Уменьшение содержания табак-специфичных нитрозаминов посредством изменения пути ассимиляции нитратов - RU2721799C2
Код документа: RU2721799C2
Чертежи

Описание
Область изобретения
Настоящее изобретение относится к модифицированным растениям табака, характеризующимся уменьшенными уровнями нитратов, табачным продуктам, получаемым из модифицированных растений табака, характеризующихся уменьшенным содержанием табак-специфичных нитрозаминов (TSNA), и способам уменьшения содержания TSNA в табачных продуктах посредством изменения экспрессии генов пути ассимиляции нитратов.
Предпосылки изобретения
Коммерческие растения табака, такие как табак Берли, накапливают в своих листьях высокие уровни свободных нитратов, которые являются нежелательными, поскольку высокие уровни нитратов связаны с образованием канцерогенных соединений, называемых табак-специфичными нитрозаминами (TSNA). TSNA представляют собой класс соединений, которые вырабатываются главным образом в процессе сушки листьев табака, хотя дополнительное образование может происходить при последующей переработке и хранении листьев, а также, возможно, путем пиросинтеза во время сгорания. Два TSNA, обнаруживаемых в высушенных листьях, N-нитрозонорникотин (NNN) и 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанон (NNK), классифицируются Международным агентством по изучению рака как канцерогенные вещества группы I (обозначение высшего уровня). Ввиду значительного объема свидетельств причастности этих соединений к развитию различных форм рака, связанных с табаком, Всемирная организация здравоохранения и другие эксперты в данной области рекомендовали реализовать предписания, направленные на обеспечение того, чтобы в будущих табачных продуктах уровни таких токсичных веществ были уменьшены. TSNA представляют собой продукты нитрозации алкалоидов табака. В отношении видов табака воздушной сушки существует общее мнение, что нитрит является средством, непосредственно отвечающим за образование TSNA. Однако из-за их клеточной токсичности уровни эндогенных нитритов, как правило, очень низки в растительных тканях. Полагают, что вместо этого подавляющее большинство нитритов, вовлеченных в образование TSNA, образуются в результате нитратредуктазной (NR) активности микробов, находящихся на поверхности листа в течение 6-10 недель процесса сушки, посредством которой часть нитратных пулов листа преобразуется в нитриты по мере распада клеточных мембран и органелл в течение этого периода.
TSNA образуются главным образом во время процесса сушки листьев, что предусматривает нитрозацию алкалоидов табака. Генетические стратегии снижения содержания и уровней TSNA в высушенных листьях были сосредоточены на целенаправленном воздействии на (1) алкалоидного (алкалоидных) предшественника (предшественников) либо (2) вовлеченное (вовлеченные) нитрозирующее (нитрозирующие) средство(средства). Большинство усилий по уменьшению содержания TSNA на уровне изменения генетических характеристик табака целенаправленно воздействуют на превращение алкалоидных предшественников в TSNA. Такие стратегии обеспечивают существенное уменьшение содержания NNN вследствие снижения экспрессии генов семейства, отвечающего за синтез его алкалоидного предшественника норникотина. Однако такие стратегии не обеспечивают уменьшение уровней всех TSNA, обнаруживаемых в табачных продуктах. Соответственно, остается значительная необходимость в уменьшении уровней всех TSNA в табачном продукте.
Краткое описание изобретения
Настоящее изобретение направлено в одном аспекте на табачный продукт, характеризующийся уменьшенными уровнями табак-специфичных нитрозаминов (TSNA), который получают из растения табака, при этом указанное растение табака модифицируют таким образом, чтобы оно содержало: (i) полинуклеотид, содержащий последовательность, кодирующую дерегулированный фермент нитратредуктазу, состоящий из нее или по сути состоящий из нее; (ii) полипептид, кодируемый полинуклеотидом, изложенным в (i); (iii) полипептид, содержащий дерегулированный фермент нитратредуктазу, состоящий из него или по сути состоящий из него; или (iv) конструкцию, вектор или вектор экспрессии, содержащие полинуклеотид, изложенный в (i), где экспрессия или активность указанной нитратредуктазы является дерегулированной по сравнению с контрольным немодифицированным растением табака и где дерегулированный фермент нитратредуктаза включает в себя (a) полипептидную нитратредуктазу, содержащую аминокислотную замену в положении, соответствующем положению 523 в SEQ ID NO: 4; или (b) полипептидную нитратредуктазу, содержащую аминокислотную замену в положении, соответствующем положению 523 в SEQ ID NO: 4, где аминокислота в положении 523 SEQ ID NO: 4 заменена аспарагиновой кислотой. Дерегулированный фермент нитратредуктаза может представлять собой фермент нитратредуктазу, который является конститутивно активным. Растение табака может относиться к виду Nicotiana tabacum, например, табачный продукт может содержать растительный материал из растения табака вида Nicotiana tabacum. Полинуклеотид, кодирующий дерегулированную нитратредуктазу, может представлять собой гетерологичный полинуклеотид, кодирующий модифицированную полипептидную нитратредуктазу. Гетерологичный полинуклеотид может быть связан с промотором, в нативных условиях не связанным с эндогенным геном нитратредуктазы. Промотор может представлять собой промотор 35S вируса мозаики цветной капусты. Полинуклеотид, кодирующий дерегулированную нитратредуктазу, может содержать полинуклеотидную последовательность SEQ ID NO: 5. Полинуклеотид, кодирующий дерегулированную нитратредуктазу, может представлять собой эндогенный ген нитратредуктазы, который был модифицирован с помощью системы редактирования генома или с помощью мутагена. Система редактирования генома может включать в себя сконструированную систему на основе CRISPR/Cas, сконструированную эффекторную нуклеазу, подобную активаторам транскрипции, сконструированную нуклеазу с «цинковыми пальцами» или сконструированную мегануклеазу. Табак может представлять собой табак Берли. Табачный продукт может характеризоваться уменьшенными уровнями табак-специфичных нитрозаминов (TSNA) по сравнению с табачным продуктом, полученным из контрольного растения табака, в котором фермент нитратредуктаза не был дерегулирован. Общий уровень TSNA можно измерить в листе растения табака, где (a) лист является свежесобранным; (b) лист является высушенным, хранящимся или переработанным; или (c) лист является высушенным на воздухе. Уровень по меньшей мере одного TSNA в табачном продукте может быть уменьшенным по сравнению с контрольным уровнем по меньшей мере одного TSNA, где по меньшей мере один TSNA выбран из группы, включающей N-нитрозонорникотин (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанон (NNK), N-нитрозоанабазин (NAB), N-нитрозоанатабин (NAT) и их комбинации. Общие уровни TSNA можно уменьшить по меньшей мере на 50%. Уровни TSNA можно измерить в дыме, получаемом при сгорании листьев растения табака. Общие уровни TSNA в дыме можно уменьшить по меньшей мере на 70%. Уровень NNN можно уменьшить на приблизительно 90%. Уровень NNK можно уменьшить на приблизительно 66%. Уровень NAB можно уменьшить на приблизительно 92%. Уровень NAT можно уменьшить на приблизительно 88%. Растение табака может дополнительно содержать модифицированный ген пути синтеза норникотина. Модифицированный ген пути синтеза норникотина может включать в себя модифицированный ген никотиндеметилазы или ген цитохрома P450. Растение табака может содержать модифицированный ген CYP82E4 или модифицированный ген CYP82E10. Модифицированный ген CYP82E4 или модифицированный ген CYP82E10 может быть инактивированным.
Настоящее изобретение в одном аспекте направлено на способ получения табачного продукта, где уровни TSNA, измеренные в дыме, полученном при сгорании листьев модифицированного растения табака, уменьшены по сравнению с уровнями TSNA, измеренными в дыме, полученном при сгорании немодифицированного растения табака. Способ включает (a) модификацию растения табака таким образом, чтобы оно содержало: (i) полинуклеотид, содержащий последовательность, кодирующую дерегулированный фермент нитратредуктазу, состоящий из нее или по сути состоящий из нее; (ii) полипептид, кодируемый полинуклеотидом, изложенным в (i); (iii) полипептид, содержащий дерегулированный фермент нитратредуктазу, состоящий из него или по сути состоящий из него; или (iv) конструкцию, вектор или вектор экспрессии, содержащие полинуклеотид, изложенный в (i), где экспрессия или активность указанной нитратредуктазы является дерегулированной по сравнению с контрольным немодифицированным растением табака и где дерегулированный фермент нитратредуктаза включает в себя (I) полипептидную нитратредуктазу, содержащую аминокислотную замену в положении, соответствующем положению 523 в SEQ ID NO: 4; или (II) полипептидную нитратредуктазу, содержащую аминокислотную замену в положении, соответствующем положению 523 в SEQ ID NO: 4, где аминокислота в положении 523 SEQ ID NO: 4 заменена аспарагиновой кислотой; (b) сбор листьев табака с указанного модифицированного растения табака и (c) получение табачного продукта из собранных листьев.
В другом аспекте в данном документе также раскрывается табачный продукт, характеризующийся уменьшенными уровнями табак-специфичных нитрозаминов (TSNA), который получают из растения табака, при этом указанное растение табака модифицируют таким образом, чтобы оно содержало: (i) полинуклеотид, содержащий последовательность, кодирующую дерегулированный фермент нитратредуктазу, состоящий из нее или по сути состоящий из нее; (ii) полипептид, кодируемый полинуклеотидом, изложенным в (i); (iii) полипептид, содержащий дерегулированный фермент нитратредуктазу, состоящий из него или по сути состоящий из него; или (iv) конструкцию, вектор или вектор экспрессии, содержащие полинуклеотид, изложенный в (i), где экспрессия или активность указанной нитратредуктазы является дерегулированной по сравнению с контрольным немодифицированным растением табака и где дерегулированный фермент нитратредуктаза включает в себя (a) усеченную полипептидную нитратредуктазу; (b) полипептидную нитратредуктазу, содержащую N-концевое усечение, или (c) полипептидную нитратредуктазу, содержащую N-концевое усечение 56 аминокислот.
В другом аспекте в данном документе также раскрывается способ получения табачного продукта, где уровни TSNA, измеренные в дыме, полученном при сгорании листьев модифицированного растения табака, уменьшены по сравнению с уровнями TSNA, измеренными в дыме, полученном при сгорании немодифицированного растения табака, при этом способ включает (a) модификацию растения табака таким образом, чтобы оно содержало: (i) полинуклеотид, содержащий последовательность, кодирующую дерегулированный фермент нитратредуктазу, состоящий из нее или по сути состоящий из нее; (ii) полипептид, кодируемый полинуклеотидом, изложенным в (i); (iii) полипептид, содержащий дерегулированный фермент нитратредуктазу, состоящий из него или по сути состоящий из него; или (iv) конструкцию, вектор или вектор экспрессии, содержащие полинуклеотид, изложенный в (i), где экспрессия или активность указанной нитратредуктазы является дерегулированной по сравнению с контрольным немодифицированным растением табака и где дерегулированный фермент нитратредуктаза включает в себя (I) усеченную полипептидную нитратредуктазу; (II) полипептидную нитратредуктазу, содержащую N-концевое усечение; или (III) полипептидную нитратредуктазу, содержащую N-концевое усечение 56 аминокислот; (b) сбор листьев табака с указанного модифицированного растения табака и (c) получение табачного продукта из собранных листьев.
Полинуклеотид, кодирующий дерегулированную нитратредуктазу, может содержать полинуклеотидную последовательность SEQ ID NO: 7.
Краткое описание графических материалов
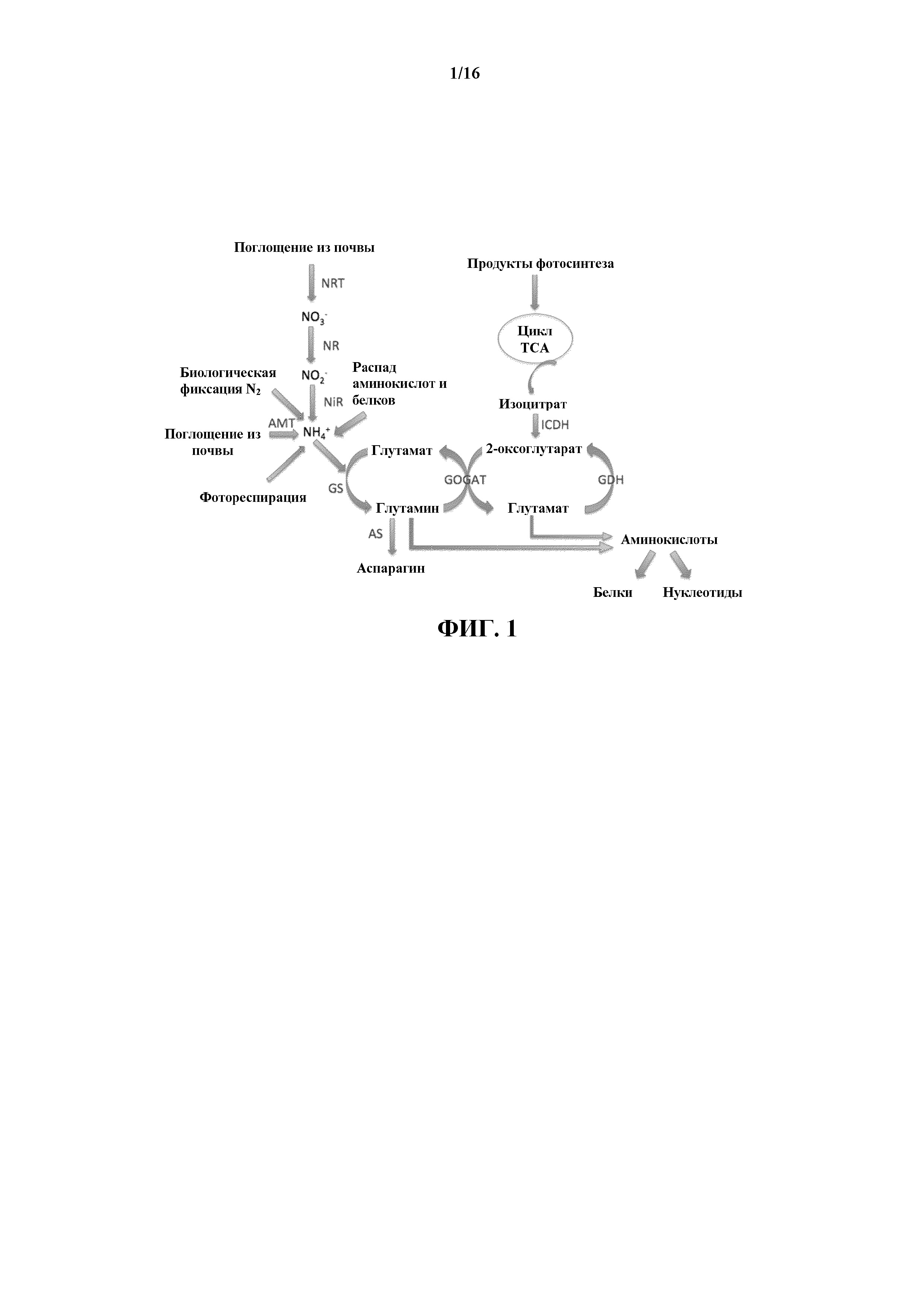
На фиг. 1 показана схема пути ассимиляции азота в высших растениях. В пути первичной N-ассимиляции нитрат (NO3-) превращается в нитрит (NO2-), затем последовательно в аммоний (NH4+) и глутамин, при этом основными ферментами, вовлеченными в эти реакции, являются нитрат- и нитритредуктаза (соответственно NR и NiR), глутаминсинтетаза (GS) и глутаматсинтаза (GOGAT). AMT - переносчики аммония; AS - аспарагинсинтетаза; GDH - NADH-зависимая глутаматдегидрогеназа; ICDH - NADH-зависимая изоцитратдегидрогеназа; NRT - переносчики нитрата; TCA - цикл трикарбоновых кислот.
На фиг. 2 показано выравнивание участков, содержащих положение 523 аминокислотной последовательности нитратредуктазы, из разных видов растений.
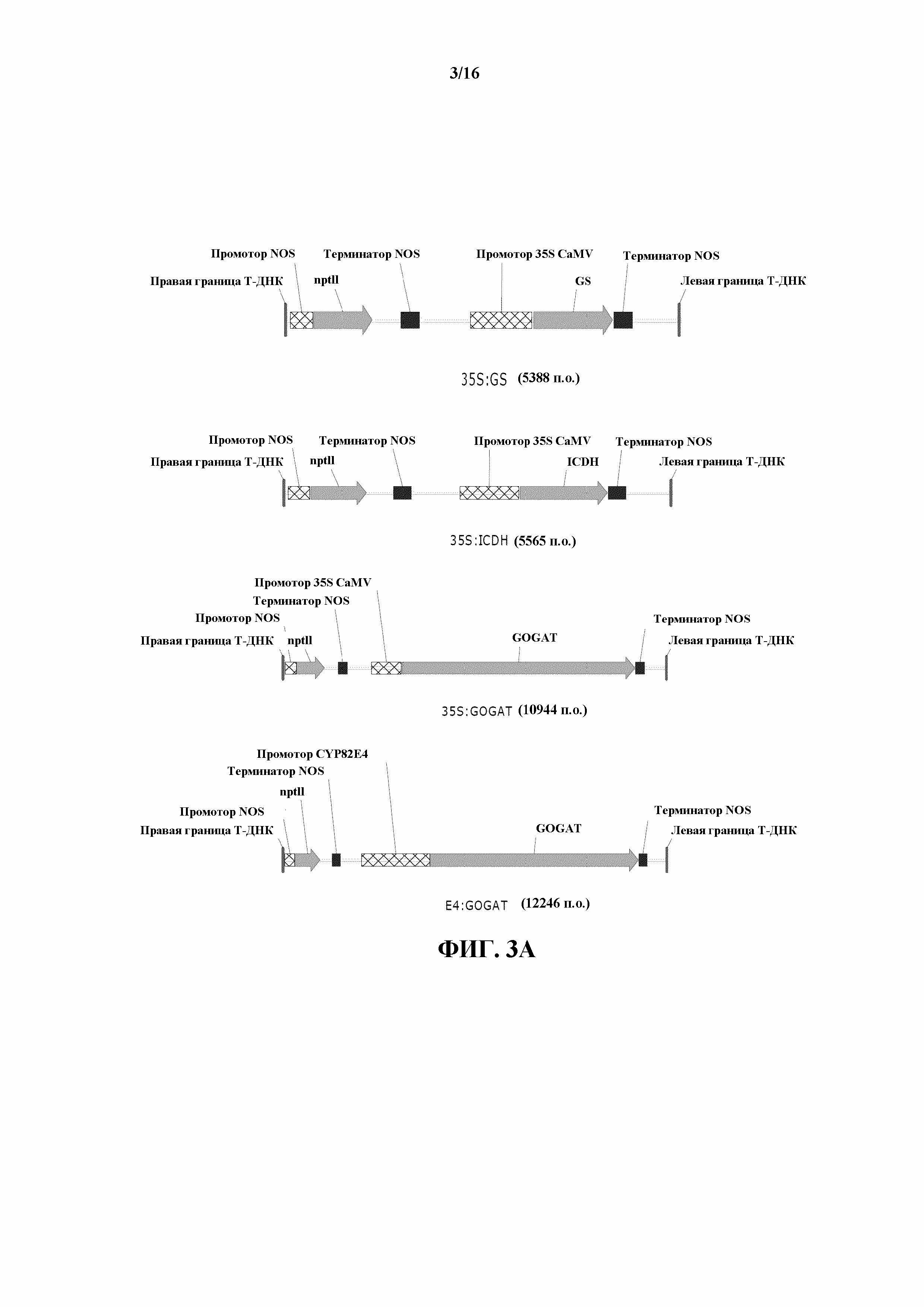
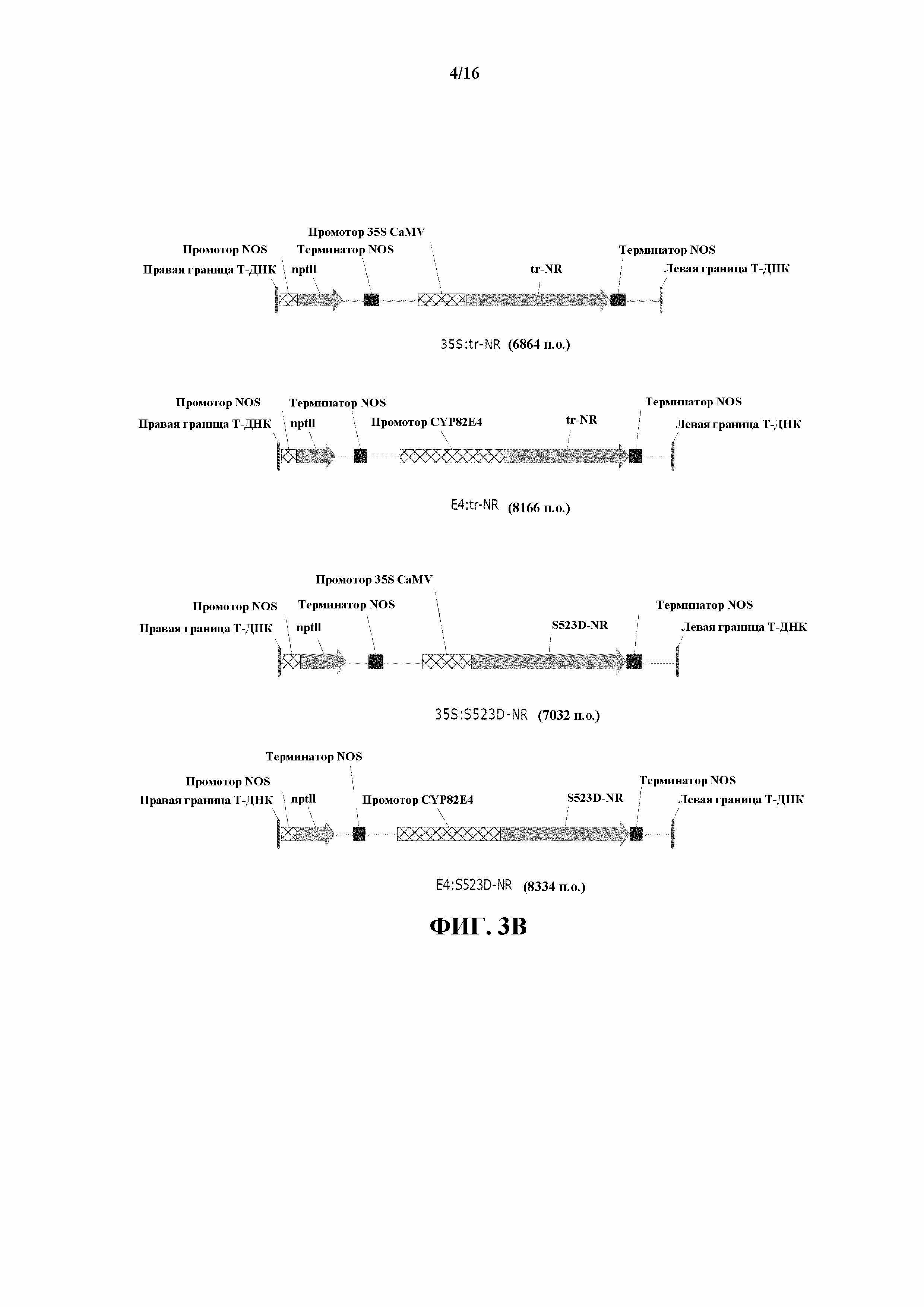
На фиг. 3A и 3B показаны карты конструкций для 35S:GS1, 35S:ICDH, 35S:GOGAT, E4:GOGAT, 35S:tr-NR, E4:tr-NR, 35S:S523D-NR и E4:S523D-NR.
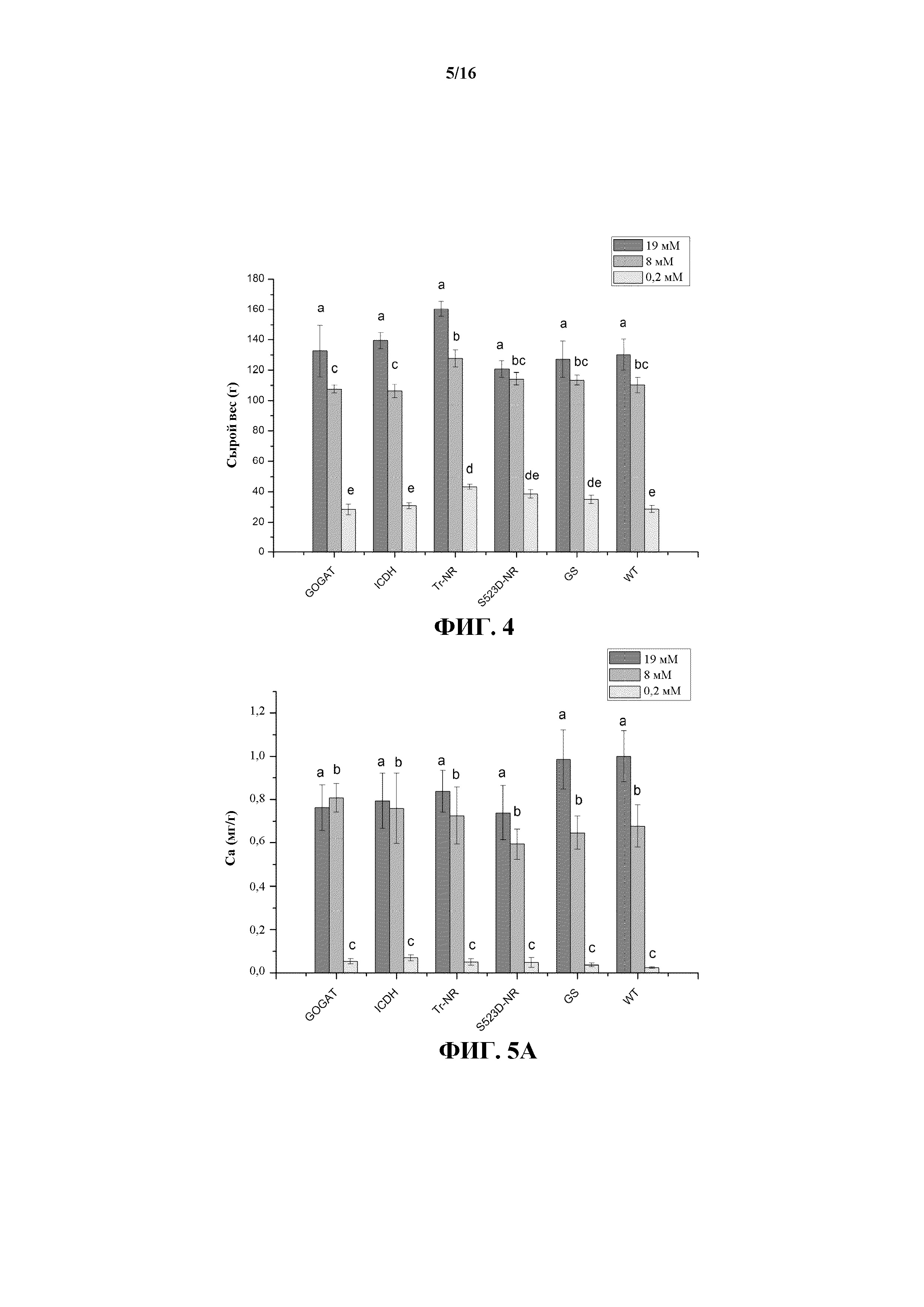
На фиг. 4 показаны значения сырого веса растений дикого типа (WT) и трансгенных линий 35S:tr-NR, 35S:S523D-NR, 35S:GS1, 35S:GOGAT и 35S:ICDH, выращиваемых при трех уровнях обработки нитратными (N) удобрениями.
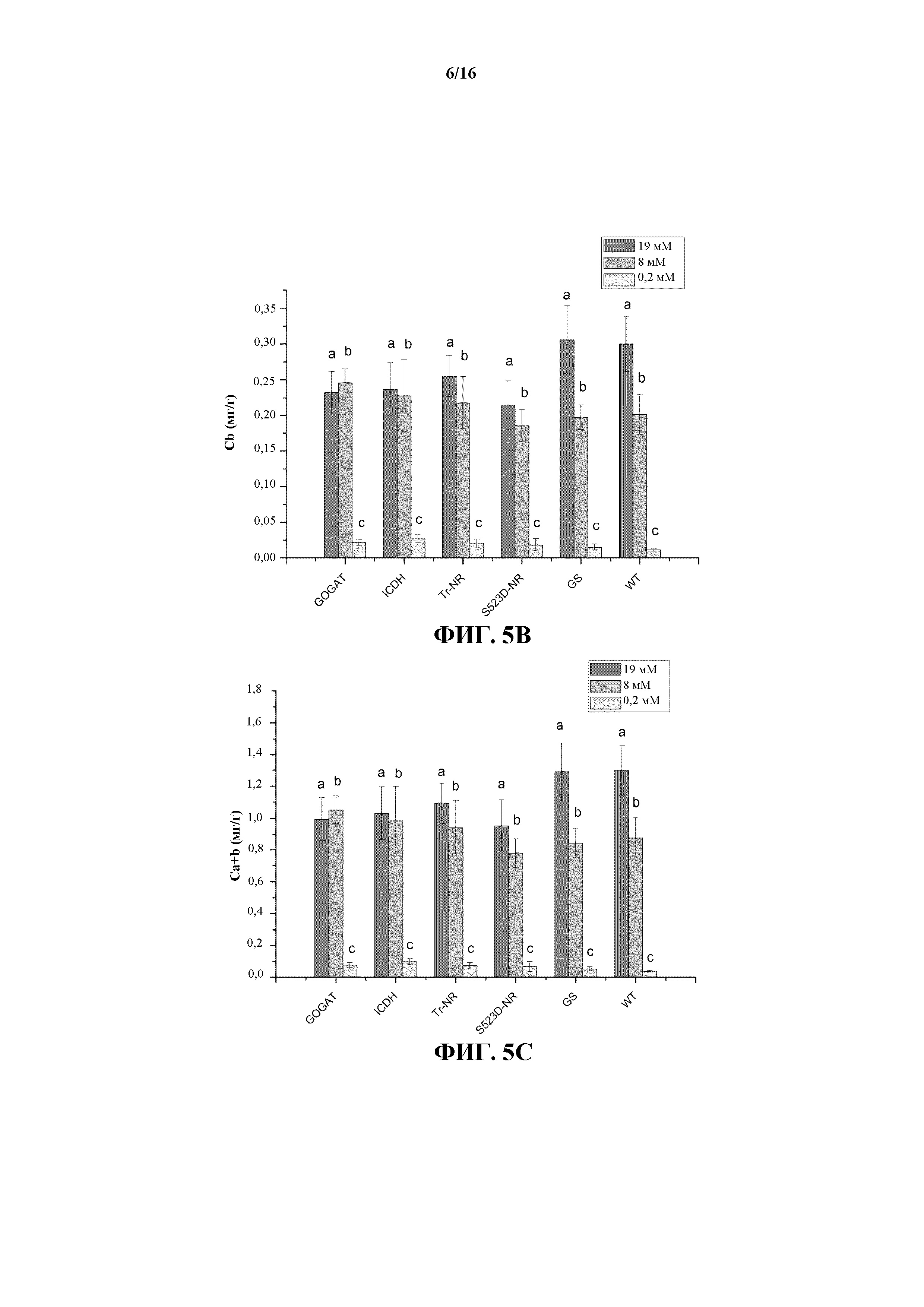
На фиг. 5A-5C показаны значения содержания хлорофилла a (Ca, на фиг. 5A), хлорофилла b (Cb, на фиг. 5B) и хлорофилла a+b (Ca+b, на фиг. 5C) для растений WT и трансгенных линий 35S:tr-NR, 35S:S523D-NR, 35S:GS1, 35S:GOGAT и 35S:ICDH, выращиваемых при трех уровнях обработки N-удобрениями.
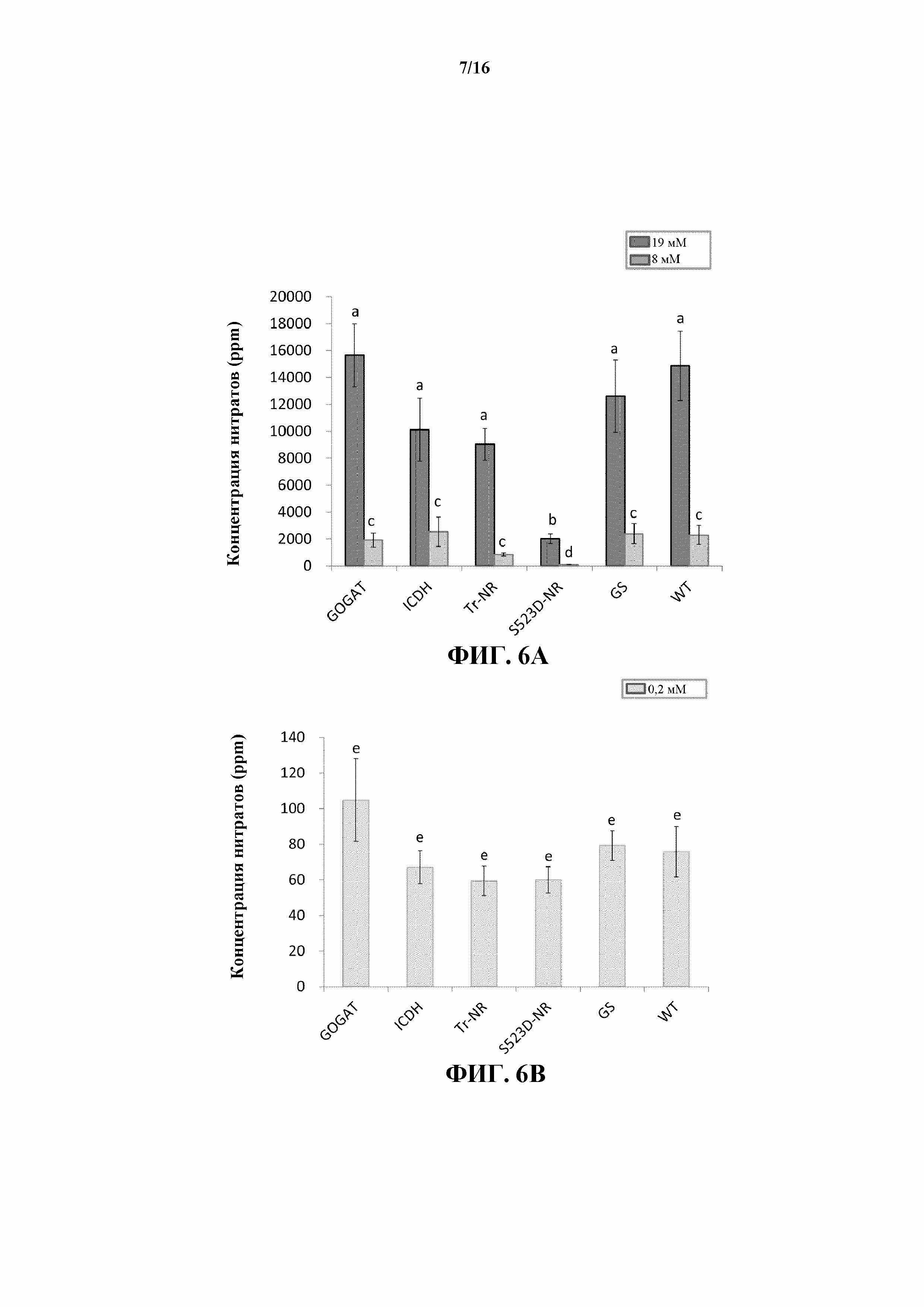
На фиг. 6A и 6B показано общее содержание нитратов в листьях растений WT и трансгенных линий 35S:tr-NR, 35S:S523D-NR, 35S:GS1, 35S:GOGAT и 35S:ICDH, выращиваемых при трех уровнях обработки N-удобрениями.
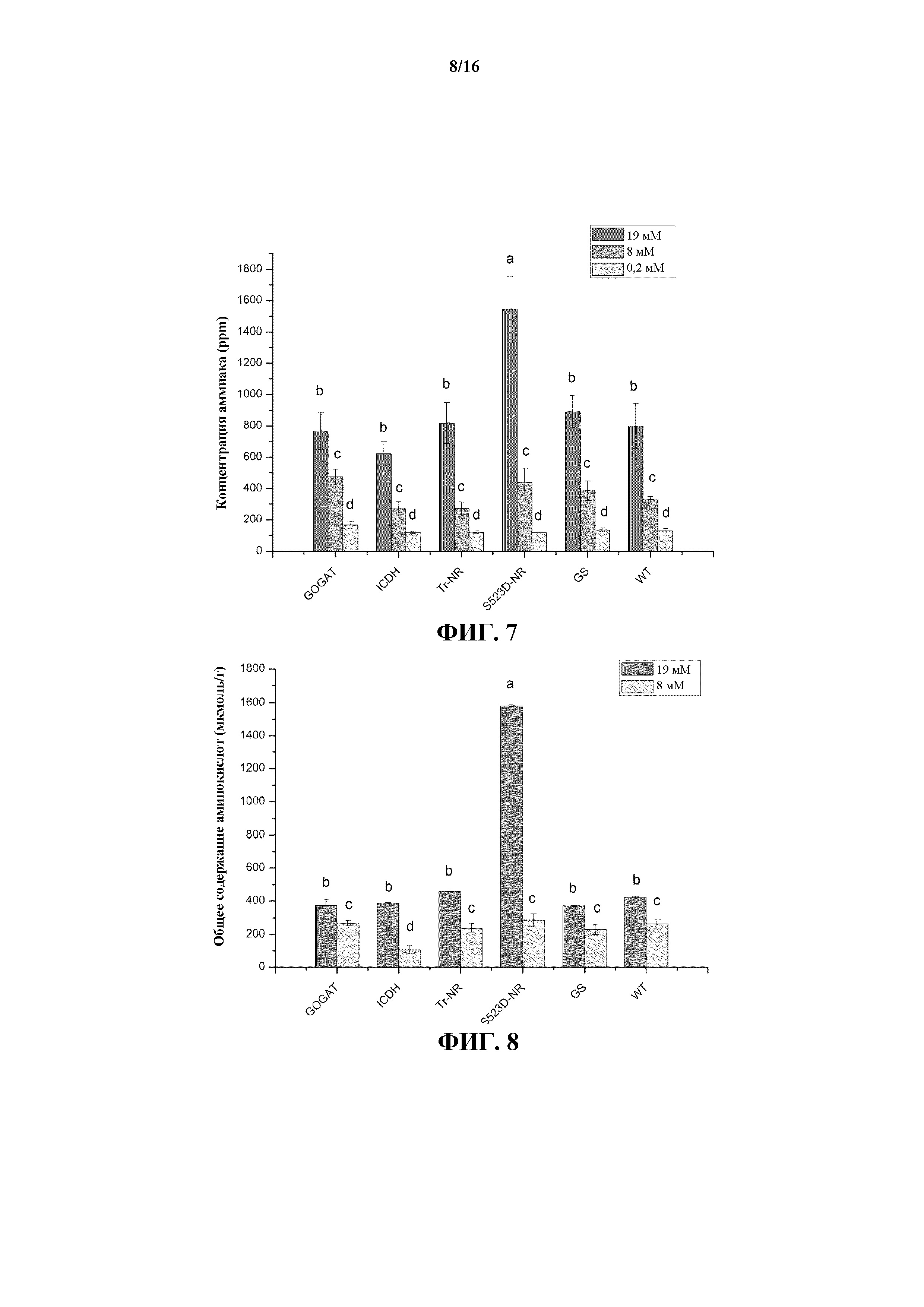
На фиг. 7 показано среднее содержание аммиака в листьях растений WT и трансгенных линий 35S:tr-NR, 35S:S523D-NR, 35S:GS1, 35S:GOGAT и 35S:ICDH, выращиваемых при трех уровнях обработки N-удобрениями.
На фиг. 8 показано общее содержание свободных аминокислот в листьях 35S:GOGAT, 35S:ICDH, 35S:tr-NR, 35S:S523D-NR, 35S:GS1 и растений WT, выращиваемых при среднем (8 мМ) и высоком (19 мМ) уровнях обработки N-удобрениями.
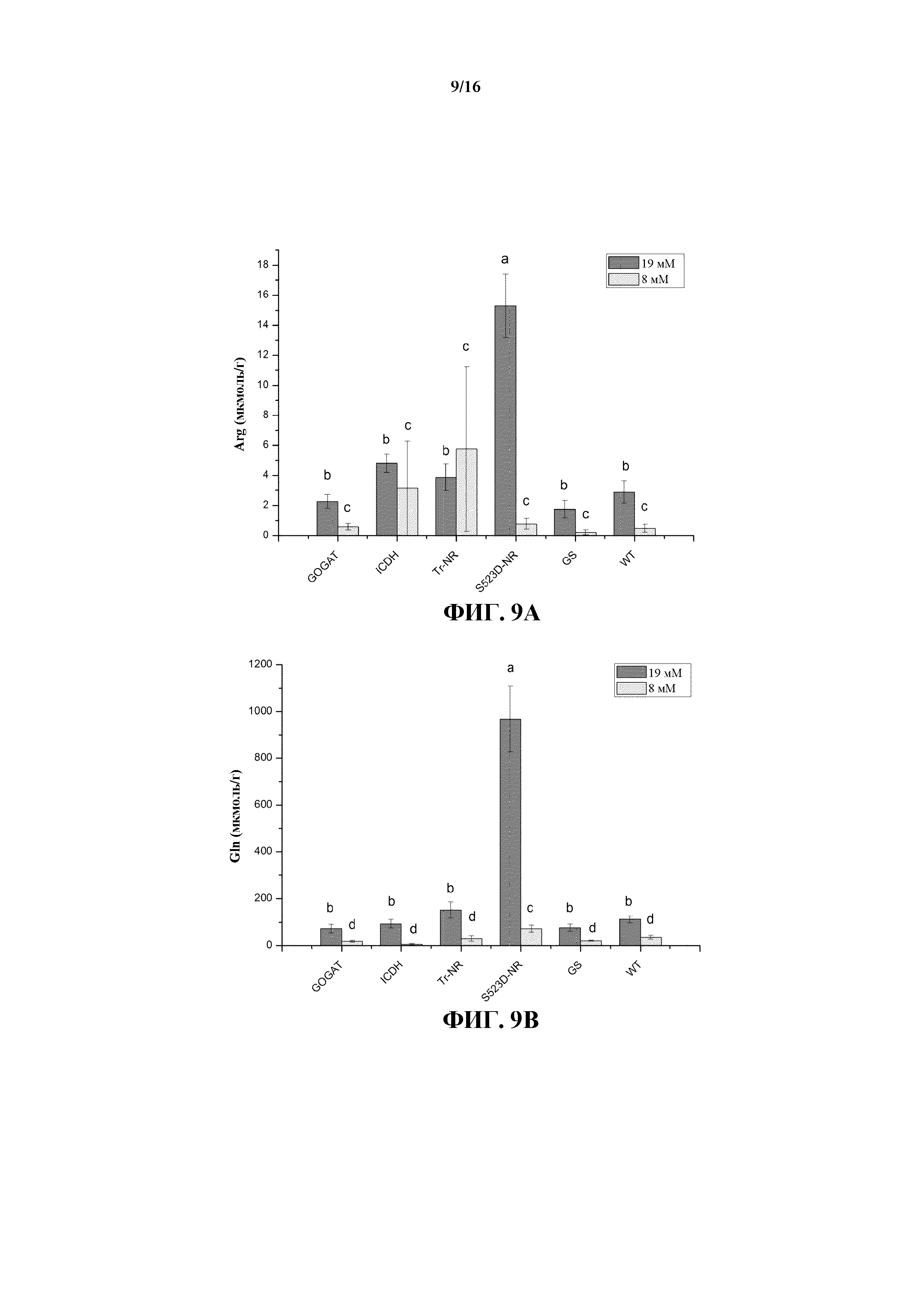
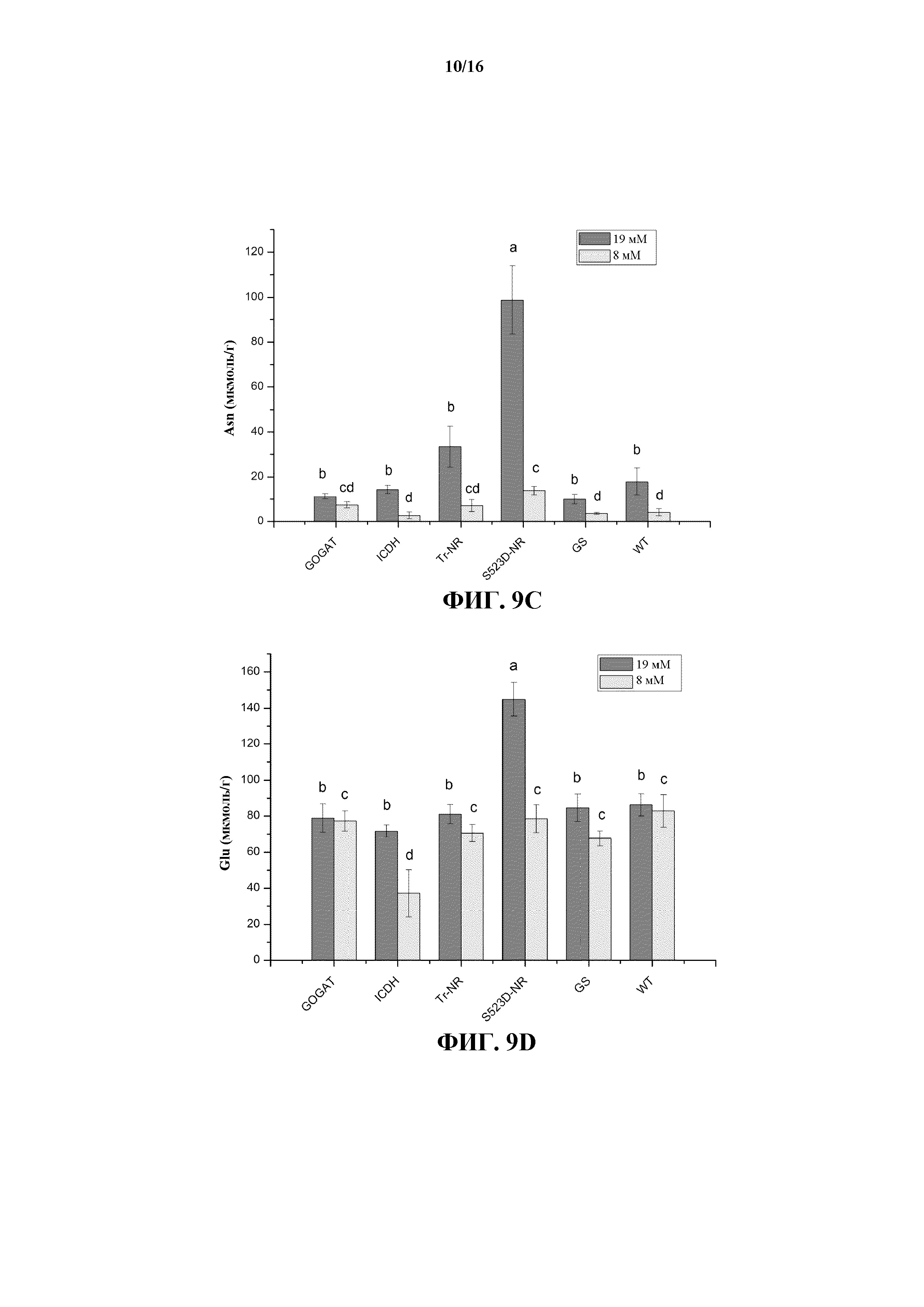
На фиг. 9A-9D показано содержание аргинина (Arg) (на фиг. 9A), глутамина (Gln) (на фиг. 9B), аспарагина (Asn) (на фиг. 9C) и глутамата (Glu) (на фиг. 9D) в листьях табака растений WT и трансгенных линий 35S:tr-NR, 35S:S523D-NR, 35S:GS1, 35S:GOGAT, а также 35S:ICDH, выращиваемых при среднем (8 мМ) и высоком (19 мМ) уровнях обработки N-удобрениями.
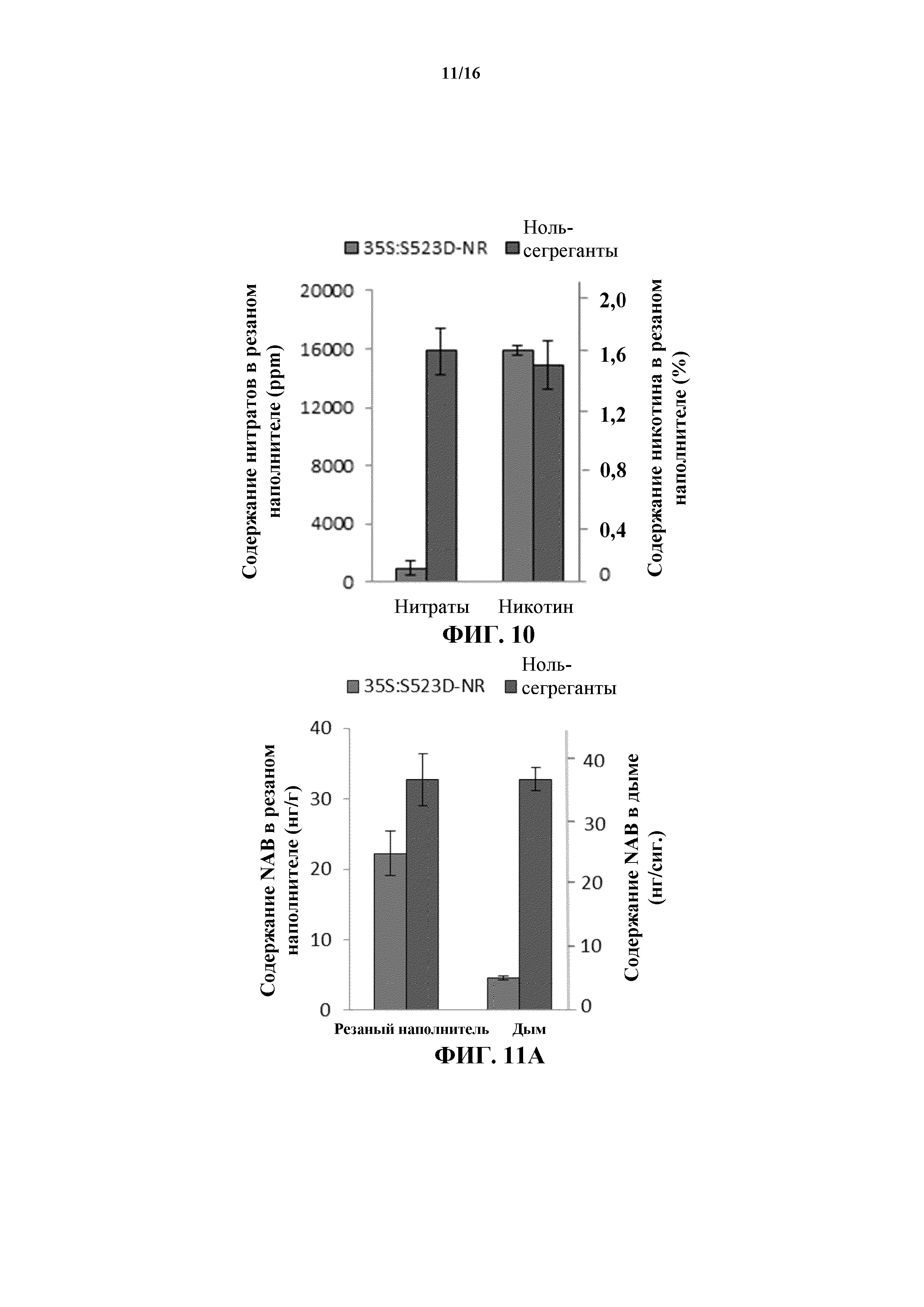
На фиг. 10 показано среднее содержание нитратов и никотина в резаном табачном наполнителе, полученном из листовой пластинки генотипов WT и 35S:S523D-NR.
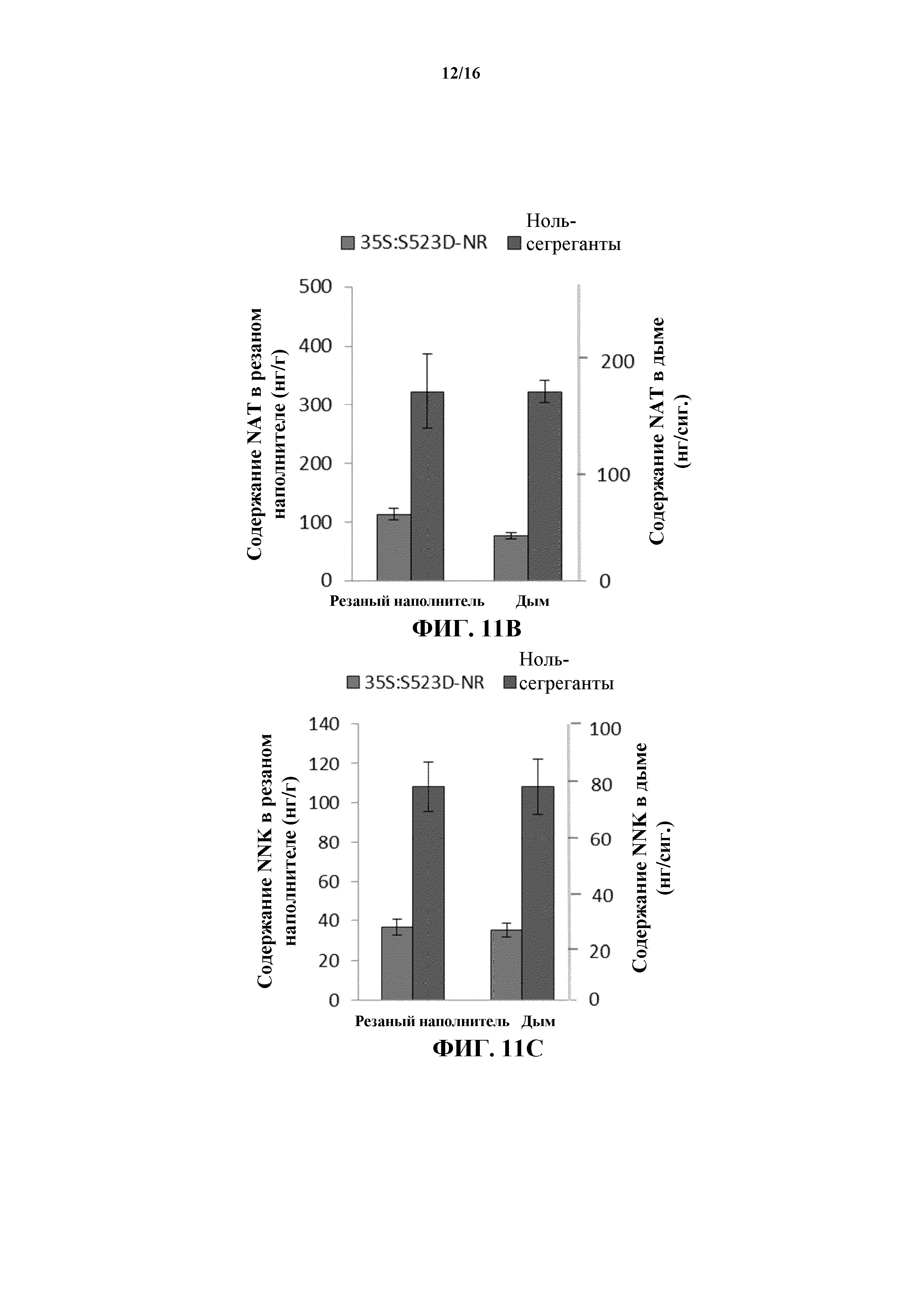
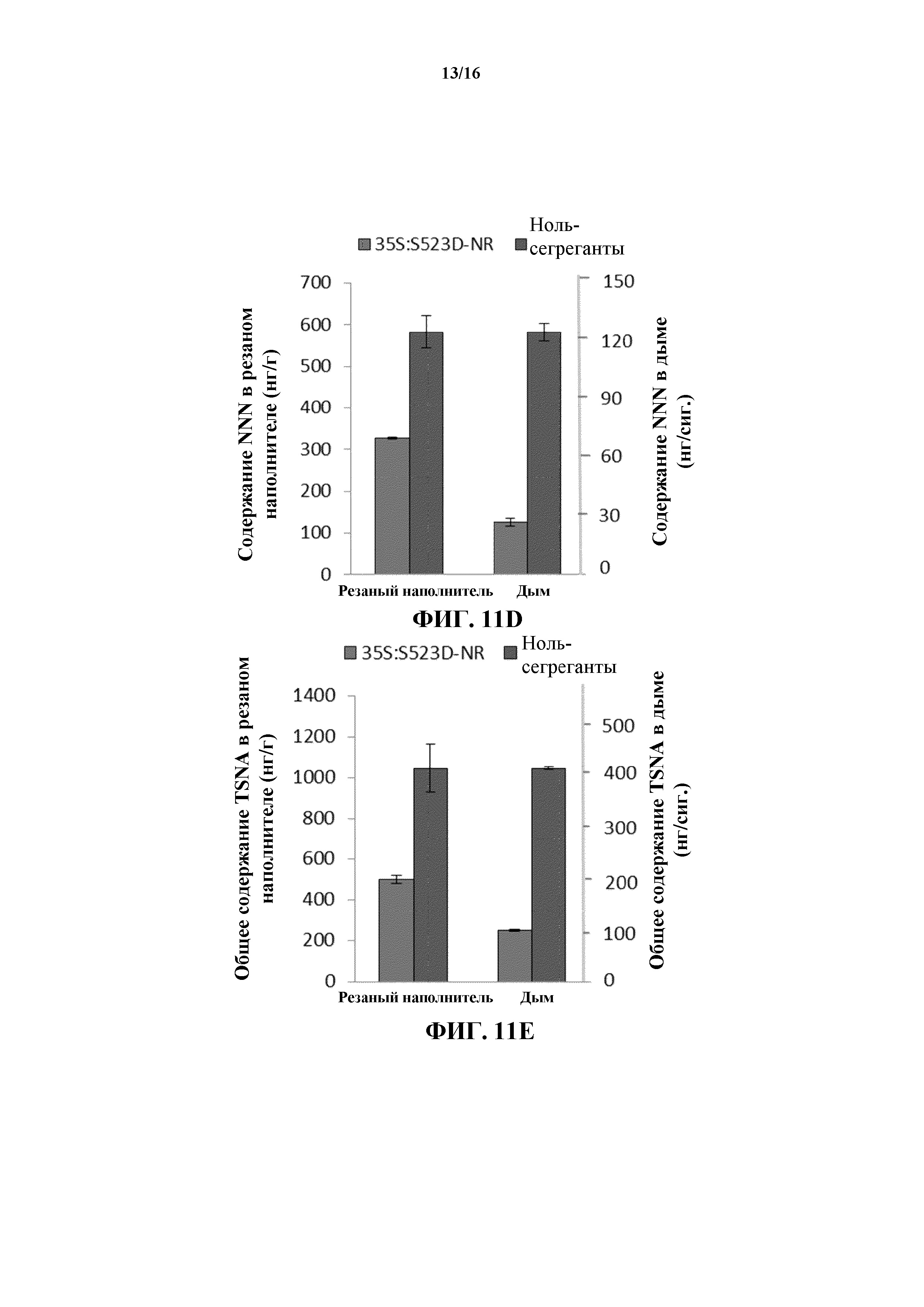
На фиг. 11A-11E показано среднее содержание NAB (на фиг. 11A), NAT (на фиг. 11B), NNK (на фиг. 11C), NNN (на фиг. 11D) и общее содержание TSNA (на фиг. 11E) в резаном наполнителе и дыме сигарет, изготовленных из листовой пластинки листьев WT и 35S:S523D-NR.
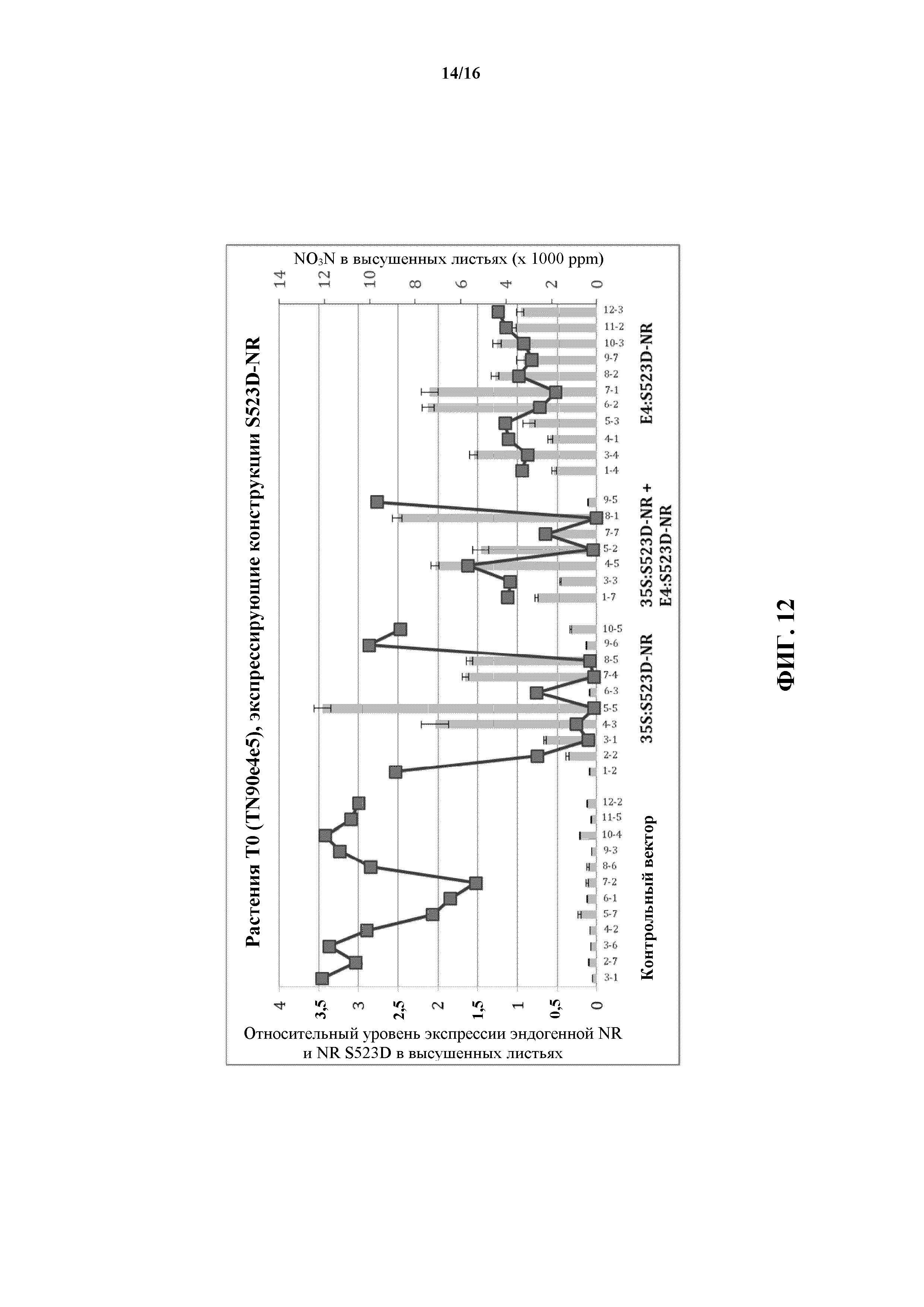
На фиг. 12 показаны концентрация нитратов и относительные уровни NR-транскрипта в обработанных этефоном листьях табака, содержащих конструкции S523D-NR под транскрипционным контролем промотора 35S CaMV и/или этефон-индуцируемого промотора CYP82E4 (E4).
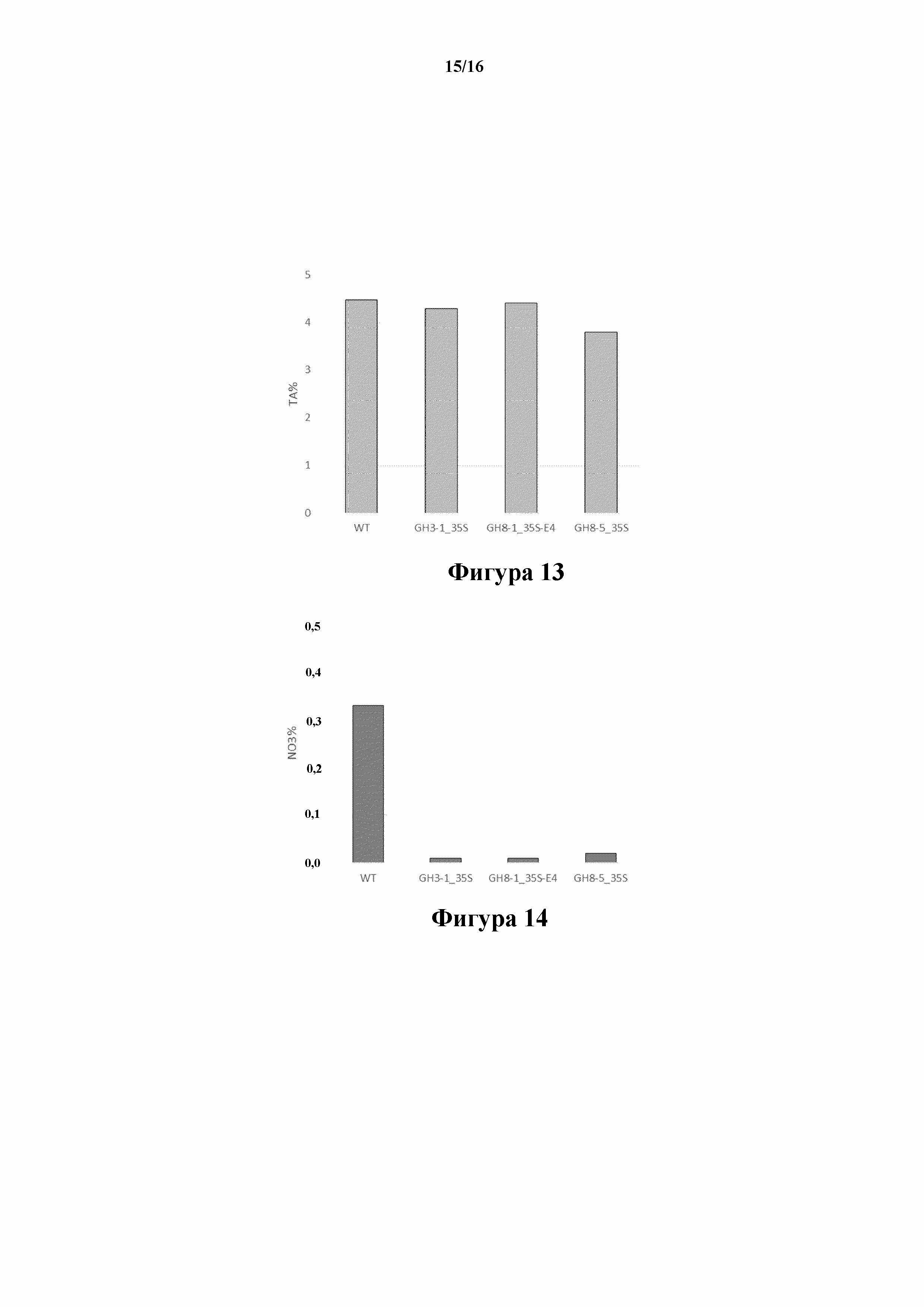
На фиг. 13 показано общее содержание алкалоидов (TA, % DW) в трансгенных линиях 35S:S523D-NR и 35S:S523D-NR/E4:S523D-NR, соответственно GH3-1, GH8-1 и GH8-5.
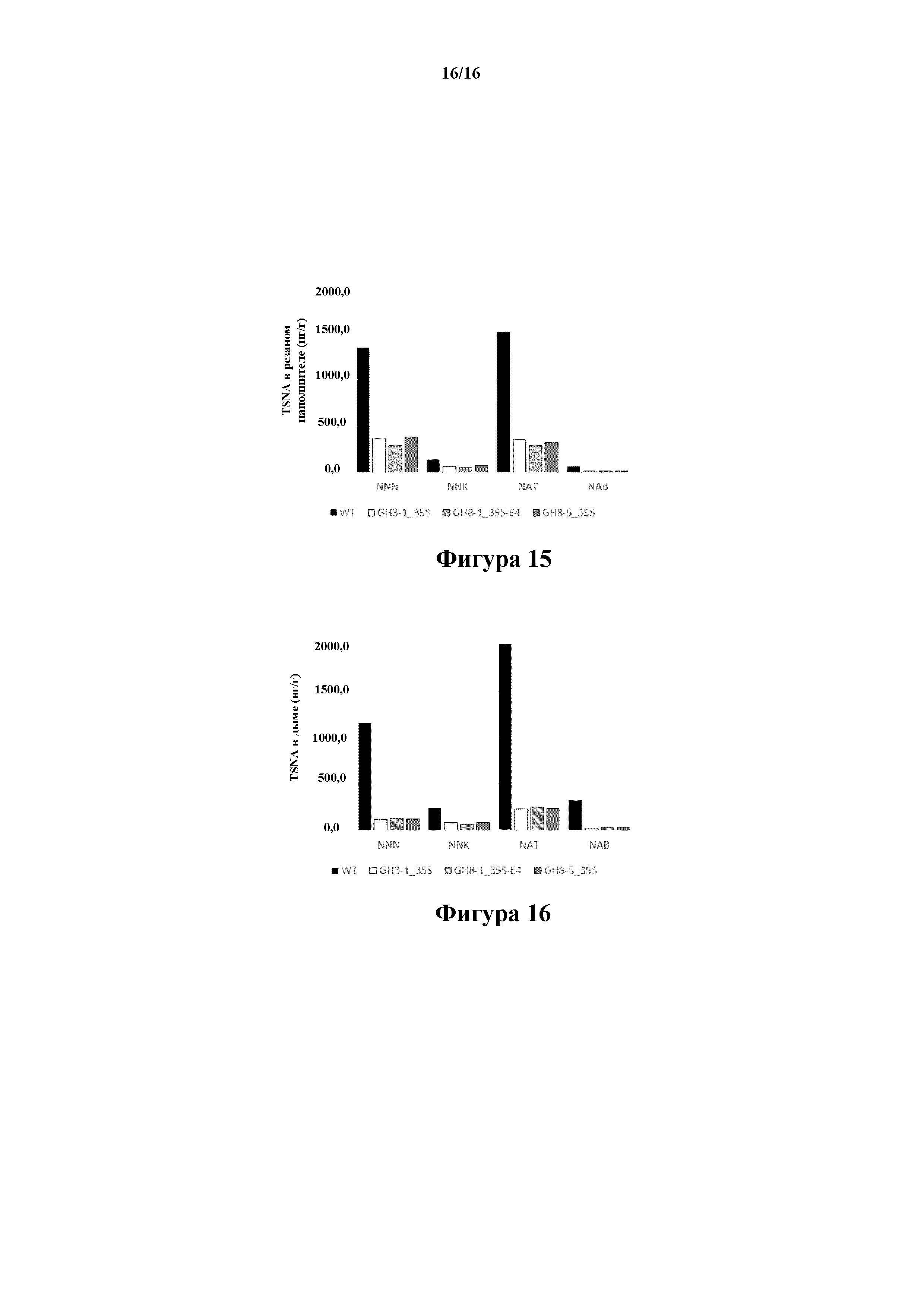
На фиг. 14 проиллюстрировано содержание нитратов (NO3, % DW) в трех трансгенных линиях 35S:S523D-NR и 35S:S523D-NR/E4:S523D-NR, соответственно GH3-1, GH8-1 и GH8-5.
На фиг. 15 показано содержание TSNA (NNN, NNK, NAT и NAB, нг/г) в резаном наполнителе из трех трансгенных линий 35S:S523D-NR и 35S:S523D-NR/E4:S523D-NR, соответственно GH3-1, GH8-1 и GH8-5.
На фиг.16 показано содержание TSNA (NNN, NNK, NAT и NAB, нг/г) в дыме сигарет, изготовленных с использованием резаного наполнителя из трех трансгенных линий 35S:S523D-NR и 35S:S523D-NR/E4:S523D-NR, соответственно GH3-1, GH8-1 и GH8-5.
Подробное описание
Настоящее изобретение в одном аспекте направлено на новую стратегию уменьшения уровней свободных нитратов в листьях модифицированных растений табака посредством изменения пути ассимиляции нитратов с уменьшением таким образом уровней всех TSNA в высушенных листьях модифицированного растения табака. Уровни свободных нитратов, а также содержание и уровни TSNA, в том числе TSNA в целом и/или конкретных TSNA, в листьях табака уменьшались с помощью дерегулированных нитратредуктаз. Путем целенаправленного воздействия на нитрозирующее средство и уменьшения количества свободных нитратов, запасаемых в листе, выработку канцерогенных TSNA в табачных продуктах сводят к минимуму. Данная технология представляет собой новый подход к уменьшению содержания TSNA, который мог бы способствовать получению в табачной промышленности табачных продуктов с более низкими уровнями TSNA, чем это возможно при применении существующих методик.
Дерегулированные нитратредуктазы были описаны у Nicotiana plumbaginofolia, например, Nussaume et al. Plant Cell 7:611-621 (1995), а также Lea et al., Plant Physiology 140:1085-1094 (2006). Уменьшение уровней нитратов, наблюдаемое у N. plumbaginofolia, значительно уступает тому, которое наблюдается у N. tabacum в данных, представленных в данном документе. Кроме того, уменьшение уровней TSNA, обусловленное дерегуляцией нитратредуктазы, ранее не было продемонстрировано.
Заголовки разделов, используемые в данном разделе, а также во всем раскрытии в данном документе, приводятся лишь в организационных целях и не предназначены для ограничения.
Определения
Если не определено иное, то все технические и научные термины, используемые в данном документе, имеют такое же значение, какое обычно понимает средний специалист в данной области. В случае противоречий данный документ, включая определения, будет иметь преимущественную силу. Предпочтительные способы и материалы описаны ниже, хотя способы и материалы, подобные или эквивалентные описанным в данном документе, могут быть применены на практике или при тестировании настоящего изобретения. Все публикации, патентные заявки, патенты и другие литературные источники, упомянутые в данном документе, включены посредством ссылки во всей своей полноте. Материалы, способы и примеры, раскрытые в данном документе, являются лишь иллюстративными и не предназначены для ограничения.
Подразумевается, что термины «включают(включает)», «включают(включает) в себя», «характеризуются», «имеет», «может», «содержат(содержит)» и их варианты, используемые в данном документе, являются открытыми переходными фразами, терминами или словами, которые не исключают возможность наличия дополнительных действий или структур. Формы единственного числа включают ссылки на множественное число, если по контексту явно не требуется иное. Настоящее раскрытие также охватывает другие варианты осуществления, «содержащие» варианты осуществления или элементы, представленные в данном документе, «состоящие из» и «по сути состоящие из» них, независимо от того, указано это явно или нет. В случае перечисления в данном документе числовых диапазонов каждое промежуточное число в них предусматривается в явной форме с той же степенью точности. Например, в случае диапазона 6-9 в дополнение к 6 и 9 предусматриваются числа 7 и 8, а в случае диапазона 6,0-7,0 в явной форме предусматриваются числа 6,0, 6,1, 6,2, 6,3, 6,4, 6,5, 6,6, 6,7, 6,8, 6,9 и 7,0.
Следующие термины, используемые во всем данном описании и формуле изобретения, имеют следующие значения.
Выражения «кодирующая последовательность» или «кодирующая нуклеиновая кислота», используемые в данном документе, означают нуклеиновые кислоты (молекулы РНК или ДНК), которые содержат нуклеотидную последовательность, кодирующую белок. Кодирующая последовательность может дополнительно содержать сигналы инициации и терминации, функционально связанные с регуляторными элементами, в том числе c промотором и сигналом полиаденилирования, способными управлять экспрессией в клетках индивидуума или млекопитающего, которому вводят нуклеиновую кислоту. Кодирующая последовательность может быть кодон-оптимизированной.
Выражения «комплементарная последовательность» или «комплементарный», используемые в данном документе, означают нуклеиновую кислоту, для которой может подразумеваться спаривание оснований по Уотсону-Крику (например, A-T/U и C-G) или Хугстину между нуклеотидами или аналогами нуклеотидов молекул нуклеиновых кислот. «Комплементарность» относится к свойству, которым совместно обладают две последовательности нуклеиновых кислот, заключающемуся в том, что в случае, когда они выровнены друг с другом антипараллельно, нуклеотидные основания в каждом положении будут комплементарны друг другу.
Выражение «конструкция», используемое в данном документе, относится к фрагменту двухнитевой рекомбинантной нуклеиновой кислоты, содержащему один или несколько полинуклеотидов. Конструкция содержит «матричную цепь», основания которой спарены с комплементарной «смысловой» или «кодирующей» цепью. Указанная конструкция может быть внедрена в вектор в двух возможных ориентациях, в той же (или смысловой) ориентации либо в противоположной (или антисмысловой) ориентации по отношению к ориентации промотора, расположенного в векторе, таком как вектор экспрессии.
Термин «контроль», используемый в данном документе применительно к контрольному растению или контрольным растительным клеткам, означает растение или растительные клетки, в которых экспрессия или активность нитратредуктазы не была модифицирована (например, увеличена или снижена), и поэтому они могут обеспечивать возможность сравнения с растением, в котором экспрессия или активность нитратредуктазы была модифицирована. Используемое в данном документе выражение «контрольное растение» означает растение, которое является практически эквивалентным тестируемому растению или модифицированному растению по всем параметрам, за исключением тестируемых параметров. Например, когда речь идет о растении, в которое был введен полинуклеотид в соответствии с настоящим изобретением, в некоторых вариантах осуществления контрольным растением является эквивалентное растение, в которое такой полинуклеотид не был введен. В некоторых вариантах осуществления контрольным растением является эквивалентное растение, в которое был введен контрольный полинуклеотид. В таких случаях контрольный полинуклеотид представляет собой полинуклеотид, для которого обуславливаемый им фенотипический эффект в отношении растения, как предполагается, является незначительным или отсутствует. Контрольное растение может содержать пустой вектор. Контрольное растение может соответствовать растению дикого типа. Контрольное растение может быть ноль-сегрегантом, где сегрегант T1 больше не имеет трансген.
Выражения «донорная ДНК» или «донорная матрица», которые используются в данном документе взаимозаменяемо, относятся к фрагменту или молекуле двухнитевой ДНК, которые содержат по меньшей мере часть гена, представляющего интерес. Донорная ДНК может кодировать полнофункциональный белок или частично функциональный белок.
Выражение «эндогенный ген», используемое в данном документе, относится к гену, который происходит из генома организма и не претерпел изменения, такого как потеря, приобретение или замена генетического материала. Эндогенный ген подвергается нормальному переносу генов и экспрессии генов.
Выражение «энхансерные последовательности», используемое в данном документе, относится к последовательностям, которые могут увеличивать экспрессию генов. Эти последовательности могут быть расположены выше, в пределах интронов или ниже транскрибируемого участка. Транскрибируемый участок состоит из экзонов и промежуточных интронов от промотора до участка терминации транскрипции. Усиление экспрессии генов может происходить посредством различных механизмов, которые включают без ограничения увеличение эффективности транскрипции, стабилизацию зрелой мРНК и усиление трансляции.
Выражение «экспрессия», используемое в данном документе, относится к выработке функционального продукта. Например, экспрессия фрагмента нуклеиновой кислоты может относиться к транскрипции фрагмента нуклеиновой кислоты (например, транскрипции, приводящей к получению мРНК или функциональной РНК) и/или трансляции мРНК в молекулу-предшественника или зрелый белок. «Сверхэкспрессия» относится к выработке продукта гена в трансгенных организмах на уровнях, которые превышают уровни выработки в ноль-сегрегантном (или нетрансгенном) организме из того же эксперимента.
Выражения «функциональный» и «полнофункциональный», используемые в данном документе, описывают белок, который обладает биологической активностью. Выражение «функциональный ген» относится к гену, транскрибируемому в мРНК, которая транслируется в функциональный белок.
Выражение «генетическая конструкция», используемое в данном документе, относится к молекулам ДНК или РНК, которые содержат нуклеотидную последовательность, кодирующую белок. Кодирующая последовательность содержит сигналы инициации и терминации, функционально связанные с регуляторными элементами, в том числе с промотором и сигналом полиаденилирования, способными управлять экспрессией в клетках индивидуума, которому вводят молекулу нуклеиновой кислоты.
Выражение «редактирование генома», используемое в данном документе, относится к такому изменению эндогенного гена, кодирующего эндогенный полипептид или белок, при котором достигается экспрессия усеченного эндогенного белка или эндогенного белка, имеющего аминокислотную замену. Редактирование генома может включать в себя замещение участка эндогенного гена, подлежащего целенаправленному воздействию, или замещение всего эндогенного гена копией гена, которая имеет усечение или аминокислотную замену, с помощью механизма репарации, такого как репарация путем гомологичной рекомбинации (HDR). Редактирование генома может также включать в себя создание аминокислотной замены в эндогенном гене посредством создания двухнитевого разрыва в эндогенном гене, который затем репарируют с помощью негомологичного соединения концов (NHEJ). Путем NHEJ можно производить добавление или делецию по меньшей мере одной пары оснований в ходе репарации, вследствие чего может создаваться аминокислотная замена. Редактирование генома может включать в себя также делецию сегмента гена посредством одновременного действия двух нуклеаз на одну и ту же нить ДНК для того, чтобы создать усечение между двумя целевыми сайтами для нуклеаз, и репарацию разрыва ДНК с помощью NHEJ.
Выражение «гетерологичный», используемое в данном документе по отношению к последовательности, означает последовательность, которая происходит из чужеродного вида или, в случае, если она происходит из того же вида, является существенно модифицированной по сравнению со своей нативной формой в композиции и/или геномном локусе вследствие преднамеренного вмешательства человека.
Выражения «репарация путем гомологичной рекомбинации» или «HDR», используемые в данном документе взаимозаменяемо, относятся к механизму репарации двухнитевых повреждений ДНК в клетках в тех условиях, когда гомологичный фрагмент ДНК присутствует в ядре, главным образом в G2- и S-фазе клеточного цикла. В ходе HDR используют донорную ДНК или донорную матрицу для управления репарацией, и ее можно использовать для создания специфических изменений последовательностей в геноме, в том числе для направленного добавления целых генов. Если донорная матрица предоставляется вместе с сайт-специфичной нуклеазой, то клеточный аппарат будет репарировать разрыв с помощью гомологичной рекомбинации, которая усиливается по величине на несколько порядков при наличии расщепления ДНК. Если гомологичный фрагмент ДНК отсутствует, то вместо этого может происходить негомологичное соединение концов.
Термины «гомология» или «подобие», используемые в данном документе, относятся к степени подобия последовательностей между двумя полипептидами или между двумя молекулами нуклеиновых кислот, сравниваемыми с помощью выравнивания последовательностей. Степень гомологии между двумя отдельными сравниваемыми последовательностями нуклеиновых кислот зависит от количества идентичных или совпадающих нуклеотидов в сопоставимых положениях.
Выражения «идентичный» или «идентичность», используемые в данном документе применительно к двум или более нуклеиновым кислотам или полипептидным последовательностям, означают, что последовательности характеризуются определенной процентной долей остатков, которые являются одинаковыми на протяжении определенного участка. Процентную долю можно рассчитать посредством оптимального выравнивания двух последовательностей, сравнения двух последовательностей на протяжении определенного участка, определения числа положений, в которых находятся идентичные остатки в обеих последовательностях, с получением числа совпадающих положений, деления числа совпадающих положений на общее число положений в определенном участке и умножения результата на 100 с получением процентного значения идентичности последовательностей. В тех случаях, когда эти две последовательности имеют разную длину или при выравнивании создается один или несколько несимметрично расположенных концов, а определенный участок сравнения включает в себя только одну последовательность, то при расчете остатки одной последовательности включаются в знаменатель, а не в числитель. При сравнении ДНК и РНК тимин (Т) и урацил (U) можно считать эквивалентными. Идентичность можно определять вручную или с помощью компьютерного алгоритма для работы с последовательностями, такого как ClustalW, ClustalX, BLAST, FASTA или алгоритм Смита-Уотермана.
Термин «увеличение» или «увеличенный», используемый в данном документе, относится к увеличению, составляющему от приблизительно 10% до приблизительно 99%, или увеличению, составляющему по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 100%, по меньшей мере 150% или по меньшей мере 200% или более от количества или активности, как, например, без ограничения, уровней нитратов или уровней TSNA.
Термин «ингибировать» или «ингибированный», используемый в данном документе, относится к уменьшению, составляющему от приблизительно 98% до приблизительно 100%, или уменьшению, составляющему по меньшей мере 98%, по меньшей мере 99% и, в частности, 100% от количества или активности, как, например, без ограничения, активности полипептида, транскрипционной активности и/или экспрессии белка.
Термин «введенный», используемый в данном документе, означает предоставление нуклеиновой кислоты (например, экспрессионной конструкции) или белка в клетку. Выражение «введенный» включает ссылку на включение нуклеиновой кислоты в эукариотическую клетку, где нуклеиновая кислота может быть включена в геном клетки, а также включает ссылку на транзиентное внедрение нуклеиновой кислоты или белка в клетку. Выражение «введенный» включает ссылки на способы стабильной или транзиентной трансформации, а также на половое скрещивание. Таким образом, выражение «введенный» применительно к внедрению фрагмента нуклеиновой кислоты (например, рекомбинантной ДНК-конструкции/экспрессионной конструкции) в клетку означает «трансфекцию», или «трансформацию», или «трансдукцию» и включает ссылку на включение фрагмента нуклеиновой кислоты в эукариотическую клетку, где фрагмент нуклеиновой кислоты может включаться в геном клетки (например, хромосомную, плазмидную, пластидную или митохондриальную ДНК), преобразовываться в автономный репликон или транзиентно экспрессироваться (например, в виде трансфицированной мРНК).
Термины «выделенный», «очищенный» или «биологически чистый», используемые в данном документе, относятся к материалу, который практически или по сути не содержит компоненты, обычно сопутствующие ему, как встречается в его нативном состоянии. Как правило, чистота и однородность определяются с помощью методик аналитической химии, таких как электрофорез в полиакриламидном геле или высокоэффективная жидкостная хроматография. Белок, который является преобладающей молекулой, присутствующей в препарате, является практически очищенным. В частности, выделенная нуклеиновая кислота по настоящему изобретению отделена от открытых рамок считывания, которые фланкируют требуемый ген и кодируют белки, отличные от требуемого белка. Термин «очищенный», используемый в данном документе, обозначает, что нуклеиновая кислота или белок дают по сути одну полосу в электрофорезном геле. В частности, это означает, что нуклеиновая кислота или белок являются по меньшей мере на 85% чистыми, более предпочтительно по меньшей мере на 95% чистыми и наиболее предпочтительно по меньшей мере на 99% чистыми.
Выделенные полинуклеотиды могут быть очищены из клетки-хозяина, в которой они встречаются в природе. Для получения выделенных полинуклеотидов могут быть использованы традиционные способы очистки нуклеиновых кислот, известные специалистам в данной области. Данный термин также охватывает рекомбинантные полинуклеотиды и химически синтезируемые полинуклеотиды.
Выражение «нитратредуктаза», используемое в данном документе, относится к ферменту, который восстанавливает нитрат (NO3-) до нитрита (NO2-). Термин «дерегулированный фермент нитратредуктаза» обычно относится к ферменту нитратредуктазе, который не подпадает под действие одного или нескольких регуляторных механизмов (например, транскрипционного, посттранскрипционного или посттрансляционного регуляторных механизмов), которые контролируют или ограничивают экспрессию или активность нитратредуктазы в контрольном немодифицированном растении табака. Таким образом, выражение «экспрессия или активность указанной нитратредуктазы является дерегулированной» обычно означает, что экспрессия или активность нитратредуктазы является увеличенной в модифицированном растении табака по сравнению с контрольным немодифицированным растением табака. Термин «полинуклеотид, кодирующий дерегулированную нитратредуктазу» включает полинуклеотиды, кодирующие модифицированную нитратредуктазу (NR) из Nicotiana tabacum, и включает полинуклеотиды, содержащие полинуклеотиды с существенной степенью гомологии (т. е. подобия последовательности) или существенной степенью идентичности с SEQ ID NO: 5 или 7; варианты полинуклеотидов, характеризующиеся по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с последовательностью SEQ ID NO: 5 или 7; фрагменты полинуклеотидов, в том числе фрагменты SEQ ID NO: 5 или 7; и фрагменты SEQ ID NO: 5 или 7 с существенной степенью гомологии (т. е. подобия последовательности) или существенной степенью идентичности с ними, характеризующиеся по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью последовательности с соответствующими фрагментами SEQ ID NO: 5 или 7, состоящие из них или по сути состоящие из них. Фрагменты могут иметь длину по меньшей мере приблизительно 20, 50, 70, 100, 200, 300, 500, 1000 или 2000 нуклеотидов, например, полинуклеотид, кодирующий дерегулированную нитратредуктазу, может содержать (i) по меньшей мере приблизительно 20, 50, 70, 100, 200, 300, 500, 1000 или 2000 нуклеотидов SEQ ID NO: 5 или 7 или (ii) последовательность, характеризующуюся по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью последовательности по меньшей мере с приблизительно 20, 50, 70, 100, 200, 300, 500, 1000 или 2000 нуклеотидов SEQ ID NO: 5 или 7. Как правило, фрагмент может сохранять биологическую активность последовательности полной длины, например, фрагмент отвечает за нитратредуктазную активность, как правило, модифицированную или дерегулированную нитратредуктазную активность. Полипептидная нитратредуктаза также содержит последовательности, обладающие достаточной или существенной степенью идентичности или подобия с SEQ ID NO: 5 или 7, чтобы кодировать полипептид, который функционирует в качестве нитратредуктазы. В одном варианте осуществления термин «полинуклеотид, кодирующий нитратредуктазу» относится к полимеру из нуклеотидов, который содержит полинуклеотид, обозначенный в данном документе как SEQ ID NO: 1 или 3, состоящие из них или по сути состоящие из них.
Термин «дерегулированная полипептидная нитратредуктаза» включает модифицированную нитратредуктазу (NR) из Nicotiana tabacum и включает полипептиды, содержащие полипептиды с существенной степенью гомологии (т. е. подобия последовательности) или существенной степенью идентичности с SEQ ID NO: 6 или 8 или варианты полипептидов, характеризующиеся по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с последовательностью SEQ ID NO: 6 или 8; фрагменты полипептидов, в том числе фрагменты SEQ ID NO: 6 или 8; и фрагменты SEQ ID NO: 6 или 8 с существенной степенью гомологии (т. е. подобия последовательности) или существенной степенью идентичности с ними, характеризующиеся по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью последовательности с соответствующими фрагментами SEQ ID NO: 6 или 8, состоящие из них или по сути состоящие из них. Фрагменты могут иметь длину по меньшей мере приблизительно 10, 30, 50, 100, 200, 300, 400, 500, 600, 700, 800 или 900 аминокислотных остатков, например, дерегулированная полипептидная нитратредуктаза может содержать (i) по меньшей мере приблизительно 10, 30, 50, 100, 200, 300, 400, 500, 600, 700, 800 или 900 аминокислот SEQ ID NO: 6 или 8 или (ii) последовательность, характеризующуюся по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью последовательности по меньшей мере с приблизительно 10, 30, 50, 100, 200, 300, 400, 500, 600, 700, 800 или 900 аминокислот SEQ ID NO: 6 или 8. Как правило, фрагмент может сохранять биологическую активность последовательности полной длины, например, фрагмент обладает нитратредуктазной активностью, как правило, модифицированной или дерегулированной нитратредуктазной активностью. Полипептидная нитратредуктаза также содержит последовательности, характеризующиеся достаточной или существенной степенью идентичности или подобия с SEQ ID NO: 6 или 8, которые функционируют в качестве нитратредуктазы. В одном варианте осуществления термин «полипептидная нитратредуктаза» относится к полимеру из аминокислот, который содержит полипептид, обозначенный в данном документе как SEQ ID NO: 2 или 4, состоит из него или по сути состоит из него.
Выражение «путь негомологичного соединения концов (NHEJ)», используемое в данном документе, относится к пути, за счет которого происходит репарация двухнитевых разрывов в ДНК путем прямого лигирования концов разрывов без необходимости в гомологичной матрице. Независимое от матрицы повторное лигирование концов ДНК с помощью NHEJ является стохастическим, подверженным ошибкам процессом репарации, в ходе которого в точку разрыва ДНК вводятся случайные микровставки и микроделеции (вставки/делеции). Данный способ можно использовать для преднамеренного разрушения, делеции или изменения рамки считывания последовательностей целевых генов. Как правило, в ходе NHEJ используют короткие гомологичные последовательности ДНК, называемые микрогомологами, для управления репарацией. Эти микрогомологи часто присутствуют в однонитевых выступах на концах двухнитевых разрывов. Если выступы полностью совместимы, то в ходе NHEJ обычно происходит точная репарация разрыва, хотя может иметь место также неточная репарация, приводящая к потере нуклеотидов, однако гораздо более распространены случаи, когда выступы несовместимы. Выражение «NHEJ, опосредованное нуклеазами», используемое в данном документе, относится к NHEJ, инициируемому после разреза двухнитевой ДНК нуклеазой.
Выражение «N-концевое усечение», используемое в данном документе, означает делецию аминокислот на N-конце или вблизи него. В некоторых вариантах осуществления делеция аминокислот может иметь место на N-конце полипептида, т. е. делеция охватывает первую аминокислоту транскрибированного или зрелого белка. В некоторых вариантах осуществления делеция аминокислот может иметь место в пределах N-конца полипептида, т. е. делеция не охватывает первую аминокислоту транскрибированного или зрелого белка, но имеет место в пределах первой половины данного полипептида.
Выражения «нуклеиновая кислота», или «олигонуклеотид», или «полинуклеотид», используемые в данном документе, означают по меньшей мере два нуклеотида, ковалентно связанных друг с другом. Описание отдельной нити также определяет последовательность комплементарной нити. Таким образом, нуклеиновая кислота также охватывает нить, комплементарную описанной отдельной нити. Многие варианты нуклеиновой кислоты могут использоваться для той же цели, что и указанная нуклеиновая кислота. Таким образом, нуклеиновая кислота также охватывает практически идентичные нуклеиновые кислоты и комплементарные им последовательности. Отдельная нить представляет собой зонд, который может гибридизироваться с целевой последовательностью в жестких условиях гибридизации. Таким образом, нуклеиновая кислота также охватывает зонд, который гибридизируется в жестких условиях гибридизации.
Нуклеиновые кислоты могут быть однонитевыми или двухнитевыми или могут содержать части как двухнитевой, так и однонитевой последовательности. Нуклеиновая кислота может представлять собой ДНК, как геномную, так и кДНК, РНК или гибридную молекулу, где нуклеиновая кислота может содержать комбинации дезоксирибо- и рибонуклеотидов, а также комбинации оснований, включающих урацил, аденин, тимин, цитозин, гуанин, инозин, ксантин, гипоксантин, изоцитозин и изогуанин. Нуклеиновые кислоты можно получить с помощью способов химического синтеза или с помощью рекомбинантных способов.
Специфичность однонитевой ДНК для гибридизации с комплементарными фрагментами определяется «жесткостью» условий реакции (Sambrook et al., Molecular Cloning and Laboratory Manual, Second Ed., Cold Spring Harbor (1989)). По мере снижения склонности к образованию ДНК-дуплексов увеличивается жесткость гибридизации. В реакциях гибридизации нуклеиновых кислот можно выбрать жесткость для содействия специфичным реакциям гибридизации (высокая жесткость), которые можно использовать для идентификации, например, клонов полной длины из библиотеки. Менее специфичные реакции гибридизации (низкая жесткость) можно использовать для идентификации родственных, но не точно соответствующих (гомологичных, но не идентичных) молекул или сегментов ДНК.
ДНК-дуплексы стабилизируются в соответствии с: (1) числом комплементарных пар оснований; (2) типом пар оснований; (3) концентрацией солей (ионной силы) в реакционной смеси; (4) температурой реакции и (5) присутствием определенных органических растворителей, таких как формамид, снижающих стабильность ДНК-дуплекса. Обычно чем длиннее зонд, тем выше температура, необходимая для надлежащего отжига. Общепринятый подход заключается в изменении температуры; более высокие относительные температуры приводят к более жестким условиям реакции.
Для гибридизации в «жестких условиях» описаны протоколы гибридизации, в которых нуклеотидные последовательности, по меньшей мере на 60% гомологичные друг другу, остаются гибридизированными. Обычно жесткие условия выбирают таким образом, чтобы температура была приблизительно на 5°C ниже, чем температура точки плавления (Tm) для конкретной последовательности при определенных значениях ионной силы и рН. Tm представляет собой температуру (при определенных значениях ионной силы, рН и концентрации нуклеиновой кислоты), при которой 50% зондов, комплементарных целевой последовательности, гибридизируются с целевой последовательностью в равновесном состоянии. Поскольку целевые последовательности обычно присутствуют в избытке, то при Tm 50% зондов заняты в равновесном состоянии.
«Жесткие условия гибридизации» представляют собой условия, которые позволяют зонду, праймеру или олигонуклеотиду гибридизироваться только со своей целевой последовательностью (например, SEQ ID NO: 1, 3, 5 или 7). Жесткие условия зависят от последовательности и будут различаться. Жесткие условия предусматривают: (1) низкую ионную силу и промывки при высокой температуре, например, 15 мМ хлорида натрия, 1,5 мМ цитрата натрия, 0,1% додецилсульфата натрия при 50°C; (2) присутствие денатурирующего средства во время гибридизации, например, 50% (об./об.) формамида, 0,1% бычьего сывороточного альбумина, 0,1% фиколла, 0,1% поливинилпирролидона, 50 мМ натрий-фосфатного буфера (750 мМ хлорида натрия, 75 мМ цитрата натрия; pH 6,5) при 42°C или (3) присутствие 50% формамида. Как правило, промывки также предусматривают 5 x SSC (0,75 M NaCl, 75 мМ цитрата натрия), 50 мМ фосфата натрия (pH 6,8), 0,1% пирофосфат натрия, 5 x раствор Денхардта, ДНК из молок лососевых рыб, подвергнутую ультразвуковой обработке (50 мкг/мл), 0,1% SDS и 10% сульфат декстрана при 42°C с промывкой при 42°C в 0,2 x SSC (хлорид натрия/цитрат натрия) и 50% формамиде при 55°C и последующей промывкой в условиях высокой жесткости, предусматривающей 0,1 x SSC, содержащий EDTA, при 55°C. Предпочтительно условия являются такими, что последовательности, по меньшей мере приблизительно на 65%, 70%, 75%, 85%, 90%, 95%, 98% или 99% гомологичные друг другу, обычно остаются гибридизированными друг с другом. Эти условия представлены в качестве примеров и не предполагаются как ограничивающие.
В «условиях умеренной жесткости» используют моющие растворы и условия гибридизации, которые являются менее жесткими, так что полинуклеотид гибридизируется со всей целевой последовательностью, ее фрагментами, производными или аналогами (например, SEQ ID NO: 1, 3, 5 или 7). Один пример предусматривает гибридизацию в 6 x SSC, 5 x растворе Денхардта, 0,5% SDS и 100 мкг/мл денатурированной ДНК из молок лососевых рыб при 55°C с последующими одной или несколькими промывками в 1 × SSC, 0,1% SDS при 37°C. Температуру, ионную силу и т. д. можно корректировать в целях соответствия экспериментальным факторам, таким как длина зонда. Были описаны другие условия умеренной жесткости (Ausubel et al., Current Protocols in Molecular Biology, Volumes 1-3, John Wiley & Sons, Inc., Hoboken, N.J. (1993); Kriegler, Gene Transfer and Expression: A Laboratory Manual, Stockton Press, New York, N.Y. (1990); Perbal, A Practical Guide to Molecular Cloning, 2nd edition, John Wiley & Sons, New York, N.Y. (1988)).
В «условиях низкой жесткости» используют моющие растворы и условия гибридизации, которые являются менее жесткими, чем для умеренной жесткости, так что полинуклеотид гибридизируется со всей целевой последовательностью, ее фрагментами, производными или аналогами (например, SEQ ID NO: 1, 3, 5 или 7). Неограничивающий пример условий гибридизации низкой жесткости предусматривает гибридизацию в 35% формамиде, 5 x SSC, 50 мМ Tris-HCl (pH 7,5), 5 мМ EDTA, 0,02% PVP, 0,02% фиколле, 0,2% BSA, 100 мкг/мл денатурированной ДНК из молок лососевых рыб, 10% (вес/об.) сульфате декстрана при 40°C с последующими одной или несколькими промывками в 2 x SSC, 25 мМ Tris-HCl (pH 7,4), 5 мМ EDTA и 0,1% SDS при 50°C. Хорошо описаны другие условия низкой жесткости, такие как условия для перекрестной гибридизации (Ausubel et al., 1993; Kriegler, 1990).
Выражение «функционально связанный», используемое в данном документе, означает, что экспрессия гена находится под контролем промотора, с которым он пространственно соединен. Промотор может быть расположен в 5'-направлении (выше) или 3'-направлении (ниже) от гена, который находится под его контролем. Расстояние между промотором и геном может быть примерно таким же, как расстояние между этим промотором и геном, который он контролирует, в гене, из которого получен промотор. Как известно из уровня техники, изменение этого расстояния можно согласованно осуществлять без потери функции промотора. Выражение «функционально связанный» относится к ассоциации фрагментов нуклеиновой кислоты в одном фрагменте таким образом, что функция одного регулируется другим. Например, промотор функционально связан с фрагментом нуклеиновой кислоты, если он способен регулировать транскрипцию данного фрагмента нуклеиновой кислоты.
Термин «растение», используемый в данном документе, относится к любому растению на любой стадии его жизненного цикла или развития, а также к его потомству. В одном варианте осуществления растение представляет собой растение табака, которое относится к растению, принадлежащему к роду Nicotiana. Выражение «растение» включает в себя ссылку на целые растения, органы растений, растительные ткани, ростки, семена растений и растительные клетки, а также потомство растений. Растительные клетки включают без ограничения клетки из семян, суспензионных культур, зародышей, меристематических участков, каллюсной ткани, листьев, корней, побегов, гаметофитов, спорофитов, пыльцы и микроспор. Предпочтительные виды, сорта, гибриды и разновидности растений табака описаны в данном документе.
Выражения «полинуклеотид», «последовательность нуклеиновой кислоты», «нуклеотидная последовательность» или «фрагмент нуклеиновой кислоты» используются в данном документе взаимозаменяемо для обозначения полимера из РНК или ДНК, который является одно- или двухнитевым и необязательно содержит синтетические, неприродные или измененные нуклеотидные основания. Нуклеотиды (обычно находящиеся в форме их 5'-монофосфата) называют однобуквенным обозначением следующим образом: «А» для аденилата или дезоксиаденилата (соответственно для РНК или ДНК), «C» для цитидилата или дезоксицитидилата, «G» для гуанилата или дезоксигуанилата, «U» для уридилата, «T» для дезокситимидилата, «R» для пуринов (A или G), «Y» для пиримидинов (C или T), «K» для G или T, «H» для A, или C, или Т, «I» для инозина и «N» для любого нуклеотида. Полинуклеотид может представлять собой без ограничения геномную ДНК, комплементарную ДНК (кДНК), мРНК или антисмысловую РНК или их фрагмент(фрагменты). Кроме того, полинуклеотид может представлять собой однонитевую или двухнитевую нуклеиновую кислоту, которая является смесью однонитевых и двухнитевых участков, гибридную молекулу, содержащую ДНК и РНК, или гибридную молекулу со смесью однонитевых и двухнитевых участков или их фрагмент(фрагменты). Кроме того, полинуклеотид может содержать трехнитевые участки, содержащие ДНК, РНК или их обе, или их фрагмент(фрагменты). Полинуклеотид может содержать одно или несколько модифицированных оснований, таких как фосфотиоаты, и может представлять собой пептидную нуклеиновую кислоту (PNA). Как правило, полинуклеотиды могут быть собраны из выделенных или клонированных фрагментов кДНК, геномной ДНК, олигонуклеотидов или отдельных нуклеотидов или комбинации вышеперечисленного. Хотя полинуклеотидные последовательности, описанные в данном документе, показаны в виде последовательностей ДНК, последовательности включают в себя соответствующие им последовательности РНК и комплементарные (например, полностью комплементарные) им последовательности ДНК или РНК, в том числе обратно комплементарные им последовательности.
Выражения «полипептид», «пептид», «аминокислотная последовательность» и «белок» используются в данном документе взаимозаменяемо для обозначения полимера из аминокислотных остатков. Данные термины применимы к полимерам из аминокислот, в которых один или несколько аминокислотных остатков представляют собой искусственный химический аналог соответствующей встречающейся в природе аминокислоты, а также к встречающимся в природе полимерам из аминокислот. Термины «полипептид», «пептид», «аминокислотная последовательность» и «белок» также подразумевают модификации, в том числе, без ограничения, гликозилирование, присоединение липидов, сульфатирование, гамма-карбоксилирование остатков глутаминовой кислоты, гидроксилирование и ADP-рибозилирование.
Выражение «промотор», используемое в данном документе, означает синтетическую или полученную природным способом молекулу, которая способна придавать, активировать или усиливать экспрессию нуклеиновой кислоты в клетке. Выражение «промотор» относится к элементу/последовательности нуклеиновой кислоты, расположенным, как правило, выше по последовательности и функционально связанным с фрагментом двухнитевой нуклеиновой кислоты. Промоторы могут быть полностью получены из участков вблизи нативного гена, представляющего интерес, или могут содержать разные элементы, полученные из разных нативных промоторов или сегментов синтетической нуклеиновой кислоты. Промотор может содержать одну или несколько специфических последовательностей, регулирующих транскрипцию, для дополнительного усиления экспрессии и/или изменения пространственной экспрессии и/или также временной экспрессии. Промотор может также содержать дистальные энхансерные или репрессорные элементы, которые могут быть расположены на расстоянии до несколько тысяч пар оснований от сайта начала транскрипции. Промотор может быть получен из источников, включающих вирусы, бактерии, грибы, растения, насекомых и животных. Промотор может регулировать экспрессию компонента гена конститутивно или дифференциально по отношению к клетке, ткани или органу, в которых происходит экспрессия, или по отношению к стадии развития, на которой происходит экспрессия, или в ответ на внешние стимулы, такие как физиологические нагрузки, патогены, ионы металлов или индуцирующие средства. Выражения «тканеспецифичный промотор» и «промотор, предпочтительный для определенной ткани», используемые в данном документе взаимозаменяемо, относятся к промотору, который экспрессируется преимущественно, но не обязательно исключительно в одном органе или ткани, но может экспрессироваться также в одной конкретной клетке. Выражение «промотор, регулируемый в процессе развития», используемое в данном документе, относится к промотору, активность которого определяется событиями, связанными с развитием. Промоторы, которые вызывают экспрессию гена в большинстве типов клеток в большинстве случаев, обычно называют «конститутивными промоторами». Индуцируемые промоторы обеспечивают избирательную экспрессию функционально связанной последовательности ДНК в ответ на присутствие эндогенных или экзогенных стимулов, например, с помощью химических соединений (химических индукторов) или в ответ на сигналы окружающей среды, гормональные, химические сигналы и/или сигналы, связанные с развитием. Примеры индуцируемых или регулируемых промоторов включают без ограничения промоторы, регулируемые светом, теплом, стрессом, наводнением или засухой, патогенами, фитогормонами, ранениями или химическими веществами, такими как этанол, жасмонат, салициловая кислота или антидоты.
Выражение «рекомбинантный», используемое в данном документе, относится к искусственной комбинации двух в иных обстоятельствах разделенных сегментов последовательности, полученной, например, посредством химического синтеза или посредством манипуляции с выделенными сегментами нуклеиновых кислот с помощью методик генной инженерии. Выражение «рекомбинантный» также включает ссылку на клетку или вектор, которые были модифицированы путем введения гетерологичной нуклеиновой кислоты, или клетку, полученную из модифицированной таким образом клетки, но не охватывает изменение клетки или вектора в результате встречающихся в природе событий (например, в результате спонтанной мутации, естественной трансформации/трансдукции/транспозиции), таких как те, которые происходят без преднамеренного вмешательства человека.
Выражение «рекомбинантная ДНК-конструкция», используемое в данном документе, относится к комбинации фрагментов нуклеиновых кислот, которые обычно не встречаются в природе вместе. Соответственно, рекомбинантная ДНК-конструкция может содержать регуляторные последовательности и кодирующие последовательности, полученные из разных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного и того же источника, но расположенные иначе, чем это обычно встречается в природе. Термины «рекомбинантная ДНК-конструкция» и «рекомбинантная конструкция» используются в данном документе взаимозаменяемо.
Термин «уменьшение» или «уменьшенный», используемый в данном документе, относится к уменьшению, составляющему от приблизительно 10% до приблизительно 99%, или уменьшению, составляющему по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99% или по меньшей мере 100% или более от количества или активности, как, например, без ограничения, уровней нитратов и уровней табак-специфичных нитрозаминов (TSNA). Термин «уменьшенный» или фраза «уменьшенное количество» предназначены для обозначения количества TSNA в модифицированном растении табака или табачном продукте, полученном из модифицированного растения табака, которое является меньшим, чем можно обнаружить в растении табака той же разновидности табака, переработанном таким же образом, которое не было модифицировано, или в табачном продукте из него. Таким образом, в некоторых случаях табак дикого типа той же разновидности, который был переработан таким же образом, используется в качестве контроля, с помощью которого путем измерений определяют, было ли достигнуто уменьшение содержания нитратов или TSNA посредством способов по настоящему изобретению, описанных в данном документе.
Выражения «регуляторные последовательности» и «регуляторные элементы», используемые в данном документе взаимозаменяемо, относятся к нуклеотидным последовательностям, расположенным выше (5'-некодирующие последовательности), в пределах или ниже (3'-некодирующие последовательности) кодирующей последовательности, которые влияют на транскрипцию, процессинг или стабильность РНК или на трансляцию связанной кодирующей последовательности. Регуляторные последовательности могут включать в себя без ограничения промоторы, лидерные последовательности, регулирующие трансляцию, интроны и распознаваемые последовательности полиаденилирования. Термины «регуляторная последовательность» и «регуляторный элемент» используются в данном документе взаимозаменяемо.
Выражение «сайт-специфичная нуклеаза», используемое в данном документе, относится к ферменту, способному к специфичному распознаванию и расщеплению последовательностей ДНК. Сайт-специфичная нуклеаза может быть сконструированной. Примеры сконструированных сайт-специфичных нуклеаз включают нуклеазы с «цинковыми пальцами» (ZFN), TAL-эффекторные нуклеазы (TALEN), системы на основе CRISPR/Cas9 и мегануклеазы.
Термин «табак», используемый в данном документе, в некоторых случаях используется в собирательном смысле для обозначения табачных культур (например, множества растений табака, выращиваемых в поле, т. е. табака, выращиваемого не методом гидропоники), растений табака и их частей, в том числе, без ограничения, корней, стеблей, листьев, цветов и семян, подготовленных и/или полученных, как описано в данном документе. Понятно, что выражение «табак» относится к растениям Nicotiana tabacum и продуктам из них. Термин «табачные продукты» в некоторых случаях относится к потребительским табачным продуктам, в том числе, без ограничения, к курительным материалам (например, сигаретам, сигарам и трубочному табаку), нюхательному табаку, жевательному табаку, жевательной резинке и леденцам, а также компонентам, материалам и ингредиентам для производства потребительских табачных продуктов. Предпочтительно данные табачные продукты производят из листьев и стеблей табака (Nicotiana tabacum), собранных с табака, обработанных, как описано выше, и нарезанных, томленых, высушенных и/или ферментированных в соответствии с традиционными методиками получения табака.
Выражения «терминатор транскрипции», «последовательности терминации» или «терминатор», используемые в данном документе, относятся к последовательностям ДНК, расположенным ниже кодирующей последовательности, включающим распознаваемые последовательности полиаденилирования и другие последовательности, кодирующие регуляторные сигналы, способные воздействовать на процессинг мРНК или экспрессию генов. Сигнал полиаденилирования обычно характеризуется осуществлением добавления трактов полиадениловой кислоты на 3'-конец предшественника мРНК.
Выражение «трансгенный», используемое в данном документе, относится к любой клетке, линии клеток, каллюсу, ткани, части растения или растению, геном которых был изменен в результате присутствия гетерологичной нуклеиновой кислоты, такой как рекомбинантная ДНК-конструкция, в том числе к исходным трансгенным объектам, а также полученным с помощью процедур полового скрещивания или бесполого размножения из исходного трансгенного объекта. Термин «трансгенный», используемый в данном документе, не охватывает изменение генома (хромосомное или внехромосомное) с помощью традиционных способов селекции растений или в результате встречающихся в природе событий, таких как случайное перекрестное опыление, инфекция, вызванная нерекомбинантным вирусом, трансформация нерекомбинантными бактериями, нерекомбинантная транспозиция или спонтанная мутация.
Выражение «трансгенное растение», используемое в данном документе, включает ссылку на растение, которое содержит в своем геноме гетерологичный полинуклеотид, т. е. растение или дерево, которые содержат рекомбинантный генетический материал, обычно не обнаруживаемый в растениях или деревьях данного типа, который был введен в рассматриваемое растение (или в предков растения) посредством манипуляции, осуществляемой человеком. Например, гетерологичный полинуклеотид стабильно интегрируют в геном таким образом, что полинуклеотид передается последующим поколениям. Гетерологичный полинуклеотид можно интегрировать в геном в отдельности или в виде части рекомбинантной ДНК-конструкции. Коммерческая разработка генетически улучшенной идиоплазмы также продвинулась к стадии введения нескольких признаков в культурные растения, часто называемой подходом пирамидирования генов. В этом подходе в растение можно ввести несколько генов, придающих разные характеристики, представляющие интерес. Пирамидирование генов можно осуществлять многими способами, в том числе, без ограничения, путем котрансформации, повторной трансформации и скрещивания линий с разными трансгенами. Таким образом, растение, выращиваемое из растительной клетки, в которую рекомбинантную ДНК вводят с помощью трансформации, является трансгенным растением, равно как и все потомство данного растения, которое содержит введенный трансген (полученное как половым, так и бесполым путем). Понятно, что термин «трансгенное растение» охватывает все растение или дерево и части растения или дерева, например, зерна, семена, цветы, листья, корни, плоды, пыльцу, стебли и т. д.
Выражение «трансгенное растение» включает также ссылки на растения, которые содержат более чем один гетерологичный полинуклеотид в своем геноме. Каждый гетерологичный полинуклеотид может придавать трансгенному растению отдельный признак.
Выражение «эффектор, подобный активаторам транскрипции» или «TALE», используемое в данном документе, относится к белковой структуре, которая распознает определенную последовательность ДНК и связывается с ней. Выражение «ДНК-связывающий домен TALE» относится к ДНК-связывающему домену, который содержит массив тандемных повторов из 33-35 аминокислот, также известных как RVD-модули, каждый из которых специфично распознает одну пару оснований ДНК. RVD-модули могут располагаться в любом порядке для сборки в массив, который распознает определенную последовательность.
Специфичность связывания ДНК-связывающего домена TALE определяется массивом RVD, за которым расположен один усеченный повтор из 20 аминокислот. ДНК-связывающий домен TALE может иметь от 12 до 27 RVD-модулей, каждый из которых содержит RVD и распознает одну пару оснований ДНК. Были идентифицированы специфичные RVD, которые распознают каждый из четырех возможных нуклеотидов ДНК (А, Т, С и G). Поскольку ДНК-связывающие домены TALE являются модульными, то повторы, которые распознают четыре разных нуклеотида ДНК, можно соединить друг с другом для распознавания любой конкретной последовательности ДНК. Эти нацеленные ДНК-связывающие домены можно впоследствии объединить с каталитическими доменами для создания функциональных ферментов, в том числе искусственных факторов транскрипции, метилтрансфераз, интеграз, нуклеаз и рекомбиназ.
Выражения «эффекторные нуклеазы, подобные активаторам транскрипции» или «TALEN», используемые в данном документе взаимозаменяемо, относятся к сконструированным белкам слияния на основе каталитического домена нуклеазы, такой как эндонуклеаза FokI, и разработанному ДНК-связывающему домену TALE, который может быть нацелен на специально синтезированную последовательность ДНК. Выражение "мономер TALEN" относится к сконструированному белку слияния с каталитическим доменом нуклеазы и разработанным ДНК-связывающим доменом TALE. Два мономера TALEN могут быть разработаны для нацеливания на целевой участок для TALEN и его расщепления.
Выражение «трансген», используемое в данном документе, относится к гену или генетическому материалу, содержащему последовательность гена, которая была выделена из одного организма и введена в другой организм. Этот ненативный сегмент ДНК может сохранять способность к обеспечению выработки РНК или белка в трансгенном организме или он может изменять нормальную функцию генетического кода трансгенного организма. Введение трансгена имеет потенциал к изменению фенотипа организма.
Выражение «вариант», используемое в данном документе по отношению к нуклеиновой кислоте, означает (i) часть или фрагмент упоминаемой нуклеотидной последовательности; (ii) последовательность, комплементарную упоминаемой нуклеотидной последовательности или ее части; (iii) нуклеиновую кислоту, практически идентичную упоминаемой последовательности нуклеиновой кислоты или комплементарной ей последовательности; или (iv) нуклеиновую кислоту, которая гибридизируется в жестких условиях с упоминаемой нуклеиновой кислотой, комплементарной ей последовательностью или практически идентичными ей последовательностями.
Выражение «вариант» по отношению к пептиду или полипептиду означает, что они отличаются по аминокислотной последовательности вставкой, делецией или консервативной заменой аминокислот, но сохраняют по меньшей мере один вид биологической активности. Вариант может также означать белок с аминокислотной последовательностью, которая практически идентична аминокислотной последовательности упоминаемого белка, который сохраняет по меньшей мере один вид биологической активности. Консервативную замену аминокислоты, т. е. замещение аминокислоты другой аминокислотой с аналогичными свойствами (например, степенью гидрофильности и распределением заряженных участков), понимают в данной области техники как обычно включающую незначительное изменение.
Термин «разновидность», используемый в данном документе, относится к популяции растений, которые обладают постоянными характеристиками, отделяющими их от других растений того же вида. Имея один или более отличительных признаков, разновидность дополнительно характеризуется очень небольшой общей изменчивостью индивидуумов в пределах такой разновидности. Разновидность зачастую является предметом коммерческих продаж.
Выражение «вектор», используемое в данном документе, относится к средству доставки на основе нуклеиновой кислоты, которое содержит комбинацию компонентов нуклеиновой кислоты для обеспечения транспорта нуклеиновой кислоты, конструкций нуклеиновых кислот и конъюгатов нуклеиновых кислот и т. п. Вектор может представлять собой вирусный вектор, бактериофаг, бактериальную искусственную хромосому или искусственную хромосому дрожжей. Вектор может представлять собой ДНК- или РНК-вектор. Подходящие векторы включают в себя эписомы, способные к внехромосомной репликации, такие как кольцевые плазмиды на основе двухнитевой нуклеиновой кислоты; линеаризованные плазмиды на основе двухнитевой нуклеиновой кислоты и другие векторы любого происхождения. Например, вектор может кодировать дерегулированный белок нитратредуктазу, содержащий аминокислотную последовательность под любым из SEQ ID NO: 5 или 7. Выражение «вектор экспрессии», используемое в данном документе, означает средство доставки на основе нуклеиновой кислоты, которое содержит комбинацию компонентов нуклеиновой кислоты для обеспечения экспрессии нуклеиновой кислоты, конструкций нуклеиновых кислот и конъюгатов нуклеиновых кислот и т. п. Подходящие векторы экспрессии включают в себя эписомы, способные к внехромосомной репликации, такие как кольцевые плазмиды на основе двухнитевой нуклеиновой кислоты; линеаризованные плазмиды на основе двухнитевой нуклеиновой кислоты и другие функционально эквивалентные векторы экспрессии любого происхождения. Вектор экспрессии содержит по меньшей мере промотор, расположенный выше по последовательности и функционально связанный с нуклеиновой кислотой, конструкциями нуклеиновых кислот или конъюгатом нуклеиновой кислоты, как определено ниже.
Выражение «цинковый палец», используемое в данном документе, относится к белковой структуре, которая распознает последовательности ДНК и связывается с ними. Домен «цинковый палец» является наиболее распространенным ДНК-связывающим мотивом в протеоме человека. Один «цинковый палец» содержит примерно 30 аминокислот, и домен обычно функционирует путем связывания с 3 последовательными парами оснований ДНК посредством взаимодействий одной боковой цепи аминокислоты с каждой парой оснований.
Выражения «нуклеаза с «цинковыми пальцами» или «ZFN», используемые в данном документе взаимозаменяемо, относятся к химерной молекуле белка, содержащей по меньшей мере один ДНК-связывающий домен «цинковый палец», эффективно связанный по меньшей мере с одной нуклеазой или частью нуклеазы, способной в полностью собранном виде расщеплять ДНК.
Если в данном документе не определено иное, то научные и технические термины, используемые в связи с настоящим раскрытием, будут иметь значения, которые обычно понятны средним специалистам в данной области. Например, любые системы номенклатуры и методики, используемые в связи с клеточной и тканевой культурами, молекулярной биологией, иммунологией, микробиологией, генетикой и химией белков и нуклеиновых кислот, а также гибридизацией, которые описаны в данном документе, хорошо известны и широко применяются в данной области техники. Значение и объем терминов должны быть ясны, однако в случае какой-либо скрытой двусмысленности определения, приведенные в данном документе, имеют преимущественную силу по сравнению с любым словарным или не относящимся к данному документу определением. Кроме того, если иное не требуется по контексту, термины в единственном числе будут включать в себя множественное число, и термины во множественном числе будут включать в себя единственное число.
Модифицированное растение табака с уменьшенным содержанием нитратов
В одном аспекте настоящее изобретение направлено на модифицированные растения табака, характеризующиеся уменьшенными уровнями свободных нитратов в их листьях. Уровни свободных нитратов уменьшаются посредством пути ассимиляции нитратов. Упрощенная схема пути ассимиляции азота в растениях показана на фиг. 1. После того, как растения подвергаются воздействию нитратов из почвы, имеют место несколько этапов, связанных с включением N, в том числе поглощение, ассимиляция, транслокация и ремобилизация. После поглощения специфическими переносчиками, расположенными на мембранах клеток корней, нитрат поступает в специфический путь ассимиляции. Вначале нитрат восстанавливается до нитрита (NO2-) ферментом нитратредуктазой (NR; EC 1.7.1.1-3). Затем нитрит восстанавливается до аммиака ферментом нитритредуктазой (NiR). Последний этап в растениях обычно бывает высокоэффективным, и в результате растительные клетки, как правило, содержат нитрит на очень низких уровнях. После восстановления нитрита до аммиака с помощью фермента глутаминсинтетазы (GS) происходит включение азота в органическую молекулу глутамина (Gln). Фермент GS функционирует совместно с ферментом глутаматсинтазой (GOGAT), образуя цикл GS/GOGAT, функцией которого является перенос аминогрупп от Gln к глутаминовой кислоте (Glu), что служит в качестве «входных ворот» для последующего перераспределения азота в другие аминокислоты и в конечном счете в белки и другие азотсодержащие макромолекулы. Еще один важный этап в данном процессе катализируется изоцитратдегидрогеназой (ICDH), ферментом, который, как полагают, отвечает за предоставление углеродного скелета 2-оксоглутарата, необходимого GOGAT для выработки Glu.
Табак (Nicotiana tabacum) имеет два высокогомологичных гена NR табака, обозначаемых как Nia1 и Nia2. Модифицированные растения табака, раскрытые в данном документе, которые характеризуются уменьшенными уровнями свободных нитратов в своих листьях, имеют модифицированный полинуклеотид, кодирующий нитратредуктазу, который кодирует дерегулированную полипептидную нитратредуктазу, т. е. дерегулированный продукт генов Nia1 и Nia2. Нитратредуктаза является высокорегулируемой как на транскрипционном, так и на посттранскрипционном уровнях. Благодаря механизмам посттрансляционной регуляции, которые включают в себя регуляцию циклом чередования света и темноты посредством фосфорилирования, сверхэкспрессия генов NR в трансгенных растениях, как правило, приводит лишь к незначительному увеличению активности NR в клетке. В некоторых вариантах осуществления дерегулированная полипептидная нитратредуктаза представляет собой эндогенную или гетерологичную полипептидную нитратредуктазу, которая является конститутивно активной. Дерегулированная полипептидная нитратредуктаза может характеризоваться потерей способности к регуляции посредством фосфорилирования вследствие утраты сайта фосфорилирования в полипептидной нитратредуктазе и поэтому не регулируется циклом чередования света и темноты. Модифицированное растение табака может иметь полинуклеотид, кодирующий дерегулированную полипептидную нитратредуктазу или дерегулированную полипептидную нитратредуктазу, SEQ ID NO: 5 или SEQ ID NO: 7 или их варианты или фрагменты. Например, дерегулированная нитратредуктаза может представлять собой усеченную полипептидную нитратредуктазу или полипептидную нитратредуктазу, имеющую аминокислотную замену, где сайт фосфорилирования удален.
В некоторых вариантах осуществления модифицированное растение табака демонстрирует практически нормальный рост и развитие растения по сравнению с немодифицированным контрольным растением. Например, модифицированное растение табака может стареть обычным образом, т. е. старение у модифицированного растения табака может быть практически таким же, как и у немодифицированного контрольного растения табака. В некоторых вариантах осуществления внешний вид модифицированного растения табака может быть практически таким же, как и у немодифицированного контрольного растения, например, через 3 или 6 месяцев после пересадки в поле. Например, содержание хлорофилла в листьях, окраска листьев, степень зрелости, количество листьев на растение, высота стебля, угол врастания листа, размер листа (ширина или длина), межузловое расстояние и/или весовое соотношение листовой пластинки и средней жилки может в модифицированных растениях табака быть практически таким же по сравнению с немодифицированными контрольными растениями.
Усеченная нитратредуктаза
Дерегулированная полипептидная нитратредуктаза может представлять собой полипептидную нитратредуктазу с N-концевым усечением, т. е. делецией на N-конце или вблизи него. Усеченный ген Nia2 табака, который кодирует белок NR, имеющий делецию 56 аминокислот в своем N-концевом домене, характеризуется значительной степенью устранения посттрансляционного контроля фермента. Делеция 56 аминокислот дает возможность ферменту нитратредуктазе быть одинаково активным как на свету, так и в темноте по сравнению с ферментом дикого типа (WT), который вследствие фосфорилирования становится инактивированным во время циклов темноты. В сочетании с действием сильного конститутивного промотора экспрессия усеченного гена нитратредуктазы может обеспечивать повышение общей активности нитратредуктазы NR в растении, но также и дерегуляцию выработки глутамина (Gln) и аспарагина (Asn).
Дерегулированная полипептидная нитратредуктаза может представлять собой полипептидную нитратредуктазу, имеющую N-концевое усечение длиной от приблизительно 10 аминокислот до приблизительно 100 аминокислот, от приблизительно 10 аминокислот до приблизительно 90 аминокислот, от приблизительно 10 аминокислот до приблизительно 80 аминокислот, от приблизительно 10 аминокислот до приблизительно 70 аминокислот, от приблизительно 10 аминокислот до приблизительно 60 аминокислот, от приблизительно 10 аминокислот до приблизительно 55 аминокислот, от приблизительно 10 аминокислот до приблизительно 50 аминокислот, от приблизительно 10 аминокислот до приблизительно 40 аминокислот, от приблизительно 10 аминокислот до приблизительно 30 аминокислот, от приблизительно 10 аминокислот до приблизительно 20 аминокислот, от приблизительно 20 аминокислот до приблизительно 100 аминокислот, от приблизительно 20 аминокислот до приблизительно 90 аминокислот, от приблизительно 20 аминокислот до приблизительно 80 аминокислот, от приблизительно 20 аминокислот до приблизительно 70 аминокислот, от приблизительно 20 аминокислот до приблизительно 60 аминокислот, от приблизительно 20 аминокислот до приблизительно 55 аминокислот, от приблизительно 20 аминокислот до приблизительно 50 аминокислот, от приблизительно 20 аминокислот до приблизительно 40 аминокислот, от приблизительно 20 аминокислот до приблизительно 30 аминокислот, от приблизительно 30 аминокислот до приблизительно 100 аминокислот, от приблизительно 30 аминокислот до приблизительно 90 аминокислот, от приблизительно 30 аминокислот до приблизительно 80 аминокислот, от приблизительно 30 аминокислот до приблизительно 70 аминокислот, от приблизительно 30 аминокислот до приблизительно 60 аминокислот, от приблизительно 30 аминокислот до приблизительно 55 аминокислот, от приблизительно 30 аминокислот до приблизительно 50 аминокислот, от приблизительно 30 аминокислот до приблизительно 40 аминокислот, от приблизительно 40 аминокислот до приблизительно 100 аминокислот, от приблизительно 40 аминокислот до приблизительно 90 аминокислот, от приблизительно 40 аминокислот до приблизительно 80 аминокислот, от приблизительно 40 аминокислот до приблизительно 70 аминокислот, от приблизительно 40 аминокислот до приблизительно 60 аминокислот, от приблизительно 40 аминокислот до приблизительно 55 аминокислот, от приблизительно 40 аминокислот до приблизительно 50 аминокислот, от приблизительно 50 аминокислот до приблизительно 100 аминокислот, от приблизительно 50 аминокислот до приблизительно 90 аминокислот, от приблизительно 50 аминокислот до приблизительно 80 аминокислот, от приблизительно 50 аминокислот до приблизительно 70 аминокислот, от приблизительно 50 аминокислот до приблизительно 60 аминокислот, от приблизительно 50 аминокислот до приблизительно 55 аминокислот, от приблизительно 55 аминокислот до приблизительно 100 аминокислот, от приблизительно 55 аминокислот до приблизительно 90 аминокислот, от приблизительно 55 аминокислот до приблизительно 80 аминокислот, от приблизительно 55 аминокислот до приблизительно 70 аминокислот или от приблизительно 55 аминокислот до приблизительно 60 аминокислот или по меньшей мере приблизительно 10 аминокислот, по меньшей мере приблизительно 20 аминокислот, по меньшей мере приблизительно 30 аминокислот, по меньшей мере приблизительно 40 аминокислот, по меньшей мере приблизительно 50 аминокислот, по меньшей мере 51 аминокислота, по меньшей мере приблизительно 52 аминокислоты, по меньшей мере 53 аминокислоты, по меньшей мере приблизительно 54 аминокислоты, по меньшей мере 55 аминокислот, по меньшей мере приблизительно 56 аминокислот, по меньшей мере 57 аминокислот, по меньшей мере приблизительно 58 аминокислот, по меньшей мере 59 аминокислот, по меньшей мере приблизительно 60 аминокислот или по меньшей мере 65 аминокислот по сравнению с SEQ ID NO: 4. Дерегулированная полипептидная нитратредуктаза может представлять собой полипептидную нитратредуктазу, имеющую N-концевое усечение 56 аминокислот.
Аминокислотная замена
Дерегулированная полипептидная нитратредуктаза может представлять собой полипептидную нитратредуктазу, имеющую аминокислотную замену в положении, соответствующем положению 523 в SEQ ID NO: 4. Мутация серинового остатка в предполагаемом сайте фосфорилирования (см. фиг. 2), которая таким образом устраняет посттрансляционный регулирующий контроль со стороны цикла чередования света и темноты, способствует увеличению уровней активной формы нитратредуктазы и поддержанию их высокими в трансгенных растениях для облегчения ассимиляции нитратов. При изменении серинового остатка (Ser) в положении 523 на аспартат (Asp) в SEQ ID NO: 4 с помощью сайт-направленного мутагенеза полипептидная нитратредуктаза больше не служит в качестве субстрата для протеинкиназ, которые обычно инактивируют данный фермент в период темноты посредством фосфорилирования данного остатка Ser. В некоторых вариантах осуществления дерегулированная нитратредуктаза включает в себя полипептид, содержащий аминокислотную замену в положении, соответствующем положению 523 в SEQ ID NO: 4. В некоторых вариантах осуществления дерегулированная нитратредуктаза включает в себя полипептид, содержащий аминокислоту, отличную от серина, в положении, соответствующем положению 523 в SEQ ID NO: 4. В некоторых вариантах осуществления дерегулированная нитратредуктаза включает в себя полипептид, содержащий аспартат в положении, соответствующем положению 523 в SEQ ID NO: 4.
a. Уменьшенные уровни нитратов
Модифицированные растения табака могут характеризоваться уменьшенными уровнями нитратов по сравнению с контрольным растением табака, таким как контрольное растение табака дикого типа. В некоторых вариантах осуществления модифицированные растения табака могут характеризоваться уменьшенными уровнями нитратов в листьях в случае выращивания при высоких уровнях, таких как 19 мМ нитратов, или средних уровнях, таких как 8 мМ нитратов, обработки нитратными удобрениями по сравнению с контрольным растением табака. В некоторых вариантах осуществления модифицированные растения табака могут характеризоваться уменьшенными уровнями нитратов в листьях в случае выращивания в полевых условиях по сравнению с контрольным растением табака. В некоторых вариантах осуществления листья являются свежесобранными. В некоторых вариантах осуществления листья являются высушенными, хранящимися или переработанными. В некоторых вариантах осуществления листья являются высушенными на воздухе.
Модифицированные растения табака могут характеризоваться по меньшей мере приблизительно 1-кратным уменьшением, по меньшей мере приблизительно 2-кратным уменьшением, по меньшей мере приблизительно 3-кратным уменьшением, по меньшей мере приблизительно 4-кратным уменьшением, по меньшей мере приблизительно 5-кратным уменьшением, по меньшей мере приблизительно 6-кратным уменьшением, по меньшей мере приблизительно 7-кратным уменьшением, по меньшей мере приблизительно 8-кратным уменьшением, по меньшей мере приблизительно 9-кратным уменьшением, по меньшей мере приблизительно 10-кратным уменьшением, по меньшей мере приблизительно 15-кратным уменьшением, по меньшей мере приблизительно 20-кратным уменьшением, по меньшей мере приблизительно 25-кратным уменьшением или по меньшей мере приблизительно 30-кратным уменьшением уровней нитратов в свежесобранных листьях модифицированного растения табака, выращиваемого при высоких уровнях обработки нитратными удобрениями, по сравнению со свежесобранными листьями контрольного табака. Свежесобранные листья модифицированных растений табака могут характеризоваться уровнями нитратов, обнаруживаемыми в свежесобранных листьях, которые составляют от приблизительно 0,5% до приблизительно 99%, от приблизительно 0,5% до приблизительно 95%, от приблизительно 0,5% до приблизительно 90%, от приблизительно 0,5% до приблизительно 85%, от приблизительно 0,5% до приблизительно 80%, от приблизительно 0,5% до приблизительно 75%, от приблизительно 0,5% до приблизительно 70%, от приблизительно 0,5% до приблизительно 65%, от приблизительно 0,5% до приблизительно 60%, от приблизительно 0,5% до приблизительно 55%, от приблизительно 0,5% до приблизительно 50%, от приблизительно 0,5% до приблизительно 45%, от приблизительно 0,5% до приблизительно 40%, от приблизительно 0,5% до приблизительно 35%, от приблизительно 0,5% до приблизительно 30%, от приблизительно 0,5% до приблизительно 25%, от приблизительно 0,5% до приблизительно 20%, от приблизительно 0,5% до приблизительно 15%, от приблизительно 0,5% до приблизительно 13%, от приблизительно 0,5% до приблизительно 10%, от приблизительно 0,5% до приблизительно 5%, от приблизительно 1% до приблизительно 99%, от приблизительно 1% до приблизительно 95%, от приблизительно 1% до приблизительно 90%, от приблизительно 1% до приблизительно 85%, от приблизительно 1% до приблизительно 80%, от приблизительно 1% до приблизительно 75%, от приблизительно 1% до приблизительно 70%, от приблизительно 1% до приблизительно 65%, от приблизительно 1% до приблизительно 60%, от приблизительно 1% до приблизительно 55%, от приблизительно 1% до приблизительно 50%, от приблизительно 1% до приблизительно 45%, от приблизительно 1% до приблизительно 40%, от приблизительно 1% до приблизительно 35%, от приблизительно 1% до приблизительно 30%, от приблизительно 1% до приблизительно 25%, от приблизительно 1% до приблизительно 20%, от приблизительно 1% до приблизительно 15%, от приблизительно 1% до приблизительно 13%, от приблизительно 1% до приблизительно 10%, от приблизительно 1% до приблизительно 5%, от приблизительно 5% до приблизительно 99%, от приблизительно 5% до приблизительно 95%, от приблизительно 5% до приблизительно 90%, от приблизительно 5% до приблизительно 85%, от приблизительно 5% до приблизительно 80%, от приблизительно 5% до приблизительно 75%, от приблизительно 5% до приблизительно 70%, от приблизительно 5% до приблизительно 65%, от приблизительно 5% до приблизительно 60%, от приблизительно 5% до приблизительно 55%, от приблизительно 5% до приблизительно 50%, от приблизительно 5% до приблизительно 45%, от приблизительно 5% до приблизительно 40%, от приблизительно 5% до приблизительно 35%, от приблизительно 5% до приблизительно 30%, от приблизительно 5% до приблизительно 25%, от приблизительно 5% до приблизительно 20%, от приблизительно 5% до приблизительно 15%, от приблизительно 5% до приблизительно 13%, от приблизительно 5% до приблизительно 10%, от приблизительно 10% до приблизительно 99%, от приблизительно 10% до приблизительно 95%, от приблизительно 10% до приблизительно 90%, от приблизительно 10% до приблизительно 85%, от приблизительно 10% до приблизительно 80%, от приблизительно 10% до приблизительно 75%, от приблизительно 10% до приблизительно 70%, от приблизительно 10% до приблизительно 65%, от приблизительно 10% до приблизительно 60%, от приблизительно 10% до приблизительно 55%, от приблизительно 10% до приблизительно 50%, от приблизительно 10% до приблизительно 45%, от приблизительно 10% до приблизительно 40%, от приблизительно 10% до приблизительно 35%, от приблизительно 10% до приблизительно 30%, от приблизительно 10% до приблизительно 25%, от приблизительно 10% до приблизительно 20%, от приблизительно 10% до приблизительно 15%, от приблизительно 10% до приблизительно 13%, от приблизительно 20% до приблизительно 99%, от приблизительно 20% до приблизительно 95%, от приблизительно 20% до приблизительно 90%, от приблизительно 20% до приблизительно 85%, от приблизительно 20% до приблизительно 80%, от приблизительно 20% до приблизительно 75%, от приблизительно 20% до приблизительно 70%, от приблизительно 20% до приблизительно 65%, от приблизительно 20% до приблизительно 60%, от приблизительно 20% до приблизительно 55%, от приблизительно 20% до приблизительно 50%, от приблизительно 20% до приблизительно 45%, от приблизительно 20% до приблизительно 40%, от приблизительно 20% до приблизительно 35%, от приблизительно 20% до приблизительно 30%, от приблизительно 20% до приблизительно 25%, от приблизительно 30% до приблизительно 99%, от приблизительно 30% до приблизительно 95%, от приблизительно 30% до приблизительно 90%, от приблизительно 30% до приблизительно 85%, от приблизительно 30% до приблизительно 80%, от приблизительно 30% до приблизительно 75%, от приблизительно 30% до приблизительно 70%, от приблизительно 30% до приблизительно 65%, от приблизительно 30% до приблизительно 60%, от приблизительно 30% до приблизительно 55%, от приблизительно 30% до приблизительно 50%, от приблизительно 30% до приблизительно 45%, от приблизительно 30% до приблизительно 40%, от приблизительно 30% до приблизительно 35%, от приблизительно 40% до приблизительно 99%, от приблизительно 40% до приблизительно 95%, от приблизительно 40% до приблизительно 90%, от приблизительно 40% до приблизительно 85%, от приблизительно 40% до приблизительно 80%, от приблизительно 40% до приблизительно 75%, от приблизительно 40% до приблизительно 70%, от приблизительно 40% до приблизительно 65%, от приблизительно 40% до приблизительно 60%, от приблизительно 40% до приблизительно 55%, от приблизительно 40% до приблизительно 50%, от приблизительно 40% до приблизительно 45%, от приблизительно 50% до приблизительно 99%, от приблизительно 50% до приблизительно 95%, от приблизительно 50% до приблизительно 90%, от приблизительно 50% до приблизительно 85%, от приблизительно 50% до приблизительно 80%, от приблизительно 50% до приблизительно 75%, от приблизительно 50% до приблизительно 70%, от приблизительно 50% до приблизительно 65%, от приблизительно 50% до приблизительно 60%, от приблизительно 50% до приблизительно 55%, от приблизительно 60% до приблизительно 99%, от приблизительно 60% до приблизительно 95%, от приблизительно 60% до приблизительно 90%, от приблизительно 60% до приблизительно 85%, от приблизительно 60% до приблизительно 80%, от приблизительно 60% до приблизительно 75%, от приблизительно 60% до приблизительно 70%, от приблизительно 60% до приблизительно 65% или по меньшей мере приблизительно 0,5%, по меньшей мере приблизительно 1%, по меньшей мере приблизительно 2%, по меньшей мере приблизительно 3%, по меньшей мере приблизительно 4%, по меньшей мере приблизительно 5%, по меньшей мере приблизительно 6%, по меньшей мере приблизительно 7%, по меньшей мере приблизительно 8%, по меньшей мере приблизительно 9%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 11%, по меньшей мере приблизительно 12%, по меньшей мере приблизительно 13%, по меньшей мере приблизительно 14%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 16%, по меньшей мере приблизительно 17%, по меньшей мере приблизительно 18%, по меньшей мере приблизительно 19%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 99% по сравнению с контрольными растениями табака, такими как растение табака дикого типа, в случае выращивания при высоких уровнях обработки нитратными удобрениями. Модифицированные растения табака могут иметь по меньшей мере приблизительно 50 ppm, по меньшей мере приблизительно 100 ppm, по меньшей мере приблизительно 500 ppm, по меньшей мере приблизительно 1000 ppm, по меньшей мере приблизительно 1500 ppm, по меньшей мере приблизительно 2000 ppm, по меньшей мере приблизительно 2500 ppm, по меньшей мере приблизительно 3000 ppm, по меньшей мере приблизительно 3500 ppm, по меньшей мере приблизительно 4000 ppm или по меньшей мере приблизительно 5000 ppm нитратов в свежесобранных листьях модифицированного растения табака, выращиваемого при средних уровнях обработки нитратными удобрениями, по сравнению со свежесобранными листьями контрольного табака.
Модифицированные растения табака могут характеризоваться по меньшей мере приблизительно 1-кратным уменьшением, по меньшей мере приблизительно 2-кратным уменьшением, по меньшей мере приблизительно 3-кратным уменьшением, по меньшей мере приблизительно 4-кратным уменьшением, по меньшей мере приблизительно 5-кратным уменьшением, по меньшей мере приблизительно 6-кратным уменьшением, по меньшей мере приблизительно 7-кратным уменьшением, по меньшей мере приблизительно 8-кратным уменьшением, по меньшей мере приблизительно 9-кратным уменьшением, по меньшей мере приблизительно 10-кратным уменьшением, по меньшей мере приблизительно 15-кратным уменьшением, по меньшей мере приблизительно 20-кратным уменьшением, по меньшей мере приблизительно 25-кратным уменьшением или по меньшей мере приблизительно 30-кратным уменьшением уровней нитратов в свежесобранных листьях модифицированного растения табака, выращиваемого при средних уровнях обработки нитратными удобрениями, по сравнению со свежесобранными листьями контрольного табака. Свежесобранные листья модифицированных растений табака могут характеризоваться уровнями нитратов, обнаруживаемыми в свежесобранных листьях, которые составляют от приблизительно 0,5% до приблизительно 99%, от приблизительно 0,5% до приблизительно 95%, от приблизительно 0,5% до приблизительно 90%, от приблизительно 0,5% до приблизительно 85%, от приблизительно 0,5% до приблизительно 80%, от приблизительно 0,5% до приблизительно 75%, от приблизительно 0,5% до приблизительно 70%, от приблизительно 0,5% до приблизительно 65%, от приблизительно 0,5% до приблизительно 60%, от приблизительно 0,5% до приблизительно 55%, от приблизительно 0,5% до приблизительно 50%, от приблизительно 0,5% до приблизительно 45%, от приблизительно 0,5% до приблизительно 40%, от приблизительно 0,5% до приблизительно 35%, от приблизительно 0,5% до приблизительно 30%, от приблизительно 0,5% до приблизительно 25%, от приблизительно 0,5% до приблизительно 20%, от приблизительно 0,5% до приблизительно 15%, от приблизительно 0,5% до приблизительно 13%, от приблизительно 0,5% до приблизительно 10%, от приблизительно 0,5% до приблизительно 5%, от приблизительно 1% до приблизительно 99%, от приблизительно 1% до приблизительно 95%, от приблизительно 1% до приблизительно 90%, от приблизительно 1% до приблизительно 85%, от приблизительно 1% до приблизительно 80%, от приблизительно 1% до приблизительно 75%, от приблизительно 1% до приблизительно 70%, от приблизительно 1% до приблизительно 65%, от приблизительно 1% до приблизительно 60%, от приблизительно 1% до приблизительно 55%, от приблизительно 1% до приблизительно 50%, от приблизительно 1% до приблизительно 45%, от приблизительно 1% до приблизительно 40%, от приблизительно 1% до приблизительно 35%, от приблизительно 1% до приблизительно 30%, от приблизительно 1% до приблизительно 25%, от приблизительно 1% до приблизительно 20%, от приблизительно 1% до приблизительно 15%, от приблизительно 1% до приблизительно 13%, от приблизительно 1% до приблизительно 10%, от приблизительно 1% до приблизительно 5%, от приблизительно 5% до приблизительно 99%, от приблизительно 5% до приблизительно 95%, от приблизительно 5% до приблизительно 90%, от приблизительно 5% до приблизительно 85%, от приблизительно 5% до приблизительно 80%, от приблизительно 5% до приблизительно 75%, от приблизительно 5% до приблизительно 70%, от приблизительно 5% до приблизительно 65%, от приблизительно 5% до приблизительно 60%, от приблизительно 5% до приблизительно 55%, от приблизительно 5% до приблизительно 50%, от приблизительно 5% до приблизительно 45%, от приблизительно 5% до приблизительно 40%, от приблизительно 5% до приблизительно 35%, от приблизительно 5% до приблизительно 30%, от приблизительно 5% до приблизительно 25%, от приблизительно 5% до приблизительно 20%, от приблизительно 5% до приблизительно 15%, от приблизительно 5% до приблизительно 13%, от приблизительно 5% до приблизительно 10%, от приблизительно 10% до приблизительно 99%, от приблизительно 10% до приблизительно 95%, от приблизительно 10% до приблизительно 90%, от приблизительно 10% до приблизительно 85%, от приблизительно 10% до приблизительно 80%, от приблизительно 10% до приблизительно 75%, от приблизительно 10% до приблизительно 70%, от приблизительно 10% до приблизительно 65%, от приблизительно 10% до приблизительно 60%, от приблизительно 10% до приблизительно 55%, от приблизительно 10% до приблизительно 50%, от приблизительно 10% до приблизительно 45%, от приблизительно 10% до приблизительно 40%, от приблизительно 10% до приблизительно 35%, от приблизительно 10% до приблизительно 30%, от приблизительно 10% до приблизительно 25%, от приблизительно 10% до приблизительно 20%, от приблизительно 10% до приблизительно 15%, от приблизительно 10% до приблизительно 13%, от приблизительно 20% до приблизительно 99%, от приблизительно 20% до приблизительно 95%, от приблизительно 20% до приблизительно 90%, от приблизительно 20% до приблизительно 85%, от приблизительно 20% до приблизительно 80%, от приблизительно 20% до приблизительно 75%, от приблизительно 20% до приблизительно 70%, от приблизительно 20% до приблизительно 65%, от приблизительно 20% до приблизительно 60%, от приблизительно 20% до приблизительно 55%, от приблизительно 20% до приблизительно 50%, от приблизительно 20% до приблизительно 45%, от приблизительно 20% до приблизительно 40%, от приблизительно 20% до приблизительно 35%, от приблизительно 20% до приблизительно 30%, от приблизительно 20% до приблизительно 25%, от приблизительно 30% до приблизительно 99%, от приблизительно 30% до приблизительно 95%, от приблизительно 30% до приблизительно 90%, от приблизительно 30% до приблизительно 85%, от приблизительно 30% до приблизительно 80%, от приблизительно 30% до приблизительно 75%, от приблизительно 30% до приблизительно 70%, от приблизительно 30% до приблизительно 65%, от приблизительно 30% до приблизительно 60%, от приблизительно 30% до приблизительно 55%, от приблизительно 30% до приблизительно 50%, от приблизительно 30% до приблизительно 45%, от приблизительно 30% до приблизительно 40%, от приблизительно 30% до приблизительно 35%, от приблизительно 40% до приблизительно 99%, от приблизительно 40% до приблизительно 95%, от приблизительно 40% до приблизительно 90%, от приблизительно 40% до приблизительно 85%, от приблизительно 40% до приблизительно 80%, от приблизительно 40% до приблизительно 75%, от приблизительно 40% до приблизительно 70%, от приблизительно 40% до приблизительно 65%, от приблизительно 40% до приблизительно 60%, от приблизительно 40% до приблизительно 55%, от приблизительно 40% до приблизительно 50%, от приблизительно 40% до приблизительно 45%, от приблизительно 50% до приблизительно 99%, от приблизительно 50% до приблизительно 95%, от приблизительно 50% до приблизительно 90%, от приблизительно 50% до приблизительно 85%, от приблизительно 50% до приблизительно 80%, от приблизительно 50% до приблизительно 75%, от приблизительно 50% до приблизительно 70%, от приблизительно 50% до приблизительно 65%, от приблизительно 50% до приблизительно 60%, от приблизительно 50% до приблизительно 55%, от приблизительно 60% до приблизительно 99%, от приблизительно 60% до приблизительно 95%, от приблизительно 60% до приблизительно 90%, от приблизительно 60% до приблизительно 85%, от приблизительно 60% до приблизительно 80%, от приблизительно 60% до приблизительно 75%, от приблизительно 60% до приблизительно 70%, от приблизительно 60% до приблизительно 65% или по меньшей мере приблизительно 0,5%, по меньшей мере приблизительно 1%, по меньшей мере приблизительно 2%, по меньшей мере приблизительно 3%, по меньшей мере приблизительно 4%, по меньшей мере приблизительно 5%, по меньшей мере приблизительно 6%, по меньшей мере приблизительно 7%, по меньшей мере приблизительно 8%, по меньшей мере приблизительно 9%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 11%, по меньшей мере приблизительно 12%, по меньшей мере приблизительно 13%, по меньшей мере приблизительно 14%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 16%, по меньшей мере приблизительно 17%, по меньшей мере приблизительно 18%, по меньшей мере приблизительно 19%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 37%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 99% по сравнению с контрольными растениями табака, такими как растение табака дикого типа, в случае выращивания при средних уровнях обработки нитратными удобрениями. Модифицированные растения табака могут иметь по меньшей мере приблизительно 50 ppm, по меньшей мере приблизительно 100 ppm, по меньшей мере приблизительно 150 ppm, по меньшей мере приблизительно 200 ppm, по меньшей мере приблизительно 250 ppm, по меньшей мере приблизительно 300 ppm, по меньшей мере приблизительно 350 ppm, по меньшей мере приблизительно 400 ppm, по меньшей мере приблизительно 450 ppm, по меньшей мере приблизительно 500 ppm, по меньшей мере приблизительно 550 ppm, по меньшей мере приблизительно 600 ppm, по меньшей мере приблизительно 650 ppm, по меньшей мере приблизительно 700 ppm, по меньшей мере приблизительно 750 ppm, по меньшей мере приблизительно 800 ppm, по меньшей мере приблизительно 850 ppm, по меньшей мере приблизительно 900 ppm, по меньшей мере приблизительно 950 ppm или по меньшей мере приблизительно 1000 ppm нитратов в свежесобранных листьях модифицированного растения табака, выращиваемого при средних уровнях обработки нитратными удобрениями, по сравнению со свежесобранными листьями контрольного табака.
Свежесобранные листья или высушенные листья модифицированных растений табака могут характеризоваться по меньшей мере приблизительно 1-кратным уменьшением, по меньшей мере приблизительно 2-кратным уменьшением, по меньшей мере приблизительно 3-кратным уменьшением, по меньшей мере приблизительно 4-кратным уменьшением, по меньшей мере приблизительно 5-кратным уменьшением, по меньшей мере приблизительно 6-кратным уменьшением, по меньшей мере приблизительно 7-кратным уменьшением, по меньшей мере приблизительно 8-кратным уменьшением, по меньшей мере приблизительно 9-кратным уменьшением, по меньшей мере приблизительно 10-кратным уменьшением, по меньшей мере приблизительно 15-кратным уменьшением, по меньшей мере приблизительно 20-кратным уменьшением, по меньшей мере приблизительно 25-кратным уменьшением или по меньшей мере приблизительно 30-кратным уменьшением уровней нитратов в свежесобранных листьях или высушенных листьях модифицированного растения табака, выращиваемого в поле. Свежесобранные листья или высушенные листья модифицированных растений табака могут характеризоваться уровнями нитратов, обнаруживаемыми в свежесобранных листьях или высушенных листьях, которые составляют от приблизительно 0,5% до приблизительно 99%, от приблизительно 0,5% до приблизительно 95%, от приблизительно 0,5% до приблизительно 90%, от приблизительно 0,5% до приблизительно 85%, от приблизительно 0,5% до приблизительно 80%, от приблизительно 0,5% до приблизительно 75%, от приблизительно 0,5% до приблизительно 70%, от приблизительно 0,5% до приблизительно 65%, от приблизительно 0,5% до приблизительно 60%, от приблизительно 0,5% до приблизительно 55%, от приблизительно 0,5% до приблизительно 50%, от приблизительно 0,5% до приблизительно 45%, от приблизительно 0,5% до приблизительно 40%, от приблизительно 0,5% до приблизительно 35%, от приблизительно 0,5% до приблизительно 30%, от приблизительно 0,5% до приблизительно 25%, от приблизительно 0,5% до приблизительно 20%, от приблизительно 0,5% до приблизительно 15%, от приблизительно 0,5% до приблизительно 13%, от приблизительно 0,5% до приблизительно 10%, от приблизительно 0,5% до приблизительно 5%, от приблизительно 1% до приблизительно 99%, от приблизительно 1% до приблизительно 95%, от приблизительно 1% до приблизительно 90%, от приблизительно 1% до приблизительно 85%, от приблизительно 1% до приблизительно 80%, от приблизительно 1% до приблизительно 75%, от приблизительно 1% до приблизительно 70%, от приблизительно 1% до приблизительно 65%, от приблизительно 1% до приблизительно 60%, от приблизительно 1% до приблизительно 55%, от приблизительно 1% до приблизительно 50%, от приблизительно 1% до приблизительно 45%, от приблизительно 1% до приблизительно 40%, от приблизительно 1% до приблизительно 35%, от приблизительно 1% до приблизительно 30%, от приблизительно 1% до приблизительно 25%, от приблизительно 1% до приблизительно 20%, от приблизительно 1% до приблизительно 15%, от приблизительно 1% до приблизительно 13%, от приблизительно 1% до приблизительно 10%, от приблизительно 1% до приблизительно 5%, от приблизительно 5% до приблизительно 99%, от приблизительно 5% до приблизительно 95%, от приблизительно 5% до приблизительно 90%, от приблизительно 5% до приблизительно 85%, от приблизительно 5% до приблизительно 80%, от приблизительно 5% до приблизительно 75%, от приблизительно 5% до приблизительно 70%, от приблизительно 5% до приблизительно 65%, от приблизительно 5% до приблизительно 60%, от приблизительно 5% до приблизительно 55%, от приблизительно 5% до приблизительно 50%, от приблизительно 5% до приблизительно 45%, от приблизительно 5% до приблизительно 40%, от приблизительно 5% до приблизительно 35%, от приблизительно 5% до приблизительно 30%, от приблизительно 5% до приблизительно 25%, от приблизительно 5% до приблизительно 20%, от приблизительно 5% до приблизительно 15%, от приблизительно 5% до приблизительно 13%, от приблизительно 5% до приблизительно 10%, от приблизительно 10% до приблизительно 99%, от приблизительно 10% до приблизительно 95%, от приблизительно 10% до приблизительно 90%, от приблизительно 10% до приблизительно 85%, от приблизительно 10% до приблизительно 80%, от приблизительно 10% до приблизительно 75%, от приблизительно 10% до приблизительно 70%, от приблизительно 10% до приблизительно 65%, от приблизительно 10% до приблизительно 60%, от приблизительно 10% до приблизительно 55%, от приблизительно 10% до приблизительно 50%, от приблизительно 10% до приблизительно 45%, от приблизительно 10% до приблизительно 40%, от приблизительно 10% до приблизительно 35%, от приблизительно 10% до приблизительно 30%, от приблизительно 10% до приблизительно 25%, от приблизительно 10% до приблизительно 20%, от приблизительно 10% до приблизительно 15%, от приблизительно 10% до приблизительно 13%, от приблизительно 20% до приблизительно 99%, от приблизительно 20% до приблизительно 95%, от приблизительно 20% до приблизительно 90%, от приблизительно 20% до приблизительно 85%, от приблизительно 20% до приблизительно 80%, от приблизительно 20% до приблизительно 75%, от приблизительно 20% до приблизительно 70%, от приблизительно 20% до приблизительно 65%, от приблизительно 20% до приблизительно 60%, от приблизительно 20% до приблизительно 55%, от приблизительно 20% до приблизительно 50%, от приблизительно 20% до приблизительно 45%, от приблизительно 20% до приблизительно 40%, от приблизительно 20% до приблизительно 35%, от приблизительно 20% до приблизительно 30%, от приблизительно 20% до приблизительно 25%, от приблизительно 30% до приблизительно 99%, от приблизительно 30% до приблизительно 95%, от приблизительно 30% до приблизительно 90%, от приблизительно 30% до приблизительно 85%, от приблизительно 30% до приблизительно 80%, от приблизительно 30% до приблизительно 75%, от приблизительно 30% до приблизительно 70%, от приблизительно 30% до приблизительно 65%, от приблизительно 30% до приблизительно 60%, от приблизительно 30% до приблизительно 55%, от приблизительно 30% до приблизительно 50%, от приблизительно 30% до приблизительно 45%, от приблизительно 30% до приблизительно 40%, от приблизительно 30% до приблизительно 35%, от приблизительно 40% до приблизительно 99%, от приблизительно 40% до приблизительно 95%, от приблизительно 40% до приблизительно 90%, от приблизительно 40% до приблизительно 85%, от приблизительно 40% до приблизительно 80%, от приблизительно 40% до приблизительно 75%, от приблизительно 40% до приблизительно 70%, от приблизительно 40% до приблизительно 65%, от приблизительно 40% до приблизительно 60%, от приблизительно 40% до приблизительно 55%, от приблизительно 40% до приблизительно 50%, от приблизительно 40% до приблизительно 45%, от приблизительно 50% до приблизительно 99%, от приблизительно 50% до приблизительно 95%, от приблизительно 50% до приблизительно 90%, от приблизительно 50% до приблизительно 85%, от приблизительно 50% до приблизительно 80%, от приблизительно 50% до приблизительно 75%, от приблизительно 50% до приблизительно 70%, от приблизительно 50% до приблизительно 65%, от приблизительно 50% до приблизительно 60%, от приблизительно 50% до приблизительно 55%, от приблизительно 60% до приблизительно 99%, от приблизительно 60% до приблизительно 95%, от приблизительно 60% до приблизительно 90%, от приблизительно 60% до приблизительно 85%, от приблизительно 60% до приблизительно 80%, от приблизительно 60% до приблизительно 75%, от приблизительно 60% до приблизительно 70%, от приблизительно 60% до приблизительно 65% или по меньшей мере приблизительно 0,5%, по меньшей мере приблизительно 1,0%, по меньшей мере приблизительно 2,0%, по меньшей мере приблизительно 3,0%, по меньшей мере приблизительно 4,0%, по меньшей мере приблизительно 4,1%, по меньшей мере приблизительно 4,2%, по меньшей мере приблизительно 4,3%, по меньшей мере приблизительно 4,4%, по меньшей мере приблизительно 4,5%, по меньшей мере приблизительно 5,0%, по меньшей мере приблизительно 5,1%, по меньшей мере приблизительно 5,2%, по меньшей мере приблизительно 5,3%, по меньшей мере приблизительно 5,4%, по меньшей мере приблизительно 5,5%, по меньшей мере приблизительно 5,6%, по меньшей мере приблизительно 5,7%, по меньшей мере приблизительно 5,8%, по меньшей мере приблизительно 5,9%, по меньшей мере приблизительно 6,0%, по меньшей мере приблизительно 7,0%, по меньшей мере приблизительно 8,0%, по меньшей мере приблизительно 9,0%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 11%, по меньшей мере приблизительно 12%, по меньшей мере приблизительно 13%, по меньшей мере приблизительно 14%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 16%, по меньшей мере приблизительно 17%, по меньшей мере приблизительно 18%, по меньшей мере приблизительно 19%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 37%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 45%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 55%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 65%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% или по меньшей мере приблизительно 99% по сравнению с контрольными растениями табака, такими как растение табака дикого типа, в случае выращивания в поле.
Гетерологичная нитратредуктаза
Дерегулированная полипептидная нитратредуктаза может кодироваться гетерологичным полинуклеотидом или геном нитратредуктазы, т. е. геном нитратредуктазы, который не является эндогенным для модифицированного растения табака. Гетерологичный ген нитратредуктазы можно ввести в геном модифицированного растения табака посредством трансформации растения, как описано ниже. Например, гетерологичный ген нитратредуктазы можно ввести в модифицированное растение табака с помощью способов стабильной трансформации или транзиентной трансформации.
Гетерологичный ген нитратредуктазы может представлять собой ген нитратредуктазы растения, гриба или водоросли. Например, гетерологичный ген нитратредуктазы может происходить из Ricinus communis, Ricinus communis, Vitis vinifera, Populus trichocarpa, Selaginella moellendorffii, Selaginella moellendorffii, Cucumis sativus, Cucurbita maxima, Cucumis sativus, Brassica napus, Populus trichocarpa, Betula pendula, Solanum tuberosum, Nicotiana tomentosiformis, Nicotiana tabacum, Solanum lycopersicum, Nicotiana benthamiana, Medicago truncatula, Nicotiana benthamiana, Brassica rapa subsp. chinensis, Beta vulgaris, Nicotiana sylvestris, другой разновидности Nicotiana tabacum, Nicotiana plumbaginifolia, Glycine max, Solanum tuberosum, Solanum tuberosum, Brassica napus, Arabidopsis lyrata subsp. lyrata, Arabidopsis thaliana, Phaseolus vulgaris, Medicago truncatula, Petunia x hybrida, Spinacia oleracea, Lotus japonicus, Cichorium intybus, Petunia x hybrida, Spinacia oleracea, Thellungiella halophile, Oryza sativa японской группы, Zea mays, Arabidopsis thaliana, Arabidopsis lyrata subsp. lyrata, Phaseolus vulgaris, Sorghum bicolor, Oryza sativa индийской группы, Oryza sativa японской группы, Hordeum vulgare subsp. vulgare, Sorghum bicolor, Oryza sativa японской группы, Glycine max, Hordeum vulgare subsp. vulgare, Sorghum bicolor, Zea mays, Heterosigma akashiwo, Chlorella vulgaris UTEX259, Chlorella vulgaris NJ-7, Chlorella variabilis, Ectocarpus siliculosus, Volvox carteri f. nagariensis, Gracilaria tenuistipitata, Dunaliella tertiolecta, Chlamydomonas reinhardtii, Dunaliella viridis, Thalassiosira pseudonana CCMP1335, Dunaliella salina, Phaeodactylum tricornutum или Ostreococcus lucimarinus CCE9901.
Гетерологичный ген нитратредуктазы может представлять собой ген нитратредуктазы, имеющий последовательность с номером доступа в GenBank XP_002513830.1, AAG30576.1, XP_002285831.1, XP_002307415.1, XP_002984312.1, XP_002972481.1, ADN96689.1, P17569.1, ADK77877.1, P39867.1, XP_002301015.1, P27783.1, BAB93534.1, 227925, P11605.1, P17570.1, BAE46746.1, ADV03139.1, BAE96752.1, ACF93242.1, ABW05098.1, 227926, P08509.2, P39870.1, AAB18985.1, AAB52786.1, P39868.1, XP_002889158.1, NP_177899.1, P39866.1, ADV03138.1, P36859.1, P23312.1, P39869.1, P43101.1, AAA33712.1, BAA13047.1, BAJ33682.1, NP_001062006.1, AAD38068.1, NP_174901.1, XP_002891229.1, P39865.1, XP_002444490.1, EEC74079.1, NP_001048253.1, P27967.1, XP_002454625.1, NP_001062009.1, P54233.1, P27968.1, XP_002454083.1, P49102.1, ACS44801.1, ABP97095.1, ABJ91208.4, EFN52691.1, CBN78746.1, XP_002955156.1, ACX31652.1, AAL79356.1, AAF17595.1, AAT72293.1, XP_002294410.1, AAP75705.1, AAV66996.1 или XP_001420098.1.
Модификация эндогенного гена нитратредуктазы
Табак (Nicotiana tabacum) имеет два высокогомологичных гена NR табака, обозначаемых как Nia1 и Nia2, т. е. два эндогенных гена нитратредуктазы. Ген дерегулированной нитратредуктазы может представлять собой эндогенный ген нитратредуктазы, который является модифицированным или подвергнутым мутагенезу с помощью системы редактирования генома или с помощью мутагена. Мутации в нуклеотидных последовательностях и полипептидах, описанных в данном документе, могут включать в себя внесенные человеком мутации или синтетические мутации либо мутации, созданные с помощью генной инженерии. Мутации в нуклеотидных последовательностях и полипептидах, описанных в данном документе, могут представлять собой мутации, которые получены или которые можно получить посредством способа, который включает этап манипуляции in vitro или in vivo. Мутации в нуклеотидных последовательностях и полипептидах, описанных в данном документе, могут представлять собой мутации, которые получены или которые можно получить посредством способа, который включает вмешательство человека.
Способы получения мутантов хорошо известны из уровня техники и могут включать мутагенез с использованием экзогенно добавляемых химических веществ, таких как мутагенные, тератогенные или канцерогенные органические соединения, например, этилметансульфонат (EMS), которые вызывают случайные мутации в генетическом материале. В качестве дополнительного примера способ может включать один или несколько этапов осуществления генной инженерии, таких как один или несколько этапов осуществления генной инженерии, которые описаны в данном документе, или их комбинации. Также можно использовать целенаправленное введение индуцированных локальных повреждений в геномах (TILLING), как описано в другом месте.
В качестве дополнительного примера способ может включать один или несколько этапов скрещивания растений. В одном варианте осуществления семена растений подвергают мутагенезу, а затем из них выращивают мутантные растения первого поколения. Затем обеспечивают возможность самоопыления растений первого поколения, и из семян растений первого поколения выращивают растения второго поколения, которые затем подвергают скринингу на наличие мутаций в их локусах. Хотя подвергнутый мутагенезу растительный материал можно подвергать скринингу на наличие мутаций, преимуществом скрининга растений второго поколения является то, что все соматические мутации соответствуют герминативным мутациям. Специалисту в данной области будет понятно, что ряд растительных материалов, в том числе, без ограничения, семена, пыльцу, растительную ткань или растительные клетки, можно подвергать мутагенезу для создания мутантных растений. Однако тип растительного материала, подвергаемого мутагенезу, может иметь значение в тех случаях, когда нуклеиновую кислоту растения подвергают скринингу на наличие мутаций. Например, если пыльцу подвергают мутагенезу до опыления не подвергаемого мутагенезу растения, то из семян, полученных в результате такого опыления, выращивают растения первого поколения. Каждая клетка растений первого поколения будет содержать мутации, созданные в пыльце; таким образом, эти растения первого поколения можно затем подергать скринингу на наличие мутаций вместо того, чтобы дожидаться получения второго поколения.
Система редактирования генома
Система редактирования генома может связываться с эндогенным геном нитратредуктазы и расщеплять его в его местоположении с образованием усеченной полипептидной нитратредуктазы или полипептидной нитратредуктазы, имеющей аминокислотную замену. Эндогенный ген нитратредуктазы можно модифицировать с помощью системы редактирования генома, которая содержит сайт-специфичную нуклеазу, связывающуюся с эндогенным геном нитратредуктазы и расщепляющую его. В системе редактирования генома может использоваться опосредованное нуклеазами негомологичное соединение концов или репарация путем гомологичной рекомбинации. Систему редактирования генома можно ввести в модифицированное растение табака с помощью способов стабильной трансформации или транзиентной трансформации.
Сайт-специфичная нуклеаза может быть сконструированной. Например, сконструированная сайт-специфичная нуклеаза может представлять собой систему на основе CRISPR/Cas9, ZFN или TALEN. Сайт-специфичная нуклеаза может связываться с эндогенным геном нитратредуктазы и расщеплять его. Система редактирования генома может включать в себя сконструированную систему CRISPR/Cas, такую как сконструированная система CRISPR/Cas-9, сконструированную эффекторную нуклеазу, подобную активаторам транскрипции, сконструированную нуклеазу с «цинковыми пальцами» или сконструированную мегануклеазу.
Мутагены
Для создания мутаций в гене нитратредуктазы можно применять мутагены, которые создают главным образом точечные мутации и короткие делеции, вставки, трансверсии и/или транзиции, в том числе химические мутагены или излучение. Мутагены включают без ограничения этилметансульфонат, метилметансульфонат, N-этил-N-нитрозомочевину, триэтилмеламин, N-метил-N-нитрозомочевину, прокарбазин, хлорамбуцил, циклофосфамид, диэтилсульфат, акриламид-мономер, мелфалан, азотистый иприт, винкристин, диметилнитрозамин, N-метил-N'-нитронитрозогуанидин, нитрозогуанидин, 2-аминопурин, 7,12-диметилбенз(a)антрацен, этиленоксид, гексаметилфосфорамид, бисульфан, диэпоксиалканы (диэпоксиоктан, диэпоксибутан и т. п.), 2-метокси-6-хлор-9[3-(этил-2-хлорэтил)аминопропиламино]акридина дигидрохлорид и формальдегид.
Комбинация с геном, отвечающим за синтез алкалоидного предшественника
Дерегулированную полипептидную нитратредуктазу, целенаправленно воздействующую на нитрозирующее средство, можно использовать в комбинации с модифицированным геном, отвечающим за синтез алкалоидного предшественника, который ингибирует выработку алкалоидного предшественника норникотина. Комбинация дерегулированной полипептидной нитратредуктазы и модифицированного гена, отвечающего за синтез алкалоидного предшественника, может вызывать еще более значительное уменьшение накопления NNN по сравнению с использованием дерегулированной нитратредуктазы или модифицированного гена, отвечающего за синтез алкалоидного предшественника, по отдельности. Модифицированный ген, отвечающий за синтез алкалоидного предшественника, может обеспечивать уменьшение содержания специфического TSNA, такого как N'-нитрозонорникотин, путем уменьшения уровней алкалоидного предшественника норникотина до наиболее низкого содержания норникотина. Модифицированный ген, отвечающий за синтез алкалоидного предшественника, может представлять собой модифицированный ген никотиндеметилазы или ген цитохрома P450. Модифицированный ген, отвечающий за синтез алкалоидного предшественника, может представлять собой модифицированный ген CYP82E4, модифицированный ген CYP82E5v2 или модифицированный ген CYP82E10 и их комбинации. В некоторых вариантах осуществления дерегулированную полипептидную нитратредуктазу можно ввести в растение табака, которое содержит нокаут-мутацию в эндогенном гене никотиндеметилазы, такое как линия скрещивания DH98-325-6#775 (Lewis et al. Phytochem. 71:1988-1998 (2010)), или в растение табака, имеющее тройную мутацию в генетическом окружении гена никотиндеметилазы. В некоторых вариантах осуществления дерегулированную полипептидную нитратредуктазу можно ввести в растение табака, которое имеет нокаут-мутацию как в гене CYP82E4, так и в гене CYP82E5 никотиндеметилазы, такое как сорт Берли TN90e4e5, который является линией, представляющей собой полученный в результате возвратного скрещивания вариант коммерческой разновидности TN90, и описан в заявке на патент США № 20120216822, включенной в данный документ посредством ссылки во всей своей полноте.
Конструкции, вектор и вектор экспрессии
В некоторых вариантах осуществления полинуклеотиды, предназначенные для введения в растение, функционально связаны с промоторной последовательностью и могут быть предоставлены в виде конструкции. Как используется в данном документе, полинуклеотид является «функционально связанным», если он находится в функциональной взаимосвязи со второй полинуклеотидной последовательностью. Например, промотор является функционально связанным с кодирующей последовательностью, если промотор соединен с кодирующей последовательностью таким образом, что он может оказывать влияние на транскрипцию кодирующей последовательности. В различных вариантах осуществления полинуклеотиды могут быть функционально связаны по меньшей мере с одним, по меньшей мере с двумя, по меньшей мере с тремя, по меньшей мере с четырьмя, по меньшей мере с пятью или по меньшей мере с десятью промоторами.
Последовательности нуклеиновых кислот могут образовывать генетическую конструкцию, которая может представлять собой вектор. Вектор может быть способен экспрессировать дерегулированную полипептидную нитратредуктазу или систему редактирования генома в клетке растения. Вектор может быть рекомбинантным. Вектор может содержать гетерологичную нуклеиновую кислоту, кодирующую дерегулированную полипептидную нитратредуктазу или систему редактирования генома. Подходящие векторы включают в себя плазмиды и векторы, полученные из вирусов. Можно использовать векторы, известные из уровня техники, которые подходят для введения в растения путем трансформации, клонирования и экспрессии белка. Вектор может быть применим для трансфекции клеток нуклеиновой кислотой, кодирующей дерегулированную полипептидную нитратредуктазу или систему редактирования генома, при этом трансформируемую клетку-хозяина культивируют и поддерживают в условиях, в которых происходит экспрессия дерегулированной полипептидной нитратредуктазы или системы редактирования генома.
Генетические конструкции могут содержать последовательность нуклеиновой кислоты, которая кодирует дерегулированную полипептидную нитратредуктазу или систему редактирования генома, раскрытые в данном документе. Генетическая конструкция может присутствовать в клетке как функционирующая внехромосомная молекула. Генетическая конструкция может представлять собой линейную минихромосому, содержащую центромеру, теломеры, или плазмиды или космиды. Генетическая конструкция может быть также частью генома рекомбинантного вирусного вектора, в том числе векторов на основе рекомбинантного вируса мозаики цветной капусты, рекомбинантного вируса табачной мозаики и рекомбинантного Х-вируса картофеля. Генетическая конструкция может быть частью генетического материала в аттенуированных живых микроорганизмах, которые живут в клетках, или рекомбинантных микробных векторах. Генетические конструкции могут содержать регуляторные элементы для экспрессии генов с кодирующих последовательностей нуклеиновой кислоты. Регуляторные элементы могут представлять собой промотор, энхансер, инициаторный кодон, стоп-кодон или сигнал полиаденилирования.
Вектор может содержать гетерологичную нуклеиновую кислоту, кодирующую дерегулированную полипептидную нитратредуктазу, и может дополнительно содержать инициаторный кодон, который может быть расположен выше последовательности, кодирующей дерегулированную полипептидную нитратредуктазу, и стоп-кодон, который может быть расположен ниже последовательности, кодирующей дерегулированную полипептидную нитратредуктазу. Инициаторный и терминаторный кодоны могут находиться внутри рамки считывания с последовательностью, кодирующей дерегулированную полипептидную нитратредуктазу. Вектор может также содержать промотор, который функционально связан с последовательностью, кодирующей дерегулированную полипептидную нитратредуктазу. Промотор, который функционально связан с последовательностью, кодирующей дерегулированную полипептидную нитратредуктазу, может в нативных условиях не быть связанным с полинуклеотидом, кодирующим дерегулированную полипептидную нитратредуктазу. Промоторы, применимые для практического осуществления настоящего изобретения, включают в себя без ограничения конститутивные, индуцируемые, регулируемые по времени, регулируемые в процессе развития, химически регулируемые, предпочтительные для определенных тканей и тканеспецифичные промоторы. В подходящем случае промотор вызывает в растении экспрессию, достаточную для получения фенотипа, описанного в данном документе. Подходящие промоторы включают в себя без ограничения промотор 35S вируса мозаики цветной капусты, промотор CYP82E4, убиквитиновый промотор, криптический конститутивный промотор tCUP, промотор Rsyn7, патоген-индуцируемые промоторы, промотор In2-2 маиса, промотор PR-1a табака, глюкокортикоид-индуцируемые промоторы, а также тетрациклин-индуцируемые и тетрациклин-репрессируемые промоторы.
Вектор может также содержать сигнал полиаденилирования, который может быть расположен ниже последовательности, кодирующей дерегулированную полипептидную нитратредуктазу. Вектор может также содержать энхансер выше последовательности, кодирующей дерегулированную полипептидную нитратредуктазу. Энхансер может быть необходим для экспрессии ДНК. Вектор может также содержать растительную точку начала репликации для поддержания вектора во внехромосомном состоянии и получения нескольких копий вектора в клетке. Вектор может также содержать регуляторную последовательность, которая может хорошо подходить для экспрессии гена в растительной клетке, в которую вводят вектор. Вектор может также содержать репортерный ген, такой как ген зеленого флуоресцентного белка («GFP»), и/или селективный маркер, такой как ген устойчивости к гигромицину («Hygro»).
Кодирующие последовательности могут быть оптимизированы для обеспечения стабильности и высоких уровней экспрессии. В некоторых случаях кодоны выбирают так, чтобы уменьшить образование вторичной структуры РНК, как, например, образующейся за счет формирования внутримолекулярных связей.
Вектор может представлять собой векторы или системы экспрессии для получения белка с помощью обычных методик и легкодоступных исходных материалов, в том числе описанных в Sambrook et al., 1989, включенной в полном объеме посредством ссылки. В некоторых вариантах осуществления вектор может содержать последовательность нуклеиновой кислоты, кодирующую дерегулированную полипептидную нитратредуктазу. Вектор может представлять собой pBI121.
Трансформация растений
Полинуклеотид, кодирующий дерегулированную нитратредуктазу, можно ввести в растительную клетку для получения трансгенного растения. Выражение «введенный в растение», используемое в данном документе в отношении полинуклеотидов, охватывает доставку полинуклеотида в растение, растительную ткань или растительную клетку с помощью любого подходящего способа доставки полинуклеотидов. Способы, подходящие для введения полинуклеотидов в растение, применимые для практического осуществления настоящего изобретения, включают без ограничения способ замораживания-оттаивания, бомбардировку микрочастицами, прямое поглощение ДНК, электропорацию, ультразвуковую обработку, микроинъекцию, перенос в растение, опосредованный растительным вирусом и опосредованный Agrobacterium. Для трансформации растения в соответствии с настоящим изобретением можно использовать любой подходящий штамм Agrobacterium, вектор или векторную систему. В некоторых вариантах осуществления полинуклеотид вводят с применением по меньшей мере одного из способов стабильной трансформации, способов транзиентной трансформации или способов, опосредованных вирусами.
Под выражением «стабильная трансформация» подразумевается, что вводимая в растение нуклеотидная конструкция интегрируется в геном растения и способна наследоваться его потомством. Например, введенную последовательность нуклеотидов и/или связанный селектируемый маркерный ген, как, например, систему редактирования генома, можно сегрегировать в последующих поколениях растений с применением традиционных методик селекции.
Под выражением «транзиентная трансформация» подразумевается, что вводимая в растение нуклеотидная конструкция не интегрируется в геном растения. Растение или растительную клетку также можно подвергнуть транзиентной трансформации таким образом, что рекомбинантный полинуклеотид не будет интегрироваться в их геном. Как правило, транзиентно трансформированные клетки утрачивают весь введенный рекомбинантный полинуклеотид или некоторую его часть при каждом клеточном делении, так что введенный рекомбинантный полинуклеотид невозможно выявить в дочерних клетках после прохождения достаточного количества клеточных делений. Например, растительные клетки можно изначально трансформировать системой редактирования генома, не содержащей селектируемый маркер, и можно выращивать на средах, не содержащих средство для отбора. В таких условиях часть обработанных клеток приобретет систему редактирования генома и будет экспрессировать систему редактирования генома транзиентно без интеграции системы редактирования генома в геном. Поскольку это не обеспечивает эффективность трансформации, то для этой последней процедуры трансформации требуется скрининг большего количества обработанных клеток для получения требуемой модификации генома.
Подходящие способы введения нуклеотидных последовательностей в растительные клетки и последующего встраивания в геном растения включают микроинъекцию (Crossway et al., Biotechniques 4:320-334 (1986)), электропорацию (Riggs et al., Proc. Natl. Acad. Sci. USA 83:5602-5606 (1986)), трансформацию, опосредованную Agrobacterium (патент США №№ 5981840 и 5563055), прямой перенос генов (Paszkowski et al., EMBO J. 3:2717-2722 (1984)) и баллистическое ускорение частиц (см, например, патенты США №№ 4945050; 5879918; 5886244; 5932782; Tomes et al., в Plant Cell, Tissue, and Organ Culture: Fundamental Methods, ed. Gamborg and Phillips (Springer-Verlag, Berlin) (1995), а также McCabe et al., Biotechnology 6:923-926 (1988)), все из которых включены в данный документ посредством ссылки во всей своей полноте.
В некоторых вариантах осуществления растение можно регенерировать или вырастить из растения, растительной ткани или растительной клетки. Можно использовать любые подходящие способы регенерации или выращивания растения из растительной клетки или растительной ткани, такие как, без ограничения, культивирование тканей или регенерация из протопластов. В подходящем случае растения можно регенерировать путем выращивания трансформированных растительных клеток на среде для индукции образования каллюса, среде для индукции образования побегов и/или среде для индукции образования корней. См., например, McCormick et al., Plant Cell Reports 5:81-84 (1986). Затем можно выращивать данные растения и опылять их одним и тем же трансформированным штаммом либо разными штаммами и идентифицировать получаемый в результате гибрид, характеризующийся экспрессией требуемой фенотипической характеристики. Можно выращивать два или более поколений, чтобы убедиться, что экспрессия требуемой фенотипической характеристики стабильно поддерживается и наследуется, и затем собирать семена, чтобы убедиться, что экспрессия требуемой фенотипической характеристики была достигнута. Таким образом, выражение «трансформированные семена», используемое в данном документе, относится к семенам, которые содержат нуклеотидную конструкцию, стабильно интегрированную в геном растения.
Разновидности табака
Разновидности табака, которые можно модифицировать в соответствии с раскрытыми способами, включают без ограничения темные разновидности (например, Берли), разновидности трубоогневой сушки или светлые разновидности (например, Вирджиния трубоогневой сушки), восточные или турецкие разновидности, например, Латакия, а также генетически модифицированные разновидности (например, Вектор 2141). Растения табака Берли обычно накапливают более высокие уровни нитратов, чем растения табака других типов, и обычно привносят непропорционально большую долю TSNA, обнаруживаемую в широко распространенных сигаретах типа «Американская мешка» (которые содержат также разновидности табака трубоогневой сушки и восточного табака).
Табачный продукт с уменьшенным содержанием табак-специфичных нитрозаминов
В одном аспекте настоящее изобретение направлено на табачный продукт, характеризующийся уменьшенным содержанием табак-специфичных нитрозаминов (TSNA). TSNA вырабатываются во время процесса сушки табака и во время курения. TSNA образуются путем реакции между нитрозирующим средством и алкалоидами, обнаруживаемыми в табаке в естественных условиях, а также представляют собой канцерогены в табачном дыме. Табачный продукт получают из модифицированного растения табака, как описано выше.
Модифицированные растения табака могут быть свежесобранными, высушенными и переработанными в табачные продукты, которые характеризуются уменьшенным канцерогенным потенциалом. Листья могут быть высушенными на воздухе, подвергнутыми огневой сушке, подвергнутыми трубоогневой сушкой или высушенными на солнце. Табак воздушной сушки подвешивают в хорошо вентилируемых хранилищах и оставляют сушиться в течение периода от четырех до восьми недель. Табак воздушной сушки имеет низкое содержание сахара, что придает табачному дыму легкий мягкий вкус, и высокое содержание никотина. Сигарный табак и табак Берли представляют собой «темные» разновидности табака воздушной сушки. Табак огневой сушки подвешивают в больших хранилищах, где огонь от сжигания древесины твердых пород оставляют при непрерывном или прерывистом медленном горении, что занимает от трех дней до десяти недель в зависимости от способа и разновидности табака. При огневой сушке получают табак с низким содержанием сахара и высоким содержанием никотина. Табак трубоогневой сушки вначале нанизывают на палки для навешивания пучков табачных листьев, которые подвешивают на расположенных в ряд жердях в хранилищах для сушки. Данные хранилища имеют дымоходы, проходящие от топочных камер с внешней подачей, благодаря чему табак высушивается теплом, не подвергаясь действию дыма, и медленно поднимается температура в ходе сушки. Процесс обычно занимает приблизительно неделю, и в ходе него получают сигаретный табак, который характеризуется высоким содержанием сахара и уровнями никотина от средних до высоких. Большинство сигарет содержит табак трубоогневой сушки, который дает более мягкий, легче вдыхаемый дым. Табак солнечной сушки томят на солнце ненакрытым. Данный способ используется в Турции, Греции и других средиземноморских странах для получения восточного табака. Табак солнечной сушки имеет низкое содержание сахара и никотина и используется в сигаретах.
Также вариантами осуществления являются табачные продукты, включающие без ограничения курительные материалы (например, сигареты, сигары, трубочный табак, ароматизированный табак для кальяна), нюхательный табак, табак, который закладывают за губу, заменители сигарет, такие как никотиновый пластырь, никотиносодержащий ополаскиватель для рта, распыляемое средство, упакованное в газовый баллон высокого давления с газом-вытеснителем, никотиносодержащую жевательную резинку и никотиносодержащий напиток, а также леденцы, полученные из указанных обработанных модифицированных растений табака. Также вариантами осуществления являются табачные продукты, которые используются в качестве жидкости для электронных сигарет или раствора никотина для электронных сигарет, персонального парогенератора или электронных систем доставки никотина.
Табачные продукты, полученные из модифицированных растений табака, описанные в данном документе, могут характеризоваться уменьшенными уровнями TSNA по сравнению с табачными продуктами, полученными из контрольного растения табака, такого как контрольное растение табака дикого типа. В некоторых вариантах осуществления табачные продукты, полученные из модифицированных растений табака, могут характеризоваться уменьшенными общими уровнями TSNA по сравнению с табачными продуктами, полученными из контрольного растения табака, такого как контрольное растение табака дикого типа. В некоторых вариантах осуществления табачные продукты, полученные из модифицированных растений табака, могут характеризоваться уменьшенными уровнями по меньшей мере одного TSNA, в том числе, без ограничения, N-нитрозонорникотина (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанона (NNK), N-нитрозоанабазина (NAB), N-нитрозоанатабина (NAT) и их комбинаций, по сравнению с табаком той же разновидности, культивируемым с помощью традиционных методик, или табачным продуктом, полученным из традиционного табака.
Табачный продукт, полученный из высушенных листьев модифицированного растения табака, может характеризоваться по меньшей мере приблизительно 5,0% уменьшением, по меньшей мере приблизительно 10,0% уменьшением, по меньшей мере приблизительно 15,0% уменьшением, по меньшей мере приблизительно 20,0% уменьшением, по меньшей мере приблизительно 25,0% уменьшением, по меньшей мере приблизительно 30,0% уменьшением, по меньшей мере приблизительно 35,0% уменьшением, по меньшей мере приблизительно 40,0% уменьшением, по меньшей мере приблизительно 45,0% уменьшением, по меньшей мере приблизительно 50,0% уменьшением, по меньшей мере приблизительно 55,0% уменьшением, по меньшей мере приблизительно 60,0% уменьшением, по меньшей мере приблизительно 65,0% уменьшением, по меньшей мере приблизительно 70,0% уменьшением, по меньшей мере приблизительно 75,0% уменьшением, по меньшей мере приблизительно 80,0% уменьшением, по меньшей мере приблизительно 85,0% уменьшением, по меньшей мере приблизительно 90,0%, по меньшей мере приблизительно 95,0% уменьшением или по меньшей мере приблизительно 99,0% уменьшением общих уровней TSNA по сравнению с контрольным табачным продуктом, полученным из высушенных листьев контрольного растения табака. Табачный продукт, полученный из высушенных листьев модифицированного растения табака, может характеризоваться уменьшением общих уровней TSNA, составляющим от приблизительно 5,0% до приблизительно 99,0%, от приблизительно 5,0% до приблизительно 95,0%, от приблизительно 5,0% до приблизительно 90,0%, от приблизительно 5,0% до приблизительно 85,0%, от приблизительно 5,0% до приблизительно 80,0%, от приблизительно 5,0% до приблизительно 75,0%, от приблизительно 5,0% до приблизительно 70,0%, от приблизительно 5,0% до приблизительно 65,0%, от приблизительно 5,0% до приблизительно 60,0%, от приблизительно 5,0% до приблизительно 55,0%, от приблизительно 5,0% до приблизительно 50,0%, от приблизительно 5,0% до приблизительно 45,0%, от приблизительно 5,0% до приблизительно 40,0%, от приблизительно 10,0% до приблизительно 99,0%, от приблизительно 10,0% до приблизительно 95,0%, от приблизительно 10,0% до приблизительно 90,0%, от приблизительно 10,0% до приблизительно 85,0%, от приблизительно 10,0% до приблизительно 80,0%, от приблизительно 10,0% до приблизительно 75,0%, от приблизительно 10,0% до приблизительно 70,0%, от приблизительно 10,0% до приблизительно 65,0%, от приблизительно 10,0% до приблизительно 60,0%, от приблизительно 10,0% до приблизительно 55,0%, от приблизительно 10,0% до приблизительно 50,0%, от приблизительно 10,0% до приблизительно 45,0%, от приблизительно 10,0% до приблизительно 40,0%, от приблизительно 20,0% до приблизительно 99,0%, от приблизительно 20,0% до приблизительно 95,0%, от приблизительно 20,0% до приблизительно 90,0%, от приблизительно 20,0% до приблизительно 85,0%, от приблизительно 20,0% до приблизительно 80,0%, от приблизительно 20,0% до приблизительно 75,0%, от приблизительно 20,0% до приблизительно 70,0%, от приблизительно 20,0% до приблизительно 65,0%, от приблизительно 20,0% до приблизительно 60,0%, от приблизительно 20,0% до приблизительно 55,0%, от приблизительно 20,0% до приблизительно 50,0%, от приблизительно 20,0% до приблизительно 45,0%, от приблизительно 20,0% до приблизительно 40,0%, от приблизительно 30,0% до приблизительно 99,0%, от приблизительно 30,0% до приблизительно 95,0%, от приблизительно 30,0% до приблизительно 90,0%, от приблизительно 30,0% до приблизительно 85,0%, от приблизительно 30,0% до приблизительно 80,0%, от приблизительно 30,0% до приблизительно 75,0%, от приблизительно 30,0% до приблизительно 70,0%, от приблизительно 30,0% до приблизительно 65,0%, от приблизительно 30,0% до приблизительно 60,0%, от приблизительно 30,0% до приблизительно 55,0%, от приблизительно 30,0% до приблизительно 50,0%, от приблизительно 30,0% до приблизительно 45,0%, от приблизительно 30,0% до приблизительно 40,0%, от приблизительно 40,0% до приблизительно 99,0%, от приблизительно 40,0% до приблизительно 95,0%, от приблизительно 40,0% до приблизительно 90,0%, от приблизительно 40,0% до приблизительно 85,0%, от приблизительно 40,0% до приблизительно 80,0%, от приблизительно 40,0% до приблизительно 75,0%, от приблизительно 40,0% до приблизительно 70,0%, от приблизительно 40,0% до приблизительно 65,0%, от приблизительно 40,0% до приблизительно 60,0%, от приблизительно 40,0% до приблизительно 55,0%, от приблизительно 40,0% до приблизительно 50,0%, от приблизительно 50,0% до приблизительно 99,0%, от приблизительно 50,0% до приблизительно 95,0%, от приблизительно 50,0% до приблизительно 90,0%, от приблизительно 50,0% до приблизительно 85,0%, от приблизительно 50,0% до приблизительно 80,0%, от приблизительно 50,0% до приблизительно 75,0%, от приблизительно 50,0% до приблизительно 70,0%, от приблизительно 50,0% до приблизительно 65,0%, от приблизительно 50,0% до приблизительно 60,0%, от приблизительно 50,0% до приблизительно 55,0%, от приблизительно 60,0% до приблизительно 99,0%, от приблизительно 60,0% до приблизительно 95,0%, от приблизительно 60,0% до приблизительно 90,0%, от приблизительно 60,0% до приблизительно 85,0%, от приблизительно 60,0% до приблизительно 80,0%, от приблизительно 60,0% до приблизительно 75,0%, от приблизительно 60,0% до приблизительно 70,0% или от приблизительно 60,0% до приблизительно 65,0% по сравнению с контрольным табачным продуктом, полученным из высушенных листьев контрольного растения табака.
Табачный продукт, полученный из высушенных листьев модифицированного растения табака, может характеризоваться по меньшей мере приблизительно 5,0% уменьшением, по меньшей мере приблизительно 10,0% уменьшением, по меньшей мере приблизительно 15,0% уменьшением, по меньшей мере приблизительно 20,0% уменьшением, по меньшей мере приблизительно 25,0% уменьшением, по меньшей мере приблизительно 30,0% уменьшением, по меньшей мере приблизительно 35,0% уменьшением, по меньшей мере приблизительно 40,0% уменьшением, по меньшей мере приблизительно 45,0% уменьшением, по меньшей мере приблизительно 50,0% уменьшением, по меньшей мере приблизительно 55,0% уменьшением, по меньшей мере приблизительно 60,0% уменьшением, по меньшей мере приблизительно 65,0% уменьшением, по меньшей мере приблизительно 70,0% уменьшением, по меньшей мере приблизительно 72,0% уменьшением, по меньшей мере приблизительно 72,5% уменьшением, по меньшей мере приблизительно 73,0% уменьшением, по меньшей мере приблизительно 75,0% уменьшением, по меньшей мере приблизительно 80,0% уменьшением, по меньшей мере приблизительно 85,0% уменьшением, по меньшей мере приблизительно 90,0%, по меньшей мере приблизительно 95,0% уменьшением или по меньшей мере приблизительно 99,0% уменьшением уровней по меньшей мере одного TSNA, в том числе, без ограничения, N-нитрозонорникотина (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанона (NNK), N-нитрозоанабазина (NAB), N-нитрозоанатабина (NAT) и их комбинаций, по сравнению с контрольным табачным продуктом, полученным из высушенных листьев контрольного растения табака. Табачный продукт, полученный из высушенных листьев модифицированного растения табака, может характеризоваться уровнями по меньшей мере одного TSNA, в том числе, без ограничения, N-нитрозонорникотина (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанона (NNK), N-нитрозоанабазина (NAB), N-нитрозоанатабина (NAT) и их комбинаций, составляющими от приблизительно 5,0% до приблизительно 99,0%, от приблизительно 5,0% до приблизительно 95,0%, от приблизительно 5,0% до приблизительно 90,0%, от приблизительно 5,0% до приблизительно 85,0%, от приблизительно 5,0% до приблизительно 80,0%, от приблизительно 5,0% до приблизительно 75,0%, от приблизительно 5,0% до приблизительно 70,0%, от приблизительно 5,0% до приблизительно 65,0%, от приблизительно 5,0% до приблизительно 60,0%, от приблизительно 5,0% до приблизительно 55,0%, от приблизительно 5,0% до приблизительно 50,0%, от приблизительно 5,0% до приблизительно 45,0%, от приблизительно 5,0% до приблизительно 40,0%, от приблизительно 10,0% до приблизительно 99,0%, от приблизительно 10,0% до приблизительно 95,0%, от приблизительно 10,0% до приблизительно 90,0%, от приблизительно 10,0% до приблизительно 85,0%, от приблизительно 10,0% до приблизительно 80,0%, от приблизительно 10,0% до приблизительно 75,0%, от приблизительно 10,0% до приблизительно 70,0%, от приблизительно 10,0% до приблизительно 65,0%, от приблизительно 10,0% до приблизительно 60,0%, от приблизительно 10,0% до приблизительно 55,0%, от приблизительно 10,0% до приблизительно 50,0%, от приблизительно 10,0% до приблизительно 45,0%, от приблизительно 10,0% до приблизительно 40,0%, от приблизительно 20,0% до приблизительно 99,0%, от приблизительно 20,0% до приблизительно 95,0%, от приблизительно 20,0% до приблизительно 90,0%, от приблизительно 20,0% до приблизительно 85,0%, от приблизительно 20,0% до приблизительно 80,0%, от приблизительно 20,0% до приблизительно 75,0%, от приблизительно 20,0% до приблизительно 70,0%, от приблизительно 20,0% до приблизительно 65,0%, от приблизительно 20,0% до приблизительно 60,0%, от приблизительно 20,0% до приблизительно 55,0%, от приблизительно 20,0% до приблизительно 50,0%, от приблизительно 20,0% до приблизительно 45,0%, от приблизительно 20,0% до приблизительно 40,0%, от приблизительно 30,0% до приблизительно 99,0%, от приблизительно 30,0% до приблизительно 95,0%, от приблизительно 30,0% до приблизительно 90,0%, от приблизительно 30,0% до приблизительно 85,0%, от приблизительно 30,0% до приблизительно 80,0%, от приблизительно 30,0% до приблизительно 75,0%, от приблизительно 30,0% до приблизительно 70,0%, от приблизительно 30,0% до приблизительно 65,0%, от приблизительно 30,0% до приблизительно 60,0%, от приблизительно 30,0% до приблизительно 55,0%, от приблизительно 30,0% до приблизительно 50,0%, от приблизительно 30,0% до приблизительно 45,0%, от приблизительно 30,0% до приблизительно 40,0%, от приблизительно 40,0% до приблизительно 99,0%, от приблизительно 40,0% до приблизительно 95,0%, от приблизительно 40,0% до приблизительно 90,0%, от приблизительно 40,0% до приблизительно 85,0%, от приблизительно 40,0% до приблизительно 80,0%, от приблизительно 40,0% до приблизительно 75,0%, от приблизительно 40,0% до приблизительно 70,0%, от приблизительно 40,0% до приблизительно 65,0%, от приблизительно 40,0% до приблизительно 60,0%, от приблизительно 40,0% до приблизительно 55,0%, от приблизительно 40,0% до приблизительно 50,0%, от приблизительно 50,0% до приблизительно 99,0%, от приблизительно 50,0% до приблизительно 95,0%, от приблизительно 50,0% до приблизительно 90,0%, от приблизительно 50,0% до приблизительно 85,0%, от приблизительно 50,0% до приблизительно 80,0%, от приблизительно 50,0% до приблизительно 75,0%, от приблизительно 50,0% до приблизительно 70,0%, от приблизительно 50,0% до приблизительно 65,0%, от приблизительно 50,0% до приблизительно 60,0%, от приблизительно 50,0% до приблизительно 55,0%, от приблизительно 60,0% до приблизительно 99,0%, от приблизительно 60,0% до приблизительно 95,0%, от приблизительно 60,0% до приблизительно 90,0%, от приблизительно 60,0% до приблизительно 85,0%, от приблизительно 60,0% до приблизительно 80,0%, от приблизительно 60,0% до приблизительно 75,0%, от приблизительно 60,0% до приблизительно 70,0% или от приблизительно 60,0% до приблизительно 65,0% по сравнению с контрольным табачным продуктом, полученным из высушенных листьев контрольного растения табака.
Резаный табачный наполнитель и табачный дым с уменьшенным содержанием табак-специфичных нитрозаминов
В одном аспекте настоящее изобретение направлено на табачный продукт, характеризующийся уменьшенным содержанием табак-специфичных нитрозаминов (TSNA) в дыме, получаемом при сгорании листьев при курении, т. е. табачном дыме. Содержание TSNA в основном потоке дыма сигарет после сгорания зависит от следующих факторов: (1) количества предсуществующих TSNA в резаном наполнителе; (2) эффективности перехода предсуществующих TSNA из резаного наполнителя в дым; (3) степени разрушения предсуществующих TSNA посредством пиролиза и (4) степени образования новых TSNA посредством пиросинтеза. Как описано выше, табачный дым получают из модифицированного растения табака, которое характеризуется уменьшенными уровнями свободных нитратов по сравнению с контрольным растением табака, таким как контрольное растение табака дикого типа.
Резаный табачный наполнитель или табачный дым, полученные из модифицированных растений табака, описанных в данном документе, могут характеризоваться уменьшенными уровнями TSNA по сравнению с резаным табачным наполнителем или табачным дымом, полученными из контрольного растения табака, такого как контрольное растение табака дикого типа. В некоторых вариантах осуществления резаный табачный наполнитель или табачный дым, полученные из модифицированных растений табака, могут характеризоваться уменьшенными общими уровнями TSNA по сравнению с резаным табачным наполнителем или табачным дымом, полученными из контрольного растения табака, такого как контрольное растение табака дикого типа. В некоторых вариантах осуществления резаный табачный наполнитель или табачный дым, полученные из модифицированных растений табака, могут характеризоваться уменьшенными уровнями по меньшей мере одного TSNA, в том числе, без ограничения, N-нитрозонорникотина (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанона (NNK), N-нитрозоанабазина (NAB), N-нитрозоанатабина (NAT) и их комбинаций, по сравнению с резаным табачным наполнителем или табачным дымом, полученными из табака той же разновидности, культивируемого с помощью традиционных методик, или табачного продукта, полученного из традиционного табака.
Резаный табачный наполнитель, полученный из модифицированного растения табака, может характеризоваться по меньшей мере приблизительно 5,0% уменьшением, по меньшей мере приблизительно 10,0% уменьшением, по меньшей мере приблизительно 15,0% уменьшением, по меньшей мере приблизительно 20,0% уменьшением, по меньшей мере приблизительно 25,0% уменьшением, по меньшей мере приблизительно 30,0% уменьшением, по меньшей мере приблизительно 32,0% уменьшением, по меньшей мере приблизительно 35,0% уменьшением, по меньшей мере приблизительно 40,0% уменьшением, по меньшей мере приблизительно 44,0% уменьшением, по меньшей мере приблизительно 45,0% уменьшением, по меньшей мере приблизительно 50,0% уменьшением, по меньшей мере приблизительно 52,0% уменьшением, по меньшей мере приблизительно 55,0% уменьшением, по меньшей мере приблизительно 60,0% уменьшением, по меньшей мере приблизительно 64,0% уменьшением, по меньшей мере приблизительно 65,0% уменьшением, по меньшей мере приблизительно 70,0% уменьшением, по меньшей мере приблизительно 72,0% уменьшением, по меньшей мере приблизительно 72,5% уменьшением, по меньшей мере приблизительно 73,0% уменьшением, по меньшей мере приблизительно 75,0% уменьшением, по меньшей мере приблизительно 80,0% уменьшением, по меньшей мере приблизительно 85,0% уменьшением, по меньшей мере приблизительно 90,0%, по меньшей мере приблизительно 95,0% уменьшением или по меньшей мере приблизительно 99,0% уменьшением общих уровней TSNA или уровней по меньшей мере одного TSNA, в том числе, без ограничения, N-нитрозонорникотина (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанона (NNK), N-нитрозоанабазина (NAB), N-нитрозоанатабина (NAT) и их комбинаций, по сравнению с контрольным резаным табачным наполнителем, полученным из контрольного растения табака.
Резаный табачный наполнитель, полученный из табачного продукта, может характеризоваться уменьшением общих уровней TSNA или уровней по меньшей мере одного TSNA, в том числе, без ограничения, N-нитрозонорникотина (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанона (NNK), N-нитрозоанабазина (NAB), N-нитрозоанатабина (NAT) и их комбинаций, составляющим от приблизительно 5,0% до приблизительно 99,0%, от приблизительно 5,0% до приблизительно 95,0%, от приблизительно 5,0% до приблизительно 90,0%, от приблизительно 5,0% до приблизительно 85,0%, от приблизительно 5,0% до приблизительно 80,0%, от приблизительно 5,0% до приблизительно 75,0%, от приблизительно 5,0% до приблизительно 70,0%, от приблизительно 5,0% до приблизительно 65,0%, от приблизительно 5,0% до приблизительно 60,0%, от приблизительно 5,0% до приблизительно 55,0%, от приблизительно 5,0% до приблизительно 50,0%, от приблизительно 5,0% до приблизительно 45,0%, от приблизительно 5,0% до приблизительно 40,0%, от приблизительно 10,0% до приблизительно 99,0%, от приблизительно 10,0% до приблизительно 95,0%, от приблизительно 10,0% до приблизительно 90,0%, от приблизительно 10,0% до приблизительно 85,0%, от приблизительно 10,0% до приблизительно 80,0%, от приблизительно 10,0% до приблизительно 75,0%, от приблизительно 10,0% до приблизительно 70,0%, от приблизительно 10,0% до приблизительно 65,0%, от приблизительно 10,0% до приблизительно 60,0%, от приблизительно 10,0% до приблизительно 55,0%, от приблизительно 10,0% до приблизительно 50,0%, от приблизительно 10,0% до приблизительно 45,0%, от приблизительно 10,0% до приблизительно 40,0%, от приблизительно 20,0% до приблизительно 99,0%, от приблизительно 20,0% до приблизительно 95,0%, от приблизительно 20,0% до приблизительно 90,0%, от приблизительно 20,0% до приблизительно 85,0%, от приблизительно 20,0% до приблизительно 80,0%, от приблизительно 20,0% до приблизительно 75,0%, от приблизительно 20,0% до приблизительно 70,0%, от приблизительно 20,0% до приблизительно 65,0%, от приблизительно 20,0% до приблизительно 60,0%, от приблизительно 20,0% до приблизительно 55,0%, от приблизительно 20,0% до приблизительно 50,0%, от приблизительно 20,0% до приблизительно 45,0%, от приблизительно 20,0% до приблизительно 40,0%, от приблизительно 30,0% до приблизительно 99,0%, от приблизительно 30,0% до приблизительно 95,0%, от приблизительно 30,0% до приблизительно 90,0%, от приблизительно 30,0% до приблизительно 85,0%, от приблизительно 30,0% до приблизительно 80,0%, от приблизительно 30,0% до приблизительно 75,0%, от приблизительно 30,0% до приблизительно 70,0%, от приблизительно 30,0% до приблизительно 65,0%, от приблизительно 30,0% до приблизительно 60,0%, от приблизительно 30,0% до приблизительно 55,0%, от приблизительно 30,0% до приблизительно 50,0%, от приблизительно 30,0% до приблизительно 45,0%, от приблизительно 30,0% до приблизительно 40,0%, от приблизительно 40,0% до приблизительно 99,0%, от приблизительно 40,0% до приблизительно 95,0%, от приблизительно 40,0% до приблизительно 90,0%, от приблизительно 40,0% до приблизительно 85,0%, от приблизительно 40,0% до приблизительно 80,0%, от приблизительно 40,0% до приблизительно 75,0%, от приблизительно 40,0% до приблизительно 70,0%, от приблизительно 40,0% до приблизительно 65,0%, от приблизительно 40,0% до приблизительно 60,0%, от приблизительно 40,0% до приблизительно 55,0%, от приблизительно 40,0% до приблизительно 50,0%, от приблизительно 50,0% до приблизительно 99,0%, от приблизительно 50,0% до приблизительно 95,0%, от приблизительно 50,0% до приблизительно 90,0%, от приблизительно 50,0% до приблизительно 85,0%, от приблизительно 50,0% до приблизительно 80,0%, от приблизительно 50,0% до приблизительно 75,0%, от приблизительно 50,0% до приблизительно 70,0%, от приблизительно 50,0% до приблизительно 65,0%, от приблизительно 50,0% до приблизительно 60,0%, от приблизительно 50,0% до приблизительно 55,0%, от приблизительно 60,0% до приблизительно 99,0%, от приблизительно 60,0% до приблизительно 95,0%, от приблизительно 60,0% до приблизительно 90,0%, от приблизительно 60,0% до приблизительно 85,0%, от приблизительно 60,0% до приблизительно 80,0%, от приблизительно 60,0% до приблизительно 75,0%, от приблизительно 60,0% до приблизительно 70,0% или от приблизительно 60,0% до приблизительно 65,0% по сравнению с контрольным резаным табачным наполнителем, полученным из контрольного растения табака.
Табачный дым, образующийся из табачного продукта, может характеризоваться по меньшей мере приблизительно 5,0% уменьшением, по меньшей мере приблизительно 10,0% уменьшением, по меньшей мере приблизительно 15,0% уменьшением, по меньшей мере приблизительно 20,0% уменьшением, по меньшей мере приблизительно 25,0% уменьшением, по меньшей мере приблизительно 30,0% уменьшением, по меньшей мере приблизительно 32,0% уменьшением, по меньшей мере приблизительно 35,0% уменьшением, по меньшей мере приблизительно 40,0% уменьшением, по меньшей мере приблизительно 44,0% уменьшением, по меньшей мере приблизительно 45,0% уменьшением, по меньшей мере приблизительно 50,0% уменьшением, по меньшей мере приблизительно 52,0% уменьшением, по меньшей мере приблизительно 55,0% уменьшением, по меньшей мере приблизительно 60,0% уменьшением, по меньшей мере приблизительно 64,0% уменьшением, по меньшей мере приблизительно 65,0% уменьшением, по меньшей мере приблизительно 70,0% уменьшением, по меньшей мере приблизительно 72,0% уменьшением, по меньшей мере приблизительно 72,5% уменьшением, по меньшей мере приблизительно 73,0% уменьшением, по меньшей мере приблизительно 75,0% уменьшением, по меньшей мере приблизительно 80,0% уменьшением, по меньшей мере приблизительно 85,0% уменьшением, по меньшей мере приблизительно 90,0%, по меньшей мере приблизительно 95,0% уменьшением или по меньшей мере приблизительно 99,0% уменьшением общих уровней TSNA или уровней по меньшей мере одного TSNA, в том числе, без ограничения, N-нитрозонорникотина (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанона (NNK), N-нитрозоанабазина (NAB), N-нитрозоанатабина (NAT) и их комбинаций, по сравнению с табачным дымом из контрольного табачного продукта, который получен из контрольного растения табака.
Табачный дым, образующийся из табачного продукта, может характеризоваться уменьшением общих уровней TSNA или уровней по меньшей мере одного TSNA, в том числе, без ограничения, N-нитрозонорникотина (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанона (NNK), N-нитрозоанабазина (NAB), N-нитрозоанатабина (NAT) и их комбинаций, составляющим от приблизительно 5,0% до приблизительно 99,0%, от приблизительно 5,0% до приблизительно 95,0%, от приблизительно 5,0% до приблизительно 90,0%, от приблизительно 5,0% до приблизительно 85,0%, от приблизительно 5,0% до приблизительно 80,0%, от приблизительно 5,0% до приблизительно 75,0%, от приблизительно 5,0% до приблизительно 70,0%, от приблизительно 5,0% до приблизительно 65,0%, от приблизительно 5,0% до приблизительно 60,0%, от приблизительно 5,0% до приблизительно 55,0%, от приблизительно 5,0% до приблизительно 50,0%, от приблизительно 5,0% до приблизительно 45,0%, от приблизительно 5,0% до приблизительно 40,0%, от приблизительно 10,0% до приблизительно 99,0%, от приблизительно 10,0% до приблизительно 95,0%, от приблизительно 10,0% до приблизительно 90,0%, от приблизительно 10,0% до приблизительно 85,0%, от приблизительно 10,0% до приблизительно 80,0%, от приблизительно 10,0% до приблизительно 75,0%, от приблизительно 10,0% до приблизительно 70,0%, от приблизительно 10,0% до приблизительно 65,0%, от приблизительно 10,0% до приблизительно 60,0%, от приблизительно 10,0% до приблизительно 55,0%, от приблизительно 10,0% до приблизительно 50,0%, от приблизительно 10,0% до приблизительно 45,0%, от приблизительно 10,0% до приблизительно 40,0%, от приблизительно 20,0% до приблизительно 99,0%, от приблизительно 20,0% до приблизительно 95,0%, от приблизительно 20,0% до приблизительно 90,0%, от приблизительно 20,0% до приблизительно 85,0%, от приблизительно 20,0% до приблизительно 80,0%, от приблизительно 20,0% до приблизительно 75,0%, от приблизительно 20,0% до приблизительно 70,0%, от приблизительно 20,0% до приблизительно 65,0%, от приблизительно 20,0% до приблизительно 60,0%, от приблизительно 20,0% до приблизительно 55,0%, от приблизительно 20,0% до приблизительно 50,0%, от приблизительно 20,0% до приблизительно 45,0%, от приблизительно 20,0% до приблизительно 40,0%, от приблизительно 30,0% до приблизительно 99,0%, от приблизительно 30,0% до приблизительно 95,0%, от приблизительно 30,0% до приблизительно 90,0%, от приблизительно 30,0% до приблизительно 85,0%, от приблизительно 30,0% до приблизительно 80,0%, от приблизительно 30,0% до приблизительно 75,0%, от приблизительно 30,0% до приблизительно 70,0%, от приблизительно 30,0% до приблизительно 65,0%, от приблизительно 30,0% до приблизительно 60,0%, от приблизительно 30,0% до приблизительно 55,0%, от приблизительно 30,0% до приблизительно 50,0%, от приблизительно 30,0% до приблизительно 45,0%, от приблизительно 30,0% до приблизительно 40,0%, от приблизительно 40,0% до приблизительно 99,0%, от приблизительно 40,0% до приблизительно 95,0%, от приблизительно 40,0% до приблизительно 90,0%, от приблизительно 40,0% до приблизительно 85,0%, от приблизительно 40,0% до приблизительно 80,0%, от приблизительно 40,0% до приблизительно 75,0%, от приблизительно 40,0% до приблизительно 70,0%, от приблизительно 40,0% до приблизительно 65,0%, от приблизительно 40,0% до приблизительно 60,0%, от приблизительно 40,0% до приблизительно 55,0%, от приблизительно 40,0% до приблизительно 50,0%, от приблизительно 50,0% до приблизительно 99,0%, от приблизительно 50,0% до приблизительно 95,0%, от приблизительно 50,0% до приблизительно 90,0%, от приблизительно 50,0% до приблизительно 85,0%, от приблизительно 50,0% до приблизительно 80,0%, от приблизительно 50,0% до приблизительно 75,0%, от приблизительно 50,0% до приблизительно 70,0%, от приблизительно 50,0% до приблизительно 65,0%, от приблизительно 50,0% до приблизительно 60,0%, от приблизительно 50,0% до приблизительно 55,0%, от приблизительно 60,0% до приблизительно 99,0%, от приблизительно 60,0% до приблизительно 95,0%, от приблизительно 60,0% до приблизительно 90,0%, от приблизительно 60,0% до приблизительно 85,0%, от приблизительно 60,0% до приблизительно 80,0%, от приблизительно 60,0% до приблизительно 75,0%, от приблизительно 60,0% до приблизительно 70,0% или от приблизительно 60,0% до приблизительно 65,0% по сравнению с табачным дымом из контрольного табачного продукта, полученного из контрольного растения табака.
Способ получения табачного продукта, характеризующегося уменьшенным содержанием TSNA
Настоящее изобретение также направлено на способы получения табачного продукта, характеризующегося уменьшенными уровнями TSNA. В некоторых вариантах осуществления способы, описанные в данном документе, можно применять для введения в растительную клетку выделенного полинуклеотида, кодирующего дерегулированную полипептидную нитратредуктазу; регенерации трансформированной клетки с получением трансгенного растения и получения табачного продукта из листьев трансгенного растения. В некоторых вариантах осуществления способы, описанные в данном документе, можно применять для введения в растительную клетку системы редактирования генома, которая целенаправленно воздействует на эндогенный ген нитратредуктазы; регенерации трансформированной клетки с получением трансгенного растения, а также получения табачного продукта из листьев трансгенного растения.
В некоторых вариантах осуществления растение можно регенерировать или вырастить из растения, растительной ткани или растительной клетки. Можно использовать любые подходящие способы регенерации или выращивания растения из растительной клетки или растительной ткани, такие как, без ограничения, культивирование тканей или регенерация из протопластов. В подходящем случае растения можно регенерировать путем выращивания трансформированных растительных клеток на среде для индукции образования каллюса, среде для индукции образования побегов и/или среде для индукции образования корней. Регенерированные растения обладают практически всеми морфологическими и физиологическими характеристиками модифицированного растения табака.
Способ уменьшения содержания табак-специфичных нитрозаминов в табачном продукте
В одном аспекте настоящее изобретение направлено на способы получения табачного продукта, характеризующегося уменьшенными уровнями TSNA по сравнению с уровнями TSNA в немодифицированном растении табака. Способ включает модификацию растения табака таким образом, чтобы оно содержало полинуклеотид, содержащий последовательность, кодирующую дерегулированный фермент нитратредуктазу, состоящий из нее или по сути состоящий из нее, полипептид, кодируемый полинуклеотидом, изложенным в (i); полипептид, содержащий кодируемую последовательность дерегулированного фермента нитратредуктазы, состоящий из нее или по сути состоящий из нее; или конструкцию, вектор или вектор экспрессии, содержащие полинуклеотид, изложенный в (i), где экспрессия или активность указанной нитратредуктазы является дерегулированной по сравнению с контрольным немодифицированным растением табака; сбор листьев табака с указанного модифицированного растения табака и получение табачного продукта из собранных листьев.
Способ уменьшения содержания табак-специфичных нитрозаминов в дыме табачного продукта
Настоящее изобретение в одном аспекте направлено на способы получения табачного продукта, где уровни TSNA, измеренные в дыме, полученном при сгорании листьев модифицированного растения табака, уменьшены по сравнению с уровнями TSNA, измеренными в дыме, полученном при сгорании немодифицированного растения табака. Способ включает модификацию растения табака таким образом, чтобы оно содержало полинуклеотид, содержащий последовательность, кодирующую дерегулированный фермент нитратредуктазу, состоящий из нее или по сути состоящий из нее, полипептид, кодируемый полинуклеотидом, изложенным в (i); полипептид, содержащий кодируемую последовательность дерегулированного фермента нитратредуктазы, состоящий из нее или по сути состоящий из нее; или конструкцию, вектор или вектор экспрессии, содержащие полинуклеотид, изложенный в (i), где экспрессия или активность указанной нитратредуктазы является дерегулированной по сравнению с контрольным немодифицированным растением табака; сбор листьев табака с указанного модифицированного растения табака и получение табачного продукта из собранных листьев.
Для специалистов в данной области будет очевидно, что другие подходящие модификации и адаптации способов по настоящему раскрытию, описанных в данном документе, легко применимы и поддаются пониманию и могут быть осуществлены с использованием подходящих эквивалентов без отступления от объема настоящего раскрытия или аспектов и вариантов осуществления, раскрытых в данном документе. При наличии подробного описания настоящего раскрытия оно теперь будет более отчетливо понятным со ссылкой на следующие примеры, которые предназначены лишь для иллюстрации некоторых аспектов и вариантов осуществления настоящего раскрытия и не должны рассматриваться как ограничивающие объем настоящего раскрытия. Раскрытия всех упомянутых в данном документе журнальных статей, патентов США и публикаций включены посредством ссылки во всей своей полноте.
Настоящее изобретение имеет несколько аспектов, иллюстрируемых следующими неограничивающими примерами.
Примеры
Пример 1
Способы и материалы
Плазмидные конструкции. Следующие пять генов, вовлеченных в путь ассимиляции нитратов (N-ассимиляцию), клонировали с помощью полимеразной цепной реакции (ПЦР): «S523D-NR» - кДНК Nia2 табака (Vaucheret et al. Plant Mol. Biol. 12:597-200 (1989)), содержащая мутацию по типу замены Ser на Asp, введенную в кодон 523 с помощью сайт-направленного мутагенеза (Lillo et al. Plant J. 35:566-573 (2003)); «tr-NR» - кДНК Nia2 с N-концевым усечением, где кодоны 2-56 были удалены с помощью сайт-направленного мутагенеза (Nussaume et al. Plant Cell 7:611-621 (1995)); «GS1» - кДНК полной длины гена Gln1-3 табака (Dubois et al. Plant Mol. Biol. 31:803-817 (1996)); «GOGAT» - кДНК полной длины гена GLT1Arabidopsis thaliana(Lancienetal.PlantJ.29:347-358(2002)) и «ICDH» - кДНК полной длины гена цитозольной изоцитратдегидрогеназы табака (номер доступа в GenBank 77944).
Для экспрессии различных конструкций у табака каждую кДНК встраивали по отдельности в бинарный вектор pBI121 (Chen et al. Mol. Breed. 11:287-293 (2003)) путем замены репортерного гена GUS внутри данного вектора соответствующей кДНК гена пути N-ассимиляции. В результате этого конструкции были помещены под транскрипционный контроль сильного конститутивного промотора 35S CaMV. Вектор экспрессии для растений pBI121 содержит ген nptII, который обеспечивает отбор трансформированных клеток с использованием антибиотика канамицина. Для конструкций, содержащих промотор CYP82E4 (E4), участок вектора размером ~850 п. о., содержащий промотор 35S, вырезали и заменяли участком размером 2,2 т. п. о., расположенным непосредственно выше стартового кодона CYP82E4 (Chakrabarti et al. Plant Mol. Biol. 66:415-427 (2008)). Полученные в результате векторы экспрессии для растений вводили в штамм EHA105 Agrobacterium tumefaciens и выращивали на среде YEP (1% бактопептон, 1% бакто-дрожжевой экстракт и 0,5% NaCl), дополненной канамицином (50 мг/л) и рифампицином (20 мг/л).
Растительные материалы. Линию скрещивания Берли DH98-325-6#775 трансформировали по отдельности конструкциями 35S:tr-NR, 35S:S523D-NR, 35S:GS1, 35S:GOGAT, 35S:ICDH, E4:tr-NR, E4:S523D-NR и E4:GOGAT. Несколько независимых трансгенных линий для каждой конструкции анализировали с помощью полуколичественной ОТ-ПЦР для идентификации отдельных растений, демонстрирующих наиболее высокие уровни экспрессии трансгенов по сравнению с геном актина табака. Ткань зеленых листьев использовали для анализа экспрессии у растений, содержащих конструкции под транскрипционным контролем промотора 35S CaMV. Для растений, содержащих промотор E4, анализ экспрессии проводили в листьях, которые были отделены и обработаны этефоном, как описано Chakrabarti et al. (2008). Для каждого индивидуума T0 с высокой экспрессией несколько растений T1 выращивали и генотипировали с использованием диагностических праймеров для когнатных трансгенов, чтобы отличить потомство, наследующее трансген, от ноль-сегрегантов. Многочисленное потомство T1 от каждого растения T0, для которого анализ на наличие указанного трансгена был положительным, снова анализировали с помощью полуколичественной ОТ-ПЦР, чтобы проверить, происходит ли адекватная передача фенотипа высокой экспрессии следующему поколению. Для получения потомства поколения T2 использовали линии, которые продемонстрировали единообразную передачу фенотипа высокой экспрессии.
Сорт Берли TN90e4e5 подвергали котрансформации конструкциями 35S:S523D-NR и E4:S523D-NR. Равные количества двух культур Agrobacterium, каждая из которых содержала одну из конструкций S523D-NR, объединяли и использовали для трансформации листовых дисков TN90e4e5. Для того, чтобы различить набор трансгенов в каждом индивидууме T0, проводили молекулярное генотипирование на основе ПЦР. Чтобы различить две конструкции, праймеры, специфичные для промоторных участков 35С или E4, объединяли в пары с праймерами, соответствующими S523D-NR. Растения с контрольным вектором получали путем трансформации TN90e4e5 конструкцией E4:GUS (Chakrabarti et al., 2008). Уровни экспрессии трансгена для каждого растения T0 определяли с помощью ОТ-ПЦР с использованием инструмента Mx3000P для количественной ПЦР в соответствии с протоколом производителя (Agilent Technologies). Данные ОТ-ПЦР с использованием праймеров, соответствующих S523D-NR, нормализовали согласно экспрессии контрольного гена фактора элонгации 1α.
Условия роста для экспериментов в камере с контролируемой средой. Трансгенные растения табака T2 из линий 35S:tr-NR, 35S:S523D-NR, 35S:GS1, 35S:GOGAT, 35S:ICDH и контроли WT выращивали в камерах с контролируемой средой в лабораторном комплексе Phytotron Университета штата Северная Каролина. Семена проращивали на торфо-песчаной смеси с соотношением 2:1. Четырехнедельные сеянцы пересаживали в отдельные ячейки и выращивали с применением режима 12 ч. дня при 26°C/12 ч. ночи при 22°C. Плотность фотосинтетического фотонного потока 1500 мкмоль м2с-1 обеспечивалась металлогалогенными и натриевыми лампами высокого давления в соотношении 1:1, а также лампами накаливания. Растения поливали один раз в день деионизированной водой и один раз в день питательным раствором, содержащим 106,23 мг/л азота, 10,41 мг/л фосфора, 111,03 мг/л калия, 54,4 мг/л кальция, 12,4 мг/л магния, 5 мг/л железа, 13,13 мг/л серы, 0,113 мг/л марганца, 0,24 мг/л бора, 0,013 мг/л цинка, 0,005 мг/л меди, 0,00003 мг/л кобальта, 0,05 мг/л молибдена и 11,04 мг/л натрия.
Примерно через два месяца после посадки 18 растений на генотип пересаживали в 6''-горшки, наполненные речным песком, стерилизованным паром. Следили за тем, чтобы отбирали индивидуумов сходного размера. Шесть растений на генотип подвергали каждому из типов обработки нитратными удобрениями. Гибель растений и/или наблюдение фенотипов с аномальной задержкой роста приводили к утрате четырех растений 35S:ICDH, двух 35S:S523D-NR и одного 35S:GS1.
В течение первых семи дней после пересадки в 6''-горшки все растения поливали питательным раствором, содержащим избыток нитратов (19 мМ NO3-), чтобы дать им возможность приспособиться к шоку от пересадки. Питательный раствор с 19 мМ нитратов содержал следующее: 5 мМ Ca(NO3)2, 2 мМ Mg(NO3)2, 5 мМ KNO3, 1 мМ KH2PO4, 0,5 мМ K2SO4, 19 мкМ H3BO3, 3,7 мкМ MnCl2, 0,3 мкМ ZnSO4, 0,13 мкМ CuSO4, 0,05 мкМ Na2MoO4 и 10,0 мкМ секвестрена 330 Fe. После семидневного периода приспособления растения из каждой генотипической группы разделяли поровну на три группы и поливали питательным раствором с очень низким содержанием нитратов (0,2 мМ NO3-), средним содержанием (8 мМ NO3-) либо высоким содержанием N (19 мМ NO3-) в течение следующих 16 дней. Для поддержания ионного и осмотического баланса при необходимости SO42- заменяли на NO3- для всех растворов с низким и средним содержанием. Таким образом, питательный раствор с низким содержанием N (0,2 мМ NO3-) содержал 0,1 мМ Ca(NO3)2, 3 мМ K2SO4, 2 мМ MgSO4 и 4,9 мМ CaSO4⋅2H2O (без Mg(NO3)2 или KNO3), а питательный раствор со средним содержанием N (8 мМ NO3-) содержала 4 мМ Ca(NO3)2, 3 мМ K2SO4, 2 мМ MgSO4 и 1 мМ CaSO4•2H2O (без Mg(NO3)2 или KNO3).
В ходе 16-дневного эксперимента каждое растение поливали три раза в день: утром 200 мл соответствующего питательного раствора, в полдень 200 мл деионизированной воды и во второй половине дня 1000 мл деионизированной воды для вымывания питательных веществ, а затем 200 мл соответствующего питательного раствора. Условия в камере представляли собой 26°C днем/22°C ночью при периоде день/ночь 12 ч./12 ч.
Анализы содержания хлорофилла. В конце периода обработки проводили анализы содержания хлорофилла в соответствии с протоколом, опубликованным в Protocol Exchange (Ni et al. (2009) doi:10.1038/nprot.2009.12) с использованием спектрофотометра DU® 640 (Beckman™ Coulter). Концентрации хлорофилла а (Ca), хлорофилла b (Cb) и хлорофилла а+b (Са+b) рассчитывали по следующим формулам (ArnonPlantPhysiol24:1-15(1949)), где V представляет собой объем экстракта (мл), а W представляет собой сырой вес листьев (г):
Ca (мг/г)=(12,7 × A663-2,69 × A645) × V/(1000 × W)
Cb (мг/г)=(22,9 × A645-4,68 × A663) × V/(1000 × W)
Са+b (мг/г)=(8,02 × A663+20,20 × A645) × V/(1000 × W)
Анализ сырого веса и получение образцов. После 16-дневного периода обработки измерения общей биомассы растений проводили путем срезания каждого растения у основания и регистрации их значений сырого веса. Непосредственно после взвешивания каждого образца у каждого растения отрезали один лист в аналогичном положении (4-й или 5-й лист от верхушки), его среднюю жилку удаляли, а остальную часть листовой пластинки замораживали в жидком азоте и хранили при -80°C для анализа содержания аминокислот. У каждого растения из аналогичного положения также собирали небольшой образец листа (~300 мг) для анализов содержания хлорофилла. Остальные листья на каждом растении впоследствии срывали с основного стебля, помещали в бумажный пакет и инкубировали в сушильном шкафу при 65°C в течение двух дней. Образцы высушенных листьев затем измельчали в тонкодисперсный порошок для анализов содержания нитратов и аммиака.
Анализы содержания нитратов, нитритов и аммиака. Для экспериментов в вегетационной камере примерно 100 мг порошка из высушенных листьев для каждого образца взвешивали и экстрагировали с использованием 10 мл деионизированной воды. После фильтрации через фильтровальную бумагу Whatman™ № 4 в образцах надосадочной жидкости анализировали концентрации нитратов и аммиака с использованием многоканального автоанализатора в лаборатории, оказывающей услуги по проведению испытаний на воздействие окружающей среды и сельскохозяйственных испытаний (EATS), Университета штата Северная Каролина. Значения количества нитратов и аммиака вычисляли в соответствии с протоколом из руководства Lachat Instruments.
Для образцов, выращенных в поле, и материалов T0, выращенных в теплице, измельченную ткань листьев анализировали на содержание нитратов в лаборатории анализа табака Университета Кентукки. Количественное определение нитратов проводили в соответствии с протоколом, изложенным в общих чертах Crutchfield и Grove (J. AOAC International 94:1898-1905 (2011)). Анализ содержания нитритов проводили в измельченных, высушенных на воздухе полевых образцах с помощью способа, описанного Crutchfield и Burton (Anal. Letters 22:555-571 (1989)).
Анализы содержания аминокислот. Образцы листьев растений, выращенных в вегетационной камере, которые были заморожены в жидком азоте, полностью высушивали путем помещения их в сублимационную сушилку на два дня. Затем высушенные образцы измельчали в жидком азоте в тонкодисперсный порошок с помощью ступки и пестика. Примерно 100 мг порошка из высушенных листьев взвешивали и отправляли в аналитическую лабораторию в Биопроизводственном центре обучения и подготовки при Университете штата Северная Каролина для измерений в рамках анализа содержания аминокислот. Вкратце, каждый образец экстрагировали с помощью 1,0 мл 80% MeOH-20% H2O и перемешивали вихревым способом с последующим центрифугированием при 20000 g. Аликвоты осветленной надосадочной жидкости объемом 200 мкл охлаждали при -80°C в течение примерно 2 ч., а затем лиофилизировали в течение ночи. Высушенный материал растворяли в 200 мкл боратного буфера (рН 8,0), затем снова разбавляли боратным буфером в соотношении 1:10. Образцы дериватизировали путем добавления 10 мкл экстрагированного образца к 70 мкл боратного буфера и 20 мкл реагента для дериватизации. Ампулы нагревали при 55°C в течение 10 минут, охлаждали до комнатной температуры, и их содержимое перемешивали вихревым способом в целях смешивания. Дериватизированные образцы анализировали с использованием сверхвысокоэффективной жидкостной хроматографии.
Анализ содержания алкалоидов и TSNA в листьях, высушенных на воздухе. Уровни никотина, норникотина, анатабина и анабазина в высушенных образцах листьев определяли количественно с использованием газового хроматографа Perkin-Elmer Autosystem XL в соответствии с ранее установленными протоколами (Jack et al. Rec. Adv. Tob. Sci. 33:58-79 (2007)). Общее содержание алкалоидов рассчитывали как сумму значений содержания никотина, норникотина, анатабина и анабазина. Процедуры количественного определения NNN, NNK, NAT и NAB проводили в соответствии со «способом 1» у Morgan et al. (Beit. Tabakforschung 23:192-203 (2004)). Общее содержание TSNA представляло собой сумму значений содержания NNN, NNK, NAT и NAB.
Экстракция и анализ TSNA в резаном табачном наполнителе и дыме. Образцы резаного табачного наполнителя экстрагировали 100 мМ раствором ацетата аммония, содержащим дейтерированные внутренние стандарты, обеспечивающие возможность количественного определения. Экстракты фильтровали с использованием шприцевого фильтра из PVDF (размер пор 0,45 мкм) и анализировали с помощью жидкостной хроматографии с тандемной масс-спектрометрией (LC-MS/MS) в сочетании с ионизацией электрораспылением (ESI). D4-N-нитрозонорникотин (D4-NNN) использовали в качестве внутреннего стандарта для NNN, тогда как D4-4-(метилнитрозамин)-1-(3-пиридил)-1-бутанон (D4-NNK) служил в качестве внутреннего стандарта для NNK, NAT и NAB.
Тестовые сигареты, изготовленные из резаного наполнителя, кондиционировали и выкуривали в соответствии с процедурой курения, указанной в способе № T-115, используемом Министерством здравоохранения Канады. Основной поток дыма собирали на фильтрах Cambridge и экстрагировали 100 мМ ацетатом аммония, содержащим стандарты D4-NNN и D4-NNK, с последующей фильтрацией с использованием шприцевого фильтра из PVDF с размером пор 0,45 мкм. Экстракты в ацетате аммония анализировали с помощью LC-MS/MS в сочетании с ESI и определяли количественно в соответствии со стандартами D4-NNN и D4-NNK.
Статистический анализ. Процедуру PROC GLM из SAS 9.1 (SAS Institute, Кэри, Северная Каролина) использовали для проведения дисперсионного анализа и расчета средних значений для каждой из включенных трансгенных линий и линий WT. Из-за неоднородности дисперсий проводили логарифмические преобразования данных с использованием натурального логарифма для аппроксимации данных сырого веса, содержания нитратов и аммиака, собранных в экспериментах в вегетационной камере, к нормальному распределению. Для всех других наборов данных использовали непреобразованные значения. Уровни значимости определяли в соответствии с множественным критерием размаха Райана-Эйнота-Габриэля-Уэлша (REGWQ).
Пример 2
Сверхэкспрессия генов пути N-ассимиляции у табака Берли
Ферменты GS1, NADH-GOGAT, ICDH или дерегулированная NR (мутация усечения 56 аминокислот либо точечная мутация S523D) сверхэкспрессировали в растениях табака Берли для определения того, обуславливала ли эктопическая экспрессия любого из этих генов пути N-ассимиляции 1) компенсацию дефицита хлорофилла и/или характерных для растений табака Берли дефектов роста, связанных с обработкой N-удобрениями, и/или 2) уменьшение уровней свободных нитратов, накапливаемых в листьях, и способствовала ли она реверсии фенотипа высокого уровня накопления нитратов, характерного для данного класса растений табака. При замене репортерного гена GUS в векторе экспрессии для растений pBI121 на гены пути N-ассимиляции каждая из этих конструкций была помещена под регулирующий контроль сильного конститутивного промотора 35S вируса мозаики цветной капусты (CaMV), а также 3'-концевой терминаторной последовательности nos гена нопалинсинтазы Agrobacterium tumefaciens (Chen et al. Mol. Breed. 11:287-293 (2003)). Схемы используемых конструкций показаны на фиг. 3A-3B.
Линию DH98-325-6#775 табака Берли трансформировали по отдельности каждой из пяти конструкций с использованием стандартных протоколов трансформации, опосредованной Agrobacterium. DH98-325-6#775 представляет собой результат отбора из популяции табака Берли, подвергнутой мутагенезу, индуцированному этилметансульфонатом (EMS), который содержал нокаут-мутацию в CYP83E4, основном гене никотиндеметилазы табака (Lewis et al. Phytochem. 71:1988-1998 (2010)). Растения табака Берли подвержены явлению, называемому конверсией никотина, при котором обычно неактивный ген CYP82E4 спонтанно активируется, что приводит к высокой доле потомства, которое накапливает крайне высокие уровни предшественника TSNA норникотина. Нокаут-мутация CYP82E4 в DH98-325-6#775 предотвращает конверсию никотина, и данную линию выбирали во избежание возможных осложнений, которые могут возникнуть при интерпретации данных о TSNA среди различных полученных трансгенных материалов, которые, вероятно, могли стать фактором в случае использования стандартного сорта Берли (из-за большой изменчивости уровней норникотина, которая, как можно было ожидать, наблюдалась бы при использовании нормального генетического окружения табака Берли).
Для обеспечения отбора растений, которые экспрессируют соответствующие трансгены на высоком уровне, по меньшей мере 10 отдельных растений T0 для каждой конструкции анализировали с помощью полуколичественной ОТ-ПЦР. Для каждой конструкции семена собирали из трех линий T0, которые демонстрировали наиболее высокие уровни накопления продуктов трансгенов. Из-за таких явлений, как сайленсинг трансгенов, или ситуаций, при которых трансген со слабой экспрессией и трансген с сильной экспрессией могут быть интегрированы в независимые локусы в одном и том же растении T0, возможно, что некоторые линии T0, которые демонстрируют высокую экспрессию трансгена, могут давать потомство, которое сегрегирует как по высокой, так и по низкой экспрессии трансгенов. Таким образом, полуколичественную ОТ-ПЦР проводили также для многочисленного потомства каждого из отобранных растений T0 с высокой экспрессией, чтобы проверить, продолжает ли потомство T1, которое унаследовало трансген(трансгены), экспрессировать их на высоком уровне. Все растения, используемые в последующих исследованиях, представляли собой растения T2, взятые из партий семян T1, где все потомство, для которого результат тестирования на наличие трансгена (посредством молекулярного генотипирования) был положительным, продолжало экспрессировать трансген на высоком уровне. Сегреганты T1, которые больше не имели трансген, использовали в качестве источника для контролей WT (ноль-сегрегантов).
Пример 3
Анализ трансгенных растений в вегетационной камере с контролируемой средой
Для изучения эффектов сверхэкспрессии генов пути N-ассимиляции в растениях Берли, выращиваемых в различных условиях доступности нитратов, три набора растений, содержащих шесть молодых растений T2 для каждого трансгенного генотипа (35S:tr-NR, 35S:S523D-NR, 35S:GS1, 35S:GOGAT и 35S:ICDH) и контроля WT, выращивали в вегетационной камере с контролируемой средой. Каждый набор растений поливали в течение 16 дней питательным раствором, содержащим 0,2 мМ, 8 мМ или 19 мМ нитратов. Однако, при выполнении эксперимента небольшое количество растений погибло или демонстрировало задержку роста и поэтому не было включено в анализ. В конце 16-дневного периода обработки между наборами растений, которые подвергали воздействию разных уровней нитратов, наблюдались сильные различия. Как и следовало ожидать, растения табака, которые поливали средой, содержащей 0,2 мМ нитратов, были хлоротическими и демонстрировали наименьшие темпы роста, тогда как растения, получавшие 19 мМ нитратов, были самыми большими и имели наиболее темный зеленый цвет. Однако в общем фенотипе роста растений, содержащих какой-либо специфический трансген, при любой указанной обработке N-удобрениями не наблюдались четкие различия по сравнению с контролями WT.
Измерения сырого веса надземных частей всех растений проводили в конце периода обработки, и их значения показаны в таблице 1 и на фиг. 4. Значения, показанные на фиг. 4, представляют собой непреобразованные средние значения ± стандартные ошибки для 4-6 растений по каждому генотипу. Статистические тестирования проводили с преобразованными данными (преобразование с использованием натурального логарифма). Для каждого уровня обработки нитратами средние значения в рамках одной и той же общей буквы значимо не отличались друг от друга (P < 0,5). Например, буква «a» применима только к растениям, обработанным 19 мМ нитратов, буквы «b» и «c» применимы только к растениям, поливаемым средой с 8 мМ нитратов, а «d» и «e» применимы только к растениям, обработанным 0,2 мМ нитратов. В таблице 1 показаны измеренные средние значения сырого веса, содержания хлорофилла, нитратов и аммиака для растений, экспрессирующих гены пути N-ассимиляции, выращиваемых при трех условиях обработки N-удобрениями, где «Ca» представляет собой хлорофилл а и «Cb» представляет собой хлорофилл b. Для растений, подвергаемых обработке нитратами на уровнях 8 мМ и 19 мМ, не наблюдались значимые различия между различными трансгенными генотипами по сравнению с контролем WT. Однако, при обработке с очень низким содержанием нитратов сырой вес растений, экспрессирующих конструкцию 35S:tr-NR, был незначительно большим, чем для WT, и это различие считалось статистически значимым.
Таблица 1
Растения табака Берли характеризуются дефицитом хлорофилла и четко отличаются от других типов табака по более светлому зеленому внешнему виду. Если какой-либо из сверхэкспрессируемых трансгенов был способен к компенсации мутаций yb1 и/или yb2, которые характеризуют типы Берли, то можно было ожидать увидеть увеличение содержания хлорофилла в данных растениях. Хотя между контролями WT и различными тестируемыми трансгенными генотипами не наблюдались какие-либо очевидные различия при простом визуальном наблюдении, в том случае, если присутствовали менее выраженные различия, проводили прямые измерения содержания хлорофилла a, b и a+b (таблица 1). Аналогично измерениям сырого веса, наблюдался очень сильный эффект обработки, при этом содержание хлорофилла положительно коррелировало с увеличением уровня обработки нитратными удобрениями. Однако, между любым трансгенным генотипом и контролями WT в рамках любого из трех типов обработки нитратами не наблюдались значимые различия в концентрациях хлорофилла (фиг. 5А-5С). Значения, показанные на фиг. 5A-5C, представляют собой средние значения ± стандартные ошибки для 4-6 растений по каждому генотипу. Для каждого уровня обработки нитратами средние значения в рамках одной и той же общей буквы значимо не отличались друг от друга (P < 0,5).
В таблице 1 также показаны средние концентрации нитратов и аммиака для различных генотипов на каждом уровне обработки нитратными удобрениями. При уровне обработки нитратами 0,2 мМ все растения демонстрировали крайне низкое накопление нитратов, и между любым трансгенным генотипом и контролями WT значимые различия не наблюдались (фиг. 6). На фиг. 6A показаны данные от растений, выращиваемых с применением обработок 19 мМ NO3-и 8 мМ NO3-; результаты для обработки 0,2 мМ NO3- показаны на фиг. 6B. Показанные значения представляют собой непреобразованные средние значения ± стандартные ошибки для 4-6 растений по каждому генотипу. Статистические тестирования проводили с преобразованными данными (преобразование с использованием натурального логарифма). Для каждого уровня обработки нитратами средние значения в рамках одной и той же общей буквы значимо не отличались (P < 0,5).
При обработках нитратами на уровнях 8 мМ и 19 мМ средняя концентрация нитратов, измеренная в растениях 35S:tr-NR, составляла соответственно лишь 37% и 60% от наблюдаемой у контролей WT. При обработке удобрениями с 8 мМ или 19 мМ нитратов происходило сильное снижение содержания нитратов, измеренное в листьях растений, экспрессирующих конструкцию 35S:S523D-NR. Растения, содержащие данную конструкцию, кодирующую дерегулированную NR, в среднем накапливали лишь приблизительно 5% от количества нитратов, присутствующего в контрольных растениях, при поливе в режиме обработки нитратными удобрениями на уровне 8 мМ. При применении обработки N-удобрениями на уровне 19 мМ растения WT в среднем содержали примерно 14900 ppm нитратов по сравнению со всего лишь ~2000 ppm, наблюдаемыми в растениях 35S:S523D-NR, что составляло почти 8-кратное уменьшение у трансгенных особей (фиг. 6А).
Концентрации аммиака в растениях, проанализированные в данном эксперименте, показаны на фиг. 7. Значения, показанные на фиг. 7, представляют собой непреобразованные средние значения ± стандартные ошибки для 4-6 растений по каждому генотипу. Статистические тестирования проводили с преобразованными данными (преобразование с использованием натурального логарифма). Для каждого уровня обработки нитратами средние значения в рамках одной и той же общей буквы значимо не отличались друг от друга (P < 0,5). Хотя уровни аммиака не изменялись столь сильно, как уровни нитратов, растения 35S:S523D-NR накапливали примерно вдвое больше данного метаболита пути N-ассимиляции по сравнению с контрольными растениями при уровне обработки нитратами 19 мМ. Однако, при низких и средних уровнях N-обработки ни один из указанных трансгенных генотипов статистически не отличался по его среднему содержанию аммиака от контролей WT или любого другого трансгенного генотипа.
Поскольку путь N-ассимиляции непосредственно приводит к биосинтезу аминокислот, то была установлена степень, в которой концентрации свободных аминокислот могут изменяться в трансгенных растениях. Независимо от трансгенного генотипа у растений, удобряемых средой с 0,2 мМ нитратов, наблюдались лишь минимальные различия в уровнях нитратов и аммиака (фиг. 6 и 7); таким образом, количественное определение содержания свободных аминокислот проводили только для растений, обработанных 8 мМ и 19 мМ нитратов. В таблице 2 показаны средние концентрации (мкмоль/г сухого веса) для всех 20 аминокислот по каждому трансгенному и контрольному генотипам, выращиваемым в условиях среднего (8 мМ) и высокого (19 мМ) уровня нитратов.
Таблица 2
Таблица 2, продолжение
Наблюдалось значительное увеличение общего содержания свободных аминокислот растениях, содержащих конструкцию 35S:S523D-NR, которые были обработаны 19 мМ нитратов (фиг. 8). Значения, показанные на фиг. 8, представляют собой средние значения ± стандартные ошибки для 4-6 растений по каждому генотипу. Для каждого уровня обработки нитратами средние значения в рамках одной и той же общей буквы значимо не отличались друг от друга (P < 0,5). Растения табака, экспрессирующие данную конструкцию, накапливали в среднем в 3,7 раза больше общего количества свободных аминокислот, чем растения WT. Хотя содержание практически каждой аминокислоты было выше в группе 35S:S523D-NR по сравнению с WT при высоком уровне обработки N, уровни Gln, Asn и Arg были особенно повышенными, демонстрируя увеличения соответственно в 8,5, 5,5 и 5,3 раза (фиг. 9). Значения, показанные на фиг. 9, представляют собой средние значения ± стандартные ошибки для 4-6 растений по каждому генотипу. Для каждого уровня обработки нитратами средние значения в рамках одной и той же общей буквы значимо не отличались друг от друга (P < 0,5). При выращивании с использованием питательного раствора с 8 мМ нитратов уровни Gln и Asn также были значительно более высокими в растении 35С:S523D-NR, чем у контроля (соответственно в 2 и 3,4 раза), однако значения общей концентрации аминокислот в двух линиях были практически идентичными (фиг. 8) из-за снижения средних уровней некоторых других аминокислот в растениях 35S:S523D-NR по сравнению с WT (таблица 2). В целом, аминокислотный профиль растений, экспрессирующих другую генетическую конструкцию, кодирующую дерегулированную NR (35S:tr-NR), был сходным с таковым у растений WT.
Глутамин является продуктом реакции, катализируемой ферментом GS. Поэтому интересно, что уровни Gln не были увеличены у растений со сверхэкспрессией гена GS (фиг. 9b). Это отличается от 2-кратного увеличения содержания Gln, наблюдаемого в случае, когда кДНК GS люцерны сверхэкспрессировалась в растениях табака, удобряемых раствором с высоким содержанием нитратов. Аналогично, уровни Glu не были значительно увеличены в трансгенных растениях со сверхэкспрессией гена, кодирующего фермент GOGAT, который непосредственно отвечает за его синтез (фиг. 9). При уровне обработки нитратными удобрениями 8 мМ растения со сверхэкспрессией ICDH демонстрировали более низкое общее содержание аминокислот (фиг. 8) и, в частности, более низкие количества Glu (фиг. 9D) по сравнению с каждым из других генотипов.
Дефицит хлорофилла представляет собой одну из характерных черт растений табака Берли и является признаком, проявление которого приписывают конкретно локусам yb1 и yb2. Все растения, независимо от того, выращивали ли их в вегетационной камере, теплице или полевых условиях, демонстрировали один и тот же светло-зеленый фенотип, который является отличительной чертой данного типа табака. В экспериментах в вегетационной камере проводили прямое измерение содержания хлорофилла в WT и трансгенных линиях, и различия между любыми из этих материалов не наблюдались. Такие результаты позволяют предположить, что сверхэкспрессия генов, связанных с путем N-ассимиляции (tr-NR, S523D-NR, GS1, GOGAT и ICDH) не дополняет или, иными словами, функционально не затрагивает мутации локусов yb1 и/или yb2, которые определяют фенотип Берли.
Хотя не было обнаружено никаких доказательств, что сверхэкспрессия любого из тестируемых трансгенов пути N-ассимиляции способна преодолевать дефекты роста или дефицит хлорофилла, связанные с обработкой N-удобрениями, которые характерны для растений табака Берли, сверхэкспрессия NR с дерегулированной активностью была явно эффективной в снижении уровней свободных нитратов
Пример 4
Анализ трансгенных растений, выращиваемых в полевых условиях
Конститутивная экспрессия конструкции, кодирующей дерегулированную S523D-NR, опосредовала сильное уменьшение накопления свободных нитратов в листьях при выращивании в условиях умеренного или высокого уровня обработки N-удобрениями. Также оказалось, что конструкция, кодирующая дерегулированную tr-NR, может обеспечивать незначительное уменьшение содержания нитратов. Для определения эффектов конструкций 35S:S523D-NR и 35S:tr-NR в отношении концентрации нитратов в листьях при выращивании в полевых условиях растения T2 из тех же партий семян, которые использовали в экспериментах в вегетационной камере, проращивали в подвесных ящиках и пересаживали в поле. Поскольку последствия избытка свободных нитратов в листе по отношению к образованию TSNA проявляются, прежде всего, во время старения и высушивания листа, конструкции, кодирующие дерегулированную NR, помещали под транскрипционный контроль сильного промотора, специфичного для старения. Промотор, контролирующий ген CYP82E4 никотиндеметилазы, специфично опосредует высокий уровень экспрессии гена во время старения и высушивания на воздухе. Конструкции, в которых кДНК S532D-NR и tr-NR помещены под контроль промотора CYP82E4 (E4), создавали путем первоначальной замены промотора 35S CaMV из вектора экспрессии для растений pBI121 на участок размером 2,2 т. п. о., расположенный непосредственно выше стартового кодона Met CYP82E4 для инициации трансляции, с последующей вставкой кДНК NR ниже промотора E4. В качестве контроля, представляющего собой сверхэкспрессию гена пути N-ассимиляции, который не влияет на содержание нитратов в листьях, в полевые эксперименты включали также трансгенные растения 35S:GOGAT и E4:GOGAT. Схемы этих векторов показаны на фиг. 3A-3B. Конкретные популяции T2, тестируемые в полевых условиях, которые имели конструкции под управлением промотора E4, были получены из партий семян растений T1, которые, как было показано, экспрессировали трансгены на высоком уровне при обработке соединением этефоном, индуцирующим старение (Chakrabarti et al., 2008). Аналогично исследованиям в вегетационной камере, ноль-сегреганты из растений T1, которые больше не имели трансген, использовали в качестве источника для контролей WT в полевых экспериментах.
Примерно 100 молодых растений для каждого из семи тестируемых генотипов (WT, 35S:S523D-NR, E4:S523D-NR, 35S:tr-NR, E4:tr-NR, 35S:GOGAT и E4:GOGAT) разделяли поровну и пересаживали в два местоположения в поле в Северной Каролине, используя рандомизированный полноблочный план. Растения выращивали на полевой научно-исследовательской станции вблизи города Клейтон, Северная Каролина, либо города Роки-Маунт, Северная Каролина. Поскольку значительное число растений погибло после пересадки, то общее число растений, которые выросли до зрелого состояния, находилось в диапазоне от 60 до 79 на генотип. Растения табака выращивали, у них отрезали верхушку, и их собирали в соответствии со стандартной отраслевой практикой, принятой для растений табака Берли. В зрелом состоянии собирали стебли растений, и с каждого растения срывали два листа в положении на стебле, находящемся приблизительно на расстоянии одной трети от верхушки, среднюю жилку удаляли, а остальную часть ткани листовой пластинки помещали в бумажный пакет. Затем растения подвешивали на прутьях и переносили для высушивания на воздухе в защищенную от дождя конструкцию. Материалы листьев в бумажных мешках помещали в сушильную печь и полностью высушивали при 65°C. Высушенные образцы листьев измельчали и анализировали на содержание нитратов.
Средняя концентрация нитратов в растениях T2, экспрессирующих различные гены, которые вовлечены в ассимиляцию нитратов, показана в таблице 3. Как показано в таблице 3, средние уровни нитратов, наблюдаемые для разных анализируемых генотипов, были весьма сходными между двумя местоположениями в поле (одно вблизи Клейтона, Северная Каролина, другое вблизи Роки-Маунта, Северная Каролина). Наиболее существенным наблюдением для этого набора данных было значительное уменьшение содержания нитратов в листьях растений 35S:S523D-NR. Растения табака, содержащие конструкцию 35S:S523D-NR, демонстрировали в среднем только 4,2% и 5,6% от количества нитратов, наблюдаемого в контролях WT в местоположениях Клейтона и Роки-Маунта соответственно.
Таблица 3
Статистический анализ объединенного набора данных для обоих местоположений показан в таблице 4. В таблице 4 показаны эффекты конструкций с генами, связанными с метаболизмом нитратов, в отношении содержания NO3- (N) (ppm в пересчете на сухой вес) в листьях трансгенных растений Берли поколения T2 в зрелом состоянии.Группа 35S:S523D-NR накапливала значительно меньше нитратов, чем любой другой генотип. Уровни свободных нитратов в растениях других генотипов, содержащих кДНК, кодирующую дерегулированную нитратредуктазу (35S:tr-NR, E4:tr-NR и E4:S532D-NR), также свидетельствовали о том, что они накапливали меньше нитратов, чем группа WT, хотя такое уменьшение является далеко не настолько сильным, однако только растения, содержащие конструкцию 35S:tr-NR, считались значимо отличающимися от контрольных генотипов, экспрессирующих GOGAT. Содержание нитратов в группах E4:GOGAT и 35S:GOGAT не считалось значимо отличающимся от WT.
Таблица 4
Растения, стебли которых собрали и которые переносили в хранилище для сушки, оставляли высушиваться на воздухе в течение примерно 10 недель. В конце данного периода собирали два дополнительных листа из положения в верхней трети стебля, у них отрывали среднюю жилку, и их помещали в бумажные мешки. Хотя материалы при сборе после высушивания были очень сухими, пакеты помещали в сушильное устройство при комнатной температуре с работающими вентиляторами для обеспечения полной сухости (тепло не добавляли из-за возможного влияния высокой температуры на образование TSNA). Высушенные образцы измельчали и подвергали анализу содержания нитратов. В таблице 5 показаны эффекты конструкций с генами, связанными с метаболизмом нитратов, в отношении содержания NO3- (N) (ppm в пересчете на сухой вес) в листьях трансгенных растений Берли поколения T2 после восьми недель высушивания на воздухе. Сравнение данных, содержащихся в таблице 4, с данными, представленными в таблице 5, показывает, что содержание нитратов в листовых пластинках для всех генотипов было в ~2 раза выше после высушивания на воздухе. После высушивания на воздухе в растениях табака Берли обычно наблюдается увеличение концентрации нитратов в листьях, хотя механизм данного явления неизвестен. Аналогично результатам, полученным с использованием зрелой невысушенной ткани, растения генотипа 35S:S523D-NR накапливали гораздо меньше свободных нитратов, чем любая другая генотипическая группа, и только ~4% от наблюдаемого у растений WT (таблица 5). Высушенные образцы листьев других генотипов, содержащие конструкции, кодирующие дерегулированную NR, также продолжали демонстрировать статистически значимое уменьшение содержания нитратов по сравнению с растениями WT, которое находилось в диапазоне от 70% до 77% от концентрации нитратов, наблюдаемой в группе WT.
Таблица 5
Хотя уровни нитритов в растениях, как правило, являются очень низкими, вероятно, благодаря эффективности видов активности эндогенной нитритредуктазы, интенсивный метаболизм клеточных запасов нитрата, опосредованный конститутивно экспрессируемым ферментом S523D-NR, может приводить к увеличению уровней его конечного продукта нитрита. С учетом того, что нитрит по сути представляет собой соединение, которое, как полагают, отвечает за образование TSNA во время высушивания на воздухе, важно, что генетические модификации, которые приводят к уменьшению пулов нитратов, не приводят к увеличению уровней нитритов в высушенных листьях. Чтобы определить, связана ли сверхэкспрессия какой-либо из трансгенных конструкций с изменениями содержания нитритов в листьях, проводили анализы содержания нитритов во всех собранных образцах листьев, высушенных на воздухе. В целом, уровни нитритов среди различных генотипов были сходными (таблица 6). В таблице 6 показаны эффекты конструкций с генами, связанными с метаболизмом нитратов, в отношении содержания NO2- (ppm в пересчете на сухой вес) в листьях трансгенных растений Берли поколения T2 после восьми недель высушивания на воздухе.
Таблица 6
Хотя среднее числовое значение для группы 35S:S523D-NR было самым низким из всех генотипов, оно не считалось статистически значимым по сравнению с контролями WT (хотя оно рассматривалось как значительно более низкое, чем для линии E4:GOGAT). Эти результаты демонстрируют, что конститутивная экспрессия конструкции, кодирующей дерегулированную S523D-NR, не приводит к увеличению уровня нитритов; наоборот, возможно, что данный трансген может опосредовать незначительное снижение концентраций нитритов в высушенных листьях.
Чтобы проверить, будет ли уменьшение содержания нитратов в листьях влиять на накопление TSNA или алкалоидов, подмножество вышеописанных высушенных образцов листьев генотипов WT, 35S:S523D-NR и 35S:GOGAT отобрали для дальнейшего анализа. Для каждого генотипа были выбраны в общей сложности 33 образца листьев, равномерно распределенных между двумя местоположениями (16 из Клейтона, 17 из Роки-Маунта) и отобранных из отдельных рядов, в которых были представлены все три генотипа (т. е. мертвые растения для любого из трех генотипов в пределах указанного выбранного ряда отсутствовали). Результаты анализов содержания TSNA и алкалоидов показаны в таблице 7. В таблице 7 показано содержание TSNA и алкалоидов в высушенных листьях растений WT 35S:GOGAT и 35S:S523D-NR. В совокупности общее уменьшение содержания TSNA у генотипа 35S:S523D-NR по сравнению с WT составляло 77,5%. Статистически значимые различия между содержанием любых отдельных TSNA или общим содержанием TSNA в растениях WT по сравнению с растениями 35S:GOGAT не наблюдались. Эти результаты четко демонстрируют, что конститутивная экспрессия конструкции S523D-NR приводит к существенному уменьшению содержания TSNA в высушенных на воздухе листьях табака, и это явление наиболее вероятно объясняется ее способностью к сильному уменьшению уровней свободных нитратов в листьях.
Таблица 7
Среди трех генотипов не наблюдались значимые различия ни в общем содержании алкалоидов, ни в содержании любого отдельного анализируемого алкалоида (никотина, норникотина, анабазина и анатабина). В противоположность этому, значительные различия в содержании TSNA были выявлены в материалах листьев 35S:S523D-NR с низким содержанием нитратов. В группе 35S:S523D-NR наблюдали 90% уменьшение уровней NNN по сравнению с контролями WT. Накопление N-нитрозоанатабина (NAT) и NNK в 35S:S523D-NR уменьшалось соответственно на 72,5% и 55% по сравнению с растениями WT. При том, что N-нитрозоанабазин (NAB), как правило, является наименее распространенным TSNA в высушенных листьях табака, в растениях 35S:S523D-NR присутствие данного соединения было почти невыявляемым (таблица 7).
Фенотип низкого содержания нитратов, опосредуемый конструкцией 35S:S523D-NR, был связан со значительным уменьшением содержания TSNA в высушенных листьях, согласующимся с моделью образования TSNA в растениях табака, высушенных на воздухе, в которой постулируется, что микробы, обитающие на поверхности листьев, используют пулы нитратов в листьях для выработки нитрита, который непосредственно отвечает за нитрозацию алкалоидов табака. Несмотря на то, что концентрации нитратов, алкалоидов и TSNA в растении табака существенно различались в зависимости от положения листа на стебле, конструкция 35S:S523D-NR была эффективной в опосредовании уменьшения содержания нитратов и TSNA во всем растении, поскольку значительное уменьшение содержания этих соединений наблюдалось как в выбранных листьях из верхних положений, так и в образцах резаного табачного наполнителя, полученных из остальной части растения с существенным смещением в сторону листьев из нижней части стебля. В противоположность этому, уровни никотина значительно не изменялись вследствие опосредованного 35S:S523D-NR уменьшения содержания нитратов.
Пример 5
Анализ резаного наполнителя и дыма сигарет, изготовленных из растений табака с низким содержанием нитратов
Для определения влияния фенотипа уменьшенного содержания нитратов в отношении распространенности TSNA в основном потоке дыма сигареты изготавливали из объединенных образцов оставшихся высушенных материалов листьев растений 35S:S523-NR и контролей WT. Первые два образца листьев, описанные в предыдущем разделе, брали из положения в верхней трети стебля; таким образом, в оставшейся ткани листьев, используемой для изготовления резаного наполнителя, наблюдалось существенное смещение в сторону листьев из нижних положений на стеблях растений. Поскольку относительные концентрации нитратов, никотина и TSNA различаются в зависимости от положения на стебле, то было важно независимо определить уровни этих соединений, представленных в резаном наполнителе, для того, чтобы точно интерпретировать соответствующие данные о дыме.
Как показано на фиг. 10, концентрация нитратов в резаном наполнителе, изготовленном из листовых пластинок WT, составляла в среднем 15800 ppm. В противоположность этому, в резаном наполнителе, полученном из растений 35S:S523D-NR, уровни нитратов составляли в среднем 930 ppm, и это значение составляло примерно 5,9% от наблюдаемого в наполнителе из WT, что согласовалось с соотношением, наблюдаемым для этих двух генотипов в высушенных листьях из верхней части стебля (таблица 5). Более высокие общие концентрации нитратов, измеренные в резаном наполнителе из растений WT и 35S:S523D-NR, по сравнению с показателями верхних листьев (сравните таблицу 5 и фиг. 10), согласовались с предыдущими наблюдениями того, что уровни нитратов в нижних листьях табака являются гораздо более высокими, чем обнаруживаемые в верхних листьях. На фиг. 10 все растительные материалы каждого генотипа объединяли в соответствии с местоположением (приблизительно по 30 растений 35S:S523D-NR для каждого из Клейтона и Роки-Маунта; приблизительно по 40 растений WT для каждого из двух местоположений). Показанные данные представляют собой среднее значение и стандартную ошибку для обоих местоположений. Концентрация никотина в листьях табака отображала противоположную тенденцию, при этом верхние листья демонстрировали более высокие концентрации, чем нижние листья, хотя величина разницы не была столь чрезвычайной, как наблюдалось для нитратов. Содержание никотина в резаном наполнителе из растений WT и 35S:S523D-NR составляло в среднем соответственно 1,6% и 1,7% в пересчете на сухой вес (фиг. 10) по сравнению с 2,5% и 2,4% в их соответствующих верхних листьях (таблица 7). В любом случае, несмотря на огромное влияние конструкции S523D-NR на содержание нитратов в листьях, ее конститутивная экспрессия значительно не изменяла уровни никотина в листьях.
Результаты анализов содержания TSNA в резаном табачном наполнителе показаны на фиг. 11A-11E. Аналогично наблюдениям при использовании высушенных материалов верхних листьев (таблица 7), наблюдалось значительное уменьшение содержания каждого из четырех видов TSNA при использовании материала листьев растений с низким содержанием нитратов, хотя наблюдались некоторые различия в относительной степени соответствующих уменьшений. Резаный наполнитель, полученный из растений 35S:S523D-NR, содержал соответственно на 44%, 64%, 65% и 32% меньше NNN, NNK, NAT и NAB, чем наполнитель, изготовленный из контрольных растений WT, при наблюдаемом уменьшении общего содержания TSNA, составляющем 52%. Общие уровни TSNA были несколько более высокими в материале резаного наполнителя по сравнению с высушенными верхними листьями (сравните таблицу 7 и фиг. 11), и этот результат согласуется с предыдущими наблюдениями того, что накопление TSNA является более высоким в листьях из нижних положений на стеблях, чем в листьях из верхних положений. На фиг. 11 материалы листьев каждого генотипа были объединены в соответствии с каждым из двух местоположений, как описано на фиг. 10. Показанные данные представляют собой совокупные средние значения и стандартные ошибки для двух повторностей на местоположение для содержания TSNA в резаном наполнителе и трех повторностей на местоположение для содержания TSNA в основном потоке дыма.
Данные о TSNA в основном потоке дыма, полученном из сигарет, изготовленных из резаного наполнителя из WT и 35S:S523D-NR, также показаны на фиг. 11. На удивление, уменьшение общего количества TSNA, свойственное фенотипу низкого содержания нитратов, в основном потоке дыма является даже большим, чем наблюдаемое в соответствующем резаном наполнителе. В основном потоке дыма сигарет из 35С:S523D-NR демонстрировалось уменьшение общего содержания TSNA на 76% по сравнению с сигаретами из WT, в отличие от уменьшения на 52%, наблюдаемого в резаном табачном наполнителе из них (фиг. 11E). Различия в степени уменьшения содержания TSNA между резаными табачными наполнителями из 35S:S523D-NR и WT по сравнению с их соответствующим основным потоком дыма наиболее ярко выражены для NNN и NAB. Содержание NNN в несгоревшем табачном наполнителе из растений 35S:S523D-NR было на 44% меньшим, чем в наполнителе из WT, хотя разница для основного потока дыма увеличивалась до 78% (фиг. 11D). Аналогично, уровни NAB в наполнителе из растений 35S:S523D-NR были на 32% более низкими, чем для WT, по сравнению с 86% уменьшением в основном потоке дыма (фиг. 11А). Из четырех видов TSNA с измеренным содержанием только NNK не продемонстрировал статистически значимой разницы в относительном уменьшении его содержания в резаном наполнителе по сравнению с основным потоком дыма при сравнении результатов сигарет из WT и 35S:S523D-NR. Как для резаного наполнителя, так и для основного потока дыма содержание NNK было уменьшено примерно на 67% в сигаретах, изготовленных с использованием табака из 35С:S523D-NR с низким содержанием нитратов, по сравнению с контрольным табаком WT (фиг. 11C). Относительное уменьшение уровней всех видов TSNA, кроме NNK, в основном потоке дыма сигарет из табака с низким содержанием нитратов было даже большим, чем в соответствующем несгоревшем резаном наполнителе.
Уменьшение уровней нитратов, опосредованное 35S:S523D-NR, может смягчать проблему наличия TSNA в табачных продуктах на двух уровнях: (1) за счет уменьшения выработки TSNA как таковых в высушенных листьях и (2) за счет дополнительного ингибирования процессов, посредством которых TSNA появляются в основном потоке дыма.
Пример 6
Сверхэкспрессия S523D-NR в дополнительном генетическом окружении табака Берли приводит к сильному уменьшению накопления нитратов в листьях
Векторы экспрессии для растений, содержащие конструкции 35S:S523D-NR и E4:S523D-NR, использовали для трансформации сорта Берли TN90e4e5. В дополнение к созданию растений, содержащих конструкции 35S:S523D-NR и E4:S523D-NR по отдельности, обе конструкции объединяли в одном и том же растении для определения того, можно ли добиться дополнительного уменьшения содержания нитратов вдобавок к тому, которое достигается с помощью одной конструкции. Для получения растений T0 с каждым трансгеном и комбинацией трансгенов использовали стратегию котрансформации. Для того, чтобы различить набор трансгенов в каждом полученном растении T0, проводили генотипирование на основе ПЦР с использованием 5'-праймера, специфичного для промоторных участков 35S либо E4, в паре с 3'-праймером, направленным на участок S523D-NR конструкции. Листовые диски TN90e4e5 также трансформировали конструкцией, содержащей репортерный ген GUS под транскрипционным контролем промотора E4, в качестве контрольного вектора.
После того, как растения становились достаточно большими, чтобы перенести их в почву, следующие линии T0 выращивали до зрелого состояния в теплице: E4:GUS с контрольным вектором (12 трансгенных объектов); 35S:S523D-NR (10 трансгенных объектов); E4:S523D-NR (11 трансгенных объектов) и 35S:S523D-NR+E4:S523D-NR (7 трансгенных объектов). Растения выращивали в стандартной почвенной смеси, удобряемой Multicote 4, удобрением с контролируемым высвобождением (NPK 14-14-16+микроэлементы). Непосредственно перед началом цветения два листа из средне-верхней части каждого растения собирали, средние жилки удаляли, а остальную часть листовой пластинки высушивали при 65°C. В ту же дату сбора брали два дополнительных листа из той же части каждого растения и обрабатывали этефоном для индукции старения, как описано Jack et al. (Rec. Adv. Tob. Sci. 33:58-79 (2007)). После того, как обработанные этефоном листья полностью пожелтели (7-8 дней после обработки), часть каждого листа замораживали для анализа по методу ПЦР в реальном времени (ОТ-ПЦР), тогда как остальную часть ткани листа оставляли до полного высушивания для последующего анализа содержания нитратов.
В таблице 8 показано содержание нитратов в зеленых и «искусственно состаренных» образцах листьев растений T0 (т. е. растений T0 TN90e4e5). Все 12 независимых растений с контрольным вектором E4:GUS демонстрировали высокие уровни нитратов в своих листьях в диапазоне от 3938 до 12730 частей на миллион (ppm). В целом, концентрация нитратов в зеленых листьях была весьма сходной с наблюдаемой в листьях, обработанных этефоном. В отличие от контролей E4:ВСО, пять из десяти независимых трансгенных объектов T0 35S:S523D-NR накапливали менее 1000 ppm, причем три индивидуума накапливали 300 ppm или менее как в зеленых, так и в обработанных этефоном образцах. Все 11 индивидуумов E4:S523D-NR продемонстрировали высокое накопление нитратов в зеленых листьях (более 3987 ppm).
Таблица 8
В обработанных этефоном листьях, полученных из одних и тех же растений, наблюдалось, однако, уменьшение содержания нитратов в 2-3 раза (таблица 8). Ожидается, что промотор E4, который контролирует экспрессию S523D-NR в этих растениях, является неактивным в зеленых листьях, однако становится экспрессируемым на высоком уровне во время обработки этефоном, старения и высушивания на воздухе. Два из семи растений T0, содержащих конструкции как 35S:S523D-NR, так и E4:S523D-NR, продемонстрировали очень низкие уровни нитратов в зеленых листьях (146 и 238 ppm). Низкие уровни нитратов в зеленой ткани данных растений позволяют предположить, что конструкция 35S:S523D-NR активно экспрессируется у этих двух индивидуумов. В одном из этих растений (35S+E4:S523D-NR 5-2) уровни нитратов в зеленых листьях по сравнению с листьями, обработанными этефоном, оставались практически одинаковыми. Однако, в другом растении (35S+E4:S523D-NR 8-1) свободные нитраты в листьях, обработанных этефоном, не выявлялись. Максимального уменьшения накопления нитратов можно достичь путем объединения конструкции, управляющей сильной конститутивной экспрессией S523D-NR, с конструкцией, опосредующей сильную специфичную для старения/высушивания экспрессию S523D-NR. Примечательно, что очевидные фенотипические различия между растениями T0, характеризующимися крайне низкими уровнями свободных нитратов, и индивидуумами T0, демонстрирующими нормальные уровни нитратов, не наблюдались.
Из-за таких явлений, как «эффекты положения» и сайленсинг генов, в любом указанном эксперименте по трансформации обычно наблюдают большую изменчивость в уровнях накопления транскриптов среди различных трансгенных объектов T0. Результаты анализов содержания нитратов в растениях T0 позволяли предположить, что трансген S523D-NR эффективно экспрессировался у некоторых индивидуумов, но этого не происходило не у остальных (в частности, применительно к конструкции 35S:S523D-NR). Чтобы это проверить, проводили анализ по методу ОТ-ПЦР в вышеописанных пожелтевших образцах, обработанных этефоном. Результаты анализа по методу ОТ-ПЦР с использованием тканей зеленых листьев было трудно интерпретировать в силу того, что анализ не обеспечивал установление различий между эндогенными транскриптами Nia2 и транскриптами, полученными из S523D-NR (так как их кодирующие участки отличаются только по одному кодону), и в ткани зеленых листьев контрольных растений выявляли высокий уровень нативного NR-транскрипта. В отличие от этого, в искусственно состаренных листьях, обработанных этефоном, выявляли минимальное накопление эндогенного NR-транскрипта. На фиг. 12 показаны относительные уровни NR-транскриптов (нормализованные по гену фактора элонгации 1α табака) в листьях, обработанных этефоном, каждого растения T0, приведенного в таблице 8, вместе с соответствующей концентрацией нитратов. На фиг. 12 данные ОТ-ПЦР (желтые полосы, шкала слева) демонстрируют относительное накопление NR-транскриптов (как эндогенной NR, так и S523D-NR) после нормализации по эталонному транскрипту фактору элонгации 1α табака. Красные прямоугольники обозначают концентрацию нитратов (шкала справа). Во всех растениях с контрольным вектором накопление эндогенного NR-транскрипта является относительно низким, а содержание нитратов является высоким (более 5000 частей на миллион). В растениях T0, содержащих только конструкцию 35С:S523D-NR, наблюдается очень хорошая корреляция между высоким накоплением NR-транскрипта и фенотипом низкого содержания нитратов. Все растения T0, имеющие только трансген E4:S523D-NR, демонстрировали более высокий уровень накопления NR-транскрипта, чем растения с контрольным вектором, и их соответствующие уровни нитратов также были единообразно более низкими. Неудивительно, что уровни нитратов не уменьшаются столь сильно в растениях E4:S523D-NR по сравнению с растениями с высокой экспрессией 35S:S523D-NR, с учетом того, что дерегулированный фермент NR, как ожидается, будет способствовать уменьшению пулов нитратов только после обработки этефоном в первой группе растений, а не на протяжении всей жизни растения, как можно было бы ожидать для второй группы индивидуумов. Значительная доля NR-транскриптов в растениях 5-2 и 8-1 в данной группе может проистекать из конструкции 35S:S523D-NR с учетом крайне низких уровней нитратов, наблюдаемых в образцах как зеленых, так и обработанных этефоном листьев этих растений (таблица 8).
Пример 7
При сверхэкспрессии S523D-NR в растениях T1 демонстрируется сильное уменьшение накопления нитратов в листьях
Трансгенные растения T1 в генетическом окружении TN90e4e5 (в том числе контроли, 1920 растений) из подвесных ящиков генотипировали и пересаживали в два местоположения в поле (полностью рандомизированный план полевого эксперимента). Образцы тканей листьев (листовые пластинки) с каждого растения собирали для анализа содержания нитратов, и затем растения помещали в устройство для воздушной сушки с образованием резаного наполнителя для анализов сигаретного дыма.
Данные по содержанию нитратов в зеленых листьях, собранных из среднего положения на стебле в двух местоположениях на поле, представлены в таблицах 9 и 10. В обоих случаях данные продемонстрировали существенное уменьшение содержания нитратов, обусловленное присутствием трансгена.
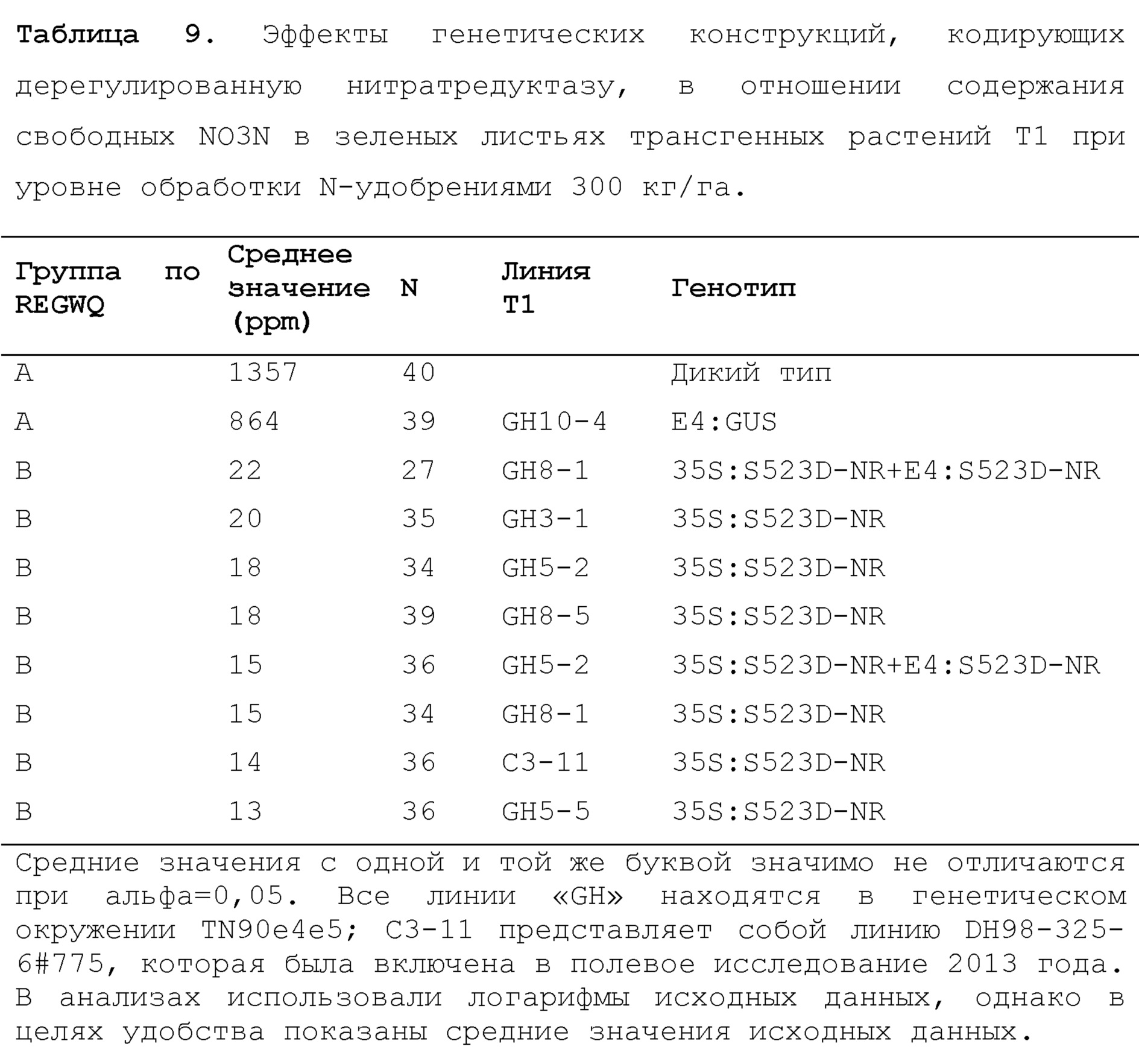
Из этого эксперимента можно сделать вывод, что экспрессия трансгена 35S:S523D-NR в более активном генетическом окружении TN90e4e5 является столь же эффективной в снижении содержания нитратов в листьях, как это наблюдалось в линии DH98-325-6#775 с худшими характеристиками.
Некоторые линии (GH3-1, GH8-1 и GH8-5) отбирали для анализа содержания нитратов и общего содержания алкалоидов в высушенных листьях (среднее положение на стебле, образцы сыпучего порошка, типичные для опытных участков с растениями). В целом, как это наблюдалось ранее, общее содержание алкалоидов и никотина не зависело от присутствия трансгена (35S:S523D-NR). Однако, в генетическом окружении TN90e4e5 наблюдалось незначительное уменьшение (15%) в одной линии GH8-5 (фиг. 13).
Как и следовало ожидать из предыдущих результатов, содержание нитратов в листьях существенно уменьшается (> 95%) в листовых пластинках за счет более активной ассимиляции нитратов. Добавление трансгена, который регулируется промотором E4, специфичным для старения, не влияет на данные, что таким образом позволяет предположить, что активность трансгенной нитратредуктазы под контролем конститутивного промотора 35S является весьма эффективной в зеленых листьях (см. таблицы 9 и 10) и достаточной для уменьшения содержания нитратов, запасаемых в листовых пластинках (фиг. 14).
Резаный наполнитель получали из высушенных листьев и анализировали на содержание TSNA. Интересно отметить, что содержание TSNA заметно уменьшалось (~72% для NNN, ~45% для NNK, ~76% для NAT и ~78% для NAB), что свидетельствует о том, что снижение содержания нитратов в зеленых листьях существенно влияет на выработку TSNA в материале резаного наполнителя (фиг. 15).
Точно так же как для линии DH98-325-6#775 с худшими характеристиками, трансформированной 35S:S523D-NR (см. пример 2), сигареты изготавливали из материала, анализируемого на фиг. 15, и выкуривали. Определяли содержание TSNA в дыме, и данные приводили на фиг. 16. Содержание TSNA в дыме уменьшалось в еще большей степени (~90% для NNN, ~66% для NNK, ~88% для NAT и ~92% для NAB), чем в резаном наполнителе, что, несомненно, обусловлено влиянием снижения нитрозации во время пиролиза. Таким образом, в целом это указывает на то, что благодаря снижению содержания нитратов в зеленых листьях с помощью сверхэкспрессии дерегулированной формы нитратредуктазы существенно уменьшается выработка TSNA в дыме коммерческого табака, такого как TN90e4e5.
Следует понимать, что приведенное выше подробное описание и сопроводительные примеры являются только иллюстративными и не должны рассматриваться как ограничивающие объем настоящего изобретения, который определяется исключительно прилагаемой формулой изобретения и ее эквивалентами.
Различные изменения и модификации раскрытых вариантов осуществления будут очевидны специалистам в данной области. Такие изменения и модификации, в том числе, без ограничения, относящиеся к химическим структурам, заместителям, производным, промежуточным продуктам, синтезам, композициям, составам или способам применения настоящего изобретения могут быть сделаны без отступления от его сущности и объема.
Для полноты картины различные аспекты настоящего изобретения изложены в следующих пронумерованных пунктах.
Пункт 1. Табачный продукт, характеризующийся уменьшенными уровнями табак-специфичных нитрозаминов (TSNA), получаемый из растения табака, при этом указанное растение табака модифицировано таким образом, что оно содержит: (i) полинуклеотид, содержащий последовательность, кодирующую дерегулированный фермент нитратредуктазу, состоящий из нее или по сути состоящий из нее; (ii) полипептид, кодируемый полинуклеотидом, изложенным в (i); (iii) полипептид, содержащий дерегулированный фермент нитратредуктазу, состоящий из него или по сути состоящий из него; или (iv) конструкцию, вектор или вектор экспрессии, содержащие полинуклеотид, изложенный в (i), и где экспрессия или активность указанной нитратредуктазы является дерегулированной по сравнению с контрольным немодифицированным растением табака.
Пункт 2. Табачный продукт по пункту 1, где дерегулированный фермент нитратредуктаза представляет собой фермент нитратредуктазу, который является конститутивно активным.
Пункт 3. Табачный продукт по пункту 1 или пункту 2, где последовательность, кодирующая дерегулированный фермент нитратредуктазу, кодирует (i) усеченную полипептидную нитратредуктазу; (ii) полипептидную нитратредуктазу, содержащую N-концевое усечение; (iii) полипептидную нитратредуктазу, содержащую N-концевое усечение 56 аминокислот; (iv) полипептидную нитратредуктазу, содержащую аминокислотную замену в положении, соответствующем положению 523 в SEQ ID NO: 4, или (v) полипептидную нитратредуктазу, содержащую аминокислотную замену в положении, соответствующем положению 523 в SEQ ID NO: 4, где аминокислота в положении 523 SEQ ID NO: 4 заменена аспарагиновой кислотой.
Пункт 4. Табачный продукт по любому из предыдущих пунктов, где полинуклеотид, кодирующий дерегулированную нитратредуктазу, представляет собой гетерологичный полинуклеотид, кодирующий модифицированную полипептидную нитратредуктазу.
Пункт 5. Табачный продукт по пункту 4, где гетерологичный полинуклеотид связан с промотором, в нативных условиях не связанным с эндогенным геном нитратредуктазы.
Пункт 6. Табачный продукт по пункту 5, где промотор представляет собой промотор 35S вируса мозаики цветной капусты или промотор CYP82E4.
Пункт 7. Табачный продукт по любому из предыдущих пунктов, где полинуклеотид, кодирующий дерегулированную нитратредуктазу, содержит полинуклеотидную последовательность SEQ ID NO: 5 или SEQ ID NO: 7.
Пункт 8. Табачный продукт по любому из пунктов 1-3, где полинуклеотид, кодирующий дерегулированную нитратредуктазу, представляет собой эндогенный ген нитратредуктазы, который был модифицирован с помощью системы редактирования генома или с помощью мутагена.
Пункт 9. Табачный продукт по пункту 8, где система редактирования генома включает в себя сконструированную систему на основе CRISPR/Cas, сконструированную эффекторную нуклеазу, подобную активаторам транскрипции, сконструированную нуклеазу с «цинковыми пальцами» или сконструированную мегануклеазу.
Пункт 10. Табачный продукт по любому из предыдущих пунктов, где табак представляет собой табак Берли, восточный табак, темный табак, табак сигарного типа и табак трубоогневой сушки.
Пункт 11. Табачный продукт по любому из предыдущих пунктов, характеризующийся уменьшенными уровнями табак-специфичных нитрозаминов (TSNA) по сравнению с табачным продуктом, полученным из контрольного растения табака, в котором фермент нитратредуктаза не был дерегулирован.
Пункт 12. Табачный продукт по пункту 11, где общий уровень TSNA измеряется в листе растения табака, где (a) лист является свежесобранным; (b) лист является высушенным, хранящимся или переработанным; или (c) лист является высушенным на воздухе.
Пункт 13. Табачный продукт по пункту 11 или пункту 12, где уровень по меньшей мере одного TSNA в табачном продукте уменьшен по сравнению с контрольным уровнем по меньшей мере одного TSNA, где по меньшей мере один TSNA выбран из группы, включающей N-нитрозонорникотин (NNN), 4-(метилнитрозамино)-1-(3-пиридил)-1-бутанон (NNK), N-нитрозоанабазин (NAB), N-нитрозоанатабин (NAT) и их комбинации.
Пункт 14. Табачный продукт по любому из пунктов 11-13, где общие уровни TSNA уменьшены по меньшей мере на 50%.
Пункт 15. Табачный продукт по пункту 11, где уровни TSNA измеряются в дыме, получаемом при сгорании листьев растения табака.
Пункт 16. Табачный продукт по пункту 15, где общие уровни TSNA в дыме уменьшены по меньшей мере на 70%.
Пункт 17. Способ получения табачного продукта, в котором уровни TSNA, измеряемые в дыме, получаемом при сгорании листьев модифицированного растения табака, уменьшены по сравнению с уровнями TSNA, измеряемыми в дыме, получаемом при сгорании немодифицированного растения табака, включающий: (a) модификацию растения табака таким образом, чтобы оно содержало: (i) полинуклеотид, содержащий последовательность, кодирующую дерегулированный фермент нитратредуктазу, состоящий из нее или по сути состоящий из нее; (ii) полипептид, кодируемый полинуклеотидом, изложенным в (i); (iii) полипептид, содержащий дерегулированный фермент нитратредуктазу, состоящий из него или по сути состоящий из него; или (iv) конструкцию, вектор или вектор экспрессии, содержащие полинуклеотид, изложенный в (i), и где экспрессия или активность указанной нитратредуктазы является дерегулированной по сравнению с контрольным немодифицированным растением табака; (b) сбор листьев табака с указанного модифицированного растения табака и (c) получение табачного продукта из собранных листьев.
Пункт 18. Табачный продукт по любому из предыдущих пунктов, где растение табака дополнительно содержит модифицированный ген пути синтеза норникотина.
Пункт 19. Табачный продукт по пункту 18, где модифицированный ген пути синтеза норникотина включает в себя модифицированный ген никотиндеметилазы или ген цитохрома P450.
Пункт 20. Табачный продукт по пункту 18 или пункту 19, где растение табака содержит модифицированный ген CYP82E4 или модифицированный ген CYP82E10.
Пункт 21. Табачный продукт по пункту 20, где модифицированный ген CYP82E4 или модифицированный ген CYP82E10 является инактивированным.
Пункт 22. Табачный продукт по любому из предыдущих пунктов, где растение табака представляет собой разновидность, выбранную из группы, включающей табак Берли, восточный табак, темный табак, табак сигарного типа и табак трубоогневой сушки.
Пункт 23. Табачный продукт по любому из предыдущих пунктов, где табачный продукт представляет собой курительный материал, нюхательный табак, жевательный табак, жевательную резинку, леденец или раствор никотина.
Пункт 24. Способ получения табачного продукта по любому из пунктов 1-23, при этом способ включает: (a) введение в растительную клетку выделенного полинуклеотида, кодирующего модифицированный ген нитратредуктазы или модифицированный ген переносчика нитрата; (b) регенерацию трансформированной клетки с получением трансгенного растения и (c) получение табачного продукта из листа трансгенного растения.
Пункт 25. Способ получения табачного продукта по любому из пунктов 1-23, при этом способ включает: (a) введение в растительную клетку системы редактирования генома, которая целенаправленно воздействует на эндогенный ген нитратредуктазы; (b) регенерацию трансформированной клетки с получением трансгенного растения и (c) получение табачного продукта из листьев трансгенного растения.
Пункт 26. Способ по пункту 25, где система редактирования генома связывается с эндогенным геном нитратредуктазы и расщепляет его с образованием модифицированного гена нитратредуктазы.
Пункт 27. Способ по пункту 25 или пункту 26, где система редактирования генома включает в себя сконструированную систему на основе CRISPR/Cas9, сконструированную эффекторную нуклеазу, подобную активаторам транскрипции, сконструированную нуклеазу с «цинковыми пальцами» или сконструированную мегануклеазу.
Пункт 28. Способ по любому из пунктов 25-27, где система редактирования генома транзиентно экспрессируется в растении табака.
Пункт 29. Способ по любому из пунктов 25-27, где система редактирования генома включена в геном растения табака.
Пункт 30. Способ по любому из пунктов 25-29, где растение табака дополнительно содержит модифицированный ген пути синтеза норникотина.
Пункт 31. Способ по пункту 30, где модифицированный ген пути синтеза норникотина включает в себя модифицированный ген никотиндеметилазы или ген цитохрома P450.
Пункт 32. Способ по пункту 30 или пункту 31, где растение табака содержит модифицированный ген CYP82E4 или модифицированный ген CYP82E10.
Пункт 33. Способ по пункту 32, где модифицированный ген CYP82E4 или модифицированный ген CYP82E10 является инактивированным.
Пункт 34. Способ по любому из пунктов 25-33, где растительная клетка представляет собой клетку разновидности табака, выбранной из группы, включающей табак Берли, восточный табак, темный табак, табак сигарного типа и табак трубоогневой сушки.
Пункт 35. Способ по любому из пунктов 25-34, где табачный продукт представляет собой курительный материал, нюхательный табак, жевательный табак, жевательную резинку, леденец или раствор никотина.
Следующие последовательности нуклеиновых кислот и аминокислотные последовательности раскрыты в перечне последовательностей, прилагающемся к данному документу.
SEQ ID NO: 1 - кодирующий участок кДНК Nia1
SEQ ID NO: 2 - предсказанная последовательность белка Nia1p
SEQ ID NO: 3 - кодирующий участок кДНК Nia2 (содержит остаток Ser в положении 523)
SEQ ID NO: 4 - предсказанная последовательность белка Nia2p
SEQ ID NO: 5 - кодирующий участок кДНК Nia2 S523D-NR
SEQ ID NO: 6 - предсказанная последовательность белка S523D-NRp (содержит остаток Asp в положении 523)
SEQ ID NO: 7 - кодирующий участок кДНК tr-NR
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Университет штата Северная Каролина
Филип Моррис Продактс С.А.
<120> УМЕНЬШЕНИЕ СОДЕРЖАНИЯ ТАБАК-СПЕЦИФИЧНЫХ НИТРОЗАМИНОВ ПОСРЕДСТВОМ
ИЗМЕНЕНИЯ ПУТИ АССИМИЛЯЦИИ НИТРАТОВ
<130> P4597PC00
<150> EP 14194798.6
<151> 2014-11-25
<150> EP 14186705.1
<151> 2014-09-26
<160> 28
<170> PatentIn, версия 3.5
<210> 1
<211> 2715
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 1
atggcggcat ctgtcgaaaa caggcagttc agtcacatag aagccggttt atcccggtct 60
ttcaagcctc ggtctgattc cccggttcgt ggctgcaact tccctccgcc caacagtact 120
aatttccaaa agaaaccaaa ttccaccatt ttccttgatt actcgtcgag tgaagacgac 180
gatgatgatg acgaaaaaaa tgagtacctt caaatgatca aaaaagggaa ttcagaatta 240
gagccatctg ttcatgacag cagggacgaa ggtaccgctg ataactggat tgaacgcaac 300
ttttccttga ttcgtctcac cggaaagcat ccatttaact ccgaaccgcc gttgaaccgt 360
ctcatgcacc acggttttat cacaccggtc ccacttcatt acgttcgtaa ccatggaccg 420
gttcccaagg gcacatggga tgactggacc gtggaagtca cgggactagt gaaacgtcct 480
atgaaattca caatggacca gttggttaac gaattccctt ccagagaatt gcccgttacg 540
cttgtgtgtg ctggcaaccg aaggaaagaa cagaacatgg ttaaacaaac cattggtttc 600
aactggggtg ccgctgccgt ttcaacaact gtatggcgcg gggtacccct acgcgctttg 660
ttaaaacggt acggtgtttt tagcaagaat aaaggggcgc ttaatgtttg cttcgaagga 720
gctgatgtct tgcccggagg cggtggttca aagtatggaa ccagcattaa gaaggaattt 780
gcaatggatc cagcacgaga tatcataata gcttacatgc agaacggaga aaaattggca 840
cccgaccacg ggtttccagt acgaatgata attccaggat tcattggagg aagaatggtg 900
aaatggataa agaggattat agtcaccacc caagaatcag acagctatta tcatttcaag 960
gacaatagag ttcttcctcc ccatgttgat gctgaacttg caaatactga agcatggtgg 1020
tacaagccag agtacatcat caatgagctc aatattaact ctgtcattac gacgccgtgt 1080
catgaagaaa ttttgcctat taacgcctgg acgactcagc gaccttacac gttgaggggc 1140
tattcttatt ctggcggagg gaaaaaagta acgcgagtag aagtgacctt ggatggagga 1200
gaaacatggc aagtttgcac actagatcac ccagagaagc ccaccaaata tggcaagtac 1260
tggtgttggt gcttttggtc actcgaggtt gaggtgttag acttgctcag tgccaaagaa 1320
attgctgttc gagcttggga tgagaccctc aatactcaac ctgagaagct tatttggaat 1380
gtcatgggaa tgatgaacaa ttgctggttc cgagtaaaga tgaatgtgtg caagcctcac 1440
aagggagaga ttggaatagt gtttgaacac ccgactcaac ctggaaacca atcaggtgga 1500
tggatggcaa aggagaggca tttggagata tcagcagagg cacctccaac actaaagaag 1560
agtatctcaa ctccattcat gaacacagct tccaagatgt actccatgtc ggaggtgagg 1620
aaacacagct ctgctgactc tgcttggatc atagtccatg gtcatatcta tgacgccacg 1680
cgtttcttga aagatcaccc cggtggttct gacagcattc tcatcaatgc tggcactgat 1740
tgcactgagg aatttgatgc aattcattct gataaggcta agaagctatt ggaggaattc 1800
aggattggtg aactcctaac tactggttac acctctgact ctcctggcaa ctccgtccat 1860
ggatcttctt ccttcagcag ctttctagca cctattaagg aacttgttcc agcgcagagg 1920
agtgtggccc tcattccaag agagaaaatc ccatgcaaac tcatcgacaa acaatccatc 1980
tcccctgatg ttaggaaatt tcgatttgca ttgccctctg aggatcaagt cttgggcttg 2040
cctgttggta aacacatctt cctctgtgcc gttattgacg ataagctctg catgcgcgcc 2100
tacacgccta ctagcacgat cgatgaggtg gggtacttcg agttggttgt caagatatac 2160
ttcaaaggaa ttcaccctaa attccccaat ggggggcaaa tgtcacaata ccttgattct 2220
ctccaattag ggtcatttct cgacgtgaaa ggtccattag gtcacattga ataccaagga 2280
aagggcaatt tcttagttca tggcaaacaa aagtttgcca agaagttggc catgatagca 2340
ggtggaacag ggataactcc agtttatcaa gtcatgcagg caattctgaa agatccagaa 2400
gatgacacag aaatgtatgt ggtctatgct aatagaacag aggatgatat tttacttaag 2460
gaagagcttg attcatgggc tgagaaaatt ccagaaaggg ttaaagtttg gtatgtggtt 2520
caagattcta ttaaagaagg atggaagtac agccttggtt ttatttcaga agccattttg 2580
agagaacata tccctgagcc atctcacaca acactggctt tggcttgtgg accacctcct 2640
atgattcaat ttgctgttaa tccaaacttg gagaagatgg gctatgacat taaggattcc 2700
ttattggtgt tctaa 2715
<210> 2
<211> 904
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 2
Met Ala Ala Ser Val Glu Asn Arg Gln Phe Ser His Ile Glu Ala Gly
1 5 10 15
Leu Ser Arg Ser Phe Lys Pro Arg Ser Asp Ser Pro Val Arg Gly Cys
20 25 30
Asn Phe Pro Pro Pro Asn Ser Thr Asn Phe Gln Lys Lys Pro Asn Ser
35 40 45
Thr Ile Phe Leu Asp Tyr Ser Ser Ser Glu Asp Asp Asp Asp Asp Asp
50 55 60
Glu Lys Asn Glu Tyr Leu Gln Met Ile Lys Lys Gly Asn Ser Glu Leu
65 70 75 80
Glu Pro Ser Val His Asp Ser Arg Asp Glu Gly Thr Ala Asp Asn Trp
85 90 95
Ile Glu Arg Asn Phe Ser Leu Ile Arg Leu Thr Gly Lys His Pro Phe
100 105 110
Asn Ser Glu Pro Pro Leu Asn Arg Leu Met His His Gly Phe Ile Thr
115 120 125
Pro Val Pro Leu His Tyr Val Arg Asn His Gly Pro Val Pro Lys Gly
130 135 140
Thr Trp Asp Asp Trp Thr Val Glu Val Thr Gly Leu Val Lys Arg Pro
145 150 155 160
Met Lys Phe Thr Met Asp Gln Leu Val Asn Glu Phe Pro Ser Arg Glu
165 170 175
Leu Pro Val Thr Leu Val Cys Ala Gly Asn Arg Arg Lys Glu Gln Asn
180 185 190
Met Val Lys Gln Thr Ile Gly Phe Asn Trp Gly Ala Ala Ala Val Ser
195 200 205
Thr Thr Val Trp Arg Gly Val Pro Leu Arg Ala Leu Leu Lys Arg Tyr
210 215 220
Gly Val Phe Ser Lys Asn Lys Gly Ala Leu Asn Val Cys Phe Glu Gly
225 230 235 240
Ala Asp Val Leu Pro Gly Gly Gly Gly Ser Lys Tyr Gly Thr Ser Ile
245 250 255
Lys Lys Glu Phe Ala Met Asp Pro Ala Arg Asp Ile Ile Ile Ala Tyr
260 265 270
Met Gln Asn Gly Glu Lys Leu Ala Pro Asp His Gly Phe Pro Val Arg
275 280 285
Met Ile Ile Pro Gly Phe Ile Gly Gly Arg Met Val Lys Trp Ile Lys
290 295 300
Arg Ile Ile Val Thr Thr Gln Glu Ser Asp Ser Tyr Tyr His Phe Lys
305 310 315 320
Asp Asn Arg Val Leu Pro Pro His Val Asp Ala Glu Leu Ala Asn Thr
325 330 335
Glu Ala Trp Trp Tyr Lys Pro Glu Tyr Ile Ile Asn Glu Leu Asn Ile
340 345 350
Asn Ser Val Ile Thr Thr Pro Cys His Glu Glu Ile Leu Pro Ile Asn
355 360 365
Ala Trp Thr Thr Gln Arg Pro Tyr Thr Leu Arg Gly Tyr Ser Tyr Ser
370 375 380
Gly Gly Gly Lys Lys Val Thr Arg Val Glu Val Thr Leu Asp Gly Gly
385 390 395 400
Glu Thr Trp Gln Val Cys Thr Leu Asp His Pro Glu Lys Pro Thr Lys
405 410 415
Tyr Gly Lys Tyr Trp Cys Trp Cys Phe Trp Ser Leu Glu Val Glu Val
420 425 430
Leu Asp Leu Leu Ser Ala Lys Glu Ile Ala Val Arg Ala Trp Asp Glu
435 440 445
Thr Leu Asn Thr Gln Pro Glu Lys Leu Ile Trp Asn Val Met Gly Met
450 455 460
Met Asn Asn Cys Trp Phe Arg Val Lys Met Asn Val Cys Lys Pro His
465 470 475 480
Lys Gly Glu Ile Gly Ile Val Phe Glu His Pro Thr Gln Pro Gly Asn
485 490 495
Gln Ser Gly Gly Trp Met Ala Lys Glu Arg His Leu Glu Ile Ser Ala
500 505 510
Glu Ala Pro Pro Thr Leu Lys Lys Ser Ile Ser Thr Pro Phe Met Asn
515 520 525
Thr Ala Ser Lys Met Tyr Ser Met Ser Glu Val Arg Lys His Ser Ser
530 535 540
Ala Asp Ser Ala Trp Ile Ile Val His Gly His Ile Tyr Asp Ala Thr
545 550 555 560
Arg Phe Leu Lys Asp His Pro Gly Gly Ser Asp Ser Ile Leu Ile Asn
565 570 575
Ala Gly Thr Asp Cys Thr Glu Glu Phe Asp Ala Ile His Ser Asp Lys
580 585 590
Ala Lys Lys Leu Leu Glu Glu Phe Arg Ile Gly Glu Leu Leu Thr Thr
595 600 605
Gly Tyr Thr Ser Asp Ser Pro Gly Asn Ser Val His Gly Ser Ser Ser
610 615 620
Phe Ser Ser Phe Leu Ala Pro Ile Lys Glu Leu Val Pro Ala Gln Arg
625 630 635 640
Ser Val Ala Leu Ile Pro Arg Glu Lys Ile Pro Cys Lys Leu Ile Asp
645 650 655
Lys Gln Ser Ile Ser Pro Asp Val Arg Lys Phe Arg Phe Ala Leu Pro
660 665 670
Ser Glu Asp Gln Val Leu Gly Leu Pro Val Gly Lys His Ile Phe Leu
675 680 685
Cys Ala Val Ile Asp Asp Lys Leu Cys Met Arg Ala Tyr Thr Pro Thr
690 695 700
Ser Thr Ile Asp Glu Val Gly Tyr Phe Glu Leu Val Val Lys Ile Tyr
705 710 715 720
Phe Lys Gly Ile His Pro Lys Phe Pro Asn Gly Gly Gln Met Ser Gln
725 730 735
Tyr Leu Asp Ser Leu Gln Leu Gly Ser Phe Leu Asp Val Lys Gly Pro
740 745 750
Leu Gly His Ile Glu Tyr Gln Gly Lys Gly Asn Phe Leu Val His Gly
755 760 765
Lys Gln Lys Phe Ala Lys Lys Leu Ala Met Ile Ala Gly Gly Thr Gly
770 775 780
Ile Thr Pro Val Tyr Gln Val Met Gln Ala Ile Leu Lys Asp Pro Glu
785 790 795 800
Asp Asp Thr Glu Met Tyr Val Val Tyr Ala Asn Arg Thr Glu Asp Asp
805 810 815
Ile Leu Leu Lys Glu Glu Leu Asp Ser Trp Ala Glu Lys Ile Pro Glu
820 825 830
Arg Val Lys Val Trp Tyr Val Val Gln Asp Ser Ile Lys Glu Gly Trp
835 840 845
Lys Tyr Ser Leu Gly Phe Ile Ser Glu Ala Ile Leu Arg Glu His Ile
850 855 860
Pro Glu Pro Ser His Thr Thr Leu Ala Leu Ala Cys Gly Pro Pro Pro
865 870 875 880
Met Ile Gln Phe Ala Val Asn Pro Asn Leu Glu Lys Met Gly Tyr Asp
885 890 895
Ile Lys Asp Ser Leu Leu Val Phe
900
<210> 3
<211> 2715
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 3
atggcggcat ctgtcgaaaa caggcagttc agtcacctag aagccggttt atcccggtct 60
ttcaagcccc ggtctgattc cccggttcgt ggctgcaact tcccttcgcc caacagtact 120
aatttccaaa agaaaccaaa ttccaccatt taccttgatt actcgtcgag tgaagacgac 180
gatgatgatg acgaaaaaaa tgagtacctt caaatgatta aaaaagggaa ttcagagtta 240
gagccatctg ttcatgacac tagggacgaa ggtaccgctg ataattggat tgaacgcaac 300
ttttccatga ttcgtctcac cggaaagcat ccatttaact ccgaaccacc gttgaaccgg 360
ctcatgcacc acggctttat cacaccggtc ccacttcatt acgttcgtaa ccatggaccg 420
gttcccaagg gcacgtggga tgactggacc gtggaagtca cgggactagt gaagcgtcct 480
atgaaattca caatggacca gttggttaac gaattccctt gtagagaatt gcccgttacg 540
cttgtttgtg ctggcaatcg aaggaaagaa cagaacatgg ttaaacaaac cattggtttc 600
aactggggcg ccgctgccgt ttcaacaacg atatggcgcg gggtacccct ccgcgctttg 660
ctaaaacggt gcggtgtttt tagcaagaat aaaggggcgc ttaatgtttg cttcgaagga 720
gctgatgtgt tgcccggagg tggtggttca aagtatggaa ccagcattaa gaaggaattt 780
gcaatggatc cagcacgaga tatcatcgta gcctacatgc agaacggaga aaaattggca 840
cccgaccacg ggtttccagt acgaatgata attccaggat tcattggagg aagaatggtg 900
aaatggataa agaggattat agtcaccacc caagaatcag acagctatta tcatttcaag 960
gacaatagag ttcttcctcc ccatgttgat gctgaacttg caaataccga agcatggtgg 1020
tacaagccag agtatatcat caatgagctt aatattaact ctgtcattac gacgccgtgt 1080
catgaagaaa ttttgccaat taacgcctgg acgactcagc gaccttacac gttgaggggc 1140
tattcttatt ctggcggagg gaaaaaagta acgcgagtag aagtgacgtt ggatggagga 1200
gaaacatggc aagttagcac actagatcac ccagagaagc ccaccaaata tggcaagtac 1260
tggtgttggt gcttttggtc actcgaggtt gaggtgttag acttgctcag tgctaaagaa 1320
attgctgttc gagcttggga tgagaccctc aatactcaac ccgagaagct tatttggaac 1380
gtcatgggaa tgatgaataa ttgctggttc cgagtaaaga tgaatgtgtg caagcctcac 1440
aagggagaga ttggaatagt gtttgagcat ccgactcaac ctggaaacca atcaggtgga 1500
tggatggcga aggagagaca tttggagata tcagcagagg cacctcaaac actaaagaag 1560
agtatctcaa ctccattcat gaacacagct tccaagatgt actccatgtc cgaggtcagg 1620
aaacacagct ctgctgactc tgcttggatc atagtccatg gtcatatcta tgacgccacg 1680
cgtttcttga aagatcaccc tggtgggact gacagcattc tcatcaatgc tggcactgat 1740
tgcactgagg aatttgatgc aattcattct gataaggcta agaagctctt ggaggatttc 1800
aggattggtg aactcataac tactggttac acctctgact ctcctggcaa ctccgtgcac 1860
ggatcttctt ccttcagcag ctttctagca cctattaagg aacttgttcc agcgcagagg 1920
agtgtggccc taattccaag agagaaaatc ccatgcaaac tcatcgacaa gcaatccatc 1980
tcccatgatg ttaggaaatt tcgatttgca ttgccctctg aggatcaagt cttgggcttg 2040
cctgttggaa aacatatctt cctctgtgcc gttattgacg ataagctctg catgcgcgct 2100
tacacgccta ctagcacgat cgatgaggtg gggtacttcg agttggttgt caagatatac 2160
ttcaaaggaa ttcaccctaa attccccaat ggagggcaaa tgtcacagta tcttgattct 2220
atgccgttag ggtcatttct cgacgtgaaa ggtccattag gtcacattga ataccaagga 2280
aagggaaatt tcttagttca tggcaaacag aagtttgcca agaagttggc catgatagca 2340
ggtggaacag gaataactcc agtgtatcaa gtcatgcagg caattctgaa agatccagaa 2400
gatgacacag aaatgtatgt ggtgtatgct aacagaacag aggatgatat tttacttaag 2460
gaagagcttg attcatgggc tgagaaaatt ccagagaggg ttaaagtttg gtatgtggtt 2520
caggattcta ttaaagaagg atggaagtac agcattggtt ttattacaga agccattttg 2580
agagaacata tccctgagcc atctcacaca acactggctt tggcttgtgg accacctcct 2640
atgattcaat ttgctgttaa tccaaacttg gagaagatgg gctatgacat taaggattcc 2700
ttattggtgt tctaa 2715
<210> 4
<211> 904
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 4
Met Ala Ala Ser Val Glu Asn Arg Gln Phe Ser His Leu Glu Ala Gly
1 5 10 15
Leu Ser Arg Ser Phe Lys Pro Arg Ser Asp Ser Pro Val Arg Gly Cys
20 25 30
Asn Phe Pro Ser Pro Asn Ser Thr Asn Phe Gln Lys Lys Pro Asn Ser
35 40 45
Thr Ile Tyr Leu Asp Tyr Ser Ser Ser Glu Asp Asp Asp Asp Asp Asp
50 55 60
Glu Lys Asn Glu Tyr Leu Gln Met Ile Lys Lys Gly Asn Ser Glu Leu
65 70 75 80
Glu Pro Ser Val His Asp Thr Arg Asp Glu Gly Thr Ala Asp Asn Trp
85 90 95
Ile Glu Arg Asn Phe Ser Met Ile Arg Leu Thr Gly Lys His Pro Phe
100 105 110
Asn Ser Glu Pro Pro Leu Asn Arg Leu Met His His Gly Phe Ile Thr
115 120 125
Pro Val Pro Leu His Tyr Val Arg Asn His Gly Pro Val Pro Lys Gly
130 135 140
Thr Trp Asp Asp Trp Thr Val Glu Val Thr Gly Leu Val Lys Arg Pro
145 150 155 160
Met Lys Phe Thr Met Asp Gln Leu Val Asn Glu Phe Pro Cys Arg Glu
165 170 175
Leu Pro Val Thr Leu Val Cys Ala Gly Asn Arg Arg Lys Glu Gln Asn
180 185 190
Met Val Lys Gln Thr Ile Gly Phe Asn Trp Gly Ala Ala Ala Val Ser
195 200 205
Thr Thr Ile Trp Arg Gly Val Pro Leu Arg Ala Leu Leu Lys Arg Cys
210 215 220
Gly Val Phe Ser Lys Asn Lys Gly Ala Leu Asn Val Cys Phe Glu Gly
225 230 235 240
Ala Asp Val Leu Pro Gly Gly Gly Gly Ser Lys Tyr Gly Thr Ser Ile
245 250 255
Lys Lys Glu Phe Ala Met Asp Pro Ala Arg Asp Ile Ile Val Ala Tyr
260 265 270
Met Gln Asn Gly Glu Lys Leu Ala Pro Asp His Gly Phe Pro Val Arg
275 280 285
Met Ile Ile Pro Gly Phe Ile Gly Gly Arg Met Val Lys Trp Ile Lys
290 295 300
Arg Ile Ile Val Thr Thr Gln Glu Ser Asp Ser Tyr Tyr His Phe Lys
305 310 315 320
Asp Asn Arg Val Leu Pro Pro His Val Asp Ala Glu Leu Ala Asn Thr
325 330 335
Glu Ala Trp Trp Tyr Lys Pro Glu Tyr Ile Ile Asn Glu Leu Asn Ile
340 345 350
Asn Ser Val Ile Thr Thr Pro Cys His Glu Glu Ile Leu Pro Ile Asn
355 360 365
Ala Trp Thr Thr Gln Arg Pro Tyr Thr Leu Arg Gly Tyr Ser Tyr Ser
370 375 380
Gly Gly Gly Lys Lys Val Thr Arg Val Glu Val Thr Leu Asp Gly Gly
385 390 395 400
Glu Thr Trp Gln Val Ser Thr Leu Asp His Pro Glu Lys Pro Thr Lys
405 410 415
Tyr Gly Lys Tyr Trp Cys Trp Cys Phe Trp Ser Leu Glu Val Glu Val
420 425 430
Leu Asp Leu Leu Ser Ala Lys Glu Ile Ala Val Arg Ala Trp Asp Glu
435 440 445
Thr Leu Asn Thr Gln Pro Glu Lys Leu Ile Trp Asn Val Met Gly Met
450 455 460
Met Asn Asn Cys Trp Phe Arg Val Lys Met Asn Val Cys Lys Pro His
465 470 475 480
Lys Gly Glu Ile Gly Ile Val Phe Glu His Pro Thr Gln Pro Gly Asn
485 490 495
Gln Ser Gly Gly Trp Met Ala Lys Glu Arg His Leu Glu Ile Ser Ala
500 505 510
Glu Ala Pro Gln Thr Leu Lys Lys Ser Ile Ser Thr Pro Phe Met Asn
515 520 525
Thr Ala Ser Lys Met Tyr Ser Met Ser Glu Val Arg Lys His Ser Ser
530 535 540
Ala Asp Ser Ala Trp Ile Ile Val His Gly His Ile Tyr Asp Ala Thr
545 550 555 560
Arg Phe Leu Lys Asp His Pro Gly Gly Thr Asp Ser Ile Leu Ile Asn
565 570 575
Ala Gly Thr Asp Cys Thr Glu Glu Phe Asp Ala Ile His Ser Asp Lys
580 585 590
Ala Lys Lys Leu Leu Glu Asp Phe Arg Ile Gly Glu Leu Ile Thr Thr
595 600 605
Gly Tyr Thr Ser Asp Ser Pro Gly Asn Ser Val His Gly Ser Ser Ser
610 615 620
Phe Ser Ser Phe Leu Ala Pro Ile Lys Glu Leu Val Pro Ala Gln Arg
625 630 635 640
Ser Val Ala Leu Ile Pro Arg Glu Lys Ile Pro Cys Lys Leu Ile Asp
645 650 655
Lys Gln Ser Ile Ser His Asp Val Arg Lys Phe Arg Phe Ala Leu Pro
660 665 670
Ser Glu Asp Gln Val Leu Gly Leu Pro Val Gly Lys His Ile Phe Leu
675 680 685
Cys Ala Val Ile Asp Asp Lys Leu Cys Met Arg Ala Tyr Thr Pro Thr
690 695 700
Ser Thr Ile Asp Glu Val Gly Tyr Phe Glu Leu Val Val Lys Ile Tyr
705 710 715 720
Phe Lys Gly Ile His Pro Lys Phe Pro Asn Gly Gly Gln Met Ser Gln
725 730 735
Tyr Leu Asp Ser Met Pro Leu Gly Ser Phe Leu Asp Val Lys Gly Pro
740 745 750
Leu Gly His Ile Glu Tyr Gln Gly Lys Gly Asn Phe Leu Val His Gly
755 760 765
Lys Gln Lys Phe Ala Lys Lys Leu Ala Met Ile Ala Gly Gly Thr Gly
770 775 780
Ile Thr Pro Val Tyr Gln Val Met Gln Ala Ile Leu Lys Asp Pro Glu
785 790 795 800
Asp Asp Thr Glu Met Tyr Val Val Tyr Ala Asn Arg Thr Glu Asp Asp
805 810 815
Ile Leu Leu Lys Glu Glu Leu Asp Ser Trp Ala Glu Lys Ile Pro Glu
820 825 830
Arg Val Lys Val Trp Tyr Val Val Gln Asp Ser Ile Lys Glu Gly Trp
835 840 845
Lys Tyr Ser Ile Gly Phe Ile Thr Glu Ala Ile Leu Arg Glu His Ile
850 855 860
Pro Glu Pro Ser His Thr Thr Leu Ala Leu Ala Cys Gly Pro Pro Pro
865 870 875 880
Met Ile Gln Phe Ala Val Asn Pro Asn Leu Glu Lys Met Gly Tyr Asp
885 890 895
Ile Lys Asp Ser Leu Leu Val Phe
900
<210> 5
<211> 2715
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 5
atggcggcat ctgtcgaaaa caggcagttc agtcacctag aagccggttt atcccggtct 60
ttcaagcccc ggtctgattc cccggttcgt ggctgcaact tcccttcgcc caacagtact 120
aatttccaaa agaaaccaaa ttccaccatt taccttgatt actcgtcgag tgaagacgac 180
gatgatgatg acgaaaaaaa tgagtacctt caaatgatta aaaaagggaa ttcagagtta 240
gagccatctg ttcatgacac tagggacgaa ggtaccgctg ataattggat tgaacgcaac 300
ttttccatga ttcgtctcac cggaaagcat ccatttaact ccgaaccacc gttgaaccgg 360
ctcatgcacc acggctttat cacaccggtc ccacttcatt acgttcgtaa ccatggaccg 420
gttcccaagg gcacgtggga tgactggacc gtggaagtca cgggactagt gaagcgtcct 480
atgaaattca caatggacca gttggttaac gaattccctt gtagagaatt gcccgttacg 540
cttgtttgtg ctggcaatcg aaggaaagaa cagaacatgg ttaaacaaac cattggtttc 600
aactggggcg ccgctgccgt ttcaacaacg atatggcgcg gggtacccct ccgcgctttg 660
ctaaaacggt gcggtgtttt tagcaagaat aaaggggcgc ttaatgtttg cttcgaagga 720
gctgatgtgt tgcccggagg tggtggttca aagtatggaa ccagcattaa gaaggaattt 780
gcaatggatc cagcacgaga tatcatcgta gcctacatgc agaacggaga aaaattggca 840
cccgaccacg ggtttccagt acgaatgata attccaggat tcattggagg aagaatggtg 900
aaatggataa agaggattat agtcaccacc caagaatcag acagctatta tcatttcaag 960
gacaatagag ttcttcctcc ccatgttgat gctgaacttg caaataccga agcatggtgg 1020
tacaagccag agtatatcat caatgagctt aatattaact ctgtcattac gacgccgtgt 1080
catgaagaaa ttttgccaat taacgcctgg acgactcagc gaccttacac gttgaggggc 1140
tattcttatt ctggcggagg gaaaaaagta acgcgagtag aagtgacgtt ggatggagga 1200
gaaacatggc aagttagcac actagatcac ccagagaagc ccaccaaata tggcaagtac 1260
tggtgttggt gcttttggtc actcgaggtt gaggtgttag acttgctcag tgctaaagaa 1320
attgctgttc gagcttggga tgagaccctc aatactcaac ccgagaagct tatttggaac 1380
gtcatgggaa tgatgaataa ttgctggttc cgagtaaaga tgaatgtgtg caagcctcac 1440
aagggagaga ttggaatagt gtttgagcat ccgactcaac ctggaaacca atcaggtgga 1500
tggatggcga aggagagaca tttggagata tcagcagagg cacctcaaac actaaagaag 1560
agtatcgata ctccattcat gaacacagct tccaagatgt actccatgtc cgaggtcagg 1620
aaacacagct ctgctgactc tgcttggatc atagtccatg gtcatatcta tgacgccacg 1680
cgtttcttga aagatcaccc tggtgggact gacagcattc tcatcaatgc tggcactgat 1740
tgcactgagg aatttgatgc aattcattct gataaggcta agaagctctt ggaggatttc 1800
aggattggtg aactcataac tactggttac acctctgact ctcctggcaa ctccgtgcac 1860
ggatcttctt ccttcagcag ctttctagca cctattaagg aacttgttcc agcgcagagg 1920
agtgtggccc taattccaag agagaaaatc ccatgcaaac tcatcgacaa gcaatccatc 1980
tcccatgatg ttaggaaatt tcgatttgca ttgccctctg aggatcaagt cttgggcttg 2040
cctgttggaa aacatatctt cctctgtgcc gttattgacg ataagctctg catgcgcgct 2100
tacacgccta ctagcacgat cgatgaggtg gggtacttcg agttggttgt caagatatac 2160
ttcaaaggaa ttcaccctaa attccccaat ggagggcaaa tgtcacagta tcttgattct 2220
atgccgttag ggtcatttct cgacgtgaaa ggtccattag gtcacattga ataccaagga 2280
aagggaaatt tcttagttca tggcaaacag aagtttgcca agaagttggc catgatagca 2340
ggtggaacag gaataactcc agtgtatcaa gtcatgcagg caattctgaa agatccagaa 2400
gatgacacag aaatgtatgt ggtgtatgct aacagaacag aggatgatat tttacttaag 2460
gaagagcttg attcatgggc tgagaaaatt ccagagaggg ttaaagtttg gtatgtggtt 2520
caggattcta ttaaagaagg atggaagtac agcattggtt ttattacaga agccattttg 2580
agagaacata tccctgagcc atctcacaca acactggctt tggcttgtgg accacctcct 2640
atgattcaat ttgctgttaa tccaaacttg gagaagatgg gctatgacat taaggattcc 2700
ttattggtgt tctaa 2715
<210> 6
<211> 904
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 6
Met Ala Ala Ser Val Glu Asn Arg Gln Phe Ser His Leu Glu Ala Gly
1 5 10 15
Leu Ser Arg Ser Phe Lys Pro Arg Ser Asp Ser Pro Val Arg Gly Cys
20 25 30
Asn Phe Pro Ser Pro Asn Ser Thr Asn Phe Gln Lys Lys Pro Asn Ser
35 40 45
Thr Ile Tyr Leu Asp Tyr Ser Ser Ser Glu Asp Asp Asp Asp Asp Asp
50 55 60
Glu Lys Asn Glu Tyr Leu Gln Met Ile Lys Lys Gly Asn Ser Glu Leu
65 70 75 80
Glu Pro Ser Val His Asp Thr Arg Asp Glu Gly Thr Ala Asp Asn Trp
85 90 95
Ile Glu Arg Asn Phe Ser Met Ile Arg Leu Thr Gly Lys His Pro Phe
100 105 110
Asn Ser Glu Pro Pro Leu Asn Arg Leu Met His His Gly Phe Ile Thr
115 120 125
Pro Val Pro Leu His Tyr Val Arg Asn His Gly Pro Val Pro Lys Gly
130 135 140
Thr Trp Asp Asp Trp Thr Val Glu Val Thr Gly Leu Val Lys Arg Pro
145 150 155 160
Met Lys Phe Thr Met Asp Gln Leu Val Asn Glu Phe Pro Cys Arg Glu
165 170 175
Leu Pro Val Thr Leu Val Cys Ala Gly Asn Arg Arg Lys Glu Gln Asn
180 185 190
Met Val Lys Gln Thr Ile Gly Phe Asn Trp Gly Ala Ala Ala Val Ser
195 200 205
Thr Thr Ile Trp Arg Gly Val Pro Leu Arg Ala Leu Leu Lys Arg Cys
210 215 220
Gly Val Phe Ser Lys Asn Lys Gly Ala Leu Asn Val Cys Phe Glu Gly
225 230 235 240
Ala Asp Val Leu Pro Gly Gly Gly Gly Ser Lys Tyr Gly Thr Ser Ile
245 250 255
Lys Lys Glu Phe Ala Met Asp Pro Ala Arg Asp Ile Ile Val Ala Tyr
260 265 270
Met Gln Asn Gly Glu Lys Leu Ala Pro Asp His Gly Phe Pro Val Arg
275 280 285
Met Ile Ile Pro Gly Phe Ile Gly Gly Arg Met Val Lys Trp Ile Lys
290 295 300
Arg Ile Ile Val Thr Thr Gln Glu Ser Asp Ser Tyr Tyr His Phe Lys
305 310 315 320
Asp Asn Arg Val Leu Pro Pro His Val Asp Ala Glu Leu Ala Asn Thr
325 330 335
Glu Ala Trp Trp Tyr Lys Pro Glu Tyr Ile Ile Asn Glu Leu Asn Ile
340 345 350
Asn Ser Val Ile Thr Thr Pro Cys His Glu Glu Ile Leu Pro Ile Asn
355 360 365
Ala Trp Thr Thr Gln Arg Pro Tyr Thr Leu Arg Gly Tyr Ser Tyr Ser
370 375 380
Gly Gly Gly Lys Lys Val Thr Arg Val Glu Val Thr Leu Asp Gly Gly
385 390 395 400
Glu Thr Trp Gln Val Ser Thr Leu Asp His Pro Glu Lys Pro Thr Lys
405 410 415
Tyr Gly Lys Tyr Trp Cys Trp Cys Phe Trp Ser Leu Glu Val Glu Val
420 425 430
Leu Asp Leu Leu Ser Ala Lys Glu Ile Ala Val Arg Ala Trp Asp Glu
435 440 445
Thr Leu Asn Thr Gln Pro Glu Lys Leu Ile Trp Asn Val Met Gly Met
450 455 460
Met Asn Asn Cys Trp Phe Arg Val Lys Met Asn Val Cys Lys Pro His
465 470 475 480
Lys Gly Glu Ile Gly Ile Val Phe Glu His Pro Thr Gln Pro Gly Asn
485 490 495
Gln Ser Gly Gly Trp Met Ala Lys Glu Arg His Leu Glu Ile Ser Ala
500 505 510
Glu Ala Pro Gln Thr Leu Lys Lys Ser Ile Asp Thr Pro Phe Met Asn
515 520 525
Thr Ala Ser Lys Met Tyr Ser Met Ser Glu Val Arg Lys His Ser Ser
530 535 540
Ala Asp Ser Ala Trp Ile Ile Val His Gly His Ile Tyr Asp Ala Thr
545 550 555 560
Arg Phe Leu Lys Asp His Pro Gly Gly Thr Asp Ser Ile Leu Ile Asn
565 570 575
Ala Gly Thr Asp Cys Thr Glu Glu Phe Asp Ala Ile His Ser Asp Lys
580 585 590
Ala Lys Lys Leu Leu Glu Asp Phe Arg Ile Gly Glu Leu Ile Thr Thr
595 600 605
Gly Tyr Thr Ser Asp Ser Pro Gly Asn Ser Val His Gly Ser Ser Ser
610 615 620
Phe Ser Ser Phe Leu Ala Pro Ile Lys Glu Leu Val Pro Ala Gln Arg
625 630 635 640
Ser Val Ala Leu Ile Pro Arg Glu Lys Ile Pro Cys Lys Leu Ile Asp
645 650 655
Lys Gln Ser Ile Ser His Asp Val Arg Lys Phe Arg Phe Ala Leu Pro
660 665 670
Ser Glu Asp Gln Val Leu Gly Leu Pro Val Gly Lys His Ile Phe Leu
675 680 685
Cys Ala Val Ile Asp Asp Lys Leu Cys Met Arg Ala Tyr Thr Pro Thr
690 695 700
Ser Thr Ile Asp Glu Val Gly Tyr Phe Glu Leu Val Val Lys Ile Tyr
705 710 715 720
Phe Lys Gly Ile His Pro Lys Phe Pro Asn Gly Gly Gln Met Ser Gln
725 730 735
Tyr Leu Asp Ser Met Pro Leu Gly Ser Phe Leu Asp Val Lys Gly Pro
740 745 750
Leu Gly His Ile Glu Tyr Gln Gly Lys Gly Asn Phe Leu Val His Gly
755 760 765
Lys Gln Lys Phe Ala Lys Lys Leu Ala Met Ile Ala Gly Gly Thr Gly
770 775 780
Ile Thr Pro Val Tyr Gln Val Met Gln Ala Ile Leu Lys Asp Pro Glu
785 790 795 800
Asp Asp Thr Glu Met Tyr Val Val Tyr Ala Asn Arg Thr Glu Asp Asp
805 810 815
Ile Leu Leu Lys Glu Glu Leu Asp Ser Trp Ala Glu Lys Ile Pro Glu
820 825 830
Arg Val Lys Val Trp Tyr Val Val Gln Asp Ser Ile Lys Glu Gly Trp
835 840 845
Lys Tyr Ser Ile Gly Phe Ile Thr Glu Ala Ile Leu Arg Glu His Ile
850 855 860
Pro Glu Pro Ser His Thr Thr Leu Ala Leu Ala Cys Gly Pro Pro Pro
865 870 875 880
Met Ile Gln Phe Ala Val Asn Pro Asn Leu Glu Lys Met Gly Tyr Asp
885 890 895
Ile Lys Asp Ser Leu Leu Val Phe
900
<210> 7
<211> 2547
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 7
atggcggcat ctgtcgaaaa caggcagttc agtcacctag aagccggttt atcccggtct 60
aattcagagt tagagccatc tgttcatgac actagggacg aaggtaccgc tgataattgg 120
attgaacgca acttttccat gattcgtctc accggaaagc atccatttaa ctccgaacca 180
ccgttgaacc ggctcatgca ccacggcttt atcacaccgg tcccacttca ttacgttcgt 240
aaccatggac cggttcccaa gggcacgtgg gatgactgga ccgtggaagt cacgggacta 300
gtgaagcgtc ctatgaaatt cacaatggac cagttggtta acgaattccc ttgtagagaa 360
ttgcccgtta cgcttgtttg tgctggcaat cgaaggaaag aacagaacat ggttaaacaa 420
accattggtt tcaactgggg cgccgctgcc gtttcaacaa cgatatggcg cggggtaccc 480
ctccgcgctt tgctaaaacg gtgcggtgtt tttagcaaga ataaaggggc gcttaatgtt 540
tgcttcgaag gagctgatgt gttgcccgga ggtggtggtt caaagtatgg aaccagcatt 600
aagaaggaat ttgcaatgga tccagcacga gatatcatcg tagcctacat gcagaacgga 660
gaaaaattgg cacccgacca cgggtttcca gtacgaatga taattccagg attcattgga 720
ggaagaatgg tgaaatggat aaagaggatt atagtcacca cccaagaatc agacagctat 780
tatcatttca aggacaatag agttcttcct ccccatgttg atgctgaact tgcaaatacc 840
gaagcatggt ggtacaagcc agagtatatc atcaatgagc ttaatattaa ctctgtcatt 900
acgacgccgt gtcatgaaga aattttgcca attaacgcct ggacgactca gcgaccttac 960
acgttgaggg gctattctta ttctggcgga gggaaaaaag taacgcgagt agaagtgacg 1020
ttggatggag gagaaacatg gcaagttagc acactagatc acccagagaa gcccaccaaa 1080
tatggcaagt actggtgttg gtgcttttgg tcactcgagg ttgaggtgtt agacttgctc 1140
agtgctaaag aaattgctgt tcgagcttgg gatgagaccc tcaatactca acccgagaag 1200
cttatttgga acgtcatggg aatgatgaat aattgctggt tccgagtaaa gatgaatgtg 1260
tgcaagcctc acaagggaga gattggaata gtgtttgagc atccgactca acctggaaac 1320
caatcaggtg gatggatggc gaaggagaga catttggaga tatcagcaga ggcacctcaa 1380
acactaaaga agagtatctc aactccattc atgaacacag cttccaagat gtactccatg 1440
tccgaggtca ggaaacacag ctctgctgac tctgcttgga tcatagtcca tggtcatatc 1500
tatgacgcca cgcgtttctt gaaagatcac cctggtggga ctgacagcat tctcatcaat 1560
gctggcactg attgcactga ggaatttgat gcaattcatt ctgataaggc taagaagctc 1620
ttggaggatt tcaggattgg tgaactcata actactggtt acacctctga ctctcctggc 1680
aactccgtgc acggatcttc ttccttcagc agctttctag cacctattaa ggaacttgtt 1740
ccagcgcaga ggagtgtggc cctaattcca agagagaaaa tcccatgcaa actcatcgac 1800
aagcaatcca tctcccatga tgttaggaaa tttcgatttg cattgccctc tgaggatcaa 1860
gtcttgggct tgcctgttgg aaaacatatc ttcctctgtg ccgttattga cgataagctc 1920
tgcatgcgcg cttacacgcc tactagcacg atcgatgagg tggggtactt cgagttggtt 1980
gtcaagatat acttcaaagg aattcaccct aaattcccca atggagggca aatgtcacag 2040
tatcttgatt ctatgccgtt agggtcattt ctcgacgtga aaggtccatt aggtcacatt 2100
gaataccaag gaaagggaaa tttcttagtt catggcaaac agaagtttgc caagaagttg 2160
gccatgatag caggtggaac aggaataact ccagtgtatc aagtcatgca ggcaattctg 2220
aaagatccag aagatgacac agaaatgtat gtggtgtatg ctaacagaac agaggatgat 2280
attttactta aggaagagct tgattcatgg gctgagaaaa ttccagagag ggttaaagtt 2340
tggtatgtgg ttcaggattc tattaaagaa ggatggaagt acagcattgg ttttattaca 2400
gaagccattt tgagagaaca tatccctgag ccatctcaca caacactggc tttggcttgt 2460
ggaccacctc ctatgattca atttgctgtt aatccaaact tggagaagat gggctatgac 2520
attaaggatt ccttattggt gttctaa 2547
<210> 8
<211> 848
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 8
Met Ala Ala Ser Val Glu Asn Arg Gln Phe Ser His Leu Glu Ala Gly
1 5 10 15
Leu Ser Arg Ser Asn Ser Glu Leu Glu Pro Ser Val His Asp Thr Arg
20 25 30
Asp Glu Gly Thr Ala Asp Asn Trp Ile Glu Arg Asn Phe Ser Met Ile
35 40 45
Arg Leu Thr Gly Lys His Pro Phe Asn Ser Glu Pro Pro Leu Asn Arg
50 55 60
Leu Met His His Gly Phe Ile Thr Pro Val Pro Leu His Tyr Val Arg
65 70 75 80
Asn His Gly Pro Val Pro Lys Gly Thr Trp Asp Asp Trp Thr Val Glu
85 90 95
Val Thr Gly Leu Val Lys Arg Pro Met Lys Phe Thr Met Asp Gln Leu
100 105 110
Val Asn Glu Phe Pro Cys Arg Glu Leu Pro Val Thr Leu Val Cys Ala
115 120 125
Gly Asn Arg Arg Lys Glu Gln Asn Met Val Lys Gln Thr Ile Gly Phe
130 135 140
Asn Trp Gly Ala Ala Ala Val Ser Thr Thr Ile Trp Arg Gly Val Pro
145 150 155 160
Leu Arg Ala Leu Leu Lys Arg Cys Gly Val Phe Ser Lys Asn Lys Gly
165 170 175
Ala Leu Asn Val Cys Phe Glu Gly Ala Asp Val Leu Pro Gly Gly Gly
180 185 190
Gly Ser Lys Tyr Gly Thr Ser Ile Lys Lys Glu Phe Ala Met Asp Pro
195 200 205
Ala Arg Asp Ile Ile Val Ala Tyr Met Gln Asn Gly Glu Lys Leu Ala
210 215 220
Pro Asp His Gly Phe Pro Val Arg Met Ile Ile Pro Gly Phe Ile Gly
225 230 235 240
Gly Arg Met Val Lys Trp Ile Lys Arg Ile Ile Val Thr Thr Gln Glu
245 250 255
Ser Asp Ser Tyr Tyr His Phe Lys Asp Asn Arg Val Leu Pro Pro His
260 265 270
Val Asp Ala Glu Leu Ala Asn Thr Glu Ala Trp Trp Tyr Lys Pro Glu
275 280 285
Tyr Ile Ile Asn Glu Leu Asn Ile Asn Ser Val Ile Thr Thr Pro Cys
290 295 300
His Glu Glu Ile Leu Pro Ile Asn Ala Trp Thr Thr Gln Arg Pro Tyr
305 310 315 320
Thr Leu Arg Gly Tyr Ser Tyr Ser Gly Gly Gly Lys Lys Val Thr Arg
325 330 335
Val Glu Val Thr Leu Asp Gly Gly Glu Thr Trp Gln Val Ser Thr Leu
340 345 350
Asp His Pro Glu Lys Pro Thr Lys Tyr Gly Lys Tyr Trp Cys Trp Cys
355 360 365
Phe Trp Ser Leu Glu Val Glu Val Leu Asp Leu Leu Ser Ala Lys Glu
370 375 380
Ile Ala Val Arg Ala Trp Asp Glu Thr Leu Asn Thr Gln Pro Glu Lys
385 390 395 400
Leu Ile Trp Asn Val Met Gly Met Met Asn Asn Cys Trp Phe Arg Val
405 410 415
Lys Met Asn Val Cys Lys Pro His Lys Gly Glu Ile Gly Ile Val Phe
420 425 430
Glu His Pro Thr Gln Pro Gly Asn Gln Ser Gly Gly Trp Met Ala Lys
435 440 445
Glu Arg His Leu Glu Ile Ser Ala Glu Ala Pro Gln Thr Leu Lys Lys
450 455 460
Ser Ile Ser Thr Pro Phe Met Asn Thr Ala Ser Lys Met Tyr Ser Met
465 470 475 480
Ser Glu Val Arg Lys His Ser Ser Ala Asp Ser Ala Trp Ile Ile Val
485 490 495
His Gly His Ile Tyr Asp Ala Thr Arg Phe Leu Lys Asp His Pro Gly
500 505 510
Gly Thr Asp Ser Ile Leu Ile Asn Ala Gly Thr Asp Cys Thr Glu Glu
515 520 525
Phe Asp Ala Ile His Ser Asp Lys Ala Lys Lys Leu Leu Glu Asp Phe
530 535 540
Arg Ile Gly Glu Leu Ile Thr Thr Gly Tyr Thr Ser Asp Ser Pro Gly
545 550 555 560
Asn Ser Val His Gly Ser Ser Ser Phe Ser Ser Phe Leu Ala Pro Ile
565 570 575
Lys Glu Leu Val Pro Ala Gln Arg Ser Val Ala Leu Ile Pro Arg Glu
580 585 590
Lys Ile Pro Cys Lys Leu Ile Asp Lys Gln Ser Ile Ser His Asp Val
595 600 605
Arg Lys Phe Arg Phe Ala Leu Pro Ser Glu Asp Gln Val Leu Gly Leu
610 615 620
Pro Val Gly Lys His Ile Phe Leu Cys Ala Val Ile Asp Asp Lys Leu
625 630 635 640
Cys Met Arg Ala Tyr Thr Pro Thr Ser Thr Ile Asp Glu Val Gly Tyr
645 650 655
Phe Glu Leu Val Val Lys Ile Tyr Phe Lys Gly Ile His Pro Lys Phe
660 665 670
Pro Asn Gly Gly Gln Met Ser Gln Tyr Leu Asp Ser Met Pro Leu Gly
675 680 685
Ser Phe Leu Asp Val Lys Gly Pro Leu Gly His Ile Glu Tyr Gln Gly
690 695 700
Lys Gly Asn Phe Leu Val His Gly Lys Gln Lys Phe Ala Lys Lys Leu
705 710 715 720
Ala Met Ile Ala Gly Gly Thr Gly Ile Thr Pro Val Tyr Gln Val Met
725 730 735
Gln Ala Ile Leu Lys Asp Pro Glu Asp Asp Thr Glu Met Tyr Val Val
740 745 750
Tyr Ala Asn Arg Thr Glu Asp Asp Ile Leu Leu Lys Glu Glu Leu Asp
755 760 765
Ser Trp Ala Glu Lys Ile Pro Glu Arg Val Lys Val Trp Tyr Val Val
770 775 780
Gln Asp Ser Ile Lys Glu Gly Trp Lys Tyr Ser Ile Gly Phe Ile Thr
785 790 795 800
Glu Ala Ile Leu Arg Glu His Ile Pro Glu Pro Ser His Thr Thr Leu
805 810 815
Ala Leu Ala Cys Gly Pro Pro Pro Met Ile Gln Phe Ala Val Asn Pro
820 825 830
Asn Leu Glu Lys Met Gly Tyr Asp Ile Lys Asp Ser Leu Leu Val Phe
835 840 845
<210> 9
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Томат; NIA; 523-533
<400> 9
Leu Lys Lys Ser Ile Ser Thr Pro Phe Met Asn
1 5 10
<210> 10
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Табак; NIA1; 518-528
<400> 10
Leu Lys Lys Ser Ile Ser Thr Pro Phe Met Asn
1 5 10
<210> 11
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Табак; NIA2; 518-528
<400> 11
Leu Lys Lys Ser Ile Ser Thr Pro Phe Met Asn
1 5 10
<210> 12
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петуния; NIA; 522-532
<400> 12
Leu Lys Lys Ser Ile Ser Thr Pro Phe Met Asn
1 5 10
<210> 13
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тыква; NIA; 530-540
<400> 13
Leu Lys Lys Ser Val Ser Thr Pro Phe Met Asn
1 5 10
<210> 14
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Береза; NIA1; 515-525
<400> 14
Leu Lys Lys Ser Val Ser Thr Pro Phe Met Asn
1 5 10
<210> 15
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Резуховидка; NIA1; 532-542
<400> 15
Leu Lys Lys Ser Val Ser Ser Pro Phe Met Asn
1 5 10
<210> 16
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Резуховидка; NIA2; 529-539
<400> 16
Leu Lys Lys Ser Val Ser Thr Pro Phe Met Asn
1 5 10
<210> 17
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рапс; NIA1; 526-536
<400> 17
Leu Lys Lys Ser Val Ser Ser Pro Phe Met Asn
1 5 10
<210> 18
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рапс; NIA2; 526-536
<400> 18
Leu Lys Lys Ser Val Ser Thr Pro Phe Met Asn
1 5 10
<210> 19
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Соя; NIA2; 504-514
<400> 19
Leu Lys Lys Ser Val Ser Ser Pro Phe Met Asn
1 5 10
<210> 20
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Фасоль обыкновенная; NIA1; 502-512
<400> 20
Leu Lys Lys Ser Val Ser Thr Pro Phe Met Asn
1 5 10
<210> 21
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Фасоль обыкновенная; NIA2; 500-510
<400> 21
Leu Lys Lys Ser Val Ser Ser Pro Phe Met Asn
1 5 10
<210> 22
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Lotus japonicus; NIA; 508-518
<400> 22
Leu Lys Lys Ser Val Ser Ser Pro Phe Met Asn
1 5 10
<210> 23
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Цикорий; NIA1; 521-531
<400> 23
Leu Lys Lys Ser Val Ser Ser Pro Phe Met Asn
1 5 10
<210> 24
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Маис; NIA1; 234-244
<400> 24
Leu Lys Arg Ser Thr Ser Thr Pro Phe Met Asn
1 5 10
<210> 25
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Ячмень; NIA1; 524-534
<400> 25
Leu Lys Arg Ser Thr Ser Thr Pro Phe Met Asn
1 5 10
<210> 26
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Ячмень; NIA2; 521-531
<400> 26
Leu Lys Arg Ser Thr Ser Thr Pro Phe Met Asn
1 5 10
<210> 27
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рис; NIA1; 527-537
<400> 27
Leu Lys Arg Ser Thr Ser Thr Pro Phe Met Asn
1 5 10
<210> 28
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шпинат; NIA; 538-548
<400> 28
Leu Lys Arg Thr Ala Ser Thr Pro Phe Met Asn
1 5 10
<---
Реферат
Изобретение относится к области биохимии, в частности к табачному материалу, обладающему уменьшенными уровнями табак-специфичных нитрозаминов (TSNA). Также раскрыт способ получения табачного материала, обладающего уменьшенными уровнями табак-специфичных нитрозаминов (TSNA). Изобретение позволяет эффективно получать табачный материал, обладающий уменьшенными уровнями табак-специфичных нитрозаминов. 2 н. и 22 з.п. ф-лы, 16 ил., 10 табл., 7 пр.
Комментарии