Иммуностимулирующие олигодезоксинуклеотиды - RU2615457C2
Код документа: RU2615457C2
Чертежи





















































Описание
Настоящее изобретение относится к иммуностимулирующим олигодезоксинуклеотидам, векторам и вакцинам, содержащим такие олигодезоксинуклеотиды, к их применению в качестве лекарственного средства, к их применению при предотвращении или борьбе с инфекционным заболеванием, к способам детекции таких олигодезоксинуклеотидов и к клеткам, которые можно использовать в этих способах.
В течение последних двух десятилетий в иммунологии было установлено, что иммунная система позвоночных обладает механизмами детекции микробной инфекции и инициации быстрой активации иммунной системы через рецептор-опосредованное распознавание уникальных характеристик патогенов, так называемых патоген-ассоциированных молекулярных паттернов (PAMP), взаимодействующих с когнатными распознающими патоген рецепторами хозяина (PRR) (Iwasaki A., Medzhitov R., 2001. Science 327, 291-295. Medzhitov R., 2009. Immunity 30, 766-775).
В настоящее время ясно, что определенные формы патогенной дезоксирибонуклеиновой кислоты (ДНК) относятся к числу таких PAMP. В 1995 году было опубликовано, что неметилированные мотивы CpG в бактериальной ДНК инициируют активацию B-клеток мыши (Krieg et al., 1995). В этом исследовании впервые описана связь между специфичным распознаванием бактериальной ДНК, содержащей иммунностимулирующие неметилированные CpG, и ранее известную супрессию CpG, а также широко распространенное метилирование CpG в ДНК млекопитающих. Показано, что наиболее эффективный стимулирующий B-клетки неметилированный оликодезоксинуклеотид CpG (CpG ODN) содержит элемент последовательности GACGTT.
Следующая значимая статья в данной области была опубликована лабораторией Shizuo Akira в Осаке/Япония (Hemmi et al., 2000). Посредством клонирования гена и подходом нокаута исследуемого гена у мышей было однозначно показано, что клеточный ответ у мышей на CpG-ODN является опосредованным Toll-подобным рецептором 9 (TLR9). Затем было показано, что CpG-ODN являются агонистами опосредованной TLR9 передачи сигналов преимущественно через путь NF каппа-B (Medzhitov, 2001). В последующее десятилетие был опубликован целый ряд исследований по темам фундаментальных исследований и по общим потенциальным иммунотерапевтическим применениям (например, описанные у Krieg, 2002, 2003, 2006, Klinman, 2004, Vollmer, 2005, Wilson et al., 2006, Kindrachuk et al., 2008, Dorn and Kippenberger, 2008, Vollmer and Krieg, 2009, Wilson et al., 2009). Ряд обзорных статей посвящен противоинфекционным применениям CpG-ODN (Krieg 2007), применению агонистов TLR9 в лечении злокачественной опухоли (Krieg, 2007, Weiner, 2009), активации TLR9 для лечения астмы и аллергии (Kline, 2007, Kline and Krieg, 2008, Fonseca and Kline, 2009) и применению в качестве адъювантов вакцины (Klinman et al., 2004, Klinman, 2006, Daubenberger, 2007, Wagner, 2009, Mutwiri et al., 2009, Klinman et al., 2009).
CpG ODN также описывали и обсуждали в качестве иммуностимулирующих средств и адъювантов вакцин при ветеринарном применении, в частности у крупного рогатого скота, свиней, овцы, собак, курицы и рыбы (Babiuk et al., 2003, Carrington and Secombes, 2006, Griebel et al., 2005, Mutwiri et al., 2003, Singh and O’Hagan, 2003, Werling and Jungi, 2003).
В области ветеринарного применения у кур описано применение олигодезоксинуклеотидов CpG, например, в вакцинах для защиты кур от болезни Ньюкасла (Linghua, 2007).
В настоящее время показано, что у курицы TLR21 действует как функциональный гомолог TLR9 млекопитающих при распознавании олигодезоксинуклеотидов CpG (Brownlie et al., 2009).
Конструирование конкретных CpG ODN в качестве иммуномодуляторов до сих пор являлось достаточно случайным. Это особенно относится к CpG ODN немлекопитающих. Причина этого является многофакторной; во-первых, не существует знаний о корреляции между иммуномодулирующими мотивами CpG для TLR человека и для TLR у не относящегося к человеку виду, не учитывая немлекопитающие виды. Во-вторых, не существует доступных клеточных систем с достаточно низким фоновым уровнем по отношению к уровню шума для избирательного тестирования эффектов очень низких концентраций CpG ODN. Кроме того, не существует доступных высокопроизводительных способов скрининга, и даже если они существуют, то не существует четкой корреляции между эффективностью in vivo по сравнению с in vitro CpG ODN в качестве иммуномодуляторов у немлекопитающих видов.
Таким образом, очевидно, что существует потребность в новых CpG ODN, которые обладают более высоким иммуномодулирующим действием и, таким образом, являются эффективными в низких дозах. Также существует необходимость в системах выбора избирательных и чувствительных CpG ODN для ветеринарных целей, для которых показана корреляция между in vitro и in vivo активностью CpG.
Одной из задач настоящего изобретения является предоставление таких новых CpG ODN.
В этом аспекте один из вариантов осуществления настоящего изобретения относится к иммуностимулирующему неметилированному олигодезоксинуклеотиду с общей формулой5’[N1]x[N7]r{N3[N4]pCG[N5]qN6}n[N8]s[N2]z3’, где каждый N1 независимо представляет собой C или G, каждый N2 независимо представляет собой C или G, N3 представляет собой T, C или G при условии, что исключена комбинация, где N3 и N4 оба представляют собой C, каждый N4 и N5 независимо представляют собой C или T, N6=A, T, G или C, N7=A, T, C или G, N8=A, T, C или G, x=3-10, z=0-10, n=2-100, p=1-6 или 1-25, если N4=T, q=1-6 или 1-25, если N5=T, r=0-8 или 1-25, если N7=T, и s=0-8 или 1-25, если N8=T, или фармацевтически приемлемой соли указанного олигодезоксинуклеотида.
"Иммуностимулирующий неметилированный олигодезоксинуклеотид" относится к олигодезоксинуклеотиду, который содержит неметилированную динуклеотидную последовательность цитидин-фосфат-гуанозин, которая стимулирует инициацию сигнальных каскадов, приводящих к активации факторов транскрипции, таких как NF-kB или фактор регуляции интерферона 3 (IRF3). Именно эта активация в свою очередь приводит к экспрессии воспалительных цитокинов и других событий клеточной активации. Участки связывания NF-kB и экспрессия генов под влиянием NF-kB в том числе описаны Schindler and Baichwal (1994).
Термин олигодезоксинуклеотид означает короткий полимер нуклеиновой кислоты дезоксинуклеотидов, т.е. молекулу, содержащую большое число дезоксирибоз, связанных с фосфатной группой и взаимозаменяемым органическим основанием. Такое органическое основание представляет собой замещенный пиримидин или замещенный пурин. Примерами являются цитозин и тимин, соответственно аденин и гуанин.
Олигонуклеотиды по изобретению могут содержать модификации. Примеры таких модификаций представляют собой, например, модификации в фосфодиэфирный межнуклеозидный мостик, расположенный на 3’- и/или 5’-конце нуклеозида. Такие модификации относятся, в том числе, к замене фосфодиэфира, например, фосфотиоатом или фосфодитиоатом.
Другие модификации представляют собой, например, замену фосфодиэфирного мостика дефосфомостиком. Примеры дефосфомостиков представляют собой метилгидроксиламинные, формацетальные и диметиленсульфоновые группы.
Другие модификации представляют собой модификации, которые относятся к замене природного нуклеозидного основания искусственным нуклеозидным основанием, таким как 5-фторцитозин, 7-деаза-7-замещенный гуанин, 7-деаза-8-замещенный гуанин, 2-тиоурацил, дигидроурацил, 5-бромцитозин, 6-замещенные цитозины, N4-замещенные цитозины.
Другие модификации представляют собой модификации, касающиеся замены сахарного звена, β-рибозного сахарного звена или β-D-2’-рибозного сахарного звена модифицированным сахарным звеном, таким как, например, L-2’-дезоксирибоза или 2’-L-арабиноза.
Пособием, дающим дополнительное представление о олигонуклеотидах, является, например, "PCR Primer: A Laboratory Manual", Second Edition, 2003, Edited By Carl W. Dieffenbach, National Institute of Allergy and Infectious Diseases, Gabriela S. Dreksler, Uniformed Services University of the Health Sciences, Cold Spring Harbor Laboratory Press ISBN2 978-087969654-2.
Структура {N3[N4]pCG[N5]qN6}n, несущая мотив CpG, представляет собой активную иммуностимулирующую составную группу ODN по изобретению. Таким образом, настоящее изобретение относится к иммуностимулирующим олигодезоксинуклеотидам, которые содержат эту так называемую "основную цепь".
Выявлено, что основная цепь олигодезоксинуклеотида по изобретению структуры {N3[N4]pCG[N5]qN6}n должен содержаться по меньшей мере два раза, предпочтительно три раза. Таким образом, n должно равняться по меньшей мере двум. Также выявлено, что активность олигодезоксинуклеотидов увеличивается при увеличении n. Этот эффект выравнивается при увеличении n. В основном число n структуры основной цепи, таким образом, должно оставлять по меньшей мере 2. Предпочтительно диапазон n составляет 3≤n≤100, лишь вследствие того факта, что чем длиннее синтетическая последовательность, тем более трудно ее получать. В практическом осуществлении предпочтительно диапазон n составляет 2≤n≤18. Более предпочтительно диапазон n составляет 3≤n≤18, даже более предпочтительно диапазон n составляет 4≤n≤18, еще даже более предпочтительно диапазон n составляет 5≤n≤18.
Определение CpG ODN по изобретению стало возможным, в том числе с использованием более избирательной системы детекции по сравнению с системами, используемыми в настоящее время для детекции активации NF-kB. Brownlie at al. (2009) описали репортерную систему NF-kB на основе люциферазы. Другие системы основаны, например, на измерении транскрипта IL-8 или секреции цитокинов, или детекции секреции NO.
В отличие от этого, в настоящем изобретении использовали систему детекции на основе секретируемой щелочной фосфатазы (SEAP). SEAP представляет собой репортерный фермент в системах млекопитающих (Yang et al., 1997). Оказалось, что эта система является неожиданно чувствительной и, кроме того, неожиданно обеспечивает близкую корреляцию между in vitro и in vivo активностями тестируемых CpG ODN. Систему SEAP использовали с пара-нитрофенилфосфатом (pNPP) в качестве субстрата.
Другим улучшением по сравнению с существующими системами являлось введение и стабильное поддержание в клетках плазмиды, несущей ген SEAP. До настоящего времени во все системах детекции использовали транзиентную трансфекцию клеток с репортерным геном. Вследствие введения и стабильного поддержания в клетках репортерного гена впервые в настоящее время можно построить кривую доза-эффект. Такая кривая является обязательной, если необходимо проводить достоверное сравнение между активностями различных CpG ODN.
Таким образом, способы и клеточные линии, подробно описанные в разделе примеры, по настоящему изобретению позволяют впервые проводить достоверное параллельное сравнение между различными CpG ODN.
Дополнительное подробное описание используемой системы приведено в разделе примеры.
Вследствие того, что настоящие способы и клеточные линии теперь позволяют проводить такие достоверные параллельные сравнения между различными CpG ODN, возможно было определить, что олигодезоксинуклеотид по изобретению, где N6= A, T или C, обладает наиболее высоким уровнем активности, чем когда N6=G. Таким образом, в предпочтительной форме такого варианта осуществления N6= A, T или C.
По той же причине в другой предпочтительной форме N3 представляет собой T или G и N6=Y (Y=C или T).
В более предпочтительной форме этого варианта осуществления N3, N4, N5 и N6=T.
Другая предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где N3, N4 и N5=T и N6=C.
Еще одна другая предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где N3 представляет собой G и N6=T.
Другая предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где N5=T и N6=C.
Также предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где N5=C, N6=C и q=1.
Другая предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где N4=Y и N5=Y.
Более предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где N4=T и N5=Y.
Даже более предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где N4=T и N5=T.
Другая форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где x представляет собой 4-7 и r=0, или N7 представляет собой A или T.
Предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где x представляет собой 6 и r=0, или N7 представляет собой A или T.
Другая форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где z представляет собой 0-6 и s=0, или N8 представляет собой A или T.
Предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотиду по изобретению, где z представляет собой 0-3 и s=0, или N8 представляет собой A или T.
В еще другой форме этого варианта осуществления N1 представляет собой G.
В предпочтительной форме этого варианта осуществления N2 представляет собой G.
Хотя существует широкий диапазон для обоих количеств 3’- и 5’-концевых нуклеотидов, выявлено, что существует оптимальный диапазон для обоих значений. Выявлено, что, если s=0, или N8 представляет собой A или T, то количество [N2] нуклеотидов, которое образует 3’-фланкирующую область основной цепи олигодезоксинуклеотида по изобретению, предпочтительно находится в диапазоне от 0 до 5 нуклеотидов, более предпочтительно от 0 до 3 нуклеотидов.
Также выявлено, что, если r=0, или N7 представляет собой A или T, то количество [N1] нуклеотидов, которые образуют 5’-фланкирующую область основной цепи олигодезоксинуклеотида по изобретению, является оптимальным в диапазоне от 4 до 7 нуклеотидов.
В наиболее предпочтительной форме этого варианта осуществления r=0, или N7 представляет собой A или T и s=0, или N8 представляет собой A или T и n=5-18, и x=4-7, и z=0-3.
Как указано выше, возможны различные виды модификаций в фосфодиэфирных межнуклеозидных мостиках, расположенных на 3-’ и/или 5’-конце нуклеозида. Но в основном в зависимости от способа синтеза обычные распространенные типы связей между двумя нуклеотидами представляют собой: фосфодиэфирные (PDE) связи и фосфотиоатные (PTO) связи. С целью улучшения стабильности и иммуностимулирующего действия CpG ODN элементарные звенья синтетических олигодезоксинуклеотидов предоставлены с фосфотиоатами, таким образом, что они формируют PTO связи.
Неожиданно выявлено, однако, что, когда только [N1] нуклеотиды и [N2] нуклеотиды связаны PTO связями и другие нуклеотиды связаны PDE связями, эффективность олигодезоксинуклеотида по изобретению значительно увеличивается (В таких случаях связь N1 с N7 (GT) является PTO, тогда как связь N8 с N2 (TG) является PDE).
Это особенно относится к случаю, когда [N1] и [N2] нуклеотиды представляют собой G.
Таким образом, другая предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотидам по изобретению, где N1 и/или N2 имеют фосфотиоатную связь, другие нуклеотиды имеют фосфодиэфирную связь.
Выявлено, что для олигодезоксинуклеотидов по изобретению получают даже более эффективный олигодезоксинуклеотид, когда N7=T и N8=T.
Таким образом, другая предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотидам по изобретению, где N7=T и N8=T. В этом случае r и s независимо находятся в диапазоне 1-25.
Не является необходимым, чтобы основная цепь олигодезоксинуклеотидов по изобретению, структура {N3[N4]pCG[N5]qN6}n, являлась идентичной для каждого n. Это значит, что олигодезоксинуклеотид по изобретению может выглядеть, в том числе как такой: {TTCGTT}{CTCGTG}{GTCGTA}. Такие серии трех различных последовательных различных основных цепей будут обозначены как гетерополимер. Участок трех идентичных копий будет называться гомополимером.
Предпочтительно олигодезоксинуклеотид по изобретению содержит гомополимер {N3[N4]pCG[N5]qN6}.
Олигодезоксинуклеотиды CpG по изобретению являются в большинстве случаев активными в наномолярных количествах как в тестовой системе in vitro, так и in vivo. Однако некоторые олигодезоксинуклеотиды CpG по изобретению являются активными даже в пикомолярных (субнаномолярных) количествах, их EC50 составляет ниже 1нМ.
Полумаксимальная эффективная концентрация (EC50) олигодезоксинуклеотида представляет собой количество олигодезоксинуклеотида, которое является необходимым для индукции количества репортерного фермента SEAP (который продуцирует окрашенный продукт, поглощающий при 405 нм) в репортерных клетках (HEK293-pNifty2-chicken TLR21 или HD11-pNifTy2Hyg), которое даст половину максимального поглощения. Если EC50 олигодезоксинуклеотида составляет менее 1нМ в этих клетках, считают, что он является активным в пикомолярных (субнаномолярных) количествах.
Показано, что многие из CpG ODN, которые соответствуют одной из четырех приведенных ниже общих формул, инициируют эффект in vitro в наномолярных количествах:
1)5’[G]x{TTCGTN6}n[G]z3’, где N6=A или T, n=5-100, x=3-10, z=0-10
2)5’[G]x{N3TCGTC}n[G]z3’, где N3=G или T, n=5-100, x=3-10, z=0-10
3)5’[G]x{TTCGCC}n[G]z3’, где n=5-100, x=3-10, z=0-10
4)5’[G]x{T[T]pCG[T]qT}n[G]z3’, где p=1-10, q=1-10, n=5-100, x=3-10, z=0-10
Для всех этих четырех формул по причинам эффективности n предпочтительно находится в диапазоне 5-18. X предпочтительно находится в диапазоне 4-9, 5-8, 6 или 7 в порядке предпочтения, и z предпочтительно представляет собой 8, 7, 6, 5, 4, 3, 2, 1 или 0 в порядке предпочтения. Где применимо p предпочтительно находится в диапазоне 1-5, и q предпочтительно находится в диапазоне 1-5.
Вполне возможно связывать олигодезоксинуклеотид по изобретению с носителем или гаптеном через реактивную химическую группу. Такое связывание усиливает иммуностимулирующее действие объединенных молекул.
Простые примеры таких компонентов представляют собой, например, дигоксигенин, аминогексил, техасский красный и биотин. Предпочтительные носители или гаптены представляют собой 3’- и 5’-меченный техасский красный и 5’-меченный дигоксигенин. В данной области хорошо известно связывание олигодезоксинуклеотидов с гаптенами/носителями.
Другой вариант осуществления изобретения относится к вектору, содержащему иммуностимулирующий неметилированный олигодезоксинуклеотид по изобретению. Такой вектор может представлять собой молекулу нуклеиновой кислоты, такую как плазмида, вирус, бактериофаг или любой другой используемый в молекулярной биологии вектор. Лишь в качестве примера: вектор, содержащий иммуностимулирующий неметилированный олигодезоксинуклеотид, например, может представлять собой молекулу ДНК, такую как плазмида, которую можно размножать в бактериях, в которые клонирован иммуностимулирующий неметилированнвй олигодезоксинуклеотид по изобретению. Такая плазмида предпочтительно содержит активный участок начала репликации, обуславливающий содержание большого количества плазмиды в хозяине. Выращивание таких бактерий в большом масштабе с последующим выделением плазмиды представляет собой альтернативу синтетическому способу получения иммуностимулирующего неметилированного олигодезоксинуклеотида по изобретению.
Одной из целей по настоящему изобретению является предоставление новых CpG ODN, которые можно использовать в качестве эффективных иммуностимулирующих компонентов в вакцинах, которые предотвращают или борются с инфекционным заболеванием, совместно с антигенным компонентом или генетической информацией, кодирующей антигенный компонент, и фармацевтически приемлемым носителем.
В основном термин антигенный компонент относится к композиции определенного состава, содержащей по меньшей мере один эпитоп, который может индуцировать, стимулировать или усиливать иммунный ответ при введении человеку или животному.
Антигенный компонент может представлять собой любые виды антигенного компонента, но предпочтительно выделенные из микроорганизма или вируса, который в своей форме дикого типа является патогенным для людей или животных.
Антигенный компонент может представлять собой целый патоген, предпочтительно в инактивированной или аттенуированной форме, экстракт патогена или иммуногенный белок патогена.
Если антигенный компонент представляет собой иммуногенный белок патогена, такой иммуногенный белок предпочтительно экспрессируется и его выделяют из культивируемых клеток in vitro.
Таким образом, другой вариант осуществления относится к вакцине для предотвращения или борьбы с инфекционным заболеванием, отличающейся тем, что указанная вакцина содержит иммуностимулирующее количество олигодезоксинуклеотида по изобретению и/или вектор по изобретению, иммуногенное количество антигенного компонента или генетической информации, кодирующей антигенный компонент, и фармацевтически приемлемый носитель.
Разумеется, что иммуностимулирующее количество олигодезоксинуклеотида и иммуногенное количество антигенного компонента строго взаимосвязаны. Одно из преимуществ настоящего изобретения заключается в том, что содержание олигодезоксинуклеотида по изобретению может снижать количество антигенного компонента, что является необходимым для предотвращения или борьбы с инфекционным заболеванием.
Количество антигенного компонента, которое является необходимым для предотвращения или борьбы с инфекционным заболеванием, называется иммуногенным количеством антигенного компонента.
Иммуностимулирующее количество олигодезоксинуклеотида представляет собой количество, которое способно уменьшать иммуногенное количество антигенного компонента, т.е. количество антигенного компонента, которое является необходимым для предотвращения или борьбы с инфекционным заболеванием.
Таким образом, в основном формулировки "иммуностимулирующее количество олигодезоксинуклеотида" и "иммуногенное количество" следует рассматривать относительно друг друга.
Само собой разумеется, что, если вакцина содержит генетическую информацию, кодирующую антигенный компонент, то количество антигенного компонента, экспрессируемого этой генетической информацией, должно быть достаточным для предотвращения или борьбы с инфекционным заболеванием, т.е. оно должно представлять собой иммуногенное количество.
Тот факт, что неметилированные олигодезоксинуклеотиды по изобретению являются иммуностимулирующими, означает, что они усиливают иммунологическую эффективность антигенных компонентов в вакцинах. По этой причине вакцины по изобретению во многих случаях содержат меньше антигенного компонента или генетической информации, кодирующей антигенный компонент, что является случаем, если олигодезоксинуклеотиды по изобретению не содержатся.
В некоторых случаях антигенный компонент, такой как без добавления иммуностимулирующих олигонуклеотидов, может обладать такими низкими иммунногенными свойствами, что следует вводить большие количества в любом случае, тем не менее, без получения желаемого иммуногенного уровня. В таких случаях антигенный компонент может содержаться в обычной высокой концентрации, однако теперь совместно с олигодезоксинуклеотидом по изобретению для получения, таким образом, желаемого уровня иммуногенности.
Таким образом, количество антигенного компонента или генетической информации, кодирующей антигенный компонент, которое следует вводить с олигонуклеотидом по изобретению, как правило, эмпирически равно или ниже количества, данного при отсутствии олигонуклеотида. Специалисту, принимающему участие в получении конкретных вакцин, известно такое количество для конкретной вакцины. Также в примерах приведено, например, подробное руководство о количестве антигенных компонентов, которое необходимо использовать, например, в трех различных инактивированных вирусных вакцинах: вакцине против вируса болезни Ньюкасла, вакцине против вируса инфекционного бронхита и вакцине против ринотрахеита индеек.
Количество олигодезоксинуклеотида по изобретению, которое необходимо вводить совместно с антигенным компонентом или генетической информацией, кодирующей антигенный компонент, зависит от выбранного олигодезоксинуклеотида и антигенного компонента.
Очень подходящее количество олигодезоксинуклеотида по изобретению, как правило, варьирует от 1 до 100 наномоль. Например, очень хорошие результаты получали in vivo с 1-10 мкг олигодезоксинуклеотидов по изобретению со средней длиной 30 дезоксинуклеотидов, для которых было показано, что они являются активными в тестах in vitro в наномолярном диапазоне.
Если олигодезоксинуклеотид выбран из группы олигодезоксинуклеотидов, которые являются активными в пикомолярном диапазоне, специалисту, будет понятно, что до тестирования наномолярных количеств следует тестировать количества ниже, возможно значительно ниже 1 наномоля, т.е. пикомолярные количества.
Вакцины по изобретению содержат фармацевтически приемлемый носитель. Природа такого носителя зависит, в том числе от пути введения. Если способ введения проводят через пероральный или интраназальный путь, носитель может являться таким простым как стерильная вода, раствор физиологической соли или буфер. Если предпочтительным путем введения инъекция, то носитель предпочтительно является изотоническим и имеет ограничения pH, которые делают его подходящим для инъекции. Тем не менее, некоторые такие носители широко известны в данной области.
Вакцины по изобретению в дополнение к антигенному компоненту или генетической информации, кодирующей антигенный компонент, и олигодезоксинуклеотиду по изобретению могут содержать адъювант. Адъюванты в основном представляют собой вещества, которые усиливают иммунный ответ хозяина неспецифическим образом.
Многие известные в данной области адъюванты являются подходящими, такие как полный и неполный адъювант Фрейнда, витамин E, неионные блокполимеры и полиамиды, такие как сульфат декстрана, карбопол и пиран, гидроксид алюминия. Также часто используемыми являются фосфат алюминия, сапонины, растительные масла, такие как токоферол и минеральные масла. Очень часто используемые эффективные адъюванты представляют собой эмульсии "масло-в-воде" и особенно эмульсии "вода-в-масле", далее также обозначаемые как адъюванты типа масло-в-воде и адъюванты типа вода-в-масле. Такие эмульсии хорошо известны в данной области. Таким образом, предпочтительно вакцина содержит адъювант типа вода-в-масле.
Предпочтительно антигенный компонент представляет собой, или его получают из вируса или микроорганизма, который в своей форме дикого типа является патогенным для птичьих.
Более предпочтительно указанные вирус или микроорганизм выбраны из группы, состоящей из вируса инфекционного бронхита, вируса болезни Ньюкасла, инфекционного бурсита (Гумборо), вируса анемии кур, птичьего реовируса, Mycoplasma gallisepticum, вируса ринотрахеита индеек, Haemophilus paragallinarum (ринита), поксвируса кур, вируса энцефаломиелита птиц, вируса синдром снижения яйценосности, вируса инфекционного ларинготрахеита, вируса герпеса индеек, видов Eimeria, Ornithobacterium rhinotracheale, Pasteurella multocida, Mycoplasma synoviae, видов Salmonella и Escherichia coli.
В другом варианте осуществления настоящее изобретение относится к иммуностимулирующему неметилированному олигодезоксинуклеотиду по изобретению для применения в качестве лекарственного средства.
В другом варианте осуществления настоящее изобретение относится к иммуностимулирующему неметилированному олигодезоксинуклеотиду по изобретению для применения при предотвращении или борьбе с инфекционным заболеванием у птичьих.
До настоящего времени во всех системах детекции использовали транзиентную трансфекцию клеток репортерным геном. Такая транзиентная система не обеспечивала достоверного параллельного сравнения эффективности CpG ODN. Как указано выше, основное усовершенствование в отношении существующих систем представляло собой введение и стабильное поддержание в клетках несущей репортерный ген плазмиды. Стабильное означает, что плазмида сохраняется в клетках после нескольких циклов клеточного деления.
Часто стабильное поддержание плазмиды получают выращиванием клеток под воздействием одного или более селективных средств, таких как антибиотики, ген устойчивости к которым содержится в плазмиде. Потеря плазмиды в дальнейшем вызывала бы гибель клетки, потерявшей плазмиду. Оставшиеся живые клетки все еще содержали бы плазмиду.
Таким образом, еще один другой вариант осуществления настоящего изобретения относится к клетке, содержащей рецептор TLR21 и плазмиду, кодирующую репортерный ген NF-kB, где плазмида является стабильно поддерживаемой в клетке. Такие клетки являются очень пригодными для использования в скрининге молекул CpG, более конкретно в скрининге молекул CpG по изобретению.
В примерах приведено подробное руководство о том, как получать такую клетку, содержащую плазмиду, кодирующую репортерный ген, который может стабильно поддерживаться в клетке.
Как указано выше, также показано, что системы детекции в зависимости от секретируемой щелочной фосфатазы (SEAP) очень подходят для используемой системы детекции.
Таким образом, предпочтительно репортерный ген представляет собой ген, кодирующий секретируемую щелочную фосфатазу.
В основном любая клетка или клеточная линия, несущая TLR21, которая обеспечивает введение и предпочтительно стабильное поддержание плазмиды, несущей репортерный ген NF-kB, предпочтительно ген SEAP, как описано выше, является подходящей для тестирования CpG ODN, специфичных к TLR21.
Предпочтительный пример такой подходящей клеточной линии для тестирования специфичных к TLR21 CpG ODN представляет собой клеточная линия HD11 курицы.
Таким образом, предпочтительно клеточная линия для применения в системе детекции представляет собой клеточную линию HD11, содержащую стабильную плазмиду, кодирующую репортерный ген.
Клеточные линии курицы, такие как клеточная линия HD11, предоставляют целый набор TLR курицы. Они могут при определенных условиях генерировать определенную фоновую активность.
Таким образом, клеточные линии, не относящиеся к птичьим, такие как клеточные линии млекопитающего, являются более предпочтительными клеточными линиями. Примером такой клеточной линии млекопитающего является клетка HEK293, в которую клонирован TLR21. Такая клеточная линия является более специфично избирательной для активирующих TLR21 сигналов.
Таким образом, более предпочтительно клеточная линия для применения в системе детекции представляет собой клеточную линию млекопитающего HEK293, содержащую стабильно поддерживаемый репортерный ген и в клетку HEK293 которой клонирован TLR21.
Еще один другой вариант осуществления настоящего изобретения относится к способу детекции иммуностимулирующих олигодезоксинуклеотидов по изобретению, где такой способ включает этапы a) контактирования олигодезоксинуклеотида с клеткой по изобретению, b) детекцию уровня продукции репортерного гена.
В предпочтительной форме этого способа продукт репортерного гена представляет собой SEAP.
Более предпочтительная форма этого варианта осуществления относится к способу детекции иммуностимулирующих олигодезоксинуклеотидов по изобретению, где клетка представляет собой клетку клеточной линии курицы HD11 или клеточной линии HEK293, в которую клонирован TLR21 курицы.
ПРИМЕРЫ
Пример 1
Клонирование гена и гетерологическая экспрессия TLR21 курицы
Современные достижения в исследовании TLR курицы позволяют предположить, что TLR21 представляет собой функциональный гомолог TLR9 млекопитающих у видов птиц (Keestra 2008, Brownlie et al., 2009).
План клонирования гена TLR21
На основании последовательности NM_001030558 базы данных Genbank синтезировали пару праймеров для полимеразной цепной реакции (ПЦР) амплификации гена TLR21 курицы:
Ga-TLR21-прямой 1
GAAGCTTACCATGATGGAGACAGCGGAGAAGGC
Ga-TLR21-обратный 1
GGCGGCCGCTACATCTGTTTGTCTCCTTCCCTG
Праймеры конструировали таким образом, чтобы предоставлять фланкирующие рестрикционные сайты клонирования (подчеркнутые) и последовательность Козака (выделено курсивом) для кодонов начала транскрипции и стоп-кодонов (выделено жирным). ОТ-ПЦР проводили с использованием этих праймеров и тотальной РНК из селезенки курицы в качестве матрицы. Продукт ПЦР ожидаемого размера (~ 3000 п.н.) клонировали в pCR2.1-Topo и секвенировали 5 независимых клонов плазмид (P1, P2, P12, P13, P14).
ДНК последовательность TLR21 курица, как использовали.
AAGCTTACCATGATGGAGACAGCGGAGAAGGCATGGCCCAGCACCAGGATGTGCCCCTCCCACTGCTGTCCACTCTGGCTGCTGCTGCTGGTGACAGTGACACTGATGCCGATGGTGCACCCGTATGGCTTTCGCAACTGCATTGAGGATGTCAAGGCACCTTTGTACTTCCGCTGCATCCAGCGCTTCCTGCAGTCGCCGGCCCTGGCAGTGTCTGACCTGCCACCACATGCCATCGCGCTCAATCTGTCATACAACAAAATGCGCTGCCTGCAGCCCTCTGCCTTTGCCCACCTGACACAGCTGCATACCCTGGACCTGACCTACAACCTCCTGGAGACCCTCTCCCCTGGTGCCTTCAATGGGCTGGGTGTGCTGGTGGTGCTGGACCTGTCTCACAACAAGCTGACCACACTTGCTGAAGGGGTGTTCAACAGCTTGGGCAACCTGTCCTCGCTGCAGGTACAACATAACCCCCTCAGCACGGTGTCACCAAGTGCTCTGCTACCCCTGGTCAACCTGCGCCGCCTGTCTCTACGGGGCGGGCGGCTGAATGGGTTGGGGGCAGTGGCAGTGGCAGTGCAGGGCTTGGCACAGCTGGAGCTGTTGGACCTATGTGAAAACAACCTGACAACGCTGGGGCCAGGCCCACCGCTACCCGCCTCGCTGCTCACCCTGCAGCTGTGCAACAACTCGCTGAGGGAGTTAGCGGGGGGCAGCCCGGAGATGCTATGGCACGTGAAGATACTCGACCTCTCCTACAACAGTATCTCACAGGCGGAGGTCTTCACCCAGCTCCACCTGCGCAACATCAGCCTGCTCCACCTGATCGGCAACCCCTTGGATGTCTTCCACCTGTTGGACATCTCTGACATCCAACCTCGCAGCCTGGATTTCTCTGGGTTGGTGCTGGGGGCTCAGGGGCTGGATAAGGTGTGCCTGAGGCTGCAGGGTCCCCAGGCCTTGCGGCGGCTGCAGCTACAACGCAACGGGCTGAAGGTGCTGCATTGTAATGCACTGCAGTTGTGTCCTGTGCTGAGAGAGCTGGACCTGTCCTGGAACCGGCTACAGCACGTGGGCTGTGCCGGCCGGCTGCTGGGCAAGAAGCAGCGGGAGAAGCTGGAAGTGCTGACAGTGGAACACAACCTGCTGAAGAAACTGCCGTCTTGCCTGGGGGCCCAGGTGCTGCCTCGGCTGTACAACATTTCCTTCCGCTTTAACCGCATCCTGACTGTTGGGCCCCAAGCCTTTGCCTACGCCCCGGCCCTGCAGGTGTTGTGGCTCAATATTAACAGCCTGGTGTGGCTGGACAGGCAGGCACTGTGGAGGCTGCACAACCTGACAGAGCTGCGCCTGGACAACAACCTGCTGACCGACCTCTATCACAACTCCTTCATTGACCTCCACAGACTGCGCACCCTCAACCTGCGCAACAACCGTGTCTCCGTCCTCTTCTCTGGTGTCTTCCAGGGGCTGGCTGAGCTGCAGACGCTGGATTTAGGGGGCAACAACTTGCGCCACCTGACTGCACAGTCACTGCAGGGGCTGCCCAAACTGCGCAGGCTGTACCTGGACCGCAACAGATTGCTGGAGGTGAGCAGCACTGTGTTCGCCCCAGTGCAGGCTACCCTGGGGGTGCTGGACCTGCGGGCCAACAACCTGCAGTACATCTCACAGTGGCTGCGCAAGCCGCCACCCTTCCGCAACCTGAGCAGCCTGTACGACCTGAAGCTGCAGGCGCAGCAGCCCTATGGACTGAAGATGCTGCCTCACTACTTCTTCCAGGGCTTGGTGAGGCTGCAGCAGCTGTCGCTGTCACAGAACATGCTGCGGTCCATCCCACCGGATGTCTTCGAGGACTTGGGCCAGCTGCGCTCCCTGGCATTGGCTGACAGCAGCAATGGGCTGCATGACCTGCCTGACGGCATCTTCAGAAACCTGGGCAACCTGCGGTTCCTGGACCTGGAGAATGCAGGGCTGCACTCGCTCACTCTGGAAGTCTTCGGCAATCTCAGCCGGCTGCAGGTGCTGCACTTGGCCAGAAACGAGCTGAAGACCTTCAATGACAGCGTTGCCAGCCGGCTGTCCTCCTTGCGCTACCTGGACCTGCGCAAGTGTCCGCTCAGCTGCACCTGTGACAACATGTGGCTGCAGGGCTGGCTGAACAACAGCCGTGTGCAGGTTGTCTACCCCTACAACTACACCTGTGGCTCACAGCACAATGCCTACATCCACAGCTTTGACACACACGTCTGCTTCCTGGACCTGGGGCTCTATCTCTTTGCTGGGACTGCACCGGCAGTGCTGCTGCTGCTGGTGGTGCCGGTGGTGTACCACCGCGCCTACTGGAGGCTGAAGTACCACTGGTACCTTCTGCGGTGCTGGGTCAACCAGCGGTGGCGGCGGGAGGAAAAGTGCTACCTCTATGACAGCTTTGTGTCCTACAATTCAGCTGATGAAAGTTGGGTGTTGCAGAAGCTGGTGCCTGAGCTGGAGCACGGTGCCTTCCGCCTCTGCTTGCACCACCGCGACTTCCAGCCGGGCCGCAGCATCATTGACAACATTGTGGATGCTGTCTACAACAGCCGGAAGACGGTGTGCGTGGTGAGCCGCAGCTACCTGCGCAGCGAGTGGTGCTCTCTAGAGGTGCAGTTGGCCAGCTACCGGCTGTTGGATGAGCGGCGTGACATCCTGGTACTGGTGCTGCTGGAGGACGTGGGTGATGCTGAGCTGTCTGCCTACCACCGCATGCGGCGGGTGCTGCTGCGGCGCACCTACCTGCGCTGGCCTCTTGACCCCGCAGCTCAGCCGCTCTTTTGGGCACGGCTGAAGAGGGCACTGAGGTGGGGAGAGGGAGGAGAGGAGGAGGAAGAAGAAGGTTTGGGTGGAGGGACGGGAAGGCCCAGGGAAGGAGACAAACAGATGTAGCGGCCGC
Трансфекция клеток НЕК293-pNifTy2-Zeo (клональная клеточная линия) посредством pcDNA3.1 (+)-нео-chiTLR21
Эмбриональные клетки почки человека (HEK) 293 были получены в 1970-х годах посредством вирусной трансформации (Graham et al., 1977) и в настоящее время являются доступными для научного сообщества посредством хранилищ клеточных линий, таких как ATCC.
pNifty2 представляет собой плазмиду, которая обеспечивает детекцию активации фактора транскрипции NF-kB, которая является основной для многих иммуностимулирующих активаций, в том числе активации Toll-подобного рецептора. Репортерный ген в pNifTy2, транскрипция/трансляция которого зависит от активации NF-kB, представляет собой секретируемую щелочную фосфатазу (SEAP). Более подробно описано в инструкции компании поставщика этой плазмиды: Invivogen. События трансформации/трансфекции pNifty2 отбирали в бактериях и клетках млекопитающих добавлением в среды для выращивания зеоцина.
Клетки HEK293 трансфицировали pNifTy2 общепринятыми способами (липофекция), выбирали стабильную клеточную линию, функциональные свойства оси NF-kB/SEAP устанавливали стимуляцией фактором некроза опухоли α человека (Sigma). Секретируемую SEAP в супернатанте культуры стимулируемых клеток определяли колориметрическим анализом в планшете для микротитрования с использованием хромогенного субстрата пара-нитрофенилфосфата (pNPP, 5 мМ) в щелочном буфере (50 мМ NaHCO3, pH9,6, 2 мМ MgCl2). Развитие окраски (λ=405 нм) наблюдали посредством спектрофотометра для прочтения планшетов для микротитрования. Такое считывание показаний также использовали для отбираемых клональных линий (способом предельного разведения) с высокими отношениями сигнала к шуму. Один из этих отбираемых клонов (модифицированный клон 11) в дальнейшем использовали для дополнительных исследований с TLR21 курицы.
pcDNA3.1(+)-neo представляет собой стандартный экспрессирующий вектор млекопитающего, приобретаемый от Invitrogen. Субклонирование гена TLR21 курицы в этот вектор проводили через фланкирующие сайты Hind III (кодон начала n) и Not I (стоп-кодон), которые вводили ПЦР (См. фиг.1).
Затем эту плазмиду трансфицировали (липофекцией) в клональную линию HEK293-pNifty2-zeo и отбирали рекомбинантные клетки добавлением зеоцина и G418 в среду для выращивания. Функциональные свойства получаемой поликлональной рекомбинантной клеточной линии оценивали стимуляцией культуры ODN-X4 и ODN-HEK1-PTO и детекцией SEAP. Затем определяли лучшие клональные линии способом предельного разведения с последующей стимуляцией и детекцией SEAP.
SEAP представляет собой репортерный фермент в системах млекопитающих (Yang et al., 1997). SEAP является секретируемой формой эмбриональной щелочной фосфатазы человека. Ее основные преимущества заключаются в высокой стабильности и очень высокой специфической активности, которые обеспечивают чувствительность и надежность детекции. Для детекции SEAP описано несколько субстратов, но выбирали экономически выгодный и надежный pNPP, поскольку продукт его реакции пара-нитрофенолат детектирует с высокой чувствительностью (ε405=18500 M-1см-1). В своих подготовках к тестированию авторы проводили кинетические анализы, поскольку они обеспечивают широкий динамический диапазон количественного определения.
Клетки HEK293-pNifTy2-Zeo трансфицировали pcDNA3.1(+)-Neo-chiTLR21 (линеризованной Pvu I) и отбирали поликлональную клеточную линию добавлением к среде 350 мкг/мл зеоцина и 600 мкг/мл G418. Тестирование функциональных свойств проводили стимуляцией клеток ODN-X4 (PDE) и ODN-HEK1 (PTO). Отбираемые клетки продуцировали секретируемую щелочную фосфатазу (SEAP), а родительская клеточная линия HEK293-pNifTy2-Zeo не продуцировала. Проводили клонирование отдельной клетки и анализировали отдельные клоны на их ответ на ODN-X4 (PDE) (GGGGGGTTCGTTTTCGTTTTCGTTGGGGG) и ODN-HEK1 (PTO) (TCGTCGTTTTGTCGTTTGTCGTT).
Из 46 клональных клеточных линий с двойной устойчивостью к зеоцину/G418 только 3 точно отвечали на стимулы ODN, тогда как для 3-4 дополнительных клеточных линий были показаны более слабые сигналы. Таки образом, 85% из отобранных клонов являлись нефункциональными.
Для всех дальнейших исследований использовали клонированную клеточную линию 38, которая продуцировала наиболее высокий SEAP сигнал считывания в ответ на стимуляцию ODN-X4 (PDE) и ODN-HEK1 (PTO). На фиг.2-5 приведен общий обзор активности SEAP различных клональных клеточных линий с двойной устойчивостью к зеоцину/G418.
Пример 2
Анализ влияния природы N3-N6 на активность
Тестировали следующие PDE CpG-ODN:

Кроме того, в качестве положительного контроля использовали вариант PDE ODN-2006 (CpG7909), PTO аналог которого является кандидатным лекарственным средством/вакциной в лечении опухолей человека, в то время как его аналог GpC использовали в качестве отрицательного контроля (ODN2006-контроль).
На фиг.6 показаны результаты, получаемые с клональной клеточной линией HD11-pNifTyhyg в экспериментах титрования, начиная с 2000 нМ.
Ранжирование активности на основании этого теста:
ODN-X4>ODN-X2>ODN-X1>>ODN-2006 (PDE)
Менее активный:
ODN-X3, ODN-X5, ODN-X6, ODN-X7, ODN-2006-контроль (PDE)
На фиг.7 показаны результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100 нМ.
Ранжирование активности на основании этого теста:
ODN-X4>>ODN-X2
Менее активный:
ODN-X1, ODN-X3, ODN-X5, ODN-X6, ODN-X7
Совокупно из этих тестов доказано, что PDE CpG ODN-X4 и не характерные (ODN-X1) мыши и (ODN-X2) человека являются наиболее эффективным реагентом в обоих клеточных линиях курицы HD11 и в гетерологической системе тестирования на основе TLR21 курицы.
Пример 3
Роль нуклеотидов непосредственно смежных с мотивов CpG
Для определения активности мотивов вариантов гексануклеотидной последовательности для клеток HD11 курицы и гетерологичного TLR21 курицы получали производные, где переставляли непосредственно соседние положения элемента CpG:
На основе мотива [TNCGNT]3

Следует отметить, что пермутация последовательностей приводит в одном случае обратно к мотиву ODN-X4 (→ODN-Y11).
На фиг.8 представлены результаты, получаемые с клональной клеточной линией HD11-pNifTyhyg в экспериментах титрования, начиная с 2000 нМ.
Ранжирование активности на основании этого теста в HD11-pNiftyhyg: ODN-Y11 (=ODN-X4)>ODN-Y15>ODN-Y12>ODN-Y9>ODN-Y3>ODN-Y16>ODN-Y7~ODN-Y6~ODN-Y10~ODN-Y14>ODN-Y8~ODN-Y5.
Менее активный: ODN-Y1, ODN-Y2, ODN-Y4, ODN-Y13.
На фиг.9 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100 нМ.
Ранжирование активности на основании этого теста в HEK293-pNifty2-pcDNA3.1-chiTLR21: ODN-Y11 (=ODN-X4)>>ODN-Y15>ODN-Y9>ODN-Y12>ODN-Y14~ODN-Y6>ODN-Y7~ODN-Y8~ODN-Y10~ODN-Y16>ODN-Y3~ODN-Y5.
Менее активный: ODN-Y1, ODN-Y2, ODN-Y4, ODN-Y13.
Совокупно из обеих систем тестирования можно сделать аналогичные заключения:
ODN-Y11, который является идентичным ODN-X4, is confirmed as наиболее сильный стимулятор макрофагов HD11 и клеток HEK293, которые гетерогенно экспрессируют TLR21 курицы. Вероятно, что дискриминационная способность клональной клеточной линии HEK293-pNifTy2-pcDNA3.1-chiTLR21 является выше, чем у HD11-pNiftyhyg.
Пример 4
Роль 3’-соседних положений элемента TpCpGpT в ODN-X4
Для дополнительного определения предпочтительных мотивов гексануклеотидной последовательности для клеток HD11 курицы и гетерологически экспрессируемого TLR21 курицы переставляли 3’-соседние положения элемента TpCpGpT в ODN-X4:
На основе мотива
На основе мотива (TTCGTN)3
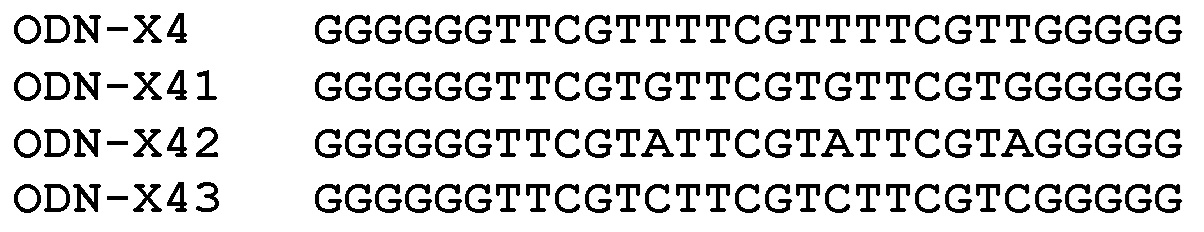
На фиг.10 представлены результаты, получаемые с клональной клеточной линией HD11-pNifTyhyg в экспериментах титрования, начиная с 2000 нМ.
Ранжирование активности на основании этого теста в HD11-pNiftyhyg: ODN-X4~ODN-X43>ODN-X42~ODN-X41.
На фиг.11 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100 нМ.
Ранжирование активности на основании этого теста в HEK293-pNifTy2-pcDNA3.1-chiTLR21: ODN-X43>>ODN-X4~ODN-X42>ODN-X41.
Пример 5
Роль 5’-соседних положений элемента TpCpGpT в ODN-X4
Для дополнительного определения дополнительных мотивов гексануклеотидной последовательности клеток HD11 курицы приставляли 5’-соседние положения элемента TpCpGpT в ODN-X4:
На основе мотива (NTCGTT)3

На фиг.12 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100 нМ.
Ранжирование активности на основании этого теста в HEK293-pNifTy2-pcDNA3.1-chiTLR21: ODN-X4>>ODN-X25>ODN-X2>ODN-X24.
Пример 6
Эффект укорочения или делеции 5’-dG6
Для дополнительной характеристики взаимосвязи структура-активность (SAR) для PDE-ODN X4 в клетках HD11 курицы и гетерологически экспрессируемого TLR21 курицы исследовали эффект укорочения или делеции 5’-dG6.

На фиг.13 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100 нМ.
Ранжирование активности на основании этого теста в HEK293-pNifTy2-pcDNA3.1-chiTLR21:
ODN-X4>ODN-X4-5’-1>ODN-X4-5’-2>ODN-X4-5’-3>>ODN-X4-5’-4>ODN-X4-5’-6≈ODN-X4-5’-5.
ODN X4-5’-4-6 являются менее активными в этом диапазоне концентраций.
Пример 7
Эффект укорочения или делеция 3’-dG5
Для дополнительной характеристики взаимосвязи структура-активность (SAR) для PDE-ODN X4 в клетках HD11 курицы и гетерологически экспрессируемого TLR21 курицы исследовали эффект укорочения или делеции 3’-dG5.

На фиг.14 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100 нМ.
Ранжирование активности на основании этого теста в HEK293-pNifTy2-pcDNA3.1-chiTLR21:
ODN-X4-5’-5≈ODN-X4-5’-4≈ODN-X4-5’-3≈ODN-X4-5’-2>ODN-X4-5’-1>ODN-X4
ODN X4-минус G, не содержащий 3’dG6 и 3’dG5, является менее активным в этом диапазоне концентраций.
Кроме того, исследовали, оказывает ли эффект добавление G в 5’-dG6 и 3’-dG5:
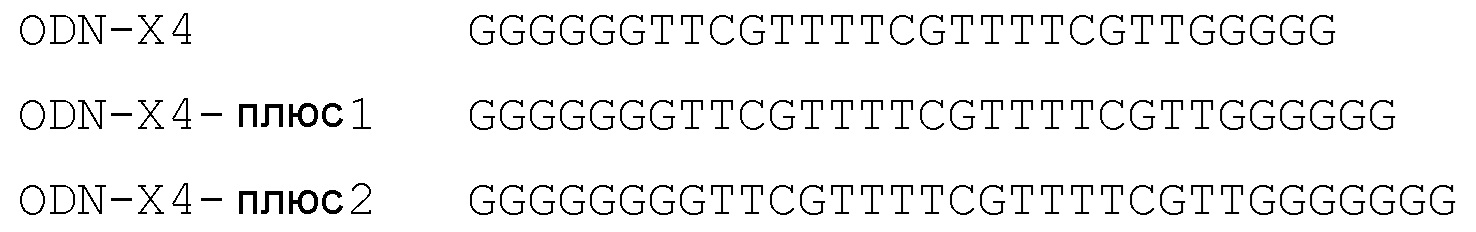
На фиг.15 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100 нМ.
Несмотря на то что добавление одного G с обеих сторон ODN-X4 не оказывали ни полезного, ни вредного воздействия на стимулирующую активность в HEK293-pNifTy2-pcDNA3.1-chiTLR21, вероятно, что добавление G приводит к молекуле более низкой активностью.
Пример 8
Замена фосфодиэфирных (PDE) связей фосфотиоатными (PTO) аналогами
Для улучшения стабильности и иммуностимулирующей способности CpG-ODN исследовании замену фосфодиэфирных (PDE) связей фосфотиоатными (PTO) аналогами. Для дополнительной характеристики этого аспекта взаимосвязи структура-активность (SAR) для PDE-ODN-X4 в макрофагах курицы HD11-pNifTyhyg и в гетерологически экспрессируемом TLR21 курицы исследовали эффект замены всех PDE связей на PTO (ODN-X4-PTO) и PDE связей на PTO только в сериях 5’-dG6 и 3’dG5 (ODN-X4-PTO-G-only).
На фиг.16 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 50 нМ.
В этой системе определения выявлена более низкая активность X4-PTO по сравнению с X4-PDE. Доказано, что X4-PTO-G-only является более высокоактивной в HEK293-pNifTy2-pcDNA3.1-chiTLR21, чем в исходном X4-PDE.
Ранжирование активности in vitro:
ODN-X4-PTO-Gтолько>ODN-X4 (PDE)>ODN-X4-PTO
Пример 9
Исследование видоспецифичности ODN-X4 (PDE)
Для исследования видоспецифичности ODN-X4 (PDE) приобретали клетки HEK293-XL-pUNO-humanTLR9, затем трансфицировали pNifTy2, определяли их ответ на описанные в литературе PTO-CpG, получали клональные функциональные клеточные линии и использовали одну из них для сравнительных исследований с HEK293-pNifTy2-pcDNA3.1-chiTLR21.
В этих сравнительных исследованиях в дополнение к ODN-X4 (PDE) использовали хорошо установленные для TLR9 человека высокоактивные PTO-ODN 2006 (=pG7909) и 2007 и их контрольные аналоги GpC.
На фиг.17 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 50 нМ.
Получали следующее ранжирование активности:
ODN-X4 (PDE)~PTO-2006>PTO-2007.
GpC контроль PTO-ODN 2006 и 2007 являлись неактивными в исследуемых в настоящем исследовании диапазонах концентраций.
На фиг.18 представлены результаты, получаемые с клональной клеточной линией HEK293XL-pUNO-huTLR9-pNifTy2 в экспериментах титрования, начиная с 50 нМ.
Получали следующий порядок ранжирования активности:
PTO-2006>PTO-2007.
GpC контроль PTO-ODN 2006 и 2007 и ODN-X4 (PDE) являлись неактивными в исследуемых диапазонах концентраций.
Этот результат доказывал видоспецифичность к курице ODN-X4 (PDE).
Пример 10
Исследования оптимального числа повторений TTCGTT
Для исследования оптимального числа повторений TTCGTT получали следующие ниже конструкции:

На фиг.19 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 20 нМ.
Для HEK293-pNifTy2-pcDNA3.1-chiTLR21 установлено следующее ниже ранжирование стимулирующей активности:
X4-hex~X4-pent>X4-quad>X4-trip (=’классическая’ X4).
X4-doub и X4-sin являлись неактивными при применяемых в настоящем исследовании тестируемых концентрациях.
Пример 11
Эффект числа разделяющих T
Для исследования эффекта числа T, разделяющих мотивы CpG, получали следующие конструкции:

На фиг.20 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 25 нМ.
Для HEK293-pNifTy2-pcDNA3.1-chiTLR21 установлено следующее ниже ранжирование стимулирующей активности:
X4-Li6~X4-Li5~X4-Li4 (=’классический’ X4)>X4-Li3>X4-Li2~X4-Li1
Пример 12
Исследования оптимального числа остатков T на границах серий dG
Для исследования оптимального числа остатков T на границах серий dG получали следующие конструкции:

На фиг.21 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 20 нМ.
Для HEK293-pNifTy2-pcDNA3.1-chiTLR21 установлено следующее ниже ранжирование стимулирующей активности:
X4-Bo4~X4-Bo3>X4-Bo2 (=’классическая’ X4)>X4-Bo1
Для дополнительной оценки оптимального числа остатков T на границах серий dG получали и (вновь) тестировали следующие (аналогичные или длиннее) конструкции:

На фиг.22 представлены результаты, получаемые с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 20 нМ.
Для HEK293-pNifTy2-pcDNA3.1-chiTLR21 установлено следующее ниже ранжирование стимулирующей активности:
X4-Bo6>X4-Bo5>X4-Bo4>X4-Bo3>X4-Bo2 (=’классическая’ X4)>X4-Bo1
Пример 13
Дополнительное исследование эффекта числа T,
граничащих с тримером основной цепи
Для исследования оптимального числа T, граничащих с тримером основной цепи получали следующие конструкции:

Вероятно, что увеличения максимальной стимуляции и "эффективной концентрации 50%" (=EC50), обусловленные добавлением дополнительных T из X4-Bo5, являются незначительными или не существуют. Однако X4-Bo10 все еще является высокоактивной. Таким образом, можно на полном основании полагать, что эффект добавления больше T нивелируется. Можно легко предполагать, что конструкции до X4-Bo20, X4-Bo25 или даже X4-Bo30 еще являются очень подходящими. См. фиг.23.
Пример 14
Дополнительное исследование эффекта числа T, "разделяющих" элементы CG
Для исследования оптимального числа T, разделяющих элементы CG, получали следующие конструкции:


Как можно видеть ранее, X4-Li1 и X4-Li2 являются неактивными в рассматриваемом диапазоне концентраций (<20 нМ). Вероятно, что пока EC50 сильно не изменяется от X4-Li3 до X4-Li7, достижение максимальной стимуляции увеличивается в этом порядке. Удивительным является скачок EC50 от X4-Li7 до X4-Li8, который также сопровождается увеличением максимальной стимуляции. X4-Li8, X4-Li9 и X4-Li10 являются примерно одинаково эффективными в отношении EC50 и максимальной стимуляции. Однако X4-Li10 еще является высокоактивной. Таким образом, можно на полном основании полагать, что эффект добавления больше T нивелируется. Можно легко предполагать, что конструкции до X4-Li20, X4-Li25 или даже X4-Li30 еще являются очень подходящими. См. фиг.24.
Пример 15
Дополнительное исследование эффекта числа повторений TTCGTT
Для исследования оптимального числа повторений TTCGTT получали следующие конструкции:


Как можно видеть ранее, X4-sing и X4-doub являются неактивными в рассматриваемом диапазоне концентраций (<20нМ). Вероятно, что достижение максимальной стимуляции значительно увеличивается от X4-trip до X4-hept, и EC50 в этом порядке также значительно снижается. В частности, заметен скачок от X4-quad до X4-pent. От X4-hept до X4-dec максимальная стимуляция увеличивается, и EC50 умеренно, но постоянно снижается. Таким образом, можно на полном основании полагать, что эффект добавления больше тримеров нивелируется. Можно легко предполагать, что конструкции до X4-X, X4-XV или даже X4-XVIII еще являются очень подходящими. Однако такие конструкции будет все более и более трудно синтезировать. См. фиг.25 и 26.
Пример 16
Дополнительное исследование эффекта типа повторяющихся тримеров
Для исследования оптимального типа повторяющихся тримеров получали следующие конструкции:

Уровни стимуляции сильно увеличивались от X4-trip до X4-I до X4-II/X4-III. Кроме того, EC50 значительно снижалась от X4 до X4-I, а затем становилась постепенно меньше до X4-III.
X4-III еще является высокоактивным. См. фиг.27.
Пример 17
Дополнительное исследование эффекта повторений TTTCGTTT
Для исследования оптимального числа остатков T на границах повторений TTCGTT получали следующие конструкции:

Аналогично как в сериях X4, X4-I-sing и X4-I-doub являются неактивными в рассматриваемом диапазоне концентраций (<20 нМ). Первым активным ODN является X4-I, и достижение максимальной стимуляции дополнительно увеличивается для X4-quad и X4-I-pent/X4-I-hex. EC50 составляет тот же порядок величины (низкая нМ) для X4-I-trip - X4-I-hex.
X4-I-hex еще является высокоактивной. См. фиг.28.
Пример 18
Дополнительное исследование мотива CG тримерного гексамера - 3’-положение границы
Для исследования оптимального мотива CG тримерного гексамера - 3’-положение границы получали следующие конструкции:

На основании этих (и более ранних) результатов ODN-X43 превосходит ODN-X4 в отношении максимальной стимуляции и значения EC50. ODN-X42 является в некоторой степени ниже в отношении максимального сигнала, но EC50 является сходным с EC50 ODN-X4.
Пример 19
Дополнительное исследование мотива CG тримерного гексамера
- определение GTCGTC
В исследовании эффективности PDE-ODN на основе ODN-X2 синтезировали следующие ODN как модификации 5’- и 3’-конца гескамера. Результаты X2, X24, X25 и X26/X4 описаны выше.

X2, X24, X25, как и X21 и X22 являются лишь слабо активными или неактивными по сравнению с X26/X4. Однако для X23 выявлена неожиданно высокая активность, превосходящая активность X26/X4.
На основании этих (и более ранних) результатах ODN-X23 превосходит ODN-X4 в отношении максимальной стимуляции и значения EC50.
Пример 20
Эффект числа мотивов ODN-X42
В основе ODN-X42 лежит тример мотива TTCGTA. Для тестирования эффекта числа мотивов исследовали число мотивов от 1 до 6:


Как видно из предшествующего эксперимента активность X4-trip и X42-trip является сравнимой. Снижение числа гексануклеотидных повторений в серии X42 приводит к потере активности (X42-sing, X42-doub), тогда как увеличение числа от 4, 5 и 6 приводит к увеличению максимального сигнала и EC50 в этом порядке с получением пикомолярной активности у X42-pent. Также стоит отметить тот факт, что начиная с X42-quad ODN превосходят ODN-X4-trip-PTO-G-only.
Можно легко предполагать, что конструкции до n=10, n=15 или даже n=18 еще являются очень подходящими. Однако такие конструкции будет все более и более трудно синтезировать. См. фиг.29.
Пример 21
Эффект числа мотивов ODN-X43
В основе ODN-X43 лежит тример мотива TTCGTC. Для тестирования эффекта числа мотивов исследовали число мотивов от 1 до 6.
Кроме того, синтезировали и тестировали варианты PTOG-only X43-trip - X43-hex.


Как видно из предшествующих экспериментов, активность X43-trip превосходит активность X4-trip. Уменьшение числа гексануклеотидных повторений в серии X43 приводит к потере активности (X43-sing, X43-doub), тогда как увеличение числа до 4, 5 и 6 приводит к увеличению максимального сигнала и EC50 в этом порядке, с получением пикомолярной активности уже у X43-quad. Также стоит отметить тот факт, что все X43-ODN, начиная с X43-trip превосходят ODN-X4-trip-PTO-G-only.
Варианты PTOG-only в сравнении с X43-trip-X43-hex являются по меньшей мере так же активными, как варианты, связанные только фосфодиэфирными связями варианты ODN.
X43-hex и X43-hex-PTOG-only являются еще высокоактивными, т.е. пределов и/или оптимума еще не достигли.
Снова можно легко предполагать, что конструкции до n=10, n=15 или даже n=18 еще являются очень подходящими. Однако такие конструкции будет все более и более трудно синтезировать. См. фиг.30 и 31.
Пример 22
Дополнительные вариации ODN-X4
С целью исследования дополнительных эффективных PDE-ODN на основе ODN-X4, синтезировали ODN с заменами динуклеотидов TT 5’- и 3’- элементов CpG на GG, AA и CC соответственно.

В тестах стимуляции HEK293-pNifty2-chiTLR21 доказано, что X4-GG, X4-AA, X4-CC, GG-X и AA-X являются неактивными в рассматриваемых диапазонах концентраций. Однако для CC-X (EC50=6,94 нМ) показана активность EC50, превосходящая в 7 раз активность X4 (EC50=52,3 нМ), а также проявляющая более высокие максимальные сигналы стимуляции. См. фиг.32.
Пример 23
Тестирование на животных мотивов CpG по изобретению
1. Введение
1.1 Задача
Для оценки того, может ли лиганд TLR (Toll-подобного рецептора), объединенный с минимальным количеством инактивированного антигена NDV клона 30, объединенного с эмульсией в/м, обеспечивать защиту против заражения вирусом болезни Ньюкасла (NDV) Herts 33/56.
2. Вещества и способы
2.1 Краткий план эксперимента
Восемнадцать групп свободных от особых патогенов (SPF) кур породы белый леггорн в возрасте 3 недель, помещенных в изоляторы, вакцинировали однократно внутримышечно (в/м) в правую грудную мышцу одним из составов, указанных в таблице 1 "Распределение по группам и дозирование". Из каждой группы из 12 животных вакцинировали только 10 кур, другие 2 птицы служили в качестве контроля. Образцы крови брали за 1 сутки до вакцинации (T=0) у 18 в случайном порядке отобранных животных (1 из каждой группы) и на T=3 неделе после вакцинации у всех животных из всех групп. После получения образцов крови на T=3 неделе после вакцинации всех кур экспериментально заражали внутримышечным (в/м) путем в мышцу правой лапы 0,2 мл (106,0 EID50) на курицу велогенным штаммом Herts 33/56 NDV. В течение периоды 14 суток после заражения кур ежедневно оценивали на появление клинических признаков инфекции NDV или смертность. Через две недели после заражения получали кровь у всех оставшихся животных, после чего животных подвергали эвтаназии. Местные реакции исследовали и оценивали макроскопически. Образцы для общепринятого гистологического исследования забирали, когда реакции и повреждения являлись видимыми.
Тестируемые вещества
2.2.1
2.2.1.1 Вакцина: 0.25% масс./масс. инактивированного NDV клон 30 в эмульсии в/м.
2.2.1.2 Лиганды: X4-PDE (Y11) - поставляемый Biolegio - Нидерланды
X4-PTO (Y11) - поставляемый TibMolBiol - Берлин - Германия
X4-PTO-G-only (Y11) - поставляемый TibMolBiol
2007-PTQ (известный из литературы) - поставляемый TibMolBiol
2.2.1.3 последовательности CpG:

PTO = фосфо(ро)тиоат (указан во множественном числе) (= устойчивый к нуклеазе); PDE = фосфодиэфир (общепринятый олигосинтез)
2.2.2 Получение вакцины
С каждым лигандом TLR каждый раз получали определенные разведения, которые добавляли к [0,25% масс./масс. NDV в эмульсии в/м]-вакцине до конечной концентрации 2.5% об./об., приводящей к дозе 1 мкг или 10 мкг на 0,5 мл (Полная доза вакцины экспериментальной вакцины, используемой в настоящем описании, содержит 8,06% масс./об. зародышевой жидкости инфицированных NDV яиц/эмульсии в/м). После добавления лиганда TLR лиганд к вакцине ее тщательно перемешивали с использованием небольшой вихревой мешалки.
("1/4 дозы инактивированного вируса болезни Ньюкасла" означает 1/4 минимального количества инактивированного NDV, для которого известно, что он обеспечивает титр антител, который способен защищать птичьих от инфекции NDV при отсутствии олигодезоксинуклеотида).
2.3 Вакцинация
Десять животных из каждой группы вакцинировали 0,5 мл вакцины в/м в правую грудную мышцу в возрасте 3 недель. Оставшихся 2 животных в каждой группе не вакцинировали, и они служили в качестве контроля.
2.4 Заражение
На 3 неделе после вакцинации всех 12 животных из всех 18 групп заражали 0,2 мл живым NDV Herts 33/56 (106,0 EID50 на курицу) через в/м путь в мышцу правой лапы.
2.5 Образцы крови
Образцы крови для серологии брали за 1 сутки до вакцинации (T=0) у 18 в случайном порядке отобранных животных (1 из каждой группы) и на T=3 неделе после первой вакцинации у всех животных. Через два недели после заражения кровь брали у всех оставшихся животных, которые выжили после заражения NDV.
2.6 Анализ HI
Уровни в сыворотке специфических к NDV антител определяли анализом подавления гемагглютинации (HI). В планшетах для микротитрования получали серийные двукратные разведения сыворотки и смешивали с равным объемом, содержащим 8 гемагглютинирующих единиц/50 мкл антигена NDV. Титры выражали в виде реципрокного самого большого разведения, которое обеспечивало полное подавление гемагглютинации эритроцитов курицы (1% (об./об.) в забуференном физиологическом растворе). Образцы считали положительными в отношении подавления гемагглютинации при разведении ≥1:2.
3. Результаты
Тиры HI NDV
Из результатов также очевидно, что титры HI NDV хорошо коррелируют с защитой. Обнаружено, что для каждого лиганда TLR, который индуцировал защиту, наиболее высокий титр HI коррелирует с наиболее высокой защитой, т.е. при 10 мкг на дозу. В противоположность этому, при наиболее высокой дозе лиганда TLR титр HI был наименьшим.
Гистология и патология
При макроскопическом исследовании участков инъекции не выявлено значительных макроскопических различий участко инъекции птиц из различных групп. Эти наблюдения свидетельствуют о том, что используемые лиганды TLR являются безопасными и что они не вызывают дополнительных побочных эффектов, например, местные реакции.
Защита/выживаемость
Из результатов очевидно, что не получали защиту только с NDV в эмульсии в/м (группа 1), тогда как в некоторых других группах от 20% до 90% птиц являлись защищенными вследствие добавления лиганда TLR к 0,25% (масс./масс.) NDV клону 30 в эмульсии в/м.
Не наблюдали защиты у невакцинированных контрольных кур (n=36).
Пример 24
Дополнительно тестирование на животных мотивов CpG по изобретению.
1. Введение
1.1 Задача
Для оценки влияния X4-Pent-PDE в комбинации с эмульсией в/м (эмульсии "вода-в-масле" на основе минерального масла) на титр антител к NDV, к IBV и к TRT у кур.
1.2 Обоснование
В этом испытании авторы исследовали, может ли добавление X4-Pent-PDE к один четвертой от полной дозы инактивированного антигена NDV, IBV или TRT, объединенного с эмульсией в/м, индуцировать титры антител, которые являются равными или выше при сравнении с полной дозой o NDV и TRT или с половиной дозы IBV.
2. Вещества и способы
2.1 Краткий план эксперимента
Группы свободных от особых патогенов (SPF) кур (n=10 на группу) породы белый леггорн в возрасте 4 недель однократно вакцинировали в/м в мышцу правой лапы одним из составов, указанных в таблице 2. Образцы крови брали до вакцинации (T=0) у 20 в случайном порядке отобранных животных и на T=4 и T=6 неделях после вакцинации у всех животных из всех групп. Для определения титров антител к NDV, к IBV и к TRT использовали сыворотку.
Тестируемые вещества
2.2.1 Тестируемые препараты
2.2.1.1 Антигены (инактивированные): NDV clone 30: полная доза вакцины содержит 8.06% масс./об. зародышевой жидкости инфицированных NDV яиц/эмульсии в/м.
IBV-249G: полная доза вакцины содержит 30 % масс./об. зародышевой жидкости инфицированных IB яиц/эмульсии в/м.
TRT: партия стандартного продукта. Полная доза вакцины содержит 100 ЕЭ/доза.
2.2.1.2 Вакцины: см. таблицу 2
2.2.1.3 Иммуностимулирующее средство:
X4-Pent-PDE: 5’-GGGGGGTTCGTTTTCGTTTTCGTTTTCGTTTTCGTTGGGGG -3’ (Eurofins MWG Operon (Germany))
2.2.2 Получение вакцины
Получали свежее предварительное разведение X4-Pent-PDE лиганда TLR и добавляли к вакцинам до конечной концентрации 2,5% об./об., приводящей к дозе 1 мкг или 10 мкг на 0,5 мл вакцины. После добавления лиганда TLR вакцину тщательно перемешивали с использованием небольшой вихревой мешалки.
2.3 Вакцинация
Животных из каждой группы вакцинировали 0,5 мл вакцины в/м в мышцу правой лапы в возрасте 4 недель.
2.4 Образцы крови
Образцы крови для серологии брали за 1 сутки до вакцинации (T=0) у 20 в случайном порядке отобранных животных и на T=4 неделе после первичной инфекции у всех животных.
2.5 Титры антител
2.5.1 Анализ HI NDV
Уровни в сыворотке специфических к NDV антител определяли анализом подавления гемагглютинации (HI). В планшетах для микротитрования получали серийные двукратные разведения сыворотки и смешивали с равным объемом, содержащим 8 гемагглютинирующих единиц/50 мкл антигена NDV. Титры выражали в виде реципрокного самого большого разведения, которое обеспечивало полное подавление гемагглютинации эритроцитов курицы (1% (об./об.) в забуференном физиологическом растворе). Образцы считали положительными в отношении подавления гемагглютинации при разведении ≥1:4 и выражали в виде 2log.
2.5.2 Анализ HI IBV
Уровни в сыворотке специфических к IB антител определяли анализом подавления гемагглютинации (HI). В планшетах для микротитрования получали серийные двукратные разведения сыворотки и смешивали с равным объемом, содержащим 8-16 гемагглютинирующих единиц/50 мкл антигена IBV-D274. Титры выражали в виде реципрокного самого большого разведения, которое обеспечивало полное подавление гемагглютинации эритроцитов курицы (1% (об./об.) в забуференном физиологическом растворе). Образцы считали положительными в отношении подавления гемагглютинации при разведении ≥1:16 и выражали в виде 2log.
2.5.3 ELISA TRT
Уровни в сыворотке специфических к TRT антител определяли стандартным ELISA. В кратком изложении 100 мкл 1:200 разведенного антигенного вещества TRT наносили в планшеты для микротитрования. Сыворотку предварительно разводили 1:100 и 1:800 и добавляли в планшеты для микротитрования. Уровни в сыворотке считали положительными при титре ≥5 и выражали в виде 2log.
2.5.4 Заключение
Из результатов таблицы 2 можно непосредственно заключить следующее:
1) 1/4 дозы вакцины против NDV при введении совместно с 10 мкг X4-Pent дают титр, который является сравнимым с титром полной дозы NDV без добавления X4-Pent.
2) 1/4 дозы объединенной вакцины против NDV/IBV при введении совместно с 10 мкг X4-Pent дает титр NDV и IBV, который является сравнимым с полной дозой объединенной вакцины против NDV/IBV без добавления X4-Pent.
3) 1/4 дозы вакцины против TRT при введении совместно с 10 мкг X4-Pent дает титр, который является сравнимым с полной дозой вакцины против TRT без добавления X4-Pent.
Результаты
ПОДПИСИ К ФИГУРАМ
Фиг.1: Карта плазмиды pcDNA3.1(+)-chiTLR21
Фиг.2-5: обзор активности SEAP различный клональных клеточных линий с двойной устойчивостью к зеоцину/G418.
Фиг.6: Эксперименты титрования, начиная с 2000нМ с клональной клеточной линией the HD11-pNifTyhyg.
Фиг.7: Результаты, полученные с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100нМ.
Фиг.8: Результаты, полученные с клональной клеточной линией HD11-pNifTyhyg в экспериментах титрования, начиная с 2000нМ.
Фиг.9: Результаты, полученные с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100нМ.
Фиг.10: Результаты, полученные с клональной клеточной линией HD11-pNifTyhyg в экспериментах титрования, начиная с 2000нМ.
Фиг.11-15: Результаты, полученные с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная со 100нМ.
Фиг.16-17: Результаты, полученные с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 50нМ.
Фиг.18: Результаты, полученные с клональной клеточной линией HEK293XL- pUNO-huTLR9-pNifTy2 в экспериментах титрования, начиная с 50нМ.
Фиг.19: Результаты, полученные с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 20нМ.
Фиг.20: Результаты, полученные с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 25нМ.
Фиг.21: Результаты, полученные с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 20нМ.
Фиг.22: Результаты, полученные с клональной клеточной линией HEK293-pNifTy2-pcDNA3.1-chiTLR21 в экспериментах титрования, начиная с 20нМ.
Фиг.23: Эффект числа T, граничащих с тримером основной цепи.
Фиг.24: Эффект числа T, разделяющих элементы CG.
Фиг.25: Эффект числа повторений TTCGTT.
Фиг.26: Эффект числа повторений TTCGTT.
Фиг.27: Эффект типа повторяющегося тримера.
Фиг.28: Эффект числа остатков T на границах повторений TTCGTT.
Фиг.29: Эффект числа мотивов ODN-X42.
Фиг.30-31: Эффект числа мотивов ODN-X43.
Фиг.32: дополнительные вариации ODN-X4.
Литературные ссылки



Реферат
Настоящее изобретение относится к биотехнологии. Предложен иммуностимулирующий неметилированный CpG-олигодезоксинуклеотид, вектор экспрессии, его содержащий, вакцина для предотвращения или борьбы с инфекционным заболеванием у птиц, содержащая указанные олигодезоксинуклеотид и/или вектор экспрессии и иммунологическое количество антигенного компонента, выделенного из патогенного для птичьих вируса или микроорганизма, а также применение олигодезоксинуклеотида в качестве лекарственного средства и для предотвращения инфекции у птичьих. Предложенный олигодезоксинуклеотид обладает высоким иммуномодулирующим действием и может быть использован в ветеринарии в качестве эффективного иммуностимулирующего компонента в вакцинах против инфекционных заболеваний птиц совместно с антигенным компонентом, выделенным из вируса или микроорганизма патогенного для птичьих. 5 н. и 24 з.п. ф-лы, 32 ил., 3 табл., 24 пр.
Комментарии