описанные в примере 1.
Вирусные белки для слияния с производным FKBP выбирали на основе двух критериев. Во-первых, интересующие белки не были обнаружены в вирионах CMV протеомным анализом (Varnum et al., 2004, J. Virol. 78: 10960), таким образом, была снижена вероятность того, что слитый с FKBP белок будет включен в вирус. Во-вторых, интересующие белки являлись необходимыми для вирусной репликации в тканевой культуре.
Используя beMAD в качестве родительского вируса, проводили слияние производного FKBP (SEQ ID NO: 12) с 12 необходимыми вирусными белками в отдельности, включая IE 1/2 (SEQ ID NO: 1), pUL37x1, pUL44, pUL51 (SEQ ID NO: 3), pUL52 (SEQ ID NO: 5), pUL53, pUL56, pUL77, pUL79 (SEQ ID NO: 7), pUL84 (SEQ ID NO: 9), pUL87 и pUL105. Также конструировали вирус с двумя различными необходимыми белками, слитыми с FKBP, при этом каждый из IE1/2 и UL51 был слит с производным FKBP (геном rdCMV с дестабилизированными IE1/2 и UL51 приведен в SEQ ID NO: 14). После конструирования все рекомбинантные ДНК BAC трансфицировали в клетки ARPE-19 и культивировали в среде, содержащей Shld-1.
Изучали зависимость роста вируса от Shld-1. Вирусы со слитыми IE1/2, UL51, UL52, UL84, UL79 и UL87 легко выживали в анализе на бляшкообразование с добавлением 2 мкМ Shld-1 (данные не представлены). Вирусы с UL37x1, UL77 и UL53 также образовывали бляшки, но бляшки были небольшими, и они росли значительно медленнее по сравнению с родительским beMAD. Увеличение концентрации Shld-1 до 10 мкМ существенно не ускоряло рост вирусов (данные не представлены). Вирусы со слитыми UL56 и UL105 не выживали, что свидетельствовало о том, что слияние этих белков нарушает их функции или экспрессию соседних генов.
Варьирующие концентрации Shld-1 использовали в дополнительных экспериментах для дальнейшей оценки вирусной репликации в присутствии или в отсутствие Shld-1. Клетки ARPE-19 инфицировали экспрессирующим gH CMV, который также содержал производное FKBP, слитое с необходимым белком, при MOI 0,01 БОЕ/мл. После инфицирования в течение 1 часа клетки дважды промывали свежей средой для удаления Shld-1 из инокулятов. Затем инокуляты добавляли к клеткам ARPE-19, культивируемым в среде, содержащей 0,05, 0,1, 0,5 или 2 мкМ Shield-1. Через семь дней после инфицирования внеклеточное потомство вируса в супернатанте собирали и титровали на клетках ARPE-19 при добавлении 2 мМ Shield-1. Титры вируса определяли при помощи анализа для определения 50% инфекционной дозы для тканевой культуры (TCID50). Вкратце, этот анализ с разбавлением позволяет рассчитать количество вируса, необходимое для гибели 50% инфицированных хозяев. Клетки ARPE-19 высевали и добавляли вирус в серийных разведениях. После инкубации процент клеточной гибели (то есть, инфицированных клеток) вручную подсчитывали и регистрировали для каждого разведения вируса. Результаты использовали для математического расчета TCID50.
Как показано на фиг. 3, эффективная репликация всех CMV, содержащих продукты слияния с FKBP, зависела от концентрации Shield-1, хотя и в разной степени. Более низкая концентрация Shield-1, как правило, приводила к снижению титра полученного потомства вируса. Среди вирусов с одиночным слиянием только UL51 и UL52 безусловно нуждались в Shield-1 для репликации. Другие вирусы с одиночным слиянием, IE1/2, UL84, UL79, и UL87, были способны производить поддающееся определению вирусное потомство в отсутствие Shield-1. Это правило было особенно строгим, когда производное FKBP было слито с UL51 или UL52.
Кинетику роста вирусов со слитыми IE1/2, UL51, IE1/2-UL51 сравнивали с кинетикой родительского вируса beMAD в присутствии или в отсутствие 2 мкМ Shld-1. Как показано на фиг. 4, в присутствии Shld-1 вирусы с одиночным или двойным слиянием демонстрировали кинетику роста, сравнимую с таковой у родительского beMAD. Однако в отсутствие Shld-1 только вирус IE1/2 был способен реплицироваться, хотя и в меньшей степени и с меньшей скоростью, чем родительский beMAD.
Степень контроля вирусной репликации в вирусе с двойным слиянием также тестировали в клетках различного типа (фиг.5). Эти клетки включали клетки пупочной вены человека (HUVEC), фибробласты MRC-5, гладкомышечные клетки аорты (AoMC), клетки скелетных мышц (SKMC) и клетки CCF-STTG1 астроцитомы. Клетки инфицировали вирусом со слитыми IE1/2-UL51 при MOI 0,01 БОЕ/клетку (за исключением CCF-STTG1, которые инфицировали при MOI 5 БОЕ/клетку), а затем инкубировали в среде в присутствии или в отсутствие Shield-1. Клетки всех типов были способны поддерживать литическую вирусную репликацию в присутствии Shield-1. Никакой продукции вируса не наблюдали в отсутствие Shield-1.
Пример 4: Иммуногенность вируса с двойным слиянием IE1/2-UL51 у животных
Иммуногенность вируса с двойным слиянием IE1/2-UL51 оценивали на мышах, кроликах и обезьянах резусах. Сначала сравнивали дозозависимый нейтрализующий ответ против вируса с двойным слиянием IE1/2-UL51 или родительского вируса beMAD у мышей (фиг.6A). Шестинедельных самок мышей BALB/c иммунизировали в недели 0 и 4 вирусом beMAD или вирусом с двойным слиянием IE1/2-UL51 в дозах, находящихся в диапазоне от 0,12 мкг до 10 мкг. Образцы сыворотки в неделю 6 собирали и анализировали микроанализом на нейтрализацию CMV на клетках ARPE-19, как описано ранее (Tang et al., Vaccine, "A novel throughput neutralization assay for supporting clinical evaluations of human cytomegalovirus vaccines" электронная публикация 30 августа 2011 г. на doi: 10.1016/j.vaccine.2011.08.086). Ответы сравнивали при дозах 0,12, 0,37, 1,1, 3,3 и 10 мкг. В диапазоне более низких доз (от 0,12 до 1,1 мкг) beMAD был немного более иммуногенным, при этом нейтрализующие антитела постоянно обнаруживались при уровнях доз выше 0,37 мкг. В диапазоне высоких доз (3,3 и 10 мкг) титры нейтрализующих антител, индуцированных обоими вирусами, были сравнимыми.
Затем сравнивали иммуногенность различных вирусов в дозе 10 мкг на кроликах. Самок кроликов NZW иммунизировали в недели 0, 3 и 8, используя 10 мкг beMAD или указанных вирусов со слитыми белками. В неделю 10 сыворотки собирали и анализировали микроанализом на нейтрализацию CMV на клетках ARPE-19 (фиг.5B). Вирус beMAD, вирусы с одиночным слиянием IE1/2 или UL51 и вирус с двойным слиянием IE1/2-UL51 были способны индуцировать значительно более высокие титры нейтрализующих антител, чем MAD169, вирус, сходный с AD169 и лишенный пентамерного gH-комплекса. Это подтвердило, что экспрессия gH-комплекса вирусом значительно повышает иммуногенность рекомбинантного CMV.
Затем иммуногенность 100 мкг вируса с двойным слиянием IE1/2-UL51 или родительского вируса beMAD тестировали на макаках-резусах. В неделю 12 сыворотки собирали и анализировали микроанализом на нейтрализацию CMV на клетках ARPE-19. GMT титров NT50 в неделю 12 (после дозы 3) составляли 11500 или 15600, соответственно. Эти титры были сопоставимы с титрами NT50, наблюдаемыми у индивидуумов, инфицированных естественным образом (фиг. 5C).
Продолжительность иммунного ответа, индуцированного вакциной вируса CMV с двойным слиянием IE1/2-UL51, была продемонстрирована на макаках-резусах. Животные были вакцинированы либо 10 мкг/дозу, либо 100 мкг/дозу вирусом с двойным слиянием IE1/2-UL51 (в расчете на общую массу белка). Также были включены препараты вакцины 10 мкг/дозу с адъювантом аморфным гидроксифосфат-сульфатом алюминия (AAHS) или ISCOMATRIX®. Вакцины вводили в недели 0, 8 и 24 макакам-резусам (n=5). Для сравнения, контрольная группа получала рекомбинантный gB в количестве 30 мкг/дозу, сформулированный с адъювантом MF59, в недели 0, 4 и 24. Средние геометрические значения для реципрокных титров NT50 (GMT) для всех групп представлены с течением времени (фиг. 7). До вакцинации не было никаких поддающихся определению титров нейтрализующих антител >40 ни у одной из обезьян. Минимальную нейтрализующую активность обнаруживали после первой дозы в неделю 4 для всех групп, с пиками титров нейтрализующих антител приблизительно в неделю 12 и неделю 28 (через четыре недели после второй и третьей вакцинации, соответственно). Пиковое значение GMT в неделю 28 для группы 100 мкг/дозу составляло 14500 (примерно в 3 раза выше, чем титр 4660 для группы 10 мкг/дозу). Адъювант ISCOMATRIX®, но не AAHS, обеспечивал адъювантное преимущество при сравнении с группой 10 мкг/дозу. Значение GMT в неделю 28 для группы ISCOMATRIX® составляло 15800, тогда как для группы AAHS составляло 3000 и для группы 10 мкг/дозу составляло 4660. Минимальную нейтрализующую активность обнаруживали в контрольной (gB/MF59) группе, с пиком GMT никогда не превышающим 200. В неделю 72 исследования, ближе к 1 году после завершения режима вакцинации в недели 0, 8 и 24, величины GMT для группы 100 мкг/дозу и группы препарата с ISCOMATRIX® поддерживались на уровнях 1400 и 3000, соответственно. В это время GMT для группы 10 мкг/дозу и группы с AAHS составляли примерно 200.
Мононуклеарные клетки периферической крови (PBMC) от макак-резусов собирали в неделю 28 (через 4 недели после дозы 3) режима вакцинации и оценивали в анализе ELISPOT на IFN-γ. Обезьян вакцинировали либо 100 мкг/дозу (фиг. 8A), либо 10 мкг/дозу (фиг. 8B-8D) вирусом с двойным слиянием IE1/2-UL51. Кроме того, препарат 10 мкг/дозу составляли либо без адъюванта (фиг. 8B), либо с адъювантами AAHS (фиг. 8C) или ISCOMATRIX® (фиг. 8D). Антигены объединенных перекрывающихся пептидов, представляющие пять антигенов HCMV, использовали для стимулирования продукции IFN-γ ex-vivo. Используемыми антигенами HCMV были IE1 и IE2 (оба вирусные регуляторные белки), а также pp65, gB и pp150 (преобладающие вирусные структурные антигены). Качество T-клеточных ответов оценивали по величине (геометрическим средним) результатов анализа ELISPOT, а также по частоте положительного ответа на вирусные антигены. До вакцинации не было никакого антиген-специфического титра в анализе ELISPOT ни у одной из обезьян (данные не представлены).
В неделю 28 средние геометрические значения ответов в анализе ELISPOT на пять антигенов HCMV (то есть, IE1, IE2, pp65, gB и pp150) составляли 186, 132, 253, 87, 257 образующих "ореол" клеток (SFC)/106 PBMC для группы 100 мкг/дозу против 21, 24, 107, 111, 33 SFC/106 PBMC для группы 10 мкг/дозу, соответственно (фиг.8A и 8B). Частоту положительных ответов в каждой группе (n=5) оценивали на основе критерия порога отсечения более чем 55 SFC/106 PBMC и более чем 3-кратного превышения антиген-специфического ответа над ответом на диметилсульфоксид (ДМСО). Число положительных ответов на антигены HCMV (то есть, IE1, IΕ2, pp65, gB и pp150) составляло 4, 4, 5, 1, 3 для группы 100 мкг/дозу против 1, 1, 5, 4, 0 для группы 10 мкг/дозу.
Влияние адъюванта ISCOMATRIX® на T-клеточные ответы на 10 мкг/дозу вируса с двойным слиянием IE1/2-UL51 показан на фиг. 8D. Средние геометрические значения ответов в анализе ELISPOT на пять антигенов HCMV (то есть, IE1, IE2, pp65, gB и pp150) составляли 114, 53, 491, 85, 113 SFC/106 PBMC, соответственно, и число положительных ответов в группе (n=5) составляло 3, 2, 5, 3, 3, соответственно. Величина и широта T-клеточных ответов в группе с адъювантом ISCOMATRIX® были сходны с таковыми в группе 100 мкг/дозу.
PBMC от животных, вакцинированных либо 10 мкг/дозу, либо 100 мкг/дозу вируса с двойным слиянием IE1/2-UL51 (в расчете на общую массу белка) с ISCOMATRIX® дополнительно анализировали путем внутриклеточного окрашивания цитокинов после стимулирования антигенами HCMV (pp65, IΕ1, IΕ2 или целого вириона HCMV). Отрицательным контролем служили клетки одной интактной обезьяны, не вакцинированной вирусом с двойным слиянием IE1/2-UL51, тогда как положительным контрольным веществом служил стафилококковый энтеротоксин типа B (SEB). На фиг. 9 показано, что отрицательный контроль демонстрировал минимальные ответы на все стимуляции антигенами, но отвечал на положительное контрольное вещество стафилококковый энтеротоксин типа B (SEB), как и ожидалось. У всех десяти вакцинированных обезьян из обеих групп наблюдали ответы на специфические для HCMV антигены с одинаковой величиной и паттернами. Средние геометрические значения для каждого антигена были рассчитаны для всех десяти обезьян. У всех обезьян наблюдали сопоставимые CD8+ (фиг. 9A) и CD4+ (фиг. 9B) T-клеточные ответы, когда их PBMC стимулировали пулами пептидных антигенов CMV (то есть, pp65, IE1 и IE2), но предпочтительно наблюдали CD4+ T-клеточные ответы при стимуляции целыми вирионами HCMV. Это не было неожиданным, так как целые вирионы являются белковыми антигенами, и вероятно, процессируются как экзогенные антигены и представляются молекулами MHC класса II CD4+ T-клеткам. Вирус с двойным слиянием IE1/2-UL51 может вызывать T-клеточные ответы как CD4+, так и CD8+ фенотипов, подобно тому, как это обычно происходит у здоровых субъектов с HCMV инфекцией.
Сравнивали различные препараты вируса с двойным слиянием IE1/2-UL51, содержащие соли алюминия, по их способности вызвать продукцию нейтрализующих антител у макак-резусов (фиг. 10). 30 мкг/дозу вируса с двойным слиянием IE1/2-UL51 формулировали либо с HNS (базовый буфер), либо с адъювантами аморфным гидроксифосфат-сульфатом алюминия (AAHS) или алюминий-фосфатным адъювантом Merck (MAPA), и вводили в недели 0 и 8. Образцы сыворотки, собранные в неделю 12, показали, что, хотя MAPA повышал продукцию нейтрализующих антител, повышение не было статистически значимым (непарный двухсторонний критерий Стьюдента).
Пример 5: Определение буферов для хранения
Вирус CMV в HBSS (сбалансированный солевой раствор Хэнка), хранящийся при -70°C до использования, разбавляли ~10× соответствующим буфером. Остаточные компоненты буфера HBSS в каждом образце включали хлорид калия 0,533 мМ, одноосновный фосфат калия 0,044 мМ, двухосновный фосфат натрия 0,034 мМ, хлорид натрия 13,79 мМ, бикарбонат натрия 0,417 мМ и глюкозу 0,1% масс./об. Затем образцы хранили при комнатной температуре или при температуре от 2°C до 8°C в течение 4 дней, или замораживали и размораживали. Для замораживания-размораживания образец сохраняли при -70°C в течение по меньшей мере 1 часа и размораживали при RT в течение 30 минут на протяжении одного или трех циклов. Стабильность образцов тестировали в день 4 при помощи анализа на проникновение вируса. Вкратце, анализ проводили, используя несколько различных разведений образца для получения кривой ответов, и значения EC50 (мкг/мл) получали на основании результатов анализа на проникновение вируса путем аппроксимации нелинейной кривой. Более низкие значения EC50 соответствуют более высокой стабильности. Значения EC50 образцов для изучения стабильности сравнивали с результатами для замороженного при -70°C контрольного образца.
Анализ на проникновение вируса позволяет измерять способность CMV инфицировать клетки ARPE-19 и экспрессировать IE1 (предранний белок 1). Анализ проводили в прозрачных 96-луночных планшетах. Специфичные для IE1 первичные антитела и биотинилированные вторичные антитела использовали для обнаружения целевых белков в фиксированных клетках, и флуоресцентный сигнал от каждой лунки количественно определяли с использованием конъюгата красителя IR Dye 800CW со стрептавидином, наряду с красителем Sapphire 700/DRAQ5 (для нормирования вклада клеток). На основании результатов строили график зависимости интегрированного отношения интенсивностей 800/700 (интегр. отношение) от концентрации CMV (общий белок, мкг/мл). Значения EC50 также получали из результатов анализа инфекционности путем аппроксимации нелинейной кривой. Поскольку инфицирование клеток ARPE-19 вирусом зависит от целостности вирусных гликопротеиновых антигенов, в частности, пентамерного gH-комплекса, значения EC50 отражают то, насколько хорошо вирусные частицы сохраняются в этих условиях.
Как видно на фиг. 11, CMV теряет инфекционность при хранении в течение четырех дней в HBSS при RT. Более того, 3 цикла замораживания-размораживания в HBSS приводили к полной потере инфекционности, которую оценивали анализом на проникновение вируса. Таким образом, HBSS не являлся оптимальным буфером для хранения CMV.
Влияние pH на стабильность CMV при комнатной температуре изучали в диапазоне pH от 3 до 8. Использовали следующие буферы: цитратный буфер (25 мМ), pH 3,0; ацетатный буфер (25 мМ), pH 4; ацетатный буфер (25 мМ), pH 5; гистидиновый буфер (25 мМ), pH 6; HEPES буфер (25 мМ), pH 7; сбалансированный солевой раствор Хэнка (HBSS), pH 7,5, и Трис-буфер (25 мМ), pH 8.
Образцы готовили разбавлением маточного раствора вируса в 10 раз соответствующим буфером. Образцы хранили при RT (25°C) в течение 4 дней. В день 4 стабильность образцов измеряли с помощью анализа на проникновение вируса. CMV в HBSS, хранящийся замороженным при -70°C, использовали в качестве контроля. Спектры в УФ/видимой области для каждого из образцов получали в момент времени 0 и в день 4 для изучения структурных изменений и агрегации, происходящей во время хранения.
Как показано на фиг. 12, 25 мМ гистидиновый буфер при pH 6 обеспечивал лучшую стабильность CMV, сохраняя более высокую инфекционность при RT по сравнению с другими тестируемыми значениями рН. Второе производное УФ-спектра указывало на схожий структурный профиль вируса при всех значениях pH (данные не представлены). Никакой существенной агрегации не наблюдали ни при одном из тестируемых значений pH, судя по результатам измерения оптической плотности при 350 нм (данные не представлены).
Влияние мочевины самой по себе или в сочетании с хлоридом натрия на стабильность вируса CMV тестировали в 25 мМ гистидиновом буфере, pH 6. Добавление одной 2% мочевины не оказывало влияния на стабильность CMV. Однако 2% мочевина в сочетании с 150 мМ NaCl повышала стабильность CMV при RT (фиг. 13).
Влияние ионной силы на стабильность CMV изучали при pH 6. NaCl в возрастающих концентрациях (0 мМ, 75 мМ, 150 мМ и 320 мМ NaCl) добавляли к 25 мМ гистидиновому буферу при pH 6. Стабильность CMV зависела от ионной силы, причем более высокая ионная сила приводила к более высокой стабильности (фиг. 14). Присутствие мочевины не имело или имело минимальное влияние на стабильность CMV (данные не представлены).
Кроме того, проверяли влияние некоторых других эксципиентов (сахарозы, сорбита, глицерина и пролина) на стабильность экспрессирующего gH CMV при комнатной температуре. Тестируемые эксципиенты добавляли к CMV в 25 мм гистидиновом буфере, pH 6, при комнатной температуре на 4 дня до того, как стабильность вируса CMV измеряли с помощью анализа на проникновение вируса. Для образцов рассчитывали значения EC50. Среди всех протестированных эксципиентов, 150 мМ NaCl отдельно или в сочетании с 9% масс./об. сахарозой обеспечивал более высокую стабильность при pH 6 (данные не представлены). Таким образом, рекомендуемым буфером для хранения CMV при RT является 25 мМ гистидин (pH 6) с 150 мМ NaCl с добавлением или без добавления 9% масс./об. сахарозы.
Изучали влияние криопротекторов на стабильность CMV в процессе замораживания-размораживания. Как указывалось ранее (фиг. 11), CMV в HBSS полностью теряет свою инфекционность, когда его подвергают трем циклам замораживания-размораживания. Различные криопротекторы (включая сахарозу, сорбит, глицерин) тестировали на способность уменьшать стресс для CMV при замораживании-размораживании. Для каждого цикла замораживания-размораживания образцы замораживали при -70°C в течение по меньшей мере 1 часа и размораживали при RT в течение 30 минут. Добавление криопротекторов приводило к повышению стабильности вируса. Кроме того, 9% масс./об. сахарозы в сочетании с 150 мМ хлоридом натрия значительно повышали стабильность вируса по сравнению с другими протестированными криопротекторами (фиг.15). Таким образом, рекомендуемым буфером для хранения CMV при -70°C или при использовании 3 циклов замораживания-размораживания является 25 мМ гистидин, 150 мМ NaCl и 9% сахарозы (буфер HNS).
Буфер HNS сравнивали с буфером HBSS с точки зрения сохранения стабильности CMV в процессе трех циклов замораживания-размораживания, при охлаждении (2-8°C) и RT (25°C). Буфер HNS обеспечивал более высокую стабильность живого вируса CMV при всех протестированных условиях хранения (данные не представлены).
Пример 6: Стабильность CMV в буфере HNS
Сток вируса CMV с двойным слиянием IE1/2-UL51 переводили в буфер HNS и хранили при -70°C до использования. Исследование стабильности проводили при концентрации 100 мкг/мл (в расчете на общее содержание белка, измеренное анализом по Брэдфорду). Маточный раствор вируса разбавляли буфером HNS до конечной концентрации вируса. Затем образцы хранили при соответствующих температурах и тестировали, как описано, в срок до 3 месяцев. Для замораживания-размораживания образцы замораживали при -70°C в течение по меньшей мере 1 часа и размораживали при комнатной температуре в течение 30 минут. Образцы отбирали в различные моменты времени и хранили замороженными при -70°C до анализа.
Общее содержание белка в образцах измеряли анализом по Брэдфорду. Общее содержание белка в образцах не изменялось на протяжении 3-месячного периода (данные не представлены).
Размер частиц CMV в образцах с течением времени контролировали путем измерения гидродинамического диаметра образца методом DLS. Данный метод позволяет отслеживать любую агрегацию или разрушение вирусных частиц с течением времени и при различных температурах хранения. Не наблюдали никаких реальных тенденций для спорадических изменений размеров частиц в конкретных образцах (данные не представлены). Результаты свидетельствовали о том, что вирусные частицы были интактными и не агрегировали при повышенных температурах.
Пример 7: Влияние условий хранения на способность вируса к проникновению и иммуногенность
Значительные изменения в титрах вирусного проникновения (значения EC50) наблюдали, подвергая образцы CMV хранению при различных температурах (данные не представлены). Хранение при -20°C приводило к снижению титров вирусного проникновения по сравнению с хранением при 2-8°C и 25°C. Титры образцов, хранящихся при 2-8°C, как установлено, были ниже по сравнению с титрами вирусного проникновения образов, хранящихся при 25°C. На основании значений EC50 температура хранения была ранжирована в следующем порядке (от максимальной стабильности к минимальной стабильности): 25°C > 2-8°C > -20°C вплоть до временной точки 1 месяц. Титры вирусного проникновения не поддавались обнаружению во временной точке 3 месяца для образцов, хранящихся при -20°C, 2-8°C и 25°C.
Исследование иммуногенности на мышах начинали в конце исследования стабильности, чтобы определить влияние температуры хранения на способность CMV вызывать продукцию нейтрализующих CMV антител. Мышей иммунизировали вакциной по 2,5 мкг на дозу в/м в день 0 и выполняли бустерную инъекцию в день 21 с последующим забором крови в день 28. Мышиную сыворотку тестировали на нейтрализующие антитела против экспрессирующего gH CMV, используя клетки ARPE 19, и титры NT50 получали путем аппроксимации нелинейной кривой.
Оценивали влияние хранения при различных температурах в течение 3 месяцев на иммуногенность CMV с двойным слиянием IE1/2-UL51. Титры NT50 зависели от температуры хранения, причем более высокие температуры приводили к снижению титров по сравнению с контролем, замороженным при -70°C, хотя и незначительно (p=0,2584, однофакторный анализ ANOVA) (фиг. 16A). Титр NT50 для препаратов, хранящихся при -20°C, был понижен менее чем в 2 раза, однако титры в анализе на проникновение вируса для этих образцов были существенно изменены по сравнению с контролем, замороженным при -70°C. Тенденция для титров NT50 образцов для изучения стабильности, хранящихся при -20°C, 2-8°C и 25°C, логически вытекала из титров CMV, полученных методом масс-ELISA для данных образцов.
Оценивали влияние хранения при различных температурах в течение 8 часов после размораживания на иммуногенность CMV с двойным слиянием IE1/2-UL51. Титры NT50 препаратов сравнивали с контролем, замороженным при -70°C. Титры NT50 не менялись (p=0,5865, однофакторный анализ ANOVA) при хранении образцов в течение 8 часов при любой из протестированных температур (фиг. 16B).
В исследовании иммуногенности на мышах изучали влияние хранения CMV с двойным слиянием IE1/2-UL51 при 25°C в течение различных периодов времени после размораживания образцов. Титры NT50 этих препаратов сравнивали с контролем, замороженным при -70°C. Титры NT50 не менялись (p=0,1848, непарный двухсторонний критерий Стьюдента) при хранении образцов при 25°C в течение срока до недели. Через 3 месяца титры NT50 падали немного более чем в 2 раза, указывая на возможные проблемы со стабильностью препарата в случае хранения при 25°C в течение более длительного времени (данные не представлены).
В исследовании иммуногенности на мышах изучали влияние 3 циклов замораживания-размораживания на CMV с двойным слиянием IE1/2-UL51, сформулированный в буфере HNS. Три цикла замораживания-размораживания (F/T) препарата CMV с двойным слиянием не оказывали влияния на иммуногенность (p=0,2103, непарный двухсторонний критерий Стьюдента) по сравнению с контролем, замороженным при -70°C (данные не представлены).
Другие варианты осуществления находятся в рамках следующей далее формулы изобретения. Хотя были представлены и описаны некоторые варианты осуществления, можно осуществлять различные модификации без отступления от сущности и объема настоящего изобретения
Реферат
Настоящее изобретение относится к биотехнологии. Предложены условно дефектный по репликации цитомегаловирус (CMV) и композиции его содержащие для применения в качестве вакцины, способ получения и применение указанного CMV при получении лекарственного средства и в медицине и применение указаной композиции для индукции иммунного ответа против CMV у пациента. Предложенный условно дефектный по репликации CMV содержит пентамерный gH-комплекс, включающий UL128, UL130, UL131, gH и gL, и нуклеиновую кислоту, кодирующую два слитых белка. Первый слитый белок состоит из IE1/2 и дестабилизирующего белка, второй слитый белок – из UL51 и дестабилизирующего белка. Дестабилизирующий белок является FK506-связывающим белком (FKBP) или его производным с аминокислотными заменами F36V и L106P. В предложенном CMV восстановлена экспрессия gH-комплекса исправлением мутации UL131 и отсутствуют IE1/2 и UL51 дикого типа. Предложенный CMV может быть использован в медицине в качестве компонента вакцин на основе аттенуированных штаммов CMV, генерирующих при этом нейтрализующие антитела и вызывающих Т-клеточные иммунные ответы. 7 н. и 12 з.п. ф-лы, 39 ил., 2 табл., 7 пр.
Комментарии