Способы и композиции для получения нуклеозидтрифосфата и рибонуклеиновой кислоты - RU2777282C2
Код документа: RU2777282C2
Чертежи













Описание
Ссылка на родственные заявки
Согласно настоящей заявке в соответствии с 35 U.S.C. 119(e) Свода законов США испрашивается приоритет находящейся на рассмотрении заявки на выдачу патента США с серийным № 62/571071, поданной 11 октября 2017 года, которая включена в настоящий документ посредством ссылки во всей своей полноте.
Предшествующий уровень техники настоящего изобретения
Рибонуклеиновая кислота (РНК) содержит повторяющиеся звенья рибонуклеотидов и участвует в ключевых клеточных процессах, в том числе в экспрессии генов и синтезе белка. Таким образом, РНК является привлекательной мишенью для модулирования фундаментальных процессов клетки, например, в РНК-вакцинах, которые индуцируют клеточный иммунный ответ. Однако экономически эффективное получение РНК в промышленном масштабе (например, от граммов до килограммов) является сложной задачей, отчасти из-за стоимости исходного материала (например, нуклеозидтрифосфатов (НТФ)) и реакционных компонентов (например, ДНК-матрицы и полимеразы). Обеспечение высококачественной РНК в коммерчески значимых масштабах требует рентабельного получения как НТФ, так и РНК.
Краткое раскрытие настоящего изобретения
Настоящий документ относится к системам, способам, композициям (например, клеткам, клеточным лизатам, реагентам и реакционным смесям) и наборам для рентабельного получения (биосинтеза) НТФ и/или РНК с использованием путей биосинтеза, разработанных для использования недорогих субстратов (например, клеточной РНК, нуклеиновых оснований, нуклеозидов, нуклеозидмонофосфатов (НМФ) и/или нуклеозиддифосфатов (НДФ)), рекомбинантных и/или эндогенных ферментов (например, киназ и/или полимераз) и источников энергии (например, НТФ, полифосфата и/или пирофосфата). Получение НТФ и/или РНК согласно некоторым вариантам осуществления достигают с использованием in vitro и/или бесклеточных лизатных систем, разработанных для минимизации (например, снижения, ингибирования и/или устранения) нежелательных ферментативных активностей, с повышением тем самым эффективности процесса и выхода желаемого конечного продукта.
В путях биосинтеза, описываемых в настоящем документе, как правило, используют полифосфаткиназу и полифосфат в качестве альтернативы ферментам эндогенного пути и источникам фосфатов.
Таким образом, некоторые аспекты настоящего раскрытия относятся к способам и композициям для получения НТФ, которые предусматривают инкубирование в реакционной смеси НДФ (например, АДФ, ЦДФ, ГДФ и/или УДФ), полифосфаткиназы (например, PPK2) и полифосфата (например, гексаметафосфата) в условиях, подходящих для получения НТФ. Как показано на фиг. 2A, PPK переносит фосфат из полифосфата в АДФ, ЦДФ, ГДФ и УДФ, что приводит к получению АТФ, ЦТФ, ГТФ и УТФ. Согласно некоторым вариантам осуществления эта реакционная смесь дополнительно содержит НДФ-киназу (например, ndk).
Другие аспекты настоящего раскрытия относятся к системам, способам, композициям и наборам для получения НТФ, которые предусматривают инкубирование в реакционной смеси НМФ (например, 5’-НМФ, такого как 5’-AMФ, 5’-ЦМФ, 5’-ГМФ и/или 5’-УМФ), полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит НМФ-киназу или НДФ-киназу (например, ndk). Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит НМФ-киназу (например, adk, cmk, gmk и/или pyrH) и НДФ-киназу (например, ndk).
Следующие аспекты настоящего раскрытия относятся к системам, способам, композициям и наборам для получения НТФ, которые предусматривают инкубирование в реакционной смеси нуклеозидов (например, аденозина, цитидина, гуанозина и/или уридина), полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу или НДФ-киназу. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и НДФ-киназу.
Следующие аспекты настоящего раскрытия относятся к системам, способам, композициям и наборам для получения НТФ, которые предусматривают инкубирование в реакционной смеси нуклеиновых оснований (например, аденина, цитозина, гуанина и/или урацила), фосфорибозилтрансферазы, фосфорибозилпирофосфата, полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу или НДФ-киназу. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и НДФ-киназу.
Согласно некоторым вариантам осуществления исходный материал (например, НМФ, НДФ и/или нуклеозиды) для биосинтеза НТФ получают из клеточной РНК. Таким образом, некоторые аспекты настоящего раскрытия относятся к системам, способам, композициям и наборам для получения НТФ, которые предусматривают (a) инкубирование в реакционной смеси клеточной РНК (например, полученной из одноклеточных или многоклеточных организмов), полинуклеотидфосфорилазы (ПНФазы) и неорганического фосфата в условиях, подходящих для получения нуклеозиддифосфатов (НДФ); (b) элиминирование ПНФазы (и необязательно элиминирование других нежелательных ферментативных активностей) и (c) инкубирование в полученной в результате реакционной смеси НДФ, полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь стадии (c) дополнительно содержит НДФ-киназу. В качестве альтернативы, способы могут предусматривать (a) инкубирование в реакционной смеси клеточной рибонуклеиновой кислоты (РНК), ПНФазы, неорганического фосфата, полифосфаткиназы и полифосфата в условиях, подходящих для получения нуклеозиддифосфатов (необязательно при этом реакционная смесь дополнительно содержит НДФ-киназу); (b) элиминирование ПНФазы и (c) инкубирование реакционной смеси в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления требуемые ферменты пути (например, полифосфаткиназа и/или НДФ-киназа) могут выдерживать условия элиминирования (например, воздействие высокой температуры или химического ингибитора), так что они сохраняют свою активность (например, по меньшей мере 50% своей активности) после воздействия условий, используемых для элиминирования (например, снижения, ингибирования и/или устранения) ПНФазы.
Другие аспекты настоящего раскрытия относятся к системам, способам, композициям и наборам для получения НТФ, которые предусматривают (a) инкубирование в первой реакционной смеси клеточной РНК и рибонуклеазы (РНКазы, например, РНКазы R или нуклеазы P1) в условиях, подходящих для получения НМФ (например, 5’-НМФ); (b) элиминирование РНКазы (и необязательно других нежелательных ферментативных активностей); и (c) инкубирование в полученной в результате реакционной смеси НМФ, полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь стадии (c) дополнительно содержит НМФ-киназу, НДФ-киназу или как НМФ-киназу, так и НДФ-киназу. В качестве альтернативы, способы могут предусматривать (a) инкубирование в реакционной смеси клеточной РНК, РНКазы, полифосфаткиназы и полифосфата в условиях, подходящих для получения НМФ (например, 5’-НМФ); (b) элиминирование РНКазы и (c) инкубирование реакционной смеси в условиях, подходящих для получения НТФ.
НТФ, получаемые в настоящем документе, используют согласно некоторым вариантам осуществления для получения РНК (например, мРНК или двухнитевой РНК). Это может быть достигнуто, например, путем добавления ДНК-матрицы и полимеразы (например, T7 РНК-полимеразы) в любую из реакционных смесей, используемых для получения НТФ, как описывается в настоящем документе. В качестве альтернативы, НТФ могут быть выделены и объединены с ДНК-матрицей и полимеразой в отдельную реакционную смесь для получения РНК. Таким образом, настоящее раскрытие также относится к способам и композициям для получения РНК.
В любом из путей биосинтеза, описываемых в настоящем документе, нуклеиновые основания, нуклеозиды, НМФ, НДФ или НТФ при использовании в качестве исходного субстрата могут быть химически синтезированы, получены ферментацией или получены другими способами.
Полифосфаткиназа, используемая в системах, реакционных смесях и способах, описываемых в настоящем документе, может быть выбрана из любых полифосфаткиназ, приведенных в таблице 2 или 12. Согласно некоторым вариантам осуществления полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
Полифосфат может быть любым полифосфатом, который служит субстратом для ферментов пути. Согласно некоторым вариантам осуществления полифосфат представляет собой гексаметафосфат.
Согласно вариантам осуществления при использовании клеточной РНК клеточная РНК включает в себя, например, рибосомальную РНК, матричную РНК и/или транспортную РНК. Клеточная РНК может быть из одноклеточного организма (например, из бактерий или дрожжей) или многоклеточного организма (например, из растений).
Ферменты путей биосинтез, применимые в соответствии с настоящим раскрытием, могут быть получены (выделены и/или очищены) из (по меньшей мере одного) клеточного лизата, полученного, например, из клеток (например, созданных методами генной инженерии клеток), которые экспрессируют ферменты пути (например, нуклеазы (такие как РНКазы и/или ПНФазы), полифосфаткиназы, НМФ-киназы, НДФ-киназу и/или полимеразы). Иллюстративные способы получения таких клеточных лизатов описываются в настоящем документе. В качестве альтернативы, реакционная смесь может содержать клеточный лизат (один клеточный лизат или смесь клеточных лизатов), полученный из клеток (например, созданных методами генной инженерии клеток), которые экспрессируют ферменты пути. То есть завершенная реакция может быть выполнена в клеточном лизате или в смеси клеточных лизатов, содержащих рекомбинантные ферменты и/или эндогенные ферменты пути, а также другие реакционные компоненты (например, полифосфат), необходимые для получения НТФ. Согласно некоторым вариантам осуществления (по меньшей мере один) очищенный фермент пути добавляют в реакционную смесь.
Для реакционных смесей, которые включают в себя клеточный(ые) лизат(ы) или ферменты, полученные из клеточного(ых) лизата(ов), может быть полезно элиминирование нежелательных нативных ферментативных активностей с использованием любого из способов элиминирования, описываемых в настоящем документе. Нежелательные нативные ферментативные активности включают в себя, например, фосфатазы, нуклеазы, протеазы, дезаминазы, оксидоредуктазы и гидролазы. Согласно некоторым вариантам осуществления нативную ферментативную активность элиминируют посредством генетической модификации, ферментативной секреции из клетки, локализации (например, периплазматического нацеливания) и/или нацеливания протеазы. Согласно другим вариантам осуществления нативную ферментативную активность элиминируют посредством температуры, pH, соли, детергента, спирта или других растворителей и/или химических ингибиторов. Согласно следующим вариантам осуществления нативную ферментативную активность элиминируют посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
Подробности некоторых вариантов осуществления настоящего изобретения изложены в прилагаемых примерах, графических материалах и подробном описании. Другие признаки, цели и преимущества настоящего изобретения станут очевидными из описания и из формулы изобретения.
Краткое описание графических материалов
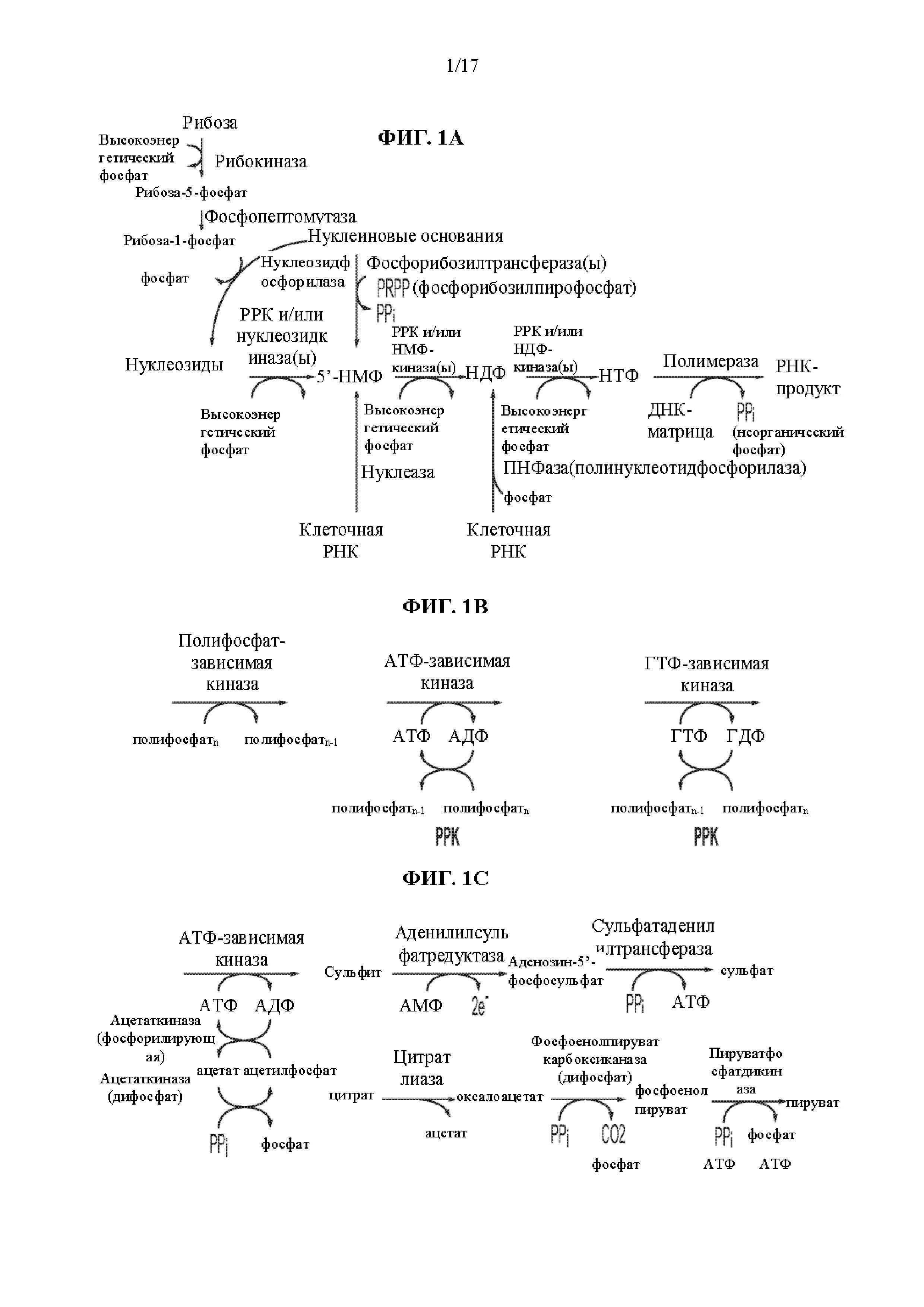
На фиг. 1A показаны пути биосинтеза для получения нуклеозидтрифосфатов (НТФ) и последующей рибонуклеиновой кислоты (РНК) с использованием нуклеотидных исходных материалов. На фиг. 1B показаны примеры стратегий высокоэнергетических фосфатов, при которых полифосфат подают в реакционную смесь. На фиг. 1С показаны примеры дополнительных стратегий высокоэнергетических фосфатов.
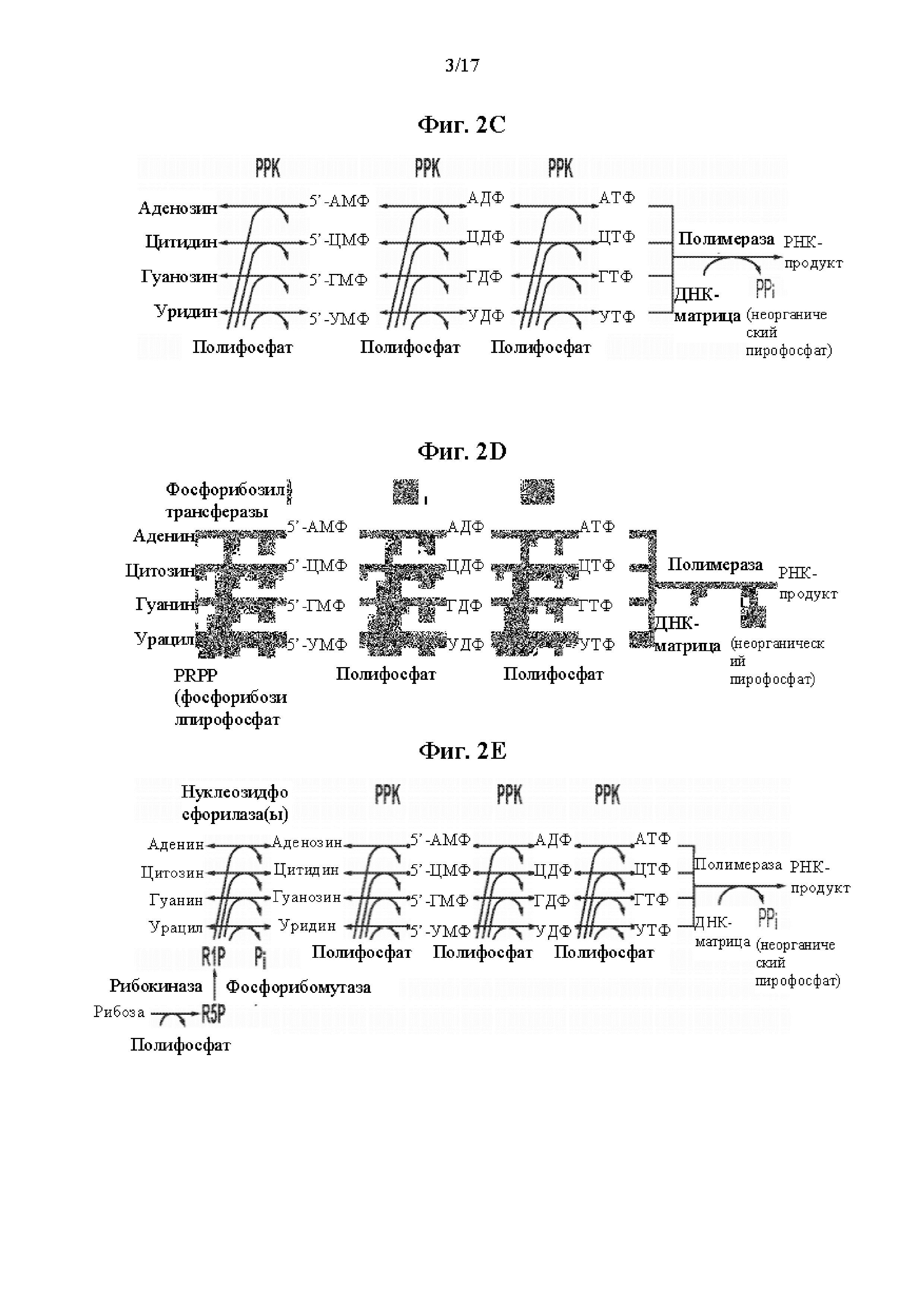
На фиг. 2А показан путь биосинтеза для получения НТФ и последующей РНК с использованием нуклеозиддифосфатов (НДФ) в качестве исходных материалов. На фиг. 2B показан путь биосинтеза для получения НТФ и последующей РНК с использованием 5’-нуклеозидмонофосфатов (5’-НМФ) в качестве исходных материалов. На фиг. 2C показан путь биосинтеза для получения НТФ и последующей РНК с использованием нуклеозидов в качестве исходных материалов. На фиг. 2D показан путь биосинтеза для получения НТФ и последующей РНК с использованием нуклеиновых оснований в качестве исходных материалов. На фиг. 2Е показан путь биосинтеза для получения НТФ и последующей РНК с использованием нуклеиновых оснований и рибозы в качестве исходных материалов.
На фиг. 3A показан путь биосинтеза для получения НТФ и последующей РНК с использованием клеточных РНК в качестве исходного материала. В этом пути полинуклеотидфосфорилазу используют для разложения клеточной РНК на НДФ. На фиг. 3B показан путь биосинтеза для получения НТФ и последующей РНК с использованием клеточных РНК в качестве исходного материала. В этом пути рибонуклеазу используют для разложения клеточной РНК на НДФ.
На фиг. 4A показан путь биосинтеза для получения НТФ и последующей РНК с использованием только полифосфаткиназ. На фиг. 4B показан путь биосинтеза для получения НТФ и последующей РНК с использованием как полифосфаткиназ (например, PPK2), так и АТФ/АДФ-зависимых киназ (например, НМФ-киназ, таких как adk, cmk, gmk и/или pyrH, и/или НДФ-киназ, таких как ndk).
На фиг. 5 показан путь биосинтеза для получения РНК исходя из 5’-НМФ.
На фиг. 6 показан путь биосинтеза для получения РНК исходя из клеточной РНК. На схеме показан пример, в котором матрицу, киназу и полимеразу добавляют в ходе реакции получения РНК.
На фиг. 7 показан путь биосинтеза для получения РНК исходя из клеточной РНК. На схеме показан пример, в котором матрица может быть добавлена в ходе фазы деполимеризации или фазы получения РНК, киназа может быть добавлена в ходе фазы деполимеризации или фазы получения РНК, и полимераза может быть добавлена в ходе фазы деполимеризации или фазы получения РНК.
На фиг. 8A показан график растворимых в кислоте нуклеотидов (мM), полученных в течение определенного времени в ходе деполимеризации РНК из лизатов E. coli с использованием надэкспрессированной РНКазы R. Растворимые в кислоте нуклеотиды измеряли с помощью поглощения УФ.
На фиг. 8B показан агарозный гель РНК-продуктов, полученных в реакциях, включающих в себя РНК-полимеразу и НМФ, полученные путем деполимеризации (- НМФ), или очищенные НМФ (+ НМФ, 4 мM каждый). Сокращения: - 2log: 2-log маркера длины ДНК (New England Biolabs), НМФ: эквимолярная смесь 5’-НМФ, РНК-пол.: термостабильная T7 РНК-полимераза, матрица 1: линейная ДНК-матрица, матрица 2: плазмидная ДНК-матрица.
На фиг. 9A показан график растворимых в кислоте нуклеотидов (мM), полученных в течение определенного времени в ходе деполимеризации очищенной РНК с использованием 1 мг/мл очищенной РНКазы R. Растворимые в кислоте нуклеотиды измеряли с помощью поглощения УФ.
На фиг. 9B показан агарозный гель РНК-продуктов, полученных в реакциях, включающих в себя РНК-полимеразу и НМФ, полученные путем деполимеризации очищенной РНК. В качестве отрицательного контроля реакцию осуществляли в отсутствие РНК-полимеразы. Сокращения: - 2log: 2-log маркера длины ДНК (New England Biolabs), НМФ: эквимолярная смесь 5’-нуклеозидмонофосфатов, РНК-пол.: термостабильная T7 РНК-полимераза, матрица 1: линейная ДНК-матрица, матрица 2: плазмидная ДНК-матрица.
На фиг. 10 показан агарозный гель РНК-продуктов, полученных с помощью бесклеточного синтеза РНК с использованием полимеразы дикого типа (W) или термостабильного мутанта полимеразы (T) при 37°C. Сокращения: - 2log: 2-log маркера длины ДНК (New England Biolabs), W: T7 РНК-полимераза дикого типа (New England Biolabs), T: термостабильная T7 РНК-полимераза, матрица 1: линейная ДНК-матрица, матрица 2: плазмидная ДНК-матрица.
На фиг. 11A показан график фактора отклика (вычисленного как отношение площади представляющей интерес dsRNA к площади коммерчески доступного внутреннего стандарта dsRNA) реакций, включающих в себя либо DgPPK2 в качестве единственной киназы, либо лизатную систему из 5 ферментов.
На фиг. 11B показаны хроматограммы HPLC dsRNA-продукта, полученного в реакциях, включающих в себя лизат DgPPK2, лизатную систему из 5 ферментов и отрицательные контроли без T7 РНК-полимеразы.
На фиг. 12A показан график растворимых в кислоте нуклеотидов (мM), полученных в течение определенного времени в ходе деполимеризации различных источников РНК с использованием очищенной РНКазы R или нуклеазы P1. Растворимые в кислоте нуклеотиды измеряли с помощью поглощения УФ.
На фиг. 12B показан график процента доступных 5’-НМФ, полученных в течение определенного времени в ходе деполимеризации РНК из E. coli или дрожжей с использованием нуклеазы P1. Процент доступных 5’-НМФ определяли с помощью LC-MS.
На фиг. 13 показаны графики нуклеомного профиля для деполимеризации РНК при разных температурах лизата из GL17-109. Показаны кумулятивные концентрации 20 аналитов. Нуклеозиды показаны в бело-крапчатом паттерне и были получены в минимальном количестве. Данные для 50°C были собраны, но не показаны.
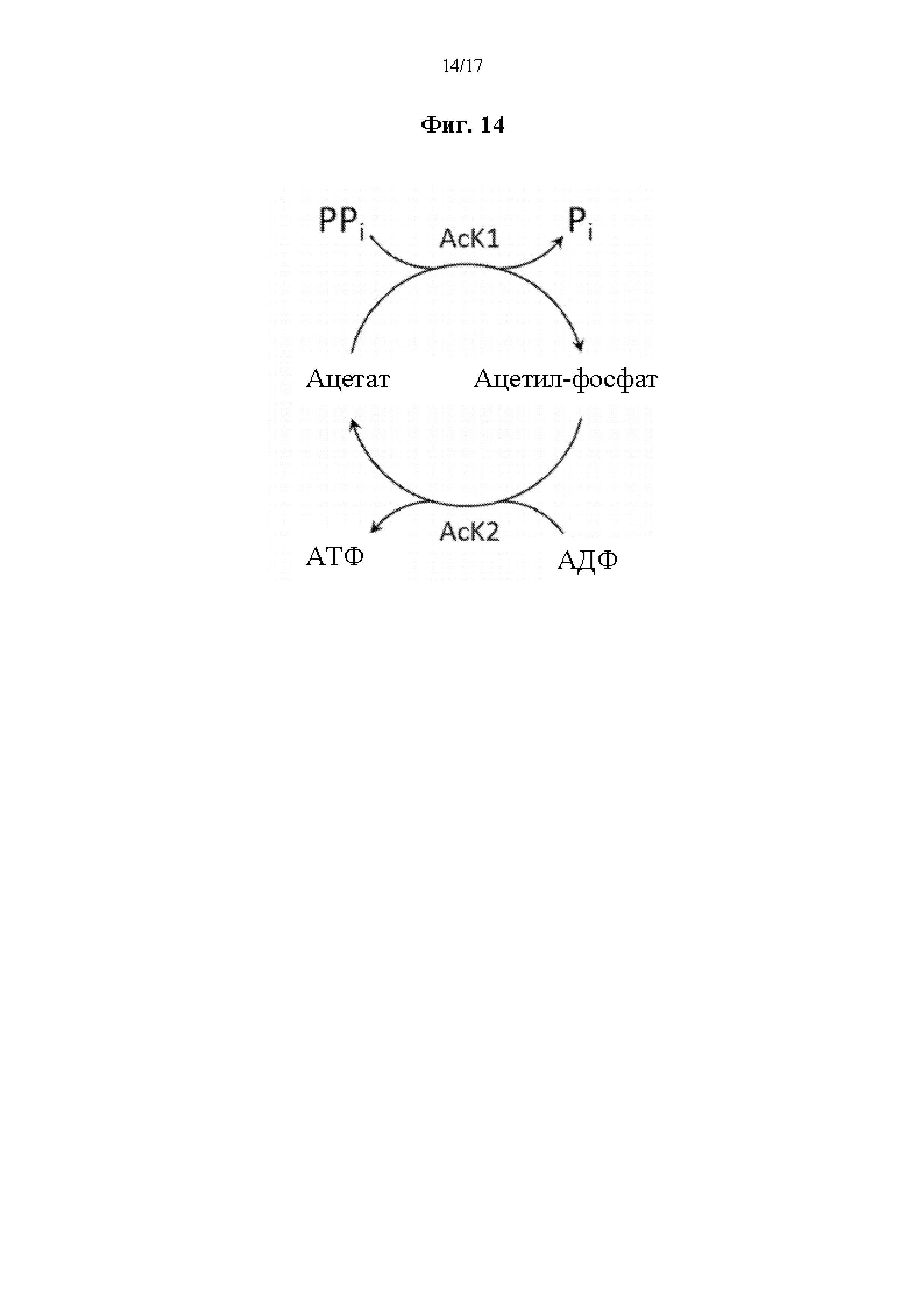
На фиг. 14 представлена схема ферментативного пути для получения АТФ из пирофосфата через циклическое фосфорилирование ацетата. Сокращения имеют следующие значения: AcK1 = первая ацетаткиназа, AcK2 = вторая ацетаткиназа, PPi = неорганический пирофосфат, Pi = неорганический фосфат, АТФ = аденозинтрифосфат, АДФ = аденозиндифосфат, и ацетил-P = ацетил-фосфат.
На фиг. 15A-15B представлена схема ферментативного пути для получения АТФ из цитрата. На фиг. 15A представлены три ферментативных реакции для получения АТФ из цитрата и пирофосфата. На фиг. 15B представлена общая химическая реакция. Сокращения имеют следующие значения: PPi = неорганический пирофосфат, PEP = фосфоенолпируват, CO2 = диоксид углерода, Pi = неорганический фосфат, АТФ = аденозинтрифосфат, и AMФ = аденозинмонофосфат.
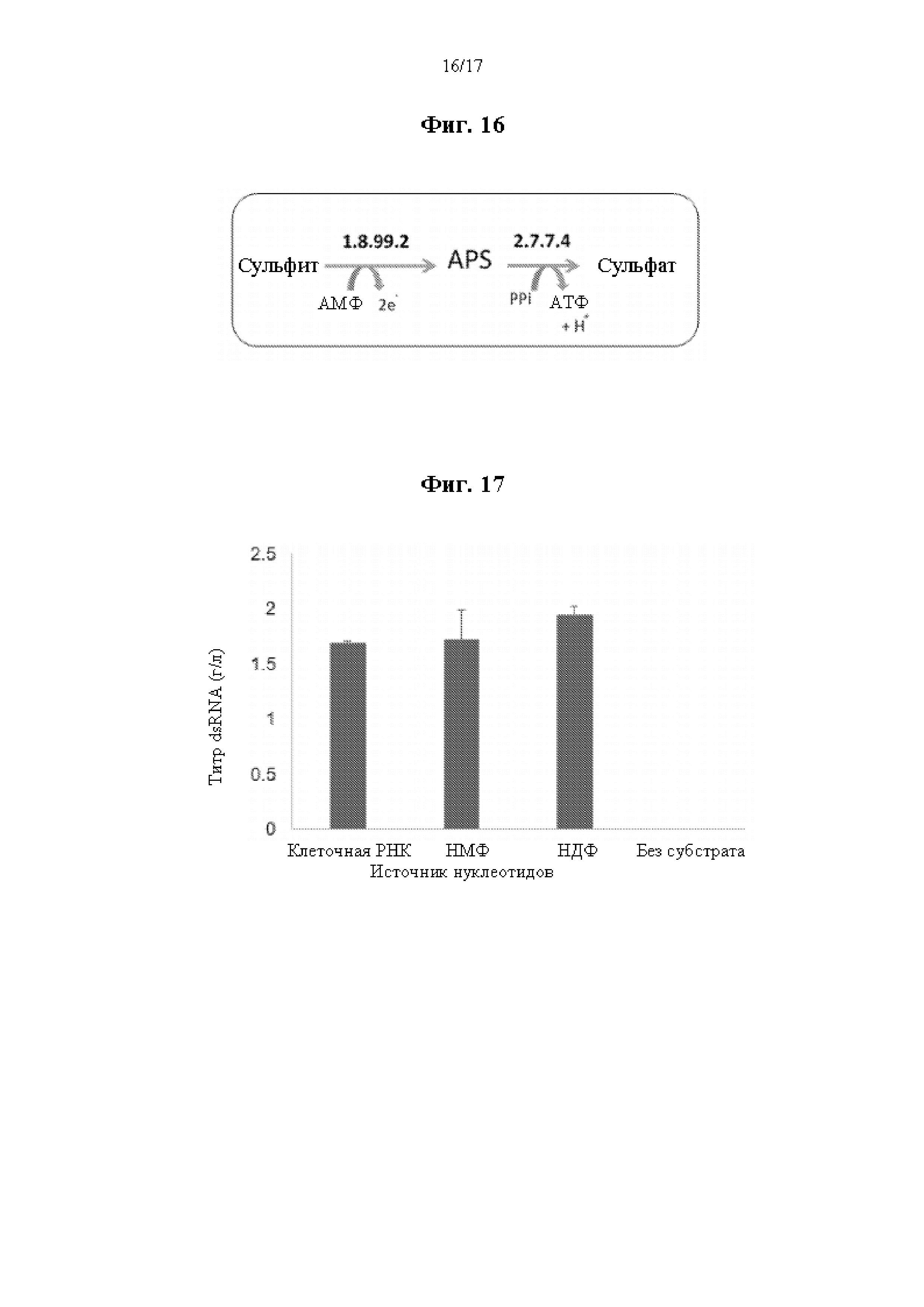
На фиг. 16 представлена схема ферментативного пути для получения АТФ из сульфита. Сокращения имеют следующие значения: АТФ = аденозинтрифосфат, AMФ = аденозинмонофосфат, APS = аденозин-5’-фосфосульфат, и PPi = неорганический пирофосфат.
На фиг. 17 показан график, демонстрирующий, что бесклеточный синтез dsRNA дает подобные титры продукта независимо от источников нуклеотидов.
На фиг. 18 показан график, демонстрирующий, что бесклеточный синтез dsRNA дает титры продукта, сравнимые с РНК-полимеразами дикого типа и термостабильными мутантными РНК-полимеразами при мезофильной температуре реакции.
На фиг. 19 показан график, демонстрирующий, что бесклеточный синтез dsRNA дает подобные титры НТФ независимо от источников нуклеотидов после 1 часа инкубации при 48°C. Для каждого источника нуклеотидов (клеточной РНК, очищенных НМФ или очищенных НДФ) в реакционную среду добавляли количество субстрата, достаточное для обеспечения приблизительно 4 мM каждого нуклеотида. Например, реакционные среды с НДФ содержали по 4 мM каждого из АДФ, ЦДФ, ГДФ и УДФ.
Подробное раскрытие настоящего изобретения
Согласно некоторым аспектам настоящее раскрытие относится к путям биосинтеза для получения НТФ и/или РНК, при которых используют экономически эффективные компоненты реакции, такие как субстраты клеточной РНК или мономерные субстраты, такие как нуклеиновые основания, нуклеозиды, НМФ или НДФ, рекомбинантные и/или очищенные ферменты пути (например, фосфорибозилтрансферазы, нуклеозидфосфорилазы, рибокиназы, фосфопентомутазы, нуклеазы, полифосфаткиназы, НМФ-киназы, НДФ-киназы, нуклеозидкиназы, РНК-полимеразы), источники высокоэнергетического фосфата (например, полифосфат) и/или ДНК-матрицы.
Компоненты реакции
Клеточная РНК. Клеточная РНК включает в себя, например, матричную РНК (мРНК), транспортную РНК (тРНК) и рибосомальную РНК (рРНК), полученные из клеточного материала (биомассы). Клеточная РНК может быть получена из любого источника клеточного материала, включающего в себя без ограничения одноклеточные организмы (например, бактерии и дрожжи) и многоклеточные организмы (например, растения и животные), либо из ферментации, либо из потока технологических отходов, например, клеточная РНК, полученная из лизата, экспрессирующего фермент (например, киназу).
Нуклеиновые основания. Нуклеиновое основание представляет собой компонент азотистого основания нуклеозида или нуклеотида. Нуклеиновые основания функционируют как фундаментальные единицы генетического кода. Нуклеиновые основания включают в себя аденин (A), цитозин (C), гуанин (G), тимин (T) и урацил (U). Нуклеиновые основания включают в себя модифицированные нуклеиновые основания, в том числе без ограничения псевдоуридин (Ψ), дигидроуридин (D) и 7-метилгуанозин (m7G).
Нуклеозиды. Нуклеозид представляет собой нуклеиновое основание, связанное с сахаром с пятью атомами углерода (например, с рибозой). Примеры нуклеозидов включают в себя аденозин, цитидин, гуанозин, тимидин и уридин.
Нуклеотиды. Нуклеотид включает в себя нуклеозид и фосфатную группу. Нуклеозид, имеющий одну фосфатную группу, представляет собой нуклеозидмонофосфат (НМФ), который включает в себя аденозинмонофосфат (AMФ), цитидинмонофосфат (ЦМФ), гуанозинмонофосфат (ГМФ), тимидинмонофосфат (TMP) и уридинмонофосфат (УМФ). Нуклеозид, имеющий две фосфатные группы, представляет собой нуклеозиддифосфат (НДФ), который включает в себя аденозиндифосфат (АДФ), цитидиндифосфат (ЦДФ), гуанозиндифосфат (ГДФ), тимидиндифосфат (ТДФ) и уридиндифосфат (УДФ). Нуклеозид, имеющий три фосфатные группы, представляет собой нуклеозидтрифосфат (НTФ), который включает в себя аденозинтрифосфат (ATФ), цитидинтрифосфат (ЦТФ), гуанозинтрифосфат (ГТФ), тимидинтрифосфат (TTФ) и уридинтрифосфат (УTФ).
Фосфорибозилтрансферазы. Фосфорибозилтрансферазы, такие как аденинфосфорибозилтрансфераза (АФРТаза), вовлекаются в путь утилизации нуклеотида в клетках, который представляет альтернативу биосинтезу нуклеотида. АФРТаза катализирует следующую реакцию в пути утилизации пуринового нуклеотида: аденин + фосфорибозилпирофосфат (PRPP) → аденозин-5’-монофосфат (AMФ) + пирофосфат (PPi).
Рибокиназы. Рибокиназа представляет собой фермент, который переносит фосфат из источника высокоэнергетического фосфата (например, АТФ или полифосфата) к D-рибозе, образующей D-рибоза-5-фосфат. Примеры включают в себя продукт гена rbsK из E. coli и продукт гена QT17_05185 из Thermus sp. 2.9.
Фосфопентомутазы. Фосфопентомутаза представляет собой фермент, который переносит фосфат в молекуле рибоза-фосфат. В частности, фосфорибомутаза катализирует обратимое взаимопревращение D-рибоза-1-фосфата и D-рибоза-5-фосфата. Примеры включают в себя продукт гена deoB из E. coli и продукт гена TM0167 из Thermotoga maritima.
Нуклеозидфосфорилазы. Нуклеозидфосфорилазы представляют собой ферменты, которые катализируют следующую обратимую реакцию: нуклеиновое основание + D-рибоза-1-фосфат ←→ нуклеозид + неорганический фосфат. Пуриннуклеозидфосфорилаза катализирует такую реакцию с пуриновыми нуклеиновыми основаниями (например, аденином, гуанином) и пуриновыми нуклеозидами (например, аденозином, гуанозином). Пиримидиннуклеозидфосфорилазы катализируют такую реакцию с пиримидиновыми нуклеиновыми основаниями (например, цитозином, урацилом) и пиримидиновыми нуклеозидами (например, цитидином, уридином). Примеры нуклеозидфосфорилаз включают в себя продукты генов deoD, xapA и УДФ из E. coli, а также TtPNPI, TtPNPII и TtPyNP, ферменты из Thermus thermophilus HB27.
Полинуклеотидфосфорилазы. Полинуклеотидфосфорилаза (ПНФаза) представляет собой бифункциональный фермент с фосфолитической активностью 3’-5’-экзорибонуклеазы и активностью 3’-терминальной олигонуклеотидполимеразы. ПНФаза способна катализировать разложение РНК на нуклеозид-5’-дифосфаты (НДФ) с использованием неорганического фосфата в качестве совместного субстрата. С помощью применения высоких концентраций неорганического фосфата при использовании ПНФазы для разложения РНК можно одновременно управлять активностью ПНФазы, снижая при этом потенциальную потерю выхода НДФ из-за активностей фосфатазы, которые могут присутствовать в реакционной смеси, поскольку известно, что неорганический фосфат ингибирует такие нежелательные активности. Согласно некоторым вариантам осуществления ПНФазу используют, необязательно в сочетании с одной или несколькими геликазами, для катализа разложения РНК на НДФ. Добавление геликазы может улучшить опосредованную ПНФазой деполимеризацию клеточной РНК за счет улучшения доступности структурированной РНК.
Нуклеазы. Нуклеазы представляют собой ферменты, которые расщепляют фосфодиэфирные связи в остове ДНК (ДНКазы) или РНК (РНКазы). Таким образом, рибонуклеазы (РНКазы) способны катализировать разложение РНК на нуклеозидмонофосфаты (НМФ). Неограничивающие примеры ферментов, которые могут быть использованы для деполимеризации РНК, как представлено в настоящем документе, представлены в таблице 1. Согласно некоторым вариантам осуществления более чем одну нуклеазу используют в реакционной смеси для деполимеризации РНК. Согласно некоторым вариантам осуществления 2, 3, 4 или 5 разных нуклеаз используют в реакционной смеси.
Таблица 1. Примеры ферментов для деполимеризации РНК
Киназы. Киназы, как правило, представляют собой ферменты, которые катализируют перенос фосфатных групп от молекулы-донора высокоэнергетического фосфата (например, АТФ, ГТФ, УТФ, ЦТФ или полифосфат, содержащий n фосфатных групп в полимере) к специфическим субстратам/молекулам. Этот процесс дает фосфорилированный субстрат и дефосфорилированную форму молекулы-донора высокоэнергетического фосфата (например, АДФ, ГДФ, УДФ, ЦДФ или полифосфат, содержащий n-1 фосфатных групп в полимере). Неограничивающие примеры киназ для применения, представленного в настоящем документе, включают в себя НМФ-киназы, НДФ-киназы, нуклеозидкиназы и полифосфаткиназы.
Полифосфаткиназы. Полифосфаткиназа представляет собой фермент, который катализирует перенос фосфатной(ых) группы(групп) от молекул-доноров высокоэнергетического фосфата, таких как полифосфат (PolyPn), к специфическим субстратам/молекулам. Этот процесс называют фосфорилированием, при котором субстрат получает фосфатную группу, а молекула-донор высокоэнергетического фосфата отдает фосфатную группу. Такая переэтерификация дает фосфорилированный субстрат и молекулу-донора фосфата, в которой отсутствует отданная фосфатная группа, такую как PolyPn-1. Полифосфаткиназы в соответствии с настоящим раскрытием согласно некоторым вариантам осуществления превращают нуклеозиды в НМФ, НМФ в НДФ и/или НДФ в НТФ. Неограничивающие примеры полифосфаткиназ представлены в таблице 2. Согласно некоторым вариантам осуществления более чем одну полифосфаткиназу используют в реакционной смеси. Согласно некоторым вариантам осуществления 2, 3, 4 или 5 разных полифосфаткиназ используют в реакционной смеси.
Таблица 2. Примеры полифосфаткиназ
Нуклеозидкиназы. Нуклеозидкиназы катализируют перенос фосфорила от молекулы-донора высокоэнергетического фосфата (например, нуклеотидтрифосфата) к R-OH акцептору, который, как правило, представляет собой 5’-гидроксильную группу сахарного фрагмента нуклеозида (например, аденозина, гуанозина, цитидина, уридина). Этот процесс превращает нуклеозид в НМФ (например, AMФ, ЦМФ, ГМФ, УМФ). Согласно некоторым вариантам осуществления нуклеозидкиназа катализирует перенос фосфата от молекулы-донора фосфата к аденозину с получением аденозинмонофосфата (AMФ). Согласно некоторым вариантам осуществления нуклеозидкиназа катализирует перенос фосфата от молекулы-донора фосфата к цитидину с получением цитидинмонофосфата (ЦМФ). Согласно некоторым вариантам осуществления нуклеозидкиназа катализирует перенос фосфата от молекулы-донора фосфата к гуанозину с получением гуанозинмонофосфата (ГМФ). Согласно некоторым вариантам осуществления нуклеозидкиназа катализирует перенос фосфата от молекулы-донора фосфата к уридину с получением уридинмонофосфата (УМФ). Неограничивающие примеры нуклеозидкиназ представлены в таблице 3. Согласно некоторым вариантам осуществления более чем одну нуклеозидкиназу используют в реакционной смеси. Согласно некоторым вариантам осуществления 2, 3, 4 или 5 разных нуклеозидкиназ используют в реакционной смеси.
Таблица 3. Примеры нуклеозидкиназ
НМФ-киназы. Нуклеозидмонофосфаткиназа (НМФ-киназа) представляет собой фермент, который катализирует перенос концевой фосфорильной группы от нуклеозидтрифосфата (НTФ), обычно АТФ, к фосфорильной группе в нуклеозидмонофосфате (например, AMФ, ЦМФ, ГМФ, УМФ). Этот процесс превращает НМФ в НДФ (например, АДФ, ЦДФ, ГДФ, УДФ). Согласно некоторым вариантам осуществления НМФ-киназа катализирует перенос фосфата от молекулы-донора фосфата к AMФ с получением аденозиндифосфата (АДФ). Согласно некоторым вариантам осуществления НМФ-киназа катализирует перенос фосфата от молекулы-донора фосфата к ЦМФ с получением цитидиндифосфата (ЦДФ). Согласно некоторым вариантам осуществления НМФ-киназа катализирует перенос фосфата от молекулы-донора фосфата к ГМФ с получением гуанозиндифосфата (ГДФ). Согласно некоторым вариантам осуществления НМФ-киназа катализирует перенос фосфата от молекулы-донора фосфата к УМФ с получением уридиндифосфата (УДФ). Неограничивающие примеры НМФ-киназ представлены в таблице 4. Согласно некоторым вариантам осуществления более чем одну НМФ-киназу используют в реакционной смеси. Согласно некоторым вариантам осуществления 2, 3, 4 или 5 разных НМФ-киназ используют в реакционной смеси.
Таблица 4A. Примеры AMФ-киназных ферментов
Таблица 4B. Примеры ЦМФ-киназных ферментов
Таблица 4C. Примеры УМФ-киназных ферментов
Таблица 4D. Примеры ГМФ-киназных ферментов
НДФ-киназы. Нуклеозиддифосфаткиназа (НДФ-киназа) представляет собой фермент, который катализирует обмен концевого фосфата между разными НДФ (например, АДФ, ЦДФ, ГДФ, УДФ) и нуклеозидтрифосфатами (НТФ) обратимым образом с получением НТФ (например, АТФ, ЦТФ, ГТФ, УТФ). Согласно некоторым вариантам осуществления НДФ-киназа катализирует перенос фосфата от молекулы-донора фосфата к АДФ с получением аденозинтрифосфата (ATФ). Согласно некоторым вариантам осуществления НДФ-киназа катализирует перенос фосфата от молекулы-донора фосфата к ЦДФ с получением цитидинтрифосфата (ЦТФ). Согласно некоторым вариантам осуществления НДФ-киназа катализирует перенос фосфата от молекулы-донора фосфата к ГДФ с получением гуанозинтрифосфата (ГТФ). Согласно некоторым вариантам осуществления НДФ-киназа катализирует перенос фосфата от молекулы-донора фосфата к УДФ с получением уридинтрифосфата (УTФ). Неограничивающие примеры НДФ-киназ представлены в таблице 5. Согласно некоторым вариантам осуществления более чем одну НДФ-киназу используют в реакционной смеси. Согласно некоторым вариантам осуществления 2, 3, 4 или 5 разных НДФ-киназ используют в реакционной смеси.
Таблица 5. Примеры НДФ-киназ
Неограничивающие примеры киназ, которые превращают НДФ в НТФ, включают в себя нуклеозиддифосфаткиназу, полифосфаткиназу и пируваткиназу. Как обсуждается в настоящем документе, термостабильные варианты упомянутых выше ферментов охватываются настоящим раскрытием. Согласно некоторым вариантам осуществления НДФ-киназа(ы) получают из Aquifex aeolicus.
Фосфорилирование НМФ в НТФ происходит согласно некоторым вариантам осуществления через путь полифосфат-зависимой киназы, при котором высокоэнергетический фосфат переносится от полифосфата к АДФ посредством полифосфаткиназы (PPK). Согласно некоторым вариантам осуществления полифосфаткиназа принадлежит семейству полифосфаткиназы 1 (PPK1), которое переносит высокоэнергетический фосфат от полифосфата к АДФ с образованием АТФ. Этот АТФ затем используется НМФ-киназой (например, АМФ-киназой, УМФ-киназой, ГМФ-киназой и ЦМФ-киназой) для превращения НМФ в когнатные им рибонуклеотиддифосфаты (НДФ). Кроме того, АТФ затем используется нуклеотиддифосфаткиназой для превращения НДФ в НТФ.
Согласно некоторым вариантам осуществления полифосфаткиназа принадлежит семейству полифосфаткиназы 2 (PPK2). Согласно некоторым вариантам осуществления полифосфаткиназа принадлежит семейству PPK2 класса I, которое переносит высокоэнергетический фосфат от полифосфата к НДФ с образованием НТФ. АТФ, продуцированный системой, используют как донор высокоэнергетического фосфата для превращения НМФ в НДФ. Согласно некоторым вариантам осуществления полифосфаткиназа принадлежит семейству PPK2 класса III, которое переносит высокоэнергетический фосфат от полифосфата к НМФ и НДФ с образованием НТФ. Согласно некоторым вариантам осуществления PPK2 класса III используют отдельно для получения НТФ из НМФ. Согласно другим вариантам осуществления PPK2 класса III используют в комбинации с другими киназами. PPK2 класса III продуцирует АТФ из АДФ, AMФ и полифосфата, который затем используется НМФ- и НДФ-киназами для превращения НМФ в НТФ.
Неограничивающие примеры PPK2 ферментов для применения в соответствии с настоящим документом перечислены в таблице 2. Таким образом, согласно некоторым вариантам осуществления PPK2 ферменты являются термостабильными. Например, ферменты PPK2 могут представлять собой термостабильные ферменты PPK2 класса III, которые способствуют синтезу АТФ, а не полифосфатной полимеризации, и превращают как АДФ, так и AMФ в АТФ. Согласно некоторым вариантам осуществления ферменты PPK2 используют для превращения полифосфата, такого как гексаметафосфат, в АТФ, при скоростях, варьирующих, например, от 10 до 800 мM в час (например, 10, 15, 20, 25, 50, 75, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750 или 800 мM в час).
Полифосфат и другие высокоэнергетические фосфаты. Молекулы высокоэнергетического фосфата (молекулы-доноры фосфата) высвобождают энергию при гидролизе высокоэнергетической связи с обеспечением тем самым источника энергии для биохимических реакций. Полифосфат (PolyPn) и другие молекулы высокоэнергетического фосфата могут быть использованы в качестве источников фосфата для получения НТФ и последующего получения РНК, как описывается в настоящем документе. PolyPn, например, содержит повторяющиеся звенья фосфата (PO4), связанные вместе общими атомами кислорода. Фосфорилирование специфических субстратов/молекул киназами в соответствии с настоящим раскрытием вовлекает донирование фосфатной группы от PolyPn с получением тем самым PolyPn-1.
Настоящее раскрытие не ограничивается числом фосфатных групп в полифосфате. Согласно некоторым вариантам осуществления PolyPn содержит по меньшей мере 3 фосфатных группы (PolyP3). Согласно некоторым вариантам осуществления PolyPn содержит по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9 или по меньшей мере 10 фосфатных групп. Согласно некоторым вариантам осуществления PolyPn представляет собой гексаметафосфат.
Другие примеры молекул высокоэнергетического фосфата включают в себя без ограничения НТФ (например, АТФ), НДФ (например, АДФ), НМФ (например, AMФ), фосфоенолпируват, 1,3-бисфосфоглицерат, фосфокреатин, фосфоенолпируват, глюкоза-1-фосфат, фруктоза-6-фосфат и глюкоза-6-фосфат. Согласно некоторым вариантам осуществления более чем один высокоэнергетический фосфат используют в реакционной смеси. Согласно некоторым вариантам осуществления 2, 3, 4 или 5 разных высокоэнергетических фосфата используют в реакционной смеси.
Матрицы. ДНК-матрица включает в себя промотор, необязательно индуцируемый промотор, функционально связанный с нуклеотидной последовательностью, кодирующей желаемый РНК-продукт, и необязательно транскрипционный терминатор. ДНК-матрицу, как правило, обеспечивают в векторе, таком как плазмида, хотя могут быть использованы другие форматы матрицы (например, линейные ДНК-матрицы, созданные с помощью полимеразной цепной реакции (ПЦР), химического синтеза или других средств, известных в уровне техники). Согласно некоторым вариантам осуществления более чем одну ДНК-матрицу используют в реакционной смеси. Согласно некоторым вариантам осуществления 2, 3, 4 или 5 разных ДНК-матриц используют в реакционной смеси.
Промотор или терминатор могут представлять собой встречающуюся в природе последовательность или созданную методами генной инженерии последовательность. Согласно некоторым вариантам осуществления созданная методами генной инженерии последовательность является модифицированной для усиления транскрипционной активности. Согласно некоторым вариантам осуществления промотор представляет собой встречающуюся в природе последовательность. Согласно другим вариантам осуществления промотор представляет собой созданную методами генной инженерии последовательность. Согласно некоторым вариантам осуществления терминатор представляет собой встречающуюся в природе последовательность. Согласно другим вариантам осуществления терминатор представляет собой созданную методами генной инженерии последовательность.
Полимеразы. Полимеразы являются ферментами, которые синтезируют полимеры нуклеиновых кислот. Полимеразы в соответствии с настоящим раскрытием включают в себя ДНК-зависимые РНК-полимеразы и РНК-зависимые РНК-полимеразы. Неограничивающие примеры полимераз представлены в таблице 6. Согласно некоторым вариантам осуществления полимераза представляет собой РНК-полимеразу, такую как T7 РНК-полимераза. Согласно некоторым вариантам осуществления более чем одну полимеразу используют в реакционной смеси. Согласно некоторым вариантам осуществления 2, 3, 4 или 5 разных полимер используют в реакционной смеси.
Таблица 6. Примеры РНК-полимераз
РНК-продукты. РНК, полученная способами, представленными в настоящем документе, может иметь любую форму РНК, в том числе однонитевой РНК (ssRNA) и двухнитевой РНК (dsRNA). Неограничивающие примеры однонитевой РНК включают в себя матричную РНК (мРНК), микроРНК (miRNA), малую интерферирующую РНК (siRNA) и антисмысловую РНК. Двухнитевая РНК в настоящем документе включает в себя полностью двухнитевые молекулы, которые не содержат однонитевую область (например, петлю или «липкий» конец), а также частично двухнитевые молекулы, которые содержат двухнитевую область и однонитевую область (например, петлю или «липкий» конец). Таким образом, короткая шпилечная РНК (shRNA) может быть получена способами в соответствии с настоящим раскрытием.
РНК, полученная способами, представленный в настоящем документе, может быть модифицирована, как описывается в настоящем документе. Согласно некоторым вариантам осуществления РНК получают согласно способу, описываемому в настоящем документе, а затем модифицируют. Согласно некоторым вариантам осуществления РНК получают согласно способу, описываемому в настоящем документе, с использованием модифицированного исходного материала. Согласно некоторым вариантам осуществления модифицированный исходный материал представляет собой модифицированное нуклеиновое основание. Согласно некоторым вариантам осуществления модифицированный исходный материал представляет собой модифицированный нуклеозид. Согласно некоторым вариантам осуществления модифицированный исходный материал представляет собой модифицированный нуклеотид.
Согласно некоторым вариантам осуществления модифицированная РНК содержит модификацию остова. В некоторых случаях модификация остова обеспечивает более продолжительное время полужизни для РНК благодаря сниженному опосредованному нуклеазой разложению. Это, в свою очередь, приводит к более продолжительному времени полужизни. Примеры подходящих модификаций остова включают в себя без ограничения фосфоротиоатные модификации, фосфородитиоатные модификации, п-этокси-модификации, метилфосфонатные модификации, метилфосфоротиоатные модификации, алкил- и арил-фосфаты (в которых заряженный кислород фосфоната замещается алкильной или арильной группой), сложные алкилфосфотриэфиры (в которых заряженный кислородный фрагмент алкилируется), модификации остова пептидной нуклеиновой кислоты (PNA), модификации остова запертой нуклеиновой кислоты (LNA) и т.п. Эти модификации могут быть использованы в комбинации друг с другом и/или в комбинации со связями фосфодиэфирного остова.
В качестве альтернативы или дополнения, РНК может содержать другие модификации, включающие в себя модификации в основании или сахарных фрагментах. Примеры включают в себя РНК, имеющую сахара, которые ковалентно присоединяются к низкомолекулярным органическим группам, отличным от гидроксильной группы в 3’-положении и отличным от фосфатной группы в 5’-положении (например, 2’-O-алкилированная рибоза), РНК, имеющую сахара, такие как арабиноза вместо рибозы. РНК также охватывают замещенные пурины и пиримидины, такие как модифицированные C-5 пропином основания (Wagner et al., Nature Biotechnology 14:840-844, 1996). Другие пурины и пиримидины включают в себя без ограничения 5-метилцитозин, 2-аминопурин, 2-амино-6-хлорпурин, 2,6-диаминопурин, гипоксантин. Другие такие модификации хорошо известны специалистам в данной области.
Пути получения НTФ
Настоящий документ относится к системам, способам, композициям и наборам для получения НТФ посредством ряда разных ферментативных путей, в каждом из которых используются источники энергии, описываемые в настоящем документе, и недорогие исходные материалы в реакционной смеси. Такие ферментативные пути могут быть расширены согласно некоторым вариантам осуществления для получения РНК (например, мРНК или двухнитевой РНК) путем добавления ДНК-матрицы и полимеразы в реакционную смесь (см., например, фиг. 1).
Следует учитывать, что любой из описываемых в настоящем документе ферментов пути (например, нуклеаз, киназ и/или полимераз) может быть получен из немодифицированных (нативных) или созданных методами генной инженерии клеток. Согласно некоторым вариантам осуществления фермент(ы) пути секретируются клетками (например, клетками, созданными методами генной инженерии для секреции фермента(ов)). Согласно другим вариантам осуществления ферменты пути получают из клеточного(ых) лизата(ов) клеток. Согласно некоторым вариантам осуществления ферменты пути являются компонентами клеточного(ых) лизата(ов) клеток, в этом случае клеточный(ые) лизат(ы) добавляется(ются) в реакционную смесь или служит(служат) реакционной смесью в биосинтетической реакции. В случаях, если клеточный(ые) лизат(ы) используется(ются) в реакционной смеси или служит(служат) реакционной смесью, клеточный лизат может быть подвергнут условиям для элиминирования нежелательных ферментативных активностей, как описано ниже, перед получением представляющего интерес продукта (НTФ и/или РНК).
Превращение НДФ в НТФ.Согласно некоторым аспектам НТФ получают с использованием НДФ в качестве субстратов, как показано на фиг. 2A. Например, способы получения НТФ могут предусматривать инкубирование в реакционной смеси НДФ, (например, 1, 2, 3 или 4) полифосфаткиназ и (например, 1, 2, 3 или 4) полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ включает в себя НДФ-киназу (см., например, таблицу 5). Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ также может включать в себя нуклеозидкиназу.
Превращение НМФ в НТФ.Согласно некоторым аспектам НТФ получают с использованием 5’-НМФ в качестве субстратов, как показано на фиг. 2B. Например, способы получения НТФ могут предусматривать инкубирование в реакционной смеси 5’-НМФ, (например, 1, 2, 3 или 4) полифосфаткиназы и (например, 1, 2, 3 или 4) полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ включает в себя НМФ-киназу (см., например, таблицу 4) и/или НДФ-киназу (см., например, таблицу 5). Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ также может включать в себя нуклеозидкиназу.
Превращение нуклеозидов в НТФ.Согласно некоторым аспектам НТФ получают с использованием нуклеозидов в качестве субстратов, как показано на фиг. 2C. Например, способы получения НТФ могут предусматривать инкубирование в реакционной смеси нуклеозидов, (например, 1, 2, 3 или 4) полифосфаткиназы и (например, 1, 2, 3 или 4) полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ также может включать в себя нуклеозидкиназу (см., например, таблицу 3), и/или НМФ-киназу (см., например, таблицу 4), и/или НДФ-киназу (см., например, таблицу 5).
Превращение нуклеиновых оснований в НТФ.Согласно некоторым аспектам НТФ получают с использованием нуклеиновых оснований в качестве субстратов, как показано на фиг. 2D. Например, способы получения НТФ могут предусматривать инкубирование в реакционной смеси нуклеиновых оснований, (например, 1, 2, 3 или 4) фосфорибозилтрансферазы, фосфорибозилпирофосфата, (например, 1, 2, 3 или 4) полифосфаткиназы и (например, 1, 2, 3 или 4) полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ также может включать в себя НМФ-киназу (см., например, таблицу 4) и/или НДФ-киназу (см., например, таблицу 5). Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ также может включать в себя нуклеозидкиназу.
Согласно некоторым вариантам осуществления в пути биосинтеза для получения НТФ и/или РНК может использоваться клеточная РНК в качестве субстрата путем либо сначала деполимеризации клеточной РНК в НДФ, либо сначала деполимеризации клеточной РНК в НМФ.
Превращение нуклеиновых оснований и рибозы в НТФ.Согласно некоторым аспектам НТФ получают с использованием нуклеиновых оснований в качестве субстратов, как показано на фиг. 2E. Например, способы получения НТФ могут предусматривать инкубирование в реакционной смеси нуклеиновых оснований, D-рибозы, рибокиназы, фосфопентомутазы, по меньшей мере одной (например, 1, 2, 3 или 4) нуклеозидфосфорилазы, по меньшей мере одной (например, 1, 2, 3 или 4) полифосфаткиназы и по меньшей мере одного (например, 1, 2, 3 или 4) полифосфата в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ также может включать в себя по меньшей мере одну НМФ-киназу (см., например, таблицу 3) и/или по меньшей мере одну НДФ-киназу (см., например, таблицу 4) и/или нуклеозидкиназу.
Превращение клеточной РНК в НТФ через НДФ. Согласно некоторым аспектам НТФ получают с использованием клеточной РНК в качестве субстрата сначала путем разрыва (разложения/деполимеризации) клеточной РНК на НДФ, а затем превращения НДФ в НТФ, как показано на фиг. 3A. Например, способы получения НТФ могут предусматривать инкубирование в реакционной смеси клеточной РНК, полинуклеотидфосфорилазы (ПНФазы) и фосфата в условиях, подходящих для получения НДФ. Для осуществления получения НТФ реакционная смесь согласно некоторым вариантам осуществления также содержит полифосфаткиназу и полифосфат. Таким образом, способы, кроме того, предусматривают инкубирование реакционной смеси в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит НДФ-киназу. Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ также может включать в себя нуклеозидкиназу.
Превращение клеточной РНК в НТФ через НМФ. Согласно некоторым аспектам НТФ получают с использованием клеточной РНК в качестве субстрата сначала путем разрыва клеточной РНК на 5’-НМФ, а затем превращения НМФ в НДФ и НДФ в НТФ, как показано на фиг. 3B. Например, способы получения НТФ могут предусматривать инкубирование в реакционной смеси клеточной РНК и рибонуклеазы в условиях, подходящих для получения 5’-НМФ. Для осуществления получения НТФ реакционная смесь согласно некоторым вариантам осуществления также содержит полифосфаткиназу и полифосфат. Таким образом, способы, кроме того, предусматривают инкубирование реакционной смеси в условиях, подходящих для получения НТФ. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит НДФ-киназу. Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ также может включать в себя нуклеозидкиназу. В качестве альтернативы, способы получения НТФ могут предусматривать инкубирование в реакционной смеси клеточной РНК, рибонуклеазы, которая расщепляет РНК на 3’-НМФ, и соответствующей фосфатазы (например, щелочной фосфатазы или других) в условиях, подходящих для получения нуклеозидов. Фосфатаза затем может быть элиминирована перед осуществлением получения НТФ.
Пути получения РНК
Как показано на фиг. 1, РНК (например, мРНК или двухнитевая РНК) могут быть получены посредством ряда разных ферментативных путей, в каждом из которых используются источники энергии, описываемые в настоящем документе, и недорогие исходные материалы в реакционной(ых) смеси(ях). Таким образом, в настоящем документе представлены системы, способы, композиции и наборы для получения РНК.
Превращение НДФ в РНК.Согласно некоторым аспектам РНК получают с использованием НДФ в качестве субстратов, как показано на фиг. 2A. Например, способы получения РНК могут предусматривать инкубирование в реакционной смеси НДФ, полифосфаткиназы, полифосфата, ДНК-матрицы и РНК-полимеразы в условиях, подходящих для получения РНК. Согласно некоторым вариантам осуществления реакционная смесь для получения РНК включает в себя НДФ-киназу (см., например, таблицу 5). Согласно некоторым вариантам осуществления реакционная смесь для получения РНК также может включать в себя нуклеозидкиназу.
Превращение НМФ в РНК.Согласно некоторым аспектам РНК получают с использованием 5’-НМФ в качестве субстратов, как показано на фиг. 2B. Например, способы получения РНК могут предусматривать инкубирование в реакционной смеси 5’-НМФ, полифосфаткиназы, полифосфата, ДНК-матрицы и РНК-полимеразы в условиях, подходящих для получения РНК. Согласно некоторым вариантам осуществления реакционная смесь для получения РНК включает в себя НМФ-киназу (см., например, таблицу 4) и/или НДФ-киназу (см., например, таблицу 5). Согласно некоторым вариантам осуществления реакционная смесь для получения РНК также может включать в себя нуклеозидкиназу.
Превращение нуклеозидов в РНК.Согласно некоторым аспектам РНК получают с использованием нуклеозидов в качестве субстратов, как показано на фиг. 2C. Например, способы получения РНК могут предусматривать инкубирование в реакционной смеси нуклеозидов, полифосфаткиназы, полифосфата, ДНК-матрицы и РНК-полимеразы в условиях, подходящих для получения РНК. Согласно некоторым вариантам осуществления реакционная смесь для получения РНК также может включать в себя нуклеозидкиназу (см., например, таблицу 3) и/или НМФ-киназу (см., например, таблицу 4) и/или НДФ-киназу (см., например, таблицу 5).
Превращение клеточной РНК в РНК через НДФ. Согласно некоторым аспектам РНК получают с использованием клеточной РНК в качестве субстрата сначала путем разрыва клеточной РНК на НДФ, как показано на фиг. 3A. Например, способы получения РНК могут предусматривать инкубирование в реакционной смеси клеточной РНК, полинуклеотидфосфорилазы (ПНФазы) и фосфата в условиях, подходящих для получения НДФ. Перед осуществлением получения РНК может быть выгодным элиминирование ПНФазы, чтобы избежать разложения конечного продукта. Таким образом, способы могут дополнительно предусматривать элиминирование ПНФазы и инкубирование в реакционной смеси или во второй реакционной смеси НДФ, полифосфаткиназы, полифосфата, ДНК-матрицы и полимеразы в условиях, подходящих для получения РНК. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит НДФ-киназу. Согласно некоторым вариантам осуществления реакционная смесь для получения РНК также может включать в себя нуклеозидкиназу.
Согласно некоторым вариантам осуществления эти ферменты пути способны выдерживать условия элиминирования, как обсуждается ниже, и, таким образом, все компоненты реакции включают в одну (одностадийную) реакционную смесь. Например, способ получения РНК может предусматривать (a) инкубирование в реакционной смеси клеточной РНК, ПНФазы, фосфата, полифосфаткиназы, полифосфата, ДНК-матрицы и полимеразы в условиях, подходящих для получения НДФ (необязательно при этом реакционная смесь дополнительно содержит НДФ-киназу), (b) элиминирование ПНФазы и (c) инкубирование реакционной смеси в условиях, подходящих для получения РНК.
Превращение клеточной РНК в РНК через НМФ. Согласно некоторым аспектам РНК получают с использованием клеточной РНК в качестве субстрата сначала путем разрыва клеточной РНК на 5’-НМФ, как показано на фиг. 3B. Например, способы получения РНК могут предусматривать инкубирование в реакционной смеси клеточной РНК и рибонуклеазы в условиях, подходящих для получения 5’-НМФ. Перед осуществлением получения РНК может быть выгодным элиминирование рибонуклеазы, чтобы избежать разложения конечного продукта. Таким образом, способы могут дополнительно предусматривать элиминирование рибонуклеазы и инкубирование в реакционной смеси или во второй реакционной смеси 5’-НМФ, полифосфаткиназы, полифосфата, ДНК-матрицы и полимеразы в условиях, подходящих для получения РНК. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит НМФ-киназу и/или НДФ-киназу. Согласно некоторым вариантам осуществления реакционная смесь для получения РНК также может включать в себя нуклеозидкиназу.
Согласно некоторым вариантам осуществления эти ферменты пути способны выдерживать условия элиминирования, как обсуждается ниже, и, таким образом, все компоненты реакции включают в одну (одностадийную) реакционную смесь. Например, способ получения РНК может предусматривать (a) инкубирование в реакционной смеси клеточной РНК, рибонуклеазы, полифосфаткиназы, полифосфата, ДНК-матрицы и полимеразы в условиях, подходящих для получения НМФ (необязательно при этом реакционная смесь дополнительно содержит НМФ-киназу и/или НДФ-киназу), (b) элиминирование рибонуклеазы и (c) инкубирование реакционной смеси в условиях, подходящих для получения РНК.
Превращение нуклеиновых оснований в РНК.Согласно некоторым аспектам РНК получают с использованием нуклеиновых оснований в качестве субстратов, как показано на фиг. 2D. Например, способы получения РНК могут предусматривать инкубирование в реакционной смеси нуклеиновых оснований, (например, 1, 2, 3 или 4) фосфорибозилтрансферазы, фосфорибозилпирофосфата, полифосфаткиназы, полифосфата, ДНК-матрицы и РНК-полимеразы в условиях, подходящих для получения РНК. Согласно некоторым вариантам осуществления реакционная смесь для получения РНК также может включать в себя НМФ-киназу (см., например, таблицу 4) и/или НДФ-киназу (см., например, таблицу 5). Согласно некоторым вариантам осуществления реакционная смесь для получения РНК также может включать в себя нуклеозидкиназу.
Превращение нуклеиновых оснований и рибозы в РНК.Согласно некоторым аспектам РНК получают с использованием нуклеиновых оснований в качестве субстратов, как показано на фиг. 2E. Например, способы получения РНК могут предусматривать инкубирование в реакционной смеси нуклеиновых оснований, D-рибозы, рибокиназы, фосфопентомутазы, по меньшей мере одной (например, 1, 2, 3 или 4) нуклеозидфосфорилазы, по меньшей мере одной полифосфаткиназы, по меньшей мере одного полифосфата, по меньшей мере одной ДНК-матрицы и по меньшей мере одной РНК-полимеразы в условиях, подходящих для получения РНК. Согласно некоторым вариантам осуществления реакционная смесь для получения РНК также может включать в себя по меньшей мере одну НМФ-киназу (см., например, таблицу 3) и/или по меньшей мере одну НДФ-киназу (см., например, таблицу 4) и/или нуклеозидкиназу.
Источники ферментов
Любые (например, один, два, три или более) или все ферменты пути, представленные в настоящем документе (например, нуклеазы, киназы, полимеразы и т.д.), могут быть эндогенными (немодифицированными) ферментами или рекомбинантными ферментами, экспрессируемыми клеткой. Согласно некоторым вариантам осуществления ферменты пути обеспечивают как компонент клеточного лизата, который включают в реакционную смесь. Согласно некоторым вариантам осуществления ферменты пути очищают от клеточного лизата, включенного в реакционную смесь. Согласно некоторым вариантам осуществления фермент пути обеспечивают как компонент клеточного лизата и фермент пути очищают от клеточного лизата. Согласно некоторым вариантам осуществления фермент пути секретируется и необязательно очищается от клеточного бульона.
Согласно некоторым вариантам осуществления фермент пути (например, нуклеазы, киназы, полимеразы и т.д.) является эндогенным ферментом, очищенным от клетки и включенным в реакционную смесь в виде очищенного фермента. Согласно некоторым вариантам осуществления фермент пути (например, нуклеазы, киназы, полимеразы и т.д.) является эндогенным ферментом, обеспеченным в виде компонента клеточного лизата, который включен в реакционную смесь. Согласно некоторым вариантам осуществления фермент пути (например, нуклеазы, киназы, полимеразы и т.д.) является рекомбинантным ферментом, очищенным от клетки и включенным в реакционную смесь в виде очищенного фермента. Согласно некоторым вариантам осуществления фермент пути (например, нуклеазы, киназы, полимеразы и т.д.) является рекомбинантным ферментом, обеспеченным в виде компонента клеточного лизата, который включен в реакционную смесь. Согласно некоторым вариантам осуществления фермент пути секретируется и необязательно очищается от клеточного бульона.
Настоящее раскрытие также относится к эндогенным ферментам и рекомбинантным ферментам, секретируемым клеткой. Таким образом, согласно некоторым вариантам осуществления фермент пути (например, нуклеазы, киназы, полимеразы и т.д.) является эндогенным ферментом, секретируемым из клетки. Согласно некоторым вариантам осуществления фермент пути (например, нуклеазы, киназы, полимеразы и т.д.) является рекомбинантным ферментом, секретируемым из клетки.
Элиминирование нежелательных ферментативных активностей
Согласно различным вариантам осуществления, представленным в настоящем документе, ферменты, полученные из клеток или лизатов клеток, которые экспрессируют ферменты пути, используют в реакционной смеси для получения НТФ и/или РНК. В этих клетках или клеточных лизатах есть ферменты, которые могут оказывать вредные воздействия на получение НТФ и/или РНК. Неограничивающие примеры таких ферментов включают в себя фосфатазы, нуклеазы, протеазы, дезаминазы, оксидоредуктазы и/или гидролазы, такие как экспрессируемые клетками Escherichia coli. Фосфатазы удаляют фосфатные группы (например, превращая НМФ в нуклеозиды, превращая НДФ в НМФ или превращая НТФ в НДФ), что снижает получение НТФ из-за футильных циклов фосфорилирования/дефосфорилирования нуклеотида. Нуклеазы расщепляют нуклеиновые кислоты на мономеры или олигомеры, что приводит к разложению РНК-продукта (например, РНКазой) и/или к разложению ДНК-матрицы (например, ДНКазой). Протеазы расщепляют белки на аминокислоты или пептиды, что разлагает ферменты пути. Дезаминазы удаляют аминогруппы, что снижает концентрации НТФ из-за превращения промежуточных соединений пути в бесполезные субстраты (например, ксантин и гипоксантин) и может приводить к мутациям в РНК-продуктах (например, Ц в У). Гидролазы (например, нуклеозидгидролаза или нуклеотидгидролаза) расщепляют нуклеозиды или нуклеотиды на основание и сахарные фрагменты, что снижает концентрации НТФ из-за необратимого разложения нуклеотидов. Оксидоредуктазы катализируют перенос электронов от одной молекулы (окислителя) к другой молекуле (восстановителю). Реакции окисления и/или восстановления могут, например, повреждать нуклеиновые основания в ДНК и/или РНК, приводя к ошибкам в транскрипции и/или трансляции, или повреждать белки или ферменты, что приводит к утрате функции.
Таким образом, согласно многим вариантам осуществления выгодно элиминировать такие нативные ферментативные активности или другие нежелательные ферментативные активности в ферментном препарате, лизате клеток и/или реакционной смеси. В настоящем документе термин «элиминирование» ферментативных активностей может означать частичной (например, элиминирование по меньшей мере 30%, 40%, 50%, 60%, 70%, 80% или 90% активности) или полной (элиминирование 100% активности) элиминирование нежелательной ферментативной активности. Как обсуждается в настоящем документе, ферментативная активность может быть элиминирована с помощью генетической модификации, условной инактивации и/или физического отделения. Также могут быть использованы другие способы элиминирования. Нежелательная ферментативная активность может быть обусловлена по меньшей мере одним (например, 1, 2, 3, 4 или 5) нативным (эндогенным) ферментом, включающим в себя без ограничения фосфатазы, нуклеазы, протеазы, дезаминазы, оксидоредуктазы и/или гидролазы.
Согласно некоторым вариантам осуществления нежелательную фосфатазную активность элиминируют в ферментативном препарате, клеточном лизате и/или реакционной смеси. Согласно некоторым вариантам осуществления нежелательную нуклеазную активность элиминируют в ферментативном препарате, клеточном лизате и/или реакционной смеси. Согласно некоторым вариантам осуществления нежелательную протеазную активность элиминируют в ферментативном препарате, клеточном лизате и/или реакционной смеси. Согласно некоторым вариантам осуществления нежелательную дезаминазную активность элиминируют в ферментативном препарате, клеточном лизате и/или реакционной смеси. Согласно некоторым вариантам осуществления нежелательную гидролазную активность элиминируют в ферментативном препарате, клеточном лизате и/или реакционной смеси.
Нежелательная(ые) (например, нативная(ые)) ферментативная(ые) активность(и) может(могут) быть элиминирована(ы) с использованием генетических, условных подходов или подходов с отделением. Согласно некоторым вариантам осуществления генетический подход используют для удаления нежелательной ферментативной активности. Таким образом, согласно некоторым вариантам осуществления клетки модифицируют для снижения или элиминирования нежелательных ферментативных активностей. Примеры генетических подходов, которые могут быть использованы для снижения или элиминирования нежелательной ферментативной активности, включают в себя без ограничения секрецию, нокауты генов и нацеливание протеазы. Согласно некоторым вариантам осуществления условный подход используют для удаления нежелательной ферментативной активности. Таким образом, согласно некоторым вариантам осуществления нежелательные ферменты, демонстрирующие нежелательные активности, остаются в ферментативном препарате, клеточном лизате и/или реакционной смеси и селективно инактивируются. Примеры условных подходов, которые могут быть использованы для снижения или элиминирования нежелательной ферментативной активности, включают в себя без ограничения изменения температуры, pH, соли, детергента, органического растворителя (например, спирта) и применение химических ингибиторов. Согласно некоторым вариантам осуществления подход с отделением/очисткой используют для удаления нежелательной ферментативной активности. Таким образом, согласно некоторым вариантам осуществления нежелательные ферменты, демонстрирующие нежелательные активности, физически удаляют из ферментативного препарата, клеточного лизата и/или реакционной смеси. Примеры подходов с отделением, которые могут быть использованы для снижения или элиминирования нежелательной ферментативной активности, включают в себя без ограничения осаждение, иммобилизацию, фильтрацию и хроматографию.
Генетические подходы. Согласно некоторым вариантам осуществления клетки, экспрессирующие фермент и/или ДНК-матрицу пути получения НТФ и/или РНК, модифицируют для снижения или элиминирования нежелательных ферментативных активностей. Согласно некоторым вариантам осуществления ген, кодирующий фермент, демонстрирующий нежелательную активность, делетируют из клеток. Согласно некоторым вариантам осуществления осуществляют мутацию гена, кодирующего фермент, демонстрирующий нежелательную активность, так, что полученный в результате генный продукт становится нефункциональным. Согласно некоторым вариантам осуществления фермент, демонстрирующий нежелательную активность, модифицируют с включением сайт-специфической последовательности распознавания протеазой в их белковую последовательность так, что на фермент можно «нацеливаться» и расщеплять для инактивации (см., например, публикацию патентного документа США № 2012/0052547 A1, опубликованную 1 марта 2012 года, публикацию международной заявки № WO2015/021058 A2, опубликованную 12 февраля 2015 года, и публикацию международной заявки № WO2012/030980, опубликованную 8 марта 2012 года, каждая из которых включена в настоящий документ посредством ссылки).
Расщепление фермента, содержащего сайт-специфическую последовательность распознавания протеазой, является результатом контакта с когнатой сайт-специфической протеазой, которая секвестируется в периплазме клетки (отдельно от целевого фермента) в ходе фазы роста клетки (например, при культивировании созданных методами генной инженерии клеток) и вводится в контакт с ферментом в ходе фазы получения АТФ (например, после лизиса клеток с получением клеточного лизата). Таким образом, созданные методами генной инженерии клетки в соответствии с настоящим раскрытием содержат согласно некоторым вариантам осуществления (i) созданную методами генной инженерии нуклеиновую кислоту, кодирующую фермент, демонстрирующий нежелательную активность, и включающую в себя сайт-специфическую последовательность распознавания протеазой в белковой последовательности фермента, и (ii) созданную методами генной инженерии нуклеиновую кислоту, кодирующую сайт-специфическую протеазу, которая расщепляет сайт-специфическую последовательность распознавания протеазой фермента и включает в себя нацеленную на периплазму последовательность. Эта нацеленная на периплазму последовательность отвечает за секвестирование сайт-специфической протеазы в периплазматическом пространстве клетки до тех пор, пока клетка не лизируется. Примеры нацеленных на периплазму последовательностей известны.
Примеры протеаз, которые могут быть использованы в соответствии с настоящим раскрытием включают в себя без ограничения аланинкарбоксипептидазу, актацин, бактериальную лейциламинопептидазу, прокоагулянт рака, катепсин B, клострипаин, цитозольную аланинаминопептидазу, эластазу, эндопротеиназу Brg-C, энтерокиназу, гастриксин, желатиназу, Gly-X карбоксипептидазу, глицилэндопептидазу, протеазу 3C риновируса человека, гиподермин C, Iga-специфическкю серинэндопептидазу, лейциламинопептидазу, лейцилэндопептидазу, lysC, лизосомальную pro-X карбоксипептидазу, лизиламинопептидазу, метиониламинопептидазу, миксобактер, нардилизин, панкреатическую эндопептидазу E, пикорнаин 2B, пикорнаин 3C, проэндопептидазу, пролиламинопептидазу, пропротеинконвертазу I, пропротеинконвертазу II, расселлизин, сахаропепсин, семеногелазу, активатор T-плазминогена, тромбин, тканевой калликреин, вирус гравировки табака (TEV), тогавирин, триптофаниламинопептидазу, активатор U-плазминогена, V8, веномбин B, веномбин BB и Xaa-pro аминопептидазу.
Условные подходы. Согласно некоторым вариантам осуществления ферментативный препарат, клеточный лизат и/или реакционная смесь включают в себя фермент, демонстрирующий нежелательную активность, которую селективно инактивируют. Согласно некоторым вариантам осуществления фермент, демонстрирующий нежелательную активность, селективно инактивируют путем воздействия на фермент условий элиминирования (например, высокой или низкой температуры, кислотного или основного значения pH, высокой концентрации соли или низкой концентрации соли, детергента и/или органического растворителя).
Согласно некоторым вариантам осуществления ферментативный препарат, клеточный лизат и/или реакционную смесь подвергают воздействию температуры, которая временно или необратимо инактивирует фермент, демонстрирующий нежелательную активность. Термин «инактивация температурой» относится к процессу нагревания или охлаждения ферментативного препарата, клеточного лизата и/или реакционной смеси до температуры, достаточной для инактивации (или по меньшей мере частичной инактивации) нативного целевого фермента. Как правило, процесс температурной инактивации предусматривает денатурацию (разворачивание) вредного фермента. Температура, при которой фермент денатурируется, варьирует у организмов. У E. coli, например, ферменты, как правило, денатурируются при температурах выше 41°C. Температура денатурации может быть выше или ниже 41°C для других организмов. Ферменты клеточного лизата, как представлено в настоящем документе, могут быть инактивированы температурой при температуре 0°C - 95°C или выше. Согласно некоторым вариантам осуществления ферменты клеточного лизата инактивируют температурой при температуре 0-90°C, 0-80°C, 0-70°C, 0-60°C, 0-50°C, 0-40°C, 0-30°C, 0-20°C, 0-10°C или 0-5°C. Согласно некоторым вариантам осуществления ферменты клеточного лизата инактивированы температурой при температуре 5-95°C, 10-95°C, 20-95°C, 30-95°C, 40-95°C, 50-95°C, 60-95°C, 70-95°C, 80-95°C или 90-95°C. Например, ферменты клеточного лизата могут быть инактивированы температурой при температуре приблизительно 40°C, 42°C, 45°C, 50°C, 55°C, 60°C, 65°C, 70°C, 75°C, 80°C, 85°C, 90°C или 95°C. Согласно некоторым вариантам осуществления ферменты клеточного лизата инактивируют температурой при температуре 50-80°C. Согласно некоторым вариантам осуществления ферменты клеточного лизата инактивируют температурой при температуре приблизительно 70°C. Согласно некоторым вариантам осуществления ферменты клеточного лизата инактивируют температурой при температуре приблизительно 60°C.
Согласно некоторым вариантам осуществления ферментативный препарат, клеточный лизат и/или реакционную смесь подвергают воздействию кислоты или основания (изменения pH), что временно или необратимо инактивирует фермент, демонстрирующий нежелательную активность. Термин «инактивация кислотой или основанием» относится к процессу доведения ферментативного препарата, клеточного лизата и/или реакционной смеси до pH, достаточного для инактивации (или по меньшей мере частичной инактивации) фермента. Как правило, процесс инактивации кислотой или основанием предусматривает денатурацию (разворачивание) фермента. pH, при котором фермент денатурируется, варьирует у организмов. У E. coli, например, нативные ферменты, как правило, денатурируются при pH выше 7,5 или ниже 6,5. pH денатурации может быть выше или ниже, чем pH денатурации для других организмов. Ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси, как представлено в настоящем документе, могут быть инактивированы основанием при pH 7,5-14, или выше. Согласно некоторым вариантам осуществления ферменты клеточного лизата инактивируют основанием при pH 8-14, 8,5-14, 9-14, 9,5-14, 10-14, 10,5-14, 11-14, 11,5-14, 12-14, 12,5-14, 13-14 или 13,5-14. Согласно некоторым вариантам осуществления ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси инактивируют основанием при pH 7,5-13,5, 7,5-13, 7,5-12,5, 7,5-12, 7,5-11,5, 7,5-11, 7,5-10,5, 7,5-10, 7,5-9,5, 7,5-9, 7,5-8,5 или 7,5-8. Например, ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси могут быть инактивированы основанием при pH приблизительно 7,5, 8, 8,5, 9, 9,5, 10, 10,5, 11, 11,5, 12, 12,5, 13, 13,5 или 14. Ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси, как представлено в настоящем документе, могут быть инактивированы кислотой при pH 6,5-0 или ниже. Согласно некоторым вариантам осуществления ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси инактивируют кислотой при pH 6,5-0,5, 6,5-1, 6,5-1,5, 6,5-2, 6,5-2,5, 6,5-3, 6,5-3,5, 6,5-4, 6,5-4,5, 6,5-5 или 6,5-6. Согласно некоторым вариантам осуществления ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси инактивируют кислотой при pH 6-0, 5,5-0, 5-0, 4,5-0, 4-0, 3,5-0, 3-0, 2,5-0, 2-0, 1,5-0, 1-0 или 0,5-0. Например, ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси могут быть инактивированы кислотой при pH приблизительно 6,5, 6, 5,5, 5, 4,5, 4, 3,5, 3, 2,5, 2, 1,5, 1, 0,5 или 0.
Согласно некоторым вариантам осуществления ферментативный препарат, клеточный лизат и/или реакционную смесь подвергают воздействию соли высокой концентрации или соли низкой концентрации (изменению концентрации соли), что временно или необратимо инактивирует фермент, демонстрирующий нежелательную активность. Термин «инактивация солью» относится к процессу доведения ферментативного препарата, клеточного лизата и/или реакционной смеси до концентрации соли, достаточной для инактивации (или частичной инактивации) фермента. Как правило, процесс инактивации солью предусматривает денатурацию (разворачивание) фермента. Концентрация соли, при которой ферменты денатурируются, варьирует у организмов. У E. coli, например, нативные ферменты, как правило, денатурируются при концентрации соли выше 600 мM. Денатурирующая концентрация соли может быть выше или ниже денатурирующей концентрации соли для других организмов. Соли представляют собой комбинации анионов и катионов. Неограничивающие примеры катионов включают в себя литий, натрий, калий, магний, кальций и аммоний. Неограничивающие примеры анионов включают в себя ацетат, хлорид, сульфат и фосфат. Ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси, как представлено в настоящем документе, могут быть инактивированы солью при концентрации соли 600-1000 мM или выше. Согласно некоторым вариантам осуществления ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси инактивируют солью при концентрации соли 700-1000 мМ, 750-1000 мМ, 800-1000 мМ, 850-1000 мМ, 900-1000 мМ, 950-1000 мМ. Согласно некоторым вариантам осуществления ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси инактивируют солью при концентрации соли 600-950 мМ, 600-900 мМ, 600-850 мМ, 600-800 мМ, 600-750 мМ, 600-700 мМ или 600-650 мМ. Например, ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси могут быть инактивированы солью при концентрации соли приблизительно 600 мМ, 650 мМ, 700 мМ, 750 мМ, 800 мМ, 850 мМ, 900 мМ, 950 мМ или 1000 мМ. Ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси, как представлено в настоящем документе, могут быть инактивированы солью при концентрации соли 400-0 мМ или ниже. Согласно некоторым вариантам осуществления ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси инактивируют солью при концентрации соли 350-0 мМ, 300-0 мМ, 250-0 мМ, 200-0 мМ, 150-0 мМ, 100-0 мМ или 50-0 мМ. Согласно некоторым вариантам осуществления ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси инактивируют солью при концентрации соли 400-50 мМ, 400-100 мМ, 400-150 мМ, 400-200 мМ, 400-250 мМ, 400-300 мМ или 400-350 мМ. Например, ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси могут быть инактивированы солью при концентрации соли приблизительно 400 мМ, 350 мМ, 300 мМ, 250 мМ, 200 мМ, 150 мМ, 100 мМ, 50 мМ или 0 мМ.
Согласно некоторым вариантам осуществления органический растворитель добавляют в ферментативный препарат, клеточный лизат и/или реакционную смесь для инактивации фермента, демонстрирующего нежелательную активность. Неограничивающие примеры органических растворителей включают в себя этанол, метанол, эфир, диоксан, ацетон, метилэтилкетон, ацетонитрил, диметилсульфоксид и толуол.
Согласно некоторым вариантам осуществления детергент добавляют в ферментативный препарат, клеточный лизат и/или реакционную смесь для инактивации фермента, демонстрирующего нежелательную активность. Неограничивающие примеры детергентов включают в себя додецилсульфат натрия (SDS), этилтриметиламмония бромид (ETMAB), лаурилтриметиламмония бромид (LTAB) и лаурилтриметиламмония хлорид (LTAC).
Согласно некоторым вариантам осуществления химический ингибитор добавляют в ферментативный препарат, клеточный лизат и/или реакционную смесь для инактивации фермента, демонстрирующего нежелательную активность. Неограничивающие примеры химических ингибиторов включают в себя ортованадат натрия (ингибитор протеинфосфотирозилфосфатаз), фторид натрия (ингибитор фосфосерил- и фосфотреонилфосфатаз), пирофосфат натрия (ингибитор фосфатазы), фосфат натрия и/или фосфат калия. Согласно некоторым вариантам осуществления химические ингибиторы выбраны из библиотеки химических ингибиторов.
Для любого из условных подходов, используемых в настоящем документе, следует учитывать, что любой из ферментов пути, присутствующий в клеточном лизате или реакционной смеси, также может быть подвергнут условиям элиминирования (например, высокой или низкой температуре, кислотному или основному значению pH, соли высокой концентрации или соли низкой концентрации, детергенту и/или органическому растворителю). Таким образом, согласно некоторым вариантам осуществления ферменты пути (например, полифосфаткиназа, НМФ-киназа, НДФ-киназа и/или полимераза) могут выдерживать условия элиминирования. Считают, что фермент выдерживает условия элиминирования, если фермент после воздействия условий элиминирования сохраняет по меньшей мере 10% (например, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90%) своей ферментативной активности (относительно ферментативной активности до воздействия условия инактивации).
Например, если нативные ферменты ферментативного препарата, клеточного лизата и/или реакционной смеси инактивируются нагреванием (например, воздействием температуры по меньшей мере 40°C или 40-95°C в течение по меньшей мере 2 минут или 2-60 минут), то ферменты пути могут быть термостабильными ферментами. Таким образом, согласно некоторым вариантам осуществления по меньшей мере одна из полифосфаткиназы, НМФ-киназы, НДФ-киназы, нуклеозидкиназы, фосфорибозилтрансферазы, нуклеозидфосфорилазы, рибокиназы, фосфопентомутазы и полимеразы является термостабильной. Фермент (например, киназа или полимераза) считают термостабильным, если фермент (a) сохраняет активность после временного воздействия высоких температур, которые денатурируют нативные ферменты, или (b) функционирует с высокой скоростью после временного воздействия температуры, от средней до высокой, при которой нативные ферменты функционируют при низких скоростях. Термостабильные ферменты известны, и неограничивающие примеры термостабильных ферментов для применения представлены в настоящем документе. Другие неограничивающие примеры ферментов пути, которые могут выдерживать условия элиминирования, также представлены в настоящем документе.
Подходы с отделением. Согласно некоторым вариантам осуществления нативный фермент, демонстрирующий нежелательную активность, физически удаляют из ферментативного препарата, клеточного лизата и/или реакционной смеси. Согласно некоторым вариантам осуществления фермент, демонстрирующий нежелательную активность, осаждают из ферментативного препарата, клеточного лизата и/или реакционной смеси. Согласно некоторым вариантам осуществления фермент, демонстрирующий нежелательную активность, фильтруют (например, на основании размера) из ферментативного препарата, клеточного лизата и/или реакционной смеси. Согласно некоторым вариантам осуществления фермент, демонстрирующий нежелательную активность, удаляют из ферментативного препарата, клеточного лизата и/или реакционной смеси путем захвата и/или хроматографии (например, с использованием дифференциальной аффинности по отношению к стационарной фазе).
Согласно некоторым вариантам осуществления фермент, демонстрирующий нежелательную активность, удаляют из ферментативного препарата, клеточного лизата и/или реакционной смеси путем аффинной хроматографии. Примеры аффинной хроматографии включают в себя без ограничения хроматографию с использованием белка А, хроматографию с использованием белка G, металл-связывающую хроматографию (например, связывающую никель хроматографию), лектиновую хроматографию и хроматографию с использованием GST.
Согласно некоторым вариантам осуществления фермент, демонстрирующий нежелательную активность, удаляют из ферментативного препарата, клеточного лизата и/или реакционной смеси путем ионообменной хроматографии. Примеры анионообменной хроматографии (AEX) включают в себя без ограничения хроматографию с использованием диэтиламиноэтила (DEAE), хроматографию с использованием четвертичного аминоэтила (QAE) и хроматографию с использованием четвертичного амина (Q). Примеры катионообменной хроматографии включают в себя без ограничения хроматографию с использованием карбоксиметила (CM), хроматографию с использованием сульфоэтила (SE), хроматографию с использованием сульфопропила (SP), хроматографию с использованием фосфата (P) и хроматографию с использованием сульфоната (S).
Согласно некоторым вариантам осуществления фермент, демонстрирующий нежелательную активность, удаляют из ферментативного препарата, клеточного лизата и/или реакционной смеси путем хроматографии с гидрофобным взаимодействием (HIC). Примеры хроматографии с гидрофобным взаимодействием включают в себя без ограничения хроматографию с использованием фенилсефарозы, хроматографию с использованием бутилсефарозы, хроматографию с использованием октилсефарозы, хроматографию с использованием каптофенила, хроматографию с использованием Toyopearl Butyl, хроматографию с использованием Toyopearl Phenyl, хроматографию с использованием Toyopearl Hexyl, хроматографию с использованием Toyopearl Ether и хроматографию с использованием Toyopearl PPG. Любые из химических веществ, подробно описанных выше, могут быть, в качестве альтернативы, использованы для иммобилизации или захвата ферментов пути.
Термостабильные ферменты
Любые из ферментов пути, представленных в настоящем документе (например, нуклеазы, киназы, полимеразы и т.д.), могут быть термостабильными ферментами. Термостабильность относится к качеству ферментов, заключающемуся в устойчивости к денатурации при относительно высокой или низкой температуры. Например, если фермент денатурируется (инактивируется) при температуре 42°C, то фермент, обладающий подобной активностью (например, киназной активностью) считают «термостабильным», если он не денатурируется при 42°C.
Фермент (например, киназу или полимеразу) считают термостабильным, если фермент (a) сохраняет активность после временного воздействия высоких температур, которые денатурируют другие нативные ферменты, или (b) функционирует при высокой скорости после временного воздействия температуры, от средней до высокой, при которой нативные ферменты функционируют при низких скоростях.
Фермент (например, киназу или полимеразу) также считают термостабильным, если фермент (a) сохраняет активность после временного воздействия низких температур, которые денатурируют другие нативные ферменты, или (b) функционирует при высокой скорости после временного воздействия температуры, от средней до низкой, при которой нативные ферменты функционируют при низких скоростях.
Согласно некоторым вариантам осуществления термостабильный фермент сохраняет более чем 10% активности после временного воздействия относительно высокой температуры (например, выше чем 41°C для киназ, полученных из E. coli, выше чем 37°C для многих РНК-полимераз), которая в других случаях денатурирует подобный (нетермостабильный) нативный фермент. Согласно некоторым вариантам осуществления термостабильный фермент сохраняет 10-100%, 25-100% или 50-100% активности после временного воздействия относительно высокой температуры, которая в других случаях денатурирует подобный (нетермостабильный) нативный фермент. Например, термостабильный фермент может сохранять 10-90%, 10-85%, 10-80%, 10-75%, 10-70%, 10-65%, 10-60%, 10-55%, 25-90%, 25-85%, 25-80%, 25-75%, 25-70%, 25-65%, 25-60%, 25-55%, 50-90%, 50-85%, 50-80%, 50-75%, 50-70%, 50-65%, 50-60% или 50-55% активности после временного воздействия относительно высокой температуры, которая в других случаях денатурирует подобный (нетермостабильный) нативный фермент. Согласно некоторым вариантам осуществления термостабильный фермент сохраняет 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или 100% активности после временного воздействия относительно высокой температуры, которая в других случаях денатурирует подобный (нетермостабильный) нативный фермент.
Согласно некоторым вариантам осуществления термостабильный фермент сохраняет более чем 50% активности после временного воздействия относительно низкой температуры (например, ниже чем 32°C для киназ, полученных из E. coli, ниже чем 32°C для многих РНК-полимераз), которая в других случаях денатурирует подобный (нетермостабильный) нативный фермент. Согласно некоторым вариантам осуществления термостабильный фермент сохраняет 50-100% активности после временного воздействия относительно низкой температуры, которая в других случаях денатурирует подобный (нетермостабильный) нативный фермент. Например, термостабильный фермент может сохранять 50-90%, 50-85%, 50-80%, 50-75%, 50-70%, 50-65%, 50-60% или 50-55% активности после временного воздействия относительно низкой температуры, которая в других случаях денатурирует подобный (нетермостабильный) нативный фермент. Согласно некоторым вариантам осуществления термостабильный фермент сохраняет 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или 100% активности после временного воздействия относительно низкой температуры, которая в других случаях денатурирует подобный (нетермостабильный) нативный фермент.
Согласно некоторым вариантам осуществления активность термостабильного фермента после временного воздействия температуры, от средней до высокой (например, 42-80°C), больше чем (например, на 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или 100% больше) активность подобного (нетермостабильного) нативного фермента.
Согласно некоторым вариантам осуществления активность термостабильного фермента после временного воздействия температуры, от средней до низкой (например, 32-0°C), больше чем (например, на 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или 100% больше) активность подобного (нетермостабильного) нативного фермента.
Активность термостабильной киназы, например, может быть измерена по количеству НМФ- или НДФ-киназы, способной к фосфорилированию. Таким образом, согласно некоторым вариантам осуществления термостабильная киназа при относительно высокой температуре (например, 42°C) превращает более чем 50% НМФ в НДФ или более чем 50% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C. Согласно некоторым вариантам осуществления термостабильная киназа при относительно высокой температуре (например, 42°C) превращает более чем 60% НМФ в НДФ или более чем 60% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C. Согласно некоторым вариантам осуществления термостабильная киназа при относительно высокой температуре (например, 42°C) превращает более чем 70% НМФ в НДФ или более чем 70% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C. Согласно некоторым вариантам осуществления термостабильная киназа при относительно высокой температуре (например, 42°C) превращает более чем 80% НМФ в НДФ или более чем 80% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C. Согласно некоторым вариантам осуществления термостабильная киназа при относительно высокой температуре (например, 42°C) превращает более чем 90% НМФ в НДФ или более чем 90% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C.
Согласно некоторым вариантам осуществления термостабильная киназа при относительно низкой температуре (например, 32°C) превращает более чем 50% НМФ в НДФ или более чем 50% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C. Согласно некоторым вариантам осуществления термостабильная киназа при относительно низкой температуре (например, 32°C) превращает более чем 60% НМФ в НДФ или более чем 60% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C. Согласно некоторым вариантам осуществления термостабильная киназа при относительно низкой температуре (например, 32°C) превращает более чем 70% НМФ в НДФ или более чем 70% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C. Согласно некоторым вариантам осуществления термостабильная киназа при относительно низкой температуре (например, 32°C) превращает более чем 80% НМФ в НДФ или более чем 80% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C. Согласно некоторым вариантам осуществления термостабильная киназа при относительно низкой температуре (например, 32°C) превращает более чем 90% НМФ в НДФ или более чем 90% НДФ в НТФ за тот же промежуток времени, необходимый для завершения подобного превращения при 37°C.
Активность термостабильной полимеразы, например, оценивают на основании точности и кинетических параметров полимеризации (например, скорости полимеризации). Таким образом, одна единица термостабильной T7 полимеразы, например, может включать 10 нмоль НТФ в нерастворимый в кислоте материал за 30 минут при температурах выше 37°C (например, при 50°C). В другом примере одна единица термостабильной T7 полимеразы, например, может включать 10 нмоль НТФ в нерастворимый в кислоте материал за 30 минут при температурах ниже 32°C (например, при 25°C).
Согласно некоторым вариантам осуществления термостабильные ферменты (например, киназы или полимеразы) могут сохранять активность (способность катализировать реакцию) при температуре от 42°C до 80°C или выше. Согласно некоторым вариантам осуществления термостабильные ферменты сохраняют активность при температуре 42-80°C, 42-70°C, 42-60°C, 42-50°C, 50-80°C, 50-70°C, 50-60°C, 60-80°C, 60-70°C или 70-80°C. Например, термостабильные ферменты могут сохранять активность при температуре 42°C, 43°C, 44°C, 45°C, 46°C, 47°C, 48°C, 49°C, 50°C, 51°C, 52°C, 53°C, 54°C, 55°C, 55°C, 56°C, 57°C, 58°C, 59°C, 60°C, 61°C, 62°C, 63°C, 64°C, 65°C, 66°C, 67°C, 68°C, 69°C, 70°C, 71°C, 72°C, 73°C, 74°C, 75°C, 76°C, 77°C, 78°C, 79°C или 80°C. Термостабильные ферменты могут сохранять активность при относительно высоких температурах в течение от 15 минут до 48 часов или больше. Например, термостабильные ферменты могут сохранять активность при относительно высоких температурах в течение 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 24, 36, 42 или 48 часов.
Согласно некоторым вариантам осуществления термостабильные ферменты (например, киназы или полимеразы) могут сохранять активность (способность катализировать реакцию) при температуре от 32°C до 0°C или ниже. Согласно некоторым вариантам осуществления термостабильные ферменты сохраняют активность при температуре 32-5°C, 32-10°C, 32-20°C, 32-25°C, 32-30°C, 30-0°C, 25-0°C, 20-0°C, 10-0°C или 5-0°C. Например, термостабильные ферменты могут сохранять активность при температуре 32°C, 31°C, 30°C, 29°C, 28°C, 27°C, 26°C, 25°C, 24°C, 23°C, 22°C, 21°C, 20°C, 19°C, 18°C, 17°C, 16°C, 15°C, 14°C, 13°C, 12°C, 10°C, 9°C, 8°C, 7°C, 6°C, 5°C, 4°C, 3°C, 2°C, 1°C или 0°C. Термостабильные ферменты могут сохранять активность при относительно низких температурах в течение от 15 минут до 48 часов или больше. Например, термостабильные ферменты могут сохранять активность при относительно низких температурах в течение 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 24, 36, 42 или 48 часов.
Неограничивающие примеры термостабильных НМФ-киназ приведены в таблицах 4A - 4D. Другие термостабильные киназы включают в себя термостабильные нуклеозиддифосфаткиназы (см., например, таблицу 5), термостабильные пируваткиназы и термостабильные полифосфаткиназы (см., например, таблицу 2). Другие термостабильные киназы охватываются настоящим раскрытием.
Неограничивающие примеры РНК-полимераз приведены в таблице 6. Другие РНК-полимеразы, в том числе термостабильные РНК-полимеразы, охватываются настоящим раскрытием.
Термостабильные РНК-полимеразы могут быть получены с помощью модификации ферментов дикого типа. Такие модификации (например, мутации) известны. Например, вариантные термостабильные T7 РНК-полимеразы могут включать в себя одну или несколько из следующих точковых мутаций: V426L, A702V, V795I, S430P, F849I, S633P, F880Y, C510R и S767G (европейские патентные документы №№ EP2377928 и EP1261696A1, каждый из которых включен в настоящий документ посредством ссылки). Согласно некоторым вариантам осуществления вариантная термостабильная T7 РНК-полимераза включает в себя мутации V426L, A702V и V795I. Согласно некоторым вариантам осуществления вариантная термостабильная T7 РНК-полимераза включает в себя мутации S430P, F849I, S633P и F880Y. Согласно некоторым вариантам осуществления вариантная термостабильная T7 РНК-полимераза включает в себя мутации F880Y, S430P, F849I, S633P, C510R и S767G. Согласно некоторым вариантам осуществления вариантная термостабильная T7 РНК-полимераза включает в себя мутации Y639V, H784G, E593G и V685A. Согласно некоторым вариантам осуществления вариантная термостабильная T7 РНК-полимераза включает в себя мутации S430P, N433T, S633P, F849I и F880Y. Другие вариантные и рекомбинантные термостабильные полимеразы охватываются настоящим раскрытием.
Согласно некоторым вариантам осуществления термостабильную T7 полимеразу используют для получения представляющей интерес РНК. Например, термостабильная T7 полимераза (например, инкубированная при температуре 37-60°C), имеющая концентрацию 0,1-5% суммарного белка, может быть использована для синтеза представляющей интерес РНК при скорости более чем 1 г/л/час (или, например, 1 г/л/час - 20 г/л/час).
Следует учитывать, что хотя во многих вариантах осуществления настоящего раскрытия описывается применение термостабильных полимераз/ферментов, могут быть использованы другие ферменты/полимеразы. Согласно некоторым вариантам осуществления полимераза может быть добавлена извне в инактивированные нагреванием клеточные лизаты, например, чтобы компенсировать любое снижение или потерю активности термостабильного фермента(ов).
Слитые ферменты
Любые из представленных в настоящем документе ферментов пути (например, нуклеаз, киназ, полимераз и т.д.) могут представлять собой отдельные ферменты, ферменты с несколькими активностями или слитые ферменты. Слитый фермент может быть создан путем соединения двух или более генов или генных сегментов, которые кодируют отдельные белки. Трансляция такого слитого гена дает один или несколько полипептидов с функциональными свойствами, полученными от каждого из оригинальных белков, например, слитого белка, который действует как нуклеаза, действует как киназа и/или действует как полимераза. Другие ферменты также могут быть экспрессированы в виде слитого белка.
Некоторые ферменты, которые существуют в природе, являются многофункциональными (например, ЦМФ-УМФ-киназы). Таким образом, термин «фермент» охватывает «ферментативные активности» независимо от того, как они обеспечиваются.
Считают, что слитый фермент «действует как нуклеаза», если фермент демонстрирует нуклеазную активность (расщепляет или деполимеризует нуклеиновую кислоту; например, РНКаза R). Считают, что слитый фермент «действует как киназа», если фермент демонстрирует киназную активность (катализирует перенос фосфатной группы от одной молекулы к другой молекуле; например, полифосфаткиназа). Считают, что слитый фермент «действует как полимераза», если фермент демонстрирует полимеразную активность (собирает нуклеотиды с получением нуклеиновых кислот; например, РНК-полимераза).
Источники энергии
Существует несколько источников энергии и фосфата, которые могут быть использованы, как представлено в настоящем документе, для получения НТФ и/или РНК. Неограничивающие примеры источников фосфата включают в себя НТФ (например, АТФ, ГТФ, УТФ, ЦТФ), полифосфат (например, гексаметафосфат) и пирофосфат (PPi). Согласно некоторым вариантам осуществления НТФ, синтезируемый химическим путем, являющийся продуктом ферментации или экстрагируемый из природного источника, включают в реакционную смесь для получения РНК. Согласно некоторым вариантам осуществления полифосфат и полифосфаткиназу включают в реакционную смесь для получения НТФ и/или РНК. Согласно некоторым вариантам осуществления ацетат, АДФ, пирофосфат и по меньшей мере две ацетаткиназы (например, ацетаткиназу (дифосфат) EC 2.7.2.12 и ацетаткиназу (фосфорилирующую) EC 7.2.1) включают в реакционную смесь для получения НТФ и/или РНК. Согласно некоторым вариантам осуществления цитрат, AMФ, пирофосфат, цитратлиазу (комплекс цитратлиазы), фосфоенолпируваткарбоксикиназу (PEPCK) или фосфоенолпируваткарбоксилазу (PEPC) и пируватфосфатдикиназу (PPDK) включают в реакционную смесь для получения НТФ и/или РНК. Согласно некоторым вариантам осуществления сульфит, AMФ, пирофосфат, аденилилсульфатредуктазу и сульфатаденилилтрансферазу включают в реакционную смесь для получения НТФ и/или РНК. Другие источники энергии также охватываются настоящим раскрытием.
Согласно некоторым вариантам осуществления источником энергии является АТФ, полученный из пирофосфата посредством циклического фосфорилирования ацетата, из пирофосфата и цитрата или из пирофосфата и сульфита. Способы получения АТФ упомянутыми выше путями описываются в настоящем документе. Краткое описание путей получения АТФ и ферментов пути представлены в приведенной ниже таблице 7.
Таблица 7. Краткое описание иллюстративных путей получения АТФ и ферментов
Получение АТФ из пирофосфата и АДФ посредством цикла фосфорилирования/дефосфорилирования ацетата
Согласно некоторым аспектам настоящее раскрытие относится к применению способов получения АТФ из пирофосфата (донора высокоэнергетического фосфата) и АДФ (элементарного акцептора энергии/фосфата) посредством цикла фосфорилирования/дефосфорилирования ацетата (см., например, фиг. 14). Первая ацетаткиназа (AcK1; EC 2.7.2.12) фосфорилирует ацетат с использованием неорганического пирофосфата (PPi), что дает ацетил-фосфат и неорганический фосфат (Pi). Затем ацетил-фосфат дефосфорилируется второй ацетаткиназой (AcK2; EC 2.7.2.1), которая переносит группу высокоэнергетического фосфата из ацетил-фосфата к АДФ и дает АТФ и ацетат. Полученный в результате ацетат затем снова подвергается фосфорилированию с помощью AcK1 с завершением тем самым цикла реакции.
Согласно некоторым вариантам осуществления способы получения АТФ из пирофосфата и АДФ предусматривают культивирование клеток, созданных методами генной инженерии для экспрессирования первой ацетаткиназы, второй ацетаткиназы или двух разных ацетаткиназ. Согласно некоторым вариантам осуществления способы предусматривают культивирование клеток, созданных методами генной инженерии для экспрессирования первой ацетаткиназы и второй ацетаткиназы. Согласно некоторым вариантам осуществления первая ацетаткиназа и вторая ацетаткиназа экспрессируются в виде одного слитого (химерного) белка.
Согласно некоторым вариантам осуществления по меньшей мере один из ферментов является термостабильным ферментом. Согласно некоторым вариантам осуществления по меньшей мере два из ферментов являются термостабильными ферментами. Согласно некоторым вариантам осуществления все ферменты являются термостабильными ферментами. Таким образом, согласно некоторым вариантам осуществления способы предусматривают культивирование клеток, созданных методами генной инженерии для экспрессирования термостабильной ацетаткиназы. Согласно другим вариантам осуществления способы предусматривают культивирование клеток, созданных методами генной инженерии для экспрессирования первой термостабильной ацетаткиназы и второй термостабильной ацетаткиназы.
Согласно некоторым вариантам осуществления способы получения АТФ из пирофосфата посредством циклического фосфорилирования ацетата предусматривают лизирование (например, термический, осмотический, механический (например, обработка ультразвуком), химический или ферментативный лизис) культивируемых клеток с получением по меньшей мере одного (например, по меньшей мере двух) клеточного лизата. Следует учитывать, что несколько клеточных лизатов (и, таким образом, несколько клеточных популяций, например, из одного и того же организма (например, бактерий) или из разных организмов (например, бактерий, дрожжей и/или растения) могут быть использованы в ферментативной реакции, как представлено в настоящем документе. Например, одна клеточная популяция может быть создана методами генной инженерии для экспрессирования первой ацетаткиназы пути получения АТФ, тогда как другая клеточная популяция может быть создана методами генной инженерии для экспрессирования второй ацетаткиназы пути получения АТФ. Таким образом, согласно некоторым вариантам осуществления способы предусматривают культивирование популяции клеток, созданных методами генной инженерии для экспрессирования ацетаткиназы, и/или культивирование клеточной популяции, созданной методами генной инженерии для экспрессирования по меньшей мере одной дополнительной ацетаткиназы. После лизиса клеток клеточные лизаты объединяют, так что ферменты присутствуют в одном клеточном лизате/реакционной смеси.
Согласно некоторым вариантам осуществления способы получения АТФ из пирофосфата посредством циклического фосфорилирования ацетата, кроме того, предусматривают нагревание клеточного(ых) лизата(ов) (или смеси клеточных лизатов) до температуры, которая инактивирует нативную ферментативную активность, но не инактивирует никакие из термостабильных ферментов пути получения АТФ, с получением инактивированного нагреванием лизата. Клеточный(ые) лизат(ы) согласно некоторым вариантам осуществления нагревают до температуры по меньшей мере 50°C. Например, клеточный(ые) лизат(ы) можно нагревать до температуры по меньшей мере 55°C, 60°C, 65°C, 70°C, 75°C, 80°C, 85°C или 90°C. Нативный фермент (или другой нетермостабильный фермент) считают неактивным согласно некоторым вариантам осуществления, если уровень его активности снижается по меньшей мере на 50%. Согласно некоторым вариантам осуществления нативный фермент (или другой нетермостабильный фермент) считают неактивным, если уровень его активности снижается по меньшей мере на 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100%.
Клеточный(ые) лизат(ы) можно нагревать на протяжении периода времени, достаточного для инактивации нативных ферментов (или других нетермостабильных ферментов) клетки. Например, клеточный(ые) лизат(ы) можно нагревать в течение по меньшей мере 2, 3, 4 или по меньшей мере 5 минут. Согласно некоторым вариантам осуществления клеточный(ые) лизат(ы) нагревают в течение более чем 5 минут. Согласно некоторым вариантам осуществления клеточный(ые) лизат(ы) нагревают в течение более чем 15 минут. Согласно некоторым вариантам осуществления клеточный(ые) лизат(ы) нагревают в течение менее чем 2 минут. Согласно некоторым вариантам осуществления клеточный(ые) лизат(ы) нагревают на протяжении периода времени, достаточного для снижения активности нативных ферментов (или других нетермостабильных ферментов) по меньшей мере на 50% (например, по меньшей мере на 60%, 70%, 80% или 90%).
После инактивации нагреванием согласно некоторым вариантам осуществления по меньшей мере один (например, по меньшей мере два или по меньшей мере три) очищенный фермент может быть добавлен в клеточный лизат/реакционную смесь. Таким образом, реакционная смесь согласно некоторым вариантам осуществления может включать в себя комбинацию ферментов, присутствующих в клеточном лизате (экспрессируемых созданной(ыми) методами генной инженерии клеткой-хозяином(клетками-хозяевами)), и по меньшей мере один очищенный фермент. По меньшей мере один очищенной фермент может представлять собой первую ацетаткиназу и/или вторую ацетаткиназу. Согласно некоторым вариантам осуществления клеточный лизат может быть охлажден (например, до 50°C) после стадии инактивации нагреванием перед добавлением очищенного(ых) фермента(ов).
Согласно некоторым вариантам осуществления способы получения АТФ из пирофосфата посредством циклического фосфорилирования ацетата также предусматривают инкубирование инактивированного(ых) нагреванием лизата(ов) в присутствии ацетата, аденозиндифосфата (АДФ) и неорганического фосфата с получением АТФ. Неорганический фосфат может представлять собой, например, пирофосфат. Могут быть использованы другие неорганические фосфаты и/или ортофосфатные полимеры, включающие в себя без ограничения триполифосфат, тетраполифосфат, пентаполифосфат, гексаметафосфат и их смеси.
Также настоящий документ охватывает клетки и клеточные лизаты, используемые для получения АТФ из пирофосфата посредством циклического фосфорилирования ацетата. Таким образом, созданная методами генной инженерии клетка (например, бактериальная клетка, дрожжевая клетка и/или растительная клетка) или клеточный(ые) лизат(ы) в соответствии с настоящим раскрытием могут включать в себя по меньшей мере одну (например, по меньшей мере две) ацетаткиназу. Согласно некоторым вариантам осуществления созданная методами генной инженерии клетка (например, бактериальная клетка, дрожжевая клетка и/или растительная клетка) или клеточный(ые) лизат(ы) в соответствии с настоящим раскрытием включают в себя по меньшей мере одну (например, по меньшей мере две) термостабильную ацетаткиназу.
Таблица 8. Иллюстративные ферменты ацетаткиназы
Получение АТФ из пирофосфата, AMФ и цитрата
Согласно некоторым аспектам настоящее раскрытие относится к применению способов получения АТФ из пирофосфата, AMФ и цитрата (см., например, фиг. 15A - 15B). Трехстадийный ферментативный путь показан на фиг. 15A. На первой стадии цитратлиаза превращает цитрат в ацетат и оксалоацетат. На второй стадии фосфоенолпируваткарбоксикиназа (PEPCK) превращает пирофосфат и оксалоацетат, образованный на первой стадии, в фосфоенолпируват (PEP), диоксид углерода (CO2) и неорганический фосфат (Pi). На третьей стадии пируватфосфатдикиназа (PPDK) превращает неорганический пирофосфат (PPi), AMФ и PEP, образованный на второй стадии, в пируват, Pi и АТФ. В объединенной химической реакции используется один моль цитрата, один моль AMФ и два моля PPi с полученияем одного моля ацетата, одного моля пирувата, одного моля CO2, двух молей Pi и одного моля АТФ (фиг. 15B). В качестве альтернативы, фосфоенолпируваткарбоксилаза (PEPC) может быть использована для катализа карбоксилирования PEP до оксалоацетата, которое может быть обратимым при определенных условиях.
Эти способы согласно некоторым вариантам осуществления предусматривают культивирование клеток, созданных методами генной инженерии для экспрессирования цитратлиазы, PEPCK (или по меньшей мере одной PEPC), PPDK или комбинации по меньшей мере двух или по меньшей мере трех из упомянутых выше ферментов. Согласно некоторым вариантам осуществления цитратлиаза и PEPCK (или PEPC), PEPCK (или PEPC) и PPDK или цитратлиаза и PPDK экспрессируются в виде одного слитого (химерного) белка.
Согласно некоторым вариантам осуществления по меньшей мере один из ферментов является термостабильным ферментом. Согласно некоторым вариантам осуществления по меньшей мере два или по меньшей мере три из ферментов являются термостабильными ферментами. Согласно некоторым вариантам осуществления все из ферментов являются термостабильными ферментами. Таким образом, согласно некоторым вариантам осуществления способы предусматривают культивирование клеток, созданных методами генной инженерии для экспрессирования термостабильной цитратлиазы, термостабильной PEPCK, PPDK или комбинации по меньшей мере двух или по меньшей мере трех их упомянутых выше термостабильных ферментов.
Согласно некоторым вариантам осуществления способы получения АТФ из цитрата предусматривают лизирование (например, термический, осмотический, механический (например, обработка ультразвуком), химический или ферментативный лизис) культивируемых клеток с получением по меньшей мере одного (например, по меньшей мере двух или трех) клеточного лизата. Следует учитывать, что несколько клеточных лизатов (и, таким образом, несколько клеточных популяций, например, из одного и того же организма (например, бактерий) или из разных организмов (например, бактерий, дрожжей и/или растения) могут быть использованы в ферментативной реакции, как представлено в настоящем документе. Например, одна клеточная популяция может быть создана методами генной инженерии для экспрессирования одного или нескольких ферментов пути получения АТФ, тогда как другая клеточная популяция (или несколько других клеточных популяций) может быть создана методами генной инженерии для экспрессирования другого (по меньшей мере одной другого) фермента пути получения АТФ. Таким образом, согласно некоторым вариантам осуществления способы предусматривают культивирование популяции клеток, созданных методами генной инженерии для экспрессирования цитратлиазы, культивирование клеточной популяции, созданной методами генной инженерии для экспрессирования PEPCK (термостабильной PEPCK), и/или культивирование клеточной популяции, созданной методами генной инженерии для экспрессирования PPDK. После лизиса клеток клеточные лизаты объединяют, так что ферменты присутствуют в одном клеточном лизате/реакционной смеси.
Согласно некоторым вариантам осуществления способы получения АТФ из цитрата, кроме того, предусматривают нагревание клеточного(ых) лизата(ов) (или смеси клеточных лизатов) до температуры, которая инактивирует нативную ферментативную активность, но не инактивирует никакой из термостабильных ферментов пути получения АТФ, с получением инактивированного нагреванием лизата. Клеточный(ые) лизат(ы) согласно некоторым вариантам осуществления нагревают до температуры по меньшей мере 50°C. Например, клеточный(ые) лизат(ы) можно нагревать до температуры по меньшей мере 55°C, 60°C, 65°C, 70°C, 75°C, 80°C, 85°C или 90°C. Нативный фермент (или другой нетермостабильный фермент) считают неактивным согласно некоторым вариантам осуществления, если уровень его активности снижается по меньшей мере на 50%. Согласно некоторым вариантам осуществления нативный фермент (или другой нетермостабильный фермент) считают неактивным, если уровень его активности снижается по меньшей мере на 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100%.
Клеточный(ые) лизат(ы) можно нагревать на протяжении периода времени, достаточного для инактивации нативных ферментов (или других нетермостабильных ферментов) клетки. Например, клеточный(ые) лизат(ы) можно нагревать в течение по меньшей мере 2, 3, 4 или по меньшей мере 5 минут. Согласно некоторым вариантам осуществления клеточный(ые) лизат(ы) нагревают в течение более чем 5 минут. Согласно некоторым вариантам осуществления клеточный(ые) лизат(ы) нагревают на протяжении периода времени, достаточного для снижения активности нативных ферментов (или других нетермостабильных ферментов) по меньшей мере на 50% (например, по меньшей мере на 60%, 70%, 80% или 90%).
После инактивации нагреванием согласно некоторым вариантам осуществления по меньшей мере один (например, по меньшей мере два или по меньшей мере три) очищенный фермент может быть добавлен в клеточный лизат/реакционную смесь. Таким образом, реакционная смесь согласно некоторым вариантам осуществления может включать в себя комбинацию ферментов, присутствующих в клеточном лизате (экспрессируемых созданной(ыми) методами генной инженерии клеткой-хозяином(клетками-хозяевами)), и по меньшей мере один очищенный фермент. По меньшей мере один очищенной фермент может быть выбран из группы, состоящей из цитратлиазы, PEPCK (или PEPC) и PPDK. Согласно некоторым вариантам осуществления клеточный лизат может быть охлажден (например, до 50°C) после стадии инактивации нагреванием перед добавлением очищенного(ых) фермента(ов).
Согласно некоторым вариантам осуществления способы получения АТФ из цитрата также предусматривают инкубирование инактивированного(ых) нагреванием лизата(ов) в присутствии цитрата, аденозинмонофосфата (AMФ) и неорганического фосфата с получением АТФ. Неорганический фосфат может представлять собой, например, пирофосфат. Неорганический фосфат может представлять собой, например, пирофосфат. Могут быть использованы другие неорганические фосфаты и/или ортофосфатные полимеры, в том числе без ограничения триполифосфат, тетраполифосфат, пентаполифосфат, гексаметафосфат и их смеси.
Также настоящий документ охватывает клетки и клеточные лизаты, используемые для получения АТФ из цитрата. Таким образом, созданная методами генной инженерии клетка (например, бактериальная клетка, дрожжевая клетка и/или растительная клетка) или клеточный(ые) лизат(ы) в соответствии с настоящим раскрытием могут включать в себя по меньшей мере один (например, по меньшей мере два или по меньшей мере три) фермента, выбранных из группы, состоящей из цитратлиазы, PEPCK (или PEPC) и PPDK. Согласно некоторым вариантам осуществления созданная методами генной инженерии клетка (например, бактериальная клетка, дрожжевая клетка и/или растительная клетка) или клеточный(ые) лизат(ы) в соответствии с настоящим раскрытием включают в себя по меньшей мере один (например, по меньшей мере два или по меньшей мере три) фермент, выбранный из группы, состоящей из термостабильной цитратлиазы, термостабильной PEPCK (или термостабильной PEPC) и термостабильной PPDK.
Таблица 9. Иллюстративное ферменты пути для получения АТФ из пирофосфата и цитрата
Получение АТФ из пирофосфата, AMФ и сульфита
Согласно некоторым аспектам настоящее раскрытие относится к применению способов получения АТФ из пирофосфата, AMФ и сульфита (см., например, фиг. 16). На первой стадии аденилилсульфатредуктаза превращает аденозинмонофосфат (AMФ) в аденозин-5’-фосфосульфат (АФС) с поглощением сульфита. На второй стадии сульфатаденилилтрансфераза катализирует превращение АФС в сульфат с образованием АТФ и поглощением пирофосфата.
Согласно некоторым вариантам осуществления способы получения АТФ из пирофосфата, AMФ и сульфита предусматривают культивирование клеток, созданных методами генной инженерии для экспрессирования аденилилсульфатредуктазы, сульфатаденилилтрансферазы или комбинации аденилилсульфатредуктазы и сульфатаденилилтрансферазы. Согласно некоторым вариантам осуществления аденилилсульфатредуктаза и сульфатаденилилтрансфераза экспрессируются в виде одного слитого (химерного) белка или бифункционального белка.
Согласно некоторым вариантам осуществления могут быть добавлены восстановители, которые служат стоками электронов. Примеры таких восстановителей включают в себя без ограничения следующие: дитиотреитол (DTT), или глутатион, или феррицианид. или дитиоэритритол. или трис-2-карбоксиэтилфосфин гидрохлорид (TCEP). Хотя отдельные ферменты будут различаться по своим предпочтениям в отношении кофакторов, могут быть случаи, когда биологические кофакторы, такие как NAD+, NADP+, NADH или NADPH, могут быть использованы ферментом для поглощения этих электронов. В этих случаях такие кофакторы также могут быть включены.
Согласно некоторым вариантам осуществления по меньшей мере один из ферментов является термостабильным ферментом. Согласно некоторым вариантам осуществления по меньшей мере два из ферментов являются термостабильными ферментами. Согласно некоторым вариантам осуществления все ферменты являются термостабильными ферментами. Таким образом, согласно некоторым вариантам осуществления способы предусматривают культивирование клеток, созданных методами генной инженерии для экспрессирования термостабильной аденилилсульфатредуктазы и термостабильной сульфатаденилилтрансферазы.
Согласно некоторым вариантам осуществления способы получения АТФ из сульфита предусматривают лизирование (например, термический, осмотический, механический (например, обработка ультразвуком), химический или ферментативный лизис) культивируемых клеток с получением по меньшей мере одного (например, 2, 3, 4 или 5) клеточного лизата. Следует учитывать, что несколько клеточных лизатов (и, таким образом, несколько клеточных популяций, например, из одного и того же организма (например, бактерий) или из разных организмов (например, бактерий, дрожжей и/или растения), могут быть использованы в ферментативной реакции, как представлено в настоящем документе. Например, одна клеточная популяция может быть создана методами генной инженерии для экспрессирования аденилилсульфатредуктазы, тогда как другая клеточная популяция (или несколько других клеточных популяций) могут быть созданы методами генной инженерии для экспрессирования сульфатаденилилтрансферазы. Таким образом, согласно некоторым вариантам осуществления способы предусматривают культивирование популяции клеток, созданных методами генной инженерии для экспрессирования аденилилсульфатредуктазы, и/или культивирование клеточной популяции, созданной методами генной инженерии для экспрессирования сульфатаденилилтрансферазы. После лизиса клеток клеточные лизаты объединяют, так что ферменты присутствуют в одном клеточном лизате/реакционной смеси.
Согласно некоторым вариантам осуществления способы получения АТФ из сульфита, кроме того, предусматривают нагревание клеточного(ых) лизата(ов) (или смеси клеточных лизатов) до температуры, которая инактивирует нативную ферментативную активность, но не инактивирует никакие из термостабильных ферментов пути получения АТФ, с получением инактивированного нагреванием лизата. Клеточный(ые) лизат(ы) согласно некоторым вариантам осуществления нагревают до температуры по меньшей мере 50°C. Например, клеточный(ые) лизат(ы) можно нагревать до температуры по меньшей мере 55°C, 60°C, 65°C, 70°C, 75°C, 80°C, 85°C или 90°C. Нативный фермент (или другой нетермостабильный фермент) считают неактивным согласно некоторым вариантам осуществления, если уровень его активности снижается по меньшей мере на 50%. Согласно некоторым вариантам осуществления нативный фермент (или другой нетермостабильный фермент) считают неактивным, если уровень его активности снижается по меньшей мере на 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100%.
Клеточный(ые) лизат(ы) можно нагревать на протяжении периода времени, достаточного для инактивации нативных ферментов (или других нетермостабильных ферментов) клетки. Например, клеточный(ые) лизат(ы) можно нагревать в течение по меньшей мере 2, 3, 4 или по меньшей мере 5 минут. Согласно некоторым вариантам осуществления клеточный(ые) лизат(ы) нагревают в течение более чем 5 минут. Согласно некоторым вариантам осуществления клеточный(ые) лизат(ы) нагревают на протяжении периода времени, достаточного для снижения активности нативных ферментов (или других нетермостабильных ферментов) по меньшей мере на 50% (например, по меньшей мере на 60%, 70%, 80% или 90%).
После инактивации нагреванием согласно некоторым вариантам осуществления по меньшей мере один (например, по меньшей мере два или по меньшей мере три) очищенный фермент может быть добавлен в клеточный лизат/реакционную смесь. Таким образом, реакционная смесь согласно некоторым вариантам осуществления может включать в себя комбинацию клеточного лизата, ферментов, присутствующих в клеточном лизате (экспрессируемых созданной(ыми) методами генной инженерии клеткой-хозяином(клетками-хозяевами)), и по меньшей мере один очищенный фермент. По меньшей мере один очищенной фермент может представлять собой первую ацетаткиназу и/или вторую ацетаткиназу. Согласно некоторым вариантам осуществления клеточный лизат может быть охлажден (например, до 50°C) после стадии инактивации нагреванием перед добавлением очищенного(ых) фермента(ов).
Согласно некоторым вариантам осуществления способы получения АТФ из сульфита также предусматривают инкубирование инактивированного(ых) нагреванием лизата(ов) в присутствии сульфита, аденозинмонофосфата (AMФ) и неорганического фосфата с получением АТФ. Неорганический фосфат может представлять собой, например, пирофосфат. Могут быть использованы другие неорганические фосфаты и/или ортофосфатные полимеры, включающие в себя без ограничения триполифосфат, тетраполифосфат, пентаполифосфат, гексаметафосфат и их смеси.
Также настоящий документ охватывает клетки и клеточные лизаты, используемые для получения АТФ. Таким образом, созданная методами генной инженерии клетка (например, бактериальная клетка, дрожжевая клетка и/или растительная клетка) или клеточный(ые) лизат(ы) в соответствии с настоящим раскрытием могут включать в себя по меньшей мере одну (например, по меньшей мере две) аденилилсульфатредуктазу и/или по меньшей мере одну сульфатаденилилтрансферазу. Согласно некоторым вариантам осуществления созданная методами генной инженерии клетка (например, бактериальная клетка, дрожжевая клетка и/или растительная клетка) или клеточный(ые) лизат(ы) в соответствии с настоящим раскрытием включают в себя по меньшей мере одну (например, по меньшей мере две, по меньшей мере три или по меньшей мере четыре) термостабильную аденилилсульфатредуктазу и/или по меньшей мере одну термостабильную сульфатаденилилтрансферазу.
Таблица 10. Иллюстративные ферменты пути для получения АТФ из пирофосфата, AMФ и сульфита
Деполимеризация клеточной РНК
Согласно некоторым вариантам осуществления клеточная РНК служит в качестве субстрата для получения НТФ и/или РНК. Деполимеризация (разложение) клеточной РНК дает в результате пул, содержащий нуклеозиддифосфаты (НДФ) или 5’-нуклеозидмонофосфаты (5’-НМФ), в зависимости от ферментов, используемых для деполимеризации.
Клеточную РНК согласно некоторым вариантам осуществления деполимеризуют в НДФ с использованием, например, полинуклеотидфосфорилазы (ПНФазы) (см., например, таблицу 1). Согласно некоторым вариантам осуществления концентрация ПНФазы, используемой в реакционной смеси, составляет 0,001-10 мг/мл (например, 0,001, 0,01, 0,1, 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1,0, 5 или 10 мг/мл). Согласно некоторым вариантам осуществления концентрация ПНФазы, используемой в реакционной смеси, составляет 0,5-5 мг/мл. Согласно некоторым вариантам осуществления концентрация ПНФазы, используемой в реакционной смеси, составляет 5 мг/мл. Согласно некоторым вариантам осуществления концентрация ПНФазы, используемой в реакционной смеси, составляет более чем 10 мг/мл.
Клеточную РНК согласно другим вариантам осуществления деполимеризуют в НМФ с использованием, например, нуклеазы (например, РНКазы R или P1 нуклеазы) (см., например, таблицу 1). В зависимости от фермента ферментативная деполимеризация РНК может давать 3’-НМФ, 5’-НМФ или комбинацию 3’-НМФ и 5’-НМФ. Поскольку невозможно полимеризовать 3’-НTФ (превратить из 3’-НДФ, которые превращаются из 3’-НМФ), ферменты (например, РНКаза R и/или P1 нуклеаза), которые дают 5’-НМФ (которые затем превращаются в 5’-НДФ, а затем в 5’-НTФ), являются предпочтительными. Согласно некоторым вариантам осуществления концентрация нуклеазы (например, РНКазы R и/или P1 нуклеазы), используемой в реакционной смеси, составляет 0,001-10 мг/мл (например, 0,001, 0,01, 0,1, 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9,1,0, 5 или 10 мг/мл). Согласно некоторым вариантам осуществления концентрация нуклеазы в реакционной смеси составляет 0,0,5-5 мг/мл. Согласно некоторым вариантам осуществления концентрация нуклеазы в реакционной смеси составляет 5 мг/мл. Согласно некоторым вариантам осуществления концентрация нуклеазы в реакционной смеси составляет более чем 10 мг/мл.
ПНФаза и/или РНКаза согласно некоторым вариантам осуществления получается из клеточного лизата или является компонентом клеточного лизата клеток, которые экспрессируют ПНФазу и/или РНКазу.
Количество клеточной РНК, необходимое для синтеза представляющего интерес РНК-продукта, может варьировать в зависимости от, например, желаемой длины и выхода РНК-продукта, а также нуклеотидной композиции РНК-продукта относительно нуклеотидной композиции исходного материала клеточной РНК. Как правило, для бактериальной клетки или дрожжевой клетки, например, содержание клеточной РНК варьирует от 5 до 50% суммарной массы клетки. Процент суммарной массы клетки может быть вычислен, например, с использованием следующего уравнения: (килограмм (кг) РНК/килограмм сухой массы клетки) × 100%.
Условия, подходящие для получения НМФ, и условия, подходящие для получения НДФ, известны в уровне техники или могут быть определены рядовым специалистом в данной области с учетом, например, оптимальных условий для активности нуклеазы (например, РНКазы), в том числе pH (например, pH 3-8), температура (например, от 15°C до 70°C), промежуток времени (например, от 5 минут до 72 часов) и концентрация соли (например, хлорид натрия, хлорид калия, ацетат натрия, ацетат калия при концентрации от 5 мМ до 1 M) реакционной смеси, а также каких-либо экзогенных кофакторов. Согласно некоторым вариантам осуществления буфер добавляют в клеточный лизат, например, для достижения определенного значения pH и/или концентрации соли. Примеры буферов включают в себя без ограничения фосфатный буфер, трисбуфер, MOPS буфер, HEPES буфер, цитратный буфер, ацетатный буфер, малатный буфер, MES буфер, гистидиновый буфер, PIPES буфер, бис-трисбуфер и этаноламиновый буфер.
Согласно некоторым вариантам осуществления реакционную смесь в ходе реакции деполимеризации РНК инкубируют в течение 24 часов при температуре 37°C. Согласно некоторым вариантам осуществления реакционную смесь в ходе реакции деполимеризации РНК инкубируют в течение 5-30 минут при температуре 37°C. Согласно некоторым вариантам осуществления реакционная смесь в ходе реакции деполимеризации РНК характеризуется pH 7,0 и инкубируется в течение 15 минут при температуре 37°C. Согласно некоторым вариантам осуществления реакционная смесь в ходе реакции деполимеризации РНК может быть инкубирована в условиях, которые обеспечивают более чем 65% превращение РНК в НДФ или РНК в 5’-НМФ. Согласно некоторым вариантам осуществления РНК превращается в НДФ или 5’-НМФ при скорости (или по меньшей мере) 50 мМ/час, 100 мМ/час или 200 мМ/час. Согласно другим вариантам осуществления реакционную смесь в ходе реакции деполимеризации РНК инкубируют при более высокой температуре (например, 50°C - 70°C), как в примере 5.
Полимеризация РНК-продукта
Согласно некоторым вариантам осуществления НТФ, либо полученный способом, представленным в настоящем документе, либо приобретенный из коммерческих источников, используют в пути биосинтеза для получения представляющего интерес РНК-продукта. ДНК, сконструированная для кодирования РНК-продукта, служит в качестве матрицы для синтеза РНК. ДНК-матрица может быть создана методами генной инженерии в некоторых случаях с наличием транскрипционного промотора, который селективно управляет транскрипцией представляющей интерес РНК. Для полимеризации РНК требуется НТФ, ДНК-матрица, содержащая транскрипционный промотор и полимераза (например, РНК-полимераза), специфическая по отношению к транскрипционному промотору. Как правило, полимераза для применения, представленного в настоящем документе, представляет собой полимеразу с одной субъединицей, является высокоселективной в отношении когнатных ей транскрипционных промоторов, характеризуется высокой точностью и является высокоэффективной.
Согласно некоторым вариантам осуществления концентрация ДНК-матрицы в реакционной смеси составляет 0,001-10 мкг/мкл. Согласно некоторым вариантам осуществления концентрация ДНК-матрицы в реакционной смеси составляет 0,001 мкг/мкл, 0,05 мкг/мкл, 0,1 мкг/мкл, 0,5 мкг/мкл, 1,0 мкг/мкл, 5 мкг/мкл или 10 мкг/мкл.
Условия, подходящие для получения РНК, известны в уровне техники или могут быть определены рядовым специалистом в данной области с учетом, например, оптимальных условий для активности полимеразы (например, T7 РНК-полимеразы), в том числе pH (например, pH 3-8), температура (например, от 15°C до 70°C), промежуток времени (например, от 5 минут до 72 часов) и концентрация соли (например, хлорид натрия, хлорид калия, ацетат натрия, ацетат калия при концентрации от 5 мМ до 1 M) реакционной смеси, а также каких-либо экзогенных кофакторов. Согласно некоторым вариантам осуществления буфер добавляют в клеточный лизат, например, для достижения определенного значения pH и/или концентрации соли. Примеры буферов включают в себя без ограничения фосфатный буфер, трисбуфер, MOPS буфер, HEPES буфер, цитратный буфер, ацетатный буфер, малатный буфер, MES буфер, гистидиновый буфер, PIPES буфер, бис-трисбуфер и этаноламиновый буфер.
Согласно некоторым вариантам осуществления реакционную смесь в ходе реакции деполимеризации РНК инкубируют в течение 0,5-24 часов при температуре 37°C. Согласно некоторым вариантам осуществления реакционную смесь в ходе реакции деполимеризации РНК инкубируют в течение 0,5-24 часов при температуре 50°C.
Клетки и клеточные лизаты
Клетки в соответствии с настоящим раскрытием согласно некоторым вариантам осуществления экспрессируют клеточную РНК, ферменты, которые деполимеризуют РНК (например, РНКазы), ферменты пути (например, рекомбинантные ферменты, такие как полифосфаткиназа) и/или полимеразы (например, РНК-полимеразы). Согласно некоторым вариантам осуществления созданные методами генной инженерии клетки включают в себя ДНК-матрицу, содержащую промотор и необязательно транскрипционный терминатор, функционально связанный с нуклеотидной последовательностью, кодирующей представляющая интерес РНК-продукт.
Согласно некоторым вариантам осуществления клетки являются созданными методами генной инженерии клетками. Созданные методами генной инженерии клетки представляют собой клетки, которые содержат созданную методами генной инженерии (например, рекомбинантную или синтетическую) нуклеиновую кислоту или являются иным образом модифицированными, так что они структурально и/или функционально отличаются от их встречающихся в природе аналогов. Таким образом, клетку, которая содержит созданную методами генной инженерии нуклеиновую кислоту, считают «созданной методами генной инженерии клеткой».
Клетка «экспрессирует» продукт, если продукт, кодируемый нуклеиновой кислотой (например, созданной методами генной инженерии нуклеиновой кислотой), продуцируется в клетке. В уровне техники известно, что экспрессия генов относится к процессу, с помощью которого генетические инструкции в форме нуклеиновой кислоты используются для синтеза продукта, такого как белок (например, фермент).
Клетки могут быть прокариотическими клетками или эукариотическими клетками. Согласно некоторым вариантам осуществления клетки являются бактериальными клетками, дрожжевыми клетками, клетками насекомых, клетками млекопитающих, клетками растений или другими типами клеток.
Бактериальные клетки в соответствии с настоящим раскрытием включают в себя без ограничения Escherichia spp., Streptomyces spp., Zymomonas spp., Acetobacter spp., Citrobacter spp., Synechocystis spp., Rhizobium spp., Clostridium spp., Corynebacterium spp., Streptococcus spp., Xanthomonas spp., Lactobacillus spp., Lactococcus spp., Bacillus spp., Alcaligenes spp., Pseudomonas spp., Aeromonas spp., Azotobacter spp., Comamonas spp., Mycobacterium spp., Rhodococcus spp., Gluconobacter spp., Ralstonia spp., Acidithiobacillus spp., Microlunatus spp., Geobacter spp., Geobacillus spp., Arthrobacter spp., Flavobacterium spp., Serratia spp., Saccharopolyspora spp., Thermus spp., Stenotrophomonas spp., Chromobacterium spp., Sinorhizobium spp., Saccharopolyspora spp., Agrobacterium spp., Pantoea spp. и Vibrio natriegens.
Дрожжевые клетки в соответствии с настоящим раскрытием включают в себя без ограничения созданные методами генной инженерии Saccharomyces spp., Schizosaccharomyces, Hansenula, Candida, Kluyveromyces, Yarrowia и Pichia.
Согласно некоторым вариантам осуществления клетки в соответствии с настоящим раскрытием представляют собой клетки Escherichia coli, клетки Bacillus subtilis, клетки Pseudomonas putida, клетки Saccharomyces cerevisiae или клетки Lactobacillus brevis. Согласно некоторым вариантам осуществления клетки в соответствии с настоящим раскрытием представляют собой клетки Escherichia coli.
Как правило, клетки культивируют. Культивирование представляет собой процесс, с помощью которого клетки выращивают в контролируемых условиях, как правило, вне их природного окружения. Например, клетки, такие как бактериальные клетки, могут расти в виде клеточной суспензия в жидком питательном бульоне, также называемом жидкой культуральной средой.
Примеры традиционно используемых бактериальных ростовых сред для Escherichia coli включают в себя без ограничения LB (лизогенный бульон) бульон Миллера (1% NaCl): 1% пептона, 0,5% дрожжевого экстракта и 1% NaCl; LB (лизогенный бульон) бульон Леннокса (0,5% NaCl): 1% пептона, 0,5% дрожжевого экстракта и 0,5% NaCl; среду SOB (супероптимальный бульон): 2% пептона, 0,5% дрожжевого экстракта, 10 мМ NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4; среду SOC (супероптимальный бульон с катаболическим репрессором): SOB + 20 мМ глюкоза; 2× бульон YT (2× дрожжевой экстракт и триптон): 1,6% пептона, 1% дрожжевого экстракта и 0,5% NaCl; среду TB (Terrific Broth): 1,2% пептона, 2,4% дрожжевого экстракта, 72 мМ K2HPO4, 17 мМ KH2PO4 и 0,4% глицерина; среду SB (супербульон): 3,2% пептона, 2% дрожжевого экстракта и 0,5% NaCl, и/или среду Korz (Korz, DJ et al. 1995).
Примеры высокоплотных бактериальных ростовых сред для Escherichia coli включают в себя без ограничения среду DNAGro™, среду ProGro™, среду AutoX™, среду DetoX™, среду InduX™ и среду SecPro™.
Согласно некоторым вариантам осуществления клетки культивируют в условиях, которые приводят к экспрессии ферментов или нуклеиновых кислот. Такие условия культивирования могут зависеть от конкретного продукта, подлежащего экспрессированию, и желаемого количества продукта.
Согласно некоторым вариантам осуществления клетки культивируют при температуре от 30°C до 40°C. Например, созданные методами генной инженерии клетки можно культивировать при температуре 30°C, 31°C, 32°C, 33°C, 34°C, 35°C, 36°C, 37°C, 38°C, 39°C или 40°C. Как правило, клетки, такие как созданные методами генной инженерии клетки E. coli, культивируют при температуре 37°C.
Согласно некоторым вариантам осуществления клетки культивируют на протяжении периода времени от 12 часов до 72 часов или больше. Например, созданные методами генной инженерии клетки можно культивировать на протяжении периода времени 12, 18, 24, 30, 36, 42, 48, 54, 60, 66 или 72 часов. Как правило, клетки, такие как созданные методами генной инженерии бактериальные клетки, культивируют на протяжении периода времени от 12 до 24 часов. Согласно некоторым вариантам осуществления клетки культивируют в течение 12-24 часов при температуре 37°C.
Согласно некоторым вариантам осуществления клетки культивируют (например, в жидкой клеточной культуральной среде) до оптической плотности, измеряемой при длине волны 600 нм (OD600), от 5 до 200. Согласно некоторым вариантам осуществления клетки культивируют до OD600 5, 10, 15, 20, 25, 50, 75, 100, 150 или 200.
Согласно некоторым вариантам осуществления клетки культивируют до плотности от 1 × 108 (OD600 < 1) до 2 × 1011 (OD ~ 200) жизнеспособных клеток/мл клеточной культуральной среды. Согласно некоторым вариантам осуществления клеток культивируют до плотности 1 × 108, 2 × 108, 3 × 108, 4 × 108, 5 × 108, 6 × 108, 7 × 108, 8 × 108, 9 × 108, 1 × 109, 2 × 109, 3 × 109, 4 × 109, 5 × 109, 6 × 109, 7 × 109, 8 × 109, 9 × 109, 1 × 1010, 2 × 1010, 3 × 1010, 4 × 1010, 5 × 1010, 6 × 1010, 7 × 1010, 8 × 1010, 9 × 1010, 1 × 1011 или 2 × 1011 жизнеспособных клеток/мл. (коэффициент перевода: OD 1 = 8 × 108 клеток/мл).
Согласно некоторым вариантам осуществления клетки культивируют в биореакторе. Под биореактором понимают просто контейнер, в котором культивируют клетки, такой как колба для культивирования, чашка или пакет, который может быть разового использования (одноразовым), автоклавируемым или стерилизуемым. Биореактор может быть изготовлен из стекла, или он может быть полимерным, или он может быть изготовлен из других материалов.
Примеры биореакторов включают в себя без ограничения биореакторы в виде резервуара с перемешиванием (например, с полным перемешиванием) и трубчатые (например, поршневые проточные) биореакторы, биореакторы с аэрирующим устройством, мембранные резервуары с перемешиванием, резервуары с перемешиванием с вращающимся фильтром, вибросмесители, реакторы с псевдоожиженным слоем и мембранные биореакторы. Режим работы биореактора может представлять собой периодические или непрерывные процессы и будет зависеть от созданных методами генной инженерии клеток, подлежащих культивированию. Биореактор является непрерывным, если потоки сырья и продукта непрерывно подают и извлекают из системы. Периодический биореактор может иметь непрерывный рециркуляционный поток, но без непрерывной подачи питательных веществ или сбора продукта. Для культур с прерывистым сбором и с подпиткой (или с подкормкой) клетки инокулируют при более низкой плотности жизнеспособных клеток в среде, которая по составу аналогична среде партии. Клеткам позволяют расти в геометрической прогрессии без каких-либо внешних манипуляций, пока питательные вещества не истощатся, и клетки не достигнут стационарной фазы роста. В этот момент для периодического процесса периодического сбора часть клеток и продукта может быть собрана, и удаленная культуральная среда пополняется свежей средой. Этот процесс может повторяться несколько раз. Для получения рекомбинантных белков и антител можно использовать периодический процесс с подпиткой. В то время как клетки растут в геометрической прогрессии, а питательные вещества истощаются, добавляют концентрированную среду подкормки (например, концентрированную в 10-15 раз основную среду) либо непрерывно, либо периодически для подачи дополнительных питательных веществ, что обеспечивает дальнейшее увеличение концентрации клеток и продолжительности производственной фазы. Свежая среда может быть добавлена пропорционально концентрации клеток без удаления культуральной среды (бульона). Чтобы обеспечить добавление среды, запускают подпитываемую культуру в объеме, намного меньшем, чем полная емкость биореактора (например, приблизительно от 40% до 50% от максимального объема).
Некоторые способы в соответствии с настоящим раскрытием направлены на крупномасштабное (коммерческого масштаба) получение РНК (например, мРНК). Для способов крупномасштабного получения клетки можно выращивать в жидкой культуральной среде в объеме от 5 литров (л) до 250000 л или больше. Согласно некоторым вариантам осуществления клетки можно выращивать в жидкой культуральной среде в объеме более чем (или равном) 10 л, 100 л, 1000 л, 10000 л или 100000 л. Согласно некоторым вариантам осуществления клетки выращивают в жидкой культуральной среде в объеме 5 л, 10 л, 15 л, 20 л, 25 л, 30 л, 35 л, 40 л, 45 л, 50 л, 100 л, 500 л, 1000 л, 5000 л, 10000 л, 100000 л, 150000 л, 200000 л, 250000 л или больше. Согласно некоторым вариантам осуществления клетки можно выращивать в жидкой культуральной среде в объеме от 5 л до 10 л, от 5 л до 15 л, от 5 л до 20 л, от 5 л до 25 л, от 5 л до 30 л, от 5 л до 35 л, от 5 л до 40 л, от 5 л до 45 л, от 10 л до 15 л, от 10 л до 20 л, от 10 л до 25 л, от 20 л до 30 л, от 10 л до 35 л, от 10 л до 40 л, от 10 л до 45 л, от 10 л до 50 л, от 15 л до 20 л, от 15 л до 25 л, от 15 л до 30 л, от 15 л до 35 л, от 15 л до 40 л, от 15 л до 45 л или от 15 л до 50 л. Согласно некоторым вариантам осуществления клетки можно выращивать в жидкой культуральной среде в объеме от 100 л до 300000 л, от 100 л до 200000 л или от 100 л до 100000 л.
Как правило, культивирование клеток сопровождается лизированием клеток. Лизирование представляет собой процесс, при котором клетки разрушаются, например, с помощью вирусных, тепловых, химических, ферментативных, механических или осмотических механизмов. Клеточный лизат представляет собой жидкость, содержащую содержимое лизированных клеток (например, лизированных созданных методами генной инженерии клеток), включающее в себя, например, органеллы, мембранные липиды, белки, нуклеиновые кислоты и вывернутые мембранные везикулы. Клеточные лизаты в соответствии с настоящим раскрытием могут быть получены путем лизиса любой популяции созданных методами генной инженерии клеток, как представлено в настоящем документе.
Клеточный лизис может нарушать тщательно контролируемые клеточные среды, что приводит к разложению и модификации белка под действием нерегулируемых эндогенных протеаз и фосфатаз. Таким образом, согласно некоторым вариантам осуществления ингибиторы протеаз, и/или ингибиторы фосфатаз, и/или ингибиторы нуклеази, и/или ингибиторы гидролаз, и/или ингибиторы дезаминаз могут быть добавлены к клеточному лизату или клеткам перед лизисом, или эти активности могут быть удалены тепловой инактивацией, инактивацией генов или нацеливанием протеазы.
Клеточные лизаты согласно некоторым вариантам осуществления могут быть объединены с питательным веществом. Например, клеточные лизаты могут быть объединены с Na2HPO4, KH2PO4, NH4Cl, NaCl, MgSO4, CaCl2. Примеры других питательных веществ включают в себя без ограничения сульфат магния, хлорид магния, оротат магния, цитрат магния, одноосновный фосфат калия, двухосновный фосфат калия, трехосновный фосфат калия, одноосновный фосфат натрия, двухосновный фосфат натрия, трехосновный фосфат натрия, одноосновный фосфат аммония, двухосновный фосфат аммония, сульфат аммония, хлорид аммония и гидроксид аммония.
Клеточные лизаты согласно некоторым вариантам осуществления могут быть объединены с кофактором. Например, клеточные лизаты могут быть объединены с аденозиндифосфатом (АДФ), аденозинтрифосфатом (ATФ), никотинамидадениндинуклеотидом (NAD+) или другими небелковыми химическими соединениями, необходимыми для активности фермента (например, неорганическими ионами и коферментами).
Объем клеточного лизата, используемого для одной реакции, может варьировать. Согласно некоторым вариантам осуществления объем клеточного лизата составляет от 0,001 до 250 м3.
Нуклеиновые кислоты
Термин «нуклеиновая кислота» означает по меньшей мере два нуклеотида, ковалентно связанных вместе, и в некоторых случаях может предусматривать фосфодиэфирные связи (например, фосфодиэфирный «остов»). Нуклеиновые кислоты (например, компоненты или части нуклеиновых кислот) могут встречаться в природе или могут быть созданы методами генной инженерии. «Встречающиеся в природе» нуклеиновые кислоты присутствуют в клетке, которая существует в природе в отсутствие вмешательства человека. Термин «созданные методами генной инженерии нуклеиновые кислоты» включает в себя рекомбинантные нуклеиновые кислоты и синтетические нуклеиновые кислоты. Термин «рекомбинантная нуклеиновая кислота» относится к молекуле, которая конструируется путем соединения молекул нуклеиновой кислоты (например, из одного и того же вида или из разных видов) и, как правило, может реплицироваться в живой клетке. Термин «синтетическая нуклеиновая кислота» относится к молекуле, которая синтезирована биологически, синтезирована химически, или синтезирована другими средствами, или амплифицирована. Синтетическая нуклеиновая кислота включает в себя нуклеиновые кислоты, которые являются химически модифицированными или модифицированными иным образом, но могут образовывать пары оснований с встречающимися в природе молекулами нуклеиновых кислот. Рекомбинантные и синтетические нуклеиновые кислоты также включают в себя те молекулы, которые возникают в результате репликации любого из упомянутых выше. Созданные методами генной инженерии нуклеиновые кислоты могут содержать части нуклеиновых кислот, которые встречаются в природе, но в целом созданные методами генной инженерии нуклеиновые кислоты не встречаются в природе и требуют вмешательства человека. Согласно некоторым вариантам осуществления нуклеиновая кислота, кодирующая продукт в соответствии с настоящим раскрытием, представляет собой рекомбинантную нуклеиновую кислоту или синтетическую нуклеиновую кислоту. Согласно другим вариантам осуществления нуклеиновая кислота, кодирующая продукт, встречается в природе.
Созданная методами генной инженерии ДНК-матрица, кодирующая РНК, как представлено в настоящем документе, может быть функционально связана с промотором, который представляет собой контрольную область нуклеиновой кислоты, по которой контролируются инициация и скорость транскрипции остальной части нуклеиновой кислоты. Промотор управляет экспрессией или управляет транскрипцией нуклеиновой кислоты, которую он регулирует.
Промотор может быть промотором, естественным образом связанным с геном или последовательностью, что может быть получено путем выделения 5’-некодирующих последовательностей, расположенных в 3’-5’-направлении кодирующего сегмента данного гена или последовательности. Такой промотор может быть эндогенным.
Согласно некоторым вариантам осуществления кодирующая последовательность нуклеиновой кислоты может находиться под контролем рекомбинантного или гетерологичного промотора, который относится к промотору, который обычно не связан с кодируемой последовательностью в его естественной среде. Такие промоторы могут включать в себя промоторы других генов; промоторы, выделенные из любой другой клетки; и синтетические промоторы или энхансеры, которые не «встречаются в природе», такие как, например, те, которые содержат различные элементы разных транскрипционных регуляторных областей и/или мутации, экспрессию которых изменяют методами генной инженерии, известными в уровне техники. Помимо получения последовательностей нуклеиновых кислот из промоторов и энхансеров синтетические последовательности могут быть получены с использованием технологии рекомбинантного клонирования и/или амплификации нуклеиновых кислот, в том числе полимеразной цепной реакции (ПЦР).
Считают, что промотор функционально связан с нуклеотидной последовательностью, если он находится в правильном функциональном положении и ориентации по отношению к нуклеотидной последовательности, которую он регулирует, чтобы контролировать («управлять») инициирование транскрипции и/или экспрессию этой нуклеотидной последовательности.
Созданные методами генной инженерии нуклеиновые кислоты в соответствии с настоящим раскрытием могут содержать конститутивный промотор или индуцируемый промотор. Согласно некоторым вариантам осуществления конститутивный промотор или индуцируемый промотор является функционально связанным с кодирующей последовательностью и необязательно с одним или несколькими транскрипционными терминаторами. Согласно некоторым вариантам осуществления кодирующая последовательность кодирует белок или РНК-продукт. Термин «конститутивный промотор» относится к промотору, который является постоянно активным в клетке. Термин «индуцируемый промотор» относится к промотору, который инициирует или усиливает транскрипционную активность, если находится в присутствии, под воздействием или в контакте с индуктором или индуцирующим средством или активирован в отсутствие фактора, который вызывает подавление. Индуцируемые промоторы для применения в соответствии с настоящим раскрытием включают в себя любой индуцируемый промотор, описываемый в настоящем документе или известный рядовому специалисту в данной области. Примеры индуцируемых промоторов включают в себя без ограничения химически/биохимически регулируемые и физически регулируемые промоторы, такие как регулируемые органическим растворителем промоторы, регулируемые тетрациклином промоторы, регулируемые стероидами промоторы, регулируемые металлом промоторы, регулируемые патогенезом промоторы, индуцируемые температурой/нагреванием, регулируемые фосфатом (например, PhoA) и регулируемые светом промоторы.
Созданная методами генной инженерии ДНК-матрица, кодирующая РНК, как представлено в настоящем документе, также может быть функционально связана с одним или несколькими транскрипционными терминаторами, которые представляют собой контрольные области нуклеиновой кислоты, которые заставляют полимеразу прекращать транскрибирование и диссоциироваться от ДНК-матрицы.
Терминатор может представлять собой одну или несколько последовательностей, естественно связанных с геном или последовательностью, что может быть получено путем выделения 3’-некодирующих последовательностей, расположенных в 5’-3’-направлении кодирующего сегмента данного гена или последовательности. Такой терминатор может быть эндогенным или созданным методами генной инженерии для повышения эффективности терминации. Эндогенные и/или созданные методами генной инженерии терминаторные последовательности из одного или нескольких источников могут быть добавлены последовательно для повышения эффективности терминации.
Кольцевые ДНК-матрицы, кодирующие РНК, могут включать в себя один или несколько транскрипционных терминаторов для минимизации или предотвращения транскрипции нематричной последовательности ДНК, например, последовательности, которая является частью плазмидного остова.
Созданные методами генной инженерии нуклеиновые кислоты могут быть введены в клетки-хозяева с использованием любых средств, известных в уровне техники, в том числе без ограничения трансформации, трансфекции (например, химической (например, фосфатом кальция, катионными полимерами или липосомами) или нехимической (например, электропорации, сонопорации, импалефекции, оптической трансфекции, гидродинамической трансфекции)) и трансдукции (например, вирусной трансдукции).
Ферменты или другие белки, кодируемые встречающейся в природе, внутриклеточной нуклеиновой кислотой, могут называться «эндогенными ферментами» или «эндогенными белками».
Композиции
Согласно некоторым вариантам осуществления реакционная смесь для получения нуклеозидтрифосфатов (НTФ) содержит нуклеозиддифосфаты (НДФ), полифосфаткиназу и полифосфат. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу и/или НДФ-киназу.
Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ содержит 5’-нуклеозидмонофосфаты, полифосфаткиназу и полифосфат. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ содержит нуклеозиды, полифосфаткиназу и полифосфат. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ содержит нуклеиновые основания, фосфорибозилтрансферазу, фосфорибозилпирофосфат, полифосфаткиназу и полифосфат. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
Согласно некоторым вариантам осуществления реакционная смесь для получения НТФ содержит нуклеиновые основания, D-рибозу, рибокиназу, фосфопентомутазу, нуклеозидфосфорилазу, полифосфаткиназу и полифосфат. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
Согласно некоторым вариантам осуществления реакционная смесь для получения рибонуклеиновой кислоты (РНК) содержит нуклеозиддифосфаты (НДФ), полифосфаткиназу, полифосфат, матрицу дезоксирибонуклеиновой кислоты (ДНК) и полимеразу. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, NMO киназу и/или НДФ-киназу.
Согласно некоторым вариантам осуществления реакционная смесь для получения РНК содержит 5’-нуклеозидмонофосфаты, полифосфаткиназу, полифосфат, матрицу дезоксирибонуклеиновой кислоты (ДНК) и полимеразу. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
Согласно некоторым вариантам осуществления реакционная смесь для получения РНК содержит нуклеозиды, нуклеозидкиназу, полифосфаткиназу, полифосфат, матрицу дезоксирибонуклеиновой кислоты (ДНК) и полимеразу. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
Согласно некоторым вариантам осуществления реакционная смесь для получения РНК содержит нуклеиновые основания, фосфорибозилтрансферазу, фосфорибозилпирофосфат, полифосфаткиназу, полифосфат, матрицу дезоксирибонуклеиновой кислоты (ДНК) и полимеразу. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
Согласно некоторым вариантам осуществления реакционная смесь для получения РНК содержит нуклеиновые основания, D-рибозу, рибокиназу, фосфопентомутазу, нуклеозидфосфорилазу, полифосфаткиназу, полифосфат, матрицу дезоксирибонуклеиновой кислоты (ДНК) и полимеразу. Согласно некоторым вариантам осуществления реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
Дополнительный вариант осуществления
Дополнительные варианты осуществления настоящего раскрытия охватываются следующими пронумерованными параграфами.
1. Способ получения нуклеозидтрифосфатов (НTФ), предусматривающий инкубирование в реакционной смеси нуклеозиддифосфатов (НДФ), полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу и/или НДФ-киназу.
2. Способ по параграфу 1, при котором НДФ включают в себя АДФ, ГДФ, ЦДФ и/или УДФ.
3. Способ по параграфу 1 или параграфу 2, при котором НДФ синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
4. Способ по любому из параграфов 1-3, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
5. Способ по параграфу 4, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
6. Способ по любому из параграфов 1-5, при котором полифосфат включает в себя гексаметафосфат.
7. Способ по любому из параграфов 1-6, при котором полифосфаткиназу, нуклеозидкиназу и/или НДФ-киназу получают из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу и/или НДФ-киназу.
8. Способ по любому из параграфов 1-7, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу и/или НДФ-киназу.
9. Способ по параграфу 8, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована.
10. Способ по параграфу 9, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
11. Способ по параграфу 9 или параграфу 10, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
12. Способ по любому из параграфов 9-11, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
13. Способ по любому из параграфов 9-12, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
14. Способ по любому из параграфов 1-13, при котором полифосфаткиназа, нуклеозидкиназа и/или НДФ-киназа могут выдерживать условия элиминирования.
15. Способ получения нуклеозидтрифосфатов (НTФ), предусматривающий инкубирование в реакционной смеси 5’-нуклеозидмонофосфатов (5’-НМФ), полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
16. Способ по параграфу 15, при котором 5’-НМФ включают в себя 5’-AMФ, 5’-ГМФ, 5’-ЦМФ и/или 5’-УМФ.
17. Способ по параграфу 15 или параграфу 16, при котором 5’-НМФ синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
18. Способ по любому из параграфов 15-17, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
19. Способ по параграфу 18, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
20. Способ по любому из параграфов 15-19, при котором полифосфат включает в себя гексаметафосфат.
21. Способ по любому из параграфов 15-20, при котором полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу получают из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
22. Способ по любому из параграфов 15-21, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
23. Способ по параграфу 22, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
24. Способ по параграфу 23, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
25. Способ по параграфу 23 или параграфу 24, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
26. Способ по любому из параграфов 23-25, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
27. Способ по любому из параграфов 23-26, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
28. Способ по любому из параграфов 15-27, при котором полифосфаткиназа, нуклеозидкиназа, НМФ-киназа и/или НДФ-киназа могут выдерживать условия элиминирования.
29. Способ получения нуклеозидтрифосфатов (НTФ), предусматривающий инкубирование в реакционной смеси нуклеозидов, полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
30. Способ по параграфу 29, при котором нуклеозиды включают в себя аденозин, гуанозин, цитидин и/или уридин.
31. Способ по параграфу 29 или параграфу 30, при котором нуклеозиды синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
32. Способ по любому из параграфов 29-31, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
33. Способ по параграфу 32, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
34. Способ по любому из параграфов 29-33, при котором полифосфат включает в себя гексаметафосфат.
35. Способ по любому из параграфов 29-34, при котором полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу получают из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
36. Способ по любому из параграфов 29-35, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
37. Способ по параграфу 36, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
38. Способ по параграфу 37, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
39. Способ по параграфу 37 или параграфу 38, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
40. Способ по любому из параграфов 37-39, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
41. Способ по любому из параграфов 37-40, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
42. Способ по любому из параграфов 29-41, при котором полифосфаткиназа, нуклеозидкиназа, НМФ-киназа и/или НДФ-киназа могут выдерживать условия элиминирования.
43. Способ получения нуклеозидтрифосфатов (НTФ), предусматривающий инкубирование в реакционной смеси нуклеиновых оснований, фосфорибозилтрансферазы, фосфорибозилпирофосфата, полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
44. Способ по параграфу 43, при котором нуклеиновые основания включают в себя аденин, гуанидин, цитозин и/или урацил.
45. Способ по параграфу 43 или параграфу 44, при котором нуклеиновые основания синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
46. Способ по любому из параграфов 43-45, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
47. Способ по параграфу 46, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
48. Способ по любому из параграфов 43-47, при котором полифосфат включает в себя гексаметафосфат.
49. Способ по любому из параграфов 43-48, при котором полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу получают из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
50. Способ по любому из параграфов 43-49, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
51. Способ по параграфу 50, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
52. Способ по параграфу 51, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
53. Способ по параграфу 51 или параграфу 52, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
54. Способ по любому из параграфов 51-53, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
55. Способ по любому из параграфов 51-54, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
56. Способ по любому из параграфов 43-55, при котором полифосфаткиназа, нуклеозидкиназа, НМФ-киназа и/или НДФ-киназа могут выдерживать условия элиминирования.
57. Способ получения нуклеозидтрифосфатов (НTФ), предусматривающий инкубирование в реакционной смеси нуклеиновых оснований, рибозы, рибокиназы, фосфопентомутазы, нуклеозидфосфорилазы, полифосфаткиназы и полифосфата в условиях, подходящих для получения НТФ, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
58. Способ по параграфу 57, при котором нуклеиновые основания включают в себя аденин, гуанидин, цитозин и/или урацил.
59. Способ по параграфу 57 или параграфу 58, при котором нуклеиновые основания синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
60. Способ по любому из параграфов 57-59, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
61. Способ по параграфу 60, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
62. Способ по любому из параграфов 57-61, при котором полифосфат включает в себя гексаметафосфат.
63. Способ по любому из параграфов 57-62, при котором рибокиназу, фосфопентомутазу, нуклеозидфосфорилазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу получают из клеток, которые экспрессируют рибокиназу, фосфопентомутазу, нуклеозидфосфорилазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
64. Способ по любому из параграфов 57-63, при котором реакционная смесь содержит лизат, полученный из клеток, которые экспрессируют рибокиназу, фосфопентомутазу, нуклеозидфосфорилазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
65. Способ по параграфу 64, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
66. Способ по параграфу 65, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
67. Способ по параграфу 65 или параграфу 66, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, органического растворителя и/или химических ингибиторов.
68. Способ по любому из параграфов 65-67, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
69. Способ по любому из параграфов 65-68, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
70. Способ по любому из параграфов 57-69, при котором рибокиназу, фосфопентомутазу, нуклеозидфосфорилазу, полифосфаткиназу, НМФ-киназу, НДФ-киназу и/или нуклеозидкиназу модифицируют для выдерживания условий элиминирования.
71. Способ получения нуклеозидтрифосфатов (НTФ), предусматривающий инкубирование в реакционной смеси клеточной рибонуклеиновой кислоты (РНК), полинуклеотидфосфорилазы (ПНФазы), неорганического фосфата, полифосфаткиназы и полифосфата в условиях, подходящих для получения НДФ и НТФ, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу и/или НДФ-киназу.
72. Способ по параграфу 71, при котором клеточная РНК включает в себя рибосомальную РНК, матричную РНК и/или транспортную РНК.
73. Способ по параграфу 61 или параграфу 72, при котором клеточная РНК происходит из одноклеточного организма или многоклеточного организма.
74. Способ по любому из параграфов 71-73, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
75. Способ по параграфу 74, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
76. Способ по любому из параграфов 71-75, при котором полифосфат включает в себя гексаметафосфат.
77. Способ по любому из параграфов 71-76, при котором ПНФазу, полифосфаткиназу, нуклеозидкиназу и/или НДФ-киназу получают из клеток, которые экспрессируют ПНФазу, полифосфаткиназу, нуклеозидкиназу и/или НДФ-киназу.
78. Способ по любому из параграфов 71-77, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют ПНФазу, полифосфаткиназу, нуклеозидкиназу и/или НДФ-киназу.
79. Способ по параграфу 78, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
80. Способ по параграфу 79, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
81. Способ по параграфу 79 или параграфу 80, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
82. Способ по любому из параграфов 79-81, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
83. Способ по любому из параграфов 79-82, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
84. Способ по любому из параграфов 79-83, при котором ПНФаза, полифосфаткиназа, нуклеозидкиназа и/или НДФ-киназа могут выдерживать условия элиминирования.
85. Способ получения нуклеозидтрифосфатов (НTФ), предусматривающий:
(a) инкубирование в реакционной смеси клеточной рибонуклеиновой кислоты (РНК), рибонуклеазы, полифосфаткиназы и полифосфата в условиях, подходящих для получения 5’-НМФ и НТФ, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
86. Способ по параграфу 85, при котором клеточная РНК включает в себя рибосомальную РНК, матричную РНК и/или транспортную РНК.
87. Способ по параграфу 85 или параграфу 86, при котором клеточная РНК происходит из одноклеточного организма или многоклеточного организма.
88. Способ по любому из параграфов 85-87, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
89. Способ по параграфу 88, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
90. Способ по любому из параграфов 85-89, при котором полифосфат включает в себя гексаметафосфат.
91. Способ по любому из параграфов 85-90, при котором рибонуклеазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу получают из клеток, которые экспрессируют рибонуклеазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
92. Способ по любому из параграфов 85-91, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют рибонуклеазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
93. Способ по параграфу 92, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
94. Способ по параграфу 93, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
95. Способ по параграфу 93 или параграфу 94, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
96. Способ по любому из параграфов 93-95, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
97. Способ по любому из параграфов 93-96, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
98. Способ по любому из параграфов 93-97, при котором рибонуклеаза, полифосфаткиназа, нуклеозидкиназа, НМФ-киназа и/или НДФ-киназа могут выдерживать условия элиминирования.
99. Способ получения рибонуклеиновой кислоты (РНК), предусматривающий инкубирование в реакционной смеси нуклеозиддифосфатов (НДФ), полифосфаткиназы, полифосфата, ДНК-матрицы, кодирующей представляющую интерес РНК, и РНК-полимеразы в условиях, подходящих для получения представляющей интерес РНК, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу и/или НДФ-киназу.
100. Способ по параграфу 99, при котором НДФ включают в себя АДФ, ГДФ, ЦДФ и/или УДФ.
101. Способ по параграфу 99 или параграфу 100, при котором НДФ синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
102. Способ по любому из параграфов 99-101, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
103. Способ по параграфу 102, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
104. Способ по любому из параграфов 99-103, при котором полифосфат включает в себя гексаметафосфат.
105. Способ по любому из параграфов 99-104, при котором полифосфаткиназу, ДНК-матрицу, полимеразу, нуклеозидкиназу и/или НДФ-киназу получают из клеток, которые экспрессируют полифосфаткиназу, ДНК-матрицу, полимеразу, нуклеозидкиназу и/или НДФ-киназу.
106. Способ по любому из параграфов 99-105, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют полифосфаткиназу, ДНК-матрицу, полимеразу, нуклеозидкиназу и/или НДФ-киназу.
107. Способ по параграфу 106, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована.
108. Способ по параграфу 107, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
109. Способ по параграфу 107 или параграфу 108, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
110. Способ по любому из параграфов 107-109, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
111. Способ по любому из параграфов 107-110, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
112. Способ по любому из параграфов 99-111, при котором полифосфаткиназа, полимераза, нуклеозидкиназа и/или НДФ-киназа могут выдерживать условия элиминирования.
113. Способ по любому из параграфов 99-112, при котором полимераза включает в себя РНК-полимеразу.
114. Способ получения рибонуклеиновой кислоты (РНК), предусматривающий инкубирование в реакционной смеси 5’-нуклеозидмонофосфатов (5’-НМФ), полифосфаткиназы, полифосфата, ДНК-матрицы, кодирующей представляющую интерес РНК, и полимеразы в условиях, подходящих для получения представляющей интерес РНК, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
115. Способ по параграфу 114, при котором 5’-НМФ включают в себя 5’-AMФ, 5’-ГМФ, 5’-ЦМФ и/или 5’-УМФ.
116. Способ по параграфу 114 или параграфу 115, при котором 5’-НМФ синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
117. Способ по любому из параграфов 114-116, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
118. Способ по параграфу 117, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
119. Способ по любому из параграфов 114-118, при котором полифосфат включает в себя гексаметафосфат.
120. Способ по любому из параграфов 114-119, при котором полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу получают из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
121. Способ по любому из параграфов 114-120, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
122. Способ по параграфу 121, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
123. Способ по параграфу 122, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
124. Способ по параграфу 122 или параграфу 123, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
125. Способ по любому из параграфов 122-124, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
126. Способ по любому из параграфов 122-125, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
127. Способ по любому из параграфов 114-126, при котором полифосфаткиназа, нуклеозидкиназа, НМФ-киназа, НДФ-киназа и/или полимераза могут выдерживать условия элиминирования.
128. Способ по любому из параграфов 114-127, при котором полимераза включает в себя РНК-полимеразу.
129. Способ получения рибонуклеиновой кислоты (РНК), предусматривающий инкубирование в реакционной смеси нуклеозидов, полифосфаткиназы, полифосфата, ДНК-матрицы, кодирующей представляющую интерес РНК, и/или полимеразы в условиях, подходящих для получения представляющей интерес РНК, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
130. Способ по параграфу 129, при котором нуклеозиды включают в себя аденозин, гуанозин, цитидин и/или уридин.
131. Способ по параграфу 129 или параграфу 130, при котором нуклеозиды синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
132. Способ по любому из параграфов 129-131, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
133. Способ по параграфу 132, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
134. Способ по любому из параграфов 129-133, при котором полифосфат включает в себя гексаметафосфат.
135. Способ по любому из параграфов 129-134, при котором полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу получают из клеток, которые экспрессируют по меньшей мере одно из полифосфаткиназы, нуклеозидкиназы, полифосфата, нуклеозидкиназы, НМФ-киназы, НДФ-киназы, ДНК-матрицы и/или полимеразы.
136. Способ по любому из параграфов 129-135, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют по меньшей мере одну полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
137. Способ по параграфу 136, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
138. Способ по параграфу 137, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
139. Способ по параграфу 137 или параграфу 138, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
140. Способ по любому из параграфов 137-139, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
141. Способ по любому из параграфов 137-140, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
142. Способ по любому из параграфов 129-141, при котором по меньшей мере одна полифосфаткиназа, полифосфат, нуклеозидкиназа, НМФ-киназа, НДФ-киназа и/или полимераза может выдерживать условия элиминирования.
143. Способ по любому из параграфов 129-142, при котором полимераза включает в себя РНК-полимеразу.
144. Способ получения рибонуклеиновой кислоты (РНК), предусматривающий инкубирование в реакционной смеси нуклеиновых оснований, фосфорибозилтрансферазы, фосфорибозилпирофосфата, полифосфаткиназы и полифосфата, ДНК-матрицы, кодирующей представляющую интерес РНК, и/или полимеразы в условиях, подходящих для получения представляющей интерес РНК, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
145. Способ по параграфу 144, при котором нуклеиновые основания включают в себя аденин, гуанидин, цитозин и/или урацил.
146. Способ по параграфу 144 или параграфу 145, при котором нуклеиновые основания синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
147. Способ по любому из параграфов 144-146, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
148. Способ по параграфу 148, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
149. Способ по любому из параграфов 144-148, при котором полифосфат включает в себя гексаметафосфат.
150. Способ по любому из параграфов 144-149, при котором фосфорибозилтрансферазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу получают из клеток, которые экспрессируют фосфорибозилтрансферазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
151. Способ по любому из параграфов 144-150, при котором реакционная смесь содержит клеточный лизат или ферментативный препарат из клеток, которые экспрессируют фосфорибозилтрансферазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
152. Способ по параграфу 151, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
153. Способ по параграфу 152, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
154. Способ по параграфу 152 или параграфу 153, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
155. Способ по любому из параграфов 152-154, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
156. Способ по любому из параграфов 152-155, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
157. Способ по любому из параграфов 144-156, при котором фосфорибозилтрансфераза, полифосфаткиназа, нуклеозидкиназа, НМФ-киназа, НДФ-киназа и/или полимераза могут выдерживать условия элиминирования.
158. Способ по любому из параграфов 144-157, при котором полимераза включает в себя РНК-полимеразу.
159. Способ получения рибонуклеиновой кислоты (РНК), предусматривающий инкубирование в реакционной смеси нуклеиновых оснований, рибозы, рибокиназы, фосфопентомутазы, нуклеозидфосфорилазы, полифосфаткиназы, полифосфата, ДНК-матрицы, кодирующей представляющую интерес РНК, и полимеразы в условиях, подходящих для получения представляющей интерес РНК, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или по меньшей мере одну НДФ-киназу.
160. Способ по параграфу 159, при котором нуклеиновые основания включают в себя аденин, гуанидин, цитозин и/или урацил.
161. Способ по параграфу 159 или параграфу 160, при котором нуклеиновые основания синтезируются химическим путем, являются продуктом ферментации или экстрагируются из природного источника.
162. Способ по любому из параграфов 159-161, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
163. Способ по параграфу 162, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
164. Способ по любому из параграфов 159-163, при котором полифосфат включает в себя гексаметафосфат.
165. Способ по любому из параграфов 159-164, при котором рибокиназа, фосфопентомутаза, нуклеозидфосфорилаза, полифосфаткиназа, нуклеозидкиназа, НМФ-киназа, НДФ-киназа, ДНК-матрица и/или полимераза происходят по меньшей мере из одного лизата, полученного из созданных методами генной инженерии клеток, модифицированных для экспрессии рибокиназы, фосфопентомутазы, нуклеозидфосфорилазы, полифосфаткиназы, нуклеозидкиназы, НМФ-киназы, НДФ-киназы, ДНК-матрицы и/или полимеразы.
166. Способ по любому из параграфов 159-165, при котором лизат реакционной смеси получен из клеток, которые экспрессируют рибокиназу, фосфопентомутазу, нуклеозидфосфорилазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
167. Способ по параграфу 166, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована.
168. Способ по параграфу 167, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
169. Способ по параграфу 167 или параграфу 168, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, органического растворителя и/или химических ингибиторов.
170. Способ по любому из параграфов 167-169, при котором нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
171. Способ по любому из параграфов 167-170, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
172. Способ по любому из параграфов 159-171, при котором рибокиназа, фосфопентомутаза, нуклеозидфосфорилаза, полифосфаткиназа, нуклеозидкиназа, НМФ-киназа, НДФ-киназа и/или полимераза являются модифицированными для выдерживания условий элиминирования.
173. Способ по любому из параграфов 159-172, при котором по меньшей мере одна полимераза включает в себя по меньшей мере одну РНК-полимеразу.
174. Способ получения рибонуклеиновой кислоты (РНК), предусматривающий:
(a) инкубирование в реакционной смеси клеточной рибонуклеиновой кислоты (РНК), полинуклеотидфосфорилазы (ПНФазы) и неорганического фосфата в условиях, подходящих для получения нуклеозиддифосфатов (НДФ);
(b) элиминирование ПНФазы и
(c) инкубирование в реакционной смеси или во второй реакционной смеси НДФ, полифосфаткиназы, полифосфата, ДНК-матрицы, кодирующей представляющую интерес РНК, и полимеразы в условиях, подходящих для получения представляющей интерес РНК, необязательно при этом реакционная смесь стадии (c) дополнительно содержит нуклеозидкиназу и/или НДФ-киназу.
175. Способ по параграфу 174, при котором клеточная РНК включает в себя рибосомальную РНК, матричную РНК и/или транспортную РНК.
176. Способ по параграфу 174 или параграфу 175, при котором клеточная РНК происходит из одноклеточного организма или многоклеточного организма.
177. Способ по любому из параграфов 174-176, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
178. Способ по параграфу 177, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
179. Способ по любому из параграфов 174-178, при котором полифосфат включает в себя гексаметафосфат.
180. Способ по любому из параграфов 174-179, при котором ПНФазу получают из клеток, которые экспрессируют ПНФазу.
181. Способ по любому из параграфов 174-180, при котором реакционная смесь стадии (a) содержит клеточный лизат, полученный из клеток, которые экспрессируют ПНФазу.
182. Способ по параграфу 181, при котором стадия (b) предусматривает элиминирование ПНФазы посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
183. Способ по параграфу 181 или параграфу 182, при котором стадия (b) предусматривает элиминирование ПНФазы, посредством чего нативная ферментативная активность ферментов в клеточном лизате или ферментативном препарате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
184. Способ по любому из параграфов 174-183, при котором полифосфаткиназу, нуклеозидкиназу, НДФ-киназу, ДНК-матрицу и/или полимеразу получают из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
185. Способ по любому из параграфов 174-183, при котором реакционная смесь стадии (c) содержит клеточный лизат, полученный из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
186. Способ по параграфу 185, при котором нативная ферментативная активность ферментов в клеточном лизате стадии (c) была элиминирована.
187. Способ по параграфу 186, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
188. Способ по параграфу 186 или параграфу 187, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
189. Способ по любому из параграфов 186-188, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
190. Способ по любому из параграфов 186-189, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
191. Способ по любому из параграфов 186-190, при котором полифосфаткиназа, нуклеозидкиназа, НДФ-киназа и/или полимераза могут выдерживать условия элиминирования.
192. Способ по любому из параграфов 174-191, при котором полимераза включает в себя РНК-полимеразу.
193. Способ получения рибонуклеиновой кислоты (РНК), предусматривающий:
(a) инкубирование в реакционной смеси клеточной рибонуклеиновой кислоты (РНК), полинуклеотидфосфорилазы (ПНФазы), неорганического фосфата, полифосфаткиназы, полифосфата, ДНК-матрицы, кодирующей представляющую интерес РНК, и полимеразы в условиях, подходящих для получения нуклеозиддифосфатов, необязательно при этом реакционная смесь дополнительно содержит нуклеозидкиназу и/или НДФ-киназу;
(b) элиминирование ПНФазы и
(c) инкубирование реакционной смеси в условиях, подходящих для получения представляющей интерес РНК.
194. Способ по параграфу 193, при котором клеточная РНК включает в себя рибосомальную РНК, матричную РНК и/или транспортную РНК.
195. Способ по параграфу 193 или параграфу 194, при котором клеточная РНК происходит из одноклеточного организма или многоклеточного организма.
196. Способ по любому из параграфов 193-195, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
197. Способ по параграфу 196, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
198. Способ по любому из параграфов 193-197, при котором полифосфат включает в себя гексаметафосфат.
199. Способ по любому из параграфов 193-198, при котором ПНФазу, полифосфаткиназу, нуклеозидкиназу, НДФ-киназу, ДНК-матрицу и/или полимеразу получают из клеток, которые экспрессируют ПНФазу, полифосфаткиназу, нуклеозидкиназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
200. Способ по любому из параграфов 193-199, при котором реакционная смесь стадии (a) содержит клеточный лизат, полученный из клеток, которые экспрессируют ПНФазу, полифосфаткиназу, нуклеозидкиназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
201. Способ по параграфу 200, при котором стадия (b) предусматривает элиминирование ПНФазы и нативных ферментативных активностей в клеточном лизате посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
202. Способ по параграфу 200 или параграфу 201, при котором стадия (b) предусматривает элиминирование ПНФазы и нативных ферментативных активностей в клеточном лизате посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
203. Способ по любому из параграфов 200-202, при котором стадия (b) предусматривает элиминирование нативных ферментативных активностей в клеточном лизате посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
204. Способ по любому из параграфов 201-203, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
205. Способ по любому из параграфов 201-204, при котором полифосфаткиназа, нуклеозидкиназа, НДФ-киназа и/или полимераза могут выдерживать условия элиминирования.
206. Способ по любому из параграфов 193-205, при котором полимераза включает в себя РНК-полимеразу.
207. Способ получения рибонуклеиновой кислоты (РНК), предусматривающий:
(a) инкубирование в реакционной смеси клеточной рибонуклеиновой кислоты (РНК) и рибонуклеазы в условиях, подходящих для получения 5’-нуклеозидмонофосфатов (5’-НМФ);
(b) элиминирование рибонуклеазы и
(c) инкубирование в реакционной смеси или во второй реакционной смеси 5’-НМФ, полифосфаткиназы, полифосфата, ДНК-матрицы, кодирующей представляющую интерес РНК, и полимеразы в условиях, подходящих для получения представляющей интерес РНК, необязательно при этом реакционная смесь стадии (c) дополнительно содержит нуклеозидкиназу, НМФ-киназу и/или НДФ-киназу.
208. Способ по параграфу 207, при котором клеточная РНК включает в себя рибосомальную РНК, матричную РНК и/или транспортную РНК.
209. Способ по параграфу 207 или параграфу 208, при котором клеточная РНК происходит из одноклеточного организма или многоклеточного организма.
210. Способ по любому из параграфов 207-209, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
211. Способ по параграфу 210, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
212. Способ по любому из параграфов 207-211, при котором полифосфат включает в себя гексаметафосфат.
213. Способ по любому из параграфов 207-212, при котором рибонуклеазу получают из клеток, которые экспрессируют рибонуклеазу.
214. Способ по любому из параграфов 207-213, при котором реакционная смесь стадии (a) содержит клеточный лизат, полученный из клеток, которые экспрессируют рибонуклеазу.
215. Способ по параграфу 214, при котором стадия (b) предусматривает элиминирование рибонуклеазы посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
216. Способ по параграфу 214 или параграфу 215, при котором стадия (b) предусматривает элиминирование рибонуклеазы посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
217. Способ по любому из параграфов 207-216, при котором полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу получают из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
218. Способ по любому из параграфов 207-216, при котором реакционная смесь стадии (c) содержит клеточный лизат, полученный из клеток, которые экспрессируют полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
219. Способ по параграфу 218, при котором нативная ферментативная активность ферментов в клеточном лизате стадии (c) была элиминирована.
220. Способ по параграфу 219, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
221. Способ по параграфу 219 или параграфу 220, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
222. Способ по любому из параграфов 219-221, при котором нативная ферментативная активность ферментов в клеточном лизате была элиминирована посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
223. Способ по любому из параграфов 219-222, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
224. Способ по любому из параграфов 219-223, при котором полифосфаткиназа, нуклеозидкиназа, НМФ-киназа, НДФ-киназа и/или полимераза могут выдерживать условия элиминирования.
225. Способ по любому из параграфов 207-224, при котором полимераза включает в себя по меньшей мере одну РНК-полимеразу.
226. Способ получения рибонуклеиновой кислоты (РНК), предусматривающий:
(a) инкубирование в реакционной смеси клеточной рибонуклеиновой кислоты (РНК), рибонуклеазы, полифосфаткиназы, полифосфата, ДНК-матрицы, кодирующей представляющую интерес РНК, и полимеразы в условиях, подходящих для получения 5’-нуклеозидмонофосфатов (5’-НМФ);
(b) элиминирование рибонуклеазы и
(c) инкубирование реакционной смеси в условиях, подходящих для получения представляющей интерес РНК.
227. Способ по параграфу 226, при котором клеточная РНК включает в себя рибосомальную РНК, матричную РНК и/или транспортную РНК.
228. Способ по параграфу 226 или параграфу 214, при котором клеточная РНК происходит из одноклеточного организма или многоклеточного организма.
229. Способ по любому из параграфов 226-228, при котором полифосфаткиназа выбрана из семейства ферментов PPK1 и семейства ферментов PPK2.
230. Способ по параграфу 229, при котором полифосфаткиназа включает в себя полифосфаткиназу 2 класса III из Deinococcus geothermalis.
231. Способ по любому из параграфов 226-230, при котором полифосфат включает в себя гексаметафосфат.
232. Способ по любому из параграфов 226-231, при котором рибонуклеазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу получают из клеток, которые экспрессируют рибонуклеазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
233. Способ по любому из параграфов 227-232, при котором реакционная смесь стадии (a) содержит клеточный лизат, полученный из клеток, которые экспрессируют рибонуклеазу, полифосфаткиназу, нуклеозидкиназу, НМФ-киназу, НДФ-киназу, ДНК-матрицу и/или полимеразу.
234. Способ по параграфу 233, при котором стадия (b) предусматривает элиминирование рибонуклеазы и нативных ферментативных активностей в клеточном лизате посредством температуры, pH, соли, детергента, спирта и/или химических ингибиторов.
235. Способ по параграфу 233 или параграфу 234, при котором стадия (b) предусматривает элиминирование рибонуклеазы и нативных ферментативных активностей в клеточном лизате посредством отделения, осаждения, фильтрации, захвата и/или хроматографии.
236. Способ по любому из параграфов 233-235, при котором стадия (b) предусматривает элиминирование нативных ферментативных активностей в клеточном лизате посредством генетической модификации, секреции ферментов из клетки и/или нацеливания протеазы.
237. Способ по любому из параграфов 234-236, при котором нативные ферментативные активности выбраны из фосфатаз, нуклеаз, протеаз, дезаминаз, оксидоредуктаз и гидролаз.
238. Способ по любому из параграфов 234-237, при котором полифосфаткиназа, нуклеозидкиназа, НМФ-киназа, НДФ-киназа и/или полимераза могут выдерживать условия элиминирования.
239. Способ по любому из параграфов 226-238, при котором полимераза включает в себя РНК-полимеразу.
Примеры
Пример 1. Бесклеточный синтез РНК, начиная либо с РНК лизата, либо с очищенной РНК E. coli
Материалы и способы
Штаммы и лизаты
Штамм BL21(DE3) E. coli трансформировали векторами pETDuet-1, кодирующими кодон-оптимизированные версии следующих ферментов: РНКазы R (K544R) из E. coli (EcRNR), УМФ-киназы из Pyrococcus furiosus (PfPyrH), ЦМФ-киназы из Thermus thermophilus (TthCmk), ГМФ-киназы из Thermotoga maritima (TmGmk), НДФ-киназы из Aquifex aeolicus (AaNdk) и полифосфаткиназы 2 класса III из Deinococcus geothermalis (DgPPK2). Все ферменты, за исключением DgPPK2, содержали N-концевые гексагистидиновые метки. Полученные в результате штаммы выращивали в процессе периодической ферментации при 37°C в среде Korz, дополненной 40 г/л глюкозы и 50 мг/л карбенициллина, до OD600 = 20, при индуцировании с помощью 0,8 мМ изопропил-D-1-тиогалактопиранозида (IPTG) и выращивали в течение дополнительного 1 часа до сбора с помощью центрифугирования. После сбора гранулы биомассы хранили при -80°С.
Гранулы биомассы затем использовали для получения клеточных лизатов. Лизаты получали путем размораживания гранул биомассы на льду, затем повторно суспендировали в 1,5 объема буфера для повторного суспендирования. Для штамма, экспрессирующего EcRNR, биомассу повторно суспендировали в 58,8 мМ двухосновного фосфата калия. Для штаммов, экспрессирующих PfPyrH, TthCmk, TmGmk и AaNdk, биомассу повторно суспендировали в 50 мМ Tris-HCl (pH 8,5) с 50 мМ NaCl. Для штамма, экспрессирующего DgPPK2, биомассу повторно суспендировали в 100 мМ MOPS-NaOH (pH 7,0). Для всех штаммов, биомассу лизировали с помощью 2-3 циклов механической гомогенизации при 15000 фунтов на квадратный дюйм при 4°C. Лизаты затем осветляли с помощью центрифугирования при 16000 × g в течение 1 часа при 4°C.
Экстракция и очистка РНК E. coli
РНК экстрагировали и очищали из лизатов E. coli с высокой плотностью (концентрация белка: 40-50 мг/мл) в соответствии с установленными протоколами (Mohanty, B. K., Giladi, H., Maples, V. F., & Kushner, S. R. (2008). Analysis of RNA decay, processing, and polyadenylation in Escherichia coli and other prokaryotes. Methods in Enzymology, 447, 3-29).
Экспрессия и очистка белка
Штамм BL21(DE3) E. coli трансформировали вектором pBAD24, кодирующим термостабильную мутантную T7 РНК-полимеразу с N-концевой гексагистидиновой меткой. Полученный в результате штамм культивировали во встряхиваемых колбах с дефлектором с использованием среды лизогенного бульона (LB), дополненной 50 мг/л карбенициллина. Культуры выращивали при 37°C со встряхиванием до OD600 = 0,6, затем индуцировали с помощью 0,2% (масса/объем) L-арабинозы и выращивали в течение дополнительных 4 часов. Биомассу затем собирали с помощью центрифугирования и хранили при -80°С до лизиса. Лизаты получали путем размораживания гранул биомассы на льду, повторного суспендирования в 1,5 объема буфера для уравновешивания/промывания (20 мМ фосфата натрия, pH 7,4, 500 мМ хлорида натрия, 30 мМ имидазола) и подвергали 2-3 циклам механической гомогенизации при 15000 фунтов на квадратный дюйм и 4°C. Лизаты затем осветляли с помощью центрифугирования. Рекомбинантный белок очищали жидкостной экспресс-хроматографией белков (FPLC) с использованием AKTAPrime Plus, оснащенного колонкой HisTrap HP (GE Healthcare Life Sciences), в соответствии со стандартными протоколами. Фракции, содержащие рекомбинантный белок, затем объединяли и заменяли буфер путем диализа в 2× забуференном фосфатом солевом растворе (PBS), дополненном 5 мМ DTT и 0,01% Triton X-100. После диализа стоковые растворы белка разбавляли равным объемом глицерина и хранили при -20°C.
Для РНКазы R E. coli выращивали культуры, индуцировали экспрессию и получали лизаты в соответствии с протоколами, описываемыми в настоящем документе. Фермент затем очищали в соответствии с протоколами, описываемыми в настоящем документе, за исключением того, что очищенный фермент подвергали замене буфера путем диализа в 2× PBS, дополненном 500 мМ NaCl, перед смешиванием с глицерином.
Получение ДНК-матрицы
Линейные ДНК-матрицы амплифицировали из синтетической ДНК с помощью ПЦР и очищали с использованием твердофазной обратимой иммобилизации (SPRI) на парамагнитных гранулах. Плазмидные ДНК-матрицы получали путем клонирования представляющей интерес последовательности в подходящий плазмидный вектор, трансформации полученной в результате плазмиды в штамм DH10b E. coli, культивирование трансформантов в среде LB и очистки плазмид с использованием наборов Plasmid Maxi или Giga (Qiagen).
Бесклеточный синтез РНК из РНК лизата
Каждый из лизатов, экспрессирующих EcRNR, TthCmk, PfPyrH, TmGmk, AaNdk и DgPPK2, разбавляли до 42 мг/мл в 50 мМ Tris, 50 мМ NaCl (pH 7,0), затем объединяли в равной пропорции. Деполимеризацию РНК в лизатах инициировали путем добавления равного объема 3 мМ этилендиаминтетрауксусной кислоты (EDTA) в 50 мМ Tris, 50 мМ NaCl (pH 7,0) и инкубирования при 37°C в течение 15 минут. За деполимеризацией наблюдали с помощью определения количества растворимых в кислоте нуклеотидов по поглощению при 260 нм. Добавляли сульфат магния и гексаметафосфат натрия в лизаты до конечных концентраций 30 мМ и 1 мМ, соответственно, затем лизаты инкубировали при 70°C в течение 15 минут для инактивации EcRNR и эндогенных ферментов E. coli. Через 15 минут температуру снижали до 50°C и собирали реакционную смесь со следующей композицией.
Таблица 11. Условия реакции
Реакционные смеси инкубировали при 50°C в течение 2 часов, обрабатывали с помощью ДНКазы TURBO (Thermo Fisher) и анализировали с помощью электрофореза на агарозном геле.
Бесклеточный синтез РНК из очищенной РНК E. coli
Очищенную РНК E. coli (приблизительно 8 г/л) деполимеризовали с помощью инкубирования с 1 г/л очищенной РНКазы R E. coli в буфере, содержащем 50 мМ Tris-HCl (pH 7,0), 50 мМ хлорида натрия и 2 мМ сульфата магния. Реакционную смесь деполимеризации инкубировали при 37°C в течение 30 минут и наблюдали с помощью определения количества растворимых в кислоте нуклеотидов по поглощению при 260 нм. Через 30 минут реакционную смесь деполимеризации объединяли с лизатами TthCmk, TmGmk, PfPyrH, AaNdk и DgPPK2, каждый из которых разбавляли в 50 мМ Tris-HCl (pH 7,0), 50 мМ NaCl и смешивали в равной пропорции. Затем добавляли сульфат магния и гексаметафосфат натрия в лизаты до конечных концентраций 30 мМ и 1 мМ, соответственно, и реакционную смесь нагревали до 70°C в течение 15 минут. После инактивации нагреванием собирали реакционные смеси, как описывается в настоящем документе, со следующей композицией.
Таблица 12. Условия реакции
Результаты
Бесклеточный синтез РНК выполняли с использованием РНК лизата в качестве субстрата. РНК из объединенных лизатов, надэкспрессирующих ферменты пути, сначала деполимеризовали с помощью надэкспрессированной РНКазы R E. coli (EcRNR), экзонуклеазы, которая продуцирует 5’-НМФ. Деполимеризация была быстрой с образованием приблизительно 14 мМ растворимых в кислоте нуклеотидов через 15 минут (фиг. 8A). Смесь лизата, содержащую ферменты пути и 5’-НМФ, затем обрабатывали нагреванием для инактивации EcRNR и эндогенных ферментов E. coli, тогда как термостабильные ферменты пути оставались активными. После инактивации нагреванием собирали реакционную смесь синтеза РНК, инкубировали при 50°C и визуализировали на агарозном геле (фиг. 8B). Реакции с двумя разными матрицами, включающие в себя линейный ПЦР-продукт (матрицу 1) и матрицу-шпильку, кодируемую в плазмиде (матрицу 2), давали отличающиеся полосы на агарозном геле (фиг. 8B). Полосы не наблюдали в условиях без РНК-полимеразы. В качестве положительного контроля реакции осуществляли при добавлении очищенных 5’-НМФ (по 4 мМ каждого из AMФ, ЦМФ, ГМФ и УМФ). Эти реакции давали продукты, которые мигрировали с тем же размером, что и при реакциях с использованием НМФ, полученных с помощью деполимеризации РНК.
Бесклеточный синтез РНК также выполняли с использованием очищенной РНК E. coli в качестве субстрата. РНК сначала инкубировали с очищенной РНКазой R E. coli (K544R) для высвобождения 5’-НМФ (фиг. 9A). Через 30 минут реакционную смесь деполимеризации объединяли со смесью лизата, содержащей ферменты пути, и обрабатывали нагреванием для инактивации EcRNR и эндогенных ферментов E. coli, тогда как термостабильные ферменты пути оставались активными. После инактивации нагреванием собирали реакционную смесь синтеза РНК, инкубировали при 50°C и визуализировали на агарозном геле (фиг. 9B). Реакции с двумя разными матрицами, включающие в себя линейный ПЦР-продукт (матрицу 1) и матрицу-шпильку, кодируемую в плазмиде (матрицу 2), давали отличающиеся полосы на агарозном геле, хотя выходы оказались более низкими для матрицы 2 (фиг. 9B). Опять же, никаких полос не наблюдали в условиях без РНК-полимеразы.
Взятые вместе, эти результаты демонстрируют, что бесклеточный синтез РНК может быть использован для получения представляющей интерес РНК из различных материалов источника НМФ, включающих в себя НМФ высокой чистоты, НМФ, полученные ферментативной деполимеризацией РНК лизата, и НМФ, полученные ферментативной деполимеризацией очищенной РНК.
Пример 2. Бесклеточных синтез РНК с использованием нетермостабильной полимеразы дикого типа или термостабильной мутантной полимеразы и очищенных НМФ в качестве субстрата
Материалы и способы
Биомассу, лизаты, очищенные белки и ДНК-матрицу получали, как описывается в настоящем документе. Бесклеточных синтез РНК выполняли по сути, как описано в примере 1, за исключением того, что EcRNR не использовали и добавляли по 4 мМ каждого из AMФ, ЦМФ, ГМФ и УМФ в реакцию.
Результаты
Бесклеточный синтез РНК также выполняли с использованием очищенных НМФ в качестве субстрата при температуре реакции 37°C. Лизаты, содержащие ферменты пути, собирали и обрабатывали нагреванием для инактивации эндогенных ферментов E. coli, тогда как термостабильные ферменты пути оставались активными. После инактивации нагреванием собирали реакционную смесь с использованием либо T7 РНК-полимеразы дикого типа, либо термостабильного мутанта, инкубировали при 37°C в течение 2 часов и визуализировали на агарозном геле (фиг. 10). Реакции с двумя разными матрицами, включающие в себя линейный ПЦР-продукт (матрицу 1) и матрицу-шпильку, кодируемую в плазмиде (матрицу 2), давали отличающиеся полосы с обеими полимеразами. При 37°С выход РНК (на основании интенсивности полос геля) оказался больше с полимеразой дикого типа, чем с термостабильным мутантом, особенно с матрицей 2. В условиях без РНК-полимеразы полосы не наблюдали.
Эти результаты демонстрируют, что РНК-полимераза, используемая для бесклеточного синтеза РНК, не обязательно должна быть термостабильной, и что РНК может быть получена с использованием Т7 РНК-полимеразы дикого типа в реакции при 37°С.
Пример 3. Бесклеточный синтез РНК с использованием полифосфаткиназы 2 класса III из Deinococcus geothermalis (DgPPK2) и НМФ в качестве субстрата
Материалы и способы
Реакционные среды бесклеточного синтеза РНК собирали, по сути, как описано в примере 2, со следующей композицией.
Таблица 13. Условия реакции
В качестве положительного контроля использовали лизатную систему из 5 ферментов, содержащую уридилаткиназу, цитидилаткиназу, гуанилаткиназу, нуклеотиддифосфаткиназу и полифосфаткиназу, в реакции согласно примерам 1-2. dsRNA, синтезированную в реакциях, очищали по адаптированному протоколу экстракции RNASwift и определяли количественно с использованием ион-парной обращеннофазной хроматографии, как описывается в настоящем документе (Nwokeji, A. O., Kilby, P. M., Portwood, D. E., & Dickman, M. J. (2016). RNASwift: A rapid, versatile RNA extraction method free from phenol and chloroform. Analytical Biochemistry, 512, 36-46).
Результаты
Бесклеточный синтез РНК выполняли в реакциях, предусматривающих DgPPK2 в качестве единственной киназы в реакции и НМФ в качестве субстрата. В качестве положительного контроля РНК синтезировали в присутствии лизатной системы из 5 ферментов, содержащей уридилаткиназу, цитидилаткиназу, гуанилаткиназу, нуклеотиддифосфаткиназу и полифосфаткиназу. Контрольную реакция осуществляли в отсутствие полимеразы.
Определяли фактор отклика для каждой реакции. Фактор отклика вычисляли как отношение площади представляющей интерес dsRNA к площади коммерчески доступного внутреннего стандарта dsRNA. Сравнение факторов отклика демонстрировало, что реакции, предусматривающие DgPPK2 в качестве единственной киназы, синтезировали ~48% синтезированной dsRNA, которую синтезировали в реакциях, предусматривающих лизатную систему из 5 ферментов (фиг. 11A). dsRNA-продукт, синтезированный в реакции с DgPPK2 и реакции с лизатной системой из 5 ферментов, анализировали с помощью HPLC. Хроматограммы HPLC dsRNA-продукта из реакций были подобными, что указывает на то, что dsRNA-продукт, продуцируемый системой только с DgPPK2, был сходен с продуктом, продуцируемым системой из 5 ферментов (фиг. 11B).
Взятые вместе, эти результаты показывают, что DgPPK2 можно использовать в качестве единственной киназы для синтеза бесклеточной dsRNA из НМФ или НДФ.
Пример 4. Деполимеризация РНК из различных источников с использованием очищенной РНКазы R или очищенной нуклеазы P1
Материалы и способы
Экстракция и очистка РНК и нуклеазы
РНК экстрагировали и очищали из лизатов E. coli с высокой плотностью (концентрация белка: 40-50 мг/мл) в соответствии с установленными протоколами (Mohanty, B. K., Giladi, H., Maples, V. F., & Kushner, S. R. (2008). Analysis of RNA decay, processing, and polyadenylation in Escherichia coli and other prokaryotes. Methods in Enzymology, 447, 3-29).
РНК из Vibrio очищали из клеточного бульона V. natriegens с использованием протокола RNASwift (Nwokeji, A. O., Kilby, P. M., Portwood, D. E., & Dickman, M. J. (2016). RNASwift: A rapid, versatile RNA extraction method free from phenol and chloroform. Analytical Biochemistry, 512, 36-46).
Полученный из дрожжей экстракт РНК приобретали из коммерческих источников. Порошок РНК характеризовался чистотой ~85% - ~90% и не требовал дополнительной очистки. РНКазу R очищали из штамма E. coli, надэкспрессирующего РНКазу R и выращиваемого до высокой плотности клеток. Белки очищали с помощью аффинной хроматографии с использованием иммобилизованных металлов с использованием колонок HisTrap HP, соединенных с системой AKTAPrime Plus FPLC (GE Healthcare). Очищенную нуклеазу P1, 5’-фосфодиэстеразу, получали из коммерческих источников.
Деполимеризация РНК с экзогенной нуклеазой
РНК E. coli, РНК V. natriegens, полученный из дрожжей порошок РНК, повторно суспендированный в не содержащей нуклеазу воде (содержание РНК 11 мг/мл), раствор РНКазы R (1 мг/мл в 300 мМ буфере на основе фосфата калия, pH 7,4, 200 мМ KCl, 2 мМ MgCl2) и очищенную нуклеазу P1 (также называемую 5’-фосфодиэстеразой) (1-2 мг/мл в 100 мМ буфере на основе фосфата калия, pH 7,4, 1 мМ ZnCl2, 10 мМ MgCl2) предварительно уравновешивали при 2°C перед инициацией реакции. В момент времени t = 0 смешивали 50 мкл РНК и 50 мкл раствора нуклеазы и реакцию инициировали путем переноса в предварительно нагретый до 37°C блок. После инициации реакционные смеси инкубировали при 37°C и периодически отбирали образцы путем переноса 10 мкл в раствор для остановки реакции с добавлением кислоты (90 мкл 0,2 M серной кислоты) на льду. После завершения времени погашенные образцы осветляли с помощью центрифугирования при 3200 × g в течении 20 минут при 2°C. Деполимеризацию сначала определяли количественно по поглощению растворимых в кислоте нуклеотидов при 260 нм. Суммарный пул нуклеотидов (например, при 100% деполимеризации) определяли с помощью щелочного гидролиза РНК: 50 мкл РНК объединяли с 150 мкл 0,2 M гидроксида калия, затем нагревали до 99°C в течение 20 минут. Затем останавливали реакцию в гидролизованных щелочью образцах и анализировали, как описывается выше. Деполимеризацию также определяли с помощью анализа LC-MS 5’-, 2’- и 3’-НМФ: в 10 мкл образца останавливали реакцию в 30 мкл 100% ацетонитрила и разбавляли в 500 мкл деионизированной воды, содержащей 10 мкM адипиновой кислоты, используемой в качестве внутреннего стандарта. Затем образец центрифугировали и пропускали через 0,2-мкм фильтр перед анализом LC-MS.
Анализ нуклеотидов
Анализ 2’-, 3’- и 5’-НМФ выполняли с помощью масс-спектрометрии и жидкостной хроматографии с использованием масс-спектрометра ABSCIEX API 5000 и стандартной Agilent 1200 HPLC, оснащенной колонкой Sequant Zinc-hilic (2,1 × 50 мм, внутренний диаметр 3 мкм). Подвижные фазы состояли из 20 мМ ацетата аммония в 90% ацетонитриле (A) и 20 мМ ацетата аммония в 10% ацетонитриле (B). Способ отделения предусматривал следующее: начальный градиент 6% B с последующим градиентом до 8,5% B в течение 600 секунд, градиентом до 13% B в течение 400 секунд с последующим градиентом от 20% B в течение 60 секунд, промывка при 50% B в течение 60 секунд и, наконец, повторное уравновешивание при 6% B в течение 220 секунд. Количественное определение выполняли при электроионизации распылением (ESI) в режиме регистрации отрицательных ионов с использованием следующих масс-спектроскопических переходов: 2’3’5’-AMФ: 346,1-134,1, 2’3’5’-УМФ: 323,0-97, 2’3’5’-ЦМФ: 322-97, 2’3’5’-ГМФ: 362,1-211. Площади пиков сравнивали со стандартными кривыми, предусматривающими очищенные соединения (приобретенные у компании Sigma-Aldrich, за исключением 2’- и 3’-ЦМФ, УМФ и ГМФ, которые приобретали в Biolog Life Science Institute), и нормализовали по внутреннему стандарту (адипиновая кислота), который вносили в образцы перед анализом. Для анализа образцов стандартные кривые получали по деионизированной воде, разбавляли внутренним стандартом и фильтровали, как в стадиях подготовки образцов, описываемых в настоящем документе.
Результаты
РНК V. natriegens расщепляли с использованием очищенной РНКазы R E. coli, а РНК E. coli и полученный из дрожжей экстракт РНК расщепляли с использованием как РНКазыR E. coli, так и нуклеазы P1. За деполимеризацией наблюдали по высвобождению растворимых в кислоте нуклеотидов. Обработка РНК E. coli и V. natriegens с помощью РНКазы R показывала зависимое от времени превращение РНК в растворимые в кислоте нуклеотиды, достигающие ~98 - ~100% деполимеризации через 30 минут инкубации (фиг. 12A). Обработка полученного из дрожжей экстракта РНК достигала ~94% деполимеризации с нуклеазой P1 (фиг. 12A). Также обработка РНК E. coli РНК с помощью нуклеазы P1 давала в результате ~90% деполимеризацию (фиг. 12A). Последующие анализы с помощью LC-MS демонстрировали высвобождение 5’-НМФ в РНК E. coli и полученном из дрожжей экстракте РНК, обрабатываемых с помощью нуклеазы P1 (фиг. 12B).
Эти результаты демонстрируют, что варьирующие источники РНК (например, РНК Vibrio, РНК E. coli и дрожжевая РНК) могут быть расщеплены до 5’-НМФ с использованием различных нуклеаз (например, РНКазы R и нуклеазы P1).
Пример 5. Эффекты температуры и инактивации лизата в отношении деполимеризации РНК
Материалы и способы
Штаммы и лизаты
Штамм GL17-086 (BL21(DE3).ΔtolC.Δph[DE3]1+2*.Δph[285p]*.ΔfhuA*.ΔlamB*. РНК::tolC) и штамм GL17-109 (BL21(DE3). ΔtolC.Δph[DE3]1+2*.Δph[285p]*.ΔfhuA* .ΔlamB*. Δrna*. ΔphoA*.ΔappA*.Δamn*.ΔnagD*.ΔushA::tolC) использовали в исследовании, описываемом в настоящем документе. Для обоих штаммов 1 л культур выращивали в условия периодического выращивания в среде KORZ (5 г/л (NH4)2SO4, 15,7 г/л K2HPO4, 4,2 г/л KH2PO4, 1,7 г/л лимонной кислоты, 0,6 г/л MgSO4, 0,1% тиамин-HCl, 0,01% плюроника, следовые количества металлов и 40 г/л глюкозы) в течение приблизительно семи часов. После выращивания клетки центрифугировали при 6000 g в течение 20 минут, а затем хранили при -80°C. Замороженную биомассу размораживали в 1,5× объеме 58,8 мМ раствора двухосновного фосфата калия и лизировали путем пропускания через гомогенизатор EmulsiFlex C3 (Avestin) на протяжении 3 циклов с рециркулированием с импульсами 15000-20000 фунтов на квадратный дюйм. Лизат осветляли путем откручивания при 15000 g в течение 1 часа при 4°C. Лизат затем хранили при -80°C в аликвотах для одного применения.
Анализ продуктов полимеризации РНК при различных температурах
Замороженные лизаты для обоих штаммов (086 и 109) инкубировали при варьирующих температурах для профилирования продуктов деполимеризации РНК и их потенциального разложения. Аликвоты лизата размораживали на льду, а затем распределяли в стрипованные пробирки для ПЦР. Затем пробирки инкубировали при 40°C, 50°C, 60°C и 70°C до часа с периодическим отбором образцов. За начальную временную точку t0 брали гашение, когда лизаты еще были на льду. Во все другие моменты времени образцы гасили в 5× объемах ацетонитрила с разбавлением 60 мкл до 500 мкл 50 мкM раствора адипиновой кислоты в 50 мМ ацетата аммония, а затем прогоняли на LC-QQQ. С использованием MRM все четыре основных нуклеотида и связанные с ними производные (АТФ, АДФ, AMФ, аденин, аденозин, ГТФ, ГДФ, ГМФ, гуанин, гуанозин, ЦТФ, ЦДФ, ЦМФ, цитидин, цитозин, УТФ, УДФ, УМФ, уридин и урацил) определяли количественно в каждой временной точке. Гидролиз лизатов основанием, при котором РНК полностью гидролизовалась до НМФ, выполняли с помощью инкубирования лизата в 3× объемах 0,2 M NaOH при 99°C в течение 20 минут. Затем нейтрализовали с помощью равного объема 150 мМ HCl и 20 мкл этого раствора разбавляли до 200 мкл 50 мМ раствора ацетата аммония с 50 мкM адипиновой кислотой. Объемы гидролизованного основанием лизата представляли 100% деполимеризации. В некоторых условиях эти лизаты инкубировали как есть, а в других условиях лизаты смешивали либо с 0,5 мг/мл нуклеазы P1 (Sigma N8630), либо с РНКазой R. РНКазу R готовили следующим образом: клетки повторно суспендировали в 1,5× объеме буфера для лизиса (50 мМ фосфата калия, pH = 7,4, 500 мМ NaCl, 20 мМ имидазола), лизировали в гомогенизаторе EmulsiFlex C3 (Avestin) за три цикла при 15000-25000 фунтов на квадратный дюйм, осветляли в течение одного часа при 16000 g, 4°C. Затем надосадочную жидкость очищали с помощью FPLC, затем диализовали в течение ночи в 2× PBS. Осажденный белок после диализа извлекали путем добавления 500 мМ NaCl, смешанного с глицерином, с получением 50% раствора глицерина для аликвотирования и хранения при -20°C.
Анализ эффектов разбавления и эффектов ослабления предварительным нагреванием в отношении деполимеризации РНК
Эксперименты осуществляли с лизатом GL17-109, полученным, как описывается в настоящем документе. Лизаты получали при широком диапазоне условий. Обычно при всех условиях реакции получали добавленный 150 мМ фосфат калия (рН 7,4), 100 мМ KCl и 0,1 мМ ZnCl2, и все следующие условия тестировали с добавлением РНКазы R, нуклеазы P1 или без добавления экзогенной нуклеазы. Оба стоковых раствора нуклеаз готовили, как описано ранее. Реакции деполимеризации проводили при 80% и 50% разбавлениях лизата в воде. Их подвергали скринингу с 0,5 или 0,31 мг/мл, соответственно, с добавленной экзогенной нуклеазой. Лизатные смеси также готовили, получая лизат нуклеазы 1 мг/мл и смешивая его с лизатом без добавления с получением 0,064 и 0,04 мг/мл нуклеазы в условиях 80% разбавления или 0,04 и 0,025 мг/мл нуклеазы в условиях 50% разбавления. Наконец, протестировали условия, при которых лизаты без добавления нуклеазы подвергали ослаблению нагреванием при 70°С в течение 15 минут, а затем смешивали либо с очищенной нуклеазой, либо с все еще активным лизатом нуклеазы 1 мг/мл. Образцы инкубировали при 37°С в течение 30 минут и отбирали образцы путем гашения в 5× объемах ацетонитрила и разбавления до 1 мл 50 мкМ раствора адипиновой кислоты в 50 мМ ацетата аммония. Затем гашенный раствор центрифугировали, фильтровали через 0,2-мкм фильтровальную пластину и фильтрат прогоняли на LC-QQQ для измерения НМФ, нуклеозидов и нуклеиновых оснований.
Результаты
Известно, что РНК-нуклеазы (РНКазы) имеют профили активности, которые охватывают температурные профили более широко, чем многие другие ферменты из мезофильных источников, иногда проявляя активность до 60°C, несмотря на то, что они происходят из организма, эволюционировавшего для роста при 37°C. Чтобы оценить влияние повышенных температур на деполимеризацию РНК в лизате E. coli, проводили два отдельных исследования. Сначала лизаты из двух отдельных штаммов инкубировали при температуре выше 37°С в течение часа с добавлением или без добавления одной из двух РНКаз - РНКазы R или РНКазы P1. Второе исследование предусматривало элиминирование лизата для удаления вредных ферментативных активностей и смешивание этого неактивного лизата с активным лизатом для изучения потенциальных воздействий на деполимеризацию РНК или смешивание экзогенных нуклеаз с инактивированным лизатом.
Улучшенную деполимеризацию наблюдали, когда лизаты инкубировали при 60°C и 70°C (фиг. 13). Деполимеризация улучшалась несколькими путями. При деполимеризации РНК преимущественно продуцируются НМФ. Однако в активном лизате эти NMP продолжают разлагаться другими ферментами с очень большим накоплением двух основных продуктов - гуанозина и урацила. Инкубирование лизатов при 60°C и 70°C значительно снижало накопление этих нуклеиновых оснований, даже в лизатах, которые не получали экзогенную РНКазу R или нуклеазу Р1 (фиг. 13). Поскольку каждый моль нуклеинового основания коррелирует с необратимой потерей НМФ из РНК, это продемонстрировало улучшенную стабильность НМФ в лизатах. Кроме того, когда в этих температурных условиях добавляют экзогенную нуклеазу, наблюдается значительное улучшение накопления суммарного НМФ, как показано на фиг. 13, при 70°С с добавлением РНКазы R. Гидролиз основанием лизата GL17-109 показал максимальную концентрацию 32,6 мМ НМФ. С поправкой на разбавление добавленной нуклеазы накопление ~22 мМ НМФ, наблюдаемое при 70°C с РНКазой R, составляет выход 75%. Принимая во внимание то, что тРНК представляет приблизительно 15% пула суммарной РНК, и что она недоступна для этих нуклеаз, это будет составлять 94% выход всех доступных РНК. Деполимеризация в GL17-086 была хуже по всем показателям, чем GL17-109, вероятно, из-за большей фосфорилитической активности (данные не показаны).
Влияния ослабления предварительным нагреванием и разбавлений лизата показывали небольшое преимущество, но только при сильнее разбавленном лизате. В этом исследовании получали два типа лизатов - лизат «реагент», который содержит РНК, подлежащую деполимеризации, и лизат «катализатор», содержащий экзогенную нуклеазу (РНКазу R или нуклеазу P1). Многие разные условия оценивали, как показано в приведенной ниже таблице 14, чтобы определить их влияние на деполимеризацию РНК. Лизаты, которые разбавляли только до 80%, показывали сходные характеристики деполимеризации, независимо от того, были ли части лизата ослаблены нагреванием или нет. Однако лизаты, которые разбавляли до 50%, показывали себя немного лучше, если большая часть РНК была в инактивированном лизате. Во всех условиях, тестируемых при этом разбавлении, наблюдали улучшение среднего выхода деполимеризации на 2,7%. Характеристика реакции деполимеризации в этих условиях не показывает существенного улучшения, но является эквивалентной или только немного лучше, что оставляет возможность открытой для осуществления ослабления деполимеризации предварительным нагреванием, если этого требуют другие части процесса, например, прекращение роста каких-либо нелизированных клеток, которые могут оставаться в культуре, или снижение содержания белка в лизате, если стадия гранулирования предусматривается после тепловой обработки.
Таблица 14. Краткое описание выходов деполимеризации РНК в варьирующих смесях лизатов, нуклеаз и инактивированных лизатов. Процентные выходы представлены на основе 32,6 мМ НМФ, как определяли количественно посредством гидролиза основанием РНК в лизате.
Пример 6. Бесклеточное получение РНК с использованием различных источников нуклеотидов
Материалы и способы
Порошок дрожжевой РНК, полученный из коммерческих источников, растворяли в воде при 45-60 г/л и деполимеризовали с использованием 1,2 г/л нуклеазы P1 при 70°C, pH 5,5-5,8, в течение 1 часа в присутствии 0,05 мМ хлорида цинка. Полученный в результате деполимеризованный материал осветляли с помощью центрифугирования и фильтровали с использованием фильтра 10 кДа MWCO. Полученный в результате поток содержал 5’-нуклеотидмонофосфаты (НМФ) при суммарной концентрации ~90-100 мМ (~20-25 мМ каждого из AMФ, ЦМФ, ГМФ и УМФ).
Производные E. coli BL21(DE3), несущие полученные из pBAD24 векторы, кодирующие отдельные киназные ферменты (TthCmk, PfPyrH, TmGmk, AaNdk и DgPPK2), культивировали при ферментациях со средой Korz, дополненной 50 мг/л карбенициллина, с использованием стандартных методик [Korz, D. J., Rinas, U., Hellmuth, K., Sanders, E. A., & Deckwer, W. D. (1995). Simple fed-batch technique for high cell density cultivation of Escherichia coli. Journal of biotechnology, 39(1), 59-65]. Экспрессию белка индуцировали путем добавления L-арабинозы. После сбора готовили лизаты в 60 мМ фосфатном буфере с использованием гомогенизации под высоким давлением с получением смесей приблизительно 40 г/л суммарного белка.
Производные E. coli BL21(DE3), несущие полученные из pUC19 векторы, содержащие одну или несколько транскрипционных матриц (при этом каждая состоит из T7 промотора, целевой последовательности и одного или нескольких транскрипционных терминаторов), кодирующих последовательность двухнитевой РНК 524 п. о. Клетки культивировали при ферментациях со средой Korz с использованием стандартных методик [Phue, J. N., Lee, S. J., Trinh, L., & Shiloach, J. (2008). Modified Escherichia coli B (BL21), a superior producer of plasmid DNA compared with Escherichia coli K (DH5alpha). Biotechnology and bioengineering, 101(4), 831]. После сбора создавали лизаты гомогенизацией под высоким давлением, разбавляли и обрабатывали нагреванием в соответствии с подобными процедурами.
Производные E. coli BL21(DE3), несущие полученные из pBAD24 векторы, кодирующие ферменты термостабильной T7 РНК-полимеразы, культивировали, индуцировали экспрессию ферментов и получали лизаты с использованием подобных процедур. Полимеразные ферменты частично очищали с использованием двух стадий фракционирования с сульфатом аммония.
Реакционные смеси собирали в 30 мМ фосфатном буфере, pH 7, согласно таблице 15.
Таблица 15. Условия реакции
При сборке бесклеточных реакционных смесей лизаты, содержащие киназные ферменты и ДНК-матрицу, разбавляли, объединяли в равной пропорции и смешивали с реакционными добавками, такими как сульфат магния и гексаметафосфат натрия. Лизаты инкубировали при 70°C в течение 15 минут для инактивации других ферментативных активностей при сохранении активностей надэкспрессируемых киназ. Бесклеточные реакции инициировали путем добавления РНК-полимеразы, инкубировали при 48°C в течение 1 часа и анализировали в соответствии с установленными протоколами [Nwokeoji, A. O., Kilby, P. M., Portwood, D. E., & Dickman, M. J. (2016). RNASwift: A rapid, versatile RNA extraction method free from phenol and chloroform. Analytical biochemistry, 512, 36].
Результаты
Бесклеточные реакции синтеза РНК осуществляли с использованием клеточной РНК, эквимолярной смеси нуклеозид-5’-монофосфатов (AMФ, ЦМФ, ГМФ, УМФ) или эквимолярной смеси 5’-нуклеозиддифосфатов (АДФ, ЦДФ, ГДФ, УДФ). Подобные титры dsRNA-продукта получали для каждого источника нуклеотидов (фиг. 17).
Эти результаты демонстрируют, что бесклеточные реакции, описываемые в настоящем документе, могут быть использованы для синтеза РНК из многих источников нуклеотидов, в том числе клеточной РНК, нуклеозид-5’-монофосфатов и нуклеозиддифосфатов.
Пример 7. Бесклеточное получение РНК с использованием РНК-полимеразы дикого типа
Материалы и способы
Производные E. coli BL21(DE3), несущие полученные из pBAD24 векторы, кодирующие меченные гексагистидином ферменты термостабильной или дикого типа T7 РНК-полимеразы, культивировали, индуцировали экспрессию фермента и получали лизаты с использованием процедур, описываемых в настоящем документе. Полимеразные ферменты очищали с помощью жидкостной экспресс-хроматографии белков (FPLC), как описывается в настоящем документе. Бесклеточные реакции осуществляли, как описывается в настоящем документе, за исключением того, что реакции осуществляли в диапазоне температур (37-48°C) в течение 2 часов. Титры dsRNA-продукта определяли количественно, как описывается в настоящем документе.
Результаты
Бесклеточные реакции синтеза РНК осуществляли с использованием T7 РНК-полимеразы дикого типа или термостабильного мутанта (фиг. 18). Реакции выполняли с продуцируемым термостабильным мутантом dsRNA-продуктом при 37°C и 48°C. Для сравнения реакции выполняли с продуцируемым полимеразой дикого типа продуктом при 37°C, но не 48°C.
Эти результаты демонстрируют, что для бесклеточных реакций, описываемых в настоящем документе, не требуются термостабильные РНК-полимеразы, при условии инкубирования при соответствующих температурах.
Пример 8. Бесклеточное получение НТФ с использованием различных источников нуклеотидов
Материалы и способы
Бесклеточные реакции осуществляли, как описано в примере 6, за исключением того, что не использовали лизат ДНК-матрицы и РНК-полимеразу. Нуклеотиды анализировали с помощью HPLC с использованием адаптации опубликованных способов [de Korte, D., Haverkort, W. A., Roos, D., & van Gennip, A. H. (1985). Anion-exchange high performance liquid chromatography method for the quantitation of nucleotides in human blood cells. Clinica chimica acta; international journal of clinical chemistry, 148(3), 185].
Результаты
Бесклеточные реакции для получения нуклеотид-5’-трифосфатов (НTФ: АТФ, ЦТФ, ГТФ и УТФ) осуществляли с использованием клеточной РНК, эквимолярной смеси нуклеозид-5’-монофосфатов (AMФ, ЦМФ, ГМФ, УМФ) или эквимолярной смеси 5’-нуклеозиддифосфатов (АДФ, ЦДФ, ГДФ, УДФ). Подобные титры каждого НТФ получали для каждого источника нуклеотида (фиг. 19).
Эти результаты демонстрируют, что бесклеточные реакции, описываемые в настоящем документе, также можно использовать для получения НТФ, просто исключая РНК-полимеразу и ДНК-матрицу. Подобно бесклеточным реакциям, продуцирующим РНК, описанным в примере 6, в бесклеточных реакциях, продуцирующих НТФ, можно использовать несколько источников нуклеотидов, в том числе клеточную РНК, нуклеозид-5’-монофосфаты и нуклеозиддифосфаты.
Ссылки
1. Maekewa K., Tsunasawa S., Dibo G., Sakiyama F. 1991. Primary structure of nuclease P1 from Penicillium citrinum. Eur. J. Biochem. 200:651-661.
2. Volbeda A., Lahm A., Sakiyama F., Suck D. 1991. Crystal structure of Penicillium citrinum P1 nuclease at 2.8-A resolution. EMBO J. 10:1607-1618(1991).
3. Romier C., Dominguez R., Lahm A., Dahl O., Suck D. 1998. Recognition of single-stranded DNA by nuclease P1: high resolution crystal structures of complexes with substrate analogs. Proteins 32:414-424.
4. Cheng Z.F., Deutscher M.P. 2002. Purification and characterization of the Escherichia coli exoribonuclease RNase R. Comparison with RNase II. J. Biol. Chem. 277:21624-21629.
5. Zilhao R., Camelo L., Arraiano C.M. 1993. DNA sequencing and expression of the gene rnb encoding Escherichia coli ribonuclease II. Mol. Microbiol. 8:43-51.
6. March P.E., Ahnn J., Inouye M. 1985. The DNA sequence of the gene (rnc) encoding ribonuclease III of Escherichia coli. Nucleic Acids Res. 13:4677-4685.
7. Chen S.M., Takiff H.E., Barber A.M., Dubois G.C., Bardwell J.C., Court D.L. 1990. Expression and characterization of RNase III and Era proteins. Products of the rnc operon of Escherichia coli. J. Biol. Chem. 265:2888-2895.
8. Robertson H.D., Webster R.E., Zinder N.D. 1968. Purification and properties of ribonuclease III from Escherichia coli. J. Biol. Chem. 243:82-91.
9. Molina L., Bernal P., Udaondo Z., Segura A., Ramos J.L.2013. Complete Genome Sequence of a Pseudomonas putida Clinical Isolate, Strain H8234. Genome Announc. 1:E00496-13; and Cheng, Z.F. and M.P. Deutscher. 2002. Purification and characterization of the Escherichia coli exoribonuclease RNAse R. Comparison with RNAse II. J Biol Chem. 277(24).
10. Even S., Pellegrini O., Zig L., Labas V., Vinh J., Brechemmier-Baey D., Putzer H. 2005. Ribonucleases J1 and J2: two novel endoribonucleases in B. subtilis with functional homology to E. coli RNase E. Nucleic Acids Res. 33:2141-2152.
11. Li de la Sierra-Gallay I., Zig L., Jamalli A., Putzer H. 2008. Structural insights into the dual activity of RNase J. Nat. Struct. Mol. Biol. 15:206-212.
12. Ball T.K., Saurugger P.N., Benedick M.J. 1987. The extracellular nuclease gene of Serratia marcescens and its secretion from Escherichia coli. Gene 57:183-192.
13. Biedermann K., Jepsen P.K., Riise E., Svendsen I. 1989. Purification and characterization of a Serratia marcescens nuclease produced by Escherichia coli. Carlsberg Res. Commun. 54:17-27.
14. Shlyapnikov S.V., Lunin V.V., Perbandt M., Polyakov K.M., Lunin V.Y., Levdikov V.M., Betzel C., Mikhailov A.M. 2000. Atomic structure of the Serratia marcescens endonuclease at 1.1 A resolution and the enzyme reaction mechanism. Acta Crystallogr. D 56:567-572.
15. Zuo Y., Deutscher M.P. 2002. Mechanism of action of RNase T. I. Identification of residues required for catalysis, substrate binding, and dimerization. J. Biol. Chem. 277:50155-50159.
16. Zuo Y., Zheng H., Wang Y., Chruszcz M., Cymborowski M., Skarina T., Savchenko A., Malhotra A., Minor W. 2007. Crystal structure of RNase T, an exoribonuclease involved in tRNA maturation and end turnover. Structure 15:417-428.
17. Huang S., Deutscher M.P. 1992. Sequence and transcriptional analysis of the Escherichia coli rnt gene encoding RNase T. J. Biol. Chem. 267:25609-25613.
18. Chauhan A.K., Miczak A., Taraseviciene L., Apirion D. 1991. Sequencing and expression of the rne gene of Escherichia coli. Nucleic Acids Res. 19:125-129.
19. Cormack R.S., Genereaux J.L., Mackie G.A. 1993. RNase E activity is conferred by a single polypeptide: overexpression, purification, and properties of the ams/rne/hmp1 gene product. Proc. Natl. Acad. Sci. U.S.A. 90:9006-9010.
20. Motomura, K., Hirota, R., Okada, M., Ikeda, T., Ishida, T., & Kuroda, A. (2014). A New Subfamily of Polyphosphate Kinase 2 (Class III PPK2) Catalyzes both Nucleoside Monophosphate Phosphorylation and Nucleoside Diphosphate Phosphorylation. Applied and Environmental Microbiology, 80(8), 2602-2608. http://doi.org/10.1128/AEM.03971-13.
21. Elkin, S. R., Kumar, A., Price, C. W., & Columbus, L. (2013). A Broad Specificity Nucleoside Kinase from Thermoplasma acidophilum. Proteins, 81(4), 568-582. doi.org/10.1002/prot.24212.
22. Hansen, T., Arnfors, L., Ladenstein, R.,

23. Ota, H., Sakasegawa, S., Yasuda, Y., Imamura, S., & Tamura, T. (2008). A novel nucleoside kinase from Burkholderia thailandensis: a member of the phosphofructokinase B-type family of enzymes. The FEBS journal, 275(23), 5865.
24. Tomoike, F., Nakagawa, N., Kuramitsu, S., & Masui, R. (2011). A single amino acid limits the substrate specificity of Thermus thermophilus uridine-cytidine kinase to cytidine. Biochemistry, 50(21), 4597.
25. Henne A., Brueggemann H., Raasch C., Wiezer A., Hartsch T., Liesegang H., Johann A., Lienard T., Gohl O., Martinez-Arias R., Jacobi C., Starkuviene V., Schlenczeck S., Dencker S., Huber R., Klenk H.-P., Kramer W., Merkl R., Gottschalk G., Fritz H.-J. 2004. The genome sequence of the extreme thermophile Thermus thermophilus. Nat. Biotechnol. 22:547-553.
26. Tan ZW, Liu J, Zhang XF, Meng FG, Zhang YZ.Nan Fang Yi Ke Da Xue Xue Bao. 2010. Expression, purification and enzymatic characterization of adenylate kinase of Thermus thermophilus HB27 in Escherichia coli.Jan;30(1):1-6.
27. Maeder D.L., Weiss R.B., Dunn D.M., Cherry J.L., Gonzalez J.M., DiRuggiero J., Robb F.T. 1999. Divergence of the hyperthermophilic archaea Pyrococcus furiosus and P. horikoshii inferred from complete genomic sequences. Genetics 152:1299-1305.
28.
29.
30.

31. Feil, H., Feil, W. S., Chain, P., Larimer, F., DiBartolo, G., Copeland, A., … Lindow, S. E. (2005). Comparison of the complete genome sequences of Pseudomonas syringae pv. syringae B728a and pv. tomato DC3000. Proceedings of the National Academy of Sciences of the United States of America, 102(31), 11064-11069. doi.org/10.1073/pnas.0504930102.
32. Song, S., Inouye, S., Kawai, M., Fukami-Kobayashi, K., Gδ, M., & Nakazawa, A. (1996). Cloning and characterization of the gene encoding Halobacterium halobium adenylate kinase. Gene, 175(1), 65-70.
33. Masui R., Kurokawa K., Nakagawa N., Tokunaga F., Koyama Y., Shibata T., Oshima T., Yokoyama S., Yasunaga T., Kuramitsu S. Complete genome sequence of Thermus thermophilus HB8. Submitted (NOV-2004) to the EMBL/GenBank/DDBJ databases.
34. Ng, W. V., Kennedy, S. P., Mahairas, G. G., Berquist, B., Pan, M., Shukla, H. D., … DasSarma, S. (2000). Genome sequence of Halobacterium species NRC-1. Proceedings of the National Academy of Sciences of the United States of America, 97(22), 12176-12181.
35. Marco-Marin C., Escamilla-Honrubia J.M., Rubio V. 2005. First-time crystallization and preliminary X-ray crystallographic analysis of a bacterial-archaeal type UMP kinase, a key enzyme in microbial pyrimidine biosynthesis. Biochim. Biophys. Acta 1747:271-275.
36. Marco-Marin C., Escamilla-Honrubia J.M., Rubio V. 2005. First-time crystallization and preliminary X-ray crystallographic analysis of a bacterial-archaeal type UMP kinase, a key enzyme in microbial pyrimidine biosynthesis. Biochim. Biophys. Acta 1747:271-275.
37. Jensen, K. S., Johansson, E., & Jensen, K. F. (2007). Structural and enzymatic investigation of the Sulfolobus solfataricus uridylate kinase shows competitive UTP inhibition and the lack of GTP stimulation. Biochemistry, 46(10), 2745-2757.
38. Nelson K.E., Clayton R.A., Gill S.R., Gwinn M.L., Dodson R.J., Haft D.H., Hickey E.K., Peterson J.D., Nelson W.C., Ketchum K.A., McDonald L.A., Utterback T.R., Malek J.A., Linher K.D., Garrett M.M., Stewart A.M., Cotton M.D., Pratt M.S. Fraser C.M. 1999. Evidence for lateral gene transfer between Archaea and Bacteria from genome sequence of Thermotoga maritima. Nature 399:323-329.
39. Riley, M., Staley, J. T., Danchin, A., Wang, T. Z., Brettin, T. S., Hauser, L. J., … Thompson, L. S. (2008). Genomics of an extreme psychrophile, Psychromonas ingrahamii. BMC Genomics, 9, 210. doi.org/10.1186/1471-2164-9-210.
40. Ishibashi, M., Tokunaga, H., Hiratsuka, K., Yonezawa, Y., Tsurumaru, H., Arakawa, T., & Tokunaga, M. (2001). NaCl-activated nucleoside diphosphate kinase from extremely halophilic archaeon, Halobacterium salinarum, maintains native conformation without salt. FEBS letters, 493(2-3), 134.
41. Polosina, Y. Y., Zamyatkin, D. F., Kostyukova, A. S., Filimonov, V. V., & Fedorov, O. V. (2002). Stability of Natrialba magadii NDP kinase: comparisons with other halophilic proteins. Extremophiles: life under extreme conditions, 6(2), 135.
42. Polosina, Y. Y., Zamyatkin, D. F., Kostyukova, A. S., Filimonov, V. V., & Fedorov, O. V. (2002). Stability of Natrialba magadii NDP kinase: comparisons with other halophilic proteins. Extremophiles: life under extreme conditions, 6(2), 135.
43. Udaondo, Z., Molina, L., Daniels, C.,

44.

45. Brune, M., Schumann, R., & Wittinghofer, F. (1985). Cloning and sequencing of the adenylate kinase gene (adk) of Escherichia coli. Nucleic Acids Research, 13(19), 7139-7151.
46. Pel, H. J., de Winde, J. H., Archer, D. B., Dyer, P. S., Hofmann, G., Schaap, P. J., … & Andersen, M. R. (2007). Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nature biotechnology, 25(2), 221.
47. Magdolen, V., Oechsner, U., & Bandlow, W. (1987). The complete nucleotide sequence of the gene coding for yeast adenylate kinase. Current genetics, 12(6), 405.
48. Pedersen, S., Skouv, J., Kajitani, M., & Ishihama, A. (1984). Transcriptional organization of the rpsA operon of Escherichia coli. Molecular & general genetics: MGG, 196(1), 135.
49. Smallshaw, J., & Kelln, R. A. (1992). Cloning, nucleotide sequence and expression of the Escherichia coli K-12 pyrH gene encoding UMP kinase. Genetics (Life Sci. Adv.), 11, 59-65.
50. Liljelund, P., Sanni, A., Friesen, J. D., & Lacroute, F. (1989). Primary structure of the S. cerevisiae gene encoding uridine monophosphokinase. Biochemical and biophysical research communications, 165(1), 464.
51. Gentry, D., Bengra, C., Ikehara, K., & Cashel, M. (1993). Guanylate kinase of Escherichia coli K-12. The Journal of biological chemistry, 268(19), 14316.
52. Konrad, M. (1992). Cloning and expression of the essential gene for guanylate kinase from yeast. The Journal of biological chemistry, 267(36), 25652.
53. Hama, H., Almaula, N., Lerner, C. G., Inouye, S., & Inouye, M. (1991). Nucleoside diphosphate kinase from Escherichia coli; its overproduction and sequence comparison with eukaryotic enzymes. Gene, 105(1), 31.
54. Besir, H., Zeth, K., Bracher, A., Heider, U., Ishibashi, M., Tokunaga, M., & Oesterhelt, D. (2005). Structure of a halophilic nucleoside diphosphate kinase from Halobacterium salinarum. FEBS letters, 579(29), 6595.
55. Deutscher, M. & Reuven N. (1991). Enzymatic basis for hydrolytic versus phosphorolytic mRNA degradation in Escherichia coli and Bacillus subtilis. PNAS, 88, 3277-3280.
56. Nwokeji, A. O., Kilby, P. M., Portwood, D. E., & Dickman, M. J. (2016). RNASwift: A rapid, versatile RNA extraction method free from phenol and chloroform. Analytical Biochemistry, 512, 36-46.
57. Mohanty, B. K., Giladi, H., Maples, V. F., & Kushner, S. R. (2008). Analysis of RNA decay, processing, and polyadenylation in Escherichia coli and other prokaryotes. Methods in Enzymology, 447, 3-29.
58. Korz, D. J., Rinas, U., Hellmuth, K., Sanders, E. A., & Deckwer, W. D. (1995). Simple fed-batch technique for high cell density cultivation of Escherichia coli. Journal of biotechnology, 39(1), 59-654.
59. Phue, J. N., Lee, S. J., Trinh, L., & Shiloach, J. (2008). Modified Escherichia coli B (BL21), a superior producer of plasmid DNA compared with Escherichia coli K (DH5alpha). Biotechnology and bioengineering, 101(4), 831.
60. de Korte, D., Haverkort, W. A., Roos, D., & van Gennip, A. H. (1985). Anion-exchange high performance liquid chromatography method for the quantitation of nucleotides in human blood cells. Clinica chimica acta; international journal of clinical chemistry, 148(3), 185.]
Последовательности
Deinococcus geothermalis DSM 11300 PPK2
MQLDRYRVPPGQRVRLSNWPTDDDGGLSKAEGEALLPDLQQRLANLQERLYAESQQALLIVLQARDAGGKDGTVKHVIGAFNPSGVQVSNFKVPTEEERAHDFLWRIHRQTPRLGMIGVFNRSQYEDVLVTRVHHLIDDQTAQRRLKHICAFESLLTDSGTRIVKFYLHISPEEQKKRLEARLADPSKHWKFNPGDLQERAHWDAYTAVYEDVLTTSTPAAPWYVVPADRKWFRNLLVSQILVQTLEEMNPQFPAPAFNAADLRIV (SEQ ID NO: 1)
Meiothermus ruber DM 1279 PPK2
MGFCSIEFLMGAQMKKYRVQPDGRFELKRFDPDDTSAFEGGKQAALEALAVLNRRLEKLQELLYAEGQHKVLVVLQAMDAGGKDGTIRVVFDGVNPSGVRVASFGVPTEQELARDYLWRVHQQVPRKGELVIFNRSHYEDVLVVRVKNLVPQQVWQKRYRHIREFERMLADEGTTILKFFLHISKDEQRQRLQERLDNPEKRWKFRMGDLEDRRLWDRYQEAYEAAIRETSTEYAPWYVIPANKNWYRNWLVSHILVETLEGLAMQYPQPETASEKIVIE (SEQ ID NO: 2)
Meiothermus silvanus DSM 9946 PPK2
MAKTIGATLNLQDIDPRSTPGFNGDKEKALALLEKLTARLDELQEQLYAEHQHRVLVILQGMDTSGKDGTIRHVFKNVDPLGVRVVAFKAPTPPELERDYLWRVHQHVPANGELVIFNRSHYEDVLVARVHNLVPPAIWSRRYDHINAFEKMLVDEGTTVLKFFLHISKEEQKKRLLERLVEADKHWKFDPQDLVERGYWEDYMEAYQDVLDKTHTQYAPWHVIPADRKWYRNLQVSRLLVEALEGLRMKYPRPKLNIPRLKSELEKM (SEQ ID NO: 3)
Thermosynechococcus elongatus BP-1 PPK2
MIPQDFLDEINPDRYIVPAGGNFHWKDYDPGDTAGLKSKVEAQELLAAGIKKLAAYQDVLYAQNIYGLLIIFQAMDAAGKDSTIKHVMSGLNPQACRVYSFKAPSAEELDHDFLWRANRALPERGCIGIFNRSYYEEVLVVRVHPDLLNRQQLPPETKTKHIWKERFEDINHYERYLTRNGILILKFFLHISKAEQKKRFLERISRPEKNWKFSIEDVRDRAHWDDYQQAYADVFRHTSTKWAPWHIIPANHKWFARLMVAHFIYQKLASLNLHYPMLSEAHREQLLEAKALLENEPDED (SEQ ID NO: 4)
Anaerolinea thermophila UNI-1 PPK2
MGEAMERYFIKPGEKVRLKDWSPDPPKDFEGDKESTRAAVAELNRKLEVLQERLYAERKHKVLVILQGMDTSGKDGVIRSVFEGVNPQGVKVANFKVPTQEELDHDYLWRVHKVVPGKGEIVIFNRSHYEDVLVVRVHNLVPPEVWKKRYEQINQFERLLHETGTTILKFFLFISREEQKQRLLERLADPAKHWKFNPGDLKERALWEEYEKAYEDVLSRTSTEYAPWILVPADKKWYRDWVISRVLVETLEGLEIQLPPPLADAETYRRQLLEEDAPESR (SEQ ID NO: 5)
Caldilinea aerophila DSM 14535 PPK2
MDVDRYRVPPGSTIHLSQWPPDDRSLYEGDKKQGKQDLSALNRRLETLQELLYAEGKHKVLIILQGMDTSGKDGVIRHVFNGVNPQGVKVASFKVPTAVELAHDFLWRIHRQTPGSGEIVIFNRSHYEDVLVVRVHGLVPPEVWARRYEHINAFEKLLVDEGTTILKFFLHISKEEQRQRLLERLEMPEKRWKFSVGDLAERKRWDEYMAAYEAVLSKTSTEYAPWYIVPSDRKWYRNLVISHVIINALEGLNMRYPQPEDIAFDTIVIE (SEQ ID NO: 6)
Chlorobaculum tepidum TLS PPK2
MKLDLDAFRIQPGKKPNLAKRPTRIDPVYRSKGEYHELLANHVAELSKLQNVLYADNRYAILLIFQAMDAAGKDSAIKHVMSGVNPQGCQVYSFKHPSATELEHDFLWRTNCVLPERGRIGIFNRSYYEEVLVVRVHPEILEMQNIPHNLAHNGKVWDHRYRSIVSHEQHLHCNGTRIVKFYLHLSKEEQRKRFLERIDDPNKNWKFSTADLEERKFWDQYMEAYESCLQETSTKDSPWFAVPADDKKNARLIVSRIVLDTLESLNLKYPEPSPERRKELLDIRKRLENPENGK (SEQ ID NO: 7)
Oceanithermus profundus DSM 14977 PPK2
MDVSRYRVPPGSGFDPEAWPTREDDDFAGGKKEAKKELARLAVRLGELQARLYAEGRQALLIVLQGMDTAGKDGTIRHVFRAVNPQGVRVTSFKKPTALELAHDYLWRVHRHAPARGEIGIFNRSHYEDVLVVRVHELVPPEVWGRRYDHINAFERLLADEGTRIVKFFLHISKDEQKRRLEARLENPRKHWKFNPADLSERARWGDYAAAYAEALSRTSSDRAPWYAVPADRKWQRNRIVAQVLVDALEAMDPRFPRVDFDPASVRVE (SEQ ID NO: 8)
Roseiflexus castenholzii DSM 13941 PPK2
MYAQRVVPGMRVRLHDIDPDANGGLNKDEGRARFAELNAELDVMQEELYAAGIHALLLILQGMDTAGKDGAIRNVMLNLNPQGCRVESFKVPTEEELAHDFLWRVHRVVPRKGMVGVFNRSHYEDVLVVRVHSLVPESVWRARYDQINAFERLLADTGTIIVKCFLHISKEEQEQRLLARERDVSKAWKLSAGDWRERAFWDDYMAAYEEALTRCSTDYAPWYIIPANRKWYRDLAISEALVETLRPYRDDWRRALDAMSRARRAELEAFRAEQHAMEGRPQGAGGVSRR (SEQ ID NO: 9)
Roseiflexus sp. RS-1 PPK2
MHYAHTVIPGTQVRLRDIDPDASGGLTKDEGRERFASFNATLDAMQEELYAAGVHALLLILQGMDTAGKDGAIRNVMHNLNPQGCRVESFKVPTEEELAHDFLWRVHKVVPRKGMVGVFNRSHYEDVLVVRVHSLVPEHVWRARYDQINAFERLLTDTGTIIVKCFLHISKDEQEKRLLAREQDVTKAWKLSAGDWRERERWDEYMAAYEEALTRCSTEYAPWYIIPANRKWYRDLAISEVLVETLRPYRDDWQRALDAMSQARLAELKAFRHQQTAGATRL (SEQ ID NO: 10)
Truepera radiovictrix DSM 17093 PPK2
MSQGSAKGLGKLDKKVYARELALLQLELVKLQGWIKAQGLKVVVLFEGRDAAGKGSTITRITQPLNPRVCRVVALGAPTERERTQWYFQRYVHHLPAAGEMVLFDRSWYNRAGVERVMGFCTEAEYREFLHACPTFERLLLDAGIILIKYWFSVSAAEQERRMRRRNENPAKRWKLSPMDLEARARWVAYSKAKDAMFYHTDTKASPWYVVNAEDKRRAHLSCIAHLLSLIPYEDLTPPPLEMPPRDLAGADEGYERPDKAHQTWVPDYVPPTR (SEQ ID NO: 11)
Thermus thermophilus Adk
MDVGQAVIFLGPPGAGKGTQASRLAQELGFKKLSTGDILRDHVARGTPLGERVRPIMERGDLVPDDLILELIREELAERVIFDGFPRTLAQAEALDRLLSETGTRLLGVVLVEVPEEELVRRILRRAELEGRSDDNEETVRRRLEVYREKTEPLVGYYEARGVLKRVDGLGTPDEVYARIRAALGI (SEQ ID NO: 12)
Thermus thermophilus Cmk
MRGIVTIDGPSASGKSSVARRVAAALGVPYLSSGLLYRAAAFLALRAGVDPGDEEGLLALLEGLGVRLLAQAEGNRVLADGEDLTSFLHTPEVDRVVSAVARLPGVRAWVNRRLKEVPPPFVAEGRDMGTAVFPEAAHKFYLTASPEVRAWRRARERPQAYEEVLRDLLRRDERDKAQSAPAPDALVLDTGGMTLDEVVAWVLAHIRR (SEQ ID NO: 13)
Pyrococcus furiosus PyrH
MRIVFDIGGSVLVPENPDIDFIKEIAYQLTKVSEDHEVAVVVGGGKLARKYIEVAEKFNSSETFKDFIGIQITRANAMLLIAALREKAYPVVVEDFWEAWKAVQLKKIPVMGGTHPGHTTDAVAALLAEFLKADLLVVITNVDGVYTADPKKDPTAKKIKKMKPEELLEIVGKGIEKAGSSSVIDPLAAKIIARSGIKTIVIGKEDAKDLFRVIKGDHNGTTIEP (SEQ ID NO: 14)
Thermotoga maritima Gmk
MKGQLFVICGPSGAGKTSIIKEVLKRLDNVVFSVSCTTRPKRPHEEDGKDYFFITEEEFLKRVERGEFLEWARVHGHLYGTLRSFVESHINEGKDVVLDIDVQGALSVKKKYSNTVFIYVAPPSYADLRERILKRGTEKEADVLVRLENAKWELMFMDEFDYIVVNENLEDAVEMVVSIVRSERAKVTRNQDKIERFKMEVKGWKKL (SEQ ID NO: 15)
Aquifex aeolicus Ndk
MAVERTLIIVKPDAMEKGALGKILDRFIQEGFQIKALKMFRFTPEKAGEFYYVHRERPFFQELVEFMSSGPVVAAVLEGEDAIKRVREIIGPTDSEEARKVAPNSIRAQFGTDKGKNAIHASDSPESAQYEICFIFSGLEIV (SEQ ID NO: 16)
Все литературные источники, патенты и заявки на выдачу патентов, раскрываемые в настоящем документе, включены посредством ссылки в отношении объекта изобретения, для которого каждые из них цитируются, что в некоторых случаях может охватывать весь документ.
Формы единственного числа, используемые в настоящем документе в описании и в формуле изобретения, если явно не указано иное, следует понимать как означающие «по меньшей мере один».
Также следует учитывать, что, если явно не указано иное, в любых способах, заявленных в настоящем документе, которые предусматривают более одной(одного) стадии или действия, порядок стадий или действий способа не обязательно ограничен порядком, в котором стадии или действия акты способа изложены.
В формуле изобретения, а также в приведенном выше описании все переходные фразы, такие как «содержащий», «включающий в себя», «несущий», «имеющий», «предусматривающий», «вовлекающий», «держащий», «состоящий из» и т.п., следует понимать как открытые, то есть означающие включающие в себя без ограничения. Только переходные фразы «состоящий из» и «состоящий по сути из» должны быть закрытыми или полузакрытыми переходными фразами, соответственно, как указано в Руководстве по процедурам патентной экспертизы Патентного ведомства США, раздел 2111.03.
Термины «приблизительно» и «по сути» предшествующие числовому значению, означают ± 10% от приведенного числового значения.
При представлении диапазона значений каждое значение между верхним и нижним границами диапазона специально рассматривается и описывается в настоящем документе.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Гринлайт Байосайенсиз, Инк.
<120> СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПОЛУЧЕНИЯ НУКЛЕОЗИДТРИФОСФАТА И
РИБОНУКЛЕИНОВОЙ КИСЛОТЫ
<130> G0830.70025WO00
<140> Еще не передано
<141> Одновременно с настоящим
<150> US 62/571,071
<151> 2017-10-11
<160> 16
<170> Патентная версия 3.5
<210> 1
<211> 266
<212> PRT
<213> Deinococcus geothermalis
<400> 1
Met Gln Leu Asp Arg Tyr Arg Val Pro Pro Gly Gln Arg Val Arg Leu
1 5 10 15
Ser Asn Trp Pro Thr Asp Asp Asp Gly Gly Leu Ser Lys Ala Glu Gly
20 25 30
Glu Ala Leu Leu Pro Asp Leu Gln Gln Arg Leu Ala Asn Leu Gln Glu
35 40 45
Arg Leu Tyr Ala Glu Ser Gln Gln Ala Leu Leu Ile Val Leu Gln Ala
50 55 60
Arg Asp Ala Gly Gly Lys Asp Gly Thr Val Lys His Val Ile Gly Ala
65 70 75 80
Phe Asn Pro Ser Gly Val Gln Val Ser Asn Phe Lys Val Pro Thr Glu
85 90 95
Glu Glu Arg Ala His Asp Phe Leu Trp Arg Ile His Arg Gln Thr Pro
100 105 110
Arg Leu Gly Met Ile Gly Val Phe Asn Arg Ser Gln Tyr Glu Asp Val
115 120 125
Leu Val Thr Arg Val His His Leu Ile Asp Asp Gln Thr Ala Gln Arg
130 135 140
Arg Leu Lys His Ile Cys Ala Phe Glu Ser Leu Leu Thr Asp Ser Gly
145 150 155 160
Thr Arg Ile Val Lys Phe Tyr Leu His Ile Ser Pro Glu Glu Gln Lys
165 170 175
Lys Arg Leu Glu Ala Arg Leu Ala Asp Pro Ser Lys His Trp Lys Phe
180 185 190
Asn Pro Gly Asp Leu Gln Glu Arg Ala His Trp Asp Ala Tyr Thr Ala
195 200 205
Val Tyr Glu Asp Val Leu Thr Thr Ser Thr Pro Ala Ala Pro Trp Tyr
210 215 220
Val Val Pro Ala Asp Arg Lys Trp Phe Arg Asn Leu Leu Val Ser Gln
225 230 235 240
Ile Leu Val Gln Thr Leu Glu Glu Met Asn Pro Gln Phe Pro Ala Pro
245 250 255
Ala Phe Asn Ala Ala Asp Leu Arg Ile Val
260 265
<210> 2
<211> 280
<212> PRT
<213> Meiothermus ruber
<400> 2
Met Gly Phe Cys Ser Ile Glu Phe Leu Met Gly Ala Gln Met Lys Lys
1 5 10 15
Tyr Arg Val Gln Pro Asp Gly Arg Phe Glu Leu Lys Arg Phe Asp Pro
20 25 30
Asp Asp Thr Ser Ala Phe Glu Gly Gly Lys Gln Ala Ala Leu Glu Ala
35 40 45
Leu Ala Val Leu Asn Arg Arg Leu Glu Lys Leu Gln Glu Leu Leu Tyr
50 55 60
Ala Glu Gly Gln His Lys Val Leu Val Val Leu Gln Ala Met Asp Ala
65 70 75 80
Gly Gly Lys Asp Gly Thr Ile Arg Val Val Phe Asp Gly Val Asn Pro
85 90 95
Ser Gly Val Arg Val Ala Ser Phe Gly Val Pro Thr Glu Gln Glu Leu
100 105 110
Ala Arg Asp Tyr Leu Trp Arg Val His Gln Gln Val Pro Arg Lys Gly
115 120 125
Glu Leu Val Ile Phe Asn Arg Ser His Tyr Glu Asp Val Leu Val Val
130 135 140
Arg Val Lys Asn Leu Val Pro Gln Gln Val Trp Gln Lys Arg Tyr Arg
145 150 155 160
His Ile Arg Glu Phe Glu Arg Met Leu Ala Asp Glu Gly Thr Thr Ile
165 170 175
Leu Lys Phe Phe Leu His Ile Ser Lys Asp Glu Gln Arg Gln Arg Leu
180 185 190
Gln Glu Arg Leu Asp Asn Pro Glu Lys Arg Trp Lys Phe Arg Met Gly
195 200 205
Asp Leu Glu Asp Arg Arg Leu Trp Asp Arg Tyr Gln Glu Ala Tyr Glu
210 215 220
Ala Ala Ile Arg Glu Thr Ser Thr Glu Tyr Ala Pro Trp Tyr Val Ile
225 230 235 240
Pro Ala Asn Lys Asn Trp Tyr Arg Asn Trp Leu Val Ser His Ile Leu
245 250 255
Val Glu Thr Leu Glu Gly Leu Ala Met Gln Tyr Pro Gln Pro Glu Thr
260 265 270
Ala Ser Glu Lys Ile Val Ile Glu
275 280
<210> 3
<211> 268
<212> PRT
<213> Meiothermus silvanus
<400> 3
Met Ala Lys Thr Ile Gly Ala Thr Leu Asn Leu Gln Asp Ile Asp Pro
1 5 10 15
Arg Ser Thr Pro Gly Phe Asn Gly Asp Lys Glu Lys Ala Leu Ala Leu
20 25 30
Leu Glu Lys Leu Thr Ala Arg Leu Asp Glu Leu Gln Glu Gln Leu Tyr
35 40 45
Ala Glu His Gln His Arg Val Leu Val Ile Leu Gln Gly Met Asp Thr
50 55 60
Ser Gly Lys Asp Gly Thr Ile Arg His Val Phe Lys Asn Val Asp Pro
65 70 75 80
Leu Gly Val Arg Val Val Ala Phe Lys Ala Pro Thr Pro Pro Glu Leu
85 90 95
Glu Arg Asp Tyr Leu Trp Arg Val His Gln His Val Pro Ala Asn Gly
100 105 110
Glu Leu Val Ile Phe Asn Arg Ser His Tyr Glu Asp Val Leu Val Ala
115 120 125
Arg Val His Asn Leu Val Pro Pro Ala Ile Trp Ser Arg Arg Tyr Asp
130 135 140
His Ile Asn Ala Phe Glu Lys Met Leu Val Asp Glu Gly Thr Thr Val
145 150 155 160
Leu Lys Phe Phe Leu His Ile Ser Lys Glu Glu Gln Lys Lys Arg Leu
165 170 175
Leu Glu Arg Leu Val Glu Ala Asp Lys His Trp Lys Phe Asp Pro Gln
180 185 190
Asp Leu Val Glu Arg Gly Tyr Trp Glu Asp Tyr Met Glu Ala Tyr Gln
195 200 205
Asp Val Leu Asp Lys Thr His Thr Gln Tyr Ala Pro Trp His Val Ile
210 215 220
Pro Ala Asp Arg Lys Trp Tyr Arg Asn Leu Gln Val Ser Arg Leu Leu
225 230 235 240
Val Glu Ala Leu Glu Gly Leu Arg Met Lys Tyr Pro Arg Pro Lys Leu
245 250 255
Asn Ile Pro Arg Leu Lys Ser Glu Leu Glu Lys Met
260 265
<210> 4
<211> 300
<212> PRT
<213> Thermosynechococcus elongatus
<400> 4
Met Ile Pro Gln Asp Phe Leu Asp Glu Ile Asn Pro Asp Arg Tyr Ile
1 5 10 15
Val Pro Ala Gly Gly Asn Phe His Trp Lys Asp Tyr Asp Pro Gly Asp
20 25 30
Thr Ala Gly Leu Lys Ser Lys Val Glu Ala Gln Glu Leu Leu Ala Ala
35 40 45
Gly Ile Lys Lys Leu Ala Ala Tyr Gln Asp Val Leu Tyr Ala Gln Asn
50 55 60
Ile Tyr Gly Leu Leu Ile Ile Phe Gln Ala Met Asp Ala Ala Gly Lys
65 70 75 80
Asp Ser Thr Ile Lys His Val Met Ser Gly Leu Asn Pro Gln Ala Cys
85 90 95
Arg Val Tyr Ser Phe Lys Ala Pro Ser Ala Glu Glu Leu Asp His Asp
100 105 110
Phe Leu Trp Arg Ala Asn Arg Ala Leu Pro Glu Arg Gly Cys Ile Gly
115 120 125
Ile Phe Asn Arg Ser Tyr Tyr Glu Glu Val Leu Val Val Arg Val His
130 135 140
Pro Asp Leu Leu Asn Arg Gln Gln Leu Pro Pro Glu Thr Lys Thr Lys
145 150 155 160
His Ile Trp Lys Glu Arg Phe Glu Asp Ile Asn His Tyr Glu Arg Tyr
165 170 175
Leu Thr Arg Asn Gly Ile Leu Ile Leu Lys Phe Phe Leu His Ile Ser
180 185 190
Lys Ala Glu Gln Lys Lys Arg Phe Leu Glu Arg Ile Ser Arg Pro Glu
195 200 205
Lys Asn Trp Lys Phe Ser Ile Glu Asp Val Arg Asp Arg Ala His Trp
210 215 220
Asp Asp Tyr Gln Gln Ala Tyr Ala Asp Val Phe Arg His Thr Ser Thr
225 230 235 240
Lys Trp Ala Pro Trp His Ile Ile Pro Ala Asn His Lys Trp Phe Ala
245 250 255
Arg Leu Met Val Ala His Phe Ile Tyr Gln Lys Leu Ala Ser Leu Asn
260 265 270
Leu His Tyr Pro Met Leu Ser Glu Ala His Arg Glu Gln Leu Leu Glu
275 280 285
Ala Lys Ala Leu Leu Glu Asn Glu Pro Asp Glu Asp
290 295 300
<210> 5
<211> 281
<212> PRT
<213> Anaerolinea thermophila
<400> 5
Met Gly Glu Ala Met Glu Arg Tyr Phe Ile Lys Pro Gly Glu Lys Val
1 5 10 15
Arg Leu Lys Asp Trp Ser Pro Asp Pro Pro Lys Asp Phe Glu Gly Asp
20 25 30
Lys Glu Ser Thr Arg Ala Ala Val Ala Glu Leu Asn Arg Lys Leu Glu
35 40 45
Val Leu Gln Glu Arg Leu Tyr Ala Glu Arg Lys His Lys Val Leu Val
50 55 60
Ile Leu Gln Gly Met Asp Thr Ser Gly Lys Asp Gly Val Ile Arg Ser
65 70 75 80
Val Phe Glu Gly Val Asn Pro Gln Gly Val Lys Val Ala Asn Phe Lys
85 90 95
Val Pro Thr Gln Glu Glu Leu Asp His Asp Tyr Leu Trp Arg Val His
100 105 110
Lys Val Val Pro Gly Lys Gly Glu Ile Val Ile Phe Asn Arg Ser His
115 120 125
Tyr Glu Asp Val Leu Val Val Arg Val His Asn Leu Val Pro Pro Glu
130 135 140
Val Trp Lys Lys Arg Tyr Glu Gln Ile Asn Gln Phe Glu Arg Leu Leu
145 150 155 160
His Glu Thr Gly Thr Thr Ile Leu Lys Phe Phe Leu Phe Ile Ser Arg
165 170 175
Glu Glu Gln Lys Gln Arg Leu Leu Glu Arg Leu Ala Asp Pro Ala Lys
180 185 190
His Trp Lys Phe Asn Pro Gly Asp Leu Lys Glu Arg Ala Leu Trp Glu
195 200 205
Glu Tyr Glu Lys Ala Tyr Glu Asp Val Leu Ser Arg Thr Ser Thr Glu
210 215 220
Tyr Ala Pro Trp Ile Leu Val Pro Ala Asp Lys Lys Trp Tyr Arg Asp
225 230 235 240
Trp Val Ile Ser Arg Val Leu Val Glu Thr Leu Glu Gly Leu Glu Ile
245 250 255
Gln Leu Pro Pro Pro Leu Ala Asp Ala Glu Thr Tyr Arg Arg Gln Leu
260 265 270
Leu Glu Glu Asp Ala Pro Glu Ser Arg
275 280
<210> 6
<211> 270
<212> PRT
<213> Caldilinea aerophila
<400> 6
Met Asp Val Asp Arg Tyr Arg Val Pro Pro Gly Ser Thr Ile His Leu
1 5 10 15
Ser Gln Trp Pro Pro Asp Asp Arg Ser Leu Tyr Glu Gly Asp Lys Lys
20 25 30
Gln Gly Lys Gln Asp Leu Ser Ala Leu Asn Arg Arg Leu Glu Thr Leu
35 40 45
Gln Glu Leu Leu Tyr Ala Glu Gly Lys His Lys Val Leu Ile Ile Leu
50 55 60
Gln Gly Met Asp Thr Ser Gly Lys Asp Gly Val Ile Arg His Val Phe
65 70 75 80
Asn Gly Val Asn Pro Gln Gly Val Lys Val Ala Ser Phe Lys Val Pro
85 90 95
Thr Ala Val Glu Leu Ala His Asp Phe Leu Trp Arg Ile His Arg Gln
100 105 110
Thr Pro Gly Ser Gly Glu Ile Val Ile Phe Asn Arg Ser His Tyr Glu
115 120 125
Asp Val Leu Val Val Arg Val His Gly Leu Val Pro Pro Glu Val Trp
130 135 140
Ala Arg Arg Tyr Glu His Ile Asn Ala Phe Glu Lys Leu Leu Val Asp
145 150 155 160
Glu Gly Thr Thr Ile Leu Lys Phe Phe Leu His Ile Ser Lys Glu Glu
165 170 175
Gln Arg Gln Arg Leu Leu Glu Arg Leu Glu Met Pro Glu Lys Arg Trp
180 185 190
Lys Phe Ser Val Gly Asp Leu Ala Glu Arg Lys Arg Trp Asp Glu Tyr
195 200 205
Met Ala Ala Tyr Glu Ala Val Leu Ser Lys Thr Ser Thr Glu Tyr Ala
210 215 220
Pro Trp Tyr Ile Val Pro Ser Asp Arg Lys Trp Tyr Arg Asn Leu Val
225 230 235 240
Ile Ser His Val Ile Ile Asn Ala Leu Glu Gly Leu Asn Met Arg Tyr
245 250 255
Pro Gln Pro Glu Asp Ile Ala Phe Asp Thr Ile Val Ile Glu
260 265 270
<210> 7
<211> 294
<212> PRT
<213> Chlorobaculum tepidum
<400> 7
Met Lys Leu Asp Leu Asp Ala Phe Arg Ile Gln Pro Gly Lys Lys Pro
1 5 10 15
Asn Leu Ala Lys Arg Pro Thr Arg Ile Asp Pro Val Tyr Arg Ser Lys
20 25 30
Gly Glu Tyr His Glu Leu Leu Ala Asn His Val Ala Glu Leu Ser Lys
35 40 45
Leu Gln Asn Val Leu Tyr Ala Asp Asn Arg Tyr Ala Ile Leu Leu Ile
50 55 60
Phe Gln Ala Met Asp Ala Ala Gly Lys Asp Ser Ala Ile Lys His Val
65 70 75 80
Met Ser Gly Val Asn Pro Gln Gly Cys Gln Val Tyr Ser Phe Lys His
85 90 95
Pro Ser Ala Thr Glu Leu Glu His Asp Phe Leu Trp Arg Thr Asn Cys
100 105 110
Val Leu Pro Glu Arg Gly Arg Ile Gly Ile Phe Asn Arg Ser Tyr Tyr
115 120 125
Glu Glu Val Leu Val Val Arg Val His Pro Glu Ile Leu Glu Met Gln
130 135 140
Asn Ile Pro His Asn Leu Ala His Asn Gly Lys Val Trp Asp His Arg
145 150 155 160
Tyr Arg Ser Ile Val Ser His Glu Gln His Leu His Cys Asn Gly Thr
165 170 175
Arg Ile Val Lys Phe Tyr Leu His Leu Ser Lys Glu Glu Gln Arg Lys
180 185 190
Arg Phe Leu Glu Arg Ile Asp Asp Pro Asn Lys Asn Trp Lys Phe Ser
195 200 205
Thr Ala Asp Leu Glu Glu Arg Lys Phe Trp Asp Gln Tyr Met Glu Ala
210 215 220
Tyr Glu Ser Cys Leu Gln Glu Thr Ser Thr Lys Asp Ser Pro Trp Phe
225 230 235 240
Ala Val Pro Ala Asp Asp Lys Lys Asn Ala Arg Leu Ile Val Ser Arg
245 250 255
Ile Val Leu Asp Thr Leu Glu Ser Leu Asn Leu Lys Tyr Pro Glu Pro
260 265 270
Ser Pro Glu Arg Arg Lys Glu Leu Leu Asp Ile Arg Lys Arg Leu Glu
275 280 285
Asn Pro Glu Asn Gly Lys
290
<210> 8
<211> 269
<212> PRT
<213> Oceanithermus profundus
<400> 8
Met Asp Val Ser Arg Tyr Arg Val Pro Pro Gly Ser Gly Phe Asp Pro
1 5 10 15
Glu Ala Trp Pro Thr Arg Glu Asp Asp Asp Phe Ala Gly Gly Lys Lys
20 25 30
Glu Ala Lys Lys Glu Leu Ala Arg Leu Ala Val Arg Leu Gly Glu Leu
35 40 45
Gln Ala Arg Leu Tyr Ala Glu Gly Arg Gln Ala Leu Leu Ile Val Leu
50 55 60
Gln Gly Met Asp Thr Ala Gly Lys Asp Gly Thr Ile Arg His Val Phe
65 70 75 80
Arg Ala Val Asn Pro Gln Gly Val Arg Val Thr Ser Phe Lys Lys Pro
85 90 95
Thr Ala Leu Glu Leu Ala His Asp Tyr Leu Trp Arg Val His Arg His
100 105 110
Ala Pro Ala Arg Gly Glu Ile Gly Ile Phe Asn Arg Ser His Tyr Glu
115 120 125
Asp Val Leu Val Val Arg Val His Glu Leu Val Pro Pro Glu Val Trp
130 135 140
Gly Arg Arg Tyr Asp His Ile Asn Ala Phe Glu Arg Leu Leu Ala Asp
145 150 155 160
Glu Gly Thr Arg Ile Val Lys Phe Phe Leu His Ile Ser Lys Asp Glu
165 170 175
Gln Lys Arg Arg Leu Glu Ala Arg Leu Glu Asn Pro Arg Lys His Trp
180 185 190
Lys Phe Asn Pro Ala Asp Leu Ser Glu Arg Ala Arg Trp Gly Asp Tyr
195 200 205
Ala Ala Ala Tyr Ala Glu Ala Leu Ser Arg Thr Ser Ser Asp Arg Ala
210 215 220
Pro Trp Tyr Ala Val Pro Ala Asp Arg Lys Trp Gln Arg Asn Arg Ile
225 230 235 240
Val Ala Gln Val Leu Val Asp Ala Leu Glu Ala Met Asp Pro Arg Phe
245 250 255
Pro Arg Val Asp Phe Asp Pro Ala Ser Val Arg Val Glu
260 265
<210> 9
<211> 290
<212> PRT
<213> Roseiflexus castenholzii
<400> 9
Met Tyr Ala Gln Arg Val Val Pro Gly Met Arg Val Arg Leu His Asp
1 5 10 15
Ile Asp Pro Asp Ala Asn Gly Gly Leu Asn Lys Asp Glu Gly Arg Ala
20 25 30
Arg Phe Ala Glu Leu Asn Ala Glu Leu Asp Val Met Gln Glu Glu Leu
35 40 45
Tyr Ala Ala Gly Ile His Ala Leu Leu Leu Ile Leu Gln Gly Met Asp
50 55 60
Thr Ala Gly Lys Asp Gly Ala Ile Arg Asn Val Met Leu Asn Leu Asn
65 70 75 80
Pro Gln Gly Cys Arg Val Glu Ser Phe Lys Val Pro Thr Glu Glu Glu
85 90 95
Leu Ala His Asp Phe Leu Trp Arg Val His Arg Val Val Pro Arg Lys
100 105 110
Gly Met Val Gly Val Phe Asn Arg Ser His Tyr Glu Asp Val Leu Val
115 120 125
Val Arg Val His Ser Leu Val Pro Glu Ser Val Trp Arg Ala Arg Tyr
130 135 140
Asp Gln Ile Asn Ala Phe Glu Arg Leu Leu Ala Asp Thr Gly Thr Ile
145 150 155 160
Ile Val Lys Cys Phe Leu His Ile Ser Lys Glu Glu Gln Glu Gln Arg
165 170 175
Leu Leu Ala Arg Glu Arg Asp Val Ser Lys Ala Trp Lys Leu Ser Ala
180 185 190
Gly Asp Trp Arg Glu Arg Ala Phe Trp Asp Asp Tyr Met Ala Ala Tyr
195 200 205
Glu Glu Ala Leu Thr Arg Cys Ser Thr Asp Tyr Ala Pro Trp Tyr Ile
210 215 220
Ile Pro Ala Asn Arg Lys Trp Tyr Arg Asp Leu Ala Ile Ser Glu Ala
225 230 235 240
Leu Val Glu Thr Leu Arg Pro Tyr Arg Asp Asp Trp Arg Arg Ala Leu
245 250 255
Asp Ala Met Ser Arg Ala Arg Arg Ala Glu Leu Glu Ala Phe Arg Ala
260 265 270
Glu Gln His Ala Met Glu Gly Arg Pro Gln Gly Ala Gly Gly Val Ser
275 280 285
Arg Arg
290
<210> 10
<211> 282
<212> PRT
<213> Roseiflexus castenholzii
<400> 10
Met His Tyr Ala His Thr Val Ile Pro Gly Thr Gln Val Arg Leu Arg
1 5 10 15
Asp Ile Asp Pro Asp Ala Ser Gly Gly Leu Thr Lys Asp Glu Gly Arg
20 25 30
Glu Arg Phe Ala Ser Phe Asn Ala Thr Leu Asp Ala Met Gln Glu Glu
35 40 45
Leu Tyr Ala Ala Gly Val His Ala Leu Leu Leu Ile Leu Gln Gly Met
50 55 60
Asp Thr Ala Gly Lys Asp Gly Ala Ile Arg Asn Val Met His Asn Leu
65 70 75 80
Asn Pro Gln Gly Cys Arg Val Glu Ser Phe Lys Val Pro Thr Glu Glu
85 90 95
Glu Leu Ala His Asp Phe Leu Trp Arg Val His Lys Val Val Pro Arg
100 105 110
Lys Gly Met Val Gly Val Phe Asn Arg Ser His Tyr Glu Asp Val Leu
115 120 125
Val Val Arg Val His Ser Leu Val Pro Glu His Val Trp Arg Ala Arg
130 135 140
Tyr Asp Gln Ile Asn Ala Phe Glu Arg Leu Leu Thr Asp Thr Gly Thr
145 150 155 160
Ile Ile Val Lys Cys Phe Leu His Ile Ser Lys Asp Glu Gln Glu Lys
165 170 175
Arg Leu Leu Ala Arg Glu Gln Asp Val Thr Lys Ala Trp Lys Leu Ser
180 185 190
Ala Gly Asp Trp Arg Glu Arg Glu Arg Trp Asp Glu Tyr Met Ala Ala
195 200 205
Tyr Glu Glu Ala Leu Thr Arg Cys Ser Thr Glu Tyr Ala Pro Trp Tyr
210 215 220
Ile Ile Pro Ala Asn Arg Lys Trp Tyr Arg Asp Leu Ala Ile Ser Glu
225 230 235 240
Val Leu Val Glu Thr Leu Arg Pro Tyr Arg Asp Asp Trp Gln Arg Ala
245 250 255
Leu Asp Ala Met Ser Gln Ala Arg Leu Ala Glu Leu Lys Ala Phe Arg
260 265 270
His Gln Gln Thr Ala Gly Ala Thr Arg Leu
275 280
<210> 11
<211> 274
<212> PRT
<213> Truepera radiovictrix
<400> 11
Met Ser Gln Gly Ser Ala Lys Gly Leu Gly Lys Leu Asp Lys Lys Val
1 5 10 15
Tyr Ala Arg Glu Leu Ala Leu Leu Gln Leu Glu Leu Val Lys Leu Gln
20 25 30
Gly Trp Ile Lys Ala Gln Gly Leu Lys Val Val Val Leu Phe Glu Gly
35 40 45
Arg Asp Ala Ala Gly Lys Gly Ser Thr Ile Thr Arg Ile Thr Gln Pro
50 55 60
Leu Asn Pro Arg Val Cys Arg Val Val Ala Leu Gly Ala Pro Thr Glu
65 70 75 80
Arg Glu Arg Thr Gln Trp Tyr Phe Gln Arg Tyr Val His His Leu Pro
85 90 95
Ala Ala Gly Glu Met Val Leu Phe Asp Arg Ser Trp Tyr Asn Arg Ala
100 105 110
Gly Val Glu Arg Val Met Gly Phe Cys Thr Glu Ala Glu Tyr Arg Glu
115 120 125
Phe Leu His Ala Cys Pro Thr Phe Glu Arg Leu Leu Leu Asp Ala Gly
130 135 140
Ile Ile Leu Ile Lys Tyr Trp Phe Ser Val Ser Ala Ala Glu Gln Glu
145 150 155 160
Arg Arg Met Arg Arg Arg Asn Glu Asn Pro Ala Lys Arg Trp Lys Leu
165 170 175
Ser Pro Met Asp Leu Glu Ala Arg Ala Arg Trp Val Ala Tyr Ser Lys
180 185 190
Ala Lys Asp Ala Met Phe Tyr His Thr Asp Thr Lys Ala Ser Pro Trp
195 200 205
Tyr Val Val Asn Ala Glu Asp Lys Arg Arg Ala His Leu Ser Cys Ile
210 215 220
Ala His Leu Leu Ser Leu Ile Pro Tyr Glu Asp Leu Thr Pro Pro Pro
225 230 235 240
Leu Glu Met Pro Pro Arg Asp Leu Ala Gly Ala Asp Glu Gly Tyr Glu
245 250 255
Arg Pro Asp Lys Ala His Gln Thr Trp Val Pro Asp Tyr Val Pro Pro
260 265 270
Thr Arg
<210> 12
<211> 186
<212> PRT
<213> Thermus thermophilus
<400> 12
Met Asp Val Gly Gln Ala Val Ile Phe Leu Gly Pro Pro Gly Ala Gly
1 5 10 15
Lys Gly Thr Gln Ala Ser Arg Leu Ala Gln Glu Leu Gly Phe Lys Lys
20 25 30
Leu Ser Thr Gly Asp Ile Leu Arg Asp His Val Ala Arg Gly Thr Pro
35 40 45
Leu Gly Glu Arg Val Arg Pro Ile Met Glu Arg Gly Asp Leu Val Pro
50 55 60
Asp Asp Leu Ile Leu Glu Leu Ile Arg Glu Glu Leu Ala Glu Arg Val
65 70 75 80
Ile Phe Asp Gly Phe Pro Arg Thr Leu Ala Gln Ala Glu Ala Leu Asp
85 90 95
Arg Leu Leu Ser Glu Thr Gly Thr Arg Leu Leu Gly Val Val Leu Val
100 105 110
Glu Val Pro Glu Glu Glu Leu Val Arg Arg Ile Leu Arg Arg Ala Glu
115 120 125
Leu Glu Gly Arg Ser Asp Asp Asn Glu Glu Thr Val Arg Arg Arg Leu
130 135 140
Glu Val Tyr Arg Glu Lys Thr Glu Pro Leu Val Gly Tyr Tyr Glu Ala
145 150 155 160
Arg Gly Val Leu Lys Arg Val Asp Gly Leu Gly Thr Pro Asp Glu Val
165 170 175
Tyr Ala Arg Ile Arg Ala Ala Leu Gly Ile
180 185
<210> 13
<211> 208
<212> PRT
<213> Thermus thermophilus
<400> 13
Met Arg Gly Ile Val Thr Ile Asp Gly Pro Ser Ala Ser Gly Lys Ser
1 5 10 15
Ser Val Ala Arg Arg Val Ala Ala Ala Leu Gly Val Pro Tyr Leu Ser
20 25 30
Ser Gly Leu Leu Tyr Arg Ala Ala Ala Phe Leu Ala Leu Arg Ala Gly
35 40 45
Val Asp Pro Gly Asp Glu Glu Gly Leu Leu Ala Leu Leu Glu Gly Leu
50 55 60
Gly Val Arg Leu Leu Ala Gln Ala Glu Gly Asn Arg Val Leu Ala Asp
65 70 75 80
Gly Glu Asp Leu Thr Ser Phe Leu His Thr Pro Glu Val Asp Arg Val
85 90 95
Val Ser Ala Val Ala Arg Leu Pro Gly Val Arg Ala Trp Val Asn Arg
100 105 110
Arg Leu Lys Glu Val Pro Pro Pro Phe Val Ala Glu Gly Arg Asp Met
115 120 125
Gly Thr Ala Val Phe Pro Glu Ala Ala His Lys Phe Tyr Leu Thr Ala
130 135 140
Ser Pro Glu Val Arg Ala Trp Arg Arg Ala Arg Glu Arg Pro Gln Ala
145 150 155 160
Tyr Glu Glu Val Leu Arg Asp Leu Leu Arg Arg Asp Glu Arg Asp Lys
165 170 175
Ala Gln Ser Ala Pro Ala Pro Asp Ala Leu Val Leu Asp Thr Gly Gly
180 185 190
Met Thr Leu Asp Glu Val Val Ala Trp Val Leu Ala His Ile Arg Arg
195 200 205
<210> 14
<211> 225
<212> PRT
<213> Pyrococcus furiosus
<400> 14
Met Arg Ile Val Phe Asp Ile Gly Gly Ser Val Leu Val Pro Glu Asn
1 5 10 15
Pro Asp Ile Asp Phe Ile Lys Glu Ile Ala Tyr Gln Leu Thr Lys Val
20 25 30
Ser Glu Asp His Glu Val Ala Val Val Val Gly Gly Gly Lys Leu Ala
35 40 45
Arg Lys Tyr Ile Glu Val Ala Glu Lys Phe Asn Ser Ser Glu Thr Phe
50 55 60
Lys Asp Phe Ile Gly Ile Gln Ile Thr Arg Ala Asn Ala Met Leu Leu
65 70 75 80
Ile Ala Ala Leu Arg Glu Lys Ala Tyr Pro Val Val Val Glu Asp Phe
85 90 95
Trp Glu Ala Trp Lys Ala Val Gln Leu Lys Lys Ile Pro Val Met Gly
100 105 110
Gly Thr His Pro Gly His Thr Thr Asp Ala Val Ala Ala Leu Leu Ala
115 120 125
Glu Phe Leu Lys Ala Asp Leu Leu Val Val Ile Thr Asn Val Asp Gly
130 135 140
Val Tyr Thr Ala Asp Pro Lys Lys Asp Pro Thr Ala Lys Lys Ile Lys
145 150 155 160
Lys Met Lys Pro Glu Glu Leu Leu Glu Ile Val Gly Lys Gly Ile Glu
165 170 175
Lys Ala Gly Ser Ser Ser Val Ile Asp Pro Leu Ala Ala Lys Ile Ile
180 185 190
Ala Arg Ser Gly Ile Lys Thr Ile Val Ile Gly Lys Glu Asp Ala Lys
195 200 205
Asp Leu Phe Arg Val Ile Lys Gly Asp His Asn Gly Thr Thr Ile Glu
210 215 220
Pro
225
<210> 15
<211> 207
<212> PRT
<213> Thermotoga maritima
<400> 15
Met Lys Gly Gln Leu Phe Val Ile Cys Gly Pro Ser Gly Ala Gly Lys
1 5 10 15
Thr Ser Ile Ile Lys Glu Val Leu Lys Arg Leu Asp Asn Val Val Phe
20 25 30
Ser Val Ser Cys Thr Thr Arg Pro Lys Arg Pro His Glu Glu Asp Gly
35 40 45
Lys Asp Tyr Phe Phe Ile Thr Glu Glu Glu Phe Leu Lys Arg Val Glu
50 55 60
Arg Gly Glu Phe Leu Glu Trp Ala Arg Val His Gly His Leu Tyr Gly
65 70 75 80
Thr Leu Arg Ser Phe Val Glu Ser His Ile Asn Glu Gly Lys Asp Val
85 90 95
Val Leu Asp Ile Asp Val Gln Gly Ala Leu Ser Val Lys Lys Lys Tyr
100 105 110
Ser Asn Thr Val Phe Ile Tyr Val Ala Pro Pro Ser Tyr Ala Asp Leu
115 120 125
Arg Glu Arg Ile Leu Lys Arg Gly Thr Glu Lys Glu Ala Asp Val Leu
130 135 140
Val Arg Leu Glu Asn Ala Lys Trp Glu Leu Met Phe Met Asp Glu Phe
145 150 155 160
Asp Tyr Ile Val Val Asn Glu Asn Leu Glu Asp Ala Val Glu Met Val
165 170 175
Val Ser Ile Val Arg Ser Glu Arg Ala Lys Val Thr Arg Asn Gln Asp
180 185 190
Lys Ile Glu Arg Phe Lys Met Glu Val Lys Gly Trp Lys Lys Leu
195 200 205
<210> 16
<211> 142
<212> PRT
<213> Aquifex aeolicus
<400> 16
Met Ala Val Glu Arg Thr Leu Ile Ile Val Lys Pro Asp Ala Met Glu
1 5 10 15
Lys Gly Ala Leu Gly Lys Ile Leu Asp Arg Phe Ile Gln Glu Gly Phe
20 25 30
Gln Ile Lys Ala Leu Lys Met Phe Arg Phe Thr Pro Glu Lys Ala Gly
35 40 45
Glu Phe Tyr Tyr Val His Arg Glu Arg Pro Phe Phe Gln Glu Leu Val
50 55 60
Glu Phe Met Ser Ser Gly Pro Val Val Ala Ala Val Leu Glu Gly Glu
65 70 75 80
Asp Ala Ile Lys Arg Val Arg Glu Ile Ile Gly Pro Thr Asp Ser Glu
85 90 95
Glu Ala Arg Lys Val Ala Pro Asn Ser Ile Arg Ala Gln Phe Gly Thr
100 105 110
Asp Lys Gly Lys Asn Ala Ile His Ala Ser Asp Ser Pro Glu Ser Ala
115 120 125
Gln Tyr Glu Ile Cys Phe Ile Phe Ser Gly Leu Glu Ile Val
130 135 140
<---
Реферат
Группа изобретений относится к биотехнологии, в частности к способам получения РНК. Инкубируют клеточную РНК и рибонуклеазу или нуклеазу P1 для получения 5’-нуклеозидмонофосфатов (5’-НМФ). Проводят элиминирование рибонуклеазы или нуклеазы. Затем инкубируют 5’-НМФ с полифосфаткиназой (PPK), полифосфатом, матрицей ДНК, кодирующей представляющую интерес РНК, и РНК-полимеразой. Клеточная РНК включает рибосомальную, матричную и/или транспортную РНК. В одном варианте на стадии инкубирования 5’-НМФ в реакционную смесь дополнительно вносят по меньшей мере одну НМФ-киназу и по меньшей мере одну НДФ-киназу. PPK может представлять собой фермент PPK2 класса III из Deinococcus geothermalis с SEQ ID NO: 1. Элиминирование проводят посредством температуры, pH, соли, детергента, спирта, химических ингибиторов, отделения, осаждения, фильтрации, захвата и/или хроматографии. Изобретения обеспечивают эффективный бесклеточный синтез РНК из многих источников нуклеотидов, в том числе клеточной РНК, нуклеозид-5’-монофосфатов и нуклеозиддифосфатов, при этом не требуются термостабильные РНК-полимеразы. 4 н. и 26 з.п. ф-лы, 19 ил., 8 пр.
Комментарии